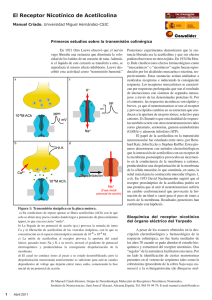
La insulina inicia su acción
al interaccionar con su
receptor
específico
ubicado en la membrana
celular. Este receptor es
una glucoproteína que
pertenece a la familia de
receptores para factores
de
crecimiento
con
actividad específica de
quinasas de Tirosina. El receptor de la insulina es tetramérico, es decir, consta de cuatro
subunidades; dos subunidades Alfa (α) y dos subunidades Beta (β); las subunidades α son
periféricas y por ende son las que interaccionan con la insulina. Tal interacción induce un
cambio conformacional en dichas subunidades, que a su vez permite que una molécula de ATP
se enlace a las subunidades β y así se active la propiedad tirosina quinasa intrínseca que
caracteriza al receptor de insulina. La actividad tirosina quinasa permite que una subunidad β
fosforile los residuos de tirosina de la subunidad β continua, y viceversa dándose así un
mecanismo de fosforilación cruzada. Una vez fosforilados la subunidad β de los receptores de
la insulina, interaccionan con una proteína citoplasmática llamada Sustrato de receptor de
insulina (IRS-1) mediante un dominio PTB; dicho sustrato es fosforilado por las mismas
subunidades β del receptor de insulina, específicamente en los residuos de tirosina. El IRS-1 a
su vez actúa con otra proteína, la Fosfoinositol 3 quinasa (PI3K), la cual es un heterodímero
que consta de una subunidad reguladora y de una subunidad catalítica. Las subunidades
reguladoras son proteínas adaptadoras que contienen dos dominios SH2, los cuales permiten
su unión a las proteínas IRS-1. La interacción entre ambas proteínas provoca cambios
alostéricos en la conformación de la subunidad reguladora dando por resultado la activación
de la subunidad catalítica de PI3K y por ser éste una quinasa tendrá la capacidad de fosforilar
otros compuestos. La PI3K, fosforila al lípido de membrana Fosfoinositol 4,5 bifosfato (PIP2) y
es transformado en Fosfoinositol 3, 4, 5 trifosfato (PIP3). Éste último estando fosforilado, es
reconocido por el dominio PH de una proteína quinasa dependiente de fosfoinositoles (PDK);
dicha interacción induce un cambio conformacional en la PDK que permite su activación. La
PDK, estando activa, puede actuar sobre la proteína Proteinquinasa B (PKB). Dicha proteína
quinasa B se encuentra interaccionando con una molécula de PIP3 por lo cual se encuentra
anclada a la membrana plasmática y adyacente a la PDK, así esta ultima fosforilara a la PKB
induciéndole un cambio conformacional que permitirá su desacoplamiento de la membrana y
su activación encargándose de dversos mecanismos como: favorecer la translocación del
citoplasma a la membrana plasmática de la célula de vesículas que contienen transportadores
de glucosa GLUT-4 en tejidos como el adiposo y el muscular, permitiendo así el incremento de
glucosa en el medio intracelular; activando fosfodiesterasas que convertirán el AMPc en AMP
lineal y; activando enzimas con actividad fosfatasa para que éstas desfosforilen compuestos
según se requieran en período postpandrial.
 0
0