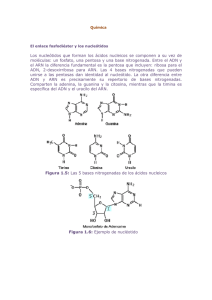
ESTRUCTURA DE LOS ÁCIDOS NUCLÉICOS Módelo de la Doble Hélica Difracción de Rayos X: ADN-B J. Watson y F. Crick Composición de los Ácidos nucleicos. Proporciones de las Bases Nitrogenadas: Reglas de Chargaff (1950). El Modelo de la Doble Hélice: Watson y Crick (1953). Alternativas al Modelo de la Doble Hélice. Propiedades físico-químicas de los ácidos nucleicos. Densidad de los Ácidos Nucleicos. Desnaturalización: Temperatura de Fusión. Absorbancia a 260 nm. Cinética de la Renaturalización: Curvas Cot. Hibridación de los ácidos nucleicos. Secuenciación del ADN: Método Didesoxi y Método Automático. COMPOSICIÓN QUÍMICA DE LOS ÁCIDOS NUCLEICOS Miescher en 1871 aisló del núcleo de las células de pus una sustancia ácida rica en fósforo que llamó "nucleína". Un año más tarde, en 1872, aisló de la cabeza de los espermas del salmón un compuesto que denominó "protamina" y que resultó ser una sustancia ácida y otra básica. El nombre de ácido nucleico procede del de "nucleína" propuesto por Miescher. Cuando se realiza la hidrólisis completa de los ácidos nucleicos, se obtienen tres tipos de componentes principales: Azúcar, en concreto una pentosa. Bases nitrogenadas: púricas y pirimidínicas. Ácido fosfórico. El azúcar, en el caso de los ácidos desoxirribonucleicos (ADN) es la 2-desoxi-D-ribosa y en el caso de los ácidos ribonucleicos (ARN) es la D-ribosa. Pentosas Ácido fosfórico Las bases nitrogenadas que forman parte de los ácidos nucleicos son de dos tipos, púricas y pirimidínicas. Las bases púricas derivadas de la purina (fusión de un anillo pirimidínico y uno de imidazol) son la Adenina (6-aminopurina) y la Guanina (2-amino-6-hidroxipurina). Las bases pirimidínicas (derivadas de la pirimidina) son la Timina (2,6-dihidroxi-5-metilpirimidina o también llamada 5-metiluracilo), Citosina (2-hidroxi-6-aminopirimidina) y Uracilo (2,6-dihidroxipirimidina). Las bases nitrogenadas que forman normalmente parte del ADN son: Adenina (A), Guanina (G), Citosina y Timina (T). Las bases nitrogenadas que forman parte de el ARN son: Adenina (A), Guanina (G), Citosina (C) y Uracilo (U). Por tanto, la Timina es específica del ADN y el Uracilo es específico del ARN. Bases Púricas Bases Pirimidínicas Además de las bases nitrogenadas anteriormente descritas, se han encontrado otras bases nitrogenadas en algunos virus o formando parte de algunos tipos especiales de ARNs. Ejemplos de algunas de estas bases púricas poco corrientes son: Hipoxantina, Xantina, 2-metiladenina, 6metil-aminopurina. Entre las bases pirimidínicas podríamos citar la 5-metilcitosina (propia del ADN) y la 5-hidroximetil citosina (HMC) que sustituye a la citosina en los fagos T-pares. En los ARN transferentes (ARN-t) que intervienen en el proceso de traducción de proteínas se encuentran la Ribotimidina, Dihidrouridina, Seudouridina e Inosina (I). La unión de la base nitrogenada a la pentosa recibe el nombre de nucleósido y se realiza a través del carbono 1’ de la pentosa y los nitrógenos de las posiciones 3 (pirimidinas) o 9 (purinas) de las bases nitrogenadas mediante un enlace de tipo N-glucosídico. La unión del nucleósido con el ácido fosfórico se realiza a través de un enlace de tipo éster entre el grupo OH del carbono 5’ de la pentosa y el ácido fosfórico, originando un nucleótido. Los nucleótidos son las unidades o monómeros utilizados para construir largas cadenas de polinucleótidos. Nucleósido = Pentosa + Base nitrogenada. Nucleótido = Pentosa + Base nitrogenada + Ácido fosfórico. Polinucleóotido = Nucleótido + Nucleótido + Nucleótido + .... Nucleótido Tanto los nucleótidos como los nucleósidos pueden contener como azúcar la D-ribosa (ribonucleótidos y ribonucleósidos) o la pentosa 2-desoxi-D-ribosa (desoxirribonucleótidos y desoxirribonucleósidos). Además, los nucleótidos pueden tener 1, 2 ó 3 grupos fosfato unidos al carbono 5’ de la pentosa, existiendo por tanto, nucleótidos 5’ monofosfato, nucleótidos 5’ difosfato y nucleótidos 5’ trifosfato. En algunos casos el ácido fosfórico se une a la pentosa por el carbono 3’, existiendo nucleótidos 3’ monofosfato, difosfato o trifosfato según el número de grupos fosfato que posea. La terminología empleada para referirse a los nucleósidos y nucleótidos es la siguiente: Base Nitrogenada Nucleósido Nucleótido Adenina Adenosina Ácido Adenílico Guanina Guanidina Ácido Guanílico Citosina Citidina Ácido Citidílico Timina Timidina Ácido Timidílico Uracilo Uridina Ácido Uridílico Los nucleótidos se unen entre si para formar largas cadenas de polinuclóetidos, esta unión entre monómeros nucleótidos se realiza mediante enlaces fosfodiéster entre los carbonos de las posiciones 3’ de un nucleótido con la 5’ del siguiente. Polinucleótido PROPORCIONES DE LAS BASES NITROGENADAS: REGLAS DE CHARGAFF Al principio se pensaba que los ácidos nucleicos eran la repetición monótona de un tetranucleótido, de forma que no tenían variabilidad suficiente para ser la molécula biológica que almacenara la información. Sin embargo, Chargaff (1950) demostró que las proporciones de las bases nitrogenadas eran diferentes en los distintos organismos, aunque seguían algunas reglas. Estas reglas de Chargaff se cumplen en los organismos cuyo material hereditario es ADN de doble hélice y son las siguientes: REGLAS DE CHARGAFF PARA ADN DE DOBLE HÉLICE La proporción de Adenina (A) es igual a la de Timina (T). A = T . La relación entre Adenina y Timina es igual a la unidad (A/T = 1). La proporción de Guanina (G) es igual a la de Citosina (C). G= C. La relación entre Guanina y Citosina es igual a la unidad ( G/C=1). La proporción de bases púricas (A+G) es igual a la de las bases pirimidínicas (T+C). (A+G) = (T + C). La relación entre (A+G) y (T+C) es igual a la unidad (A+G)/(T+C)=1. Edwin Chargaff Sin embargo, la proporción entre (A+T) y (G+C) era característica de cada organismo, pudiendo tomar por tanto, diferentes valores según la especie estudiada. Este resultado indicaba que los ácidos nucleicos no eran la repetición monótona de un tetranucleótido. Existía variabilidad en la composición de bases nitrogenadas. En la siguiente tabla se observan las proporciones de las bases nitrogenadas en algunos organismos. Procedencia del ADN A G C T 5-Me-C Timo de Bovino 28,2 21,5 21,2 27,8 1,3 Esperma de bovino 28,7 22,2 20,7 27,3 1,3 Germen de trigo 27,3 22,7 16,8 27,1 6,0 Saccharomyces 31,3 18,7 17,1 32,9 - Escherichia coli 26,0 24,9 25,2 23,9 - Mycobacterium tuberculosis 15,1 34,9 35,4 14,6 - ØX174 24,3 24,5 18,2 32,3 - T3 23,7 26,2 27,7 23,5 - T5 30,3 19,5 19,5 30,8 - T7 32,4 18,3 32,4 17,0 HMC A G Virus ARN C U Mosaico del tabaco (TMV) 29,8 25,4 18,5 26,3 Mosaico amarillo nabo 22,6 17,2 38,0 22,2 Poliomielitis 28,6 24,0 22,0 25,4 Encéfalo miocarditis del ratón 27,3 23,5 23,2 25,9 Reovirus Tipo 3 28,0 22,3 22,0 27,9 Tumor de las heridas 31,1 18,6 19,1 31,3 De la observación de la tabla anterior pueden extraerse las siguientes conclusiones: Todos los ADN estudiados cumplen la relación A=T y G=C, excepto el ADN del bacteriofago ØX174. El ADN de este virus es de una sola hélice. En los virus ARN no se cumple la equimolaridad de las bases excepto en el caso del virus del Tumor de las heridas y de los Reovirus que tienen ARN de doble hélice. En estos virus se cumple que A=U y G=C, además se cumple que A+G/U+C=1. El fago T2 y los otros fagos T-pares (T4 y T6) en vez de citosina tienen hidroximetil-citosina (HMC). Algunos organismos tiene en su ADN una pequeña proporción de 5-metil-citosina (5-Me-C) que sustituye a la citosina. Igualmente, en la siguiente tabla puede observarse como la proporción A+T/G+C varia de un organismo a otro. Organismo Tejido A+T/G+C Escherichia coli - 1,00 Diplococcus pneumoniae - 1,59 Mycobacterium tuberculosis - 0,42 Levadura - 1,79 Paracentrolus lividus (erizo mar) Esperma 1,85 Arenque Esperma 1,23 Médula ósea 1,33 Hombre Timo 1,52 Hombre Hígado 1,53 Hombre Esperma 1,52 Rata Por tanto, la proporción A+T/G+C es específica de cada organismo y como veremos más adelante cuando hablemos de las propiedades físico- químicas de los ácidos nucleicos, dicha proporción está relacionada con la densidad y la temperatura de fusión. EL MODELO DE LA DOBLE HÉLICE: WATSON Y CRICK (1953) Una vez demostrado que los ácidos nucleicos eran los portadores de la información genética, se realizaron muchos esfuerzos encaminados a determinar su estructura con exactitud. Watson y Crick (1953) fueron los primeros investigadores en proponer una estructura para los ácidos nucleicos y su labor investigadora se vio recompensada con el Premio Nobel en 1962, Premio Nobel que compartieron con M. H. F. Wilkins y que se les concedió por sus descubrimientos en relación con la estructura molecular de los ácidos nucleícos y su significación para la transmisión de la información en la materia viva.. Para realizar su trabajo emplearon dos tipos de datos ya existentes. Francis H. C. Circk James D. Watson Maurice H. F. Wilkins Por un lado, utilizaron los datos obtenidos varios años antes por Chargaff (1950), relativos a la composición de bases nitrogenadas en el ADN de diferentes organismos. El otro tipo de datos eran los procedentes de estudios de difracción de rayos X sobre fibras de ADN. Para determinar la estructura tridimensional o disposición espacial de las moléculas de ADN, se hace incidir un haz de rayos X sobre fibras de ADN y se recoge la difracción de los rayos sobre una película fotográfica. La película se impresiona en aquellos puntos donde inciden los rayos X, produciendo al revelarse manchas. El ángulo de difracción presentado por cada una de las manchas en la película suministra información sobre la posición en la molécula de ADN de cada átomo o grupo de átomos. Mediante esta técnica de difracción de rayos X se obtuvieron los siguientes resultados: Las bases púricas y pirimidínicas se encuentran unas sobre otras, apiladas a lo largo del eje del polinucleótido a una distancia de 3,4 Å. Las bases son estructuras planas orientadas de forma perpendicular al eje (Astbury, 1947). El diámetro del polinucleótido es de 20 Å y está enrollado helicoidalmente alrededor de su eje. Cada 34 Å se produce una vuelta completa de la hélice. Existe más de una cadena polinucleotídica enrollada helicoidalmente (Wilkins et el. 1953, Frankling y Gosling, 1953). Difracción de Rayos X: ADN-B Rosalin Franklin Basándose en estos dos tipos de datos Watson y Crick propusieron su Modelo de estructura para el ADN conocido con el nombre de Modelo de la Doble Hélice. Las características del Modelo de la Doble Hélice son las siguientes: El ADN es una doble hélice enrollada helicoidalmente “a derechas” (sentido dextrorso). Algo parecido a dos muelles entrelazados. Enrollamiento de tipo plectonémico: para separar las dos hélices es necesario girarlas como si fuera un sacacorchos. Módelo de la Doble hélice: ADN-B J. Watson y F. Crick Cada hélice es una serie de nucleótidos unidos por enlaces fosfodiéster en los que un grupo fosfato forma un puente entre grupos OH de dos azúcares sucesivos (posiciones 3’ de un azúcar y 5’ del siguiente). Las dos hélices se mantienen unidas mediante puentes o enlaces de hidrogeno producidos entre las bases nitrogenadas de cada hélice. Siguiendo los datos de Chargaff (1959), la Adenina de una hélice aparea con la Timina de la hélice complementaria mediante dos puentes de hidrógeno. Igualmente, la Guanina de una hélice aparea con la Citosina de la complementaria mediante tres puentes de hidrógeno. Par A-T Par G-C Pares A-T y G-C Las dos hélices por razones de complementaridad de las bases nitrogenadas son antiparalelas, teniendo secuencias de átomos inversas. Una hélice lleva la secuencia 5’P → 3’ OH , mientras que la hélice complementaria sigue la secuencia de átomos 3’OH → 5’P. El diámetro de la doble hélice es de 20 Å. Las bases nitrogenadas son estructuras planas perpendiculares al eje de la doble hélice y están apiladas unas sobre otras a una distancia de 3,4 Å. Cada 10 bases, cada 34 Å se produce una vuelta completa de la doble hélice (360º). Las bases se encuentran en sus configuraciones cetónicas, cumpliendo así las reglas de apareamiento A-T y G-C. La secuencia de bases nitrogenadas puede ser cualquiera, no existe ninguna restricción. Además, la estructura en doble hélice propuesta por Watson y Crick (1953) sugería varías propiedades importantes del material hereditario: Las reglas de complementaridad de las bases nitrogenadas A-T y G-C sugieren un forma sencilla de replicación del material hereditario. Esta forma sencilla de replicación se denomina método Semiconservativo. Cuando el ADN se replica sus dos hélices se separan y cada una de ellas sirve de molde para sintetizar una nueva hélice siguiendo las reglas de apareamiento de las bases nitrogenadas. La mutación a nivel molecular consistiría en un cambio en la secuencia de bases nitrogenadas del ADN. Al no existir ninguna restricción en la secuencia de bases nitrogenadas, el ADN poseía la suficiente variabilidad como para ser el material hereditario. Además, esta estructura sugería la existencia de algún código que permitiera pasar de la secuencia lineal de bases nitrogenadas en el ADN a la secuencia lineal de aminoácidos en las proteínas. ALTERNATIVAS AL MODELO DE LA DOBLE HÉLICE El modelo de la Doble Hélice propuesto por Watson y Crick está basado en estudios del ADN en disolución (hidratado). La denominada forma B ó ADN-B tiene un mayor interés biológico ya que es la que presenta el ADN en interacción con las proteínas nucleares. Además de la forma B, existen otras estructuras posibles que puede presentar el ADN. Algunas de estas alternativas son las siguientes: ADN-B: ADN en disolución, 92% de humedad relativa, se encuentra en soluciones con baja fuerza iónica se corresponde con el modelo de la Doble Hélice. ADN-A: ADN con 75% de humedad, requiere Na, K o Cs como contraiones, presenta 11 pares de bases por giro completo y 23 Å de diámetro. Es interesante por presentar una estructura parecida a la de los híbridos ADN-ARN y a las regiones de autoapareamiento ARN-ARN. ADN-C: ADN con 66% de humedad, se obtiene en presencia de iones Li, muestra 9+1/3 pares de bases por giro completo y 19 Å de diámetro. ADN-Z: doble hélice sinistrorsa (enrollamiento a izquierdas), 12 pares de bases por giro completo, 18 Å de diámetro, se observa en segmentos de ADN con secuencia alternante de bases púricas y pirimidínicas (GCGCGC), debido a la conformación alternante de los residuos azúcar-fosfato sigue un curso en zig-zag. Requiere una concentración de cationes superior a la del ADN-B, y teniendo en cuenta que las proteínas que interaccionan con el ADN tienen gran cantidad de residuos básicos sería posible que algunas convirtieran segmentos de ADN-B en ADN-Z. Las posiciones N7 y C8 de la Guanina son más accesibles. ADN-A ADN-B ADN-Z ADN-A ADN-Z ADN-B ADN con enrollamiento paranémico: Las dos hélices se pueden separar por traslación, cada hélice tiene segmentos alternantes dextrorsos y sinistrorsos de unas cinco bases. Uno de los principales problemas del modelo de la doble hélice (ADN-B) es el enrollamiento plectonémico, para separar las dos hélices es necesario girarlas como un sacacorchos, siendo necesario un gran aporte energético. ADN triple hélice o ADN-H: "In vitro" es posible obtener tramos de triple hélice intercalando oligonucleótidos cortos constituidos solamente por pirimidinas (timinas y citosinas) en el surco mayor de una doble hélice. Este oligonucleótido se une a pares de bases A-T y G-C mediante enlaces de hidrógeno tipo Hoogsteen que se establecen entre la T o la C del oligonucleótido y los pares A-T y G-C de la doble hélice. No se sabe la función biológica del ADN-H aunque se ha detectado en cromosomas eucarióticos. Triple hélice: pirimidinas Triple hélice: purinas Triple Hélice ADN cuadruplexo: "In vitro" se han obtenido cuartetos de Guanina (ADN cuadruplexo) unidas mediante enlaces tipo Hoogsteen, empleando polinucleótidos que solamente contienen Guanina (G). Los extremos de los cromosomas eucarióticos (telómeros) tienen una estructura especial con un extremo 3' OH de cadena sencilla (monocatenario) en el que se repite muchas veces en tandem una secuencia rica en Guaninas. Se piensa que el ADN cuadruplexo telomérico serviría para proteger los extremos cromosómicos de la degradación enzimática. Ejemplo de secuencia telomérica rica en guaninas (G): 5´P TTGGGTTGGGGTTGGGG...............TTGGGG 3'OH Cuartetos de Guanina Además, de las alternativas anteriormente citadas es necesario tener en cuenta que no todos los organismos vivos tienen como material hereditario ADN de doble hélice, algunos virus tienen ADN de hélice sencilla, ARN de una y de doble hélice. Palíndromos: plegamiento o apareamiento de una hélice consigo misma. El palíndromo también es una figura gramatical que se lee igual en los dos sentidos, por ejemplo: DABALE ARROZ A LA ZORRA EL ABAD. Existe ADN palindrómico de hélice sencilla y de hélice doble. En el palíndromo de doble cadena la secuencia de bases se lee igual en dirección 5’ P→ 3’OH en ambas cadenas. Secuencias palindrómicas Palíndromos en ADN de una y doble hélice Existen algunos virus cuyos ácidos nucleicos son de una sola hélice: el ADN de los fagos ØX174 y M13 es circular de hélice sencilla, sin embargo, se ha comprobado que una pequeña parte resiste la acción de enzimas que digieren específicamente ADN de hélice sencilla. Por tanto, estas regiones resistentes de ADN presentan complementaridad interna o autoapareamiento, formando ADN duplex o doble hélice, teniendo secuencias palíndrómicas. Una situación semejante se ha observado en el ARN de hélice sencilla del bacteriofago MS2 (3569 ribonucleotidos y tres genes). En este caso, el gen de la proteína de la cubierta del virus presenta segmentos que tienen complementaridad interna (autoapareamiento) que se pliegan sobre si mismos formando horquillas (regiones de ARN de doble hélice). ARN Fago MS2: proteína de la cápside El ARN transferente (ARN-t) que transporta los aminoácidos y el ARN ribosómico (ARNr) que intervienen en el proceso de traducción, son ácidos ribonucleicos de una sola hélice y también presentan autoapareamiento. Esquema ARN-transferente Esquema ARN-ribosómico de Tetrahymena PROPIEDADES FÍSICO-QUÍMICAS DE LOS ÁCIDOS NUCLEICOS Las principales propiedades físico-químicas de los ácidos nucleicos que vamos a considerar son las siguientes: Densidad de los ácidos nucleicos. Desnaturalización de los ácidos nucleicos: Temperatura de fusión (Tm). Absorbancia a 260 nm. Cinética de Renaturalización: Curvas Cot. Hibridación de los ácidos nucleicos. DENSIDAD DE LOS ÁCIDOS NUCLEICOS Densidad: existe una relación lineal entre el contenido en G+C y la densidad del ADN determinada en un gradiente de densidad. A mayor contenido en G+C mayor densidad posee el ADN. Cuanto mayor es el contenido en (G+C) mayor es la densidad. Meselson y col. (1957) desarrollaron una técnica de centrifugación en gradiente de densidad. Cuando se centrifuga a alta velocidad un solución densa (saturada) de un soluto de bajo peso molecular (ClCs 7,7 M, sucrosa, etc.) se produce un equilibrio entre dos fuerzas opuestas, la de difusión del soluto y la fuerza de sedimentación, como consecuencia se produce un gradiente de densidad que aumenta en la dirección de la fuerza centrifuga (aumenta a medida que avanzamos hacia el fondo del tubo de centrifuga). Si añadimos al centrifugar una molécula de ADN, está migrará hacia el fondo del tubo hasta llegar al punto en que la densidad del soluto de bajo peso molecular (la densidad del ClCs) coincide con la densidad del ADN centrifugado (densidad de flotación). Posteriormente, es posible aislar las moléculas de ADN de diferente densidad practicando un orificio en el fondo del tubo y sacando varias fracciones diferentes. También es posible pinchar el tubo con una jeringa, justo en la posición de la banda y extraer el ADN contenido de esa zona del tubo. Centrifugación en gradiente de CsCl Basándose en múltiples estudios de la densidad de los ADNs de diferentes organismos y de su composición en bases nitrogenadas, se ha establecido una fórmula empírica que relaciona la densidad de flotación (ρ) con el contenido en G+C expresado en moles por ciento. Está fórmula es la siguiente: ρ = 1,660 + 0,00098(G+C). DESNATURALIZACIÓN: TEMPERATURA DE FUSIÓN Desnaturalización: la proporción A+T/C+G está relacionada en primer lugar con la estabilidad de la molécula de ADN de doble hélice. Cuanto mayor es el contenido en G+C de una molécula, mayor cantidad de pares G-C presentará, como consecuencia tendrá una mayor cantidad de triples enlaces y, por consiguiente, será necesario suministrar una mayor cantidad de energía a esa doble hélice para separar sus dos hebras (desnaturalización o fusión del ADN). Cuanto mayor es el contenido en G+C mayor cantidad de calor que hay que suministrar a un ADN de doble hélice para desnaturalizarlo. La temperatura de fusión (Tm) necesaria para desnaturalizar la mitad del ADN de una mezcla (punto medio de la reacción ADN doble hélice ® ADN hélice sencilla) esta directamente relacionada con el contenido en G+C, a mayor contenido en G+C mayor temperatura de fusión (Tm). Relación entre el contenido en (G+C) y Tm Curvas de desnaturalización de diferentes ADNs ABSORBANCIA A 260 nm Absorbancia a 2.600 Å: El estado físico de los ácidos nucleicos está relacionado con su capacidad de absorción de la luz ultravioleta (UV) a 2.600 Å. El menor grado de absorción se produce en estado de doble hélice, la absorción aumenta cuando se produce la desnaturalización pasando a estado de hélice sencilla (efecto hipercrómico, aumento de la absorbancia) y, por último, si degradamos este ADN de hélice sencilla a nivel de nucleótidos libres, de nuevo aumenta la absorbancia. Esquema efecto hipercrómico Por tanto, la absorbancia a 2.600 Å se puede utilizar como una medida del estado físico de la molécula de ADN. Las curvas de fusión tienen forma de S observándose un aumento brusco de la absorbancia al llegar a una temperatura determinada, la temperatura de fusión. CINÉTICA DE RENATURALIZACIÓN: CURVAS CoT Velocidad de renaturalización: la velocidad de renaturalización del ADN de un organismo está relacionada con su complejidad. La complejidad se define como la suma del número de nucleotidos que tiene cada tipo de secuencia sin tener en cuenta el número de veces que esta repetida. El ADN de los virus y las bacterias en su mayoría sólo tiene secuencias que están una sola vez en el genoma (secuencias únicas). Sin embargo, los organismo más complejos, como los eucariontes, presentan distintos tipos de secuencias en su genoma. Los eucariontes tienen secuencias únicas (SU), secuencias de bajo número de copias (SBNC, 2-10 copias), secuencias repetidas (SR, alrededor de 102 copias) y altamente repetidas (SAR, 103 a 106 copias). Tipo de Secuencia Nº de repeticiones Nº de nucleótidos A (SU) 1 5.700 B (SU) 1 7.530 C (SBNC) 4 3.720 D (SBNC) 6 4.350 E (SR) 200 500 F (SAR) 10.000 300 G (SAR) 1.000.000 200 Complejidad 22.300 Si imaginamos un organismo eucarionte con los tipos de secuencias indicados en la tabla anterior, su complejidad sería la suma del número de nucleótidos de cada tipo de secuencia (22.300) sin tener en cuenta el número de veces que está repetida cada una de ellas. No es difícil imaginar que la velocidad de renaturalización del ADN (paso de dos hélices sencillas a una doble hélice) este relacionada con el número de veces que esta repetida una determinada secuencia. Supongamos que tenemos dos organismos hipotéticos distintos, ambos con la misma cantidad de ADN (1.000.000 de pares de nucleótidos). El organismo A posee 1.000 secuencias únicas diferentes, cada una con una longitud media de 1.000 nucleótidos. El organismo B tiene una secuencia de 100 nucleótidos de longitud que está repetida 10.000 veces. Si extraemos el ADN de estos dos organismos por separado, lo desnaturalizamos y las piezas de ADN desnaturalizado de cada organismo se ponen en condiciones de renaturalizar (tamaño uniforme de los fragmentos de ADN, entre 250 y 450 pb, temperatura, condiciones iónicas, etc), es evidente que el ADN del organismo B renaturalizará mucho antes, más rápidamente, que el del organismo A, ya que es mucho más probable que la secuencia que está repetida 10.000 veces encuentre otra complementaria en el medio de reacción para formar la doble hélice. Las curvas de renaturalización o curvas Cot, de organismos que carecen de secuencias repetidas como virus y bacterias, tienen forma de S, tienen un sólo punto de inflexión o punto medio de la reacción. Todas las secuencias son únicas y tienen la misma probabilidad de encontrar a su complementaria para formar la doble hélice. Curvas Cot de virus y bacterias Curvas Cot de un eucarionte Las curvas Cot de organismos eucariontes con diferentes tipos de secuencias, poseen varios puntos de inflexión. Cada uno de ellos correspondiente a un tipo de secuencias (únicas, moderadamente repetidas, y altamente repetidas). Como en el caso de las curvas de fusión, también se utiliza como medida el punto medio de la reacción de renaturalización, es decir, el Cot ½ o tiempo al que se ha reasociado la mitad del ADN. El Cot ½ es directamente proporcional a la complejidad del ADN del organismo. Valores de Cot ½ bajos (10-4 a 10-1) corresponden a secuencias altamente repetidas, valores de Cot ½ comprendidos entre 10 y 102 corresponden a secuencias moderadamente repetidas y las secuencias únicas o de bajo número de copias dan lugar a valores de Cot ½ altos (mayores de 103) . HIBRIDACIÓN DE LOS ÁCIDOS NUCLEICOS Las técnicas de desnaturalización y posterior renaturalización se utilizan para conseguir la hibridación de ácidos nucleicos, es decir, una vez separadas las dos hebras del ADN doble hélice, es posible volver a formar una doble hélice con una hebra de ADN que sea complementaria (hibridación de ADN con ADN) o volver a formar una doble hélice con un segmento de ARN que contenga la secuencia complementaria (hibridación de ADN con ARN). Endonucleasas de restricción Las técnicas de hibridación (desnaturalización y posterior renaturalización) permiten, por ejemplo, identificar el gen que ha dado lugar a un determinado ARN-mensajero. Si se aísla un mensajero determinado en una célula, es posible hibridar este ARN con el ADN de la célula previamente fragmentado mediante el uso de endonucleasas de restricción. Las endonucleasas de restricción de tipo II son enzimas que reconocen secuencias cortas de ADN (de 4 a 6 pares de bases) de tipo palindrómico y cortan por el interior de la secuencia diana a la misma altura en las dos hélices de ADN (corte simétrico) o a distinto nivel en cada hélice (corte asimétrico). Eco RI Hae III Dichas endonucleasas de restricción fueron descubiertas por Werner Arber, Hamilton O. Smith y Daniel Nathans en la bacteria E.coli , trabajos por los que recibieron el premio Nobel. Estas enzimas forman parte del sistema de defensa (sistema de modificación-restricción) de las bacterias frente a la entrada de ADN exógeno en su interior. Existen cientos de endonucleasas diferentes que reconocen distintas secuencias cortas de tipo palíndrómico. En la siguiente tabla se indican algunas de las endonucleasas más frecuentes: Bacteria de la que procede Secuencia que reconoce (5'→ 3') y lugar de corte (¯) Eco RI Escherichia coli 5' G¯AATTC 3' (asimétrico) Eco RII Escherichia coli 5' ¯CCTGG 3' (asimétrico) Hae III Haemophilus aegyptus 5' GG¯CC 3' (simétrico) Hin dII Haemophilus influenzae 5' GTPi¯Pu AC 3' (simétrico) Hin dIII Haemophilus influenzae 5' A¯AGCTT 3' (asimétrico) Hpa I Haemophilus parainfluenzae 5' GTT¯AAC 3' (simétrico) Hpa II Haemophilus parainfluenzae 5' C¯CGG 3' (asimétrico) Ava I Anabaena variabilis 5' CGPu¯PiCG 3' (simétrico) Endonucleasa restricción de La utilización de diferentes endonucleasas y la separación por tamaños de los fragmentos producidos mediante electroforesis permite construir lo que se denomina Mapas de restricción. Cuando el ADN de un organismo eucarionte se fragmenta con una endonucleasa de restricción, se producen cientos de miles o incluso hasta millones de fragmentos. Dichos fragmentos se separan por tamaños mediante electroforesis en geles de agarosa, una vez se parados por tamaños se transfieren los fragmentos a una membrana de nylon en condiciones desnaturalizantes y de forma que permanezcan en la misma posición. Los fragmentos desnaturalizados y pegados a la membrana se pueden renaturalizar o hibridar utilizando una segmento de ADN o de ARN (habitualmente denominado sonda) marcado radiactivamente. Dicha sonda, hibridará solamente con aquellos fragmentos de ADN que contengan la secuencia complementaria. La técnica que permite transferir los fragmentos de ADN desnaturalizados a una membrana de nylon y posteriormente hibridar con una sonda de ADN marcada se denomina Técnica Southern (hibridación ADN-ADN), cuando la hibridación se realiza con una sonda de ARN marcada se denomina Northern (hibridación ADN-ARN). El empleo combinado de endonucleasas de restricción y de diferentes sondas de ADN de secuencia única marcadas permite obtener Polimorfismos para Longitudes de Fragmentos de Restricción (RFLPs), muy utilizados en la construcción de mapas de ligamiento. Hibridación "in situ" Las técnicas de hibridación "in situ" permiten localizar un segmento concreto de ADN (por ejemplo un gen) en una posición determinada de un cromosoma eucariótico. Es posible obtener preparaciones citológicas de cromosomas en metafase mitótica o en otras fases del ciclo, desnaturalizar el ADN de los cromosomas en la propia preparación citológica y posteriormente hibridar con la pieza de ADN deseada marcada. Actualmente, el marcaje suele ser de tipo fluorescente, denominándose dicha técnica Hibridación "in situ" mediante fluorescencia (abreviadamente FISH). También, es posible marcar todo el ADN de un genomio o juego de cromosomas completo y realizar una Hibridación "in situ" Genómica (abreviadamente GISH) que permite averiguar si los cromosomas de un determinado genomio están presentes. Otra aplicación, consiste en detectar determinados cromosomas o regiones cromosómicas concretas utilizando sondas o piezas de ADN marcadas mediante fluorescencia y procedentes solamente del cromosoma deseado, de esta manera, se obtiene lo que se denomina Pintura Cromosómica. FISH trigo (Triticum intermedium) GISH: Híbrido (Liliaceae) Pintura cromosómica (ratón) Pintura cromosómica (humanos) SECUENCIACIÓN DEL ADN: MÉTODO DIDESOXI Y MÉTODO AUTOMÁTICO El análisis más detallado de la estructura del ADN consiste en averiguar la secuencia de nucleótidos. A lo largo del tiempo se han desarrollado diferentes métodos para obtener la secuencia de nucleótidos del ADN, sin embargo, actualmente los métodos más utilizados son el de secuenciación automática y el método enzimático de terminación de cadena de Sanger también conocido por el método didesoxi. Método enzimático de terminación de cadena o método didesoxi de Sanger Para obtener la secuencia de bases nitrogenadas de un segmento de ADN por el método enzimático de terminación de cadena, se necesitan los siguientes compuestos: El ADN molde o segmento de ADN que se desea secuenciar. Para poder secuenciar un segmento de ADN, previamente se necesita tener gran cantidad de ese fragmento, y por tanto, hay que clonarlo en un vector apropiado. Además, debe estar en estado de hélice sencilla. Un enzima que replique el ADN, normalmente la ADN Polimerasa I del bacteriofago T4. La ADN Polimersa I del fago T4 emplea como molde ADN de hélice sencilla y siguiendo las reglas de complementaridad de las bases nitrogenadas va añadiendo nucleótidos a partir de un cebador o "primer". Un cebador o "primer" que suele ser un oligonucleótido corto de alrededor de 20 bases de longitud necesario para que la ADN polimerasa I comience a añadir nucleótidos por el extremo 3' OH. Este cebador debe poseer una secuencia de bases complementaria a la del fragmento de ADN que se desea secuenciar. Debido a que la secuencia de nucleótidos del segmento que se quiere secuenciar es desconocida, se emplea un "primer" con secuencia complementaria al vector empleado para clonar el fragmento de ADN, además, este cebador procede de una región del vector muy cercana al punto de inserción del ADN problema cuya secuencia se conoce. El "primer" utilizado suele marcarse radiactivamente. Los cuatro nucleótidos trifosfato (dATP, dCTP, dGTP y dTTP). A veces en vez de marcar radiactivamente el cebador, se marca radiactivamente uno de los cuatro nucleótidos trifosfato en cada reacción. Por último, se necesitan nucleótidos didesoxi (ddATP, ddTTP, ddCTP y ddGTP). Los nucleótidos didesoxi son nucleótidos modificados que han perdido el grupo hidroxilo de la posición 3' de la desoxirribosa. Estos nucleótidos pueden incorporarse a la cadena de ADN naciente, pero no es posible que se una a ellos ningún otro nucleótido por el extremo 3'. Por tanto, una vez incorporado un nucleótido didesoxi se termina la síntesis de la cadena de ADN. Breve descripción del método enzimático de terminación de cadena En primer lugar, deben realizarse en cuatro tubos diferentes, cuatro mezclas de reacción. Cada mezcla de reacción contiene los cuatro nucleótidos trifosfato (dATP, dCTP, de dTTP y dGTP), ADN polimerasa I, un cebador marcado radiactivamente y un nucleótido dideoxi, por ejemplo ddATP, a una concentración baja. El nucleótido didesoxi utilizado (ddATP en este ejemplo) competirá con su homólogo (dATP) por incorporarse a la cadena de ADN que se está sintetizando, produciendo la terminación de la síntesis en el momento y lugar donde se incorpora. Por este sistema, en cada mezcla de reacción se producen una serie de moléculas de ADN de nueva síntesis de diferente longitud que terminan todas en el mismo nucleótido y marcadas todas radiactivamente por el extremo 5' (todas contienen en el extremo 5' el cebador utilizado). Los fragmentos de ADN de nueva síntesis obtenidos en cada mezcla de reacción se separan por tamaños mediante electroforesis en geles verticales de acrilamida muy finos (0,5 mm de espesor) y de gran longitud (cerca de 50 cm) que permiten distinguir fragmentos de ADN que se diferencian en un solo nucleótido.Los productos de cada una de las cuatro mezclas de reacción se insertan en cuatro calles o carriles diferentes del gel. Una vez terminada la electroforesis, el gel se pone en contacto con una película fotográfica de autorradiografía. La aparición de una banda en una posición concreta de la autorradiografía en una de las cuatro calles nos indica que en ese punto de la secuencia del ADN de nueva síntesis (complementario al ADN molde) está la base correspondiente al nucleótido didesoxi utilizado en la mezcla de reacción correspondiente. Esquema de las electroforesis de las reacciones de secuenciación Autorradiografía Teniendo en cuenta que el ADN de nueva síntesis crece en la dirección 5' → 3', si comenzamos a leer el gel por los fragmentos de menor tamaño (extremo 5') y avanzamos aumentando el tamaño de los fragmentos (hacia 3'), obtendremos la secuencia del ADN de nueva síntesis en la dirección 5' → 3'. Breve descripción del método automático de secuenciación La principal diferencia entre método enzimático de terminación de cadena y el método automático de secuenciación radica, en primer lugar en el tipo de marcaje. En el método automático en vez de radiactividad se utiliza fluorescencia y lo habitual es realizar cuatro mezclas de reacción, cada una con nucleótido trifosfato (dTTP) marcado con un fluorocromo distinto. Este sistema permite automatizar el proceso de manera que es posible leer al mismo tiempo los ADNs de nueva síntesis producto de las cuatro mezclas de reacción. La segunda diferencia radica en el sistema de detección de los fragmentos de ADN. La detección del tipo de fluorescencia correspondiente a cada reacción se lleva a cabo al mismo tiempo que la electroforesis, de manera que los fragmentos de ADN de menor tamaño que ya han sido detectados se dejan escapar del gel, permitiendo este sistema aumentar el número de nucleótidos que se pueden determinar en cada electroforesis y, por consiguiente, en cada secuenciación. El siguiente esquema representa de forma abreviada el método automático de secuenciación. Esquema del método automático de secuenciación En la siguiente figura se muestra un ejemplo de una secuencia obtenida por el método automático de secuenciación. Cuando aparece la letra N significa que no ha sido posible determinar el nucleótido existente en esa posición de la secuencia. Secuencia obtenida por métodos automáticos