Mendez_Gastroenterologia_2a_c29_SECRECIONES
Anuncio

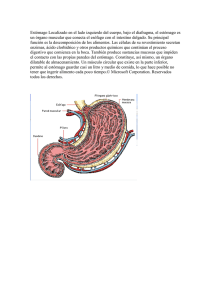
Sección VII Estómago 29. Anatomía del estómago: secreciones normales y anormales Norberto C. Chávez Tapia Nahum Méndez-Sánchez Misael Uribe 30. Dispepsia funcional Fernando Rojas Mendoza 31. Gastritis Graciela E. Castro Narro 32. Enfermedad ulceropéptica Francisco Javier Bosques Padilla Manuel Alejandro Martínez Vázquez 33. Síndrome de Zollinger-Ellison Jesús Kazuo Yamamoto-Furusho Nahum Méndez-Sánchez Norberto C. Chávez Tapia Misael Uribe 34. Cáncer gástrico Lizbeth López Carrillo Luis F. Oñate Ocaña 35. Tumores gástricos diferentes del adenocarcinoma Daniel Motola Kuba Estela Pérez Bustos Ricardo Sosa Sánchez Dan Green Renner Gabriela Alvarado Luna Capítulo 29 Anatomía del estómago: secreciones normales y anormales Norberto C. Chávez Tapia, Nahum Méndez-Sánchez, Misael Uribe Contenido Embriología • Anatomía • Histología • Secreciones gástricas constituye la curvatura menor. La rotación de este órgano hasta su orientación adulta concluye hacia la séptima semana. Embriología Anatomía El estómago forma parte del intestino anterior embrionario, además de la faringe, el aparato respiratorio inferior, el esófago, el duodeno hasta la entrada del colédoco, el hígado, el páncreas y el árbol biliar. Aparece hacia la cuarta semana de gestación y se observa como una dilatación de la parte distal del intestino anterior, suspendida entre los mesenterios dorsal y ventral. A medida que se prolonga, el borde dorsal se desarrolla con mayor rapidez, gira 90 grados hacia la izquierda y se transforma en la curvatura mayor. Por su parte, el borde ventral se vuelve hacia la derecha y El estómago se divide en zonas según los tipos de células mucosas presentes en su interior y las relaciones con las estructuras próximas. Como las características histológicas y fisiológicas de la mucosa gástrica varían con arreglo a su situación en el estómago, es importante tener un conocimiento general de las zonas anatómicas gástricas (fig. 29-1). Así, por ejemplo, el antro no contiene células secretoras de ácido, mientras que el cuerpo posee gran Porción cervical Esófago Porción torácica Hígado Porción abdominal Estómago Bazo Ángulo cólico izquierdo Colon transverso Vesícula biliar Colon transverso Colon descendente Yeyuno Íleon Colon sigmoide Recto Colon ascendente Orificio y válvula ileocecal Ciego Apéndice vermiforme Ano Figura 29-1. Vista general del tracto digestivo, desde la boca hasta el ano. 293 294 Sección VII Estómago Escotadura cardial Esófago Curvatura menor Fondo A. coronaria estomáquica (gástr. izq.) Rama esofágica A. gástricas cortas Bazo A. esplénica A. esplénica Incisura angular Cuerpo T. celiaco A. hepática Epiplón menor A. pilórica (gástr. der.) Curvatura mayor A. gastroduodenal Epiplón mayor Duodeno Píloro A. gastroepiploica izquierda Antro pilórico Figura 29-2. Vista anterior de la cara externa del estómago. A. supraduodenal número de células parietales y el fórnix tiene pocas. Las células productoras de gastrina abundan en el antro, pero son escasas en el fórnix y el cuerpo. Estas consideraciones adquieren importancia tanto quirúrgica como en el tratamiento de la enfermedad ulcerosa péptica. La zona cardial es la parte del estómago adyacente a la unión esofagogástrica. Su borde cefálico está bien delimitado en su interior por la transformación del epitelio escamoso en glándulas gástricas. El fondo o fórnix es la parte del estómago que se localiza por encima de una línea horizontal que va desde la unión gastroesofágica a la curvatura mayor. El fondo carece de límite anatómico apreciable a simple vista y el histológico tampoco está bien definido. El cuerpo del estómago se encuentra entre el fondo y un punto de la curvatura menor denominado incisura angular. El borde distal del fondo es inconstante y cambia con la edad, así que es inexistente a simple vista una limitación anatómica precisa del borde corporo-antral (fig. 29-2). El antro empieza donde termina el cuerpo y se prolonga hasta el píloro (fig. 29-3). La unión del píloro con el duodeno se encuentra marcada por una incisura externa y también por una vena constante (de Mayo) que lo atraviesa a este nivel. Orificio cardial del estómago Esfínter pilórico Conducto pilórico Antro pilórico Crestas (pliegues) longitudinales Figura 29-3. Vista anterior de la cara interna del estómago. Figura 29-4. A. gastroepiploica derecha Arterias del estómago y el bazo. Riego arterial El riego sanguíneo arterial del esófago distal, del estómago y del duodeno proximal tiene lugar, sobre todo, a través de ramas procedentes del tronco celiaco (fig. 29-4). Sin embargo, el riego arterial de los órganos abdominales superiores es muy variable y la imposibilidad de descubrir una vascularización anómala puede llevar a cometer errores fatales en el planteamiento quirúrgico de la resección del estómago, el duodeno o el páncreas. Por lo regular, el tronco celiaco da origen a tres ramas: la coronaria estomáquica o gástrica izquierda, la esplénica y la hepática, aunque se debe decir que esta descripción anatómica normal sólo se presenta en 55% de la población. El diámetro del propio tronco puede variar entre pocos milímetros y 2 cm. Existe la posibilidad de que el tronco celiaco sea incompleto, es decir que parte del riego sanguíneo hepático proceda de la mesentérica superior (12%) o de las arterias gástricas izquierdas (23%). Además, el tronco celiaco puede no dar origen a la gástrica izquierda (3.5%), a la arteria hepática (5.5%) o a la arteria esplénica (1.5%). La fuente principal del riego sanguíneo del estómago es el tronco celiaco. Existen cuatro arterias principales: las arterias gástricas izquierda y derecha, que corren por la curvatura menor, y las arterias gastroepiploicas izquierda y derecha, que corren por la curvatura mayor. Además, un gran aporte sanguíneo del estómago proximal proviene de la arteria frénica inferior y de las arterias gástricas cortas (o vasos cortos), que son ramas de la esplénica. La arteria gástrica izquierda puede dar origen a la arteria hepática izquierda. Microcirculación El riego arterial del estómago procede de los vasos de mediano tamaño que ya se mencionaron, cuyas paredes constan de una túnica muscular, de la íntima y la adventicia. Al llegar al órgano Capítulo 29 penetran en la serosa, se ramifican y continúan por debajo de ella; en su momento, cada una de estas ramas ingresan en la pared muscular hasta la submucosa, sin enviar nuevos vasos a la muscular. En la submucosa se forma un extenso plexo capilar que dirige sus ramificaciones hacia la capa muscular y la mucosa. Esta rica red vascular existe en todo el estómago, excepto a lo largo de la curvatura menor. Aquí, los vasos que atraviesan la pared muscular dan origen a otras ramas, que no dan origen a la amplia red capilar submucosa. Este hecho se propone como causa de las úlceras situadas a lo largo de la pared gástrica. Aunque se ha estimado su tamaño con base en cálculos experimentales en animales por medio de microesferas, son estudios que por el momento están sin confirmación en los seres humanos. Sin embargo, por métodos indirectos se demostró la existencia de anastomosis, que son importantes en términos anatómicos y fisiológicos para la regulación del riego gástrico. Drenaje venoso El drenaje venoso del estómago es similar en su distribución al riego arterial. Las venas gástricas izquierda y derecha drenan hacia la vena porta. La vena gastroepiploica derecha drena a la vena mesentérica superior y la vena gastroepiploica izquierda drena a la vena esplénica. Las venas viscerales carecen de válvulas; por tanto, ofrecen una buena fuente de riego sanguíneo colateral cuando se obstruye cualquier porción del sistema porta. Drenaje linfático Se demostró la existencia de cuatro zonas gástricas, que drenan grupos ganglionares linfáticos concretos. Los ganglios que se distribuyen alrededor de la arteria gástrica izquierda drenan el esófago distal, el cardias, la curvatura menor hasta la incisura angular y una parte del fondo muy próxima al cardias (zona I). El drenaje de la zona II se origina en la parte antral de la curvatura menor y el conducto pilórico. Los ganglios que drenan esta zona acompañan a la arteria gástrica derecha. La zona III corresponde a la parte proximal de la curvatura mayor, donde se distribuyen los ganglios gastroepiploicos izquierdos, que se vacían en los ganglios linfáticos esplénicos. La zona IV corresponde a la parte distal de la curvatura mayor distal hasta el píloro, donde se localizan los ganglios linfáticos gastroepiploicos derechos. Estos últimos drenan en los ganglios subpilóricos, que son posteriores al píloro. Toda la linfa del estómago se vacía por último en los ganglios celiacos, que reciben también la linfa procedente del hígado, el bazo y el páncreas. La linfa de los ganglios celiacos drena a través del conducto torácico en el sistema venoso a la altura del cuello, en el punto donde se unen las venas yugular interna y subclavia izquierdas. Por esta razón, gran parte de las afecciones malignas abdominales, en particular del estómago, se pueden manifestar con una metástasis ganglionar linfática supraclavicular izquierda (ganglio de Virchow o Ewald). Inervación El estómago tiene inervación tanto parasimpática (nervio vago) como simpática (plexo celiaco). El nervio vago se origina en Anatomía del estómago: secreciones normales y anormales 295 núcleos localizados en el piso del cuarto ventrículo y atraviesa el cuello para entrar al mediastino, donde se divide en varias ramas alrededor del esófago. Estas ramas se vuelven a unir a la altura del hiato esofágico para formar el nervio vago derecho (posterior) e izquierdo (anterior). La gran mayoría de las fibras vagales son aferentes y conducen estímulos del tubo digestivo hacia el encéfalo. Las fibras vagales eferentes se originan en el núcleo dorsal del vago, localizado en el bulbo, y realizan sinapsis con neuronas de los plexos mientérico y submucoso. Estas neuronas utilizan acetilcolina como neurotransmisor, influenciando las funciones motoras y secretoras gástricas. Por otro lado, el aporte simpático procede de sus núcleos torácicos que se extienden entre T5 a T10, cuyas fibras hacen sinapsis en la cadena homónima y desde allí nacen fibras que conforman los nervios esplácnicos mayor y menor que se dirigen hacia los ganglio celiacos. Las fibras posganglionares siguen a las arterias para inervar el estómago. El sistema nervioso entérico se encuentra conformado por los plexos autónomos de Auerbach y Meissner. En estos plexos existen neuronas colinérgicas, serotoninérgicas y peptidérgicas, cuyo papel persiste sin comprenderse por completo. Musculatura La musculatura del estómago está constituida por tres capas: interna, media y externa. La capa muscular más interna es incompleta y más densa en el fondo que en el antro. Las fibras musculares descienden de manera oblicua desde el fondo hasta la curvatura mayor; su ausencia en la curvatura menor crea una depresión longitudinal que se denomina magenstrasse. Las fibras musculares internas se mezclan con las circulares medias en la curvatura mayor. Las fibras internas se unen a la capa muscular circular del esófago en la unión gastroesofágica. La capa circular media es la única completa de toda la pared gástrica. Sus fibras van desde la curvatura menor hasta la mayor y aumentan en número desde el fórnix hasta el antro. Las fibras circulares se concentran más en el antro y se encargan de una de las funciones antrales más importantes: el vaciamiento gástrico. El mecanismo de la “bomba antral” y el esfínter pilórico ayudan a establecer la velocidad del vaciamiento gástrico, en particular de los alimentos sólidos. La capa muscular longitudinal externa del estómago es la continuación de las fibras musculares longitudinales del esófago. Éstas se prolongan de manera distal desde el esófago, en gran parte a lo largo de las curvaturas menor y mayor, lo que deja buena parte de las paredes anterior y posterior del estómago con una relativa carencia de fibras musculares longitudinales. Las fibras de las curvaturas menor y mayor convergen en la región del píloro y forman una vaina circular, que se continúa con la capa muscular longitudinal del duodeno. Histología La pared gástrica está constituida por mucosa, submucosa, muscular propia y serosa. A su vez tiene dos compartimientos, 296 Sección VII Estómago la superficie foveolar y el compartimiento glandular. El compartimiento foveolar es la capa más superficial que limita la luz gástrica y se encuentra formada por células epiteliales (células foveolares) que contienen gránulos de mucina. En un plano más profundo se encuentran las células mucosas del cuello, que cuentan con una menor cantidad de mucina y son las progenitoras de las células epiteliales de la superficie. El compartimiento glandular varía en relación con su localización topográfica. Las glándulas del cardias contienen únicamente células secretoras de moco; las glándulas oxínticas (fondo y cuerpo) contienen células parietales, células principales y algunas células endocrinas; las glándulas pilóricas (o antrales) contienen células endocrinas y células secretoras de moco. Las características de cada tipo celular se muestran en el cuadro 29-1. Secreciones gástricas La regulación de la secreción ácida del estómago implica mecanismos complejos, así como diversas vías neuronales, hormonales y de señalización paracrina que actúan de forma directa en la célula parietal y de forma indirecta al modular la secreción de gastrina y agentes de acción paracrina como la histamina y la somatostatina. La gastrina es el principal estimulante de la secreción ácida durante la ingesta de alimento. Se libera a partir de las células G del antro y su efecto primordial se debe a la liberación de histamina a partir de células parecidas a las enterocromafines. Después, la histamina se une a los receptores H2 en la membrana basolateral de la célula parietal, lo que resulta en un incremento de las concentraciones de cAMP; el contexto anterior desencadena el flujo de cloro e hidrógeno desde la superficie apical de la célula a través de los canales del cloro y bajo la acción de la ATP-asa de H+-K+. Una vez que se activa por los secretagogos (como gastrina, histamina y acetilcolina), la ATP-asa de H+-K+ se transloca desde las vesículas tubulares del citosol hacia la superficie apical de la célula, lo que resulta en la secreción ácida hacia la luz gástrica. La secreción gástrica de ácido facilita la digestión de proteínas, la absorción del hierro, calcio, vitamina B12 y previene el sobrecrecimiento bacteriano y las infecciones entéricas. Regulación central de la secreción de ácido El núcleo motor dorsal del vago en el bulbo raquídeo y los núcleos paraventriculares en el hipotálamo tienen un papel preponderante en la integración de las vías aferentes y eferentes. La estimulación vagal inducida por la exposición al frío o por la inyección intracisternal de hormona liberadora de tirotropina activa las neuronas colinérgicas en el estómago, con el consecuente incremento de la secreción gástrica ácida. Otro mecanismo importante que participa en la regulación del ácido conlleva la activación de los receptores transitorios potenciales vaniloides (transient receptor potential vanilloid, TRPV) tipo 1 y se observa que la inyección intraventricular de capsaicina o el endocanabinoide anandamida estimulan la secreción gástrica de ácido mediante activación vagal. Regulación periférica de la secreción ácida En la regulación de la secreción ácida participan gran variedad de péptidos en la mucosa gástrica (gastrina, péptidos liberadores de gastrina, grelina, orexina, péptido natriurético auricular y leptina), transmisores de tipo gaseoso (óxido nítrico) y los nutrimentos que se ingieren. Estimulantes Los elementos estimulantes incluyen gastrina, péptidos liberadores de gastrina, grelina, orexina y L-aminoácidos. Gastrina Ésta se sintetiza en las células G del antro y favorece la secreción ácida, en particular por la liberación de histamina a partir de células parecidas a las enterocromafines que se localizan en el cuerpo Cuadro 29-1. Localización y función de los tipos celulares gástricos Células Localización Función Parietal Cuerpo Secreción de ácido y factor intrínseco Mucosas Cuerpo y antro Producción de moco Principales Cuerpo Secreción de pepsina Epiteliales Difusa Secreción de moco, bicarbonato y prostaglandinas Tipo enterocromafín Cuerpo Histamina G Antro Gastrina D Cuerpo y antro Somatostatina Interneuronas gástricas mucosas Cuerpo y antro Péptido liberador de gastrina Neuronas entéricas Difusa Péptido relacionado con el gen de la calcitonina, otros Endocrinas Cuerpo Grelina Capítulo 29 Anatomía del estómago: secreciones normales y anormales 297 y el fondo gástrico. La gastrina también promueve el crecimiento de la mucosa. Diversas neoplasias son sensibles a la acción de la gastrina, como es el caso de los tumores carcinoides, así como de los cánceres de estómago, colon, páncreas y pulmón. Los receptores activados de la proliferación de peroxisomas (PPAR, por sus siglas en inglés) son receptores nucleares que en fechas recientes demostraron su importancia en la secreción gástrica ácida al ser agonistas directos de las células G, aunque su papel es incierto. Péptido natriurético auricular El péptido natriurético auricular se localiza en las células enterocromafines de la mucosa gástrica. El porcentaje de producción es mayor en el fondo, intermedio en el antro y en menor grado en el cuerpo gástrico. La producción endógena de péptido natriurético auricular estimula la secreción de somatostatina en el antro y fondo gástricos. En el antro, la estimulación de somatostatina se vincula con la inhibición de la secreción de gastrina, y en el fondo, con la de histamina. Péptidos liberadores de gastrina Los péptidos liberadores de gastrina son neuropéptidos que se encuentran en la mucosa gástrica antral y que, como su nombre lo indica, estimulan la secreción de gastrina. Éstos se sintetizan a partir de una gran molécula precursora que es convertida en pro-GRP1-125, el cual es procesado en dos productos terminales: el GRP1-27 y el GRP18-27. Leptina La leptina se ubica en particular en las células principales. Los receptores de leptina se localizan a lo largo de todo el intestino delgado tanto en la membrana apical como basolateral de los enterocitos. La degradación de la leptina que se secreta a la luz gástrica está protegida por una proteína que la une a su correspondiente dominio extracelular del receptor de leptina. Cuando se encuentra en la luz gástrica, su principal función es estimular al estómago de manera indirecta a través del vago, mientras que en el intestino actúa a través de la membrana apical de los enterocitos. Grelina En los humanos, el gen que codifica a la grelina se localiza en el cromosoma 3, en el locus 3p25-26, y consiste en cuatro exones y tres intrones. La proteína madura se codifica en los exones 1 y 2, mientras que la transcripción del gen se procesa por corte y empalme en un sitio alterno, lo cual permite la producción de dos diferentes mRNA maduros y, por último, de dos péptidos: la grelina y la des-Gln14-grelina, los cuales conservan una gran homología entre diversas especies de mamíferos. El precursor de la grelina (prepro-grelina) se compone de 117 aminoácidos (y carece de Gln en su posición 14), comparte homología con el precursor de la motilina, lo que supone un proceso ontogénico similar. Hasta el momento se desconoce con precisión si tiene un efecto directo en la secreción gástrica de ácido. Orexina La orexina A y la orexina B son péptidos hipotalámicos que incrementan la ingesta; se unen a dos receptores relacionados tipo proteína G a los que activan. En ratas, se demostró que al antagonizar este sistema se inhibe la secreción ácida estimulada por pentagastrina y la secreción basal, lo que sugiere su papel en la fisiología gastrointestinal. Aminoácidos Los L-aminoácidos (L-fenilalanina, L-triptófano, L-leucina) estimulan la secreción de gastrina a partir de las células G y la secreción ácida a partir de las células parietales, quizás a través de la activación de receptores sensibles al calcio. La activación de estos receptores activa de forma secundaria a la ATP-asa de H+-K+ de modo independiente. Inhibidores Los mecanismos periféricos inhibidores incluyen el péptido natriurético auricular, leptina, péptidos derivados del glucagon y óxido nítrico. Péptidos derivados de proglucagon Los cambios postranscripcionales de diversos genes dan como resultado el péptido parecido al glucagon tipo 1 y tipo 2, glicentina y oxitomodulina. Estos péptidos se pueden secretar de forma conjunta en respuesta a la ingesta de nutrimentos para inhibir la secreción gástrica. Óxido nítrico El óxido nítrico, un gas sintetizado a partir de L-arginina, funciona como neurotransmisor, mensajero de señalización intracelular o agente paracrino. Estudios recientes con animales indican que sus efectos son contradictorios, aunque al parecer disminuye la secreción ácida inducida por histamina, lo cual se acompaña de un incremento del cGMP en las células parietales. Secreción gástrica de pepsinógeno Los jugos gástricos de los vertebrados contienen proteasas, las cuales intervienen en la digestión de las proteínas provenientes de la dieta. Se clasifican en cuatro grupos, con base en sus propiedades inmunoquímicas o enzimáticas: pepsina A, pepsina B, pepsina C y quimosina. Las pepsinas A, B y C se encuentran en los jugos gástricos de los vertebrados adultos, mientras que la quimosina se observa sólo en neonatos. Como miembros de la clase de proteinasas aspárticas, estas proteinasas gástricas tienen una actividad óptima con pH ácidos, las inhibe la pepestatina y poseen dos residuos activos de aspartato que deben ser catalizados. Las formas cimógenas de la pepsina A, pepsina B, gastricsina y quimosina se conocen como pepsinógeno A, pepsinógeno B, progastricsina y proquimosina, respectivamente. Los cimógenos gástricos se sintetizan y almacenan en las células principales de la mucosa gástrica. Luego de la ingestión de 298 Sección VII Estómago alimentos, los cimógenos se secretan hacia el lumen del estómago, donde se convierten en sus respectivas formas activas a causa del pH ácido de la luz gástrica. La porción activa de la enzima del cimógeno consta de dos dominios. Los cimógenos comienzan a transformarse en su forma activa con un pH de 5.0; esta dependencia del pH se debe a las interacciones electrostáticas entre los residuos básicos en los prosegmentos y los residuos ácidos en la porción activa de la enzima. Es importante señalar que la tasa de activación de los cimógenos gástricos depende del pH, la temperatura, la fuerza de unión iónica y la concentración del cimógeno. Estudios in vitro en las células principales indican que los agentes que incrementan el cAMP en la célula principal aumentan la secreción de pepsinógeno (secretina, péptido intestinal vasoactivo, prostaglandina E2 e isoproterenol); dicha secreción se puede estimular también por medio de agentes que elevan las concentraciones intracelulares de Ca2+ (carbacol, colecistocinina, leucotrieno B4). Los agentes que regulan las concentraciones de Ca2+ estimulan además la sintetasa de óxido nítrico al incrementarlo y, en conjunto, amplifican el efecto estimulante del Ca2+. Otros efectores son acetilcolina (vía receptores muscarínicos M3), colecistocinina (receptor CCKA) e histamina (receptor H2). En modelos in vivo, sustancias análogas a la acetilcolina, histamina, gastrina y secretina estimulan la secreción de pepsinógeno, mientras que la somatostatina la inhibe (fig. 29-5). Secreción gástrica de lipasa Además de las enzimas proteolíticas, se produce asimismo secreción de enzimas lipolíticas como la lipasa gástrica humana, enzima compuesta por 397 aminoácidos con propiedades que varían poco respecto de la lipasa pancreática; su pH óptimo es de H + ATP-asa de K+-H+ Célula parietal cAMP M3R Acetilcolina H2R CCC2R Gastrina Histamina ¿M1R? CCC2R HDC Células tipo enterocromafín Célula G Histidina Figura 29-5. Mecanismos moleculares que intervienen en la secreción acidogástrica. 5.4, la inhiben las micelas ácidas biliares y no requiere colipasa. La secreción de pepsinógeno estimula la secreción de lipasa. Secreción alcalina y mucosa La mucosa del estómago y el duodeno se encuentra cubierta por una capa continua y adherente de gel mucoso viscoso y elástico, el cual constituye una barrera física de baja permeabilidad a la pepsina u otras macromoléculas entre la luz y la superficie apical de las células. Este gel viscoso está formado por glucoproteínas (mucinas), las cuales constituyen de 3 a 5% de su peso, mientras que el 95% restante corresponde a agua con pequeñas cantidades de lípidos, ácidos nucleicos y proteínas que incluyen inmunoglobulinas. En el estómago (antro, fondo y cardias), el gen de mucina MUC-5 se aprecia en la superficie de las células epiteliales y el gen MUC-6 se encuentra en las células del cuello. Tanto en el estómago como en el duodeno existe evidencia demostrativa de que la capa de gel mucoso adherente y el bicarbonato secretado a partir del epitelio superficial proveen la primera línea de defensa en contra de la acidez luminal y de la pepsina. Al parecer, la capa de moco no es obstáculo para el transporte de ácido y pepsina proveniente de las criptas gástricas. Sin embargo, la secreción de hidrogeniones secretados por las células oxínticas dentro de las criptas gástricas parece atravesar pequeños canales dentro del gel que se forman a expensas de la presión hidrostática que impera en el interior de las glándulas gástricas. Más aún, se propone que el moco secretado por las células de las criptas gástricas transporta protones y pepsinógeno de las criptas hacia la luz. El espesor de la capa de moco varía de 80 a 280 μm, al margen de marcadas diferencias topográficas del epitelio superficial. El pH a través de la capa de gel mucoso se midió in vivo e in vitro y es tan bajo como 2.0 a 3.0 en la porción vecina a la luz, pero se incrementa de forma progresiva hasta alcanzar valores de pH cercanos al neutral en la superficie de las células epiteliales.14,15 La mayor resistencia al daño inducido por ácido y que mantiene el gradiente de pH durante los periodos de secreción ácida gástrica se debe quizá a la “marea alcalina”. Aunque la neutralización del ácido por el bicarbonato en la superficie mucosa provee la primera línea de protección contra el ácido, también intervienen otros mecanismos. La neutralización intracelular del H+ ocurre por medio del HCO3– que entra por el mecanismo cotransportador basolateral de NaHCO3 (NBC) y por la exportación de protones que tiene lugar mediante el intercambio de Na+/H+ (NHE-1); de esa manera se corrige la disminución del pH intracelular. La baja permeabilidad relativa de la membrana apical a los H+ y su baja conductancia en las vías paracelulares son mecanismos adicionales que protegen la superficie gástrica. La secreción alcalina por el fondo y el antro gástricos muestra características similares, que incluyen mecanismos idénticos en sensibilidad a factores inhibidores y estimulantes, lo que sugiere que la superficie de las células epiteliales (presente tanto en el fondo como en el antro) es el origen de este tipo de secreciones. La secreción se incrementa por el estímulo colinérgico (que Capítulo 29 acompaña por ejemplo al consumo de alimentos), por diversos tipos de PGE, y por la presencia de un pH luminal bajo, mientras que la secreción de HCO3– se inhibe por estímulo simpático mediado a través de los receptores adrenérgicos alfa-2. El intercambio de Cl–/HCO3– es el mecanismo principal para la salida de HCO–3 al gel mucoso. La recaptación celular de álcalis se encuentra mediada por cotransporte (NBC) de Na+-HCO3–. Como hecho interesante, tanto NBC-1 como NBC-2 se encuentran expresados en las células epiteliales de la superficie gástrica más aún que en las células parietales. Su presencia facilita la recaptación del HCO3– por las células epiteliales. El pH intraluminal, el sistema nervioso intrínseco y extrínseco, la producción local de eicosanoides y algunas hormonas son los principales factores que regulan la secreción de HCO3–. La influencia del estímulo simpático en la secreción de HCO3– es importante; la noradrenalina y la adrenalina inhiben la secreción de HCO3– y sus efectos pueden verse limitados por el uso de fentolamina, lo que sugiere que las células secretoras de HCO3– se regulan por medio de receptores adrenérgicos alfa. Sin embargo, la activación del sistema nervioso simpático libera neuropéptido Y así como noradrenalina. Estudios recientes muestran que su presencia inhibe la secreción estimulada por PGE2 y péptido intestinal vasoactivo. La administración de antagonistas del neuropéptido Y1 incrementa la secreción basal de HCO3–. Anatomía del estómago: secreciones normales y anormales 299 Secreción gástrica de Na+, K+ y Cl– Además de H+, otros dos cationes se encuentran en el jugo gástrico: Na+ y K+. La concentración de Na+ es inversamente proporcional a la de H+. Debido a que las células parietales no excretan sodio, es un excelente marcador de la secreción extraparietal. La secreción de potasio es de 2.5 a 4.5 veces su concentración en el plasma; aunque parte de este potasio proviene de la secreción extraparietal, la mayor parte procede de la secreción activa de las células parietales. El cloro se secreta de forma paralela al ácido y durante la fase activa de secreción alcanza valores superiores a los plasmáticos. Secreción de factor intrínseco El factor intrínseco es una glucoproteína que segregan las células parietales. Tanto los efectores estimulantes como inhibidores producen los mismos efectos en la secreción de factor intrínseco, aunque la duración de la respuesta secretora de factor intrínseco es más corta que la secreción ácida. Debido a los efectos del factor intrínseco en la absorción de la vitamina B12, que consisten en unirse a la misma para formar un complejo que permite su absorción en el íleon, su deficiencia provoca malabsorción de esta vitamina y condiciona anemia perniciosa. REFERENCIAS 1. Eisenberg M, Fondacaro P, Dunn D. Applied anatomy and anomalies of the stomach. En: Haubrich WS, Schaffner F, Berk JE (eds). Bockus gastroenterology. Philadelphia: WB Saunders, 1995:561-581. 2. Guyton AC, Hall JE. Gastrointestinal physiology. En: Guyton AC, Hall JE (eds). Textbook of medical physiology. Philadelphia: WB Saunders, 2000:718-769. 3. Aihara T, Nakamura E, Amagase K, Tomita K, Fujishita T, Furutani K, et al. Pharmacological control of gastric acid secretion for the treatment of acid-related peptic disease: past, present, and future. Pharmacol Ther, 2003;98:109-127. 4. Cohen SJ. Studies on the secretion of gastric juice. J Biol Chem, 1920;41: 257-261. 5. Hollander F. Studies in gastric secretion. II. A comparison of criteria of acidity used in the investigation. J Biol Chem, 1931;91:481-492. 6. Hollander F, Cowgill GR. Studies in gastric secretion. I. Gastric juice of constant acidity. J Biol Chem, 1931;91:151-182. 7. Hollander F. Studies in gastric secretion. IV. Variations in the chlorine content of gastric juice and their significance. J Biol Chem, 1932;97: 585-604. 8. Hubbard RS, Munford SA, Tyner J. The relationship between gastric secretion and the alkaline tide in urine. J Biol Chem, 1933;101:781-785. 9. Hollander F. Studies in gastric secretion. V. The composition of gastric juice as a function of its acidity. J Biol Chem, 1934;104:33-42. 10. Lindstrom E, Chen D, Norlen P, Andersson K, Hakanson R. Control of gastric acid secretion: the gastrin-ECL cell-parietal cell axis. Comp Biochem Physiol A Mol Integr Physiol, 2001;128:505-514. 11. Gardner JD, Ciociola AA, Robinson M. Measurement of meal-stimulated gastric acid secretion by in vivo gastric autotitration. J Appl Physiol, 2002;92:427-434. 12. Barocelli E, Ballabeni V. Histamine in the control of gastric acid secretion: a topic review. Pharmacol Res, 2003;47:299-304. 13. Richter C, Tanaka T, Yada RY. Mechanism of activation of the gastric aspartic proteinases: pepsinogen, progastricsin and prochymosin. Biochem J, 1998;335(Pt 3):481-490. 14. Flemstrom G, Isenberg JI. Gastroduodenal mucosal alkaline secretion and mucosal protection. News Physiol Sci, 2001;16:23-28. 15. Yang H. Central and peripheral regulation of gastric acid secretion by peptide YY. Peptides, 2002;23:349-358. 16. Hou W, Schubert ML. Gastric secretion. Curr Opin Gastroenterol, 2006;22:593-598.