Modificaciones de los principios Mendelianos Archivo
Anuncio

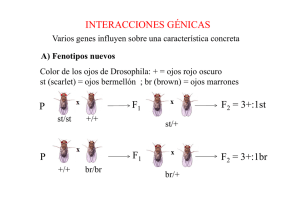
Unidad 11. MODIFICACION DE LOS PRINCIPIOS MENDELIANOS Documento elaborado con fines docentes por: GUSTAVO LOZANO CASABIANCA Biólogo M. Sc. Profesor asociado Escuela de Nutrición y Dietética Universidad de Antioquia YOENIS GARCÍA HERRERA Biólogo M. Sc. Profesor de cátedra Escuela de Nutrición y Dietética Universidad de Antioquia SEBASTIAN GARCÍA RESTREPO Estudiante Instituto de Biología Universidad de Antioquia CLARA I. ORTIZ RAMIREZ Bióloga Estudiante de maestría en Biología Universidad de Antioquia En el siglo XX, luego del redescubrimiento del trabajo de Mendel, las investigaciones fijaron su atención en las muchas formas en que los genes influyen en el fenotipo. Los tipos de herencia descritos en éste apartado se investigaron cuando los datos observados no se adecuaban con las proporciones mendelianas esperadas. Esto llevó a proponer hipótesis que modificaban y ampliaban los principios mendelianos y fueron comprobadas con cruces especiales (1). Dominancia incompleta y codominancia De manera contraria a los cruces mendelianos introducidos en la unidad anterior, un cruce entre padres con caracteres alternativos algunas veces pueden dar lugar a descendientes con fenotipo intermedio. Por ejemplo al realizar cruces de plantas de “boca de dragón” (Antirrhinum majus) con parentales de flores rojas y parentales de flores blancas, los descendientes expresan flores rosadas. Dado que ni el color rojo, ni el blanco es dominante, dicha situación suele denominase dominancia incompleta (Tabla 12) (1). Tabla 12. Dominancia incompleta. Ejemplo para el color de las flores de “boca de dragón” Genotipo Cruce (Fenotipo) R1R1x R2R2 P1 (roja) x (blanca) R1R2 F1 (rosa) F1 x F1 R1R2x R1R2 ¼ R1R1 (roja) ½ R1R2 F2 (rosa) ¼ R2R2 (blanca) Los casos bien definidos de dominancia incompleta, son relativamente raros. Sin embargo, aun cuando la dominancia completa sea aparente, un examen cuidadoso de los niveles del producto génico revela a menudo un nivel intermedio de la expresión génica (1). Por otra parte, si dos alelos de un único gen son responsables ambos de la expresión de determinado genotipo o fenotipo, puede surgir una situación diferente tanto de la dominancia/recesividad de alelos como de la dominancia incompleta. Tal es el caso de la expresión conjunta e independiente de ambos alelos en el heterocigoto la cual se denomina codominancia (1). Un ejemplo de codominancia está dado por el grupo sanguíneo MN. Se caracteriza por una glicoproteína de membrana de los eritrocitos que proporcionan inmunidad a los individuos. Hay dos formas de ésta glicoproteína: M y N. Un individuo puede presentar una de ella o las dos (Tabla 13). Tabla 13. Codominancia en el grupo sanguíneo MN Los alelos se denominan LM y LN Genotipo Fenotipo LMLM M LMLN MN LNLN N Alelos múltiples Si consideramos que un gen almacena mucha información, podemos entender que las mutaciones pueden afectar dicho gen de muchas maneras. Un solo cambio tiene el potencial de dar lugar a un alelo diferente al original. Por esto, para cualquier gen, el número de alelos presentes en los individuos de una población no necesariamente está limitado a dos. En ocasiones de en un mismo gen se encuentran tres o más alelos alternativos en la población, y por lo tanto se habla de la presencia de alelos múltiples, los cuáles dan lugar a un modo de herencia característico. Estos alelos múltiples solo pueden estudiarse en poblaciones. Cualquier individuo de un organismo diploide tiene, como mucho, dos loci génicos homólogos, que pueden estar ocupados por alelos diferentes del mismo gen. Sin embargo, en los miembros de una especie, puede haber muchas formas alternativas del mismo gen (1). El grupo sanguíneo ABO El caso más simple de alelos múltiples es aquel en el que hay tres alelos alternativos de un gen. Esta situación se presenta en la herencia del grupo sanguíneo ABO de la especie humana. El sistema ABO, como el grupo MN, se caracteriza por la presencia de antígenos en la superficie de los glóbulos rojos. Sin embargo, los antígenos A y B son distintos de los antígenos MN y se encuentran bajo el control de un gen diferente, localizado en el cromosoma 9 (1). El fenotipo ABO de cualquier individuo se averigua mezclando una muestra de sangre con antisuero que tenga los anticuerpos A o B, si hay antígeno en la superficie de los glóbulos rojos de dicha persona, reaccionará con el correspondiente anticuerpo se producirá la coagulación o aglutinación de los glóbulos rojos. Cuando se examinan los glóbulos rojos de un individuo de este modo, puede aparecer uno de los cuatro fenotipos. Cada individuo tiene o el antígeno A (fenotipo A), el antígeno B (fenotipo B), los antígenos A y B (fenotipo AB) o ninguno de ellos (fenotipo O) (1). En 1924 se propuso que la herencia de estos fenotipos dependía de tres alelos de un mismo gen. Esta hipótesis se basó en estudios de los grupos sanguíneos de muchas familias distintas. Aunque se pueden utilizar notaciones diversas, se utilizarán los símbolos: IA, IB, e IO para los tres alelos. La letra I se refiere a Isoaglutinógeno (otro término para antígeno) (1). Si se asume que los alelos IA y IB son responsables de la producción de los antígenos A y B respectivamente, y que el IO es un alelo que no produce ningún antígeno A o B detectable, se puede hacer una lista de los distintos genotipos posibles y de sus correspondientes fenotipos (Tabla 14). Tabla 14. Genotipos correspondientes a los fenotipos expresados en seres humanos. Genotipo Antígeno IA IA A IA IO A IB IB B Fenotipo A B IB IO B IA IB A, B AB IO IO Ninguno O Nótese en la tabla que los alelos IAeIB son dominantes sobre el alelo IO, pero codominantes entre sí. En la actualidad se acepta totalmente la hipótesis de que los grupos sanguíneos ABO en las poblaciones humanas están controlados por tres alelos (1). Herencia Poligénica Luego del redescubrimiento del trabajo de Mendel, diversos experimentos revelaron que muchos caracteres fenotípicos se encuentran, a menudo, bajo el control de más de un gen. Esto permitió entender que la influencia de los genes sobre el fenotipo es mucho más compleja que la sugerida por Mendel a partir de sus experimentos (1). En lugar de que un único gen controle el desarrollo de características concretas del organismo, quedó claro que caracteres como, por ejemplo, el color de los ojos, el color del pelo o la forma del fruto están influidos por muchos genes, lo que ahora es conocido como poligenia (1). Para describir éste, y otros fenómenos, se utiliza el término interacción génica (1). Este término se usa a menudo para describir la idea de que varios genes influyen sobre una característica concreta. Sin embargo, esto no significa que dos o más genes, o sus productos, interactúen necesariamente de manera directa para dar lugar a un fenotipo concreto, más bien, la función celular de numerosos productos génicos contribuyen al desarrollo de un fenotipo común. Por ejemplo, el desarrollo de un órgano como el ojo de un insecto es extraordinariamente complejo y da lugar a una estructura con manifestaciones fenotípicas múltiples, descritos de la manera más sencilla como una forma, tamaño, textura y color específicos (1). Epistasis Algunos de los ejemplos de interacción génica son los que presentan el fenómeno de la epistasia, la cual se presenta cuando la expresión de un gen enmascara o modifica la expresión de otro gen (1). Los genes implicados en la expresión de una misma característica fenotípica algunas veces lo hacen de una manera antagonista, como cuando ocurre enmascaramiento, y en otros casos ejercen su influencia de manera complementaria o cooperativa (1). Por ejemplo, la presencia en homocigosis de un alelo recesivo puede evitar, o anular, la expresión de otro alelo en un segundo locus; en este caso los alelos del primer locus se denominan epistáticos con respecto a los del segundo locus, que son enmascarados y se denominan hipostáticos, así por ejemplo los alelos que determinan el albinismo, los cuales siguen un patrón de herencia mendeliano recesivo simple son epistáticos respecto a los diferentes alelos que determinan la cantidad de melanina que produce un individuo en su piel (1). Pleiotropía La situación inversa a lo planteado anteriormente, consiste en los fenómenos donde la expresión de un solo gen tiene múltiples efectos fenotípicos. Para ilustrar éste caso podemos considerar el siguiente ejemplo: El Síndrome de Marfan es una enfermedad que se origina por una mutación autosómica dominante en el gen que codifica la proteína fibrilina del tejido conjuntivo. Dado que la proteína está ampliamente distribuida en muchos tejidos del organismo, se esperarían múltiples efectos a causa de dicha mutación (1). La fibrilina es importante para la integridad del cristalino y de los huesos, y para el revestimiento de los vasos sanguíneos. Por esto, una persona con el fenotipo asociado al Síndrome de Marfan puede sufrir dislocación de cristalino, aumento del riesgo de aneurismas aórticos y alargamientos de los huesos largos de las extremidades (1), otro ejemplo de pleiotropía es el albinismo en el cual un solo par de alelos tiene incidencia en la pigmentación de ojos, piel y cabello. Existen muchos otros ejemplos que podrían citarse. Basta decir que si se analizan cuidadosamente, la mayoría de las mutaciones presentan más de una manifestación cuando se expresan (1). Genes ligados Investigaciones realizadas a principios del siglo XX revelaron que ciertos genes no se transmitían de acuerdo con la ley de la transmisión independiente. En lugar de ello, parecía que dichos genes segregaban como si estuvieran ligados o unidos (1). Hoy sabemos que la mayoría de los cromosomas albergan un gran número de genes, y se dice que los genes que hacen parte del mismo cromosoma están ligados y evidencian ligamiento en los cruces. También, que los genes ligados no son libres de transmitirse independientemente debido a que la unidad de transmisión en la meiosis es el cromosoma, y no el gen (1). Herencia ligada al sexo En muchas especies animales y vegetales uno de los sexos posee un par de cromosomas diferentes que están implicados en la determinación del sexo. Por ejemplo en la especie humana, los varones tienen un cromosoma X y un cromosoma Y, mientras que las mujeres tienen dos cromosomas X. Aunque se reconocen ciertos genes específicos en el cromosoma Y humano, éste carece de copias de la gran mayoría de genes presentes en el cromosoma X. Es por esto que los genes situados en el cromosoma X presentan patrones únicos de herencia en comparación con los genes autosómicos, situación que se describe con el término ligamiento al cromosoma X o herencia ligada al sexo (1). La diferencia en los patrones de herencia resultantes de los genes presentes en X pero ausentes en Y, da lugar a una modificación de las proporciones mendelianas (1). Ligamiento al X en la especie humana En la especie humana se han reconocido muchos genes, junto con sus respectivos fenotipos, que están ligados al cromosoma X. Estos se pueden identificar en las genealogías debido a su patrón de herencia (1). Entre algunos genes ligados al cromosoma X que afectan la salud están los que controlan dos tipos de hemofilia; distrofia muscular, genes reguladores de enzimas tales como la glucosa-6-fosfato deshidrogenasa y la hipoxantin-guanosinfosforibosiltransferasa, cuya ausencia en el organismo se presenta en pacientes con el Síndrome de Lesch-Nyhan (caracterizado por disfunción neurológica, trastornos cognitivos y de conducta y aumento o sobreproducción de ácido úrico). Debido al modo en que se transmiten los genes ligados al X, patrones de herencia inusuales pueden ir asociadas con anomalías recesivas ligadas al X, cuando se las compara con anomalías autosómicas. Por ejemplo, si una anomalía ligada al X debilita o es letal para el individuo afectado antes de que se reproduzca, la anomalía se da mayoritariamente en los varones. Esto es así porque los alelos letales o deletereos permanecen en la población gracias a las mujeres heterocigóticas que son “portadoras” y que no manifiesta el rasgo. Ellas pasan el alelo a la mitad de sus hijos varones, que desarrollan la enfermedad debido a que son hemicigotos. Las mujeres heterocigotas también pasan el alelo a la mitad de sus hijas que, como sus madres, se convierten en portadoras, pero no desarrollan la enfermedad (1). Herencia limitada e influenciada por el sexo (el sexo del individuo influye en el fenotipo) En algunos casos, la expresión de un fenotipo concreto se encuentra estrictamente limitada a un sexo; en otros, el sexo de un individuo influye en la expresión de un fenotipo que no está limitado a uno u otro sexo. Esta distinción diferencia la herencia limitada por el sexo de la herencia influenciada por el sexo (1). En ambos tipos de herencia, los genes autosómicos son responsables de la existencia de fenotipos alternativos, pero la expresión de estos genes depende de la constitución hormonal del individuo. Así, el genotipo heterocigoto puede manifestar un fenotipo en los machos y el alternativo en las hembras. Por ejemplo, en aves de corral, la cola y las plumas del cuello son a menudo muy diferentes en hembras y en machos, demostrando herencia limitada por el sexo. El plumaje del gallo es más largo, curvado y puntiagudo que el de la gallina (1). Otro caso de herencia limitada por el sexo se refiere a los genes autosómicos responsables de la producción de leche en el ganado vacuno. Independientemente del genotipo que influye en la cantidad de leche producida, estos genes obviamente se expresan sólo en las vacas (1). Entre los casos de herencia influenciada por el sexo se encuentra el patrón de calvicie en la especie humana, la formación de los cuernos en ciertas razas de ovejas, y ciertos patrones de pelaje en el ganado vacuno. En tales casos, son genes autosómicos los responsables de los fenotipos alternativos manifestados por hembras y machos, pero la expresión de estos genes depende de la constitución hormonal de los individuos. Así, el genotipo heterocigoto puede manifestar un fenotipo en un sexo y el fenotipo alternativo en el otro (1). Por ejemplo, la calvicie en la especie humana, en donde el pelo es muy fino, o no hay, en la parte superior de la cabeza se hereda como se ilustra en la Tabla 15: Tabla 15. Influencia del sexo en la heredabilidad de la calvicie Fenotipo Genotipo hembra macho BB Calvo Calvo Bb No calvo Calvo bb No calvo No calvo Aun cuando las mujeres pueden presentar calvicie, este fenotipo es mucho más prevalente en los varones. Cuando las mujeres heredan el genotipo BB, el fenotipo es mucho menos pronunciado que en los varones y se expresa más tarde (1). Influencia del ambiente en el fenotipo La expresión fenotípica no siempre es el reflejo directo del genotipo. Hasta ahora se ha asumido que el genotipo de un organismo se expresa siempre directamente en su fenotipo, por ejemplo que los guisantes homocigotos para el alelo recesivo siempre serán cortos. Se ha tratado el tema de la expresión del gen como si los genes actuasen en un sistema cerrado en el que la presencia o ausencia de productos funcionales determina directamente el fenotipo global de un individuo (2). La situación es realmente mucho más compleja. La mayoría de los productos génicos actúan en el medio interno de la célula y las células interactúan unas con otras de varios modos. Además, el organismo debe sobrevivir ante diversas influencias ambientales. Así, la expresión del gen y el fenotipo resultante son modificados a menudo por la interacción entre el genotipo concreto del individuo y el ambiente interno y externo (2). El grado de influencia ambiental puede variar desde inapreciable hasta débil o muy elevado. Las interacciones débiles son mucho más difíciles de detectar y documentar, y conducen al conflicto “naturaleza-crianza” no resuelto, en el que los científicos debaten la importancia relativa de los genes frente al ambiente (2). Efectos de la temperatura Debido a que la actividad química depende de la energía cinética de las sustancias que reaccionan (que a su vez depende de la temperatura circundante), podemos esperar que la temperatura influya en los fenotipos. Así por ejemplo hay plantas, que produce flores rojas cuando se cultiva a 23°C y flores blancas a 18°C. Se dice que las mutaciones que están afectadas por la temperatura son condicionales y se denominan sensibles a la temperatura. Se conocen ejemplos en los virus y en una serie de organismos, como bacterias, hongos y Drosophila sp. En casos extremos, un organismo portador de un alelo mutante puede expresar un fenotipo mutante cuando se cultiva a una temperatura, pero expresan el fenotipo silvestre cuando se cultiva a otra temperatura. Este tipo de efecto es útil para estudiar mutaciones que interrumpen procesos esenciales en el desarrollo y por ello son normalmente perjudiciales para el organismo (2). Efectos de la nutrición Otro ejemplo en donde el fenotipo no es el reflejo directo del genotipo del organismo es el de las mutaciones nutricionales. En los microorganismos, son muy frecuentes las mutaciones que evitan la síntesis de moléculas nutritivas, como cuando se inactiva una enzima esencial de una ruta metabólica. Un microorganismo que lleve tal mutación se denomina auxótrofo. Si no se puede sintetizar el producto final de una ruta bioquímica, y si dicha molécula es esencial para el crecimiento y el desarrollo normales, la mutación impedirá el crecimiento y puede ser incluso letal (2). En la especie humana se conocen una serie de circunstancias ligeramente diferentes. La presencia o ausencia de ciertas sustancias en la dieta, que los individuos normales pueden consumir sin perjuicios, pueden afectar gravemente a individuos con constituciones genéticas anormales. A menudo, una mutación puede evitar que un individuo metabolice una sustancia que normalmente se encuentra en la dieta. Por ejemplo, los afectados por el trastorno genético fenilcetonuria no pueden metabolizar el aminoácido fenilalanina. Aquellos con galactosemia no pueden metabolizar la galactosa. Sin embargo, si se reduce drásticamente, o se elimina, la ingestión de la molécula correspondiente en la dieta, el fenotipo asociado puede mejorarse (2). El caso, bastante corriente, de la intolerancia a la lactosa, por el que los individuos no toleran el azúcar lactosa de la leche, es un ejemplo de los principios generales implicados. La lactosa es un disacárido que representa el 7% en la leche humana y 4% en la leche de la vaca. Para metabolizar lactosa, la especie humana necesita la enzima lactasa, que escinde el disacárido. En los primeros años de vida se producen cantidades adecuadas de lactasa. Sin embargo, en muchas personas, el nivel de ésta enzima cae pronto drásticamente. Estos individuos, cuando llegan a adulto, no pueden tolerar la leche (2). Esta situación es particularmente prevalente en esquimales, africanos, asiáticos y nativos americanos, aunque no se limita sólo a estos. En algunas de estas culturas la leche normalmente se convierte en queso, mantequilla y yogur, en donde la cantidad de lactosa se reduce significativamente y casi se pueden eliminar sus efectos adversos. En muchos lugares se dispone comercialmente de leche baja en lactosa y, para ayudar la digestión de otros alimentos que contienen lactosa, la lactasa es ahora un producto comercial que se puede ingerir (2). REFERENCIAS 1. Klug WS, Cummings MR, Spencer CA. Conceptos de Genética. 8a ed. Madrid: Pearson Educación; 2006. 2. Audesirk T, Audesirk G, Byers BE. Biología, la vida en la tierra, con fisiología. 9a ed. México: Pearson Educación de México, S.S de C.V; 2012.