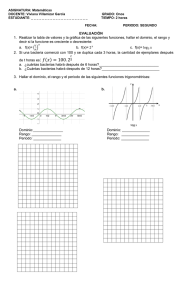
Apunte - Biología - Microbiología
Anuncio

MICROBIOLOGIA INTRODUCCIÓN I; PRINCIPALES MICROORGANISMOS 1 Priones y viroides. 2 Virus. 3 Procariontes II; HISTORIA DE LA MICROBIOLOGÍA 1 Acercamiento a los MO. 2 Teoría de la generación espontánea (TGE). 3 Conocimiento de los MO en el siglo XVIII. 4 Nacimiento de la microbiología en el siglo XIX. 5 Desarrollo de la técnica microbiológica. 6 Microbiología del suelo. 7 Quimioterapia antimicrobiana. III; OBSERVACIÓN DE MICROORGANISMOS A/ Sistemas de lentes 1 Sistemas de lentes. 2 Tipos de microscopía. B/ Preparaciones y muestras 1 Tipos de observaciones. 2 Toma de muestras. 3 Técnica de la gota pendiente. 4 Preparación de muestras teñidas. 5 Tinción simple. 6 Tinción diferencial. 7 Tinción Gram. 8 Tinción en caliente. 9 Tinción ácido alcohol resistente (AAR), o de Ziehl - Niessen. C/ Diferencias observables entre microorganismos 1 Tamaño de los microorganismos. 2 Principales formas de bacterias. 3 División y disposición en colonias de los cocos. 4 Las bacterias gigantes. IV; AISLAMIENTO Y CULTIVO DE MICROORGANISMOS A/ Medios de cultivo 1 Generalidades. 1 2 Requisitos nutritivos del medio de cultivo. 3 Otros compuestos del medio de cultivo. 4 Tipos de medios de cultivo. B/ Técnicas de aislamiento 1 El cultivo puro. 2 Obtención de cultivos puros. 3 Obtención de colonias por agotamiento de asa en placa. 4 Obtención de unidades formadoras de colonias (UFC) por divisiones sucesivas. 5 Obtención de colonias por filtración e incubación de muestras. 6 Microorganismos que no crecen en medios de cultivo. C/ Conservación de cultivos 1 Incubación de cultivos. 2 Métodos de conservación de cultivos. 3 Colecciones de cultivos. Parte I; LOS MICROORGANISMOS PROCARIONTES INTRODUCCIÓN A/ Evolución de la célula. B/ La célula bacteriana. 1 Generalidades. 2 Analisis de la composición de Escherichia coli. 3 Elementos constantes en Bacteria. 4 Elementos frecuentes en Bacteria. 5 Elementos siempre ausentes en Bacteria. I; ESTRUCTURA Y FUNCIONAMIENTO DE LA CÉLULA BACTERIANA. A/ La Cubierta de la Célula Bacteriana; Membrana Plasmática. 1 La Membrana Plasmática de Procariontes. 2 Membrana plasmática de Bacteria. 3 Permeabilidad. 4 Obtención y Acumulación de Energía. 5 Percepción del medio. 6 Exportación de proteínas sintetizadas por ribosomas. 7 Separación de los cromosomas durante división celular. B/ La Cubierta de la Célula Bacteriana; Pared Celular. 2 1 La pared celular en procariontes. 2 La pared celular Gram -. 3 La pared celular Gram +. 4 Los Peptidoglucanos. 5 La membrana externa y el Periplasma de Gram -. 6 Biosíntesis del Peptidoglucano. 7 Las Bacterias Acido-Alcohol Resistentes (AAR), o micobacterias. 8 Casos especiales. C/ El interior de la Célula Bacteriana; La Información Genética. 1 El Nucleoide. 2 Replicación del DNA. 3 Organización de los Genes y Transcripción en procariontes. 4 Regulación de la Transcripción. D/ El interior de la Célula Bacteriana; El Citoplasma. 1 MreB; Citoesqueleto Baceriano. 2 Ribosomas 70S y Traducción del mRNA a Proteínas. 3 Inclusiones del Citoplasma. E/ Estructuras y Apéndices externos de la Célula Bacteriana. 1 El Glicocalix. 2 Fimbria y Pili. 3 Apéndices implicados en la movilidad bacteriana; Los Flagelos Bacterianos. 4 Endoflagelos. F/ Movilidad Bacteriana. 1 Tipos de Movilidad. 2 Movilidad Flagelar. 3 Quimiotaxis; Percepción Sensorial relacionada con Movimiento Flagelar. 4 Movilidad Deslizante. G/ Ciclos Celulares de las Bacterias. 1 La División Celular en Bacteria. 2 Procesos de Diferenciación Celular; Las Endosporas. 3 Esporulación de las Endosporas. 4 Genes Spo; Los Genes de la Esporulación. 5 Germinación de las Endosporas. 6 Otros procesos de Diferenciación Celular; Caulobacter. 7 Otros procesos de Diferenciación Celular; Mixobacterias. 8 Otras Bacterias que desarrollan formas de resistencia. II; EL CRECIMIENTO BACTERIANO. A/ Introducción. 1 Introducción. 2 Generalidades sobre crecimiento bacteriano. 3 Las Técnicas de medida. 3 B/ Las Técnicas de Medida de Densidad de una Bacteriana. 1 Método directo por recuento directo al microscopio. 2 Método directo por diluciones seriadas. 3 Método directo por rastreo de poblaciones por incremento de la masa celular. 4 Método directo por el peso de la población. 5 Métodos indirectos. Población C/ Estudio del Crecimiento de una Población. 1 La curva de crecimiento poblacional. 2 Expresión matemática del crecimiento poblacional. 3 Importancia y técnicas industriales del seguimiento de una población bacteriana. D/ Efectos de los Factores Ambientales sobre el Crecimiento. 1 Interés Industrial. 2 La Temperatura. 3 El pH. 4 La Osmolaridad. 5 Tolerancia al Oxígeno. E/ Control del Crecimiento Bacteriano. 1 Generalidades. 2 La Esterilización. 3 La Pasteurización. 4 La Filtración. 5 Desinfección y Antisepsia. 6 Agentes Antimicrobianos Quimioterapeuticos. III; PARTICULARIDADES BACTERIANO. DEL METABOLISMO A/ Introducción al Metabolismo en Procariontes. 1 El Metabolismo; Generalidades. 2 Tipos de Metabolismos generales. 3 Tipos de Rutas Metabólicas Bacterianas. 4 El Poder Reductor. B/ Nutrientes y Vías de Asimilación. 1 Los Macronutrientes. 2 Los Micronutrientes. 3 Asimilación de Nutrientes. 4 Incorporación del Nitrógeno. 5 Incorporación del Azufre. 6 Incorporación del Fósforo. C/ Energía, Precursores Metabólicos, Poder Reductor. 4 1 Energía. 2 Precursores Metabólicos. 3 Poder Reductor. D/ Fuentes de Energía en Bacterias. 1 Metabolismo Quimioheterótrofo. 2 Cadenas de Transporte Electrónico de Bacterias. 3 Metabolismo Fotótrofo. 4 La Fotosíntesis. 5 La Autotrofía. 6 Metabolismo Quimiolitótrofo. E/ Bacterias Quimiolitótrofas. 1 Generalidades. 2 Las Bacterias Nitrificantes (oxidantes del nitrógeno). 3 Las Bacterias Sulfooxidantes (oxidantes del azufre). 4 Las Bacterias Férricas o Ferrooxidantes (oxidantes del hierro). 5 Drenaje ácido de las minas. 6 Bacteria oxidantes de Hidrógeno (H2). IV; GENÉTICA BACTERIANA; ADAPTACIÓN Y MUTACIÓN. A/ Mutaciones. (Láminas A) 1 Tipos de Mutaciones Génicas. 3 Agentes Mutágenos. 4 Mecanismos celulares de Reparación. 5 Mutaciones con Cambio en el Fenotipo y su detección en el laboratorio. B/ Plásmidos y Transposones. (Láminas C) 1 Plásmidos. 2 Transposones y Transposición. 3 Generalidades sobre el Intercambio de Material Genético entre Bacterias. C/ Transformación y Transducción. (Láminas D) 1 Transformación. 2 Transducción. C/ Conjugación. (Láminas E) 1 Generalidades sobre la Conjugación en Gram negativas. 2 El proceso de Conjugación. 3 Cepas Hfr. 4 Otros plásmidos, comunicación entre Gram positivas y entre Bacterias y Vegetales. C/ Introducción a la Ingeniería Genética. (Láminas B) 1 Generalidades sobre Ingeniería Genética; Enzimas de Restricción. 2 Vectores de Clonación; Plásmidos Artificiales. 3 Otros Vectores de Clonación. 5 4 Técnicas de Rastreo. Parte II; VIROLOGÍA I; INTRODUCCIÓN A LOS VIRUS. A/ Características Generales (Láminas F). 1 Definición. 2 Estructura General; El Core o Genoma Vírico. 3 Estructura General; La Cápsida y La Envuelta. 4 El Cultivo de Virus. B/ La Replicación de los Virus. 1 Ciclo de Replicación Vírica. 2 La Clasificación de Baltimore. II; Tipos de Virus. A/ Virus Bacteriófagos (Láminas G). 1 Clasificación General de Bacteriófagos. 2 Los Fagos de la Serie T. 3 El Fago Lambda. 4 El Fago Mu. 5 Fagos con ADN mc. 6 Fagos con ARN. B/ Virus Animales (Láminas H). 1 Generalidades sobre la Infección de virus a células animales. 2 Efectos Citopáticos de la infección (ECP). 3 Evasión del Sistema Inmune. 4 Virus Animales de ADN. 5 Virus de ARN. 6 Retrovirus y Hepadnavirus. C/ Virus Vegetales (Láminas I). 1 Generalidades sobre la Infección de virus a células vegetales. D/ Otros Agentes Infecciosos Acelulares (Láminas J). 1 Viroides y Virusoides. 2 Priones. 6 Parte III; LOS MICROORGANISMOS EN LA NATURALEZA Y EN LA INDUSTRIA I; Interacciones entre Microorganismos. A/ Tipos de Simbiosis (Láminas K). 1 Generalidades; la Simbiosis. 2 Mutualismo. 3 Parasitismo. B/ Respuesta del Sistema Inmune ante infección por Parásitos (Láminas K). 1 Evasión de la Respuesta Inmune. 2 La Respuesta Inmune; Defensas Inespecíficas. 3 La Respuesta Inmune; Defensas Específicas. II; Taxonomía Bacteriana. A/ Principios de Taxonomía Bacteriana (Láminas L). 1 Generalidades. 2 Taxonomía fenética. 3 Filogenia Bacteriana. III; MICROORGANISMOS DE ESPECIAL INTERÉS. A/ Microorganismos Patógenos (Láminas M). 1 Generalidades. 2 Bacterias de Transmisión Aérea. 3 Bacterias Transmitidas por Picadura de Artrópodos. 4 Bacterias de Transmisión Sexual. 5 Bacterias Transmitidas por el Agua y los Alimentos. 6 Otros tipos de Transmisión. B/ Microorganismos de Interés Industrial (Láminas N). 1 Generalidades, Microbiología industrial. 2 Cultivos Industriales. 3 Industria Farmaceutica (médica). 4 Industria Agroalimentaria. 5 Elaboración de Alimentos. 6 Biodeterioro. 7 INTRODUCCIÓN Microbios o microorganismos (MO): Seres microscópicos no visibles, generalmente unicelulares. Presentan una relación Superficie/Volumen muy elevada, alimentándose y respirando por difusión. Presentan un metabolismo rápido, ciclos vitales muy cortos y una elevada capacidad de dispersión (ubicuidad). Pueden presentar fotosíntesis, quimiolitotrofía, metanogénesis, y habitar medios de condiciones muy duras, como T elevadas, altas P o aridez. Microbiología: Estudio de los MO. Requiere métodos de trabajo específicos. I; PRINCIPALES MICROORGANISMOS Los microorganismos no conforman un grupo taxonómico filogenéticamente. Son, de hecho, microorganismos seres evolutivamente muy distantes, que solo tienen en común su reducido tamaño. Entidades acelulares: Algunos MO son acelulares, y generalmente infecciosos. Son los priones, viroides y virus. Organismos celulares: La mayoría de los MO son células procariontes, como Bacteria y Achaea, o eucariontes, del grupo de las algas, los hongos o protozoos. 1 Priones y viroides: Los priones son proteínas que en su forma natural (PRPc) se encuentra en el SN de varios mamíferos. La forma patógena (PRPSc) se contagia por contacto, y es responsable de enfermedades degenerativas neuronales contagiosas. Los viroides son secuencias circulares de ARN, de comportamiento infeccioso. Provoca muchas enfermedades en vegetales. 2 Virus: Entidades acelulares, de tamaño mucho menor que el de las bacterias, formadas por: Ácido nucleico (DNA o ARN). Una cápsida, formada por uno o varios tipos de proteínas acopladas en estructuras geométricas. Una cubierta lipídica con proteínas obtenida de la mbr de una C infectada. Carecen de auntonomía: carecen de maquinaria de reproducción del genoma y metabolismo. Parasitan los de las C durante las infecciones (parásitos intracelulares). 3 Procariontes La amplia mayoría de los MO son procariontes unicelulares. Los procariontes se dividen en dos dominios, bacteria y archaea, filogenéticamente muy distantes. Se distinguen de los organismos eucariontes a distintos niveles: C de pequeño tamaño y organización interna simple. Carecen de mbr nuclear y compartimentacíon Car. No presentan CE. Ribosomas 70S. Generalmente su sencillez estructural condiciona su reducido tamaño. Como excepción, se conocen dos especies de bacterias gigantes. 8 II; HISTORIA DE LA MICROBIOLOGÍA 1 Acercamiento a los MO: Los MO pasan desapercibidos a la vista. Solo se pudo detectar su presencia gracias a enfermedades infecciosas, fermentación (láctea, vino, cerveza) y conservación de alimentos, elaboración de encurtidos… es decir, a sus efectos sobre la materia. Hipócrates, médico de la Antigua Grecia, nombró miasmas a los agentes patógenos que se transmitían entre las personas. 2 Teoría de la generación espontánea (TGE): Fuente de controversia que acompaña a la microbiología durante siglos. Según la TGE, algunos organismos vivos pueden originarse a partir de la materia no viva. Aristóteles fue el primero en plantear que podía suceder así al observar la aparición de de gusanos en la materia en descomposición de cadáveres. Van Helmont defendía que podían crearse ratones por generación espontánea colocando en el desván carne putrefacta envuelta en trapos. Francesco Redi desmiente por primera vez, en el siglo XVII, la TGE utilizando una mosquitera. Sin embargo, la carne no infestada de gusanos ni ratones acababa descomponiéndose debido a los MO. Finalmente Pasteur desmiente la TGE, gracias a su experimento en medio aséptico con su matraz en cuello de cisne. Demuestra que la putrefacción depende de los gérmenes del aire: la vida microbiana proviene de gérmenes en partículas en suspensión en el aire (También endosporas). 3 Conocimiento de los MO en el siglo XVIII: Van Leeuwenhoek, pañero, fue la primera persona que pudo ver MO, gracias a las primeras lentes de grandes aumentos que utilizaba para contar hilos. Observa los bacilos móviles en infusiones y les da en nombre de animáculos. The Royal Society recibe sus cartas y descripciones pero no le dan veracidad pues no es científico. Se comienzan a observar animáculos en infusiones (agua + materia orgánica muerta). Aparecen Needhan (espontaneista) y Spallantzani (antiespontaneista), que trabajan con infusiones en matraces esterilizados para confirmas o desmentir respectivamente la TGE. Ambos creen que sus experimentos confirman sus ideas. Las formas esporuladas y pequeñas entradas de aire en los matraces hacen muy difíciles estos experimentos. Jenner, médico inglés, realiza la primera variolación; aplicación empírica de la vacunación, contra la viruela. Injecta pústulas de animales infectados (vacas), de forma a inmunizar a personas sanas. 4 Nacimiento de la microbiología en el siglo XIX: Se comienza a utilizar desde el punto de vista quirúrgico la asepsia o antisepsia. Fue, en parte, gracias al joven ginecólogo de Viena, Semmelweiss. El hospital en que trabajaba estaba dividido en una zona para pobres cuyos partos atendían comadronas, y otra para ricos donde los médicos se hacían cargo de los partos. Asoció la mayor frecuencia de defunciones en partos de la zona de los ricos, y que los médicos, que tratan con cadáveres y enfermos sin utilizar asepsia ni higiene, concluyendo que transmitían los MO a las madres en los partos. Fue expulsado del hospital. Lister, cirujano inglés, fue el primero en utilizar la técnica aséptica en sus operaciones, con alcoholes. Louis pasteur, químico francés, es el fundador de la Microbiología como ciencia. El gobierno francés le encarga trabajos de análisis de vinos franceses para exportar, que se picaban en el trayecto. Asi descubre los MO. 9 Estudia también la cerveza, descubriendo bacterias anaerobias y levaduras de la fermentación alcohólica. Descubre así también la vida anaerobia. Pone de manifiesto el origen microbiano de muchas enfermedades contagiosas. Desmiente la TGE ganando un premio de la academia francesa. Descubre la primera vacuna, para la rabia. Inocula el virus de la rabia de la medula espinal de animales afectados. La prueba sobre un niño en estado terminal, y logra milagrosamente salvar su vida. Descubre la pasteurización como técnica de conservación de alimentos, que no llega al punto de la esterilización. Expone el producto a 100ºC durante 10 minutos y lo hermetiza. Robert Koch identifica gracias a su postulado agentes causantes de enfermedades como la tuberculosis, el cólera… Según su postulado, un tipo de MO es causante de una enfermedad concreta. Se dedica al aislamiento y cultivo de bacterias. Aislamiento del MO. Obtención del cultivo puro. Inocular en animal sano de experimentación Si el animal sano desarrolla la enfermedad, el MO purificado es responsable de ella. Se puede aislar de nuevo el MO a partir del animal inoculado de experimentación. Descubre con su postulado MO responsables de enfermedades como el ántrax (Bacillus anthracis), o de la devastadora tuberculosis (Microbacterium tuberculosis). Esta última es una bacteria muy difícil de cultivar por sus altos requerimientos en nutrientes y el lento crecimiento poblacional, y de observar pues no se tiñe fácilmente. 5 Desarrollo de la técnica microbiológica: Los primeros medios de cultivo fueron líquidos, como las infusiones. Son cultivos más difíciles de observar y valorar. El propio pasteur no dominaba la actual metodología del cultivo puro. Robert Koch es el primero en utilizar cultivos sólidos. Logra hacer desarrollarse colonias por mitosis de una única bacteria sobre rodajas de patatas. Las colonias forman grandes grumos fáciles de aislar; no se dispersan por flotación. Sin embargo, es difícil aislar estos cultivos de contaminación. Un discipulo de Koch inventa el cultivo sólido en placas de Peyer sobre gelatinas. Actualmente se utilizan las placas de Petri sobre cultivos sólidos de agar-agar extraído de algas, pero es un derivado de la técnica de Peyer. 6 Microbiología del suelo: Martinus Beijerinck descubre los MO relacionados con la fertilidad del suelo. Crea los cultivos de enriquecimiento, no puros, extraídos del suelo. Las condiciones de cultivo hacen proliferar determinados grupos de MO desplazando a las otras poblaciones. Descubre las bacterias fijadoras de nitrógeno: Estas convierten el nitrógeno molecular en compuestos nitrogenados fértiles para la vida. Son esenciales en los suelos para la subsistencia de muchas plantas. (N2 -> 2 NH3 -> Incorporación en aminoácidos). Por ejemplo, Rhizobium vive en simbiosis en los nódulos reticulares de la raíz de leguminosas, fijando N 2. También funda los principios básicos de la virología gracias al mosaico del tabaco, infección dañina para la economía. Gracias a diminutos poros logra filtrar MO a los que llama virus filtrables. Por entonces se denominaba virus a un agente patógeno. Sergei Winogradsky descubre los ciclos de renovación del nitrógeno y del azufre y muchas bacterias relacionadas. Descubre también la quimiolitotrofía: Metabolismo autótrofo no dependiente de la luz. 7 Quimioterapia antimicrobiana: Paul Ehrlich inicia la búsqueda de la bala mágica, fármaco nocivo selectivamente contra las bacterias o MO infecciosos, o C cancerosas. Crea el Salvarsan para tratamiento de la sífilis. Este 10 medicamento derivado de arsénicos contenía metales pesados nocivos para bacterias, pero es responsable de muchos efectos secundarios. Domagk descubre que las sulfamidas refrenan el crecimiento bacteriano buscando colorantes que las tiña. Crea el Prontosil, con efecto bacteriostático, no bactericida, que bloquea el metabolismo bacteriano. Alexander Fleming descubre finalmente la penicilina, primer antibiótico, en 1928. Descubre este producto de secreción bactericida del Hongo Penicillum gracias a placas Petri accidentalmente contaminadas por este moho. Observó a simple vista Halos de inhibición. La extracción del producto resulta muy cara y escasa, y hasta 1940 no comienza la era de comercialización de estos productos. Waksman descubre la estreptomicina, antibiótico eficaz contra la tuberculosis, ante la que la penicilina no surgía efecto. Actualmente la mayoría de antibióticos se obtienen a partir de Actinomicetos, MO del suelo. 11 III; OBSERVACIÓN DE MICROORGANISMOS A/ Sistemas de lentes 1 Sistemas de lentes: Los microscopios están formados por dos lentes de aumento (objetivo y ocular) cuyos valores se multiplican para obtener el aumento del microscopio, y un condensador que dirige y condensa en la muestra los rayos del foco de luz (ópticos) o electrones (electrónicos). El aumento para observar bacterias gira entorno a 1000x (Por ejemplo; objetivo 100x, ocular 10x). El poder de resolución d del sistema de lentes es la distancia mínima que separa dos puntos claramente distinguibles. Depende de (longitud de onda de la luz), n (indice de refracción del medio) y (Semiángulo del cono de luz). d = (0,5.) / (n.sen Al presentar un d mucho menor, los microscopios electrónicos permiten muchos más aumentos que los ópticos. Son necesarios para la observación de los virus. Profundidad de campo: Espesor de muestra enfocado. Área de campo: Diámetro del espacio enfocado. 2 Tipos de microscopía: La microscopía óptica utiliza como fuente un haz de luz visible. Debido a su elevada tiene mayor d. De campo claro: Con muestras teñidas. De campo oscuro: Para muestras pequeñas. Fondo oscuro, muestra aparece iluminada por refracción de un haz de luz desviado. De contraste de fases: Para observación de estructuras internas. Alto contraste producido por desfase de rayos de luz incidentes. De contraste de interferencia diferencial: observaciones 3D. De fluorescencia: Sin haz de luz. Técnicas de inmunofluorescencia; anticuerpos unidos a fluorocromo. Muestras clínicas. La microscopía electrónica utiliza como fuente un haz de electrones. Permite enfocar en aumentos mucho mayores gracias a su menor d. De transmisión: Observación de estructuras internas en cortes ultra finos. De barrido: Observación de superficies. B/ Preparaciones y muestras 1 Tipos de observaciones: Observación en fresco: MO vivos, no teñidos, en gota pendiente. Observación de preparaciones teñidas: MO muertos. Se realiza frotis, fijación y tinción. Existen varios tipos de tinción distintos (Simple, negativa, diferencial) 2 Toma de muestras: Se utiliza la técnica aséptica para obtener cultivos puros: Se trabaja en ambiente estéril, y se manipula a un radio de 20cm del mechero Bunzen. 12 Se utiliza un asa de siembra para tomar la muestra. Se forma por un mango y un filamento enroscado que debe ser flameado para esterilizarlo. También se flamea la boca del tubo de ensayo donde va a ser depositada la muestra: Cultivo joven en medio líquido. 3 Técnica de la gota pendiente: Se observan los MO vivos del cultivo joven, pudiéndose valorar la movilidad (perdida a las 48h), la pureza del cultivo, las divisiones celulares o procesos de esporulación. Pueden utilizarse algunos colorantes no tóxicos como azul de metileno. Se utiliza un porta excavado, con una depresión central. La muestra se coloca sobre el cubre con el asa de siembra, todo esterilizado. Se coloca el cubre sobre el porta de forma que la muestra quede en la depresión. Se emplea microscopia óptica de campo claro, campo oscuro o contraste de fases. La carencia de tinción y el movimiento dificultan la observación. 4 Preparación de muestras teñidas: Frotis: Película fina de suspensión de C o MO. Se coloca una muestra concentrada y una gota de agua en el centro del porta. Se extiende la mezcla y se deja secar al aire. Fijación: Adhesión de la muestra al porta. Para bacterias es suficiente pasar la muestra por el mechero bunzen, gracias a la PC. Otras C requien métodos más complejos. Tinción: Aplicación de colorantes sobre la muestra. Se pueden aplicar mordientes que refuerzan la unión colorante – muestra. 5 Tinción simple: Se utiliza un único colorante. Se aplica directamente sobre la muestra fijada, después se lava con agua. Permite mejor observación de formas, tamaños y agrupaciones. Colorantes: Azul de metileno, Cristal Violeta, Safranina, Fuscina (…) Técnica de tinción negativa con Nigrosina: Partículas de carbón, que al ser demasiado grandes para penetrar y teñir al MO deja el fondo negro y los MO iluminados. 6 Tinción diferencial: Permite diferenciar distintos tipos celulares gracias a la utilización conjunta de varios colorantes distintos. A continuación veremos distintos tipos de tinción diferencial. 7 Tinción Gram: Principal tinción diferencial. Permite separar las bacterias actuales en dos grandes grupos: Gram + y Gram -. Metodología: Cristal violeta (colorante fundamental) – 2 min. Lugol (mordiente) – 1 min. Alcohol (decoloración) – 30 seg. Lavado con agua. Safranina (colorante de contraste, rojo) – 1 min. Lavado con agua y secado. El cristal violeta solo colorea las bacterias Gram + que aparecen de color violeta rosáceo. Las bacterias Gram – aparecen de color rojo, teñidas por la safranina. 8 Tinción en caliente: 13 rojo. - Se utiliza para teñir las endosporas bacterianas. Metodología: Verde malaquita (colorante fundamental, hidrosoluble) – 3 min. Aplicado sobre mechero bunzen, en caliente, sin llegar a herbir. Decoloración y lavado con agua. Safranina (colorante de contraste) – 1 min. Lavado y secado. Las esporas aparecen teñidas en verde sobre el cuerpo vegetativo bacteriano que aparece La presencia de endosporas es determinante a la hora de la clasificación de las bacterias. 9 Tinción ácido alcohol resistente (AAR), o de Ziehl - Niessen: Permite diferenciar las mycobacterias. Se utiliza como prueba diagnóstica para la tuberculósis. Metodología: Fuscina fenicada (colorante fundamental) – 3-5 min. Aplicado en caliente. Decoloración con ácido nítrico (33%) – alcohol. Lavado con agua. Azul de metileno (colorante de contraste) – 1-2 min. Las mycobacterias presentan acido-alcohol-resistencia debido a la alta cantidad de grandes y complejos lípidos en su PC (ceras), y aparecen de color fuscia. Se denominan AAR +. Son Gram -. Las otras bacterias y C pierden el fuscia durante la decoloración, apareciendo azules. Se trata de AAR-. C/ Diferencias observables entre microorganismos 1 Tamaño de los microorganismos: Las bacterias son generalmente muy pequeñas. Se miden en micras y para su observación es necesario un sistema de lentes equivalente a 1000x. Los eucariotas son de mucho mayor tamaño. Pueden ser pluricelulares y medirse en milímetros. Los virus son los más pequeños. Solo son observables con microscopio electrónico. Se miden en nanómetros. Los POX virus, de la viruela, son los más grandes: Pueden verse con microscopio óptico. 2 Principales formas de bacterias: Los Cocos son las bacterias de forma esférica. No suelen presentar flagelos ni movimiento. Los Bacilos son bacterias alargadas. Son las que presentan flagelos más frecuentemente. Los Espirilos son bacilos con forma curvada. Las Espiroquetas son bacterias muy alargadas y en espiral. Pueden observarse en microscopio de campo oscuro. Las Bacterias filamentosas son muy alargadas, con aspecto similar a una cuerda. Algunas bacterias (cocos, espirilos) presentan prosteca o tallo, para fijarse al sustrato. Presentan un ciclo caracteristico donde se alterna la forma de C nadadora con flagelo y C sésil adherida al sustrato mediante el apéndice. Existen otras formas especiales. (Formas geométricas, pentágonos). 3 División y disposición en colonias de los cocos: Estreptococos: Presentan un único plano de división formando cadenas. 14 Tétradas: Presentan dos planos de división. Forman cuadrados de 4 C o tétradas. Sarcinas: Presentan tres planos de división y forman cubos. Estrafilococos: Presentan tres planos de división y forman grandes agrupaciones deformes o racimos. 4 Las bacterias gigantes: Mientras las dimensiones de E. coli son 1,5 x 2 m y las del género Bacillus 1,5 x 4 m, se conocen dos especies de bacterias gigantes cuyas dimensiones son 8 x 50 m: Son mayores que los paramecios. Hasta su descubrimiento se pensaba que los procariontes tenían una limitación de tamaño mucho mayor. Una de las bacterias gigantes es quimiolitótrofa, la otra endosimbionte del pez cirujano. 15 IV; AISLAMIENTO Y CULTIVO DE MICROORGANISMOS A/ Medios de cultivo 1 Generalidades: La obtención de cultivos puros y viables requiere la utilización de técnicas determinadas, como la técnica aséptica. Siembra: Se deposita un inóculo, o pequeña porción de muestra en un medio de cultivo. Incubación: Se procuran y mantienen las condiciones físico-químicas adecuadas para la proliferación del inóculo de la muestra. Medio de cultivo: Disolución acuosa, generalmente sólida, de nutrientes. Suele depositarse en placas de Petri, matraces o tubos de ensayo. Los medios de cultivo se pueden comprar preparados y deshidratados. Se disuelve el preparado en agua y se lleva a ebullición durante 3 minutos. 2 Requisitos nutritivos del medio de cultivo: El medio de cultivo debe contar con distintos componentes en función del tipo de microorganismo que se desea cultivar. Estos son los principales requisitos: Fuente de carbono: Orgánico (azúcares) en heterótrofos, Bicarbonato o CO 2 en autótrofos. Fuente de N, P y S: Sales minerales. Cationes: Ca2+, Na+, Mg2+, K+. Oligoelementos en baja concentración: Fe2+, Cl Factores de crecimiento, compuestos orgánicos (vitaminas, aminoácidos). Agente solidificante: AGAR AGAR, Agar bacteriológico. E coli, por ejemplo, crece con sólo azucares y sales. Las bacterias lácticas, sin embargo, precisan de factores de crecimiento. Para obtener medios de cultivo nutritivo se suelen utilizar los siguientes componentes: Sales minerales: NaCl, NH4+, KCl … Extractos: Caldos de carne, levaduras o soja. Peptonas: Digeridos proteicos animales o vegetales, aportan aminoácidos. Fluidos corporales: En caso de cultivo de bacterias patógenas. 3 Otros compuestos del medio de cultivo: Solución tampón: para evitar cambios bruscos en el pH, y mantener su valor en 6 < pH < 7. En el caso de bacterias acidófilas o alcalinófilas, pH < 6 o pH >7 respectivamente. Indicadores de pH: Controlando su valor se pueden localizar reacciones fisiológicas como la fermentación de lactosa que implica un descenso del pH. Colorantes para localizar reacciones fisiológicas, como el púrpura de bromocresol, que pasa de morado a amarillo cuando tiene lugar la fermentación del sustrato. Se utiliza para seleccionar colonias fermentadoras de azúcares en placas de Petri (zonas amarillas). Se pueden emplear agentes selectivos creando medios de cultivo selectivos. Inhiben el crecimiento de ciertos grupos determinados de MO para aislamiento de bacterias. Por ejemplo, se emplean agentes reductores como triglicolato o cisteína, que captan el O 2, aislando bacterias anaeróbias o microerófilos. 4 Tipos de medios de cultivo: Los cultivos pueden ser líquidos, semisólidos o sólidos. Suelen presentarse en placas de Petri, si son líquidos, en matraces. Suelen ser conservados en tubos de ensayo incliados, aumentando la superficie de exposión. 16 Generales: Contienen nutrientes que permiten el crecimiento de cualquier muestra bacteriana. Enriquecidos: Contienen además compuestos adicionales requeridos para ciertos cultivos especiales, como plasma sanguíneo para ciertos MO patógenos. Selectivos: Permiten el crecimiento selectivo de determinado grupo de bacterias, inhibiendo el crecimiento de las otras. Por ejemplo, cristal violeta impide crecimiento de Gram +. En preswencia de nitrógeno, solo se desarrollan las bacterias capaces de fijarlo. Diferenciales: Permiten diferenciar visualmente ciertas colonias de bacterias, como en el caso de indicadores de pH o colorantes. Especiales o de identificación: Se utilizan para diferenciar y clasificar bacterias, recurriendo a diferencias fisiológicas y metabólicas como fermentación, respiración, o proteolisis. Son cultivos enriquecidos y diferenciales, que permiten comprobar si un MO da determinada reacción. B/ Técnicas de aislamiento 1 El cultivo puro: El cultivo puro es un cultivo donde solo prolifera una única estirpe de MO. Suelen obtenerse a partir de una célula microbiana por sucesivas mitosis. Para obtener el cultivo puro debemos aislar el MO que deseamos estudiar de una muestra natural, que es una población mixta. 2 Obtención de cultivos puros: Colonia: Formación macroscópica con morfología característica de la especie, que tiene por origen una única célula UFC (unidad formadora de colonias). A partir de la inoculación de una colonia obtendremos un cultivo puro. Por medio de cultivos de enriquecimiento, selectivos o diferenciales, podemos favorecer selectivamente la proliferación de determinadas colonias de MO, o diferenciarlas con mayor facilidad. Se pueden obtener cultivos puros por micromanipulación, a partir de una única C madre. 3 Obtención de colonias por agotamiento de asa en placa: Después de esterilizar el asa de siembra, se inocula, y se descarga en la placa Petri. A continuación se limpia y esteriliza el asa, se inocula de la primera descarga y de nuevo se realiza una segunda descarga. Después de repetir el proceso, en la últimas descargas quedan muy pocas C y muy separadas, de este modo después de incubar se obtienen colonias visiblemente separadas, y se pueden inocular selectivamente obteniendo cultivos puros. 4 Obtención de unidades formadoras de colonias (UFC) por divisiones sucesivas: Se parte de un cultivo joven en medio líquido. Las poblaciones bacterianas se encuentran en concentraciones muy altas. Se procede a realizar diluciones sucesivas de la muestra en sueros nutritivos. Después de n diluciones, se inocula 1 ml del último tubo en un medio de cultivo en placa, extendiéndose con asa Drigalski de vidrio. Después de incubación, aparecen UFC colonias visiblemente separadas, originadas en una sola célula. Efectivo con muestras más cargadas que en el caso de agotamiento de asa. Técnica de profundidad: En ocasiones se coloca el medio de cultivo sobre sobre la dilución y se homogeneizan de modo que las colonias no quedan todas en la superficie del medio de cultivo. 5 Obtención de colonias por filtración e incubación de muestras: Esta técnica se utiliza con muestras líquidas. Pueden ser cultivos jóvenes o diluciones en sueros. 17 La muestra se deposita en un recipiente sin fondo sobre un filtro en un portafiltros. El conjunto se dispone sobre un matraz con sistema de succión. Al activar la bomba de succión, todos los desechos caen al matraz, mientras el filtro retiene todos los MO, salvo los virus. Las bacterias retenidas en el filtro se llevan a medio de cultivo. Se obtienen colonias por incubación. Esta técnica también se utiliza para la esterilización de líquidos. 4 Microorganismos que no crecen en medios de cultivo: Treponema pallidum: Responsable de la sífilis. Se utilizan conejos. Microbacterium leprae: Responsable de la lepra. Se utilizan armadillos. Parásitos intracelulares obligados, como Riquetsia, Clamydia o virus. Se utilizan cultivos de otros tipos celulares para su proliferación en ellos. C/ Conservación de cultivos 1 Incubación de cultivos: Tanto para la obtención de colonias como para la conservación de cultivos es esencial incubarlos y mantenerlos a temperatura, pH y condiciones nutritivas adecuadas para la población estudiada. Para incubación de bacterias fotosintéticas es precisa la luz. Para bacterias anaerobias estrictas es esencial la carencia de O2, que las resulta tóxico. Las anaerobias facultativas no lo necesitan, pero pueden desarrollarse en su presencia. Para los cultivos de anaeróbios se utilizan jarras de anaerobios y catalizadores que eliminan el O2. Pueden producir gases como CO. Estos cultivos suelen sellarse con una capa de parafina estéril. Las bacterias microaerófilas de la fermentación de la lactosa requieren una baja [O 2]. Las bacterias aerobias estrictas necesitan el O2. Se utilizan para los cultivos aerófilos recipientes especiales estériles con alta aireación o planchas de aireación. Los cultivos bimembres son cultivos especiales de bacterias simbióticas que necesitan otras especies para subsistir, como las bacterias bacteriolíticas, consorcios de simbiosis altamente dependiente, o cultivos de virus. 2 Métodos de conservación de cultivos: Es suficiente realizar continuas resiembras en medios estériles para conservar durante temporadas cortas los cultivos. Si se agotan los nutrientes o acumulan tóxicos en el medio, los MO no subsisten. La refrigeración a 4ºC permite con un bajo coste conservar un cultivo durante un mes, reduciendo el metabolismo y la tasa de crecimiento de las poblaciones. La congelación permite conservar hasta 5 meses un cultivo, si se añaden agentes protectores para las C conservadas. La liofilización permite deshidratar al vacío las C sin alterarlas. Puede servir para conservar cultivos hasta durante 20 años. Los MO con formas criptobióticas como endosporas o quistes pueden subsistir varios años. 3 Colecciones de cultivos: Existen colecciones de cultivos de MO vivos, purificados y numerados, que se conservan generalmente liofilizados. ATCC (American Type Cultura Collection) es la colección de referencia. La Colección del Instituto Pasteur es la mayor de Europa. 18 Una de las más destacables en España es la Colección Española de Cultivo de la universidad de Valencia. 19 Parte I; LOS MICROORGANISMOS PROCARIONTES INTRODUCCIÓN A/ Evolución de la célula: Todos los tipos celulares encuentran su origen evolutivo en la célula procarionte. A partir de ese ancestro algunas C buscaron el camino de la individualidad, resultando organismos siempre unicelulares. Para seguir esta estrategia la estructura procarionte fue suficiente. Se encuentran dos líneas o dominios de procariontes claramente diferenciables: Bacteria y Archaea. Con esta estrategia evolutiva las C se comportan como individuos, y deben hacer frente a los cambios ambientales. Presentan mayor respuesta a cambios de pH o T. Presentan uniformidad celular: Gran capacidad de adaptación al ambiente cambiante. Otras C buscaron el camino de la cooperatividad, formando organismos pluricelulares. La estructura eucarionte, más compleja, triunfó en este propósito. Las células unidas conservan con más facilidad un ambiente estable y un medio de crecimiento adecuado. Se especializan en tejidos. Por sus procesos fisiológicos y su estructura similares, se piensa que las ancestrales Archaea dieron lugar a la aparición de eucariontes por un lado, y bacterias actuales por otro. B/ La célula bacteriana: 1 Generalidades: Las Bacterias actuales son seres primitivos unicelulares, que aparecieron hace unos 3000 Ma. C estructuralmente muy simples. Sin compartimentación ni núcleo celular. Presentan rápida interacción con el ambiente y una alta y rápida capacidad de adaptación (Uniformidad celular). Su reproducción tiene lugar mayoritariamente por fisión binaria. Su variabilidad tiene origen fundamentalmente en la mutación. En ocasiones puede tener lugar recombinación de material genético. Son seres haploides. Llevan a cabo la expresión génica por procesos muy distintos a los eucariontes. 2 Analisis de la composición de Escherichia coli: - 50% Proteínas: Constitutivas (estructurales). Inducibles. (encimáticas, sólo se sintetizan en determinadas circunstancias). - 25% RNA: 80% RNAr. 15% ARNt. 5% ARNm. - 3% DNA: Cromosomas circulares. Pequeños plásmidos. - 22% Otros: 41,4% Lípidos. 15,9% Lipopolisacárido (Sólo en Gram -). 20 11,4% Peptidoglucano (Pared celular bacteriana, en mayor concentración en Gram +). 11,4% Glucógeno. 20% Metabolitos, Vitaminas, Iones. 3 Elementos constantes en Bacteria: Pared Celular Bacteriana (PCB): Cubierta exterior de la membrana plasmática, responsable del mantenimiento de la morfología y presión osmótica Car. Membrana Car lipídica. Citoplasma. Nucleoide o Cromosoma Bacteriano. Ribosomas esparcidos en el citoplasma. Durante crecimiento activo aumenta su concentración considerablemente. Plásmidos con genes aventajados selectivamente, pero no indispensables para la supervivencia de la C. 4 Elementos frecuentes en Bacteria: Cápsula de polisacáridos externa a la PCB. Flagelos bacterianos para la movilidad. Fimbrias y Pili para adhesión al sustrato y recombinación de material genético. Inclusiones: Reservas de carbono y otros nutrientes. 5 Elementos siempre ausentes en Bacteria: Doble membrana nuclear. Compartimentación intracelular (Orgánulos). Citoesqueleto. 21 I; ESTRUCTURA Y FUNCIONAMIENTO DE LA CÉLULA BACTERIANA: A/ La Cubierta de la Célula Bacteriana; Membrana Plasmática: 1 La Membrana Plasmática de Procariontes: Rodea y delimita la célula. En caso de Bacteria es el sistema de relación entre ser vivo y medio externo. Mesosomas: Se trata de invaginaciones y plegamientos de la membrana. Aumentan la superficie de intercambio con el medio. Se piensa que sirven de anclaje para los cromosomas durante la división celular. Existen diferencias apreciables entre las membranas de los dos grupos de procariontes: Bacteria: Siempre es una Bicapa Lipídica. Es similar a Eucariontes; la unión Glicerol – 2 ácidos grasos es de tipo Ester Archaea: Puede ser Bicapa o Monocapa. La unión Glicerol – 2 cadenas Isoprenoides (Lípidos derivados de Isopreno) es de tipo Eter. En Archaea, Fitanilo es un Isoprenoide frecuente. En caso de bicapa, se forma de Glicerol Dieter Bifitanilo. En caso de monocapa se producen enlacer entre las moléculas de Fitanilo, formandose Glicerol Tretraeter Dibifitanilo. BICAPA MONOCAPA Enlace entre Dibifitanilos La monocapa lipídica de algunos miembros de Archaea proporcionan mayor resistencia para la vida en ambientes extremos, como alta T o bajo pH. Estas condiciones no pueden atacar los enlaces tetraeter monocapa. Los esteroles presentes en membranas de Archaea son los Hopanoides. 2 Membrana plasmática de Bacteria: Se trata de una Bicapa lipídica en Modelo de Mosaico fluído. Los esteroles presentes en membranas bacterianas son los Hopanoides. Presentan diversas proteínas ancladas a la membrana: Integrales y Periféricas. Entre las integrales o transmembrana se encuentran transportadores de nutrientes y desechos, proteínas exportadoras a pared celular o periplasma, y proteínas sensoras de percepción del entorno. Entre las periféricas se encuentran las proteínas del periplasma, las asociadas a cadenas de transporte electrónico (funciones mitocondriales), y las quinasas transductoras de señales, relacionadas con las proteínas transmembrana sensoras. La membrana de la Bacteria cumple más funciones que las de las membranas plasmáticas de eucariontes pluricelulares. Al delimitar Organismo/Entorno, debe cubrir más necesidades celulares. Son descritas a continuación. 22 3 Permeabilidad: El transporte de nutrientes al interior y la eliminación de los desechos en el entorno son mediados por prot transportadoras. Se han descubierto las Acuoporinas, que facilitan los Intercambios por Osmosis (H2O). No permiten el paso de sustancias grandes, ni de iones (polares). Difusión Simple: Moléculas que pueden atravesar la membrana. Relación lineal, depende sólo de [SINT] y [SEXT]. Caso de Canales (acuoporina) y Sistemas de Uniporte selectivo. Difusión facilitada: Moléculas que necesitan transportadores para atravesar la membrana. Depende de [SINT] y [SEXT], pero también de la Saturación: La velocidad del transporte se estabiliza cuando todos los transportadores están ocupados; es el nivel de saturación. Transporte activo: No es espontáneo, y requiere suministro energético. Se produce en contra del gradiente de concentraciones: Sistemas de simporte y antiporte: Transporte en contra de grad se acopla a otro a favor de grad, utilizando su energía. Sistema Fosfotransferasa (translocación): Se utiliza para importar azúcares al int Car. Durante el proceso. El nutriente es químicamente modificado cuando atraviesa la membrana. La energía proviene del fosfanolpiruvato; éste cede su P y queda Piruvato. Una cadena de kinasas llevan P hasta el transportador y lo activan. Durante el transporte, los azúcares quedan fosforilados. EXT INT Glucosa Glucosa-6-fosfato Transportador Transportadores ABC: Polímeros transmembrana que hidrolizan ATP como fuente de energía. 4 Obtención y Acumulación de Energía: Síntesis de ATP (Respiración/Fotosíntesis). Acumulación de Energía en forma de Gradiente Quimioosmótico de H + (ATPasas transductoras de protones). Suministro de Energía (Movimiento Flagelar, Transporte activo transmembrana). 5 Percepción del medio: Recepción de Señales extracelulares: Kinasas. El dominio interno se fosforila ante llegada de la señal externa. Transducción de señales para provocar la respuesta celular. Por ejemplo, E. coli puede reconocer en el entorno si se encuentra en agua o en un intestino, debido a la diferente P osmótica. 6 Exportación de proteínas sintetizadas por ribosomas: Componentes del Peptidoglucano Proteínas periplásmicas 7 Separación de los cromosomas durante división celular: No hay membrana nuclear. 23 B/ La Cubierta de la Célula Bacteriana; Pared Celular: 1 La pared celular en procariontes: La mayoría de procariontes poseen una pared celular, de estructura totalmente distinta de la Vegetal, pero de funciones similares. Como excepción, las archaeas Hipertermófilas y las bacterias Micoplasmas no presentan PC. La C procarionte debe soportar una alta presión osmótica. Además de esto, la PC mantiene la morfología Car. Archaea presenta gran variedad estructural en su PC. Se forma de proteínas, glucoproteínas y polisacáridos. Dentro del Dominio Bacteria, se distinguen tres tipos estructurales de PC: Gram –. Gram +. Ácido-Alcohol Resistentes. La PC bacteriana suele tener carga neta nula defendiendo a la bacteria ante fagocitosis. 2 La pared celular Gram -: En la región externa se encuentra una membrana bicapa lipídica de LPS (Lipopolisacáridos) en el exterior, y fosfolípidos en la hemimbr interna, y con proteínas ancladas. Entre esta mbr externa y la mbr plasmática se encuentra el Periplasma. En posición central, en el espacio periplasmático, se encuentra una cubierta de Peptidoglucano: Se considera PC propiamente dicha a esta cubierta. Las lipoproteínas median la unión entre la mbr externa y el peptidoglucano. 3 La pared celular Gram +: Se forma de un recubrimiento general, grueso, de peptidoglucano, unida a la mbr plasmática. Hay abundantes Ácidos Teicoicos; son polialcoholes (unidades repetidas de Glicerol-Ribitol) que actúan como antígenos bacterianos. Los ácidos lipoteicoicos median la unión PC – mbr Car. Existen algunas proteínas asociadas a PC. 4 Los Peptidoglucanos: Se trata de largas cadenas de azúcares formadas por repeticiones del disacárido formado por N-Acetilglucosamina (NAG) y N-Acetilhurámico (NAH): NAG-NAH, por uniones (1,4). El Disacárido Tetrapeptídico es la unidad básica del Peptidoglucano. Se produce la unión de un tetrapéptido en NAH del disacárido. El tetrapéptido es similar pero distinto en los dos tipos de PC: GRAM + NAH - L-Ala – D-Glu – L-Lys – D-Ala GRAM - NAH - L-Ala – D-Glu – H-Dap – D-Ala H-Dap: ácido meso diaminopimélico Se produce la unión lateral (1,4) entre los disacáridos tetrapeptídicos, formándose una larga cadena de peptidoglucano. Las cadenas de peptidoglucando dispuestas paralelamente pueden formar enlaces interpeptídicos, formandose una malla tridimensional de peptidoglucano, resistente, que recubre toda la superficie celular. Las uniones interpeptídicas son distintas en cada caso: GRAM + L-Lys – Gly(5) – D-Ala GRAM H-Dap – D-Ala 24 La Lisozima es una defensa ante las bacterias pues rompe el tetrapéptido que conecta entre sí las cadenas de peptidoglucano. 5 La membrana externa y el Periplasma de Gram -: Un Lipopolisacarido (LPS) se forma de un Lípido A, con varios ácidos grasos que se sitúan en la región hidrofóbica de la mbr externa. Este lípido con propiedades antigénicas es el causante de la Fiebre; Se trata de una endotoxina (Toxina estructural de la célula bacteriana). El Lípido A del LPS se encuentra unido a un núcleo polisacarídico, unido a su vez a un polisacárido específico que se proyecta hacia el exterior. Esta última cadena de sacáridos es muy variada y adquiere función de antígeno bacteriano para Gram -. La conexión Entorno-Periplasma es facilitada por proteínas Porina de la mbr externa. Se trata de polímeros de 3 subunidades, que forman tres canales transmembrana. Facilitan la difusión no selectiva a través de la mbr externa. Se encuentran también proteínas receptoras de señales específicas. 6 Biosíntesis del Peptidoglucano: Citoplasma: Síntesis de precursores: NAG y NAH-pentapéptido. Activación energética de precursores: Uridildifosfato (UDP) es un transportador que se une la los N-Acetil. Transporta los precursores selectivamente a través de Bactoprenol, prot transmbr de la mbr plasmática, hasta el periplasma. Periplasma: 1. Polimerización: Se forma la unión (1,4) entre NAG y NAH formándose un Disacárido Pentapeptídico. 2. Transpeptidación: Los enlaces polipeptídicos son modificados por la enzima Transpeptidasa. Queda una cadena de repeticiones de Disacáridos Tetrapeptídicos. La energía de la transpeptidación es utilizada para la formación de las uniones peptídicas entre cadenas de Peptidoglucano. La Penicilina inhibe la Transpeptidasa. Autolisinas: Se encargan de cortar los Peptidoglucanos preexistentes, permitiendo la incorporación de Peptidoglucanos recién sintetizados y el crecimiento celular. En el caso de Cocos, se diferencian Zonas de Crecimiento: regiones específicas donde se produce la acción de autolisinas y el crecimiento de la PC. En las bacterias Gram +, los Peptidoclucanos son sintetizados en la región más próxima a la mbr plasmática, mientras las capas externas se van perdiendo. En Gram – Deben incorporarse a la PG varios productos sintetizados por la célula, como los LPS. Existen estructuras específicas que facilitan la salida de los precursores. Las más frecuente son las Uniones Bayer. Las Uniones Bayer: Entre hemimembrana externa plasmática y hemimembrana interna de la mbr externa. 7 Las Bacterias Acido-Alcohol Resistentes (AAR), o micobacterias: Grupo responsable de la tuberculosis. Presentan un modelo de PC distinto al de Gram + y Gram -. La mbr plasmática está recubierta por una capa de Peptidoglucano. Esta, a su vez, está recubierta por tres capa externas; de la más interna a la más externa son: Glúcidos: Arabinogalactanos. Glucolípidos: Lipoarabinomanano. Lípidos: Ácidos micólicos (enormes lípidos). Ralentizan la entrada de nutrientes por su espesor y efecto hidrofóbico. Las micobacterias presentan un crecimiento muy lento, debido a su compleja PC. 25 8 Casos especiales: Los Micoplasmas: Carecen de PC y son resistentes a la penicilina. Tienen forma esférica deformable. Presentan en su mbr plasmática esteroles similares a los Eucariontes. Las Archaeas Termófilas o Termoplasmas viven a 80-90 ºC y tampoco presentan PC. Presentan una mbr plasmática particular. Protoplastos: La Lisozima digiere sus Peptidoglucanos y su pared celular. Generalmente el protoplasto abandona los restos de su PC digerida, y no la necesita para mantener su presión osmótica. En algunos casos puede tener lugar la Lisis celular. C/ El interior de la Célula Bacteriana; La Información Genética: 1 El Nucleoide: La célula procarionte no presenta compartimentación ni núcleo celular. El Nucleoide es una molécula circular de DNA bacteriano: Es su único cromosoma. No rodeado por mbr. Como excepciones, hay bacterias que presentan membrana nuclear, varios cromosomas, o cromosomas lineales. El Nucleoide o Cromosoma bacteriano constituye el genoma de la bacteria. Los Plásmidos son moléculas de DNA, que suelen ser más pequeñas. No son parte esencial del genoma bacteriano, pero suelen aportan ventajas selectivas a sus portadores. Pueden autorreplicarse. El cromosoma bacteriano suele encontrarse empaquetado por proteínas no Histonas. En Dominio Archaea son similares a Histonas y se produce superenrollamiento positivo. En Bacteria se produce Superenrollamiento Negativo del cromosoma bacteriano, mediado por enzimas Girasas (tropoisomerasas), que lo estabiliza. Otras proteínas empaquetadoras asociadas se unen al DNA superenrrollado, formando una estructura tipo estrellas, cuyos brazos reciben el nombre de Dominios. 2 Replicación del DNA: El Modelo de Replicación más común en Cromosomas Bacterianos es el modelo o semiconservativo. La replicación se inicia en un punto concreto Ori C del cromosoma circular, y es Bidireccional: tiene lugar en ambos sentidos simultáneamente terminando en el punto Ter C. Se diferencian la Cadena Lider leída de 5’ a 3’, y la cadena retardada en la que se forman los fragmentos de Okazaki. Girasas: desenrollan el DNA permitiendo su lectura y replicación, introduciendo superenrollamiento positivo. Helicasas: Abren la doble hélice de DNA. Algunas proteínas de unión a DNA monocatenario se unen y mantienen las cadenas separadas. DNA Polimerasas (I, II y III) leen y replican la información de las Bases Nitrogenadas. Reconocen y dependen de un fragmento Iniciador de DNA de su cadena molde. Tienen capacidad para reparar sus errores. RNA Primasas: Leen los fragmentos iniciadores formando el primosoma: Sintetizan Fragmentos Cebadores de RNA. DNA Polimerasa III continúa la síntesis de DNA de 5’ a 3’ a partir del primosoma. DNA Polimerasa I corta los fragmentos cebadores y los reemplaza por DNA. DNA Ligasa enlaza los fragmentos de DNA. DNA Polimerasa II tiene la función exclusiva de reparación de errores. 26 Durante etapas de división, antes del fin de la síntesis de un cromosoma comienza la replicación del cromosoma hijo. Los procariontes son siempre Haploides pero pueden presentar varias copias de su cromosoma. Salvo algunas excepciones en Archaea, Los procariontes no suelen tener intrones en sus genes, y nunca presentan sistema mitótico o del huso. 3 Organización de los Genes y Transcripción en procariontes: Los genes de procariontes se encuentran organizados en Operones, o conjuntos de Genes que son transcritos todos en bloque. Un mRNA va a transportar la información de varios genes a los Ribosomas. Todas las proteínas correspondientes a un mismo operón están implicadas en una misma función. En la región de Iniciación de la Transcripción presentan un Fragmento Operador. Y antes un Centro Promotor, de reconocimiento y unión de RNA Polimerasa. (TATA box). OPERÓN: Centro Promotor Gen 1 Operador Gen 3 Gen 2 Gen 5 Gen 4 El factor reconoce y se une a los Centros Promotores, activándolos, y permitiendo a RNA Polimerasa unirse al inicio del operón. El Operador, no siempre presente, sirve como Regulador. Algunos represores pueden unirse, inhibiendo la acción de RNA Polimerasa. Se habla de ADN Policistrónico, pues algunos genes se encuentran en la hebra principal, y otros en la complementaria. 4 Regulación de la Transcripción: Los genes inducibles (operones) sólo son sintetizados cuando se vuelven necesarios para la célula, en funcion de la presencia o ausencia de determinadas sustancias en el medio. Su transcripción debe ser regulada. La regulación tiene lugar de tres modos: Mediante factores (existen distintos tipos). Control Negativo: Hay represión de la transcripción. Puede ser por Represión o Inducción. Control Positivo: Hay expresión de Operones. Control Negativo por Represión: Se forma un correpresor que indica la presencia de una sustancia que hace inútil la traducción de cierto operón. Al unirse al correpresor, el represor queda activado y se une al fragmento Operador impidiendo que avance RNA Polimerasa. Control Negativo por Inducción: Se forma un correpresor que indica la presencia de una sustancia que hace útil la traducción de cierto operón. Al unirse al correpresor, el represor queda inactivado y liberado del fragmento Operador al que estaba inicialmente unido, permitiendo así que avance RNA Polimerasa. Control Positivo: Se forma un inductor que indica que la transcripción es necesaria. Al unirse al Activador lo activa permitiendo que se una al DNA antes del Centro promotor, y facilitando la unión de RNA Polimerasa. Proteínas Kinasas transmembrana sensoras: Son las que detectan la presencia de ciertas sustancias del entorno que implican regulación de la información genética. Al unirse al sustrato quedan fosforiladas en su dominio externo. Se desencadena una cadena de fosforilación de Kinasas que puede terminar el la activación o inhibición de la transcripción. 27 D/ El interior de la Célula Bacteriana; El Citoplasma: 1 MreB; Citoesqueleto Baceriano: MreB es una proteína similar a la Actina de Eucariontes, que polimeriza formando filamentos que hacen las veces de citoesqueleto bacteriano. Forman bandas en el interior de la membrana celular, sobretodo en Bacilos. 2 Ribosomas 70S y Traducción del mRNA a Proteínas: Son complejos de proteínas y rRNA de tres tipos, ligeramente inferiores a los de eucariontes. Hay dos subunidades 50S y 30S. Forman pequeños gránulos en el citoplasma bacteriano. 50S: rRNA 23S y rRNA 5S. 30S: rRNA 16S. Durante la traducción, el codón iniciador siempre codifica un aa especial que luego será eliminado; f-Met (formil metionina). Los polirribosomas son estructuras tipo pluma que se forman a lo largo de un mRNA cuando está siendo transcrito simultáneamente por numerosos Ribosomas. La Iniciación de la traducción está marcada por una secuencia de iniciación al comienzo del Operón de mRNA: Secuencia de Shine-Dalgarno. Se une primero la subunidad 30S del Ribosoma al mRNA, y luego la 50S. El proceso gasta GTP, e intervienen proteínas Factores de Iniciación. La Elongación es mediada por los tRNA que activan a los aa y los unen al péptido biosintetizado. tRNA pasa por el sitio A (Aceptor), P (Peptidil) y E (Salida) de la subunidad 50S. El proceso gasta GTP, e intervienen proteínas Factores de Elongación. La Terminación de cada gen está marcada por un Codón de terminación. Intervienen factores de Terminación. Al final del operón se forman bucles de ARN que impiden al Ribosoma que siga la traducción. Algunos antibióticos actúan selectivamente contra los Ribosomas 70S. Ciertas toxinas bacterianas inhiben la acción de factores de elongación Eucariontes, impidiendo la síntesis de proteínas. 3 Inclusiones del Citoplasma: Son los gránulos de mayor tamaño del citoplasma bacteriano. Suelen ser acúmulos de sustancias de reserva, que se forman ante la abundante presencia de un elemento nutritivo en el medio. Carbono: Nunca se almacena en forma de lípidos. Suele ser Gucógeno o PHB (Poli-Hidroxibutirato) Fósforo: Polifosfatos Azufre: Es acumulado en bacterias que necesitan azufre como fuente de E o poder reductor. Estas inclusiones pueden estar rodeadas por capas de lípidos (no membrana) o proteínas. Otras inclusiones concretas: Carboxisomas: Cristalizaciones de Enzima Rubisco en bacterias autótrofas. Vesículas de gas: En bacterias acuaticas, son impermeables y regulan la flotación. Al tomar o expulsar gas permiten a la bacteria situarse a la profundidad óptima. Se forman de aire y dos tipos de proteínas: GVP-A (Dan volumen a la vesícula) y GVP-C (estabilizan a GVP-A). Magnetosomas: Se encuentran en algunos organismos marinos que viven entre los sedimentos. Se trata de acúmulos de Magnetita (Fe3O4). Fototactismo: Algunas bacterias utilizan las vesículas de gas para situarse a la profundidad donde reciben la de luz que puede serles útil. Magnetotaxis: Los magnetosomas permiten a las bacterias detectar el campo magnético terrestre para determinar su desplazamiento entre los sediementos marinos. 28 E/ Estructuras y Apéndices externos de la Célula Bacteriana: 1 El Glicocalix: Se trata de una cubierta externa y bastante impermeable presente en muchas bacterias. Su espesor es variable y se forma fundamentalmente de Glucoproteínas y Polisacáridos, pero puede variar en función de la especie y los nutrientes presentes en el medio. Se encuentran dos tipos distinguibles de Glucocaliz: Cápsula: Estructura rígida y definida. Capa Mucosa: Flexible y amorfa. Una de sus principales funciones es la de adherencia entre integrantes de Colonias y acumulación de nutrientes y agua evitando desecación, por la formación de Biopelículas o Biofilms, que proporcionan además adherencia a, por ejemplo, las rocas de los ríos. Otra importante función es la protección de la célula ante bactericidas o fagocitosis, proporcionando mayor resistencia en hospedadores. Cuanto más espesa es la cápsula, más dificulta la fagocitosis A menudo, en una misma especie, la cepa virulenta es aquella con mayor espesor de cápsula (Ejemplo: Bacillus anthracis). 2 Fimbria y Pili: Fimbria son apéndices cortos y numerosos en la cubierta bacteriana. Sus funciones no son bien conocidas, pero entre ellas está probablemente la de adhesión a tejidos de hospedadores. La Proteína Adhesina de las Fimbrias es fundamental en bacterias patógenas. Algunas Fimbrias especialmente reciben el nombre de Pili. Pili Sexuales o Conjugativos, muy largos y escasos, permiten la conjugación genética entre dos bacterias de la misma especie. Las bacterias son capaces de transferirse copias de Plásmidos a traves del Pili. Durante la conjugación, se da el nombre F+ a la célula donante, y F- a la receptora. Otros Pili están también relacionados con la adhesión de bacterias patógenas a los tejidos. Fimbria (y Pili) presentan receptores para el anclaje de Fagos. 3 Apéndices implicados en la movilidad bacteriana; Los Flagelos Bacterianos: Los Flagelos son apéndices mucho más grandes que Fimbria. Son largos y finos, y están unidos a la célula por un extremo. Son distintos a los flagelos de Eucariontes. Existen varios tipos de Flagelación en función de la disposición de los flagelos: Monotrica: 1 flagelo en disposición polar. Lofotrica: Dos penachos de flagelos en polos opuestos celulares. Peritrica: Flagelos dispuestos sobre toda la superficie celular. El Flagelo se forma de un Filamento, un Gancho, y una base anclada a la membrana celular. El Filamento es una estructura helicoidal, con longitud de onda constante para una misma especie. El movimiento se basa en la rotación del filamento sobre sí mismo a modo de hélice de barco. La Base, anclada a la membrana, es responsable de la rotación del filamento. Se forma por un eje central y un sistema de anillos transversales (4 en Gram -; 2 en Gram +). Incluyen además proteínas Fli entre el 1er y 2do anillo, y proteínas Mot externas a los anillos. En Gram -, los dos primeros anillos (S, M) se encuentran anclados a la membrana plasmática de la bacteria, con las proteínas Mot y Fli. El tercer anillo (P) se encuentra a la altura de la capa de Peptidoglucano. El último anillo (L) se encuentra a la altura de la membrana externa. Las Proteínas Mot permiten la entrada de H+ al interior celular. De tal manera logran aprovechar el gradiente quimioosmótico como energía para el movimiento. 29 Las Proteínas Fli son las conmutadoras del motor: Se encargan de transformar esa energía en el giro de los anillos S ,M, asegurando la rotación del flagelo. Determina si el giro es Horario o Antihorario. El Gancho es una estructura formada por un único tipo de proteína, que une el Eje de la Base con el Filamento del Flagelo. Esta ligeramente curvado, de acuerdo con la forma helicoidal del Filamento. El Filamento está formado de proteína Flagelina. Es helicoidal, y tiene una específica para cada especie en función del tipo de flagelina. Se encarga de transformar la energía de rotación de los anillos en movimiento celular. 4 Endoflagelos: Son Flagelos Internos especiales de Espiroquetas. Se trata de Filamentos Axiales responsables de la morfogénesis y movimientos específicos de este tipo de Bacteria. E/ Movilidad Bacteriana: 1 Tipos de Movilidad: La movilidad bacteriana puede ser de tres tipos: Por Vesícula de Gas (medio acuoso). Flagelar (medio acuoso). Deslizante (sobre sólidos). Suelen orientar sus movimientos en función de su percepción sensorial, en busca de nutrientes o evitando sustancias tóxicas. 2 Movilidad Flagelar: En Función del Giro de Fli, el movimiento es CW (Horario) o CCW (Antihorario). Las Bacterias Monotricas pueden nadar, parar y cambiar de dirección. Cuando giran CCW nadan hacia delante. Cuando giran CW van marcha atrás. Las bacterias Peritricas tienen un modo de nado más complejo. Se produce una alternancia en el tiempo entre CCW/CW que se traduce en una alternancia Carrera/Voltereta. CCW: Carrera; la bacteria nada en línea recta. Hay Enlazamiento de los flagelos: Todos se disponen paralelamente. CW: Voltereta o Giro; la bacteria cambia aleatoriamente la dirección de nado. Los flagelos se separan y empujan en direcciones contrarias. 3 Quimiotaxis; Percepción Sensorial relacionada con Movimiento Flagelar: La Quimiotaxis es la capacidad de percepción de Nutrientes y Repelentes Químicos en el entorno. No se trata simplemente de la percepción de presencia o ausencia de sustancias, sino de su gradiente de concentraciones. Gracias a la Quimiotaxis las bacterias pueden nadar a favor de grad de nutrientes o en contra de grad de repelentes. Cuando no se detecta gradiente, Carrera/Voltereta se alternan aleatoriamente. Al detectarse un gradiente, las volteretas se hacen menos frecuentes cuando la carrera se produce en la dirección deseada por la bacteria, por ejemplo, huyendo de sustancias tóxicas. A pesar de detectar grad, poseen unicamente un tipo de receptor para la sustancia. La célula toma muestras del medio cada poco tiempo, varias durante una carrera, pudiendo evaluar si la concentración ha aumentado o disminuido, hasta que alcanza la saturación de los receptores. Se comparan resultados en el tiempo, no en el espacio. 30 Receptores de Metilo: Son un grupo de receptores de Quimiotaxis. Al unirse la sustancia analizada, el dominio interno del R se metila produciéndose la transducción. Proteínas Che: Relacionadas con cascadas metabólicas, regulan la movilidad flagelar bacteriana. Son activadas cuando al ser fosforiladas. CheY – P actúa sobre Fli estimulando la rotación CCW. En su presencia aumenta la longitud de la Carrera, o disminuye la frecuencia de Volteretas. Cuando CheY no tiene P, aumenta la frecuencia de Volteretas. Bacterias Flageladas Capilar con Nutrientes Capilar con Repelentes Durante la división celular de las Bacterias Flageladas, la síntesis de los Flagelos es muy compleja, e implica varias decenas de genes. Se sintetizan, por este orden Eje y Anillos profundos, Mot y Fli, Anillos externos, Garfio, Filamento. Durante la síntesis del Filamento, Flagelina es Biosintetizada en el Citosol y sale a través del eje central, que es un Tubo Hueco, pudiendo así alcanzar con relativa facilidad el extremo terminal del Filamento para ensamblarse ahí. Si se produce una rotura del Flagelo, Puede regenerarse por el mismo proceso. 4 Movilidad Deslizante: Es, al igual que la Flagelar, uno de los tipos de movilidad más frecuentes. Sin embargo, está menos estudiado este último proceso. Se trata de un movimiento más lento, similar al arrastre, que precisa de una superficie sólida. En Gram -, se sitúan, en contacto al nivel del peptidoglucano, dos tipos de proteínas: Uno de ellos en la membrana plasmática, el otro en la externa. Por rotación, las proteínas internas desplazan a las externas, que a su vez se anclan al medio sólido. Mbr Plasmática Proteína Interna Peptiodoglucano Proteína Externa Mbr Externa Movimiento Bacteriano 31 F/ Ciclos Celulares de las Bacterias: 1 La División Celular en Bacteria: El tipo de división más común es la Fisión Binaria, aunque en algunos casos se producen divisiones asimétricas, como gemación o crecimiento polar. La Fisión Binaria de las Bacterias suele tener lugar en tres pasos básicos: 1. Replicación del Cromosoma. 2. Síntesis de nuevo material: De la Cubierta celular (mbr, PC). Ribosomas en Inclusiones. Plásmidos. Flagelos. Enzimas y Metabolitos. 3. Formación del Divisoma, y del Septo, generalmente transversal, y separación de las células. El Divisoma de las Bacterias adquiere la funcionalidad de Citocínesis. También se forma en Mitocondrias y Cloroplastos. Intervienen diversas proteínas: Fts Z: Se encuentran dispersas en el citoplasma durante crecimiento activo. Es de estructura similar a la Tubulina de Eucariontes. Durante la división se dispone formando un anillo a nivel del secto, en la cara interna de la membrana plasmática. Otras proteínas son responsables de la síntesis de de Peptidoglucano y mbr plasmática. Una vez formado el Divisoma, el anillo contractil de Fts Z sufre una constricción que lleva a la separación física de ambas células. Una mutación en Fts Z puede conllevar la aparición de minicélulas: Normalmente Fts Z encuentra justo el medio de la Célula para formar el anillo. Algunas Bacterias tienen varios cromosomas: Deben ser repartidos equitativamente. Sin embargo, las bacterias no tienen aparato mitótico y se desconoce la naturaleza del proceso. 2 Procesos de Diferenciación Celular; Las Endosporas: Se trata de los procesos mediante los cuales se produce la formación de tipos celulares morfológica y fisiológicamente distintos al tipo celular más frecuente. Las Endosporas son formas de resistencia bacterianas, frecuentes en Gram + del suelo, como el género Bacillus. Se forman y maduran en el interior de la célula, y posteriormente son liberadas al entorno. Se forma exclusivamente una por célula. La Endospora contiene la información genética de la célula, y al germinar dará lugar a un nuevo cuerpo vegetativo, con capacidad de división por Fisión Binaria. Sin embargo, los genes que se expresan en la endospora son distintos que en el cuerpo vegetativo. Se diferencian las Esporas No Deformantes, más pequeñas que la célula que las contiene, de las Esporas Deformantes, de mayor tamaño que el cuerpo vegetativo bacteriano. Clostridium: Género causante del Tétanos, son anaeróbios estrictos. Las formas esporuladas pueden soportar, sin embargo, la presencia de oxígeno. Las Endosporas son, de hecho, la forma de resistencia biológica más inalterable conocida hasta la fecha: Termorresistencia: Soporta hasta 100 ºC (Asepsia: 20’’, 120 ºC). Resistencia a Rayos UV. Resistencia a compuestos químicos tóxicos. Resistencia a la desecación. Resisten condiciones adversas durante muchos años, incluso siglos. Los motivos de su alta resistencia también son varios: Son metabólicamente inertes. Germinan sólo en medios adecuados. 32 Son impermeables. Para su tinción se precisa de técnicas especiales a altas T. Presentan alto nivel de deshidratación en el citosol Poseen proteínas protectoras contra radiaciones en el citosol: SASP. Sirven como fuente de carbono durante la germinación. Además de deshidratación y proteínas SASP, en el citosol de pH especialmente ácido se encuentra un compuesto característico que es acumulado: Ácido Dipicolínico. Se asocia a Ca 2+. Citosol PROTOPLASTO o NUCLEO PC + Mbr Plasmática CORTEX: Peptidoglucanos similares PC de Gram + CUBIERTA: Proteica EXOSPORIO: Proteico, No siempre presente Estructura de la Endospora Bacteriana Estas formas celulares fueron responsables de las principales causas de controversias durante la época de la Teoría de la Generación Espontánea. 3 Esporulación de las Endosporas: Es el proceso de esporulación mediante el cual la Célula comienza la Difereciación Celular y formación de Endosporas. El proceso es inducido por la elevada concentración de productos Tóxicos o la carencia de Nutrientes. La esporulación requiere energía por lo que se inicia antes de la desaparición total de los nutrientes, o saturación letal de tóxicos. En poblaciones grandes, las células emiten Señales de Densidad Celular cuando detectan condiciones que favorecen la Esporulación. Son sustancias responsables de la comunicación celular bacteriana: Cuando su concentración en el medio sobrepasa un determinado umbral, las células de la población, que poseen receptores para dichas sustancias, inician la Esporulación. El proceso comienza de modo similar a la división celular, pero es Asimétrica: C Madre Vegetativa: Tamaño normal. Preespora: Pequeño tamaño. Después de la formación del Septo, las dos células hijas no se separan: Se produce una lisis del Peptidoglugano en el Septo y la Célula Madre invagina a la Preespora, la cual comienza su maduración como Endospora. Tiene lugar en varios pasos: 1. Síntesis del Cortex a partir de dos tipos de Peptidoglucano. 2. Síntesis de la cubierta y el Exosporio proteicos. 3. Síntesis del Ácido Dipicolínico, que se asocia en forma de sal con Ca 2+, y SPSA, pequeñas proteínas específicas de la espora. 4. Deshidratación del Citosol. 5. Detención del Metabolismo. 6. Lisis de la Célula y liberación de la Espora. 33 4 Genes Spo; Los Genes de la Esporulación: La expresión de los Genes Spo está regulada por señales externas. Estas desencadenan cascadas de fosforilación de Kinasas que terminan desencadenando la síntesis de Factores específicos. Fosforilación Kinasa A Fosforilación Spo OF Fosforilación Spo OA Fosforilación Su expresión es regulada por medio de cuatro Factores . Mientras en C vegetativas se encuentra A, al iniciarse la esporulación: Célula Madre: E y K. Preespora: F (Tempranos) y G (Tardíos). Regulan la expresión selectiva de los Genes Spo da lugar respectivamente a: Célula Madre: Cortex, Cubierta, Ácido dipicolínico. Preespora: Peptidoglucano del Núcleo. Proteínas receptoras responsables de la Germinación. 5 Germinación de las Endosporas: Cuando los receptores de la Espora perciben las condiciones óptimas, estimulan la Germinación. Suelen percibir presencia de Nutrientes. Se producen cambios metabólicos y fisiológicos típicos del cuerpo vegetativo, dejando de ser células retráctiles. Se reactiva el Metabolismo. Las proteínas de la Espora sirven como fuente inicial de Carbono. En la endospora hay acumulación de fosfoglicerato y fosfoenolpiruvato, pero no ATP. La célula vegetativa, rehidratada, abandona la cubierta de la espora. El proceso tiene lugar en dos etapas: Germinación (pocos minutos): Hidratación; Activación metabólica; Hidrólisis del Cortex; Pérdida del Ác Dipicolínico; Síntesis de ATP a partir del fosfoglicerato. Pérdida de la capacidad de resistencia. Crecimiento Celular (2 horas) 6 Otros procesos de Diferenciación Celular; Caulobacter: No forma esporas, pero presenta un ciclo vital y procesos de diferenciación celular muy especiales en Bacteria. Las Células hijas presentan Flagelos y Fimbrias. Son Células de vida libre, Nadadoras. Tiene lugar un Envejecimiento Celular, caso raro entre bacterias: La celula pierde fimbrias y flagelos, y desarrolla una Prosteca, o prolongación Celular, Con un Botón Mucoso de Fijación al Sustrato en el extremo. Sólo las células viejas, Ancladas al sustrato, tienen capacidad de división: Se forma un complejo de C hija y C madre, ancladas al sustrato, llamado Célula Predivisional. Posteriormente la C hija madura, se desprende y comienza a nadar en el medio. Caulobacter son oligotrofas: viven en lugares con bajas concentraciones de nutrientes. 7 Otros procesos de Diferenciación Celular; Mixobacterias: Mixobacterias es un grupo de bacterias particular: Presentan Movimiento Social formando Cerpos Fuctíferos, en ocasiones visibles. Son un grupo numeroso e importante. El cuerpo fructífero produce Mixosporas (distintas a las Endosporas), de resistencia. En condiciones adecuadas, germinan, dando lugar a cuerpos vegetativos que proliferan por División Celular. 34 Ante carencia de nutrientes, las Mixobacterias producen señales de comunicación celular, que conlleva la agregación celular de los cuerpos vegetativos que se desplazan por deslizamiento, dando lugar a los grandes Cuerpos Fructíferos, de morfologías características según cada especie. Son especiales, pues presentan Comunicación Celular y formación coordinada de Sistemas Multicelulares. A diferencia de las endosporas, las Mixosporas no son células Hijas, sino algunos Cuerpos Vegetativos que proceden a la Diferenciación Celular. Presentan menor resistencia que las Endosporas. 8 Otras Bacterias que desarrollan formas de resistencia: Azotobacter: Quistes. Cianobacterias: Heterocistos (Fijan nitrógeno Atmosférico). 35 II; EL CRECIMIENTO BACTERIANO: A/ Introducción: 1 Introducción: Muchos de las ideas planteadas en esta sección pueden aplicarse también al resto de microorganismo. Se Plantean la acción de factores ambientales en el crecimiento y el control del crecimiento. Los estudios es crecimiento microbiano encuentran múltiples aplicaciones: Proliferación de MO patógenos (medicina). Deterioro de materia orgánica (alimenticia). Detener o Anular el crecimiento de MO (antisepsia). Producción industrial de fermentados (alimenticia). El crecimiento bacteriano puede plantearse como crecimiento celular o crecimiento poblacional. 2 Generalidades sobre crecimiento bacteriano: El crecimiento celular es el aumento del tamaño celular o Biomasa. Tiene lugar la Biosíntesis, Polimerización y Ensamblaje de los componentes de la célula. El crecimiento poblacional suele tener lugar en bacterias por fisión binaria, y es por lo tanto un crecimiento geométrico exponencial. Al cabo de n generaciones: Nº = 2n. En diez generaciones se puede pasar de 200 células a millones de ellas. Suceden alrededor de 2000 reacciones previas a la división celular. Sin embargo, el número de células (Nº) puede doblarse en 20-30 minutos. El crecimiento poblacional puede representarse aritméticamente, obteniendo como resultado una curva con aumento exponencial (N / t). También puede representarse semi-logarítmicamente (log N / t), de forma que obtendremos una recta más facil de estudiar, en lugar de una curva. N Log N Tiempo 3 Las Técnicas de medida: Se diferencian principalmente dos tipos de técnicas de medida de la densidad de una población celular. Se trata de las técnicas Directas e Indirectas. Las técnicas directas de medida se basan en recuentos de parámetros intrínsecos de la población, como los recuentos directos del número de células, o el cálculo de la masa celular de la población. 36 Este tipo de recuentos no suelen ser exactos; siempre deben emplearse métodos estadísticos para evitar y descartar posibles errores. Pueden emplearse para controles de calidad. Las técnicas indirectas de medida se basan en la detección de cambios inducidos por la población bacteriana en el medio en el que viven, como cambios en la concentración de oxígeno en aerobias (disminuye), en la concentración de proteína y ácido nucleico (aumenta), o en el consumo de sustrato disminuyen iones y nutrientes. B/ Las Técnicas Bacteriana: de Medida de Densidad de una Población 1 Método directo por recuento directo al microscopio: Existen portaobjetos excavados micrométricos especialmente habilitados para esta técnica directa, una de las más sencillas pues sólo precisa de éste portaobjetos y un microscopio. Se deben realizar inoculaciones de un volumen constante y recontar el Nº de células por volumen de medio. Por ejemplo, contando el Nº por mm, se puede calcular por mm 2, luego por cm3 y así por mL. Esta técnica no permite diferenciar las células viables de aquellas que están muertas. Por ello no es la más adecuada para analizar infecciones o alimentos, donde las células viva son las únicas importantes. 2 Método directo por diluciones seriadas: Inicialmente se realiza un banco de diluciones de la muestra. Suele diluirse 10 veces en cada paso. Posteriormente, se siembra 1 mL de cada dilución, por extensión, en una Placa Petri donde se incuba, o se vierte la dilución en el medio líquido, antes de vertirlo en la placa y dejarlo enfriar, de modo que las UFC aparezcan tanto en superficie como en el interior de la placa. El medio utilizado puede ser general o selectivo. Se descartan las placas con poblaciones demasiado numerosas o demasiado escasas. El óptimo se encuentra entre a 30 y 500 UFC. Se debe tener en cuenta no solo el número de UFCs, sino también el número de dilución, es decir la concentración de muestra natural en la dilución. Dado que una UFC proviene por división de una sola bacteria (en teoría) podemos contar a simple vista el número de bacterias que había en el mililitro de dilución incubada. Número dilución Número UFCs de 10-1 10-2 10-3 10-4 10-5 de Incontables (14.287) Incontables (1.431) 159 17 2 En este caso: 159.103 UFC / mL. 3 Método directo por rastreo de poblaciones por incremento de la masa celular: Este método, más fiable, precisa de instrumental específico: un colorímetro. Se realiza una Turbidimetría. Al chocar con las células de la muestra, parte de la luz emitida es desviada, de tal modo que a mayor densidad celular, menor será el foco de luz transmitida. Emisor MUESTRA Foco de luz incidente Foco de luz transmitida Receptor 37 A = log (I0 / I) = . l. C A: Absorbancia; % de luz adsorbida. (T = Transmitancia, % de luz transmitida; T = 100 - A). = Coeficiente de extinción molar. l = Distancia recorrida por el foco. C = Concentración (Nº de células). 4 Método directo por el peso de la población: Puede realizarse por peso húmedo. El peso húmedo celular de una población puede calcularse separando las células de su medio por medio de centrifugación o filtración. Las diferencias entre poblaciones más o menos densas es pequeño, por lo que la técnica no es muy fiable. Puede realizarse por peso seco. Después de centrifugación o filtración se somete a la muestra a la desecación, a 80 ºC durante 12 horas. Las diferencias en peso seco celular son sensiblemente superior y notablemente más significativas que las anteriores. 5 Métodos indirectos: Suelen precisar de equipamientos informáticos avanzados, capaces de detectar la presencia de moléculas. A pesar de ser caros son bastante precisos y no alteran la población de la muestra. Algunas cuantifican productos moleculares de la célula, como proteínas o ácidos nucleicos. Otras cuantifican el consume de nutrientes y de oxígeno (Respirometría). Nº de Células Nº de Células Concentración de productos de la célula Concentración de nutrientes del medio C/ Estudio del Crecimiento de una Población: 1 La curva de crecimiento poblacional: Las curvas de crecimiento se realizan gracias al seguimiento de la densidad celular de una población, sobre un cultivo Bacht; es decir, en medio aislado. Se obtienen por Turbidimetría o por Recuento de Viables, pues las células muertas no nos interesan. En la curva se distinguen claramente cuatro fases distintas descritas a continuación. Fase de latencia: Periodo durante el cual las células se adaptan a su nuevo medio de cultivo. Se trata de una fase corta durante la cual no hay crecimiento de la población. Será tanto más larga como grande la diferencia del nuevo medio con el medio anterior. Fase exponencial de crecimiento: La población, cuyas células están adaptadas, comienzan su fase de crecimiento exponencial. El medio abunda en nutrientes y las bacterias ya se han adaptado a él. 38 Fase estacionaria: Se inicia cuando uno de los nutrientes comienza a escasear, y/o los desechos metabólicos comienzan a acumularse en el medio. Mientras algunas células aún se dividen, otras comienzan a morir de modo que se alcanza un equilibrio, y la cantidad de células viables parece mantenerse constante. Fase de muerte celular: En un medio poco nutritivo y plagado de residuos tóxicos, las células del cultivo Bacht comienzan a morir. La población desaparece también exponencialmente, pero a menos velocidad. Fase de Latencia Fase de Crecimiento exponencial Fase de Muerte Celular Fase Estacionaria Curva de crecimiento: Representa N (tamaño de la población) en función del tiempo. Por ejemplo, durante el proceso de Pasteurización de la leche se produce una reducción de la tasa de microorganismos, que se encuentran en una fase de latencia prolongada por la refrigeración. Sólo dos horas fuera de la nevera pueden servir a las bacterias de la leche para superar la barrera de la fase de latencia. 2 Expresión matemática del crecimiento poblacional: La utilización de una representación semi-logarítmica, recta, facilita de nuevo el estudio del crecimiento, durante la fase de crecimiento exponencial. La velocidad de crecimiento de la población en dicha fase coincide con la pendiente de la recta, y recibe el nombre de Tasa de Crecimiento: . 39 depende de varios factores, como la especie de bacteria, el tipo de medio de cultivo y su compatibilidad (pH, POSM, presencia de oxígeno…), y de la concentración en nutrientes del medio de cultivo. La tasa de crecimiento se define matemáticamente como la variación del número de individuos (N) respecto al tiempo (t): = N / t = . N - Si integramos la expresión, y luego tomamos antilogaritmos, obtenemos: ln N = ln N0 + . t N = N0 + et Por otro lado encontramos el Tiempo de Generación; tg, otro parámetro que también depende de la especie de bacteria y, dentro de cada especie, del tipo de medio de cultivo. Recibe también el nombre de tiempo de duplicación, pues es el tiempo que tarda una célula, desde que aparece, en dividirse en dos. Coincide con el tiempo que tarda en duplicarse la población: N = N0 . 2n (partiendo de una célula; N = 2n) Se realiza una determinación gráfica de tg, gracias a la fase de crecimiento exponencial de la curva de crecimiento, observando el tiempo que tarda la población en doblar su densidad. N 2.104 Aquí, tg = 3h 104 6 9 t (horas) Generalmente, en condiciones óptimas, tg varía entre 1 y 6 horas. Existe una clara relación entre tg y , de manera que a mayor tasa de crecimiento, menor será el tiempo de generación, y viceversa: N = N0 + et ; N = 2 . N0 2 . N0 = N0 + etg 2 = etg ; es decir, ln 2 = tg = ln 2 /tg 40 3 Importancia y técnicas industriales del seguimiento de una población bacteriana: El control del crecimiento de poblaciones bacterianas es muy importante, en estudios de laboratorio. Se utilizan poblaciones de células, y no solo bacterianas, para el estudio de encimas, metabolismo o producción industrial. Suelen utilizarse aparatos tecnológicos muy específicos, entre los cuales destaca el Quimiostato. Se trata un aparato para el cultivo de microorganismos capaz de mantener las condiciones óptimas fisico-químicas en el medio de cultivo. Está provisto de un sistema de renovación del medio, de forma que su volumen se mantenga constante. El cultivo se mantiene así en fase de crecimiento exponencial. La tasa de crecimiento puede ser regulada a su vez por medio de la Tasa de Dilución (D) de nutrientes en el medio renovado (opuesto a la concentración), o por medio de la Velocidad de Flujo de medio de cultivo fresco. De ambas formas podemos limitar la concentración de nutrientes máxima en el medio, de manera que controlamos la densidad a la que la población del medio iniciará la fase estacionaria, en la que quedará estable. Nunca se inicia la fase de muerte en este estado. Funcionamiento del Quimiostato Cuando D es baja, hay un aporte grande de nutrientes (su concentración es alta), pero sin embargo el caudal necesario es bajo. La acumulación de tóxicos y grandes concentraciones de nutriente puede ser letal para las células, y por lo menos alarga su tg. Por lo tanto hay mayor cantidad de células eluyentes que nuevas células que aparecen. 41 Cuando, sin embargo, D es muy alta, el caudal de medio de cultivo Fresco (= Q Efluente) es necesariamente más grande, y por lo tanto hay más células que eluyen con el medio, que células que aparecen por mitosis, a pesar de su bajo tg. Recibe el nombre de Lavado (Washout). En condiciones de equilibrio (Steady state) (D = ), la densidad celular permanece constante, pues las velocidades de aparición o tg (Doubling time), y de elución de células es igual. Así, finalmente, se puede controlar N en las condiciones de equilibrio, regulando la concentración de nutriente limitante (es decir D dentro de las condiciones de equilibrio). A mayor [nutriente], mayor N. D/ Efectos de los Factores Ambientales sobre el Crecimiento: 1 Interés Industrial: El estudio de la influencia de factores ambientales sobre el crecimiento de cada grupo bacteriano adquiere un papel importante para la ciencia, pero también para la industria. Para el estudio de las áreas de distribución de seres vivos en la tierra, las bacterias suelen presentar áreas naturales cosmopolitas, prueba de su gran capacidad de adaptación. Sin embargo, algunas bacterias se han especializado para cubrir algunos nichos con factores climáticos y edáficos muy extremos. Su conocimiento puede ayudar en la medicina y la industria alimenticia para lograr mayor rentabilidad, recreando dichas condiciones en un Quimiostato. También es útil en el control de crecimiento de determinada poblaciones microbianas, para el desarrollo de técnicas de asepsia, antisepsia y control alimentario. 2 La Temperatura: Por lo general, a mayor temperatura, más rápido suceden los procesos metabólicos en las células. Las temperaturas que estimulan exponencialmente el crecimiento de una población fluctúan entre una T mínima y una T óptima. En la primera el crecimiento es mínimo mientras en la segunda el crecimiento es máximo ( máximo). A partir de T óptima, y en adelante, se produce un descenso brusco de la población hasta la T máxima. Cada especie bacteriana tiene sus propias T mínima, T óptima y T máxima. A partir de T máxima, el T mín T opt T max crecimiento cesa drásticamente. A temperaturas inferiores a la T mínima, la membrana, no fluida, dificulta el acceso de los nutrientes al interior celular. El gradiente de protones, no funcional o inestable, no permite la síntesis de ATP. Las altas temperaturas dañan irreversiblemente las proteínas, macromoléculas y estructuras de la célula. La alteración de la membrana es responsable de un colapso de la membrana y la lisis térmica de la célula. Aunque generalmente la T óptima fluctua entre 30 y 40 ºC, como hemos visto, muchas bacterias se han adaptado a medios extremos, entre ellos temperaturas extremas, por lo alto o por lo bajo. - Conociendo la T óptima de un tipo bacteriano, podemos cultivarlo a dicha T para obtener la máxima m, o a temperaturas muy superiores (superiores a la máxima) para lograr la esterilización. 42 Psicrófilos 0 5 10 15 20 Mesófilos 30 35 40 Termófilos 50 60 Extremotermófilos 70 80 90 100 110 (ºC) Temperatura En esta tabla encontramos los principales grupos de bacterias en función de su espectro de temperaturas de crecimiento. Se encuentra, resaltado, en cada caso, la temperatura media óptima. Las bacterias Psicrófilas estrictas suelen encontrarse en océanos, polos y el la alta montaña. Son capaces de sobrevivir en pequeñas burbujas de agua líquida a 0 ºC en el hielo, pero no pueden dividirse a tan bajas temperaturas. Pueden ser Endolíticas (viven en rocas) Pueden proliferar fácilmente a la temperatura normal de una nevera, pero no soportan T > 15 º C. Las bacterias Psicrotolerantes pueden subsistir en refrigerados e incluso congelados, pero con más dificultad para proliferar. Pueden sin embargo soportar T > 15 ºC. Pueden vivir en lugares con mucha amplitud térmica día/noche. Las Adaptaciones al Frío suelen estar relacionadas con lípidos de membrana y ácidos grasos con más insaturaciones; predominancia de -hélice sobre -laminar en las proteínas; y predominancia de aminoácidos polares. Las bacterias Mesófilas son las más comunes, pues se adaptan a las temperaturas actuales más comunes en la superficie de nuestro planeta. Pueden encontrarse en aguas dulces, suelos y sueros. Las bacterias Termófilas se encuentran en ambientes naturales de elevadas temperaturas, zonas de insolación intensa como desiertos, y zonas volcánicas. Pueden proliferar en calefacciones y agua caliente de lavado o industrial. También pueden aparecer en pilas de compostage y pilas de ceniza de carbón. Las bacterias Extremotermófilas suelen ser Archaea, pero tambien hay alguna Bacteria. A menudo son Quimiolitótrofas. Se encuentran en regiones de calentamiento geotérmico volcánico, como los Geighers o volcanes de profundidades oceánicas, a menudo en zonas sulfatadas. También en algunos ambientes artificiales industriales. A partir de los 100 ºC, el agua sólo es líquida a muy altas presiones. Las bacterias capaces de aguantar semejantes condiciones extremas no solo soportan, sino que necesitan estas altas presiones para subsistir. Se trata de bacteria Barófilas (Barofilia) que habitan también las profundidades oceánicas. Las Adaptaciones al Calor son básicamente las inversas a las adaptaciones al frío. Además hay abundancia de Chaperoninas que repliegan aquellas proteínas que pierden, debido a la actividad causada por el calor, su estructura nativa. Por otro lado, el ADN suele encontrarse compactado por 43 proteínas tipo Histona en Archaea (distintas de las Eucariontes) que evitan que se desnaturalice el ADN, haciéndolo Termoestable. Acumulan mayores concentraciones de soluto en el medio intercelular para aumentar su T de ebulición. Por último, los Enlaces tipo Eter entre lípidos Isoprenoides característica de la membrana celular de Archaea aumenta también su termoestabilidad. 3 El pH: Igual que con la temperatura, cada grupo de bacterias soporta un pH que fluctúa entre uno niveles mínimo, óptimo, y máximo. Las bacterias Acidófilas soportan y necesitan pH < 5,5, como muchos alimentos, suelos volcánicos (Archaea Hipertermófilas), o los jugos gástricos (patógenas). Soportan metodos de conserva de alimentos como el ácido láctico, las vinagretas y los encurtidos. Las bacterias Acidotolerantes son bacterias Neutrófilas capaces de soportar también condiciones ácidas. Las bacterias Neutrófilas son aquellas que soportan y necesitan pH entorno a 7. Las bacterias Alcalófilas soportan y necesitan pH > 8,5. Suelen encontrarse en suelos y lagos alcalinos, y pueden resistir a desinfectantes como el jabón y el amoniaco. Este tipo de bacterias suelen ser también Halófilas, pues viven en medios con altas concentraciones en ClNa. 4 La Osmolaridad: De nuevo, la presión osmótica de los medios líquidos y húmedos es un nuevo limitante para la conquista de nichos, pero la capacidad de adaptación de las bacterias triunfa de nuevo. Distintos grupos de bacterias resisten [NaCl], mínimas, máximas y óptimas, distintas. La Halofília se define como la necesidad de una alta concentración de sal en el medio de cultivo. 44 La mayoría de bacterias del continente son No Halófilas. Estas no presentan ninguna adaptación a medios con alta concentración de nutrientes, y lisan en medios de alta osmolaridad. Se trata por lo tanto de organismos oligotrofos. Los Oligotrofos presentan transportadores de nutrientes de alta afinidad pero baja especificidad. Por ello en aguas muy cargadas de nutrientes la célula tiende a hincharse de nutrientes y estallar con una Aw int < Aw ext. La mayoría de bacterias son Halotolerantes. Las bacterias Halófilas suelen vivir en mares y océanos. Las bacterias Halófilas Extremas son, de nuevo, generalmente, Archaea. Adquieren su principal importancia en las conservas en salazón, desecados como la mermelada, y ahumados. Cuando las bacterias se encuentran en un medio con Aw (actividad de agua, inversa de la P osmolar) ditinta de su Aw interior, y no tienen capacidad de Osmorregulación, se produce plasmolisis y la célula muere. Algunas células tienen capacidad de Osmorregulación, pudiendo adsorber (transportar) no tanto agua como iones, hasta alcanzar el equilibrio Aw ext = Aw int. Se biosintetizan y activan canales y proteínas transportadoras que introducen iones, como el K+, aminoácidos, o alcoholes. Estas bacterias pueden aguantar mayores fluctuaciones en la [NaCl] del medio. Otra alternativa de Omorregulación o adaptación a las altas osmolaridades se basa en la síntesis de solutos compatibles en el interior celular, que no son tóxicos pero aun así disminuyen el Aw int. Puede tratarse de biosíntesis de azúcares, alcoholes, o aminoácidos. A menudo las bacterias emiten señales celulares que estimulan por Quórum Sensing la expresión de operones de Osmorregulación, que activan a su vez la biosíntesis de transportadores o enzimas específicas. El Choque Osmótico es un cambio brusco de la osmolaridad del medio. Resulta nocivo para las bacterias que no presentan ningún sistema de respuesta por Osmorregulación. 5 Tolerancia al Oxígeno: La presencia o ausencia de Oxígeno es el último principal condicionante de la capacidad de un grupo de bacterias para colonizar y proliferar en un medio. Las bacterias Aerobias son aquellas que precisan la presencia de oxígeno en el medio de cultivo en el que crecen. Las bacterias Anaerobias facultativas son bacterias que pueden vivir en presencia de oxígeno, y utilizarlo en su metabolismo, tanto como prescindir de él. Las bacterias Anaerobias aerotolerantes no necesitan el oxígeno, pero sin embargo pueden tolerar su presencia en el medio de cultivo. Suelen presentar metabolismos fermentativos, y no tienen cadenas de transporte de electrones para el oxígeno, por lo que no forman sus formas tóxicas. Las bacterias Anaerobias estrictas sólo son capaces de subsistir en medios anaerobios, pues el oxígeno les resulta tóxico. En las cadenas de oxidación (respiración) el oxígeno acepta electrones produciendose formas tóxicas. O2 Dioxígeno O2Anión Superóxido O22Anión Peróxido Muchas bacterias producen eficientes enzimas Detoxificantes que eliminan las formas tóxicas del oxígeno, acceptando los e- en la respiración aerobia, formando agua y dioxígeno. Entre ellas se encuentran Catalasa (H2O2), Peroxidasa (H2O2), y Superóxido Dismutasa (O2-, produce H2O2). Las bacterias que no presentan estas enzimas mueren en presencia de O 2. Existen medios y técnicas de cultivo específicas para cultivar bacterias Anaerobias. Pueden utilizarse medios de cultivo semisólidos en tubos de ensayo. El oxígeno alcanza sólo las capas superficiales, donde viven organismos aerobios. En el medio viven los microaerófilos: el 45 oxígeno, si bien presente, se encuentra en bajas concentraciones. En el fondo del tubo se dan las condiciones anaerobias estrictas. En otros casos se utilizan catalizadores; compuestos reductores que atrapan y se combinan con el oxígeno libre, bajando su concentración, y produciendo gases inertes. Estos productos pueden introducirse, junto con las placas de cultivo, en Jarras de Anaerobios: Recipientes específicamente diseñados, de cierre hermético. Se pueden colocar también sistemas indicadores de Red-Ox, que cercioren la ausencia de oxígeno. Para Anaerobios estrictos extremos se utilizan Cabinas anaerobias: Se trata de cámaras totalmente carentes de oxígeno, con gases inertes en su interior, que permiten la manipulación de los cultivos desde el exterior. E/ Control del Crecimiento Bacteriano: 1 Generalidades: El hombre debe a menudo aplicar un control sobre el crecimiento microbiano, por ejemplo, para frenar epidemias y enfermedades, evitar el deterioro de los alimentos, o la producción de fermentados. - Se diferencian tres tipos fundamentales de control sobre el crecimiento microbiano: - Esterilización: Proceso de destrucción de toda forma de vida viable presente, inclusas aquellas metabólicamente inactivas, como endosporas. Desinfección: Destrucción de las formas de vida vegetativas de microorganismos patógenos o indeseados. No destruye las formas esporuladas. Antisepsia: Desinfección de tejidos vivos, sin dañar dichos tejidos con productos tóxicos. Suele emplearse en pre-cirugía y desinfección de heridas. Por otro lado se encuentran distintos Agentes de Control: Agentes Físicos Agentes Químicos Calor seco Calor Húmedo Filtración Radiación Ultrasonidos Microondas Óxido de Etileno Lejía Metales Pesados Arsénicos Alcoholes y Fenoles Alcohol Acidificado Agua Oxigenada Permenganato Halógenos (Cloro, Yodo) Sulfamidas Antibióticos 46 Por último, los Agentes Químicos pueden dividirse por su acción Bactericida (1) o Bacteriostática (2) y Bacteriolítica (3): N (1) totales N viables (2) totales N (3) totales viables t t viables t 2 La Esterilización: En los laboratorios de microbiología, el material debe ser esterilizado para evitar que se altere el medio o los microorganismos estudiados. La Esterilización Preparativa es esencial para asegurarse de que el material de trabajo es estéril, evitando la contaminación de muestras. La Esterilización Final es importante después de trabajar, para eliminar los microorganismos con los que hemos trabajado. Se debe seleccionar un método de desinfección que no dañe el material de nuestros instrumentos. Por lo general, el método más eficaz es por calor húmedo (transmitido por vapor de agua) en el Autoclave. Sin embargo, algunos materiales no resisten el calor, por lo que es conveniente utilizar Radiaciones, Ultrasonido, Microondas u Óxido de Etileno. El Autoclave es un aparato capaz de generar unas condiciones de Sobrepresión: altas presión y temperatura. Se pueden alcanzar temperaturas de 120 ºC sin ebulición del medio acuoso. (Generalmente; 121 ºC; 15-20 minutos; 1,1 atm). Esteriliza material y líquidos en corto tiempo. En algunos casos, como la esterilización del asa de siembra, se emplea el calor seco por Incineración. La esterilización de ambientes es también importante en algunos laboratorios, y en los Quirófanos de los hospitales. Para ello se suele utilizar sistemas con radiación UV que provocan la formación masiva de dímeros de Timina, entre otras mutaciones perjudiciales.. La Tyndalización es un tipo de esterilización fraccionada, para material no resistente a temperaturas superiores a 100 ºC (100ºC, 30 minutos, en ciclos de 24 horas). Se utiliza un Autoclave. Este calor no es capaz de destruir endosporas, pero sí erradicar las formas vegetativas. Después de aplicar el calor húmedo, se deja un reposo de 24 h para que germinen las formas de resistencia, y de nuevo se aplica un ciclo de calor. El proceso debe repetirse durante tres días. 3 La Pasteurización: Se trata de un tipo de desinfección especial para alimentos. Se Utiliza un Horno Pasteur. No destruye endosporas, y sólo elimina una parte importante del número de células vegetativas viables. (63-66 ºC; 30 minutos. HTST: 72 ºC; 15 segundos). La leche pasteurizada puede aguantar dos o tres días fuera de la nevera. La leche puede ser un foco de transmisión de patógenos, como Salmonella tify, Tuberculosis o Brucelosis. La cerveza de botella suele estar Pasteurizada (la de barril no, de ahí su sabor distinto). El vino también puede estar Pasteurizado. Cada tipo de bacteria tiene su propia sensibilidad al calor, y su Tasa de Muerte Térmica. Este parámetro es calculado para diseñar sistemas de esterilización y desinfección industrial, en función de las poblaciones. 47 4 La Filtración: Las células, lejos de ser destruídas, son separadas de el medio líquido en el que se encuentran, quedando retenidas en un filtro. Sólo se aplica a fluídos. No puede considerarse esterilización pues los poros dejan pasar a los Virus, pero sí desde el punto de vista bacteriano. El sistema de filtración se basa en un matraz con succión, sobre cuyo cuello se colocan, por este orde, un adaptador de goma, una base, una plataforma de vidrio, un filtro de membrana y un embudo. El líquido que llega al matraz es esteril. A menudo, para manipular microorganismos patógenos, se utiliza una Campana de Siembra. Un filtro de radiación UV esteriliza el aire interno de la campana. 5 Desinfección y Antisepsia: Para rebajar notablemente N de las poblaciones de microorganismos patógenos suelen emplearse Agentes Químicos. En particular, para antisepsia, se suelen elegir aquellos que son menos dañinos para nuestros tejidos. Si no, en general, se emplean los más potentes y menos corrosivos. Los Metales Pesados son muy eficaces, pero relativamente tóxicos, pues son venenos enzimáticos. Entre ellos encontramos Sal de plata (Ag), Mercromina (Cr), SO 4Cu (Cu), Arsénicos (As, predecesor de los antibióticos, utilizado por Darwin)… Los Halógenos (Cl, I) son agentes oxidantes que en baja concentración no son dañinos para los tejidos. Entre ellos Betadine (I), e hipoclorito o lejía (Cl). El Peróxido de Hidrógeno (H2O2), o Agua oxigenada, tóxica para bacteriasanaerobias, en muy empleada para desinfectar heridas. Los Alcoholes y Fenoles disuelven los lípidos de membrana y desnaturalizan las proteínas. Son también muy utilizados en antisepsia. Entre ellos se encuentra el alcohol 96º y el alcohol Isopropílico. El Coeficiente Fenol (CF) es un parámetro que evalúa la eficacia de los Agentes Desinfectantes Químicos. Se utiliza como desinfectante de referencia el Fenol (CF = 1). Para probarlo se utilizan poblaciones de Staphylococcus aureus y Salmonella typhi. Se toman muestras entre 5 y 10 minutos y se calcula CF. Los Agentes Bacteriostáticos no eliminan, simplemente impiden la proliferación de la población bacteriana. No suelen ser tóxicas para el humano. Son muy importantes para las industrias Farmaceutica, cosmética, Alimentaria, y para la prevención del biodeterioro. Los Agentes Conservantes mantienen las condiciones Bacteriostáticas desde el momento en que el producto es envasado. Entre ellos destacan las Salazones, Mermeladas, Ahumados, Fermentación Láctica, Encurtidos, Ácido Acético (vinagre) y Desecados. 6 Agentes Antimicrobianos Quimioterapeuticos: Se trata de productos útiles para tratar epidemias e infecciones, y cuya toxicidad es selectiva para determinados microorganismos sin afectar a los pacientes. Se trata de las Sulfamidas, y principalmente de los Antibióticos (Bála Mágica). Las propiedades de las Sulfamidas fueron descubiertas en 1920. Inhiben la síntesis de una vitamina, el ácido fólico, necesario para la bacteria. Los húmanos no sintetizamos esta vitamina aunque debemos ingerirla. Posteriormente se descubrió el primer antibiótico; la Penicilina. Esta generación de fármacos hacen que, hoy en día, muchas enfermedades parezcan mucho menos letales de lo que realmente son. El antibiótico se define como un Agente Bioquímico con acción antimicrobiana específica, que generalmente ha sido producido por otro MO con fines de competencia ecológica. Actualmente los Antibióticos naturales son químicamente modificados en laboratorios, para aumentar su eficacia o 48 reducir su toxicidad para el humano. Reciben el nombre de Antibióticos de Amplio Espectro aquellos que son efectivos tanto con bacterias Gram – como Gram +. Existen varios grupos de Antibióticos. Entre ellos destacan: -Lactámicos: Por ejemplo, Ácido-6-AminoPenicilánico. Aminoglucósidos: Son aminoazúcares con enlaces glicosílicos, más eficaces con bacterias Gram – como la tuberculosis. Afectan a la subunidad Ribosomática 30S. Macrólidos: Presentan anillos Lactona unidos a núcleos de azúcares. Por ejemplo la Eritromicina, empleada contra Legionella, y en pacientes alérgicos a la Penicilina. Ataca la subunidad ribosómica 50S. Tetraciclinas: Amplio Espectro. Ataca la subunidad ribosómica 30S. Muchos antibióticos pueden tener efectos secundarios como sordera o ennegrecimiento dental. Las bacterias pueden adquirir resistencia a antibióticos por procesos genéticos de intercambio de información, dando lugar a nuevas Cepas resistentes. A pesar de no deber abusar de los antibióticos, no debemos dejar de hacer uso de ellos. 49 III; PARTICULARIDADES BACTERIANO: DEL METABOLISMO A/ Introducción al Metabolismo en Procariontes: 1 El Metabolismo; Generalidades: Por su mecanismo de acción, las Enzimas permiten múltiples reacciones vitales en el interior de toda célula. Este facilita la Biosíntesis y Degradación de múltiples compuestos inprescindibles, además de la producción de Enlaces de Alta Energía biológica. Estas reacciones tienen su Energía de activación, y a menudo ni siquiera son espontáneas. Por ello la acción catalizadora de la enzima se hace indispensable, regulando la velocidad de dichas reacciones. El Metabolismo y las Enzimas relacionadas hacen viable la vida de toda Célula, Procarionte o Eucarionte. Muchas Enzimas necesitan la unión de un Cofactor (inorgánico) o un Coenzima (orgánico) para permitir su actividad. Otras necesitan utilizar enlaces de Alta energía para poder catalizar determinadas reacciones. ATP (Adenosina TriFosfato) tiene un enlace de alta energía en el tercer grupo Fosfato, unido en posición . Al igual, Fosfoenolpiruvato, acumulado en endosporas, contiene energía química en la unión del fosfato. También, por ejemplo, Acetyl Fosfato. Fosfoenolpiruvato: OO- P O- Enlace de Alta Energía O C CH 2 COO- Acetyl Coenzima A (Acetyl CoA) posee también un enlace de alta energía; el enlace Tiol formado entre el grupo Acetyl y el átomo de Sulfuro. Todas estas moléculas se caracterizan por contener energía química en potencia, que es cedida a las reacciones Catalizadas Enzimáticamente que lo requieran. 2 Tipos de Metabolismos generales: Antigua Clasificación: Anabolismo: Biosíntesis. Requiere Energía, Materia y Poder Reductor. Catabolismo: Degradación. Libera Energía. Oxidación. Actual Clasificación de las Rutas Metabólicas: Productoras de Energía. Productoras de Precursores Metabólicos. Biosintéticas. 3 Tipos de Rutas Metabólicas Bacterianas: Productoras de Energía: Oxidación de Compuestos Orgánicos. Fase Luminosa de la Fotosíntesis. Oxidación de Compuestos Inorgánicos (Bacterias Quimiolitotróficas). Como vimos anteriormente, las Bacterias son capaces de sintetizar ATP gracias a la energía transmembrana debida al Gradiente Iónico. Se crea una Fuerza Protomotriz, y un Potencial Eléctrico. 50 También pueden obtener ATP Metabólicamente, por fosforilación a nivel de Sustrato con enlaces de alta energía. Obtención de Precursores Metabólicos: Ruta de la Glucolisis. Ruta de Entner – Doudoroff. Rutas Fosfato. Ciclo de Krebs (TCA) Ciclo de Calvin (Autótrofos). TCA inverso (Autótrofos). Las Bacterias necesitan asimilar compuestos Inorgánicos. Fundamentalmente C a partir de la Glucosa, pero también O, N, H, P y S. 4 El Poder Reductor: Se trata de la indispensable capacidad de Oxidarse de ciertos compuestos para reducir otros en las Rutas Metabólicas. Estos compuestos pueden perder Protones, y pos lo tanto ceder sus electrones. Existen transportadores de e- que aseguran que el electrón es cedido al compuesto debido. Por ejemplo, el coenzima NAD+/NADH: Muchas enzimas tienen dos Sitios de Unión; uno para el sustrato y otro para NAD+. Durante la reacción se oxida NAD+ a NADH, y el Sustrato es reducido. Son similares los casos de NADP+/NADPH y de FADH+/FADH2. El Poder Reductor es esencial para los procesos de Biosíntesis, y también para las cadenas de transporte de electrones de la Respiración Celular. FUENTE DE CARBONO TIPOS TRÓFICOS EN BACTERIA FUENTE DE ENERGÍA Inorgánico (CO2) Autótrofos / Litótrofos Orgánico Heterótrofos / Quimiótrofos Luz Fotótrofos Fotoautótrofos* I Fotolitótrofos Fotoheterótrofos Compuestos Químicos Quimiótrofos Quimioautótrofos Quimiolitótrofos Quimioheterótrofos* II Quimioorganótrofos *I: Similares a Eucariontes Fotosintéticos. *II: Similares a Eucariontes Heterótrofos. Desechos FUENTE DE ENERGÍA Movimiento Adaptaciones CATABOLISMO Poder Reductor Energía ANABOLISMO FUENTE DE MATERIA Componentes celulares 51 B/ Nutrientes y Vías de Asimilación: 1 Los Macronutrientes: Los Macronutrientes son los principales Nutrientes que las Bacterias deben absorber del Medio. Suelen ser los mismos en TODAS las Células. Además de ellos necesitan Micronutrientes, en menor nivel de requerimiento, y por último, Factores de Crecimiento: Sustancias orgánicas sencillas no sintetizadas por la bacteria, pero que es tomada del entorno. CARBONO: De compuestos orgánicos u inorgánicos (CO2). HIDRÓGENO: Para el poder reductor, suelen tomarlo del Agua. OXÍGENO: Aceptor final de e- en respiración aerobia, también lo toman del agua. NITRÓGENO: Elemento clave de muchas proteínas y Bases Nitrogenadas. FÓSFORO: En enlaces de Alta energía, Nucleótidos y Fosfolípidos. AZUFRE: En aminoácidos indispensables para muchas proteínas, Vitaminas, CoA, donadores de e (fotosíntesis), y aceptores de e- (respiración). 2 Los Micronutrientes: El requerimiento es mucho menor, es decir, en mucha menor concentración. HIERRO: El más fundamental de todos ellos. Interviene en la Fotosíntesis, Fijación del N, Actividad de numerosas Enzimas y Proteínas, Citocromos, Muchas bacterias forman Sideróforos: Estructuras especializadas que unen Fe con facilidad, secuestrando el Fe del hospedador, pues presenta mayor afinidad. POTASIO: Para muchas enzimas implicadas en Biosíntesis de proteínas. MAGNESIO: Para Actividad Enzimática y Estabilización de Ribosomas, Membrana Plasmática y Ácidos Nucleicos. CALCIO: Importante para la Pared Celular y esporulación de Endosporas. SODIO: Para bacterias Halófitas y Halófitas estrictas (muchas Achaeas). COBRE: Para Citocromo C y Plastocianina (Fotosíntesis). OLIGOELEMENTOS: Requerimiento aún más reducido, mínimo. Cobalto, Zinc, Niquel… 3 Asimilación de Nutrientes: Los Precursores Metabólicos suelen contener ya el C, H y O. Las Células Bacterianas deben incorporar por Vías de Asimilación particulares los otros Macronutrientes; N, S y P. Además deben incorporar los Micronutrientes y Oligoelementos. 4 Incorporación del Nitrógeno: Las principales fuentes de N orgánicas son grupos Amino, Imino y Heterociclos. Las principales fuentes de N inorgánicos, que deben ser incorporados, son el Nitrógeno molecular (0), Nitritos(-5), Nitratos(-5) y Amonio(-3). La incorporación tiene lugar en forma de Amonio, cuyo estado de ionización es -3. Nitratos y Nitritos son transformados en Amonio por una Reducción Asimilatoria metabólica de ciertas bacterias Fijadoras de Nitrógeno: Nitrato-Reductasa Asimilatoria NO3 Nitrato Nitrito-Reductasa Asimilatoria NO2 Nitrito NH4+ Amonio 52 El Nitrógeno Molecular también puede ser transformado en Amonio, en las bacterias Fijadoras de Nitrógeno: Nitrogenasa N2 Nitrógeno NH4+ Amonio La incorporación del Amonio Puede realizarse por dos vias: Directa o Indirecta. Cuando en Amonio está en alta concentración, se realiza por vía directa. Lo hace la Enzima Glutamato dh, que no gasta energía. En caso contrario, se hace por vía directa, por medio de la Glutamin Sintetasa, de mucha mayor afinidad pero que gasta ATP, formando ácido Glutámico. 5 Incorporación del Azufre: Muchas bacterias obtienen S orgánico del Medio- Las principales fuentes exógenas de S inorgánico son transformadas en SH2, de tal forma que pueden ser asimiladas. La asimilación de sulfatos requiere altos Poder Reductor y gasto Energético. SH2 solo se encuentra estable en medios anaeróbicos. Tiene lugar por medio de la siguiente Reducción Asimilatoria: Acetilserina Sulfohidrolasa SH2 + Acetilserina Cisteína 6 Incorporación del Fósforo: Se asimila a partir de Fosfatos, sin gastos Energéticos ni de Poder Reductor. Es incorporado generalmente por medio de Rutas Catabólicas. Tiene lugar en Procesos de Fosforilación de ATP, a partir del ión Fosfato inorgánico. La membrana celular es impermeable a Fosfatos orgánicos. Puede producir encimas que catalizan la Hidrólisis Extracelular de dichos compuestos, produciendo ión Fosfato que es absorbido. Fosfatasa Alcalina; 5-Nucleotidasa Fosfato Orgánico Ión Fosfato C/ Energía, Precursores Metabólicos, Poder Reductor: 1 Energía: Existen cuatro tipos de Vías para generar energía: Heterótrofos: Metabolismo Respiratorio. Metabolismo Oxidativo. Autótrofos: Metabolismo Fotótrofo. Metabolismo Quimiolitótrofo. Hay, además, dos Vías para generar ATP: Gradientes Quimiosmóticos. Fosforilación Oxidativa de Respiración / Fosforilación de Fotosíntesis. Fosforilación a nivel de Sustrato: Fermentación, Glucolisis, Ciclo TCA (Krebs) y otros. 53 2 Precursores Metabólicos: Existen 13 Precursores Metabólicos fundamentales para toda Bacteria. Estos son procesados por las rutas del Metabolismo Central, ya sean reductoras u oxidativas. Algunos de los principales precursores metabólicos son: Glucosa-6-P Fructosa-6-P Ribosa-5-P Eritrosa-5-P Triosas Fosfato 3-Fosfoglicerato Fosfoenolpiruvato Acetil CoA Glucólisis: Glucosa-6-P Piruvato (6 precursores) Ciclo TCA: Acetil CoA CO2 (3 precursores) Vía de las Pentosas Fosfato: Glucosa-6-P CO2 (2 precursores) 3 Poder Reductor: Depende de dos sustancias transportadoras de protones: NADH y NADPH. Son indispensables en Rutas de Biosíntesis. En heterótrofos respiratorios, ceden su e- en la cadena de transporte electrónico, para la síntesis de ATP D/ Fuentes de Energía en Bacterias: 1 Generalidades: La fuente universal de energía Celular es el ATP. El ATP de la célula puede obtenerse por distintas vías: Respiración, Fermentación o Fotosíntesis. 1 Metabolismo Quimioheterótrofo: Utiliza Compuestos Orgánicos. La Respiración es la principal fuente de E, Precursores y P Red. En su lugar, puede tener lugar la Fermentación. Su Fuente primaria son los Glúcidos, también utilizan Ácidos Grasos por -oxidación, y ante carencia de estos, pueden utilizar Aminoácidos de proteínas, formando Acetil CoA. La oxidación de Glucosa en Piruvato, puede darse por: Glucolisis, Ruta de las Pentosas Fosfato o Ruta de Entner – Doudoroff. La oxidación de Acetil CoA en el ciclo TCA forma ATP y Poder Reductor dando lugar a CO 2. La oxidación completa del sustrato donador de e- por Respiración, hasta el aceptor (O, N, S) tiene lugar a partir de Acetil CoA y otros precursores, formando E. Biosíntesis Compuestos Orgánicos CO2 Flujo Electrónico E celular 54 2 Cadenas de Transporte Electrónico de Bacterias: Las Cadenas de Transporte Electrónico tiene lugar a nivel de la hemimembrana interna celular, y está relacionado con proteínas asociadas a la membrana. Se trata de la Oxidación completa del sustrato. Son muy similares a las que ocurren en las Mitocondrias, pero pueden variar ligeramente entre Bacterias. Respiración: El aceptor de electrones puede ser O2 (Aeróbica) o S, N… (Anaeróbica) Fermentación: El aceptor de electrones es Endógeno y Orgánico. En estas Oxidaciones, los Dadores pueden ceder H + y e- o ceder sólo el e- y liberar el protón al medio. NADH suele ser el principal dador de electrones. Hay una cadena de aceptores que se transfieren los e- de unos a otros: NADH Deshidrogenasa: Captan H+ de NADH y transfieren a Flavoproteínas. Flavoproteínas: Aceptan H+ y ceden sólo e-. Proteínas Fe/S (No Hético): Sólo transfieren e-. Quinonas: Liposolubles. Aceptan H+ y sólo transfieren e-. Pueden desplazarse en la mbr. Citocromos: Proteínas con grupo prostético Hemo. Hay varios tipos. Transportan e- hasta el aceptor final: generalmente Dióxido. La Energía es así acumulada gracias a una fuerza protón-motriz. Puede ser transformada en ATP por Bombas ATPasas, utilizada para simporte con protones o para movimiento flagelar… 3 Metabolismo Fotótrofo: En Fotótrofos la Energía es obtenida por medio de Fosforilación de Fotosíntesis. La Energía fundamental tiene por origen la luz del sol. Luz - Flujo electrónico E celular; ATP Fotoheterótrofos: Compuestos Orgánicos - Grad Protónico Biosíntesis Fotoautótrofos: CO2 Biosíntesis El proceso de la Fotosíntesis puede generar energía por medio de Fosforilación Cíclica o No Cíclica. 4 La Fotosíntesis: Síntesis de ATP por Fotofosforilación a partir de la energía de la luz solar. La Fotofosforilación, presente en Cianobacterias, puede ser no cíclica o cíclica en función del medio habitado: No cíclica: Similar a la fotosíntesis vegetal. La luz excita los Fotosistemas II (P680) y I (P700). Hay producción de Energía y Poder Reductor. Cíclica: Sólo funciona el Fotosistema I (P700). Produce Energía pero no Poder Reductor. La base del proceso es la excitación de la clorofila por la luz del sol. Esto facilita el transporte de e , que suelen ser cedidos a NAD+ o NADP+. El donador exógeno de electrones es el H2O. Las Bacterias Fotosintéticas suelen ser bacterias Gram – unicelulares o filamentosas, que poseen estructuras celulares específicas: Los Tilacoides. 55 Los Tilacoides son sistemas internos de membrana independientes, dispuestos paralelamente a la membrana citoplasmática. Actúan como Centros Antena, Centros de Reacción y Cadenas de transporte de electrones. En los Centros de Reacción se encuentra la Clorofila A. Los Ficobilisomas son gránulos de los Tilacoides con Ficobilinas: Se trata de pigmentos de captación de luz (Ficocianina, Ficoentrina). Estas Bacterias suelen poseer vesículas de gas para disponerse en el agua a la altura perfecta para captar su de luz óptima. También suelen presentar gránulos de Cianoficina como reserva de N. Fotosistema II P680 FOTOSÍNTESIS OXIGÉNICA Fotosistema I Luz 2 H2O O2 ; 4H+ P700 Luz NADP+ NADPH El éxito evolutivo de estas formas de vida permite su crecimiento y expansión en gran variedad de nichos: Sus requerimientos son tan básicos como Agua y Luz solar. Gracias a ello comenzó la acumulación de O2 atmosférico, haciendo de Cianobacter un grupo fundamental de la Historia Natural del planeta. Esto supuso una etapa de selección masiva: extinción de especies para las cuales el oxígeno es tóxico, y adaptación a la respiración oxigénica. Bacterias Acuáticas; Cohabitación en estratos, sin competencia: Cianobacterias (Aerobias, Amplio espectro). Bacterias verdes del azufre (Anaerobias, en acúmulos de sulfuro). Bacterias púrpuras, (Anaerobias de infrarrojos en acúmulos de sulfuro). El agua puede ejercer como filtro potente para la luz. Por ello en un único estanque pueden convivir distintos tipos bacterianos en estratos distintos, en función de sus necesidades. 7 La Autotrofía: La mayoría de bacterias con fotofosforilación son bacterias autótrofas, capaces de sintetizar su propia materia orgánica. Deben incorporar el carbono inorgánico en su materia: CO2 - CH2O Para ello, pueden emplear distintos mecanismos: Ciclo de Calvin: Tiene lugar en los carboxisomas; gránulos de acumulación de enzima RubisCo cristalizada. TCA inverso: Carboxilaciones reductivas. Vía TCA en sentido inverso. 56 6 Metabolismo Quimiolitótrofo: CO2 Biosíntesis Compuesto Inorgánico Flujo electrónico NO3SO42O2 E celular; ATP La Quimiolitotrofía es un tipo de metabolismo exclusivo de Bacteria y Arquea. A menudo las permite ser autótrofas (por ciclo de Calvin o TCA inverso), o simplemente utilizar la quimiolitotrofía como fuente de energía (ATP) por fosforilación oxidativa a partir de un donador de e inorgánico. El aceptor de electrones suele ser O2 (aerobias) en Bacteria y otros compuestos moleculares (anaerobia) en Arquea. Los compuestos orgánicos pueden inhibir el crecimiento de este tipo de bacterias. E/ Bacterias Quimiolitótrofas: 1 Generalidades: Existen distintos tipos de bacterias quimiolitotrofas en función de los compuestos inorgánicos que oxidan: Nitrificantes. Sulfooxidantes. Ferrooxidantes. Oxidantes de Hidrógeno Molecular. 2 Las Bacterias Nitrificantes (oxidantes del nitrógeno): Son bacterias quimiolitotrofas estrictas, muy sensibles a la materia orgánica. Son las que presentan mayor especificidad; existen distintos grupos, cada uno especializado en la oxidación de distintas formas del Nitrógeno: Bacterias Nitrosas NH3 Amoniaco Bacterias Nítricas NO2Nitrito NO3Nitrato La nitrificación completa del amoniaco precisa de la actuación secuencial de ambos grupos de bacterias. Estas bacterias, acidófilas, no crecen ante presencia de materia orgánica. Son frecuentes en las aguas residuales urbanas. Las bacterias oxidantes del amoniaco en nitrito requieren de la acción de dos enzimas exclusivas de este grupo: eCitocromo C NH3 + O2 Amonio monooxigenasa - NH2OH + H2O NO2Hidroxilamina oxidoreductasa La energía generada en el proceso es relativamente baja (- 65 kCal / mol). 57 El Nitrito es un compuesto tóxico, y las bacterias capaces de utilizarlo y oxidarlo son especiales. Poseen una enzima integral de membrana, exclusiva de este grupo: NO2- + H2O NO3- + 2H+ Nitrito oxidasa - La energía generada en el proceso es baja (- 18 kCal / mol). 3 Las Bacterias Sulfooxidantes (oxidantes del azufre): Presentan también especificidad ante el sustrato, pero no tan marcado como en nitrificantes. Utilizan como dador de electrones compuestos de azufre S reducidos; SH -, S0, S2O32-. SH- cede sus electrones a FP, en un nivel más avanzado de la cadena respiratoria. El azufre elemental y S2O32- los ceden, sin embargo, al Citocromo C. A menudo las bacterias oxidantes del azufre presentan gránulos de acumulación de azufre elemental S0. Las arqueas sulfooxidantes son un importante grupo de oxidantes de azufre, y suelen fijar el CO2 por el TCA inverso. En Bacteria, esta fijación suele tener lugar por el Ciclo de Calvin. 4 Las Bacterias Férricas o Ferrooxidantes (oxidantes del hierro): Son bacterias quimiolitotrofas acidófilas que oxidan el Ferroso (Fe II) en Férrico (Fe III), en anaerobiosis pues sino la oxidación es espontánea. EXTERIOR PERIPLASMA MEMBRANA 2 Fe 2+ CITOPLASMA H2O Rusticianina Cyt c e- 2 Fe 3+ H+ Cyt a1 ½ O2 + 2 H+ H+ Al aumentar la concentración de H+ intracelular se genera un gradiente electroquímico en la membrana de la célula. Ante un exceso, el pH disminuye y la célula puede morir. La formación de agua evita un pH excesivamente bajo. Este tipo de oxidación suministra muy poca energía. Se forma ATP a partir del gradiente electroquímico. 5 Drenaje ácido de las minas: Tanto las bacterias sulfooxidantes como las férricas suelen proliferar en las minas de Pirita (S2Fe). Destruyen la pirita de las minas y la contaminan con óxido de hierro, a lo que se debe las aguas de color rojo que manan. Thiobacillus sulfoxidans oxidan el azufre de la pirita formando H + y haciendo que disminuya el pH. Thiobacillus ferroxidans oxidan Fe II en Fe III, y este último sedimenta unido a tres átomos de hidroxilo: Fe3+ Fe(OH)3 La Biolixilación es una técnica de extracción de minerales alternativa, que se hace más reentable para la extracción de compuestos en baja concentración (Cu, manganeso, o pirita en baja concentración). Se realiza la oxidación del mineral por acción bacteriana. 58 6 Bacteria oxidantes de Hidrógeno (H2): El H2 es el donador de electrones para la respiración. Se establece así un gradiente de protones que sirve para la síntesis de ATP. La reacción es mediada por la Hidrogenasa, una enzima citoplasmática. 59 IV; GENÉTICA BACTERIANA; ADAPTACIÓN Y MUTACIÓN: A/ Mutaciones: (Láminas A) 1 Tipos de Mutaciones Génicas: Se define mutación como un cambio en las BN originales del individuo y/o su línea genealógica. Las poblaciones de Bacterias presentan una alta tasa de crecimiento, y las mutaciones juegan un papel esencial. Cepa Silvestre: Población cuyos individuos conservan el patrimonio genético original de su especie. Cepa Mutante: Población cuyos individuos comparten una mutación que les diferencia de la cepa silvestre de la que provienen. El origen de esta mutación se encuentra en una sola célula que ha formado una UFC. Cepa Revertida: Población cuyos individuos son fieles al patrimonio genético original de su especie debido a una reversión, pero no lo han conservado de la cepa silvestre. Una Reversión es una mutación que aparece en una cepa mutante, de manera que permite la reaparición del genoma de la cepa silvestre. Ocurre con relativa frecuencia en caso de cepas con mutación puntual, pero es raro en otros casos. Reversión auténtica: El triplete que reaparece es exactamente igual que el original. Reversión equivalente: Aparece un triplete distinto al original, pero que codifica el mismo Nucleótido. La Mutaciones pueden ser Espontáneas si ocurren naturalmente, o pueden ser Inducidas por agentes mutágenos. De tal manera podemos obtener cepas mutantes en cultivo a partir del Aislamiento de UFC mutantes cultivadas. Las ADN polimerasas de Bacterias tienen capacidad de reparar mutaciones y errores en la replicación. La tasa de mutación en bacterias es 10-7 – 10-11 mutaciones/pares de Bases (es decir, 10-4 – 10-8 Mutaciones/Gen/Generación). En virus estas proteínas no tienen capacidad de reparación, y se encuentran niveles de mutación más elevados (10-1 – 10-5 Mutaciones/Gen/Generación), y por ello es difícil encontrar vacuna para muchos virus, como el de la Gripe, pues sus proteínas de membrana cambian constantemente. Las Mutaciones Puntuales cambian un Nucleótido por otro. Pueden ser transiciones (de purina a purina, de pirimidina a pirimidina; ej: de AT a GC) o transversiones (de purina a pirimidina, de pirimidina a purina; ej: de AT a CG). Las Mutaciones de Desfase actúan por inserción o deleción de un nucleótido. Desfasan y alteran así el patrón de lectura de los tripletes, provocando la modificación de toda la proteína. Las Mutaciones a nivel de Código pueden ser Silenciosas (no alteran la proteína), Sin Sentido (la codificación de la proteína queda truncada por la aparición de un codón STOP), o Con Cambio de Sentido, Conservada (el cambio provoca una proteína distinta con la misma actividad), o Radical (la proteína pierde su actividad natural). De tal manera que, a nivel de fenotipo, se distinguen las Mutaciones Silenciosas (la proteína mantiene su funcionalidad) y las Mutaciones con Cambio en el Fenotipo (la proteína cambia o pierde su funcionalidad). 3 Agentes Mutágenos: Entre ellos se encuentran Agentes Mutágenos Físicos, entre los cuales hay radiaciones no ionizantes, como rayos UV, de poca capacidad mutagénica (forma Dímeros de T, de C), y las ionizantes, como los rayos X o , muy potentes pero que entrañan peligro ya que ionizan el agua de las células. 60 Entre los Agentes Mutágenos Químicos destacan los Análogos de Base, 2-Aminopurina y 5Bromouracilo, que pueden unirse a las otras Bases Nitrogenadas del ADN. Pueden ser reparados por ADN polimerasa. 5-Bromouracilo presenta Tautomería Ceto-Enol, de forma que puede reemplazar a T uniendose a A en forma normal Ceto, pero si tautomeriza la forma enol facilita la unión de G, de forma que al ser corregido el error se introduce una C, y por lo tanto induce una Mutación Puntual de Transición (de AT a GC). 3-Aminopurina es un análogo de A, que puede unirse a T y cambiar de conformación facilitando la unión de C. Al ser reparada la doble hélice se introduce G en el lugar que ocupa 3Aminopurina, de manera que se induce una Mutación Puntual de Transición. Los otros agentes químicos reaccionan con el ADN. Los agentes desaminantes provocan transiciones por desaminación. Los agentes intercalantes, como el Bromuro de Etilo, se intercalan entre las bases produciendo inserciones o deleciones. Los agentes Alquilantes provocan transiciones por mutilación o etilación, y a diferencia de los demás hacen su efecto por simple contacto con el cultivo durante determinado tiempo. Los agentes oxidantes modifican las bases pudiendo causar Transversiones. Los agentes que impiden el apareamiento de las bases actúan facilitando, en algunos fragmentos, la desaparición de determinadas purinas (sitios apurínicos). Los Agentes Mutágenos Biológicos son fundamentalmente Transposones: Se trata de fragmentos de ADN que son insertados en la cadena inicial. Pueden acarreas transversiones o deleciones. Su inserción es facilitada por algunos Virus Bacteriófagos. 4 Mecanismos celulares de Reparación: Las células procariotas presentan tres principales mecanismos de reparación del ADN, capaces de reparar las mutaciones poco después de su formación: Reparación directa: Mecanismo de reparación de mutaciones concretas. Por ejemplo, Fotoliasa rompe los enlaces covalentes de los dímeros de Timina causados por las radiaciones UV. Sistema de Escisión-Sintesis-Ligación: Primero se detecta el fragmento de ADN modificado, por ejemplo por presencia de dímeros de Timina. A continuación, una Endonucleasa corta y elimina dicho fragmento. A continuación, DNA polimerasa sintetiza de nuevo el fragmento eliminado a partir de la Hebra guía. Finalmente, Ligasa reestablece los enlaces Fosfodiester. Sistema SOS: Se trata de un tipo de reparación por recombinación, que es activado cuando ambas hebras han sido modificadas. Necesita la activación de varios genes. 5 Mutaciones con Cambio en el Fenotipo y su detección en el laboratorio: Los Mutantes Nutricionales son aquellos con los que se trabaja más frecuentemente en el Laboratorio. Presentan una Mutación con Cambio Fenotípico que afecta a una Enzima fundamental en Rutas de Biosíntesis del Metabolismo. De tal manera que estos mutantes pueden crecer exclusivamente en un medio en el que estén presentes los Productos de la reacción catalizada por dicha Enzima. De distinguen de esta manera dos tipos de Mutantes, en función de si son o no Nutricionales: Protótrofos: Mutantes NO Nutricionales. Auxótrofos: Mutantes Nutricionales. Por ejemplo, en el caso de síntesis de Histidina, los protótrofos son (His +), y los auxótrofos son (His -). Estos tipos de Mutantes, idénticos entre sí morfológicamente, son sin embargo fáciles de diferenciar en medios de cultivo. Para nuestro ejemplo anterior: o En Medio Completo crecen Auxótrofos y Protótrofos. o En Medio Mínimo sólo crecen Protótrofos. o En Medio Mínimo + Histidina crecen tanto Protótrofos como Auxótrofos (His-). 61 Los Mutantes Resistentes a Drogas y Antibióticos pueden haber desarrollado sistemas que rompen o impiden la entrada al interior celular de dichas sustancias. Para su detección se utiliza un Medio Completo + Antibiótico. Por ejemplo en Medio Completo + Streptomicina sólo se desarrollan las bacterias Streptomicina-resistentes. Los Mutantes Estructurales han desarrollado cambios en la estructura celular. Por ejemplo puede haber desarrollado o no una cápsula. En ocasiones las bacterias con cápsulas más gruesas son resistentes a Antibióticos pues dificultan su penetración en la célula. Los Mutantes Restrictivos o Condicionales son Cepas Mutantes que sólo expresan la mutación a determinadas temperaturas. Para la Generación y Contraselección de Mutantes Auxótrofos en el laboratorio, los Microbiólogos proceden de la siguiente manera (XV, XVI): 1. Exposición del caldo de cultivo con la bacteria en tubo de ensayo a un agente mutágeno, durante el tiempo respectivo que necesita cada agente para ser efectivo. Eliminación posterior del Agente. 2. Plaquear en Medio Completo de forma a obtener UFC discretas. Se desarrollan todas las colonias. Gracias a un fieltro replica-placas se pueden obtener copias de las colonias, con situación en cada placa exactamente idéntica a la original. Deben hacerse dos copias en placas distintas: 3. Plaquear en Medio Selectivo, por ejemplo Medio Mínimo + Histidina. En este medio sólo crecerán Protótrofos y Auxótrofos (His -). 4. Plaquear en Medio Mínimo. Sólo crecerán las Bacterias Protótrofas. 5. Comparando los resultados de cada placa se pueden identificar las colonias de la placa original que son Auxótrofas, en este caso para His. Se puede aislar la colonia. La selección puede también realizarse Generación y Selección Directa de Mutantes resistentes, por realización de un Antibiograma (generalmente utilizado para valorar la efectividad de un antibiótico): Para ello, después de la exposición al agente mutágeno, se cultiva el microorganismo en una placa en Medio Completo, en forma de Césped. La adición de un papel de filtro impregnado con el Atibiótico (por ejemplo Streptomicina) permite el crecimiento exclusivo de las colonias que han desarrollado resistencia (XVII). A mayor halo de inhibición, más efectividad del Antibiótico. Para la Detección de Revertientes Nutricionales, debemos partir de una población Auxótrofa aislada y cultivada. Primero es expuesta al agente mutágeno. A continuación es incubada sobre un Medio Mínimo. De esta manera sólo podrán desarrollarse las UFC revertientes Protótrofas (XIX). En todos estos experimentos debe prepararse una placa testimonial que permita detectar la proporción de mutantes espontáneos e inducidos. Para ello se repite el experimento con otra placa cuyas bacterias no han sido sometidas a la acción de agentes mutágenos. Así podemos restar, de la Mutación Total de nuestra placa, el número de Mutación espontánea (XVIII). El Test de Ames permite la detección de la capacidad mutagénica de sustancias estudiadas. El estudio de cada sustancia se basa en un cultivo de una cepa de Salmonella (His -). Estas bacterias son cultivadas en un Medio Mínimo que incluye un papel de filtro impregnado con la sustancia estudiada. En esta placa sólo podrán crecer los revertientes Protótrofos, de forma que a cuanto más crezcan, mayor es la capacidad del agente estudiado. Para comparar se realiza el mismo experimento con un papel de filtro impregnado sólo en agua, de forma a poder observar y restar el número de Revertientes Espontáneos (XX). 62 B/ Plásmidos y Transposones: (Láminas C) 1 Plásmidos: Se define como elementos genéticos Extracromosómicos y transmisibles, generalmente circulares (cerrado covalentemente o superenrollado negativamente por tropoisomerasas), con capacidad de autorreplicación. Pueden ser tan grandes como el cromosoma o mucho más pequeños. No incluyen genes vitales, aunque por lo general otorgan ventajas selectivas para las bacterias que lo contienen. Se presenta una buena definición en (I) (II,III). La transmisión de Plásmidos, generalmente por Conjugaciónm, otorga a una población una ventaja selectiva, como puede ser por ejemplo la resistencia a Antibióticos. Se trata de una transmisión genética Horizontal, muy exclusiva. Los Plásmidos son Compatibles cuando se conservan durante muchas generaciones, e Incompatibles cuando son eliminados tempranamente. La replicación del Plásmido es independiente a la del cromosoma, aunque pueden estar acopladas. En algunos casos el Plásmido puede integrarse al cromosoma. Se distinguen tres principales tipos de replicación, que dependen fundamentalmente del tamaño de plásmido. Esisten varios mecanismos de clasificación de los Plásmidos: Plásmidos Conjugativos: Transmisibles por conjugación, contienen genes tra. Plásmidos no Conjugativos: No contienen genes tra. Pueden ser transmitidos por otro tipo de mecanismo. En función del Control de la replicación vegetativa: Control estricto del número de copias. Frecuente en Plásmidos de bajo número de copias; una o dos. Control no estricto del número de copias. Por el fenotipo que codifican: Confieren Resistencia (Drogas o Antibióticos). Bacteriocinogénicos (Síntesis de Bacteriocinas). De Virulencia (propios de Estirpes o Cepas virulentas, Síntesis de Factores de Virulencia). Factores de Colonización (Síntesis de Enzimas que permiten utilizar distintos metabolitos como fuente de Nutrientes). Plásmidos Crípticos (función desconocida). La Nomenclatura de los Plásmidos sigue el modelo: pXX000, donde p se refiere a plásmido, XX hace referencia a la Bacteria que lo porta, y 000 es el número de kPB (kilo Pares de Bases). Por ejemplo, pR100 es un plásmido de E. coli de 100 kPB. Gen tet: resistencia a Tetraciclina (en Transposón Tn10). pBR322 es un Plásmido Artificial creado por Ingeniería Genética. Esta disciplina se centra en aprovechar la capacidad multiplicativa e Integrativa genética de los plásmidos, para la obtención de bacterias que sintetizan determinadas proteínas deseadas. 2 Transposones y Transposición: También llamados Elementos Transponibles (Tn), son entidades genéticas discretas que pueden formar parte tanto de Plásmidos como del Cromosoma Bacteriano. Son capaces de cambiar de locus, por un salto llamado Transposición. En cada extremo de un Transposón se encuentra su Secuencia de Inserción (IS); repeticiones terminales invertidas de entre 700 y 2000 pares de bases. Las IS presentan un único gen que codifica una proteína Transposasa (enzima implicada en la transposición). El Transposón es de mayor longitud y complejidad que su secuencia de inserción, y contiene información adicional a la necesaria para transposición. En cuanto a la nomenclatura, a cada Transposón y sus respectivas Secuencias de Inserción se les adjudica un número. Por ejemplo, en el Transposón de pR100: Tn10; IS10. 63 La Transposición es el salto de un Transposón y sus Secuencias de Inserción entre dos locus genéticos. Se distinguen dos tipos de Transposición: Conservativa: El elemento transponible salta de un locus a otro distinto. El número de transposones permanece constante. Replicativa: Una copia del elemento transponible salta a un nuevo locus, quedando una copia en cada localidad. El número de transposones se duplica. Tiene lugar en varios pasos: 1. Localización de la Secuencia Diana: La tranposición no sucede entre dos locus aleatorios. Primero la Transposasa debe reconocer una secuencia específica del DNA donde se insertará el elemento transponible; la Secuencia Diana (XV). 2. Corte Asimétrico de la Secuencia Diana: La misma enzima, con actividad endonucleasa, produce cortes asimétricos en dicha secuencia. Este corte asimétrico tiene lugar de manera que al final de todo el proceso queda duplicada la Secuencia Diana a ambos lados del Transposón. 3. Corte de Transposón: Sólo tiene lugar en caso Conservativo. Es el corte mediado por Transposasas del ADN adyacente al lugar de origen del Transposón (XVII). 3'. Cointegración y Resolución: Sólo tiene lugar en el caso Replicativo, y es facilitado por la enzima Resolvasa (XVII). Primero se forma la Estructura Cointegrado; una estructura mixta que integra dos copias del transposón unidas en cada extremo a hebras distintas de DNA (una para cada locus), de manera que las hebras de origen y destino quedan unidas. Se produce Recombinación y Resolución (o separación) de las dos copias en hebras distintas. Por ejemplo, pR1 contiene un transposón (Tn10) con genes de Transposasa, Resolvasa y un gen de Resistencia. Existen otros elementos genéticos móviles, propios de Virus Bacteriófagos, similares a transposones. Se insertan en la secuencia del DNA de bacterias infectadas. El mecanismo de translación recibe el nombre de Transducción. 3 Generalidades sobre el Intercambio de Material Genético entre Bacterias: Estos elementos genéticos descritos anteriormente son los principales elementos que pueden ser intercambiados entre bacterias distintas. Pueden hacerlo por tres procesos distintos: Transformación: Entrada de DNA exógeno al interior de una célula bacteriana. Este material puede tener como origen otra célula bacteriana lisada. Transducción: Dos bacterias pueden intercambiar material genético a traves de un Bacteriófago. Conjugación: Complejo mecanismo especializado para la transferencia de Plásmidos completos o partes del ADN cromosómico. Se produce por un contacto estrecho entre las bacterias que intercambian información genética. 64 65 C/ Transformación y Transducción: (Láminas D) 1 Transformación: Este proceso se produce de forma natural, pero también puede ser forzado en laboratorios con fines industriales de la Ingeniería Genética. El Experimento de Griffith (IV, V) con la bacteria Steptoccocus phneumoniae, Diplococo Gram +, demostró que se producía un intercambio de material genético entre dos Cepas, una virulenta (mortal para ratones) y otra no virulenta. En concreto se diferenciaban colonias lisas (S) virulentas, con cápsulas bien desarrolladas que dificultaban la fagocitosis, y colonias rugosas (R), con una cápsula muy delgada, y casi exentas de propiedades patógenas. Al inocular en un ratón la cepa S muerta por calor, resultaba benigna. Sin embargo la cepa S muerta + cepa R viva, inoculada en un Ratón era Mortal. La Cepa R viva ha debido tomar el ADN de las células muertas de la cepa S, adquiriendo así sus genes de resistencia/virulencia que otorgan una cápsula más gruesa. Para demostrar que dichas células habían tomado el ADN de las células muertas, y no su ARN o sus proteínas, Avery, MacLeod y McCarty desarrollaron pocos años después tres ensayos, similares a los de Griffith, donde se añadían a los productos de la Cepa L muerta Proteasas, Ribonucleasas y Desoxiribonucleasas respectivamente (VI). El hecho de que fuese únicamente en presencia de Desoxirribonucleasas en el que las bacterias de la Cepa R no adquirían una cápsula más gruesa puso de manifiesto que eran los genes, y no otros componentes de las células muertas, los que eran transferidos de unas a otras bacterias. El proceso que permite la llegada del ADN de una a otra bacteria es complejo y no siempre sucede satisfactoriamente (VII). 1. La bacteria donadora es sometida a condiciones de Degradación y lisis. Su ADN es liberado al medio en forma de Exogenotes. 2. Adsorción: El Exogenote (fragmento de DNA de la bacteria donante) penetra en la célula receptora en Estado de Competencia, transformándose en Endogenote. Sólo entra una hebra monocatenaria. 3. Recombinación: En Endogenote monocatenario se integra por recombinación en el cromosoma de la bacteria receptora, llamada entonces Merozigoto. A menudo el Endogenote no es recombinado con el propio ADN de la bacteria receptora, y es digerido y degradado en el interior del Merozigoto. Sólo si el ADN Exógeno es recombinado con el Genóforo (cromosoma), la transformación es totalmente efectiva. En otros casos el Exogenote permanece como elemento extracromosómico, cointegrado con parte de la información genética propia de la bacteria, recibiendo entonces el nombre de Diploide Parcial. Esto sucede de manera natural solamente en determinadas bacterias, llamadas Bacterias con Capacidad de Transformación Natural (VIII). Estado de Competencia: Es el único estado en el que las bacterias con capacidad de transformación natural se muestran receptivas a la entrada y recombinación de ADN exógeno. Es posible inducir en laboratorio este estado en algunas bacterias sin capacidad natural. Son distintos entre bacterias Gram + y -. Este estado es desencadenado por procesos de Quórum Sensing en bacterias Gram + (IX), y por señales específicas independientes de la densidad celular en Gram -. En B. subtilis, ComX es el Autoinductor, que en altas concentraciones desencadena el proceso. La Adsorción, y Digestión o Recombinación del Endogenote es mediada por una serie de proteínas específicas (hasta 100 distintas): En un primer paso actúan unas proteínas tansmembrana de reconocimiento y unión al exogenote, que facilitan la adsorción. A menudo reconocen secuencias concretas, de desde unos pocos pares de bases hasta 20 kPB. Si no se produce reconocimiento, el Exogenote es destruido por acción de Nucleasas; en bacterias Gram + se produce en el exterior celular, y en bacterias Gram – se produce en el espacio Periplasmático. 66 A continuación, durante la penetración, actúa una Endonucleasa que rompe las hebras bicatenarias dejando entrar solamente a una de ellas (Haemophilus es la única bacteria que permite la entrada de cadenas bicatenarias). En el Interior celular, la hebra monocatenaria es muy inestable y se rompe si no es unida a determinadas proteínas específicas de Competencia de unión al DNA monocatenario, propias del estado de Competencia. Esta fase, durante la cual el fragmento de DNA es monocatenario recibe el nombre de Fase de Eclipse; si la célula es lisada en este instante, estos fragmentos de DNA no pueden transformar a otra bacteria (Haemophilus carece de esta fase). La proteína Rec A es responsable de recombinar el exogenote con el Genóforo, formándose así finalmente el Merozigoto. La Transformación Artificial es la transducción forzada en bacterias sin capacidad natural. Puede realizarse a través de Virus, o por otras técnicas de tratamiento químico (CaCl 2 + choque térmico a 42 ºC, se induce estado de competencia en el 10 % de la población) o Electroporación (descargas de alto voltaje en cortos pulsos, más eficaz pues se induce estado de competencia en el 50 % de la población) mediante las cuales se realizan poros a través de la pared celular y la membrana. Este reordenamiento de la cubierta celular facilita la entrada de Exogenotes bicatenarios de gran importancia en la industria para la ingeniería genética. La Transfección es el caso especial en el que el Exogenote no proviene directamente de otra bacteria, sino que lo hace a través de un Bacteriófago. Es el caso por ejemplo de E. coli, sin capacidad de transformación natural. Es el tipo de transferencia descubierta más recientemente, también llamada Transducción. 2 Transducción: Fue descubierta por Lederberg y Zinder, gracias a su experimento (XVI). Introdujeron en un Tubo en U dos cepas de Salmonella typhimurium (una Met -, Thr +, Leu +, Thi +, y la otra Met +, Thr -, Leu -, Thi -), separadas por una membrana impermeable a bacterias. Al observarse que también se producía recombinación genética sin haberse forzado lisis celular, ni haber contacto entre las bacterias de cepas distintas, dedujeron que debía existir un agente capaz de transportar los exogenotes de una cepa a la otra, filtrable a través de la membrana. Actualmente se conocen dos procesos de Transducción: La transducción Generalizada, facilitada por Bacteriófagos Líticos y la transducción Especializada, facilitada por Bacteriófagos atemperados. Durante la Transducción Generalizada, los Bacteriófagos Líticos primero infectan a las bacterias, a continuación se reproducen en su interior, y finalmente son liberados por lisis de la bacteria infectada (XVIII): 1. Injección del DNA en la Bacteria infectada. 2. Este DNA exógeno secuestra la maquinaria de la célula para replicarse, transcribirse y traducirse. Además el DNA propio de la bacteria es fragmentado. De esta forma el virus se multiplica por ensamblaje de los productos. En algunos casos se forman virus que albergan fragmentos aleatorios de DNA propio de la bacteria que han infectado. Estos virus bacteriófagos defectuosos reciben varios nombres: Partícula Defectuosa, Pseudovirión o Partícula Transductante. 3. Cuando los virus con DNA bacteriano injectan su DNA a otra célula bacteriana puede producirse la Transfección: Este fragmento cualquiera de DNA exógeno puede ser eliminado, permanecer inerte o Recombinarse con el Genóforo, como si se tratase de un Endogenote de Transformación. Es un proceso raro y poco exitoso. Durante la Transducción Especializada, los Bacteriófagos Atemperados pueden presentar dos estados, y presentan dos fases respectivas para cada estado (XXII,XXIII): 67 Estado Lítico: En esta fase el virus produce una infección lítica, idéntica a la de los fagos líticos. Estado Lisogénico: En esta fase el virus produce una infección, pero no lisa la célula ni destruye su DNA; Su DNA se permanece latente en el interior de la célula. Sucede de la siguiente forma: 1. El Bacteriófago injecta su DNA. 2. El DNA se integra en el Genóforo propio de la bacteria, en forma de Profago. De tal manera que se replica junto con él y está presente en todas bacterias hijas. 3. El DNA del Profago se desintegra o escinde del Genóforo, y lo destruye. Se puede lograr por inducción en laboratorio, por ejemplo por sometimiento a rayos UV. Si la escisión es Normal se inicia una fase lisogénica normal. Si la escisión es Incorrecta se forman un gran número de Viriones que recombinan una región de ADN del virus y otra de la bacteria. 4. Estos últimos Bacteriófagos defectivos tienen la capacidad de producir una infección en una nueva bacteria con la ayuda de un fago helper, o auxiliar, que se encuentra en forma de Profago en el interior de la bacteria. Sin los genes de este fago auxiliar normal, el fago defectivo no puede llevar a cabo la infección. En este caso, sólo los genes bacterianos adyacentes al sitio de integración del Profago podrán ser transferidos. Además, su transferencia siempre está acompañada del genoma vírico. Un buen ejemplo bien estudiado de Transducción Especializada es el ciclo del Fago Lambda, que infecta E. coli (XX, XXI). Los extremos ATT del DNA del Profago se recombinan en la zona ATT del Genóforo (sitio de integración), adyacente al operón Gal (para degradación de Galactosa). La escisión normal se produce de nuevo entre los sitios ATT. La escisión incorrecta es el resultado de la recombinación de dos regiones homólogas que no son los sitios ATT, y tiene como resultado un virión que incluye parte del DNA vírico, y parte del Bacteriano inclusos los genes Gal. Este bacteriófago defectuoso recibe el nombre de Fago Lambda-d-Gal. 68 69 C/ Conjugación: (Láminas E) 1 Generalidades sobre la Conjugación en Gram negativas: Se trata de un proceso descrito por primera vez en una cepa de E. coli por Lederberg y Tatum en 1946, con lo que ganaron un premio Nobel en 1958 (I, II, III). Se basaron en dos cepas auxótrofas distintas, una para Metionina y Biotina (A), y la otra para Treonina y Leucina (B). Fueron cultivadas ambas en Medio Mínimo, en placas distintas, y a continuación ambas juntas (A + B) en la misma placa también en Medio Mínimo. En el primer caso no se desarrolló ninguna colonia, ya que es muy difícil la aparición de revertientes para dos mutaciones distintas. En el último caso obtuvieron un cesped: debía tratarse de Recombinantes Protótrofos. El posterior experimento en Tubo en U demostró que el proceso necesitaba de un contacto estrecho entre las bacterias. Por otro lado, el experimento en un medio con ADNasa daba resultado normal, y demostraba que el fragmento de ADN transferido nunca se encuentra en forma de Exogenote, libre en el medio. Se distinguen en función de la conjugación distintas cepas bacterianas (X): La Cepa F + es aquella que contiene y puede transferir los genes estudiados, antiguamente denominada cepa macho. Estas cepas deben además contener un Factor de Fertilidad (F’’); se trata del plásmido pF, que es de hecho el material transferido (Lámina C-IX). La Cepa F – es la cepa receptora, que puede aceptar el plásmido de una F +, antiguamente denominada cepa hembra. Las Cepas Hfr (High frecuency of recombination), aisladas y descubiertas más recientemente, son cepas de muy alta frecuencia de recombinación. IS3 Región tra Tn1000 IS3 pF Ori T IS2 Resistencia Ori V CC o SE-; 100kPB. Las secuencias de inserción IS3 de pF se encuentran también en el Genóforo de E. coli, por lo que ambos pueden recombinarse, y tal vez esto este relacionado con el origen de este plásmido. La región tra contiene los genes tra, imprescindibles para la conjugación. Sólo los plásmidos con esta región tra pueden ser transferidos. Codifican unas 60 proteínas implicadas en distintos pasos del proceso de transferencia: Formación de Pili conjugatorios, Estabilización de los complejos de Agregación, Replicación del ADN y Transferencia del ADN a través del puente citoplasmático. 2 El proceso de Conjugación: Se trata de un proceso complejo que tiene lugar en varios pasos: 1. Formación de Agregados de Conjugación; complejos de agregación celular de bacterias de cepas F + y F -. 2. Las bacterias de Cepa F + generan un Pili Conjugatorio o Sexual que crece por polimerización de proteína Pilina F, y contacta con un receptor celular de una bacteria de la cepa F -, formando un lazo entre ambas bacterias. De este modo la pareja queda establecida. 70 3. Comienza la despolimerización de Pilina, de manera que la longitud del Pili se reduce progresivamente, y las bacterias se aproximan hasta que se produce contacto entre sus superficies. 4. Una vez producido contacto físico, se forma, en el lugar que ocupaba el Pili, un puente citoplasmático estrecho y selectivo. 5. Una cadena madre es transferida a través dicho puente de la bacteria F + a la F -. Este proceso es simultáneo al proceso de replicación del mismo plásmido en F +, generalmente según el modelo de círculo rodante (VIII), aunque puede ocurrir también según el modelo theta (IX). La Exclusión Superficial es el mecanismo dispuesto para evitar la recombinación entre Cepas F + o F -. Los genes tra son responsables de este mecanismo por el cual las Bacterias F + rechazan la unión del Pili conjugatorio, mientras las F - son incapaces de producir dicho Pili. La Conjugación implica un elevado coste energético, y además algunos bacteriófagos se sirven del Pili Conjugatorio para producir su infección. Por ello existe un Control Negativo que es ejercido por Quórum Sensing sobre las cepas F +. Asimismo, después de varias generaciones, comienza la producción de Autoinductores (moléculas señal) que inhiben la conjugación; cuando la mayor parte de la población ha adquirido los genes de resistencia y se la transmitirá a su descendencia, seguir emitiendo Pili Sexuales es un riesgo innecesario. Esta situación, en la que la mayor parte de las bacterias de la población son de la Cepa F + y continúan emitiendo Pili, recibe el nombre de Zigosis Letal: Una misma F – puede ser taladrada por varios Pili que tratan de establecer contacto, de forma que puede morir por lisis. La proporción F +/F – debe ser adecuada. La Replicación de Plásmidos según el modelo en Círculo Rodante y su Transferencia son facilitadas por un gran número de proteínas tra. Comienza con la escisión de una sola hebra del plásmido F en Ori T, por acción de una Endonucleasa específica. El extremo 5’ de la hebra cortada, que contiene en el extremo los genes de Resistencia, se dirige hacia el puente citoplasmático de unión a la bacteria F -. Según la hebra cortada es desenrollada y va siendo transferida, la hebra complementaria sirve como molde para su replicación (VIII) (Láminas C-VII, IX). 3 Cepas Hfr: La particularidad principal de estas cepas es que su pF es capaz de insertarse en el Genóforo a partir de las secuencias IS2 e IS3, de tal manera que actúa como Episoma (XI). Por ello, durante la conjugación F + transfiere no solo el plásmido F, sino también el resto de sus genes. Para ello son necesarios alrededor de unos 100 minutos. Este proceso es de hecho utilizado para calcular, en minutos, el tamaño de los plásmidos. (XVI; Genóforo de E. coli en minutos) Se trata de un proceso complicado, ya que las dos bacterias deberán permanecer en contacto durante más de una hora. A menudo tiene por ello lugar un proceso de Conjugación Interrumpida, en cuyo caso solo es transferida una parte de los genes, hasta un momento en el que el proceso se detiene y las células se separan. En función del sentido de la transferencia se transferirán así unos u otros genes (XII). Los genes tra se encuentran al final del fragmento transferido, por lo que en caso de Conjugación Interrumpida estos genes esenciales para pF no son transferidos. Este pequeño plásmido sin genes tra contiene los genes de resistencia y algunos genes cromosómicos. Por otro lado este Episoma puede ser cortado del Genóforo, por Escisión Correcta originando de nuevo el pF, o por Escisión incorrecta, de forma que se forma el plásmido F’ que actuará del mismo modo que el plásmido F siempre y cuanto contenga los genes tra (XIV, XV). El individuo que contiene el pF’ es un Merodiploide pues contiene dos copias de algunos genes del Genóforo. 71 El cruzamiento Hfr x F – es de hecho utilizado para la realización de Mapeos Cromosómicos, por medio de experimentos de Conjugación Interrumpida. Gracias a los pequeños plásmidos formados en cada caso podemos interprestar y deducir la posición de los genes en el Genóforo. Por ejemplo, podemos basarnos en una Cepa Hfr StrpS (sensible a streptomicina) para cruzarla con una Cepa F – StrpR (resistente). Se realizan varias Conjugaciones Interrumpidas con distintos intervalos de tiempo, repitiendo el procedimiento, y se agita violentamentamente para romper las uniones entre células. Finalmente se plaquea en un Medio Mínimo + Streptomicina, de forma que las Hfr no puedan crecer. Utilizando marcadores en un Medio Mínimo para los distintos genes podemos observar por que orden y al cabo de cuanto tiempo pasa cada marcador a la F –. Por ejemplo, para determinar la posición de los genes Thr, Leu y Lac utilizamos cepas Hfr: Thr +, Leu + y Lac +, y cepas F –: Thr -, Leu - y Lac -. Si las cepas F – pueden crecer en Medio Mínimo + Streptomicina significa que son recombinantes, y que el tiempo mínimo de Conjugación en el que estas bacterias comienzan a crecer corresponde con la distancia en minutos entre Ori T y dichos genes (XVI, XVII, XVIII). F+ x F– Hfr x F Hfr x F Hfr x F - F+ (escisión correcta) Hfr (escisión incorrecta) F’ (interrumpido) F- F’ + F - F’ 4 Otros plásmidos, comunicación entre Gram positivas y entre Bacterias y Vegetales: Existen otros plásmidos en Gram - que no son pF. Son plásmidos conjugativos e incluyen por lo tanto regiuón tra. Por ejemplo, en Enterobacterias, pR100 y pR1. Incluyen además genes de Resistencia a antibióticos, y que permiten la utilización de fuentes de carbono alternativas. La comunicación en Gram + es ligeramente distinta y mucho menos frecuente. No se ha descrito Pili Conjugativo. La agregación y contacto Físico entre F +/F - se produce por secreción de feromonas (pequeños péptidos). Se forman directamente los puentes citoplasmáticos. Presente en Bacillus, Streptococcus y Chlostridium. Agrobacterium tumefaciens es una bacteria responsable de la formación de agallas (tumoraciones) en el tallo de muchos vegetales. Esta patología infecciosa es producida por la transferencia del plásmido Ti al núcleo de las células vegetales. pTi es transferido por medio de un puente citoplasmático, y puede integrarse en el genoma de la célula infectada, expresando genes que codifican determinadas hormonas vegetales responsables del crecimiento incontrolado. PROBLEMA CONJUGACIÓN Se parte de una cepa Hfr de Eschericia coli que es sensible a la tetraciclina y prototrofa para Prolina (Pro) e Histidina (His), y se cruza con una cepa receptora resistente a tetraciclina pero auxotrofa para los mismos marcadores. Responde a las siguientes cuestiones: ¿Qué es una cepa Hfr? ¿Qué proceso genético se producirá entre ambas? ¿Qué característica debe cumplir la cepa receptora para que se produzca dicho fenómeno? Define qué es un mutante auxotrofo ¿Cómo seleccionarías recombinantes prototrofos? 72 73 C/ Introducción a la Ingeniería Genética: (Láminas B) 1 Generalidades sobre Ingeniería Genética; Enzimas de Restricción: Muchos procesos de transferencia de información genética entre bacterias son fundamentales para la industria, ya que permiten la modificación de procesos naturales por el humano con fines productivos. Se define como la formación In-Vitro de nuevas combinaciones de material genético por medio de la inserción de un ADN de interés, en un Vehículo Genético o Vector de Clonación, de tal manera que tras su introducción en un organismo huésped este ADN se pueda multiplicar y propagar, y pueda expresar sus genes, cuyos productos nos interesan. Las Enzimas de Restricción de las bacterias son indispensables para llevar estos procesos a buen término. Se trata de Enzimas Endonucleasas creadas exclusivamente por bacterias que reconocen secuencias pudiendo cortarlas de forma específica (I). Estas enzimas forman parte del Sistema RM (restricción-modificación) natural de las bacterias para defenderse ante ADN exógeno no reconocido. Este sistema tiene además la capacidad de modificación por metilación del ADN propio, con el fin de evitar que dichas enzimas corten el ADN propio. Se distinguen, en función de su forma de cortar la cadena de ADN, tres tipos de Enzimas de Restricción: Tipo I: (ATP, Mg2+) Reconoce secuencias asimétricas, produciendo cortes a más de 100 pares de bases de estas. Tipo II: (Mg2+) Reconoce una secuencia Palindrómica (simétrica) realizando un corte exacto en ella. Puede ser un corte Protuberante (desigual) (I) o Romo (liso, recto). En caso de corte protuberante, el extremo cohesivo puede ser 5’ o 3’ (se trata del más corto de la secuencia palindrómica). Tipo III: (ATP, Mg2+) Reconoce secuencias asimétricas, produciendo cortes a unos 30 pares de bases de estas. Entre ellas, las más interesantes para la Ingeniería genética son las de Tipo II. El tipo de corte que realiza (Romo o Protuberante de extremo cohesivo 5’ o 3’), pues debemos lograr el mismo corte exacto en las secuencias de ADN de interés y donde vamos a insertar este. Se suelen utilizar cortes protuberantes En cuanto a su nomenclatura, alude a la bacteria en la que ha sido aislada por primera vez, por ejemplo: Eco R I (Escherichia coli, Cepa R, nº1). Bam H I (Bacillus amilolicuefaciens, Cepa H, nº1). El primer paso es estudiar la secuencia del gen en el que estamos interesados, y detectar todas las Secuencias de Restricción con que cuenta para cada enzima. Este trabajo está informatizado. Así podemos seleccionar la Enzima adecuada para escindir un gen concreto deseado, sin destruirlo, que deberá ser la misma que escinda el ADN del Vector de Clonación, u otra que realice cortes con extremos cohesivos idénticos. Seleccionamos de entre los fragmentos el que nos interesa, aislandolo por cromatografía o electroforesis. El Vector de Clonación es el elemento genético que nos permite introducir nuestro fragmento de ADN de interés en un organismo huésped. Se suelen emplear Plásmidos Artificiales, Baceriófagos, Cósmidos (plásmido + bacteriófago) o Vectores de Alta Capacidad. Finalmente podremos así clonar nuestro ADN en el Vector, que deberá ser cortado con extremos idénticos a los del fragmento de interés. Ambos fragmentos serán recombinados con ayuda de una ADN ligasa. Por último se introduce el Vector clonado en un Organismo Huésped: Bacterias, Levaduras, y en ocasiones células eucariontes vegetales o animales. 74 En bacterias, el plásmido es introducido por transformación, forzando un estado de competencia artificial gracias a elecroporación o utilizando CaCl 2. Debido al Sistema RM (Rec), la mayoría de bacterias modifica o escinde el fragmento de ADN introducido como método de defensa. Estas reciben el nombre de Rec +. Las pocas bacterias que llegan al final del proceso y contienen el plásmido en su interior son Rec -; el Sstema RM debe ser eliminado de las bacterias utilizadas. En células Eucariontes, esta información puede ser introducida por dos vías: Mediante infección por Virus, o con Pistola de Genes. En el caso de levaduras puede también lograrse por electroporación. 2 Vectores de Clonación; Plásmidos Artificiales: Los Plásmidos Naturales presentan una serie de desventajas: Para comenzar pueden presentar región tra y ser movilizables. No nos interesa la transferencia horizontal de genes, sino simplemente la transferencia vertical (a la descendencia). Además no contienen marcadores seleccionables en placa, como Resistencia a Antibióticos. No suelen poder introducir más de 10 kPB. Por ello se generan Plásmidos Artificiales, como pBR 322 (V), o pUC 18. Para comenzar, debemos eliminar los genes tra para evitar la recombinación. La Región Polilinker es una región que se incluye siempre en los plásmidos artificiales, con genes de resistencia (o nutricional), en la que se pueden insertar los fragmentos de interés ya que posee una gran cantidad de secuencias de escisión para múltiples enzimas. Región Polilinker Para comprobar que un plásmido ha sido clonado correctamente, la Inactivación Insercional de la Región Polilinker, por inserción del fragmento de interés, facilita el reconocimiento de plásmidos que han insertado correctamente dicho fragmento, ya que las bacterias portadoras pierden su resistencia (o capacidad nutricional) (VI). Por ejemplo, la Inactivación Insercional de la región polilinker que contiene el gen Lac Z para -Galactosidasa, clonados e insertados en bacterias que son cultivadas en Medio X-Gal, es responsable de la formación de colonias bacterianas de color azul, mientras que las que contienen el gen Lac-Z intacto forman colonias de color blanco, visiblemente distinguibles (VII, VIII). 3 Otros Vectores de Clonación: Los Fagos son de fácil manejo pero sólo aceptan genes de pequeño tamaño (10kPB). Los Fagos - Cósmidos son más difíciles de manejar, pero permiten trabajar con ADN de mayor tamaño (20-30 kPB). 75 El Fago Lambda produce en el interior celular un gran número de copias de su información genética y de las proteínas de su cápsida. Sin embargo el tamaño de su cápsida es limitado y no puede contener mucha información. Para utilizarlo en ingeniería genética deben ser eliminadas partes no indispensables de su ADN para poder introducir el fragmento de interés (IX). Se ha insertado un gen -Gal con Región Polilinker en la cadena de ADN de Fagos Lambda. Deben ser cultivados en una placa sobre césped de bacterias, de forma que se crean halos de lisis (X). El Cósmido es un vector plasmídico con Sitios Cos (extremos cohesivos del genoma del Fago Lambda), de tal manera que dichos plásmidos son empaquetados en los viriones y fagos (IX). Los Vectores de Alta Capacidad son vectores para fragmentos de más de 100 kPB. Entre ellos se encuentran los Cromosomas Bacterianos Artificiales (BAC) basados en el plásmido pF, y los Cromosomas Artificiales de Levaduras (YAC) derivados de un cromosoma de levadura. 4 Técnicas de Rastreo: Se trata de técnicas previstas para diferenciar las bacterias que han incorporado correctamente el fragmento de ADN de interés y no lo han modificado, al final del proceso. Puede realizarse por Resistencia a Antibióticos, utilizando un vector que contiene un gen de Resistencia independiente de la región Polilinker (V, VI; Resistencia a Ampicilina). Sólo las bacterias que han incorporado el Vector podrán crecer en un Medio con dicho Antibiótico. Puede realizarse por Hibridación del plásmido y en ADN bacteriano, inducida por Sondas, combinado con el método de Resistencia a Antibióticos. Puede también realizarse un Rastreo por Ensayo Inmunológico. Se utilizan Anticuerpos Marcados específicos para una proteína. 76 77 Parte II; VIROLOGÍA I; INTRODUCCIÓN A LOS VIRUS: A/ Características Generales (Láminas F): 1 Definición: Los virus fueron descubiertos durante el siglo XIX, 50 años antes del descubrimiento del microscopio electrónico, como agentes infecciosos responsables de enfermedades transmisibles, de muy pequeño tamaño (10-100 nm, Bacterias 1-10m), y capaces de atravesar filtros para bacterias e incapaces de crecer en medios de cultivo artificiales destinados al cultivo rutinario de bacterias. Actualmente se definen como Sistemas Vivos Acelulares que contienen ADN o ARN como información genética rodeada por una cápsida de proteínas y otras macromoléculas, y en algunos casos una membrana plasmática externa, similar a las células que infecta. Además, en su Estadio Extracelular Inerte carecen de metabolismo, y por lo tanto deben secuestrar el metabolismo de las células que infecta para llevar a cabo su replicación. Sus células huésped pueden ser Bacterias (Bacteriófagos), o células Vegetales o Animales. Están formados por una Envuelta y una Nucleocápsida proteica, que contienen en su interior el Core o Genoma Vírico. Este último es determinante sobre cómo y dónde un Virus va a llevar a cabo una infección para replicarse (reproducirse). 2 Estructura General; El Core o Genoma Vírico: El Core puede ser de varios tipos: ADN mc (monocatenario, ss), ADN dc (dicatenario, ds), ARN mc o ARN dc. En general su ciclo replicativo en el interior de la célula implica tanto ADN como ARN. Algunos Virus injectan su Core en el Núcleo Celular, especialmente los virus de ADN. Los virus de ARN pueden injectar su ADN en el Citosol. Entre los Virus Vegetales, los más frecuentes son ARN mc, en Fagos lo más frecuente es sin embargo ADN dc. En algunos casos, especialmente Fagos, encontramos Bases Nitrogenadas modificadas, como por ejemplo Hidroximetilcitosina. Los virus aprovechan el flujjo de información genética propio de la célula para integrar en él su propia información, produciendose así la infección. Utilizan tanto sus mecanismo de Replicación como los de Transcripción y de Traducción. Entre los virus de ARN, su ARN presenta una estructura similar a la de los ARNm de las células Eucariontes: con Cap en su extremo 5’ y Poli-A en el extremo 3’, de tal manera que se produce reconocimiento ribosómico y Traducción. Los virus de ARN mc pueden presentar polaridad negativa ARN mc(-) o positiva ARN mc(+). Estos últimos sirven directamente como ARNm 3 Estructura General; La Cápsida y La Envuelta: La Cápsida es la estructura proteica que envuelve al Core. Las proteínas que la componen están codificadas por el propio Core y son sintetizadas por la maquinaria de la célula infectada, durante la infección. Existe una gran variedad, pero la principal clasificación se realiza en función de su Forma: Icosahédrica: Virus esféricos (VIII, IX, X). Helicoidal: Virus cilíndricos (V, VI, VII). 78 Compleja: Por ejemplo muchos Fagos, que presentan Simetría Binal: Cola Helicoidal e Cabeza Icosahédrica (XV, XVI). Los Virus Cilíndricos son estrechos y alargados (lMAX = 300 nm; D = 15-18 nm). Su cápsida se forma por ensamblaje helicoidal de Protómeros (monómeros proteicos) alrededor del Ácido Nucleico (V). El ensamblaje es regulado por el virus, aunque pueden influir las proteínas Chaperonas propias de la célula. Los Virus Esféricos se forman por una estructura en Icosahedro, elegida por la naturaleza por ser la más efectiva. Presenta 12 Vértices, 30 Aristas y 20 Caras. Cada subunidad proteica es denominada Capsómero, y está formado por varios Protómeros. Los Capsómeros más comunes son los Pentámeros (5 protómeros iguales o distintos) y los Hexámeros (6 protómeros iguales o distintos). De hecho, por lo general, los Pentámeros (P) ocupan los Vértices mientras los Hexámeros (H) ocupan las Aristas (IX). Calculando obtenemos un total de 240 monómeros. Existen Virus Esféricos de distintos tamaños, que se diferencian también por su número de Capsómeros. Por ejemplo, el virus más pequeño, Parvovirus, solo se forma por 12 Capsómeros, mientras que Adenovirus (XI) se forma por unos 252 Capsómeros. Poxvirus presenta un tipo de Cápsida especial, de forma Ovoide. Se trata de una Nucleocápsida en forma de disco bicóncavo, rodeada por una envuelta de doble membrana. Presenta dos cuerpos elípticos laterales, son partes no rellenas por un esqueleto proteico de Capsómeros (XIV). Los virus no siempre presentan Envuelta, y esto permite clasificarlos en dos grupos: Envueltos o Desnudos. La Envuelta de naturaleza de Bicapa Lipídica, que deriva de la membrana plasmática de la célula huésped de la que ha salido. Esta envuelta suele tener insertadas Glicoproteínas que han sido sintetizadas a partir del Core del Virus. Suelen ser amorfas y deformables. Son especialmente frecuentes en virus animales, y nunca se encuentran en Virus Cilíndricos. 4 El Cultivo de Virus: El cultivo de cada tipo de virus es distinto en función del tipo de célula que infectan. Los más desarrollados son los cultivos de Bacteriófagos por la facilidad del cultivo de las células bacterianas y su importancia en ingeniería genética, y el cultivo de virus animales por su importancia para la Medicina. El cultivo de Virus Animales se lleva a cabo por dos principales técnicas: Cultivo en Huevo Embrionario: Es el más barato y sencillo (XX). Cultivo en Cultivos Celulares Primarios, en monocapa o en suspensión (XXI, XXII). Suelen utilizarse células musculares. El Cultivo en Monocapa es similar a un césped bacteriano. Las células cubren la superficie del medio, facilitando la adsorción del Virus. El virus es directamente inoculado sobre el medio, y las alteraciones que provoca sobre las células pueden observarse por microscopía óptica. El procedimiento por Cultivo en Suspensión es similar pero se dificulta la observación de las alteraciones celulares, ya que permite la proliferación excesiva de las células cultivadas. Las principales alteraciones observables son la modificación de la morfología de las células, la pérdida o fusión de los núcleos celulares y la formación se Sincitios (XXIII, XXIV, XXV, XXVI, XXVII). Los sincitios facilitan la dispersión del virus a distintas células. Existen dos principales líneas celulares utilizadas en laboratorio: Hela (Helen L.). Vero (riñon de mono verde americano). El Cultivo de Virus Vegetales suele realizarse por infección de la planta completa, siendo la opción más sencilla y barata, aunque puede también llevarse a cabo en cultivos celulares primarios en 79 monocapa o suspensión. Sin embargo el cultivo vivo permite además la observación de las alteraciones que se producen a nivel de los órganos. El Cultivo de Bacteriófagos se realiza por infección de bacterias vivas en un cultivo en Césped. La infección puede reconocerse a simple vista por la formación de los Halos de Lisis (Calvas) que aparecen cuando los Virus abandonan la Célula. Estas calvas corresponden a células UFC que han sido infectadas. El Fago se multiplica lisándo la célula y expandiéndose concentricamente a otras. El cultivo tiene lugar en tres fases: 1. Adsorción de los virus en la superficie bacteriana. 2. Recubrimiento del cultivo en placa con Agar de Cobertura que facilita la multiplicación de los virus. 3. Incubación de la placa a 37 ºC. B/ La Replicación de los Virus: 1 Ciclo de Replicación Vírica: El virus debe introducir su Core en la célula, e inducirla a sintetizar todos los componentes necesarios para formar muchos viriones, que se transformarán en virus al abandonar de nuevo la célula. De hecho, desde que el virus infecta una célula hasta que comienzan a liberar viriones transcurre un tiempo de latencia o Fase de Eclipse (20-30 min en bacterias, 8-40 horas en eucariontes). Durante ese tiempo los componentes del virus se encuentran por separado y en el interior de la célula infectada. Todo este proceso tiene lugar en varias fases comunes: 1. Adsorción/Fijación del Virus 2. Penetración 3. Expresión/Replicación 4. Ensamblaje/Encapsidación 5. Liberación de los Viriones La Adsorción/Fijación es el reconocimiento y unión del virus sobre la superficie de la célula. Mientras en células vegetales el virus entra por abrasión de la pared celular o a través de roturas, los virus animales y los fagos necesitan de un proceso de reconocimiento de receptores celulares que permite su adsorción. Los virus con envuelta utilizan Glicoproteínas transmembrana, mientras que los virus desnudos se sirven de las proteínas de la cápsida. En concreto, para las células animales, los virus presentan Tropismo, es decir, no pueden infectar cualquier célula y son específicos de determinados tejidos. Pueden dispersarse por medio de formación de Sincitios de una célula a otra adyacente. La Penetración del virus al citosol celular puede producirse de formas distintas. Los Fagos insertan su Ácido Nucleico por Injección directa, muy rara en eucariontes (excepción: Poliovirus). En vegetales el virus completo penetra en las células por abrasiones, pudiendo extenderse a través de los plasmodesmos y los tejidos vasculares. Se desencapsida en el interior de la célula. En animales los virus pueden penetrar por distintos mecanismos en función de si son envueltos o desnudos: La Endocitosis de Virus tanto desnudos como envueltos implica la formación de una concavidad internamente recubierta por Clatrina. El virus desnudo es endocitado y desencapsidado en el interior de la vesícula, de forma que sólo el Core abandona la vesícula y llega al citosol. El virus desnudo suele fusionar su propia envuelta con la membrana de la vesícula, de forma que la cápsida es liberada al citoplasma. Posteriormente se produce la desencapsidación del Core y su liberación. Los Virus Envueltos suelen penetrar por simple fusión de la envuelta con la membrana celular, de forma que la cápsida es liberada al citoplasma. Posteriormente se produce la desencapsidación del Core y su liberación. 80 La Expresión/Replicación es el paso fundamental, objetivo principal de la infección. La Expresión de los genes del Core permite la síntesis de las proteínas necesarias para la Cápsida, y en su caso las otras proteínas y enzimas necesarias para la infección. El virus debe adaptarse tanto a la maquinaria como a la compartimentación de su célula huésped para poder reproducirse. El Virus organiza la expresión de sus genes en orden estricto: 1. Genes Tempranos: Enzimas y Proteínas encargadas del secuestro de la maquinaria de la célula. 2. Genes Tardíos: Proteínas estructurales del Virión. La Replicación constituye la formación de múltiples copias del ácido nucleico que constituye el Genoma Vírico. Es un proceso esencial ya que se trata de una parte constitutiva y esencial de los viriones, y debe ser encapsidada y posteriormente injectada en otra célula. Existe una clasificación de los virus que los diferencia fundamentalmente por su tipo de ácido nucleico y su ciclo de Expresión/Replicación; la clasificación de Baltimore. La fase de Ensamblaje/Encapsidación sucede de formas distintas en virus Icosaédricos y Helicoidales. En Helicoidales los protómeros se unen en primer lugar en forma de discos; interaccionan con las secuencias Pac del ácido nucleico, reconociéndolas y uniéndose a ellas. El ácido Nucleico se enrolla de forma helicoidal y va incorporándose en el hueco central simultáneamente a la Encapsidación. Al final, el ácido nucleico forma un Loop para evitar que se pierda la estructura (XL, XLI). En Icosaédricos, primero los Protómeros se unen formándo los Capsómeros, y a continuación se forma la cápsida, por su ensamblaje, con ayuda de proteínas y enzimas víricas que pueden penetrar en la cápsida pero finalmente son liberadas. La cápsida madura debe ser digerida por proteasas (proteínas de maduración), para transformarse finalemente en cápsida madura (XLII). En Fagos con simetría Binal primero se forma la cápsida icosahédrica madura, y finalmente se forma el cuello y a partir de ahí la cápsida cilíndrica que corresponde a la cola. La Liberación es el último proceso del ciclo, y de él depende que el virus sea envuelto o desnudo (excepto algunos casos como Herpesvirus, cuya envuelta proviene de la membrana interna Nuclear, de manera que entra en el Retículo endoplasmático y es exocitado por vesículas a través del aparato de Golgi (XL)). De hecho los virus desnudos suele ser liberados por lisis de la célula infectada. Es el caso más común en Fagos. Los virus envueltos suele abandonar la célula por Gemación o Exocitosis, de manera que la envuelta procede de la propia membrana de la célula infectada. Este es el caso más común en virus animales. Para ello debe ponerse en marcha todo un proceso de expresión de los genes que codifican las Glicoproteínas de la envuelta. Esta síntesis tiene lugar en el Retículo Endoplasmático, y sus productos pasan al aparato de Golgi , desde donde son transportadas mediante vesículas e insertadas en la membrana celular, en puntos discretos (no se difuminan por la membrana). El virus abandona la célula en los puntos exactos donde estas vesículas han descargado las Glicoproteínas, donde además es importante que no se encuentran las Glicoproteínas de membrana propias de la célula (XXXVII, XXXVIII). En caso de liberación por gemación, la célula infectada termina también muriendo, y puede ser debido a diversos motivos: Por el secuestro de la maquinaria, por la gran cantidad de proteínas virales tóxicas acumuladas, por alteraciones cromosómicas (algunos virus insertan su ADN), por la presencia de proteínas de membrana víricas que hacen que el sistema inmunitario reconozca y ataque a la célula huésped, o por el alto volumen de los viriones contenidos, que hacen estallar a la célula. 2 La Clasificación de Baltimore: Clasificación antigua de los Virus, que los agrupa en siete clases (en números romanos), en función de el tipo de Core que presentan, y por lo tanto de su ciclo de Expresión/Replicación (XXXVI). 81 La Clase I se compone de Virus de ADN bc. Se trata, por ejemplo, de muchos Bacteriófagos y de Herpesvirus. Su Expresión tiene lugar por simple Transcripción a ARNm, mediada por ARN polimerasa (ADN dep), y Traducción a Proteína. Para su Replicación entrar en acción una ADN polimerasa (ADN dep). Este mecanismo es el más sencillo, ya que es idéntico al mecanismo propio de Expresión y Replicación de la célula. Algunos virus de la Clase I injectan su información en el núcleo, pero otras, como Poxvirus, lo llevan a cabo en el citosol, gracias a la codificación de sus propias enzimas ADN polimerasas. La Clase II se compone de los Virus de ADN mc. Es una clase poco frecuente, y se encuentra especialmente entre los Fagos. Esta hebra ADN sirve como molde para ADN polimerasa, de forma que esta sintetiza la hebra complementaria. Una vez en forma de ADN bc los procesos son idénticos a los de la Clase I. Algunos tienen también sus propias ADN polimerasas. La Clase III se compone de los Virus de ARN bc. Entre ellos encontramos Reovirus. Los virus suelen evitar en la medida de lo posible los ARN bc, pues estos suelen ser detectados por el Interferón y la célula puede defenderse. Se utiliza la cadena de polaridad (-), que va a ser leída y replicada por una ARN polimerasa (ARN dep) o Transcriptasa Viral generándose la cadena de polaridad (+) que ejerce como ARNm para la Expresión. Esta misma enzima, que no existe en eucariontes por lo que debe ser parte del Genoma Vírico, facilita desde la cadena (+) la Replicación de la cadena (-). La Clase IV se compone de los virus de ARN mc (+). Por ejemplo, Picornavirus. Esta cadena es utilizada directamente para la Expresión. Para la Replicación debe generarse una copia de la cadena complemetaria (-), a partir de la cual puede procederse a la generación de nuevas cadenas (+). Siempre evitan la formación de ARN bc. Ambos pasos deben ser facilitados por una ARN polimerasa (ARN dep) propia del Core. La Clase V se compone de los virus de RNA mc (-). La integran una gran cantidad de virus patógenos, como por ejemplo los Virus de la Gripe. Esta cadena es complementaria a la cadena ARNm (+), por lo que para su Expresión debe ser replicada también por la enzima Transcriptasa Viral formándose la cadena (+), que sirve tanto de molde para la Expresión, como para la replicación de la cadena (-). La Clase VI se compone de los virus de ARN mc (+), y son conocidos como Retrovirus. Estos son un grupos de virus especiales, entre los que se encuentra el VIH (SIDA), cuyo ciclo de Expresión/Replicación es especialmente complejo y necesita de una enzima especial exclusiva del Genoma Vírico de los Virus de esta Clase. Se trata de las enzimas ADN polimerasas (ARN dep), más conocidas como Retrotranscriptasas, capaces de reconstruir a partir de ARN mc (+) la cadena correspondiente de ADN bc, por un proceso de hibridación ARN/ADN similar a una Transcripción en sentido inverso. Este ADN bc va a ser integrado en el Genoma propio de la célula huésped, de forma que va a ser transcrito a ARNm sirviendo para la replicación ya que es la cadena (+), como para la Expresión. La Clase VII se compone de virus de ADN bc, pero se comportan de manera distinta a los de la Clase I. Entre ellos destacan VHB (Hepatitis B) y Hepadnavirus. Este ADN es transcrito a ARNm, y de ahí traducido a proteínas, llevándose así a cabo la Expresión. La Replicación tiene lugar también a partir de las hebras de ARNm, por medio de enzimas Retrotranscriptasas que forman la primera cadena de ADN. A continuación se crea la cadena complementaria. 82 83 II; Tipos de Virus: A/ Virus Bacteriófagos (Láminas G): 1 Clasificación General de Bacteriófagos: Para la clasificación y recuento de Fagos se puede proceder mediante dos técnicas distintas: Microscopía Electrónica o Multiplicación sobre Placa en células huésped bacterianas cultivadas. En este último caso podemos identificar su presencia a simple vista, gracias a los Halos de Lisis (III, IV). Además el cultivo de este tipo de células es fácil y barato. Si en Bacterias hablamos de UFC, en virus, un Halo o Placa de Lisis proviene también de un solo virus viable: UFP (Unidad Formadora de Placa). Suelen estar clasificados en tres grupos: ADN bc, ADN mc, y ARN (mc o dc). Estos últimos son los menos numerosos (II). Los Fagos de ADN bc son los más frecuentes. Presentan simetría Binal (VI). Presentan una Cabeza Icosaédrica, que en su base tiene un ensanchamiento, se trata de un Collar que une la Cabeza al Cuello, el cual a su vez une esto con una Cola Helicoidal rodeada por una Vaina. La Cola presenta también un ensanchamiento en su extremo: se trata de la Placa Basal, de forma hexagonal, que sirve de anclaje para las Fibras de la Cola. Entre ellos encontramos: Serie T par. Serie T impar. Fago Mu. Fago Lambda. Los fagos de ADN mc pueden ser Helicoidales o Icosaédricos. Los Fagos de ARN, ya sean mc o bc, son por lo general Icosaédricos. 2 Los Fagos de la Serie T: Se trata de un grupo de Fagos Líticos, carentes de Ciclo Lisogénico. Por ejemplo, T4 es un virus de la Serie T par de alta complejidad, formado por un Core de 178 kPB que codifica para unas 250 proteínas distintas, 50 % de las cuales son estructurales. La Adsorción tiene lugar por medio de Receptores presentes en las fibras de la Cola, que reconocen proteínas de la Pared Celular Bacteriana. Al producirse reconocimiento, la Placa basal se ancla a la pared celular (VII). La Penetración precisa de un cambio conformacional en la Placa Basal y las Fibras de la cola. Esto permite la salida del ADN, que es forzada por una contracción de la Vaina de la cola, de tal manera que el Ácido Nucleico es Injectado al interior de la célula. Existen acúmulos de ATP en la Cola que permiten dicha contracción. En muchos fagos existen además enzimas Lisozimas que facilitan la formación de un poro a través de la Pared Celular, que facilita a su vez la injección (VII). La Expresión de los genes está organizada en fases temprana y tardía. La Síntesis de Tránscritos Tempranos comienza aproximadamente 2 minutos después de la injección. Se forman proteínas y enzimas importantes para la infección. Por un lado se forman Nucleasas que destruyen el ADN celular, secuestrando su maquinaria y utilizando los productos como precursores para la síntesis de sus propios ácidos nucleicos. Por otro lado se encuentran enzimas que modifican las RNA polimerasas bacterianas con ayuda de factores sintetizados por los fagos de manera que estas reconozcan los Promotores del ADN Vírico. La Síntesis de Tránscritos Tardíos suelen comenzar pasados unos 5-9 minutos desde la Injección. Son proteínas implicadas en la formación de los Viriones: Protómeros de cada región del Fago, Copias del ADN, y Enzimas que facilitan su correcto ensamblaje. De tal manera que pasados unos 22 minutos de la Injección, la célula bacteriana es susceptible de lisar y liberar multitud de Viriones (X, XI). En algunos casos los Fagos abandonan la baceria por gemación. 84 La Replicación es un proceso no menos complejo. Estos Fagos T pares (T2, T4 y T6) presentan Ácidos Nucleicos con Permutación Circular y Redundancia Terminal. La redundancia terminal consiste en la repetición de una secuencia de 2-6 kPB en los dos extremos. Por ejemplo, las secuencias AB se repiten en los extremos del virus: ABCDEFGHAB. Además, diferentes virus T4 muestran diferentes ordenaciones de sus genes (diferentes permutaciones circulares), un virus tiene la secuencia ABCDEFGHAB y otro tiene la secuencia CDEFGHABCD, pudiéndose encontrar otro que tenga la secuencia EFGHABCDEF. Por esta causa, los mapas genéticos en T4 resultan ser circulares, a pesar de tener como material hereditario ADN doble hélice lineal. Durante la Replicación, varias cadenas de ADN bc se unen así formando Concaterios, estas estructuras superiores deben ser fragmentadas en partes iguales, cada un de las cuales está destinada a un Virión. El Ensamblaje es un proceso tan complejo como la estructura de los propios virus. Primero tiene lugar la encapsidación del ADN en las Cápsidas Icosaédricas, a las que posteriormente se une el Collar. Por otro lado la polimerización de la Cola, que incluye Placa Basal, Cola, Vaina y Cuello. La síntesis de las Fibras e la Cola tiene lugar también independientemente, y estas se unen al Complejo en último lugar (XIV). Las Enzimas de Restricción propias de Sistema RM deben ser evitadas por los Fagos de DNA bc. Los Fagos han desarrollado para ello diversas estrategias. El sistema más común en la Serie T par es la incorporación de Bases Nitrogenadas en su ADN que no son reconocidas por estas enzimas, entre las que destaca Hidroximetilcitosina (XII). Para ello producen enzimas especiales capaces de Metilar, Glicosilar y Modificar sus propias Bases. Pueden también producir proteínas capaces de inhibir a dichas enzimas bacterianas. Los Fagos de la Serie T impar no presentan Vaina y son incapaces de contraer la cola. A pesar de ello son muy similares a los de la Serie T par. 3 El Fago Lambda: Se trata de otro Fago de Simetría Binal, caracterizado por no poseer Vaina de la Cola, y además por presentar una única Fibra de la cola (XV). Sus Ácidos Nucleicos presentan extremos Cohesivos y Permutación Circular. Reciben el nombre de Extremos Cos (cohesivos). La circularización solo tiene lugar en el interior de la célula infectada, en ciclo Lítico (XVII). Por el conjunto de sus propiedades es muy utilizado en Ingeniería Genética. Se trata de un Fago Atemperado, que comparte un ciclo Lítico y un ciclo Lisogénico en el que se integra en el Genóforo bacteriano y replica a la par que la célula, en forma de Profago, pudiendo ser activado el ciclo Lítico, proceso que ocurre bajo intensa regulación. Para establecer el ciclo Lisogénico el metabolismo de la célula infectada no debe ser alterado. 85 El control de los ciclos Lítico/Lisogénico se debe a la competencia entre las proteínas cI y Cro, y su síntesis está fuertemente controlada. A mayor concentración de la primera se lleva a cabo el ciclo Lisogénico, y viceversa. cI proporciona las proteínas necesarias para la integración del Genoma Vírico en regiones concretas y reconocidas previamente del Genoma Bacteriano (XX). La activación del ciclo Lítico se produce gracias a determinados factores que dañan el ADN bacteriano. La proteína Rec A reconoce y rompe la proteína cI mientras la concentración de Cro va en aumento. Entonces se activa Ecisionasa, una enzima capaz de escindir y liberar el ADN del Profago, formándose ADN con Permutación Circular (XIX). El genoma de este Fago puede ser leído en ambos sentidos, de forma que a cada uno le corresponden Promotores y proteínas distintas. Se trata de una estrategia de compactación de la información en el menor espacio de muy compleja regulación (XIX). De tal manera, además de distinguirse los transcritos tempranos (1) y tardíos (2), se hace la diferencia entre los Transcritos Izquierdos L (left) a partir de los PL (promotor izquierdo) en sentido antihorario, y los Transcritos Derechos R (right) a partir de los PR. Encontramos por lo tanto (XVIII): L1: Transcritos Izquierdos Tempranos. Entre ellos cI. R1: Transcritos Derechos Tempranos. L2: Transcritos Izquierdos Tardíos. Entre ellos Cro. R2: Transcritos Derechos Tardíos. La Replicación del ADN Circularizado tiene lugar según el modelo de Círculo Rodante gracias a la maquinaria que utiliza la Bacteria para la replicación de sus propios plásmidos pequeños circulares. Su Expresión puede llevarse a cabo también por la vía propia de la Célula Bacteriana. La Replicación según el modelo del Círculo Rodante se inicia con la liberación del extremo Cebador 3’ OH de la hebra externa, de forma que la interna permanece circularizada, y sirve como molde. Puede acoplarse la síntesis lineal y continua de muchas copias y la simultánea síntesis de sus hebras complementarias. El resultado es una larguísima hebra de ADN bc que contiene mútiples copias del Core. Esta cadena deberá ser fragmentada e incluida en un virión. 4 El Fago Mu: Este Fago Atemperado es muy similar a los de la Serie T par, y se caracteriza por su mecanismo biológico de transposición. Su Genoma posee en cada extremo una región de ADN bacteriano (host DNA): Izquierda: Región host DNA pequeño y constante. Derecha: Región host DNA variable. Este fragmento transporta un fragmento de la información de la última bacteria que ha infectado, y permite que el Core sea insertado en una secuencia diana concreta del Genóforo de la bacteria, sirviendo esto como Mecanismo de Replicación de su DNA bc. El DNA del fago actúa como un Transposón que se duplica y salta a distintas regiones del Genoma de la Bacteria. Durante el ciclo Lítico los distintos fragmentos, unidos a partes distintas del genoma de la bacteria, son cortados y encapsidados (XXIV). 5 Fagos con ADN mc: Su ciclo Replicativo es similar al de Fagos con ADN bc, con la diferencia que en un primer paso el ADN mc debe ser replicado para formarse ADN bc (XXVII, XXVIII). El Fago X174 presenta una cápsida Icosaédrica. Su Genoma presenta Permutación Circular, y Replicación según el Modelo del Círculo Rodante (XXIX). Presenta el Fenómeno de los Genes Solapantes. Su genoma es de muy pequeño tamaño pero contiene una gran cantidad de información comprimida en una misma hebra en un mismo sentido. Los distintos genes se encuentran codificados bajo distintas Pautas de Lectura. Cuando los codones no se corresponden, aunque se lea la misma 86 hebra, el resultado es completamente distinto, pudiendo solapar los genes en una misma hebra (XXVI). GAG TGA TGT AAT GTC T Stop GA GTG ATG TAA TGT CT Stop G AGT GAT GTA ATG TCT Start Ejemplo: Misma cadena leída con tres Pautas de Lectura distintas. El Fago M13 presenta una cápsida Helicoidal. Sólo es capaz de penetrar a las células bacterianas a través de Pilis Conjugatorios (XXX). No produce lisis, y la liberación de los Viriones tiene lugar por Gemación. Determinadas Proteínas Glicosiladas deben intercalarse en la membrana para permitir la exocitosis del virus. En la membrana citoplasmática, la proteína A recubre el virus. A pesar de ello, el virus no es envuelto por membrana lipídica (XXXI). 6 Fagos con ARN: El Fago MS2 es el más conocido y estudiado dentro de este grupo. Se compone de ADN mc (+) de pequeño tamaño. Su ciclo de Replicación es el común para la Clase IV de Baltimore. (XXXIII, XXXIV). 87 B/ Virus Animales (Láminas H): 1 Generalidades sobre la Infección de virus a células animales: Nos centraremos en los virus que infectan al humano, ya que son con diferencia los mejor estudiados, aunque se sabe que muchos de ellos crean las mismas patologías o patologías similares en otros animales. Son los virus más difíciles de estudiar, y sin embargo son los mejor conocidos por su especial importancia en Medicina. Como vimos anteriormente, los virus animales sólo pueden infectar determinadas células del organismo, por lo que la infección está restringida a uno o pocos órganos. De tal manera distinguimos varios tipos de posibles células diana en un organismo: Células Permisivas: La infección y el ciclo lítico pueden llevarse a cabo. Células Semipermisivas: Sólo permiten el desarrollo de determinadas fases de los ciclos virales, pero no permiten la expresión de los Genes Tardíos. Por ejemplo, Papilomavirus es el virus animal responsable de la aparición de Verrugas. Las células superiores del Epitelio Pluriestratificado son Permisivas. Las células mitóticas de este epitelio, en la capa más profunda, son semiopermisivas. El resto de células son inaccesibles a este virus. Del mismo modo, distinguimos varios tipos de infecciones: Abortiva: La célula controla y evita la infección. El virus fracasa. Lítica: El virus logra replicarse y terminar el ciclo Lítico en la célula hospedadora. Los virus no suelen lisar todas la células que infectan; se trata de una estrategia de supervivencia ya que dichas células son más que su medio de vida, su propia maquinaria metabólica vital. Persistente: Las células son fábricas de Viriones que son liberados por gemación. Es el caso más frecuente. El ciclo replicativo sucede lenta y continuamente, y termina matando a la célula. Latente: El virus detiene su multiplicación y permanece inactivo en el interior celular. Puede permanecer integrado en un cromosoma o en forma de Plásmido o Episoma. La multiplicación activa se inicia después de un tiempo de latencia. Por ejemplo Herpesvirus es un virus animal que puede permanecer latente varios años. No mata a sus células hospedadoras. Transformación Maligna Tumoral: Estos virus reciben el nombre de Virus Oncogénicos (no todos los tumores malignos están causados por virus). Inducen tumores por mecanismos no muy bien conocidos. El Genoma Vírico de este tipo de virus incluye Oncogenes que intervienen en el control del Ciclo Celular produciendo crecimiento celular incontrolado. Presentan en concreto Promotores que se insertan en determinados genes aumentando su expresión. Los Prooncogenes son genes propios de la célula, muy similares a los oncogenes y de los que estos podrían haber derivado. El primer Virus Oncogénico descubierto es el Virus del Sarcoma de Rouss. Actualmente se sabe que son Ontogénicos: VHB (hepatitis B), VPH (papiloma humano), VEB (Epstein-Barr, Mononucleosis), VLTH (Retrovirus Linfotrófico humano, Leucemia). Los virus susceptibles de causar tumores no lo hacen ni necesariamente, ni inmediatamente después de la infección. 2 Efectos Citopáticos de la infección (ECP): Los ECP son las alteraciones que producen los virus a nivel celular y tisular. Un mismo virus puede causar distintos ECP, en células distintas. Son muy útiles para la realización de diagnósticos. Suelen inhibir la síntesis de proteínas y replicación del ADN del hospedador. A menudo alteran la membrana celular formándose Agregados Celulares, o Sincitios si hay fusión de las células, como por ejemplo VRS (respiratorio sincitial). Pueden producirse acúmulos de proteínas virales estructurales, muchas de las cuales son además tóxicas para la célula animal. Conducen a la formación de Cuerpos de Inclusión en Núcleo celular o en el Citosol. Por ejemplo CMV (III) (citomegalovirus) es responsable de la formación de Inclusiones en Ojo de Buey, y del consecuente aumento del volumen celular. El Virus de la Rabia 88 forma grandes cuerpos de inclusión en el Cerebro, que son responsables de los síntomas psiquiátricos y la posible muerte del individuo. Pueden verse a simple vista en Biopsia. Pueden producir Alteraciones Cromosómicas por la integración de Provirus. Por ejemplo el VIH (inmunodeficiencia humana). Pueden producir Tumoraciones Malignas por alteraciones del ciclo celular. Por ejemplo por interacción con la proteína p53. Además de la lisis los virus presentan múltiples mecanismos para alterar o destruir las células infectadas. Sin embargo, en la mayoría de los casos las células infectadas presentan en su membrana los antígenos propios del virus, y son reconocidas y destruidas por el Sistema Inmunitario del organismo hospedador. 3 Evasión del Sistema Inmune: El cuerpo de los animales dispone de un complejo sistema de defensa ante infecciones de toda clase (V). Los virus han desarrollado una serie de sistemas de evasión (VI). La Deriva Antigénica pretende evadir el sistema de reconocimiento y unión de anticuerpos, basándose en engañar a los sistemas de memoria inmunitaria. La variación de los antígenos en las nuevas cepas que aparecen, o incluso durante el propio transcurso de la infección de un mismo organismo evita los mecanismos de Inmunidad Adquirida. Rinovirus, el Virus RNA de la gripe, presenta cada invierno hasta 100 antígenos distintos. Su ARN polimerasa (ARN dep) propia no tiene capacidad de reparación, y esto explica la alta tasa de error de ru Replicación (entorno a 1 nucleótido de cada 105). El Establecimiento de Persistencia, propio de toda infección latente en células de baja capacidad de respuesta inmune evita la activación de dicho sistema. Algunos virus quedan latentes durante toda la vida del organismo, y no se reactivan nunca. Por ejemplo, este sistema es utilizado por Herpesvirus. La Inmunotolerancia es un factor determinante frente a una infección. Muchos virus infectan solamente a organismos con sistemas inmunes inmaduros, pero no infecta o no crea síntomas graves en personas adultas. Puede llegar a ser letal en Neonatos. Por ejemplo la Rubéola, que en adultos sólo crea algunos sarpullidos. La Inmunosupresión puede ser causada por el ataque directo de los virus a las propias células del sistema inmunitario, estos se replican en su interior y posteriormente la destruyen. La Inmunodepresión por este u otros motivos facilita la infección virus. Por ejemplo el VIH provoca un importante descenso en la presencia de Linfocitos T, debilitando la barrera inmunitaria del organismo infectado, facilitando la penetración de nuevos virus distintos en otras células. El Bloqueo de la presentación del Antígeno es un mecanismo utilizado por muchos virus que ocultan sus antígenos de manera que evitan ser reconocidos por Anticuerpos. 4 Virus Animales de ADN: Existen Virus Animales de las siete Clases de Baltimore. Veremos a continuación algunos ejemplos descritos en la tabla (VII) (VIII). Sus respectivos ciclos de Replicación se corresponden con la Clase de Baltimore a la que pertenecen. Herpesvirus Humano (VHH): Grupo de Virus de DNA bc con Permutación Circular. Envueltos por una membrana deformable que adquieren en el Núcleo de la última célula infectada, y con Cápsida Icosaédrica. Su ciclo de Replicación tiene algunas peculiaridades (XIII). Tiene lugar en el Núcleo como en la mayoría de Virus DNA, pero su encapsidación y liberación tienen también lugar en el Núcleo, de manera que el virus adquiere su envuelta en la Membrana Nuclear Interna pasando así al Retículo Endoplásmico, de ahí al aparato de Golgi, siendo exocitado en una vesícula. El Herpesvirus Labial fue antiguamente nombrado Herpes Simplex (VHS). El Herpesvirus Genital antiguamente recibía el nombre de Varicela-Zoster (VVZ), pues su manifestación inmediata es la Varicela, pero puede permanecer latente en las neuronas durante toda la vida del hospedador y 89 activarse causando los Herpes Zoster genitales. Otros Herpesvirus son los antiguamente conocidos como Epstein-Barr (VEB), causante de la Mononucleosis o Enfermedad del beso, Citomegalovirus (CMV), que afecta sólo a inmunodeprimidos pudiendo causar aborto espontáneo, o Sarcoma de Kaposi (VSK). Poxvirus: Virus de DNA bc de muy gran tamaño llegando incluso a ser visibles en microscopía electrónica, y de estructura peculiar. Presentan una Cápsida Bicóncava (XX) que encierra el Core, y esta se presenta Envuelta. En el espacio que queda entre la Cápsida y la Envuelta se encuentran dos Cuerpos Laterales cuya función es tal vez la contención de algunas proteínas necesarias para los primeros pasos de la infección. Este virus nunca penetra en el núcleo y completa su ciclo de Replicación en el Citosol, por lo que debe contener sus propias enzimas ARN polimerasas (ADN dep) (XXIII). Entre ellos encontramos el Virus de la Viruela, que en humanos fue erradicada por vacunación poco antes de 1980, pero que pueden ser transmitidos de animales a humanos. También se encuentra el Virus del Molusco Contagioso, similar al herpes, que se contagia por contacto o por transmisión sexual. Adenovirus: Virus de DNA bc, Cápsida Icosaédrica y Desnudos. Causan Enfermedades Respiratorias y Gastroenteritis. Son responsables de la Fiebre Faringoconjuntival frecuente por contagio en piscinas públicas. Su DNA está provisto de proteínas unidas a 5’. Se replica la cadena (-) de tal manera que se forman cadenas de DNA mc que pueden circularizarse gracias a sus proteínas de 5’, sirviendo como molde para la hebra complementaria, y evitando la formación de Fragmentos de Okazaki. Este método de replicación es idéntico para Poliomavirus y Papilomavirus, por las fuertes relaciones de parentesco entre las tres. Se trata de un grupo de virus con Core de pequeño tamaño, que presentan una fibra de reconocimiento en cada vértice (12 en total) (Láminas F - XI). Poliomavirus: Virus de DNA bc, Cápsida Icosaédrica y Desnudos, similares a Adenovirus. SV40 es un Poliomavirus Oncogénico en muchos simios. Produce infecciones latentes en células semipermisivas durante largos periodos de tiempo, pudiendo integrar su Genoma en los cromosomas o dejándolo libre en el núcleo. Producen proteínas que alteran el ciclo celular formando así Tumoraciones Malignas que pueden resultar beneficiosas para su propia replicación (XXIV, XV). Papilomavirus: Virus de DNA bc, Cápsida Icosaédrica y Desnudos, similares a Adenovirus. Se distinguen dos tipos por los síntomas que causan: Mucosos y Cutáneos. Los Cutáneos son responsables de la formación de Verrugas en el epitelio estratificado de la piel, por alteraciones de las células Queratinizadas permisivas. Las células basales son semipermisivas pero no suelen producirse tumoraciones malignas. Los Mucosos son responsables de enfermedades de transmisión sexual muy frecuentes y además son Oncogénicos, como el Cáncer de Cuello de Útero o las Verrugas Genitales. La vacuna está aún en desarrollo. Parvovirus: Virus de DNA mc, de Cápsida Icosaédrica de tamaño muy reducido (los más pequeños conocidos) y Desnudos. Su ciclo de replicación se corresponde con la Clase II de Baltimore. Algunos, como B19, causan erupciones cutáneas en niños (XXIX). 5 Virus de ARN: Los Virus de ARN presentan varias características comunes. Presentan gran variabilidad antigénica. Suelen presentar un ARN policistrónico (Un mismo ARNm codifica varios genes y proteínas). Este ARN es traducido contínuamente, formando Poliproteínas, las cuales van a ser escindidas a continuación por acción de Proteasas. Además, suelen presentar dentro de una misma Cápsula varias moléculas de ARN: Core Segmentado. Estas moléculas se unen y replican de forma contínua en la célula huésped. Su Replicación tiene lugar por lo general en el Citoplasma (excepción: Ortomixovirus), al igual que su ensamblaje. Los virus ARN mc (-) son siempre Desnudos. En ocasiones excepcionales, dos virus distintos pueden mezclar su información dando lugar a nuevos Viriones híbridos, durante una Coinfección, si los virus son similares. Esto representa un riego importante para Humanos y Animales. 90 Reovirus: Virus de ARN bc de la Clase III de Baltimore. Son virus Desnudos de Cápsida Icosaédrica. Entre ellos destacan los Rotavirus, responsables de Gastroenteritis, de gran influencia a nivel mundial (XXXIV, XXXV). Presentan una doble Cápsida que les otorga una protección extra contra el bajo pH generado por los jugos gástricos. Al alcanzar el intestino alteran el balance de electrolitos creando los síntomas de la Gastroenteritis: pérdidas de agua y desecación. Su penetración es directa y su descapsidación sucede en el citosol (XXXV, XXXVI). Picornavirus: Virus de ARN mc (+) de la Clase IV de Baltimore. Son virus Desnudos de Cápsida Icosaédrica. Entre ellos se encuentran los Rinovirus (resfriados), Hepatovirus (hepatitis A), y Enterovirus (enfermedades respiratorias; Poliovirus responsable de la Poliomielitis, Echovirus, Coxsackievirus) (XXXVII, XXXVIII). Forman ARNm policistrónicos y Poliproteínas que deben ser escindidas (XXXIX, XL). Calcivirus: Virus de ARN mc (+) de la Clase IV de Baltimore. Son virus Desnudos de Cápsida Icosaédrica, causantes de la Hepatitis E. Dentro de este grupo encontramos los Norovirus (Norwalk) causantes de Gastroenteritis en Adultos, que desaparece a los 2 o 3 días. Coronavirus: Virus de ARN mc (+) de la Clase IV de Baltimore. Son virus Desnudos de Cápsida Icosaédrica. Causantes del Síndrome Respiratorio Agudo y Severo (SRAS) muy común en China, en humanos y otros mamíferos (XLVIII, XLIX, L). Togavirus y Flavivirus: Virus de ARN mc (+) de la Clase IV de Baltimore. Son virus Desnudos de Cápsida Icosaédrica. Se agrupan dentro de los Arbovirus, transmitidos por la picadura de Artrópodos como Mosquitos o Garrapatas (XLII, XLIII, XLIV). Los Hospedadores intermediarios y definitivos naturales son diversos Vertebrados, y en ocasiones el humano puede ser hospedador accidental por picadura. Rhabdovirus: Virus de ARN mc (-) de la Clase V de Baltimore. Son virus siempre Envueltos, de Cápsida Helicoidal, en forma de bala (XLVI). Son los causantes de la Rabia transmitida por mordedura de animales. Los Cuerpos de Negri son Inclusiones formadas por estos virus, responsables directas de los efectos psiquiátricos de la Rabia, que pueden tardar en ser graves un año desde la infección. La infección inicial tiene lugar en el Músculo liso cerca de la mordedura, y puede extenderse y proliferar especialmente en localizaciones concretas en el encéfalo, como Córnea, Retina, Glándulas Salivares y Sistema Nervioso Central donde causa los síntomas propios de la Rabia. En las fases finales de la infección el individuo presenta estado comatoso que puede llevarle a la muerte. Los cuerpos de Negri aparecen en la Autopsia como agregados de michocápsidas formadas en el citosol de las células (LXII, LXIII, LXIV, LXV). Paramixovirus y Ortomixovirus: Virus de ARN mc (-) de la Clase V de Baltimore. Se agrupan dentro de los Mixovirus, caracterizados por presentar una Cápsida Helicoidal Envuelta enrollada en una membrana amorfa, más o menos esférica (XLVI). Los Paramixovirus incluyen a los virus del Sarampión, la Parotiditis, Respiratorio Sincitial (VRS) causante de Bronquiditis en niños, o Parainfuenza (VPI) (LXVI. Los Ortomixovirus se caracterizan por presentar un ARN segmentado en el interios de la Cápsida, y por su Envuelta especialmente amorfa. Incluye el Virus Influenza (VI) responsable de la Gripe y de Core segmentado en ocho fragmentos. Su Cápsida se constituye por dos tipos de proteínas diferentes (Hemaglutinina, Neuraminidasa). Constituyen una excepción dentro de los virus de ARN ya que su Replicación tiene lugar en el interior del Núcleo (LXIX, LXX, LXXI). Bunyavirus: Virus de ARN mc (-) de la Clase V de Baltimore. Presentan una Cápsida Helicoidal Envuelta enrollada en una membrana amorfa, más o menos esférica, como los anteriores, pero son de menor tamaño. Presentan un ARN segmentado en tres fragmenos; pequeño (S), mediano (M) y grande (L). Se conocen más de 200 especies distintas, y generalmente son transmitidos por picaduras de Artrópodos. Hantavirus es el más estudiado y peligroso, ya que produce un 50 % de mortalidad, con especial influencia en Estados Unidos. Es responsable de Síndrome Pulmonar, y transmisible por contacto con roedores (LXXII, LXXIV, LXXV, LXXVI). 91 Arenavirus: Virus de ARN mc (-) de la Clase V de Baltimore, Envuelto. Presenta en Microscopía Electrónica aspecto arenoso (XLVI, LXXII). Presenta su ARN en dos fragmentos; pequeño (S) y grande (L). Afectan al sistema nervioso causando fiebre hemorrágica. 6 Retrovirus y Hepadnavirus: Retrovirus: Componen la Clase VI de Baltimore, con ARN mc, cuyo ciclo de Replicación se caracteriza por la actividad de la enzima Retrotranscriptasa capaz de transformar el ARN en ADN. VIH es el principal representante, pero existen otros Retrovirus, como por ejemplo el Virus del Sarcoma de Rose, o el Virus del Linfotrópico Humano (VLH). VIH es un virus con Cápsida Icosaédrica, Envuelto. La envuelta presenta proteínas imprescindible para el reconocimiento de las Células. En el interior de la Cápsida se encuentran dos fragmentos de RNA mc (+) y la enzima Retrotranscriptasa (LI). Durante la fase de Adsorción/Fijación, se produce el reconocimiento de las proteínas de la Envuelta gp120 y los Receptores CD4 de la superficie de los Linfocitos T. Este reconocimiento precisa además de la presencia del Correceptor CCR5 (LVIII). La descapsidación tiene lugar sobre la membrana nuclear, de manera que el RNA mc (+) entra al núcleo junto con las enzimas que van a retrotranscribir esta cadena a una de ADN bc, que finalmente quedará integrada en el Genoma del ADN del propio hospedador gracias a otra enzima; Integrasa. Podrá permanecer así en forma de Provirus durante varios años. Su transcripsción y Traducción se produce por medio de ARN policistrónico y Poliproteínas. El ensamblaje tiene lugar en el Citosol, y el Virión es liberado por Exocitosis (LII, LIII, LIV, LV). VIH-1 es el principal que afecta a huimanos, y proviene del Chimpancé del Sur de África. VIH-2 es un Retrovirus de Mangabey tinado, un simio de África Centro-occidental, muy estudiado. Durante los primeros pasos de la infección se producen los síntomas de la Primoinfección por la desetabilización del sistema inmune, pero puede cursar como una gripe normal. Sin embargo el virus queda latente dentro de los Linfocitos, de tal manera que pasados unos años comienza la fase SIDA durante la cual el sistema inmune queda fuertemente debilitado por la carencia de LT4. Esto es Letal ya que permite la coinfección (infección secundaria) de muchos otros parásitos ante los que no se opone resistencia Química, que llevan al individuo a la muerte. Hepadnavirus: Es el único representante de la Clase VII de Baltimore. Contiene una molécula de DNA bc parcialmente mc, y Retrotranscriptasa. Es responsable de la Hepatitis B (VHB). Engaña al sistema inmune pues codifica la región de su Genoma en cantidades superiores a las necesarias. Estas partículas víricas vacías o Pseudoviriones actúan como señuelo dando trabajo a las células del sistema inmune para entretenerlo. Contiene genes solapados. Además introduce en el núcleo de las células tres fragmentos de distinto tamaño; el mayor de ellos se llama Pregenoma, y genera los ARNm que pasan al Citosol. 92 93 C/ Virus Vegetales (Láminas I): 1 Generalidades sobre la Infección de virus a células vegetales: Estos son bastante similares a los Virus Animales, aunque son menos estudiados y conocidos. Sobretodo han sido estudiados por los daños que pueden causar en Agricultura, especialmente en Cítricos ya que son de gran importancia en nuestra región y a la vez muy susceptibles de contraer infecciones. Su cápsula es mayoritariamente Helicoidal, aunque puede ser Icosaédrica. Suelen presentar un Core de ARN, ya sea mc o bc (especialmente ARN mc (+)). Sólo se encuentran dos tipos de virus Vegetales con ADN; Caulimovirus y Geminivirus. Además suelen presentar un ARN fragmentado, lo cual sólo ocurre en virus animales de ARN mc (-) (I). A diferencia de los virus animales, los virus vegetales no necesitan Receptores en las membranas de la células infectadas, ya que su penetración se produce a través de heridas o abrasiones causadas por el viento o la fauna, especialmente las picaduras de animales, es decir, donde se producen roturas en la Pared Celular Vegetal, barrera muy difícil de franquear. Carecen por tanto de fase de Adsorción/Fijación. En ocasiones poseen desarrollados métodos de infección, como la capacidad de proliferar en las glándulas salivares de insectos chupadores de savia, como pulgones o chinches. La expansión del virus a las demás células del organismo está facilitado por los propios sistemas de comunicación celular y de vascularización de la materia. Una vez infectada la primera célula, los virus pueden dispersarse a otras a través del Floema rápidamente a todo el organismo o a través de los Plasmodesmos, de célula en célula. El efecto de la infección sobre la célula es letal, debido a Lesiones Cloróticas (la célula muere por degradación de sus cloroplastos). Al extenderse a toda la planta suelen causar Manifestaciones Sistémicas, como por ejemplo el enanismo, el marchitamiento moteado de distintos órganos, o el mosaicismo (distintos colores en las hojas). De hecho los virus vegetales suelen ser conocidos por sus manifestaciones patólógicas en los órganos vegetales. Tobamovirus; VMT (TMV): También conocido como Virus del Mosaico del Tabaco, es un virus de Simetría Helicoidal, Desnudo, y con ARN mc (+), Clase IV de Baltimore. Tiene un extremo Cap 5’, por lo que puede servir directamente como ARNm. Presenta cuatro genes fundamentales: MTH (Metiltransferasa y ARN helicasa, sintetiza el extremo Cap), RNP (Ribonucleoproteína viral, ARN polimerasa (ARN dep)), MP (Proteína de Movimiento que favorece la transmisión por Plasmodesmos), y CP (Proteína de Cápsida). La Acumulación de CP es responsable del aspecto de Mosaico de las hojas infectadas: se forman inclusiones citoplasmáticas (III, V). Virus del Mosaico de la Coliflor; VMC: Se trata de un Caulimovirus de ADN, similar al virus de la Hepatitis B, de la Clase VII de Baltimore. De hecho, al igual que Hepadnavirus introduce su ADN en el Núcleo, pero posee una Retrotranscriptasa que Replica en el citosol el ADN a partir del ARNm. 94 95 D/ Otros Agentes Infecciosos Acelulares (Láminas J): 1 Viroides y Virusoides: Los Viroides son Moléculas desnudas de ARN mc (+) circularizado, causantes de manifestaciones sitemáticas en vegetales, que se pensaba que eran causadas por virus, como el Tubérculo Filiforme de la Patata y el Enanismo del Crisantemo. La transmisión se produce de una a otra planta, directamente, o por ejemplo a través del Polen. Varias regiones del ARN mc pueden aparear dando un aspecto parcialmente bicatenario. Penetran en el Nucleolo de la célula vegetal, donde pueden llegar a acumularse hasta unas 2000 copias. Necesitan la proteína viral ARN polimerasa (ARN dep), pero no la codifican por lo que se piesa que debe presentarse una coinfección con virus. Se piensa que estas moléculas o los Transposones pueden ser el origen de los ancestros ARN de la vida (I, II, III). Los Virusoides son aún menos conocidos que los Viroides. Se definen como Virus Satélite vegetales. Se trata de moléculas de ARN circular de muy bajo PM y alto contenido en C-G. Se encuentra incluída en una Cápsida muy sencilla y reducida, constituída por un único tipo de Proteína. No presentan capacidad de Autorreplicación, y al igual que los anteriores se piensa que precisan de una coinfección por virus. Algunos se han encontrado de hecho dentro de los propios Virus Vegetales, como el Virus Aterciopelado del Tabaco. 2 Priones: Los Priones son Partículas Proteicas sin Ácido Nucleico que pueden infectar células animales, y que puede transmitirse de unos a otros. Son específicas para cada hospedador, y de hecho cada uno produce sus propias Proteínas Priónicas Naturales o Celulares (PrP C, predominancia de -hélice), ligeramente distintas entre las especies de Mamíferos, y propia del Sistema Nervioso. Por simple contacto con una Proteína Priónica Aterada o Scrapie (PrP Sc, predominancia de -laminar), todas las proteínas Naturales alteran su conformación o Estructura Terciaria. Se piensa que el origen de la forma alterada se encuentra en las Ovejas y Cabras, donde se manifiesta como Enfermedad Scrapie. Estos Ovinos son utilizados para realizar piensos por un lado, y pueden servir de alimento para algunos animales salvajes. La Enfermedad Esponjiforme Bovina (EEB) es transmitida a las Vacas a través del Pienso. Otros animales presentan Desgaste Crónico debido también al consumo de dicho pienso, como son Ciervos, Renos, Alces, Visones de Granja, Felinos en cautividad. Las Encefatopatías Espongiformes reciben su nombre de la similitud que presenta un corte hitológico de Cerebro infectado con una esponja. Aparecen agujeros debido a la degradación de la Materia Gris (pérdida de Neuronas). El origen de la enfermedad en humanos se debe principalmente al consumo de Carne de Vaca infectada. Puede manifestarse en distintas formas. Kuru: por consumo de Carne humana de misionarios (canibalismo). Más frecuente en consumo de mujeres (consumen Vísceras y Cráneo) que de varones (músculos). Insomnio Familiar Letal: Los individuos se duermen y no despiertan nunca. Suele deberse a la transmisión por electrodos en el Psiquiatra. Enfermedad de Creutfeldt-Jackob o de las Vacas Locas: Debido al consumo de carne de vaca infectada (o mutaciones en el gen PrPC). GSS: Debido al uso médico de hormonas y tejidos de cadáveres. 96 97 Parte III; LOS MICROORGANISMOS EN LA NATURALEZA Y EN LA INDUSTRIA I; Interacciones entre Microorganismos: A/ Tipos de Simbiosis (Láminas K): 1 Generalidades; la Simbiosis: Se trata de la asociación intima entre dos organismos, de manera que a menudo desarrollan dependencia vital. El microorganismo siempre sale beneficiado por el suministro de alimento y la protección que le otorga su Hospedador. Se distinguen Endosimbiontes (intracelulares) y Ectosimbiontes (extracelulares). Se diferencian también (III): Simbiosis Positivas (al menos un organismo sale beneficiado), como Mutualismo, Protocooperación o Comensalismo. Simbiosis Negativas (al menos un individuo sale perjudicado), como la Predación, el Parasitismo, o la Competitividad. El Mutualismo es la Simbiosis Positiva más importante y sorprendente, en la que ambos organismos salen beneficiados. Se diferencia de la Protocooperación porque sus integrantes desarrollan dependencia con su simbionte y no pueden desarrollarse en su ausencia. Se trata por ejemplo de los Líquenes. El Comensalismo es la relación que beneficia a unos Microorganismos, pero que es indiferente al Hospedador o el otro organismo que interacciona. El Parasitismo es la Simbiosis Negativa más extrema, pues a pesar de ser beneficiosa para los microorganismos parásitos (infecciosos) los otros organismos resultan dañados. Puede llegar a ser letal para los Hospedadores. El Hospedador se comporta como Foco de Infección. 2 Mutualismo: Es el caso más frecuente y extremo de Simbiosis positiva. Se establece una relación beneficiosa para ambos seres que la integran, de tal manera que la evolución les lleva a desarrollar una dependencia mutua. Los Líquenes son un ejemplo de Mutualismo entre Hongos (Ascomicetos o Basidiomicetos) y Algas (Cianobacterias, Xantofitas, Clorofitas). Las algas proporcionan metabolitos orgánicos sintetizados por la Fotosíntesis mientras el hongo aisla el sistema evitando que se deseque en medios atmosféricos donde las algas por si solas no pueden llegar, y además digiere su sutrato proporcionando a las algas materia mineral. Otros Microorganismos que pueden establecer Simbiosis con algas son; Paramecios, Protistas Ciliados o Amoebas. Dan lugar a distintos seres: Zooclorelas (con algas Clorofitas), Zooxantelas (con algas Xantofitas) o Cianelas (con Cianobacterias). Las algas proporcionan metabolitos orgánicos sintetizados por la Fotosíntesis mientras el animal presenta capacidad de desplazamiento y puede movilizar a todas las algas contenidas hacia los lugares de mayor luz. Existen también relaciones de Mutualismo entre Microorganismos y Vegetales. Por ejemplo las plantas Leguminosas establecen relaciones con bacterias (Rhizobium, Bradyrhyzobium, Azorhyzobium) capaces de fijar el Nitrógeno Mineral Atmosférico. Se Forman Nódulos Radiculares que precisan tanto de genes bacterianos como vegetales (por ejemplo, la Leghemoglogina es una proteína sintetizada por ambas). La planta es capaz de desarrollarse correctamente en un sustrato 98 pobre en Nitrógeno, gracias al N2 atmosférico. Otras simbiosis planta bacteria fijadora de Nitrógeno son Aliso-Frankia, y Azolla-Cianobacteria (II). Las Micorrizas (Ectomicorrizas, Endomicorrizas) son relaciones de Mutualismo entre Plantas (frecuentemente Coníferas) y Hongos. Los hongos aprovechan la raíz para crecer en su interior, y producen sustancias que estimulan su crecimiento y ramificación. La mayor amplitud de la raíz facilita la mayor captación de nutrientes, especialmente en suelos pobres. Existen también muchos casos de Mutualismo entre Microorganismos y Animales. Los Gusanos Tubiformes Gigantes de las profundidades marinas son un buen ejemplo. Son bentónicos y viven a 2500 metros de profundidad, en la zona Batipelágica. Viven en las chimeneas oceánicas, en condiciones de anoxia, altos niveles de Sulfuros y temperaturas alrededor de 100 ºC. Albergan en su interior bacterias capaces de oxidar el Sulfuro de Hidrógeno (SH) utilizando dicha energía para llevar a cabo la Fijación de Carbono inorgánico (CO2) en el Ciclo de Clavin, gracias a su Metabolismo Quimiolitótrofo. Muchos Metabolitos Orgánicos sintetizados por las bacterias sirven para alimentar a Gusano, mientras este proporciona un nicho protegido para las bacterias (IV). Los Rumiantes ingieren grandes cantidades de Celulosa, de muy difícil digestión. La Flora Intestinal es el conjunto de Bacterias que viven en simbiosis en el aparato digestivo y participan de forma coordinada en la digestión de estas sustancias en Azúcares, Acidos Grasos, generando a partir de ellos Acetato, Butirato, Propionato, que sirve como nutrientes al animal, y Metano y CO 2 que se desprenden en forma de Gases. Todas las Termitas presentan una fuerte dependencia de Trichonympha, un Protozoo Flagelado que forma parte de su Flora Intestinal Normal, y que también facilita la hidrólisis de la celulosa de la madera de la que se alimentan. Las Bacterias de los Géneros Photobacterium y Vibrio son productoras de Bioluminiscencia: Sintetizan la Proteína Luciferasa. Viven como Endosimbiontes en un gran numero de animales variados, especialmente de las profundidades oceánicas donde no alcanza la luz del sol. El Humano también presenta relaciones Mutualistas (en ocasiones de Comensalismo) con Bacterias y Hongos diversos, que reciben como conjunto el nombre de Microbiota Humana. Estas relaciones han sido estudiadas con especial ahínco, y se han sacado a la luz mutitud de Microorganismos que se encuentran naturalemente en determinadas partes de los diversos tejidos (Piel, Aparato Digestivo, Aparato Urogenital, Tracto Respiratorio Superior, esteril por debajo de la Laringe). Estos Microorganismos nos protegen ante invasiones de otros que pueden ser patógenos por competencia y producción de Bacteriocinas (Antibióticos). La gran diversidad de la Microbiota humana, estudiada a fono, nos da una idea de lo frecuente y habitual que son este tipo de relaciones con animales (VII, VIII, IX, X, XI). Entre la Microbiota Humana del Intestino Grueso se produce un caso de Comensalismo: encuentran dos bacterias que deben convivir: E. coli, anaerobia facultativa, y Brevibacterium, anaerobia estricta. Primero debe producirse la colonización de la primera que elimina el oxígeno, permitiendo así la llegada de otras bacterias, comensalistas de E.coli, como Brevibacterium. Esta última también se encuentra en las glándulas sudoríparas de la Piel, produciendo sustancias queconfieren el mal olor del sudor pueden servir como feromonas. Son también utilizadas para la maduración del Queso. 3 Parasitismo: Entre los parásitos podemos diferenciar dos principales tipos: Endoparásitos: Viven en el interior del organismo, no necesariamente de sus células. Ectoparásitos: Viven en el exterior del organismo, adherido a su cubierta o tegumento. Algunos integrantes de la Microbiota Humana pueden convertirse en Parásitos cuando proliferan excesivamente. Así mismo Brevibacterium puede llegar a causar una infección. 99 Todo proceso infeccioso tiene como finalidad la Reproducción del Parásito, ya que el Hospedador representa su Medio Óptimo y en muchas ocasiones su única Fuente de Alimento. En un ser vivo infeccioso, su forma parásita representa siempre un paso fundamental de su ciclo vital. Los procesos infecciosos presentan siempre varias fases comunes: 1. Adherencia (Reconocimiento; Adehsina Bacteriana, Fimbrias, Glicocalix…). 2. Penetración (en caso de Endoparásito en el organismo, si es Endosimbionte en la célula, incluso en vesículas, en algunos casos los parásitos son ectosimbiontes pero sus factores de toxicidad entran en la célula). 3. Reproducción, Crecimiento, Colonización o Diseminación (dispersión por el organismo). 4. Adaptación y Evasión del Sistema Inmune. 5. Producción de daños (Toxicidad o Invasividad). Las vías de diseminación de los Parásitos depende a la vez de su propia naturaleza, y de la del Hospedador infectado. Entre las más comunes en animales destacamos la vía Linfática, la vía Sanguínea y por Contiguidad de los tejidos. Shigella presenta un modo de diseminación peculiar entre dos células: utiliza proyecciones de Actina. Los daños tisulares pueden ser efectos secundarios (inclusiones por agregación de proteínas estructurales, secuestro de maquinaria celular) aunque se debe en la mayoría de caso a la producción de Toxinas (factores de toxicidad). Entre estas encontramos dos tipos: Endotoxinas: Proteínas estructurales que pueden aumentar su concentración con la muerte del parásito, como los Lipopolisacáridos de bacterias Gram -, o fragmentos de Peptidoglicano de las Gram +. Exotoxinas: Proteínas solubles y termoestables secretadas por las bacterias, a menudo propias de Cepas y codificadas por plásmidos o incluso propias de Bacteriófagos. Pueden actuar Localmente como las Endotoxinas, o actuar Globalemente accediendo al torrente de circulación Sanguínea. Son por ejemplo las toxinas Botulínica, Diftérica y Estafilocócica. La Virulencia de una Especie o Cepa de Parásitos depende del conjunto de proteínas que produce; tanto las proteínas estructurales y de adhesión, como endotoxinas y exotoxinas, y de su capacidad de Evadir al Sistema Inmune. B/ Respuesta del Sistema Inmune ante infección por Parásitos (Láminas K): 1 Evasión de la Respuesta Inmune: Existe una gran variedad de mecanismos para su evasión. Los virus tienen sus propios mecanismos, que ya hemos visto en Virus Animales (Deriva Antigénica…). Algunas Bacterias producen Proteasas para Inmunoglobina A (rompen los anticuerpos). Las bacterias (o cepas) con Cápsula (pared celular engrosada) presentan especial protección ante la Fagocitosis de Macrófagos, como Haemophilus influenzae. Algunas bacterias producen Bacteriocina, y pasan desapercibidas al sistema inmune como si se tratase de parte de la Microbiota Normal. Las bacterias con Sideróforos presentan gran afinidad por el hierro del organismo, pudiendo debilitarlo. El organismo ha desarrollado sistemas de defensa y protección de sus átomos de Hierro, como el Grupo Prostético Hemo, y las Apoproteínas que lo contienen. 2 La Respuesta Inmune; Defensas Inespecíficas: Las Defensas Inespecificas que presenta pasivamente el organismo y que suponen la primera barrera a flanquear por los Parásitos. Para comenzar, los endoparásitos deben superar por ejemplo la primera barrera física: el recubrimiento del organismo (piel, pared celular…) (XX). La Fiebre es el aumento de la temperatura corporal en respuesta a una infección, para dificultar la proliferación del parásito. 100 El Sistema del Complemento es un sistema termolabil del plasma sanguíneo, en el que participan una gran variedad de proteínas que son activadas en cascada por complejos procesos, cuyo principal fin es la destrucción de microorganismos infecciosos (XX, XXIV). Existen tres vías de activación: Vía Cásica: Activación por reconocimiento Antígeno-Anticuerpo. Vía Alternativa: Activación por reconocimiento de Endotoxinas, Polisacáridos, Lipopolisacáridos, Ácidos Teitoicos por Inmunoglobinas. Vía de las Lectinas: Reconocimiento de Manosa en la superficie de Microorganismos por Proteínas de unión a Manosa. Este sistema puede actuar por tes vías distintas: o Destrucción Directa del Microorganismo: Mecanismo llevado a cabo por un conjunto de Proteínas que se adhieren a la superficie del microorganismo: Las Proteínas del complejo de Ataque. Causan poros en la pared celular y membrana de los parásitos, lisando sus células (XX). o Reclutamiento de Células Inflamatorias. o Opsonización: Marcaje de las células parásitas de tal manera que puedan ser reconocidas y Fagocitadas. Los marcadores de membrana son proteínas del Complejo C3b, Anticuerpos, o la combinación de ambos (XXII). La Inflamación es la reacción localizada frente a un proceso traumático. Las células infectadas producen señales químicas que facilitan el reclutamiento de células inflamatorias de la sangre (Neutrófilos, también Monolitos y Macrófagos) por el tejido. Estas células atraviesan los capilares donde las células Endoteliales se encuentran activadas por los estímulos químicos. Entonces se produce un aumento localizado de la temperatura, del flujo sanguíneo y de la dilatación capilar (XIX). La Destrucción por Fagocitosis en la Ingestión o Endocitosis de Partículas Virulentas, Por Macrófagos, Monocitos y Neutrófilos. Este mecanismo puede se dependiente o independiente de la Opsonización si se reconocen proteínas propias de la célula infecciosa y no marcadores del sistema inmune, pero la capacidad de reconocimiento aumenta considerablemente en presencia de Opsoninas (Anticuerpos, C3b). Las bacterias reconocidas son fagocitadas. El fagosoma se fusiona con un lisosoma donde las partículas infecciosas serán digeridas. Algunas bacterias pueden subsistir en el interior de Fagolisosomas (XXIII). La Producción de Citoquinas por el Complejo Interferón son señales químicas que pueden activar muchas vías específicas (Celular) e inespecíficas de respuesta inmune, y funcionan como mediadores por quimiotaxis para señalizar la inflamación. Las Citoquinas son Lipoproteínas solubles liberadas por Monolitos y Linfocitos T. Su liberación es inducida generalmente por estímulos inespecíficos ante infecciones, aunque en ocasiones es inducida por estímulos específicos (XXV). 3 La Respuesta Inmune; Defensas Específicas: Este sistema es mucho más complejo, y presenta capacidad de memoria, de forma que permite una amplificación de la respuesta ante una segunda infección. Se distinguen Inmunidad Celular e Inmunidad Humoral (XXVI). La Inmunidad Humoral se basa en la producción de Anticuerpos por linfocitos B. Se trata de IgM e IgG (Inmunoglobinas circulantes y a nivel local). Se trata de la huella inmunológica, pues sus niveles sirven para calcular cuanto tiempo ha pasado desde que se ha producido la infección (más IgG en infección antigua, más IgM en infección reciente). Las múltiples funciones se los anticuerpos están descritas en (XVII). La Inmunidad Celular es el Atatque directo y destrucción por medio de proteínas digestivas de Linfocitos T a las células Infecciosas. 101 II; Taxonomía Bacteriana: A/ Principios de Taxonomía Bacteriana (Láminas L): 1 Generalidades: Disciplina científica en continuo cambio. El concepto de especie en bacterias es especialmente difícil de definir, y ha sido puesto en duda en numerosas ocasiones, especialmente durante la revolución molecular, que dio un vuelco radical a la taxonomía cladista, y notablemente a la clasificación, nomenclatura e Identificación de taxones bacterianos. Especie bacteriana: Conjunto de Cepas similares que difieren lo suficiente de otros grupos de Cepas (molecular y morfológicamente) como para asegurar su reconocimiento como unidad taxonómica básica. La Taxonomía Fenética o Numérica se basa en caracteres fenotípicos para clasificarlos por medio de matrices y dendogramas. Este proceso está computarizado. No se basa en la evolución. La Taxonomía Filogenética se basa en la evolución y utiliza para ello los datos moleculares, especialmente los genéticos. 2 Taxonomía fenética: Este tipo de clasificación no está basada en la evolución, pero es muy útil en taxonomía bacteriana para establecer el grado de diferencias entre dos cepas distintas. Se pueden calcular los coeficientes de Semejanza y de Coincidencia. Deben analizarse un alto número de caracteres, y solo adquieren validez los resultados de diferencias claramente marcadas. Entre la gran variedad de caracteres que pueden ser empleados encontramos dos criterios básicos: Clásicos: Caracteres morfológicos, metabólicos y algunos genéticos (morfología del Genóforo, presencia de Plásmidos, capacidad de Transformación o Conjugación). Moleculares: Entre ellas detacan los siguientes carácteres: El contenido en G+C del ADN: Sólo cuando la diferencia detectada es significativa, ya que si es similar no indica proximidad necesariamente (en otros seres vivos no es útil pues presentan muy poca variación entre ellos, X). La Hibridación de Ácidos Nucleicos: Se desnaturalizan las cadenas de dos cepas y se hibridan. La capacidad de hibridación entre las cadenas se corresponderá con las homologías entre ambas. Los valores de hibridación inferiores al 70 % indican que las dos cepas son muy susceptibles de pertenecer a distintas especies. Al ser inferiores al 25 % las cepas son de género distinto (XI, XII, XIII). La Secuenciación del ADN: Por medio de la secuenciación del Genoma completo o de Genes completos y la comparación computarizada de ambas nos sirve también para valorar la similitud entre dos cepas. ADN distintos pueden codificar proteínas similares o de misma función. Para valorar se calcula nº diferencias / nº Bases Nitrogenadas (XIV). 3 Filogenia Bacteriana: Se basa fundamentalmente en los criterios moleculares, especialmente en la secuenciación, que sirve para trazar los Árboles filogenéticos de los Genes, representandose así las relaciones evolutivas entre las especies de bacterias distintas. Este es el objetivo de la Taxonomía Filogenética. La secuenciación de los ARNr 16S de las distintas especies bacterianas adquieren un papel importante ya que estas moléculas presentan una distribución universal, una función fundamental en todas las bacterias, y a pesar de ser muy estables evolutivamente soportan un 1-2 % de variación cada 50 Millones de Años. Se estudian especialmente determinadas regiones especialmente representativas, y existe un elaborado banco de datos público (XVII, XVIII, XIX, XX). 102 103 III; MICROORGANISMOS DE ESPECIAL INTERÉS: A/ Microorganismos Patógenos (Láminas M): 1 Generalidades: Nos centraremos aquí en las Bacterias Patógenas, sabiendo que también existen Hongos y Animales Parásitos, y otros agentes infecciosos que ya hemos visto como los virus, viroides y priones. Las bacterias patógenas se clasifican en cuatro grupos en función de sus Vectores de Transmisión: Agua y Alimentos, Picadura de Artrópodos, Transmisión Sexual y Transmisión Aérea (I). Se diferencian también dos Mecanismos de Transmisión: Vertical y Horizontal (II). 2 Bacterias de Transmisión Aérea: Existe una grandísima variedad de Enfermedades causadas por Bacterias Infecciosas capaces de transmitirse por vía Aerea entre humanos (III). La Faringitis (-itis; inflamación) tiene un origen 30 % bacteriano y 70 % Vírico. La Faringitis Streptocócica es causada por S. pyogenes, un coco Gram + que también es responsable de Amigdalitis Streptocócica (Faringoamigdalitis) (IV, V). Algunas cepas presentan Toxina Eritrogénica, una toxina codificada por un Fago Lisogénico responsable de la Escarlatina (lengua aframbuesada). Se evita con antibióticos como la Penicilina. La Neumonía es una infección del Parenquima Pulmonar, cursada con tos y dolor pleural, y causada por una gran variedad de Bacterias Gram + y -, entre las que destaca Streptococcus pneumoniae y Pseudomonas (VI). La Difteria es una faringitis Pseudomembranosa causada exclusivamente por la bacteria Corynebacterium diphteriae. Las células de la Pseudomembrana son destruídas por la Toxina Diftérica, de forma que aumenta su volumen obstruyendo las vías respiratorias. Se trata de una Exotoxina tipo A-B (subunidades A Catalítica y B de Reconocimiento y unión a células diana) que penetra en las células inhibiendo la síntesis de proteínas y causando Necrosis. Puede extenderse por el torrente sanguíneo. La Difteria Severa son las manifestaciones sistémicas de la expansión de la bacteria o su toxina por todo el organismo. Algunos órganos diana son especialmente afectados, causando Conjuntivitis, o Cuello de Toro. La vacuna DTP (Difteria/Tétanos/Pertusis) que se encuentra incluída en el calendario obligatorio de vacunaciones. Esta vacuna incluye toxoides (toxinas inactivadas con formol) y bacteria B. pertussis muertas (VII). La Tosferina es una enfermedad que cursa con una tos muy intensa en ataques incontrolables y con secreciones gelatinosas. La bacteria Bordetella pertussis, bacilo Gram -, produce varias toxinas que alteran el tracto respiratorio, y por ello en la Vacuna DTP se utiliza la bacteria completa. Entre ellas se encuentran Toxina pertussis (altera la comunicación celular), Adenilato Ciclasa (inhibe a los leucocitos), Toxina Dermonecrótica (destrucción local de tejidos), y la Citotoxina traqueal. Afectan localmente a los tejidos respiratorios (VIII). La Tuberculosis es causada por unos peculiares bacilos Gram +, del grupo de las Micobacterias, que se caracterizan por su abundancia en ácidos Micólicos en su Pared Celular que las hace resistentes a la coloración Ácido/Alcohol: Mycobacterium tuberculosis. Se trata de una enfermedad altamente relevante y reemergente, pues entre 30 y 50 % de personas infectadas por VIH en fase SIDA la padecen, o padecen otras enfermedades relacionadas con Micobacterias. Esta enfermedad cursa en dos fases (IX, X, XI): 1. Primoinfección: Colonización de la bacteria por vía aerea, a traves de los aerosoles y escreciones salivares de un individuo infectado. Al llegar al pulmón se produce su Fagocitosis por Macrófagos alveolares. Estas bacterias en el interior del Fagosoma inhiben la unión de un Lisosoma Digestivo, e invaden los Macrófagos utilizándolos a modo de Caballo de Troya. A partir de ahí pueden dispersarse a través de la Linfa y de la Sangre. Forman Granulomas 104 (Tubérculos o Infiltrados) que aparecen como nubes blanquecinas en Radiografía; se trata de Agregados de Macrófagos, Monolitos, Neutrófilos… Estos granulomas pueden quedar cubiertos de Fibrina o Calcificados, quedando en su interior la bacteria latente. El 90 % de los infectados supera esta fase y acaba con la bacteria. 2. Tuberculosis Pulmonar Progresiva: Al dispersarse los Macrófagos infectados pueden formar Granulomas en distintos órganos diana, como el Riñón. En este avanzado estado se forman Cavernas o Agujeros en el pulmón, y se produce la destrucción de diversos tejidos. Los infectados muestran manifestaciones clínicas graves como dificultad para respirar, pérdida de peso, esputo sanguinolento y cansancio. La Tuberculosis Miliar es característica de enfermos en fase SIDA y muestra un sistema inmunne suprimido. Se producen agregados de micobacterias por medio de Ácidos Micólicos; efecto cordón. La Lepra es una infección que puede transmitirse tanto por vía aerea como por contacto directo con las heridas de la piel de una persona infectada. Es causada por Mycobacterium leprae, un Bacilo Hansen no cultivable en medios artificiales. Para estudiarlo se cultiva en Armadillos, y se realiza una tinción directa en corte histológico del tejido infectado. Su crecimiento es muy lento y su tg muy elevado: la incubación puede alargarse hasta 6 años desde contacto. Si la enfermedad no es tratada el deterioro es progresivo a lo largo de los años. Sus células Diana son Células Epiteliales cutáneas, Linfocitos, y Células de Shwann del Sistema Nervioso Periférico. La enfermedad puede ser tratada con mezclas de Antibióticos (pueden desarrollar dresistencia) pero el tratamiento se prolonga durante más de dos años. Cursa, al igual que la anterior, en varias fases con distintas manifestaciones (XII): 1. Lepra Difusa: Despigmentación por lesiones cutáneas (manchas planas sin color). Esta fase inicial puede evolucionar de dos formas dependiendo de la respuesta inmune. 2. Lepra Tuberculoide: Ante una respuesta inmune normal o debil. Proliferación en Fagosomas y formación de Granulomas similares a Tuberculosis. Avanzan las lesiones cutáneas y comienzan las deformidades en la cara. Comienza la afectación al sistema nervioso periférico causando atrofia muscular y pérdidas de sensibilidad en las extremidades. Puede evolucionar a (3.). 3. Lepra Lepromatosa: Ante una respuesta inmune muy debiliotada. Se produce el avance de todas las lesiones. Puede producirse pérdida completa de extremidades y partes de la cara, incluídos huesos y cartílagos (por ejemplo, de la nariz) dando lugar al aspecto leonino. La Legionellosis es la infección característica del Bacilo Gram – Legionella pneumophila, bacteria pleomórfica capaz de cambiar de forma durante la infección, y transmisible por vía aerea, a través de diminutas gotas de agua dispersadas con el viento. Sus modos de transmisión más frecuentes son a través del aire acondicionado y de las fuentes públicas de ciudades. Recibe este nombre (enfermedad de los Legionarios) ya que el primer caso se dio durante una convención de legionarios en Philadelphia, debido al aire acondicionado del Hotel donde se albergaron. Ataca al tracto respiratorio infectando el parénquima pulmonar (que carece de Microbiota). En individuos sanos el sistema inmune es capaz de controlar la infección, pero en inmunocomprometidos o ancianos con otras coinfecciones respiratorias causa alteraciones más graves, pudiendose incluso dispersar por el torrente sanguíneo y alcanzar riñón, hígado, bazo, sistema nervioso… Es muy difícil de erradicar ya que puede desarrollarse en el interior de protozoos de aguas estancadas, como Amoebas, y puede formar Quistes de Resistencia en aguas de las fuentes y aires acondicionado (XIII). La Meningitis es causada principalmente por Neisseria meningiditis, diplococo Gram – que forma parte de la Microbiota Humana del tracto respiratorio superior; 30 % de la población porta la bacteria en Nasofaringe. Su acción es muy rápida, pudiendo producir la muerte en sólo 6 o 7 horas. También puede debersa a la infección de otros Neumococos y Meningococos como Sterptococcus pneumoniae, S. agalactiae, o Haemophilus influenzae serotipo b (capsulada), todas ellas transmitidas por el aire. Cursa con fiebre. Se produce el bloqueo de la Barrera Hematoencefálica, ya que la inflamación de las Meninges dificulta la circulación de la sangre en el cerebro. Puede diseminarse por 105 el torrente circulatorio y afectar a otros órganos diana, formando por ejemplo Exantemas Cutáneos o Petequias; manchas violáceas en la piel que ponen de manifiesto la presencia de la bacteria en la sangre, también conocido como Meningococcemia, difícil de atacar y capaz de producir la muerte (XIV). 3 Bacterias Transmitidas por Picadura de Artrópodos: La Peste es causada por el bacilo Gram – Yersinia pestis, y produjo la muerte del 30 % de la población mundial durante la Edada Media; se trataba de una enfermedad característica urbana transmitida por roedores de ciudad, entre los cuales se dispersaba a través de la picadura de la Pulga. Algunas ratas murieron debido a la infección, pero la especie rápidamente desarrolló resistencia. Generalmente la transmisión de la Rata al Humano es mediada por pulgas que pican a la primera y a continuación al segundo, de tal manera que el humano es un Hospedador intermediario accidental. Actualmente es propia del Ciclo Selvático, y sus reservorios y Hospedadores Intermediarios se encuentran controlados. Al infectar, esta bacteria puede establecerse y proliferar en los Ganglios Linfáticos, y dispersarse a través del sistema Liunfático. Es capaz de desarrollarse en el interior de Macrófagos sin ser Fagocitada. Forman Bubones, o hinchamientos dolorosos con virulencia de los Ganglios Linfáticos. Presenta distintas manifestaciones en función de los tejidos infectados de forma secundaria (XVI, XVII): Peste Neumónica: En el tracto respiratorio. Único caso donde tener lugar un contagio directo humano-humano por vía aerea. Peste Negra: En el Epitelio Pluriestratificado de la Piel. Forma Necrosis del tejido y ennegrecimiento de las extremidades. Peste Septicémica: Formación de Múltiples Bubones en diversos tejidos. Tifus, Fiebre de las Montañas Rocosa: Se trata de dos manifestaciones sitémicas producidas por la infección de Rickettsias, Parásitos Endosimbiontes (intracelulares obligados), no cultivables en laboratorio sobre medios Artificiales. Las personas infectadas suelen presentar Perixantemas cutáneos, y si no son tratadas a tiempo, la dispersión de la bateria por la sangre puede dañar otros órganos diana como son Hígado, Bazo y Riñón. Presenta una elevada mortalidad, de alrededor del 6-30 %, y puede ser transmitida por una gran variedad de artrópodos como Garrapatas y Pulgas (XVIII). La Enfermedad de Lyne es una infección de fuerte incidencia en Estados Unidos, y muy estudiada por ello, causada por la Espiroqueta Borrelia burgdorferi. Las espiroquetas son por lo general poco infecciosas, y difícilmente observables; para ello se suele recurrir a Microscopía óptica de Campo Oscuro. Es transmitida por la Garrapata, y encuentra reservorios selváticos en Roedores silvestres. La bacteria penetra por el punto de picadura directamente a la sange, y en este punto forma un Eritema Migrans: enrojecimiento y endurecimiento de la piel que aumenta de tamaño concentricamente. Desaparece espontáneamente por lo que puede parecer una picadura inflamada, pero si no es tratado, pasados unos meses del Eritrema pueden aparecer otras complicaciones como Artritis o complicaciones neurológicas. 4 Bacterias de Transmisión Sexual: La Sífilis es causa por la espiroqueta Treponema pallidum, no cultivable en medios artificiales, por lo que se debe realizar un diagnóstico Xenológico (ensayo inmunológico). Se transmite por contacto sexual, por medio de la heridas y abrasiones que a menudo se producen durante el propio acto. Se manifiesta en varias fases (XXII, XXIII): 1. Sífilis Primaria: Entre las dos semanas y los dos meses de contraer la infección se forma el Chancre; una úlcera indolora, elevada y dura, que desaparece antes de las 6 semanas. En mujeres esta suele ser interna y difícil de detectar, pudiendo pasar fácilmente desapercibida. La infección suele ser superada después de esta fase, aunque pueden llegar al torrente sanguíneo e infectar otros órganos si no es debidamente tratada. 106 2. Sífilis Secundaria: Puedes formarse Exantemas Cutáneos por toda la piel (Periodo Florido). Pueden presentarse otros síntomas sistémicos. 3. Sífilis Terciaria: Si la enfermedad no es tratada, se producen granulomas en diversos órganos, que alteran su función. Puede aparecer Neurosífilis si alcanza el sistema nervioso (similar a Meningitis). Además esta bacteria puede atravesar la barrera placentaria, causando las malformaciones del feto e incluso el aborto propios de la Sífilis Congénita. La Gonorrea (Gonococia, Blenorragia) es la infección del diplococo Gram – Neisseria gonorrhoeae, una enfermedad de transmisión sexual (ETS) de relativamente alta incidencia, debido a la facilidad de contagio por contacto sexual heterosexual (en un solo contacto el hombre tiene 20 % de posibilidad de contraerla, la mujer 50 %). No se desarrolla memoria inmunológica frente a todos sus tipos, por lo que puede haber reinfección. Sobre todo en mujeres puede cursar de forma asintomática. Puede aparecer Uretritis Gonocócica y Disuria (dolor y ardor al miccionar) con exhudado purulento poco detectable, puediéndose confundir con una inflamación urinaria. El uso de anticonceptivos vía oral altera la mucosa genital facilitando su proliferación. En varones es más facil de identificar. Se produce Uretritis con Disuria y exhudado purulento de fácil identificación. Además pueden aparecer manifestaciones sistemáticas como Faringitis gonocócica o Proctitis si la bacteria se expande por la sangre. Además puede diseminarse en el Aparato Urogenital (Útero, Ovario, Trompas de Falopio) y causar esterilidad. No puede atravesar la barrera placentaria, pero a menudo se produce una transmisión congénita o vertical durante el parto por contacto, produciendose entre otros Oftalmia Neonatal (conjuntivitis severa) (XXIV, XXV). El Linfogranuloma Venéreo es causado por un grupo de patógenos intracelulares obligados, no cultivables en laboratorio, y causantes de múltiples infecciones. Chlamydia trachomatis, responsable de esta enfermedad, produce Uretritis no gonocócica, pero frecuentemente Postgonocócica (se manifiesta después de Gonorrea). Se trata en concreto de la formación de dolorosos Bubones, por acumulación bacteriana, en los ganglios inginales, produciendo hinchazón en la ingle. Esta bacteria presenta un ciclo vital característico, de forma que se replica en el interior de las células, y a continuacón las lisa para su liberación (similar a los virus). Su diagnóstico se basa en la observación en microscopía óptica de los Cuerpor elementales, o los Cuerpos Reticulados (XXVI, XXVII). 5 Bacterias Transmitidas por el Agua y los Alimentos: Dentro de este grupo se diferencian por el tiempo de infección y por lo síntomas causados tres tipos de infecciones: Infección alimentaria: proliferación de la bacteria viva. Se manifiesta después de un periodo de incubación. Toxoinfección alimentaria: proliferación de la bacteria y liberación de toxinas. Las manifestaciones pueden comenzar pocas horas después de la ingestión. Intoxicación alimentaria: ingestión de un alimento donde las bacterias han proliferado y producido toxinas. La intoxicación se manifiesta poco tiempo después de la ingestión. La Gastroenteritis es una una Infección alimentaria. que puede ser causada por una gran diversidad Enterobacterias, como E. coli, Shigella (agua), Salmonella (Microbiota de animales de granja), Yersinia (Microbiota de animales de granja), o Campylobacter (no enterobacteria, agua). Producen Fermentación de la Glucosa, y se distiguen por la fermentación de la lactosa en dos tipos: Coliformes (Lac+), pertenecientes a la Microbiota Humana, y No Coliformes (Lac -), estas últimas siendo responsables de este tipo de infección. Además, las anaerobias facultativas reducen el Nitrato a Nitritos. Por ejemplo, E. coli puede ser Coliforme y parte de la Microbiota, puede ser enterohemorrágica (carne de vacuno mal cocinada), o enterotoxigénica (síndrome del vijero, agua). Otras Infecciones alimentarias son Salmonelosis, producida por Salmonella, o Listeriosis, producida por Listeria monocytogenes. 107 El Cólera es una Toxoinfección relacionada con el consumo de agua, causada por Vibrio cholerae. La bacteria no penetra a través del intestino, pero produce una toxina tipo A-B (Toxina Colérica) en la luz del tubo digestivo que excita la actividad de Adenilato Ciclasa, produciendo cAMP intracelular. Esto produce la alteración del balance de electrolitos, la salida masiva de agua al intestino, y la consecuente deshidratación del organismo que puede llegar a perder hasta 20 litros por día. Se manifiesta por las heces en agua de arroz, en líquido similar al agua de cocción del arroz (XXX, XXXI). Las Fiebres Tifoideas y Paratifoideas son Infecciones causadas por Salmonella typhi y S. parathyfi transmitidas por el consumo de agua. Pueden pasar al torrente circulatorio, encontrando un Reservorio en la Vesícula biliar. La Intoxicación Alimentaria Estafilocócica es una Intoxicación producida por el coco Gram + Staphylococcus aureus capaz de producir hasta 7 exotoxinas termoestables. Se puede contagiar a través de cremas, dulces y ensaladas, y se multiplica en muchos alimentos no refrigerados. Entre 1 y 7 horas después de la ingestión se presentan síntomas similares a los de Gastroenteritis, acompañados por un fuerte dolor abdominal. El Botulismo es una Intoxicación severa producida por la Toxina Botulínica, liberada por el bacilo anaerobio estricto Gram + Clostridium botulinum, formador de esporas. Presenta una distribución Ubiquista y prolifera en alimentos conservados en condiciones anaerobias, como los enlatados. Al germinar las esporas en dichas condiciones, comienza la producción de toxinas y la intoxicación del alimento. Se trata de Exotoxinas tipo A-B, son en concreto Neurotoxinas, muy potentes en baja concentración, que pueden llegar a producir la muerte por parada cardiovascular. Bloquean la liberación de Neurotransmisor Acetilcolina en las sinapsis Neuro-Musculares inhibiendo la contracción, de tal manera que en personas intoxicadas suele producir Parálisis Flácida (XXXII). 6 Otros tipos de Transmisión: El Tétanos es la toxoinfección causada por los bacilos Clostridium tetanii, anaerobios estrictos formadores de esporas deformantes en palillo de tambor. Es transmitida por esporas a través de heridas infectadas, que al cicatrizar proporcionan el ambiene anaerobio necesario para su germinación y proliferación. Entonces comienza la producción de la Toxína Tetánica, una exotoxina tipo A-B que también actua como Neurotoxina, y tiene el efecto contrario al de la Toxina Botulínica; impide la recaptación de la Acetilcolina por inhibición de la liberación de la Glicina por la neurona inhibidora, y provoca Parálisis Espástica, muy dolorosa, y Risa Sardónica (contracción permanente de la cara). El Tétanos Neonatal puede ser transmitido por materiales no esterilizados utlizados para cortar el cordon umbilical (XXXIII). 108 109 B/ Microorganismos de Interés Industrial (Láminas N): 1 Generalidades, Microbiología industrial: Los microorganismos adquieren una especial importancia en multitud de procesos industriales relacionados especialmente con la Industria Alimentaria, la Industria Farmaceutica, y la Depuración de Aguas Residuales. La selección de los microorganismos se basa en unos criterios básicos como su cultivo fácil y barato, su rápido crecimiento, su alto rendimiento, su carácter no patógeno. Además la Ingeniería Genética jueg aquí un importante papel, de modo que se seleccionan muchos microorganismos bacterianos por su susceptibilidad a la manipulación genética. La función de los Microorganismos es distinta en cada caso, pero se distinguen tres procesos fundamentales que hacen de los microorganismos grandes fábricas industriales (II): Producción de Células: Por ejemplo levaduras para la industria alimentaria. Productos de Bioconversión: Utilización de la maquinaria metabólica de los microorganismos para catalizar la síntesis de determinados productos a partir de sustratos de fácil obtención, muy difil de llevar a cabo en el laboratorio por síntesis química. Productos Derivados: Utilización de los productos naturales sintetizados por los Microorganismos. Sirve para la obtención de Antibióticos, Enzimas, Alcohol (fermentaciones). Se distinguen los metabolitos primario y secundarios en función de la fase de crecimiento de la población en la que se maximiza su síntesis a partir de Glucosa. Esta distinción debe ser tenida en cuenta para la mayor efectividad del proceso (III): Los Metabolitos Primarios, como los productos de la Fermentación, crecen mientras lo hace la población durante la Fase Exponencial de Crecimiento. Este es el mismo criterio a tener en cuente para la Producción de Células. Los Metabolitos Secundarios, como muchos Antibióticos, no son esenciales para el crecimiento y su producción se maximiza durante la Fase Estacionaria. 2 Cultivos Industriales: Para rebajar los costes al máximo se suelen utilizar como nutrientes para cultivos de Gran Volumen, en la medida de lo posible, los productos de desecho de otras industrias, en especial del campo agroalimentario (IV). Las Fuentes de Carbono más comunes son las Melaza (líquidos densos, subproductos de la cristalización del azúcar), u otros subproductos como semillas o sueros. Como Fuentes de Nitrógeno suelen utilizarse Amoniaco, Sales de Amonio y Nitratos, subproductos de los mataderos y fábricas de harina de Soja. Los Fermentadores Industriale son enormes recipientes provistos para el cultivo industrial de Microorganismos. Antes de poner en funcionamiento un Fermentador industrial, que puede contener entre 100.000 y 500.000 litros, deben realizarse ensayos y comprobar que el proceso funciona correctamente. El Fermentador precisa de Palas de Agitación que pueden producir espumas, debe estar provisto de Antiespumantes, y disponer de sistemas de Aireación y Refrigeración (V). Para ponerlo en marcha se procede por Escalado Industrial, aumentando progresivamente el volumen del cultivo (VI): 1. Tubo de ensayo, Matraz o Erlen-Meyer para valorar el proceso y su rendimiento. 2. Fermentador de laboratorio, utilizado como simulador del proceso a nivel industrial. Sirve para la regulación de los parámetros de tasa de aireación, temperatura y pH óptimos… 3. Fermentador de Planta Piloto, con capacidad de hasta 3.000 litros, donde se analiza y comprueba el correcto funcionamiento del proceso en cultivos de gran volumen. 4. Fermentador industrial. Aún durante el proceso de producción industrial, deben seguir llevandose a cabo controles y seguimiento. 110 3 Industria Farmaceutica (médica): Dentro de este campo destacan la producción de Antibióticos por los géneros bacterianos Streptomyces y Bacillus, y los hongos Penicillium. Suelen ser productos derivados, aunque a menudo son modificados químicamente, como la Penicilina (VIII, IX, X). También se utilizan bacterias modificadas por Ingeniería genética para la producción de Enzimas y Hormonas Humanas o animales, como por ejemplo Insulina, o Esteroides. A menudo se obtienen por Bioconversión, como es el caso de muchos esteroides como la Cortisona y Hormonas sexuales (XVI). Para ello pueden utilizarse Microorganismos en cultivo, inmovilizados, o incluso sus enzimas. En caso de proteínas se pueden obtener como productos, gracias a la ingeniería genética. Son generalmente Metabolitos Secundarios. El producto deseado debe ser purificado y en su caso cristalizado, a partir del medio del Fermentador. Dentro de la Producción de enzimas destaca la producción de Extremoenzimas (enzimas de Extremófilos) capaces de trabajar en condiciones de temperaturas o pH extremos, que pueden ser utilizadas para diagnósticos médicos, como por ejemplo Tac polimerasa. Para su fabricación no es necesaria la refrigeración del Fermentador. 4 Industria Agroalimentaria: Existen una gran variedad de productos y procesos dentro de esta industria que precisan de la acción de los Microorganismos. Entre ellos destaca la producción aditivos para muchos alimentos. Entre ellos encontramos: Ácidos Orgánicos: Son utilizados como acidulantes y conservantes. Ácido Láctico, producido por Bacterias Lácticas y algunos Hongos. Ácido Acético, producido por Acetobacter y Glicconobacter, es un acidulante muy importante para Encurtidos y Mayonesas. Aminoácidos: Modifican el sabor de los alimentos. Glutamato (monosódico), saborizante muy utilizado en sopas, con sabor a carne (Avecrem). Muy utilizado en oriente. Glicina: Suavizante utilizado para contrarrestar el Ácido Acético. Aspartamo (Asp + Fenilalanina), edulcorante. Lisina, aditivo nutritivo fundamental en alimentación animal, añadido a los piensos. Polisacáridos: Emulsionantes y Estabilizantes, utilizados durante los procesos de producción. Xantato. Alginato, producido por Pseudomonas y Azotobacter. Vitaminas: Componentes Nutricionales que pueden actuar como conservantes. Vitamina C, o ácido ascórbico, antioxidante y conservante, producido por Erwinia y Acetobacter. 5 Elaboración de Alimentos. Además de Producción industrial, los Microorganismos son componentes fundamentales en los procesos de elaboración de alimentos, principalmente debido a las Fermentaciones Láctica y Alcohólica (Etanólica y Formólica). La Fermentación Láctica puede ser de varios tipos, por sus productos: Homoláctica; Producción de Lactato Vía Glucolítica. Heteroláctica: Producción de Lactato, además de otros productos como Etanol y CO2, Vía Glucolítica. Esta fermentación puede ser llevada a cabo por distintas bacterias, a distintas temperaturas. Clasificamos las fermentación láctica por su temeratura óptima: Mesófila: entre 20 y 30 ºC, Matequilla. Termófila: 45 ºC, Yogur. 111 En ocasiones la Fermentación heteroláctica puede ser Terapeutica si entre sus productos se encuentran compuestos nutritivos o ventajosos, como Bifidobacterium. Los Bífidus ayudan a mantener el equilibrio intestinal, mejora la tolerancia a la lactosa y ayudan a eliminar los excesos de colesterol. Entre las principales bacterias que realizan la fermentación láctica se encuentran Lactobacillus, Lactococcus, Bifidobacterium, Streptococus. Asimismo, para evitar el biodeterioro la leche debe ser Homogeneizada, Estabilizada y finalemente Pausteurizada antes de su comercialización. La producción de Quesos implica un proceso de maduración a 8-10 ºC que dura varios años, en el que intervienen principalmente los Hongos Penicillium, pero tambien algunas bacterias como Brevibacterium. Los encurtidos son también productos vegetales fermentados por Microorganismos (XIV). Muchas enzimas son también producidas con fines agroalimentarios, para la producción de alimentos (XV). 6 Biodeterioro: Se trata de la infección de un producto, no de un ser vivo. El producto es degradado y metabolizado por los microorganismos descomponedores de la materia: por su crecimiento y proliferación, o más comúnmente por alteraciones químicas. El proceso puede ser Asimilatorio si el microorganismo capta los metabolitos que procesa, o Desasimilatorio si los metabolitos producidos son liberados al medio de cultivo (XVII, XVIII). Controlar estos procesos es muy util tanto para evitar la degradación y alteración indeseada de alimentos y productos orgánicos útiles, y la oxidación y obstrucción de cañerías metálicas, como para la correcta y completa degradación de muchos productos de desecho como sucede en la depuración de aguas sucias. En este proceso intervienen una serie de factores que podemos clasificar en dos grupos (XIX): Intrínsecos: Condiciones propias del material. Extrínsecos: Factores del medio, como temperatura, pH y humedad. Entre los productos susceptibles de Biodeterioro destacan productos de origen vegetal, animal, metales y piedras, y alimentos y aguas contaminadas (XX). El Biodeterioro de metales y piedras es un proceso combinado fisico-químico y microbiológico. En Metales puede dar lugar a corrosión; manchas, oxidación, perforación e incluso transformación en gelatina. Tiene lugar en dos principales pasos: 1. Obturación: crecimiento de microorganismos y degradación en condiciones anaeróbicas. Crecimiento de bacterias Sulfato-reductoras que producen SH2. 2. Corrosión. En piedra tiene lugar un proceso mixto de meteorización y erosión y la acción de Microorganismos que pueden facilitar la solubilización de la piedra. En el Biodeterioro de Alimentos y Aguas contaminadas participan una gran variedad de microorganismos. Se producen un gran número de alteraciones del producto final, como reblandecimiento, turbidez, gases, al igual que alteraciones organolécticas en la acidez y sabor de los alimentos, pudiendo incluso quedar intoxicados. El deterioro de productos alimenticios debe ser controlado, ya que puede ser responsable de grandes pérdidas económicas. 112