Metabolismo de lipoproteínas
Anuncio

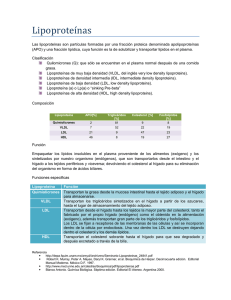
2013 Metabolismo de lipoproteínas Bqca. Patricia N. Goicoechea – Aux. doc. Bqca. María Victoria Aguirre – Profesora titular QUÍMICA BIOLÓGICA II Química Biológica II, FaCENA, UNNE METABOLISMO DE LIPOPROTEÍNAS DIGESTIÓN DE LÍPIDOS Los lípidos de la dieta incluyen triacilgliceroles, fosfolípidos, colesterol, ésteres de colesterol y vitaminas liposolubles (A, D, E y K). La digestión de estos nutrientes insolubles en agua está favorecida por la presencia de sales biliares que forman pequeñas micelas suspendidas en el contenido intestinal, y ofrecen una gran superficie para el ataque de enzimas hidrolíticas. Triacilgliceroles El proceso de degradación comienza en el estómago, catalizado por la lipasa gástrica. Ataca las uniones éster de ácidos grasos en los carbonos primarios del glicerol, los productos son ácidos grasos libres, 1,2-diacilgliceroles y 2-monoacilgliceroles. Cuando el contenido gástrico pasa al duodeno, se encuentra con las sales biliares que favorecen la emulsión de lípidos. Las sales biliares forman una capa alrededor de las moléculas hidrófobas, reducen la tensión superficial y permiten la dispersión de las grasas en finas partículas que se estabilizan en el medio acuoso. La presencia de otras moléculas anfipáticas como fosfolípidos, ayudan a formar las micelas. La colipasa desplaza a las sales biliares para permitir la acción de la lipasa pancreática con los triacilgliceroles. Los productos finales de la digestión de grasas son ácidos grasos libres y 2-monoacilgliceroles, se liberan muy pequeñas cantidades de glicerol. Fosfolípidos La fosfolipasa A2 presente en la micela cataliza la hidrólisis de la función éster en la posición 2 para dar lisofosfolípido y ácido graso libre, la acción enzimática requiere de sales biliares. Ésteres de colesterol Son escindidos en colesterol y ácido graso por colesterol esterasa. Esta enzima depende de la presencia de sales biliares e hidroliza ésteres de triacilgliceroles y ésteres de vitaminas A, D y E. ABSORCIÓN DE LÍPIDOS La mayor parte de la grasa ingerida es degradada a 2-monoacilglicerol, compuesto que ingresa al enterocito donde se realiza una importante actividad metabólica. La principal vía de síntesis de triacilglicéridos parte de ácidos grasos y 2-monoacilglicerol. Los ácidos grasos se unen a la coenzima A para dar el compuesto activado acil-coenzima A, la reacción es catalizada por la tioquinasa y la energía es provista por hidrólisis de ATP: Tioquinasa ACIDO GRASO + CoA ACIL CoA ATP AMP+PPi Otra vía de síntesis de triacilglicéridos y fosfolípidos es la vía del ácido fosfatídico y requiere glicerol-3-fosfato. 2 Química Biológica II, FaCENA, UNNE Éste se forma por fosforilación de glicerol catalizada por la gliceroquinasa o por reducción de Dihidroxiacetonafosfato (proveniente de la glucólisis) catalizada por la glicerofosfato deshidrogenasa. Una pequeña parte de glicerol libre y ácidos grasos de cadena corta (menos de 10 Carbonos) son liberados a la luz intestinal, difunden pasivamente a través de las membranas y pasan a los capilares del sistema porta. Estos triacilgliceroles neosintetizados, son incluidos con una pequeña porción de colesterol en partículas lipoproteicas (Quilomicrones) encargadas del transporte de lípidos exógenos. El hígado también realiza intensa síntesis de TRIACILGLICEROL que son enviados a la circulación mediante lipoproteínas de muy baja densidad, responsables del transporte de lípidos endógenos. En los capilares sanguíneos estos lípidos sufren hidrólisis total formando ácidos grasos y glicerol que pasan a las células. El glicerol es metabolizado en los tejidos con capacidad para fosforilarlo, los ácidos grasos pueden ser oxidados en los distintos tejidos para obtener energía o ser almacenados en tejido adiposo como una importante reserva de energía. Las grasas poseen ventajas sobre los hidratos de carbono porque su valor calórico es más del doble: Grasas 9kcal por g Carbohidratos y proteínas 4kcal por g Además el contenido de agua en depósitos de grasa es muy reducido comparándolo con el glucógeno. Las grasas constituyen la forma más concentrada de proveer y almacenar energía química potencial. TRANSPORTE DE LIPOPROTEÍNAS EN PLASMA La grasa absorbida de la dieta y los lípidos que se sintetizan en el hígado deben ser transportados entre los tejidos y órganos para su uso y almacenamiento. Como los lípidos son insolubles en agua, el problema se resuelve combinando: Lípidos no polares: Triacilgliceroles, Ésteres de Colesterol, Ácidos Grasos Libres Lípidos anfipáticos: Fosfolípidos, Colesterol libre Proteínas De esta manera se vuelven solubles en el plasma. ESTRUCTURA DE LIPOPROTEÍNAS Las lipoproteínas son partículas globulares que presentan un núcleo no polar de triacilgliceroles y ésteres de colesterol rodeados por una cubierta de fosfolípidos anfipáticos, proteínas y moléculas de colesterol libre. La figura muestra la composición de la monocapa de membrana de fosfolípidos de 2 colas con las cabezas hacia la 3 Química Biológica II, FaCENA, UNNE superficie externa, algunas moléculas de colesterol dispersas, poros de proteínas que atraviesan el espesor de la membrana. La porción proteica se conoce con el nombre de Apoproteínas, algunas forman parte de la partícula y no se eliminan, mientras que otras están libres y pueden ser transferidas. CLASES DE LIPOPROTEÍNAS PLASMÁTICAS Debido a que la grasa es menos densa que el agua, la densidad de las lipoproteínas disminuye en la medida que aumenta la proporción entre lípidos y proteínas. Se identificaron 4 tipos de lipoproteínas muy importantes fisiológicamente y en el diagnóstico clínico: Derivados de la absorción intestinal de triacilglicerol y otros lípidos. Obtenidas del hígado para la exportación de triacilglicerol Representan la etapa final del catabolismo de VLDL Intervienen en el metabolismo de VLDL, Quilomicrones y en el transporte invertido del Colesterol Los triacilgliceroles predominan en Qm y VLDL, mientras que Colesterol y fosfolípidos son más abundantes en LDL y HDL. Además se separan de acuerdo a su movilidad electroforética (ver fig.). APOLIPOPROTEÍNAS – FUNCIONES Las apolipoproteínas revisten la superficie de las lipoproteínas. - Forman parte de la estructura de las lipoproteínas: Las principales apolipoproteínas de HDL son las Apo A. Las LDL, IDL y VLDL tienen principalmente apolipoproteína B100, una de las cadenas polipeptídicas más largas que se conocen: 4536 aa. Los Qm contienen una forma truncada de Apo B100 llamada Apo B48, formada del mismo ARN mensajero e idéntica en el 48% de la porción inicial. Las Apo CI – CII y CIII son más pequeñas y libremente transferibles entre varias lipoproteínas. - Son cofactores de enzimas: Apo AI para Lecitin colesterol acil transferasa Apo CII para Lipoproteínlipasa - O inhibidores de enzimas: Apo AII y CIII para Lipoproteínlipasa Apo CII para Proteínas de transferencia de éster de colesterilo - Actúan como ligando para la interacción con receptores de lipoproteínas en tejidos: Apo AI para receptor de HDL. Apo B100 y Apo E para receptor de LDL. Además la Apo E se encuentra en Qm, Qm remanentes, VLDL, IDL. 4 Química Biológica II, FaCENA, UNNE VIA EXÓGENA DEL METABOLISMO DE LIPOPROTEÍNAS Formación de Quilomicrones: Los ácidos grasos libres son absorbidos por las células de la mucosa intestinal y combinados con glicerol para formar TAG, los cuales son empaquetados con pequeñas cantidades de colesterol esterificado en una capa de fosfolípidos, colesterol libre y Apo B48 para formar los Qm, en el núcleo hidrofóbico se incorporan también las vitaminas liposolubles absorbidas de la luz intestinal. Las Apo B48 sintetizada en el retículo endoplásmico rugoso se incorpora a las lipoproteínas en el retículo endoplásmico liso (sitio principal de síntesis de TAG), luego en el complejo de Golgi se adicionan residuos de carbohidratos. Los Qm nacientes viajan en vesículas desde el complejo de Golgi a la membrana basolateral, son secretados por exocitosis al espacio intersticial y llegan a capilares linfáticos, luego al conducto torácico y de ahí a la vena subclavia. Las partículas de 90 a 100 nm de diámetro y densidad menor a 0,95 g/ml, aparecen 1 hora después de la ingestión de lípidos y continúan ingresando por varias horas, se requieren más de ocho horas de ayuno hasta la desaparición en sangre. La presencia de Qm otorga al plasma un aspecto turbio o lechoso, debido al alto contenido de TAG. Al ingresar a la circulación, reciben Apoproteínas C y E transferidas desde HDL. En el interior de los capilares los Qm se unen a la Lipoproteinlipasa (LpL) (anclada en endotelio de los capilares sanguíneos por cadenas de proteoglucanos con carga negativa), enzima activada por Apo CII, que cataliza la hidrólisis de TAG contenidos en el interior de la partícula. Los ácidos grasos liberados pasan rápidamente a las células subyacentes, los principales sitios de actividad de la LpL son los capilares de tejido adiposo, miocardio, músculo esquelético y glándula mamaria. Los ácidos grasos captados se utilizan para sintetizar triacilgliceroles y almacenarlos en tejido adiposo, para oxidarlos y obtener energía en músculo cardíaco y esquelético o para secretarlos en glándula mamaria. El otro producto de la hidrólisis es el GLICEROL, tomado del plasma para ser metabolizado principalmente por los hepatocitos. Este proceso de lipólisis reduce el tamaño de los Qm, la partícula pierde gran parte de su masa original, aumenta la proporción de colesterol y la cubierta anfipática resulta grande. El exceso de fosfolípidos de superficie es transferido a HDL, a las cuales se les devuelve las apoproteínas CI y CII, formándose los Qm remanentes. Éstos se disocian del endotelio capilar, vuelven a circular para ser captados por el hígado. Los Qm remanentes poseen un diámetro de 45 a 50 nm, densidad menor a 1,006 y están relativamente enriquecidas con ésteres de colesterol y Apoproteínas B48 y 5 Química Biológica II, FaCENA, UNNE E. Los hepatocitos tienen receptores para Apo E, que fijan los remanentes y los internan por endocitosis para hidrolizar y metabolizar sus ésteres de colesterol y triacilgliceroles, la vida media en circulación es menor a una hora. TG de la dieta INTESTINO DELGADO VIA ENDÓGENA DEL METABOLISMO DE LIPOPROTEÍNAS Lipoproteínas de muy baja densidad: VLDL Los triacilgliceroles sintetizados en el hepatocito son incorporados a partículas VLDL junto con ésteres de colesterol. La principal proteína de cubierta es la Apo B100. Las partículas completas son secretadas por exocitosis hacia los espacios de Disse, alcanzan la luz de capilares sinusoides y llegan al torrente sanguíneo, donde sufren cambios similares a los Qm. Estas partículas de diámetro comprendido entre 30 y 90nm, densidad de 0,95 a 1,06g/ml, transportan principalmente TAG y circulan en plasma durante 4horas. Primero las VLDL reciben apoproteínas C y E procedentes de HDL, luego en capilares de tejidos extrahepáticos son sometidas a la acción de la Lipoproteinlipasa activada por Apo CII, que extrae ácidos grasos y glicerol al fragmentar los triacilgliceroles para el metabolismo energético de los tejidos o para su depósito en tejido adiposo además estas lipoproteínas cambian triacilgliceroles por ésteres de colesterol con HDL. Las VLDL reducen su tamaño, sobran fosfolípidos que son transferidos a HDL, retornan a éstas las Apo CII, con lo cual se interrumpe la acción de la Lipoproteinlipasa. Los cambios sufridos por estas partículas las convierten en IDL. El tamaño se reduce a un diámetro de 20 a 25 nm, con una densidad de 1,006 a 1,019 g/dl, sus apoproteínas son B100 y E, la vida media en plasma es de 2 a 5 horas. 6 Química Biológica II, FaCENA, UNNE Los receptores existentes en el hepatocito captan más de la mitad de IDL mediante el receptor de LDL (Apo B100/E) y las internan por endocitosis. Como no tienen Apo CII, la Lipoproteinlipasa no actúa sobre sus triacilgliceroles, sin embargo siguen siendo hidrolizados por la LIPASA HEPÁTICA (enzima extracelular localizada en la pared de los capilares sinusales del hígado, insensibles a la regulación por Apo C). La apoproteína E es devuelta a HDL y las VLDL se convierten en LDL. Lipoproteínas de baja densidad: LDL Las LDL son las principales lipoproteínas aterogénicas, representan el producto final de las VLDL. La densidad es de 1,019 a 1,063 g/ml y su tamaño reducido de 20 a 25 nm, contienen ésteres de colesterol y pequeñas cantidades de triacilgliceroles, la vida media en plasma es de 2,5 días. En su superficie se encuentra la Apoproteína B100, la cual es el ligando para el receptor LDL presente en hígado y muchos tejidos extrahepáticos. Las LDL son captadas por los receptores, introducidas por endocitosis y sus componentes hidrolizados por enzimas lisosomales. Los productos resultantes: ácidos grasos, aminoácidos y colesterol pasan al citosol. El colesterol es incorporado a membranas y a algunas células de la corteza suprarrenal, ovarios y testículos, también es utilizado para síntesis de hormonas esteroides. El exceso es reesterificado por acción de la acil CoA colesterol aciltransferasa (ACAT) y almacenado en la célula, los receptores LDL son reciclados a la membrana plasmática. Las cifras plasmáticas de LDL están controladas por la actividad de los receptores para LDL de los hepatocitos, cuya expresión aumenta o disminuye con el contenido intracelular de colesterol en el hígado. 7 Química Biológica II, FaCENA, UNNE Lipoproteínas de alta densidad: HDL Se sintetizan y secretan en hígado y en menor proporción en el intestino delgado. Sin embargo, las Apo C y E se sintetizan en hígado y se transfieren desde HDL hepática hasta la HDL intestinal cuando esta última entra al plasma. Las HDL nacientes son bicapas de fosfolípidos (fosfatidilcolina), Apolipoproteínas A, C, E y colesterol. Son partículas pequeñas con un diámetro a veces menor a 5 nm y hasta 20nm, con una densidad de 1,063 a 1,21 g/dl son consideradas antiaterogénicas. Actividades de HDL: - Transferencia de apoproteínas Las Apo A son las principales proteínas de las HDL y permanecen siempre unidas a estas partículas. Las Apo C y E son transferidas a Qm y VLDL. La Apo CII es un factor activador de la LpL, que retorna a las HDL cuando cumple su función. La Apo E cedida a VLDL regresa a HDL cuando las IDL pierden su contenido de triacilgliceroles. - Transporte invertido del colesterol A través de la pared de los capilares de tejidos extrahepáticos, las HDL interactúan con la membrana plasmática de las células subyacentes en un proceso en el que interviene Apo AI. El colesterol intracelular es movilizado a la superficie de las células y transferido a la partícula de HDL, éste es rápidamente esterificado por Lecitina-colesterol-aciltransferasa (LCAT), activada por Apo AI. LCAT Colesterol + Fosfatidilcolina Éster de colesterol + lisofosfatidilcolina (Lecitina) (Lisolecitina) Los ésteres de colesterol se desplazan al interior hidrófobo de la bicapa, la lisolecitina se transfiere a la albúmina plasmática y las HDL aumentan su tamaño pasando de la forma discoide a la esférica. Estas lipoproteínas transfieren ésteres de colesterol a Qm y VLDL mediada por la Proteína de transporte de ésteres de colesterol (CETP), de esta manera la LCAT interviene en la eliminación del exceso de colesterol no esterificado de lipoproteínas y tejidos. En intercambio por el colesterol esterificado, las lipoproteínas ricas en triacilgliceroles Qm y VLDL, entregan TAG a las HDL. Los ésteres de colesterol son finalmente tomados por el hígado con los remanentes de Qm y las IDL para completar el transporte invertido del colesterol. Por otra parte las HDL proveen colesterol a tejidos esteroidogénicos uniéndose a receptores especiales transfiriendo colesterol a las células directamente. - HDL2 y HDL3 La HDL3 más pequeña acepta colesterol proveniente de los tejidos mediante el Transportador-1 de casete de enlace con ATP (ABC-1), que permite que el colesterol sea transportado a través de la membrana. El colesterol es esterificado mediante LCAT por lo que aumenta el tamaño de la partícula para formar HDL2 menos densa. Debido a la transferencia de ésteres de colesterol a Qm y VLDL (como mencionamos anteriormente) y la posterior hidrólisis de TAG y fosfolípidos por 8 Química Biológica II, FaCENA, UNNE acción de la Lipasa hepática, estas lipoproteínas se convierten en HDL3 con capacidad para adquirir más colesterol de células periféricas. Remoción de HDL Una pequeña proporción de HDL es captada por receptores LRP en hepatocitos y retirada de la circulación. Cualquier excedente de la Apo AI se destruye en el riñon. SÍNTESIS EN IMÁGENES DE METABOLISMO DE LIPOPROTEÍNAS 9 Química Biológica II, FaCENA, UNNE RECEPTORES Receptor de LDL Se encuentra en casi todas las células Reconoce como ligando a Apo B-100 presentes en VLDL, IDL y LDL La síntesis de receptores es regulable (down regulation): cuando aumenta el colesterol intracelular, se inhibe la producción de receptores. Receptor de remanentes LRP Presente en hígado, cerebro, placenta Reconoce como ligando a Apo E La síntesis no es afectada por niveles intracelulares de colesterol. Receptor recolector de residuos Fijan LDL modificadas químicamente Captación por macrófagos de proteínas alteradas Receptor de HDL Presente en células adiposas, endotelio, tejidos esteroidogénicos, fibroblastos. Ligan Apolipoproteínas A-I, A-II, A-IV La interacción HDL-Receptor inicia señales que promueven la transferencia de colesterol intracelular a la membrana y de ahí a HDL Bibliografía Murray, Robert K (2004). Harper: Bioquímica ilustrada. 16a Edición. México: Manual Moderno. Blanco, Antonio (2006). Química Biológica. 8a Edición. Editorial El Ateneo. Voet, Donald Voet, Judith; Pratt, Charlotte (2007). Fundamentos de Bioquímica. 2 a Edición. Editorial Panamericana. 10