la raíz estructura primaria - Anatomía de Espermatófitas
Anuncio

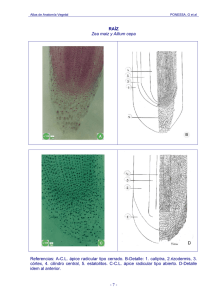
LA RAÍZ ESTRUCTURA PRIMARIA Introducción. Las raíces son por lo general órganos subterráneos, a veces son aéreas, como en el “clavel de aire” Tillandsia sp. (Bromeliaceae) y muchas otras epífitas. Las raíces son propias de las cormófitas (Pteridófitas y Espermatófitas). Las Talófitas poseen estructuras semejantes a las raíces que reciben el nombre de rizoides. La raíz es el órgano vegetativo de la planta adaptado para la absorción y conducción de agua con sales minerales. Además fija la planta al substrato y almacena sustancias de reserva elaboradas en los órganos verdes. Otra función es la reproducción agámica o multiplicación de las plantas (e.g., raíces gemíferas de la “batata” Ipomoea batatas L., Convolvulaceae). El sistema radical tiene su origen en la radícula del embrión. Esta radícula posee el tejido meristemático que originará los tejidos primarios de la raíz y produce su alargamiento o crecimiento en longitud. RADÍCULA DEL EMBRIÓN. Escala: 1 mm En las Dicotiledóneas el sistema radical es alorrizo, es decir, que está formado por raíces de diferente diámetro que provienen de la radícula del embrión. La radícula del embrión se transforma en una raíz principal y a partir de ella nacen raíces secundarias. Existen dos formas del sistema alorrizo: 2012 Pivotante o Axonomorfo: cuando la raíz principal penetra profundamente en el suelo y sus raíces secundarias –originadas en el periciclo de la raíz- alcanzan poca extensión y diámetro. Fasciculado: Cuando la raíz principal es superada por las raíces secundarias (no confundir con fasciculado de origen adventicio). En las Monocotiledóneas el sistema radical es homorrizo es decir, está formado por raíces de aproximadamente el mismo diámetro de origen adventicio. Las raíces adventicias se originan en cualquier parte de la planta (tallo, hojas, etc.), menos en el embrión. En las monocotiledóneas el embrión posee radícula, pero su extensión y vida es corta, siendo rápidamente reemplazadas por las raíces adventicias. El sistema homorrizo adventicio, por su forma se le llama en cabellera o fasciculado y es característico de los pastos de la familia Poaceae o Gramíneas. Los sistemas radicales crecen preferentemente en las capas superficiales del suelo, sin embargo, las raíces de algunas plantas dicotiledóneas (alorrizo), principalmente en zonas desérticas, penetran hasta 30 m de profundidad tratando de alcanzar el nivel freático subterráneo. ZONAS DE LA RAÍZ EN SENTIDO LONGITUDINAL Observaciones. La raíz se diferencia del tallo porque no posee nudos, entrenudos y hojas, por lo tanto, la yema apical de la raíz se diferencia de la yema apical del tallo porque tampoco presenta nudos, entrenudos ni primordios foliares u hojas juveniles. Cualquiera sea el tipo de raíz, embrionales o adventicias, una raíz con crecimiento primario, presenta las siguientes zonas. Por favor, téngase en cuenta que se debe estudiar en sentido inverso, ya que el crecimiento se inicia en el ápice de la raíz y se extiende hacia la superficie de la tierra donde a través del cuello se une al tallo. Cuello (lugar de unión de la raíz con el tallo) 2012 Zona de diferenciación o pilífera: a nivel de esta zona se visualizan los tejidos primarios adultos, que constituyen la estructura primaria de la raíz. En la superficie y originados en la rizodermis (recordar meristemoides) se pueden observar los pelos radiculares o pelos absorbentes. El pelo absorbente se caracteriza por ser unicelular, con pared celular delgada y el núcleo generalmente se desplaza hacia el ápice adoptando –a la madurez del peloposición subapical. Estos pelos generalmente son de corta vida y a medida que van muriendo se forman otros. La función de los pelos de la raíz es aumentar la superficie de absorción de agua con sales minerales. Zona de alargamiento: las células derivadas (meristemas derivados) sufren alargamiento, ocurre un cierto número de divisiones y comienzan a diferenciarse los tejidos primarios. Esta zona es intermedia entre la zona meristemática propiamente dicha y la zona de tejidos diferenciados. Zona meristemática: comprende el meristema o cono vegetativo de la raíz que de acuerdo a la teoría de los histógenos está compuesto por zonas de células generadoras: dermatógeno (d), periblema (p) y pleroma (pl) que poseen la propiedad de dividirse por mitosis y producen los meristemas derivados: protodermis, meristema fundamental y procambium respectivamente. Las células de esta región se dividen activamente para dar origen a los tejidos primarios, excepto en un sector denominado centro de quiescencia o centro de reposo. La función es producir crecimiento primario en longitud. Caliptra o cofia: se origina en el dermatógeno en las Dicotiledóneas o en el caliptrógeno en Monocotiledóneas. Sus células son ricas en almidón y con abundantes dictiosomas, cuyas vesículas transportan mucílagos que facilitan el desplazamiento de la raíz en el suelo. La función de esta zona es proteger la zona meristemática. 2012 La caliptra es un tejido protector del cono vegetativo, está formado por células vivas que continuamente se renuevan. Imagen del cono vegetativo y caliptra. Detalle de células con mucílagos en la periferia de la caliptra. Escala: 100 µm. Cono vegetativo de la raíz de Lotus sp. (Fabaceae). Mitosis en las células meristemáticas del cono vegetativo que darán los meristemas derivados y luego los tejidos primarios a nivel de la zona pilífera. 2012 ESTRUCTURA PRIMARIA DE LA RAÍZ En un corte transversal (CT) de la raíz a nivel de la zona pilífera, vemos la estructura primaria. En la misma, podemos distinguir desde la periferia hacia el centro: la rizodermis, la corteza primaria (exodermis, parénquima cortical y endodermis) y el cilindro vascular (periciclo, xilema primario, floema primario y médula). Ver el Texto de Tejidos de protección: rizodermis, exodermis y endodermis, en este sitio RIZODERMIS: como se ha expresado es generalmente una capa de células alargadas, continua, con paredes muy delgadas y protoplasto, totalmente adaptadas a su función de absorción, por lo tanto carente de cutícula y estomas. En ocasiones como se ha indicado puede ser pluristrata, en el velamen de las orquídeas, su función es la absorción y acumulación de agua. La rizodermis, para aumentar la superficie de absorción, en la zona de diferenciación de los tejidos primarios presenta los pelos absorbentes. Estos están caracterizados por su pared muy delgada y el desplazamiento del núcleo hacia el ápice. Los pelos generalmente son de corta vida pero continuamente son reemplazados por otros. En algunos casos son persistentes y también los hay pluricelulares. A continuación se puede visualizar una rizodermis con pelos absorbentes rotos ya que son muy delicados en su estructura y sobre la derecha el detalle de un pelo absorbente unicelular con pared celular muy delgada 2012 Imagen de rizodermis y pelos absorbentes Detalle de pelo absorbente de Iris sp. Escala: 50 µm. CORTEZA PRIMARIA Exodermis: tejido de protección subepidérmico formado por una o varias capas de células del parénquima cortical, más o menos hexagonales en el corte transversal, con una laminilla de suberina en sus paredes que luego es cubierta por capas de celulosa, a veces, la pared alcanza considerable espesor y se lignifica (por ejemplo muchas raíces de gramíneas). Su función es protección de la raíz evitando la pérdida de agua. Con el tiempo esta capa o capas reemplazan la rizodermis. La imagen del corte transversal de la raíz de “maíz” muestra una exodermis incipiente, con paredes aún poco engrosadas. Escala: 50 micrómetros. Parénquima cortical: frecuentemente las células poseen una disposición radial, y se van formando capas hacia el interior, es decir que la endodermis que es la capa más interna sería la más joven. Las células parenquimáticas poseen paredes delgadas, normalmente son incoloras, ya que es un parénquima tipicamente de reserva. Sin embargo, como veremos más adelante presenta muchas variaciones relacionadas al hábitat y a las necesidades de la planta. Si la raíz está expuesta a la luz forma cloroplastos (raíces de epífitas o de plantas acuáticas), ocasionalmente parte del parénquima puede esclerificarse ya sea en sus capas externas o en sus capas más internas junto a la endodermis (esto ocurre en monocotiledóneas, nunca en dicotiledóneas o en gimnosperma donde la endodermis se desprende con el crecimiento secundario). Las 2012 células del parénquima cortical pueden también presentar idioblastos cristalíferos o taníferos y estructuras secretoras como tubos laticíferos, conductos lisígenos o esquizógenos y/o cavidades secretoras. A veces, el parénquima cortical es aerenquimático, por ejemplo en plantas hidrófitas y también en gramínas de ambientes relativamente secos. La presencia de cavidades o lagunas aerenquimáticas es propio de las Panicoideae, mientras que no están presentes en las Poaeoideae (= Festucoideae) (Ver http://www.biologia.edu.ar/botanica/tema20/index20.htm) En las Gramíneas varias capas tanto del parénquima periférico como el interno próximo a la endodermis puede estar esclerificado (Ver texto que trata tejidos de elaboración y reserva en este sitio). Imagen del corte transversal (CT) de la raíz de Iris sp., parénquima cortical con pequeños espacios intercelulares. Escala: 200 µm. Endodermis: tejido de protección interno. Generalmente este tejido está formado por una capa de células que pasan por tres estadíos de engrosamiento de sus paredes. Durante el primer estadío (endodermis primaria) se produce la incrustación de la pared celular con sustancias fenólicas y grasas en forma de cinta sobre las caras radiales y transversales, formando la llamada ‘banda de Caspary’. En el corte transversal se observa como ‘puntos de Caspary’. 2012 Corte transversal de la raíz de “vara de oro” Solidago chilensis Meyen (Asteraceae) mostrando las células de la capa de endodermis con banda de Caspary (flecha blanca) y los puntos de Caspary (flechas negras). Escala: 50 µm. La banda de Caspary se forma en todas las raíces, tiene gran importancia fisiológica, por ejemplo en la circulación de la savia bruta. La circulación de la savia puede ser aploplástica (por las paredes celulares) y simplástica (por el citoplasma). La savia bruta penetra en los pelos absorbentes y por la corteza puede circular de ambas maneras (apoplástica y simplástica). Al llegar a la endodermis la savia bruta (agua con sales minerales) es rechazada por las sustancias hidrófobas de la banda de Caspary y es obligada a la circulación simplástica. Para ello debe atravesar la plasmalema que ejerce su acción selectiva. De esa manera la planta controla los iones que pasan y los iones que no pasan (Mauseth, 1988; Valla, 2004), para alcanzar luego las tráqueas y ser trasladada hacia los órganos verdes. Otro aspecto interesante que relata (Esau, 1982) es la unión que se produce entre la plasmalema y la banda de Caspary. Esto hace que cuando se produce deshidratación de la planta por ende de los tejidos y células, se demore la contracción del protoplasma celular, ya que la plasmalema no se desprende de la banda de Caspary. Es decir, retarda el paso hacia la plasmólisis celular. Plasmólisis celular: En la célula viva es la contracción del protoplasma como consecuencia de la pérdida de agua, se pone de manifiesto por la pérdida de turgencia celular y marchitamiento de la planta. La plasmólisis es reversible si se facilita la absorción de agua. El segundo estadío (endodermis secundaria) consiste en el depósito de una lámina de suberina que cubre internamente todas las paredes de la célula. El tercer estadío de engrosamiento (endodermis terciaria) consiste en el depósito de capas de celulosa sobre la suberina. Este depósito ocurre principalmente en las paredes radiales y tangencial interna de cada célula. Esta gruesa pared considerada secundaria 2012 puede tener puntuaciones y frecuentemente se lignifica. A este estadío se le suele denominar ‘cuadro de Caspary’. ESTA IMAGEN GIRARLA PARA QUE COINCIDA CON LA SIGUIENTE MOSTRANDDO LA U ABIERTA HACIA LA PERIFERIA Corte transversal de raíz de Iris sp. mostrando el tercer engrosamiento de las células de la endodermis. Escala 200 µm. Tinción con safranina Algunas células, principalmente ubicadas frente a los cordones de xilema primario no sufren este último engrosamiento, se llaman “células de paso”. Corte transversal de raíz de Iris sp. mostrando las células del parénquima cortical con sus paredes delgadas. La capa de células de la endodermis con el tercer engrosamiento en las paredes tangencial interna y radiales (observe que la cara tangencial externa permanece delgada). También se ve una célula de paso, sin dicho engrosamiento, ubicada frente al cordón de xilema. Escala: 100 µm. 2012 El tercer estadío de engrosamiento de la endodermis solamente ocurre en las raíces de las Angiospermas-Monocotiledóneas, que solamente tienen crecimiento primario y la endodermis permanece formando parte de la raíz. Mientras que en las AngiospermasDicotiledóneas y en las Gimnospermas al producirse la formación del felógeno en el periciclo y desarrollar la peridermis elimina la endodermis y restantes tejidos corticales primarios, o cuando la peridermis no se forman tan profunda la endodermis es aplastada. CILINDRO CENTRAL O CILINDRO VASCULAR Periciclo: comprende generalmente una capa de células de tipo parenquimático con propiedades meristemáticas que dan origen a las raíces laterales, a yemas adventicias y en raíces con crecimiento secundario a parte de los meristemas laterales (cambium vascular y cambium suberógeno). Generalmente la capa de periciclo forma un anillo continuo, pero puede ser discontinuo cuando a ese nivel se forma el xilema o el floema. El periciclo puede ser pluristrato, por ejemplo en Agave, Celtis, Morus y Gimnospermas. En plantas acuáticas o parásitas puede faltar y en raíces de monocotiledóneas puede engrosar sus paredes y lignificarse. La siguiente imagen corresponde al corte transversal de raíz de Iris sp. mostrando la capa de células del periciclo donde algunas células han comenzado a sufrir engrosamiento de sus paredes avanzando en un proceso de esclerificación. Se observan los cordones de xilema (Xi) que alternan con los cordones de floema (Fl). La tinción es con violeta de cresilo. Escala: 100 µm. 2012 2012 Xilema y floema primarios: en la imagen siguiente teñida con safranina de la misma raíz de Iris sp. se visualiza todo el cilindro central (Periciclo, xilema primario, floema primario y médula). Escala 200 µm. Los tejidos xilema (Xi) y floema (Fl) se encuentran formando cordones dispuestos en forma alterna y radiada. En la imagen se distinguen los vasos pequeños del protoxilema (hacia el periciclo) y los vasos grandes del metaxilema (hacia el centro de la raíz). Las paredes de los vasos son secundarias lignificadas por eso se han coloreado intensamente. Alternando con el xilema se ven los cordones de floema (Fl) cuyas células poseen pared primaria, se observan delgadas y blancas ya que está formado por tubos cribosos y células anexas, no contiene fibras. El protofloema también se ubica hacia el periciclo y el metafloema hacia el centro de la raíz. En el centro de la raíz las células del parénquima medular han engrosado sus paredes y se impregnaron con lignina, formando una médula lignificada o esclerificada. Los cordones de floema se ubican próximos al periciclo y no se extienden demasiado hacia el centro de la raíz mientras que los cordones de xilema frecuentemente se extienden hasta el centro de la raíz. También recordemos que el protoxilema está formado por vasos de menor diámetro y con engrosamientos secundarios parciales de sus paredes (anillados y espiralados) y en el metaxilema predominan los vasos de mayor diámetro con mayor engrosamiento secundario de sus paredes (reticulados, escalariformes y punteados). Médula: formada por parénquima, a veces sus células sufren engrosamiento de la pared como se observa en la imagen previa, otras veces sufren lisis y forman una raíz fistulosa (hueca); en algunas especies es aerenquimática, en otras puede ser reemplazada por uno o más vasos del metaxilema. 2012 En Monocotiledóneas es frecuente que el centro de la raíz este ocupado por una amplia médula parenquimática o esclerificada, por ejemplo en el “maíz”. Estela. Es el nombre dado a la columna formada por el cilindro central (periciclo, tejidos de conducción y médula), tanto de la raíz como del tallo. Las raíces pueden tener una protostela sin médula en el centro o una sifonostela con médula en el centro. En ambos casos se forma una actinostela por la disposición estrellada de los cordones xilemáticos. Así la estructura primaria de la raíz alterna y radiada de acuerdo a la teoría estelar es una actinostela. De acuerdo al número de cordones o polos de xilema hay diferentes tipos de raíz. Si hay un cordón es monarca, por ejemplo, Trapa natans L. (Lythraceae), dos cordones de xilema la raíz es diarca (lino, zanahoria). Si hay tres cordones de xilema es triarca (arveja); cuando son cuatro se llama tetrarca (ranúnculo, vicia), cuando son cinco es pentarca (trigo) y cuando son muchos se denomina poliarca (maíz). 2012 En las Gimnospermas son diarcas o poliarcas. En las Angiospermas son: en Dicotiledóneas diarcas a tetrarcas. en Monocotiledóneas pentarcas o poliarcas. Raíz DIARCA En los siguientes esquemas se muestran diferentes maneras que se usarán en la representación del xilema primario y floema primario. Esquema 1. se indica con palabras la ubicación del protoxilema y protofloema hacia la periferia y el metaxilema y metafloema hacia el centro de la raíz. Esquema 2. El floema se ha indicado con punteado (símbolo de Metcalfe & Chalk, 1950) para representar este tejido. El xilema se ha representado cada vaso por un círculo. Observar que los círculos de menor diámetro, que corresponden al protoxilema están ubicados hacia la periferia, mientras que los de mayor diámetro (metaxilema) se encuentran hacia el centro de la raíz. Esquema 3. Para representar ambos tejidos se usaron los Símbolos de Metcalfe & Chalk (1950). Punteado para el floema, Rayado para el xilema. Blanco para el parénquima, vemos entonces que en el centro hay abundante parénquima medular. ¿Cómo determinamos si una raíz es diarca, triarca, tetrarca, pentarca, hexarca o poliarca? Contamos los cordones de xilema -normalmente es bien visible la ubicación del protoxilema- y vemos que los siguientes esquemas corresponden a raíces TETRARCA PENTARCA Y POLIARCA En la raíz Pentarca, un vaso o tráquea reemplazó la médula. 2012 Formación de las raíces secundarias o laterales Las raíces secundarias se forman en el periciclo. De acuerdo al número de cordones de xilema y floema primario lo hacen en diferentes lugares (Esau, 1976) Si la raíz es diarca se forma entre los cordones de xilema y floema. En las raíces tri y tetrarca lo hace frente al cordón de xilema. En monocotiledóneas donde generalmente las raíces son poliarcas las raíces laterales se forman frente a los cordones de floema. Ver los siguientes esquemas. La estructura de la raíz lateral es igual a la de la raíz principal. Imagen de raíz triarca mostrando la salida de una ramificación o raíz secundaria frente al cordón de xilema. Escala: 500 micrómetros la vista general. Vista detallada del cilindro central y ramificación de la misma raíz. Escala: 200 micrómetros. 2012 Formación de las raíces adventicias Las raíces adventicias se originan en cualquier parte del vegetal menos de la radícula del embrión. Las raíces adventicias pueden tener origen endógeno o exógeno. Endógeno: en el periciclo, en el cambium, en el parénquima interfascicular en los tallos herbáceos, jóvenes o en el parénquima de los radios medulares en los tallos leñosos. Exógeno: en la epidermis y capas subepidérmicas, por ejemplo en el borde foliar. 2012 Esquema de estructura primaria completa. Endodermis con banda de Caspary Imagen de la estructura primaria completa de la raíz de maíz. Exodermis multistrata lignificada, Parénquima cortical aerenquimático. Endodermis con cuadro de Caspary. Estructura poliarca Las raíces dentro de su estructura general, presentan variaciones como formas de adaptación a diferentes ambientes, formas de nutrición, etc. Velamen de las orquídeas: se encuentra en las raíces expuestas de orquídeas epífitas o en orquídeas terrestres. Macroscopicamente estas raíces se observan blancas. El velamen está formado por una epidermis de la raíz o rizodermis pluristrata, cuyas células son muertas con engrosamientos en sus paredes. Su función es acumular agua y cuando se deshidratan están llenas de aire. 2012 Corte transversal de la raíz de Oncidium sp. (Orchidaceae), mostrando el velamen. Escala: 200 µm. Algunos autores sostienen que su función es de protección mecánica, esta puede ser otra función además de impedir el exceso de pérdida de agua. Raíz de epífita. Tillandsia sp., (Bromeliaceae) con características xeromórficas. Rizodermis seguida de varias capas parenquimáticas y un grueso anillo interno de esclerénquima hasta la endodermis con paredes engrosadas. En el cilindro central todo el parénquima esclerificado, en especial la médula. 2012 Las raíces mesomórficas serían las que corresponden a la estructura más general, ya que se forman en ambientes de condiciones moderadas de temperatura con una disponibilidad constante de humedad. La estructura presenta abundante parénquima cortical. La endodermis presenta una banda de Caspary poco engrosada. Las raíces xeromórficas, presentan un parénquima cortical reducido y una endodermis con banda de Caspary con notable engrosamiento. En las monocotiledóneas una exodermis con paredes incluso lignificadas como se puede ver en la raíz de “formio” (Phormiun tenax J.R. Forst. & G. Forst. (Xanthorrhoeaceae) Imagen de la raíz de “formio” en corte transversal mostrando el engrosamiento de las paredes de las células de la exodermis. Escala: 100 micrómetros. Imagen de la endodermis con cuadro de Caspary y el cilindro central con el parénquima totalmente lignificado de la raíz de “formio”. Escala 100 micrómetros. 2012 En las Poaceae suele haber varias capas de exodermis lignificada y un parénquima cortical con amplias cavidades aerenquimáticas que pueden ser esquizogénicas o lisigénicas. Si son esquizogénicas presentan un aspecto regular (posiblemente las de las que vemos en la raíz de maíz). Cuando estas cavidades se forman por descomposición más o menos extensa de las células parenquimáticas (lisigénesis) presentan membranas rotas, son irregulares en forma y distribución. Las lagunas corticales también son frecuentes en las raíces de otras monocotiledónes como las Cyperaceae, Palmeras, etc. Imagen del corte transversal de la raíz de “maíz” Zea mays L. (Poaceae) donde se observan las lagunas corticales. Escala: 200 µm. 2012 Imagen del corte transversal de la raíz de “grama” Chloris sp. (Poaceae), mostrando las grandes lagunas en el parénquima cortical. También se observa el engrosamiento sufrido por la exodermis y las capas del parénquima de próximas a la endodermis. Escala: 100 µm. Las raíces hidromórficas de plantas de hábitat acuático y palustres frecuentemente presentan parénquima cortical aerenquimático. Sin embargo, también se presenta con frecuencia en Poaceae de hábitats relativamente secos (como se ha mostrado para maíz y grama). Corte transversal de raíz de Sagittaria montevidensis Cham. & Schltdl. (Alismataceae) mostrando la corteza aerenquimática. Escala 500 micrómetros. 2012 Neumatóforos: son raíces con geotropismo negativo dotadas de un notable aerénquima que rodea al mínimo cilindro central, actuando como oxigenadores en plantas que viven en suelos pantanosos. Por ejemplo: imágenes de neumatóforos de Ludwigia peploides (Kunth) P.H.Raven (Onagraceae), donde al pequeño cilindro central lo rodea una red de células alargadas que dejan amplias cámaras aerenquimáticas. Escalas: 500 µm, se observa oscuro el mínimo cilindro central; 200 µm, se observa la red parenquimática de células especializadas. 2012 Parénquima secretor en las raíces. Un ejemplo de tejidos secretores en las raíces lo hallamos en las Gimnospermas, por ejemplo los conductos resiníferos de los pinos. En las Angiospermas también pueden hallarse tejidos secretores en la raíz, como se observa en la siguiente imagen con una estructura secretora en el parénquima cortical de la raíz de “vara de oro”. Corte transversal de raíz de “vara de oro” Solidago chilensis Meyen (Asteraceae). Escala: 50 micrómetros. 2012 BIBLIOGRAFÍA Cosa, M.T. & N. Dottori. 2010. Adaptaciones anatómicas de plantas medicinales a la diversidad de ambientes. X Simposio Argentino y XIII Simposio Latinoamericano de Farmacobotánica. Dimitri, M.J. & E.N. Orfila. 1985. Tratado de Morfología y Sistemática Vegetal. Ed. Acmé Esau, K. 1976. Anatomía Vegetal. Ed. Omega Esau, K. 1982. Anatomía de las plantas con semilla. Ed. Hemisferio Sur Fahn, A. 1985. Anatomía Vegetal. Ed. Pirámide Font Quer, P. 1965. Diccionario de Botánica. Ed. Labor Mauseth, J. 1988. Plant anatomy. Benjamin/Cummings Strasburger, E. et al. 1994. Tratado de Botánica. Ed. Omega Valla, J.J. 2004. Botánica. Morfología de las plantas superiores. Ed. Hemisferio Sur. J. Raisman, AM González y colab. Hipertextos del Área de Biología Vegetal. Universidad Nacional de Nordeste. http://www.hiperbotánica.net/ 2012