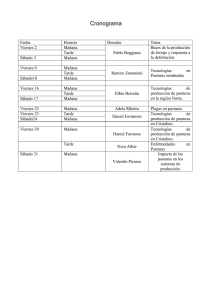
dinámica poblacional del campo natural y mejoramientos
Anuncio

1
DINÁMICA POBLACIONAL DEL CAMPO NATURAL Y
MEJORAMIENTOS EXTENSIVOS
Ing.Agr.Daniel Formoso1
Introducción
El campo natural constituye el 71% del territorio nacional (MGAP, 2000)
siendo la base alimentaria de las dos principales especies de herbívoros
domésticos (ovinos y bovinos). Ambas especies comparten distintos sitios de
pastoreo, modificando la estructura de la vegetación (Altesor et al., 2005;
Olmos et al., 2005), según la densidad y relación utilizadas (Ayala y Bermúdez,
2005; Berretta, 2005.; Boggiano et al., 2005; Formoso, 2005). Estas
modificaciones estructurales se manifiestan a través de cambios florísticos con
diferente grado de intensidad según el tipo de suelo (Formoso y Gaggero,
1990; Berretta et al., 1990; Formoso y Colucci, 2003; Rodríguez et al., 2003).
Sin embargo, Baeza et al. (2006) plantean que la respuesta al disturbio
causado por el pastoreo es expresada de manera más rápida por los cambios
funcionales (producción primaria neta aérea, reciclado de nutrientes) y no tanto
por los cambios florísticos. Por esa razón, estos autores realizaron una
caracterización funcional de la vegetación basada en el Índice Verde
Normalizado (IVN), identificando seis tipos funcionales de ecosistemas. Entre
los atributos seleccionados se encontraba el mes de mayor IVN, siendo el mes
de abril para el Basalto y el mes de noviembre para el Cristalino central, por
ejemplo. Estos atributos responden a un determinado sustrato geomorfológico,
que sostiene diversas comunidades integradas por poblaciones específicas.
Por consiguiente, la investigación puede plantearse sobre una
determinada jerarquía ecológica (ecosistema ← comunidad ← población),
siendo las poblaciones y su dinámica el motivo del presente trabajo cuyo
objetivo principal es proporcionar una introducción al tema.
Las poblaciones, principios, propiedades y aplicaciones.
Una población se define como “..un grupo de individuos de la misma
especie que vive en un área de suficiente espacio para permitir una dispersión
y/o un comportamiento migratorio normal en donde los cambios numéricos son
determinados en gran medida por los procesos de nacimientos y muertes”
(Berryman, 2002). Este autor destaca que el concepto de población es para los
ecólogos lo que el concepto de especie es para los taxónomos y sistemáticos.
La dinámica de una población puede ser afectada por dos tipos de
procesos: exógenos y endógenos (Uzal y Nores, 2004). Un proceso exógeno
puede causar cambios en una determinada población, pero no es afectado por
dichos cambios. La lluvia, la temperatura, el tipo de suelo, la topografía; son
1
Técnico del Secretariado Uruguayo de Lana (Área Investigación & Desarrollo)
2
procesos exógenos que pueden alterar la relación de las variables que operan
en la dinámica de una población y de este modo alterar las propiedades del
sistema.
Los procesos endógenos en cambio, son afectados por los cambios que
inducen y por ello son considerados parte de la estructura del sistema. Estos
procesos se conocen como procesos de retroalimentación (positiva o negativa),
siendo de primer orden (o sistema de dimensión uno) cuando el efecto de
retroalimentación es de una variable consigo misma, o de segundo orden (o
sistema de dimensión dos) cuando el efecto de retroalimentación involucra dos
variables.
En general, las poblaciones están caracterizadas por dinámicas simples
de primer y segundo orden (Lima, 2001), algunas hasta de tercer orden, pero
ninguna de las estudiadas por un orden superior a tres (Berryman, 2003).
En un sistema dinámico, los procesos de retroalimentación negativa
regulan las variables, retornándolas rápidamente hacia su valor original,
manteniendo la población estabilizada. En general, estos procesos son de
primer orden. Cuando se producen retrasos en la acción de la retroalimentación
negativa, la estructura de la población se encuentra afectada por más de una
variable, tornándose inestable y manifestando ciclos. A su vez, la
retroalimentación positiva es un proceso desestabilizador de la dinámica
poblacional (Berryman et al., 1987), siendo la población humana uno de los
ejemplos más característicos por su continuo crecimiento.
Las poblaciones están regidas por cinco principios (Berryman, 2003),
entendiéndose por principio “una ley científica general o fundamental, de la cual
se derivan otras leyes” (Heamig, 2006). Estos principios son los siguientes: 1.
El crecimiento geométrico (exponencial), denominado también Ley de Malthus;
2. Cooperación (denominado también efecto Allee); 3. Competencia (o principio
de Verhulst); 4. Especies interactuantes; y el 5. Factores limitantes.
A título de ejemplo, se realiza una breve presentación del primero y tercer
principio, por considerarlos apropiados para esta introducción.
El primer principio, o de crecimiento exponencial, corresponde a la teoría
de Thomas Malthus, clérigo inglés del siglo XVIII. Según este principio, cuando
las tasas de natalidad o mortalidad de una población son constantes, la
población crecerá (o decrecerá) a una proporción exponencial. Malthus
pensaba que la población humana crecería exponencialmente, mientras que
los recursos (alimentos) lo harían aritméticamente.
El modelo que describe el crecimiento exponencial (ec.1) asume que los
recursos son ilimitados por lo que no existe competencia, que cada organismo
tiene iguales posibilidades de crecimiento o muerte independientemente de la
edad y estado. Por estas razones también se conoce como modelo denso
independiente (Fig. 1):
3
N t = N 0 e rt
ec. 1
Nt: Cantidad de individuos en el tiempo t
N0: cantidad inicial de individuos
r: tasa instantánea de incremento por capita (nacimientos – muertes)
t: unidad de tiempo (en general un año)
e: base de los logaritmos naturales
Figura
1.
Representación
gráfica
del
crecimiento
exponencial (denso independiente) de una población teórica
En contraposición al crecimiento exponencial, se asume que en algún
momento la población alcanza un límite máximo porque los recursos son
finitos. Una vez alcanzado este límite, los individuos comenzarán a competir
entre sí, reduciéndose la tasa de crecimiento per cápita como consecuencia del
incremento en la densidad de la población. Esta propuesta fue desarrollada por
Pierre Verhulst, matemático belga, quien en 1839 introdujo el concepto de
“auto-limitación poblacional”. Este modelo de crecimiento también se conoce
como denso dependiente (Fig. 2).
La ecuación (ec.2) que describe al modelo supone que a medida que la
densidad de la población alcanza un determinado límite, el crecimiento se
desacelera tendiendo a cero (nacimientos = muertes).
N t = N 0 e {Rt }
ec.2
4
R: representa la tasa de incremento per capita, que puede ser calculada a partir
de los valores observados, o mediante métodos estadísticos (p.ej. regresión
lineal). Es una función del tamaño poblacional.
t: unidad de tiempo (en general un año)
e: base de los logaritmos naturales
Figura 2. Representación gráfica del crecimiento logístico
(denso dependiente) de una población teórica.
La importancia del cálculo de la función R por métodos estadísticos
reside en la introducción de una serie de parámetros con significado biológico
que permiten analizar la estructura de las poblaciones (Berryman y Turchin,
2001; Münster-Swendsen y Berryman, 2005) . De esta manera, R adquiere la
siguiente expresión (Berryman y Millstein, 1994)(ec.3):
⎡ ⎛ N(t-1) ⎞Q ⎤
⎟⎟ ⎥
R = A x ⎢1- ⎜⎜
K
⎢⎣ ⎝
⎠ ⎥⎦
ec.3
A: representa la máxima tasa de incremento per capita cuando la población se
encuentra a muy bajas densidades [desde (t-1) a t].
Q: es el coeficiente de curvatura que puede producirse si las relaciones denso
dependientes no son lineales. Si Q=1, la relación es lineal, Q>1, la relación es
convexa, mientras que si Q<1, la relación es cóncava.
K: es denominado la capacidad de carga del sistema, o la cantidad total de
recursos dentro del sistema dividido por los requerimientos mínimos de cada
individuo que lo integran. La capacidad de carga puede ser modificada por el
manejador del sistema mediante el agregado de nutrientes, cambios en el
espacio disponible para los individuos o reacomodo de otras poblaciones.
5
Una vez que el crecimiento de una población alcanza su capacidad de carga, la
población debería mantenerse estable y en equilibrio. Esta situación supone la
ausencia de otras poblaciones de variaciones ambientales. Sin embargo, en las
condiciones naturales, las poblaciones se encuentran relacionadas con otras
poblaciones, la capacidad de carga no es estática, hay factores ambientales
que inciden en la dinámica poblacional. Por lo tanto, la población comienza a
fluctuar/oscilar (Münster-Swendsen y Berryman, 2005) (Fig.3).
Figura 3. Fluctuaciones de una población teórica entorno a
su capacidad de carga.
Cuando estas fluctuaciones se registran cuantitativamente y ordenadas
en el tiempo, se dispone de una serie cronológica o temporal (Fig. 4). El
objetivo de analizar este tipo de información, es el conocimiento de su patrón
de comportamiento, lo que permite prever su futuro.
6
Figura 4. Serie temporal de la densidad (kg de biomasa
disponible ha-1) de la población de Lotus Rincón estimada
en invierno sobre un mejoramiento de campo en Cerro
Colorado (dpto. de Florida)
Las series temporales tienen la particularidad de sus datos o sea, son muchas
muestras de tamaño unidad (kg, número, densidad) procedentes de diversas
variables con distribución y características desconocidas, por lo que su análisis
requiere una estadística apropiada. En el estudio de las poblaciones, el punto
destacable es encontrar la estructura del sistema y el signo y orden de los
procesos de retroalimentación (Lima, 2001), ajustando un modelo a la función
R para relacionar a la población en estudio con otros componentes del
ecosistema.
En el país, y específicamente en el tema de pasturas, las series
cronológicas publicadas no son tan extensas (Berretta y Bemhaja, 1998) como
para ser analizadas con técnicas robustas que requieren al menos información
superior a 15 años. Esto no elimina la posibilidad de acceder a su estructura
con procesos más sencillos, pero de gran utilidad para complementar el
conocimiento agronómico que se posee y fortalecer las estrategias de manejo
con la finalidad de mantener o aumentar la productividad del recurso sin
deteriorarlo.
Referencias
ALTESOR A., PIÑEIRO G., LEZAMA F., RODRÍGUEZ C., LEONI E., BAEZA
S., PARUELO J.M. 2005. El efecto del pastoreo sobre la estructura y el
funcionamiento de las praderas naturales uruguayas: ¿qué sabemos y cómo podemos
usar ese conocimiento para manejarlas mejor? Seminario de Actualización Técnica en
Manejo de Campo Natural. INIA, Serie Técnica 151:21-32.
AYALA W., BERMÚDEZ R. 2005. Estrategias de Manejo en Campos Naturales sobre
Suelos de Lomadas en la Región Este. INIA, Serie Técnica 151.:41-50.
BAEZA S., PARUELO J.M., ALTESOR A. 2006. Caracterización Funcional de la
Vegetación del Uruguay mediante el uso de Sensores Remotos. INTERCIENCIA.
31(5):382-388.
BERRETTA E.J. 2005. Producción y Manejo de la Defoliación en Campos Naturales de
Basalto. INIA, Serie Técnica 151.:61-73.
BERRETTA E.J., BEMHAJA M. 1998. Producción Estacional de Comunidades
Naturales sobre Suelos de Basalto de la Unidad Queguay Chico. INTA. Serie Técnica
102.:11-31.
BERRETTA E.J., LEVRATTO J.C., SAMIT W.S., BEMHAJA M., PITTALUGA
O., SILVA J.A., CLARIDGET J.B., GUERRA J.C. 1990. Efecto del Sistema
de Pastoreo, relación Lanar/Vacuno y Carga Animal sobre la Producción y Utilización
de Pasturas Naturales. I. Evolución de la Vegetación en Pastoreo Continuo y Rotativo a
7
igual Dotación y Relación Lanar/Vacuno 2/1. In II Seminario Nacional de Campo
Natural. Tacuarembó.(Ed. Hemisferio SUR.):291-298.
BERRYMAN A. 2002. Population: a central concept for ecology? Oikos 97(3):439-442.
BERRYMAN A. 2003. On principles, laws and theory in population ecology. Oikos.
103(3):695-701.
BERRYMAN A., MILLSTEIN J.J. 1994. Population Analysis System. Version 4.0.
Ecological System Analysis. Pullman. Washington.
BERRYMAN A., STENSETH N. Chr., ISAEV A.S. 1987. Natural Regulation of
Herbivorous Forest Insect Populations. Oecologia (Berlin). 71:174-184.
BERRYMAN A., TURCHIN P. 2001. Identifying the Density Dependent Structure
Underlying Ecological Time Series. Oikos. 92(2):265-270.
BOGGIANO P., ZANONIANI R., MILLOT J.C. 2005. Respuestas del Campo Natural
a Manejos crecientes de Intervención. INIA, Serie Técnica 151.:105-113.
FORMOSO D. 2005. La Investigación en Utilización de Pasturas Naturales sobre Cristalino
desarrollada por el Secretariado Uruguayo de la Lana. INIA, Serie Técnica 151.:51-59.
FORMOSO D., COLUCCI P.E. 2003. Vegetation Changes in Native Plant Communities
in Basaltic Deep and Shallow Soils after 10 Years of Grazing Exclosure in Uruguay. In
International Rangelands Congress. Durban. South Africa.
FORMOSO D., GAGGERO C. 1990. Efecto del Sistema de Pastoreo y la relación
Ovino/Vacuno sobre la Producción de Forraje y la Vegetación del Campo Nativo. In II
Seminario Nacional de Campo Natural. Tacuarembó.(Ed.Hemisferio SUR.):299-310.
HEAMIG P.D. 2006. Leyes de la Ecología Poblacional. Ecología. Info. #23.
LIMA M. 2001. The Dynamics of Natural Populations: Feedback Structures in fluctuating
Environments. Revista Chilena de Historia Natural. 74(2).
MGAP. 2000. Censo General Agropecuario. Ministerio de Ganadería, Agricultura y Pesca.
Montevideo, Uruguay.:montevideo.mgap.gub.uy.
MÜNSTER-SWENDSEN M., BERRYMAN A. 2005. Detecting the Causes of
Population Cycles by Analysis of R-funtions: the Spruce Needle-Miner, Epinotia tedella,
and its Parasitoids in Danish Spruce Plantations. Oikos. 108(3):495-502.
OLMOS F., FRANCO J., SOSA M. 2005. Impacto en las Prácticas de Manejo en la
Productividad y Diversidad de Pasturas Naturales. INIA, Serie Técnica 151.:93-104.
RODRIGUEZ C., LEONI E., LEZAMA F., ALTESOR A. 2003. Temporal Trends in
Species Composition and Plant Traits in Natural Grasslands of Uruguay. Journal of
Vegetacion Science. 14:433-440.
UZAL A., NORES C. 2004. Endogenous and Exogenous constraints in the Population
Changes of Wild Boar (Sus scrofa Linnaeus, 1758). Galemys 16 (nº especial).83-98.
8
El presente trabajo es el complemento de la charla correspondiente a la
jornada InterCREA del 16 de marzo de 2007 en el departamento de Florida.