herencia ligada, influida y limitada al sexo
Anuncio

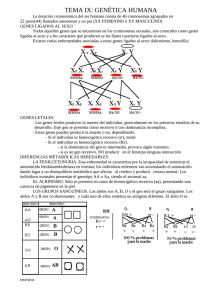
A- Determinación y diferenciación sexual Antes de comenzar a desarrollar los mecanismos de determinación y diferenciación sexual, será necesario conocer la diferencia entre ambos. Llamamos determinación del sexo al proceso por el cual se define el sexo de un organismo. La diferenciación sexual es, por su parte, el proceso por el cual el individuo manifiesta su fenotipo sexual, mediante la expresión de los caracteres sexuales primarios y secundarios. I. Determinación Genética del Sexo 1) Determinación Genética Críptica Críptico significa oculto, enmascarado, escondido. En este caso, esta palabra alude a que aún no son conocidos los mecanismos genéticos que conducen a la determinación del sexo en algunos organismos. Sí puede hablarse igualmente de determinación genética del sexo, porque el hecho de que un organismo reaccione siempre de la misma manera ante similares circunstancias, confirma la existencia de una información genética precisa que produce siempre la misma respuesta frente a un mismo estímulo. Un ejemplo de este tipo de determinación se presenta en el gusano marino Bonellia viridis. La hembra tiene un cuerpo redondeado, similar al de una ciruela pequeña, verde, con una trompa ciliada y larga, bifurcada en el extremo. El macho, que es diminuto, vive en el interior de la hembra como parásito, por poseer todos los órganos internos atrofiados, con excepción del reproductor; así el macho fecunda internamente los óvulos de la hembra, y cuando eclosionan los huevos, éstos se transforman en larvas natatorias. Al alcanzar cierto desarrollo, comienzan a caer en el fondo del mar; si en su caída las larvas se encuentran con la trompa ciliada de una hembra y se depositan sobre ella, se transforman en machos; si llegan al fondo, se transforman en hembra. Tras algunos estudios se llegó a demostrar que existe una sustancia no proteica en el interior de la trompa femenina que tiene poder masculinizante. Otro ejemplo de determinación genética críptica es el del caimán. Sus huevos tienen un período sensible a la temperatura entre los días 7 y 21 de incubación, de manera que nacerán hembras si la temperatura es inferior a 30ºC y machos si es superior a 40ºC. Este tipo de influencia de la temperatura en un momento específico durante la incubación sobre la determinación sexual, es transpolable a algunos otros reptiles y peces. 2) Determinación Sexual por Haplodiploidía Este tipo de determinación es característico de ciertas especies de la clase Insecta de los órdenes Himenópteros (abejas, avispas, hormigas), Homópteros (pulgones, cochinillas), Tisanópteros (trips) y Coleópteros (vaquitas, cascarudos, gusanos blancos o de suelo). En algunas especies de estos órdenes, la determinación del sexo no está relacionada con la presencia de cromosomas sexuales, sino que los machos son originados de los huevos sin fecundar, mientras que las hembras se originan de los huevos fecundados; por lo tanto los machos son haploides y las hembras diploides. En un principio, éste pareció ser el factor por el cual el sexo quedaba determinado. Sin embargo, la diploidía no determina el sexo femenino de por sí, sino que genera heterocigosis para un locus específico, de forma tal que al ser diploides (es decir al poseer dos juegos de cromosomas), en uno de ellos se presentan dos alelos diferentes de un serie alélica de nueve: esto determina su feminidad. Los machos por ser haploides, nunca poseen dos alelos diferentes para ese locus específico, sino tan solo uno: esto determina su masculinidad. 1 3) Determinación Sexual Cromosómica Como su nombre lo indica, en este tipo de determinación están implicados los cromosomas sexuales, sin embargo, no todos los genes que ambos poseen cumplen necesariamente la función sexual. Dentro del genomio, los cromosomas sexuales son dos homólogos especiales por presentar dimorfismo. Ambos provienen de un origen en común, pero luego de millones de generaciones, a través del proceso evolutivo, uno de ellos ha ido perdiendo cromatina y por lo tanto genes, quedando así con abundante heterocromatina. Este cromosoma es llamado Y, y en función de su forma es un cromosoma telocéntrico. Su homólogo en cambio, ha conservado sus genes, de manera que posee poca heterocromatina, casi en la misma proporción que un autosoma. Este cromosoma es llamado X y es acrocéntrico o submetacéntrico. En humanos se conocen 300 genes presentes en el cromosoma X y solo unos pocos en el cromosoma Y. • Clases de determinación sexual cromosómica En función de que un individuo presente dos cromosomas X o uno X y otro Y, las especies poseen estos diferentes sistemas de determinación cromosómica sexual: a) Sistema XX – XY El sexo femenino posee dos cromosomas sexuales X, es por ello que es llamado sexo homogamético: en todas las gametas poseerá entre el juego cromosómico, al cromosoma sexual X. El sexo masculino posee un cromosoma sexual X y un cromosoma sexual Y; es llamado entonces sexo heterogamético: la mitad de sus gametas poseerá entre sus cromosomas al X, y la otra mitad de sus gametas poseerá al cromosoma Y: X El 100% de las gametas posee el cromosoma X. XX X Sexo homogamético (sólo una clase de gametas). Un 50% de las gametas posee el cromosoma X. El otro 50% posee el cromosoma Y. Sexo heterogamético (dos clases de gametas). X XY Y Figura nº1: Distribución de los cromosomas sexuales en gametas. En cada generación se espera una proporción sexual de 1:1 en la descendencia. En el siguiente tablero de Punnet, se podrá determinar esto. Siempre hemos utilizado alelos en sus entradas. Ahora lo utilizaremos análogamente con los cromosomas XX y XY. 2 X Y X XX XY X XX XY 50% 50% Figura nº2: Probabilidad sexual de una descendencia. Este sistema de determinación sexual se presenta en Nemátodos, Moluscos, Equinodermos, algunos Artrópodos, Mamíferos y diversos géneros de ranas de los Anfibios. b) Sistema ZW - ZZ En algunas especies, el sexo heterogamético es el femenino, y el homogamético es el masculino. Por convención, para este tipo de determinación donde la constitución cromosómica sexual es XY en hembras y XX en machos, el cromosoma X es llamado Z, y el Y es llamado W, de manera tal que los machos poseen los cromosomas sexuales ZZ y las hembras, ZW. Estas especies pertenecen a algunos de los géneros de Aves, Reptiles, sapos de los Anfibios e Insectos (en especial Lepidópteros). En cuanto a las proporciones de las gametas y proporción sexual aparecida en la descendencia, las apreciaciones son las mismas que para el sistema XX – XY. c) Sistema XX - XO En algunas especies en donde el sexo heterogamético es el masculino, el cromosoma Y puede estar ausente. En ese caso, la connotación genética sexual es XO (la ausencia del cromosoma Y se simboliza con O). Así, los machos presentan un número impar de cromosomas, siendo su número 2n – 1. Este es el caso de algunas especies de los órdenes Hemípteros, y de los Ortópteros (langostas y tucuras) de la clase Insecta. d) Sistema ZO - ZZ En este sistema el sexo heterogamético es el femenino, de manera tal que son en las hembras en las que se halla ausente uno de los homólogos del par de cromosomas sexuales. Por eso se dice que su constitución cromosómica es ZO, y son ellas ahora las que poseen en sus células un número impar de cromosomas. • Inactivación del cromosoma X en mamíferos, compensación de dosis o teoría de Luon Debido a que el hombre y otros individuos de sexo masculino mamíferos poseen en su constitución cromosómica un solo cromosoma X, y que la hembra posee dos, es necesaria la existencia de un mecanismo que compense esta desigualdad, de manera que las dosis de los genes presentes en el cromosoma X sean iguales tanto en machos como en hembras. oEs este el motivo por el cual se inactiva uno de los cromosomas X en las células somáticas de las hembras. Este proceso ocurre en las primeras etapas de la división celular mitótica de los embriones, aproximadamente cuando la mórula presenta cerca de las 16 células. Esta inactivación consiste en una fuerte condensación producida de uno de los cromosomas X, de 3 modo tal que en el estado de interfase, se observa en el núcleo de las células somáticas de las hembras, un corpúsculo denominado cuerpo de Barr. Este corpúsculo corresponde así, al mencionado cromosoma X fuertemente replegado, con sus extremos unidos formando un anillo, convertido en heterocromatina. De esta forma sus genes no son transcriptos y queda solo uno de los cromosomas X como funcional. No siempre se inactiva el mismo cromosoma en todas las células; suponiendo una hembra heterocigota para un carácter fenotípicamente visible, en algunas células puede inactivarse en cromosoma que porta el alelo recesivo, y en las otras, puede inactivarse el cromosoma que porta el alelo dominante de ese carácter. Teniendo en cuenta que luego de la inactivación se suceden divisiones mitóticas, y que las nuevas células permanecen posicionalmente cerca, se forman dos zonas diferentes: aquellas en donde se expresarán las células con fenotipo dominante, y aquellas en donde se expresarán las células con el fenotipo recesivo. Así pueden observarse “parches” de dos diferentes fenotipos. Este tipo de patrón descripto a consecuencia de la inactivación azarosa de uno u otro cromosoma, se denomina patrón en mosaico. Cromosoma X de un progenitor Cromosoma X del otro progenitor A1 A2 Cigoto Inactivación de un cromosoma A1 Inactivación del otro cromosoma Mitosis A2 A1 Mitosis A1 A2 Mitosis A2 Mitosis A1 A2 Mitosis Grupo de células que expresan el alelo A2 A1 A2 Mitosis A1 A2 Mitosis Grupo de células que expresan el alelo A1 Mosaico Adulto Figura nº: Inactivación del cromosoma X en mamíferos. Figura nº3: Inactivación del cromosoma X en mamíferos. Uno de los casos más conocidos es de las gatas heterocigotas de coloración calicó, que sobre el pelaje blanco (dado por un gen autosómico), aparecen parches anaranjados y negros. Estas gatas son producto del cruzamiento entre gatas portadoras de un alelo negro por cada cromosoma X, y machos portadores de un alelo anaranjado en su único cromosoma X. La 4 posibilidad de nacimiento de hembras es del 50%, y de que éstas sean calicó, es del 100%. XX XY X X Y XY XY X XX XX Inactivación del X X Figura Nº4: Inactivación de uno u otro cromosoma X para cada individuo hembra . El cromosoma X representa el portador del alelo anaranjado. El cromosoma X representa el portador del alelo negro. cromosoma X Cuerpo de Barr Inactivación del cromosoma X Cuerpo de Barr Figura nº5: Gata calicó. En seres humanos existe una enfermedad codificada por un gen recesivo d ligado al cromosoma X, llamada displasia ectodérmica anhidrótica Los varones portadores del alelo recesivo, carecen de glándulas sudoríparas. La mujer heterocigota, a causa de la inactivación azarosa uno de sus cromosomas X, es un mosaico de sectores D y d aparecidos en todo el cuerpo, tal como se muestra en la figura siguiente. Figura nº6: Mosaicismo somático de tres generaciones de mujeres heterocigotas para la displasia ecto dérmica anhidrótica. La ausencia de las glándulas sudoríparas se manifiesta por un color de epidermis más oscuro. 5 • Conformación de los cromosomas sexuales En los cromosomas sexuales pueden diferenciarse dos zonas: un segmento homólogo el cual se aparea normalmente en la profase meiótica o también denominado por esta razón segmento apareante, teniendo entonces la posibilidad de poder intercambiar material genético y un segmento no homólogo que no se aparea y es llamado segmento diferencial. Segmento Apareante: porción homóloga con genes pseudoauto sómicos. Segmento Diferencial: porción no homóloga con genes completamente ligados al sexo. - Cromosoma Y Detalle de la región apareante: no consta de todo el brazo corto, sino del extremo distal del mismo. Cromosoma X Figura nº 7: Esquema representativo de los cromosomas sexuales. II. Diferenciación sexual Hasta ahora hemos visto que la constitución cromosómica XX en mamíferos determina el sexo femenino, y que la constitución XY, determina el sexo masculino. Trataremos de responder ahora cuál es el mecanismo por el cual se produce diferenciación una vez determinado el sexo, es decir, por qué el hecho de que una determinada constitución cromosómica haga que un organismo se diferencie en forma femenina o masculina. Primero será necesario distinguir la diferenciación primaria o gonadal de la secundaria o extragonadal. Cuando hablamos de diferenciación primaria o gonadal, como su nombre lo indica, aludimos a la formación de gónadas: ovarios o testículos. La diferenciación secundaria o extragonadal, alude en cambio a la formación de útero, vagina, vestíbulo vaginal, clítoris y labios mayores y menores en hembras, y a la formación de epidídimo, conductos deferentes, vesículas seminales, conductos eyaculadores, próstata, escroto y pene en machos. El origen embrional de estos distintos constituyentes de los aparatos sexuales, se resume en la figura nº8. Todos estos constituyentes manifiestan los caracteres sexuales primarios en los mamíferos. Los caracteres sexuales secundarios se verán más adelante puesto que son gobernados por otro tipo de herencia. Si el proceso de desarrollo embrional es normal, la diferenciación gonadal estará en concordancia con la constitución cromosómica del individuo. Una vez diferenciadas las gónadas, transcurridas entre 6 y 7 semanas desde el momento de la concepción, éstas producen hormonas sexuales (estrógeno los ovarios y andrógeno los testículos), y son ellas las que a su vez darán origen a la diferenciación secundaria, que será congruente con el sexo gonadal. 6 Ovarios Gónada embrionaria indiferenciada Testículos Epidídimo Conductos deferentes Vesículas seminales Wolff (machos) Conductos de Trompas de Falopio Utero Vagina Müller (hembras) Machos Próstata Escroto Pene Hembras Vestíbulo vaginal Clítoris Labios menores y mayores Gernitalia externa Fifura nº8: Origen embrional de los diferentes constituyentes de los aparatos sexuales femenino y masculino de los mamíferos. • Diferenciación Primaria en Mamíferos El hecho de que la presencia del cromosoma Y en la constitución cromosómica de un individuo haga que éste se desarrolle como macho, pone de manifiesto que este cromosoma es portador de información genética que es decisiva en la diferenciación testicular. En efecto, el cromosoma Y posee una pequeña región en su brazo corto, llamada TDF (factor determinante de los testículos) que a su vez está situada en el segmento diferencial. Dentro de esta región, existe un gen que codifica para la formación de testículo, llamado SRY (región Y determiante del sexo). Este gen, hace que la gónada embrional indiferenciada, comience a diferenciarse en testículo, las etapas iniciales de tal proceso ocurre en las células que darán origen a las células de Sertoli (ver figura nº9). Gónadas Ovarios C. Germinales Testículos C. Somáticas C. C.de deSoporte Soporte (C Teca) (C. Teca) C. de Tejido Conectivo C. Germinales C. Somáticas C.C. dede Soporte Soporte (C.(C. dede Sertoli) Sertoli) C. de Tejido Conectivo Figura Nº9: Constitución de las gónadas. Las zonas sombreadas indican el sitio del inicio del proceso de diferenciación sexual Cuando la constitución cromosómica de un mamífero es en cambio XX, la gónada embrional se diferencia en ovario; el inicio de esta diferenciación ocurre en las células teca. 7 Como el cromosoma Y no está presente, no hay formación de testículo, puesto que tampoco está presente el gen SRY de la región TDF. Sin embargo, la formación del ovario también está codificada por genes. En este caso, es el gen Od (determinante del ovario) el responsable de que se produzca la diferenciación primaria en hembras. Se trata de un gen presente en el cromosoma X, por lo cual también se halla en los individuos de sexo masculino. Ocurre que en estos últimos no se expresa porque si está presente el gen SRY, éste actúa antes que el gen Od para permitir que los individuos XY se desa rrollen como machos. Una vez iniciado el proceso de diferenciación primaria masculina, la diferenciación primaria femenina queda inhibida. Tiempo Si está presente, actúa el gen SRY Las gónadas indiferenciadas se convierten en testículos. inhibición Actúa el gen Od (siempre presente) Las gónadas indiferenciadas se convierten en ovarios. SEXO MASCULINO SEXO FEMENINO Figura Nº10: Momentos de diferenciación de la diferenciación de gónadas. • Diferenciación Secundaria en Mamíferos Por otra parte los embriones de ambos sexos de los mamíferos tienen formados los conductos de Wolff y de Müller. Cuando un individuo posee en su constitución cromosómica los cromosomas XY, en el embrión, los conductos de Müller degeneran pasivamente, mientras que los de Wolff predominan para transformarse en epidídimo, conductos deferentes, vesículas seminales y conductos eyaculadores. Esta regresión de los conductos de Müller es inducida por una hormona peptídica antimülleriana sintetizada en las células de Sertoli. La síntesis de esta hormona está regulada por un gen llamado MIS (Müllerian Inhibiting Substance o sustancia inhibidora de los conductos de Wolff), y dicho gen a su vez está regulado por el gen SRY. El gen MIS es autosómico. Los conductos de Wolff, en cambio, se desarrollan por acción de la testosterona. Una vez producida la diferenciación primaria, es decir una vez que se desarrollan los testículos, éstos, específicamente sus células de Leydig, producen testosterona. Esta testosterona induce a un represor (que mientras las gónadas permanecen indiferenciadas se encuentra impidiendo que los conductos de Wolf se desarrollen) para que deje de impedir la transcripción de los genes controladores del desarrollo de las conductos de Wolff, produciéndose así la diferenciación sexual secundaria masculina. En cambio, en un individuo con constitución cromosómica XX, los conductos de Wolff son los que degeneran pasivamente, quedando los de Müller para transformarse en trompas de Falopio, útero y vagina. Este proceso al contrario de lo que ocurre con los individuos de sexo masculino, no necesita inductor. Como los individuos femeninos no poseen cromosoma Y, no hay gen SRY, no se forman los testículos y no hay secreción de testosterona, por lo que el represor de los genes responsables del desarrollo de los conductos de Wolff, al no recibir inducción, permanece reprimiéndolos. Podemos decir entonces, que el proceso de diferenciación sexual secundaria o extragonadal, están determinados por un sistema genético tal que el desarrollo masculino o femenino corresponden al estado inducido o no inducido respectivamente de un único sistema regulador, en el que actúa como inductor la testosterona que se produce como 8 consecuencia de la diferenciación primaria o gonadal, que a su vez depende de la presencia del gen SRY. Células de Sertoli XY gen SRY Testículo Hormona Antimülleriana Células de Leydig Testosterona Inductor ARN mensajero i+ Gen regulador Desarrollo de los Regresión de los Coductos de Wolff Conductos de Müller Gen estructural Diferenciación Sexual Secundaria Masculina Figura nº11: Control genético de la diferenciación sexual primaria y secundaria en la especie humana B- Caracteres relacionados con el sexo Los caracteres ligados al sexo, son caracteres gobernados por genes que se encuentran en el segmento diferencial de los cromosomas sexuales. Ese segmento diferencial, al no sufrir apareamiento, y por lo tanto no realizar crossing-over, posee genes que son heredados siempre con el cromosoma sexual, y por eso se dice que los caracteres que gobiernan están ligados al sexo. Los genes situados en los loci del segmento homólogo de los cromosomas sexuales no necesariamente se heredan con el cromosoma, no están ligados a él, puesto que pueden sufrir crossing-over e intercambiarse así con el alelo correspondiente de su homólogo. Por ello se dice que esos genes están incompletamente ligados al sexo, y que su herencia es pseudoautosómica, es decir que los genes de este segmento se comportan como si estuviesen en autosomas por el hecho ya mencionado de poder intercambiarse sus alelos debido al crossing-over que realizan con sus homólogos. a) Caracteres ligados al sexo del segmento diferencial del cromosoma Y En el hombre, los genes que gobiernan los caracteres ligados al sexo que se hallan en el sector diferencial del cromosoma Y, son llamados genes holándricos (holo: todo y andro: varón). Son heredados solamente de padres a hijos varones, ya que como hemos visto, éstos determinan su sexo ante la presencia del cromosoma Y. En el hombre, aparte del gen SRY, se ha demostrado la presencia de un gen que codifica para otro carácter como es el de la 9 presencia/ausencia de pelos en el borde de la oreja. Cuando este carácter se halla en estado dominante, el borde de la oreja aparece sin pelos, cuando se halla en estado recesivo, el borde de la oreja presenta pelos. Este fenotipo es frecuente en las poblaciones de la India, y es conocido con el nombre de hipertricosis. (figura nº12). Figura nº12: Borde de la oreja peludo. Existen otros genes que probablemente estarían en este segmento, pero no se puede afirmar con certeza. Estos serían genes que influyen en la estatura, en el crecimiento de los dientes, y en la velocidad de maduración. Cuando en cambio, es el sexo heterogamético es el femenino, los genes que gobiernan los caracteres ligados al sexo se hallan en el sector diferencial del cromosoma W, son llamados hológinos (holo: todo y gyne: mujer). Son heredados solamente de madres a hijas hembras, ya que éstas determinan su sexo ante la presencia del mencionado cromosoma W. b) Caracteres ligados al sexo del segmento diferencial del cromosoma X El primer caso que se ha encontrado de herencia ligada al cromosoma X, es el del gen que gobierna el color de ojos en la mosca Drosophila melanogaster. El color de ojos rojos es el fenotipo salvaje, alelo simbolizado como w+. A partir del cruzamiento de una hembra de ojos rojos con un macho de ojos blancos, se obtiene una descendencia 100% de ojos rojos. Y del cruzamiento de una hembra de ojos rojos y de un macho de ojos rojos, ambos F1, se obtiene que el 100% de las hembras es de color de ojos rojos, y que entre los machos, el 50% es de color de ojos rojos, y el 50% restante es de color de ojos blancos. Esto se demuestra en la figura siguiente: Parentales w+ w w+ Filial 1 w w+ x w+ x w w+ w+ w+ w w+ w+ w+ w+ w+ w+ w+ w w+ w w+ w w Figura nº13: Fenotipos y sexo respectivo de los individuos F1 y F2 a partir de cruzamientos en los que participa un gen ligado al sexo. Los esquemas representados son llamados cromosómico–génicos, porque representan los cromosomas portando sus genes respectivos. Los cromosomas X son representados con una línea simple, mientras que los cromosomas Y tienen además un segmento menor inclinado que alude al brazo más corto. 10 Como puede observarse, en la primer generación se cumple la Ley de Mendel. En la segunda generación, se produce una relación 3:1, pero como todos los individuos que presentan color de ojos blancos son machos, no se puede inferir que la mutación white se comporte como un carácter mendeliano. Es importante notar, para el ejemplo de la segunda generación, que cuando en la descendencia femenina se presenta el alelo w, éste siempre se ve eclipsado por el alelo w+ que porta el otro homólogo X. Sin embargo, cuando el alelo w aparece en la descendencia masculina, se expresa en su fenotipo: en el cromosoma Y este alelo está ausente. A pesar de que el color blanco se manifieste en el fenotipo siendo recesivo, como este alelo se presenta solo, no puede hablarse de dominancia, sino de pseudodominancia. Entonces la pseudodominancia es el fenómeno por el cual, un solo alelo recesivo en el genomio, es capáz de expresarse en el fenotipo ante la ausencia del otro alelo. Tampoco se puede hablar de heterocigosis ni de homocigosis. Cuando el alelo recesivo se expresa por estar situado en un cromosoma que no posee homólogo, o en la porción no homóloga de un cromosoma sexual, estamos ante la situación de hemicigosis, y se dice entonces que el genotipo es hemicigota. Veamos qué ocurre cuando el cruzamiento de los parentales es inverso, es decir, cuando se produce entre hembras de ojos blancos y machos de ojos rojos: w w w+ Parentales x w+ w w+ w w Descendencia. w w+ w w- Figura nº14: Patrón de Herencia Cruzada. Ahora, el 50% de la descendencia es de color de ojos rojos, y corresponde al 100% de las hembras. El otro 50% es de color de ojos blancos, y se trata del 100% de los machos. Como puede verse, la descendencia femenina posee el fenotipo paterno y la masculina, el fenotipo materno. Este tipo de situación describe un patrón denominado patrón de herencia cruzada, que es la consecuencia del cruzamiento de un individuo de sexo homogamético homocigota recesivo para un gen ligado al sexo, con un individuo de sexo heterogamético, portador en su cromosoma X, de ese mismo gen en estado dominante, por lo cual su descendencia femenina recibe las características paternas de aquel gen, y su descendencia masculina recibe las características maternas. Otro caso análogo que podríamos citar es el de la tonalidad de la coloración de las alas de las mariposas. La tonalidad oscura domina sobre la clara. La diferencia es que como hemos visto, poseen el sistema ZW – ZZ de determinación cromosómica sexual, pero sin embargo, las consideraciones para el patrón de herencia cruzada son iguales. También el de las barras más oscuras aparecidas en las plumas de las gallinas, donde el fenotipo barrado es 11 dominante sobre el no barrado. Recordamos que su tipo de determinación cromosómica sexual también pertenece al mismo sistema que el de los Lepidópteros. En la especie humana, el daltonismo, la distrofia muscular de Duchenne y la hemofilia, son otros ejemplos de caracteres recesivos con loci en el segmento diferencial del cromosoma X. El daltonismo es la incapacidad de diferenciar los colores rojo y verde. Las personas que padecen esta enfermedad, (que también es conocida como “ceguera a los colores rojo y verde”), los ven como un solo color. Las mujeres son homocigotas recesivo, y los hombres son hemicigotas. De esta forma las mujeres pueden presentar tres genotipos, y sólo uno de ellos representa esta incapacidad. Los hombres poseen solo dos genotipos posibles y es uno de ellos dos que manifiesta el daltonismo. Esto explica porqué hay más hombres con daltonismo que mujeres y cómo las mujeres pueden transmitir el daltonismo sin padecerlo. Mujeres Genotipo Proporción Sin Daltonismo Con Daltonismo Hombres Genotipo Proporción DD Dd 75% D 50% dd 25% d 50% Figura nº8: Fenotipos daltónicos y sanos y sus proporciones en mujeres y hombres. La hemofilia (ausencia o mal funcionamiento de una proteína responsable de la coagulación de la sengre) y la distrofia muscular de Duchenne (enfermedad mortal que debilita y atrofia los músculos a temprana edad), poseen las mismas características que las del daltonismo. En el hombre, existen pocos casos de fenotipos ligados al cromosoma X y dominantes. Uno de ellos es la hipofosfatemia (raquitismo resistente a tratamientos con vitamina D). En este caso, observando la figura nº16, puede verse que el porcentaje de mujeres que padecen esta enfermedad es mayor que el de los hombres. Estos últimos siguen presentando dos genotipos posibles de los cuales solo uno manifiesta la hipofosfatemia. Las mujeres presentan otra vez tres genotipos, pero ahora solo el doble recesivo es el que no manifiesta la enfermedad. Mujeres Genotipo Proporción Con Hipofosfatemia Sin Hipofosfatemia Hombres Genotipo Proporción HH Hh 75% H 50% hh 25% h 50% Figura nº16: Fenotipos con hipofosfatemia y sanos y sus proporcioones en hombres y mujeres. Otro ejemplo de gen ligado al cromosoma X, es el “síndrome de femenización testicular”. Se trata de un fenotipo poco frecuente, ya que se presenta a razón de un caso cada 65.000 hombres. Las personas que lo padecen, son cromosómicamente varones, es decir que poseen los cromosomas sexuales XY. Sin embargo se desarrollan como mujeres: tienen genitalia externa femenina, vagina ciega y carecen de útero. Son estériles. Este 12 síndrome no es reversible con el tratamieneto con andrógenos, por ello también se denomina como "síndrome de insensibilidad a los andrógenos". En la causa de esa insensibilidad, está implicada la diferenciación sexual. Si hacemos una revisión de lo dicho en ese punto, recordaremos que en un individuo de sexo masculino, la testosterona producida por las células de Leydig del testículo una vez producida la diferenciación primaria, induce a un represor que en su estado no inducido impide la transcripción de los genes que codifican para el desarrollo de los conductos de Wolff, y que el mencionado represor está codificado a su vez por el gen i+. Cuando un hombre padece el síndrome de feminización testicular,la testosterona formada en las células de Leydig, no induce al represor del gen que permite que se desarrolen los conductos de Wolff, y por lo tanto éstos degeneran en lugar de evolucionar. Esto se debe a que este represor es diferente, ya que el gen que lo codifica ha mutado a su forma alélica Tfm; esta mutación genera un represor diferente capaz de impedir la transcripción de genes, pero incapaz de dejar de hacerlo ante la presencia de la testosterona. Por otro lado, la hormona antimülleriana sintetizada por las células de Sertoli parece detener su acción, puesto que en estos individuos XY, se efectúa la diferenciación sexual secundaria femenina. XY gen SRY Células de Sertoli Hormona Antimülleriana Células de Leydig ¿? Testículo Testosterona no non inducción Represor mutado no inducible i+ Tfm Gen Regulador Figura Degeneración de los Conductos de Wolff Desarrollo de los Conductos de Müller no transcripción Gen Estructural Diferenciación Sexual Secundaria Femenina Defectuosa Figura nº18: Mecanismo genético provocado por la mutación del gen i+ codificador del represor de los genes que permiten la degeneración de los conductos de Wolff. Figura nº17: Cuatro hermanos con el síndrome de feminización testicular, irreversible con tratamiento con andrógenos. 13 c) Caracteres limitados por el sexo Estos caracteres están gobernados por genes que se expresan solamente en un sexo aunque estén presentes en ambos. Para que un gen sea considerado limitado a un sexo, su penetrancia debe ser cero. Estos genes se hallan en autosomas o en la región apareante de los cromosomas sexuales, y codifican para caracteres sexuales secundarios. He aquí algunos ejemplos: • • En el sexo masculino: Distribución facial del vello (en el hombre) Producción de esperma (en todas los organismos superiores) Tono grave del mugido (en vacunos) Pelos menos suaves y más largos (en vacunos) Plumaje brillante y llamativo (en gallináceas) Rayas en la pelusa de los pollitos (en gallináceas) Presencia de cuernos (en cabras y antílopes) En el sexo femenino: Formación de glándulas mamarias (en mamíferos) Producción de leche (en mamíferos) Distribución uniforme de la grasa (en vacunos) Coloración blanca en élitros (mariposas). Veamos un ejemplo en la expresión de la cola emplumada en pollos, que solo es penetrante en el sexo masculino: Fenotipos Genotipos Gallos Gallinas GG Plumaje de gallina Plumaje de gallina Gg Plumaje de gallina Plumaje de gallina gg Plumaje de gallo Plumaje de gallina Figura nº18: Expresión de fenotipos y genotipos en ambos sexos de genes limitados por el sexo. d) Caracteres influídos por el sexo Los genes que gobiernan estos caracteres aparecen también en ambos sexos, pero se dan más en uno que en otro. La expresión de la dominancia o de la recesividad de los alelos de los loci influídos por el sexo está invertida en machos y hembras. Por ejemplo, el gen de la calvicie en humanos muestra dominancia en el hombre pero actúa un forma recesiva en la mujer. Fenotipos Genotipos Hombres Mujeres CC calvo calvo Cc calvo no calvo cc no calvo no calvo Figura nº19: Expresión de fenotipos y genotipos en ambos sexos de genes influídos por el sexo. 14 Otros ejemplos que pueden mencionarse: • • • • • Presencia de cuernos en carneros. Coloración de pelaje de algunas razas de vacunos. Braquidactilia o cortedad del dedo índice. Presencia de barba de chivos. Cierto tipo de mechón de pelo blanco en el hombre. Tanto para los caracteres influídos como para los caracteres limitados por el sexo, es necesaria la presencia de hormonas sexuales a fin de que los genes puedan expresarse. 15