Rigor Mortis
Anuncio

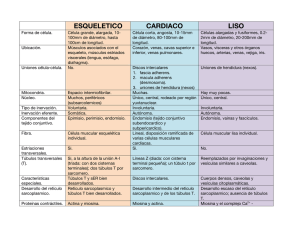
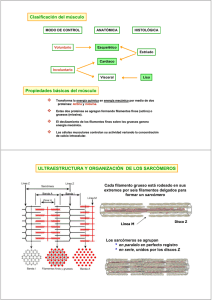
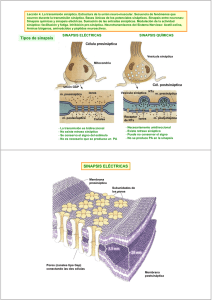
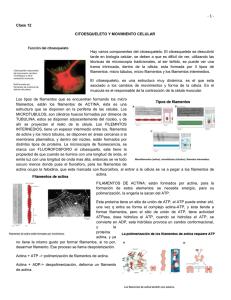
RIGOR MORTIS INTRODUCCIÓN El rigor mortis es un fenómeno que tiene lugar unas horas después del fallecimiento. Consiste en una contracción muscular continuada debida a un déficit de ATP que hace que el cadáver presente una rigidez palpable durante varias horas, o incluso días. CONTRACCIÓN MUSCULAR El músculo esquelético está formado por fibras musculares (miofibrillas), las cuales tienen en su interior dos tipos de filamentos: el filamento delgado, formado por actina, y el filamento grueso, formado por miosina. Estos filamentos están parcialmente entrecruzados, formando bandas H, A e I. Las bandas I contienen filamentos de actina, las bandas A contienen filamentos de miosina y los extremos de los filamentos de actina, y las bandas H se componen de filamentos de miosina únicamente. Los discos Z se encargan de anclar los filamentos de actina. Estos discos se extienden por todas las miofibrillas uniendo unas con otras. La porción de miofibrillas que queda entre los dos discos Z se llama sarcómero y son las unidades de contracción muscular. La contracción muscular se inicia en el sistema nervioso central desde donde se envía un impulso nervioso a una neurona que sinapta la placa motora del músculo en cuestión. Esta neurona motora libera acetilcolina que se une a sus receptores de las fibras musculares provocando una entrada masiva de sodio; produciendo así una despolarización celular, un potencial de acción. La despolarización activa los canales de dihidropiridinas de los túbulos T y los de rianodinas del retículo sarcoplásmico, canales de calcio voltaje−dependientes, provocando así una liberación de Ca2+ desde el retículo sarcoplásmico. Los iones calcio se filtran en los sarcómeros y se unen a troponina. Ésta está formada por tres dominios: un lugar de unión para el calcio, otro para la tropomiosina y el último para la actina. En situación de reposo las cabezas de miosina no están unidas al filamento de actina porque la tropomiosina tapa los lugares de unión. Cuando no hay contracción la miosina está unida a ADP−Pi. La unión de Ca2+ a la troponina hace que cambie la conformación de ésta y levanta la tropomiosina lo que hace que la actina muestre los lugares de unión para las cabezas de la miosina. Al unirse la miosina a la actina libera Pi quedando unida al ADP. En esta situación la miosina pierde afinidad por el ADP y lo libera produciendo un golpe de potencia. Las cabezas de miosina unidas a la actina giran unos 30º. Al producirse un desplazamiento de los filamentos, los sarcómeros se acortan provocando la contracción del músculo esquelético. Una molécula de ATP se une a la miosina, lo cual induce al desacoplamiento con la actina. Posteriormente la hidroliza, quedando unido a ADP−Pi, volviendo a la situación inicial antes de producir una nueva contracción. El músculo se relaja cuando termina el potencial de acción iniciado a nivel de unión neuromuscular, entonces las bombas de Ca2+ del retículo sarcoplásmico reintroducen el Ca2+ que se había liberado en su interior. La troponina vuelve a su conformación inicial separando la miosina y la actina. RIGOR MORTIS 1 Al morir se produce una parada de la circulación sanguínea, impidiendo así el recambio de gases. Entonces deja de producirse la captación de O2, que es esencial para la formación de ATP en el proceso de la fosforilación oxidativa (respiración) en las mitocondrias. Puesto que ya no se puede obtener ATP mediante esta vía aeróbica, que es la que proporciona una mayor cantidad, la célula utiliza otras fuentes de ATP que proporcionan una menor cantidad de ATP de una manera rápida. En el sistema fosfogénico la enzima creatin−quinasa hidroliza el fosfato de la fosforilcreatina, (compuesto que es almacenado para reponer ATP) y lo une al ADP; esta fuente de ATP sólo puede mantener la contracción durante unos 8−10 segundos. El sistema glicógeno−ácido láctico usa las reservas de glicógeno de los músculos. A través de este metabolismo anaeróbico el glicógeno es degradado y crea ATP y ácido láctico. Este método proporciona más ATP que el sistema fosfogénico pero es más lento que éste. Los iones de Ca2+ salen del retículo sarcoplásmico, que no es capaz de retenerlos y entran en el sarcoplasma desde el medio extracelular de modo que inundan los sarcómeros. Entonces se inicia el ciclo de contracción que sigue su curso normal hasta el momento del desacoplamiento de la miosina y la actina, que se detiene por la falta de ATP. Así, la fibra no se relaja y queda contraída. El rigor mortis se inicia en un intervalo comprendido entre 10 minutos y varias horas después de morir dependiendo de varios factores como la temperatura, el nivel de estrés antes de fallecer, etc. No obstante, su momento álgido tiene lugar entre las 12 y 24 horas post−mortem. En primer lugar, se ven afectados los músculos faciales y después se esparce por todo el cuerpo. Los músculos permanecen contraídos de uno a tres días, pero después los tejidos se degradan por la acción de los enzimas digestivos lisosomales, que se liberan. Así, la contracción decae y el cadáver pierde la rigidez. BIBLIOGRAFÍA − Strayer, L. Bioquímica. 3ª edición. Ed. Reverté. • Ira Fox, S. Human phisiology. 6ªedición. Ed. Mc Graw Hill. • Guyton,Arthur C; Hall, John E. Tratado de fisiología médica. 9ª edición. Ed. Mc Graw Hill. • Berne, Robert M; Levy, Matthew N. Fisiología. 2ª edición. Ed. Harcourt Brace. • Matthews; Van Holden, Ahern. Bioquímica. 3ª edición. Ed. Addison Wesley. cwx.prenhall.com/bookbind/pubbooks/martinidemo/chapter10/medialib/CH10/html/ch10_focus.html savell−j.tamu.edu/conversion.html health.howstuffworks.com/muscle6.htm www.nurseminerva.co.uk/dying.htm http://www.paramedicinstitute.com/galery4.asp 2