Guia Musculo como Efector
Anuncio

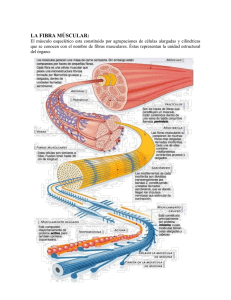
GUIA DE COMPLEMENTACION DE CONTENIDOS 3º Medio Común MUSCULOS COMO EFECTORES Vías Eferentes Estructuras de respuestas: los efectores El sistema nervioso es capaz de elaborar respuestas frente a los cambios ambientales las cuales son llevadas a cabo por estructuras especializadas llamadas efectores. Las células de los efectores son también células excitables que ante la llegada de un potencial de acción por la neurona motora recibirán los efectos de una sinapsis química, puesto que todas las células efectoras son postsinápticas. Algunos órganos efectores son: a.- Músculo Esquelético. b.- Músculo Liso. c.- Glándulas Exocrinas. d.- Glándulas Endocrinas La Contracción Muscular Se conocen dos formas fundamentales de contraerse los músculos, la contracción tónica y la contracción tetánica. La contracción tónica es la propia del tejido muscular liso o involuntario. En este tipo de músculos, una vez contraídos no hay gasto de energía, de manera que es una forma apropiada para mantener durante mucho tiempo la duración de la contracción; en algunos casos, el músculo tónico contraído adopta una notable rigidez, como sucede en el músculo abductor de las valvas de los moluscos bivalvos (almejas, ostras). La contracción del músculo liso es lenta y sostenida. Por el contrario, la contracción tetánica, propia de los músculos estriados o voluntarios, es muy rápida, y no se puede sostener más que mediante un aporte constante de energía. Se conoce bastante bien el mecanismo de la contracción muscular tetánica, estudiada en vertebrados. El tejido muscular constituye entre el 40 a 50 por ciento del peso total del cuerpo y cumple con las siguientes funciones: 1. Movimientos (reflejos y movimientos voluntarios). 2. Mantención de la postura corporal. 3. Producción de calor. Figura 1.82: (1) Contracción isotónica. El músculo al contraerse, se acorta y engruesa, realizando un trabajo eficaz. (2) Contracción isométrica. El músculo al contraerse, no cambia de longitud ni de espesor, solamente se endurece. Características de los músculos: Excitabilidad: Capacidad del tejido muscular de responder a los estímulos. Contractilidad: Capacidad del tejido muscular de generar de manera activa la fuerza que pueda contraer a la fibra, cuando el estímulo es suficiente (cambian su longitud). Extensibilidad: Este término alude a la propiedad de los músculos para estirarse sin sufrir daño. Elasticidad: Capacidad del tejido muscular para recuperar su forma y longitud original después de la contracción o extensión. Tonicidad: A través de la cual el músculo queda siempre semi contraído, ejerciendo de modo permanente una acción sobre los huesos a los que está adherido. Tabla 1.7 Tipos de músculos Para comprender mejor el funcionamiento muscular, estudiaremos el músculo esquelético o voluntario. En los vertebrados, cada músculo esquelético puede considerarse un órgano. Sus fibras alargadas, denominadas fibras musculares, se organizan en haces, llamados fascículos, envueltos en tejido conectivo. Cada fibra de músculo esquelético es una larga célula cilíndrica con muchos núcleos. La membrana plasmática, denominada sarcolema en el caso de la fibra muscular, presenta múltiples extensiones hacia el interior de la fibra que forman un conjunto de túbulos, conocidos como túbulos transversos. El citoplasma se denomina sarcoplasma y su retículo endoplasmático liso, retículo sarcoplásmico. Cada una de estas fibras contiene miofibrillas que corren a todo lo largo de la fibra muscular. Éstas se hallan constituidas por dos tipos de estructuras, muy pequeñas, los miofilamentos, los cuales son de dos tipos: filamentos de miosina y de actina. Los miofilamentos de miosina son gruesos. Cada filamento grueso consta de unas 300 moléculas de una proteína llamada miosina, conformadas a modo de palos de golf enrollados entre sí. Las colas de las moléculas de miosina adyacentes están dispuestas en paralelo y forman el cuerpo del filamento grueso. Las dos proyecciones de cada molécula de miosina se denominan cabezas de miosina o puentes cruzados, las cuales presentan sitios de unión para actina y sitios de unión para ATP. Figura 1.83: Estructura del músculo esquelético. Los miofilamentos delgados se extienden desde los puntos de anclaje en las líneas Z. Su componente principal es la proteína actina; en cada molécula de actina hay un sitio de unión a miosina donde puede insertarse una cabeza de esta proteína. También contienen las proteínas troponina (que presenta sitios de unión para calcio) y tropomiosina. En los músculos relajados, la miosina no puede unirse a la actina, porque la tropomiosina bloquea el sitio de inserción, a su vez, la tropomiosina se mantiene en su sitio gracias a la troponina. Estos miofilamentos están ordenados en compartimientos llamados sarcómeros, que son las unidades básicas de la contracción muscular. Cada sarcómero consiste en filamentos de miosina y actina que se superponen longitudinalmente en las fibras musculares, formando un patrón de bandas transversales o estrías característico del músculo estriado. Los sarcómeros están unidos en sus extremos por un entrelazamiento de filamentos, denominados línea Z. Cientos de sarcómeros conectados extremo con extremo constituyen una miofibrilla. Figura 1.84: Estructura de una fibra muscular a.- Unidad Motora y la Unión Neuromuscular La unidad funcional de los músculos de los vertebrados es la unidad motora, la que está formada por una sola neurona y el grupo de células musculares inervadas por su axón, las cuales se contraerán cuando se transmita el impulso nervioso por la neurona motora. En tanto, la unión neuromuscular se refiere a la sinapsis de un axón de la neurona motora y una fibra muscular. En la unión neuromuscular, el axón de la neurona motora se divide en diversos botones sinápticos. Dentro del citosol de cada uno se encuentran cientos de vesículas sinápticas, cada una llenas de moléculas de acetilcolina, un neurotransmisor que se libera en la unión neuromuscular. El punto de contacto entre la neurona motora y el sarcolema de la fibra muscular se denomina placa motora terminal. Figura 1. 85: Estructura de la placa motora. Si una unidad motora es aislada y estimulada con breves descargas eléctricas de creciente intensidad, no habrá respuesta hasta que se alcance cierta intensidad umbral (Ley del Todo o Nada). En contraste, el Músculo entero, compuesto de muchas unidades motoras individuales, puede responder en forma graduada según el número de unidades motoras que se contraen en un determinado tiempo. b.- Fisiología de la contracción muscular La contracción muscular ocurre cuando los sarcómeros y, por lo tanto, las fibras musculares, se acortan. Este acortamiento se verifica cuando los filamentos de actina y miosina se deslizan uno sobre otro, incrementando su superposición. Este dispositivo actuaría como cremallera, en el cual los puentes, al ir cambiando de sitio en sitio, impulsarían las moléculas de actina. Para que se produzca la contracción muscular, es necesario que el potencial de acción penetre profundamente al músculo. Esta penetración se efectúa a través de los Túbulos T. Estos túbulos T penetran a través de las fibras musculares, cruzando las miofibrillas justamente en aquellos puntos en que los filamentos de actina y miosina se entrecruzan. Cada vez que el Potencial de Acción difunde a través de los túbulos T, se liberan iones calcio desde el Retículo Sarcoplásmico hacia las vecindades de las miofibrillas. Estos iones se enlazan directa y fuertemente con la troponina y hacen que los complejos troponina-tropomiosina se alejen de los sitios donde la miosina se une a la actina. Una vez "libres" estos sitios, se inicia el ciclo de contracción. Figura 1.87: Modelo de contracción muscular Ciclo de contracción muscular Cuando una neurona motora transmite un impulso nervioso, libera al neurotransmisor acetilcolina en la hendidura sináptica. • La acetilcolina se une a receptores de cada fibra muscular, produciendo la apertura de canales iónicos, que permiten que ingresen a través de la membrana iones pequeños entre los que resalta el Na+. • El ingreso de Na+ modifica el potencial de membrana y dispara el potencial de acción muscular, que a continuación se propaga por el sarcolema y el sistema de túbulos T. • El efecto de la acetilcolina dura muy poco, dado que este neurotransmisor es eliminado rápidamente por la acción de la enzima acetilcolinesterasa. Ello da fin a la generación de potenciales de acción muscular. • Al propagarse el potencial de acción muscular a través del sarcolema a los túbulos T, se abren los canales de liberación de calcio en la membrana del Retículo sarcoplásmico. Como resultado, los iones de calcio salen al citosol, alrededor de los filamentos gruesos y delgados. • Los iones de calcio se combinan con la troponina y hacen que modifiquen su forma. Este cambio de conformación ocasiona que el complejo troponinatropomiosina se aleje de los sitios de unión de miosina en la actina, que entonces quedan expuestos. • La cabeza de miosina contiene una zona de unión al ATP y una enzima (adenosintrifosfatasa) que degrada al ATP en ADP (adenosintrifosfato) y fosfato. Esta reacción de degradación le confiere energía a la cabeza de miosina (debemos tener en claro que los productos de la hidrólisis del ATP todavía se hallan unidos a la cabeza de miosina). • La cabeza de miosina energizada se enlaza a sus sitios de unión en la actina y luego libera el grupo fosfato y el ADP desencadenándose la fase de deslizamiento, haciendo que la miosina experimente un cambio de conformación y provocando que la cabeza de miosina gire. • Al girar, la cabeza de miosina genera una fuerza, que tira del filamento de actina hacia el centro del sarcomero, con lo cual se desliza el filamento delgado sobre el grueso. Al término de la fase de deslizamiento, la cabeza de miosina permanece firmemente unida a la actina, hasta que se enlaza con otra molécula de ATP, la cual se adhiere a la cabeza de miosina. Así ésta se separa de la actina. • El ciclo de contracción se repite cuando la enzima adenosintrifosfatasa de la miosina degrada de nuevo al ATP. Esta reacción reorienta la cabeza de miosina y transfiere energía del ATP a dicha cabeza, que de nuevo está lista para combinarse con otro sitio receptor de miosina en un punto más distal del filamento delgado. El ciclo de contracción se repite una y otra vez, siempre y cuando haya ATP disponible y los niveles de calcio sean suficientemente altos cerca del filamento delgado. El movimiento continuo de las cabezas de miosina aplica la fuerza que acerca entre sí a las lineas Z, con lo que se acerca el Sarcómero. De esta manera, se contraen las miofibrillas y se acorta la fibra muscular. Después de la contracción, las fibras musculares vuelven a su estado de reposo. La acetilcolina presente en la hendidura sináptica es desactivada por la enzima acetilcolinesterasa. Entonces se bombean los iones de calcio de regreso al interior del retículo sarcoplásmico por transporte activo. Sin iones de calcio, los sitios activos en los filamentos de actina vuelven a estar cubiertos por el complejo troponina-tropomiosina. Los filamentos de actina se deslizan de regreso a su posición inicial, y ocurre la relajación del músculo. Figura 1.88: cambios en el relajado y contraído sarcómero c.- Metabolismo muscular A menudo se exige a las células musculares que realicen trabajo extenuante, de modo que deben contar con un gran suministro de energía. La fuente inmediata de energía necesaria para la contracción es el ATP. La contracción muscular requiere tanto ATP que las células musculares no pueden disponer del suficiente. En las moléculas de ATP, sólo puede almacenarse energía suficiente para unos pocos segundos de actividad intensa. Sin embargo, las células musculares tienen un compuesto de almacenamiento de energía de reserva, denominado Creatinfosfato (CP) o Fosfocreatina. El Creatinfosfato provee de grupos fosfatos ricos en energía para transformar el ADP en ATP. La energía acumulada en la forma de fosfatocreatina se transfieren al ADP, según sea necesario. La enzima implicada en esta reacción reversible es la creatinfosfoquinasa (CPK). Si la reserva de ATP en una célula es alta, el equilibrio de la reacción se dirige de tal manera que el grupo fosfato pasa del ATP a la Creatina. Si la reserva de ATP disminuye, el equilibrio se dirige en la dirección contraria, pasando los grupos fosfatos del Creatinfosfato al ADP. Sin embargo, durante el ejercicio vigoroso, el suministro de fosfocreatina dura unos 15 segundos, lo que basta para períodos breves de actividad máxima, como una carrera de 100 metros. Cuando los depósitos de ATP y CP se agotan, las células musculares deben reponer estos compuestos ricos en energía. El combustible utilizado para ello es el glucógeno, un polisacárido de elevado peso molecular formado por cientos de miles de unidades de glucosa. El glucógeno que había estado en almacenamiento es degradado para obtener glucosa, la que a su vez se degrada en la respiración celular. Cuando se dispone de suficiente oxigeno, de la glucosa se obtiene energía que se requiere para producir ATP y CP en las cantidades necesarias. Durante un ejercicio intenso, el aparato cardiovascular no es capaz de aportar oxígeno en cantidad suficiente para satisfacer las necesidades de las células musculares en rápido metabolismo. Esto da por resultado una deuda de oxígeno. En tales condiciones, las células musculares son capaces de degradar las moléculas de glucosa en forma anaeróbica por cortos períodos. La fermentación láctica es un método para generar ATP con rapidez, pero no en grandes cantidades. El agotamiento de ATP da por resultado contracciones más débiles y fatiga muscular. La acumulación de ácido láctico, un producto de desecho, también contribuye a la fatiga muscular.