El núcleo Archivo - Universidad de Antioquia
Anuncio

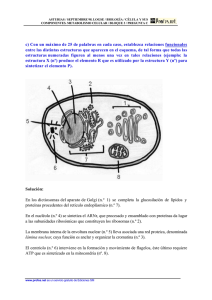
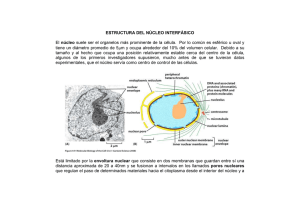
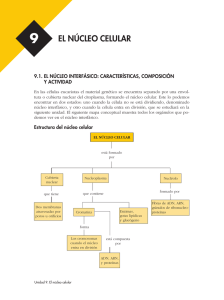
UNIDAD 2. EL NÚCLEO Documento elaborado con fines docentes por: GUSTAVO LOZANO CASABIANCA Biólogo M. Sc. Profesor asociado Escuela de Nutrición y Dietética Universidad de Antioquia VIVIANA MARTÍNEZ BETANCUR Bióloga. M. Sc. Universidad de Antioquia SANDRA MILENA JARAMILLO JARAMILLO Nutricionista Dietista Universidad de Antioquia La existencia del núcleo es la característica principal que diferencia las células eucariotas de las procariotas (1). El núcleo ocupa alrededor del 10% del volumen celular y en él está confinado casi todo el ADN en las células eucariotas (2). Por contener el genoma celular, el núcleo sirve de almacén de la información genética y como centro de control celular. La replicación del ADN, la transcripción y el procesamiento del ARN ocurren en el interior del núcleo, y sólo la última etapa de la expresión génica (traducción) tiene lugar en el citoplasma (1). CROMOSOMAS Y CROMATINA En los organismos eucariotas el ADN está distribuido en un conjunto de diferentes cromosomas que se encuentran en el núcleo. Cada uno de ellos está formado por una sola molécula de ADN muy larga asociada con proteínas que pliegan y empaquetan esta fina hebra formando una estructura más compacta. El complejo de ADN y proteínas unidas se denomina cromatina. Muchas de las proteínas asociadas al ADN de los cromosomas están implicadas en su empaquetamiento, pero algunas de ellas están involucradas en la expresión génica, en la replicación y en la reparación del ADN (3, 4). Las proteínas que se unen al ADN para formar los cromosomas eucariotas son generalmente clasificadas en histonas y no histonas. La unidad de la cromatina, el nucleosoma, consiste en ~146 pb de ADN que se enrolla sobre un núcleo de proteínas formado por 8 histonas. La cromatina está formada por muchos nucleosomas que pueden observarse como una cadena de rosario en el que cada nucleosoma está unido a otro por medio de un ADN denominado de enlace o linker. Cada núcleo de proteínas está formado por dos copias de cada una de las cuatro histonas nucleosómicas: H2A, H2B, H3 y H4. La histona H1 está encargada de interactuar con el ADN linker (2, 4). Los cromosomas están compuestos de varias regiones fácilmente identificables: Centrómero: Es una región especializada del cromosoma que está encargado de proveer sitios de unión para la segregación durante la mitosis y la meiosis, asegurando así la correcta distribución de los cromosomas a las células hijas (1, 5). Cromátidas hermanas: El ADN celular se duplica durante la interfase formando dos copias antes de que comience la mitosis; cuando esta empieza, la condensación de la cromatina conduce a la formación de cromosomas visibles en el microscópico óptico, que consisten en dos cromátidas hermanas idénticas que se mantienen unidas por el centrómero (Figura 2.1) (1). Telómeros: Corresponden a las secuencias situadas al final de los cromosomas eucariotas, los cuales desempeñan un papel crítico en la estabilidad de estos y en la división celular (6). Figura 2.1 Cromosoma mitótico en metafase Durante la interfase, cuando las células no están en división, el material genético ocupa un área en la cual no se pueden distinguir los cromosomas individuales y está disperso por todo el núcleo. Posteriormente, el empaquetamiento y la compactación de la cromatina durante la mitosis originan cromosomas visibles en la metafase (2). En la interfase se puede encontrar el núcleo con dos tipos de cromatina: La eucromatina corresponde a las regiones en donde las fibras están menos condensadas, tiene un aspecto disperso y ocupa la mayor parte del nucleoplasma. Además es activa transcripcionalmente (2, 7). La heterocromatina corresponde a regiones altamente condensadas, que han sido asociadas a genes inactivos. Se localiza en diferentes sitios a lo largo del cromosoma, incluyendo los centrómeros y telómeros (2, 7). EL GEN La existencia de un organismo depende de la capacidad de las células para almacenar, reparar y transmitir la información genética necesaria para originarlo y mantenerlo. La información hereditaria pasa de una célula a otra por medio de la división celular y de una generación de organismos a la siguiente mediante las células reproductivas. Estas instrucciones son almacenadas durante toda la vida de la célula en los genes, dicha información contiene tanto los elementos que determinan las características del individuo como las de la especie (2). En términos moleculares, un gen es definido como la secuencia completa del ácido nucleico que es necesaria para la síntesis de un producto génico funcional sea polipéptido o ARN (8). El gen consiste de las regiones codificadoras llamadas exones, y algunas veces regiones no codificantes denominados intrones (2). La mayoría de los intrones no especifican la síntesis de un producto celular, sin embargo desempeñan papeles importantes en el control de la expresión génica. El gen completo se transcribe para producir una molécula de ARN en la que los intrones se han separado mediante splicing (corte de intrones y empalme de exones), por lo que sólo los exones se encuentran incluidos en el ARNm. La presencia de intrones permite que los exones de un gen se unan en distintas combinaciones, resultando en la síntesis de distintas proteínas a partir del mismo gen; este proceso ocurre con frecuencia en los genes eucariotas y se denomina splicing alternativo (1, 9). El gen consiste además de los promotores y las regiones control necesarias para la transcripción, procesamiento y, si es aplicable, traducción del gen. Además estas regiones son responsables de asegurar que el gen se transcriba en el momento y en el tipo celular adecuado (2, 9). ENVOLTURA NUCLEAR El núcleo está delimitado por una envoltura nuclear formada por dos membranas y la lámina nuclear que proporciona un armazón estructural al núcleo y que separa el contenido del núcleo del citoplasma actuando como una barrera selectiva que impide el libre paso de las moléculas entre el interior nuclear y el citoplasma. Las membranas nucleares reciben el nombre de membrana nuclear interna y externa; esta última se continúa con la membrana del retículo endoplasmático (RE) por lo que hay una comunicación directa entre el espacio intermembrana y el lumen del RE y además posee ribosomas adheridos a la superficie (1). Subyacente a la membrana nuclear interna se localiza la lámina nuclear, una red fibrosa que proporciona soporte estructural al núcleo; está compuesta de lamininas que son proteínas de los filamentos intermedios. La lámina nuclear se une a la cromatina a través de las histonas H2A y H2B además de otras proteínas cromatínicas. Las lamininas se extienden también formando una red a través del interior del núcleo; muchas proteínas que funcionan en la síntesis y transcripción del ADN se unen a las lamininas (1). Las moléculas pequeñas, incluso algunas proteínas, pueden pasar libremente de manera pasiva a través de la envoltura nuclear en ambas direcciones por medio de canales acuosos abiertos de aproximadamente 9 nm de diámetro. La mayoría de proteínas y ARN, sin embargo, no son capaces de pasar por estos canales abiertos. Ellos utilizan el complejo de poro nuclear para atravesar la envoltura mediante un proceso activo, en el que son reconocidos y transportados selectivamente en una dirección específica (1). Los complejos de poro nucleares permiten un intercambio controlado de moléculas entre el núcleo y el citoplasma. Este complejo es una estructura muy grande con un diámetro de aproximadamente 120 nm; está compuesta de 30 - 50 proteínas distintas llamadas nucleoporinas con una simetría de octámero organizada alrededor de un canal central grande, que es la vía que utilizan las proteínas y los ARN para atravesar la envoltura nuclear. (Figura 2.2) (1, 10). Figura 2.2 Modelo del complejo del poro nuclear El tráfico de proteínas y ARN a través de los poros es altamente selectivo y no sólo mantiene la composición interna del núcleo sino que tiene un papel clave en la regulación de la expresión génica. Las moléculas de ARN que son sintetizadas en el núcleo deben ser transportadas de manera eficiente al citoplasma, donde intervienen en la síntesis de proteínas. Por otro lado, las proteínas necesarias para las funciones nucleares (ej. factores de transcripción) deben entrar en el núcleo procedentes de los lugares de síntesis en el citoplasma (1, 10). NUCLEOLO Es la estructura más destacada del núcleo, es esférica sin una membrana que lo delimite; está rodeada por nucleoplasma. Contiene concentraciones elevadas de ARN y de proteínas (2). Su función principal es la transcripción del ARN ribosomal (ARNr), procesamiento del pre-ARNr, modificaciones del ARNr y el ensamblaje de los ribosomas (11). El nucléolo está compuesto por macromoléculas incluyendo los propios genes ARNr, el pre-ARNr, los ARNr maduros, las enzimas procesadoras del ARNr, las subunidades proteicas de los ribosomas y algunos ribosomas parcialmente ensamblados (3). Los ribosomas son las unidades fundamentales de la síntesis peptídica (2). Están compuestos por diferentes moléculas de ARNr y más de 50 proteínas organizadas en una subunidad grande y en una pequeña (12). Las subunidades ribosomales y las moléculas de ARNr son designadas comúnmente en svedbergs (S), una medida de la tasa de sedimentación de partículas suspendidas centrifugadas bajo condiciones estándar (12). Los ribosomas eucariotas contienen 4 tipos diferentes de ARNr: ARN 18S, 28S, 5.8S y 5S. Los ARNr 5.8S, 18S y 28S son transcritos como una única unidad en el nucléolo por la ARN polimerasa I, dando lugar a un ARN precursor ribosómico que es procesado para originar los ARNr. El ARNr 5S se transcribe fuera del nucléolo por la ARN polimerasa III (1). Las células necesitan gran cantidad de ribosomas para satisfacer la necesidad de síntesis de proteínas (1); es por esto que las células que están fabricando cantidades especialmente grandes de proteína poseen un nucléolo de gran tamaño que puede alcanzar el 25% del volumen nuclear total (3). BIBLIOGRAFIA 1. Cooper GM, Hausman RE. La célula. 4a ed. Madrid: Marbán; 2008. 2. Patiño Grajales PJ, Ossa Londoño JE, McEwen JG. Biología de la célula. Medellín: Biogénesis. 2006. 3. Alberts B, Bray D, Hopkin K, Jonson A, Lewis J, Raff M, Roberts K, Walter P. Introducción a la biología celular. 2a ed. España: Panamericana; 2006. 4. Lavrov SA, Kibanov MV. Noncoding RNAs and Chromatin Structure. Biochemistry (Mosc). 2007; 72(13): 1422-1438 5. Dalal Y, Furuyama T, Vermaak D, Henikoff S. Structure, dynamics, and evolution of centromeric nucleosomes. PNA. 2007; 104(41): 15974–15981. 6. Gilson E, Londoño-Vallejo A. Telomere Length Profiles in Humans. Cell Cycle. 2007; 6(20): 2486-2494. 7. Wakimoto BT. Beyond the Nucleosome: Epigenetic Aspects of Position–Effect Variegation in Drosophila. Cell. 1998; 93: 321–324. 8. Spieth J, Lawson D. Overview of gene structure. WormBook. 2006; 18: 1-10 9. Wessagowit V, Nalla VK, Rogan PK, McGrath JA. Normal and abnormal mechanisms of gene splicing and relevance to inherited skin diseases. J Dermatol Sci. 2005; 40(2): 73–84. 10. Lim R, Aebi U, Fahrenkrog B. Towards reconciling structure and function in the nuclear pore complex. Histochem Cell Biol. 2008; 129:105–116. 11. Goić B, Bustamante J, Miquel A, Alvarez M, Vera MI, Valenzuela P, Burzio LO. The nucleoprotein and the viral RNA of infectious salmon anemia virus (ISAV) are localized in the nucleolus of infected cells. Virology. 2008; 379: 55–63. 12. Lodish H, Berk A, Zipursky LS, Matsudaira P, Baltimore D, Darnell J. Biología celular y molecular. 4a ed. Madrid: Panamericana; 2002. 13. Koberna K, Malinsky J, Pliss A, Masata M, Vecerova J, Fialova M, Bednar J, Raska I. Ribosomal genes in focus: new transcripts label the dense fibrillar components and form clusters indicative of ‘Christmas trees’ in situ. J Cell Biol. 2002; 157: 743–748. 14. Puvion-Dutilleul F, Bachellerie J.P, Puvion E. Nucleolar organization of HeLa cells as studied by in situ hybridization. Chromosoma. 1991 ; 100: 395–409. 15. Sato S, Yano H, Makimoto Y, Kaneta T, Sato Y. Nucleolonema as a fundamental substructure of the nucleolus. J Plant Res. 2005; 118: 71–81.