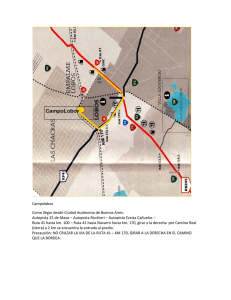
Efectos de la construcción de una autopista sobre una población de
Anuncio

Efectos de la construcción de una autopista sobre una población de avutardas: Influencia en la distribución espacial y efectos en la abundancia y la productividad AURORA TORRES MORENO DIRECTOR: CARLOS PALACÍN MOYA TUTOR ACADÉMICO: JAVIER SEOANE PINILLA 1 de febrero de 2010 MÁSTER OFICIAL EN ECOLOGÍA Índice Página Introducción .............................................................................................................................. 1 Objetivos e hipótesis ................................................................................................................. 4 Materiales y métodos ............................................................................................................... 5 Especie de estudio .................................................................................................................. 5 Área de estudio ....................................................................................................................... 6 Censos y muestreos de productividad .................................................................................... 8 Análisis de distancias a la R2 ................................................................................................. 9 Análisis BDACI de tendencias poblacionales ...................................................................... 12 Análisis BDACI de familias ................................................................................................. 15 Resultados ............................................................................................................................... 15 Modelos para las distancias a la R2 ...................................................................................... 15 Distribución de las observaciones en bandas ....................................................................... 19 Tendencias poblacionales de las zonas de impacto y control ............................................... 21 Productividad ........................................................................................................................ 24 Extinciones locales ............................................................................................................... 25 Discusión ................................................................................................................................. 26 Distancia a la autopista ......................................................................................................... 26 Abundancia y productividad ................................................................................................. 29 Otros estudios necesarios...................................................................................................... 34 Conclusiones ........................................................................................................................... 35 Bibliografía ............................................................................................................................. 35 Anexo.……………………………………………………………….……………………………………………40 40 Efectos de la construcción de una autopista sobre una población de avutardas: Influencia en la distribución espacial y efectos en la abundancia y la productividad Aurora Torres Moreno ([email protected]) Resumen: Cada vez es mayor la densidad de carreteras como consecuencia de un creciente desarrollo de las redes de transporte. Este aspecto es especialmente notorio en zonas periurbanas, por lo que en éstas resulta clave la planificación y gestión. En la mayoría de los estudios que evalúan efectos de las carreteras desde un enfoque poblacional, la evidencia de efectos está seriamente comprometida por la debilidad en el diseño de estudio. En este trabajo se analizaron los efectos de la construcción de una autopista sobre una población de avutarda común (Otis tarda), una especie globalmente amenazada, en un espacio protegido de la Comunidad de Madrid mediante el análisis de una serie temporal (1997-2009) de datos demográficos, que incluyeron las fases previa y posterior a la construcción de la infraestructura. Se realizaron modelos aditivos generalizados (GAMs) para examinar el efecto de la distancia a la carretera sobre la probabilidad de presencia de la especie, y se empleó el diseño BACI (Before-AfterControl-Impact) en el análisis de tendencias poblacionales y de número de grupos familiares. Los resultados obtenidos indican que la autopista ha tenido efectos negativos sobre el uso del espacio de la población. Desde el inicio de la construcción, la especie tiende a evitar una banda de aproximadamente 500 m. En primavera la distancia fue menor, siendo el efecto perturbador de la carretera aparentemente secundario en comparación con la marcada fidelidad a las zonas de exhibición. Respecto a las tendencias demográficas, el número de individuos ha disminuido progresivamente desde la inauguración de la carretera hasta un 50%, a diferencia de las zonas control, donde la abundancia de individuos se ha mantenido o incrementado. Los efectos sobre la presencia de grupos familiares no fueron tan evidentes, probablemente debido a la elevada variabilidad natural del éxito reproductivo de la especie. Palabras clave: aves esteparias, distancias umbral, BACI, efectos de carreteras, tendencias poblacionales Abstract: It is an evident fact nowadays the increasingly road density as a result of a growing development in the transport network. This subject is especially obvious in peri-urban areas, so that planning and management are key issues in them. In most of the studies that evaluate road effects on biological diversity with a population approach effects, evidence is seriously difficult because of the weakness of study design. In this study were analyzed the effects of a highway construction on a great bustard (Otis tarda) population, a globally endangered species, in a protected area of central Spain, through time series analysis (1997-2009) of demographic data, including the previous and subsequent phases to the infrastructure construction. We built generalized additive models (GAMs) to check the effect of road distance over probability of species presence, and it was applied the BACI (Before-AfterControl-Impact) design to analyze population trends and number of reproductive success. Our results suggest that the highway has had some negative effects on population spatial use. We realized that, since the beginning of the infrastructure construction, the species tended to avoid a band about 500 m from the roadside. In spring the avoided band was narrower, as the road disturbing effect was apparently secondary as compared to the strong site-fidelity to the exhibition grounds. Concerning the demographic trends, the number of individuals has reduced gradually since the road opening up to 50%, in contrast to control sites, where the abundance of individuals has remained or increased. Effects on number of family groups weren’t so evident, probably due to high natural variability of reproductive success in this species. Key words: BACI, population trends, road effects, steppe birds, threshold distances Agradecimientos: Este estudio se ha llevado a cabo dentro del proyecto CGL2005-04893/BOS financiado mediante un contrato HENARSA-CSIC. Los censos que he utilizado han sido financiados por la Comunidad de Madrid. He sido muy afortunada de poder contar con una extensa y rica base de datos, sin la que hubiera sido imposible realizar este trabajo. En su obtención ha participado mucha gente, a todos gracias. En mi entorno, mis amigos me han ayudado a desconectar y así disfrutar más de estos proyectos, y mi padre no sólo ha sido un enorme apoyo, sino también un atento revisor de este trabajo. Para ellos ya me he convertido en “la niña avutarda”. En general, debo agradecer la ayuda prestada a todo el equipo desde mi incorporación, entendiendo mi dedicación al master. Agradezco a Juan Carlos Alonso su confianza y que haya compartido conmigo sus conocimientos. No puedo olvidarme de las risas con Carolina Bravo y con Carlos Ponce, que aunque están siempre hasta arriba no pueden evitar transmitir lo que disfrutan con su trabajo ¡Ánimo en la recta final! Sobre todo quiero dar las gracias a mis queridos tutores, que han sido mis guías en todo este proceso, con una actitud ejemplar. Javier Seoane ha sido para mí uno de los grandes descubrimientos de la Universidad. Siento que nuestras charlas se extendieran, casi siempre, más de la cuenta, mea culpa. Si siempre he tenido ganas de aprender, ahora más todavía. Carlos Palacín, con sus frecuentes consejos y refranes me ha hecho mucho más fácil dar mis primeros pasos en el museo. A él debo agradecerle especialmente su paciencia e implicación en este trabajo, que ha disfrutado al menos tanto como yo. Gracias a mi madre (también a mi padre) estoy tan seducida por la naturaleza, así que a su memoria vaya dedicado mi primer trabajo en el mundo de la ecología. Introducción El crecimiento demográfico y la demanda progresiva de conexiones en el territorio están conduciendo al desarrollo de redes de transporte cada vez más complejas y de mayor envergadura. Consecuentemente, su papel como elemento estructurador en la configuración y articulación del espacio viene siendo mayor. En el caso de la Comunidad de Madrid, la red de carreteras se ha ido incrementando de forma notable en las últimas décadas. Concretamente, desde 1995 hasta 2007, el número de vías de gran capacidad (autopistas de peaje, carreteras de doble calzada y autovías) se duplicaron llegando a alcanzar casi 1000 km de longitud (Dirección General de Carreteras, 2010). Estas circunstancias son consecuencia, en gran parte, del crecimiento que ha experimentado la ciudad de Madrid desde los años sesenta. En ese tiempo ha llegado a saturar el espacio interior de su tradicional área urbana, al tiempo que se ha proyectado hacia el exterior mediante un modelo de crecimiento en el que coexisten la anexión de los núcleos menores de la periferia inmediata y la conurbación en los ejes de las grandes autopistas radiales (Serrano et al., 2006). Además, al igual que en otras ciudades españolas y europeas, en los últimos años se constata una progresiva dispersión de la urbanización sobre el territorio (Serrano et al., 2006). Actualmente, el conocimiento, el planeamiento y la gestión de estas zonas cada vez más humanizadas se considera un aspecto crítico para la sociedad y su futuro (Forman et al., 2002). Los efectos de las carreteras sobre la diversidad biológica, así como los mecanismos capaces de alterar a las comunidades faunísticas, son cada vez más estudiados (ver revisión de Forman et al., 2003). Entre estos últimos, han sido descritos: (1) el ruido, (2) la contaminación atmosférica, de suelos y aguas, (3) la mortalidad por colisión con el tráfico, (4) el efecto barrera, y (5) la pérdida de hábitat por ocupación directa (Fig. 1). Estos podrían influir, directa o indirectamente, en un incremento de la mortalidad, en la pérdida de hábitat más allá de la superficie ocupada por la carretera, o en la subdivisión de las poblaciones. En cuanto a la distribución espacial, parece que el efecto de la distancia a la carretera podría ser una importante variable a considerar, ya que la influencia de la vía podría extenderse más allá de la superficie de ocupación inmediata, pudiendo alcanzar desde pocos cientos de metros a varios kilómetros en función de las características del objeto de estudio y del paisaje (van der Zande et al., 1980; Reijnen et al., 1996; Forman y Deblinger, 2000). En este sentido, un estudio realizado en Holanda demostró que la diversidad de aves, la presencia de especies 1 IMPACTOS DE LAS CARRETERAS Ruido Contaminación Mortalidad por tráfico Barrera Inaccesibilidad Ocupación directa de hábitat Pérdida del hábitat disponible Incremento de la mortalidad Subdivisión de la población Podrían traducirse en cambios en: Distribución espacial Tamaño poblacional Éxito reproductivo Figura 1. Representación esquemática de los principales mecanismos y efectos asociados a las carreteras (Modificado a partir de Jaeger et al., 2005). especialistas y la densidad total de población fueron menores en hábitats adyacentes a las carreteras con tráfico abundante (Reijnen et al., 1995; 1996). Este aspecto es frecuentemente estudiado una vez que la carretera ha sido construida, es decir, desconociendo la situación anterior a la construcción de la misma (p. ej. Forman et al., 2002; Palomino y Carrascal, 2007), debido a que en la mayoría de los casos no existe la información previa necesaria. Respecto a las tendencias poblacionales, en algunas ocasiones están muy relacionadas con los cambios ambientales inducidos por el ser humano (p. ej. Tucker y Heath, 1994; revisión en Newton, 1998; Donald et al., 2001). En una reciente revisión de los efectos de carreteras sobre la abundancia animal (Fahrig y Rytwinski, 2009) se ha puesto de manifiesto que los estudios donde se han documentado efectos negativos de las carreteras son cinco veces más frecuentes que aquellos que registraron efectos positivos. No obstante estos datos deben ser considerados con prudencia. En general, en anfibios y reptiles aparecieron efectos negativos. Las aves mostraron principalmente efectos negativos o ausencia de efectos, con algunos casos de efectos positivos. Los micromamíferos fundamentalmente mostraron efectos positivos o ausencia de efectos. Y finalmente en los grandes mamíferos predominaron efectos negativos. 2 Además, para entender algo más de la dinámica poblacional es necesario conocer los posibles efectos sobre ciertos parámetros demográficos como la productividad, supervivencia, proporción de reproductores, que nos pueden dar las claves de cómo se regula la especie (Newton, 1998). Los estudios que evalúan los efectos de las carreteras desde un enfoque poblacional son cada vez más numerosos (Fahrig y Rytwinski, 2009), pero en muchos de ellos la evidencia de efectos está seriamente comprometida por la debilidad en el diseño de estudio. En muchos casos no se realiza un análisis directo de los efectos de las carreteras, sino que se incluyen variables predictoras relacionadas con las carreteras, como distancia a las mismas, densidad o longitud. En estos casos los trabajos no fueron diseñados con la intención de cuantificar los efectos de carreteras. Uno de los mejores métodos de análisis de este tipo de estudios es el diseño BACI (beforeafter-control-impact; Underwood y Chapman, 2003; Roedenbeck et al., 2007), en el que un cierto parámetro es estudiado durante bastantes años, tanto antes como después de una alteración, en zonas control y en zonas presumiblemente alteradas. A pesar de ello, este tipo de estudio es excepcional en la literatura de efectos de carreteras, tan sólo hay unos pocos estudios BA (before-after), pero que empleen este tipo de diseño para ver diferencias en la abundancia animal (Fahrig y Rytwinski, 2009), probablemente porque no se suele contar con la información espacial y temporal adecuada. Consecuentemente, se exigen estudios bien diseñados de los efectos de las carreteras en la abundancia animal y en otros parámetros, con el objetivo de mejorar la fortaleza de los trabajos y poder ser más empleados en la gestión y en la planificación de infraestructuras. La mayoría de estudios que se han realizado en medios forestales, húmedales y praderas (p. ej. Forman et al. 2002; Palomino y Carrascal, 2007) y son muy escasos los trabajos realizados en zonas agroesteparias, a pesar de que están siendo bastante afectadas por el trazado de infraestructuras dadas sus características geomorfológicas. En el presente trabajo se analizan los efectos de la construcción y funcionamiento de una autopista sobre una población de avutarda común, Otis tarda (Linnaeus, 1758), en un espacio protegido de la Comunidad de Madrid, a partir de una larga serie de datos que incluyen las fases previa y posterior a la construcción de la infraestructura. Por una parte, se analiza el efecto de la distancia a la carretera y se exploran las posibles variaciones estacionales sobre 3 ese efecto, en función del ciclo anual de la especie, cuestión que todavía no ha sido abordada en ningún trabajo. Por otra, en este trabajo se va a aplicar, por primera vez, un diseño BACI para estudiar los posibles efectos de la autopista sobre la abundancia y la productividad de avutardas. Esto podría permitir, en su caso, diferenciar los cambios producidos por la carretera de los debidos a fluctuaciones que formen parte de las tendencias reales de la población (Cody y Smallwood, 1996; Primack, 1998), así como detectar posibles alteraciones de su estructura demográfica. Se trata, por tanto, de un estudio retrospectivo no manipulativo. Hay dos amenazas principales para la conservación de la especie; la intensificación agrícola y el desarrollo urbanístico y de infraestructuras, este último especialmente notable en la Comunidad de Madrid (Palacín, 2007). La intensificación agrícola ha recibido, en general, más interés (p. ej. Bro et al., 2004; Wretenberg et al., 2007), sin embargo otros tipos de influencia humana han sido menos explorados. Objetivos e hipótesis En este trabajo testamos la hipótesis nula de la ausencia de efecto de la construcción y funcionamiento de una autopista, la Radial 2 (R2), en la distribución espacial, el tamaño poblacional y la productividad de la población de avutardas comunes en las proximidades de la autopista. Por tanto, el objetivo principal de la tesis de maestría es determinar si ha habido efectos o no sobre los aspectos mencionados anteriormente y, si ha sido así, evaluar su magnitud. Para alcanzarlo se abordan los siguientes objetivos específicos: 1. Estudiar los efectos sobre la distribución en las zonas próximas a la R2, tratando de comprobar si hay diferencias en la distancia a la que se encuentran las localizaciones de avutarda de la carretera antes, durante y después de su construcción. Además se explora la variabilidad estacional de la especie y de los grupos familiares. 2. Evaluar los efectos sobre el tamaño poblacional en las zonas próximas a la R2 mediante la elaboración de índices de tendencias poblacionales en zonas de impacto y zonas control, también en todas las fases, es decir, utilizando un diseño BACI de 12 años de duración. 3. Analizar los efectos sobre la productividad, entendida como número de familias, aplicando de nuevo el diseño BACI. 4 Materiales y métodos Especie de estudio La avutarda común, es una de las aves paleárticas de mayor tamaño y está considerada como la más pesada del mundo con capacidad de vuelo. Además presenta un acusado dimorfismo sexual en tamaño y coloración, alcanzando pesos de 12-18 kg en machos y de 4-5 kg en hembras (Alonso et al., 2009). Como en muy pocas aves, el sistema reproductivo es de tipo lek (Johnsgard, 1991; del Hoyo et al., 1996; Morales, 2000), que consiste en un sistema de apareamiento poligínico en el que un conjunto de machos se exhiben en determinados lugares siendo visitados por las hembras para el apareamiento en primavera. El resto del año viven por separado y forman bandos unisexuales. Los pollos nacen a finales de mayo-primeros de junio y las madres permanecen junto a ellos los meses de verano, cuando son más vulnerables, para luego incorporarse a un grupo de hembras. El tamaño de familia, que en septiembre en esta especie oscila entre 1 y 3 pollos por hembra, presenta muy poca variación de unos años a otros (Martín, 2009). Otras características biológicas importantes, de cara a la valoración de efectos, son el hecho de ser una especie longeva, en la que los individuos de mayor edad conocidos alcanzaron 12 años (datos propios), y que puede ser extremadamente filopátrica y refractaria a la colonización de nuevas áreas por un marcado efecto de atracción coespecífica (Alonso et al., 2004). Esta especie tiene una buena detectabilidad por su tamaño, comportamiento y por las características del paisaje en el que se encuentra, lo que ha propiciado la realización de censos fiables de la población y de otros parámetros como la productividad (Alonso et al., 2005). Aunque España dispone de más de la mitad del total mundial estimado de esta especie con 27500-30000 individuos (Palacín y Alonso, 2008), se trata de una especie globalmente amenazada y ha sido catalogada como vulnerable a nivel mundial (IUCN, 2006) así como en el ámbito nacional (Palacín et al., 2004). A escala global, se considera que la tendencia poblacional es decreciente (Birdlife International, 2004) estando amenazada su supervivencia en diversos países europeos y en Marruecos (Palacín y Alonso, 2008). La tendencia del conjunto de la población española de avutardas no es negativa, a pesar de la paulatina reducción del área ocupada por la especie, observándose una mayor concentración de individuos en áreas con hábitat de alta calidad. En la Comunidad de Madrid, subsisten 5 aproximadamente, 1500 individuos distribuidos en 15 leks o grupos reproductores distribuidos por el este y el sur de de la Comunidad (Alonso et al., 2006). Para este estudio se han considerado dos unidades poblacionales de avutardas del este de la Comunidad de Madrid. La unidad de Valdetorres-Talamanca de Jarama es la que se encuentra más al norte de la Comunidad de Madrid y ocupa unas 6297 ha. Está constituida por dos leks distintos y es, después de Daganzo-Camarma, la población local que más ejemplares de avutarda común contiene dentro del área de estudio (427±86 individuos). En esta población se encuentra el lek en el que se concentran mayor número de machos y hembras en primavera de toda el área de estudio. La unidad de Daganzo-Camarma es la que ocupa una mayor superficie, aproximadamente 11661 ha, y la que mayor número total de avutardas comunes alberga en primavera (583±64 individuos). Se compone de 6 leks distintos y entre casi todos ellos existe continuidad espacial. Área de estudio El área de estudio está localizada en el este de la Comunidad de Madrid y en el oeste de la provincia de Guadalajara, en el Área de Importancia para las Aves (Important Birds Area, IBA) no. 074 “Talamanca-Camarma” (40º40’ N 3º25’ O, 52000 ha, 792 msnm) a 25 km de Madrid (Fig. 2). Se caracteriza por un clima mediterráneo semiárido, con precipitaciones medias anuales de 350-400 mm, y un largo periodo de sequía estival. La zona presenta un paisaje agroestepario de relieve suave, dedicado principalmente al cultivo extensivo de cereales, sobre todo trigo (Triticum aestivum), cebada (Hordeum vulgare) o avena (Avena sativa), con sistema de rotación tradicional y pequeñas manchas de leguminosas, olivares y viñedos. Otros usos que caracterizan la zona son el ganadero, fundamentalmente rebaños de ovejas que son conducidos por pastores y perros, y el cinegético, basado en la caza de perdiz roja (Alectoris rufa; Linnaeus, 1758), conejo (Oryctolagus cuniculus; Linnaeus, 1758) y liebre europea (Lepus europaeus; Pallas, 1778). En este espacio existen varias figuras de la Red Natura 2000: dos Zonas de Especial Protección para Aves (ZEPA) “Estepas cerealistas de los ríos Jarama y Henares” (Madrid) y “Estepas cerealistas de campiña” (Guadalajara), y el Lugar de Interés Comunitario (LIC) “Cuenca de los ríos Jarama y Henares”. Además, debido a la riqueza y abundancia de aves ha sido clasificado como una de las zonas de máxima importancia (hotspot) de aves esteparias de la Península Ibérica (Traba et al., 2007). 6 Figura 2. Localización de la zona de estudio. La autopista R2 es la línea de trazo rojo. El área en color verde claro corresponde a la IBA 074 “Talamanca-Camarma”. Las dos áreas en verde oscuro corresponden a la ZEPA 139 “Estepas cerealistas de los ríos Jarama y Henares” (en la Comunidad de Madrid) y la ZEPA 167 “Estepas cerealistas de campiña” (en la provincia de Guadalajara). La zona alberga la población de avutardas más importante de Madrid (65%), es decir, algo más de 900 ejemplares (Alonso et al., 2006). También es un territorio relevante para otras aves esteparias como el aguilucho pálido (Circus cyaneus; Linnaeus, 1758) y cenizo (C. pygargus; Linnaeus, 1758), cernícalo primilla (Falco naumanni; Fleischer, 1818), sisón común (Tetrax tetrax; Linnaeus, 1758) o ganga ortega (Pterocles orientalis; Linnaeus, 1758). Al sur y sureste de la IBA, próximo a la localidad de Alcalá de Henares, discurre la R2 y un tramo común de la R2 y M50. En total abarca una longitud de 34 km en el interior de la IBA y la anchura promedio es aproximadamente de 25 m, con dos calzadas. Se trata de una autopista de peaje que conecta la M50 y Guadalajara y transcurre prácticamente paralela a la autovía 7 del nordeste (A2). Las obras de construcción se iniciaron en octubre de 2001 y tras dos años fue inaugurada el 6 de octubre de 2003. Tiene un tráfico promedio de 9345 vehículos/día (datos de Intensidad media diaria de tráfico facilitados por HENARSA, 2009). Censos y muestreos de productividad Los datos necesarios para realizar el estudio se han obtenido principalmente de dos fuentes: censos poblacionales y muestreos de productividad, que han aportado la información necesaria para el análisis de tendencias poblacionales y de parámetros demográficos, así como de localización de individuos. En ambos casos se ha utilizado una serie temporal de censos que van desde 1997 hasta 2009, realizados con una metodología homogénea. Excepto en el estudio de las tendencias poblacionales que se excluyó 1997 ya que no fue completo en algunas zonas utilizadas. La selección de los censos y/o muestreos empleados, que define 4 temporadas (Tabla 1), viene justificada por los siguientes aspectos: 1. Censo de invierno (diciembre/enero): Dado que la especie es migradora parcial y diferencial en el área de estudio (Palacín et al., 2009), el censo refleja la cantidad de aves invernantes, que está conformada por individuos sedentarios o exclusivamente invernantes. 2. Censo de primavera (marzo): Es el más adecuado para determinar la cantidad de individuos potencialmente reproductores debido a que al final de este mes es cuando se producen las mayores agregaciones de avutardas en torno a sus zonas de exhibición. 3. Muestreo productividad de julio: Permite obtener la cantidad de familias (hembras con pollos) cuando los pollos alcanzan entre 1 y 2 meses de edad y todavía no han empezado a dispersarse. 4. Muestreo productividad de septiembre: Tiene por objeto registrar la cantidad de familias que permanecen hasta final del verano, habiendo superado los pollos la fase de mayor mortalidad juvenil. En esta época son detectados más fácilmente. A partir de este momento empiezan a incrementarse los movimientos. El número de pollos avistados durante los censos de julio y septiembre deben considerarse como valores relativos que permiten comparaciones válidas entre zonas y entre años, pero no son una muestra absoluta del número de jóvenes nacidos en una población. La cantidad de 8 hembras que se observan en septiembre es siempre considerablemente inferior a la registrada durante la primavera en todas las poblaciones, ya que en ese mes pueden aún permanecer en zonas de cría alejadas de los lugares habituales con presencia de la especie, sin integrarse en bandos. Por este motivo los datos de julio y septiembre son considerados muestreos, a diferencia de los censos de primavera e invierno que son absolutos. Tabla 1. Resumen de los censos (C) o muestreos (M) utilizados para cada objetivo. Distancias Individuos C. invierno C. primavera M. julio M. septiembre Familias Tendencias poblacionales Individuos Parámetros demográficos Familias C. primavera M. septiembre M. septiembre Se ha llevado a cabo una compilación exhaustiva de los mismos, en total 23 censos y 26 muestreos, en los que se ha empleado un total de 132 días de trabajo de campo. Todos fueron revisados con el fin de trabajar con datos sólidos y homogéneos. En total se registraron 1517 observaciones y un total de 15689 avutardas. Todos los seguimientos fueron llevados a cabo por observadores con experiencia en censar avutardas, con conocimiento del área de estudio y una rigurosa metodología. Se realizó un recorrido sistemático de la superficie a prospectar con 2 o 3 equipos de 2 personas operando en vehículo todo-terreno con prismáticos y telescopios (20-60x), a baja velocidad y realizando frecuentes paradas (ver detalles en Alonso et al., 2005). Durante el censo se cartografiaron todas las aves vistas en mapas a escala 1:50.000 ó 1:25.000. Para hacer que los resultados fueran comparables entre años se usó un recorrido predeterminado. Estos datos vienen siendo tomados regularmente por el equipo, y desde el inicio de mi colaboración, en el verano de 2008, he podido participar en los censos y muestreos. Análisis de distancias a la R2 Para este apartado, se ha definido como zona de estudio una banda de 2 km en torno a la R2, teniendo en cuenta todas las localizaciones detectadas entre 1997 y 2009. Se seleccionó dicha distancia para asegurar que los efectos percibidos puedan atribuirse a la cercanía inmediata de la autopista y no a la proximidad de núcleos urbanos u otras alteraciones de origen humano que podrían aparecer si se incluyen áreas de estudio más amplias. 9 Los análisis de distancias se han realizado mediante dos aproximaciones: (1) utilizando todas las observaciones de censos y/o muestreos como unidad de análisis, y considerando la temporada para explorar la variabilidad estacional, y (2) con las observaciones de familias correspondientes a una sola temporada, ya que podría observarse un efecto diferente debido a los requerimientos ecológicos de las mismas (lugares seguros y con abundancia de alimento). En primer lugar, se ha realizado un análisis con la distancia como variable cualitativa, tomando para ello 4 bandas desde la carretera con una anchura de 0,5 km (Fig. 3). Teniendo en cuenta la frecuencia de observaciones en cada banda se han construido los histogramas para cada fase (antes, durante y después) con las frecuencias relativas, y se han testado las diferencias entre fases mediante una X2, a partir de una tabla de contingencia de 4x3, con el paquete “Rcmdr” en el programa estadístico R v. 2.10.1 (R Development Core Team, 2005). Figura 3. Mapa de las bandas en torno a la R2. Los puntos naranjas representan las localizaciones de avutardas durante el periodo 1997-2009. A continuación, tomando la distancia como variable cuantitativa, se han construido Modelos Aditivos Generalizados (GAMs; Hastie y Tibshirani, 1990). Se tratan de una extensión semiparamétrica de los Modelos Lineales Generalizados (GLMs) cuya fórmula básica es: Logit(p) = a0 + f1x1 + f2x2 + … + fkxk Donde la p representa la presencia o ausencia, y las f1 a fk son funciones de suavizado de las k variables predictoras (Guisan et al., 2002). En comparación con otros métodos, es suficientemente sensible para modelar las respuestas de las especies de forma satisfactoria 10 (Suarez-Seoane et al., 2002). A diferencia de los GLMs, que pueden expresarse más fácilmente en forma de una ecuación, la interpretación de los modelos GAM requiere la representación gráfica del modelo. Los modelos se han realizado con la presencia/ausencia de la especie en el área de estudio con errores binomiales y función logit usando como variable predictora la distancia a la carretera. A partir de estos modelos se obtiene una función matemática que permite su extrapolación a una superficie continua, en nuestro caso de probabilidad de presencia. Realmente las denominadas ausencias son pseudoausencias, pues se han obtenido al lanzar puntos aleatorios sobre la superficie que hemos estimado de hábitat mediante el siguiente proceso: para estimar la superficie de hábitat potencialmente utilizable se seleccionaron todas las observaciones de los censos y muestreos de las cuatro temporadas a partir de 1997 que se encontraban a distancias no superiores a 2 km (Fig. 4.a.). Figura 4. Esquema del procedimiento de selección de las zonas de utilización de la especie para generar las pseudoausencias: a) georreferen-ciación de las observaciones de censos y muestreos (19972009) en una banda de 2 km a la R2, b) áreas de campeo mediante el método Kernel al 99%, c) delimitación de las zonas, d) alteriones humanas registradas en la zona para su exclusión, y e) áreas de máxima utilización definitivas. 11 A continuación se estimó el área de campeo de la especie en la zona utilizando como procedimiento análitico el método fijo de Kernel (fixed Kernel; Worton, 1989) que es una función no paramétrica utilizada para estimar la distribución de un conjunto de localizaciones, creando unos contornos espaciales alrededor de las áreas con la misma intensidad de uso. Se han creado áreas que representan el 99% de probabilidad de uso con validación cruzada de mínimos cuadrados (Seaman y Powell, 1997) (Fig. 4.b.). Seguidamente, se delimitaron las zonas (Fig. 4.c.) y se eliminaron las superficies de alteraciones humanas que solapaban con los polígonos de Kernel obtenidos (Fig. 4.d.) anteriores al periodo de construcción de la carretera. Finalmente, ya que al realizar el Kernel se habían sobrepasado las bandas de estudio en ciertas zonas se ajustó con la capa del mismo, obteniendo finalmente una superficie fiable de hábitat utilizable para las avutardas (Fig. 4.e.). Para finalizar se tomó un banda de 100 m en torno a cada observación y se evitaron esas superficies al lanzar los puntos aleatorios. Siempre se obtuvo el mismo número de pseudoausencias que de presencias con el fin de que el diseño resultara equilibrado. Para todos los análisis de información geográfica se ha empleado la extensión Hawths Tools (Beyer, 2004) para ArcGis v. 9.2. Una vez con todos los datos se generaron tres modelos, uno para cada fase (antes, durante y después) con la librería mgcv en R v. 2.10.1., que como estimador no paramétrico univariante usa el suavizado mediante splines, es decir, genera curvas definidas por partes mediante polinomios. Los splines se emplean sobre todo para aproximar formas complicadas en los ajustes de curvas. Análisis BDACI de tendencias poblacionales La clave del diseño BACI está en que las zonas presumiblemente afectadas por un impacto se comparan con zonas control, tanto antes como después de que ocurra. Si no se consideraran zonas control la posibilidad de que un cambio observado fuera causado por algún otro fenómeno no podría ser excluida (Schmitt y Osenberg, 1996). Del mismo modo, si solo existieran datos de zonas de impacto y control posteriores al fenómeno de estudio las diferencias observadas solo serían válidas si ambas fueran idénticas en ausencia de impacto, Un subtipo del BACI es el diseño BDACI (before-during-after-control-impact; Roedenbeck et al. 2007), en el que también se analizan los efectos durante la construcción de una infraestructura (Fig. 5). Este procedimiento incrementa el conocimiento acerca de la reversibi lidad de los efectos que podrían ocurrir durante la fase de construcción pero que podrían ir disminuyendo después. 12 Figura 5. Esquema del diseño BDACI (before-during-after-control-impact). En este caso los efectos son evaluados en zonas control y de impacto antes, durante y después de la construcción de la autopista. Basado en Roedenbeck et al., 2007. En este estudio, la zona de impacto (ZI) fue delimitada como una banda de 2 km en torno a la R2. Las dos zonas de control seleccionadas (Fig. 6), se encuentran a 7.3 y 19.5 km de distancia de la autopista (calculada como la distancia al centroide de cada zona), poseen el mismo hábitat y son ambientalmente homogéneas a la zona de impacto. La Zona Control 1 (ZC1) pertenece a la misma unidad poblacional de avutardas que la zona de impacto (se tiene constancia de la existencia de movimientos de individuos, sobre todo hembras, entre varios leks a lo largo del mismo periodo reproductor y de cría; Martín, 2009), comienza a 2 km de la misma, y sólo considera las observaciones de la unidad que se encontraron a más de 4 km a la carretera. Esta distancia ha sido también escogida en otros trabajos basados en el diseño BACI (p. ej. Bro et al., 2004). La Zona Control 2 (ZC2), pertenece a otra unidad poblacional de avutardas de la IBA y está separada de la autopista por una distancia superior a 10.6 km. Para detectar los posibles cambios de tendencia del tamaño poblacional en las zonas de impacto y control se han analizado los censos de la población reproductora en la IBA en el periodo de estudio (1998-2009) con el programa TRIM v. 3.40 (TRends and Indices for Monitoring data; Pannekoek y van Strien, 2001), que calcula índices de población que representan el cambio interanual, suponiendo la variación relativa del tamaño poblacional total. Para ello calcula la ratio entre el tamaño de población de cada año y el año 1998 que hemos tomado como base. El programa analiza series temporales de datos mediante regresión loglineal y ha sido diseñado para tener en cuenta tanto el problema de la sobredispersión 13 como el de la autocorrelación temporal en el número de aves observadas (Pannekoek y van Strien, 2001). Esta técnica ha sido comúnmente utilizada para analizar series temporales de datos de aves (p. ej. Seoane y Carrascal, 2007; Wretenberg et al., 2007; Fasola et al., 2009). Los modelos TRIM pueden tener en cuenta años de cambio de la tendencia (changes points), es decir, de la pendiente, y covariables categóricas, que agrupan sitios en base a una serie de rasgos. En este estudio se ha incorporado la covariable “zona” con tres niveles: “ZI”, “ZC1” y “ZC2”, para comprobar si existen diferencias en las tendencias poblacionales de las zonas de impacto y las zonas control. En algunos modelos se han Figura 6. Localización de las zonas control 1 (verde) y 2 (azul) calculadas mediante el método de Kernel al 99%. La línea negra continua corresponde a la banda de 4 km a la seleccionado los años de cambio de manera directa para tratar de comprobar si la tendencia difiere antes y después de cada uno de ellos, carretera y la discontinua es la banda de 2 km tomando 1998, 2001 (construcción) y 2003 que delimita la zona de impacto. (apertura al tráfico) o 1998 y 2003. En otros se ha efectuado una selección de años de cambio por etapas, en la que se va testando la significación de los cambios de tendencia antes y después de cada año de cambio, de tal forma que si no alcanza un cierto umbral es eliminado del modelo. De las posibles combinaciones se han derivado una serie de modelos de tendencia lineal (linear switching trend models). Mediante este procedimiento se generaron unos índices poblacionales calculados a partir de la suma de efectivos totales observados del censo de marzo. A continuación, se determinó cuál fue el mejor modelo teniendo en cuenta el AIC (Criterio de Información de Akaike; Burnham y Anderson, 2002) y la significación estadística mediante test de verosimilitud (Likelihood ratio). El AIC es una estima de la distancia relativa entre un modelo ajustado y el mecanismo desconocido que genera los datos observados (Burnham y Anderson, 2002), por lo que no debe considerarse como valor absoluto, pues 14 como tal no es interpretable, sino que deben tenerse en cuenta los valores relativos al conjunto de modelos considerado. No se ha tenido en cuenta la mortalidad por atropello como causa de variación en el número de individuos ya que durante el periodo de estudio no se registró ningún suceso de este tipo. Análisis BDACI de familias Para este análisis se utilizaron los datos del muestreo de productividad de septiembre, ya que es en esta temporada cuando mejor se puede estimar el número de pollos, e inherente a ello el número de familias, porque se observan con mayor facilidad. Se ha seguido un diseño similar al anterior, con las mismas zonas de impacto y control (Fig. 6). En este análisis hemos decidido trabajar con familias, como unidad de muestreo, porque este parámetro nos da una idea del número de hembras que han criado con éxito, ya que el pollo habrá superado el periodo de máxima vulnerabilidad. Las familias, aunque suelen ser de un pollo en la mayoría de los casos, pueden llegar hasta tres. Es preferible trabajar con esta unidad que con la productividad ya que, aparte de que como zona de impacto no estamos seleccionando los núcleos reproductivos completos, la productividad en la avutarda es una medida con una enorme variabilidad (Palacín, 2007). Para comprobar si existen diferencias significativas o no entre las zonas de impacto y control, así como entre las fases de estudio se empleará un Análisis de la varianza simple (ANOVA; Reid et al., 2006). Resultados Modelos para las distancias a la R2 En todos los casos en los que fue analizada la fase previa a la construcción la distancia no llegó a explicar más de un 2.8% de la variabilidad en la presencia de avutardas, excepto en julio, con 5.5%, y en ningún caso resultó ser significativa, lo que indica que en ese periodo no había una distribución determinada en el territorio a lo largo del eje estudiado de 2 km. Los modelos GAM para las distancias anuales a la R2 indican que esta variable es significativa tanto durante la construcción como una vez finalizada (Tabla 2) y es ligeramente más explicativa en la primera (9.6%) que en la segunda (7.7%). Tabla 2. Tabla de devianza de los GAMs (errores binominales) para las distancias anuales, estacionales y de familias. Estos modelos relacionan la presencia/ausencia de localizaciones de 15 avutardas comunes con la distancia a la R2. Se da la devianza residual como medida de la complejidad del modelo, la devianza explicada como medida de su capacidad explicativa, el tamaño muestral “n” y los resultados del test de X2 para la variable distancia. Modelos Fase Devianza residual Devianza explicada (%) n X2 p-valor Antes Durante Después 900 359 1027 0,8 9,6 7,7 661 288 805 4,9 27,1 57,0 0,154 0,000* 0,000* Antes Durante Después Antes Durante Después Antes Durante Después Antes Durante Después 102 41 156 297 98 273 159 68 241 319 134 354 2,8 22,5 12,7 1,3 5,6 4,7 5,5 15,0 7,3 0,5 13,0 8,3 83 40 130 217 76 207 128 61 189 233 111 279 2,2 8,2 13,8 2,7 4,4 10,3 7,0 7,8 14,8 0,8 11,7 22,0 0,392 0,043* 0,016* 0,329 0,116 0,024* 0,080 0,051 0,003* 0,637 0,061 0,002* Antes Durante Después 138 44 121 1,7 24,0 9,2 101 44 111 2,3 9,3 9,4 0,236 0,030* 0,074 Anuales: Estacionales: Invierno Primavera Julio Septiembre Familias: Códigos de significación: * inferior a 0,05. Los resultados obtenidos indican que la probabilidad de presencia de las aves disminuye a medida que nos acercamos a la carretera (Fig. 7), evitando una distancia umbral de 500 m en fase de construcción y entre los 400 y los 800 m en la fase posterior. En esta última también se observa una oscilación a partir de los 1700 m. A pesar de que en todas las temporadas la variable distancia resultó significativa en la fase posterior a la construcción se encontraron diferencias en cuanto a su capacidad explicativa (Tabla 2) y su forma (Fig. 8). 16 Durante Después -1 -2 -3 -5 -4 s(Distancia) 0 1 2 Antes 0 500 1000 1500 0 500 1000 1500 0 500 1000 1500 2000 Distancia a la carretera (m) Figura 7. Estimación no paramétrica de las contribuciones aditivas de la distancia a la R2 con los datos anuales. Los valores del eje “y” corresponden al predictor linear, que se interpreta como la probabilidad de presencia de la especie. Se usó un suavizado spline con intervalo de confianza al 95%. En invierno la devianza explicada por los modelos para las fase de construcción fue 22.5%, y para la de utilización 12.7%, resultando la distancia significativa en ambas. Durante la construcción aumentó notablemente la probabilidad de presencia en torno a 1 km de distancia, y después parece detectarse una distancia umbral de entre 500 y 600 m a la R2 con mayor probabilidad de ausencias (Fig. 8). En julio y septiembre los modelos muestran resultados parecidos. Los modelos más explicativos corresponden a la fase de construcción (15 y 13%), estando la variable distancia en niveles cercanos a la significación y presentando una distancia entre 500 y 700 m en la que se ha reducido la probabilidad de presencia. En la fase posterior los modelos son menos explicativos aunque la variable distancia resulta muy significativa, situándose la distancia umbral entre 600 y 700 m. En cuando a los modelos GAM para las observaciones de familias en septiembre el de la fase de construcción es el más explicativo (24%) y la variable distancia resulta significativa (Tabla 2). Sin embargo en la fase de funcionamiento de la R2 el modelo es menos explicativo aunque parece que la distancia sigue teniendo cierta influencia, siendo casi significativo. A partir de la visualización de los resultados parece que el modelo ha cambiado considerablemente respecto a la fase anterior, de tal forma que en la fase de construcción, la distancia a la que aumenta la probabilidad de presencia notablemente es desde los 600 m, 17 Durante Después Invierno 0 500 1000 1500 0 500 1000 1500 0 500 1000 1500 -3 -4 0 500 1000 1500 0 500 1000 1500 0 500 1000 1500 1 2 Primavera Julio 0 500 1000 1500 0 500 1000 1500 0 500 1000 1500 -5 -4 -3 -2 -1 0 1 2 -4 -3 -2 -1 0 s(Distancia) s(Distancia) -2 -1 0 1 2 -4 -3 -2 -1 0 1 2 Antes Septiembre 0 500 1000 1500 0 500 1000 1500 0 500 1000 1500 2000 Distancia a la carretera (m) Figura 8. Estimación no paramétrica de las contribuciones aditivas de la distancia a la R2 con los datos estacionales. Los valores del eje “y” corresponden al predictor linear, que se interpreta como la probabilidad de presencia de la especie. Se usó un suavizado spline con intervalo de confianza al 95%. mientras que en la fase posterior a medida que nos alejamos de la carretera aumenta la probabilidad de presencia (Fig. 9). 18 Durante Después 0 -1 -2 -3 -5 -4 s(Distancia) 1 2 Antes 0 500 1000 1500 0 500 1000 1500 0 500 1000 1500 2000 Distancia a la carretera (m) Figura 9. Estimación no paramétrica de las contribuciones aditivas de la distancia a la R2 con los datos de familias. Los valores del eje “y” corresponden al predictor linear, que se interpreta como la probabilidad de presencia de la especie. Se usó un suavizado spline con intervalo de confianza al 95%. Distribución de las observaciones en bandas En ninguno de los conjuntos de datos analizados parece que exista un gradiente entre las observaciones registradas y la distancia a la carretera (Fig. 10). El 66.4% de las observaciones se ha situado entre los 500 y 1500 m de la carretera. En general, ha disminuido un 53.4% la proporción de observaciones registradas en la banda de 0 a 500 m durante la construcción de la autopista y un 56.9% después, habiendo aumentado en la banda de 500 a 1000 m un 34.3% y un 54.6% respectivamente. Los datos anuales muestran diferencias significativas entre la fase y las bandas (X2 = 21.17, g.l. = 6, p = 0.002). En este caso, antes de la construcción de la autopista se observaba una distribución de las observaciones bastante compensada entre las bandas, pero ya en la construcción la proporción de observaciones encontradas en la banda próxima a la carretera se reduce a más de la mitad y se incrementa en la banda de 500 a 1000 m. Al igual que en los análisis anteriores de modelos GAM si se exploran las variaciones estacionales en el efecto de la distancia a la carretera aparecen diferencias. Las dos temporadas en los que no se encontraron diferencias significativas entre la fase y la banda fue en primavera (X2 = 4.29, g.l. = 6, p = 0.637) y en julio (X2 = 5.89, g.l. = 6, p = 0.4356). En este último se registraron sólo un 10.7% de las observaciones antes de la construcción de la carretera en las zonas próximas a la carretera. Esa proporción se reduce 19 0,5 Invierno 0,4 0,4 0,3 0,3 0,2 0,2 0,1 0,1 0 Frecuencia relativa 0,5 Primavera 0 Septiembre Julio 0,4 0,4 0,3 0,3 0,2 0,2 0,1 0,1 0 0 Anual Familias 0,4 0,4 0,3 0,3 0,2 0,2 0,1 0,1 0 0 ANTES DURANTE DESPUÉS 0-500 m 500-1000 m ANTES 1000-1500 m DURANTE DESPUÉS 1500-2000 m Figura 10. Frecuencias relativas de observaciones registradas por banda para los tres periodos de estudio con cada conjunto de datos (anuales, estacionales y de familias). cuando se inician las obras (5.1%) y una vez se inaugura la autopista vuelve a ser prácticamente igual de utilizada (9.7%). En invierno las diferencias fueron significativas (X2 = 9.669, g.l. = 3, p = 0.022) y en septiembre prácticamente también (X2 = 12.36, g.l. = 6, p = 0.054), apuntando en la misma dirección que los GAMs. En el análisis de los datos invernales sólo se tuvieron en cuenta las fases anterior y posterior, porque para la fase de construcción la muestra de observaciones resultó insuficiente. En el caso de las familias las diferencias entre las bandas y las fases resultaron muy significativas (X2 = 24.31, g.l. = 6, p < 0.001), encontrándose una gran reducción de la utilización de esta banda en las fases de construcción y funcionamiento de la autopista. 20 Tendencias poblacionales de las zonas de impacto y control Los índices de población calculados para 600 ZC2 500 cada zona por los modelos TRIM 400 ajustados (Fig. 11), arrojan resultados desiguales para la fase previa y posterior 300 a la construcción. 200 100 La fase previa a la construcción de la 0 autopista (1998-2001) coincide con un 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 Número de individuos 600 ZC1 aumento del número de individuos en la ZI y en la ZC2 hasta el año 2000. 500 400 Un año después, el número de 300 individuos desciende en todas las zonas. 200 En septiembre de 2001 se inicia la 100 construcción de la carretera y desde ese 0 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 600 IMPACTO ZI 500 año hasta su inauguración en octubre de 2003 se aprecia un crecimiento poblacional en las tres zonas, si bien es 400 más evidente en la ZI donde pasa de 203 300 a 363 individuos y en la ZC2 que 200 aumenta de 281 a 411 individuos. La 100 ZC1, en general, presenta variaciones de menor magnitud, aumento el número de 0 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 efectivos de 163 a 197. Figura 11. Tendencias del tamaño poblacional estimado como la suma de individuos, derivados de los índices predichos por TRIM para cada zona. Las flechas indican años con un cambio significativo de la tendencia (ver “Materiales y métodos”). Todos los modelos se ajustan a los datos (p > 0.2) y son los mejores con incrementos de AIC de 166 (ZI), 39.3 (ZC1) y 74.3 (ZC2). Las líneas punteadas corresponden a los momentos de inicio de la construcción e inauguración de la R2. Durante la fase posterior a la construcción las tendencias presentaron menor variabilidad (menor número de puntos de cambio significativos). Así pues, la población de la ZI descendió notablemente de 363 a 179 individuos (2003-2008), mientras que en la ZC1 la población 21 aumentó y se mantuvo en torno a 220 individuos (2003-2008) y la ZC2 la población siguió una tendencia creciente hasta 567 individuos (2003-2009). El último año (2009) se registró un cambio de tendencia en la zona de impacto y ZC1. Todos los modelos construidos con el conjunto de datos que incorporaron la covariable “zona” indicaron que era muy significativa y mejoraron el ajuste del modelo a los datos observados (Tabla 3). Además, según los valores del AIC todos los modelos que no la introducen son bastante peores que los que la tienen en cuenta. Tabla 3. Resumen de los modelos realizados. Valores significativos de la Likelihood Ratio indican que el modelo no se ajusta a los datos, mientras que por encima de p = 0.05 el modelo se ajusta. Se muestran los resultados del test de Wald que indicó la significación de la covariable en los modelos que la incorporaron. Modelo Sin covariable 3 años de cambio 2 años de cambio años de cambio por etapas Con covariable 3 años de cambio 2 años de cambio años de cambio por etapas AIC Likelihood Ratio p-valor (LR) Años de cambio de la tendencia Test Wald p-valor (W) 336.4 342.4 396.37 404.39 <0.001* <0.001* 1998,2001,2003 - - 239.3 297.31 <0.001* - - 64.9 127.3 112.87 181.28 <0.001* <0.001* 33.88 26.58 <0.001* <0.001* -7.1 4.92 0.554 400.44 <0.001* 1998,2003 1998,1999*,2001*, 2003* 1998,2001,2003* 1998,2003* 1998*,1999*,2000*, 2001*,2002*,2003*, 2005*,2006*,2008* Códigos de significación: * valores significativos. El único modelo que se ajustó a los datos observados fue el que incorporaba la covariable “zona” y en el que se habían seleccionado los años de cambio de la tendencia significativos por etapas, de tal forma que solo ha excluido dos años, 2004 y 2007. Si los modelos sólo incorporan 2 o 3 puntos de cambio correspondientes a las fases los modelos no se llegan a ajustar a los datos. En cuanto a los valores obtenidos del AIC aunque suelen ser positivos también se pueden obtener valores negativos, en ese caso el mejor modelo es el más negativo. Por tanto el mejor modelo es el que ha considerado casi todos los puntos de cambio (Fig. 12.a), seguido del modelo con 3 puntos de cambio (Fig. 12.b) y finalmente el de 2, que es el peor de los modelos que consideran la zona. 22 Índices del Modelo TRIM ANTES DURANTE DESPUÉS ANTES DURANTE DESPUÉS 2 a 1,8 b 1,6 1,4 1,2 1 0,8 0,6 1998 2000 2002 2004 2006 2008 1998 2000 2002 2004 2006 2008 Figura 12. Índices de población predichos por los modelos TRIM para los tres niveles de la covariable “zona”: zona de impacto (línea discontinua), zona control 1 (línea verde) y 2 (línea azul). El modelo a) es el que mejor se ajusta a los datos observados (p = 0.554) y procede de la selección de puntos de cambios de tendencia significativos por etapas, y el modelo b) con los puntos de cambio seleccionados para cada etapa (1998: antes, 2001: durante, 2003: después) que no se ajusta a los datos (p < 0.001). Efecto de la zona estadísticamente significativo en ambos modelos (p < 0.001). Para el mejor modelo se han calculado las tasas de cambio (Tabla 4). En la mayoría de los casos son anuales y solo han sido bianuales cuando no ha habido cambios significativos en la tendencia de un año a otro (2004 y 2007). Tabla 4. Resultados del modelo de tendencias poblacionales con TRIM que incorpora la covariable “Zona” y se ha seleccionado los puntos de cambio de tendencia significativos por etapas, eliminado exclusivamente 2004 y 2007. Valores positivos de la tasa de cambio indican incremento de efectivos y valores negativos indican descenso. Periodo Tasa de cambio Impacto (%) Tasa de cambio ZC1 (%) Tasa de cambio ZC2 (%) Test Wald p-valor 1998-1999 1999-2000 2000-2001 2001-2002 2002-2003 2003-2005 2005-2006 2006-2008 2008-2009 74 -3 -33 9 63 -14 2 -17 12 3 -19 -9 56 -19 8 2 -6 -32 14 -44 -28 24 17 1 30 -2 10 47.26 22.14 16.67 38.59 27.66 58.75 17.66 23.23 18.24 0.000* 0.000* 0.001* 0.000* 0.000* 0.000* 0.001* 0.000* 0.000* Códigos de significación: * valores significativos. 23 Parece que el inicio de la construcción no ha supuesto un cambio de signo de la tendencia, con un balance de crecimiento en todas las zonas. Sin embargo, el año de inauguración si ha supuesto un cambio importante de la tendencia en la ZI, a diferencia de las zonas control. Productividad A lo largo del periodo de estudio se producen grandes oscilaciones del número de familias en todas las zonas (Fig. 13), por lo que las desviaciones típicas de los datos son elevadas. Sin embargo se observa un comportamiento bastante parecido en todas las zonas, de tal forma que años en los que aumenta el número de familias generalmente va a coincidir la situación en las tres zonas, aunque difiriendo en la magnitud. El número total de familias registrado en la zona de impacto durante el periodo de estudio fue de 129, notablemente inferior que en los controles (237 en ZC1 y 289 en ZC2). No se observaron diferencias significativas en el número de familias de las distintas fases (resultados del ANOVA simple: F=0.46; p=0.767) en ninguna de las zonas, ya que la 1999 2001 2003 ZC2 ZC1 2005 2007 25 20 15 10 50 45 40 35 30 25 20 15 10 5 0 1997 de familias del número Media de familias Número Número de familias interacción no resultó significativa (resultados del ANOVA simple: F=0.02; p=0.979). 2009 Impacto ZC1 ZC2 Impacto Figura 13. Tendencias del número de familias en el periodo Figura 14. Valores medios en de estudio para las zonas control y la zona de impacto. intervalos de confianza al 95% del número de familias en cada zona durante el periodo de estudio. Aun así, el número medio de familias fue menor en la fase posterior a la construcción tanto en la zonas de impacto (de 10 a 8 familias/año) como en ZC1 (de 20 a 17 familias/año), mientras 24 que en ZC2 el número se incrementó (de 21 a 24 familias/año). Durante la fase de construcción el número de familias se incrementó en la zona de impacto (14 familias/año) y disminuyó en las zonas control (17 familias/año en ZC1 y 19 familias/año en ZC2). El factor “zona” resultó significativo (resultados del ANOVA simple: F=7.73; p=0.002), siendo la zona de impacto significativamente diferente de las dos zonas control, pero no éstas entre sí (Fig. 14). No obstante, el ajuste a los datos fue débil (R2 = 0.37). Extinciones locales Tras la construcción de la R2 se han registrado los siguientes procesos de extinción local en la zona de impacto (banda de 2 km; Fig. 15): (1) Paracuellos de Jarama, que era utilizada como área de invernada y de reproducción por unas 25 aves, debido a la pérdida directa de hábitat por la construcción de una urbanización que ha ocupado la mayor parte de la superficie en la que se localizaban individuos. Unido a ello, la construcción de la R2 ha delimitado un pequeño espacio de terreno agrícola de poco más de 1 km de ancho. El último contacto se detectó en octubre de 2004, una hembra con dos pollos. Y (2) sur de Meco, zona utilizada por 21 avutardas hasta 2004. La última observación es de un bando de 14 hembras en marzo de 2005. En las zonas de control no se registraron extinciones locales de la especie. Figura 15. Localización de los núcleos desaparecidos desde la construcción de la carretera (color verde). La línea negra discontinua corresponde a la banda de 2 km a la R2 que delimita la zona de impacto y las zonas con la malla son superficies de desarrollo urbano en los últimos años. 25 Discusión Distancia a la autopista La zona en la que se ha construido la R2 parece que, aunque era utilizada de manera habitual, no concentraba la mayor parte de las observaciones, sino que se distribuían a lo largo de los 2000 m que se han escogido como área de estudio. Con la construcción de la autopista, el hábitat utilizable por la especie, en la zona sur de la misma, no llega más allá de una distancia de 345 m desde la infraestructura, por lo que es probable que acceder a estas zonas no les resulte rentable. De hecho, una vez iniciada la construcción de la R2 no se han vuelto a registrar observaciones en la zona sur de la autopista, excepto en el núcleo de Paracuellos de Jarama, hasta su desaparición en 2004. Las mayores modificaciones en la distribución de la especie tuvieron lugar en los primeros 1000 m de distancia. En la fase de construcción y funcionamiento de la R2 podría haberse producido un desplazamiento de la especie de la banda de 0-500 m a la de 500-1000 m, ya que en la primera disminuyó considerablemente el número de observaciones mientras que en la segunda aumentaron, situándose la distancia umbral en torno a los 600 m en la fase de construcción y 500 m en la fase de funcionamiento, aunque estos valores presentaron diferencias según el conjunto de datos considerado. A partir de los 1000 m las oscilaciones encontradas en los modelos GAM podrían estar más asociadas a otras variables ambientales que estuvieran determinando la probabilidad de presencia, ya que no responden a un gradiente en el que la probabilidad de presencia aumente de forma continua de 0 a 2000 m. La distancia umbral hallada es superior a la encontrada en un estudio realizado en zonas periurbanas, con una intensidad media diaria de vehículos comparable (10000 vehículos/día), en la que el efecto en aves de medios abiertos se extendía hasta los 365 m (Forman y Deblinger, 2000). De forma similar, en estudios sobre el uso del espacio de la avutarda que consideraron variables relacionadas con las carreteras se describen distancias mínimas de alejamiento de 100 m en carreteras poco transitadas de Navarra (Onrubia et al., 1998) y de 300 m en carreteras transitadas de Valladolid (Pérez et al., 1990, citado en Onrubia et al., 1998). Además se han realizado estudios con especies de la misma familia (Otididae), que también habitan medios abiertos (esteparios y semidesérticos), el sisón común y la avutarda hubara (Chlamydotis undulata; Jacquin, 1748). En una población de sisón común en Portugal se observó que el efecto de una autopista era notable hasta los 600 m (Dias, 2004) y en estudios de selección de hábitat de la avutarda hubara también se demostró la preferencia por 26 áreas alejadas a más de 500-600 m de vías de comunicación (Le Cuziar et al., 2005; Carrascal et al., 2006). Estas cifras se ajustan bastante a las encontradas para la avutarda común en la R2, pareciendo que existe un rechazo común de las especies de esta familia a las zonas próximas a carreteras. Los modelos GAM que se realizaron con las distancias a la R2 para cada temporada fueron interesantes no sólo para detectar la existencia de efectos, sino para demostrar la existencia de una variabilidad estacional de la respuesta dentro de una misma especie. Este aspecto puede estar relacionado con la selección de hábitat diferencial de la avutarda a lo largo del año, de tal forma que selecciona activamente el microhábitat (parcela), eligiendo en cada momento del año la más apropiada por la abundancia de recursos tróficos, pero teniendo en cuenta el reparto temporal de otras actividades (reproducción, reposo, alimentación) y la presencia de molestias (Palacín, 2007). En invierno el efecto de la distancia fue bastante claro, de tal forma que los individuos, tanto en la fase de construcción como de funcionamiento, tendían a evitar una distancia de 600 m desde la R2. Durante esta temporada las avutardas, al igual que otros animales gregarios que viven en agrosistemas, están alimentándose para fortalecerse de cara al periodo reproductor, por lo que es posible que busquen zonas tranquilas, alejadas de la carretera, para desempeñar esta tarea. Al igual que la avutarda, el sisón tiende evitar la proximidad a las carreteras en temporada invernal (Silva et al., 2004). También se ha demostrado que las avutardas migradoras del centro peninsular seleccionaron en esta época del año lugares más alejados de las carreteras que en las zonas de reproducción (Palacín, 2007). En cambio, en la época reproductora ninguno de los modelos construidos fue lo suficientemente explicativo como para considerar que hay un efecto importante de la distancia. Se trata de la temporada en la que menos influye el efecto de la distancia a la autopista en la distribución de la población. Este resultado no es compartido con otros estudios, en los que la distancia a carreteras sí fue un criterio significativo de la presencia de aves forestales (Palomino y Carrascal, 2007) y de las zonas de reproducción del alcaraván común (Burhinus oedicnemus; Linnaeus, 1758; Green et al., 2000), que se encuentran a distancias superiores a 3 km. En el caso de avutarda, durante esta temporada los machos dedican casi la totalidad del tiempo a la exhibición, mientras que la búsqueda de alimento queda reducida a unas pocas horas al día. Así que, es probable que los efectivos presentes en el área de estudio durante la época nupcial estén priorizando su fidelidad a las zonas de 27 exhibición, cuya ubicación podría estar condicionada por otros factores ambientales (datos propios sin publicar). Esto supone que durante esta temporada soportarían las molestias asociadas a la autopista, manteniendo su distribución tradicional. Estas diferencias entre invierno y primavera coinciden con los estudios de áreas de invernada de hembras migradoras en el centro peninsular (Palacín, 2007), en los que se detectaron distancias mínimas mayores en invierno (1324 m) que en primavera (1027 m), con un registro de distancia mínima de 424 m, aunque las diferencias no fueron significativas. En hembras migradoras, en cambio, sí aparecen diferencias significativas (1697 m en invierno y 1065 m en primavera). Sin embargo, hay que tener en cuenta que en ese trabajo se consideraron varias tipos de carreteras, no sólo autopistas, y que la densidad de estas vías podría influir bastante en el resultado final. En julio, según la distribución que presentaba la especie en el área de estudio, previa a la construcción de la R2, los contactos ya se situaban a una cierta distancia de donde posteriormente se ubicaría la autopista. De tal forma que los cambios se han encontrado entre los 1000-2000 m, pero se ha mantenido una probabilidad de presencia baja hasta los 500-600 m de distancia y alta en torno a los 1000 m, que parece ser la zona de preferencia de los grupos de hembras y las familias que se encuentran en la zona durante esta temporada, siendo escasa la presencia de machos, que después de la estación reproductora migran a zonas de temperaturas más suaves a pasar el verano, generalmente fuera de la Comunidad de Madrid (Palacín et al., 2009). En septiembre, la situación es parecida a la de invierno, con un área de exclusión un poco más reducida en torno a los 400 m. En esta temporada también se confirmó que los grupos familiares evitaron la carretera, en mayor medida que el conjunto de individuos. Así pues, existiría un gradiente tal que la probabilidad de presencia, muy baja hasta los 500 m aumentaría progresivamente hasta los 1200 m, donde ya se mantendrían niveles elevados. Este comportamiento podría estar asociado a una mayor búsqueda de tranquilidad por parte de las madres para cuidar a sus pollos, alejadas en la medida de lo posible de molestias y/o amenazas. Entre las posibles causas que podrían explicar el efecto de la distancia a la carretera, el tráfico y el ruido, han sido considerados los factores más determinantes de los patrones de aves (van der Zande, 1980; Forman et al., 2002). Sin embargo, los niveles de ruido en la zona son muy elevados, dada la proximidad de los aeropuertos de Torrejón de Ardoz y de Barajas, por tanto este factor podría no ser la causa directa de la ausencia de individuos en la banda más próxima 28 a la carretera. En cuanto a la intensidad de tráfico, variaciones en la misma podrían estar relacionadas con la tendencia a evitar zonas próximas a la carretera. Dicha intensidad, no presenta grandes oscilaciones a lo largo del año (7437±3152 vehículos/día en enero, 9009±3724 vehículos/día en marzo, 10116±3218 vehículos/día en julio y 9321±3100 vehículos/día en septiembre), por lo que parece que la variabilidad estacional se asociaría más a una respuesta a los requerimientos de la especie. Además, cabe la posibilidad de que la calidad del alimento hubiera empeorado en las zonas próximas a la carretera como consecuencia de la contaminación asociada a la misma (Tabor, 1974; Bolsinger y Flückinger, 1989; Forman y Alexander, 1998). En este sentido, las temporadas en las que la alimentación constituye el principal objetivo, las aves se desplazarían a zonas donde la calidad fuera mayor. Finalmente, a pesar del reducido porcentaje de variabilidad explicada en los modelos, los resultados parecen satisfactorios teniendo en cuenta que sólo se ha considerado una variable: la distancia a la R2. La presencia/pseudoausencia podría no ser estrictamente una variable binomial, quizás de esta forma se incrementara la variabilidad que pudieran explicar los modelos. Abundancia y productividad En la serie temporal estudiada las tendencias demográficas han presentado bastantes cambios significativos de pendiente, lo que ha supuesto que los modelos realizados no se ajustaran a los datos exclusivamente con los años de cambio correspondientes a las fases. Las fluctuaciones podrían ser explicadas, al menos en parte, por la notable variabilidad del éxito reproductor en esta especie (Alonso et al., 2003; Palacín, 2007), como se desprende del análisis de los grupos familiares. Por tanto es necesario disponer de varios años para cada periodo, tanto antes como después de la construcción, si se pretenden alcanzar resultados consistentes. De otra forma los resultados obtenidos podrían conducir a error. Aún así la gran variabilidad ha supuesto una mayor dificultad a la hora de analizar estos factores y extraer conclusiones. La puesta en funcionamiento de la autopista podría haber tenido un efecto negativo en la ZI, ya que desde la inauguración de la R2 ha habido un descenso progresivo de la abundancia de avutardas comunes en esta banda (50.1%). Tal situación parece atribuible a la autopista porque las tendencias en las zonas control para este periodo han sido diferentes. En la zona 29 control más próxima (ZC1) el número de efectivos se mantuvo en cierta estabilidad (tasas de cambio no superiores al 8%), con un ligero incremento del número de individuos hasta la primavera de 2008. En la zona control más alejada (ZC2) el tamaño poblacional se incrementó un 27.6% de forma escalonada, llegando a concentrar 567 individuos en 2009. Esa forma de crecimiento probablemente haya estado relacionada con las altas productividades de 2004 y 2007, que se habrían traducido en el crecimiento del número de efectivos en 2006 y 2009, respectivamente, ya que un intervalo de 2 años corresponde, aproximadamente, al tiempo de dispersión juvenil de la especie (Martín, 2001). Por tanto, los resultados sugieren que si no se hubiera construido la R2 se habría mantenido o incrementado. En el último año se produjo un cambio en el signo de la tendencia de la ZI (+12%) y la ZC1 (-32%), que podría estar relacionado con un flujo de individuos entre estas zonas y la banda de 2 km que las separa, puesto que ambas pertenecen a la misma unidad. En cambio, durante la fase previa y de construcción de la autopista, las tendencias poblacionales han sido más parecidas entre zonas. Las fluctuaciones encontradas en el periodo anterior a la construcción son comunes con las registradas para el conjunto de poblaciones del centro peninsular (Martín, 2009), por lo que la elección de las zonas control parece que ha sido correcta. Esta fase se caracterizó por un periodo de crecimiento hasta 1999, relacionado con el éxito reproductivo de 1997 (73 familias en el área de estudio). A continuación, se produjo un gran descenso en 2001, como consecuencia de los valores bajos de productividad del año anterior (17 familias; Alonso et al., 2003). En la fase de construcción se produjo un gran aumento de la abundancia de individuos en la ZI (44.1%) y en la ZC2 (31.6%), mientras que la ZC1, que en general presentó menor variabilidad, el incremento total de individuos fue menor (17.3%). Por tanto, parece que las obras de construcción de la R2 no han tenido un efecto, al menos inmediato, sobre la abundancia de individuos. Ya que las tendencias encontradas en las dos zonas control difieren sustancialmente, parece confirmarse la necesidad de seleccionar controles a escalas diferentes, porque a priori suele desconocerse el alcance de los efectos y, por tanto, la escala de trabajo adecuada. En cuanto a los resultados de los grupos familiares, a pesar de ser un factor que ha presentado mucha variabilidad, parece que los momentos de alta y baja productividad han coincidido entre zonas. Respecto a la división en fases, no se detectaron grandes cambios en el número medio de familias en ninguna zona, aunque en la fase posterior de la R2 se encontraron valores más bajos en la ZI y la ZC1, posiblemente extensibles a toda la unidad, mientras que 30 en la ZC2 el número de familias fue ligeramente superior, coincidiendo con un mayor número de efectivos totales. Estos resultados son parecidos a los encontrados para el mosquitero musical (Phylloscopus trochilus; Linnaeus, 1758), en el que no se detectaron diferencias en el éxito reproductivo entre zonas de impacto y control, si bien el número de pollos también fue inferior en las proximidades de la carretera (Reijnen y Foppen, 1994). Por tanto, a pesar de la presencia de la carretera parece que algunas hembras siguen criando con éxito. En la fase de construcción el número de familias fue relativamente alto en la ZI, situación que se mantuvo hasta 2004. Sin embargo, a diferencia de cómo suele ser habitual, el incremento de la productividad no supuso un incremento del número de individuos en la ZI. No obstante, el número de familias es bastante inferior a los cambios producidos en el tamaño poblacional, así que este factor sólo explicaría una pequeña parte. Parece que la tendencia poblacional, además de estar seguramente modelada por cambios ambientales, también se ha visto afectada negativamente por la construcción de la autopista. La magnitud de los cambios en las tendencias poblacionales no puede ser exclusivamente atribuida a la productividad existente en cada zona, así que deben buscarse otras explicaciones. Según Fahrig y Ritwinski (2009) hay dos categorías principales de especies que muestran efectos negativos de las carreteras en la abundancia: aquellas vulnerables a las perturbaciones del tráfico y las vulnerables a la mortalidad de carreteras. Este aspecto es fundamental para saber si la especie evita la autopista o si la disminución de efectivos es atribuible a la mortalidad por atropello (Karlsson et al., 2007). Prácticamente la totalidad de vertebrados terrestres son susceptibles de sufrir bajas por esta causa, pero casos en especies como la avutarda son esporádicos y de repetición impredecible. Muestra de ello fue un informe elaborado a escala nacional en el que se detectaron atropellos en 251 especies de vertebrados pero no avutardas (PMVC, 2003). Por tanto, esta especie pertenecería al primer grupo. Las poblaciones que viven en zonas afectadas por carreteras son menos propensas a recibir individuos de otras zonas (Jaeger et al., 2005). Este aspecto podría explicar el descenso poblacional en la ZI, porque los jóvenes de avutarda común, al igual que otras especies, cuando se dispersan tienden a asentarse como adultos en zonas de mejor calidad (Reed y Dobson, 1993; Danchin y Wagner, 1997). Por tanto, es posible que la ZC2, esté siendo seleccionada por los individuos como zona reproductora por presentar una calidad de hábitat 31 relativamente mejor, ya que se encuentra más alejada de la carretera y de las perturbaciones asociadas. Este comportamiento fue comprobado en un estudio con mosquitero musical, en el que los machos de zonas con hábitat de baja calidad, inducido por una autopista, se dispersaban cuantitativamente más para reproducirse que los de zonas de alta calidad (Reijnen y Foppen, 1994). En realidad, la cantidad del hábitat utilizable para la especie que se habría perdido debido a la construcción y a los efectos de la infraestructura estaría en torno 25.6% en la ZI y a un 6.9% si tenemos en cuenta la unidad demográfica (calculada considerando que el efecto de la distancia alcanza, al menos, los 500 m, y tomando la superficie de las zonas correspondientes a las extinciones locales). Por tanto, parece que hay una pérdida sustancial de hábitat en la ZI que movería a la especie a buscar zonas más extensas. Junto con la calidad de los territorios, el patrón de atracción de coespecíficos detectado en esta especie (Lane et al., 2001) podría ser uno de los factores más potentes que explicaran el crecimiento poblacional de la ZC2. No obstante, no puede descartarse la entrada de nuevos ejemplares desde poblaciones exteriores a la IBA (como las existentes al sur de la Comunidad de Madrid o en Toledo). Sin embargo, en la fase de construcción de la autopista también parecía esperable encontrar efectos en la abundancia o en la productividad y en cambio las tendencias se mantienen, con valores positivos. A este respecto caben dos posibilidades, que las avutardas no perciban como molestas las actividades realizadas en el proceso de construcción (las avutardas están adaptadas y toleran a la presencia de maquinaria agrícola; Sastre et al., 2009) o bien que haya un retraso en la respuesta que repercuta sobre la tendencia poblacional de la especie. En otros estudios con aves de medios abiertos se ha sugerido que el tráfico de vehículos, y no la presencia de la carretera en sí, puede ser la causa principal de los efectos observados (van der Zande et al., 1980; Illner, 1992). Pero la construcción también conlleva el tráfico de camiones, movimiento de tierras y demás actividades que podrían considerar molestas. En cuanto al retraso en la respuesta, se ha estudiado que la mayor probabilidad de las especies de asentarse en zonas donde estén presentes sus congéneres probablemente no les favorecería en un rápido ambiente cambiante, de tal forma que podría haber un cierto periodo temporal en el que los individuos seguirían asentándose en hábitats que fueron tradicionalmente de buena calidad pero que han empezado a deteriorarse (Danchin y Wagner, 1997). Por tanto, una vez que el deterioro es más patente y la especie empieza a percibir cambios notorios en la calidad del medio, podría iniciarse un flujo de individuos hacia otras zonas. En este sentido resultaría 32 trascendental la dimensión temporal para evaluar los efectos de la autopista, de otra forma se estimaría el efecto a corto plazo pero se subestimarían los efectos a largo plazo (Houlahan y Findlay, 1997). En cuanto a las extinciones locales detectadas, probablemente no sean directamente atribuibles a la construcción y funcionamiento de la autopista. Sin embargo, es conocido que el desarrollo de grandes infraestructuras promueve otras alteraciones humanas, como el desarrollo paralelo de urbanizaciones, que además irá asociado a una serie de perturbaciones (Bolger et al., 1997; Hennings y Edge, 2003; Fraterrigo y Wiens, 2005). En el caso de Paracuellos se produjo una pérdida directa de hábitat utilizable en torno al 50%, poco después de la inauguración de la R2, quedando el espacio restante de hábitat utilizable distribuido en una banda de aproximadamente 1 km, que podría ser insuficiente para satisfacer los requerimientos ecológicos de la especie. La desaparición del núcleo de la zona sur de Meco podría estar más asociada a la construcción de la R2, ya que no se ha llevado a cabo un gran desarrollo urbanístico en las inmediaciones. El hecho de que las últimas observaciones se hayan detectado en 2004 y 2005, es decir, 3-4 años después del inicio de las obras y 1-2 tras la apertura del tráfico, sugiere que podría haber un retraso temporal entre la construcción de la R2 y el conjunto de extinciones locales (Findlay y Bourdages, 2000). Es poco probable que las avutardas regresen a estas áreas por el marcado patrón de atracción de coespecíficos, pues limita la capacidad de la especie para establecerse en un nuevo territorio aunque el hábitat sea adecuado, restringiendo su distribución a los lugares tradicionales de ocupación (Lane et al., 2001). Desde el punto de vista de la conservación es muy interesante conocer las consecuencias de la autopista a nivel poblacional, sobre todo en una zona de distribución de avutardas amenazada por el desarrollo urbanístico y el trazado de infraestructuras, a pesar de tratarse de un espacio protegido. Los resultados apuntan que la zona de impacto y posiblemente la unidad demográfica de Daganzo-Camarma está disminuyendo, y se está produciendo una concentración de individuos en la unidad de Valdetorres-Talamanca. Esta circunstancia puede conllevar una menor variabilidad genética, con más posibilidades de endogamia. Además incrementa el riesgo de la población por estocasticidad demográfica. Los resultados obtenidos tienen al menos 5 implicaciones importantes de cara su aplicabilidad: 1) la contribución al conocimiento de la tolerancia de la especie a las carreteras y, a su vez, de la sensibilidad de la distribución espacial y de varios parámetros demográficos; 33 2) dado que para una rápida detección de los efectos deberían seleccionarse los atributos que muestren respuestas más rápidas a estresores de origen humano (Findlay y Bourdages, 2000), en este caso los efectos se manifestaron primero en la banda de exclusión, a continuación en la abundancia y posteriormente (también más dudosa) en el número de grupos familiares; 3) parece razonable que al construir una carretera próxima a una poblaciones de avutarda amenazada se mantenga una distancia mínima de 600 m, para reducir o evitar los efectos de la carretera, sobre todo si se trata de una vía de gran capacidad; 4) se recomienda la protección de las zonas de exhibición de la ZI próximas a la carretera, que estarán sometidas a mayor estrés, pudiendo tener consecuencias en el apareamiento y la nidificación; 5) incremento del cuerpo de evidencias que indican que las poblaciones podrían exhibir un retraso sustancial en la respuesta a los efectos (Doak, 1995; Lignon y Stacey, 1996). No obstante, hay que ser conscientes de que en el planeamiento, la localización y la evaluación de alternativas de carreteras se tienen en cuenta bastantes criterios, entre ellos los beneficios socioeconómicos y otros riesgos ecológicos. Aún así, el conocimiento de los efectos encontrados debería ser tenido en cuenta en las primeras etapas de desarrollo de este tipo de proyectos. Otros estudios necesarios Este trabajo aborda los efectos de la construcción y funcionamiento de la R2 en la distribución espacial, la abundancia y la productividad de acuerdo a los objetivos y métodos explicados anteriormente. Durante su elaboración se han identificado limitaciones y nuevas cuestiones que podrían tratarse en otros trabajos. Para estudiar en mayor profundidad los efectos en la distribución espacial, una vez abordada la capacidad explicativa de los modelos GAM para las distancias, sería conveniente evaluar la capacidad predictiva de los mismos. Además, debería plantearse la incorporación de otras variables ambientales para comprobar la importancia relativa del efecto de la distancia. Además, dada la longitud del periodo analizado parece razonable abordar el análisis de los cambios de uso del suelo en las zonas de estudio para tener un mejor conocimiento de los efectos exclusivamente asociados a la construcción y funcionamiento de la R2. En particular, algunos autores (p. ej. Houlahan y Findlay, 2003) reconocen el interés de estudiar si la pérdida de hábitat en las zonas próximas a la carretera es directa, como consecuencia de las perturbaciones asociadas a la carretera, o indirecta, si el tipo de hábitat se hubiera modificado. 34 Conclusiones En resumen, las principales conclusiones de los resultados obtenidos son las siguientes: 1. La autopista ha tenido un efecto notable sobre la distribución espacial de la especie, tanto para el conjunto de individuos como para las familias, comprobando una reducción de la probabilidad de presencia a una distancia umbral que oscila entre los 400 y 750 m, en función de los requerimientos de la especie (reproductivos, alimentación y cuidado) en cada temporada. 2. La aplicación del diseño BDACI ha resultado eficaz para detectar un efecto negativo acumulativo de la construcción de la carretera en la abundancia, con una reducción del 50% en una banda de 2 km en torno a la R2, y menores efectos sobre la productividad. En ningún caso se registraron efectos positivos. 3. En la zona de impacto de la carretera han sido identificados dos procesos de extinción local de la especie, mientras que no fue detectado ningún caso en las zonas control. 4. Los efectos no parecen tener un carácter inmediato, sino que la respuesta es progresiva. En cualquier caso no se registraron efectos positivos o reversibilidad en los efectos. Bibliografía Alonso, J.C., Magaña, M., Alonso, J.A., Palacín, C., Martín, C.A. Martín, B. 2009. The most extreme sexual size dimorphism among birds: Allometry, selection and esrly juvenile development in the Great Bustard. The Auk 126(3): 657-665. Alonso, J.C., Martín, C.A., Alonso, J.A., Palacín, C., Magaña, M., Lane, S.J. 2004. Distribution dynamics of a great bustard metapopulation throughout a decade: influence of conspecific attraction and recruitment. Biodiversity and Conservation 13: 1659-1674. Alonso, J.C., Martín, C.A., Palacín, C., Magaña, M., Martín, B. 2003. Distribution, size and recent trends of the Great Bustard Otis tarda population in Madrid región, Spain. Ardeola 50(1): 21-29. Alonso, J.C., Palacín, C., Magaña, M., Martín, C.A., Ponce, C. y Bautista, L.M. 2006. La población de avutardas de la Comunidad de Madrid: censo de individuos reproductores y productividad en el año 2006. Informe inédito para la Consejería de Medio Ambiente y Ordenación del Territorio de la Comunidad de Madrid. Madrid. Alonso, J.C., Palacín, C., Martín, C.A. 2005. La Avutarda Común en la Península Ibérica: población actual y método de censo. SEO/BirdLife. Madrid. Beyer, H.L. 2004. Hawth’s Analysis Tools for ArcGis. Dispoible en: http:www.sapatialecology.com/htools. Acceso el 15/04/2009. 35 BirdLife International. 2004. Threatened Birds of the World 2004. CD-ROM. BirdLife International. Cambridge. Bolger, D.T., Scott, T.A., Rotenberry, J.T. 1997. Breeding bird abundance in an urbanizing landscape in coastal southern California. Conservation Biology 11: 406-421. Bolsinger, M. y Flückinger, W. 1989. Ambient air pollution induced changes in amino acid pattern of phloem sap in hostplants: relevance to aphid infestation. Environmental Pollution 56: 209-216. Bro, E., Mayot, P., Corda, E., Reitz F. 2004. Impact of habitat management on grey partridge populations: assessing wildlife cover using a multisite BACI experiment. Journal of Applied Ecology 41: 846-857. Burnham, K.P., Anderson, D.R. 2002. Model selection and multi-model inference. A practical information-theoric approach. Springer. Nueva York. Carrascal, L.M., Seoane, J., Palomino, D., Alonso, C.L. 2006. Preferencias de hábitat, estima y tendencias poblacionales de la avutarda hubara Chlamydotis undulata en Lanzarote y la Graciosa (Islas Canarias). Ardeola 53(2): 251-269. Cody, M.L., Smallwood, J.A. 1996. Long-term Studies of Vertebrate Communities. Academic Press. San Diego. Danchin, E. y Wagner, R. 1997. The evolution of coloniality: the emergence of new perspectives. Trends in Ecology and Evolution 12: 342-347. Del Hoyo, J., Elliot, A., Sargatal, J. 1996. Handbook of the Birds of the World. Lynx. Barcelona. Dias, G.M. 2004. Impacte a Auto-Estrada A2 na Zona de Protecção Especial de Castro Verde: Efeitos na Distribuição e Densidade de Sisão (Tetrax tetrax). Mestrado en Gestão de Recursos Naturais. Universidade técnica de Lisboa. Dirección General de Carreteras, Red de carreteras, Madrid, serie 1990-2007. http://www.ine.es/jaxi.menu.do?type=pcaxis&path=/t10/a109/a01/&file=pcaxis Acceso el 03/01/2010. Doak, D. 1995. Source-sink models and the problem of habitat degradation: general models and applications to the Yellowstone grizzly. Conservation Biology 9: 1380-1395. Donald, P.E., Green, R.E., Heath, M.F. 2001. Agricultural intensification and the collapse of Europe’s farmland birds populations. Proceedings of Royal Society of London 268: 2529. Fahrig, L., Rytwinski, T. 2009. Effects of roads on animal abundance: an empirical review and synthesis. Ecology and Society 14(1): 21. Fasola, M., Rubolini, D., Merli, E., Boncompagni, E., Bressan, U. 2010. Long-term trends of heron and egret populations in Italy, and the effects of climate, human-induced mortality, and habitat on population dynamics. Population Ecology 52: 59-72. Findlay, C.S., Bourdages, J. 2000. Response time of wetland biodiversity to road construction on adjacent lands. Conservation Biology 14: 86-94. Forman, R.T.T., Alexander, L.E. 1998. Roads and their major ecological effects. Annual Review of Ecology and Systematics 29: 207-231. Forman, R.T.T., Deblinger, R.D. 2000. The ecological road-effect zone of a Massachusetts (USA) suburban highway. Conservation Biology 14: 36-46. 36 Forman, R.T.T., Reineking, B. y Hersperger, A.M. 2002. Road traffic and nearby grassland bird patterns in a suburbanizing landscape. Environmental Management 29: 782-800. Forman, R.T.T., Sperling, D., Bissonette, J.A., Clevenger, A.P., Cutshall, C.D., Dale, V.H., Fahrig, L., France, R., Goldman, C.R., Heanue, K., Jones, J.A., Swanson, F.J., Turrentine, T. y Winter, T.C. 2003. Road Ecology. Science and Solutions. Island Press. Washington. Fraterrigo, J.M., Wiens, J.A. 2005. Bird communities of the Colorado Rocky Mountains along a gradient of exurban development. Landscape and Urban Planning 71: 263-275. Green, R.E., Tyler, G.A., Bowden, G.R. 2000. Habitat selection, ranging behaviour and diet of the Stone curlew (Burhinus oedicnemus) in southern England. Journal of Zoology 250: 161-183. Guisan, A., Edwards, T.C., Hastie, T. 2002. Generalized linear and generalized additive models in studies of species distributions: setting the scene. Ecological Modelling 157: 89-100. Hastie, T.J., Tibshirani, R.J. 1990. Generalized Additive Models. Chapman & Hall. Londres. Hennings, L.A., Edge, W.D. 2003. Riparian bird community structure in Portland Oregon: Habitat, urbanization, and spatial scale patterns. The Condor 105: 288-302. Houlahan, J.E., Findlay, C.S. 2003. The effects of adjacent land use on wetland amphibian species richness and community composition. Canadian Journal of Fisheries and Aquatic Sciences 60: 1078-1094. Illner, H. 1992. Effect of roads with heavy traffic on grey partridge (Perdix perdix) density. Gibier Faune Sauvage 9: 467-480. Jaeger, J.A.G., Bowman, J., Brennan, J., Fahrig, L., Bert, D., Bouchard, J., Charbonneau, N., Frank, K., Gruber, B., Tluk von Toschanowitz, K. 2005. Predicting when animal populations are at risk from roads: an interactive model of road avoidance behavior. Ecological Modelling 185: 329-348. Johnsgard, P.A. 1991. Bustards, Hemipodes and Sandgrouse. Birds of Dry Places. Oxford University Press. Oxford. Karlsson, J., Broseth, H., Sand, H., Andrén, H. 2007. Predicting occurrence of wolf territories in Scandinavia. Journal of Zoology 272: 276-283. Lane, S.J., Alonso, J.C., Martín, C. 2001. Habitat preferences of great bustard Otis tarda flocks in the arable steppes of central Spain: are potentially suitable areas unoccupied? Journal of Applied Ecology 38: 193-203. Le Cuziat, J., Lacroix, F., Roche, P., Vidal, E., Médail, F., Orhant, N., Béranger, P.M. 2005. Landscape and human influences on the distribution of the endangered North African houbara bustard (Chlamydotis undulata undulata) in Eastern Morocco. Animal Conservation 8: 143-152. Lignon, J.D., Stacey, P.B. 1996. Land use, time lags and the detection of demographic changes: the case of the Acorn Woodpecker. Conservation Biology 10: 840-845. Martín, B. 2009. Dinámica de población y viabilidad de la avutarda común en la Comunidad de Madrid. Tesis doctoral. Universidad Complutense de Madrid. Martín, C.A. 2001. Dispersión y estructura genética de la población de avutardas de la Comunidad de Madrid. Tesis doctoral. Universidad Autónoma de Madrid. 37 Morales, M.B. 2000. Ecología reproductiva y movimientos estacionales en la avutarda (Otis tarda). Tesis doctoral. Universidad Complutense de Madrid. Newton, I. 1998. Population limitation in birds. Academic. Londres. Onrubia, A., Saénz de Buruaga, M., Osborne, P., Baglione, V., Purroy, F.J., Lucio, A.J., Campos, M.A. 1998. Viabilidad de la población navarra de avutardas. Consultora de Recursos Naturales. Vitoria. Palacín C., Alonso, J.C., Alonso, J.A., Martín, C.A., Magaña, M., Martín, B. 2009. Differential Migration by Sex in the Great Bustard: Possible Consequences of an Extreme Sexual Size Dimorphism Ethology 115: 617-626. Palacín, C., Alonso, J.C., Martín, C.A., Alonso, J.A., Magaña, M., Martín, M. 2004. Avutarda Común (Otis tarda). En: Libro Rojo de las Aves de España, Madroño, A., González, C., Atienza, J.C. (Eds), pp. 209-213. SEO/BirdLife-Dirección General de Conservación de la Naturaleza, Ministerio de Medio Ambiente, Madrid. Palacín, C., Alonso, J.C. 2008. An updated estimate of the world status and population trends of the great bustard Otis tarda. Ardeola 55: 13-25. Palacín, C. 2007. Comportamiento migratorio de la Avutarda Común en la Península Ibérica. Tesis doctoral. Universidad Complutense de Madrid. Palomino, D., Carrascal, L.M. 2007. Threshold distances to nearby cities and roads influence the bird community of a mosaic landscape. Biological Conservation 140: 100-109. Pannekoek, J., van Strien, A. 2001. TRIM 3 Manual (TRends & Indices for Monitoring data). Statistics Netherlands. Voorburg. Pérez, C., Pérez, M.F. 2005. Distribución y abundancia de la población de Avutarda Común (Otis tarda) en la provincia de Valladolid en los años 2003-2004-2005. Informe inédito para el Servicio Territorial de Medio Ambiente de la Junta de Castilla y León. Valladolid. PMVC 2003. Mortalidad de vertebrados en carreteras. Proyecto provisional de seguimiento de la mortalidad de vertebrados en carreteras (PMVC). Documentos Técnicos de Conservación 4: 1-136. Primack, R.B. 1998. Essentials of Conservation Biology. Sinauer Associates. Sunderland. R Development Core Team. 2005. R: A language and environment for statistical computing. R Foundation for Statistical Computing. Viena. Reed, J.M., Dobson, A.P. 1993. Behavioural constraints and conservation biology. Trends in Ecology and Evolution 8: 253-256. Reid, J.M., Bienal, E.M., Bienal, S., McCracken, D.I., Monagham, P. 2006. Spatial variation in demography and population growth rate: the importance of natal location. Journal of Animal Ecology 74: 1201-1211. Reijnen, R., Foppen, R. 1994. The effects of car traffic on breeding bird populations in woodland. I. Evidence of reduced habitat quality for willow warbles (Phylloscopus trochilus) breeding close to highway. Journal of Applied Ecology 31: 85-41. Reijnen, R., Foppen, R., Meeuwsen, H. 1996. The effects of traffic on the density of breeding birds in Dutch agricultural grassland. Biological Conservation 75: 255-260. 38 Reijnen, R., Foppen, R., ter Braak, C., Thissen, J. 1995. The effects of car traffic on breeding bird populations in woodland. III. Reduction of density in relation to the proximity of main roads. Journal of Applied Ecology 32: 187-202. Roedenbeck, I.A., Fahrig, L.,Findlay, C.S., Houlahan, J.E., Jaeger, J.A.G., Klar, N., KramerSchadt, S., van der Grift, E.A. 2007. The Rauischholzhausen Agenda for Road Ecology. Ecology and Society 12: 11. Sastre, P., Ponce, C., Palacín, C., Martín, C., Alonso, J.C. 2009. Disturbances to great bustards (Otis tarda) in central Spain: human activities, bird responses and management implications. European Journal of Wildlife Research 55: 425-432. Schmitt, R.J., Osenberg, C.W. 1996. Detecting ecological impacts: Concepts and applications in coastal habitats. Academic Press. Londres. Seaman, D.E., Powell, R.A. 1997. An evaluation of the accuracy of Kernel density estimators for home range analysis. Ecology 77(7): 2075-2085. Seoane, J., Carrascal, L.M. 2007 Interspecific differences in population trends of Spanish birds are related to hábitat and climatic preferences. Global Ecology and Biogeography 17: 111-121. Serrano, M., Gago, C., Antón, J. 2006. Impacto territorial de las carreteras orbitales de la Comunidad de Madrid. CERSA. Madrid. Silva, J.P., Pinto, M., Palmeirim, J.M. 2004. Managing landscapes for the Little bustard Tetrax tetrax: lessons from study of Winter hábitat selection. Biological Conservation 17: 521-528. Suárez-Seoane, S., Osborne, P.E., Alonso, J.C. 2002. Large-scale habitat selection by agricultural steppe birds in Spain: identifying species-habitat responses using generalized additive models. Journal of Applied Ecology 39: 755-71. Tabor, R. 1974. Earthworms, crows, vibrations and motorways. New Scientist 62: 482-483. Traba, J., García de la Morena, E.L., Morales, M.B, Suárez, F. 2007. Determining high value areas for steppe birds in Spain: hot spots, complementarity and the efficiency of protected areas. Biodiversity and Conservation 16: 3255-3275. Tucker, G.M., Heath, M.F., 1994. Birds in Europe: Their Conservation Status. BirdLife International. Cambridge. Underwood, A.J., Chapman, M.G. 2003. Power, precaution, Type II error and sampling design in assessment of environmental impacts. Journal of Experimental Marine Biology and Ecology 296: 49-70. van der Zande, A.N., ter Keurs, J., van der Weijden, W.J. 1980. The impact of roads on the densities of four bird species in an open field hábitat-evidence of a long distance effect. Biological Conservation 18: 299-321. Worton, B.J. 1989. Kernel methods for estimating the utilization distribution in home-range studies. Ecology 70: 164-168. Wretenberg, J, Lindström, A., Svensson, S., Pärt, T. 2007. Linking agricultural policies to population trends of Swedish farmland birds in different agricultural regions. Journal of Applied Ecology 44: 933-941. 39 Anexo Lista de colaboradores en los censos (por orden de participación) Juan C. Alonso López Carlos Palacín Moya Carlos A. Martín de la Calle Marina Magaña Ramos Beatriz Martín Morcuende Carlos Ponce Cabas Luis M. Bautista Sopelana Carolina Bravo Párraga Simon J. Lane Enrique Martín Montes Manuel B. Morales Prieto