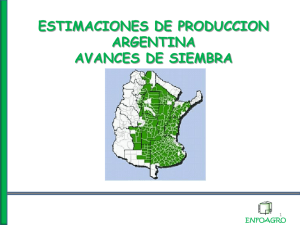
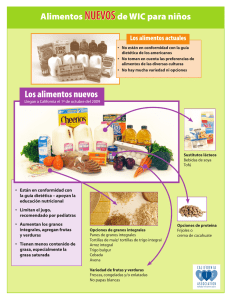
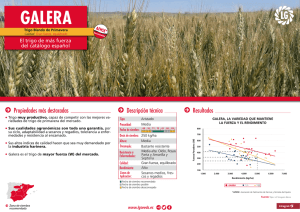
Rendimiento de Trigo: Tesis sobre Densidad de Siembra y Arreglo Espacial
Anuncio

Universidad Austral de Chile Facultad de Ciencias Agrarias Escuela de Agronomía Impacto de dos densidades de siembra y arreglo espacial contrastante sobre el rendimiento de trigo (Triticum aestivum L.) y sus determinantes fisiológicos Memoria presentada como parte de los requisitos para optar al título de Ingeniero Agrónomo Adolfo Hermann Hott González Valdivia – Chile 2022 PROFESOR PATROCINANTE: _______________________________________ Daniel Calderini R. Ingeniero Agrónomo, Dr. Cs. Agr. Instituto de Producción y Sanidad Vegetal (IPSV) PROFESOR COPATROCINANTE: _______________________________________ Patricio Sandaña G. Ingeniero Agrónomo, Dr. Agr. Instituto de Producción y Sanidad Vegetal (IPSV) PROFESOR INFORMANTE: _______________________________________ Fabián Carrasco G. Ingeniero Agrónomo. Instituto de Producción y Sanidad Vegetal (IPSV) i INDICE DE MATERIAS Capítulo Página RESUMEN 1 SUMMARY 2 1 INTRODUCCIÓN 3 2 REVISIÓN BIBLIOGRÁFICA 7 2.1 Antecedentes generales del cultivo de trigo 7 2.2 Respuesta del cultivo de trigo a la densidad de siembra 8 2.3 Intercepción de radiación de trigos con densidades y arreglos 10 espaciales contrastantes 2.4 Concentración de clorofila y proteína 12 3 MATERIALES Y MÉTODOS 15 3.1 Ubicación del experimento 15 3.2 Material vegetal 15 3.3 Tratamientos y Diseño Experimental 15 3.4 Manejo del experimento 17 3.5 Muestreos y mediciones 18 3.6 Análisis de estadístico de resultados 20 4 PRESENTACIÓN DE RESULTADOS 21 ii 4.1 Condiciones climáticas durante la temporada 2019-2020 y la 21 fenología de los genotipos 4.2 Rendimiento, biomasa, índice de cosecha y altura de planta de los 22 genotipos evaluados en ambas densidades de siembra 4.3 Componentes numéricos del rendimiento en vástagos principales y 25 macollos 4.4 Dinámica de macollaje 27 4.5 Dinámica de la captura de radiación 29 4.6 Índice de área foliar 30 4.7 Eficiencia en el uso de la radiación 31 4.8 Peso específico foliar 33 4.9 Lectura SPAD 33 4.10 Nitrógeno absorbido y concentración de proteína en granos 34 5 DISCUSION DE RESULTADOS 37 6 CONCLUSIONES 43 7 REFERENCIAS BIBILIOGRÁFICAS 45 iii INDICE DE CUADROS Cuadro 1 Página Rendimiento, biomasa aérea, índice de cosecha (IC), en PANTERA– 24 INIA y WBLL 1*2/KUKUNA creciendo bajo condiciones de densidad convencional y baja densidad. 2 Altura de planta en vástago principal (Altura VP) y, en los macollos 25 (Altura Mac) y promedio de planta (Altura de planta), en PANTERA– INIA y WBLL 1*2/KUKUNA creciendo bajo condiciones de densidad convencional y baja densidad. 3 Número de granos total por unidad de superficie (NG), peso de mil 26 granos (P1000), número de espigas por unidad de superficie y número de granos por espiga, en PANTERA–INIA y WBLL 1*2/KUKUNA en densidad convencional y baja densidad. 4 Número máximo de macollos (N° Máximo macollos) y número final de macollos (N° Final macollos), en PANTERA–INIA 29 y WBLL 1*2/KUKUNA creciendo bajo condiciones de densidad convencional y baja densidad. 5 Nitrógeno absorbido en granos, capotillos, tallos y hojas, en 35 PANTERA–INIA y WBLL 1*2/KUKUNA creciendo bajo condiciones de densidad convencional y baja densidad. 6 Concentración de proteína en grano, en PANTERA–INIA y WBLL 1*2/KUKUNA creciendo bajo condiciones de densidad convencional y baja densidad. 36 iv INDICE DE FIGURAS Página Figura 1 Respuesta de rendimiento al aumento en la densidad de plantas de un 8 cultivo de trigo. 2 EUR fotosintéticamente activa (RFA) calculada como la pendiente de la 11 regresión entre la biomasa aérea y la RFA interceptada, expresada en gramos (g) de biomasa por unidad de energía de radiación. 3 Diseño experimental del experimento. Los bloques son representados 16 por las líneas punteadas. Las parcelas principales son las líneas solidas de color azul que representan los genotipos de trigo, mientras que dentro de cada parcela se indican los niveles de densidad de siembra: 44 plantas m-2 y 300 plantas m-2. 4 Foto tomada de Ferrante et al. (2012). Distribución de semillas de trigo 17 en cintas adhesivas (a). Cintas adhesivas distribuidas en la estación experimental (b). Trigo emergido en la estación experimental (c). 5 Precipitación mensual (a) durante enero de 2019 hasta febrero de 2020 21 y radiación solar (b) y temperaturas mínima y máxima (c) desde siembra hasta cosecha del experimento. 6 Biomasa aérea desde la emergencia del cultivo en (a) PANTERA–INIA 23 (círculos) y (b) WBLL 1*2/KUKUNA (cuadrados) en densidad convencional (símbolos abiertos) y baja densidad (símbolos cerrados). 7 Número de granos m-2 en PANTERA–INIA (círculos) y WBLL 1*2/KUKUNA, (cuadrados) en función al número de espigas m -2 (a) y número de granos espigas-1. 27 v 8 Dinámica de aparición de macollos por planta desde emergencia del 28 cultivo en (a) PANTERA–INIA (círculos) y (b) WBLL 1*2/KUKUNA (cuadrados) en densidad convencional (símbolos abiertos) y baja densidad (símbolos cerrados). 9 Dinámica de la radiación interceptada en función del tiempo térmico 30 desde emergencia del cultivo en (a) PANTERA–INIA (círculos) y (b) WBLL 1*2/KUKUNA (cuadrados). En densidad convencional (símbolos abiertos) y baja densidad (símbolos cerrados). 10 Radiación interceptada y el índice de área foliar en (a) PANTERA–INIA 31 (círculos) y (b) WBLL 1*2/KUKUNA (cuadrados). En densidad convencional (símbolos abiertos) y baja densidad (símbolos cerrados). 11 Biomasa aérea y la radiación interceptada acumulada desde 32 emergencia hasta antesis en (a) PANTERA–INIA (círculos) y (b) WBLL 1*2/KUKUNA (cuadrados) y desde antesis hasta madurez de cosecha en (c) PANTERA–INIA (círculos) y (d) WBLL 1*2/KUKUNA (cuadrados). En densidad convencional (símbolos abiertos) y baja densidad (símbolos cerrados). 12 Peso específico foliar hasta madurez fisiológica en (a) PANTERA–INIA 33 (círculos) y (b) WBLL 1*2/KUKUNA (cuadrados). En densidad convencional (símbolos abiertos) y baja densidad (símbolos cerrados). 13 Lectura SPAD hasta madurez fisiológica en (a) PANTERA–INIA 34 (círculos) y (b) WBLL 1*2/KUKUNA (cuadrados). En densidad convencional (símbolos abiertos) y baja densidad (símbolos cerrados). 14 Número de espigas por metro cuadrado en PANTERA–INIA y WBLL 1*2/KUKUNA. En densidad convencional y baja densidad. 38 vi 15 Componentes numéricos del rendimiento en PANTERA–INIA (círculos) y WBLL 1*2/KUKUNA (cuadrados), en función al número de granos m-2 (a) y peso de 1000 granos m-2. 40 1 RESUMEN El trigo (Triticum aestivum L.) es uno de los cultivos más importantes a nivel mundial debido a su contribución a la alimentación humana y a su capacidad adaptativa a casi todas las regiones del planeta. En Chile, el trigo es el cultivo más importante y tradicional de la agricultura del país, en términos de superficie sembrada y producción. Experimentos en la estación experimental de la Universidad Austral de Chile, con densidades de siembra muy bajas (44 pl m-2) no mostraron diferencias en el rendimiento. El rendimiento de granos resulta de un óptimo balance de tres componentes: el número de espigas por área, el número de granos por espiga y el peso individual de los granos. En la presente tesis, se realizó un experimento en la estación Experimental Agropecuaria Santa Rosa (Universidad Austral de Chile), con el objetivo de cuantificar el efecto de dos densidades de siembra contrastantes, con diferente arreglo espacial, sobre el rendimiento en dos genotipos de trigo. El experimento se realizó, bajo manejo agronómico óptimo y en las condiciones de alto potencial de rendimiento del sur de Chile. En el experimento se utilizaron dos genotipos de trigo pandero primaveral, los cuales corresponden a una línea de la población WAMI (Wheat Association Mapping Initiative), WBLL1*2/KUKUNA y a la variedad comercial PANTERA-INIA (Instituto de Investigación Agropecuaria), bajo dos densidades de siembra con diferente arreglo espacial. Se cuantificó el rendimiento y los componentes numéricos, la producción de macollaje, biomasa y sus determinantes fisiológicos (intercepción de radiación y eficiencia de uso de radiación) y, por último, la concentración de proteína en los granos de trigo e índice de cosecha de nitrógeno. Los rendimientos alcanzados por ambas densidades de siembra presentaron diferencias significativas. Siendo mayor la densidad convencional sobre baja densidad de siembra. Mientras que la concentración de proteína en los granos, la baja densidad de siembra logró superar la densidad convencional significativamente. 2 SUMMARY Wheat (Triticum aestivum L.) is one of the most important crops worldwide due to its contribution to human nutrition and its adaptive capacity to almost all regions of the planet. In Chile, wheat is the most important and traditional crop in the country's agriculture, in terms of planted area and production. Experiments in the experimental station of the Austral University of Chile, with very low stocking densities (44 pl m-2), did not show differences in yield. The grain yield results from an optimal balance of three components: the number of ears per area, the number of grains per ear and the individual weight of the grains. In the present thesis, an experiment was carried out at the Santa Rosa Agricultural Experimental Station (Austral University of Chile), with the objective of quantifying the effect of two contrasting planting densities, with different spatial arrangements, on the yield in two wheat genotypes. The experiment was carried out under optimal agronomic management and in the conditions of high yield potential in southern Chile. In the experiment, two genotypes of spring tambourine wheat were used, which correspond to a line of the WAMI (Wheat Association Mapping Initiative) population, WBLL1*2/KUKUNA and the commercial variety PANTERA-INIA (Agricultural Research Institute), under two planting densities with different spatial arrangements. Yield and numerical components, tillering production, biomass and their physiological determinants (radiation interception and radiation use efficiency) and, finally, the protein concentration in wheat grains and nitrogen harvest index were quantified. The yields achieved by both planting densities showed significant differences. Conventional density being higher over low planting density. While the protein concentration in the grains, the low sowing density managed to significantly exceed the conventional density. 3 1 INTRODUCCIÓN El trigo (Triticum aestivum L.) es uno de los cultivos más importantes a nivel mundial debido a su contribución a la alimentación humana y a su capacidad adaptativa a casi todas las regiones del planeta. Se estima que la demanda de trigo aumentará un 50% para 2050, debido al crecimiento de la población y los cambios en sus hábitos alimentarios (CIMMYT, 2019). El consumo de trigo abarca a gran cantidad de poblaciones, tanto urbanas como rurales, cuyas dietas giran en torno a este cultivo; de ahí su importancia para asegurar la alimentación de la población mundial. En Chile, el trigo es el cultivo más importante y tradicional de la agricultura del país, en términos de superficie y producción. Es cultivado en casi todos los agroecosistemas de Chile, desde la región de Coquimbo hasta la región de Los Lagos. De acuerdo con datos de ODEPA (2020), en la temporada 2019/20 se sembraron 183.073 has, las que generaron una producción de 1.086.140 toneladas de grano, con rendimiento promedio de 5.900 kg ha-1. La región de Los Ríos participa con un 8,2% de la producción nacional, con un rendimiento promedio con 8.600 kg ha-1. El manejo de los cultivos incluye diferentes prácticas y entre ellas la densidad de siembra es de gran importancia para maximizar el rendimiento y enfrentar estreses bióticos (enfermedades fúngicas, malezas e insectos), abióticos (limitación hídrica, nutricional y solar) y otras contingencias como la tendedura. La respuesta del rendimiento de trigo a la densidad de siembra muestra un amplio rango de variación. En algunos experimentos, la producción de granos por unidad de superficie tiende a mantenerse a partir de 100 pl m-2 (Fischer, 2020), especialmente cuando las condiciones de crecimiento son favorables (riego y alta fertilidad). El Instituto de Investigaciones Agropecuarias (INIA) recomienda a los productores, dosis entre 320 y 350 pl ha-1, para la variedad Pantera Clearfiled, para alcanzar rendimientos promedio de 8.200 kg ha-1 en el Borde Costero de la Región de La Araucanía (INIA, 2014). Sin embargo, se ha reportado para el Sur de Chile que densidades de siembra de 44 y 350 pl m-2 alcanzaron rendimientos similares, 4 cuando el arreglo espacial de las plantas es equidistante en la baja densidad (Bustos et al., 2013). El rendimiento de granos resulta de un óptimo balance de tres componentes numéricos del rendimiento: el número de espigas por unidad de área, el número de granos por espiga y el peso de los granos (Evans et al., 1975). Rendimientos similares pueden lograrse en un amplio rango de densidades de siembra debido a que los cereales con capacidad de macollar logran compensar los componentes numéricos del rendimiento. Entre los cereales inverno-primaverales, estas compensaciones son evidentes en trigo, avena, cebada, centeno y triticale. Cuando se usan densidades de siembra menores a las recomendadas por el INIA, esta plasticidad se expresa claramente, por lo tanto, con baja densidad de plantas y dependiendo de la capacidad del genotipo y la condición ambiental, el número de macollos por planta es mayor que con densidades más altas (Vera, 2011). En Australia, se ha demostrado que aumentos en la densidad de siembra de 75 a 150 pl m-2 tiene poca influencia sobre el rendimiento (Duggan et al., 2005). Lo mismo encontraron Turner et al. (1994). Estos últimos autores evaluaron densidades de siembra entre 45 y 135 pl m-2. Ambos experimentos fueron realizados con cultivares invernales, los que, a diferencia de los primaverales, poseen un ciclo de cultivo más largo, un mayor periodo de macollaje y, por lo tanto, una mayor producción potencial de macollos y número de espigas por unidad de superficie. Experimentos realizados en Valdivia (Chile), con cuatro genotipos de trigo primaveral bajo densidades de siembra contrastantes (44 y 350 plantas m-2), mostraron que la baja densidad de siembra alcanzó un rendimiento y número de granos similar a la condición de alta densidad. Sin embargo, sí se presentaron diferencias en los componentes del número de granos, donde, el número de espigas m-2 fue menor en la baja densidad, a pesar de un mayor número de espigas por planta, pero este componente fue compensado con un mayor número de granos espiga-1 (Bustos et al., 2013). La competencia es el proceso de gran importancia en la regulación de las respuestas del cultivo a la densidad. Ésta, se presenta cuando las plantas comparten recursos que están provistos en forma insuficiente para satisfacer la demanda potencial de cada planta individual, lo cual es la situación común de los sistemas agrícolas. Esta competencia provoca la reducción del crecimiento y rendimiento individual por planta, dependiendo 5 del nivel de reducción de la cantidad de individuos por unidad de superficie y la disponibilidad de recursos. Por lo tanto, la correcta elección de la densidad de siembra maximiza la utilización de los recursos por unidad de área y el rendimiento, sacrificando el crecimiento de cada planta individual (Satorre et al., 2003). En este aspecto, la densidad de siembra es una de las prácticas de manejo que determina la capacidad de utilización de los recursos por parte del cultivo, pudiendo llegar a afectar de manera importante la captura y utilización de radiación, agua y nutrientes. La elección de densidad tiene por objetivo maximizar el rendimiento y también la calidad de los granos. La principal característica que define la calidad y el valor nutricional del trigo son el contenido de proteínas del grano y su composición en aminoácidos (Troccoli et al., 2000). El porcentaje de proteínas en granos tiende a disminuir significativamente en altas densidades de siembra cuando existe déficit de nitrógeno en el cultivo (Lemaire et al., 2005). La densidad de siembra debe garantizar coberturas vegetales uniformes y elevadas desde etapas tempranas, y especialmente durante el período crítico para la determinación del número de granos y el rendimiento, comprendido entre 20 días antes y 10 días después de antesis (Fischer et al., 1985). Por ello, se debe determinar la densidad del cultivo en función de las condiciones agroecológicas de cada lugar (Satorre et al., 2003). En la presente tesis se cuantificó el efecto de dos densidades de siembra contrastantes, con diferente arreglo espacial, sobre el rendimiento en dos genotipos de trigo. El experimento se realizó bajo manejo agronómico óptimo y en las condiciones de alto potencial de rendimiento del sur de Chile. 6 1.1 HIPOTESIS (i) - Baja densidad de siembra (44 pl m-2) con arreglo espacial equidistante, logra rendimiento de grano similar al de la densidad convencional (300 pl m-2) en trigos primaverales, debido a compensaciones entre el número de espigas y de granos por espiga, generando una mayor partición de biomasa (Índice de cosecha), en ambientes de alto potencial de rendimiento del sur de Chile. (ii) - La baja densidad de siembra (44 pl m-2) con arreglo espacial equidistante, favorece la concentración de proteína en granos sobre la densidad convencional (300 pl m-2), asociado al aumento del índice de cosecha de nitrógeno. 1.2 OBJETIVO GENERAL Cuantificar el efecto de dos densidades de siembra contrastantes (44 y 300 pl m-2) sobre el rendimiento, la producción de biomasa aérea, sus determinantes fisiológicos y la concentración de proteína de los granos en dos genotipos de trigo primaverales bajo manejo agronómico óptimo en las condiciones de alto potencial de rendimiento del sur de Chile. 1.3 OBJETIVOS ESPECIFICOS Determinar el rendimiento y sus componentes numéricos en respuesta a densidades de siembra y arreglos espaciales contrastantes en dos genotipos de trigo bajo condiciones de manejo agronómico óptimo. Evaluar el efecto de la densidad de siembra y el arreglo espacial contrastantes sobre la producción de biomasa y sus determinantes fisiológicos (intercepción de radiación y eficiencia de uso de radiación) en dos genotipos primaverales de trigo. Estudiar la dinámica de aparición de macollos de trigo bajo densidades de siembra y arreglos espaciales contrastantes. Cuantificar la concentración de proteína de los granos, de dos genotipos de trigo en respuesta a la densidad de siembra y el arreglo espacial en condiciones de manejo agronómico óptimo. 7 2 REVISIÓN BIBLIOGRÁFICA 2.1 Antecedentes generales del cultivo de trigo El inicio de la agricultura fue originalmente descrito como la revolución neolítica, uno de los acontecimientos más importantes, y de mayor transcendencia, ocurridos en la historia del ser humano como especie (Merino, 2017). Para el trigo, es probable que su cultivo se iniciase alrededor del 12.000 antes de la actualidad (a.a.) y en algún lugar del creciente fértil, actual Mesopotamia, región asiática comprendida entre los ríos Tigris y Éufrates. Trabajos realizados con marcadores moleculares han demostrado que todas las formas cultivadas del trigo tienen su origen en las montañas del sudeste de Turquía, desde donde se distribuyeron hacia el norte y el sur de Mesopotamia alrededor del 10.000 a.a. El cultivo del trigo en América fue introducido por la colonización inglesa en las tierras conquistadas. En Chile por los españoles que llegaron en el siglo XVI (Mellado, 1998). El trigo pertenece a la división Magnoliphyta, clase Liliopsida, orden Poales (Graminales), familia Poaceas (Gramíneas), subfamilia Festucoidae, tribu Triticaceae (Hordeae), género Triticum. Este cultivo comprende alrededor de 30 tipos de trigo que tienen suficientes diferencias genéticas como para ser consideradas especies distintas o subespecies (Mac Key, 2005). Aproximadamente el 90-95% del trigo cultivado en el mundo corresponde al trigo común, el que es utilizado mayormente para la elaboración de harina para pan y una gran variedad de masas. De esta forma, el trigo provee más nutrientes a la población mundial que cualquier otra fuente de alimentación por sí sola. El consumo de pan aumenta en países subdesarrollados y es particularmente importante como fuente de carbohidratos, proteínas y vitaminas B y E (Pomeranz, 1987, citado por Peña, 2002). En Latinoamérica, Chile es el país con mayor consumo de pan, con un promedio de 96 kilos por persona al año (Murcia, 2017). 8 A nivel mundial, los países con mayor producción y superficie cultivada de trigo son: China, con una producción anual de 133.601.131 toneladas en 23.732.560 has., India, con una producción de 103.596.230 toneladas y un área cultivada de 29.318.790 has. y Federación de Rusia, que produce 74.452.692 toneladas anuales en 27.558.617 has. (FAO, 2019). En conjunto equivalen al 40,7% de la producción mundial y 37,3% de la superficie total. 2.2 Respuesta del cultivo de trigo a la densidad de siembra La respuesta de trigo a un amplio rango de densidades de siembra se expresa mediante un modelo asintótico (Figura 1), tanto para el rendimiento de grano como para la biomasa aérea. La producción de granos por unidad de superficie tiende a mantenerse a partir de ciertos rangos de densidad de siembra, debido a la capacidad compensatoria y la plasticidad fenotípica del trigo, aunque este valor puede ser variable entre ambientes. Esto demuestra que, al aumentar la densidad de siembra, el rendimiento por planta disminuye, pero el mayor número de plantas por unidad de superficie compensa la producción final. Figura 1. Respuesta de rendimiento al aumento en la densidad de plantas de un cultivo de trigo, en la temporada 1996/97 (triángulos) y 1997/98 (cuadrados). Fuente: Whaley et al. (2000). 9 Al igual que en otros cultivos, el tipo de respuesta del rendimiento a la densidad también depende de las características ambientales y genotípicas. Sin embargo, el rendimiento por planta, a muy bajas densidades en ambientes productivos, dependerá más de las diferencias genéticas entre los cultivares estudiados que de las condiciones ambientales (Satorre et al., 2003). Lo que se busca al estudiar el efecto de la densidad, es conocer de qué forma afecta a los componentes del rendimiento cómo: número de espigas por unidad de superficie, número de granos por espiga y el peso promedio de los granos. La capacidad que tienen estos componentes de variar por efecto de las condiciones ambientales, se denomina plasticidad. Cuando mayor sea la plasticidad de un componente del rendimiento, más sensible es a sufrir cambios ante variaciones ambientales y de manejo. Entre ellos, el peso de grano es considerado un componente de baja plasticidad, ya que tiende a mantenerse estable ante diferentes densidades. Por otra parte, el número de espigas por planta y el número de granos por espiga, muestran variación en función de la densidad de siembra, por lo que se los considera componentes de alta plasticidad (Slafer et al., 2014), por lo tanto, estos componentes muestran la mayor compensación para el rendimiento. Distintos estudios han demostrado la capacidad del cultivo de trigo para compensar las reducciones en el número de plantas por unidad superficie, esto principalmente mediante la formación de espigas provenientes de macollos. La generación de estas estructuras son el resultado, principalmente, de señales fotomorfogénicas en la canopia (relación rojo/rojo lejano) y la disponibilidad de asimilados, agua y nutrientes (Evers et al., 2006). Las diferentes densidades generan cambios en el ambiente de la canopia, además de la interacción lumínica como, por ejemplo, altas densidades de siembra producen alta competencia intraespecífica por recursos como luz, agua y nutrientes durante las fases críticas. En ambientes de baja precipitaciones y poca fertilidad de suelo, esto afecta fuertemente el rendimiento final. También aumentan el riesgo de tendedura y daño por insectos o enfermedades en ambientes más productivos. La interacción entre las condiciones ambientales, el manejo agronómico y el genotipo determinarán finalmente el rendimiento por unidad de superficie (Satorre et al., 2003). Estudios realizados por Bustos et al. (2013) mostraron que el rendimiento de trigo y sus componentes no presentaron diferencias significativas a densidades de siembra muy 10 contrastantes (44 y 350 plantas m-2). Sin embargo, sí se presentaron diferencias en los componentes del número de granos, donde, el número de espigas m -2 fue menor en densidades bajas, pero este fue compensado con un mayor número de granos espiga -1. Sin embargo, esos resultados se obtuvieron solo en una temporada sin poder cuantificar la interacción genotipo y ambiente, por lo que no hubo evidencia de que alguno de ellos tuviera un comportamiento distinto a las diferentes densidades de siembra. Cabe destacar que ese estudio se realizó sin limitación hídrica o de nutrientes. 2.3 Intercepción de radiación de trigos con densidades y arreglos espaciales contrastantes La radiación incidente varía con la ubicación geográfica (latitud) y con la época del año. Aun cuando la magnitud de la radiación incidente está fuera del manejo agronómico, ciertas prácticas culturales como la elección de la fecha de siembra y del genotipo permiten adecuar el ciclo del cultivo para maximizar su captura. No toda la radiación solar es utilizada por las plantas para la fotosíntesis, ya que se estima que de la radiación directa incidente el 45% es radiación fotosintéticamente activa (400 a 700 nm). A esto es necesario adicionar la contribución de la radiación difusa, por lo que normalmente se considera radiación fotosintéticamente interceptada al 50% de la radiación incidente (Sinclair et al., 1999). En el manejo de los cultivos de alta producción, uno de los objetivos principales es lograr que el cultivo intercepte la mayor proporción de la radiación solar incidente, debido a que la misma es la fuente de energía de la fotosíntesis. Por lo tanto, la producción de materia seca de los cultivos dependerá de esa captura de radiación y otros recursos. La tasa de crecimiento de un cultivo (TCC: biomasa acumulada por unidad de superficie y de tiempo), resulta del equilibrio de dos procesos contrapuestos: la fotosíntesis (ganancia de CO2) y la respiración (pérdida de CO2). Debido a que la fracción del carbono fijado que se destina a respiración depende de la cantidad total de biomasa, aumentos en la radiación interceptada por el cultivo, se traducen en aumentos proporcionales en la tasa de crecimiento. 11 Si bien es importante lograr que el cultivo capte la mayor cantidad posible de radiación incidente, es también primordial lograr que la misma sea convertida eficientemente en materia seca. Es así como surge el concepto de eficiencia del uso de la radiación (EUR). La EUR fotosintéticamente activa (RFA) es calculada como la pendiente de la regresión entre la biomasa aérea y la RFA interceptada, y se expresa en gramos (g) de biomasa por unidad de energía de radiación (Satorre et al., 2003). Esta relación da cuenta de la cantidad de materia seca total producida por unidad de radiación solar interceptada. En la Figura 2, en el sur de Chile Bustos et al. (2013) muestra que la radiación interceptada desde emergencia es mayor en densidad convencional sobre baja densidad, pero el valor de EUR para baja densidad fue superior a la densidad convencional. Figura 2. EUR fotosintéticamente activa (RFA) calculada como la pendiente de la regresión entre la biomasa aérea y la RFA interceptada, expresada en gramos (g) de biomasa por unidad de energía de radiación. Las mayores diferencias en EUR se deben al tipo de metabolismo del carbono (especies C3 vs. especies C4) y el tipo de materia seca producida (hidratos de carbono, proteínas o lípidos), por el diferente costo de síntesis de estos compuestos (Stöckle et al., 2009). Los valores obtenidos de EUR están relacionados con características del cultivar y 12 prácticas de manejo como la estructura del cultivo, sistema de labranza, etc. Al respecto, Jamieson et al. (1995) informaron que la EUR fue afectada por el ciclo del cultivar y por el distanciamiento entre hileras. Por otro lado, Mirassón et al. (2008) concluyeron que los distanciamientos mayores a 0,175 m reducen la EUR, siendo los cultivares de ciclo intermedio los de mejor comportamiento en cuanto a rendimiento. Chen et al. (2008), en experimentos conducidos en Montana (USA), concluyeron que el trigo acumula mayor cantidad de biomasa y a tasa más alta cuando es sembrado a espaciamientos menores a 0,175 m y que la corrección de la densidad de siembra no mejora los rendimientos de trigos sembrados a mayores espaciamientos. El aumento del espaciamiento entre surcos permite conservar el agua para las etapas reproductivas más críticas. Así, en situaciones de lluvias escasas durante el ciclo del cultivo, los rindes dependerán del agua almacenada en el suelo al momento de la siembra (Robinson, 1978). A las mismas conclusiones arribaron Tompkins et al. (1991) para el oeste de Canadá e indicaron que el número de espigas m-2 es el responsable del aumento del rendimiento. Siemens (1963), en experimentos en Canadá, observó que los espaciamientos amplios aumentan significativamente la cantidad de macollos, sin embargo, los máximos rendimientos se obtuvieron con espaciamientos convencionales. 2.4 Concentración de clorofila y proteína Las principales características que definen el valor nutricional y la calidad del trigo son el contenido y, composición de proteínas, tipo de proteínas y aminoácidos en el grano (Troccoli et al., 2000). El contenido de proteínas de granos presenta una alta variabilidad, dependiendo del genotipo y las condiciones ambientales de crecimiento, principalmente con la dosis y época de aplicación del fertilizante nitrogenado, el nitrógeno residual presente en el suelo, disponibilidad hídrica durante el cultivo y la temperatura desde antesis hasta madurez fisiológica (Bénétrix et al., 2001). Para obtener el porcentaje de proteína se utiliza 5,7 como factor de conversión del nitrógeno a proteína en granos de cereales (Teller. 1932). La cantidad de nitrógeno (N) en el suelo es variable a través del tiempo y en su disponibilidad para el cultivo, encontrándose en forma de compuesto orgánico o en forma mineral. Para ser utilizado por los cultivos, el N orgánico no está disponible a diferencia 13 del N mineral. Este último, se encuentra disuelto en la solución del suelo, pudiendo ser absorbido por las raíces de las plantas. Sin embargo, el N orgánico es capaz de transformarse gradualmente en N mineral mediante el proceso de mineralización, representando un aporte parcializado del N mineral disponible para el uso de las plantas. Durante la temporada de crecimiento se asume que los cultivos solo utilizan el N mineral en condiciones sin fertilización nitrogenada, por lo tanto, se le considera como el índice de disponibilidad más confiable. Esta aproximación no considera la eficiencia de absorción del cultivo, por lo que es necesario establecer si existe variación en la eficiencia de absorción de N entre los distintos genotipos (Quiñones, 2007). El aumento del número de plantas por unidad de superficie afecta no solo la capacidad de intercepción de luz, sino que también, el aprovechamiento de recursos del suelo, al menos durante las primeras etapas del cultivo (Quiñones, 2007). En las fases tempranas del ciclo del trigo, poco después de la emergencia, las altas densidades de siembra presentan una mayor demanda de los recursos del suelo, debido a la mayor cantidad de plantas por unidad de superficie. Si bien, a medida que el cultivo avanza en su ciclo, las bajas densidades de siembra expresan sus mecanismos de compensación, formando macollos que cubren los espacios vacíos de la canopia. De este modo, la absorción de nutrientes y agua por planta genera un aumento significativo, igualando en la demanda de recursos del suelo a los cultivos de alta densidad. Aun así, en un ambiente con restricciones, la tasa de crecimiento dependerá más de la disponibilidad de recursos que de la densidad de plantas (Castellarín, 2009) y de la capacidad y duración del macollaje del cultivo. Estudios realizados por Arduini et al. (2006) presentaron mayor captación de N en los tratamientos con densidades más alta sobre los de baja densidad. Esta comparación podría plantear que el aprovechamiento de los nutrientes del suelo por parte de los cultivos podría estar influenciado por otro factor espacial además del número de plantas m-2. Estudios realizados por Auld et al. (1983) mostraron que los cultivos con un arreglo espacial regular de plantas (relación entre la distancia de plantas sobre hilera y entre hilera 1:1) presentaron mayor rendimiento y producción de biomasa que los tratamientos con de plantas en distribución rectangular, e incluso entre rendimiento y rectangularidad espacial los autores encontraron una relación negativa. Por lo tanto, en la capacidad de absorción de N del suelo por parte del cultivo de trigo, podría jugar un papel importante el arreglo espacial. 14 Al fertilizar el suelo con nitrógeno, la clorofila tiende a aumentar su concentración (Daughtry et al., 2000) y al aumentar la concentración de clorofila en la hoja se producen hojas de color verde más intenso (Varinderpal-Singh et al., 2011) y mayores rendimientos (Ning et al., 2012). Una buena disponibilidad de N asegura un buen crecimiento foliar, lo que permite al cultivo optimizar la captura de recursos como radiación y su conversión a materia seca; alcanzando así alta producción de biomasa y rendimiento (Escobar et al., 2012). Mohkum et al. (2011) señalaron que las aplicaciones de N aumentaron significativamente el índice de área foliar del cultivo, lo que garantizó su óptimo estado fisiológico en los momentos decisivos para la determinación del número de granos y durante el período de llenado de estos. El estado nutricional del cultivo puede ser evaluado a través de la medición del contenido de clorofila de la hoja realizada mediante clorofilómetro (Daughtry et al., 2000). Dichas lecturas se relacionan a su vez, directamente con el rendimiento de la planta, y por lo tanto se podrían utilizar para poder diagnosticar suficiencia o deficiencia de requerimiento de nitrógeno de los cultivos (Pagani et al., 2009). 15 3 MATERIALES Y MÉTODOS 3.1 Ubicación del experimento. El experimento se realizó durante la temporada 2019-2020 en la Estación Experimental Agropecuaria Austral (EEAA), perteneciente a la Universidad Austral de Chile, ubicada en las coordenadas de latitud 39°47’19’’ N y longitud 73°14’05’’ O, en el sector Cabo Blanco, a 4 km al norte de la ciudad de Valdivia, Región de Los Ríos. El suelo es de origen volcánico (trumao), correspondiente la serie Valdivia (Duric Hapludand). 3.2 Material vegetal Se utilizaron dos genotipos de trigo pandero primaveral, los cuales corresponden a una variedad comercial PANTERA-INIA del Instituto de Investigación Agropecuaria de Chile y una línea (WBLL1*2/KUKUNA) del programa de mejoramiento del Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT). Estos genotipos se denominarán de ahora en adelante como Pantera y WBLL, respectivamente. 3.3 Tratamientos y Diseño Experimental El experimento consistió en los siguientes tratamientos: (i) genotipo (dos genotipos de trigo, Pantera y WBLL), y (ii) densidad de siembra con diferente arreglo espacial: convencional (300 plantas m-2 con ordenamiento rectangular, 15 cm de entre hilera y 2,2 cm de sobre hilera) y baja densidad (44 plantas m-2 dispuestas en un ordenamiento equidistante, hileras separadas a 15 cm y la misma distancia entre las plantas sobre la hilera). Estos tratamientos se denominarán de ahora en adelante como densidad convencional y baja densidad, respectivamente. 16 El diseño del experimento fue un diseño en parcelas divididas, distribuido en bloques completos al azar con tres repeticiones, donde la parcela principal fueron los genotipos de trigo y las subparcelas consistieron en las dos densidades de siembra con diferente arreglo espacial (44 plantas m-2 y 300 plantas m-2) (Figura 3). Las subparcelas tuvieron un tamaño de 1,95 x 3,0 m (5,85 m2, 13 hileras), donde se sembraron los genotipos con entre hilera de 0,15 m. Entre las parcelas y entre los bloques se dejarán pasillos de 1,5 m. La superficie total del experimento fue de 102,8 m2 (13,7 x 7,5 m). Figura 3. Diseño experimental del experimento. Los bloques son representados por las líneas punteadas. Las parcelas principales son las líneas solidas de color azul que representan los genotipos de trigo, mientras que dentro de cada parcela se indican los niveles de densidad de siembra: 44 plantas m-2 y 300 plantas m-2. 17 3.4 Manejo del experimento El sitio experimental tuvo como cultivo antecesor maíz de grano sembrado a una densidad de 100.000 plantas ha -1, sin fertilización, con el objetivo de remover la mayor cantidad posible de nitrógeno del suelo para homogeneizar la disponibilidad de nitrógeno a la siembra del experimento. Al finalizar el ciclo el maíz se cosechó la biomasa, removiendo completamente de los residuos de cultivo. Previo a la siembra del experimento se tomaron muestras de suelo para evaluar el pH y contenido de nutrientes del suelo para decidir la fertilización. Los requerimientos de fertilización para la siembra fueron, 100 kg de nitrógeno (N) ha-1, 157 kg de fósforo (P) y 93 kg de potasio (K), y por último 180 kg de N ha-1 en inicio de elongación de entrenudos, en Zadoks 3,1 (Zadoks et al., 1974). La fuente de N que se utilizó fue CAN 27, en P fue superfosfato triple (SFT) y en K fue muriato de potasio (KCl). La aplicación de enmiendas se realizó con al menos un mes de anticipación a la siembra. La siembra del experimento se llevó a cabo el 5 de septiembre de 2019, y se realizó por medio de cintas adhesivas de papel (cinta de enmascarar) para ambas densidades utilizadas, donde se distribuyeron las semillas de trigo y luego se cubrieron con papel sanitario para asegurar la calidad de siembra y distribución de plantas. Figura 4. Distribución de semillas de trigo en cintas adhesivas (a). Cintas adhesivas distribuidas en la estación experimental (b). Trigo emergido en la estación experimental (c). Foto tomada de Ferrante et al. (2012). 18 El control de malezas se realizó utilizando herbicidas de pre-emergencia, y luego se realizó un segundo control con un herbicida post-emergente, cuando la población de malezas fue baja se optó por un control manual sobre las parcelas. Se aplicaron regularmente fungicidas de acción curativa/preventiva e insecticidas. Durante el periodo del experimento, las parcelas fueron regadas para evitar déficit hídrico a través de un riego por goteo. Para el control de aves se estableció una malla de monofilamento de 0,3 mm con el fin de evitar el daño en los granos. 3.5 Muestreos y mediciones Durante todo el ciclo del cultivo se hizo un seguimiento de la fenología hasta madurez de cosecha. Se realizaron nueve muestreos destructivos de biomasa aérea durante el ciclo, incluida la cosecha. El tamaño de los muestreos fue de 0,5 m lineales (0,075 m2) en la densidad convencional y 0,9 m lineales (0,0135 m2, 6 plantas) en las parcelas de baja densidad, con la excepción del muestreo final que fue de 1 m lineal en las parcelas de densidad convencional (0,15 m2) y 1,05 m lineales (0,1575 m2, 7 plantas) en las parcelas de baja densidad. En cada muestreo se separaron las plantas en sus órganos (granos, espigas, tallos, hojas y materia senescente; cuando existan). Se registró el número de plantas, vástagos y espigas m-2. Posteriormente las muestras vegetales fueron secadas en un horno a 60 °C por 48 horas. El peso de las muestras vegetales se midió en una balanza electrónica (Mettler, Toledo XP205DR, Greifensee, Suiza), siendo la sumatoria de los órganos la biomasa aérea total. Se calculó la superficie foliar medida a partir de las láminas verdes, mediante el uso de un medidor de área foliar (LI 3100, Licor Inc., Lincoln NE, USA), y la superficie muestreada, para obtener el índice de área foliar (IAF). También se midió la radiación incidente y trasmitida a nivel del suelo o a nivel del estrato verde de la canopia una vez por semana al medio día en días despejados. Para ello, se utilizó un radiómetro de 1 m de longitud (LI 191 S, Licor Inc., Lincoln NE, USA). Para calcular la EUR se realizó la regresión lineal de la biomasa aérea acumulada en función de la PAR interceptada (IPARa) acumulada entre inicio de macollos y la antesis y entre 19 la antesis y la madurez fisiológica. Se calculó el coeficiente de extinción lumínico mediante la ley de Beer (Hipps et al., 1983), relación entre la radiación interceptada y el índice de área foliar. Además, se evaluó indirectamente la concentración de clorofila de las hojas mediante SPAD (SPAD 502, Minolta, Spectrun Technologies Inc., Illinois, USA) en la última hoja completamente expandida y en dos posiciones de la hoja de 4 plantas seleccionadas al azar, una vez a la semana. La dinámica de macollaje se evaluó en cada parcela desde el inicio de macollaje, i.e. 2,1 (Zadoks et al., 1974), hasta madurez fisiológica, mediante el seguimiento del número de macollos en tres plantas a intervalos regulares de tiempo (cada 4 días). En la cosecha final del experimento, luego de madurez fisiológica (<37,5% de humedad de granos; Calderini et al., 2000), se midió la altura de planta en seis plantas por repetición, separando entre vástagos principales y macollos. Las espigas fueron trilladas, obteniéndose por separado el grano y capotillo (estructuras de la espiga que no son grano), para ser pesados en una balanza electrónica (Mettler, Toledo XP205DR, Greifensee, Suiza). Se determinó el número de granos, contándose la totalidad de la muestra colectada mediante el contador de granos (Automatic Seed Counter, VTS Group S.A., Windhof, Bélgica). El peso de mil granos se determinó como la razón entre el peso de la muestra total de granos y el número de granos, multiplicados por 1.000. Se calculó el índice de cosecha (IC) como la proporción entre la MS cosechada como grano y la MS seca total de la parte aérea del cultivo. Por otra parte, la biomasa de espigas en antesis y el número de granos a cosecha permitieron calcular la eficiencia de fructificación, como el cociente entre ambos. Para la determinación de la concentración de proteína en grano se llevó a cabo a partir de la multiplicación entre el factor de conversión 5,7 (Teller, 1932) y la concentración de nitrógeno. El porcentaje de nitrógeno fue medido por el método de Dumas, también conocido como método de combustión (Dumas, 1831) en plantas completas. Durante el periodo de crecimiento del cultivo se recopiló la información climática en la estación meteorológica de la EEAA, ubicada a 230 metros del experimento. La información climática obtenida consistió en la temperatura, precipitaciones y radiación 20 solar. Se consideró como periodo de crecimiento del cultivo desde la siembra el 5 de septiembre del año 2019 hasta la cosecha el 2 de febrero del año 2020. 3.6 Análisis de estadístico de resultados Los análisis estadísticos de los resultados se realizaron por medio de ANOVA para parcelas sub-divididas, con el software estadístico R, utilizando el paquete Agricolae, considerando el genotipo como la parcela principal y la densidad de siembra como la subparcela. La asociación entre las variables se realizó mediante análisis de regresión lineal y no lineal utilizando el software GraphPad Prism 7.0. 21 4 PRESENTACIÓN DE RESULTADOS 4.1 Condiciones climáticas durante la temporada 2019-2020 y la fenología de los genotipos La radiación solar y las temperaturas máximas y mínimas registradas en la Estación Experimental Agropecuaria Austral (EEAA), se muestran en la Figura 5. La temperatura promedio entre la emergencia y antesis del cultivo fue de 11,4°C y durante el llenado de granos (desde antesis hasta madurez fisiológica) 16°C. La radiación solar promedio entre emergencia y antesis fue de 19,2 MJ m-2 y 16 MJ m-2 durante el llenado de granos, para Pantera y WBLL, respectivamente. b) Precipitación Acumulada (mm) a) c) 50 T ºC Máx T ºC Min 40 Madurez fisiológica Antesis 30 20 10 0 -10 0 20 40 60 80 100 120 140 160 Días desde siembra Figura 5: Precipitación mensual (a) durante enero de 2019 hasta febrero de 2020 y radiación solar (b) y temperaturas mínima y máxima (c) desde siembra hasta cosecha del experimento. 22 Las precipitaciones no tuvieron una distribución uniforme a lo largo del ciclo del cultivo, ya que se concentraron en los primeros meses de primavera, durante la fase vegetativa del cultivo y disminuyeron hacia el verano, durante la segunda mitad del llenado de granos. La suma de la precipitación desde siembra hasta madurez fisiológica fue de 338 mm. Por otra parte, las temperaturas y la radiación solar tuvieron un incremento hacia el verano. La fenología fue similar entre Pantera y WBLL, tanto en densidad convencional como en baja densidad. En ambos genotipos la fecha de emergencia fue el 15 de septiembre, la antesis el 01 de diciembre y la madurez fisiológica el 20 de enero de 2020. 4.2 Rendimiento, biomasa, índice de cosecha y altura de planta de los genotipos evaluados en ambas densidades de siembra El rendimiento de grano en la densidad convencional no mostró diferencias (p>0,05) entre genotipos, alcanzando 1.467 y 1.498 g m-2 en Pantera y WBLL, respectivamente. Por otra parte, la densidad de siembra afectó (p<0,01) el rendimiento. En baja densidad, Pantera alcanzó 1.080 g m-2 y WBLL 1.262 g m-2, los que fueron 26,4 y 15,8% menores que en densidad convencional, respectivamente. Al igual que en densidad convencional, no hubo diferencias (p>0,05) entre los genotipos. Tampoco se encontró interacción (p>0,05) entre el genotipo y la densidad de siembra (Cuadro 1). La biomasa área en la densidad convencional alcanzó 3.022 y 2.910 g m-2 en Pantera y WBLL, respectivamente, sin presentar diferencias (p>0,05) entre ellos. En baja densidad, las biomasas aéreas de Pantera y WBLL también fueron similares (p>0,05): 2.293 y 2.324 g m-2, respectivamente. A lo largo de todo el periodo del cultivo, y en ambos genotipos, la biomasa aérea fue mayor (p<0,001) en la densidad convencional (Figura 6) y no se encontró interacción (p>0,05) entre el genotipo y la densidad de siembra en este atributo del cultivo (Cuadro 1). 23 a) b) PANTERA-INIA 4000 Biomasa aérea (g m-2) Madurez fisiológica 44 pl m-2 3500 -2 300 pl m 3000 2500 Antesis 2000 1500 1000 500 0 0 250 500 750 1000 1250 1500 1750 2000 2250 2500 Tiempo térmico desde emergencia (ºC d) Figura 6. Biomasa aérea desde la emergencia del cultivo en (a) PANTERA–INIA (círculos) y (b) WBLL 1*2/KUKUNA (cuadrados) en densidad convencional (símbolos abiertos) y baja densidad (símbolos cerrados). En cuanto al IC, no se registraron diferencias (p>0,05) entre genotipos ni entre densidades, pero si se encontró interacción (p<0,05) entre estas fuentes de variación, donde Pantera mostró un valor levemente mayor del IC en la densidad convencional con respecto a la baja densidad (49 y 47%, respectivamente), mientras que en la densidad convencional WBLL mostró un IC levemente menor que en baja densidad, i.e. 51 y 54%, respectivamente (Cuadro 1). 24 Cuadro 1. Rendimiento, biomasa aérea, índice de cosecha (IC), en PANTERA–INIA y WBLL 1*2/KUKUNA creciendo bajo condiciones de densidad convencional y baja densidad. Genotipo Densidad -2 Rendimiento -2 Biomasa -2 IC (plantas m ) (g m ) (g m ) (%) 300 1.467 3.022 49 44 1.080 2.293 47 300 1.498 2.910 51 44 1.262 2.324 54 Genotipo N.S N.S N.S Densidad ** *** N.S N.S N.S * PANTERA-INIA WBLL1*2/KUKUNA Genotipo: Densidad El análisis de varianza mostró que la altura de planta se diferenció (p<0,05) entre los genotipos. Pantera y WBLL alcanzaron 79,9 y 96,4 cm, respectivamente, en densidad convencional. En baja densidad, la altura fue menor en ambos genotipos, alcanzando 77,1 cm en Pantera y 86,6 cm en WBLL (Cuadro 2). WBLL se caracterizó por su mayor altura, superando (p<0,05) en un 17 y 11% a Pantera, en densidad convencional y baja densidad, respectivamente. La altura del vástago principal y los macollos también mostraron diferencias (p<0,05) entre Pantera y WBLL, en ambas densidades de siembra. Entre densidades contrastantes, solamente el vástago principal mostró diferencias (p<0,01), donde Pantera y WBLL alcanzaron 82,2 y 102 cm, respectivamente, en densidad convencional y en baja densidad, Pantera midió 78,7 cm y WBLL 90,4 cm. 25 Cuadro 2. Altura de planta en vástago principal (Altura VP) y, en los macollos (Altura Mac) y promedio de planta (Altura de planta), en PANTERA–INIA y WBLL 1*2/KUKUNA creciendo bajo condiciones de densidad convencional y baja densidad. Genotipo Densidad Altura de Altura VP Altura Mac (plantas m ) planta (cm) (cm) (cm) 300 79,9 82,2 78,3 44 77,1 78,7 76,9 300 96,4 102 89,5 44 86,6 90,4 85,9 Genotipo * * * Densidad * ** N.S N.S * N.S -2 PANTERA–INIA WBLL 1*2/KUKUNA Genotipo: Densidad 4.3 Componentes numéricos del rendimiento en vástagos principales y macollos El NG (m-2) en la densidad convencional fue de 29.402 y 26.128 en Pantera y WBLL, respectivamente, sin que hubiera diferencias estadísticas (p>0,05) entre los genotipos. En baja densidad, Pantera estableció 21.270 granos m-2 y WBLL 19.606, sin diferencias estadísticas (p>0,05) entre ellos. A pesar de los altos números de granos m-2 alcanzados por los genotipos en baja densidad, donde estuvieron por debajo (p<0,01) de los valores logrados en densidad convencional en 38,2 y 33,3% en Pantera y WBLL, respectivamente. No se encontró interacción (p>0,05) entre el genotipo y la densidad de siembra para este carácter del cultivo (Cuadro 3). Por otra parte, el P1000 sí mostró diferencias significativas entre los genotipos y densidades de siembra (p<0,05 y p<0,01, respectivamente). En densidad convencional, el P1000 alcanzado por Pantera fue de 49,9 g y 57,3 g en WBLL, mientras que en baja densidad los valores fueron 50,9 g y 64,3 g en ambos genotipos respectivamente, lo que representó un incremento de 2,0 y 10,9%, respectivamente (Cuadro 3). El genotipo WBLL mostró mayor potencial de peso grano que Pantera, independientemente de la densidad. Por lo tanto, esta fue la causa de la interacción (p<0,01) entre genotipo y densidad de siembra para este atributo del cultivo. 26 Cuadro 3. Número de granos total por unidad de superficie (NG), peso de mil granos (P1000), número de espigas por unidad de superficie y número de granos por espiga, en PANTERA–INIA y WBLL 1*2/KUKUNA en densidad convencional y baja densidad. Genotipo PANTERA-INIA WBLL1*2/KUKUNA Genotipo Densidad Genotipo: Densidad Densidad (plantas m-2) NG (Granos m-2) P1000 (g m-2) Espigas (Espigas m-2) NG (Granos espiga-1) 300 44 300 44 29402 21270 26128 19606 49,9 50,9 57,3 64,3 671 404 600 298 44 53 44 66 N.S *** N.S * ** ** * *** * N.S *** ** A pesar de que el NG no difirió entre los genotipos, el número de espigas m-2, mostró diferencias (p>0,05) ya que entre Pantera logró mayor cantidad de espigas m-2 que WBLL en ambas densidades. También se encontraron diferencias (p<0,001) para este componente del NG, entre la densidad convencional y la baja densidad. Pantera, en densidad convencional, alcanzó el mayor número de espigas (671 espigas m-2), mientras que WBLL en baja densidad presentó el menor número (298 espigas m-2), lo que se refleja en la interacción (p<0,05) entre el genotipo y la densidad de siembra (Cuadro 3). La Figura 7a muestra la asociación positiva entre el número de granos y el número de espigas por metro cuadrado considerando ambos genotipos y densidades. Por otra parte, el número de granos por espiga no presentó diferencias (p>0,05) entre los genotipos, ya que en densidad convencional el número de granos por espiga fue de 44 en ambos genotipos. En baja densidad fue de 53 y 66 granos por espiga en Pantera y WBLL, respectivamente (Cuadro 3). En baja densidad, este componente superó significativamente (p<0,001) al de alta densidad, con 9 y 22 granos por espiga más que en densidad convencional en Pantera y WBLL, respectivamente (Figura 7b). 27 a) Figura 7. b) Número de granos m-2 en PANTERA–INIA (círculos) y WBLL 1*2/KUKUNA, (cuadrados) en función al número de espigas m-2 (a) y número de granos espigas-1 en densidad convencional (símbolos abiertos) y baja densidad (símbolos cerrados). 4.4 Dinámica de macollaje La cantidad de espigas por planta y por unidad de superficie está ligada a la dinámica de macollaje del cultivo de trigo. Por ese motivo, se analiza aquí dicha dinámica. Para ello el macollaje fue evaluado considerando el número máximo de macollos aparecidos y el número final de macollos por planta, además de la evolución del número de macollos que se muestra en la Figura 8. El momento en el cual comenzó el macollaje fue a los 17 días (163,7°Cd) desde la emergencia en todos los tratamientos, independiente del genotipo y de la densidad de siembra (Figura 8a y b). 28 a) b) Figura 8. Dinámica de aparición de macollos por planta desde emergencia del cultivo en (a) PANTERA–INIA (círculos) y (b) WBLL 1*2/KUKUNA (cuadrados) en densidad convencional (símbolos abiertos) y baja densidad (símbolos cerrados). Entre Pantera y WBLL no se encontraron diferencias (p>0,05) en el número máximo de macollos aparecidos. Solamente el número final de macollos por planta mostró diferencias (p<0,05) entre los genotipos, donde Pantera superó en un 33% y 59% a WBLL en la densidad convencional y en baja densidad, respectivamente (Cuadro 4). La densidad de siembra si afectó (p<0,001) el número máximo de macollos aparecidos y el número final de macollos por planta. Esto confirma la gran capacidad compensatoria que posee el trigo. La densidad convencional presentó un mayor número de plantas establecidas por metro lineal, pero la baja densidad mostró un mayor número de macollos por planta desde los 33 días (324°Cd) desde la emergencia, en Pantera y WBLL. El número máximo de macollos en densidad convencional fue 3,3 y 2,1 en Pantera y WBLL, respectivamente, mientras que en baja densidad alcanzaron valores de 11,9 y 7,1 macollos por planta en esos mismos genotipos. Las plantas en baja densidad superaron 3,5 veces la cantidad de macollos registrados en densidad convencional. El número final de macollos por planta alcanzado a los 120 días (1.531°Cd) desde la emergencia, en densidad convencional, fue 1,2 y 0,4 en Pantera y WBLL, respectivamente, y en baja densidad Pantera alcanzó el valor de 9,3 y 5,5 en el caso de WBLL, observándose en baja densidad 2,7 veces más macollos finales que en la densidad convencional. 29 La interacción entre el genotipo y la densidad de siembra fue significativa (p<0,05 y p<0,01, respectivamente) tanto para el número máximo como final de macollos por planta, lo que indica que el macollaje de estos genotipos se comportó de manera diferencial en respuesta a la densidad (Cuadro 4). Cuadro 4. Número máximo de macollos (N° Máximo macollos) y número final de macollos (N° Final macollos), en PANTERA–INIA y WBLL 1*2/KUKUNA creciendo bajo condiciones de densidad convencional y baja densidad. Genotipo PANTERA-INIA WBLL1*2/KUKUNA Densidad (plantas m-2) N° Máximo macollos (planta-1) N° Final macollos (planta-1) 300 44 300 44 3,3 11,9 2,1 7,1 1,2 9,3 0,4 5,5 N.S *** * * *** ** Genotipo Densidad Genotipo: Densidad 4.5 Dinámica de la captura de radiación La capacidad de macollaje y el área foliar de las plantas son clave, entre otros caracteres, para la captura de radiación por parte del cultivo. Este determinante fisiológico de la biomasa fue evaluado en la presente tesis mediante la intercepción de radiación por parte del cultivo (IR) junto a la eficiencia en el uso de la radiación (EUR), el otro determinante fisiológico de la producción de biomasa. La densidad convencional en Pantera y WBLL, alcanzó la intercepción completa de radiación a los 67 días (737°Cd) desde la emergencia. Esto ocurrió aproximadamente en espigado y 11 días previo a la antesis. En el caso de la baja densidad, la captura completa de la radiación se logró a los 80 días (922°Cd) desde emergencia en ambos genotipos, durante el llenado de grano (Figura 9). 30 a) Figura 9. b) Dinámica de la radiación interceptada en función del tiempo térmico desde emergencia del cultivo en (a) PANTERA–INIA (círculos) y (b) WBLL 1*2/KUKUNA (cuadrados). En densidad convencional (símbolos abiertos) y baja densidad (símbolos cerrados). 4.6 Índice de área foliar La captura de radiación depende del índice de área foliar (IAF) y el coeficiente de extinción lumínico (k). El IAF alcanzó el valor critico (95% de intercepción de radiación) con valores de 5,9 y 3,2 en densidad convencional y baja densidad respectivamente, en Pantera y WBLL (Figura 10). El IAF en antesis para la densidad convencional fue de 5,1 y 5,0 en Pantera y WBLL, respectivamente, mientras que en baja densidad Pantera y WBLL lograron 3,7 y 2,8 (Figura 10). Una sola función se pudo ajustar para la relación entre la radiación interceptada en porcentaje y el IAF (Figura 10). Por lo tanto, y como era de esperar, el coeficiente de extinción lumínico no se vio alterado por la densidad de siembra en ninguno de los genotipos. 31 a) b) Figura 10. Radiación interceptada y el índice de área foliar en (a) PANTERA–INIA (círculos) y (b) WBLL 1*2/KUKUNA (cuadrados). En densidad convencional (símbolos abiertos) y baja densidad (símbolos cerrados). 4.7 Eficiencia en el uso de la radiación La EUR fue evaluada en las fases de pre-antesis y post-antesis en ambos genotipos y densidades de siembra. En densidad convencional la EUR pre-antesis fue 3,1 y 3,0 g MJ-1 en Pantera y WBLL, respectivamente, mientras que en baja densidad Pantera y WBLL, también alcanzaron los mismos valores, i.e. 3,0 g MJ-1 ambos genotipos (Figura 11a y b). En la fase de post-antesis, la EUR en densidad convencional fue de 2,9 y 2,6 g MJ-1 en Pantera y WBLL, respectivamente. En baja densidad Pantera alcanzó un valor levemente menor 2,4 g MJ-1 y WBLL el mismo que en densidad convencional, i.e. 2,6 g MJ-1 (Figura 11c y d). La baja densidad logró mantener una misma EUR respecto a la densidad convencional hasta antesis y durante el llenado de granos fue levemente menor que en densidad convencional en el caso de Pantera. 32 a) b) EUR Densidad convencional = 3,1 g MJ-1 EUR Baja densidad = 3 g MJ-1 EUR Densidad convencional = 3 g MJ-1 EUR Baja densidad = 3 g MJ-1 c) d) EUR Densidad convencional = 2,9 g MJ-1 EUR Baja densidad = 2,4 g MJ-1 EUR Densidad convencional = 2,6 g MJ-1 EUR Baja densidad = 2,6 g MJ-1 Figura 11. Biomasa aérea y la radiación interceptada acumulada desde emergencia hasta antesis en (a) PANTERA–INIA (círculos) y (b) WBLL 1*2/KUKUNA (cuadrados) y desde antesis hasta madurez de cosecha en (c) PANTERA–INIA (círculos) y (d) WBLL 1*2/KUKUNA (cuadrados). En densidad convencional (símbolos abiertos) y baja densidad (símbolos cerrados). 33 4.8 Peso específico foliar La EUR es afectada por el peso específico foliar y por ello se evaluó ese atributo en la presente tesis. En la Figura 12 se muestra la dinámica del peso específico foliar desde emergencia hasta madurez de cosecha del cultivo. Los valores del peso específico foliar, en densidad convencional fueron entre 44 y 76 g m-2 y 43 y 73 g m-2 en Pantera y WBLL, respectivamente. En baja densidad, se registró en Pantera entre 56 y 65 g m-2 y en WBLL desde 61 hasta 92 g m-2. Los valores fueron mayores en baja densidad a lo largo de casi todo el ciclo de cultivo de ambos genotipos (Figura 12). a) b) Figura 12. Peso específico foliar hasta madurez fisiológica en (a) PANTERA–INIA (círculos) y (b) WBLL 1*2/KUKUNA (cuadrados). En densidad convencional (símbolos abiertos) y baja densidad (símbolos cerrados). 4.9 Lectura SPAD Al Igual que la densidad, la concentración de clorofila en las hojas es un atributo importante para la EUR y también para la calidad de los granos. Los valores de concentración de clorofila, estimados a través de SPAD, alcanzaron un rango de 47,2 55,3 y 43,4 - 49,9 en densidad convencional de Pantera y WBLL, respectivamente 34 (Figura 13). En baja densidad los valores de Pantera fluctuaron entre 53,2 - 55,8 y 50,2 - 54,1 en WBLL. Estos valores demuestras que la baja densidad superó levemente la concentración de clorofila de la densidad convencional a lo largo del ciclo de cultivo (Figura 13). b) a) Figura 13. Lectura SPAD hasta madurez fisiológica en (a) PANTERA–INIA (círculos) y (b) WBLL 1*2/KUKUNA (cuadrados). En densidad convencional (símbolos abiertos) y baja densidad (símbolos cerrados). 4.10 Nitrógeno absorbido y concentración de proteína en granos El N absorbido total, y en órganos particulares del cultivo, no mostraron diferencias (p>0,05) entre Pantera y WBLL como se muestra en el Cuadro 5. Tampoco se encontró interacción entre el genotipo y la densidad de siembra (p>0,05) para los indicadores de la economía del nitrógeno del cultivo. El N absorbido en granos, tallos y hojas fue superior (p<0,01, p<0,05 y p<0,001, respectivamente) en densidad convencional sobre la baja densidad. Los capotillos fueron los únicos órganos donde fue mayor (p<0,05) en baja densidad sobre la densidad convencional (Cuadro 5). La partición del nitrógeno en la biomasa aérea (ICN) no presentó diferencias (p>0,05) entre los genotipos, ya que en densidad convencional el ICN fue de 77,1 y 77,8% en 35 Pantera y WBLL, respectivamente. En la baja densidad fue de 77,6 y 82,3% en Pantera y WBLL, respectivamente (Cuadro 5). En baja densidad, este porcentaje superó significativamente (p<0,01) al de alta densidad, en 0,6 y 5,5% superior sobre la densidad convencional en Pantera y WBLL, respectivamente. Cuadro 5. Nitrógeno absorbido en granos, capotillos, tallos y hojas, en PANTERA– INIA y WBLL 1*2/KUKUNA creciendo bajo condiciones de densidad convencional y baja densidad. Genotipo Densidad -2 (pl m ) N abs N abs N abs N abs N abs ICN total granos capotillos tallos Hojas (%) -2 -2 -2 -2 -2 (g m ) (g m ) (g m ) (g m ) (g m ) 300 42,3 32,5 2,77 4,37 2,61 77,1 44 34,6 26,8 3,34 2,79 1,67 77,6 300 38,9 30,4 2,07 3,90 2,64 77,8 44 32,0 26,3 2,27 2,09 1,33 82,3 Genotipo N.S N.S N.S N.S N.S N.S Densidad ** ** * ** *** * N.S N.S N.S N.S N.S N.S PANTERA-INIA WBLL1*2/KUKUNA Genotipo: Densidad La concentración de proteína en grano no mostró diferencias entre los genotipos (p>0,05), ni interacción entre el genotipo y la densidad de siembra (Cuadro 6). Por el contrario, la baja densidad de siembra incrementó (p<0,05) la concentración de proteína en grano por sobre la densidad convencional, fue superior a la densidad convencional en un 1,6 y 0,4% en Pantera y WBLL, respectivamente (Cuadro 6). 36 Cuadro 6. Concentración de proteína en grano, en PANTERA–INIA y WBLL 1*2/KUKUNA creciendo bajo condiciones de densidad convencional y baja densidad. Genotipo Densidad -2 PANTERA-INIA WBLL1*2/KUKUNA Concentración de proteína (plantas m ) en grano (%) 300 12,6 44 14,2 300 11,5 44 11,9 Genotipo N.S Densidad * Genotipo: Densidad N.S 37 5 DISCUSIÓN DE RESULTADOS El objetivo de la presente tesis fue cuantificar el efecto de dos densidades de siembra contrastantes, y con diferente arreglo espacial; sobre el rendimiento, la producción de biomasa aérea, sus determinantes fisiológicos y la concentración de proteína de los granos en dos genotipos de trigo primaverales de alto potencial de rendimiento. Esta evaluación se realizó bajo manejo agronómico óptimo en las condiciones agroclimáticamente favorables del sur de Chile. Entre Pantera y WBLL no se encontraron diferencias en rendimiento. Sin embargo, la densidad de siembra sí mostró diferencias contrariamente a lo esperado en base a resultados anteriores. En esta tesis se encontró que la densidad convencional logró rendimientos significativamente mayores a los de baja densidad con una diferencia de 387 y 236 g m-2 en Pantera y WBLL, respectivamente. Posiblemente, la igualdad de rendimientos esperada entre las densidades de siembra se vio afectada por la fecha de siembra. El experimento de Bustos et al. (2013), en el que la baja densidad (44 pl m-2) sí logró igualar los rendimientos de la alta densidad (350 pl m-2), la fecha de siembra fue el 22 de agosto, mientras que, en la presente tesis, la siembra se realizó dos semanas después (5 de septiembre), reduciendo así la capacidad de macollaje (Figura 14). 38 Figura 14. Número de espigas por metro cuadrado en PANTERA–INIA y WBLL 1*2/KUKUNA. En densidad convencional y baja densidad. El rendimiento del cultivo es la resultante de los componentes numéricos del mismo (Cuadro 1). Los componentes del rendimiento dependen de factores genéticos, a menudo ligados a la precocidad del cultivar y su relación con la capacidad de producir macollos (duración del período entre inicio y término de macollaje), factores ambientales de manejo como la radiación fotosintéticamente activa capturada por el cultivo y la disponibilidad de agua y nutrientes, y de factores de manejo agronómicos como la fecha de siembra, densidad de siembra y manejo de la fertilización, y la neutralización de elementos fitotóxicos como el pH y la concentración de aluminio en el suelo, entre otras causas (Evans et al., 1975). Dentro del modelo numérico, el número de granos por unidad de superficie es el principal determinante del rendimiento (Calderini et al., 1995; Slafer et al., 1996). Esta afirmación ha sido corroborada en el presente estudio, los mayores rendimientos se alcanzaron con mayor número de granos por metro cuadrado (Figura 14a). Los componentes del número de granos por unidad de superficie dependen del número de plantas m -2, el número de espigas por planta y el número de granos por espiga. El número de plantas constituyó una de las fuentes de variación de la presente tesis y los otros dos fueron condicionados por este tratamiento. El número de espigas por planta y el número de grano por espiga fueron mayores en baja densidad, sin embargo, no lograron generar un rendimiento que 39 sea similar al de la densidad convencional. El menor número de espigas por unidad de superficie de la baja densidad apoyaría la sugerencia que el menor rendimiento de esta densidad de siembra comparada con la convencional se debió a un periodo de macollaje más corto por la fecha de siembra, y más tardía que en el experimento de Bustos et al. (2013). El otro componente del rendimiento es el peso promedio de los granos. En el tratamiento de baja densidad, este componente se incrementó un 2,0 y 12,2% en Pantera y WBLL, respectivamente. A pesar de que el número de granos por unidad de superficie es la variable que mejor explica el rendimiento, cambios en el peso de los granos pueden influir en el rendimiento final del cultivo de trigo, una vez establecido el número final de granos por unidad de superficie (Figura 14b). Sin embargo, el incremento del P1000 registrado en la presente tesis no logró compensar el menor NG en la siembra de 44 pl m-2. El mayor peso de grano pudo deberse a que los ovarios de las flores en desarrollo alcanzaron mayor peso y tamaño al momento de antesis como se ha mostrado anteriormente en trigo (Calderini y Reynolds, 2000; Hasan et al., 2007) y en otros cultivos como cebada (Scott et al., 1983), maravilla (Castillo et al., 2017) y sorgo granífero (Yang et al., 2009). 40 b) Rendimento (g m-2) a) Figura 15: Componentes numéricos del rendimiento en PANTERA–INIA (círculos) y WBLL 1*2/KUKUNA (cuadrados), en función al número de granos m-2 (a) y peso de 1000 granos m-2 en densidad convencional (símbolos abiertos) y baja densidad (símbolos cerrados). Estudios previos han mostrado distintas respuestas del rendimiento de trigo a los niveles de densidad de plantas, las cuales, tuvieron relaciones positivas, negativas e invariabilidad (Nakano et al., 2009; Bustos et al., 2013; Zhang et al., 2016). Esto ha demostrado un claro efecto asintótico, en donde existe un valor critico de densidad de plantas en el que se mantendrá el máximo rendimiento. En la presente tesis, es importante destacar que los cultivos llegaron al periodo crítico de la determinación del número de granos (Fisher, 1985) en condiciones diferentes. El IAF en la densidad convencional, tanto en pre-antesis como en post-antesis, fue superior al alcanzado en la de baja densidad, con valores máximos de 6,5. Por otra parte, en baja densidad fue entre 3 y 4. Estudios realizados en la Estación Agropecuaria Santa Rosa, con densidades de siembra convencionales, han obtenido valores de IAF de 6 (Bustos et al., 2013), mientras que, en baja densidad Bustos et al. (2013) reportaron un valor máximo de 4,5. La competencia intraespecífica y las modificaciones en el ambiente lumínico de los genotipos en alta densidad de siembra determinan que la expansión foliar y el macollaje disminuyan, finalizando con una menor captura de radiación en los órganos más 41 cercanos al suelo por el sombreamiento de las hojas superiores, los tallos y las espigas. Esto genera la senescencia de los órganos vegetativos (Satorre et al., 2003). El principal determinante de la IR es el IAF (Thorne et al., 1988 citados por Carcova et al., 2003), la IR entre Pantera y WBLL fue similar durante todo el periodo del cultivo, no obstante, entre densidades contrastantes, la densidad convencional alcanzó la intercepción completa de radiación (95%) en menor tiempo que en baja densidad. La EUR es la pendiente de la relación entre la biomasa aérea acumulada y la radiación interceptada acumulada (Sinclair et al., 1999). La EUR registrada en los tratamientos de esta tesis, no influyó de la misma manera que la IR sobre la producción de biomasa. En la fase de pre-antesis, la EUR fue de alrededor de 3 g MJ-1 m-2 en la densidad convencional y en la baja densidad. Ese valor de EUR estuvo por debajo los 3,4 g MJ-1 m-2 registrados por Bustos et al. (2013). En post-antesis, la EUR disminuyó en ambas densidades respecto de pre-antesis, donde la EUR en densidad convencional fue de 2,9 y 2,6 g MJ-1 m-2 en Pantera y WBLL, respectivamente. En baja densidad los valores de EUR fueron 2,4 y 2,6 g MJ-1 m-2 en esos genotipos, respectivamente. Esos valores fueron menores a los 3,2 g MJ-1 m-2, registrados por Bustos et al. (2013). Considerando que los determinantes fisiológicos de la producción de biomasa son la intercepción de radiación (IR) y la eficiencia en el uso de la radiación (EUR), la densidad convencional, también mostró una tasa de producción de biomasa aérea ligeramente superior a la de baja densidad durante todo el período del cultivo, dando como resultado, una mayor biomasa aérea a cosecha en densidad convencional. Las mediciones de concentración de clorofila mediante el SPAD es un método simple y útil para estimar indirectamente la capacidad fotosintética, al estar ésta relacionada con el contenido en clorofilas de las hojas (Gummuluru et al., 1989). Los niveles de SPAD fueron levemente más altos en ambos genotipos (Pantera y WBLL) en baja densidad durante pre-antesis y post-antesis. En la presente tesis también se evaluó la economía del nitrógeno en ambos genotipos y densidad de siembra. Coincidiendo con la producción de biomasa aérea y el rendimiento, la absorción total de N en la biomasa aérea fue mayor en la densidad convencional, sin que se observaran diferencias entre los genotipos. 42 La calidad del grano, entendida como concentración de proteína en grano, entre densidades de siembra contrastantes y con diferente arreglo espacial, mostró diferencias, las que son respaldadas por resultados anteriores. En esta tesis se encontró que la baja densidad logró una concentración de proteína en grano significativamente mayor a los de densidad convencional con una diferencia de 1,6 y 0,4% en Pantera y WBLL, respectivamente. Lo mismo ocurrió con el ICN, donde la baja densidad de siembra también superó significativamente la densidad convencional, sin presentar diferencias entre Pantera y WBLL. Es probable que la mayor concentración de proteína en grano se deba una mayor partición de este nutriente (ICN) hacia las estructuras reproductivas. Sin embargo, también se debe considerar que el mayor rendimiento logrado por la densidad convencional genera un efecto de dilución del nitrógeno de los granos en una mayor cantidad de materia seca. Lemaire et al. (2005) propone que las plantas expuestas a menor densidad de siembra presentan un mayor índice de cosecha de nitrógeno, debido al mayor desarrollo foliar, a diferencia de las plantas expuestas a densidades convencionales, donde presentan un mayor desarrollo de tallos. Los tejidos foliares tienen una mayor concentración de N, en comparación con los vástagos, generando un mayor índice de cosecha de nitrógeno, por ende, una mayor producción de proteína en baja densidad de siembra. 43 6 CONCLUSIONES La igualdad de rendimientos esperada entre las densidades de siembra con diferente arreglo espacial no se verificó en esta tesis. Posiblemente esto fue debido a la fecha de siembra más tardía que en experimentos anteriores como el de Bustos et al. (2013). El número de granos por unidad de superficie fue el principal determinante del rendimiento, donde la densidad convencional superó a la baja densidad de siembra en un 26,4% como promedio de los dos genotipos evaluados. Los rendimientos alcanzados por Pantera y WBLL en baja densidad de siembra fueron potenciales, esto se debió también a su gran capacidad compensatoria de macollaje, superando a la densidad convencional, en un 3,5 y 2,7 veces el número máximo y final de macollos por planta, respectivamente. La baja densidad de siembra logró un mayor P1000 por sobre la densidad convencional, pero este incremento no fue suficiente para compensar el menor número de granos por unidad de superficie. La IR fue el principal determinante fisiológico de la producción de biomasa, donde la densidad convencional alcanzó la intercepción completa de la radiación en menor tiempo térmico. Sin embargo, la eficiencia de intercepción de radiación (radiación interceptada en términos porcentuales) mostró la misma dinámica entre las densidades de siembra y genotipos. La EUR registrada en los tratamientos de esta tesis, no fue afectada por la densidad de siembra. En la fase de pre-antesis ambas densidades de siembra alcanzaron la misma EUR y en post-antesis, la EUR disminuyó en ambas densidades respecto de pre-antesis. 44 La baja densidad de siembra superó significativamente a la densidad convencional en la partición del nitrógeno en la biomasa aérea y en la concentración de nitrógeno en grano. 45 7 REFERENCIAS BIBLIOGRAFICAS ABBATE, P.; ANDRADE, F. y CULOT, J. 1995. The effects of radiation and nitrogen on number of grains in wheat. Journal of Agricultural Science. 124: 351-360. ARDUINI, I., MASONI, A., ERCOLI, L. y MARIOTTI, M. 2006. Grain yield, and dry matter and nitrogen accumulation and remobilization in durum wheat as affected by variety and seeding rate. European Journal of Agronomy. 25: 309 – 318. AULD, B., KEMP, D. y MEDD, R. 1983. The Influence of spatial arrangement on grain yield of wheat. Crop & Pasture Science. 34: 99 – 108. BÉNÉTRIX, F. y AUTRAN, J.C. 2001. Protein synthesis in grains and seeds. En: Nitrogen Assimilation by Plants. Physiological, Biochemical and Molecular Aspects, J.F. Morot-Gaudry. (Ed), Science Publishers Inc., Enfield, NH, pp 343-360. BUSTOS, D.V., HASAN, A.K., REYNOLDS M.P. y CALDERINI. D.F. 2013. Combining high grain number and weight through a DH-population to improve grain yield potential of wheat in high-yielding environments. Field Crops Research. 145: 106115. CALDERINI, D., DRECCER, M. y SLAFER, G. 1995. Genetic improvement in wheat yield and associated traits. A re-examination of previous results and the latest trends. Plant Breeding. 114: 108-112. CALDERINI, D., DRECCER, M. y SLAFER, G. 1997. Consecuences of breeding on biomass, radiation-use efficiency in wheat. Field Crops Research. 52: 271-281. CALDERINI, D. F., ABELEDO, L. G., y SLAFER, G. A. 2000. Physiological maturity in wheat based on kernel water and dry matter. CARCOVA, J., ABELEDO, G. y LOPEZ-PEREIRA, M. 2003. Análisis de la generación del rendimiento: Crecimiento, partición y componentes. In: Satorre, E., Benech Arnold, R., Slafer, G., De La Fuente, E., Miralles, D., Otegui, M. y Savin, R. Producción de granos. Bases funcionales para su manejo. Universidad de Buenos Aires, Facultad de Agronomía. pp 75-95. 46 CASTELLARÍN, J. 2009. Influencia de las prácticas de manejo sobre la generación del rendimiento en el cultivo de trigo. Revista Trigo (Argentina). (40): 23 – 27. CAVIGLIA, O. P. y SADRAS, V.O. 2001. Effect of nitrogen supply on crop conductance, water and radiation use efficiency of wheat. Filed crops Res. 69: 259-266. CENTRO INTERNACIONAL DE MEJORAMIENTO DE MAÍZ Y TRIGO (CIMMYT). 2016. Investigación del trigo. (Online) <https://www.cimmyt.org/es/nuestro-trabajo/trigo/>. CHEN, CH., NEILL, K., WICHMAN, D. y WESCOTT, M. 2008. Hard red spring wheat response to row spacing, seeding rate nitrogen. Agronomy Journal 100: 12961302. CHILE, INSTITUTO DE INVESTIGACIONES AGROPECUARIAS, MINISTERIO DE AGRICULTURA (INIA). 2014. Variedades de Trigo Recomendado por INIA para la Región de La Araucanía. (Online). <http://biblioteca.inia.cl/medios/biblioteca/informativos/NR40225.pdf>. CHILE, OFICINA DE ESTUDIOS Y POLÍTICAS AGRARIAS (ODEPA). 2020. Boletín de Cereales, Producción, precios y comercio exterior de trigo, maíz y arroz (On line). <https://bibliotecadigital.odepa.gob.cl/bitstream/handle/20.500.12650/70454/Bolet in-cereales202008.pdf> 9p. CIAMPITTI, I. y VYN, T. 2012. Physiological perspectives of changes over time in maize yield dependency on nitrogen uptake and associated nitrogen efficiencies: A review. Field Crops Research 133: 48–67. DAUGHTRY, C., WALTHALL, C., KIM, M., BROWN DE COLSTOUN, E., AND MCMURTREY, J. 2000. Estimating Corn Leaf Chlorophyll Concentration from Leaf and Canopy Reflectance. Remote Sensing of Environment 74: 229-239. DUGGAN, B., RICHARDS, R., VAN HERWAARDEN, A. y FETTEL, N. 2005. Agronomic evaluation of a tiller inhibition gene (tin) in wheat. I. Effect on yield, yield components, and grain protein. Australian Journal of Agricultural Research 56 (2): 169-178. DUMAS, JB., LETTRE DE M., DUMAS A M., GAY-LUSSAC, SUR LES PROCE ́DE S ́ DE L’ANALYSE ORGANIQUE. Ann Chim Phys 1831; 2(47): 198–215. 47 EGLI, D. 2017. Seed Biology and Yield of Grain Crops. 2 ed. Lexington, USA. CABI publishing. 233 pp. ELLIS, R.; SALAHI, M. y JONES, S. 1999. Yield-density equations can be extended to quantify the effect of applied nitrogen and cultivar on wheat grain yield. Annals of Applied Biology. 134: 347–352. ESCOBAR, A. y COMBE, L. 2012. Senescence in field-grown maize: From flowering to harvest. Field Crops Research 134: 47–58. EVANS, L., WARDLAW, I. y FISCHER, R. 1975. Chapter 5: Wheat. p. 101-149. In Evans, L. T. (Ed.) Crop physiology. Cambridge University Press, Cambridge. 374 p. EVERS, j., VOS, J., ANDRIU, B. y STRUIK, P. 2006. Cessation of Tillering in Spring Wheat in Relation to Light Interception and Red: Far-red Ratio. Annals of Botany, volume 97, Issue 4, Pages 649-658. FISCHER, R. 1985. Number of kernels in wheat crops and the influence of solar radiation and temperature. Journal of Agricultural Science. 105: 447–461. FISCHER, R. 2020. Breeding wheat for increased potential yield: Contrasting ideas from Donald and Fasoulas, and the case for early generation selection under nil competition. Field Crops Research. Volume 252, 2020l, Article 107782. GALLAGHER, J. N. y BISCOE, B. 1978. Radiation absorption, growth and yield of cereals. J. Agric. Sci. 91: 47-60. GIFFORD, R., THORNE, J., HITZ, W. y GIAQUINTA, R. 1984. Crop Productivity and Photoassimilate Partitioning. Science Vol. 225. no. 4664, pp. 801 – 808. GUMMULURU, S., HOBBS, S. L. A. y JANA, S. 1989. Physiological responses of drought tolerant and drought susceptible durum wheat genotypes. PhotoSynthetica 23: 479-485. HIPPS, L. E., ASRAR, G. y KANEMASU, E. T. 2005. Assessing the interception of photosynthetically active radiation in winter wheat. Agricultural Meteorology, 28 (1983) 253—259. 48 HIREL, B., MARTIN, A., TERCÉ, T., GONZALEZ, M. y ESTABILLO, J. 2005. Physiology of maize I: A compreshensive and integrated view of nitrogen metabolism in a C4 plant. Physiologia Plantarum 124: 167-177. JAMIESON, P.D., MARTIN, R.J. y FRANCIS, J.S. 1995. Drought influences on grain yields of barley, wheat and maize. New Zeal J. Crop Hort Sci. Vol: 23:55-66. LADHA, J., PATHAK, H., KRUPNIK, T., SIX, J. y VAN KESSEL, C. 2005. Efficiency of fertilizer nitrogen in cereal production: retrospects and prospects. Advances in Agronomy 87: 85-156. LEMAIRE, G.; AVICE, J.; KIM, T. y OURRY, A. 2005. Developmental changes in shoot N dynamics of lucerne (Medicago sativa L.) in relation to leaf growth dynamics as a function of plant density and hierarchical position within the canopy. Journal of Experimental Botany. 56: 935–943. MAC KEY, J. 2005. Wheat: Its concept, evolution, and taxonomy. En: Durum Wheat Breeding: Current Approaches and Future Strategies, C. Royo, M.M. Nachit, N. DiFonzo, J.L. Araus, W.H. Pfeiffer, G.A. Slafer (eds.), vol. 1, The Haworth Press, Inc., New York, pp. 3-61. MERINO, M. 2017. La Revolución Neolítica: ¿por qué adoptar la agricultura y una forma de vida sedentaria? ResearchGate. 7 p. MELLADO, M. 1998. Análisis del cultivo del trigo en Chile durante el siglo veinte. Agricultura Técnica. 58(3): 230-240. MIRASSÓN, H., FARALDO, M.L., FERRERO, C., BREVEDAN, R., FIORETTI, M.E., FERNÁNDEZ, M., ZINGARETTI, O., DE DURANA, J., ANDREOTTI B., y MARTÍNEZ, P. 2008. Crecimiento y desarrollo de trigo de diferentes ciclos sembrados a 3 espaciamientos entre hileras, bajo siembra directa en la región semiárida pampeana. Actas VII Congreso Nacional de Trigo V Congreso Nacional de Cereales de Siembra otoño-invernal y I Encuentro del MERCOSUR. Julio 2008. Santa Rosa. MOHKUM, H., AHMAD, A., WAJID, A. y AKHTER, J. 2011. Maize response to time and rate of nitrogen application. Journal Botanical Pakistán 43: 1935-1942. 49 MURCIA, J.L. 2017. Tendencias internacionales en panadería y bollería. Pan artesano y masas congeladas confluyen en una oferta tan variada como atractiva. Volumen 2: 48-53. NAKANO, H. y MORITA, S. 2009. Effects of seeding rate and nitrogen application rate on grain yield and protein content of the bread wheat cultivar ‘Minaminokaori’ in southwestern Japan. Plant Production Science. 12: 109-115. NING, T., ZHENG, Y., HA, H., JIANG, G. y LI, Z. 2012. Nitrogen uptake, biomass yield and quality of intercropped spring- and summer-sown maize at different nitrogen levels in the North China Plain. Biomass and Bioenergy 47:1-8. ORGANIZACIÓN DE LAS NACIONES UNIDAD PARA LA ALIMENTACIÓN Y LA AGRICULTURA, (FAO), (on line) Consultado en el año 2019. <http://faostat.fao.org/site/567/DesktopDefault.aspx?PageID=567#ancor>. PAGANI, A., ECHEVERRÍA, H., ANDRADE, F. y SAINZ, H. 2009. Characterization of Corn Nitrogen Status with a Greenness Index under Different Availability of Sulfur. Agronomy Journal 101: 315- 322. PEÑA, R. 2002. Wheat for bread and other foods. In: Bread wheat, Improvement and production. Organización de las naciones Unidas para la agricultura y alimentación (FAO). Roma, Italia. QUIÑONES, F. 2007. Variación de la producción de materia seca y eficiencia de absorción de nitrógeno en cultivares antiguos y modernos de trigo (Triticum aestivum) a dos niveles de disponibilidad de nitrógeno en suelos volcánicos. Licenciado en Agronomía. (On line). Valdivia. Universidad Austral de Chile. Facultad de Ciencias Agrarias. 85 http://cybertesis.uach.cl/tesis/uach/2007/faq.7v/doc/faq.7v.pdf>. p. (13 < de enero 2016). ROBINSON, R.G. 1978. Production and culture. In: J.F. Carter (Ed) Sunflower Science and Technology nº 19 Agronomy Serie. ASA, CSSA, SSSA, Madison WI, Pp. 89143. 50 SATORRE, E., BENECH, R., SLAFER, G., DE LA FUENTE, E., MILLARES, D., OTEGUI, D. y SAVÍN, S. 2003. Producción de granos: Bases funcionales para su manejo. Editorial Facultad de Agronomía Universidad de Buenos Aires, Argentina. 783 p. SATORRE, E.; BENECH, R.; SLAFER, G.; DE LA FUENTE, E.; MIRALLES, D.; OTEGUI, M. y SAVIN, R. 2004. Producción de granos: bases funcionales para su manejo. Buenos Aires. Editorial Facultad de Agronomía, Universidad de Buenos Aires. 298 – 311. SAVIN, R. y SLAFER, G. 1991. Shading effects on the yield of an Argentinian wheat cultivar. Journal of Agricultural Science. 116: 1-7. SLAFER, G., CALDERINI, D. y MIRALLES, D. 1996. Yield Components and compensation in wheat: Opportunities for further increasing yield potential. In: Reynolds M, Rajaram S y McNab A. Increasing yield potential in wheat: 70 Breaking the barriers. Ciudad Obregón, México; CIMMYT Int. Simposio. pp 101- 134. SLAFER, G., SAVIN, R. y SADRAS, V. 2014. Coarse and fine regulation of wheat yield components in response to genotype and environment. Field Crops Research. 157: 71 – 83. SIEMENS, L.B. 1963. The effects of varying row spacing on agronomic and quality characteritics of cereal and flax. Can J. Plant Sci. 43, 119-130. SINCLAIR, T. y MUCHOW, R. 1999. Radiation use efficiency. Advances in Agronomy. 65: 217-265. STÖCKLE, C.O., KEMANIAN, A.R., 2009. Crop radiation capture and use efficiency: a framework for crop growth analysis. In: Calderini, D.F., Sadras, V.O. (Eds.), Crop Physiology - Applications for Genetic Improvement and Agronomy. Academic Press, San Diego, pp. 145–170. SZÉLES, A., MEGYES, A. y NAGY, J. 2012. Irrigation and nitrogen effects on the leaf chlorophyll content and grain yield of maize in different crop years. Agricultural Water Management 107: 133– 144. TAKAHASHI, T. y NAKASEKO, K., 1993. Seasonal changes in distribution of intercepted photosynthetically active radiation for layer and dry matter production in spring wheat canopy. J. Crop Sci., 62: 313-318. 51 TELLER, G.L. 1932. Non-protein nitrogen compounds in cereals and their relation to the nitrogen factor for protein in cereals and bread. Cereal Chemistry 9, 261-274. THORNE, G.N., PEARMAN, I., DAY, W. y TODD, A.D., 1988. Estimation of radiation interception by winter wheat from measurements of leaf area. J. Agric. Sci., 110: 101-108. TOMPKINS, D.K., HUULTGREEN, G.E., WRIGHT, A.T. y FOWLER, D.B. 1991. Seed rate and Row Spacing of No-Till Winter Wheat. Agron J 83:684-689. TROCCOLI, A., BORRELLI, G.M., DE VITA, P., FARES, C. y DI FONZO, N. 2000. Durum wheat quality: A multidisciplinary concept. J. Cereal Sci. 32: 99-113. TURNER, N.H., PRASERTSAK, P. y SETTER, T.L. 1994. Plant spancing, density and yield of wheat subjected to postanthesis water deficit. Crop Science 34: 741-748. VARINDERPAL-SINGH, YADVINDER-SINGH, BIJAY-SINGH, THIND, H., KUMAR, A. y VASHISTHA, M. 2011. Calibrating the leaf colour chart for need based fertilizer nitrogen management in different maize (Zea mays L.) genotypes. Field Crops Research 120: 276–282. VERA, R. E. 2011. Efecto de la dosis de siembra sobre el rendimiento y la calidad de trigo candeal (triticum turgidum l. ssp. durum) variedad llareta INIA, en condiciones de secano mediterráneo. Universidad de Chile, Chile. 39 p. ZADOKS, J., CHANG, T. y KONZAK, C. 1974. A decimal code for the growth stages of cereals. Weed Research. 14(6): 415-421. ZHANG, Y., DAI, X., JIA, D., LI, H., WANG, Y., LI, C., XU, H. y HE, M. 2016. Effects of plant density on grain yield, protein size distribution, and breadmaking quality of winter wheat grown under two nitrogen fertilisation rates. European Journal of Agronomy. 73: 1–10.