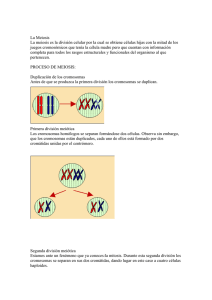
“AÑO DEL DIALOGO Y LA RECONCILIACIÓN NACIONAL” UNIVERSIDAD NACIONAL AUTÓNOMA DE CHOTA ESCUELA PROFESIONAL DE INGENIERÍA AGROINDUSTRIAL TEMA: LA MEIOSIS ASIGNATURA: BIOLOGIA PROFESOR: QUESQUEN CHANCAFE, Cristian AUTORES: MEJIA BURGA, Celeny Yohana TARRILLO TARRILLO, Rocío Milagros Chota-Perú 2018 INDICE INTRODUCCIÓN --------------------------------------------------------------------------------------5 OBJETIVOS---------------------------------------------------------------------------------------------6 Objetivos generales Objetivos específicos MARCO TEÓRICO ------------------------------------------------------------------------------------7 3.1 PRIMERA DIVISIÓN DE LA MEIOSIS------------------------------------------------8-13 Interfase I Profase I Leptoteno Cigotene Paquitene Diplotene Diasinisis Estado difuso Metafase I Anafase I 3.2 SEGUNDA DIVISIÓN DE MEIOSIS-------------------------------------------------------14 Profase II Metafase II Anafase II Telofase II FUNCIÓN-------------------------------------------------------------------------------------------15 CONCLUSIONES--------------------------------------------------------------------------------16 BIBLIOGRAFÍA------------------------------------------------------------------------------------17 2 DEDICATORIA: De manera muy especial dedico este informe a Dios el, cual fue el creador de todas las cosas, el que nos ha dado las fortalezas para continuar en nuestros estudios, a nuestros a queridos padres y hermanos por el apoyo moral y económico constantemente para mi formación profesional; a todos los docentes que cada día participan con su labor educativa y eficiente, siendo exigentes en el aprendizaje del alumno y a todas esas personas que me brindaron su apoyo para poder logar mis objetivos trazados, en especial al profesor, Cristian Quesquén Chancafe por haber guiado con el desarrollo de este trabajo y llegar a culminar del mismo. 3 AGRADECIMIENTO Expreso mi sincero agradecimiento en primer lugar a Dios, por haberme guiado en la elaboración del presente informe. A nuestros queridos padres por su gran apoyo incansable e insuperable día a día para llegar a nuestras metas, por valorarnos y por sus consejos para lograr ser unos profesionales de éxito con muchos sueños empresariales. Nuestro cordial agradecimiento a la “UNIVERSIDAD NACIONAL AUTÓNOMA DE CHOTA”, en donde nos estamos formando como profesionales en la especialidad de Ing. agroindustrial durante el tiempo transcurrido. Al Profesor, Cristian Quesquen Chancafe por el desarrollo para la realización de dicha monografía. A todos los profesores, quienes compartieron sus experiencias, nuevos conocimientos, nuevas ideas, inquietudes y por exigirnos día a día a superarnos más y ser los mejores a través de su apoyo incondicional. 4 I. INTRODUCCION La meiosis es un proceso de división celular por el que a partir de una célula madre diploide (2n) se obtienen cuatro células hijas haploides (n). Durante la meiosis se producen dos divisiones celulares consecutivas conocidas como meiosis I y meiosis II. La primera de las divisiones, que es más compleja que la segunda, es una división reduccional en la cual se pasa de una célula diploide (con 2n cromosomas) a dos células haploides (con n cromosomas) cada una de ellas con 2n cromátidas. La segunda división es mucho más sencilla y similar a una división mitótica, y en ella a partir de las dos células haploides (n) anteriormente formadas se obtienen cuatro células haploides (n) con n c cromátidas cada una de ellas. 5 II. OBJETIVOS OBJETIVO GENERAL conocer un poco más a cerca de la meiosis y también saber sus etapas OBJETIVOS ESPECIFICOS Recordar las etapas y la importancia de la meiosis. Diferenciar entre estos procesos, reconociendo características propias de cada uno de ellos. 6 las III. MARCO TEORICO La importancia de la meiosis reside en que se trata del recurso por el cual se producen las gametas que participan de la reproducción sexual. En efecto, las células somáticas habituales de todos los seres procariotas incluyen, en condiciones normales, material genético que se encuentra duplicado, aunque no redundante. Estas células se designan en forma convencional como diploides, lo cual suele simbolizarse como 2n. En el proceso de meiosis, las células participantes presentan dos divisiones celulares consecutivas, tras lo cual se producen cuatro elementos celulares que incluyen la mitad del material genético inicial. Estas células finales se llaman haploides y se denominan como 1n ó n por convención. En el contexto de la profase, los componentes de cada uno de los pares de cromosomas se emparejan para dar lugar a una recombinación de su contenido genético. En una etapa subsiguiente, conocida como metafase, los cromosomas resultantes de este fenómeno se ubican en un plano central, para luego migrar hacia los polos celulares en la etapa denominada anafase. Por consiguiente, cada una de las células originadas durante esta sucesión de etapas (conocidas en forma conjunta como meiosis I) cuenta con la mitad del genoma de la célula original. En la posterior meiosis II, estas células haploides se dividen para dar lugar a nuevos elementos celulares, cuya maduración final dará lugar a óvulos y espermatozoides. Además de asegurar la reproducción sexual, la importancia de la meiosis consiste en asegurar la variabilidad genética, dado que el proceso de recombinación del contenido genético permite que los descendientes originados a partir de las gametas conserven gran parte de las características de sus ancestros, pero dando al nuevo individuo un perfil absolutamente único y distintivo. A diferencia de la natural partenogénesis o de la artificial clonación, en la cual la reproducción surge de una nueva mitosis, la meiosis produce ejemplares realmente diferentes a sus padres, abriendo la posibilidad de nuevas adaptaciones y relaciones con el entorno biológico y, en el caso de los seres humanos, del ambiente psicológico y social. 7 ETAPAS DE LA MEIOSIS 3.1 Primera división de la meiosis • Interfase I Puede ser variable en su duración, incluso puede faltar por completo de manera que tras la telofase I se inicia sin interrupción la segunda división meiótica. Aun habiendo un periodo de interfase no se produce nunca síntesis de ADN, es decir no hay periodo S. Profase I La profase I constituye un largo y complejo proceso durante el cual se produce el sobrecruzamiento y se preparan los cromosomas para reducir su número a la mitad. Leptoteno Los cromosomas aparecen filamentos y enmarañados en el núcleo. Cada cromosoma está constituido por dos cromátidas. La disposición en el núcleo puede aparecer totalmente anárquico o bien mostrar cierta polarización como si los telómeros de los cromosomas tendieran a concluir en una región determinada. Cigotene Los cromosomas homólogos empiezan a aparease, hasta lograr el apareamiento cromomero a cromomero la espiralizacion es mayor, 8 aunque los bivalentes aun no se individualizan en esta etapa se forma el complejo sinaptonémico, estructura que permite el apareamiento de los cromosomas homólogos. El complejo sinaptonémico es una estructura tripartita que aparece en los estadios de cigoteno, paquitene y luego desaparece, estando relacionado con el sobrecruzamiento meiótico. El complejo sinaptonémico está formado por los elementos laterales y un elemento central. La zona comprendida entre los elementos laterales y el central es menos a los electrones y se la denomina espacio central, estando atravesada por finos filamentos o fibrilias. Poco antes de que dos cromosomas comiencen a aparecerse aparece elemento axial, cuando dos elementos axiales se aproxima a una distancia conveniente empiezan a aparecer la estructura tripartida de modo q la región central del complejo sinaptonémico oscila entre 900 a 1200 A el central entre 200-300 Ay los laterales unos 500 A. La total del complejo sinaptonémico en neurosporaes de 50 A, la cual supone solo 0.3% de la longitud total de ADN, mientras que el maíz es de un 0,014 a un 0.017%. En todos los casos el complejo sinaptonémico empieza y termina en la membrana nuclear. La formación y composición química de los complejos sinaptonémicos se observa en la figura. Paquitene La espiralizacion progresiva de los cromosomas hace que los bivalentes queden individualizados uno unos de otros. Si las células somáticas de la especie tienen 2n cromosomas, en paquitene aparecen n bivalentes (apareados los dos homólogos). En este estadio los cromesomeros se hace visibles siendo constates el número, tamaño y posición, lo que ´permite identificar a cada bivalente. Es corriente en paquitene que algunos cromomeros se manifiesten como zona heteropignoticas positivas, llamadas knobs o nudos que se utilizan como maracadores cromosómicos. En esta etapa lo fundamental desde el punto de vista genético es que ocurre el crossing over o sobrecruzamiento (intercambio de cromatina entre cromátidas no hermanas de cromosomas homologas) y siendo 9 uno de los factores que produce variabilidad genética en las especies ya que permite intercambiar información genética entre los cromosomas paternos y maternos. Diplotene Continua el acortamiento de los cromosomas. Desaparece los complejos sinaptonemicos los bivalentes se mantienen apareados. En diplotene dejan de existir las causas o fuerzas que mantenían apareados a los cromosomas homólogos y comienzan a separarse los centrómeros de manera que se hacen visibles las estructuras cuadruples (dos cromáticas de cada cromosoma) en cada bivalente. En este estadio se observan los quiasmas. Aceptándose en la actualidad que el quiasma es la expresión citológica del sobrecruzamiento. En algunos organismos, especialmente en animales, los cromosomas diplotenicos aperecen difusos, siendo un caso extremo los cromosomas plomosos (lampbrush) de los oocitos de anfibios. Diacinisis Los cromosomas continúan espiralizándose y acortándose de manera que los bivalentes van perdiendo su forma alargada para poder pasar a tener forma redondeada, los bordes son más nítidos y los centrómeros homólogos inician la coorientacion, es tienden a situarse a ambos lados de la placa ecuatorial. Al final de la diasenisis, desaparecen el nucléolo y la membrana nuclear. A este estadio se le conoce como el de la terminilizacion de los quiasmas, actualmente existen evidencias experimentales que afirman que no existe el movimiento o desplazamiento de los quiasmas en la forma propuesta Darlington, sino que debe sustituirse por la resolución de los quiasmas. En este caso el quiasma a originado en paquitene por el sobrecruzamiento, mantiene unidos a los cromosomas homólogos, sin cambiar de posición durante diplotene y diasinisis hasta que, posiblemente por efecto de la topoisomerasa II, se produce su resolución al final de la metafase I. Sin embargo, puede haber cosas en la que se produzca una resolución precoz de los quiasmas se separan antes de tiempo los pares de 10 homólogos y aparecen los cromosomas como univalentes en metafase I. Estado difuso La existencia de un estado difuso en la profase de la primera división meiótica a sido demostrado tanto en las plantas como animales. Moens en 1964 al analizar secuencialmente la profase I del tomate demostró que después de paquitene los cromosomas de cada bivalente tendían a separarse quedando unidos por varios puntos, los cromosomas después perdían su avidez cromática a la vez que se extendían por todo el núcleo constituyendo algo así como una malla de fibras cromosómicas débilmente teñidas, a este lo llamo difuso. Después se observó en maíz Ascobolus, Basidiomsetes, Prunus, Lilium, remolacha, acelga, etc. Y en el reino animal en saltamonte, ratón, mono, grillos, etc. Una forma particular del estado difuso podría considerarse al estado de dictiotena en la ogenesis de la mujer y otros mamíferos como el ratón. En el caso particular de la mujer, la meiosis se inicia en los fetos femenino hacia los tres meses de gestación, deteniendo su desarrollo cuando alcanza el estado de diplotene partir del cual los oocitos entran en una fase de estado difuso (dictiotena). En estado permanecen hasta que llegada la madures sexual, cada vez madura un ovulo previo reanudando de la meiosis a partir de la diasenisis. Por consiguiente, la dictiotena puede durar entre 12 a 45 años. Metafase I Desaparece totalmente el nucléolo y la membrana nuclear. Los bivalentes alcanzan su máximo grado de contracción. Los centrómeros quedan perfectamente coorientados a ambos lados de la placa ecuatorial e si insertan en las fibras del huso. Anafase I En anafase I se produce la emigración de n cromosomas a cada polo, es decir tiene lugar la radion del número cromosómico. Si se trata de un individuo eterosigota Aa y ha habido sobre cruzamiento entre el centrómero y locus A es posible que un cromosoma anafasico lleve en un cromatidio en ungen A y en el otro su alelo a. 11 La diferencia esencial entre anafase I y un anafase mitótico estriva en que en anafase I se separan en n cromosomas no homologas en cada polo y en anafase mitótica en polo se encuentran dos n cromátidas no homologas en cada polo y en anafase mitótica en cada polo se encuentra 2n cromátidas (2n cromosomas con una sola cromátida). Telofase I Termina la emigración de los cromosomas agrupándose en los respectivos polos celulares. Los cromosomas se despiralizan y reaparece al nucléolo y la membrana nuclear se produce la citocinesis dando lugar a dos células diadas. En dicotiledóneas las células de la diada permanecen unidas mientras que en la monocotilodedoneas y en animales por lo general se produce la citocinesis de la diada y posteriormente cada célula por separado continua la segunda división meiótica. Interfase I Puede ser variable en su duración, incluso puede faltar por completo de manera que tras la telofase I se inicia sin interrupción la segunda división meiótica. Aun habiendo un periodo de interfase no se produce nunca síntesis de ADN, es decir no hay periodo S. 12 3.2 Segunda división de la meoisis La segunda división meiótica es esencialmente una mitosis, aunque con pequeñas diferencias. Profase II La característica más peculiar es la aparición de los cromosomas con sus cromatidios separados en forma de cruz. La separación de los cromatidios (manteniéndose solo unidos por el centrómero) se produce por la eliminación de las proteínas de cohesión. Metafase II Se disponen los n cromosomas, generalmente muy contraídos y con forma de cruz, en la placa ecuatorial. Anafase II Se separan n cromatidios a cada polo. Para que los cromatidios puedan migrar, previamente deben adquirir un centrómero propio, denominándolo a partir de ese momento cromatidio organizado como cromosoma. Telofase II Se termina la emigración de las cromátidas hacia los polos. Los cromosomas inician la desespiralización, aparece el nucléolo y la membrana nuclear, tiene lugar la citocinesis y se obtiene cuatro células con n cromosomas cada uno. A estas cuatro células se las suele denominar tétradas. 13 14 IV. FUNCIÓN DE LA MEIOSIS La meiosis cumple la función de producir gametos o células haploides (espermatozoides u óvulos) que no pueden vivir independientemente. Un espermatozoide se fusiona con un óvulo para producir una célula diploide, el óvulo fertilizado o cigoto, que mediante divisiones celulares mitóticas da origen a un nuevo organismo multicelular. Reducir el número de cromosomas (diploide a haploide) para que los gametos (células sexuales), al unirse durante la fecundación, vuelvan a formar células somáticas diploides. Promover la diversidad genética entre los individuos producidos para que la selección natural escoja los mejor adaptados. 15 V. CONCLUSIONES Que todas las células de cualquier planta o animal han surgido a partir de una única célula inicial el ovulo fecundado por in proceso de división En la meiosis es cuando en la fecundación se une dos gametos, la célula resultante, llamada cigoto, contiene toda la dotación doble de cromosomas. La mitad de estos cromosomas proceden de un progenitor y la otra mitad del otro. 16 VI. BIBLIOGRAFIA David T. Suzuki, Anthony J. F. Griffiths, Jeffrey H. Miller, Richard C. Lewontin Genética INTERAMERICANA-McGRAW-HILL, Madrid, 1992 Anthony J. F. Griffiths, William M. Gelbart, Jeffrey H. Miller, Richard C. Lewontin Genética moderna INTERAMERICANA-McGRAW-HILL, Madrid, 2000 Bruce Alberts, Dennis Bray, Julian Lewis, Martin Raff, Keith Roberts, James D. Watson Biología molecular de le célula EDICIONES OMEGA, Barcelona, 1987 Don W. Fawcett, M.D. Tratado de Histología NTERAMERICANAMcGRAW-HILL, Madrid, 1988 Barbara Young, John W. Heath Wheater's Histología funcional HARCOURT, Madrid, 2000 17