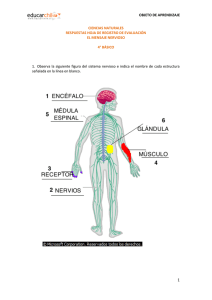
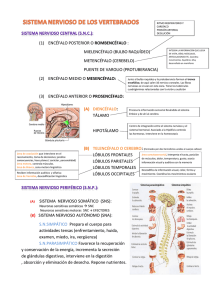
Unidad 01
Anuncio

Unidad 1 • La Psicología del Desarrollo como Ciencia CONCEPTO Y OBJETO DE LA PSICOLOGÍA DEL DESARROLLO La Psicología del desarrollo podemos definirla como la ciencia que estudia el cambio de comportamiento humano a lo largo del ciclo vital. Conviene hacer algunas precisiones en tomo a algunos de los términos más fundamentales empleados en esta definición. - Según cuanto acabamos de decir, el objeto o campo de estudio de esta ciencia es el cambio de comportamiento. La naturaleza de este cambio difiere, con todo, de otro tipo de cambios de naturaleza psicológica, como puede ser el provocado por la psicoterapia o el aprendizaje, limitados a un lapso de tiempo más corto. El cambio a que se refiere la Psicología del desarrollo es de una mayor trascendencia. - La entidad de observación de dicho cambio son los seres humanos. Lo cual no significa que ignore la existencia de una Psicología del desarrollo animal, cuyos datos pueden ser de gran utilidad en la Psicología del desarrollo de la persona. - El cambio se refiere al comportamiento del hombre. En relación con este cambio. asumimos la matización de Coll (1979) que engloba bajo este término a la conducta externa observable y a la conducta interna que es inferida por el observador. - Un elemento importante que recoge también la definición es el referente al lapso de tiempo en que se desarrolla este cambio: el ciclo vital. La edad, concreción del aspecto temporal de nuestra definición, no tiene ninguna entidad como variable explicativa, predictiva u optimizadora en sí misma (Baltes, 1983), sino que adquiere importancia en la medida en que es indicadora de un grupo muy amplio de variables y puede ser utilizada como parámetro organizador de los cambios desarrollados a través del cielo vital. Lo importante no es el curso temporal, indica Bijou y Batir (1967); sino los modos concretos de llenar ese curso temporal. Es necesario añadir una matización más al concepto de cambio, objeto de estudio de la Psicología del desarrollo: Este cambio se da en un contexto social, ambiental, ecológico, cultural determinado. Si tenemos en cuanta que este contexto cambia con el tiempo, debemos entender que la psicología del desarrollo se ocupa de algo más que del estudio del cambio en un momento histórico determinado; en ella se contempla la perspectiva de posibles patrones de cambio evolutivo en relación con los diferentes tipos de interacción que puedan establecerse entre las personas y la diversidad de contextos en que estas puedan vivir. LA PSICOLOGÍA DEL DESARROLLO Y LAS TEORÍAS CIENTÍFICAS Psicología genética Corresponde a G. Stanley Hall (1844 -1924) el título de "padre de la psicología del desarrollo". Hall fundó la primera revista científica sobre psicología del desarrollo, el Pedagogical Seminary (1893), la cual se llamó después The Journal of Genetic Psychology (1927). Fue él quien introdujo en la psicología norteamericana el método de los cuestionarios, y junto con sus estudiantes elaboró más de un centenar de cuestionarios sobre una gama muy amplia de tópicos acerca de la niñez y de la vejez. Entre los alumnos prominentes de Hall están H. H. Goddard (trabajo con retrasados), L. M. Terman (pruebas de inteligencia), y Arnold Gesell (desarrollo motor y mental). En su obra más importante, Adolescence: Its Psychology, and Its Relations to Physiology, Anthropology, Sociology, Sex, Crime, Religion, and Education (1904), Hall propuso la teoría de que "la ontogenia recapitula la filogenia". En otras palabras, Hall sostuvo que el desarrollo del individuo (ontogénesis) repite las diversas fases que se verifican en la evolución de la especie (filogénesis). El creía que el período prenatal reflejaba la fase acuática de la evolución humana; y que la preescolaridad (0-4 años) recapitula la fase animal. Comparaba el reflejo involuntario de la prensión del niño recién nacido con el mismo reflejo del primate. La niñez (4-8 años) refleja las culturas primitivas, de caza y pesca. La juventud (8-12 años) repite los períodos salvajes o primitivos del desarrollo cultural, y Hall creía que durante esa edad los niños son especialmente sensibles a las repeticiones, a la disciplina y al entrenamiento. La adolescencia (12-25 años) repite el idealismo del siglo XVIII, la rebelión contra los mayores, la pasión y emotividad, y la entrega a las metas. En la adolescencia tardía, el individuo repite los inicios de la civilización actual. La teoría de la recapitulación de Hall fue más allá de las implicaciones de la teoría de la evolución, en su punto de vista de que la adaptación es como un proceso más o menos completamente gobernado por factores internos del organismo en lugar de un proceso de interacción entre el organismo y el ambiente. En todo caso, su teoría produjo entre los primeros psicólogos del desarrollo y entre algunos contemporáneos una orientación madurativa muy marcada. Los consejos que Hall daba a los padres también reflejaban con toda claridad su punto de vista madurativo. Aconsejaba a los padres que toleraran la conducta socialmente inaceptable de sus hijos, porque, después de todo, estos niños no hacían otra cosa que repetir las diversas etapas del desarrollo evolutivo de la especie. Teoría psicoanalítica Sigmund Freud (1856--1939) fue el primero que propuso una teoría sistemática de la personalidad. El impacto que la teoría de Freud ha tenido en la cultura occidental es tan enorme que apenas hay alguien para quien el nombre de Freud no sea familiar. Freud era médico neurólogo de profesión, y fue precisamente el trabajo que llevó a cabo en neurología lo que lo acerco cada vez más a la formulación del psicoanálisis como método terapéutico. La teoría psicoanalítica sobre el desarrollo de la personalidad, a su vez, nació directamente de la terapia psicoanalítica. Fuente: Reproducido de J. P. Scott, Early experience and the organization of behavior. © 1968 por Wadsworth Publishing Company, Inc. Reproducido con el permiso del autor y de Brooks/Cole Publishing Co. (Monterey, Calif.: 1968). Fig. 1-1. Sonrisas y llanto en los lactantes humanos en relación con los períodos de desarrollo. El punto máximo de respuestas de sonrisa ocurre aproximadamente a los cuatro meses y es característico del período de socialización inicial, durante el cual se forman los primeros apegos sociales. psicólogos del desarrollo y entre algunos contemporáneos una orientación madurativa muy Enarcada. Los consejos que Hall daba a los padres también reflejaban con toda claridad su punto de vista madurativo. Aconsejaba a los padres que toleraran la conducta socialmente inaceptable de sus hijos, porque, después de todo, estos niños no hacían otra cosa que repetir las diversas etapas del desarrollo evolutivo de la especie. Teoría psicoanalítica Sigmund Freud (1856-1939) fue el primero que propuso una teoría sistemática de la personalidad. El impacto que la teoría de Freud ha tenido en la cultura occidental es tan enorme que apenas hay alguien para quien el nombre de Freud no sea familiar. Freud era médico neurólogo de profesión, y fue precisamente el trabajo que llevó a cabo en neurología lo que lo acerco cada vez más a la formulación del psicoanálisis como método terapéutico. La teoría psicoanalítica sobre el desarrollo de la personalidad, a su vez, nació directamente de la terapia psicoanalítica. La teoría de Freud provocó mucha controversia con respecto a tenias como el papel de los instintos en la conducta humana, la estructura de la mente, la función de los procesos conscientes e inconscientes, las diversas etapas sucesivas de la personalidad, y las diversas relaciones intrincadas entre padres e hijos, todo lo cual, según Freud, determina la personalidad del individuo. La teoría de Freud sobre el desarrollo de la personalidad es una teoría de secuencia de etapas. Dentro de cada etapa del individuo debe resolver un conflicto de desarrollo. Si el conflicto no es resuelto, el individuo puede quedar fijado a una etapa determinada. Freud estaba persuadido de que ciertos tipos de personalidad adulta reflejaban dichas fijaciones. Etapas psicosexuales: La teoría psicoanalítica divide la etapa oral (aproximadamente de 0 a 2 años) en dos períodos: oral-pasivo y oral-sádico. Durante la primera infancia, la energía de la libido (una fuerza motivadora innata) se dirige a la boca y a la parte alta del sistema digestivo del lactante. La madre se convierte en el objeto primario de gratificación en cuanto que le proporciona al niño el alimento con el pecho o con la botella. Cuando sobreviene el destete y el niño empieza a usar la taza, sobreviene también un conflicto, en cuanto que el niño se ve obligado a renunciar a la fuente primaria de gratificación. La fijación durante la etapa oral puede producir un carácter oral-pasivo, es decir una persona dependiente y exigente, o un carácter oral-sádico, una persona sarcástica, independiente, agresiva y hostil. La etapa anal corresponde en líneas generales a los años en que el niño empieza a caminar. Durante esta etapa, la energía de la libido se dirige a la región anal, cuando el pequeño empieza a lograr el autocontrol del intestino y la vejiga. Las prácticas de entrenamiento para el retrete proporcionan la base para un conflicto. Conforme a la teoría psicoanalítica, la fijación en la etapa anal produce un carácter analretentivo o un carácter anal-agresivo. Las personalidades anal-retentivas se preocupan exageradamente del orden, son suspicaces y rígidas, y contraen muchos y diversos patrones obsesivocompulsivos de conducta. Las personalidades anal-agresivas son lo opuesto; desaliño, la indiferencia a la rutina, y el descuido personal, constituyen sus señas características. La etapa fálica cubre aproximadamente de los tres a los cinco años. Durante estos años, la fuente de energía de la libido vira (le la región anal a la región genital. El conflicto de desarrollo se presenta ahora al pretender identificarse con el adulto apropiado del mismo sexo. A este conflicto con frecuencia se le llama complejo de Edipo para los niños y complejo de Electra para las niñas. Al principio, tanto los niños corno las niñas se apegan a la madre. Sin embargo, este apego coloca al joven varón en una posición de rivalidad directa con su padre, que es más poderoso que él, por el cariño de la madre. El pequeño varón contrae una ansiedad de castración, o el miedo de que su padre le vaya a causar algún daño a su pene si continúa la competencia activa por el cariño de la madre. El conflicto se resuelve cuando el pequeño varón reprime (es decir, cuando retrae hacia el inconsciente) sus sentimientos sexuales hacia su madre y sus sentimientos de rivalidad hacia su padre. Simultáneamente, se identifica con su padre, interiorizando la conducta, las actitudes y los valores de su padre. Como la joven obviamente no tiene pene, no puede tener miedo a la castración, pero el hecho mismo de no tener pene la lleva a desear tenerlo. Como tampoco su madre tiene pene, la joven mujer se encuentra confusa entre su deseo del amor de su madre y el deseo de tener el pene de su padre. En ambos casos, ya sean niños o niñas, la solución del complejo produce dos resultados claves. Primero, echa a andar y prácticamente completa el proceso de identificación con el progenitor del mismo sexo. Segundo, da origen a la aparición del superego, que consiste en la conciencia y el ego ideal. Una de las funciones principales del superego, como estructura de la personalidad, es restringir la libre expresión de los impulsos del id. El id, que está presente desde el momento de nacer, es la fuente de la energía motivativa. Opera conforme al principio de placer, buscando la gratificación inmediata de las necesidades. El tercer componente estructural de la personalidad, el ego, actúa como mediador entre el id y el superego. El ego opera de acuerdo con el principio de realidad. A continuación del período fálico, sobreviene un período de latencia, el cual, después de la pubertad, es reemplazado por la etapa genital de la madurez sexual. ¿Qué es lo que la teoría freudiana aportó a la psicología del desarrollo? Fue la primera teoría sistemática sobre el desarrollo de la personalidad y la primera teoría de altura que atribuyó una importancia muy especial a los primeros años del desarrollo humano. Además, como lo veremos, dio origen a un esfuerzo muy intenso para reconciliar dos tradiciones científicas opuestas, el conductismo y la teoría psicoanalítica. Tal vez el punto más débil de la teoría freudiana está en que no tiene en cuenta para nada el principio de refutación -dicho de otra manera, la teoría psicoanalítica no toma en cuenta los datos empíricos que pudieran contradecir los postulados básicos de la teoría o las predicciones que se suelen derivar de la teoría. Además, la teoría freudiana no dio la debida importancia a las funciones del ego ni tomó en cuenta debidamente las influencias sociales y culturales que se ejercen sobre el desarrollo de la personalidad. Los teorizantes psicoanalíticos contemporáneos, especialmente Erik Erikson, están procurando mezclar las etapas psicosexuales de desarrollo con los contextos sociales y culturales. Erik Erikson: Mientras que Freud hizo hincapié en los determinantes biológicos de la personalidad y en las funciones del id y del superego, Erikson destaca el desarrollo de la personalidad a la luz de los antecedentes históricos y culturales del individuo. Además, Erikson atiende principalmente al desarrollo de las funciones del ego. Para Erikson, la personalidad nunca llega a estar fija ni a ser final, sino que está constantemente sujeta al cambio. Erikson usa el término freudiano de energía de la libido para indicar la fuerza reguladora que impulsa al desarrollo epigenético. Erikson sostiene que la organización de la personalidad se verifica a través de una serie de etapas, sus "ocho etapas del hombre". Cada etapa plantea a un conflicto que tiene que resolverse antes de que se pueda avanzar a la siguiente. Cada etapa es un punto clave o un punto de elección, en la organización de la personalidad. Un elemento central en la lactancia y en la pre escolaridad es la necesidad del lactante para desarrollar el sentido de confianza en su propio ambiente. Este sentido de confianza prepara al individuo para entrar en una conducta autónoma y de propia iniciativa. En el capítulo 10 volveremos sobre la teoría de Erikson. Por ahora sin embargo, conviene advertir que hay señales de que la teoría "psicosocial" de Erikson sobre la personalidad se puede comprobar empíricamente con mayor facilidad que la teoría de Freud. Funcionalismo Wundt definió la psicología como el estudio de la experiencia consciente, y desarrolló el método de la introspección para investigar los elementos de la mente. Estaba persuadido de que el conocimiento detallado de la experiencia inmediata proporcionaría los datos apropiados para el análisis de los elementos mentales. Partiendo de estos elementos, se podían descubrir las reglas o leyes que integran y regulan a los mismos elementos. A esta corriente psicológica se le conoce como estructuralismo. Como se creía que cada elemento introspeccionado de cualquier experiencia concreta era una sensación (de rojo, de redondez, de suavidad, de vacío, etc.), el estructuralista se encontró ante la ingente tarea de tener que identificar un número casi infinito de elementos mentales. El funcionalismo fue una reacción directa en contra del enfoque estructuralista. En E.U.A., el funcionalismo se propagó gracias, principalmente a los trabajos de William James (1842-1910) y de John Dewey (1859 a 1952). El tema central del funcionalismo es que los procesos mentales y el comportamiento son mecanismos que usa el organismo para adaptarse a su ambiente. Algunos funcionalistas defienden que los procesos mentales y el comportamiento son aspectos inseparables de la adaptación, considerando como activos tanto al organismo como al ambiente. Para estos funcionalistas, la adaptación es un proceso activo, un organismo activo que interactúa con un ambiente activo. Otros funcionalistas conciben la adaptación de manera diferente. Con John Watson a la cabeza, surgió un funcionalismo radical que rechaza el estudio del consciente y define la psicología exclusivamente como la ciencia de la conducta. Behaviorismo (Conductismo): El behaviorismo de Watson tomó como su modelo básico el asociacionismo de estímulo-respuesta e hizo hincapié en los determinantes ambientales del comportamiento. Watson aceptó el postulado empiricista de Locke de que la mente, al nacer, es como una "tabula rasa" (una pizarra en blanco), totalmente dependiente del ambiente para imprimir en ella algún contenido. Según esta postura, el organismo es un objeto pasivo que el ambiente modela más o menos por completo. Watson fue más allá de los datos que poseía, especialmente al rechazar la doctrina funciónalista "común" de que el organismo es un agente activo en el proceso de adaptación. Todavía en la actualidad se advierten concepciones activas y concepciones pasivas de la organización de la conducta. Los que se fijan más en la naturaleza activa del organismo se dice que favorecen la interpretación organísmica del desarrollo de la conducta, mientras que los que se fijan más en la naturaleza pasiva del organismo se dice que favorecen la interpretación mecanicista. Es muy posible que ninguna de las posturas extremas sea totalmente correcta. En lugar de concebir lo activo y lo pasivo como posturas irreconciliables, tal vez será mejor aplicarlas como modelos explicativos cuando lo exijan los fenómenos concretos de desarrollo que se están investigando. ¿En qué forma influyó Watson en la psicología del desarrollo? Su libro sobre el cuidado de los recién nacidos y de los niños fue el primer intento que se hizo aplicar los resultados de la investigación experimental a la práctica de la crianza infantil (Watson, 1928). Al hacer esto, impulsó la persuasión que empezaba a aparecer de que la conducta de los niños y de sus padres se podía estudiar por métodos experimentales. El tema central del conductismo de Watson; que la conducta se podía modelar mediante el control eficaz y directo del ambiente, apuntaló el punto de vista de que la crianza infantil era un proceso unidireccional, de padres a hijos. Esta teoría se convirtió en el postulado clave de la teoría del aprendizaje social, que surgió como un intento para reconciliar la teoría psicoanalítica con la teoría del aprendizaje general. APARTADO 1-2 Puntos de vista opuestos sobre la naturaleza humana o modelos opuestos de desarrollo Postura mecanicista Postura organísmica Organismo positivo Organismo activo Metáfora básica: Organismo vivo Holístico: Cada una de las partes deriva su significado del todo Teleológica (tiene alguna finalidad) Cambio de conducta: Las estructuras y las funciones cambian durante el desarrollo (epigenético) y Reciprocidad entre estructura y función Metáfora básica: Máquina Elementarísmo: El todo se puede predecir por sus partes Mecanicista Cambio de conducta: Lo determina una causa eficiente material Antecedente-consecuente: efecto Causa Discontinua: Los cambios en las partes o en la Continua: El comportamiento actual se organización de las partes dan por resultado puede predecir por el comportamiento un todo con nuevas propiedades sistemáticas; las nuevas propiedades son nuevas en el anterior en sentido aditivo sentido de que no se pueden predecir por la Causalidad: Material, eficiente suma de las partes Unidireccional Causalidad: Material, eficiente, final, formal Recíproca, interactiva Fuente: Adaptado de Reese y Overton (1970); Overton y Reese (1973), con el debido permiso de los autores y del Academic Press, Nueva York y Londres. Teoría del aprendizaje social: La teoría del aprendizaje social ha tenido muchos exponentes, incluyendo a Bandura y Walters (1963), Bijou y Baer (1961), y Whiting y Child (1953). Sin embargo, Robert Sears (por ejemplo, Sears y Dollard, 1941; Sears, Maccoby y Levin, 1957) merece especial mención por haber estimulado el estudio empírico de los conceptos de la teoría psicoanalítica. La teoría del aprendizaje social empieza por la premisa de que todo comportamiento es aprendido. La adquisición de la conducta, según la concibe la teoría del aprendizaje social, se puede comparar a una cadena en la que cada nuevo comportamiento está vinculado con un comportamiento anterior. El desarrollo se concibe como un proceso continuo, pero también como un proceso aditivo, el que el todo es igual a la suma de sus partes. Además, la teoría del aprendizaje social sostiene que la primera conducta y personalidad social se aprenden, principalmente mediante las prácticas de crianza infantil. Especulando a la luz de la premisa de que la socialización del niño es un proceso básicamente unidireccional, de los padres hacia los hijos, la teoría del aprendizaje social considera la diada madre-hijo como la unidad fundamental de comportamiento que hay que estudiar. (Solamente hasta hace muy poco el papel del padre en la crianza infantil ha llegado a ser un tema de estudio positivo). La contigüidad es vista como un determinante importante del aprendizaje. Contigüidad se refiere a una relación temporal entre los eventos del estímulo. Cuando dos estímulos diferentes ocurren íntimamente vinculados en el tiempo, la respuesta a uno de los estímulos se va asociando poco a poco con el otro estímulo. Cuando esta asociación llega a ser suficientemente fuerte, ambos estímulos logran producir la misma respuesta. El reforzamiento también es considerado como otro determinante importante del aprendizaje. El reforzamiento se refiere a un cambio en la probabilidad de que ocurra cierta conducta como función de un evento estímulo que sigue a ese comportamiento. (Ver capítulo 6 para una exposición más amplia de estos procesos de aprendizaje). Sin embargo, la teoría del aprendizaje social no ha sido sólo una teoría de aprendizaje. Es también un intento explícito para aplicar la teoría del aprendizaje y los métodos experimentales de investigación al estudio de las conductas sociales y de las características de personalidad que según la teoría psicoanalítica, nacen de la relación padres-hijos. Por lo tanto, los autores del aprendizaje social emprenden la investigación experimental de tópicos tales como el desarrollo de la agresión; los efectos que tiene el conflicto, el castigo, y otros aspectos de la crianza infantil en la conducta social y en la personalidad del niño; y de las prácticas específicas de crianza infantil que se supone que son importantes en el aprendizaje de la identificación del papel del sexo. Los teorizantes del aprendizaje social insistieron en que los conceptos psicoanalíticos fueran definidos en función de la conducta. Por ejemplo, estos autores definen la dependencia en función de muchos comportamientos, incluyendo el contacto físico y la búsqueda de ayuda, aprobación, o atención. Opinan que la dependencia psicológica del niño con respecto a su madre, surge de su dependencia biológica inicial. A la madre se le considera como un elemento instrumental para reducir la tensión en el lactante en cuanto que le proporciona un reforzamiento primario en forma de alimento, comodidad, y protección contra toda amenaza de daño. En el proceso de satisfacer estas necesidades, la madre llega a asociarse -gracias a la contigüidad- con los reforzamientos primarios. Dicho de otra manera, adquiere el valor de un reforzamiento secundario, de suerte que su conducta social o su mera presencia refuerzan y conservan la conducta del niño. Se puede examinar el fortalecimiento o debilitamiento de las conductas dependientes estudiando los métodos que usan los padres para reforzar la conducta de sus niños. Pero la teoría del aprendizaje social no es una simple extensión del conductismo watsoniano. Más aún, casi desde sus principios, la teoría del aprendizaje social rechazó la postura mecanicista extremista de Watson. Esto tal vez aparece más claro a la luz del método de aprendizaje social que se adoptó para el estudio de la identificación. Se daba por supuesto que la identificación con el modelo apropiado para el papel del sexo no se lograba mediante la enseñanza parental directa. Más bien, buena parte de la identificación se aprende en forma vicaria, observando la conducta de otros. Este aprendizaje exige un organismo activo, capaz de evaluar el comportamiento de otros con relación a si mismo. La teoría del aprendizaje social fomentó un estudio intenso sobre el desarrollo infantil y las prácticas de crianza. Alentó la idea de que se podía enseñar a los padres a criar más eficazmente a los niños y de que se podía lograr comprender el desarrollo del niño mediante rigurosos métodos de estudio. Tal vez su error más grave fue el de haber concebido la socialización del niño como un proceso unidireccional o de un solo sentido, descuidando así la capacidad del niño para provocar y mantener un comportamiento social en los adultos. Como veremos en los capítulos 9 y 10, muchos teorizantes del desarrollo conciben actualmente la socialización como un proceso recíproco en el que los padres y los hijos se influyen mutuamente, aunque no necesariamente en el mismo grado. Cuadro 1-1. Secuencias de las etapas de Piaget, que describe los orígenes de la causalidad durante el período sensorimotor de la inteligencia Primera etapa: Causalidad global. A partir de las primeras semanas de vida hasta cerca de los cuatro meses, el lactante interpreta el mundo en una forma egocéntrica. No distingue entre el yo y el no yo, y no reconoce que los objetos que no están al alcance de su vista tengan existencia permanente. Segunda etapa: Sentimientos de eficacia. Desde aproximadamente los cinco meses hasta los siete, el lactante desarrolla hábitos de reflejos simples en los que están asociados las imágenes y los sonidos, como en la alimentación. Pero todavía no deduce un estímulo a base de la presencia de otro. Funciona completamente en virtud de una estimulación inmediatamente presente. Para él no existe ninguna otra. El sentido que el lactante tiene de la causalidad se reduce a sentimientos vagos de eficacia cuando percibe que algún esfuerzo da por resultado algún efecto deseado. Tercera etapa: Causalidad magico-fenomenalista. Aproximadamente a los ocho o nueve meses, el lactante empieza a percibir la causalidad como algo externo y espacial, pero siempre en función de la acción propia. El niño descubre que uno puede intentar producir algún efecto, pero no logra entender que el contacto físico y espacial es necesario entre la causa y el efecto. De esta manera, el lactante opera como por magia sobre sus alrededores para lograr efectos fenoménicos o ingenuamente percibidos que resultan estar vinculados por la contigüidad con el esfuerzo personal. Cuarta etapa: Exteriorización y objetivización elemental de la causalidad. Abarcando aproximadamente el período entre los 9 y 11 meses, el cuarto período presenta al niño distinguiendo ya entre el yo y los objetos que se imponen a su percepción y que son distintos del yo. El niño empuja hacia un lado la mano de otra persona que sostiene un objeto deseado, dando muestras así de que comprende la causalidad externa. El egocentrismo del niño disminuye. Quinta etapa: Objetivización y ubicación espacial verdadera de las causas. El niño que empieza a andar cae en la cuenta de que el (o ella) es solamente uno de los objetos dentro de todo un mundo de objetos, y que las causas operan directamente sobre los objetos para producir efectos. En esta quinta etapa, que va desde los 12 hasta los 15 meses aproximadamente, el niño percibe correctamente la causalidad externa de los efectos pero no sabe como evocarlos a su antojo. Sexta etapa: Causalidad abstracta o representativa. Aproximadamente entre los 18 meses, y los dos años, el niño logra abstraer la realidad. El niño puede reconstruir mentalmente una causa partiendo del efecto presente. El uso del lenguaje ya se ha desarrollado, lo cual permite al niño tomar en cuenta estímulos que no están inmediatamente presentes. Las experiencias pasadas tienen ahora cierto significado. Fuente: Adaptado y reproducido de C. F. Monte, Psychology's scientific endeavor (Nueva York; Praeger, 1975), p. 27. Con el permiso de Christopher F. Monte, © 1975, y de Praeger Publishers, Inc. LA PSICOLOGÍA DEL DESARROLLO Y LAS TEORÍAS CIENTÍFICAS Psicología genética Corresponde a G. Stanley Hall (1844-1924) el título de "padre de la psicología del desarrollo". Hall fundó la primera revista científica sobre psicología del desarrollo, el Pedagogical Seminary (1893), la cual se llamó después The Journal of Genetic Psychology (1927). Fue él quien introdujo en la psicología norteamericana el método de los cuestionarios, y junto con sus estudiantes elaboró más de un centenar de cuestionarios sobre una gama muy amplia de tópicos acerca de la niñez y de la vejez. Entre los alumnos prominentes de Hall están H. H. Goddard (trabajo con retrasados), L. M. Terman (pruebas de inteligencia), y Arnold Gesell (desarrollo motor y mental). En su obra más importante, Adolescence: Its Psychology, and Íts Relations to Physiology, Anthropology, Sociology, Sex, Crime, Religion, and Education (1904), Hall propuso la teoría de que "la ontogenia recapitula la filogenia". En otras palabras, Hall sostuvo que el desarrollo del individuo (ontogénesis) repite las diversas fases que se verifican en la evolución de la especie (filogénesis). El creía que el período prenatal reflejaba la fase acuática de la evolución humana; y que la pre escolaridad (0-4 años) recapitula la fase animal. Comparaba el reflejo involuntario de la prensión del niño recién nacido con el mismo reflejo del primate. La niñez (4-8 años) refleja las culturas primitivas- de caza y pesca. La juventud (8-12 años) repite los períodos salvajes o primitivos del desarrollo cultural, y Hall creía que durante esa edad los niños son especialmente sensibles a las repeticiones, a la disciplina y al entrenamiento. La adolescencia (12-25 años) repite el idealismo del siglo XVIII, la rebelión contra los mayores, la pasión y emotividad, y la entrega a las metas. En la adolescencia tardía, el individuo repite los inicios de la civilización actual. La teoría de la recapitulación de Hall fue más allá de las implicaciones de la teoría de la evolución, en su punto de vista de que la adaptación es como un proceso más o menos completamente gobernado por factores internos del organismo en lugar de un proceso de interacción entre el organismo y el ambiente. En todo caso, su teoría produjo entre los primeros Epistemología genética Jean Piaget (nació en 1896) ha empleado más de 50 años en el estudio del desarrollo de la inteligencia. Piaget con frecuencia llama a su método epistemología genética, que significa el desarrollo de diversos modos de conocer el mundo exterior. Según Piaget, dos son las funciones que caracterizan a todo organismo vivo- la organización y la adaptación. El desarrollo intelectual consiste de cambios progresivos o secuenciales en la estructura de la organización. Las estructuras o sistemas cognoscitivos cambian con la adaptación. NIVELES DE ESTUDIO DEL CAMBIO EVOLUTIVO DEL COMPORTAMIENTO HUMANO La Psicología del desarrollo, en su estudio del cambio de comportamiento, puede abordar éste desde tres enfoques distintos: descriptivo, explicativo e intervencionista. a) En su enfoque descriptivo la Psicología del desarrollo mantiene varios puntos teóricos importantes: - La descripción puede centrarse en el análisis de dos clases de varianza: en el cambio que tiene lugar en el individuo a través del ciclo vital (cambio intra individual) como en las diferencias que existen en el cambio de los diferentes individuos cuando las condiciones propias de cada uno van evolucionando con el tiempo (cambio interindividual en el cambio intra individual). - Una descripción de la evolución del comportamiento humano ha de ser pluralista; este pluralismo lleva al relativismo. Cada investigación descriptiva debe considerarse en relación con los aspectos individuales generacionales o ambientales en que se ha realizado. - La enorme variación existente en el cambio ontogenético y generacional puede reducirse mediante la determinación de secuencias en el cambio. Esta determinación de secuencias ha de realizarse teniendo en cuenta la multidimensionalidad del cambio y del patrón específico del cambio de cada sujeto. b) En su enfoque explicativo la Psicología del desarrollo parte también de varios supuestos teóricos importantes: - El cambio evolutivo se opera en función de una serie de variables tanto personales como ambientales. Objeto de la Psicología del desarrollo será, pues, identificar los distintos patrones de influencia en dicho cambio. Esta tarea es de naturaleza compleja si se tiene en cuenta que en una perspectiva dialéctica el cambio de conducta puede ser inducido por los cambios de contexto, pero que a su vez este cambio también puede modificar los propios contextos (Baltes y Schaie, 1973). - Este cambio evolutivo se basa también en la acción conjunta de una serie de leyes, continuas unas, discontinuas otras, que rigen las relaciones entre variables independientes y dependientes y que operan de modo multidimensional. - Los factores explicativos dependientes del tiempo han de entenderse de un modo dinámico y como potencialmente cambiantes. La existencia de cambios multidireccionales y truncados sugiere que los antecedentes conductuales difieren de un período a otro, por lo que parece lógico concluir que la explicación no es ya un proceso fijo -algo que se hace de una vez para todas-, sino que ha de irse haciendo y cambiando progresivamente (Vega, 1981). c) En su enfoque intervencionista, la Psicología del desarrollo contempla el cambio a través del ciclo vital como modificable y optimizable gracias a la intervención, corrección y prevención, según los casos y esto hasta tal punto, que este enfoque se ha convertido en la cuestión central de la Psicología del desarrollo (Vuyk, 1981). Una intervención a lo largo del ciclo vital descansa, según Lerner y Ryff (1878) sobre tres supuestos: - Dada la multiplicidad y el relativismo de los factores que influyen en el cambio evolutivo, la intervención es compleja. Por lo que hay que tener en cuenta qué conductas, qué individuos, en qué contextos y en qué períodos temporales es mejor intervenir para obtener mejores resultados evolutivos. - No existe una única forma de intervención, sino que los métodos y herramientas analíticas que optimicen ésta, están en dependencia de los contextos individual social e histórico. - La intervención no ha de servir sólo para corregir posibles desviaciones de conducta, sino fundamentalmente para prevenir y optimizar. Conocida una regularidad en el cambio, puede optimizarse éste en vez de esperar a que surjan las deficiencias y luego resolverlas. Como conclusión de esta consideración de los tres posibles enfoques de la Psicología del desarrollo, cabe añadir que han supuesto, según el orden en que los hemos expuesto, un avance en la reorganización de esta ciencia. La optimización, última en la secuencia histórica de sus preocupaciones, ha pasado a convertirse en la actualidad en la cuestión central de la Psicología del desarrollo y la que estimula y guía, en definitiva, la investigación descriptiva y explicativa (Baltes y Willis, 1977; Lerner y Ryff, 1978). TEORÍAS SOBRE EL DESARROLLO A través de la Historia de la Psicología del desarrollo ha habido diversos intentos teóricos de explicar este desarrollo. No todos estos intentos han logrado siempre el mismo nivel de validez explicativa. Cada uno de ellos, sin embargo, debe considerarse sumamente positivo en cuanto, frecuentemente, ha impulsado investigaciones que de otro modo no se habrían realizado, contribuyendo considerablemente, en este sentido, al progreso de la ciencia psicológica. Hasta tal punto esto es cierto que sería difícil, pensamos, entender alguna de las más recientes teorías sobre el desarrollo psicológico sin conocer previamente aquéllas que les han precedido y sobre las que de algún modo se han fundado. Estos intentos teóricos pueden agruparse en dos grandes grupos o categorías: uno de ellos orientado fundamentalmente a estudiar el curso del desarrollo. Se trata de teorías que consideran el proceso de desarrollo como una sucesión de períodos o fases. En general son teorías que han encontrado muy buena acogida entre los pedagogos. Su margen de validez ha ido, en general, disminuyendo a través de los años, debido al creciente número de datos que no cabe interpretar a la luz de sus principios. El otro grupo está orientado, fundamentalmente, por el contrario, al estudio de la naturaleza del proceso evolutivo y sus principios básicos. Se trata de teorías que niegan, en general, la existencia de períodos en el desarrollo, o al menos rehuyen su estudio, centrándose más bien en los procesos que lo constituyen. 1.1 Teorías sobre el curso del desarrollo Dos son las posiciones teóricas diferentes que cabe señalar aquí: la de los que consideran el desarrollo como sucesión de fases y la de los que lo consideran como crecimiento. 1.1.1. El desarrollo como sucesión de fases Idea común a las teorías que entienden el desarrollo como sucesión de fases es el supuesto de que este desarrollo no es continuo ni gradual, sino que introduce en su curso períodos bien definidos. Entre las teorías más conocidas de este grupo está la psicoanalítica de Freud, la madurativa de Gesell y la genético cognitiva de Piaget. a) Teoría psicoanalítica. Con objeto de precisar la naturaleza de las operaciones, procesos y cualidades psíquicas, así como sus relaciones, Freud (1951) propone una estructura de la personalidad, la cual, en su forma acabada (forma adulta) está constituida por tres instancias: el Ello, el Yo y el Super-Yo. Cada instancia es a su vez, estructura, función y lugar de operaciones y procesos psicológicos específicos de orientación muy determinada. Genéticamente, el Yo es una diferenciación del Ello, y el Super-Yo una diferenciación del Yo. Se forman sucesivamente en las diferentes etapas de la evolución del niño y determinan los diferentes estadios de su desarrollo. El desarrollo de la personalidad es un proceso de tensión entre las tres instancias. El Ello es la más primitiva de estas tres instancias psíquicas y está constituido por un sistema de necesidades dotadas de una particular potencia y energía. Entre estas necesidades, Freud sitúa en primer plano los instintos. Los instintos son numerosos, pero Freud los reduce a dos: el instinto de vida o eros (que comprende los instintos de conservación de sí y de la especie, el amor de sí y el amor objetal) y el instinto de muerte o destrucción, que representa, según Freud, esa tendencia de todo estado evolutivo a volver al estado anterior que le ha precedido en la vida. La energía respectiva de estos dos instintos fundamentales es la libido y la fuerza destructiva o agresividad. En tanto que instancia psíquica, el Ello es inconsciente. No conoce leyes ni tiene en cuenta la realidad, sino la satisfacción de sus propios deseos mediante la descarga de la tensión que éstos originan en la persona. La formación del Yo se realiza a partir del Ello. La dirección de su actividad está orientada a la búsqueda del placer, como el Ello, del cual se origina; pero teniendo en cuenta al mismo tiempo sus deseos de auto conservación en el seno de una realidad que le presenta también, a su vez, una serie de exigencias, necesidades y amenazas; lo que lleva al Yo a frenar sus impulsos, a modificarlos, a canalizarlos y a suprimir algunos que se presentan como peligrosos para esta autoconservación. Tareas, pues, del Yo, son la defensa y su adaptación al medio. La estructura del Yo responde a esta doble tarea. Está formada de conocimiento y de defensa. Su actividad cognoscitiva la constituyen las percepciones externas e internas y todos los procesos intelectuales que permiten al Yo conocer las modificaciones que se originan tanto en el mundo interior como en el exterior. Su actividad defensiva consiste en utilizar todos aquellos mecanismos de defensa formados a través de los diferentes estadios de la constitución del Yo. El Super-Yo es la tercera instancia de la estructura psíquica y la ultima en aparecer en la persona. Es una diferenciación del Yo del cual se llega a escindir constituyéndose en instancia independiente. La modificación esencial del Yo que prepara el nacimiento del Super-Yo se produce en el estadio del complejo de Edipo. La resolución de este complejo consiste en la renuncia al progenitor del sexo contrario como objeto del amor libidinal y a la interiorización por la que el niño se identifica con el progenitor del mismo sexo. El Super-Yo, según esto, es el heredero del complejo de Edipo y resulta de las identificaciones que liquidan dicho complejo. Una vez constituido el Super-Yo se comporta como representante de las fuerzas opresoras y represivas parentales y asume la función de vigilar sobre el Yo, darle órdenes, dirigirle, amenazarle con castigos, etc. Los fenómenos de auto observación, autocrítica, los sentimientos de remordimiento y culpabilidad, el masoquismo moral, están relacionados con esta instancia de la persona. La actividad del Super-Yo puede ser pre consciente o inconsciente; sobre todo, inconsciente. Entre las tres instancias de la estructura psíquica las relaciones son de naturaleza conflictual tanto en su génesis como en su funcionamiento adulto. Entre el Ello y el Yo, quien a la vez que intenta satisfacer los deseos del Ello, debe frenarlos o reprimirlos. Entre el Yo y el Super-Yo quien se opone al primero y le domina, a la par que le ofrece seguridad. Los estadios psicoanalíticos están relacionados con la génesis de esta estructura psíquica y el desplazamiento de la libido a través de una serie de órganos específicos para obtener placer. Primero se da el estadio oral, posteriormente el anal y finalmente el genital. La crítica más grave que se ha dirigido contra la teoría de Freud sobre el desarrollo es la sobreestima que éste hace de la naturaleza biológica e instintiva del hombre. Aunque haya sido así, Freud, con todo, no ha dejado de tomar en cuenta la influencia de los factores sociales. Su teoría considera secundarios los factores ambientales frente a las tendencias congénitas; pero en modo alguno niega su importancia. Los conceptos morales, las aspiraciones y ambiciones de cada sociedad entran a formar parte del individuo a través del desarrollo del Super-Yo. Posteriormente, el psicoanálisis modificando su posición ortodoxa que otorgaba tanta importancia a los instintos y al determinismo biológico en el proceso evolutivo, ha dirigido su atención hacia otras funciones psíquicas a través de los estudios de la cultura (Wilhelm Reich, 1933; Erikson, 1950) y sociológicos (Karen Horney, 1937; Fromm, 1941; Sullivan, 1948). b) Teoría madurativa de Gesell. Para Gesell (1880-1961) la maduración ocupa un lugar preponderante en la concepción del desarrollo. La maduración está ligada en primer lugar al desarrollo nervioso y en segundo lugar a la noción de constitución. El niño posee tendencias y rasgos constitucionales, la mayor parte innatos, que determinan hasta un cierto punto qué y cómo se puede aprender. Estos rasgos y tendencias son a la vez raciales y familiares. El niño toma posesión de esta doble herencia por el proceso de maduración. Esta importancia por la maduración no excluye en absoluto la influencia del medio y en particular el medio social. Los factores internos y los factores externos son inseparables. Maduración y medio social se compenetran e interrelacionan desde el nacimiento. El crecimiento humano -dice Gesell- es sensible a las influencias culturales a partir del nacimiento. La personalidad comienza a tomar forma bajo la influencia de otras personalidades. No existe, pues, en la concepción de Gesell, el determinismo organicista que a veces se le ha reprochado. Si Gesell se inclinó a subrayar con énfasis el papel de la maduración en el desarrollo, fue como reacción contra los excesos de los conductistas. Pero no por desconocimiento o subestima de la influencia del medio social como tal. La acción de ambos factores sobre el desarrollo se traduce en leyes que determinan las orientaciones, sucesión y encadenamiento de estadios. Gesell ha puesto en evidencia cómo, pese a las contingencias debidas a los diversos medios, existe un orden riguroso y constante en la sucesión de los estadios de desarrollo. La morfogénesis del comportamiento humano -dice- comprende coordenadas que normalmente jamás son perturbadas. c) Teoría genético-cognitiva de Piaget. Piaget concibe el desarrollo como un proceso de adaptación con el medio en el cual distingue dos aspectos: un componente afectivo o energético y un componente cognoscitivo o estructural. Este último es el que acapara la atención y los estudios de Piaget. ¿Cómo se desarrolla para el maestro ginebrino el proceso cognoscitivo? Varias son las ideas que explican, según él, este desarrollo: - La idea básica subyacente es que las funciones permanecen a lo largo del desarrollo infantil, mientras que las estructuras cambian sistemáticamente. - El término de "estructura" se refiere a un conjunto de elementos reunidos en una totalidad con unas propiedades propias en tanto que totalidad. En otras palabras, es un conjunto de acciones ú operaciones interrelacionadas que forman un comportamiento de conjunto. Por ejemplo, un niño ve un sonajero y lo coge. La estructura de esta conducta incluye los medios (mirar, alcanzar, asir) y el fin (estimulación del objeto en la mano). Cada uno de estos factores está relacionado con el otro. A esta relación es a lo que Piaget llama estructura. A cada unidad estructural la denomina "esquema". - El término "función" se refiere al modo de interactuar el sujeto con su medio. Las funciones básicas del desarrollo son dos: la adaptación y la organización. Ambas funciones son heredadas biológicamente y no cambian a través del desarrollo, por lo que Piaget las denomina "invariables funcionales". - La adaptación como proceso de desarrollo cognitivo consiste, como el proceso de adaptación biológica, en un proceso de equilibrio entre dos funciones: la asimilación y la acomodación o ajuste. Se produce una asimilación siempre que un organismo utiliza algo de su ambiente y se lo incorpora. La ingestión de alimentos puede ser un buen ejemplo biológico. Se produce asimilación psicológica siempre que el sujeto incorpora una nueva experiencia cognoscitiva a las estructuras cognoscitivas ya existentes. Pero así como el alimento queda modificado en la asimilación biológica, la estimulación psicológica también queda modificada al ser experimentada por el sujeto. Por ejemplo, el conocimiento que se obtiene de los objetos en la percepción, no es algo similar a una copia fotográfica sino que se halla impregnado de significado; es decir, los datos obtenidos de dicho objeto se hallan organizados en categorías en función de dimensiones tales como el tiempo, el espacio, la familiaridad, la belleza, etc. Se produce una acomodación cuando una estructura cognoscitiva cambia bajo el efecto de las experiencias asimiladas. Por ejemplo, al niño que se le da a chupar un biberón (asimilación) llega al fin a darse cuenta de que ciertos movimientos de la lengua son más aptos para la succión de la leche que otros (acomodación). Este nuevo esquema de comportamiento tiene, pues, que ser aprendido. Si no existiera asimilación, no se producirían nunca variaciones ni, por tanto, desarrollo. - La experiencia que adquiere el sujeto es otro de los factores invocados por Piaget para explicar el desarrollo cognitivo. La experiencia puede ser diversa: está la del simple ejercicio que supone la presencia de objetos frente a los cuales se ejerce la acción, pero sin que esa experiencia o comportamiento sea extraído necesariamente de dichos objetos. Como, por ejemplo, la succión que se vuelve más hábil al repetirse desde los primeros días. Está la experiencia física por la que el sujeto llega a descubrir ciertas propiedades de los objetos, por ejemplo, su peso, disociándolas de las otras ya conocidas. Y está la experiencia lógico-matemática, que consiste también en actuar sobre los objetos. Pero el conocimiento a que conduce no está extraído de los objetos sino de las acciones que se ejercen sobre los objetos. Es decir, de las propiedades que la acción del sujeto ha introducido en ellos y que antes no les pertenecían o eran independientes de dichos objetos. Por ejemplo, el orden, la clasificación, la suma o cualquier otra operación lógico-matemática que al ordenar, sumar o clasificar ha introducido en los objetos. - Para Piaget, los tres factores clásicos que son la maduración, el medio físico y el medio social, son necesarios pero insuficientes para explicar la génesis de las estructuras mentales. Hace falta añadir un cuarto factor que es el de la equilibración. Si tomamos como ejemplo la adquisición de la noción de substancia permanente en los objetos a través de sus diversos cambios, tenemos que la maduración es necesaria, pero no suficiente ya que no todos los niños la adquieren a la misma edad. La manipulación de los objetos puede contribuir a su descubrimiento. Pero puesto que la substancia es una forma vacía, inaccesible a los sentidos, tampoco esta experiencia es suficiente. Para explicar esta adquisición hay que recurrir a la equilibración, entendiendo por tal "la sucesión probabilística de fases de estrategias": la primera es más probable al principio; la segunda llega a ser más probable en función de los resultados de la primera, pero no al principio; la tercera llega a ser más probable en función de la segunda, pero no antes, etc. Un ejemplo hará comprender esto: cuando se transforma una bolita de arcilla en una salchicha, ¿cómo llega el niño a comprender la permanencia de la substancia a través del cambio de forma? Según Piaget, el niño en este proceso seguiría cuatro etapas: en la primera no considera nada más que una dimensión; en la segunda se da cuenta de la otra y oscila entre ambas; en la tercera descubre la relación de compensación entre las dos dimensiones y llega al presentimiento de la solidaridad a través de las transformaciones; en la cuarta el sujeto comprende que la transformación puede ser invertida o que las transformaciones se compensan en virtud de esa solidaridad. Este paso de una etapa a la siguiente es un factor de equilibración. El proceso de desarrollo cognitivo está constituido por una serie de equilibraciones que van desde el conocimiento senso-motor hasta la inteligencia formal. - Piaget concibe entonces el desarrollo como un proceso continuo de organización y reorganización de estructuras, de modo que cada nueva organización integra en sí misma a la anterior. Aunque tal proceso es continuo, sus resultados no lo son; resultan cualitativamente diferentes a lo largo del tiempo. Por tal motivo Piaget ha decidido dividir el curso total de desarrollo en unidades denominadas períodos, sub períodos y estadios. No obstante debe tenerse muy presente que cada una de estas porciones de desarrollo es descrita en función de lo que el niño puede hacer en aquel momento. Objeciones a las teorías del desarrollo como sucesión de fases Se han levantado muchas objeciones en torno a estas teorías de fases o estadios referidas ya a su validez y posibilidad de explicar satisfactoriamente y sin contradicciones el curso del desarrollo, ya a su interés práctico para la pedagogía y la enseñanza. Con respecto a su validez explicativa cabe señalar tres tipos de objeciones: la primera se refiere a las bases generales y determinantes de sus divisiones en fases o estadios; se estima que los procesos de maduración son endógenos y tienen por causa factores internos inherentes al individuo. Esta concepción de fijación de las características del proceso evolutivo a cargo de los factores de maduración resulta, aparte de unilateral, incompatible con gran número de comprobaciones sobre la profunda influencia de los factores ambientales. La segunda objeción se refiere a la dificultad por parte de estas teorías en asimilar adecuadamente numerosos datos empíricos. Como, por ejemplo, el acceso, mediante el aprendizaje en ciertos estadios del desarrollo, a ciertos estados que, según estas teorías de grados y fases, no cabría esperar, sino en estadios posteriores del desarrollo. Estados que, a juzgar por las investigaciones longitudinales, no son transitorios. La tercera objeción se refiere a la incapacidad de estas teorías para presentar datos seguros sobre los límites y duración de los períodos de desarrollo. Lo que ha llevado, en la práctica, a la formulación de una gran serie de grados y fases según el punto de partida adoptado. Un segundo grupo de objeciones a estas teorías se refiere, decíamos, a su pretendida eficacia y utilidad para la pedagogía. Es cierto que estas teorías de grados y fases han ofrecido a los profesores que siguen su enseñanza dividida en cursos, una orientación de trascendental importancia por cuanto han constituido el punto de partida para su programación. Esto no obstante, si relacionamos las conclusiones de estas teorías con los datos objetivos, fácilmente se comprenderá que hoy en día no se puede seguir dándoles vigencia. Porque si bien es cierto que las teorías de grados y fases del desarrollo, al destacar los peligros de la enseñanza prematura, han contribuido a evitar la sobre valoración de las posibilidades de la pedagogía, por otra parte han puesto trabas e incluso impedido el estímulo de determinados procesos evolutivos, con ciertos procedimientos pedagógicos. 1.1.2. El desarrollo como crecimiento Los teóricos pertenecientes a este grupo tratan de representar el desarrollo psíquico de modo análogo a los cambios que se observan en la esfera somática. De ahí que le denominen crecimiento. Así como las sucesivas modificaciones de los rasgos corpóreos pueden configurarse, gracias a las formas que adopta su curso, en función del progreso, del mismo modo es posible interpretar por medio de gráficos, las variaciones psíquicas que tienen lugar en la infancia, adolescencia y edad adulta. Al principio, se trazaron curvas de crecimiento de la edad intelectual (Anastasi, 1965); posteriormente se amplió su uso a otros aspectos del desarrollo evolutivo: funciones sensoriales y psicomotrices, lenguaje, pensamiento, conducta social, etc. Se comparó la forma de estas curvas por decenios y logró demostrarse que, en lo esencial, había una amplia concordancia con las funciones del crecimiento. La gráfica presentaba al principio una rápida subida, se hacía luego, con el correr de los años, más lenta, alcanzaba un punto culminante y finalmente acusaba, al cabo de un cierto espacio de tiempo, un curso regresivo. El concepto teórico fundamental, pues, de este modelo de desarrollo, es que tanto el crecimiento físico como el desarrollo psíquico son procesos correlativos que se producen en el mismo organismo y deben entenderse del mismo modo, es decir, como proceso de crecimiento. El concepto de crecimiento no comprende únicamente el aumento de peso y tamaño, sino también modificaciones estructurales que pueden ser, por ejemplo, de proporción y complejidad (Olson 1953). A la luz de las más recientes investigaciones psicológicas, hoy apenas cabe sostener la hipótesis de una amplia sincronización de los distintos aspectos físicos y psíquicos del desarrollo. Y en cuanto a este modo de entenderse, el desarrollo es una cuestión que continúa siendo más que discutible. 1.2. Teorías sobre la naturaleza del proceso evolutivo y sus principios básicos Las teorías que vamos a examinar a continuación se refieren no tanto al curso del desarrollo, cuanto a su naturaleza y procesos fundamentales que atribuyen especiales características a la serie de modificaciones. Pueden distinguirse tres posiciones más o menos opuestas: la de los que destacan el papel de los factores genéticos o endógenos, la de los que consideran como más importante la influencia del medio y la posición de los que consideran que el desarrollo es el resultado de la interacción del individuo con su medio ambiente. 1.2.1. El desarrollo como fruto de la acción de factores genéticos o endógenos Se trata de un modelo biológico creado para la comprensión del modo de ser característico de los procesos de desarrollo. Se considera que, lo mismo que en biología los procesos de progresiva diferenciación e integración constituyen un fenómeno fundamental del desarrollo somático de los seres vivos, los procesos básicos del desarrollo pueden entenderse también como una creciente diferenciación y centralización. En lo biológico, esta diferenciación y consiguiente centralización se ha conectado al progreso del sistema nervioso central. Pero pretender referirla al sistema psíquico, es actualmente algo muy discutido; de hecho es insostenible. Uno de los teóricos más representativos de este grupo es Werner. Según el punto de vista de este teórico, la diferenciación en el sistema psíquico supone, ante todo, un afinamiento de las funciones y manifestaciones psíquicas. Se da en éstas una especialización creciente y clara, a partir de la unidad primitiva indiferenciada. El proceso de centralización, contrapuesto al de diferenciación, conduce a la estructuración jerárquica de la psique y se refleja en la aparición de nuevas funciones de mayor fuerza centralizadora: los sentimientos pasan a depender de los procesos intelectuales, las percepciones se sujetan a los conceptos y a la actividad del pensamiento, etc. Werner (1959), como otros defensores de este peculiar enfoque, ha ido a la caza de hechos que apoyen sus concepciones en el estudio del desarrollo de diversos tipos de conducta, sobre todo de la percepción y el pensamiento en los niños. Sus supuestos teóricos, con todo, no se han visto confirmados por las recientes investigaciones psicológicas. 1.2.2. El desarrollo como fruto de la acción de factores externos Cabe distinguir, entre las teorías que han enfatizado esta acción externa, dos enfoques más representativos: el de la teoría del aprendizaje y el del modelo del cuño. a) La teoría del aprendizaje. Postula que los cambios más importantes que se producen a lo largo del desarrollo humano son fruto del aprendizaje mediante el ejercicio. Punto importante es que la conducta del sujeto en desarrollo puede considerarse, entonces, como un conjunto de respuestas en interrelación con una serie de estímulos. Existen tres tipos de aprendizaje: el clásico, descrito por Pavlov (1968), el instrumental, descrito por Thorndike (1898), el operante, descrito por Skinner (1953) y el observacional, descrito por Bandura (1977). Según esta teoría, el cambio evolutivo puede ser debido, entonces, a la asociación de la nueva conducta (condicionada) con otra ya establecida en el sujeto (incondicionada); al ejercicio y refuerzo discriminativo de la nueva conducta; o a la imitación de un modelo, seguido de refuerzo. Según Bijou y Baer (1967), dos de los teóricos más representativos de este modelo, las leyes del aprendizaje permiten clasificar cualquier respuesta del niño, del joven y del adulto y estudiar su conducta basándose únicamente en criterios observables y objetivos. Es innegable la importancia que tiene el aprendizaje en el desarrollo de la persona desde la edad más temprana. La adquisición de muchas de las conductas humanas puede considerarse, en efecto, dentro de amplios límites, fruto del aprendizaje: habilidades psicomotrices, emociones, actitudes, deseos y valores e incluso muchas de las habilidades intelectuales. Esto no obstante, pensamos que no se debe infravalorar, como hacen los psicólogos del aprendizaje, el papel que desempeña en el desarrollo la herencia y la maduración biológica que limitan la cantidad y calidad de las respuestas que la persona puede aprender. Ningún ser humano puede aprender a nadar exactamente como un pez, ni a correr como una gacela. De manera semejante, el niño de un año no podrá aprender ciertas habilidades propias de los seis, el de tres entender el cálculo matemático y el mongólico comprender la teoría atómica. Los principios de la teoría del aprendizaje también encuentran límites a la hora de explicar la adquisición de determinadas conductas superiores más complejas, como son, por ejemplo, la cognitiva y la verbal. b) La teoría del cuño. Las modernas investigaciones han reservado el modelo del cuño para explicar la aparición de ciertas conductas entre los animales por la acción del medio. En la acuñación el papel agente se reserva, pues, para el ambiente exterior; mientras que al animal se le reserva un papel más bien pasivo. Por ejemplo, el hecho de que en determinadas especies de aves pueda provocarse, durante un espacio relativamente corto, después de su salida del huevo, determinadas conductas instintivas que luego perdurarán toda la vida. En condiciones normales, los estímulos que intervienen proceden, por lo general, de otros animales de la misma especie o de cosas de importancia vital. Al espacio de tiempo durante el cual se consigue esta acuñación de la conducta lo denomina Lorenz (1935) "período sensible", que en la mayoría de los casos oscila alrededor de las 24 horas siguientes al nacimiento. La aplicación de este modelo, por algunos psicólogos, para explicar el desarrollo humano, no es posible sino con muchas cautelas. Puede aplicarse para explicar la adquisición de determinados tipos de comportamiento que persisten en la infancia por la acción del medio, como ha tratado de hacerlo la psicología profunda con respecto a determinadas fijaciones, o la psicología social, con respecto a la acuñación de ciertas normas sociales, tradiciones y esquemas de conducta. En todos estos casos, con todo, a diferencia de la acuñación en los animales, se trata, por lo general, del moldeo de tipos de conducta que no obedecen a la acción más o menos penetrante del medio ambiente, por una única vivencia o una serie de vivencias de corta duración, sino por experiencias de acción bastante prolongadas y relativamente constantes (Nickel, 1976). La acuñación, como proceso que tiene lugar sin intervención deliberada del propio sujeto, quien es más bien un elemento pasivo, una placa que queda impresionada, es un modelo no válido para explicar el desarrollo humano. Por lo menos para las conductas que se adquieren después de los primeros años de vida en cuya adquisición el individuo participa, por lo general, de modo activo. 1.2.3. El desarrollo como interacción entre la persona y el medio Característica común de este grupo de teorías es el entender que el desarrollo no debe considerarse como fruto de la maduración, ni tampoco como una consecuencia de la estimulación del medio, sino como una interacción entre el individuo y su medio cultural. Vamos a fijarnos de modo particular en dos teorías diferentes de este grupo: la teoría de Vygotsky y la teoría del Ciclo Vital. a) Teoría de Vygotsky. Fue hacia los años treinta cuando Vygotsky expuso una teoría que denominó "histórico-cultural" y en la que pretendía explicar el desarrollo humano. Sus tesis fundamentales son estas dos: - En primer lugar, Vygotsky (1979) afirma que las funciones psíquicas superiores del hombre (percepción, atención, memoria, pensamiento), son el resultado de la asimilación individual de los productos de la cultura humana. El desarrollo de estas funciones psíquicas se lleva a cabo en estrecha conexión con la formación de nuevas estructuras funcionales del cerebro. Dichas estructuras han sido denominadas por Luria (1976) sistemas funcionales cerebrales y se forman durante la vida del individuo permitiéndole adaptar su conducta a las exigencias de la sociedad. Vygotsky fue uno de los primeros en plantearse la hipótesis sobre el carácter mediatizado que poseen las funciones psíquicas. Es decir, que lo mismo que existe en el trabajo del hombre una utilización de instrumentos que le mediatizan en sus relaciones con el medio, así en el desarrollo de los procesos psíquicos existe una mediación, debida al sistema específico de señales que son los símbolos y los signos, a los que denomina instrumentos psíquicos. Estos sistemas de señales permiten al hombre establecer una relación con los demás hombres; pero también a través de ellos se lleva a cabo una reestructuración de las funciones psíquicas. - Una segunda hipótesis de Vygotsky se refiere al origen de los procesos intelectuales. Vygotsky y sus continuadores, Leontiev, Zoporozhetz, Luria, El konin, Galperín, Davidof, mantienen que la conciencia se halla mediatizada por la actividad práctica, externa y material del individuo. Leontiev, en un estudio detallado de este problema, llega a la conclusión de que los procesos de conciencia y la actividad externa no son procesos de orden diferente, sino que, por el contrario, son formas de una misma actividad del individuo, en la cual, una de ellas, la interna, está originada por la otra, la externa. Es decir, la actividad material externa se interioriza y adquiere la forma de actividad mental interna. Específicamente y en relación con el desarrollo psíquico del niño, Vygotsky (1979) distingue un área de desarrollo efectivo y otra de desarrollo potencial. Por desarrollo efectivo entiende el conseguido por el niño como resultado de un específico resultado del desarrollo, ya realizado. Un simple control demuestra, añade Vygotsky, que este nivel de desarrollo no indica el verdadero estado de desarrollo del niño: con ayuda de los adultos puede imitar un gran número de acciones que superan su capacidad actual; puede hacer mucho más de lo que es capaz con su actitud de acción independiente. La diferencia entre el nivel de las tareas que es capaz de realizar con una actividad independiente y el nivel de las tareas realizables con ayuda de los adultos, define el área de desarrollo potencial del niño. Lo que el niño puede hacer hoy con ayuda de los adultos, lo podrá hacer mañana por sí solo. La enseñanza es precisamente la responsable de generar esta área de desarrollo potencial; de estimular y activar los procesos internos en el marco de las interacciones. b) Teoría del Ciclo Vital. Esta teoría agrupa a investigadores como Baltes (1983), Schaie (1970) Nesselroade y Reese (1973). Sus principales hipótesis y proposiciones sobre la naturaleza del desarrollo son las siguientes: - El desarrollo es un proceso que tiene lugar a lo largo dé toda la vida. Ningún período de edad es superior a otro para regular la naturaleza del desarrollo. - Este desarrollo es multidimensional. Se ha encontrado, en efecto, que el curso del desarrollo es diverso según las conductas del individuo. O lo que es lo mismo, que diferentes clases o dominios de la conducta (por ejemplo, los componentes de la memoria o las dimensiones de la inteligencia) revelan diferentes cursos de cambio ontogenético. El proceso de desarrollo no es un movimiento único dirigido hacia una mayor eficacia, como un crecimiento en aumento. Sino que más bien el desarrollo tiene lugar, a lo largo de la vida, como una ocurrencia conjunta de ganancias (crecimiento) y pérdidas (declive). - El desarrollo psicológico presenta una gran plasticidad interindividual o modificabilidad. En relación con las condiciones y experiencias vitales de un individuo, su curso evolutivo podrá adoptar múltiples formas. La necesidad de conocer el rango de esta plasticidad es tan importante como el conocer el curso medio del desarrollo relacionado con la edad. - El desarrollo individual puede también cambiar sustancialmente en función de las condiciones socioculturales presentes en un determinado periodo histórico y el modo cómo estas cambian a través del tiempo. Pero a su vez el desarrollo individual es un antecedente del desarrollo futuro de la especie humana. Es decir, la naturaleza de dicho desarrollo individual a lo largo del ciclo vital tiene consecuencias directas en las generaciones subsiguientes. - El estudio del desarrollo hay que abordarlo en un contexto interdisciplinar (Psicología del desarrollo, Antropología, Sociología, etc.). Cualquier explicación del desarrollo basada en una sola de estas disciplinas resultaría incompleta. Los psicólogos deben ayudarse de estas disciplinas para describir y explicar de modo comprensivo el desarrollo de la conducta. Unidad 2 • Desarrollo, del Sistema Nervioso a lo Largo del Ciclo Vital INTRODUCCIÓN La edad pone su sello en nuestra conducta y en la de todos los mamíferos. Los pasos, progresión y orden de los cambios son especialmente prominentes en la vida más temprana. Sin embargo, las ciencias de la vida enfatizan ahora el cambio como una característica del ciclo vital entero. El cambio es una propiedad inevitable de los estados biológicos. Shakespeare lo puso de manifiesto en «Así es si así os parece» cuando dijo: ... de hora a hora, maduramos y maduramos, y luego, de hora a hora, decaemos y decaemos. Hay dos calendarios que proporcionan el marco temporal del desarrollo del encéfalo. En el cal . tul: anterior usábamos el marco temporal de la evolución -el curso del desarrollo del sistema nervioso a través de millones de años-. En la primera parte de este capítulo, nos centraremos en el desarrollo estructural del encéfalo durante el curso típico de la vida de un individuo. En capítulos posteriores se aportan ejemplos de los cambios conductuales y fisiológicos que acompañan al desarrollo del encéfalo. Veremos cómo progresan las características de los encéfalos adultos, descritas en los Capítulos 2 y 3, durante la vida desde el útero hasta la tumba. Por ejemplo, la fertilización de un óvulo lleva a un cuerpo con un sistema nervioso que contiene miles de millones de neuronas con un increíble número de conexiones. La ocurrencia de este proceso es extraordinaria. Durante el apogeo de crecimiento prenatal del sistema nervioso humano, las neuronas surgen con una tasa ¡de 500.000 por minuto! Hay muchos problemas a considerar, incluyendo el modo en que se forman las células nerviosas y cómo se controla el establecimiento de conexiones entre las diversas regiones del sistema nervioso. ¿Se desarrolla el sistema nervioso mediante un proceso intrínsecamente guiado que sigue obligatoriamente el anteproyecto «escrito en los genes»? ¿Cómo ayuda la experiencia a guiar el sistema nervioso que está originándose? El crecimiento y desarrollo de un sistema nervioso es un proceso intrigante, especialmente en relación con la ontogenia de la conducta. Muchas teorías psicológicas enfatizan el desarrollo e intentan asignar pesos relativos a los papeles de la naturaleza y la cultura. ¿Hasta qué punto es importante la vida temprana para la posterior conducta cognitiva y emocional? Los estudios del desarrollo estructural y funcional del encéfalo pueden proporcionar sugerencias sobre estos aspectos. Al principio El camino desde un óvulo fertilizado hasta un organismo maduro es excesivamente complicado. Imaginemos, si podemos, el número de neuronas del encéfalo humano maduro. Las estimaciones recientes oscilan entre 100.000 millones y un billón, la última imagen ofrecida por Kandel y Schwartz (1985). Además, estos miles de millones de células muestran patrones específicos de organización altamente ordenados -un pasmoso logro de procesos del desarrollo y de la evolución-. Se están investigando muchos aspectos del sistema nervioso, que van desde las influencias químicas hasta las formas en que la experiencia afecta al «establecimiento de conexiones» en el sistema nervioso. Peso del encéfalo desde el nacimiento hasta la vejez Las medidas del peso del encéfalo en diversas etapas de la vida proporcionan un índice de su desarrollo. El peso debe considerarse como una especie de resumen de muchos procesos del desarrollo. Un estudio debido a Dekaban y Sadowsky (1978) da un retrato definitivo del peso del encéfalo humano a lo largo del ciclo vital. Este estudio se basa en medidas de los encéfalos de 5.826 personas, seleccionadas de entre 25.000 casos de varias ciudades. Los investigadores pesaron los encéfalos de individuos cuyas muertes se debieron a causas que no ejercieran influencias importantes sobre el encéfalo. La Figura 4-1 muestra los cambios con la edad en los pesos de los encéfalos en machos y hembras. Adviértase el rápido incremento en los primeros cinco años. El peso del encéfalo está en su apogeo entre los 18 y los 30 años, tras los cuales hay una disminución gradual. Ahora veamos como empieza el encéfalo el camino de su desarrollo. Figura 4-1 Peso del encéfalo humano como una función de la edad. Adviértase que la escala de edad se ha expandido para los primeros cinco años con intención de mostrar los datos más claramente durante este período de crecimiento rápido. (Adaptado de Dekaban y Sadowsky, 1978.) Emergencia de la Otro resumen del desarrollo del encéfalo lo proporciona el examen de los forma encefálica cambios de su forma global. Un nuevo ser humano comienza cuando un espermatozoide de aproximadamente 60 micrometros (µm) de largo atraviesa las paredes de un óvulo de 100-150 µm de diámetro. Este evento comienza un programa de desarrollo que lleva a un nuevo individuo. El comienzo de este programa tiene lugar en las trompas de Falopio, un conducto que llega hasta el ovario (Figura 4-2). Esta unión da como resultad,) una célula con 46 cromosomas, el número normal de los humanos. Estos cromosomas contienen el proyecto genético completo del nuevo individuo. El principio del programa de desarrollo es una rápida división celular. En doce horas la célula simple se ha dividido en dos células, y después de tres días se llega a convertir en un pequeño conglomerado de células homogéneas, como un racimo de uvas, de aproximadamente 200 µm de diámetro. Durante este período la esfera de células se ha ido moviendo hacia el útero, donde llega después de varios días. Entonces, el líquido de la cavidad uterina penetra en la esfera de células y las separa en dos grupos: 1) un grupo celular externo que da origen a la placenta y 2) una masa celular interna que se convierte en el embrión propiamente dicho. En el interior de la esfera de células se forma una cavidad. En esa etapa nuestro organismo se denomina Figura 4-2 Resumen de las etapas tempranas de desarrollo embrionario. La secuencia comienza a la izquierda con la liberación de un óvulo maduro (1) y su fertilización por el espermatozoide en la trompa de Falopio (2)-(3). El huevo fertilizado o zigoto (4) comienza a dividirse (5)-(7) en la trompa de Falopio. Cuando se ha formado un conglomerado de células homogéneas -la mórula (8)- la separación de capas celulares forma la etapa de blástula (9-10) y comienza el proceso de la implantación en el útero (11)-(12). La duración de estos procesos es de aproximadamente una semana para los seres humanos. Figura 4-3 Desarrollo embrionario humano entre la segunda y la cuarta semanas. (a) El embrión se ha comenzado a implantar en la pared uterina, constando de dos capas celulares. (b) La formación de las tres capas celulares y el engrosamiento de una de ellas -el ectodermo- lleva al desarrollo de la placa neural. (c) Comienzo del surco neural. (d) El surco neural se ha cerrado a lo largo de la longitud del embrión;el cierre del surco en el extremo anterior del neuroporo anterior concluye en el primordio del encéfalo. blástula (este término viene del griego blastos, «brote» o «capullo», y de la palabra griega para vejiga, para indicar un organismo hueco en embrión). Al final de la primera semana la blástula está implantada en la pared uterina, y las células placentarias se extienden en esta pared. Durante la segunda semana el embrión humano emergente muestra tres estratos celulares distintos. Estas capas son el comienzo de todos los tejidos del embrión (Figura 4-3). El sistema nervioso se desarrollará en la capa más externa, llamada ectodermo (de las palabras griegas para «externo» y para «piel»). Según se engrosan las capas celulares, crecen en un delgado disco oval. En el nivel ectodérmico de este disco, se marca una posición media por un surco -el surco primitivo-. En el extremo cefálico de este surco hay un grueso grupo de células. Esta etapa se da dos semanas después de la fertilización. Entonces se forman engrosamientos de ectodermo a ambos lados de la posición media. Estas son los pliegues neurales. El surco entre ellas se conoce entonces como el surco neural. La sucesión de eventos comienza a ser ahora más rápida. Las invaginaciones neurales se unen y convierten el surco en el tubo neural. En la parte anterior del tubo neural se evidencian tres subdivisiones. Estas subdivisiones corresponden a los futuros encéfalo anterior (prosencéfalo), encéfalo medio (mesencéfalo) y encéfalo posterior (rombencéfalo). (Recordar estas regiones del Capítulo 2, Figura 2-7.) La cavidad del tubo neural finaliza arriba, en el lugar donde se hallan los ventrículos encefálicos y los pasajes qué los conectan. (La morfología del sistema ventricular fue descrita en la Figura 2-4.) Al final de la octava semana, el embrión humano muestra los primordios rudimentarios de la mayoría de órganos corporales. La rápida evolución del desarrollo del encéfalo durante este período se refleja en el hecho de que al final de la octava semana la cabeza es la mitad del tamaño total del embrión. (Adviértase que el desarrollo humano se denomina embrión durante las primeras diez semanas después de la fertilización; posteriormente se llama feto.) La Figura 4-4 presenta una secuencia de visiones del desarrollo prenatal del encéfalo humano desde la semana 10 a la 41. Aspectos celulares del desarrollo del sistema nervioso Cuatro mecanismos celulares delicadamente controlados subyacen a los cambios anatómicos globales del sistema nervioso durante la vida fetal y embrionaria: 1) proliferación celular, 2) migración celular, 3) diferenciación celular y 4) muerte celular. Estos acontecimientos se producen con diferentes tasas y en diversos momentos en partes distintas del tubo neural. (Aquí nos centraremos en el sistema nervioso; sin embargo, se dan acontecimientos similares en la formación de otros órganos.) Proliferación celular La producción de células nerviosas se denomina proliferación celular. Las células nerviosas tienen su inicio corno una simple capa celular a lo largo de la superficie interna del tubo neural. Inicialmente las paredes del tubo neural están compuestas por una población similar de células. Estas células forman gradualmente una capa fuertemente empaquetada, la capa ventricular de células (células ventriculares), las cuales se dividen (Figura 4-5). (La capa ventricular también se denomina capa ependimal.) Cada célula se divide dando lugar a células «hijas», las cuales, a su vez, se dividen también. Todas las neuronas y laglía se derivan de células que se originan en la capa ventricular. La evidencia reciente sugiere que algunas células de la capa ependimal dan origen a la glía, y otras a las neuronas. La separación entre estos dos tipos de células tiene lugar muy pronto en la organización de la capa ependimal. En la mayoría de los mamíferos el proceso de formación de las células neurales en la capa ventricular continúa hasta el nacimiento, pero se añaden muy pocas después de éste (Rakic, 1974). En algunas regiones encefalicas se produce la aparición postnatal de células nerviosas. Por ejemplo, en el cerebelo humano surgen neuronas durante meses después del nacimiento. Hay una «fecha de nacimiento» característica de especie para cada parte del encéfalo de un animal. Esto significa que hay un programa cronológico ordenado para el desarrollo del encéfalo, y que es posible fijar los días aproximados del desarrollo en los que se forman determinados grupos de neuronas. Por supuesto, dada la complejidad de los encéfalos de los vertebrados, es bastante difícil trazar la vía del desarrollo celular desde la pequeña población inicial del células ependimales. Los descendientes desaparecen en la multitud. Sin embargo, en los sistemas nerviosos de algunos invertebrados más simples con muy pocas neuronas, pueden trazarse los linajes celulares de forma más fácil y completa. Figura 4-4 Representaciones laterales del encéfalo humano durante el desarrollo fetal. Adviértase el proceso gradual de desarrollo de los giros y surcos. Los números muestran las edades de la gestación en semanas. Los encéfalos se muestran a un tercio del tamaño real. (De J. C. Larroche, Capítulo 11, Parte II, «The development of the central nervous system during intrauterine life». Figura 1 [p. 258] y Figura 2 [p. 259]. En F. Falkner [ed.], Human development [Philadelphia, Pa.: W. B. Saunders, 1966].) Migración celular Las neuronas del sistema nervioso en desarrollo están siempre en movimiento. En alguna etapa las células nerviosas que se forman en la capa ventricular a través de división mitótica comienzan a desplazarse. Este proceso se conoce como migración celular. Las células nerviosas en esta etapa, se conocen como neuroblastos. Adquieren cortas extensiones en los extremos de la «cabeza» y la «cola». Algunas descripciones de las células migradoras las comparan con un sendero de hormigas activa (Figura 4.6). En los primates la migración de células nerviosas en la mayoría de regiones del encéfalo está prácticamente completa en el nacimiento. Pero en los encéfalos de las ratas, las células nerviosas continúan migrando en algunas regiones durante varias semanas después de nacer. Figura 4-5 Proliferación de los precursores celulares de las neuronas y la glia. La parte (a) muestra una pequeña sección de la pared del tubo neural en una etapa temprana del desarrollo embrionario cuando sólo se ven las capas ventricular (V) y marginal (M). Posteriormente, como se muestra en (b), se desarrolla una capa intermedia (1) según se hace más gruesa la pared. La parte (c) muestra la migración de los núcleos de las neuronas desde la capa ventricular hacia las capas externas. Algunas células, sin embargo, vuelven a la capa ventricular y continúan la división, y entonces las células hijas migran a las capas externas. Durante esta fase del desarrollo del encéfalo las células no se mueven sin propósito, de una forma azarosa. Las claves sobre el proceso de la migración celular vienen de los estudios que emplean sustancias radiactivas que llegan a incorporarse en las células antes de la migración. Estas sustancias «identifican» la célula de modo que puede ser seguida y pueden perfilarse claramente sus vías migratorias. Elegantes estudios de este proceso llevados a cabo por Rakic (1985) muestran que algunas células en el encéfalo en desarrollo se mueven a lo largo de la superficie de un tipo de célula glial inusual que aparece bastante pronto. Estas células gliales se extienden desde la superficie interna del sistema nervioso emergente hasta la externa (Figura 47). La vía glial actúa como una serie de alambres guía, con cada célula nerviosa nuevamente creada progresando a lo largo de su longitud. Esta serie de alambres guía reciben el nombre de glia radial. Algunas células nerviosas que se forman más tardíamente migran de manera diferente. Estas células son atraídas a las Figura 4-6 Migración de los precursores de las células de Purkinje del cerebelo del pollo. A los 8 días, estas células aparecen como un gran grupo de «hormigas» fluyendo lejos de la. región de formación inicial. A los 11 días, han comenzado a formar una capa definida. Esta capa llega a estar más desplegada a los 14 días, y a los 19 días es una única capa de células sobre una extensa superficie de tejido que ha comenzado a formar pliegues. (De Levi-Montalcini, 1963.) superficies de las neuronas. Rakic (1985) ha descrito ejemplos de este mecanismo migratorio en el cerebelo, donde las células nerviosas de reciente formación migran a lo largo de los axones de las células nerviosas formadas más tempranamente. De hecho, algunas neuronas progresan lateralmente a lo largo de los axones horizontales y entonces siguen la glía hacia abajo en la corteza cerebelosa. Algunos trastornos del desarrollo del encéfalo implican fallos en el mecanismo de migración celular, que conduce a una población de células vastamente reducida o con una disposición desordenada. La migración de las células y el crecimiento externo de las extensiones celulares nerviosas, tales como dendritas y axones, también implican varias sustancias químicas. La adhesión de partes es importante para este proceso; las moléculas que promueven la adhesión de los elementos en desarrollo del sistema nervioso han sido descritas por algunos investigadores, los cuales los denominan MAC -moléculas de adhesión celular(Edelman, 1984). Diferenciación celular Al principio, las nuevas células nerviosas no poseen más parecido con las células nerviosas maduras del que poseen con la:; células de otros órganos. Una vez que las células alcanzan sus destinos, :;in embargo, comienzan a adquirir la apariencia distintiva de las neuronas características de sus regiones particulares. Este proceso es la diferenciación celular. La Figura 4-8 muestra el progresivo despliegue de l ::s células de Purkinje de la corteza cerebelosa. El crecimiento exterior de las dendritas de estas células aparece después del comienzo de su alineación en una capa única. Lentamente se forman más y más ramificaciones que expanden progresivamente la superficie receptiva de la célula de Purkinje. El por qué comienza este proceso de expansión de Figura 4-7 Muy pronto en el desarrollo, las células de la glía radial se expanden por la extensión de los hemisferios cerebrales emergentes como se muestra en la parte superior. Ellas actúan como alambres guía para la migración de las neuronas, como se muestra en la ampliación de la página anterior. Una ulterior ampliación, a la derecha, muestra una neurona migrando a lo largo de una fibra glial radial simple. (De Cowan, 1979, después de Rakic.) las dendritas, sigue siendo un misterio. Se conocen algunas influencias. Por ejemplo, la auto-organización intrínseca es un factor ciertamente importante; las células nerviosas en cultivos de tejidos crecen de un modo típico aunque sean privadas de algunas conexiones usuales (por ejemplo, Seil, Kelley y Leiman, 1974). Sin embargo, muchos estudios de investigación contemporáneos también muestran que el ambiente neural Figura 4-8 Desarrollo de las células de Purkinje :n el cerebelo humano en varias edades fetales y postnatales, mostrando la diferenciación de la forma. (De Zecevic and Rakic, 197(3.) influye en la diferenciación de las células nerviosas. Cualquier región dada del sistema nervioso maduro contiene un grupo de células nerviosas que pueden incluir dos o más tipos. Por ejemplo, en la corteza cerebelosa hay células de Purkinje y células granulares. Sin embargo, todas las células que migran a una región son neuroblastos que al principio se veían exactamente iguales. Así, un neuroblasto dado tiene el potencial de transformarse en uno o varios tipos diferentes de células nerviosas. Las múltiples potencialidades del neuroblasto en crecimiento parecen estar programadas de una forma ordenada en cualquier región. Una regla general que refleja este orden es: en una región que está organizada en capas (por ejemplo, la corteza cerebral o la corteza cerebelosa), se producen primero grandes células, seguidas de células pequeñas. Así, en el cerebelo, las grandes células de Purkinje se forman primero. Cuando se han alineado en una fila, los neuroblastos que se convertirán en las pequeñas células granulares comienzan a migrar. La consecución de la forma típica de una neurona depende en parte de los determinantes de la célula individual y en parte de las influencias de las células vecinas. Algunas partes de una célula dada crecen de un forma típica sin más influencia que el ambiente. Otros componentes parecen reponder a características del ambiente en el encéfalo, tales como la presencia de otras células. Muerte celular Por extraño que pueda parecer, la muerte celular es una fase crucial del desarrollo del encéfalo, especialmente durante las etapas embrionarias. De hecho, en algunas regiones del encéfalo y la médula espinal, la mayoría de las células nerviosas mueren durante el desarrollo prenatal. En la Figura 4-9 se muestra un ejemplo de esta clase de fenómenos del desarrollo. Este proceso está influido por varios factores. Entre ellos está el tamaño del campo de la superficie corporal, que se conectará en última instancia a una región del sistema nervioso central. Por ejemplo, en los renacuajos, si un investigador amputa una pata antes de que se hayan formado las conexiones de la médula espinal mueren muchas más motoneuronas espinales en desarrollo de las que morirían si la pata hubiese permanecido en su posición. Contrariamente, añadiendo una pata extra -lo que es posible en anfibios- se reduce apreciablemente la pérdida usual de células, así la médula espinal madura, en este ejemplo, tiene un número de neuronas mayor de lo usual. Estas observaciones sugieren que la diana de una población de células nerviosas en desarrollo -el lugar a que ellas se conectan- influye en la supervivencia de estas células. Algunos investigadores han sugerido que durante el desarrollo hay competición entre células para conectarse a estructuras diana tales como otras células nerviosas u órganos, como por ejemplo los músculos. De acuerdo con esta visión, aquellas células que establecen conexiones rápidamente permanecen; aquéllas sin lugar para realizar conexiones sinápticas mueren. Otro determinante de la progresión y extensión de la muerte celular es el nivel de ciertas sustancias químicas naturales. Por ejemplo, en algunos sistemas nerviosos de invertebrados, la muerte celular es inducida por la acción de hormonas concretas. Este tipo de muerte de células nerviosas se observa durante la metamorfosis de los insectos. Truman (1983) ha descrito la muerte de células nerviosas en los sistemas nerviosos de las mariposas inducida por las hormonas que producen la transformación de oruga a Figura 4-9 Patrones de la muerte de células nerviosas durante el desarrollo temprano del sistema nervioso del pollo. (a) Columna motora lateral. (Adaptado de Hamburger, 1975.) (b) Ganglio ciliar. (Adaptado de Landmesser y Pilar, 1974.) (c) Núcleo troclear del tronco encefálico. (Adaptado de Cowan y Wenger, 1967.) mariposa. Las células deMauthner de algunos anfibios cambian como resultado de la secreción de hormonas tiroideas. Las células degeneran cuando el estilo de vida del animal cambia de acuático a terrestre. La muerte de células nerviosas puede también efectuar un emparejamiento numérico entre el desarrollo de poblaciones celulares. Por ejemplo, consideremos un caso hipotético, la población de células nerviosas A contiene 100 células. Sus axones se extienden hacia la población de células nerviosas B, que consta de 50 células. El excesivo número de células nerviosas en la población A asegura la conexión a las células nerviosas de la población B, pero probablemente habrá un grupo de células que no serán necesarias después de haber forjado los lazos efectivos. El resultado: un grupo de células de la población A muere. El mecanismo mediador puede ser la capacidad de tomar un factor de «supervivencia» del ambiente de la población blanco. Algunos investigadores han sugerido también que la muerte celular puede eliminar «conexiones incorrectas». Sin embargo, los análisis críticos de esta interesente sugerencia no permiten su confirmación (Lance-Jones, 1984). Ciertamente, parece que la muerte de las células nerviosas durante el desarrollo es un mecanismo para «esculpir» el sistema nervioso en desarrollo. Procesos ulteriores del desarrollo El encéfalo humano incrementa cuatro veces su peso y tamaño entre el nacimiento y la madurez. En gatos, conejos, ratas y otros animales se hacen evidentes cambios semejantes desde la infancia hasta la edad adulta (Tabla 4-1). ¿Qué tipo de cambios estructurales postnatales se dan para que se produzca este crecimiento en peso y tamaño del encéfalo? Vamos a considerar cuatro tipos de cambios estructurales a nivel celular que caracterizan el desarrollo del encéfalo durante los períodos postnatales tempranos. Mielinización El desarrollo de una vaina alrededor de los axones -un proceso llamado mielinización- cambia enormemente la velocidad a la que éstos conducen los mensajes. Esto tiene un fuerte impacto en la conducta, ya que afecta profundamente el orden temporal de los acontecimientos en el sistema nervioso. Desafortunadamente hay pocos estudios que empleen al mismo tiempo técnicas modernas biológicas y conductuales para relacionar los atributos biológicos del sistema nervioso con cambios en la conducta. Así, el trabajo de relacionar cambios en la conducta con la mielinización permanece aún abierto en la necesidad de retomarse. En humanos, la fase más intensa de mielinización se da poco después del nacimiento. (Sin embargo, algunos investigadores creen que la mielina puede añadirse a los axones durante toda la vida.) Los primeros tractos nerviosos que se mielinizan en el sistema nervioso humano se encuentran en la médula espinal. Desde aquí, la mielinización se extiende sucesivamente al encéfalo posterior, el encéfalo medio y el encéfalo anterior. La mielinización más temprana del sistema nervioso periférico es evidente en los nervios craneales y raquídeos alrededor de las 24 semanas después de la concepción. En la corteza cerebral, las zonas sensoriales se mielinizan antes que las motoras; correspondientemente, las funciones sensoriales maduran antes que las motoras. Tabla 4-1 Incremento en el peso del encéfalo de algunos mamíferos desde el nacimiento hasta la madurez Especies Recién nacido Adulto Incremento (g) (g) (%) Cobaya 2,5 4 60 Humano 335 1.300 290 Gato 5 25 400 Conejo 2 10,5 425 Rata 0,3 1,9 530 Fuente: De Altman (1967). Formación de sinapsis y dendritas En las células nerviosas, los mayores cambios que se producen entre el nacimiento y la madurez tienen lugar en las ramificaciones y las conexiones entre neuronas. La Figura 4-8 ya nos ha mostrado el enorme aumento de la amplitud de las dendritas, que parece implicar procesos distintos a aquellos que se dan en el crecimiento de los axones. En el ápice de las dendritas hay conos en crecimiento, que son extremos abultados de los que emergen las extensiones. Algunos investigadores han encontrado incluso conos de crecimiento de las dendritas en animales adultos. Esto puede estar relacionado con Figura 4-10 Desarrollo postnatal de las sinapsis. (a) Ganglio cervical superior de la rata (según Smolen, 1981). (b) Corteza visual de la rata (Blue y Parnavelas, 1983). (c) Corteza visual humana (Huttenlocher et al., 1982). el excitante descubrimiento de que la elongación de la: dendritas puede continuar a lo largo de la vida en respuesta a demandas funcionales. Las sinapsis se incrementan a una tasa rápida, particularmente en las dendritas (Figura 4-10). En muchas células nerviosas, las sinapsis se forman en la espinas dendríticas. Las espinas en sí mismas proliferan rápidamente después del nacimiento. Estas conexiones pueden verse afectadas por la experiencia postnatal, como veremos en el Capítulo 17. Para proporcionar apoyo a las necesidades metabólicas del expandido árbol dendrítico, el cuerpo celular nervioso aumenta enormemente su volumen. Producción de neuronas después del nacimiento Tradicionalmente, muchos investigadores de la ontogenia del sistema nervioso han creído que la mayoría de los mamíferos tienen en el nacimiento todas las células nerviosas que poseerán. Flan explicado el crecimiento postnatal del encéfalo enteramente en términos de crecimiento en el tamaño de neuronas y la suma de células no neurales (gliales). En los años recientes, sin embargo, se ha modificado esta creencia, en primer lugar debido a que actualmente parece que durante un tiempo después del nacimiento se añaden pequeñas neuronas. Algunos investigadores han sugerido incluso que el nacimiento puede inducir una aceleración en la tasa de producción de estas pequeñas células. Sin embargo, esta visión no ha ganado un apoyo amplio. Otros investigadores han argüido que es la madurez del encéfalo la que determina el momento del nacimiento. La corriente más ampliamente aceptada es que todas las grandes neuronas que contendrá el encéfalo se hallan ya en el nacimiento. Sin embargo, hay unas pocas regiones alrededor de los ventrículos encefálicos, llamadas zonas subventriculares, en las que la división mitótica de los precursores de las células nerviosas sigue siendo evidente después del nacimiento. Varias regiones del encéfalo de las ratas, incluyendo el bulbo olfatorio y el hipocampo, parecen añadir pequeñas neuronas derivadas de esta región. De hecho, se ha dicho (Graziadei y Monti-Graziadei, 1978) que las células nerviosas del órgano terminal olfatorio son reemplazadas a lo largó de la vida. El dogma de que no se incorporan nuevas neuronas al sistema nervioso adulto está fuertemente desafiado por algunas excitantes investigaciones nuevas de Nottebohm (1987) sobre la neurología dei canto de los pájaros (discutido en los Capítulos 11 y 18). Brevemente, se sabe que el desarrollo del canto de los pájaros en los machos de algunas especies está bajo el control hormonal, dependiente de la hormona masculina testosterona. En un trabajo anterior Nottebohm determinó las partes del encéfalo del pájaro que, son responsables del aprendizaje y la realización del canto. Durante el curso de este trabajo, advirtió que al menos una parte del circuito encefálico relevante para el canto del pájaro es mayor en primavera y se reduce a la mitad del tamaño en el invierno. Este cambio estacional en el tamaño de una región del encéfalo está relacionado con los niveles de hormona masculina y la conducta de canto. Parte de esta variación estacional en el tamaño de la región del encéfalo está ocasionada por variaciones estacionales en la amplitud y la ramificación dendrítica. Sin embargo, Nottebohm ha presentado evidencia de que el incremento en la primavera está también relacionado con la suma de nuevas neuronas. Aparentemente, las nuevas neuronas se firman en la región subventricular y migran a la cercana región de control vocal del tronco encefálico. En el curso de un año se hacen evidentes importantes ganancias y pérdidas en el número de neuronas. Estas observaciones sugieren la necesidad de reexaminar la creencia de que el encélalo adulto no incorpora nuevas neuronas. Como mínimo, tales investigaciones renovadas podrían apoyar un mejor conocimiento de las condiciones que llevan al usual cese de suma de neuronas en la edad adulta. Formación de células gliales Las células gliales se desarrollan de las mismas poblaciones de células inmaduras que las neuronas. Las influencias que determinan si la célula se desarrolla en una neurona o en una célula glial siguen siendo un misterio. A diferencia de las neuronas, las células gliales continúan proliferando a lo largo de la vida. A veces este proceso puede llegar a ser aberrante, dándose tumores gliales (gliomas) del encéfalo. La producción de glia continúa durante más tiempo que la producción de neuronas y muestra su mayor cambio más tarde. De hecho, la fase más intensa de proliferación glial en muchos animales se da después del nacimiento, cuando las células gliales se añaden a partir de células inmaduras localizadas en las zonas subventriculares. Ejemplos de la formación de regiones neurales Cualquier región del encéfalo está caracterizada per una disposición distintiva de las células nerviosas y sus procesos. En alguna:; regiones, como la corteza cerebelosa y la cerebral, las células nerviosas están distribuidas en diversas capas. La discusión de los factores del desarrollo que están involucrados en la adquisición de la forma característica de una región proporciona un enfoque de la complejidad de la ontogenia del encéfalo. El conjunto del desarrollo de cada región del encéfalo sigue una precisa escala temporal; algunas partes del esquema parecen estar relacionadas con las interacciones mutuas de las células en la región en desarrollo. Para mostrar cómo una región particular adquiere su forma ordenada característica, vamos a tomar como ejemplo las cortezas cerebelosa y cerebral. Formación de la corteza cerebelosa Como vimos en el Capítulo 2, el corteza cerebelosa adulta consiste en una estructura con numerosos pliegues (llamados folia) y una disposición laminar (en capas), como sigue: 1. Una capa molecular externa con pequeñas células y una banda de fibras (axones). 2. Una capa media de grandes células (la capa de las células de Purkinje). 3. Una capa profunda y gruesa de células muy pequeñas -la capa de las células granulares. Vimos esta disposición laminar del cerebelo adulto en la Figura de Referencia 28. La migración inicial de las células que forman el cerebelo implica a las células de Purkinje, que al principio son pequeñas y dispersas pero que más tarde llegan a formar una capa única y uniforme de grandes células (Figura 4-11). En los seres humanos, las células de Purkinje crecen más deprisa entre el final del embarazo y un año después del nacimiento. En la rata, crecen más rápidamente justo después del nacimiento, entre los olías 2 y 30. Después de la salida a escena de las células de Purkinje, se forman las células más pequeñas del cerebelo. Inicialmente su patrón de migración 'as toma de la superficie granular externa. Entonces descienden alrededor y pasan por entre las células de Purkinje para constituir la población celular mí s profunda. El hecho de que en la rata la mayor parte del desarrollo del cerebelo se dé después del nacimiento ha permitido que este proceso sea fácil de estudiar. Cualquier tratamiento, incluyendo la exposición leve a rayos X, interfiere los neuroblastos en reciente desarrollo, pero no daña a las células que están ya diferenciadas o que son relativamente maduras. Figura 4-11 Dibujos esquemáticos del desarrollo de las células en el cerebelo de la rata, enfatizando la maduración de las células de Purkinje. Comienza por la capa granular externa (CGE), a la que las células migran durante el desarrollo cerebeloso temprano. Hacia el cuarto día postnatal, las células de Purkinje están dispuestas en una capa simple. Del día 4 al día 7 se desarrollan las células en cesta (mostrado en marrón el día 8) y la dendrita principal de las células de Purkinje crece hacia la superficie del cerebelo. Desde el día 8 hasta el 11 se forman las células estrelladas (mostradas en marrón en el día 12) y crecen las principales ramificaciones de las dendritas de las células de Purkinje. Del día 12 en adelante, se forman un gran número de células Granulares (mostradas en marrón en el bloque del día 15); los cuerpos granulares migran hacia abajo, más allá de las células de Purkinje, pero sus axones forman las fibras paralelas que sinaptan con las pequeñas ramificaciones espinosas del árbol dendrítico de Purkinje. (Adaptado de Altman, 1976.) Figura 4-12 Migración de células en la corteza cerebral de la rata. Las células corticales de la rata se originan y comienzan su migración antes del nacimiento. Las células que se originan antes migran a las capas más profundas de la corteza; las células producidas pocos días antes del nacimiento constituyen las capas celulares superiores (II y IR). La migración de células a las capas superiores continúa durante varios días después del nacimiento. (Adaptado de Berry, Rogers y Eayrs, 1964.) Se desarrollan en secuencia tres poblaciones diferentes de pequeñas células: las células en cesta, las células estrelladas y las cébalas granulares. Altman (1976) expuso a ratas jóvenes a rayos X en el curso d-: unos pocos días, lo que detuvo el desarrollo de uno o más tipos de estas pequeñas células en los cerebelos de las ratas. Esto tuvo consecuencias desastrosas para el desarrollo de la gran ramificación dendrítica de las células de Purkinje. Si administramos breves irradiaciones cada día de de el día 4 hasta el (lía 7 evitamos la formación de las células en cesta. Esto a su vez evita el posterior crecimiento de la dendrita principal de las células d; Purkinje; las dendritas principales pueden crecer en cualquier dirección y llegar a enrollarse. La irradiación entre los días 8 y 11 evita la formación de las células estrelladas y además interfiere con el crecimiento de las ramas principales de las dendritas de la célula de Purkinje. Finalmente, la irradiación do los días 12 a 15 reduce el número de células granulares y disminuye la compleja ramificación del árbol dendrítico de las células !e Purkinje (Altman, 1976). La irradiación también produce efectos conduciuales; interfiere con el mantenimiento de la postura y se ha informado de deterioro en el aprendizaje de laberintos (Pellegrino y Altman, 1977). Formación de la corteza cerebral Los 50.000 millones de neuronas de la corteza cerebral humana están dispuestas en capas, variando las células de cada una en forma y tamaño. Esta disposición en capas muestra variaciones en diferentes partes del cerebro. Estas variaciones se han empleado para definir los límites de diferentes regiones corticales. Aquí veremos la forma en que crece y desarrolla su disposición en capas la neocorteza cerebral. Si examinamos el tubo neural cerrado de un embrión humano al final de la tercera semana después de la fertilización, revela una zona de células alrededor de las superficies internas. Esta temprana proliferación de células en el extremo rostral culmina en la formación de la lámina cortical, el comienzo de la corteza cerebral. La intensa división celular en este extremo continúa produciendo células que llegarán a ser con el tiempo las neuronas de la corteza cerebral. Esta rápida proliferación continúa hasta el sexto mes de vida fetal, momento en el cual la corteza cerebral posee su dotación completa de neuronas. Entonces éstas se alinean en estratos, aunque apenas se asemejan a las capas corticales del cerebro adulto. La formación de capas corticales en la corteza cerebral sigue un proceso regular, aunque el mecanismo que lo guía sigue siendo controvertido. Las células que se forman a lo largo de la superficie ventricular (ependimal) migran lejos de ella. Cada nueva célula migra más allá que aquellas que nacieron antes (Figura 4-12). Estas nuevas células se mueven más cerca de la superficie cortical. Las células más viejas se encuentran en los e., tratos más profundos. El tiempo de generación -el ciclo mitótico para la producción de una célula cortical- es de alrededor de 11 horas y permanece constante a lo largo del desarrollo de la corteza. Sin embargo, el tiempo de migración el intervalo entre el nacimiento de la célula y la llegada a su posición final-es progresivamente mayor, durando alrededor de cinco días en el último grupo de nuevas células corticales. La fase más intensa de crecimiento dendrítico y formación de sinapsis en la corteza cerebral se da después del nacimiento. La elaboración del desarrollo cortical postnatal humano se ilustra en la Figura 4-13. ¿Por qué las conexiones neurales van hacia donde van? Todos los miembros de una especie generan tipos similares de células nerviosas. Además, estas células tienen una disposición característica. Este orden se evidencia en la forma en que las células se agrupan juntas en diferentes regiones encefálicas. Una característica estructural que es de particular interés para la conducta es la ordenación y la especificidad de las conexiones entre células individuales y entre regiones. La conducta adaptativa de cualquier animal depende claramente de cómo se halla «conectado» -esto es, la forma en que están formadas las conexiones-. El proceso de formación de conexiones ¿es invariable, especificado por mecanismos controlados por la maquinaria genética? Existen tres respuestas habituales a esta pregunta que pueden resumirse como sigue: 1. Las principales conexiones que se forman durante el desarrollo están muy fuertemente especificadas por mecanismos innatos. 2. Los aspectos detallados de las conexiones centrales pueden modificarse por el entrenamiento y la experiencia. 3. Hay una intensa competencia entre las neuronas individuales y entre grupos de neuronas para formar conexiones, con lo que si algunas unidades son inhibidas o eliminadas, se toman sus conexiones por neuronas adyacentes. Vamos a examinar algunas de las investigaciones que han dado lugar a estas conclusiones. Los miles de millones de células nerviosas, todas creciendo a la vez, dirigen de algún modo la realización de conexiones apropiadas con otras y la formación de los intrincados circuitos que median la conducta compleja. Una neurona envía fuera un axón de menos de un milímetro de largo. Otra neurona envía su axón a lo largo de un tracto particular de más de un metro. Al final de su vía, cada axón forma conexiones en lugares específicos dentro de una región específica del sistema nervioso. Por otra parte, algunos axones terminan sobre zonas particulares de las dendritas de células nerviosas específicas. Cuando se piensa sobre esta formación de vías y conexiones en el sistema nervioso, parece como si cada célula nerviosa tuviese instrucciones sobre una dirección particular, un lugar en el que debe establecer conexiones. ¿Cómo podemos dar cuenta de las conexiones altamente ordenadas que se forman durante el desarrollo del sistema nervioso? La investigación pionera de este complejo problema fue hecha por Roger Sperry, un neuropsicólogo y neuroembriólogo americano que fue galardonado con el Premio Nobel en 1981. En los años cuarenta comenzó una serie de observaciones experimentales de los sistemas visuales de anfibios y peces. Estos estudios aprovechaban las notables capacidades de estos animales para regenerar tejido, incluso tejido nervioso. Como veremos, muchos estudios del desarrollo neural se Figura 4-13 (a) Desarrollo del engrosamiento de la corteza cerebral humana. (De Rabinowicz, 1986.) (b) Visión histológica de la corteza cerebral en el desarrollo temprano humano. Panel de la izquierda, un mes; panel medio, seis meses; panel de la derecha, 24 meses. (De Conel, 1939, 1941 y 1959.) interesan en el sistema visual, en parte debido a que la visión juega un importante papel en la conducta, y en parte por la ordenada proyección espacial del campo visual desde la retina a través de los centros visuales del encéfalo (Figura 4-14). Más tarde veremos que el desarrollo del sistema visual en los mamíferos está influido claramente por la experiencia temprana con estímulos visuales. Disposición del sistema visual de los anfibios Para comprender algunas de las principales observaciones experimentales de Sperry, vamos a considerar brevemente la distribución del sistema visual en anfibios y peces. La retina es una población de elementos fotosensibles que proporcionan un mapa del mundo visual (Figura 4-14). Los axones eferentes de las células nerviosas de la retina constituyen el nervio óptico. En los anfibios estas fibras atraviesan al lado opuesto del encéfalo y terminan de forma ordenada en una estructura llamada tectum óptico. Este es un centro neural fundamental para la visión en estos animales. La superficie del tectum proporciona, en algún sentido, un mapa de la retina. Así un objeto en un lugar determinado del mundo exterior excita un lugar particular de la retina, lo que a su vez activa un lugar específico del tectum óptico. Casi parece como si cada punto de la retina «supiese», o llegase a estar enterado del curso del desarrollo, lo que le Figura 4-14 Esquema del sistema visual de la rana. El campo visual está representado por la flecha grande frente a la cabeza de la rana. Las partes laterales del campo estimulan las partes nasales de las retinas; la parte central del campo estimula las partes temporales de las retinas. Las eferencias de la retina (los axones de las células ganglionares) se dirigen al tectum en el lado opuesto de la cabeza. Las partes temporales de la retina proyectan a las partes rostrales del tectum y las partes nasales de la retina lo hacen a las regiones más caudales. Las partes superiores de la retina (no mostradas en esta ilustración), también llamadas dorsales, se dirigen a las porciones laterales del tectum y las partes más bajas de la retina (ventrales) van a las porciones mediales del tectum. llevaría a conectar con el tectum óptico. Podríamos imaginarnos que hay algún tipo de etiqueta en cada axón creciente que le explica donde ir. Los estudios iniciales de Sperry (y otros) no requerían herramientas anatómicas o fisiológicas sofisticadas. Los experimentadores dejaron que la conducta visual de los anfibios les indicara las conexiones del encéfalo que están relacionadas con las percepciones del campo visual. Muchos anfibios muestran respuestas altamente estereotipadas a estímulos visuales. Se orientan correctamente hacia pequeños objetos en movimiento y los atacan propulsando la lengua, especialmente cuando los objetos se parecen a pequeños insectos, como moscas. Por lo tanto, sus respuestas pueden ser empleadas para revelar la representación del campo visual del encéfalo. Especificidad de las conexiones retino tectales En los experimentos iniciales Sperry cortó el nervio óptico y observó la forma en que reaparecía la conducta guiada visualmente. Después de que las fibras hubieron crecido hacia atrás, hacia el tectum óptico (un período de meses), los animales eran capaces de realizar la conducta con la misma exactitud que al principio. ¿Qué implicaciones tiene la restauración de esta respuesta conductual para el restablecimiento de las conexiones entre la retina y el tectum óptico? Deben considerarse dos alternativas posibles: 1. Los axones que vuelven a crecer entran en el tectum en un laberinto de conexiones azarosas, y la experiencia (éxito o error en la localización de alimento) determina la supervivencia de las conexiones que informan de localizaciones en el espacio; es decir, las redes son reeducadas sobre la localización de objetos. 2. Las fibras del nervio crecen hacia atrás a sus posiciones originales en el tectum óptico y simplemente restablecen el mapa original del mundo visual. Varios experimentos hoy clásicos permitieron a Sperry elegir entre estas alternativas. Lo que hizo fue rotar los ojos de un tritón 180 grados. Esto invirtió completamente el campo visual del tritón. Arriba y abajo, derecha e izquierda estaban invertidos. Después de la regeneración de las conexiones visuales, se vio que la conducta del animal estaba invertida: cuando se presentaba un pequeño señuelo en la mitad superior del campo visual, la lengua era lanzada hacia abajo. Cuando se presentaba el señuelo cerca de la porción nasal del eje horizontal, el animal apuntaba al lado. Estas inversiones de conducta persistieron durante años a pesar de que fueran marcadamente inadaptativas, aparentemente no había reeducación de la conducta de localización. Estas observaciones llevaron a Sperry a concluir que la regeneración de axones ópticos vuelve a conectar sus posiciones originales en el tectum. Recreaban un patrón de conexiones ordenadas. La explicación de Sperry ha venido a ser conocida como la doctrina de la neuroespecificidad. El arguyó que durante la diferenciación de las células de la retina, cada célula adquiría una identidad única, llegaba a ser específica, se pensaba que poseía un etiqueta que la relacionaba con una cierta posición del campo visual del animal (Figura 4-15a). Los axones que salían de estas células, de acuerdo con Sperry, eran bioquímicamente únicos. Cuando alcanzaban el tectum, buscaban células que tuviesen una identidad química similar. Hay así un apareamiento de células de acuerdo con una etiqueta química. Límites de la neuroespecificidad Supongamos que extendemos el concepto de neuroespecificidad hasta que cubra la génesis de todas las conexiones del sistema nervioso; esto es, supongamos que mantenemos que existe virtualmente una total rigidez, una específica ción genética de las conexiones neurales. Tendríamos muchos problemas. Uno obvio es que la limitada capacidad de información de los genes convierte en poco posible la noción de etiquetas químicas diferentes para cada neurona. Además, debe quedar alguna capacidad genética sobrante para la planificación de órganos como los brazos, las piernas y el corazón. Se han sugerido alternativas a la total especificación individual, empleando gradientes químicos determinados por muy pocas sustancias. La Figura 415b presenta una de estas visiones. CUADRO 4-1 Degeneración y regeneración del tejido nervioso Cuando se lesiona una célula nerviosa madura, pueden darse varias formas de regeneración. Sin embargo, en los sistemas nerviosos de los mamíferos es rara la completa restitución de la célula nerviosa lesionada. La Figura del Cuadro 4-1 ilustra varias formas características de degeneración y regeneración en el sistema nervioso central y en el periférico. La lesión cercana al cuerpo celular de la neurona produce una serie de cambios que producen la eventual destrucción de la célula. Este proceso se denomina degeneración retrógrada. La sección del axón a cierta distancia del cuerpo celular produce la pérdida de la parte distal del axón (la parte que está separada de la continuidad con el cuerpo celular). Este proceso se llama degeneración anterógrada o walleriana. La parte del axón que permanece conectada al cuerpo celular puede volver a crecer. Varios axones lesionados en el sistema nervioso periférico vuelven a crecer rápidamente. Desde la parte del axón que todavía permanece conectada al cuerpo de la célula nerviosa crecen brotes que avanzan lentamente hacia la periferia. Algunos animales tienen una ventaja envidiable. Después de una lesión en el encéfalo, varios peces y anfibios parecen ser capaces de regenerar grandes partes del encéfalo mismo. Desde un punto de vista experimental, nuestro interés en la regeneración del sistema nervioso central reside fundamentalmente en el hecho de que la regeneración implica procesos que parecen similares al desarrollo original. Estudiando la regeneración podemos aumentar nuestra comprensión de los procesos originales del crecimiento del sistema nervioso. Desde un punto de vista terapéutico, estos estudios pueden ayudar a los científicos a aprender cómo inducir reparaciones y nuevos crecimientos de tejido neural lesionado en humanos. Figura de Cuadro 4-1 Tipos de degeneración de células nerviosas (a) Antes de la lesión (b) y (c) degeneración anterógrada; la lesión causa la pérdida de la sección distal del axón. (d) y (e) Degeneración retrógrada. La lesión también produce degeneración del cuerpo celular, produciendo en algunos casos una atrofia completa. (f) Degeneración transneuronal. La pérdida de aferencias puede producir cambios en otras células de la vía. (g) Recuperación. Los axones lesionados pueden hacer brotar nuevas terminaciones. Varios experimentos han aportado también dudas sobre la probabilidad de que haya una completa especificación predeterminada de todas las conexiones neurales. Estos experimentos han mostrado que hay alguna plasticidad en las conexiones retinotectales. Tal plasticidad o adaptabilidad sugiere que, como mínimo, es posible que las neuronas tectales puedan ser capaces de reespecificar sus propiedades durante el período de regeneración. Buenos ejemplos son los llamados experimentos de disparidad de tamaño. En uno de estos experimentos, Yoon (1979) mostró que cuando se extraía la mitad del tectum, toda la retina era representada en la mitad restante. Esto sugiere que las conexiones en el tectum son susceptibles de modificación. Figura 415 (a) Hipótesis de la neuroespecificidad de las determinaciones de las conexiones neurales. Los lugares específicos de la superficie del receptor se representan mediante letras diferentes. Las conexiones en el encéfalo se establecen en los lugares con la misma letra (a con A, b con B, etc.). Cada posición see halla codificada por una sustancia química o un estado único. (b) Gradientes químicos como determinantes de las conexiones neurales. Aquí las conexiones no están definidas por una sustancia química específica única sino por gradientes de dos dimensiones (mostrados aquí por los gradientes negro y marrón). Todos los lugares pueden ser identificados por una posición sobre los dos gradientes. Los experimentos sobre la especificidad retino-tectal también revelan que hay un tipo de proceso de reconocimiento que puede implicar agentes químicos específicos. Los intentos de determinar los detalles de este proceso han dirigido la atención a las características moleculares del reconocimiento químico. Determinantes del crecimiento y el desarrollo del encéfalo La emergencia de la forma, disposición y conexiones del encéfalo en desarrollo está influida por muchos estados internos y externos. En el caso del desarrollo, como en el caso de algunos otros temas que consideraremos en capítulos posteriores, es útil pensar en determinantes directos y en influencias moduladoras. Un determinante directo (o factor intrínseco) es aquel que está implicado en los procesos básicos que producen o controlan un fenómeno. En el desarrollo del sistema nervioso, son (determinantes directos cierto:; genes y los procesos que controlan. Por ejemplo, algunos tipos de neuronas expresan información genética para formar espinas dendríticas, mientras que otros tipos de neuronas en el mismo organismo nunca forman espinas. Una influencia moduladora (o factor extrínseco) es aquella que puede facilitar o inhibir los procesos básicos pero que no los controla directamente. En el desarrollo del sistema nervioso, moduladores como la nutrición y la experiencia pueden influir en la velocidad y extensión del desarrollo. Por ejemplo, en neuronas que forman espinas dendríticas, los factores moduladores determinan cuantas espinas se formarán. La efectividad de las influencias moduladoras de,-)ende críticamente de la etapa del desarrollo en que se dan. Un estado bioquímico particular presente en la vida temprana de un embrión puede ejercer influencias muy distintas a las del mismo estado presente en el desarrollo fetal o las etapas postnatales tempranas. En esta sección discutiremos algunos ejemplos de factores intrínsecos y extrínsecos del desarrollo de la estructura del encéfalo. No haremos un listado exhaustivo, sino que describiremos solamente unos pocos fenómenos bien investigados. Determinantes genético: Los psicólogos han mostrado durante mucho tiempo el papel de la genética en una variedad de conductas de muchas especies animales. Más recientemente, la investigación ha comenzado a explorar el control genético de la anatomía y la fisiología del sistema nervioso como parte del programa de comprensión de la forma en que los genes influyen y controlan la conducta (Hall, Greenspan y Hams, 1982; Wimer y Wimer, 1985). Por supuesto, los genes no trabajan aisladamente. La mayoría cl-, este trabajo se verá como la interacción de instrucciones genéticas con otras influencias del desarrollo. Los procedimientos de cría selectiva se han empleado durante años por los científicos y los granjeros para obtener animales distintos. Estas técnicas también han sido usadas por los investigadores en genética de la conducta, los cuales han explorado cambios de conducta a través de generaciones. El empleo de estas técnicas con animales más simples ha comenzado a relacionar efectos genéticos sobre el sistema nervioso con efectos sobre la conducta. Bentley (1976) mostró que los cantos de llamada de los grillos tienen intrincados patrones que pueden ser manipulados por cría selectiva. Estos patrones de canto cambian de manera distinta en función del tipo particular de genes que sean introducidos mediante aparcamiento selectivo a lo largo de varias generaciones. Los registros de las neuronas del sistema nervioso de los grillos revelan que la variabilidad de los cantos controlada genéticamente está directamente relacionada con el impacto de genes en el cambio de las disposiciones de las redes neuronales. Una técnica de cría inusual es la que produce animales idénticos genéticamente llamados clones, cuyo empleo es conocido principalmente por las películas de ciencia ficción y terror. ¡Pero la vida imita la ficción! Los estudios de la genética del desarrollo del sistema nervioso emplear criaturas idénticas. Los investigadores desarrollan estos animales mediante reproducción asexual, con lo que toda la descendencia tiene los mismos genes. Empleando clones de saltamontes, Goodman (1979) comparó la uniformidad y la variabilidad en el crecimiento y el desarrollo de diferentes neuronas. Aunque la forma básica de las células mayores mostraba una uniformidad considerable, muchas neuronas de saltamontes clonados mostraban diferencias en las conexiones neuronales entre individuos «idénticos». Si la herencia es idéntica, ¿eso significa que las conexiones neurales han de ser idénticas? Para estudiar esta cuestión, los gemelos idénticos humanos o de cualquier otro mamífero no son sujetos muy útiles, ya que sus sistemas nerviosos son demasiado complejos y no podemos encontrar la misma célula para comparar en dos individuos. Algunos investigadores abordaron esta cuestión con un pequeño crustáceo, Daphnia, bien conocido por muchos poseedores de acuarios (Macagno, Lopresti y Levinthal, 1973). La hembra de Daphnia puede reproducirse sin ser fertilizada por el macho, produciendo líneas de crías hembra genéticamente idénticas (o clones). Además, la Daphnia tiene un número fijo de neuronas que pueden identificarse bajo el microscopio. El ojo contiene exactamente 176 neuronas sensoriales que establecen contactos sinápticos con exactamente 110 neuronas del ganglio óptico. Por otra parte, una determinada neurona sensorial hace contacto con sólo unas pocas neuronas específicas del ganglio. Sin embargo, el número exacto de sinapsis establecido entre una neurona sensorial particular y una neurona específica del ganglio puede variar en una proporción mayor de 3 a 1 de uno a otro individuo Daphnia dentro de un clon. Incluso entre los lados derecho e izquierdo del ojo de un individuo, donde están localizadas simétricamente neuronas «gemelas», una de ellas puede establecer más sinapsis que la otra. Así tanto dentro de un individuo como entre individuos Daphnia, las neuronas con exactamente la misma herencia difieren en el número de sus conexiones sinápticas. De forma similar, la forma de las ramificaciones del axón difiere en un mismo sujeto y con otros sujetos Daphnia clonados, como se muestra en la Figura 4-16. En los vertebrados es más difícil encontrar neuronas idénticas para comparar conexiones sinápticas. Los investigadores que estudiaron esta cuestión en Daphnia lo intentaron con un pez que se reproduce partenogénicamente, como Daphnia, produciendo hijas que son idénticas genéticamente entre sí y con su madre. En este pez el sistema nervioso es complejo, pero cada animal tiene una sola célula de Mauthner gigante en cada lado del encéfalo. El examen microscópico mostró que, aunque el patrón de ramificación dendrítica de la célula de Mauthner es similar de individuo a individuo en un clon, hay diferencias individuales en el detalle de la ramificación y de las sinapsis. Así, el descubrimiento en Daphnia podría extenderse al menos a las células de Mauthner del encéfalo del pez (Levinthal, Macagno y Levinthal, 1976). Entre los mamíferos genéticamente idénticos, las diferencias en el sistema nervioso son incluso mayores. Así, en cepas altamente endógamas de ratón, donde todos los individuos del mismo sexo son esencialmente idénticos genéticamente, una región específica del encéfalo (como la zona del hipocampo) muestra un pequeño porcentaje (le diferencias entre los individuos en el número de neuronas que llega a tener (Wimer et al., 1976). Se han hecho observaciones indirectas de este tipo en gemelos idénticos humanos, aunque tales gemelos a menudo difieren en tamaño en el nacimiento, y sus encéfalos difieren al menos tanto como lo hacen los de los ratones endógamos. Figura 4-16 La misma neurona en cuatro gemelos idénticos de un clon de insectos. En las dos columnas se muestran dos ejemplos de patrón de ramificación. El patrón de variabilidad es mayor desde un animal genéticamente idéntico a otro, que entre el del lado izquierdo y el lado derecho en un mismo individuo. (Macagno et al., 1973.) Algunas evidencias indirectas indican que para los gemelos idénticoshumanos el patrón de ramificación de las terminaciones nerviosas de la piel debe diferir. La evidencia está en que incluso los Í gemelos idénticos tienen diferencias en sus huellas dactilares, aunque sus huellas son más similares que entre gemelos fraternos. La piel de las yemas de les dedos está ricamente inervada, con lo que las diferencias en el patrón de la:; crestas de la piel debe significar diferencias en la distribución de las terminaciones nerviosas. Además, las pequeñas glándulas sudoríparas de las yemas de los dedos tienen sus aberturas a lo largo de las crestas de la piel. Las glándulas sudoríparas están unidas a terminaciones nerviosas que controlan la secreción de sudor, por lo que de nuevo las diferencias en el patrón de crestas deben implicar diferencias en las localizaciones de las terminaciones nerviosas que van a las glándulas sudoríparas. Este ejemplo indica que entre los seres humanos, como entre otros animales, la identidad de la herencia no significa identidad en cada detalle del sistema nervioso. Quisiéramos advertir que el patrón de huellas dactilares se forma durante el cuarto mes de embarazo, con lo que las diferencias individuales están bien determinadas antes del nacimiento. Las diferencias ulteriores entre los sistemas nerviosos de los gemelos idénticos pueden deberse a respuestas a experiencias diferentes, como se verá en los Capítulos 16 y 17 cuando consideremos los erectos de la experiencia y el aprendizaje en la anatomía del sistema nervioso. Mutantes genéticos A veces la naturaleza, con la ayuda de los investigadores produce animales inusuales que muestran un cambio repentino en la estructura genética, una mutación que está relacionada con marcados cambios anatómicos o fisiológicos. Los mutantes -animales que presentan estos cambios- son interesantes de estudiar debido a que sus características genéticas repentinamente cambiadas pueden ser bastante específicas y llamativas. Esto nos proporciona evidencia de los controles genéticos de su desarrollo que son más sutiles en otros animales. Por ejemplo, Greenspan y Quinn (1984) describieron mutantes de la mosca de la fruta Drosophila que tenían problemas d, memoria. Estos mutantes -afectivamente llamadlos «Zopenco», «Amnésico» y «Nabo»--- fracasaban en el aprendizaje o podían aprender pero olvidaban rápidamente-. La evidencia reciente apunta a que déficit bioquímicos en estos mutantes podrían dar cuenta de los fallos de la memoria (Dudai, 1988). Discutiremos estos estudios en el Capítulo 17 cuando analicemos los mecanismos neurales del aprendizaje y la memoria. Muchos mutantes de Drosophila tienen defectos muy específicos en alguna parte del sistema nervioso (Hall y Greenspan, 1979). El importante número de investigaciones sobre los mutantes de Drosophila deriva de la abundancia de mutaciones específicas, cada una ele ellas implicando cl deterioro de un proceso del desarrollo distinto. Por ejemplo, un mutante letal de Drosophila, «Mellado», tiene un sistema nervioso agrandado a causa del exceso de producción de células precursoras. Los estudios de este mutante pueden capacitar a los investigadores para obtener un mejor conocimiento de los procesos que controlan el número de células producidas durante el desarrollo embriológico temprano. Aproximadamente 150 mutaciones del ratón implican al sistema nervioso (Sidman, Green y Appel, 1965). En estos animales los defectos específicos aparecen durante el desarrollo del sistema nervioso. En algunos ratones no crecen regiones particulares del encéfalo. Otros muestran desajustes anatómicos específicos, como alguna característica de la mielinización o de la disposición de las células en sus alineaciones típicas. Los mutantes de un grupo, especialmente intrigante para los investigadores, tienen todos trastornos debidos a genes simples que afectan el desarrollo postnatal del cerebelo. Los nombres de estos animales -Vacilante, Tambaleante y Tejedor-- reflejan el deterioro motor que los caracteriza. El impacto de estos genes en el tamaño y disposición del cerebelo se ilustra en la Figura. 4-17. El cerebelo de Vacilante muestra una disposición anormal de las células. No hay capas Figura 4-17 Mutaciones cerebelosas del ratón. La columna izquierda muestra cortes del cerebelo en un ratón normal a tres niveles de magnificación (x25, x66, y x250). La columna media muestra visiones comparables en el mutante Tejedor. Adviértase la ausencia casi completa de células granulares, mientras que el alineamiento de las células de Purkinje (flechas) es normal. La columna de la derecha muestra secciones del mutante cerebelar Vacilante. Es evidente el marcado desarreglo de la estratificación acostumbrada de células. Ambos mutantes muestran una reducción global del cerebelo (Leiman). características en el cerebelo, hipocampo y corteza cerebral. Sorprendentemente, aunque las células de estas regiones están en posiciones anormales, muchas de sus conexiones son las apropiadas (Cavincss, 1980). El cerebelo de Tejedor tenía muchas menos células granulares que un cerebelo normal, lo que podría originarse del fallo de estas células para migrar o formar conexiones adecuadas. La atrofia del cerebelo también es evidente en Tambaleante, de acuerdo con Sotelo (1980), el cual mostró que este animal era inca paz de formar conexiones sinápticas entre las células granulares y las células de Purkinje. El axón de las células granulares -las fibras paralelas- llega cerca de la superficie dendrítica de la célula de Purkinje, pero las especializaciones postsinápticas simplemente no se desarrollan. Cada uno de estos mutantes de ratón muestran alteración debida a un único gen, relacionado con el desarrollo de un tipo específico de célula. Los estudios de estos animales nos ofrecen un conocimiento más profundo de los procesos del desarrollo neuronal y de sus consecuencias conductuales. Influencias bioquímicas El encéfalo se compone de muchos grupos celulares distintos que se desarrollan en momentos diferentes. Las reglas que orquestan la emergencia de esta compleja estructura son indudablemente elaboradas. Sin embargo, una noción que comparten todos los investigadores es que el proceso lo regulan varias sustancias corporales. Esta sección proporciona ejemplos de dos tipos de condiciones bioquímicas que regulan el crecimiento neural. Un ejemplo de control intrínseco es el factor de crecimiento nervioso (FCN), una sustancia que parece controlar el desarrollo de una clase particular de células nerviosas. Un ejemplo de influencia bioquímica extrínseca es el papel de la nutrición en el crecimiento del encéfalo. Factor de crecimiento nervioso Hace más de veinte años, los investigadores descubrieron una sustancia que afecta marcadamente al crecimiento de las neuronas de los ganglios espinales y de los ganglios del sistema nervioso simpático (Levi-Montalcini, 1982). Esta sustancia es el factor de crecimiento nervioso. Su descubrimiento les valió a Levi-Montalcini y Cohen el Premio Nobel en 1986. Originalmente, el FCN se encontró en una variedad de lugares inusuales, incluyendo las glándulas salivares del ratón, ciertos tumores de la piel y el veneno de la serpiente. Más recientemente, las precisas técnicas bioquímicas han revelado su presencia en el sistema nervioso. Los investigadores establecieron que si administraban FCN a un feto animal, se obtenía una formación de ganglios simpáticos con muchas mas células de lo usual. Estas células eran también mayores y poseían muchos procesos (Figura 4-18). Si lo administraban postnatalmente, el FCN producía un agrandamiento de las células del sistema nervioso simpático. Más recientemente se ha visto que el factor de crecimiento nervioso puede invertir los efectos degenerativos de una droga que destruye selectivamente las células del encéfalo que contienen determinados transmisores sinápticos. Parte del interés en el FCN surge de la posibilidad de que sea un ejemplo de mecanismo de control en el desarrollo del sistema nervioso. Puede haber muchas Figura. 4-18 Efectos del factor de crecimiento nervioso (FCN). La figura superior muestra un ganglio espinal creciendo aislado en el exterior del cuerpo (in vitro) sin la presencia de FCN. La fotografía inferior es de un ganglio espinal creciendo bajo circunstancias similares pero con FCN añadido a la solución en que está bañado. Esta figura muestra una marcada proliferación de procesos axonales que parten en todas las direcciones. (De R. Levi-Montalcini, Science, vol. 143 [enero de 1964]: 105-110, Figuras 1, 2 y 10. Copyright (o 1964 de la Asociación Americana. para el Progreso de Ciencia.) sustancias, cada una controlando un tipo particular de célula en un período del desarrollo específico. Nutrición, crecimiento y desarrollo del encéfalo No todo el mundo comparte la bueno suerte de tener una nutrición adecuada. El hambre periódica afecta a muchos, y este problema se hace más urgente según el crecimiento de la población amenaza con colmar las fuentes de alimentos de muchas naciones. Durante muchos años se ha creído que el encéfalo era menos susceptible a los efectos de la dieta que otras partes del cuerpo. Ciertamente, es verdad que el encéfalo adulto se ve mucho menos afectado por la dieta o la sobrealimentación de lo que lo son la mayoría de los otros órganos. Pero hoy en día existe evidencia de que la malnutrición deteriora el encéfalo, especialmente durante el desarrollo temprano. De hecho, varias formas de malnutrición que se dan durante los períodos críticos del desarrollo del encéfalo en humanos y otros animales pueden producir cambios irreversibles en la estructura del encéfalo (Winick, 1976). Relacionar estos cambios encefálicos con la conducta es una tarea complicada. Es duro desenmarañar los efectos de las desventajas sociales de los efectos de las deficiencias dietéticas, ya que la mayoría de estos estudios implican a madres e hijos que viven en circunstancias empobrecidas (Balderston et al., 1981). Hemos aprendido algo de los efectos de la malnutrición temprana por los estudios que comparaban niños os desnutridos con parejas control que no tenían deficiencia nutricional temprana. Los estudios en México, Chile, Yugoslavia y Sudáfrica han mostrado que la desnutrición reduce posteriormente la ejecución en muchos tipos de tests de capacidad mental (Tizard, 1974). La desnutrición se da más comúnmente en familias que viven en una clase de pobreza que también supone otras potentes barreras al desarrollo del niño. Sin embargo, muchos de estos estudios muestran que los posteriores deterioros conductuales dependen de la época de la vida en que el niño estuvo en un período de malnutrición. Los niños tienen una mayor capacidad de recuperación conductual si la malnutrición se da más tarde en la vida que si se da pronto. Una amplia gama de rehabilitaciones nutritivas y conductuales pueden contrarrestar los efectos de severas malnutriciones tempranas, especialmente si comienzan a la edad de dos anos :y son mantenidos en la adolescencia (Nguyen, Meyer y Winick, 1977; Winick, Meyer y Harris, 1975). Esta investigación se hizo con huérfanos coreanos que fueron adoptados por familias americanas de clase media. Todos los niños provenían de orfanatos y eran menores de cinco años cuando fueron adoptados. El estudio se hizo retrospectivamente; esto es, estaba basado en registros disponibles cuando los niños eran adolescentes. Los niños se dividían en tres grupos de acuerdo a sus alturas a la edad de admisión en la agencia: 1. Desnutridos severos, más allá del tercer percentil (de acuerdo con las normas coreanas). 2. Desnutridos moderados, de los percentiles tercero a vigésimo cuarto. 3. Bien nutridos, en el vigésimo quinto percentil o por encima. Los grupos desnutridos se desarrollaron bien en sus familias adoptivas, llegando todos a exceder las normas coreanas de peso y altura, aunque no alcanzaron las normas americanas. Respecto al CI y el análisis de los logros escolares, la media de los tres grupos adoptados a la edad de dos años, después excedía las medias americanas (Figura 4-19). Entre aquellos que se adoptaron después de los dos años, los niños del grupo 1 (los que sufrían severa desnutrición precoz) no alcanzaban suficientemente las normas americanas, pero los niños de los otros dos grupos sí lo hacían. Aunque Figura 4-19 Efectos sobre el crecimiento, la inteligencia y los logros, de la adopción temprana y tardía de niños desnutridos. Los grupos de nutrición: 1, desnutridos severos a su admisión en la agencia de adopción; 2, desnutridos moderados, y 3, bien nutridos. (Los resultados de la adopción temprana proceden de Winick, Meyer y Harris, 1975; los resultados de la adopción tardía provienen de Nguyen, Meyer y Winick, 1977.) Figura 4-20 Tasa de desarrollo del encéfalo en relación con el nacimiento. La escala de tiempo es distinta para diversos animales, oscilando entre días y meses. Esta figura muestra que los períodos máximos de desarrollo del encéfalo son bastante diferentes para animales distintos. La rata, por ejemplo, muestra principalmente un incremento postnatal del peso del encéfalo. En contraste, el principal desarrollo del encéfalo del cobaya se produce antes del nacimiento. (Adaptado de Dobbing, 1972.) persistían algunas diferencias relacionadas con la desnutrición precoz, estas diferencias eran más bien pequeñas. Este es un estudio importante debido a que demuestra que los efectos de varias desnutriciones precoces pueden ser superados si la rehabilitación comienza pronto y es mantenida. Figura 4-21 Ilustración esquemática de diferentes tipos de efectos de la experiencia sobre el desarrollo del encéfalo (a) versus (b). La experiencia puede inducir cambios, aquí vemos el crecimiento de un grupo de terminales axónicos. (c) versus (d). La experiencia puede modular el desarrollo, incrementando aquí el crecimiento para alcanzar antes la meseta. (e) versus (f). La experiencia puede mantener el crecimiento. Aquí las terminaciones disminuyen a menos que haya experiencia. En los estudios neuroanatómicos y neuroquímicos se ha enfatizado la importancia crítica de la desnutrición precoz con respecto al deterioro posterior de las capacidades mentales. Dobbing (1976) remarcó el hecho de que el encéfalo es lo más vulnerable a la desnutrición durante el período de rápido crecimiento encefálico. Estos períodos varían para animales distintos (Figura 4-20). En humanos el período de crecimiento más rápido del encéfalo y, de acuerdo con Dobbing, de máxima vulnerabilidad a la malnutrición se da en la última fase del embarazo y los primeros meses de vida postnatal. Una malnutrición similar en el adulto produce efectos despreciables. Los experimentos con animales han mostrado algunos efectos permanentes de la desnutrición durante los períodos tempranos de crecimiento rápido del encéfalo. Se ven afectados cl tamaño y el peso del cuerpo así como la estructura del encéfalo y la conducta (Dobbing, 1974). Por ejemplo, el tamaño del cerebelo en las ratas es especialmente sensible a la desnutrición postnatal, sensorial iniciada poco después del nacimiento y continuada durante varias semanas puede producir la atrofia de algunas de las células en desarrollo: «si no lo utilizas, lo pierdes». Privación visual y falta de uso Hay gente que no ve claramente las formas con un ojo, incluso si está sano y la imagen de la forma se enfoca en la retina. Este deterioro de la visión se conoce como ambliopía (de las palabras griegas para «sombrío» o «embotado» y para «visión»). Se puede observar un ejemplo de este trastorno en la gente con un «ojo perezoso»: un ojo vuelto hacia dentro o hacia fuera (ojo cruzado). Algunos niños nacen con este tipo de mal alineamiento de los ojos. Estos niños «ven doble» en vez de ver una imagen fundida. Si el ojo desviado no se alinea quirúrgicamente antes de que la persona alcance la infancia tardía, se deteriora la visión. En el momento en que la persona alcanza la edad adulta, hay una supresión virtualmente total del patrón de visión del ojo Figura 4-22 Desarrollo de la corteza visual del gato. El desarrollo sináptico se produce más intensamente desde los días 8 al 37 después del nacimiento, un período durante el cual el uso puede tener profundas influencias. Adviértase también que el peso del encéfalo y el volumen celular aumentan de forma paralela y preceden al desarrollo sináptico. (Adaptado de Cragg, 1975.) ha sugerido que durante el desarrollo temprano de una persona, los axones que representan inputs de cada ojo «compiten» por los lugares sinápticos. Las sinapsis activas, utilizadas, llegan a ser conexiones efectivas y predominan sobre las sinapsis inactivas, que no se han empleado. Los investigadores también proponen una explicación como esta para dar cuenta de la ambliopía producida por desalineamiento de los ojos. Se produjo un animal réplica de esta condición humana cortando los músculos de un lado del ojo de gatos jóvenes (Hubel y Wiesel, 1965). El histograma de dominancia ocular de estos animales revela que las células de la corteza visual muestran una sensibilidad binocular enormemente reducida. Hay una proporción mucho mayor de excitación por estimulación del ojo darecho o el izquierdo de la que se da en los animales control. Este efecto se debe a que después de la intervención, las células de la corteza visual no reciben inputs sincrónicos de ambos ojos. Figura 4-23 (a) Histograma de dominancia ocular de células de la corteza visual de gatos adultos normales. (b) Histograma de dominancia ocular después de la no alineación ocular temprana; esto es, estrabismo. (c) Histograma de dominancia ocular que sigue a la privación monocular durante el periodo critico temprano. (d) Histograma de dominancia ocular que sigue a la privación binocular. (Adaptado de Hubel y Wiesel, 1965; Wiesel y Hubel, 1965.) Exposición tempranaa a patrones visual Al nacimiento la corteza visual es bastante inmadura, y la mayoría de las sinapsis aún no se han formado. Esto da lugar a la cuestión de si la experiencia temprana afecta al desarrollo de la corteza visual. La evidencia citada en la sección previa muestra que la ausencia de utilización persistente da lugar a cambios en la estructura y la respuesta de las vías visuales. La modificabilidad del encéfalo en desarrollo también se evidencia cuando los animales se exponen a ciertos patrones durante el desarrollo temprano. Los experimentos en que los patrones visuales se manipulan de forma temprana en la vida del animal hará empleado patrones como líneas horizontales o verticales (Blakemore, 1976), un campo de estas líneas vistas a través de anteojos (Hirsch y Spinelli, 1971) o pequeños puntos de luz (Pettigrew y Freeman, 1973). Este es un campo muy controvertido. Algunos grupos de investigación indican resultados que difieren de los de otros. Sin embargo, el peso de los resultados más frecuentes sugiere que estas diversas experiencias visuales durante los períodos de vida críticos tempranos, modifican las respuestas de las células nerviosas en la corteza visual. El período sensible para estos efectos es el mismo que para producir efectos de privación monocular. De acuerdo con una revisión detallada (Movshon y Van Sluyters, 1981), la variedad de experimentos y resultados de este área no puede proporcionar una conclusión simple o uniforme. Varios resultados apoyan la hipótesis de que la estimulación sensorial es necesaria para inducir el desarrollo del sistema visual, modular el mantenimiento del desarrollo o mantener el desarrollo que está programado genéticamente. Parece además que la experiencia puede jugar cada uno de los tres papeles hipotéticos en el desarrollo neural. Experiencias no visuales Los efectos de las experiencias tempranas en el encéfalo pueden ser producidos también mediante la manipulación de inputs sensoriales no visuales -los pelos del bigote de una rata, por ejemplo-. Thomas Woolsey y colaboradores (1976, 1981) encontraron un agrupamiento único che células nerviosas en una región de la corteza cerebral do la rata que recibía inputs desde las vibrisas (bigotes). La disposición de estos pelos sobre la piel es característica. Los pelos se hallan alineados de forma similar en todos los animales de la misma especie (ver Figura 4-24). En la región de la corteza donde están representadas las vibrisas, Woolsey observó agrupaciones de células que llamó barriles, debido a que la forma en que estaban dispuestas las hacia parecer paredes de un barril. La Figura 4-24 muestra también que la distribución de estos barriles corticales corresponde al mapa de las vibrisas. Si se secciona alguna vibrisa entre uno y cuatro días después del nacimiento, sus barriles corticales no se desarrollan. Sin embargo, los barriles que representan vibrisas adyacentes intactas tienden a agrandarse. La manipulación de la capacidad de un animal para oler también afecta a su encéfalo durante las etapas del desarrollo. Los estudios de Meisami (1978) mostraron que los dos orificios nasales de las ratas son relativamente independientes, con interconexiones evidentes sólo en la faringe. Cuando se bloquea un orificio de forma temprana, Ia rata puede respire r, pero la mucosa olfatoria -la superficie sensorial nasal- - no es estimulada e el lado ocluido. Después de someter a la rata a esta oclusión durante algunas semanas, Meisami comparó el crecimiento de los bulbos olfatorios de las ratas -la región del encéfalo que recibe inputs de los receptores sensoriales nasales. Había una mareada diferencia en tamaño entre los bulbos que recibían inputs de los orificios tapados y de los normales. El bulbo olfatorio conectado al orificio obstruido mostraba atrofia. Hemos dado sólo unos pocos ejemplos de los muchos experimentos que muestran cómo los estímulos sensoriales influyen en el desarrollo de la estructura y la función del encéfalo. Los efectos difieren en función de variables como la edad del sujeto, la duración de la experiencia y la estimulación dada. Envejecimiento del encéfalo El paso del tiempo nos proporciona una acumulación de alegrías y penas -quizás riqueza y fama- y un declinamiento progresivo de muchas de nuestras capacidades. Los cambios con la edad parecen ser inevitables en los sistemas biológicos. Vamos a revisar primero algunas de las características del envejecimiento normal. Veremos después algunas de las exageraciones patológicas del proceso de envejecimiento, especialmente la enfermedad de Alzheimer. Envejecimiento normal Muchos aspectos estructurales funcionales cambian a lo largo del ciclo de la vida humana. Aunque respondiendo más lentamente, parece inevitable que con la edad, muchas de nuestras capacidades cognitivas muestren pequeños cambios a través del período adulto hasta que alcanzamos una edad avanzada. ¿Qué le ocurre a la estructura del encéfalo des-.le la adolescencia hasta el día en que tenemos un pequeño olvido y caminamos más indecisamente? ¿La estructura del encéfalo cambia constantemente a lo largo del ciclo vital de cualquier animal? Los datos de las autopsias humanas nos dan algunas pistas sobre cómo cambia el encéfalo progresivamente durante la edad adulta. Los cambios en la estructura del encéfalo que acompañan al envejecimiento pueden verse a diferentes niveles, desde las estructuras subcelulares hasta la morfología global del encéfalo. Las diferencias en el peso del encéfalo se han examinado a menudo en relación con el envejecimiento. Durante' años se ha cuestionado la relevancia del envejecimiento en estos cambios de peso, ya que es difícil distinguir los cambios debidos al envejecimiento de los cambios que se originan de estados enfermizos que llevan rápidamente a la muerte. Un excelente estudio reciente eliminó estos factores & confusión (recordar la Figura 4-1). Los cambios son muy pequeños hasta la edad de 45 años, después de la cual el peso del encéfalo comienza a disminuir significativamente. El peso del encéfalo de los ancianos humanos es el 7-8% menor que el máximo peso del adulto (Creasy y Rapaport, 1985). El curso de estos cambios es el mismo para hombres y mujeres, incluso aunque las mujeres vivan generalmente de siete a diez años más que los hombres. Los datos también enfatizan que el envejecimiento es un estado variable. El declive es evidente en todas Figura 4-24 Localización de los pelos del bigote y los barriles corticales en ratones jóvenes. A la derecha se muestra la disposición de los barriles en la corteza somatosensorial. (a) Cada barril recibe su input desde un único pelo del lado opuesto del hocico del ratón. (b), (c). Si se destruye una línea de pelos poco después del nacimiento (como se indica por los puntos marrones), se verá que la correspondiente línea de barriles de la corteza desaparece y los barriles adyacentes se agrandan. (d) Si se destruyen todos los pelos, desaparecerá el grupo entero de barriles. La ilustración está basada en el trabajo de Thomas A. Woolsey de la Escuela Universitaria de Medicina de Washington. (De W. M. Cowan, The Development of the Brain. Copyright (o 1979 por Scientific American, Inc. Todos los derechos reservados.) las personas, pero es exagerada en algunas. A algunos invetigadores esto les sirve para enfatizar la contribución genética del envejecimiento y refuerzan la idea de que si se quiere vivir mucho hay que buscar padres y abuelos que hayan vivido mucho. Frecuentemente se observa que los pliegues de la corteza cerebral de los encéfalos de personas mayores se han atrofiado y que los ventrículos laterales se han agrandado. Estos cambios son mareados, sin embargo, sólo en los casos severos de la enfermedad de Alzheimer, lo que discutiremos brevemente. Una medida común de la estructura empleada en los estudios del envejecimiento encefálico es el número de células neurales y filiales en volúmenes concretos de tejido. Los investigadores toman regiones específicas y cuentan el número de células de varias áreas, empleando tejido procedente de personas que han muerto a diferentes edades. Estos estudios sugieren que los cambios celulares comienzan en la tercera década y son específicos de regiones particulares. Más importante ineluso que la reducción del número de células es la pérdida de conexiones sinápticas, lo que es especialmente relevante en las regiones frontales. Los barridos de TEP de personas mayores añaden una nueva perspectiva a los cambios del envejecimiento. [,os estudios de ancianos normales revelan que el metabolismo encefálico permanece casi constante. Esto está en marcado contraste con la disminución del metabolismo encefálico en la enfermedad de Alzheimer. Emplearemos dos regiones del sistema motor piara mostrar lo diferentes que pueden ser los efectos del envejecimiento. En la corteza motora un tipo de neurona grande -la célula de Betz- comienza a cambiar sobre los 50 años, y cuando la persona alcanza los 80, muchas de estas células virtualmente han degenerado del todo (Scheibel, Tomiyasu y Sehoibel, 1977). En contraste, otras células implicadas en el circuito motor -aquellas del área del tronco encefálico llamada oliva inferiorpermanecen iguales en número durante al menos ocho décadas de vida. En los sistemas nerviosos jóvenes, las lesiones de muchas partes del encéfalo y la médula espinal, inducen el nuevo crecimiento de los axones y la formación de nuevas conexiones (Capítulo 16). Pero en los adultos, aunque también se observa la regeneración axonal, ésta es mucho menos vigorosa. Además, varios investigadores (por ejemplo, Scheff, Bernardo y Cotman, 1978) han mostrado que los encéfalos de ratas viejas son mucho menos capaces de desarrollar colaterales axónicas después de lesiones ele un tracto del encéfalo. Así, parece que los encéfalos de animales viejos son menos capaces que los encéfalos de los jóvenes para compensar anatómicamente la reducción progresiva de células y sinapsis. Enfermedad de Alzheimer: una exageración patológica del envejecimiento Desde principios de este siglo, la población de 65 años de edad en los Estados Unidos se ha multiplicado por ocho. En el año 2000 habrá al menos 30 millones de personas en este grupo de edad. La mayoría de las personas que alcanzan esta edad llega a ella felizmente, con vidas productivas, aunque a un paso más lento del que los caracterizaba años atrás. Sin embargo, se hace sorprendentemente claro que hay un número creciente de personas mayores a los que la edad les trae una agonía particular, el trastorno llamado enfermedad de Alzheimer, después de que un neurólogo describiese por primera vez un tipo de demencia que aparecía antes de los 65 años. Hoy se considera que este trastorno es el mismo que otra forma de demencia que aparece de manera más tardía en la vida, la demencia senil. Hoy en día, al menos casi 2 millones de americanos de 65 años de edad sufren de enfermedad de Alzheimer, y el progresivo envejecimiento de nuestra población significa que este número de afectados será mayor en los próximos 20 a 40 años. La enfermedad de Alzheimer se caracteriza por una disminución progresiva del funcionamiento intelectual. Comienza como una pérdida de memoria de los acontecimientos recientes. Eventualmente este deterioro de memoria llega a abarcarlo todo, es tan extensivo que los pacientes de Alzheimer no pueden mantener ninguna forma de conversación, ya que olvidan rápidamente el contexto y la información previa. Preguntas simples -«¿en qué año estamos?», «¿quién es el presidente de los Estados Unidos?» o ¿dónde está usted ahora?»- se hacen imposibles de responder. El declinar cognitivo es progresivo y ralentizado. Con el tiempo, los pacientes se desorientan y se pierden fácilmente en entornos familiares. Un manual reciente, «El día de treinta y seis horas» es un comentario sobre los graves problemas de asistencia a los que han de enfrentarse las familias de pacientes de Alzheimer. Las observaciones del conjunto del encéfalo de los pacientes revelan una extraordinaria atrofia cortícal especialmente evidente en las áreas frontal, temporal y parietal. Los estudios del metabolismo del encéfalo de estos pacientes son especialmente reveladores. Los barridos con TEP que siguen a la administración de glucosa marcada radiactivamente, muestra una marcada reducción del metabolismo oxidativo en la corteza parietal posterior y algunas porciones del lóbulo temporal. Una reducción de la utilización de glucosa precede a la aparición de deterioros cognitivos más severos (Foster et al., 1984). Los estudios microscópicos de los encéfalos de los pacientes de Alzheimer revelan un grupo de cambios celulares característicos. Algunas células muestran anormalidades de los neurofilamentos, denominados ovillos neurofibrilares. Se trata de espirales anormales de filamentos que forman una disposición enredada en la célula (Figura 4-25b). Los estudios histológicos también muestran terminales axónicas en degeneración, llamadas placas seniles (Figura 4-25c). Cada placa contiene; en su interior una sustancia llamada amiloide, compuesta de una proteína inusual. El número de placas seniles se relaciona directamente con la magnitud del deterioro cognitivo. Los investigadores también han advertido estos cambios celulares, típicos de la enfermedad de Alzheimer, en los encéfalos de pacientes de síndrome de Down. Recientemente los investigadores se han centrado en los cambios de un grupo de células del encéfalo anterior que pueden resultar una clave para la comprensión fundamental de la enfermedad de Alzheimer. En varios estudios anatómicos se ha descrito una extraordinaria pérdida de neuronas en una región subcortical denominada núcleo basal de Meynert (Figura 4-25a). Los axones de estas células se extienden a Figura 4-25 (a) Localización de los núcleos basales del encéfalo anterior y distribución de los axones colinérgicos. (b) Ovillos neurofibrilares vistos en una sección transversal de la corteza cerebral de una persona mayor. Se señala un ejemplo con la flecha. (c) Placas seniles de la corteza cerebral de un paciente mayor. (Fotografías cortesía de F. J. Seil.) muchas regiones corticales y las células contienen acetilcolina, una sustancia empleada para la transmisión de la actividad neural a otras células (discutido en el Capítulo 6). Las deficiencias del sistema de acetilcolina son características de los encéfalos de pacientes ;en la enfermedad de Alzheimer, pudiendo estar implicados otros neurotransmisores químicos. Los casos reportados por Coyle, Price y Delong (1983) mostraron una disminución consistente y marcada en el número de células de esta región, lo que no se apreciaba en áreas del encéfalo inmediatamente adyacentes. Esto no es simplemente un cambio exagerado que se ve rutinariamente en el envejecimiento, ya que los individuos que envejecen normalmente muestran poca pérdida de neuronas en esta área según se van haciendo mayores (Chui et al., 1984). Las causas de la enfermedad de Alzheimer permanecen cubiertas de misterio. En algunos pacientes es evidente un factor familiar, especialmente en aquellos que muestran un inicio temprano de demencia (que ce comienza entre los 40 y los 60 años de edad). Un informe reciente documentó la presencia de la enfermedad de Alzheimer en 52 miembros de una familia seguida durante varias generaciones. El estudio de la familia sugirió una forma dominante de herencia de la enfermedad de Alzheimer (Nee et al., 1983). Sin embargo, los factores hereditarios no son lo suficientemente evidentes en la gran mayoría de casos. Algunos investigadores se han centrado en la posibilidad de que el trastorno implique un agente transmisible, como un virus o una partícula subviral (Price, Whitehouse y Struble, 1985). Esta noción está fundamentada por los datos de investigación que muestran el papel de virus inusuales en enfermedades degenerativas del encéfalo tales como el kuru. Puesto que las sales de aluminio localizadas en la corteza cerebral producen ovillos neurofibrilares en animales experimentales, algunos investigadores han sugerido que las concentraciones tóxicas del metal pueden ser responsables de la enfermedad de Alzheimer. La investigación del aluminio también se inspira en el descubrimiento de concentraciones relativamente grandes de aluminio en los encéfalos de los pacientes de Alzheimer. Sin embargo, este cambio puede originarse como una consecuencia del trastorno más bien que como una causa de él. Otra hipótesis sobre el origen de la enfermedad de Alzheimer enfatiza el fenómeno autoinmune. De acuerdo con esta perspectiva, podrían originarse en el cuerpo del paciente anticuerpos que atacarían selectivamente a neuronas que contuviesen acetilcolina. Una excitante sugerencia reciente enfatiza que el factor de crecimiento nervioso es esencial para la supervivencia de las células que contienen acetilcolina. Las deficiencias en FCN pueden ser la causa primaria de la enfermedad de Alzheimer, de acuerdo con Hefti y Weiner (1986). La investigación intensiva se centra ahora en las causas y el tratamiento de la enfermedad. El envejecimiento de nuestra población sugiere que no hay que dejar pasar mucho tiempo antes de que nos enfrentemos a un enorme problema de salud pública. Se ha sugerido como terapia para ésta y otras enfermedades degenerativas del encéfalo el reemplazar las células del encéfalo perdidas mediante trasplantes; ver Cuadro 4-2. Desarrollo anómalo del encéfalo y alteraciones de la conducta El hecho de que los procesos que guían el desarrollo del encéfalo humano sean tan numerosos, intrincados y complejos también significa, desafortunadamente, que hay muchas formas en que pueden darse erróneamente. Los numerosos factores que CUADRO 4-2 Trasplantes o injertos de encéfalo: ¿una ayuda para el futuro? La investigación en cirugía del sistema nervioso a veces hace que el presente parezca el futuro. Hemos crecido acostumbrados a los trasplantes de corazón, cambios de riñón, injertos de córnea y operaciones similares. Pero ¿que hay de los trasplantes encefálicos? Los periodistas preguntaron una vez a Christian Barnard, el primer cirujano que trasplantó un corazón humano (en 1967), que pensaba sobre un trasplante encefálico, El apuntó todas las imponentes dificultades técnicas: conexiones de axones, vasos sanguíneos, nervios y todo lo demás. Entonces pareció retroceder ante la verdadera idea al advertir que tal operación sería llamada en realidad un trasplante de cuerpo. (Las transformaciones de las cabezas cambiadas en el Capítulo 1.) Poco tiempo atrás los trasplantes encefálicos o de cuerpo eran irreales, materia de la ciencia ficción. Sin embargo, los límites de lo real se han extendido un poco más allá, con las demostraciones del aislamiento exitoso de la totalidad del encéfalo de un chimpancé por White (1976). Este investigador fue capaz de mantener un encéfalo aislado durante al menos uno o dos días mediante su conexión a máquinas que le proporcionaban oxígeno y nutrientes en la circulación. De forma más inmediata, acontecimientos menos quijotescos para humanos, provienen de trabajos a menor escala, trasplantes de pequeñas porciones de encéfalo como injertos. ¿Puede extraerse una parte del encéfalo de un animal y donarla a un segundo animal? Esta perspectiva es particularmente importante para la posible compensación de trastornos del encéfalo que impliquen deficiencias de sustancias químicas específicas generadas en ciertas regiones del encéfalo. Este campo se ha desarrollado muy rápidamente. El trabajo experimental de los años ochenta muestra claramente que los trasplantes encefálicos son factibles y que el tejido añadido llega a formar parte de los circuitos del encéfalo receptor (Sladek y Gásh, 1984). De hecho estamos aunando esfuerzos sistemáticos para reemplazar áreas degeneradas del encéfalo de humanos mediante técnicas de injerto, como muestran algunos ejemplos de la literatura de investigación animal general. Estos estudios no sólo muestran que las nuevas células llegan a ser parte del «cableado» del encéfalo huesped: los trasplantes de encéfalo en varios estudios animales pueden también corregir funciones deterioradas producidas por lesiones encefálicas. La investigación muestra que de algún modo el tejido del encéfalo es mucho más fácil de trasplantar que muchos otro:; tejidos corporales, ya que el tejido nervioso es menos probable que sea rechazado por la acción del sistema inmunológico. La mayoría de los estudias de trasplantes encefálicos han implicado la inserción de una pequeña parte de tejido dentro de una cavidad del encéfalo, como el ventrículo o la superficie del cerebro. El tejido donante proviene de encéfalos de embriones o fetos animales. Algunas técnicas de trasplante más recientes han implicado la inyección de células nerviosas embrionarias disociadas, en las regiones encefálicas más profundas. Esta técnica de inyección implica una suspensión de células flotando en una solución después de la interrupción de sus conexiones por medios mecánicos o químicos (Björklund y Stenevi, 1984). La Figura de Cuadro 4-2 muestra la secuencia de pasos implicados en la técnica de suspensiön celular. En un número notable de experimentos se ha visto la recuperación funcional que se produce tras los trasplantes encefálicos. Uno de tales experimentos con ratas examinó el impacto de un trasplante encefálico en la realización de una tarea de laberinto en T que implicaba una alternancia espacial. Después de lesionar la corteza cerebral frontal, las ratas ejecutaban pobremente esta tarea. Sin embargo, se restauraba su ejecución cuando se les trasplantaba tejido cortical frontal de animales fetales en sus regiones frontales (Labbe, Firl, Mufson y Stein, 1983). Las compensaciones de los déficit motores ha sido un tipo común de test empleado para fundamentar las ventajas funcionales de los trasplantes encefálicos. Algunos de estos Figura de Cuadro 4-2 Ejemplo de un tipo de procedimiento de trasplante. Se extraen las partes del encéfalo y se separan las células, y después se inyectan directamente como una suspensión en el encéfalo. (Adaptado de Björklund et al., 1983.) tests pretenden hacer un modelo de la enfermedad de Parkinson, un trastorno incapacitante encefálico que implica la destrucción de células en un área del tronco encefálico (la sustancia negra). Estas células contienen una sustancia química -dopamina- que es importante para el funcionamiento apropiado de los circuitos encefálicos que controlan el movimiento. Como paso inicial en la producción de un modelo en roedores de la enfermedad de Parkinson, se destruye la sustancia negra de un lado del tronco encefálico. Esta operación lleva una reducción en el nivel de dopamina similar al observado en los pacientes con enfermedad de Parkinson. Los trastornos motores producidos por esta operación incluyen rotación del cuerpo y asimetrías posturales. La inyección de injertos de sustancia negra obtenidos de animales fetales lleva a la recuperación de los déficit motores y a un incremento del nivel encefálico de dopamina (Björklund et al., 1981; Gage y Björklund, 1984; Perlow et al., 1979). Tipos similares de trasplantes de sustancia negra han ayudado también a la recuperación de trastornos motores que se dan en ratas viejas (Gage et al., 1983). Esto ha originado la perspectiva de que los trasplantes encefálicos pueden ser útiles en el tratamiento de algunas de las degeneraciones del encéfalo asociadas a la enfermedad de Alzheimer. Hay en día, el trasplante de células y tejidos en el encéfalo se ve como una oferta prometedora para la terapia. Se ha abierto una nueva ventana para el tratamiento de algunas de las más angustiosas de las aflicciones humanas: aquellas que surgen de la muerte de las células del encéfalo. controlan el desarrollo del encéfalo -los que gobiernan la proliferación, migración y diferenciación celular y la formación de sinapsis- están sujetos a fallos que pueden tener consecuencias catastróficas para la conducta adaptativa. La magnitud de este problema se refleja en la incidencia de trastornos que producen marcados deterioros cognitivos. En los Estados Unidos aproximadamente 3,6 niños de cada 1.000 entre las edades de 5 y 17 años tienen niveles de CI por debajo de 50. En esta sección discutiremos algunos ejemplos relacionados con estados controlados genéticamente y con las condiciones maternales prenatales. Estados controlados genéticamente Los ejemplos de estados controlados genéticamente que producen trastornos del desarrollo se centran en las acciones de genes mutantes y anomalías cromosómicas. Acciones de genes mutantes Muchos trastornos metabólicos afectan profundamente el desarrollo del encéfalo. Algunos de ellos, que están asociados a herencia recesiva, generalmente aparecen muy pronto en la vida. En esta categoría tenemos alrededor de cien trastornos diferentes incluyendo alteraciones en el metabolismo de las proteínas, carbohidratos o lípidos. Característicamente, el c efecto genético consiste en la ausencia de un enzima particular que controla algún paso bioquímico crítico en la síntesis o la degradación de un producto corporal vital. Hay dos resultados principales de los déficit enzimáticos que pueden afectar a los estados metabólicos y estructurales del encéfalo: 1) ciertos compuestos alcanzan niveles tóxicos y 2) los compuestos necesarios pare funciones o estructuras no son sintetizados. Pondremos un ejemplo de la primera clase; se trata de la fenilcetonuria (FCU), un trastorno hereditario recesivo del metabolismo de las proteínas que anteriormente concluía comúnmente en retraso mental. Una de cada 50 personas es un portador heterocigótico, y uno de cada 10.000 nacimientos es una víctima afectada (homocigoto). El defecto básico es la ausencia de un enzima necesario para metabolizar la fenilalanina, un aminoácido presente en muchos alimentos. La lesión encefálica producida por la fenilcetonuria problablemente se debe a que hay un enorme exceso de fenilalanina. Las altas concentraciones de fenilalanina en la sangre de los recién nacidos puede tener diversos orígenes y consecuencias. La FCU es un ejemplo de este tipo de trastornos, y las perspectivas más recientes han sugerido un origen genético más complejo que el considerado hasta ahora (Rowley, 1984; Scriver y Clow, 1980). El descubrimiento de la FCU señaló la primera vez, que un error innato de metabolismo estaba asociado a retraso mental. Actualmente hay métodos de protección, requeridos por la ley de los Estados Unidos y de muchos otros países, que informan del nivel de fenilalanina en los niños a los pocos días después del nacimiento. Esto es importante ya que puede prevenirse el deterioro del encéfalo mediante la administración de dieta:, de bajo contenido en fenilalanina. La evidencia reciente sugiere que el control dietético de la fenilcetonuria es crítico durante los primeros años, especialmente antes de los dos años, y que la dieta puede relajarse durante la edad adulta. Sin embargo, los estudios recientes muestran que las madres con FCU conductualmente normales tienen un alto porcentaje de hijos retrasados mentalmente. Esto puede estar relacionado con los niveles de fenilalanina de la madre, aunque el tratamiento dietético durante el embarazo no parece reducir estos efectos fetales (Kolodny y Cable, 1981). El éxito en el tratamieto de la fenilcetonuria encendió el entusiasmo para la investigación sobre el análisis y posible tratamiento de muchas otras formas de retraso mental controlado por los genes que influyen en los procesos metabólicos. El análisis cromosómico, las técnicas bioquímicas y los modos de visualización fetal son herramientas poderosas que están proporcionando una mejor predicción y tratamiento de este tipo de trastornos. Un extraordinario trastorno neurológico de herencia dominante -la enfermedad de Huntington- se describe en el Cuadro 4-3. Anomalías cromosómicas Uno de cada 200 nacimientos vivos exhibe algún tipo de anomalía cromosómica: o un número anormal de cromosomas (usualmente 45 ó 47 en lugar de 46) o bien modificaciones en la estructura del cromosoma. Generalmente los trastornos que no implican a los cromosomas sexuales tienen un impacto más profundo en la conducta que aquellos que sí los implican. La forma más común de trastorno cognitivo debido a una anomalía cromosómica es el síndrome de Down. El trastorno asociado con el 95% de estos casos es un cromosoma extra, el número 21 (de aquí la designación de trisomía 21). Este trastorno está fuertemente relacionado con la edad de la madre y el momento de la concepción (Tabla 4-2). Las disfunciones conductuales son bastante variadas. En la mayoría de casos de síndrome de Down el CI es muy bajo, pero algunos individuos alcanzan un Cl de 80. Las anomalías del encéfalo en las personas con síndrome de Down son también variadas. Las biopsias recientes de la corteza cerebral de pacientes con síndrome de Down muestran una formación anormal de las espinas dendríticas. Un modelo recién descubierto en el ratón que implica un cromosoma extra, produce cambios estructurales y conductuales análogos a los producidos por el síndrome de Down en humanos (Epstein, 1986). La investigación de este modelo está proporcionando valiosas indicaciones de cómo los cromosomas extra producen anormalidades estructurales y conductuales. Condiciones maternas prenatales Incluso en el ambiente protejido de la matriz, el embrión y el feto no son inmunes a lo que está teniendo lugar en el cuerpo de la madre. Condiciones maternas, como las infecciones víricas, exposición a drogas y malnutrición, son muy probablemente causantes de trastornos en el desarrollo del feto. Considere mos ejemplos de trastornos que se derivan de dos de estas condiciones. Exposición a drogas durante el embarazo El ambiente maternal como un determinante del desarrollo del encéfalo ha originado recientemente un nuevo campo: la teratología conductual (Teratología es el estudio de las deformaciones, del griego teras, «monstruo»). Quienes trabajan en este campo están especialmente interesados en los efectos conductuales patológicos de las drogas ingeridas durante el embarazo. El fuerte uso de drogas activas conductualmente en los años recientes ha centrado la atención en su conexión con varios trastornos del desarrollo. CUADRO 4-3 Enfermedad de Huntington: una historia genética de detectives Una vez durante los primeros años del siglo XIX, una mujer que vivía en la orilla del lago Maracaibo en Venezuela, se vio afectada por una enfermedad que ha afectado a muchos de sus más de 3.000 descendientes. Esta comunidad ha llegado a ser una parte importante de una historia genética de detectives que se ha desvelado a través del uso de observaciones internacionales mezcladas con técnicas contemporáneas neurológicas y genéticas. Para comenzar esta historia es importante advertir que el esquema genético que guía el crecimiento y desarrollo del encéfalo puede ser letal para muchos humanos, ya que determina intensamente la ocurrencia de trastornos particulares. Cientos de enfermedades se han atribuido a deficiencias genéticas. En algunos casos estos trastornos aparecen pronto en la vida; en otros casos el desarrollo temprano del encéfalo se da normalmente, pero de repente, durante la edad adulta, aparece un profundo trastorno del encéfalo con poco o ningún aviso. En el ejemplo anterior, los descendientes de una persona afligida están obsesionados por la perspectiva de sucumbir eventualmente ante un trastorno que ven desarrollado en un padre o en un pariente más viejo. Uno de los más crueles de estos trastornos controlados genéticamente es la enfermedad de Huntington. George Huntington fue un joven médico cuya única publicación (en 1884) fue una descripción de una extraña aflicción motora de un familiar próximo. Observó correctamente que se trataba de un trastorno neurológico heredado que se transmitía de generación a generación. Hoy sabemos que está enfermedad se transmite por un gen dominante simple por lo que cada hijo de una víctima tiene una probabilidad del 50% de desarrollar la enfermedad. Usualmente hace su primera aparición entre los 30 y 45 años de edad con lo que la mayoría de los enfermos ya han tenido hijos sin saber si en última instancia sucumbirían al trastorno. Desgraciadamente, esto asegura la continuación de generaciones de individuos afectados, a menos que se pueda desarrollar alguna técnica que pueda informar a los descendientes de los pacientes sobre su susceptibilidad genética a la enfermedad. La enfermedad de Huntington se advierte al principio en cambios conductuales muy sutiles: pequeñas sacudidas de la cara y una cierta torpeza. La sutilidad se pierde rápidamente y pasa a ser una continua corriente de espasmos involuntarios que afectan a todo el cuerpo. Lo- movimientos sin objeto de los ojos, movimientos espasmódicos de las piernas y el retorcimiento del cuerpo hacen complicada la respiración y la alimentación. Con bastante frecuencia se hace evidente una profunda demencia; en un pequeño porcentaje de pacientes, los cambios cognitivos son los signos más tempranos de la emergencia del trastorno. La marca neuroanatómica de la enfermedad de Huntington es la destrucción del núcleo caudado, un componente esencial de los sistemas cerebrales que controlan el movimiento. Un cuadro muy triste, cuya comprensión eludió a los investigadores durante muchos años. La primera perspectiva de; identificar individuos de riesgo para la enfermedad de Huntington surgió cuando los investigadores comenzaron a estudiar el pueblo de pescadores de Venezuela en que se encontraban muchos pacientes. Los patrones de endogamia en este pueblo aislado aseguraban que el caso simple que apareció inicialmente 150 años atrás se hubiese multiplicado muchas veces y ahora abarcase al menos 100 casos actuales y varios miles con riesgo de enfermedad. Los investigadores han compilado elaboradas historias del linaje de virtualmente todos los individos de este pueblo y obtuvieron muestras de piel y de sangre. Los datos de este grupo combinados con los obtenidos de víctimas de la enfermedad de Huntington de los Estados Unidos han llevado a algunos notables descubrimientos genéticos. Los científicos que trabajan con las herramientas analíticas más modernas de la genética bioquímica han descubierto un marcador genético de esta enfermedad en la molécula de ADN (Gusella et al., 1983; Folstein et al., 1985). La identificación de este marcador permite estudios que posibilitarán la localización del locus del gen que falta y las vías que conducen a la destrucción del encéfalo. El análisis mediante el marcador permite que los individuos susceptibles sean identificados antes de que enfermen. Claramente, esta identificación genética conlleva riesgos personales, pero también permite la perspectiva de ayudar a las personas a planificar mejor su futuro. Quizás las técnicas de ingeniería genética nos proporcionarán finalmente una intervención adecuada para prevenir la expansión de esta horrible enfermedad. Tabla 4-2 Riesgo de niños con síndrome de Down relacionado con la edad Edad de la madre al nacimiento del niño Riesgo de sindrome de Down Menos de 30 1 ; 1500 30 –34 1 ; 1,000 35 – 39 1 ; 300 40 – 44 1 ;100 45 o más 1 ; 40 Fuente: Karp (1976). Síndrome alcohólico fetal Aunque la investigación reciente ha demostrado el potencial del alcohol para afectar al crecimiento y al desarrollo fetal, hay una larga historia de la relación entre el alcohol y el embarazo, que data desde los tiempos clásicos de griegos y romanos. Aristóteles proclamaba que «las mujeres locas... borrachas... ofrecen posteriormente niños como elías mismas, taciturnos y lánguidos» (citado en Abel, 1982). Por ahora, la verdad de esta observación está bien apoyada por abundantes estudios de investigación. Los niños nacidos de madres alcohólicas muestran un perfil distintivo de trastornos anatómicos, fisiológicos y conductuales que hoy se conoce como el «síndrome alcohólico fetal» (Abel, 1984; Colangelo y Jones, 1982). Los importantes efectos anatómicos de la exposición fetal al alcohol incluyen cambios distintivos en las características faciales (por ejemplo, puente nasal hundido, forma alterada de la nariz) y en las características de los párpados. Las deficiencias en el crecimiento intrauterino son particularmente evidentes debido a que los niños nacidos de madres alcohólicas presentan menos altura y peso en el nacimiento. Pocos de estos niños se recuperan en los años siguientes al nacimiento (Colangelo y Jones, 1982). El problema más común asociado con el síndrome alcohólico fetal es el retraso mental, que puede variar en severidad aunque es una característica persistente. No se ha establecido aún un umbral de alcohol para este síndrome, pero está claro que puede darse con una ingesta relativamente moderada de alcohol durante el curso del embarazo. Además de retraso mental, los niños con síndrome alcohólico fetal muestran otros signos neurológicos. Se aprecian comúnmente hiperactividad, irritabilidad, temblor y otros síntomas de inestabilidad motora. Los investigadores tienen que establecer si estos efectos están fundamentalmente mediados por el alcohol, sus metabolitos tóxicos o por los efectos del alcohol en la salud metabólica y nutricional de la madre. Otra posibilidad es un efecto de los lazos circulatorios entre la madre y el hijo. Este síndrome puede no ser distintivo del alcohol; el empleo intenso de marihuana parece ejercer un efecto -similar sobre cl crecimiento y el desarrollo fetal (Hingson et al., 1982). Dos calendarios para el desarrollo encefálico Ahora vamos a intentar ofrecer conjuntamente las investigaciones sobre el desarrollo del encéfalo a lo largo de dos escalas de tiempo enormemente distintas: las semanas y meses de crecimiento de un individuo y los millones de años de evolución. Podemos emplear la analogía de las contribuciones distintas, pero igualmente esenciales, de un arquitecto de un carpintero en la construcción de una casa. El arquitecto, al preparar los planos, apela a una larga historia de conocimiento humano sobre las estructuras que satisfacen las necesidades básicas humanas: descanso, trabajo, recreo, alimentación, cuidado de los niños, etc. La estructura debe ser confortable, segura y suficiente, y debe concordar con las inclinaciones de la comunidad. El carpintero tiene que emplear estos planos para construir la casa, traduciendo la información bidimensional dada en el proyecto en una estructura tridimensional. En cierto número de ocasiones durante el proceso de construcción, serán necesarios el juicio y la interpretación del carpintero. Así, dos casas construidas por diferentes carpinteros a partir de los mismos proyectos no serán idénticas. Otra razón para las diferencias en las casas es que los materiales disponibles para sus construcciones pueden no ser exactamente los mismos. El arquitecto intenta prever algunos de los problema, de construcción e incluir factores de seguridad en los planos, así que pequeñas desviaciones o errores no deteriorarán seriamente la seguridad o utilidad de la construcción acabada. No somos los primeros en utilizar esta analogía. Una inteligencia anónima señaló que un bebé es el objeto más complicado de construir por un trabajador no especializado. Y el científico psicólogo de la información J. C. R. Licklider caracterizó a Dios como un gran arquitecto pero un trabajador descuidado. Los planos para la construcción del encéfalo tienen ciertas características que debemos señalar y comentan: 1. Los planos nuevos nunca comienzan de la riada. Evidentemente, los planos más antiguos son reutilizados y modificados para ajustarse a situaciones específicas. 2. No todos los detalles están especificados. Parte del programa está implícito en la lista de materiales y métodos de construcción. Los planos serían desesperanzadoramcnte complejos y voluminosos si tuviese que especificarse cada detalle. 3. Se realizan concesiones para la interacción entre los materiales y el ambiente. Un arquitecto sabe cómo se desgastarán ciertos guijarros en un clima dado para producir una apariencia deseada o cómo la plantación de un jardín estabilizará el sucio y embellecerá la ubicación de la casa. Así, también, los planos genéticos para el encéfalo toman ventaja de la información provista por el ambiente. Estos planes tienen en cuenta la interacción entre el organismo en desarrollo y su ambiente. La reutilización y la modificación sucesivas de los planos genéticos significan que las etapas embriológicas de desarrollo de todos los vertebrados son similares. Los tubos neurales tempranos de los embriones de una rana, una rata o una persona se parecen mucho. Además, Pis divisiones básicas del encéfalo son las mismas en todas estas formas. Sin embargo, la estructura global ha sido aumentada de escala en los mamíferos, y especialmente en los primates, y algunas partes han : do ampliadas en relación con otras. El código genético no parece tener cabida para i oda la información necesaria para especificar el diagrama de conexiones completo para cada parte del 'sistema nervioso. Consigue cierta economía mediante la aplicación de la misma información a muchas partes distintas de la estructura. Así el mismo gen puede especificar aspectos de los circuitos neurales en diferentes áreas del encéfalo. Cualquier mutación del gen puede producir además una disposición anormal de neuronas en la corteza cerebelosa y en la corteza cerebral. También, ciertas hormonas estimulan el crecimiento de las conexiones neurales a través del sistema nervioso, como veremos en el Capítulo 7. Y algunos detalles finos del establecimiento de conexiones parecen no estar especificados, sino que simplemente son trabajados localmente. La economía de las instrucciones genéticas y la adaptación a las circunstancias individuales se logran contando con que el ambiente suministrará cierta información necesaria para el desarrollo. Cada especie ha evolucionado en relación a un nicho ecológico particular, y su programa de desarrollo utiliza el ambiente como una fuente de información y estimulación. Así, por ejemplo, la mayoría de los vertebrados están expuestos a patrones de estimulación visual de forma temprana después del nacimiento. Con el tiempo de exposición, el plan básico del sistema visual se deja de lado. Pero la información de las conexiones detalladas y el mantenimiento de los circuitos visuales precisan inputs del ambiente. La coordinación precisa de los inputs de los dos ojos requiere un fino ajuste del sistema. Hay demasiadas variables en la estructura de los ojos que serían extraordinariamente costosas de especificar genéticamente para proporcionar un perfecto ajuste de las dos imágenes retinales. El programa de los genes ha llegado hasta vosotros a través de millones de años de pruebas de ensayo y error, pero esto tiene sus límites. De este modo, son necesarios ciertos ajustes antes de que el individuo comience una operación, como hablar. Pequeños desajustes de las dos imágenes retinales pueden compensarse con un «reajuste» menor de las conexiones visuales centrales. Pero si el desajuste entre los dos ojos es demasiado grande, como cuando los ojos están cruzados, entonces habitualmente se suprime el input de un ojo. De este modo se evita la visión doble. La capacidad de aprender de nuestro ambiente y de nuestra experiencia nos capacita para ajustarnos a ambientes y estilos de vida particulares. (Los Capítulos 16 y 17 considerarán los mecanismos biológicos del aprendizaje y la memoria.) Ahora podemos ver que los calendarios a corto y a largo plazo proporcionan perspectivas complementarias del desarrollo del sistema nervioso y la conducta. Apelaremos a ambas perspectivas para aclarar las relaciones entre encéfalo y la conducta en muchas áreas de la psicología fisiológica. Resumen: Aspectos fundamentales 1. Los acontecimientos embriológicos tempranos en la formación del sistema nervioso implican una secuencia intrínsecamente programada de procesos celulares: a) producción de células nerviosas (proliferación celular), b) desplazamiento de células lejos de las regiones de división mitótica (migración celular, c) adquisición por parte de las células nerviosas, de formas distintivas (diferenciación celular) y d) pérdida de algunas células (muerte celular). 2. Los cambios fetales postnatales del encéfalo implican la mielinización de los axones y el desarrollo de las dendritas y las sinapsis. Aunque muchas neuronas ya están presentes en el nacimiento, la mayor parte del desarrollo de las sinapsis en humanos se da después del nacimiento. 3. La neuroespecificidad es la teoría que indica que la formación de las vías neurales y las sinapsis siguen un plan innato que especifica las relaciones precisas entre los axones en crecimiento y las células diana particulares. La extensión en que están determinadas genéticamente las conexiones específicas es materia habitual de controversia. 4. Entre los muchos determinantes del desarrollo del encéfalo están a) la información genética, b) los factores de crecimiento, como los factores de crecimiento nervioso, y c) la nutrición. 5. La experiencia afecta al crecimiento y el desarrollo del sistema nervioso. Esto se observa mediante experimentos en que los animales sufren privación sensorial durante períodos críticos tempranos de su desarrollo. Los resultados indican que la experiencia puede inducir y modular la formación de sinapsis y puede también mantenerlas. 6. El encéfalo continúa cambiando a lo largo de la vida. La edad madura conlleva pérdida de neuronas y conexiones sinápticas en algunas regiones del encéfalo. En algunas personas los cambios son más severos que en otras, y 7. Pueden darse varios tipos de desarrollos anómalos dei encéfalo como consecuencia de trastornos controlados genéticamente. Algunos son trastornos metabólicos, como la fcnilccfonuria (FCU), e implican una incapacidad corporal para sintetizar un enzima particular. Otros trastornos hereditarios, como la enfermedad de Huntington, aparecen sólo en la edad adulta. Cada d :recto está gobernado probablemente por un solo gen. 8. Algunas formas de retri'so mental, como el síndrome de Down, están relacionadas con trastornos en los cromosomas, en este caso particular un número excesivo de ellos. 9. Los deterioros del desarrollo fetal que llevan al retraso mental pueden deberse a drogas, como el alcohol o la marihuana, si son empleadas durante el embarazo. Lecturas recomendadas Greenough, W. T., y Juraska, J. M. (Eds.) (1986). Developmental neuropsychobiology. Orlando, Fla.: Academic Press. Hopkins, W. G., y Brown, M. C. (1984). Development of nerve cells and their connections. Cambridge, England: Cambridge University Press. Jacobson, M. (1978). Developmental neurobiology. New York: Plenum, los cambios patológicos se dan en la condición conocida como enfermedad de Alzheimer o demencia senil. Lund, R. D. (1978). Development and plasticity of Me brain. New York: Oxford University Press. Purves, D., y Lichtman, J. W. (1985). Principies of neural development. Sunderland, Mass.: Sinauer. Spreen, O., Tupper, D., Risser, A., Tuoko, H., y Edgell, D. (1984). Human developmental neuropsychology. New York: Oxford University Press. Unidad 3 • Bases Genéticas y de Desarrollo de la Conducta Los rasgos dominantes asociados con la presencia de un cromosoma Y adicional incluyen altura inusitada, episodios de agresividad excesiva e inteligencia mínima. Sin que a nadie sorprenda el hecho, en algunos de los más famosos criminales de nuestro tiempo se ha encontrado un cromosoma Y adicional. Un caso entre los primeros de esta índole fue el de Robert Tait, un australiano que en 1962 fue convicto por haber dado muerte con una porra a una anciana de 81 años, en una vicaría donde él había ido en busca de ayuda. En 1965, Daniel Hugon, un establero francés de 31 años de edad, estranguló brutalmente a una prostituta parisiense, aparentemente sin motivo alguno. Durante el curso de su juicio se le halló un cromosoma Y adicional. En abril de 1969, John Farley de seis pies y ocho pulgadas de estatura, y 240 libras de peso, apodado "El alegre gigante verde"; a causa de su extraordinaria dotación física y "Juanote el malo'; por sus arrebatos violentos, confesó haber golpeado, estrangulado, violado y mutilado a una mujer de Queens, Nueva York. Fue defendido sobre la basa de que, debido a la presencia de un cromosoma Y adicional en sus células, él no tenía discernimiento ni control de sus acciones y fue hallado por lo tanto no culpable "en razón de su demencia causada por un desequilibrio cromosómico". (Jarvik y otros, 1973, p. 675) Estos ejemplos aterradores son una demostración clara de la importancia de los factores genéticos en la conducta, sin hacer mención del carácter excéntrico de nuestro sistema legal. Los individuos que poseen un tipo genético de XYY son del sexo masculino con el sobrante de un cromosoma Y. El tipo genético masculino no normal es "XY" el tipo genético femenino normal, XX. La evidencia actualmente refuerza el concepto de que este cromosoma Y adicional es mucho más común entre hombres convictos de crímenes violentos que entre hombres de la población general. El conjunto genético ciertamente influye en la conducta como lo indica el síndrome XYY. Más aún, es posible argumentar que la agresión humana puede "residir" en el cromosoma Y. Las mujeres son mucho menos agresivas que los hombres, por lo menos en lo que a violencia física se refiere. Hay dos explicaciones posibles: los hombres son más agresivos y las mujeres lo son menos porque la sociedad enseña estos papeles sociales, y (2) hay una determinación genética inherente a 13 violencia en los hombres. Los datos del síndrome XYY parecen dar cierto apoyo a la hipótesis genética. La importancia del síndrome se extiende mucho más allá del problema de la violencia considerado en sí mismo. Las anormalidades cromosómicas descubiertas anteriormente se han relacionado con anormalidades físicas o con retardo mental clasificación burda y global de defectos. La persona que tiene el síndrome de Klinefelter (tipo genético XXY) es de sexo femenino con muchas características masculinas - la "mujer" atleta de aspecto masculino. La persona que tiene el síndrome de Down (mongolismo) posee un cromosoma adicional no ligado al sexo y es retardado mental además de tener ciertos rasgos y defectos físicos característicos. Se trata de anormalidades físicas e intelectuales relativamente importantes. Sin embargo, el tipo genético XYY produce una anormalidad que es primordialmente conductual - la agresión. ¿Cómo conduce el cromosoma Y a la violencia? Los cromosomas son simplemente, después de todo, mezclas de sustancias químicas. El cromosoma Y (masculino), sea dicho de paso, es físicamente mucho más pequeño que el cromosoma X (femenino) Puede describirse como regresivo o como más dominante, dependiendo tal vez de los propios cromosomas de quien opina. De todos modos, la combinación XY produce un individuo de sexo masculino normal y la combinación XX un individuo de sexo femenino normal, con todas las diferencias físicas y conductuales normales. El cromosoma Y adicional exagera varias características masculinas - por ejemplo la altura y la contextura muscular (algunos podrían agregar así mismo, la inteligencia reducida) - además de la agresión. Esto indica que la estructura física y la actividad glandular son en cierta forma controladas por los cromosomas X y Y El desarrollo cerebral mismo - el crecimiento y estructura del sistema nervioso - está bajo el control genético y recibe la influencia de las hormonas. Hay sin embargo todavía, grandes vacíos en la forma como nosotros entendemos que los cromosomas o sea el material genético, controlan el crecimiento y desarrollo y a fin de cuentas la conducta de los seres humanos. Cada cromosoma está compuesto de una gran cantidad de genes, última unidad estructural de la herencia. Las estructuras y funciones de todos los organismos están determinadas por los genes. Realmente los genes tan sólo determinan las estructuras de las sustancias químicas denominadas proteínas. Estas son de dos tipos: proteínas estructurales y enzimas. Las proteínas estructurales del organismo y las enzimas controlan las reacciones fisicoquímicas del organismo. Teóricamente todas las funciones biológicas y conductuales pueden retrotraerse a la acción genética. En la práctica sin embargo, al desarrollarse el organismo a partir del momento en que se fertiliza el huevo, los efectos genéticos interactúan con el ambiente. La distinción decisiva se hace entre el genotipo (la estructura genética subyacente de un organismo) y el fenotipo (expresión externa de la acción genética como resultado de la interacción con el ambiente) Entre los animales superiores, toda conducta es fenotípica. En este capítulo examinaremos los fundamentos de la genética, las bases genéticas de la conducta (materia que es objeto de un nuevo campo fascinante denominado genética conductual), el desarrollo del sistema nervioso y la influencia relativa de la estructura genética y del ambiente en la conducta. GENÉTICA BÁSICA Las leyes de la herencia Aunque Darwin postuló que la selección natural operaba por la herencia de las características que demostraban adaptabilidad, el mecanismo era oscuro. Por el tiempo en que Darwin estaba formulando la versión final del Origen de las Especies, un monje austriaco, Gregor Johan Mendel, experimentaba con la hibridación de los guisantes. Los hallazgos de Mendel publicados en 1866, lo condujeron a las hipótesis de que (1) la herencia se transmite por unidades independientes y heredables; (2) cuando cada padre aporta la misma clase de unidad, la progenie presenta este carácter, pero cuando cada padre suministra una clase diferente de unidad, se forma un híbrido y cuando los híbridos forman sus propias células reproductivas, "se liberan" nuevamente las dos clases diferentes de unidades; y (3) las unidades hereditarias no se afectan por la experiencia de un individuo. En tales hipótesis formuló Mendel las leyes básicas de la herencia-la noción de los genes-pero por más de tres décadas permaneció desconocido su trabajo. En un lapso de cinco meses durante el año de 1900; tres botánicos europeos redescubrieron el trabajo de Mendel y lo pusieron cara a cara con la biología. El más conocido entre ellos era Hugo De Vries, el cual hizo hincapié en el concepto de las mutaciones como responsables del desarrollo de especies nuevas. Los estudiosos modernos de la genética no insisten en la mutación en la medida de De Vries y sus seguidores. Todos son conscientes sin embargo de que aunque el material genético de una población se halle en proceso continuo de mezcla por razón de los apareamientos, el único camino para que be introduzcan caracteres completamente nuevos es el de la mutación. La base de la herencia La ciencia de la genética empezó con el redescubrimiento de la obra de Mendel. Se sabía ya que el núcleo de cada célula contiene cierto número de pares de finas estructuras filiformes que, se reproducen por sí mismas cuando se divide la célula y a las cuales se llamó : cromosomas... Se sabía que las diferentes especies poseen números diferentes de pares-la mosca frutera, 8 cromosomas ó 4 pares estructuralmente similares; el ser humano normal, 46 cromosomas ó 23 pares (Figura 2.1). Se sabía también que cuando se forma un huevo o una célula espermática, no se incorpora en la nueva célula sexual sino un cromosoma de cada par, de tal manera que la fertilización del huevo por parte del espermatozoide produce un individuo nuevo con una complementación total de pares de cromosomas. En años posteriores se hizo evidente mediante los esfuerzos de hombres tales como Thomas Hunt Morgan y H. J. Muller, que las unidades hereditarias de Mendel, los genes, dependen de los cromosomas. Se descubrió que si por medio de radiación se producen mutaciones de los genes, se forman organismos anormales. Se demostró que los cromosomas constan principalmente de dos sustancias: ácido desoxirribonucleico (DNA) y proteínas. De los dos, el DNA es el más estable, nunca abandona el núcleo y es capaz de reproducirse por sí mismo. Tiene la propiedad fundamental de los genes. Según estudios posteriores, la totalidad del DNA tiene la misma composición química. Consta de un azúcar puro (desoxirribosa), fosfato y cuatro bases: adenina, guanina, timina y citosina (A, G, T, C,). El siguiente gran paso ocurrió con el descubrimiento de J. D. Watson y F. H. C. Crick respecto de la estructura de la molécula del DNA. Utilizando la cristalografía de rayos X, Watson y Crick propusieron el modelo de doble hélice (Figura 2.2). Según este modelo, la molécula consta de dos ramales mutuamente enrollados en espiral. El sostén de cada ramal consiste en un arreglo continuo de azúcar-fosfato-azúcar-fosfato. Las bases A, G, T y C están ligadas a dichos sostenes y a su vez se unen entre sí, vinculando en tal forma los dos ramales. Aunque las bases pueden presentarse en cualquier secuencia a lo largo de un ramal, existe una relación fija entre los ramales com- Figura 2.1 Cromosomas humanos masculinos tomados de un glóbulo blanco. Se usó un procedimiento que tiñe selectivamente solo los cromosomas de la célula. Los cromosomas aquí mostrados se encontraban en el proceso de división. El homólogo de cada uno de ellos se encuentra localizado en alguna otra parte de la microfotografía. plementarios. La adenina se une siempre con la timina y la guanina siempre con la citosina. El código genético está contenido en la secuencia de las bases. Dicho código determina las propiedades y efectos de cada gene. La capacidad del DNA para reproducirse por sí mismo depende de la combinación específica de ligaduras de las bases (Figura 2.3). Cuando los ramales se separan durante la división de las células, son capaces de producir copias idénticas de la molécula original DNA seleccionando sus compañeros apropiados. Los mecanismos de la herencia El DNÁ nunca abandona el núcleo. Los genes ejercen su acción directa determinando las proteínas que deben producirse en los ribosomas (estructuras especializadas que se hallan fuera del núcleo en el citoplasma de la célula). El mecanismo de traducción del código genético (secuencia de las bases en la molécula DNA) en una proteína implica ácido ribonucléico (RNA), sustancia que en su estructura es similar al DNA pero está presente en toda la célula. Una clase especial de RNA denominada RNA mensajero, transfiere el código del DNA al ribosoma, donde otro RNA el RNA de transferencia, probablemente actúa en la recolección de los aminoácidos particulares requeridos para la elaboración de la proteína codificada por el DNA (Figura Figura 2.2 El modelo de doble hélice para la duplicación del ADN propuesto por Watson y Crick. Los dos cabos del ADN se enroscan entre sí formando una doble hélice. Durante la división la molécula se desenrolla, las bases se separan quedando una base de cada par ligada a cada cabo. Tal como se muestra aquí, se forma un cabo complementario, que origina después del fenómeno de duplicación, dos moléculas de ADN. (Tomado de Stent, Gunther S. Molecular Biology of Bacterial Víruses. Copyright ©1963 por W. H. Freeman y Co.) Figura 2.3 Un segmento de ADN que muestra las combinaciones de enlaces entre las cuatro bases que lo forman: adenina, guanina, timina y citosina. Como se muestra aquí, la adenina solo se enlaza con la timina y la guanina solo con la citosina. (Segundo dibujo tomado de Lerner, 1. Michael. Heredity, Evolution and Society. Copyright © 1968 por W.H. Freeman y Co.) 2.4). Los tipos de proteínas como también los lugares y oportunidades de su conformación, determinan el sexo de un individuo, el color de su piel e indudablemente indican si se trata del un gusano de la tierra, una mosca frutera, un ratón o un hombre. Por lo general la formación del RNA ocurre en ocasiones en que las células realizan funciones fisiológicas, pero no mientras se están multiplicando. Durante estos períodos los dos ramales de la hélice ADN están desenrollados. Solamente uno de ellos produce RNA en forma activa. Realiza tal función seleccionando bases complementarias del material celular, pero remplaza la timina (T) por uracila (U). Cuando se termina la cadena de RNA, se desprende del DNA, abandona el núcleo de la célula y se adhiere a los ribosomas (Figura 2.4). Este es el RNA mensajero. El RNA de transferencia lleva aminoácidos a los sitios de los ribosomas, donde tales aminoácidos se recogen en proteínas - en esencia, largas cintas de, aminoácidos enganchados unos con otros. Hay 20 aminoácidos que forman la mayor parte de las proteínas. Hay cuatro bases diferentes de RNA y se requieren tres para especificar un aminoácido. Tomando cada vez tres de las cuatro bases, se llega a un total de 64 aminoacidos-cantidad considerablemente superior a los propios 20 requeridos. Finalmente, las proteínas y enzimas estructurales se forman en los ribosomas. Los genes de un rasgo particular se presentan en lugares específicos de cromosomas específicos. Los dos cromosomas que forman cada par tienen dentro de sí lugares homólogos. Cuando se forma el huevo o la célula espermática solamente se incluye un cromosoma de cada par y así tan sólo uno de los dos genes presentes en los lugares homólogos del padre queda incluido. Cuando ocurre la fecundación, la célula posee de nuevo cromosomas apareados, pero el 50 por ciento de los genes es de cada uno de los padres. Los diversos genes (secuencias de las bases) que puede haber en un solo lugar cualquiera se denominan alelos. Si el cigote recién formado (huevo fertilizado) hereda el mismo alelo en determinado lugar de los dos cromosomas homólogos, se dice que el individuo es homocigótico en cuanto a ese gene; si diferentes alelos están presentes en los dos lugares homólogos, el individuo es heterocigótico respecto de ese gene. En el caso más simple, cuando un gene en determinado lugar es Figura 2.4 Mecanismo de traslación del código genético a una proteína: La información de la molécula de ADN se transcribe a una molécula de ARN (1) Por un proceso de apareamiento se forma un cabo complementario de ARN (llamado mensajero) (2). El ARN mensajero transfiere el código del ADN a los ribosomas en el citoplasma (3). Los ribosomas son los elementos donde se realiza la síntesis de proteínas, los ARN llamados de transferencia y los aminoácidos ya enlazados (mostrados aquí como números encerrados en rectángulos) se alinean sobre el ARN mensajero en una secuencia particular (4). Estos aminoácidos se combinan formando cadenas de polipéptidos que constituyen las proteínas. (Adaptación de Niremberg, M. W. El código genético: II. Copyright © 19(33 por Scientific American, Inc. Reservados todos los derechos.) dominante (R) y otro es recesivo (r), el genotipo de cualquier miembro de la población puede ser RR, Rr, o rr. Supóngase que R produce ojos rojos y r ojos grises, en la mosca frutera. Cuando dos miembros heterocigóticos de la población, ambos de ojos rojos se aparean (RrXRr), las proporciones esperadas de los genotipos de la cría serán de 1:2:1 (RR:Rr:rr) y la proporción de fenotipos será de 3:1 (esto es, tres de cada cuatro tendrán los ojos rojos; Figura 2.5). Lo que aquí importa anotar es que se mantiene la variabilidad en la población. Más aún, siendo que uno u otro cromosoma de cualquier par puede ser aportado al cigote y dado que la integración de los cromosomas es aleatoria, se incrementa en gran manera la variabilidad genética a través de la reproducción sexual. En el último ejemplo, la acción genética dominante es completa. No es posible por el simple examen de los ojos (fenotipo) distinguir la mosca homocigótica de ojos rojos (RR) respecto de la mosca heterocigótica de ojos rojos (Rr). En muchos casos `el predominio es incompleto. Un ejemplo importante desde el punto de vista médico es la anemia de células falciformes, enfermedad hereditaria más común entre los negros de la costa central de África o entre aquellos cuyos antepasados vinieron de aquella vasta región. El mal se caracteriza por una forma anormal de hoz en las células rojas de la sangre, la cual causa en ellas la formación de un precipitado. Se presenta co- Figura 2.5 Ejemplo de la acción de los genes dominantes y recesivos en la peterminación del color de :os ojos (fenotipo) en la mosea de la fruta. Cuando dos moscas son heterócigas por el color de los ojos, la proporción de fenotipos en su descendencia será de 3:1 esto es, tres de cada cuatro tendrán los ojos rojos. Sin embargo la proporción de los genotipos es de 1:2:1 de modo que cuando tres moscas tienen ojos rojos solo una de ellas es homóciga por el color (rojo) de los ojos. mo resultado de la acción de un solo gene. Un individuo que sea homocigótico respecto de ese gene, padece una enfermedad grave; la mayor parte muere en la niñez, y los que sobreviven son enfermos crónicos de por vida. En los Estados Unidos solamente un 0,25 por ciento aproximado de la población negra tiene tal enfermedad en la forma homocigótica, pero aproximadamente el 9 por ciento es heterocigótico respecto del gene anormal. Ciertas tribus africanas tienen hasta un 40 por ciento de su población heterocigótica y un 4 por ciento homocigótica. En los individuos heterocigóticos respecto del gene de célula falciforme, el predominio del gene normal es incompleto. En circunstancias ordinarias tales personas se presentan llenas de salud, pero enferman al encontrarse en regiones de altura considerable donde la sangre no recibe oxígeno suficiente. Así pues, los portadores que tienen un gene normal y un gene de célula falciforme ordinariamente no se ven afectados, pero responden en forma anormal al impacto del oxígeno escaso. Lo que causa desconcierto en tal enfermedad es su gran incidencia en ciertas regiones de África. Todo lo que sabemos acerca de la evolución, indica que tal gene debilitador no debe existir en una proporción tan alta de la población. La razón de su persistencia se aclaró cuando se observó que las regiones de África donde la anemia de células falciformes es más común, son así mismo las regiones donde la malaria está más extendida y reviste mayor gravedad. La correlación es muy estrecha. En el cinturón central de África, las tribus que viven en regiones ajenas a la malaria no tienen virtualmente anemia de células falciformes. Los individuos heterocigóticos respecto del gene de célula falciforme tienen mejores posibilidades de sobrevivir que las personas normales o los homocigóticos respecto del gene anormal. Un epílogo halagüeño de esta historia lo constituye el hecho de que la incidencia del gene de células falciformes se halla en descenso continuado entre los negros de América donde la anormalidad no tiene valor adaptativo. Este es, en efecto, un ejemplo de la evolución humana en su curso actual. La variabilidad genética es importante en cualquier población. Es necesaria no sólo para el desarrollo de especies nuevas, sino también para la supervivencia de una especie existente que se enfrenta aun ambiente de continua mutación. La variabilidad de la fusión genética de una población aumenta por razón de procesos tales como la elección aleatoria en la reproducción, las mutaciones (arreglos repetidos de las bases de la molécula DNA), el entrecruzamiento (rompimiento de cromosomas e intercambio de partes entre ares, lo cuales permiten combinaciones nuevas de genes en un cromosoma dado) y la existencia de más de dos alelos para determinado lugar cromosómico. Bases Genéticas De La Conducta Los genes no actúan directamente sobre la conducta. Intervienen tan sólo para regular la producción de proteínas responsables de la estructura y actividades enzimáticas de un organismo. Sin embargo, desconocemos la base estructural de la conducta en la mayoría de los casos. Por ejemplo, sabemos que un caballo desprovisto de patas no puede correr y conocemos ciertos aspectos de la morfología de los huesos y de los músculos la cual se correlaciona con la mayor velocidad de los caballos. Pero esto no basta; ningún criador de caballos podría ir adelante si basara su negocio exclusivamente en los rasgos anatómicos de las patas de los animales. Otros rasgos-el temperamento, la coordinación neuromuscular, la tenacidad-cuentan mucho en un buen caballo de carreras. En lugar de considerar únicamente las patas, los criadores intentan aparear los animales que han demostrado éxito en el hipódromo. De manera semejante, los investigadores de las bases genéticas de otras conductas realizan sus estudios, sea examinando individuos que ostentan una conducta particular, sea analizando el linaje de individuos que difieren en una conducta dada. Deben destacarse dos puntos relacionados con el estudio de la genética conductual. Primero, este enfoque no deja de lado los efectos del ambiente en la conducta. Por el contrario, estudia la interacción de los genes y el ambiente tratando de cuantificar la porción de variabilidad observada en un rasgo de determinada población, la cual resulta de las diferencias genéticas entre los individuos y aquélla que es producto de las diferencias ambientales. El porcentaje de variación de un rasgo que se debe a diferencias de genotipo se ha denominado heredabilidad de un rasgo. Esta es una medida o índice numérico que indica la medida en que se hereda un rasgo, característica o tendencia conductual. Segundo, la gran mayoría de las características conductuales no depende de un solo gene que actúa en una forma totalmente común o completamente particular. Más bien las conductas dependen de muchos lugares donde la contribución de cada gene es pequeña y acumulativa. Estos sistemas se denominan poligénicos. Estudios experimentales acerca de la manipulación genética Tratándose de animales es posible disponer las condiciones de laboratorio en tal forma que el ambiente se mantenga razonablemente constante y así mismo pueda practicarse la selección artificial (1) midiendo una característica de conducta particular en una población de laboratorio, (2) seleccionando animales de los extremos superior e inferior y aparcando animales dentro de los extremos (los de puntaje alto con los de puntaje alto y los de puntaje bajo con los de pontaje bajo) y (3) probando las crías y continuando en cada generación subsiguiente el apareamiento entre los extremos. Si en tales condiciones ambientales hay influencia genética en la variabilidad de la conducta, las dos poblaciones diferirán de cada generación sucesiva hasta agotarse la escala de variabilidad genética. Los primeros estudios que utilizaron la selección natural fueron dirigidos por Robert Tryon en Berkeley. Tryon escogió el aprendizaje de laberinto en las ratas. Después de medir la ejecución de laberinto en una muestra no seleccionada, apareó los ejemplares más rápidos ("brillantes en el laberinto") entre sí, e igualmente los más lentos ("lerdos de laberinto") con los más lentos. Habiéndose cumplido tal procedimiento por ocho generaciones, los dos grupos no presentaron casi ningún traslape (Figura 2.6). Los experimentos de cría selectiva realizados con ratas y ratones han demostrado que gran cantidad de rasgos conductuales pueden variarse por medio de la manipulación genética. Las características estudiadas incluyen aprendizaje de laberinto, emotividad medida por la tendencia a defecar en una situación de campo abierto, actividad medida por el recorrido realizado en una rueda giratoria, agresividad, proporción de aprendizaje de evitación y susceptibilidad a ataques inducidos por sonidos. Muchos investigadores se han vuelto recientemente hacia la mosca frutera Drosophila para el estudio de las influencias genéticas en la conducta. La Drosophila puede educarse para una conducta foto táctica-acercamiento versus no acercamiento a una fuente de luz-o para una conducta geotáctica-tendencia a subir o bajar en un punto de elección. En un estudio fototáctico realizado en la mosca frutera, Hirsch y Boudreau (1958) cruzaron animales que tendían a acercarse a una fuente de luz (fototaxis positiva) y cruzaron animales que huían de ella. Después de 29 generaciones estimaron que aun en este organismo relativamente simple sólo el 57 por ciento de la variación estaba determinado en forma genética. Figura 2.6 Crianza selectiva de ratas con habilidad en el aprendizaje de un laberinto. Esta habilidad se midió en términos del número total de llegadas a un callejón sin salida durante 19 intentos. Existe una diferenciación progresiva entre los sujetos "talentosos" y los "mediocres" de la generación F2 a la F8. En Moss,F.A. (ed.) Comparative Psychology New York: Prentice-Hall 1942, pp. 330-365). Además de los experimentos de selección artificial, gran parte de nuestros conocimientos acerca de las influencias genéticas en la conducta de los animales (incluyendo al hombre) proviene de comparaciones de diferentes familias o razas. Consideremos -por ejemplo, las diferencias conductuales entre las razas de perros conocidos. Scott (1964) comparó cinco razas de perros en 34 rasgos conductuales y calculó el porcentaje de variabilidad (heredabilidad) atribuible a diferencias de raza. El promedio de heredabilidad para los 34 rasgos fue del 27 por ciento, exactamente igual al de los 10 rasgos físicos que midió. Se ha hecho procrear gran cantidad de clases de ratones dentro de la misma línea familiar para facilitar el examen de las diferencias raciales en la conducta. Aislando una población y apareando continuamente parientes cercanos, puede reducirse radicalmente la variabilidad genética de una población. Mediante el aislamiento y la reproducción libre de mezclas, pueden producirse familias en las cuales los individuos son genéticamente uniformes y diferentes de los de otras familias. Se ha descubierto que las familias de ratones de este tipo difieren en cierta cantidad de rasgos conductuales que incluyen agresividad, actividad locomotora, aprendizaje, preferencia alcohólica, conducta de atesoramiento, cantidad de crías, conducta exploratoria, resistencia a la depresión y conducta de apareamiento. Aunque se hacen muchas críticas al enfoque de la reproducción lineal [dentro de la misma familia] con base principalmente en el hecho de que tal cantidad de homocigóticos producidos nunca podrían presentarse en una población natural próspera, este método nos ha ayudado inmensamente a comprender las influencias genéticas en la conducta. El grado de actividad locomotora suministra un ejemplo claro de una característica conductual heredada en familias lineales de ratones (Figura 2.7). Se probaron los niveles generales de actividad de seis familias diferentes colocando los animales sobre una reja dividida en cuadros. La actividad se determinó por el número de cuadros cruzados en un período de cinco minutos. Como se observa en la Figura 2.7, las seis familias presentaron grandes diferencias en el grado de actividad locomotora. Resultó igualmente diferente la actividad dentro de una familia en cuanto a la medida de la variabilidad. La familia marcada en la gráfica superior no presenta virtualmente actividad alguna y se observa poca variabilidad. Por el contrario, la familia marcada en la gráfica más baja presenta poca actividad pero una variabilidad considerable. Thiesen (1972) ha demostrado que tales diferencias de familia en la actividad se mantienen en una gran variedad de situaciones de prueba durante años de reproducción. La actividad se presenta como un rasgo conductual estable, genéticamente determinado. La emotividad parecería ser un aspecto mucho más complejo de la conducta que la actividad. Sin embargo, utilizando como simple indicio de emotividad cierta cantidad de operaciones defecatorias en una situación de prueba en campo abierto, Lindzey y otros (1960) lograron demostrar que la capacidad de respuestas emotivas difiere según las diversas familias de ratones y como tal permanece durante años de procreación. La emotividad, por lo menos en familias lineales de ratones es un rasgo relativamente estable y marcadamente determinado por factores genéticos. Un ejemplo curioso de reproducción lineal se encuentra en el ratón bailador. Se creía que cl desarrollo de tal familia se remontaba hasta el año 80 A.C. en China, y que probablemente tiene una genealogía continua que se extiende hasta el presente. La conducta de los bailadores es realmente sorprendente: giran con rapidez durante largos períodos sin presentar indicios de vértigo. El defecto genético se expresa claramente en un daño del sistema vestibular y del oído interno. La susceptibilidad a los ataques audiogénicos es otro ejemplo. Ciertas familias de ratones, frente a determinados sonidos, sufren ataques de efectos similares a los de la epilepsia, que les causan frecuentemente la muerte. No se requiere que el sonido CUADROS ENTRADAS Figura 2.7 Grado de actividad locomotriz en seis familias lineales de ratones. La actividad locomotriz se definió como el número de cuadros cruzados durante períodos de cinco minutos. La frecuencia indica el número de animales de cada familia que realizó un número dado de cruces. Nótense las diferencias en el grado de actividad locomotriz entre las distintas familias así como las diferencias en variabilidad de la actividad dentro de las mismas familias. (McClearn, datos sin publicar). sea fuerte; el retintín de las llaves es un estímulo efectivo para que se produzca el ataque. Este rasgo resulta estar bajo el control de varios genes. Efectos de la reproducción lineal o selectiva Como se habrá dado cuenta el lector, sin duda alguna, los que padecen ataques, o los ratones bailadores, no son adaptables. En general, la reproducción lineal tiende a resultados que se manifiestan en defectos tanto físicos como conductuales. Por cada diez crías selectivas de ratones iniciadas por los criadores, mueren por lo menos nueve y los efectos deletéreos de la reproducción lineal son evidentemente más graves aún en el hombre que en el ratón. En general, el grado de parentesco entre las parejas humanas se relaciona directamente con los efectos perjudiciales en los niños. En la población general, la tasa de mortalidad entre los niños de diez años de edad es de 24 por cada 1000. En los niños de matrimonios entre primos es de 81 por cada 1000 (Thiessen, 1972). En un estudio realizado con niños japoneses en edad escolar se descubrió que los niños de primos en primer grado son menos aptos realmente en todas las medidas (se utilizaron 17i) que los niños de control, de padres no relacionados genéticamente (Schull and Neel, 1965). Estos datos aparecen en la Tabla 2.1. En comparación con el grupo de control, los niños empezaban a caminar y hablar en edad más tardía, demostraban menos vitalidad física, alcanzaban puntajes inferiores en pruebas de inteligencia y daban menos rendimiento en la escuela. Causan mayor impresión aún los efectos perjudiciales en los niños producto de los llamados incestos nucleares (hermano con hermana, padre con hija). Adams y Neel (1967) compararon 18 de tales niños con otros de padres no emparentados, iguales en edad, peso, inteligencia y situación socioeconómica. De los niños nacidos de incesto, 5 murieron, 2 eran retardados mentales, 1 tenía el paladar hendido y 3 presentaban inteligencia mínima. Solamente 7 de los 18 parecían moderadamente normales. De los niños de control, 1 presentaba un defecto físico menor y todos los demás eran normales. El efecto devastador en la viabilidad, salud e inteligencia de los niños causado por el apareamiento entre parientes parece haber sido reconocido por la mayoría de las sociedades primitivas. Indudablemente, ninguna sociedad o cultura de que tengamos conocimiento permite el incesto. Los Faraones del Antiguo Egipto que produjeron una línea notablemente débil de gobernantes por incesto, constituyeron la excepción en su cultura. Lindzey (1967) hace la anotación interesante de que el tabú universal del incesto puede haber sido el resultado de la selección natural al actuar en las culturas humanas primitivas. Todas aquellas tribus que no impidieron el incesto desaparecieron hace mucho tiempo. Efectos de la hibridación El vigor de los híbridos está al extremo contrario de la reproducción selectiva. Cuando se juntan dos razas separadas, los hijos tienden a ser más vigorosos que los padres. Este vigor de los híbridos ha sido del conocimiento de los agricultores desde hace mucho tiempo. Todo el grano que se produce hoy es híbrido. Los cereales híbridos son más robustos y echan espigas más largas que las familias que les dieron origen. No hay evidencia clara acerca del vigor híbrido en el hombre. El autor quisiera mencionar los pueblos de Polinesia como ejemplo, pero no existen datos reales. La enfermedad de células falciformes es un ejemplo de matices especiales. En las áreas palúdicas del África los niños heterocigóticos respecto del gene de célula falciforme tienen, en cuanto a la reproducción, una ventaja sobre los individuos homocigóticos y normales. Un caso más desconcertante es el de la enfermedad de Tay-Sachs, forma devastadora de deficiencia mental infantil en el individuo homocigótico respecto del gene apropiado. Se presenta principalmente entre la población judía Ashkenázica. En forma suficientemente extraña, los individuos heterocigóticos respecto del gene presentan una ligera ventaja reproductiva (6 por ciento) sobre los normales (Dobzhansky, 1964). Tabla 2.1 Efectos de la reproducción lineal sobre la conducta de niños escolares Japoneses Promedios de Efecto de la los hijos de reproducción primos en lineal primer grado Sexo Promedio de los niños control Edad en que caminaron (meses) M F 14.06 13.62 14.19 14.07 0.13 0.45 Edad en que hablaron (meses) M F 11.81 10.38 12.60 10.82 0.79 0.44 M F 57.37 55.10 54.94 52.52 2.06 2.06 M F 58.67 57.01 55.34 53.46 2.76 2.76 M F 3.09 3.28 2.95 3.10 0.10 0.10 Características Puntajes de inteligencia Puntaje de realización Puntaje Verbal Grado Escolar: Lenguaje Datos Tomados de Schull, W. J., y Neel, J. V. Los efectos de la reproducción lineal sobre niños Japoneses. Nueva York: Harper & Row Publishers, Inc. (1965). Estudios en gemelos La mayor parte de la investigación acerca de las influencias genéticas en la conducta humana ha utilizado gemelos, comparando en general gemelos (esto es, mellizos idénticos) o monocigóticos (MZ) y mellizos o dicigóticos (DZ) del mismo sexo. La razón de tal enfoque consiste en que los gemelos MZ son genéticamente idénticos mientras los mellizos DZ no son en el campo genético más similares que los hermanos comunes. Así pues cualesquiera diferencias entre gemelos MZ deben ser completamente ambientales, mientras las diferencias entre mellizos DZ son tanto hereditarias como ambientales. En la suposición de que los efectos del ambiente sean iguales en los dos grupos, las diferencias entre los gemelos MZ pueden sustraerse de las que existen entre los mellizos DZ, y el resultado debe ser una medida de los efectos de las diferencias genéticas. En esta forma se han realizado numerosos estudios acerca de puntajes en pruebas de inteligencia como también en variables de personalidad. Normalmente los investigadores han encontrado componente genético pequeño pero confiable en las variables de la personalidad y un componente genético mayor en los puntajes de pruebas de inteligencia. En este capítulo volveremos sobre este punto más adelante. Consecuencias conductuales de los defectos Ja un solo gene Ciertos tipos de deficiencia mental grave resultan de los defectos existentes en un solo lugar genético. Por ejemplo, una enfermedad conocida como fenilquetonuria (PKU) se caracteriza por un retardo mental grave y la presencia de ácido fenilpirúvico en la orina. La fenilquetonuria resulta de la herencia de un doble gene recesivo. El individuo que es homocigótico respecto del gene produce cantidades insuficientes de una sustancia requerida para la conversión normal de la fenilalanina (aminoácido común presente en muchos alimentos de proteínas) en tirosina. Se cree que el proceso mismo de la enfermedad resulta de los aumentos de niveles en los tejidos y la sangre, del ácido fenilpirúvico y otros subproductos no-normales de la fenilalanina, los cuales actúan como toxinas en el cerebro en desarrollo. Cuando el infante afectado tiene de cuatro a seis meses de edad, ha aparecido el daño cerebral grave e irreversible. Actualmente es posible detectar la fenilquetonuria en asunto de horas después del nacimiento y prevenir la grave deficiencia mental suministrándole al niño una dieta baja en fenilalanina. La mayor parte de los hospitales examinan rutinariamente a cada niño acerca de la fenilquetonuria dentro de los dos días siguientes al nacimiento. Hoy es así mismo posible identificar a los portadores heterocigóticos del gene recesivo y advertir a tales personas mediante consejo genético acerca del peligro para sus hijos antes del nacimiento o aún de la concepción. Actualmente se sabe que se transmiten genéticamente muchas otras enfermedades metabólicas que causan deficiencia mental. Dos de ellas, la idiocia amaurótica infantil (enfermedad de Táy-Sachs) y la idiocia amaurótica juvenil resultan de la herencia de un doble gene recesivo de un solo lugar e implican mal funcionamiento del metabolismo lípido. Los efectos de la primera son notorios a los pocos meses de edad; los de la segunda entre los dos y los diez años. La corea de Huntington, caracterizada por deterioro mental progresivo como también por movimientos involuntarios, ilustra el hecho de que la deficiencia mental genéticamente causada no necesariamente aparece al principio de la vida. Esta enfermedad se transmite por un gene dominante y ataca entre los 30 y los 50 años. Consecuencias conductuales y físicas de las anormalidades cromosómicas La anormalidad en la complementación cromosómica es responsable de ciertos desórdenes mentales y físicos. El ejemplo más conocido es el síndrome de Down (llamada también mongolismo por tener sus víctimas los ojos sesgados ). Además de los ojos sesgados y otros rasgos morfológicos, las personas afectadas son marcadamente retardadas (el Cl está generalmente entre 20 y 60). El síndrome de Down que es la causa aislada determinable más común del retardo mental, tiene una ocurrencia aproximada de 1 por cada 500 a 600 nacimientos y representa el 10 por ciento aproximado de las personas de reconocido retardo mental en los Estados Unidos. Es causada por una anormalidad cromosómica llamada trisomía -21. A veces en lugar de la complementación general de 46 cromosomas (23 pares), la persona tiene 47, incluyendo un cromosoma adicional del par número 21. A veces, durante la formación de las células sexuales, se han intercambiado partes de los cromosomas 21 y 15 y la persona afectada porta el cromosoma adicional 21 como parte del par número 15. Aunque se desconocen las razones por las cuates se presenta el material genético adicional, se ha demostrado que es u. s probable que el síndrome de Down ocurra en niños de mujeres cuya edad supera el promedio gestatorio. Los efectos de las anormalidades de número y clase de cromosomas sexuales en la estructura y personalidad humanas han despertado gran interés. Normalmente un individuo de sexo masculino tiene un cromosoma X y un cromosoma Y, mucho más corto que el X. La hembra normal tiene dos cromosomas X. La persona que tiene el síndrome de Klinefelter tiene un cromosoma Y y dos X (genotipo XXY) y es fenotípicamente masculina con cierto grado de feminización, reducido impulso sexual y a veces retardo mental. Se ha demostrado que algunas "mujeres" soviéticas, atletas de habilidades físicas superiores a las de las mujeres normales, tienen el síndrome de Klinefelter, situación que crea problemas tanto de definición como de relaciones internacionales. Aunque se ha informado que muchas de las personas que tienen el síndrome de Klinefelter tienen problemas de personalidad, no es posible desenmarañar la causa y el efecto y los datos no son claros. El síndrome de Klinefelter es desconcertantemente común pues se presenta en 1 de cada 400 nacimientos. La persona que tiene el síndrome de Turner tiene un cromosoma X normal pero carece del cromosoma Y (genotipo XO). Tal individuo es fenotípicamente femenino, pero es bajo de estatura y tiene características sexuales atenuadas. Se presenta normal en inteligencia general pero presenta una deficiencia específica en discriminaciones espaciales. Tal síndrome no es muy frecuente: 1 caso en 3000 nacimientos. Tal vez la que causa mayor curiosidad entre las anormalidades cromosómicas sexuales es el síndrome XYY (p.33). Los individuos de genotipo XYY tienden a ser masculinos, de gran estatura, musculados, retardados, de mucha agresividad. El síndrome se encontró inicialmente entre prisioneros y muchos estudios han reducido sus muestras a prisioneros de gran estatura y con antecedentes de episodios violentos-limitación que obviamente introduce un prejuicio de muestreo. El hecho es que las investigaciones realizadas con tales prisioneros revelan en su mayoría una incidencia de genotipo XYY de un 2 a un 12 por ciento. En la población normal la incidencia resulta ser de un 0.1 por ciento aproximado (McClearn, 1970; Jarvik y otros, 1973). A pesar de lo inadecuado de los datos hay cierta base para creer que un cromosoma Y adicional aumenta tanto las características masculinas como las agresivas. No es difícil extrapolar de este síndrome extremo a influencias genéticas mucho más sutiles del cromosoma Y normal en la conducta a evasiva. En general, los números anormales de cromosomas sexuales dan como resultado anormalidades de orden tanto físico como conductual. Sería sorprendente lo contrario. La investigación que se propone correlacionar la anormalidad cromosómica con la desviación conductual acaba de empezar y hasta el momento se ha ocuparlo tan sólo de adiciones y remociones cromosomas completos. Existen sin duda alguna, muchas anormalidades a nivel de genes individuales que todavía no logran detectarse con el microscopio. Característicos conductuales poligánicas Cuando una propiedad o característica presenta en una población una distribución continua o cuantitativa, ello se debe probablemente a ocurrencias que se presentan en más de un lugar genético. Es un problema poligénico. La genética cuantitativa se ocupa del estudio de tales características. El enfoque básico consiste en determinar el grado de relación o correlación con medidas cuantitativas tales como el CI o las pruebas de personalidad para individuos emparentados entre sí o carentes de tal relación. Las comparaciones más importantes han sido las de gemelos (MZ), las de simples mellizos (DZ) y las de individuos sin parentesco. Los resultados de tales comparaciones indican con suma claridad que rasgos tales como la esquizofrenia, la psicosis maníacodepresiva y la inteligencia son altamente heredables (114cClearn and DeFries, 1973). Esquizofrenia. Según los datos es cada vez más claro que la esquizofrenia es un mal genético. La esquizofrenia es la forma de enfermedad mental de mayor generalización y gravedad. Se presenta en un 1 por ciento aproximado de la población en todos los países y culturas del mundo, sin importar su carácter avanzado o primitivo. La esquizofrenia se caracteriza por varios grados de desorden en los procesos del pensamiento, por alucinaciones y delirios y desintegración de la personalidad. Aunque no hay acuerdo acerca de los criterios exactos de diagnóstico, la característica más ampliamente compartida es la del pensamiento desordenado. Actualmente no tenemos conocimiento de las posibles anormalidades químicas, nerviosas o biológicas que dan como resultado la esquizofrenia; el hecho es que no hay duda de que existen. Aunque los factores ambientales, culturales y sociales pueden ser importantes, particularmente en el desencadenamiento de episodios esquizofrénicos, la enfermedad es fundamentalmente psicobiológica. La gran mayoría de los estudios de gemelos idénticos versus fraternos ha indicado una concordancia mucho mayor en la esquizofrenia de los gemelos idénticos. Los valores típicos van de 42 a 86 por ciento para gemelos 9 a 15 por ciento para mellizos fraternos ( McClearn, 1970). Así pues, si un gemelo idéntico resulta con esquizofrenia hay por lo menos iguales posibilidades de que en el otro gemelo aparezca también la enfermedad. Un estudio más convincente de la heredabilidad de la esquizofrenia en niños adoptados, sin parentesco de por medio, fue realizado por Heston (1966). Identificó 47 niños de madres a quienes habían diagnosticado esquizofrenia en un hospital estatal. Los niños fueron separados de sus madres en los primeros días después de nacidos y colocados en hogares adoptivos. Se utilizó cerro control un grupo de niños adoptados a la misma edad, provenientes de madres libres de esquizofrenia. De los 47 niños de madres esquizofrénicas, 5 llegaron a tener la enfermedad y la mitad presentaron claras inhabilidades psicosociales. El grupo de control no presentó esquizofrenia y la incidencia de dificultades psicosociales resultó de un nivel mucho más bajo. Aunque hemos caracterizado la esquizofrenia como un desorden causado por anormalidades poligénicas, no todas las autoridades en la materia están de acuerdo con tal definición. Algunos consideran que puede explicarse por un solo gene recesivo; otros creen que representa diversas anormalidades de diferentes genes, diagnosticadas todas como esquizofrenia; algunos también opinan que es el resultado de defectos de dos o más genes. Finalmente, otros persisten en discutir hasta qué punto la esquizofrenia tiene una base claramente genética (ver McClearn and DeFries, 1973, acerca de tales posibilidades). Inteligencia. Acalorados debates se han llevado a cabo en los últimos años con relación al grado en que se hereda la inteligencia. Buena parte de la emotividad radica en la afirmación de que existen diferencias raciales genéticas en materia de inteligencia. Estos dos problemas deben separarse. Un repaso a los datos puede conducir solamente a la conclusión de que la inteligencia medida por pruebas típicas de inteligencia, está significativamente determinada por factores genéticos. Por el contrario, no estamos en posibilidad actualmente de interpretar los datos relacionados con la raza y la inteligencia. En cualquier estudio acerca de la base genética de la inteligencia, deben establecerse ciertos puntos que son evidentes. Primero, la inteligencia es una idea-una construcción teórica que tiene casi tantos significados como autoridades han escrito al respecto. Para medir la inteligencia tenemos que medir algún tipo de ejecución. Las medidas de utilización más generalizadas son las pruebas típicas de CI tales como la Stanford-Binet. La inteligencia medida por tal prueba de CI no es ni más ni menos que un puntaje de prueba. No tiene valor independiente, pero sí tiene relaciones y correlaciones con otras clases de ejecución. Segundo, el puntaje del Cl es un puntaje general de prueba que "per se" no especifica clases determinadas de habilidades. No es necesariamente un pronóstico de éxito en la vida. Muchas personas de éxito no tienen un CI alto y muchos individuos que sí lo tienen, fracasan estruendosamente. Hay sin embargo una gran relación general entre la ejecución de una prueba de Cl y el desempeño en la vida. Quien tiene un nivel de CI muy inferior al normal no logrará en la mayoría de las situaciones lo que consigue una persona normal. La correlación entre el Cl y los grados escolares es de un 0,5 aproximado-relación significativa pero no extraordinariamente estrecha. La relación general entre el CI y el éxito adaptativo aparece indicada en el estudio de largo alcance acerca de niños superdotados que inició Terman en 1921. Hizo el seguimiento de 1500 niños, todos los cuales tenían un CI de 140 o superior. Como grupo estaban definitivamente por encima del dio en salud, desarrollo, éxito literario y científico, situación económica, y no diferían del promedio en cuanto a salud mental o ajuste emocional. Tercero, el ambiente influye de manera importante en la ejecución de una prueba de CI. Si un individuo se somete a prueba en diversas ocasiones, durante un período de años, no son raras las fluctuaciones de 20 puntos en el CI. Los niños que pasan de un ambiente pobre a otro mejor presentan puntajes superiores de CI. Una parte importante de la ejecución de una prueba de CI depende de la experiencia cultural y del aprendizaje. Muchos puntos de las pruebas típicas de CI suponen familiaridad con la cultura occidental de la clase tedia. Se supone que los niños están familiarizados con cosas tales como el dinero, los almacenes, los carros y las casas. Otro factor de importancia decisiva es la maduración. Los niños que sufren de desnutrición crónica realizan las pruebas del CI en forma deficiente. Ninguno de estos factores ha de causar sorpresa. La prueba del CI es una medición de ejecución, la cual está bajo el influjo de todos los factores que inciden en el desempeño humano. Hechas todas estas aclaraciones, ¿tiene algo de sorpresa realmente que los puntajes de pruebas de Cl indiquen heredabilidad altamente significativa? El hecho de indicarlo así sugiere que los factores genéticos desempeñan un papel muy importante en la determinación de la ejecución de las pruebas de CI. En la Tabla 2.2 aparece un resumen de los datos acerca de la inteligencia de gemelos idénticos (MZ) y fraternos (DZ). Los gemelos idénticos que se crían juntos son idénticos en el desempeño escolar y casi idénticos en CI. La correlación de CI sigue siendo alta cuando se crían separadamente (por lo menos a partir de los seis meses de edad), pero el desempeño escolar es irás variable. Hay la crítica común a tales datos de Tabla 2.2 Correlaciones de fenotipos de fenotipos físicos y conductuales Datos tomados de Burt (1966) según se citan en McClearn (1970). Reimpreso con autorización, de "Behavioral Genetics" por G.E. McClearn, Ann. Rev. Genetics 4. Copyright © 1970 por Annual Reviews, Inc. Reservados los derechos. que los gemelos idénticos MZ criados separadamente van a hogares adoptivos de nivel socioeconómico similar. Burt (1966) hizo la evaluación del caso correlacionando la situación ocupacional de las dos familias en que se criaba cada uno de los gemelos idénticos MZ. La correlación resultó ligeramente negativa. Así pues, la alta correlación de CI respecto de gemelos idénticos MZ, criados separadamente, no puede explicarse por niveles socioeconómicos similares de los hogares adoptivos: no había similitud entre los niveles socioeconómicos. Según trabajos recientes acerca de la base genética de la inteligencia resulta reforzada la heredabilidad de diferentes clases de habilidades. Las habilidades que, definidas ampliamente, parecen presentar una fuerte determinación genética son las que se miden por las subpruebas de información, vocabulario, ordenamiento de: dibujos, símbolos de dígitos de la Escala de inteligencia de Wechsler para niños (WISC), prueba de CI de amplia utilización. Por el contrario, los resultados de las subpruebas de Memoria y Razonamiento demostraron que tales habilidades tenían poca o ninguna heredabilidad (Vandenberg, 1967, 1968). Estos resultados se basan en un análisis estadístico cuidadoso de la ocurrencia de una diferencia significativa entre gemelos idénticos MZ y mellizos fraternos DZ en las diversas subpruebas. En cuanto a los factores genéticos subyacentes, el trabajo de Vandenberg sugiere que hay por lo menos cuatro unidades genéticas diferentes las cuales corresponden a las cuatro habilidades heredables. El número de componentes genéticos varía sin embargo con el tipo de prueba que se utilice. Los cálculos del número total de lugares genéticos implicados en la inteligencia, varían de 3 ó 4 a más de 70. Evidentemente el problema no se ha resuelto. Por el contrario, los puntajes extremadamente altos de Cl pueden estar bajo el control de genes aislados y los CI anormalmente bajos pueden así mismo representar acciones de genes independientes, como en el caso de los niños aquejados de fenilquetonuria. Un problema de interés potencial para la evolución futura del hombre se relaciona con la posibilidad de que la inteligencia esté declinando en la raza humana. Muchos estudios han demostrado que existe una relación inversa significativa entre el CI y el tamaño de la familia. Aparece la conclusión de que continuamente nacen más individuos de Cl inferior, lo cual conduce a calcular que la declinación de CI por generación alcanza un promedio de 2 a 4 puntos. Dado que la humanidad ha existido en la forma del moderno Homo sapiens durante dos mil ;veneraciones por lo menos, o nuestros antepasados remotos eran increíblemente inteligentes o hay algo errado en la inferencia. De hecho, los datos provenientes de pruebas de largo alcance realizadas con niños no indican disminución alguna del CI. La explicación aparece en la Tabla 2.3. Los individuos de bajo Cl tienen en forma significativa menos niños: muchos de CI extremadamente bajo se hallan hospitalizados desde la niñez y no tienen oportunidad alguna de aparearse. Haciendo cuentas, el Cl de la población como uno todo se presenta relativamente estable. Importancia de la estructura genética para la conducta La estructura genética influye en cierto número de características conductuales humanas. En el caso de graves perturbaciones mentales tales como la esquizofrenia o el síndrome de Down, la estructura genética parece desempeñar un gran papel. Pero ¿hasta qué punto explica la variabilidad de las características conductuales humanas en la población normal? Obviamente es imposible una respuesta a esta pregunta pero todas las evidencias indican que su papel puede no ser dominante. En la evolución de los mamíferos, especialmente del hombre, la selección natural ha reforzado la plasticidad conductual. El hombre ha sido seleccionado en parte por razón de su habilidad para cambiar rápidamente su conducta según los cambios ambientales de menor cuantía: Más aún, el hombre recuerda la efectividad de los cambios conductuales. En un sentido muy real, la presión' selectiva sobre el hombre se ha ejercido sobre su habilidad para aprender. Tabla 2.3 Tasas de Reproducción de sujetos con diferente C.I. C.I. Número 0-55 56-70 71-85 86-100 101-115 116-130 131+ 29 74 208 583 778 269 25 1.4 2.5 2.4 2.2 2.3 2.4 3.0 No. niños Porcentaje Datos tomados de Higgins et. al. (1962). Reimpreso con autorización, de "Behaviord Genetics" por G. E. McClearn, Ann. Rey. Genetics 4. Copyright © 1970 por Annual Reviews, Inc. Reservados todos los derechos. DESARROLLO DEL SISTEMA NERVIOSO El crecimiento, el desarrollo y el avance físicos se hallan bajo fuerte control genético desde el momento en que el huevo es fecundado hasta que muere el organismo. El ambiente desempeña así mismo un papel. Al nacer, crece bruscamente la importancia del ambiente externo. Con anterioridad a tal evento, el ambiente que rodea a la madre tiene poco efecto sobre el feto en desarrollo, mientras no se presente una catástrofe de proporciones considerables. Pero se considera de -gran importancia el ambiente intrauterino del embrión desde el momento de la concepción, aunque actualmente se sabe poco acerca de su influencia en su desarrollo tanto normal como anormal. Ocho días aproximadamente después de fecundado, el huevo humano se prende a la pared del útero y se divide en dos células. A partir de este momento se acelera la tasa de multiplicación celular. El embrión se divide en tres capas germinales de células-endodermo, ectodermo y mesodermo. Las células del endodermo forman eventualmente órganos internos, las del mesodermo forman los músculos del esqueleto y las del ectodermo se convierten en piel y sistema nervioso. El ser humano se denomina embrión durante los dos primeros meses y feto desde ese entonces hasta el nacimiento. El corazón empieza a latir a finales del primer mes, el riñón comienza a excretar orina hacia el tercer mes y alrededor del quinto mes se inician movimientos vigorosos del esqueleto-el feto patea, se retuerce y salta. Los pulmones se desarrollan temprano pero están llenos de fluido y carecen esencialmente de función hasta el nacimiento. Un niño prematuro, con ayuda puede sobrevivir si tiene al menos siete meses y un peso aproximado de cinco libras. Desarrollo embrionario Diferenciación de las células. El crecimiento y diferenciación del sistema nervioso ilustra el hecho de que en todas las etapas del desarrollo tanto la estructura como la conducta representan una interacción compleja del conjunto genético y el ambiente. Al empezar la vida de un embrión, las células de los tejidos destinados a convertirse en sistema nervioso son de aspecto semejante al de otras células. Luego, durante el proceso por el cual las células embrionarias se dividen en las capas del endodermo, mesodermo y ectodermo (Figura 2.8) se forma una estructura de tejido mesodérmico en forma de bastón, inmediatamente debajo de una parte del ectodermo y se alarga desde la cabeza casi hasta el otro extremo del embrión. Este bastón axial, denominado notocordio sirve como operador para la diferenciación de las células ectodérmicas de encima en tejido nervioso. Sin el notocordio las células ectodérmicas permanecen sin diferenciarse. Si debajo de otros tejidos se coloca un trozo de notocordio-por ejemplo, debajo de presuntos tejidos estomacales -empieza a formarse un segundo sistema nervioso. A su vez, un presunto tejido nervioso puede convertirse en tejido estomacal si se le trasplanta en forma apropiada. Por lo tanto, el sistema nervioso en su etapa propiamente inicial depende del ambiente. Figura 2.8 Desarrollo inicial del canal y el tubo neural en el embrión humano. A) El comienzo del canal neural se forma a partir del envolvimiento del tejido ectodérmico. B) Desarrollo posterior del canal neural. C) El canal neural ha comenzado a cerrarse. D) Continuación del cierre del canal neural, a partir de la línea- media en dirección caudal y rostral. E) Vista lateral del embrión mostrando el cierre completo del tubo neural. (Adaptación de Buchanan, A.R. Functional Neuroanatomy. 4a. ed. Filadelfia: Lea ó: Febiger, 1961). En esta etapa se dice que las células son indeterminadas en cuanto a que se las puede hacer desarrollar en diversas direcciones. Una vez que tales células entran en contacto con el notocordio, ocurre la determinación y se convierten en tejido nervioso aún en el caso de que se les trasplante. El proceso por el cual el notocordio causa la determinación se denomina inducción: el notocordio induce al ectodermo superpuesto a convertirse en tejido nervioso. La inducción, aunque al respecto es poco lo que entendemos, se presenta como uno de los fenómenos más fundamentales del desarrollo. Actúa siempre para limitar o especificar las oportunidades de desarrollo de un grupo de células mediante su interacción con otro grupo. Al determinarse más y más, las células se diferencian lentamente en tejidos y sistemas orgánicos que constituyen el organismo adulto. La diferenciación es pues todo este proceso durante el cual un grupo relativamente no especializado de células se moldea en corazón, riñón, estómago, músculo o cerebro. Etapas del desarrollo. En 1842 Karl Ernst von Baer, teórico evolucionista predarwiniano, estableció el principio de que las primeras etapas de los animales superiores se asemejan a las primeras de los inferiores. Dos décadas más tarde, Ernst Haeckel replanteó este principio con el concepto de que un individuo durante su ontogenia (crecimiento) atraviesa una serie de formas que se asemejan a sus antepasad -)s adultos; de aquí la expresión "la ontogenia recapitula la filogenia". La frase implica que la historia de la evolución (filogenia) puede estudiarse directamente a través del desarrollo de un organismo individual. La evidencia actual indica que von Baer tenía razón hasta cierto punto: los animales del mismo filum se desarrollan generalmente en forma similar hasta el punto en que se apartan según sus propias especializaciones. Las bolsas branquiarias de un embrión de mamífero nunca se desarrollan hasta llegar a ser branquias funcionales como las de un pez adulto, pero claramente se asemejan a las bolsas branquiarias de un embrión de pez. En forma similar, el desarrollo cerebral de un embrión humano y el de un pez, anfibio o pájaro, guardan entre sí notable semejanza hasta un momento en que cada uno empieza a desarrollar sus propias especializaciones. Las primeras etapas de la formación del sistema nervioso son perfectamente similares en todos los embriones vertebrados (Figura 2.8 y 2.9). Poco después que el ectodermo superior entra en contacto con el notocordio, una placa nerviosa se convierte en rasgo prominente de la superficie superior. Gradualmente se forma un canal longitudinal en la parte media del embrión y se extiende lentamente hacia ambos extremos mientras se elevan los bordes de la placa. Los bordes se elevan progresivamente y el canal se profundiza hasta que se encuentran y funden sus bordes, formando el tubo nervioso (Fig 2.8). El cierre ¿,el tubo, así la formación del canal . parte media del embrión hacia ambos extremos en forma simultánea. Al cerrarse el tuco, ciertos grupos pequeños de células que hay en los bordes no se incorporan en el tubo y se apartan formando así la cresta nerviosa. En sus primeras etapas, el canal nervioso está constituido por una simple capa de células que se dividen rápidamente. Al cerrarse el tubo nervioso, la división celular ocurre todavía primariamente en el espacio interno del tubo, pero un número creciente de células nuevas empieza a emigrar hacia la parte externa del tubo, formando así tres capas diferentes. La capa interna contiene células que están todavía dividiéndose. La capa media consta de cuerpos celulares que están empezando a formar la materia gris o regiones nucleares del sistema nervioso. En la mayoría de los casos tales células nunca vuelven a dividirse; esta región contiene las mismas células que se encuentran en el organismo adulto. La capa externa permanece temporalmente libre de células y sólo la forman las fibras de las células en las dos zonas restantes. Pronto sin embargo, en algunas áreas del cerebro, dicha capa llega a contener células que han continuado su emigración. Desde que empieza a formarse la placa nerviosa, es más amplio su extremo anterior (frente) y al cerrarse el tubo aparecen tres combas denominadas las tres regiones cerebrales primarias (Figura 2.9). La región más delantera, el prosencéfalo, da origen a la corteza cerebral y a otras regiones cerebrales superiores (Figura 2.9; ver también el Capítulo 3 donde se explican estos términos anatómicos). El mesencéfalo permanece sin dividirse y da origen al mesencéfalo adulto. El romboencéfalo da origen al cerebelo y regiones inferiores del tallo cerebral. Por medio del crecimiento diferencial en estas regiones, el cerebro adquiere lentamente su forma adulta. Aún en estas primeras etapas, las partes del cerebro no son completamente independientes entre sí y dependen de las estructuras que las rodean para inducir su diferenciación normal. Por ejemplo, si en las primeras etapas se Figura 2.9 Desarrollo del cerebro humano desde la tercera semana hasta el nacimiento, vista lateral. A- En el embrión de tres semanas, las tres regiones primarias del cerebro se encuentran ya formadas. B• En el embrión de cuatro semanas el cerebro anterior, el cerebro medio y el posterior han incrementado su desarrollo. C- Embrión de siete semanas; aquí el cerebro anterior ya se ha diferenciado en telencéfalo y diencéfalo y el cerebro posterior en metencéfalo y mielencéfalo. D- En el feto de once semanas el cerebelo está claramente diferenciado. E- El feto de seis meses presenta un alto desarrollo de los hemisferios cerebrales. F- En el niño recién nacido el surco y el giro están ya bien definidos en los hemisferios cerebrales. (Adaptado de Patten, B.M: Human Embriology, 2a ed. New York: McGraw-Hill, 1953). remueve completamente la parte posterior, se rota 180 grados y se vuelve a colocar en tal forma que la parte anterior quede en contacto con el cordón espinal y el extremo posterior empalme con el mesencéfalo, se desarrolla un cerebro normal. Estas células pueden ser inducidas por su ambiente a formar regiones nucleares completamente distintas que tengan funciones diferentes. Mientras la división celular está todavía ocurriendo en forma acelerada en la superficie, las neuronas migratorias empiezan a conformarse en grupos, constituyendo los núcleos del cerebro adulto. Durante esta fase del desarrollo, la diferenciación se está llevando a cabo aun ritmo acelerado en todo el sistema nervioso. No solamente se están formando los núcleos cerebrales; los tractos de fibras están igualmente comenzando a conectar los núcleos; algunas de las células de la cresta nerviosa se están conformando en ganglios espinales, enviando sus fibras a la médula espinal y hacia fuera, en dirección a la piel; y los tallos ópticos han crecido fuera del diencéfalo para formar las copas ópticas en sus extremos. Estas copas ópticas inducen la formación de una lente a partir del ectodermo superpuesto. El ojo se forma por lo tanto directamente del tejido cerebral. Regulación del desarrollo. Gran parte de esta diferenciación aparece autocontrolada. Los movimientos musculares empiezan aun faltando la entrada sensorial y los núcleos cerebrales han de madurar aislados de sus respectivos órganos sensoriales motores. Sin embargo, después de establecido el modelo básico del sistema nervioso su periferia adquiere cada vez mayor importancia. Por ejemplo, los centros nerviosos se forman con cierto exceso de células en comparación con lo que se observa en el organismo maduro. Por el tiempo en que una región forma conexiones con la periferia (esto es, órganos y receptores que están fuera del cerebro), se degeneran muchas de las células de tal manera que el número final es el normal. Sin embargo, si se remueve el órgano periférico (por ejemplo, el ojo) se degenera un número todavía mayor de células en tal forma que la región cerebral se atrofia (se contrae); a la inversa, si se amplía en alguna forma el órgano periférico más allá de su tamaño ordinario, se degeneran menos células de lo normal. Este efecto se ha demostrado en el tallo cerebral y médula espinal respecto de los núcleos tanto motores como sensores en muchos vertebrados. Es interesante anotar que el momento en que la periferia empieza a tener esta influencia en el desarrollo nervioso aparece en correlación con el momento en que los órganos periféricos se hacen funcionales. Esta observación no permite sin embargo la conclusión de que en todos los casos es necesaria la función para el establecimiento de las conexiones nerviosas. Si se hacen crecer partes del sistema nervioso en cultivo de tejidos, se forman conexiones entre muchas neuronas. El papel de la actividad funcional en los sistemas sensorial y motor en la regulación del desarrollo futuro del sistema nervioso y de la conducta es actualmente uno de los puntos más controvertidos en la neurología del desarrollo: Según unos, existe primordialmente una relación unidireccional en la cual el desarrollo del sistema nervioso no depende de la retroalimentación proveniente del ambiente; es algo preprogramado para desarrollarse de manera autónoma siguiendo un curso determinado. Según otros, se trata de una relación recíproca en la cual el desarrollo del sistema nervioso es modulado continuamente por estímulos ambientales y el desarrollo normal del sistema nervioso depende de esta modulación. Víctor Hamburger y sus colegas, experimentando con la conducta de embriones de pollo, han destacado los aspectos autónomos del desarrollo de la conducta. Comenzando por la descripción cuidadosa de los tipos y tasas de movimiento en cada etapa del desarrollo anterior al nacimiento, estos autores lograron describir los efectos de las diversas operaciones. Por ejemplo, removiendo parte de la médula espinal o seccionándola cerca del cerebro, encontraron que cada nivel de la médula puede iniciar conductas en las estructuras corporales asociadas. El principal efecto de estas operaciones era el de reducir la cantidad total de actividad sin cambiar sustancialmente el patrón de movimientos. La influencia de la estimulación sensorial de parte de los músculos y de la piel se estudió en experimentos en los que se hizo una deaferentación. La remoción de las entradas sensoriales de la piel y de los músculos no afecta seriamente los patrones de conducta a partir del tercer día embrionario, tiempo en el que se inicia la conducta hasta el día 17, tres días antes del nacimiento. Aunque estos experimentos no logran eliminar los efectos de la estimulación temprana en los patrones de conducta de la incubación o posteriores a ella, ciertamente muestran que buena parte del desarrollo de la conducta puede ocurrir en ausencia de la estimulación sensorial. Las influencias hormonales en el desarrollo cerebral y conductual son de importancia decisiva. Varios estudios han demostrado que el exceso de la hormona de la tiroides, la tiroxina, estimula en forma anormal la maduración prematura de algunas áreas nerviosas y el desarrollo anticipado de muchos reflejos. Una cantidad muy reducida de tiroxina al principio del desarrollo puede conducir a efectos devastadores como demora en la maduración de muchas conductas reflejas, crecimiento nervioso reducido, patrones anormales de ondas cerebrales y reducida habilidad de aprendizaje. Los efectos de la tiroxina dependen en su mayoría, del suministro adecuado de la hormona durante un período decisivo específico del desarrollo del organismo (en la rata, los primeros 15 días posteriores al nacimiento). Si por este tiempo la hormona no se halla presente, no pueden remediarse las deficiencias. (Esto contrasta con la situación en la edad adulta. Los déficits conductuales producidos por la remoción de la glándula tiroides en un adulto pueden remediarse por medio de prescripción hormonal.) La presencia de hormonas sexuales masculinas, andrógenos, durante un período decisivo del desarrollo embrionario, influye en los procesos nerviosos que determinan la aparición de características conductuales masculinas o femeninas. Una de las demostraciones más dramáticas fue realizada por Young y sus colaboradores inyectándoles andrógenos a tres monas durante la preñez. En tres de tales casos la cría genéticamente femenina resultó seudohermafrodita (poseían además de los femeninos, órganos sexuales masculinos incompletos). Tales experimentos se comentarán más ampliamente en el Capítulo 9. Desarrollo postnatal Al nacer, el niño posee cierto número de conductas reflejas y un cerebro en situación de funcionamiento amplio si no completo. Sin embargo, los reflejos no se hallan aún bajo el control de la corteza cerebral, la más alta región del cerebro. Por ejemplo, el reflejo de prehensión es muy fuerte por la época del nacimiento: un niño recién nacido puede sostener su peso completo, cogido de una barra. Tal reflejo se debilita considerablemente, sin embargo, un mes más tarde. Un reflejo de fuerza semejante se encuentra en el mono recién nacido, pero conserva su fuerza y aparentemente queda bajo control voluntario unos 20 días después del nacimiento. Tal reflejo es decisivo para la supervivencia de los monos, como también sin duda alguna de los primeros antepasados del hombre. Si el recién nacido no logra prenderse del pelo de su madre mientras cuelga de ella, pierde su posición cuando ella se desplaza y al. caer puede perder la vida. Las ondas cerebrales generadas en la corteza cerebral (Capítulo 4) pueden registrarse desde el cráneo, que cubre el cerebro del recién nacido. De hecho, las ondas cerebrales del feto pueden registrarse aún desde el abdomen de a :padre. Sin embargo, o, la maduración estructural de la corteza cerebral requiere varios años y la corteza cerebral y otras estructuras cerebrales superiores cambian continuamente durante toda la vida del hombre. CONTROVERSIA DE NATURA Y NURTURA La psicobiología del desarrollo es el estudio del crecimiento y desarrollo del cerebro y la conducta. Dado que todos los organismos crecen de células individuales, todos los aspectos de la biología y la conducta tienen una historia del desarrollo. Una investigación completa de la psicobiología del desarrollo supone una revisión total de la psicobiología. En parte por esta razón, muchos psicólogos consideran que de suyo el desarrollo no constituye realmente un campo distinto de estudio. Antes bien, si alguien se interesa por determinado fenómeno tal como el aprendizaje instrumental, el desarrollo de la conducta como resultado del crecimiento y de la experiencia es simplemente parte del asunto. Muchos psicólogos biológicamente orientados sostienen un punto de vista opuesto; tienden a favorecer la noción de que una parte importante del desarrollo conductual es preprogramada, esto es, que depende del crecimiento y desarrollo del sistema nervioso. Determinadas clases de aprendizaje pueden ocurrir solamente cuando los sustratos cerebrales y corporales apropiados se han desarrollado. Viene al caso el siguiente pasaje de Daniel Lehrman: Un gatito recién nacido puede rehuir un choque eléctrico y puede presentar una respuesta motora a un sonido; pero los intentos de desarrollar una asociación condicionada simple entre el sonido y la retirada al choque no tienen éxito mientras el animalito no haya cumplido dos semanas. Sería sin embargo prematuro concluir de aquí que el gatito no puede aprender gran cosa durante sus primeros 10 días. Mi colega Jay Rosenblatt, al observar los gatitos con sus madres, notó que cuando aquellos tienen 2 ó 3 días de nacidos, cada gatito se alimenta de determinado pezón de la madre y tiene un camino definido para llegar hasta él. En ciertas circunstancias puede demostrarse que cuando el gatito tiene de 7 a 8 días de edad, ha desarrollado lo que tiene visos de derechos de territorialidad en torno al pezón; muestra cierta tendencia a colgarse del pezón de "su pertenencia" cuando un compañero lo provoca, pero suelta el pezón equivocado si se había prendido a este último. Esa es pues una clase de aprendizaje de cierta importancia y complejidad, que puede observarse en el gatito durante sus primeros días de vida. La diferencia que existe entre el científico que no puede demostrar aprendizaje en un gatito antes de cumplir éste sus diez días y el que puede demostrarlo cuando el animalito tiene tres días de nacido, es, considero yo, la diferencia que existe entre el investigador que se empeña en saber cuándo puede aprender el gatito algo que el experimentador desea que aprenda y aquél que brega por descubrir cuándo puede aprender el gatito algo que el gatito quiere aprender. (Leherman, 1971, p. 465) Estos conceptos encontrado, acerca de la importancia del crecimiento y desarrollo tienen sus orígenes en la controversia de natura y nurtura. Los psicólogos conductistas tienden a recalcar la importancia de la experiencia pasada y del aprendizaje; los psicobiólogos destacan el papel del crecimiento y el desarrollo genéticamente controlado del organismo. Muchos estudiantes del desarrollo quisieran hoy enterrar el problema de la natura y nurtura. Así, los organizadores de un importante simposio acerca de la psicobiología del desarrollo (Tobach, Aronson, and Shaw, 1971) esperaban evitar "la conceptualización de natura y nurtura, por ser una falsa dicotomía". Se desanimaron sin embargo con "el problema" cuando se desató durante la conferencia una polémica "acerca de la herencia y el ambiente, lo cual no se había programado". La razón de que persista el problema de la natura y nurtura es simple. Es quizá el más fundamental e importante de todos los interrogantes humanos. Los filósofos y eruditos lo han debatido desde los albores de la historia. Constituye una diferencia de marcada realidad. Si la conducta violenta y agresiva de los humanos es primordialmente aprendida a través de la experiencia, puede controlarse por medio de un aprendizaje adecuado. Si es primordialmente genética, el control social debe ser completamente diferente. Los individuos genéticamente violentos deben ser aislados (por ejemplo, en prisiones) de sus eventuales víctimas. Cualquier rehabilitación social bien intencionada será inútil. La violencia no puede eliminarse de la raza humana sino por medio de técnicas genéticas - por medio de apareamiento selectivo (eugenesia) o, por medio de la ingeniería genética (alteración física directa de los genes, del plasma germinal básico en sí mismo). La enfermedad mental proporciona otra buena explicación al hecho de que no haya todavía desaparecido la controversia acerca de la natura y la nurtura. Actualmente se favorece el concepto de que muchas formas de neurosis, desde condiciones benignas tales como el excesivo rubor hasta temores graves y persistentes que pueden incapacitar totalmente a una persona, son primordialmente objeto de aprendizaje a través de la experiencia. Presumiblemente pueden tratarse mediante la aplicación de principios y técnicas de aprendizaje. Por otra parte, las psicosis graves se presentan como ampliamente genéticas. El tratamiento y prevención de la esquizofrenia no pueden realizarse únicamente por medio de terapias de aprendizaje. Deben utilizarse métodos biológicos, bioquímicos y finalmente genéticos. Muchos problemas fundamentales de psicología y psicobiología pueden remontarse al campo de la natura y la nurtura en relación con el desarrollo humano. Esto virtualmente se aplica a todos los procesos psicológicos complejos tales como la percepción, la integración sensoriomotora, los procesos del pensamiento y aún el lenguaje. Un concepto extremista presenta el cerebro del infante como una pizarra limpia, una tabula rasa en que la experiencia escribe lo que será la mente humana. En el extremo contrario está el concepto de que la mente humana del adulto viene preprogramada en el cerebro del infante para desenvolverse y desarrollarse con el cree el siento, independientemente de la experiencia. La verdad como toda verdad, se halla en algún punto medio de los dos extremos. Exploraremos con mayor amplitud estos problemas del desarrollo en los capítulos pertinentes. La objeción principal a este problema de la natura y la nurtura es la que lo califica de extremada simplificación. Ningún rasgo humano es completamente genético y ninguno es tampoco fruto exclusivo de la experiencia. Los genes y el ambiente interactúan por lo menos desde el momento en que el huevo es fecundado. Los criminales violentos no son siempre violentos; los esquizofrénicos no andan siempre desorientados. La vida es toda ella, una interacción continua entre el plasma germinal y el mundo. La psicobiología moderna se interesa más por investigar todas las fuentes de la conducta que por escoger entre la natura y la nurtura. El análisis, enfocado a la heredabilidad de las características como lo practica la genética conductual contemporánea, proporciona una imagen más realista de la importancia relativa de las influencias hereditarias y ambientales y de sus interacciones, en el crecimiento y desarrollo de las habilidades, conducta y personalidad del hombre. Nos encontramos sin embargo muy lejos todavía de entender la base genética aún de conductas tan simples y fijas como el patrón de canto del grillo. No somos capaces todavía de describir la cadena de fenómenos que van desde el gene hasta el canto del grillo y eso para no mencionar la que va del gene a las conductas humanas complejas, como la violencia, la esquizofrenia o la inteligencia. RESUMEN Los genes son las unidades estructurales de la herencia. Mediante la determinación de las estructuras de las proteínas producidas en los ribosomas, los genes determinan las estructuras y funciones de un organismo. Los genes interactúan así mismo con el ambiente. El genotipo de un organismo expresa su estructura genética subyacente. El fenotipo es la expresión externa de la acción del gene, como resultado de la interacción con el ambiente. Las tres hipótesis formuladas por Mendel constituyen las leyes básicas de la herencia: (1) la herencia se transmite por unidades independientes y heredables (los genes); (2) cuando cada uno de los padres aporta la misma clase de unidad, los hijos manifiestan así mismo esta característica, pero cuando cada padre aporta una clase diferente de unidad, se forma una cría híbrida; y (3) la experiencia de un organismo no afecta los genes. El DNA es la sustancia fundamental de los genes. Se compone de desoxirribosa, fosfato y cuatro bases: adenina, guanina, timina y citosina. Watson y Crick propusieron el modelo de doble hélice para la estructura del DNA. La molécula está compuesta de dos ramales, envueltos el uno sobre el otro. Las bases adhieren al sostén de cada ramal y luego se ligan entre sí. Aunque las bases pueden presentarse en cualquier secuencia a lo largo de un solo ramal, hay una relación constante entre los dos ramales complementarios de tal manera que la adenina se liga siempre con la timina y la guanina con la citosina. En dicha secuencia de las bases está contenido el código genético. La capacidad del DNA para reproducirse por sí mismo depende de las combinaciones específicas de ligazón entre las bases. El mecanismo de traducción del código genético en proteína envuelve RNA. El RNA mensajero transmite el código del DNA al ribosoma. En el ribosoma, el RNA de transferencia actúa para reunir los aminoácidos requeridos para la elaboración de las proteínas codificadas por el DNA.. Los genes para un rasgo específico se presentan en lugares específicos de determinados cromosomas. Cada par de cromosomas contiene lugares homólogos. Un cigote se caracteriza como homocigótico si se hereda el mismo alelo en determinado lugar de ambos cromosomas. Se dice que es heterocigótico cuando dos alelos diferentes se hallan en lugares homólogos. Varios procesos diferentes contribuyen a la variabilidad de la integración genética. La complementación aleatoria durante la reproducción, la mutación, el entrecruzamiento y la existencia de más de dos alelos en determinado lugar cromosómico, sirven todos para aumentar la variabilidad de la integración genética. Dicha variabilidad genética es definitiva para el desarrollo de especies nuevas y para la supervivencia de una especie en un ambiente de mutación continua. Si los genes no afectan directamente la conducta, sí sirven para regular la producción de proteínas, las cuales son responsables de las actividades estructurales y enzimáticas de un organismo. La primera preocupación de los científicos que trabajan en el campo de la genética conductual es la de la interacción entre los genes y el ambiente. Estos científicos intentan cuantificar la cantidad de variabilidad observada en un rasgo dentro de una población, como resultante de las diferencias genéticas entre los individuos y la cantidad proveniente de las diferencias ambientales. Las conductas son en su mayoría poligénicas; dependen de muchos lugares cromosomáticos, siendo la contribución de cada, gene pequeña y acumulativa. Con la utilización de animales se han realizado muchos estudios en situaciones de laboratorio donde el ambiente se mantiene relativamente constante. De esta manera es posible practicar la selección artificial y determinar los efectos de la estructura genética en diversas conductas. Los experimentos de apareamiento selectivo han demostrado que es posible hacer variar muchos rasgos conductuales mediante la manipulación genética. Se han establecido así mismo comparaciones entre familias o razas de animales. Además de las investigaciones experimentales de los efectos de la manipulación genética en la conducta animal, buena paste de la investigación productiva se ha ocupado de las bases genéticas de la deficiencia mental acentuada. La fenilquetonuria, la idiocia amaurática infantil, la idiocia amaurática juvenil y la corea de Huntington constituyen anormalidades que implican un solo lugar genético. La fenilquetonuria, por ejemplo, se debe a la herencia de un doble gene recesivo que causa una deficiencia en la producción de una sustancia que convierte la fenilalanina en tirosina. Los defectos de la complementación cromosómica producen así mismo deficiencias mentales. Quien tiene el síndrome de Down por ejemplo, porta 47 cromosomas en lugar de los 46 ordinarios y sufre en consecuencia un grave retardo mental. Las anormalidades en los números y clases de cromosomas sexuales afectan también la estructura y personalidad del hombre. El síndrome de Klinefelter, el de Turner y el síndrome XYY se consideran como resultados de anormalidades en los cromosomas sexuales. Las características poligénicas o sean las producidas por acciones de más de un lugar genético, son de importancia en campos tales como el de la esquizofrenia y la inteligencia. Las características poligénicas se han sometido a estudio mediante la determinación del grado de relación, en medidas cuantitativas de ejecución, entre parientes y no parientes. Las comparaciones de gemelos idénticos MZ y fraternos DZ con individuos libres de parentesco han sido de gran ayuda en el estudio del grado de heredabilidad de diversos rasgos. La evidencia actual indica que la esquizofrenia es de carácter genético. La inteligencia medida por las pruebas típicas de CI, parece determinarse también en forma significativa por factores genéticos. Sin embargo, factores ambientales como la experiencia cultural, el aprendizaje y la etapa de maduración, afectan todos la ejecución de pruebas de CI. Aunque el ambiente desempeña un papel importante después del nacimiento del organismo, el crecimiento y el desarrollo físico permanecen bajo un fuerte control genético desde la fecundación del huevo hasta la muerte del organismo. El desarrollo embrionario del sistema nervioso ilustra la interacción de la acción genética y el ambiente en el desarrollo de la estructura y la conducta. Las células del tejido que se convierte en sistema nervioso son originalmente similares a todas las demás células del embrión. Cuando se forman las tres capas germinales (ectodermo, mesodermo y endodermo) desarrolla así mismo el notocordio. Este determina la diferenciación de las células ectodérmicas en tejido nervioso por un proceso denominado inducción, en el cual el desarrollo de un grupo de células actúa para especificar el desarrollo de otro grupo de células por medio de su interacción. En las primeras etapas del desarrollo cerebral de los vertebrados se forma cierto número de estructuras. Cuando el ectodermo superior entra en contacto con el notocordio, se desarrolla la placa nerviosa a lo largo de la superficie superior. Con la elevación de los bordes de la placa nerviosa, se forma el tubo nervioso y se constituyen tres capas de células. Al cerrarse el tubo nervioso, se desarrollan las tres regiones cerebrales principales (prosencéfalo, mesencéfalo y romboencéfalo). El prosencéfalo se desarrolla formando la corteza cerebral y otras regiones cerebrales superiores. El mesencéfalo permanece sin dividirse y el romboencéfalo se desarrolla formando el cerebelo y el tallo cerebral. Mediante el crecimiento diferencial en estas tres regiones, adquiere el cerebro su forma adulta. A través de todo el sistema nervioso ocurre también una diferenciación acelerada. Se constituyen los núcleos cerebrales empiezan los tractos de fibras a conectar los núcleos, de las células de la cresta nerviosa se forman los ganglios espinales y del tejido cerebral se forma el ojo. Gran parte de tal diferenciación parece ser autocontrolada. Sin embargo, una vez establecido el patrón básico del sistema nervioso, adquiere mayor importancia su periferia. Es interesante anotar que cuando la periferia comienza a afectar el desarrollo nervioso, se hacen también funcionales los órganos periféricos. Las influencias hormonales en el desarrollo cerebral y conductual son de suma importancia. Se ha demostrado que una cantidad excesiva de tiroxina estimula de manera anormal la primera maduración de algunas áreas nerviosas y numerosos reflejos. Una cantidad reducida de tiroxina tiene un efecto negativo en el desarrollo cerebral y conductual. El desarrollo normal depende del suministro adecuado de tiroxina durante un período decisivo del desarrollo. Si la hormona está ausente durante tal período, no pueden remediarse las deficiencias en época posterior. Una de las perennes controversias en neuropsicología es la de natura versus nurtura. Desde el punto de vista de la nurtura en su forma extremada, el cerebro del infante es una tabula rasa en que la experiencia construye la mente humana. Desde el extremo contrario arguyen los partidarios de la natura que la mente adulta de los humanos está preprogramada en el cerebro del infante para desarrollarse con el crecimiento, en forma independiente de la experiencia. El rígido contraste de estos dos conceptos constituye una simplificación exagerada, dado que ningún rasgo humano es completamente genético o producto exclusivo de la experiencia. Los genes y el ambiente interactúan desde el momento de la concepción. Unidad 4 • Periodo Prenatal Es bien sabido que la fertilización o fecundación ocurre cuando un óvulo y un esperma se combinan. Pero lo que no se sabe tan bien es que nuestros conocimientos sobre el mecanismo de la concepción son relativamente recientes. Los antiguos griegos sabían muy bien que el coito estaba relacionado con la producción de la prole, pero no sabían que existían los óvulos y los espermatozoides. Se creía que el varón implantaba "la semilla de la vida" en la mujer, la cual servía solamente de depósito de alimento para el organismo en crecimiento. En el siglo XVII, un científico holandés, Regnier De Graaf, descubrió que la mujer producía un óvulo. Poco después, otro científico holandés, van Leeúwenhoek, descubrió el esperma. A pesar del descubrimiento de las dos células humanas germinales o gametos, la teoría de la fertilización siguió siendo muy parecida a la de los antiguos griegos. No fue sino hasta el siglo XIX cuando se determinó finalmente que la unión entre el óvulo y el esperma era esencial para la creación de un nuevo individuo. BIOLOGÍA DE LA REPRODUCCIÓN Mitosis y meiosis de las células Todas las células del cuerpo proceden de la combinación del esperma y del óvulo (cigoto), mediante un proceso de división de las células llamado mitosis. Cada cigoto contiene cromosomas. Estos a su vez, están compuestos de genes, los cuales son el material genético que pasa de una generación a otra. La mitosis de las células, da por resultado una copia idéntica del cigoto original. Esto se verifica mediante una serie de etapas: 1 Los cromosomas se duplican, formando "cromosomas dobles" o un conjunto pareado de cromosomas. 2 Los cromosomas dobles se alinean horizontalmente a lo largo de la línea ecuatorial de la célula. 3 Los miembros de cada conjunto pareado se separan y se alejan del centro hacia los extremos opuestos de la célula. 4 La célula empieza a estrecharse por la mitad y a formar dos células. 5 La membrana nuclear circunvala y encierra a cada una de las células que están completamente separadas, de manera que cada célula "nueva" contiene ahora un grupo idéntico de cromosomas. Los genes consisten dé ácido desoxirribonucleico (DNA), ácido ribonu-cléico (RNA) y proteína. El DNA, qué sé encuentra en el núcleo dé la célula, es el mecanismo central de control que regula la producción dé proteínas. Una de sus misiones es producir RNA en el núcleo de la célula. Se producen dos formas dé RNA: RNA mensajero y RNA de transferencia. El RNA mensajero deja el núcleo y se dirige a un ribosoma qué está situado en el citoplasma qué rodea al núcleo. El mensaje que lleva contiene el calendario apropiado para que los aminoácidos se reúnan y produzcan proteínas. El RNA dé transferencia tiene él oficio de trasladar a los aminoácidos al lugar del mensaje. Cada RNA de transferencia lleva solamente un aminoácido específico al mensaje. Cuando un RNA dé transferencia con un aminoácido se encuentra con un RNA mensajero en el ribosoma, entonces empieza la producción dé proteína. Hay que tener en cuenta que él DNA sirve de regulador mediante el cual los aminoácidos sé eslabonan para producir proteínas. Como estas substancias y su actividad sé derivan dé los cromosomas, se ve claramente qué la clave genética del individuo está íntimamente vinculada con la composición dé DNA, RNA y proteína de los cromosomas. En él ser humano normal, cada célula es diploide, lo cual significa qué contiene pares dé cromosomas -23 pares, o un total de 46 cromosomas. Uno dé los pares determina el sexo de la prole, mientras qué los 22 pares restantes desempeñan otras funciones que no tienen nada que ver con la determinación del sexo. Cada uno de los progenitores aporta 50% de cromosomas al nuevo organismo. Pero, ¿cómo es esto posible? Vimos qué en la mitosis dé las células el cigoto se divide para crear dos células "hijas", cada una dé las cuales contiene una ración completa de cromosomas. Evidentemente, se requiere otro tipo dé división celular para producir células qué contengan un haploide, o la mitad del número completo de cromosomas. Esté tipo de división celular se llana meiosis. En la división meiótica de la célula, los cromosomas se aparean y se juntan en un punto llamado sinapsis. Cada gen de un cromosoma se aparea con su gen correspondiente del otro cromosoma. Durante la meiosis se verifican dos fenómenos que garantizan efectivamente que el nuevo individuo sea genéticamente único. Uno de los fenómenos, el sobrecruzamiento, ocurre cuando los cromosomas se aparcan. El otro la segregación, se verifica cuando los cromosomas empiezan a separarse. Al sobre cruzarse, parte de los genes que están ensartados en un cromosoma se intercambian con genes que están ensartados en otro cromosoma (véase fig. 2-1). En la segregación, los cromosomas se separan, Fig. 2-1. "Sobrecruzamiento". Durante la formación de los gametos masculinos y femeninos, los cromosomas se entrelazan. Cuando los cromosomas se desprenden, las secciones de los dos cromosomas se intercambian. El sobrecruzamiento es uno de los factores que aseguran la exclusividad genotípica para todos los individuos, con excepción de los gemelos idénticos. La segregación quiere decir que el movimiento hacia los polos es al azar. Por lo tanto, por lo menos tres factores aseguran la individualidad genética: (1) la clave genética única sobre los cromosomas seleccionada de alguna parte del material genético parental; (2) el sobrecruzamiento mediante el cual el material genético en los cromosomas se reorganiza por sí mismo; y (3) la segregación, mediante la cual los cromosomas se mueven al azar de un polo de la célula que se está dividiendo al otro. El proceso mediante el cual se forma el esperma y los óvulos se llama gametogénesis o gametogenia. A la gametogénesis del esperma se llama espermatogénesis, mientras que a la de los óvulos se le llama ovogénesis. Durante la mitosis de la célula en los testículos, se separan ciertas células (espermatogonias). La espermatogénesis se refiere al proceso de división de la célula mediante la cual las espermatogonias se convierten en espermatozoides, o esperma, un proceso que requiere aproximadamente dos meses y medio para completarse. El espermatocito primario todavía contiene el número diploide de cromosomas. Una meiosis ulterior de la célula produce dos espermatocitos secundarios, cada uno con un número haploide de cromosomas; es decir, cada una contiene 22 autosomas (cromosomas que no tienen nada que ver con el sexo) más un cromosoma X o Y determinante del sexo. La ovogénesis tiene lugar en el ovario. A partir aproximadamente de la sexta semana prenatal en el feto femenino, se forman pequeños grupos de células (folículos primarios) mediante la mitosis de la célula. Los folículos primarios contienen los gametos femeninos u ovogonias. Durante el resto del desarrollo prenatal, la ovogénesis sigue adelante solamente hasta la formación de ovocitos. En este punto se detiene la meiosis de la célula, y vuelve de nuevo solamente cuando se presenta la ovulación a partir de la pubertad. Inmediatamente antes de la ovulación el ovocito primario completa su división, produciendo dos células hijas. el ovocito secundario y el cuerpo polar En este momento, la ovogénesis difiere de la espermatogénesis. Mientras que cada espermatocito secundario en definitiva produce dos células espermas, cada ovocito secundario produce solamente un óvulo. Los cuerpos polares se desintegran en el transcurso del proceso. El ovocito secundario continúa con su meiosis de la célula hasta que se completa el óvulo momentos antes de la fertilización. Es, mediante este proceso que un óvulo haploide se encuentra con un esperma haploide y se crea un nuevo individuo diploide. El sexo del individuo lo determina el cromosoma que se adhiere a los 22 autosomas. Los cromosomas diploides maternos que determinan el sexo son idénticos (XX), mientras que los cromosomas diploides paternos que determinan el sexo son diferentes (XY). Las letras X y Y son símbolos estándar para designar a los cromosomas que determinan el sexo porque efectivamente tienen una apariciencia muy semejante a la de los otros cromosomas. (El cromosoma X se parece materialmente a la letra X; el cromosoma Y, sin embargo, parece más bien una V que una Y). Los espermatozoides X o Y masculinos que quedan después de la meiosis de la célula determinan finalmente el sexo de la nueva prole; es decir, el sexo del producto lo determina el padre. Un cromosoma X del padre y un cromosoma X de la madre producen un producto femenino, mientras que un cromosoma Y del padre y un cromosomas X de la madre producen un producto masculino. Fenotipo y genotipo Los cromosomas contienen las unidades genéticas de la herencia. Estas estructuras son los genes, que existen en dos formas o alelomorfos. Los genes determinan el genotipo o la composición fundamental hereditaria del individuo. El genotipo se deduce porque no se puede observar. Por otra parte, el genotipo ciertamente contribuye para el fenotipo o características observables del organismo. Un principio fundamental de la genética del comportamiento es que cada organismo individual es un organismo único, distinto de los demás, excepto los gemelos idénticos. Recordemos que está exclusividad esta asegurada por procesos tales como el sobrecruzamiento y la segregación. Los fenotipos son el resultado de la interacción del genotipo con el ambiente del organismo. En otras palabras, la herencia y el ambiente interactúan para producir las características que se observan en el organismo, incluyendo el comportamiento. En algunos casos el fenotipo refleja características dominantes, mientras que en otros casos refleja características recesivas. Los conceptos de dominante y recesivo se derivan de la acción de los alelomorfos. Si D representa un alelomorfo de un gen y r otro alelomorfo del mismo gen, tendremos cuatro combinaciones posibles de los dos alelomorfos del gen: DD, Dr, rD y rr. Cualquiera que sea la combinación que resulte, recordemos que un alelomorfo procede del esperma (haploide) y uno procede del óvulo (haploide), de suerte que el par final de alelomorfos recibido de los dos padres contiene una de las cuatro combinaciones posibles, dependiendo de cuál se recibió de cada uno de los progenitores. Si el fenotipo Dr o rD del individuo es igual que DD, se dice que el carácter D es dominante. Si el fenotipo del individuo expresa rr, se dice que el rasgo es recesivo. El color de los ojos es un ejemplo claro de características dominantes y recesivas. Los ojos pardos son dominantes, los ojos azules son recesivos. Así, cuando B (ojos pardos) y b (ojos azules) se combinan (BB, Bb, bB, bb), las combinaciones Bb y bB se asemejan a BB, y no a bb. Es decir, se puede esperar que tres de cuatro niños tendrán ojos pardos. Siempre que un B (dominante) esté presente, sus características se manifestarán en el fenotipo. Si ambos progenitores reflejan fenotípicamente características recesivas (es decir, si los dos tienen ojos azules), es de suponerse que toda la prole tendrá ojos azules. Si uno de los progenitores tiene ojos azules (bb) y el otro tiene ojos pardos (Bb), dos de los niños pueden tener ojos azules, y dos pueden tener ojos pardos. Nótese que hemos expresado la probabilidad de que resulten ojos de un determinado color. Insistimos en esto porque la probabilidad de cada uno de los niños de tener un determinado color de ojos es independiente de la probabilidad de otro niño de tener el mismo color de ojos. Por consiguiente, no podemos afirmar con certeza que tres de cada cuatro niños tendrán un cierto color de ojos, sólo que es probable que tengan un cierto color. En realidad debido a la variabilidad genética, algunos de los niños pueden inclusive tener ojos verdes u ojos grises. Vulnerabilidad biológica Por alguna razón que todavía ignoramos, se concibe un número mayor de productos masculinos que femeninos (Rhodes, 1965): aproximadamente 120-150 varones por cada 100 mujeres. Una posible explicación es que los espermas Y son más ligeros y más rápidos que los espermas X, con la cual aumenta la probabilidad de que el esperma Y llegue primero al óvulo. Sin embargo, al final del período prenatal, la proporción se reduce a aproximadamente 105 ó 100. Además, las estadísticas de mortalidad muestran que mueren más varones que mujeres en cada edad que se haya estudiado durante los primeros 20 años de vida. Todo esto demuestra con toda claridad que los varones son más vulnerables biológicamente que las mujeres. La vulnerabilidad biológica se refiere al grado de mortalidad o de enfermedad asociado con la tolerancia de un organismo con respecto a la presión biológica o ambiental. Mayor mortalidad, morbilidad más frecuente o ambas en los varones esta asociada con muchos factores, incluyendo la susceptibilidad a las enfermedades infecciosas; infecciones cardiovasculares, renales y gastrointestinales; malformaciones congénitas; anormalidades cromosómicas; incompatibilidad de la sangre; y eclampsia (Allen y Diamond, 1954; Rhodes, 1965). Anomalías cromosómicas ligadas al sexo: En muchos casos el defecto se puede deber a genes defectuosos en los cromosomas que determinan el sexo. Ejemplos comunes de este tipo de defectos cromosómicos son Recordemos que los complementos normales de cromosomas para varones y mujeres son XY y XX, respectivamente. Algunas veces ocurren adiciones o substracciones. En el síndrome de Turner la mujer tiene sólo 45 cromosomas (le falta un cromosoma X). Las personas que padecen este mal carecen fenotípicamente de características sexuales secundarias, son de estatura baja, y tienen ovarios subdesarrollados. El síndrome de Klinefelter caracteriza a los varones fenotípicos (XXY). Estas personas tienen glándulas sexuales, masculinas o femeninas, subdesarrolladas, glándulas mamarias crecidas y con frecuencia son retrasados mentales. Anomalías autosómicas: No todas las anomalías cromosómicas afectan a los cromosomas determinantes del sexo. En la fenilcetonuria (FCU) al individuo le falta una enzima que es necesaria para convertir la fenilalanina (una proteína que se encuentra en la leche) en ácido fenilpirúvico. Se forma una sustancia tóxica debido a un metabolismo incompleto en la sangre, y el desenlace inevitable es el retraso mental, a menos que se trate el padecimiento. En la actualidad, a todos los recién nacidos se, les hace la prueba sistemática de la FCU ya sea mediante análisis de sangre o la prueba del papel tornasol a la orina. El tratamiento implica colocar al lactante con dieta especial. Este tratamiento es un excelente ejemplo de cómo una intervención ambiental puede neutralizar un defecto genéticamente programado. Una de las anomalías que ocurren con mayor frecuencia es el Síndrome de Down, una de cuyas formas se conoce con el nombre de trisomía 21 (el nombre popular de esta anomalía es mongolismo). El niño con Down tiene 47 cromosomas (el sobrante se añade al par 21 de los cromosomas, de ahí trisomía 21), es típicamente un retardado mental, y se le identifica fenotípicamente por la apariencia muy poco usual del cráneo, la cara, la lengua, los ojos, las manos, los pies y el cuello. Cuando aparecen en el cigoto menos de 44 autosomas, la enfermedad se conoce con el hombre de monosomía, una enfermedad siempre mortal. Las anomalías cromosómicas se deben a diversas causas, incluyendo la nodisyunción, la translocación, la deleción e inversión. La nodisyunción resulta cuando los cromosomas no logran separarse durante la meiosis de la célula. Por lo tanto, un gameto tiene los dos cromosomas, mientras que el otro no tiene ninguno. En la translocación, una parte de uno de los cromosomas se adhiere a otro cromosoma. Se cree que el síndrome de Down lo causa la nodisyunción o la translocación. En éste último caso, el cromosoma 21 se "engancha" al cromosoma 15 o viaja montado sobre él. El portador tiene los 23 pares normales pero están mal ordenados. Si el miembro del par 15 (con el 21 enganchado a él) y los 21 libres restantes se dan a un compañero que también ofrece un 21, entonces la prole tendrá tres cromosomas número 21 en lugar de los dos normales. Algunas veces una parte de un cromosoma desaparece durante la división de la célula, mientras que otras veces ocurre una inversión de las partes de algún cromosoma. Uno de los aspectos más interesantes de la sexualidad humana es el grado en que "la naturaleza" se ha esforzado por proporcionar condiciones óptimas para la unión del esperma y del óvulo. Por ejemplo, la ovulación (desprendimiento del óvulo del ovario y entrada en la trompa de Falopio) ocurre en un momento relativamente predecible durante el transcurso de cada ciclo menstrual. El tiempo que pasa entre el desprendimiento del óvulo y su descenso a la trompa de Falopio es aproximadamente de seis horas. El esperma puede vivir entre 12 y 36 horas. Por consiguiente el embarazo puede ocurrir inclusive cuando el coito precede a la ovulación. Además, la naturaleza ha proporcionado obstáculos casi insuperables contra la probabilidad de que el óvulo se pueda escapa: de la fertilización. El óvulo es una de las células más grandes en el cuerpo humano. ?e un tamaño aproximado de 0.14 de mm de diámetro (Correr, 1944). Para buscar este blanco, se descargan de 200 a 500 millones de espermas en cada eyaculación de fluido seminal. El número de células de esperma que se descargan está en función de la cantidad de fluido seminal que se emite. Esta cantidad fluctúa entre 2.5 y 5 ml, con aproximadamente 100 millones de esperma por cada ml. Varios miles de estos espermas llegarán a cruzar efectivamente la barrera cervical y buscaran literalmente el óvulo. Sin embargo, solamente un esperma, de entre toda esta abundancia, podrá penetrar al óvulo para llevar a cabo la fertilización. Si la fertilización se verifica efectivamente, el óvulo continúa su viaje de tres o cuatro días por la trompa de Falopio, hasta incrustarse finalmente en la pared uterina. Si no se verifica la fertilización, la mayor parte de la cubierta uterina es expulsada con sangre durante la menstruación. ETAPAS DEL DESARROLLO PRENATAL Los nueve meses de la gestación normal se dividen típicamente en tres etapas: el período del óvulo, el período del embrión y el período del feto. Período del óvulo (desde la concepción hasta la segunda semana prenatal) Durante el período del óvulo, el cigoto fecundado hace su descenso de tres a cuatro días por la trompa de Falopio y entra en el útero. Ahora el cigoto existe como un blastocisto que se mueve con toda libertad para todos lados. Empieza la mitosis de la célula, y los cromosomas se duplican y se separan, una y otra vez. La principal tarea de desarrollo de este período es la implantación del blastocisto en la pared uterina, un acontecimiento que ocurre aproximadamente entre 24 y 36 horas después de la entrada en el útero. Una vez implantado en la pared del útero, la sangre materna circunda totalmente al blastocisto. El trofoblasto, o tejido exterior del blastocisto, forma dos partes. Una parte se convierte en material nutritivo para el embrión, mientras que la otra se convierte en placenta y el cordón umbilical. Habiéndose incrustado en la membrana del útero, el organismo está ahora relativamente seguro dentro del ambiente uterino, y ya se satisfacen las condiciones para el crecimiento placentario y la inserción del cordón umbilical. Período del embrión (de la segunda a la octava semana prenatal) El período del embrión se caracteriza por la diferenciación continua del cigoto hasta formar tres capas: el ectodermo, el mesodermo y el endodermo. Del ectodermo emergerá finalmente el sistema nervioso, la piel y glándulas de la piel, el pelo y las uñas. Del mesodermo procederán la musculatura, el esqueleto, los sistemas circulatorio y excretorio, así como algunas porciones del sistema reproductivo. Del endodermo sale la mucosa del sistema digestivo, las trompas de Eustaquio, la tráquea, los bronquios, los órganos vitales y glándulas, así como otras partes del sistema reproductivo. Además de establecer la diferenciación de las capas, en el período embrionario señala el tiempo cuando se desarrolla la placenta y se forma la inserción umbilical (25 a 50 cm) entre la placenta y el embrión. Aproximadamente entre el decimoctavo y vigésimo primer día se empieza a formar el corazón, y para el final de la tercera semana prenatal ya se puede percibir un leve latido cardiaco. El saco y el líquido amniótico rodean al organismo para protegerlo contra el daño físico. La característica crítica de este período es la histogénesis, es decir, la diferenciación de las partes. Es precisamente durante este período cuando las intrusiones ambientales en el proceso del desarrollo pueden producir un daño particularmente grave y permanente al organismo. Período del feto (desde los dos meses hasta el final) El período del feto se caracteriza por el desarrollo continuo y el crecimiento de los sistemas básicos y por el reemplazo del cartílago por células óseas. La característica más importante de desarrollo es la morfogénesis u organización de las estructuras en desarrollo. El período comprende un vínculo muy crítico en el desarrollo prenatal: el punto de viabilidad. Este se refiere al tiempo, por lo general, 26 semanas de gestación, después del cual el feto tiene una probabilidad razonable de sobrevivir si naciera. Aunque, por lo general, se mide por semanas de gestación (las cuales inclusive en la actualidad son difíciles de determinar con precisión), el punto de viabilidad puede establecerse con mayor precisión tomando en cuenta factores tales como el peso al nacer y la longitud del feto. Con base en estudios de miles de niños nacidos en un hospital de Moscú, Makeyeva (1959) sugiere que 1,000 gramos de peso al nacer y por lo menos 35 centímetros de longitud, constituyen los límites mínimos para fundar la probabilidad de que sobreviva un prematuro. En cualquier caso, la vigésima sexta semana todos los sistemas principales ya están desarrollados, y los meses restantes se dedican al crecimiento y fortalecimiento continuo del feto como preparación para la entrada que habrá de hacer a un ambiente definitivamente muy distinto del medio prenatal. FACTORES QUE DETERIORAN EL AMBIENTE PRENATAL Hemos visto cómo diversas anomalías cromosómicas afectan al organismo en desarrollo. Pero además de las anomalías cromosómicas, hay otros muchos factores ambientales que pueden degradar al organismo prenatal y frustrar sus oportunidades de desarrollo normal. Con frecuencia es muy difícil aislar al agente activo específico que puede viciar el ambiente prenatal. Una de las razones de esto es el grado de interacción que existe entre el feto y la madre. No obstante, una vasta literatura señala con diversos grados de certeza, a varias categorías significativas de insultos que degradan el ambiente fetal. Veamos algunas de estas categorías. Desnutrición Prácticamente todo el mundo está consciente de la importancia que tiene la nutrición para nuestra vida. Además, probablemente todos nosotros hemos visto ejemplos de niños, ya sea en Biafra, en la América rural o citadina o en muchas otras partes del mundo, que sufren espantosamente de una falta intensa de alimento. Más aún, algunos "expertos" creen que la desnutrición bien puede ser el más importante de todos los factores ambientales que afectan negativamente el desarrollo prenatal. ¿Qué se entiende por desnutrición? Algunos investigadores distinguen dos clases de desnutrición (Hurlock, 1964). Una de estas, el hambre cualitativa se refiere a la cantidad inadecuada de vitaminas, aminoácidos, etc., esenciales para el crecimiento y desarrollo normales. La otra, el hambre cuantitativa, se refiere a la cantidad insuficiente de alimento, pero que satisface los requisitos mínimos de vitaminas y de otros elementos nutritivos. Otros investigadores distinguen tres niveles o grados de desnutrición (Gómez y otros, 1956). La desnutrición de primer grado connota pesos corporales de 76 a 90% del que tiene un lactante normal de peso promedio. La desnutrición de segundo y tercer grados connotan un peso corporal de 62-75% y menor de 61% respectivamente, con respecto a las normas establecidas. Naeye (1970) defiende la opinión de que no toda desnutrición depende directamente de la cantidad de alimento que se consume. Por ejemplo, algunos desordenes, clínicos relacionados con la desnutrición incluyen en la madre, desórdenes placentarios y uterinos, nacimientos múltiples y alimentaciones inadecuadas que tiene su origen en la pobreza. Se ha visto que la desnutrición cualitativa está asociada con el nacimiento prematuro, con la longitud y peso anormales del recién nacido, y con la desnutrición fetal que se manifiesta en niños que son demasiado pequeños para su edad gestacional (Antonov, 1947; Gruenwald, 1970). También se ha logrado vincular la tasa lenta de crecimiento intrauterino con una mortalidad más elevada y con una mayor susceptibilidad en relación a la morbilidad (Drillien, 1970). En general, una desnutrición grave parece tener efectos degradantes directos sobre el encéfalo y el sistema nervioso en desarrollo. En una serie de experimentos con niños africanos mal nutridos, los resultados indicaron que la desnutrición que sufrían durante los dos primeros años de vida producía un menor perímetro cefálico, reducción en el tamaño del encéfalo, anormalidades en la actividad de las ondas encefálicas y deterioro en el sistema visual (Stock y Smythe, 1968). Al estudiar los efectos de la desnutrición sobre los encéfalos en desarrollo de nueve lactantes chilenos que murieron por inanición, Winick y Rosso (1969) encontraron un menor peso encefálico y anormalidades en la composición de proteínas, RNA y DNA del encéfalo. Una hipótesis es que la desnutrición ejerce su influjo más degradante sobre el organismo precisamente durante el último trimestre del embarazo y durante los primeros años postnatales (Dobbing, 1970), cuando el encéfalo está en su período de más rápido crecimiento. En general, las deficiencias nutricionales se han traducido en deficiencia mental, inestabilidad nerviosa, parálisis cerebral, falta de peso al nacer, la longitud deficiente al nacer, raquitismo, debilidad física generalizada y muerte. . Enfermedades infecciosas Las enfermedades infecciosas constituyen un peligro potencial para el feto en desarrollo. Esto es especialmente verdadero durante el primer trimestre del desarrollo prenatal. Casi todos hemos oído decir que la rubéola puede ser peligrosa durante los primeros meses prenatales. No hay que despreciar estos datos. Se calcula que fueron centenares los lactantes que nacieron en el estado de Michigan durante la epidemia de rubéola de 1964-1965 que fueron gravemente afectados por el insulto prenatal. Por ejemplo, se registró una de las frecuencias más elevadas de hidrocefalia durante dicho brote. Otras enfermedades infecciosas que se sabe que tienen efectos degradantes incluyen las enfermedades venéreas: sífilis y gonorrea- y la poliomielitis. Las enfermedades infecciosas han estado asociadas con una frecuencia elevada de muertes al nacer, abortos, ceguera, deficiencia mental, sordera, microcefalia y la sordomudez. Incompatibilidad sanguínea Hay razones muy sólidas para exigir que los futuros esposos se sometan a una prueba de sangre antes del matrimonio. El proteger a la propia prole descubriendo y curando alguna infección venérea es mucho más importante que cualquier bochorno temporal que pueda uno sufrir. Además de identificar las infecciones venéreas o de cualquier otro tipo, las pruebas de sangre suelen poner en alerta a los padres y a los parteros acerca de ciertos desórdenes sanguíneos que tal vez requieran después un cuidado especial prenatal o postnatal. Uno de estos desórdenes, la eritroblastosis fetal, lo causa el factor Rh, llamado así por los monos Rhesus que se usaron en la investigación que aisló la incompatibilidad de la sangre. Todas las personas son Rh-positivas o Rh-negativas: aproximadamente 85% de los caucásicos, 93% de los negros, y casi todos los orientales, son Rh-positivos. Por consiguiente estas personas tienen un antígeno de los eritrocitos determinado genéticamente (el antígeno Rh, o factor rhesus), el cual, cuando es compatible con el feto, no causa problema alguno. Sin embargo, cuando el factor Rh fetal y el factor Rh materno difieren, surgen muchas dificultades. El antígeno Rh es en realidad un grupo de seis antígenos -C. D, E, e, d, e- y es el factor D el que lleva a la clasificación de Rh+ o Rh-. Si la combinación de alelomorfos contiene un antígeno D, la designación Rh será positiva, pero si la combinación de alelomorfos contiene una d, la designación Rh será negativa. Cuando una madre Rh-negativa tiene un feto Rhpositivo, la mezcla de sus sangres produce anticuerpos contra el antígeno Rh del feto, los cuales pasan el filtro placentario y provocan destrucción de los eritrocitos del feto. Por el contrario, la sangre fetal mezclada con la sangre de la madre causará sensibilización por el antígeno fetal Rh en su sistema sanguíneo. Si no se corrige, este trastorno conducirá a problemas todavía mayores en el siguiente embarazo. Afortunadamente, la incompatibilidad Rh puede tratarse. Para los lactantes, se pueden hacer transfusiones de intercambio de la sangre del feto in útero, eliminando de esta manera la incompatibilidad entre los factores Rh fetal y materno. En el caso de las madres, la inmunización con inmunoglobulina Rh dos a tres días después del parto evitará la formación de los anticuerpos Rh. Una vez inmunizada, la sangre materna deja de ser un problema para el siguiente embarazo. Sin embargo, si la situación no se resuelve, el feto puede ser prematuro, puede abortarse o nacer muerto (Stevenson, 1973). Medicamentos Debido al uso cada vez más extenso de medicamentos en nuestra cultura, deberá ponerse mucha atención a los efectos posiblemente nocivos que puedan tener en el organismo en desarrollo. La tragedia de la talidomida a principios de la década de los 60s es un ejemplo doloroso del camino menos indicado para descubrir los efectos nocivos de las drogas. Muchas embarazadas, principalmente en Alemania, que estaban tomando este tranquilizador, produjeron una prole con diversas deformidades físicas (por ejemplo, desarrollo incompleto de los brazos, de las piernas o de ambas extremidades). El sufrimiento físico y emocional que causa este tranquilizador particular justifica el que se prevenga a todos contra el uso de este medicamento durante el embarazo. Se ha descubierto que los narcóticos atraviesan fácilmente la placenta. Taussig (1962) encontró que los barbitúricos causan sufrimiento fetal. El organismo del recién nacido ciertamente no está preparado para combatir el sufrimiento de la abstinencia de la heroína adquirida en el útero. Se ha encontrado que muchos agentes que se usan como anestésicos y analgésicos atraviesan la placenta y entran en la corriente sanguínea del feto. Aun cuando hay ciertas pruebas de que el LSD suele causar una alteración estructural en los leucocitos de la sangre, los estudios de los efectos de la heroína y drogas derivadas sobre el feto todavía no son definitivos en la actualidad. No deberá sorprender que la nicotina, una droga extremadamente tóxica, puede resultar nociva para el feto. Ya en 1938 se encontró que fumar acelera el ritmo cardiaco del feto (Sontag y Richards, 1938). En un estudio, a fumadoras empedernidas se les privó de cigarrillos por un período de 24 horas. Luego se les ofreció a las embarazadas un cigarrillo. Inclusive antes de que la embarazada llegara a encender el cigarrillo, el ritmo cardiaco del feto se aceleraba, sugiriendo que la respuesta emocional y hormonal condicionada de la mujer influía sobre el feto (Lieberman, 1963). Se ha ligado la prematuridad al grado de tabaquismo durante el embarazo, las fumadoras más inveteradas tienen la máxima frecuencia de prematuros o de productos de bajo peso (Simson, 1949). Los estudios han demostrado que las embarazadas ingieren de tres a diez medicamentos diferentes durante el curso del embarazo, incluyendo substancias aparentemente inocuas como la aspirina (Bowes, 1970). Por desgracia, las embarazadas ingieren muchos medicamentos cuyos posibles efectos en el feto nunca se han estudiado. El consejo más seguro que se puede dar, es que se tenga un cuidado muy especial al tomar todas estas drogas durante el embarazo. Radiación Varios estudios llevados a cabo en las ciudades de Hiroshima y Nagasaki han puesto de manifiesto los efectos horrendos que tiene la extrema radiación atómica. En Hiroshima, la radiación estuvo asociada con mortinatos, aborto, malformación y bajo peso al nacer (Neel, 1953). En Nagasaki, embarazadas que se encontraban dentro de los 2,000 metros del hipocentro del estallido reportaron promedios más elevados de muertes fetales, neonatales y de lactantes. Entre los lactantes que sobrevivieron al estallido, los efectos característicos de la radiación fueron el retraso mental y el retardo en el crecimiento (Yamazaki, Wright y Wright, 1954). El estallido atómico no es la única que afecta negativamente al feto. En muchos casos, mujeres con cáncer o con tumores pélvicos requieren terapia de radio. Pequeñas dosis de un tratamiento de radiación como éste no ha producido ninguna embestida dañina en contra del feto, pero las dosis grandes deberán evitarse. Por desgracia no tenemos ninguna definición clara sobre lo que se debe considerar como una dosis excesiva en el caso de cada individuo en particular. En general, entre más al principio del embarazo se aplique la radiación, mayor es el daño potencial que se puede causar al feto. Edad materna Se ha dicho que la edad óptima para dar a luz es la comprendida entre los 20 y 29 años (Scott, 1968). El máximo porcentaje de problemas fetales relacionados con la edad de la madre se encuentra en mujeres mayores de 40 años. Por ejemplo, la frecuencia del síndrome de Down es mayor en los niños nacidos de mujeres mayores de 40. Si hay alguna relación entre la edad materna y la decadencia del sistema reproductivo femenino, es de suponer que se encontrarán más casos de sufrimiento fetal como resultado de óvulos viejos. Recordemos que todos los óvulos se forman antes del nacimiento y que en la ovulación solamente ocurre:, etapas finales de la división meiótica de las células. Por lo tanto, las mujeres de 40 años de edad, tienen óvulos también de 40 años de edad. Se puede suponer que un proceso semejante a éste se verifica en el sistema reproductivo del varón, aunque en este caso no está de por medio la edad del esperma. Cualquiera que sea la razón, es evidente que las mujeres tienen una máxima probabilidad de tener niños sanos si los dan a luz durante sus años veintes. Aquí viene muy bien una advertencia, que se puede aplicar también a todo lo que hemos dicho acerca de los factores que perjudican el ambiente prenatal. Esta llamada de alerta se refiere al uso de expresiones como "mayor frecuencia". En realidad, la mayoría de los recién nacidos a todos los niveles de edad materna son perfectamente normales. "Mayor frecuencia" connota solamente la mayor probabilidad de que ocurra un fenómeno a una edad determinada, comparada con la probabilidad del mismo a una edad de referencia convencional (digamos la edad entre los 20 y 29 años). Sin embargo, aunque los factores que se describen pueden afectar sólo a un pequeño porcentaje de fetos, son muy importantes. Cinco por ciento de todos los niños nacidos cada año en E.U.A.. o en cualquier país, son realmente una cantidad muy grande. Estado emocional materno Como no existen conexiones neuronales directas entre la madre y el feto, podría parecer poco probable que el estado emocional de la madre pudiera afectar al feto. La creencia de que los pensamientos maternos pueden influir directamente en el feto es origen de todo tipo de tabúes culturales con respecto a la mujer embarazada. Por ejemplo, en ciertas culturas a las mujeres embarazadas no se les permite manejar cuchillos por temor de que esto vaya a causar la ruptura. del umbilicus, y en muchas partes de E.U.A. existen tabúes en contra de comer ciertos alimentos, no sea que en el recién nacido vayan a aparecer lunares que tengan la misma configuración que tienen esos alimentos. Estas creencias no tienen ningún fundamento. No obstante, se ha ligado el estado emocional de la madre con la conducta fetal y postnatal. Rabin (1965) sugiere que la motivación para la paternidad bien puede ser lo que establece el tono para las futuras relaciones entre progenitor y niño, inclusive mucho antes de que el niño sea concebido. Otros investigadores han encontrado que las mujeres que no están satisfechas con su posición social o que son emocionalmente inestables, sienten menos gusto por su embarazo, y son más ansiosas y están más mal adaptadas emocionalmente durante el embarazo (Davids y Rosengren, 1962). Los efectos de ansiedad, perturbación y tristeza crónicas por el embarazo tienen mucho que ver con la hiperactividad, irritabilidad, llanto, dificultades para la alimentación y con los problemas del sueño de la prole (Sontag, 1957). Además, por lo menos, ni-; Investigador refiere que les madres de lactantes propensos a los cólicos son más tensas y ansiosas durante sus embarazos que las madres de lactantes que no son propensos a los cólicos (Lakin, 1957). Las madres de lactantes con cólico también dijeron que se sentían inadecuadas respecto a su capacidad para cuidar del futuro bebé. Como no hay conexiones neuronales directas entre madre y feto, ¿en qué forma puede ejercer su influencia en el feto el estado emocional de la madre? El influjo no puede ser un efecto directo del sistema nervioso, sino que tendría que ser un efecto indirecto en el sentido de que el hipotálamo es estimulado hasta producir substancias que con el tiempo causan una descarga de hormonas en la sangre. Las hormonas que se encuentran en la sangre materna cruzan fácilmente la placenta -aun cuando la misma sangre materna no cruza- y entran en la corriente sanguínea del feto. El desequilibro hormonal resultante muy probablemente afecta al desarrollo estructural y funcional de los órganos fetales. Es obvio que un número extraordinario de eventos pueden afectar notablemente el curso del desarrollo durante los meses prenatales. La desnutrición, la enfermedad, la incompatibilidad sanguínea, las drogas y medicamentos, la edad materna y el estado emocional de la madre, todo esto se ha visto que disgrega el desarrollo normal prenatal (Barnes, 1968; Ferreira, 1969; Stevenson, 1973). PREMATURIDAD De todos los temas relacionados con el desarrollo prenatal, el nacimiento prematuro es probablemente el que recibe la menor atención en el curso introductorio sobre desarrollo humano. Las consecuencias de un nacimiento prematuro para el desarrollo posterior ciertamente exigen que se les preste mayor atención. La idea típica que se tiene de prematuridad se reduce a que el lactante nace antes de los nueve meses de gestación. Aunque correcta en parte, esta definición no toma en cuenta para nada el pronóstico desalentador de los lactantes nacidos prematuramente. La Organización Mundial de la Salud (World Health OrganizationWHO), al reconocer que la duración de la gestación no es por sí sola un índice inadecuado de prematuridad, estableció una definición basada en el bajo peso al nacer (WHO, 1961). Se definió al lactante prematuro como aquel niño cuyo peso al nacer era menor de 2,500 granos. Por definición de prematuridad aproximadamente 8% de todos los nacimientos son prematuros. Esto significa que en E.U.A. aproximadamente 300,000 niños nacen prematuramente cada año. Estos niños constituyen casi la mitad de todas las muertes neonatales, y los que sobreviven tienen las tasas más elevadas de impedimentos físicos y psicológicos. En un tiempo se creyó que la duración de la gestación y el peso al nacer eran medidas suficientemente exactas comes ,ara hacer un buen diagnóstico y pronóstico del niño nacido prematuramente. Aun cuando esto sea verdadero en general, hay otros factores que también son importantes. Por ejemplo, Drillien (1964) opina que el peso al nacer y la duración de la gestación no toman en cuenta ciertos factores que interactúan, como por ejemplo, las diferencias geográficas y sexuales, el orden de nacimiento, la condición social, la edad de la madre, los nacimientos múltiples y lo adecuado o inadecuado del cuidado parental. En general, la prematuridad definida en función de una gestación disminuida y un bajo peso de nacimiento, se ha encontrado que está relacionada con deficiencias intelectuales, disfunción perceptual-motora, lesión encefálica, lenguaje inmaduro, parálisis cerebral, defectos visuales y dificultades con el razonamiento abstracto. Además, estos efectos se mantienen firmes aun cuando los investigadores controlan los efectos de raza, actitudes maternas, prácticas maternas de crianza y clase social (Wiener, Rider, Oppel, Fischer y Harper, 1965). En el estudio de Drillien (1964) se utilizó el procedimiento longitudinal que se usa en muchos estudios sobre la prematuridad según este método, el investigador escoge una muestra de lactantes que tienen como característica común la enfermedad condición específica que se pretende estudiar. Entonces se selecciona un grupo de control apropiado igual al primero pero sin la enfermedad de que se trata. Después se vigila a ambos grupos longitudinalmente para tomar nota de la frecuencia de la condición que interesa estudiar. Por lo tanto, si el nacimiento prematuro es el fenómeno que interesa estudiar, se escoge obviamente un grupo de niños nacidos prematuramente y se comparan con un grupo de lactantes de gestación normal. Durante el estudio longitudinal se administran diversas pruebas y exámenes para determinar la diferencia que se da entre los grupos. El medio más eficaz y razonable para resolver los problemas de prematuridad es la prevención. Makeyeva (1959) alaba mucho las técnicas profilácticas que se usan extensamente en la práctica obstétrica en la Unión Soviética paró reducir a un mínimo la prematuridad. Estos procedimientos incluyen: evitar el excesivo consumo de líquidos y alimentos sólidos, reducir el consumo de sal, restringir el trabajo mental y manual, aprovechar al máximo el aire fresco y dormir todo lo que se pueda, y además ¡usar ropa floja y cómoda! Sin embargo, aun cuando se siguieran estas normas al pie de la letra, muchos niños seguirán naciendo prematuramente. Por lo tanto, la prevención reviste otro significado: a las madres hay que ayudarlas para evitar que caigan en lo que Lindemann (1965) llama aflicción anticipatoria. La aflicción anticipatoria se refiere al hecho de que la prematuridad es nociva para madre e hijo. El nacimiento prematuro significa que la madre y el niño van a estar separados por algún tiempo. En algunos casos, este período puede extenderse a meses. Debido a que el bebé tiene un riesgo tan alto, muchos progenitores se preparan para lo peor; es decir, anticipan el fallecimiento del niño. Si éste sobrevive a menudo resulta difícil para los padres volver .a establecer sentimientos positivos hacia él. Parecen haber olvidado "casi por completo" que tuvieron un hijo. En el hogar, el prematuro puede no ayudar a los padres a que desarrollen sentimientos positivos ya que el bebé tiende a ser de cuidado difícil porque necesita más alimento, más sueño y tiene más problemas de temperamento que el niño a término. Para contrarrestar el fenómeno de la aflicción anticipatoria y el alejamiento subsiguiente entre padres y vástago, muchos hospitales solicitan que la madre (por desgracia el padre no) visite diariamente el hospital para que participe en forma activa en el cuidado y manejo de su hijo. Obviamente, no todos los niños nacidos prematuramente pasan por las situaciones devastadoras que hemos descrito aquí. No obstante, el número de lactantes que ciertamente sufren efectos penosos por haber nacido demasiado pronto o pesando extremadamente poco, es lo suficientemente grande como para justificar una investigación continua acerca de técnicas preventivas y profilácticas adecuadas con las que se puede ayudar al lactante prematuro a lograr un desarrollo normal. RESUMEN El período prenatal de la vida humana, abarca desde la concepción hasta el nacimiento. La concepción ocurre cuando los gametos del sexo (el esperma y el óvulo) se unen. Esta unión es la base del crecimiento y desarrollo del nuevo organismo, al principio mediante el proceso de la división celular llamado mitosis. Cada nueva célula normal contiene 23 pares de cromosomas, uno de los cuales es el responsable de la determinación del sexo del organismo. Los gametos sexuales pasan por otra división especial llamada meiosis. Después de la división meiótica de las células, cada esperma u óvulo contiene solamente 22 cromosomas individuales (autosomas) y un cromosoma que determina el sexo. Cuando el esperma y el óvulo se unen, el nuevo organismo tiene otra vez la ración completa de 46 cromosomas. Los genes existen en dos formas llamadas alelomorfos y están situados sobre los cromosomas. Los alelomorfos pueden ser dominantes y recesivos. Los genes son los responsables de regular el programa genético del individuo, pero este programa no es algo fijo o invariable. En todas las etapas del desarrollo el ambiente aporta algo al contenido del programa. En la mayoría de los casos solamente podemos deducir la estructura genética o el genotipo del organismo, tomando nota de las características que se observan o del fenotipo. Vulnerabilidad biológica significa el grado de morbilidad o mortalidad que está asociado con la tolerancia del organismo hacia el stress. Las anomalías cromosómicas interfieren con las oportunidades que tiene el organismo para desarrollarse normalmente. Además, son varios los factores ambientales que pueden degradar el ambiente prenatal y amenazan la supervivencia misma del feto. El período prenatal se divide en tres períodos parciales: el período del óvulo, el período del embrión y el período del feto. Aunque la mayoría de los fetos tardan de 270 a 280 días para desarrollarse, algunos niños nacen mucho antes de esto. Generalmente llamamos a estos niños prematuros. Sin embargo, se ha visto que la definición de prematuridad que se basa exclusivamente en la duración de la gestación, es inadecuada. Lo menos que se puede hacer es considerar también el peso y la longitud corporal al nacer. A pesar del cuidado prenatal moderno y de los recursos hospitalarios actuales, la prematuridad sigue siendo una amenaza para el organismo. Más aún, inclusive el cuidado especial que presta el hospital puede dar por resultado cierto grado de privación sensorial o social, o una privación de la estimulación normal que ofrece el ambiente. No obstante, los futuros padres tienen que tener muy en mente que la mayoría de los niños, cualesquiera que sean las posibles dificultades, llegan al mundo en perfecto estado de salud. Cómo llegan y qué es lo que les sucede durante el primer mes de vida será el tema que se tratará en el siguiente capítulo. APARTADO 3-1 Temas por estudiar • • • • • • • • • • ¿Cuáles son las tres fases del proceso del nacimiento y cuáles son los fenómenos asociados con cada fase? ¿Cuáles son las presentaciones anormales al nacer y cómo difieren de la presentación normal? ¿Qué influencias tienen los medicamentos para el parto sobre el comportamiento del recién nacido e sobre su desarrollo subsiguiente? ¿Qué factores deben ser considerados- al determinar si el recién nacido está en peligro o no? ¿Cuáles son algunas de las características de conducta del recién nacido? ¿En qué forma el estado o nivel de excitación afecta la responsividad del recién nacido con respecto a la estimulación ambiental? ¿Cómo se puede pacificar a un recién nacido? ¿Qué importancia tienen los ritmos biológicos para la organización de la interacción entre el lactante y el que cuida de él? ¿Cuáles son las ventajas y desventajas de la alimentación con el pecho para la organización de la interacción entre el lactante y el que cuida de él? ¿Se deberá alimentar a los lactantes según un horario fijo o según demanda libre?