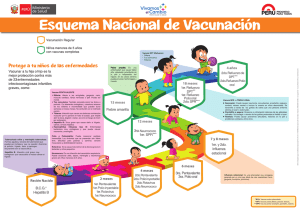
Estudios de inocuidad e inmunogenicidad protectogénica de la
Anuncio

Instituto Finlay Vicepresidencia de Investigaciones Estudios de inocuidad e inmunogenicidad protectogénica de la vacuna antimeningocócica VA-MENGOC-BC® en modelos murinos. Tesis Presentada en Opción al Grado de Doctor en Ciencias Veterinarias. Autor: Dr. Juan Francisco Infante Bourzac Tutor: Dr. Gustavo V. Sierra González Dr.C Cotutor: Dr. Carlos Bulnes Goicohea Dr.C La Habana 2000 “Año del 40 Aniversario de la Decisión de Patria o Muerte” TABLA DE CONTENIDO REFERATA Pág. INTRODUCCIÓN 2-3 Hipótesis 3 Objetivos General 3 Objetivos Parciales 3 PARTE GENERAL 3 CAPÍTULO I 3 REVISIÓN BIBLIOGRÁFICA 3 PARTE EXPERIMENTAL CAPÍTULO II MATERIALES Y MÉTODOS GENERALES 3-5 CAPÍTULO III DESARROLLO DE LOS BIOMODELOS RATÓN Y RATA PARA 6 REPRODUCIR LA ENFERMEDAD CAUSADA POR NEISSERIA MENINGITIDIS B. EXPERIMENTO 1 Introducción 6 Objetivos 6 Materiales y Métodos 6-7 Resultados y Discusión El ratón como biomodelo para la enfermedad causada por Neisseria 7-8 meningitidis B. Las ratas recién nacidas como biomodelo para reproducir la 9-10 enfermedad causada por N.meningitidis serogrupo B. 10 Conclusiones CAPITULO IV EFICACIA DE VA-MENGOC-BC EN LOS BIOMODELOS RATA Y RATÓN. 10 EXPERIMENTO 2. 11 Introducción 11 Objetivos 11-13 Materiales y Métodos Resultados y Discusión Comprobación de 13-14 la eficacia de la inmunoglobulina antimeningocóccica 14-16 BC en ratas recién nacidas y ratones Eficacia de VA-MENGOC-BC y de la Inmunoglobulina BC en ratones Balb/cj frente a diferentes cepas de Neisseria meningitidis serogrupo B aisladas en Latinoamérica. 16-17 Evaluación VA-MENGOC-BC y de la inmunoglobulina hiperinmune antimeningocóccica BC en ratones Balb/cj frente a reto con Neisseria 17-18 meningitidis de los serogrupos A, B y C. Conclusiones 18 CAPÍTULO V INOCUIDAD DEL PRODUCTO VA-MENGOC-BC EN BIOMODELOS (RATA Y RATÓN) 18 EXPERIMENTO 3 18 Introducción 18-21 Objetivos Materiales y Métodos 21-24 Resultados y Discusión 24-25 Evaluación toxicológica de VA-MENGOC-BC en ratas Spreague Dawley 25-26 Evaluación anatomopatológica de VA-MENGOC-BC en ratones Balb/cj 26-30 utilizando la vía intramuscular. 30-31 Conclusiones DISCUSION GENERAL CONCLUSIONES FINALES 31 RECOMENDACIONES ANEXOS. Agradecimientos Muchas son las personas a las cuales debo expresar mis más sinceros agradecimientos, tantas, que tal vez no serían suficientes el volumen de esta tesis, para cumplir con esa sagrada misión, no obstante, trataré de mencionar todos los que en estos momentos recuerdo, pidiendo disculpas a los que no sean nombrados por problemas de espacio o de mi memoria que pudiera no recordar y a todos esos, le expreso que estoy eternamente agradecido. Primeramente quiero reconocer muy especialmente a mi tutor Dr. Gustavo Sierra González, quien a pesar de las múltiples tareas e importantes responsabilidades, siempre tuvo tiempo para orientarme no solo con esta tesis, sino en una importante etapa de mi desarrollo profesional y al que nunca le faltó la palabra estimulante o crítica en el momento adecuado. Igualmente a mi Cotutor Dr. Carlos Bulnes Goicochea, que también con importantes responsabilidades, sacó tiempo incluso de su sagrado descanso para ayudarme dando muestras de su enorme altruismo. También debo agradecer profundamente a la Dra. Concepción Campa Huergo, quien siempre me ha dado la necesaria cuota de estímulo y confianza para desarrollar mi actividad laboral en general y esta tesis en especial. A mis compañeros de la División de Modelos Experimentales y Patología, encabezados por el Dr. Sergio Sifontes Rodríguez con su ayuda intelectualmente brillante y eficiente, al Técnico Reynaldo Oliva Hernández con mucha voluntad y capacidad de trabajo, a mí estimada amiga Viviana Pérez Amat que además de elaborar buena parte del material que sirvió de base a esta tesis, siempre me estímulo y considero todo esto como parte suya, a todos los compañeros de este gran colectivo que me ha acompañado durante estos tiempos, Mildrey, Mayo, Vismark, Arsenio, Alfredo, Heriberto, Omar, Yolanda y Dayana. En el CENSA a la Dra. Eva Marrero Faz y su esposo Dr. Octavio Fernández que como buenos amigos siempre me aconsejaron y compulsaron en la consecución de este objetivo, allí también llegue mi reconocimiento a su Directora Dra Lidia Tablada, a la Dra. Miriam Pedroso y Maritza Barreras por su revisión crítica al documento de tesis, a los Drs Jorge Luis Alvarez, Roberto Faure, Jorge Espinosa y a Figueredo. También en el Finlay al Dr. Oliver Pérez por sus adecuadas críticas al igual que a la Dra Sara Catalina Esnard y al Dr Arturo Talaveras, al Lic. Enrique Muñoz por su amistad y apoyo.A todas las queridas compañeras de la biblioteca del Instituto Finlay, Carolina, Bertha, María Victoria y Nuría al igual que al director de esa área Lic. Giberto Sotonlogo. En la Dirección de publicaciones a Lic. Orlando Gutiérrez, Candido García y a la siempre atenta Lic. Virginia Betancourt. Al Dr Tomás Verdura por su aguda y acertada crítica de la tesis, así como a la Dra. Lourdes Almeida. En este colectivo no pudiera faltar la secretaria Mayelin siempre dispuesta a brindarme su eficiente y oportuna ayuda. Mención especial merecen mis amigos Noel González, Manuel Lescay, José A. Blanco, Rafael Avilés, Gerónimo Ruíz, Roman Sánchez y Eric López. A mis amigos y profesores los Drs Oscar Viamontes, David Wiliams Cantero, Aramis Fernández, etc. A la Dra. Miriam Ribas por su excelente revisión de estilo y al Dr. Pentón por sus orientaciones. Quisiera dejar muestras de mi agradecimiento a los compañeros del CENATOX especialmente al Dr Rafael Pérez Cristiá por sus atenciones y consideración también a Juan F. Sanchez, Gema, Alfredo y a todos los demás integrantes del colectivo. Resumen En el presente trabajo se tuvieron como objetivos fundamentales comprobar la eficacia e inocuidad del producto biotecnológico VA-MENGOC-BC®, vacuna antimeningocócica contra los serogrupos B y C, para esto se utilizaron animales de laboratorio ratas y ratones procedentes del Centro Nacional para la Producción de Animales de Laboratorio (CENPALAB) de ambos sexos, adultos jóvenes en el caso de los ratones pertenecientes a la línea Balb/cj y las ratas de la línea Sprague Dawley, además se usaron ratas recién nacidas todos fueron mantenidos en condiciones convencionales de tenencia y alimentación, se logró desarrollar el biomodelo ratón mediante la aplicación de los Factores Estimuladores de la Virulencia (FEV), luego de un extenso trabajo de selección se demostró que el uso de la Dextrana - Férrica combinada con la mucina gástrica, es el más adecuado de los FEV. Este modelo resultó útil para la evaluación tanto de la vacuna VA-MENGOC-BC® y de la inmunoglobulina antimeningocócica BC, obteniéndose niveles de protección estadísticamente significativos entre los animales inmunizados activa y pasivamente con respecto a los controles, en tanto que el modelo rata recién nacida aunque, no es adecuado para la evaluación directa de la vacuna, si nos permitió conocer la eficacia de la inmunoglobulina antimeningocócica, tanto en tratamientos previos como posteriores al reto, además las ratas nos facilitaron la profundización en el conocimiento de la patogénea de la infección por Neisseria meningitidis serogrupo B (NmB) en esta especie identificando su similitud con la enfermedad meningocócica humana. Por otra parte estos modelos sirvieron para conocer la eficacia de VA-MENGOC-BCy de la inmunoglobulina frente a diferentes cepas de (NmB) aisladas de pacientes enfermos de meningitis meningocócica en Cuba, Brasil, Colombia, Chile y Argentina. También se comprobó la eficacia de ambos productos cuando fueron realizados retos con cepas de Neisseria meningitidis de los serogrupo (NmA) y (NmC) resultados de gran relevancia por demostrar el espectro de protección de VAMENGOC-BC, y de la Inmunoglobulina Antimeningocócica BC frente los serogrupos mencionados quedando de esta forma demostrada la protección cruzada que se establece entre los diferentes serogrupos de (Nm). Se realizaron estudios toxicológicos que incluyeron determinaciones de bioquímica y hematología en las cuales se obtuvieron valores situados dentro del rango establecido para esta especie línea, línea, edad y sexo, las observaciones clínicas diaria no mostraron alteraciones en el estado de las ratas, los niveles de anticuerpos frente NmB fueron demostrados en los animales vacunados mientras que los estudios anatomopatológicas, evidenciaron la inocuidad de la vacuna al no hallarse lesiones de valor diagnóstico, excepto la presencia de formaciones granulomatosas de tipo macrofágica a nivel del punto de inoculación, tanto en la vacuna como en el placebo de hidróxido de aluminio, además se comprobó la evolución de las lesiones granulomatosas en el punto de inoculación y su relación con los niveles de anticuerpo en la especie ratón. Teniendo en cuenta los resultados obtenidos en los ensayos donde se establecieron y perfeccionaron los biomodelos murinos (rata y ratón), así como la comprobación de la eficacia de VAMENGOC-BC® en los referidos biomodelos , a través de los cuales quedó demostrada su inocuidad, consistente en estudios de toxicología con el esquema propuesto para uso clínico, otro subaguda y el de evolución del granuloma en el ámbito del punto de inoculación, concluimos que VA-MENGOC-BC® es inocua y eficaz en los biomodeos murinos, lo cual brinda las bases científicas y regulatorias que garantizan el uso ampliado del producto. Introducción Entre las enfermedades infecciosas las meningitis bacterianas representan el mayor problema a que se enfrenta el hombre, para lograr el control y prevención de la morbilidad y mortalidad en la especie humana (Bugaev et al., 1990). El desencadenamiento de grandes epidemias de meningitis, la no total eficacia de los antibióticos para prevenir las graves secuelas en el Sistema Nervioso Central (SNC) de los sobrevivientes a la meningitis, y los altos porcentajes de casos fatales en recién nacidos afectados con esta enfermedad, fundamentan la necesidad de continuar los estudios tendientes a dar solución a estos problemas de salud. (Anderson et al., 1994). La enfermedad meningocócica es causada por Neisseria meningitidis (Nm) caracterizada por un proceso patológico de comienzo súbito, rápida evolución y pronóstico fatal en la mayoría de los pacientes. Se presenta en diferentes zonas geográficas del mundo, con mayor incidencia en las regiones con climas templados y tropicales; afecta sólo a los seres humanos, fundamentalmente en las edades más tempranas de la vida. La inmunización constituye el medio preventivo por excelencia (Schwartz y Moore,1989). Nm no es el único microorganismo que causa la meningitis, pero solo él es capaz de generar grandes epidemias. NmB, de modo similar a otras bacterias Gram (-) que afectan el SNC, logra provocar cerca del 100% de mortalidad en pacientes no tratados, mientras que si se realiza el tratamiento adecuadamente puede alcanzar entre el 5 y 15% de casos fatales (Saukkonen y Leinonen, 1986). La patogénesis de esta enfermedad permanece desconocida (Caputo et al., 1992). Sin embargo, se sabe que el microorganismo coloniza la nasofaringe mediante su adherencia a las células columnares no ciliadas, luego alcanza las células subepiteliales y finalmente el torrente sanguíneo donde, puede causar bacteremia, la cual se acompaña de diferentes síntomas clínicos que dependen de la inmunidad del hospedero. Los serogrupos A, B y C son los responsables del 90% de los casos de esta enfermedad (Ala`Aldeen, 1996). En el mundo y especialmente en nuestro país, cobra gran importancia la obtención de biopreparados inmunológicos dentro de los cuales podemos citar las vacunas y los sueros inmunes (sueros policlonales y anticuerpos monoclonales), los que presentan un amplio espectro de aplicación en diferentes ramas de la vida social y económica (Tamayo, 1999). A pesar de los extensos estudios realizados en décadas pasadas, los mecanismos responsables de la inmunidad natural contra meningococo no resultan claros. Los estados de protección se han relacionado con la presencia de anticuerpos bactericidas; no obstante, se piensa que también la respuesta inmune celular y la muerte de las bacterias por fagocitosis desempeñar un importante papel en la defensa del hospedero contra la enfermedad meningocócica (Frasch, 1993). Las vacunas en sentido general son consideradas una herramienta fundamental de los sistemas nacionales de salud en el mundo, debido a su importancia social en la prevención de enfermedades, por ser la práctica de salud más segura y de mejor relación costo/beneficio con respecto a otras terapias farmacéuticas tradicionales. Existen evidencias demostrativas de que la disminución de coberturas en el uso de vacunas, van unidas a la reaparición de enfermedades ya erradicadas en diferentes países (Cruse y Lewis, 1989). Por otra parte, la presencia tarde o temprano de resistencia antimicrobiana, en la mayoría de los patógenos, es otra razón importante para promover las inmunizaciones (Frasch, 1994), (Lee, 1998). Por otra parte es conocida la existencia de vacunas contra Nm basadas en el polisacárido capsular contra los serogrupos A, C, W135 y Y, las cuales ofrecen una buena, aunque relativamente corta, protección contra cada uno de los respectivos serogrupos. Estas no ofrecen protección cruzada contra el serogrupo B y para lograrla este propósito se han explorado varias estrategias de enfoques que incluyen la utilización de los polisacáridos capsulares mejorados, así como las preparaciones obtenidas a partir de las proteínas de membrana externa, de expresión constitutiva o reguladas por hierro (Brodeur y Tsang, 1986). Mundialmente se han probado múltiples vacunas contra NmB, obtenido resultados variables (Bundle y Smith 1974); sin embargo, son muchos los que se quejan de la poca efectividad que las mismas proporcionan (Wyle y Arterteins, 1972),(Zollinger et al., 1984). En la 5ta. Conferencia Internacional sobre Neisserias patógenas celebrada en 1986, se presentaron algunos resultados sobre la marcha de una vacuna contra meningococo B, anunciándose la realización de una prueba masiva de terreno, luego de la correspondiente realización de las pruebas en animales y voluntarios humanos, cuyos resultados fueron satisfactorios. Esta vacuna fue perfeccionada constituyendo un biopreparado bivalente, conocida como VA-MENGOC-BC, la cual ya ha sido comercializada en diferentes países con resultados satisfactorios. Cuba es el único país del mundo que ha desarrollado una vacuna altamente eficaz contra el serogrupo B (Sierra y Acosta, 2000); que también ofrece protección contra el serogrupo C, estos han sido históricamente responsable de grandes epidemias, (Favorova et al., 1995). Además nuestro país goza del derecho y posibilidad de producir la Inmunoglobulina Antimeningocócica eficaz contra estos serogrupos. La vacuna cubana es considerada la más efectiva contra NmB de las conocidas hasta el momento. La misma se desarrolló en la década del 80 y luego de concluir la prueba de eficacia (83 % de protección) se aplica en Cuba sistemáticamente desde 1991 a partir de los 3-5 meses de edad, (Sierra et al., 1991). La resistencia a la infección bacteriana toma muchas formas por lo que es conveniente clasificar las distintas manifestaciones de resistencia en diferentes grupos, tal como la inmunidad natural o no específica, la inmunidad activa y pasiva, mientras que los ejemplos de cada una de estas clases pueden ser demostradas de manera experimental; necesariamente no existen medios para que cada tipo de resistencia sea gobernado por mecanismos enteramente distintos. Quizás sean los modelos animales los que encuentren una respuesta a estos problemas que en el ser humano no puedan ser dilucidados por cuestiones de ética y derechos humanos. La utilización de los modelos animales para comprobar la eficacia de las vacunas en general y en especial contra Nm, ha sido estudiada por diferentes autores desde la década de los años 30 (Miller, 1933),(Braham, 1937),(Braham y Lilie, 1938), (Calver, 1976) (Brodeur et al., 1986) (Frasch, 1994), (Zollinger et al., 1995). En el Congreso Internacional sobre Neisserias patógenas celebrado en Inglaterra en 1994 se confirmó este concepto (Brandtzaeg, 1994), durante el desarrollo del mismo se realizó un taller sobre el uso de los biomodelos relacionados con estas vacunas, dejando establecido el concepto e importancia que poseen para definir el poder protector frente a diferentes serogrupos y cepas, del germen Nm. En las meningitis bacterianas, tres cuestiones fundamentales deben investigarse: ¿Cómo y bajo qué circunstancias la bacteria alcanzan el Sistema Nervioso Central (SNC)?; ¿Qué factores del hospedero y cuáles del microorganismo determinan la magnitud del daño producido?; ¿Cómo puede una de estas determinantes aplicarse para eliminar o prevenir la morbilidad y mortalidad de dichas afecciones?. Para ello los modelos animales brindan un método de acercamiento a dichas interrogantes. Por esto se han realizado múltiples intentos con el objetivo de lograr un biomodelo experimental adecuado para las meningitis bacterianas. Sin embargo, encontramos que prácticamente ninguna especie animal resulta susceptible a los principales gérmenes productores de meningitis en el hombre y la mayoría de estos requieren elevados números de bacterias para lograr que ellos sean infectados. Estas altas cargas tienden a producir endotoxemias primarias y finalmente la bacteremia resultante es transitoria con una letalidad que muchas veces es baja (Andersen, 1983). El nivel alcanzado por la industria médico-farmacéutica ha impuesto la necesidad de crear las condiciones para el desarrollo de nuevos y mejores modelos experimentales, con la finalidad de determinar los efectos tóxicos y establecer la adecuada relación riesgobeneficio en aras de garantizar la seguridad y la eficacia de los nuevos productos farmacéuticos. Los métodos “in vitro” y los nuevos modelos animales probablemente representen las direcciones principales de la investigación en el campo de la farmacotoxicología en la próxima década, y con toda seguridad, los retos más importantes de la comunidad científica en esta disciplina durante el tercer milenio. La vacuna cubana VA-MENGOC-BC® contiene dentro de su componentes principales el adyuvantes inmunológico hidróxido de aluminio el cual le confiere una importante propiedades que facilitan su eficacia. La calidad inmunológica o el logro de la inmunoestimulación efectiva, depende en muchos casos del uso de estos adyuvantes. Los cuales, son como definición preparados químicos o biológicos incorporados al antígeno e inyectados simultáneamente con él, que implican un ahorro de inmunógenos y de tiempo, al obtenerse una respuesta inmune rápida y lograrse altos títulos de anticuerpos, así como la estimulación de otros importantes elementos del sistema inmune. (Frasch, 1994). Actualmente los adyuvantes constituyen un componente imprescindible para la obtención de preparados vacunales, incluyendo las vacunas de primera generación; siendo el elemento biotecnológico más atractivo y uno de los centros de interés del Programa Nacional de Vacunas. Estas sustancias inmunopotenciadoras deben satisfacer exigencias de tolerancia e inocuidad, lo que condiciona la necesidad de buscar nuevos compuestos con estas características propiciando a su vez la obtención de bioreactivos de elevada “eficacia y efectividad” (Deng, 1998). El hidróxido de aluminio es el único adyuvante aprobado internacionalmente por la OMS para uso en humanos, entre otras cualidades por su inocuidad y por estimular la respuesta inmune, pero está lejos de ser el ideal, debido a que no es siempre efectivo, ya que solo incrementa levemente la inmunidad mediada por células y no logra la actividad inmunológica (Roitt et al., 1994). Internacionalmente, existió la tendencia de aplicar a los productos vacunales los esquemas convencionales para los estudios toxicológicos que son usados para fármacos de origen químico; sin embargo, estos no responden a los requerimientos de productos biológicos que interactúan con el sistema inmune, donde se hace necesario, ante todo, comprobar la no-existencia de procesos inmunotóxicos, a través de estudios clínicos aleatorios, así como otros a largo plazo. En épocas tan recientes como 1998 la Comunidad Económica Europea expidió las regulaciones que reglamentan y adecuan el trabajo toxicológico en vacunas (Stephene, 1998). En nuestro país, como parte de los avances científicos y biotecnológicos ocurridos en las últimas dos décadas, la toxicología experimental ha evolucionado rápidamente y muchos ensayos han sido realizados con el fin de comprobar posibles efectos tóxicos de múltiples fármacos producidos por nuestra industria médico - farmacéutica. No obstante, en relación con el sistema inmune, las experiencias son incipientes. Sin embargo, las oficinas de registro sanitario dentro y fuera del país solicitan evaluaciones toxicológicas más profundas cada día. Perfeccionar los modelos experimentales para demostrar la inocuidad y eficacia de los preparados vacunales contra Nm, es una realidad que se ha hecho patente en los principales eventos científicos internacionales, así como en las normas establecidas por los organismos regulatorios y la literatura científica relacionada con el tema. Sin embargo, no existe un modelo animal universalmente aceptado que permita una adecuada estimación de la “ protección” de candidatos vacunales contra NmB, entre otros aspectos debido a la no-existencia de los animales de laboratorio capaces de reproducir la enfermedad meningocócica como tal. Se han obtenido buenos resultados con los ratones de líneas isogénicas (C57BL/6, Balb/c, CBA, etc.) tratados con sustancias estimuladoras de la virulencia (Ashton et al., 1989). Estas sustancias basan su efecto en la capacidad que poseen para desplazar el equilibrio, existente entre la resistencia del hospedero y la virulencia del germen inclinando la balanza a favor de este último. Varias sustancias han sido utilizadas con el objetivo antes mencionado. En los laboratorios del Instituto Finlay se han obtenido los mejores resultados con una combinación de mucina y dextrana férrica. Estos FEV fueron satisfactorios en la utilización del biomodelo ratón, tanto para los estudios de protección activa con VA- MENGOC-BC, como en los de protección pasiva con la Inmunoglobulina Antimeningocócica BC, cuando se efectuaron retos con el germen Nm. Basando su efecto en el mecanismo conocido como “Inmunidad Nutricional” en el cual los individuos regulan las concentraciones de metal hierro en los fluidos corporales para de esta forma invalidar o al menos dificultar el crecimiento de los gérmenes que requieren del referido metal para su desarrollo y con este la aparición de la infección. En trabajos previos se había probado el potencial protector de los anticuerpos desarrollados en seres humanos (niños), que fueron inmunizados con la vacuna antimeningocócica (VAMENGOC-BC) (Galguera et al., 1991). También se ha empleado un modelo de ratas infantes inmunizados pasivamente con los anticuerpos obtenidos de los humanos inmunizados con (VA-MENGOC-BC)® y que posteriormente fueron sometidas a un reto mediante la inoculación intraperitoneal e intranasal de una suspensiones de NmB (Infante et al., 1997)a . A pesar de esto, las crecientes exigencias regulatorias a vencer por la referida vacuna para ser introducida en el llamado primer mundo, que establecen los requisitos para esclarecer y demostrar científicamente la inocuidad y la eficacia del producto, nos planteamos el problema de evaluar la farmacotoxicología preclínica de la vacuna antimeningocócica BC, producto único y novedoso, para lo cual ha sido necesario perfeccionar modelos murinos y establecer correlatos de protección que ofrecen una idea de lo que se espera sucederá en el humano, así como establecer marcadores adecuados de inocuidad, los cuales brindan la base científica y regulatoria para fundamentar los estudios clínicos, el registro y el uso ampliado de dicho producto. Por otra parte, los trabajos que versan sobre la toxicología de productos vacunales en el país son limitados, entre ellos se pueden mencionar los realizados por (Infante et al., 1997)b en vacunas convencionales, así como las pruebas toxicológicas específicas para las vacunas de tipo (toxoides) contra tétanos y difteria. Estos incluyen la evaluación de la potencia de las toxinas originales en ratones y curieles, pruebas de incremento de peso corporal en ratones y la prueba de reversión de toxoide a toxina, (Chuprinina et al.,1990). En el caso de VAMENGOC-BC® no existen precedentes, por lo tanto se ignora si los modelos murinos disponibles sean idóneos para evaluar la eficacia e inocuidad de nuestra vacuna. Por todo lo cual se formuló como hipótesis de trabajo la siguiente: Hipótesis: La vacuna antimeningocócica VA-MENGOC-BC es un producto inocuo de baja reactogenicidad y capaz de provocar una respuesta inmune específica protectogénica en modelos murinos. Objetivo general: Comprobar la inocuidad y capacidad de inducir una respuesta inmune específica protectogénica por parte de VA-MENGOC-BCen modelos murinos, como requerimiento preclínico para continuar el desarrollo del medicamento y lograr su registro sanitario, para lo cual nos propusimos los objetivos particulares siguientes Objetivos Particulares: - Establecer y perfeccionar los biomodelos murinos (rata y ratón) para la reproducción experimental de la infección producida por Nm. - Comprobar la eficacia de la vacuna VA-MENGOC-BCa través de la respuesta específica protectogénica en los biomodelos de rata y ratón. - Demostrar la inocuidad de VA-MENGOC-BCen los biomodelos rata y ratón. Revisión Bibliográfica - Epidemiología y biología de la enfermedad meningocócica producida por Neisseria meningitidis. El conocimiento de la genética de las poblaciones meningocócicas ha suministrado gran información para la observación del comportamiento epidemiológico de los meningococos y el curso de la enfermedad en los seres humanos, lo cual pudiera servir para la lucha y control de ellos en el mundo. Muchos microorganismos patógenos poseen una estructura poblacional clonal, así como algunas enfermedades que ocurren en un momento determinado, son causadas por pequeños números de clones patógenos. De la misma forma, la epidemiología meningocócica se caracteriza por su actividad hiperendémica periódica o epidémica, dada por la emergencia de un clon patogénico que se mueve como causa epidémica en diferentes localidades durante un tiempo. Los avances en biología molecular sobre la cadena de la polimerasa, basada en el análisis de la reacción en las secuencias de nucleótidos, han suministrado gran información para el conocimiento en la epidemiología de la enfermedad meningocócica. Así, de las secuencias de un número de genes meningocócicos de los serogrupos B y C, se ha hecho evidente que hay un intercambio horizontal considerable del material genético entre cepas, de las que se obtienen poblaciones panmísticas. - Formas clínicas de la enfermedad En este epígrafe, por la importancia que tienen en la valoración clínica y diagnóstica, así como para la parte descriptiva en la presentación de la enfermedad, nos remitimos al trabajo de Brandtzaeg, (1994). La meningitis meningocócica sin estado de choque y disfunción multiórganos es la presentación clínica más común. Después de la penetración al espacio subaracnoideo aumenta la proliferación bacteriana, que se refleja por un incremento de los LPS en el líquido cerebroespinal (LCR), los cuales a menudo son más de 100-1 000 veces superiores a los niveles medidos en muestras de plasma colectadas simultáneamente. Varias citoquinas son generadas en el LCR a niveles mucho más altos que en el plasma, reflejando las diferencias en los niveles de LPS. La relación LCR /sangre de la glucosa estuvo inversamente relacionada con los LPS del LCR, lo que evidenció la alteración del metabolismo celular del cerebro. El número de leucocitos no se correlacionó significativamente con los niveles de LPS en el LCR. La septicemia meningocócica fulminante está caracterizada por colapso vascular, disfunción renal y pulmonar, coagulación intravascular diseminada, infarto vascular, pleocitosis mínima y hemorragia intensa en la piel y adrenales. Todos los pacientes con niveles de LPS en plasma mayores que 700 µg/mL (7 EU/mL), desarrollaron un choque séptico persistente. Los LPS inducen una respuesta inflamatoria intravascular a través de un complejo interflujo de mediadores, donde las citoquinas están todas involucradas. La activación masiva del sistema de complemento, inicialmente a través de la vía alternativa, está asociada con los niveles circulantes de LPS y puede contribuir al colapso vascular. La enfermedad meningocócica también ha reflejado complicaciones durante su curso, tales como alteraciones severas de la conciencia, desórdenes neurológicos, artritis, miocarditis y necrosis extensa de los tejidos blandos (Roznovky, 1994). -Modelos animales. Concepto: Un modelo animal o biomodelo es aquel cuyos mecanismos patológicos son suficientemente similares a los de las enfermedades humanas, para las cuales el referido proceso patológico del animal pudiera servir como modelo; aclarando que la enfermedad de este animal pudiera ser similar al padecimiento humano, cuando esta ocurra en forma natural o inducida en forma experimental (MESH, 2000). También se consideran modelos animales aquellas alteraciones del estado de salud, cuyas manifestaciones clínicas no son completamente iguales a la enfermedad humana correspondiente y para la cual se utilizan. Tal es el caso de la enfermedad del ganado ovino (Ovies aries) conocida con el nombre de scrapie, que sirve de modelo a la esclerosis múltiple humana, sin embargo esta enfermedad nunca puede aparecer o su desarrollo ser inducido en un modelo animal. -Los biomodelos y el desarrollo biotecnológico actual Los animales de laboratorio están entre los más valiosos instrumentos que se utilizan en la investigación biomédica. Por eso los centros de investigación científica se preocupan por contar, dentro de sus recursos para la investigación y desarrollo, con colonias de animales de óptima calidad, cuyas características biológicas se conozcan en forma exhaustiva de manera tal que los cambios fisiológicos o patológicos que se presenten en el curso de una investigación no den lugar a errores de interpretación o lleven a confundir los resultados de un experimento con alteraciones propias de las enfermedades de los animales de experimentación. Entre las múltiples ventajas que derivan del estudio de los modelos experimentales, se mencionan como las más importantes: 1- La posibilidad de conocer la historia natural de la enfermedad, cuya etiología, patología, sintomatología y evolución pueden mantenerse en condiciones experimentales, sin la influencia de factores extraños que la modifiquen, en especial los provenientes de la terapéutica. 2- La posibilidad de reproducir las enfermedades en forma experimental, casi a voluntad, permitiendo disponer de la casuística suficiente. 3- La posibilidad de hacer estudios fisiopatológicos y patológicos, que son difíciles o inaccesibles en personas enfermas. 4- La posibilidad de utilizar medios terapéuticos (Medicamentos, cirugía, fisioterapia, etc.) cuya aplicación en la especie humana se considera peligrosa o no ética. 5- La posibilidad de estudiar factores ambientales físicos, nutricionales y genéticos que inciden en la evolución de las enfermedades. 6- La posibilidad de estudiar algunas enfermedades en líneas de animales isogénicos o (consanguíneos), lo cual ha abierto un inmenso campo de investigación en la inmunología, la cancerología y sobre todo en las enfermedades hereditarias. Las ventajas antes mencionadas se deben evaluar con todo cuidado y juzgar con riguroso sentido crítico. La llamada extrapolación de resultados no es posible en aquellos casos en que no sea probada la identidad entre la enfermedad del hombre y la del animal experimental (Cuba, 1982). Los modelos experimentales provienen de tres fuentes: a) Animales de laboratorio propiamente dicho: ratones, ratas, hámster, curieles, etc. b) Animales domésticos: vacas, ovejas, caballos, etc. c) Animales salvajes y semisalvajes: monos, ardillas, peces, etc. La amplia utilización de los animales de laboratorio se pone de manifiesto en los estudios realizados por Mora y Lomelí, (1996) que constataron en un análisis retrospectivo de 50 años (1941-1990) que el 22 % de los artículos publicados en ese periodo mencionaban el uso de los animales de laboratorio, aunque era variable la importancia relativa para cada disciplina. Así por ejemplo, uno de cada 3 artículos del área de inmunología y 1 de cada 4 en las áreas de fisiología y biología del desarrollo, involucraban el empleo de especies animales. En sentido general alrededor del 80 % de los animales usados en investigaciones biomédicas son roedores, no consanguíneos (outbred), isogénicos (inbred), mutantes, polimórficos y transgénicos. En el caso de los mutantes son modelos de enfermedades hereditarias como la obesidad, diabetes y la distrofia muscular. Los transgénicos se logran por la introducción de DNA foráneo dentro de un embrión preimplantado (Wilmat, 1997). En relación con el status sanitario, la Gnotobiología como ciencia de avanzada en la biomedicina, ha desarrollado los animales libres de patógenos específicos (SPF), los libres de todo germen (Germ-free), animales atímicos, el Knoch-out mice, el ratón con inmunodeficiencia severa combinada (SCID) (Alam et al., 1994), y finalmente la obtención de animales quiméricos y la clonación, conforman el alto nivel alcanzado por esta ciencia en la actualidad. Otros modelos que a pesar de constituir rarezas de la naturaleza y con mutaciones genéticas, merecen ser mencionadas como son el modelo Rino que padece un crecimiento anormal de la piel, los modelos de enanismo y de psoriasis, el modelo de alcoholismo IKKI-NOMI, etc. Por otra parte resulta obligado cuando se habla de animales de laboratorio, referirnos a la ética de esta actividad, gracias a la cual la biomedicina ha realizado grandes aportes a través de los animales de laboratorio logrando incrementar espectacularmente las esperanzas de vida y la calidad de esta. Dentro de la ética, lo planteado por Russel y Burch (Melius, 1997) referente a las tres importantes vías para disminuir el uso de animales de laboratorio, constituyen un principio inviolable cuando expresan: - Reemplazo: Sustituir el animal por modelos in vitro. - Reducción: Evitar la experimentación repetitiva, ajustando el número de animales al mínimo necesario que permita obtener datos estadísticamente confiables. - Refinamiento: Evitar a toda costa el sufrimiento de los animales. Estas normas éticas constituyen leyes de obligatorio cumplimiento en muchos países como es el caso de México, donde desde 1995 se elaboró el anteproyecto de Norma Oficial Mexicana (NOM) “Especificaciones Técnicas para la Producción, Cuidado y Uso de los Animales de Laboratorio”, lo cual constituyó un hito en la historia de la ciencia y la medicina de los animales de laboratorio en México, donde se refieren entre otros aspectos, al sacrificio humanitario (eutanasia) de estas especies en el contexto de las investigaciones científicas, las pruebas de control y la enseñanza superior (Proyecto de Norma Mexicana, 1996). Otro aspecto que por su importancia ocupa a los hombres de ciencia de hoy, es la alometría, disciplina que se refiere al crecimiento de los órganos y busca de aquellos parámetros, medidas o resultados que sean directamente transponibles de las especies animales al hombre y la cronofarmacología, ciencia dedicada al estudio de las influencias que los ritmos tiempo - dependientes tienen sobre la acción de los productos biológicos (ritmo circadiano), dando espacio a la siguiente pregunta: ¿Será correcto experimentar con ratones en horarios diurnos, teniendo estos hábitos de vida nocturnos?(Giraldes, 1997). - Estado de los biomodelos atendiendo a las distintas temáticas motivo de investigación en Vacunas y Sueros para uso humano en Cuba. Durante más de 10 años de intensa labor, se han desarrollado diferentes modelos animales con el objetivo de comprobar primeramente la seguridad de las vacunas y los sueros para uso humano (Cuadro I-1). Además los biomodelos han sido utilizados en la demostración de la eficacia de estos productos biológicos a través de las pruebas de potencia o de inmunogenicidad (Cuadro I-2) a la vez que otras han sido mediante confrontaciones directas llevadas acabo en animales protegidos con vacunas y sueros durante las etapas de investigación o producción (Cuadro I-3)– Las ratas SPF. Este modelo ha suministrado resultados satisfactorios en la demostración de la efectividad de VA-MENGOC-BC frente a cepas de Nm de los grupos B y C (Infante et al; 1997a), la cual se puso de manifiesto a través de los niveles de anticuerpos específicos antimeningocócicos determinados por el método de ELISA y a través de la protección contra retos frente a ambos serogrupos. Los ratones, curieles (Cavia porcellus) y conejos (Oryctolagus cuniculus) han servido para los estudios toxicológicos preclínicos de dosis única, irritación en el punto de inoculación, farmacodinámica, farmacocinética y estudios de inmunidad mediada por células en animales vacunados con VA-MENGOC-BC (Lastre et a.,; 1995), (Pérez et al., 1997) . Los ratones y curieles han sido empleados en las pruebas de inocuidad inespecífica, inmunogenicidad y anticuerpos bactericidas en las pruebas de control biológico final para el producto VA-MENGOC-BC, mostrando consistencia respecto a la seguridad y eficacia durante más de 10 años (datos no publicados). -Biomodelos para Haemophilus influenzae tipo b Ratas recién nacidas. Han demostrado sus posibilidades como modelos para el tratamiento con sueros, antibióticos y preparados vacunales, reproduciendo un cuadro lesional a nivel del sistema nervioso central, de gran similitud al que sucede en el hombre con bacteremia reconocida, con la presencia de gérmenes en líquido cefalorraquídeo y respuesta a la terapia antibiótica (Marrero, 1993)(Sifontes et al., 1996). Ratones hiperferrémicos. Este modelo con reconocido mérito por constituir un instrumento indispensable para conocer los mecanismos patogénicos de esta enfermedad y además como posible control para demostrar la eficacia de una vacuna (Infante et al., 1998). Curieles adultos con más de 450 g de peso: Por su respuesta inmunológica similar a la de un niño pequeño han demostrado su eficacia para reproducir la enfermedad meníngea producida por H. influenzae tipo b tanto en sus aspectos inmunológicos, como lesionales (Infante et al., 1999). Tienen las ventajas de una posible obtención de mayores cantidades de suero para los estudios serológicos y la demostración comparativa de posibles diferencias entre productos vacunales (Silver y Anderson, 1995). -Biomodelos para Leptospira interrogans (Leptospirosis humana) El hámster sirio (Mesocricetus auratus) o dorado: Estos animales reproducen con gran fidelidad y de manera similar, el proceso clínico patológico de la leptospirosis humana, teniendo en cuenta sus características biológicas y el conocimiento existente acerca de los aspectos sanitarios y genéticos de la especie. Por otra parte, es una de las especies recomendadas para este tipo de ensayo (Oliva et al., 1994) y también por el organismo regulatorio para el caso de las evaluaciones preclínicas de medicamentos por parte del Ministerio de Salud Pública de Cuba (Anónimo, 1993), permitiendo la evaluación de seguridad, potencia y efectividad frente a retos en hámsters con la vacuna vaxSPIRAL (bacterina trivalente contra los serovares L. canicola, L. icterohaemorrhagiae y L. pomona) Dentro de estas características se destaca clínicamente la fiebre, debilidad, inapetencia y la muerte que sucede en un período de 7 a 14 días, Anatomopatológicamente, constituyen especialmente órganos diana, el riñón y el hígado en primer término, seguido de pulmón y corazón con lesiones evidentes, así como la presencia de abundantes leptospiras identificadas por la coloraciones argénticas . Conejos, hámsters, ratones y curieles: Estos han permitido evaluar las pruebas de toxicidad en la fase preclínica por dosis única, repetida, irritación en el punto de inoculación y las pruebas de inmunopatología, para la vacuna vaxSPIRAL, cumpliendo de esta forma lo estipulado en el párrafo anterior. -Biomodelos para Vibrio cholerae (Cólera humano) Ratones recién nacidos: Son muy apropiados para conocer características importantes en la virulencia de las cepas del germen, como son la invasividad, colonización y estudios de adherencia, los cuales han sido exitosos en nuestros laboratorios (Benítez et al., 1996), (Cedré et al., 1998). Conejos adultos jóvenes: A través de la técnicas Ileal Loop y Retarded Intestinal Tied Adult Rabbit Diarrhoed (RITARD) e inmunización intraduodenal de cepas candidatas para vacunas, con el objetivo de conocer virulencia, inmunogenicidad y protección frente a reto, han sido utilizados en los ensayos cotidianos en la etapa preclínica de una futura vacuna contra el cólera. -Biomodelos para Clostridium tetani (Tétanos humano) Ratones, curieles y conejos: Han sido utilizados con el interés de desarrollar la vacuna duple y triple en las pruebas de toxicología preclínica, dosis única, repetida y punto de inoculación (Posada, 1995). Ratones y curieles: También fueron empleados para demostrar posible toxicidad residual presente en lotes a granel de la vacuna VADIF-TET y para cálculos de DL50 de la toxina tetánica, prueba de potencia de esta vacuna, etc. De esta manera también se realizaron cálculos de la dosis letal mínima para las toxinas tetánicas y diftéricas, ensayo que se basa en las propiedades de estas y de su manifestación a través de modelos experimentales sensibles, permitiéndo conocer su calidad desde el punto vista biológico. El procedimiento para ambas toxinas es igual, con la única diferencia que la toxina diftérica solo se prueba en curieles, mientras que la tetánica se verifica en ratones y curieles. -Biomodelo para Corynebacterium diphteriae (Difteria) Ratones, curieles y conejos: Con el mismo objetivo del acápite anterior se tomaron estas especies animales en las pruebas de toxicología preclínica, dosis única, repetida y punto de inoculación en la vacuna VADIF-TET (Díaz, 1995). Curieles: Se usan para la determinación de la toxina residual y para calcular la DL50 de la toxina diftérica. Curieles y ratones: Se emplean para las pruebas de potencia de esta vacuna duple bacteriana. -Biomodelos para Bordetella pertussis (Tosferina) Como complemento a la vacuna duple se adiciona el componente pertusis para lograr de esta manera la triple bacteriana. Ratones recién nacidos: Se usan para observar los efectos dermotóxicos (toxina dermonecrótica). Ratones adultos jóvenes: Se emplean para la sensibilización a la histamina. Ratones, curieles y conejos: Se utilizan para las pruebas de toxicología preclínica de vacuna duple y triple bacteriana. Ratones: Se usan en prueba de confrontación de potencia a través de la vía intracerebral. Ratón adulto: Se emplean para prueba de leucocitosis. Conejos: En pirógenos se emplea este biomodelo para todas las vacunas en las fases preclínicas y en el control final de los biopreparados. -Biomodelos para Salmonella typhi (Fiebre tifoidea) Se realizan ensayos en ratones, previamente utilizado para la selección de la línea más sensible al germen en la prueba del cálculo de la DL50 aplicando diferentes FEV (Mucina y levadura) (Cvjtanovic y Vemurak, 1965). Quedó demostrado que la línea más sensible fue la C-57/BL6. Todos los ensayos están avalados por un profundo estudio histopatológico que comprueba la eficacia del biomodelo y el objetivo final era servir como biomodelo para lograr una vacuna contra la fiebre tifoidea a base de Poli vi. La presencia de lesiones en órganos linfoides, las cuales facilitan la presentación de la enfermedad, así como los paratifomas en diferentes tejidos, fueron identificados a través de los estudios histopatológicos. Además los ratones: Nos han permitido obtener cantidades suficientes de antisueros necesarios en los estudios inmunológicos preclínicos de este preparado vacunal. -Biomodelos para estudios de inocuidad en alergenos. Los ratones de la línea OF-1 se han empleado en pruebas de inocuidad como parte del control de calidad de los alergenos de germen Stretococus aureus (Infante et al., 1996) -Los biomodelos en el desarrollo de vacunas con la utilización de la biología molecular. Expresión de antígenos vacunales en vectores vivos atenuados: Dentro de estas se encuentran el BCG, la Salmonella typhi Ty 21 a expresando la subunidad B de la toxina del cólera (Acosta y Sarmiento, 1996), (Sarmiento y Acosta, 1996). Inmunización con ADN desnudo: En Mycobacterium tuberculosis, Toxoplasma gondii, N. meningitidis (incluyendo reto en ratones), Trypanosoma cruzi (Alberti et al., 1997),(Vidal et al., 1997). Modelos experimentales de la enfermedad injerto contra huésped: Para su aplicación en el desarrollo de vacunas y productos asociados como los adyuvantes inmunológicos (Acosta et al., 1994ª ) (Acosta et al., 1994b). - Desarrollo de biomodelos para amebiasis humana: Utilizando el hámster dorado por inoculación intrahepática con el objetivo de obtener las lesiones en este órgano y probar una vacuna contra este parásito. -Biomodelos en la medicina tradicional. Los animales de laboratorio han servido como objeto de estudio en experimentos de cronobiología mediante la aplicación de la vacuna antimeningocócica VA-MENGOCBC®, y en la actualidad se acometen trabajos relacionados con la moxibustión y la acupuntura. -Biomodelos aplicados a las vacunas virales. Virus Rábico (Virus fijo CV-S). El ratón OF-1 ha sido utilizado en las investigaciones del virus rábico, para evaluar el comportamiento de inmunogenicidad y pruebas de titulación. Esta última es de gran importancia para determinar la verdadera cantidad de virus utilizado, teniendo en cuenta las reacciones clínico - patológicas de los animales como son parálisis y otros trastornos que acusan el compromiso del SNC (Núñez et al., 1997), (Almora et al., 1997). Virus Sarampión (Cepa L-16). Se acometen investigaciones en primates no humanos, monos verdes (Cercopithecus aethiops) con el objetivo de realizar las pruebas de neurovirulencia que incluyen estudios histopatológicos y la inmunogenicidad en la referida especie. Las codornices japonesas (Cotumix cotumix) han sido utilizadas en estudios de estandarización de los procesos de embriogénesis, incluyendo microbiología y morfología de estos embriones, que posteriormente sirvieron de sustrato al virus en cultivos celulares (fibroblastos de codorniz). También se usan equinos para la obtención de sueros hiperinmunes destinados a medios diagnósticos de esta enfermedad. Virus respiratorios. Se desarrollan ensayos para verificar las características higiénico morfológicas de embriones procedentes de gallinas (Gallus gallus) pertenecientes a líneas ligeras y pesadas, con vista a su utilización como substratos en los medios de cultivo del virus. Mientras tanto los conejos proporcionan sueros hiperinmunes como medio diagnóstico de los virus respiratorios. - Otras investigaciones con biomodelos. El Centro de Inmunología Molecular (CIM) investiga a partir de sangre total de equinos para la obtención de gangliósidos a partir de eritrocitos, trabajos estos relacionados con las vacunas contra el cáncer. - Papel del hierro en microbiología El hierro en microbiología desempeña un papel importante, ya que tanto a nivel del huésped, como en los microorganismos, este metal constituye un elemento esencial para sus procesos fisiológicos y su desarrollo. Precisamente, han pasado cinco décadas desde que Schade y Caroline, (1994), dieron evidencias de la presencia de hierro fuertemente ligado a las proteínas de la clara del huevo y el plasma, sugiriéndo que los ligantes pueden funcionar como bloqueadores del metal para los invasores bacterianos. De aquí en adelante han sido muchas las investigaciones que se han realizado para tratar de dar una explicación, lo más acertada posible, del papel que desempeña el hierro a nivel del huésped y los invasores. En las últimas décadas se ha puesto gran interés a la actividad de la transferrina y sus receptores en de la superficie celular en el metabolismo del hierro (Thorstensen y Romslo, 1988), por lo que tratamos de abordar varios aspectos que contribuyen al conocimiento del hierro, resultan de gran interés para el desarrollo científico. -Los sideróforos Son compuestos elaborados para solubilizar y transportar el hierro férrico en las especies microbianas. Un sistema de sideróforos está compuesto de dos partes principales: el ligante y una colocación para la captación y utilización. Ambas partes están sujetas a un dispositivo regulatorio común que se dispara según la concentración intracelular de hierro. Son considerados como uniones ferroespecíficas de bajo peso molecular; las uniones de átomos que están eslabonados al hierro, en las que predomina el oxígeno son a menudo suministradas por el ácido hidroxámico o radicales catecoles ( Neilands, 1994). -Inmunidad nutricional La inmunidad nutricional es la habilidad que tiene el huésped para privar de hierro a los microorganismos. El papel de la inmunidad nutricional hierro-específica en la protección de los huéspedes frente a las enfermedades bacterianas o fúngicas, es más importante durante el período comprendido entre la invasión y la multiplicación bacteriana. Inmediatamente después de la invasión, el huésped se hace hipoferrémico, aumentando la habilidad de la transferrina para tomar hierro de la molécula siderófora, que los microorganismos entrantes pueden sintetizar. En experimentos realizados en curieles, con la inyección IM de Cl. perfringens, se ha demostrado que inyectando por vía endovenosa el hierro de 0 a 6 horas después de la inoculación, se producen muertes dentro de las 18 horas posteriores, pero si el hierro se inyecta 8 horas después, los animales se recuperan porque han eliminado las bacterias. En particular, la inyección de hierro y la consiguiente hiperferremia baja la resistencia del huésped y disminuye el tiempo de incubación de microorganismos o productos bacterianos. No obstante, hay que considerar entre las diferentes especies, que el nivel normal de saturación de la transferrina varía. En el curiel es muy alta, en el conejo y el ratón es moderadamente alta, mientras en el ser humano y el ganado es baja. Por lo que las tres primeras especies podrían necesitar hierro para poder tener un mecanismo de defensa compensatorio alternativo. Cabría preguntarse si la inmunidad nutricional es sólo por el hierro o si pueden estar involucrados otros nutrilitos, como el fósforo inorgánico (Pi) o el zinc. En respuesta a esto se ha observado que en los seres humanos infectados con bacterias gramnegativas (no en el caso de grampositivas), se reduce la cantidad de Pi en el plasma a niveles subóptimos para el crecimiento bacteriano (Weinberg, 1974). Por otra parte, la invasión microbiana o tras la inyección de sus productos en humanos y animales, ambos se tornan rápidamente hipocincémicos (Figura I-1). Los hongos requieren más zinc que las bacterias para su crecimiento, y pacientes con altas cantidades de este metal en plasma (por administración intravenosa) o en su orina (en caso de la diabetes), tienen alta incidencia de infección por Candida en los tejidos o el tracto urinario respectivamente. El crecimiento de células tumorales requiere zinc, por lo que la respuesta hipocincémica de algunos pacientes con neoplasias podría ser un mecanismo de defensa. Además, la deficiencia dietética de zinc en ratas produce la inhibición del crecimiento de una variedad de tumores. Puesto que el crecimiento rápido de células tumorales requiere tanto hierro como zinc, ¿podría ser posible obtener remisión del crecimiento tumoral por repetidas inyecciones de agentes hipoferrémicos o hipocincémicos?. Se ha conocido que los microbios o sus productos causan una reducción en el grado de crecimiento de algunos tumores, por ejemplo, el BCG ha sido administrado a seres humanos como auxiliar experimental en la quimioterapia de tumores (Sparks et al., 1973). -Vacunas. El concepto e incluso las primeras prácticas rudimentarias de intentar prevenir las enfermedades mediante vacunación tiene ya 3000 años. Investigaciones históricas han revelado que en China e India se practicaba la variolización para proteger personas contra la viruela desde antes del año 1000 A. C. usando pequeñas cantidades de material obtenido de las pústulas de la enfermedad. Sin embargo el verdadero y decisivo paso racional y científico hacia la vacunación lo dio Edward Jenner en 1798 en Inglaterra al concluir sus trabajos y demostrar que los individuos vacunados con gérmenes de la viruela de vaca resistieron la epidemia de viruela que azotó esa región del mundo en ese tiempo; 100 años después Pasteur realizó geniales contribuciones introduciendo las vacunas contra el antrax y la rabia. A 3000 años de los rudimentos del concepto y la práctica de la inmunización, a 200 de los trascendentales descubrimientos de E. Jenner y a 100 de las contribuciones gigantescas de Pasteur, se puede asegurar que la inmunización es la medida más efectiva y de mejor balance costo-beneficio con que cuenta hoy día el arsenal médico para enfrentar el embate de numerosas y devastadoras enfermedades (Sierra y Acosta, 2000). La mayoría de las vacunas incluidas en los programas de vacunación de los diferentes países y en el Programa Ampliado de Inmunización de la Organización Mundial de la Salud (OMS) son vacunas desarrolladas hasta la década de los años 60, las cuales constituyen un grupo de preparados vacunales obtenidos por métodos empíricos y que fueron desarrollados en una época de relativo poco desarrollo de especialidades como la microbiología, bioquímica e inmunología (Levine, 1990). Estas vacunas que constituyen la columna vertebral de los programas de inmunización actuales y que han brindado un aporte de inestimable valor a la prevención de las enfermedades infecciosas se definen como vacunas convencionales. Clasificación de las vacunas convencionales Dentro de las vacunas convencionales se definen 3 grupos principales: vacunas de subunidades, vacunas de gérmenes inactivados y vacunas vivas atenuadas. a) Vacunas de subunidades Las vacunas de subunidades están constituidas por preparados basados en la presencia de un constituyente obtenido a partir de un microorganismo. Dicho constituyente es habitualmente de índole proteica, aunque algunas de estas vacunas están constituidas por polisacáridos. La selección del componente incluido en la vacuna está determinado por el conocimiento de la patogenia de la infección en cuestión, así como por el descubrimiento de los mecanismos inmunológicos asociados con la protección frente a la misma. Dentro de las vacunas de subunidades proteicas constituyen un ejemplo destacado los llamados toxoides, que no son mas que formas inactivadas de toxinas bacterianas como la tetánica y la diftérica capaces, después de su administración, de inducir anticuerpos neutralizantes antitoxina (Levine, 1990). En este grupo también se incluyen las vacunas de primera generación contra la hepatitis B que están constituidas por el antígeno de superficie del virus de la hepatitis B (HBsAg) el cual se purifica a partir del plasma de individuos infectados (Maupas, 1976). Dentro de las vacunas de subunidades polisacarídicas se encuentra la primera generación de vacunas contra el Haemofilus influenzae b, constituída por su polisacárido capsular (PRP) (Anderson, 1990) b) Vacunas de gérmenes inactivados Las vacunas de germenes inactivados están constituidas por preparados que se obtienen mediante la inactivación de los microorganismos causantes de determinada enfermedad los cuales al ser administrados muertos tienen la propiedad de inducir estados de protección. Dentro de esta categoría se incluyen el componente Pertussis de la vacuna DPT y la vacuna antipolio de virus inactivados (Galazka et al., 1984). c) Vacunas vivas atenuadas Las vacunas vivas atenuadas están constituidas por gérmenes vivos con incapacidad de producir enfermedad en individuos sanos, pero con una capacidad variable de inducir protección. Estas vacunas se obtienen mediante la utilización de microoorganismos no patógenos relacionados con el germen causante de la enfermedad o mediante la atenuación de patógenos virulentos después de pases repetidos en medios de cultivo no habituales o por especies que no constituyen sus hospederos naturales (Charles y Dougan, 1990). Esta forma de atenuación trae por consecuencia que no se conozcan las mutaciones que condujeron a su atenuación, lo que dificulta su control de calidad y mantiene siempre latente el riesgo de reversión a la virulencia. - Vacuna antimeningocócica BC VA-MENGOC-BC® Esta vacuna preparada a partir de proteínas de la membrana externa de NmB y polisacáridos de NmC, elaborada a fines de la década de los ochenta, fue capaz de controlar la epidemia de meningitis meningocócica que afectó a Cuba en esta época (Rico et al., 1996). Posteriormente ésta ha sido utilizada en diferentes países, Brasil (Milagres et al., 1994), Argentina (Dardo y Marin, 1994), Colombia (Galeano y Echevarry, 1995) y Nigeria (Almeida et al., 1997). VA-MENGOC-BC® recibió patente en Cuba en el año 1987 (Campa et al., 1987) y en otros países como Canadá, Japón, Austria, Argentina, Noruega, Federación Rusa, Estados Unidos de Norte América, y recientemente por la Oficina Europea de Patentes, estando protegida en 12 estados europeos: Alemania, Austria, Bélgica, España, Francia, Grecia, Holanda, Italia, Luxemburgo, Suecia, Suiza y Reino Unido. Hace 12 años, en 1987, se utilizó por primera vez masivamente en Cuba y en el mundo un vacuna contra el meningococo B, hasta ahora esta vacuna (VA-MENGOC-BC) es la única disponible. Durante el período 1997-1998, fueron monitoreados los eventos adversos postvacunales, demostrándose su baja incidencia (Debbagr, 1994). Durante 1996 y 1997 se han realizado nuevas campañas en el Brasil (Amazona, Espíritu Santos y Río Grande del Sur). La vacuna antimeningocócica cubana, única el mundo contra el serogrupo B, detuvo la epidemia y mantiene controlada la enfermedad meningocócica en Cuba. El balance de los resultados en la evaluación del impacto, efectividad y eventos adversos en otros países, demuestra que esta vacuna es segura y eficaz (Registro Sanitario de VA-MENGOC-BC®, 1989). La bacteria gramnegativa Neisseria es responsable de uno de cada tres casos de meningitis de tipo bacteriana en el mundo, siendo la especie humana su hospedero natural (Devoe, 1982). Las cepas de esta bacteria están subdivididas en tipos inmunológicos, de acuerdo con la presencia de epítopes específicos correspondientes a proteínas de la membrana externa (Frasch, 1985). Una de las vías para la obtención de un preparado vacunal de amplio espectro de protección, es la identificación de proteínas altamente conservadas en todas las cepas, para producir anticuerpos inmunológicamente activos (anticuerpos bactericidas). La Proteína P64K (MR 64 KDa) se encuentra presente en la membrana externa de las cepas mayormente patogénicas de Nm y es capaz de inducir anticuerpos bactericidas en su hospedero natural (Guillén, 1994), (Dlawer y Ala’Aldeen, 1996). -Proteínas de membrana externa El conocimiento de la estructura conformacional y la inmunoquímica de las proteínas de membrana externa (PME) clase 1, es una premisa importante para el uso de péptidos sintéticos a fin de inducir anticuerpos que puedan reaccionar con la proteína nativa y promover la lisis mediada por el complemento del meningococo, ya que las proteínas de la membrana externa han demostrado cierta protección contra la infección meningocócica del serogrupo B en recientes pruebas de campo (Bjne et al., 1991). Se han empleado péptidos sintéticos que contienen epítopes del subtipo P1.16b. Se ha evaluado su inmunogenicidad y la actividad biológica de los anticuerpos inducidos por ellos. En un trabajo inicial realizado por Christodoulides y Heckels, (1994), se sintetizaron péptidos lineales de 9 y 15 aminoácidos. Estos fueron usados para la inmunización con acoplamiento y sin él a una proteína portadora. Los péptidos indujeron anticuerpos con la especificidad de los epítopes deseados, pero su reacción con la proteína nativa de superficie fue pobre y no fueron bactericidas. Consecuentemente, un péptido 36 mer (que están formados por un asa 4 expuesta en la superficie completa de la proteína clase 1) fue sintetizado y sujeto a ciclización, con la intención de restringirlo a conformaciones que pudieran semejar más a la estructura del asa nativa. A diferencia del antisuero obtenido contra los péptidos lineales, los anticuerpos aumentaron por inmunización con el péptido cíclico 36 mer, pareciendo reconocer los determinantes conformacionales. Además, promovieron, a través del complemento, la muerte bacteriana de la cepa de meningococo homóloga P1.16b. El sistema de péptidos de antígenos múltiples (PAM) puede constituir una alternativa para preparar pépticos con alguna estabilidad conformacional. Inicialmente fue sintetizado un PAM octamérico que contenía el epítope P1.16b de células B, pero se encontró que no era inmunogénico en animales, lo cual vislumbró la necesidad de un epítope de células Th efectivo para tratar de inducir una respuesta inmune humoral. Entonces se sintetizó un PAM (BT-PAM) para contener el epítope P1.16b en tandem con un epítope de células Th, bien caracterizado, de la toxina tetánica según Panina-Bordignon, et al.,(1989). El antisuero de conejo no solo promovió la muerte bacteriana mediada por el complemento de la cepa meningocócica subtipo P1.16b, sino también la del subtipo P1.16a. Este estudio muestra el potencial de los péptidos sintéticos para la producción de respuestas inmunes protectoras contra Nm (Christodoulides y Heckels, 1994). De los resultados actuales con las vacunas basadas en la proteína de membrana externa experimental, se podría concluir que una vacuna perfeccionada debe dirigir la respuesta inmune específicamente hacia aquellos determinantes antigénicos que pueden inducir una respuesta inmune protectora (Zollinger et al., 1991). Según Valcárcel et al., (1991), las proteínas de la membrana externa (PME) y otros componentes, han sido evaluadas en Cuba. La experiencia mundial describe la existencia de numerosos serotipos patógenos del serogrupo B, diferentes entre sí. Se han encontrado la coexistencia de varios de ellos en algunos países. En Cuba a pesar de haber alrededor de un 90% de serotipos B:NTP1.15 (después comprobado como B4:P1.15), existen otros en menor cuantía. Debido a la pobre respuesta bactericida, su rápido declinar y el estrecho espectro del modelo de vacuna recomendado por Carl Frasch, se realizó investigaciones paralelas. Basados en estudios inmunoquímicos de los antígenos de la cepa cubana. Se demostró que varias proteínas de alto peso molecular constituían buenos inmunógenos, aun a baja concentración, en preparaciones de antígenos PME y en extractos enriquecidos, estimulando la producción de anticuerpos bactericidas con muy buena memoria reactiva. Este complejo antigénico (peso molecular 65 a 95 kDa) poseía un grupo de proteínas con unidad antigénica contra todos los serotipos del serogrupo B y capacidad bactericida cruzada contra ellas. Estos resultados llevaron a conformar un nuevo modelo de vacuna cubana constituido por vesícula de proteína de serotipo (85-88%), lipopolisacáridos (± 10%) y fosfolípidos, que funciona como presentador y adyuvante, estabilizado que presenta el complejo antigénico de alto peso molecular (CAPAM) (12 - 18%) del total de proteínas, y aportan los epítopes útiles de P1. Este CAPAM, que posee un peso molecular entre 65-95 kDa, conjuntamente el gel de aluminio que actúa como adyuvante extra, estabilizador y un polisacárido que mejora la solubilidad, conforman una vacuna polivalente que después de exámenes de laboratorio, pruebas en animales y seres humanos voluntarios fue objeto de protección como propiedad industrial en diecinueve países. -Sistema Nervioso Central (SNC). Dentro de la temática de esta tesis el SNC constituye un sistema de singular importancia, si tenemos en cuenta que la enfermedad meningocócica afecta el mismo y por tanto a los biomodelos por lo consideramos necesario revisar los aspectos desarrollados a continuación. . Composición celular del SNC. El SNC esta constituido por el parénquima celular donde se encuentran las neuronas y la neuroglia. Literalmente se llama neuroglia al tejido de sostén, pero nuestro conocimiento de las múltiples funciones de estas células, mientras que éstas se encuentran más allá de la simple función de unir el tejido. La neuroglia esta compuesta por macroglia y microglia. La macroglia está formada por los astrocitos y oligodendrocitos. El tercer tipo celular que pertenece a este tejido es conocido como células microgliares. Las células gliales aunque más numerosas que las neuronas han sido relegadas a un segundo orden de importancia. Esa perspectiva ha cambiado algo ahora y se le concede un gran peso al rol de la neuroglia en el funcionamiento normal del SNC. Los astrocitos, llamados células estrelladas, término que es más aplicable a la variedad fibrosa con su pequeño cuerpo delgado y largo proceso. Los oligodendrocitos tienen un pequeño cuerpo y un escaso proceso ramificado. De nuevo el término oligodendroblastos se reserva a una forma inmadura que aparece con poca frecuencia. Las células microgliales tienen un pequeño cuerpo y un escaso proceso ramificado en estado de reposo. El cerebro y la médula espinal son revestidos por las meninges (dura, aracnoide y pia), las cuales proporcionan protección a un compartimiento donde fluye el líquido cerebroespinal que circula a través del espacio subaracnoideo acompañada por vasos sanguíneos y la vaina de los nervios craneales y espinales. Dentro del encéfalo y la médula espinal están el sistema reticular y el canal central, los cuales son delineados por las células ependimarias y el plexo coroideo, el cual produce el líquido cefalorraquídeo (LCR). La circulación del LCR se realiza a través de los ventrículos laterales tercero y cuarto dentro del canal central o mediante las aperturas laterales en el centro del ángulo cerebelomedular situado en el espacio subaracnoideo del cerebro. El LCR en el espacio subaracnoideo desagua dentro de la vía especializada llamado aracnoide granuloso, situada en los senos venosos intracraneales, con algunos desagües dentro de los plexos venosos asociados con nervios craneales y espinales. El LCR pudiera estar en contacto con la superficie del parénquima adyacente. Algunos términos utilizados en la neuropatología. La estructura básica en el estado de espongiosis frecuentemente requiere del examen a través del microscopio electrónico, para un diagnóstico inequívoco. Gliosis es una respuesta de la neuroglia en el SNC para muchas formas de daño. Esta tiene importancia en la inflamación del SNC por isquemias, traumatismo, tóxicos y otros daños capaces de activar la neuroglia, particularmente la astroglia y la microglia. Cuando un área del SNC esta gliótica es hipercelular porque las células gliales están hipertróficas o hay proliferación celular o ambos fenómenos. El incremento del número de células puede deberse a la proliferación “in situ” o al reclutamiento de células gliales que pueden migrar. Los estudios de transplante han mostrado que los astrocitos inmaduros tienen una considerable capacidad para migrar dentro del SNC.(Goldberg y Bernstein, 1988). Los estudios de la respuesta de células gliales dentro de un rango de estímulos sugieren que la gliosis resulta por la proliferación de células y es más característico de la microglia que de la astroglia (Graeber y Bernstein, 1988),(Matsumoto et al., 1992) Microgliosis es la proliferación de células microgliales en la cual estas toman un aspecto alargado y delgado perfil. Por tradición cuando hay una disposición poco específica de estas células reactivas, se denominan racimos focales y gliosis arborescente incluyendo la microglia, donde ellas se agregan en la capa dendrítica de las células cerebelosas de Purkinge. La gliosis que produce la generación de macrófagos es comúnmente completada por la circulación, como ha podido demostrarse por la respuesta disminuida, después de la irradiación de la médula osea (Morshead y Van der Kooy, 1990). La reactividad astrocítica es la astrocitosis o astrogliosis. Estos términos son frecuentemente usados, aunque en la astrogliosis concurre un incremento en el número de procesos fibrosos ricos en filamentos. La astroglia y la microglia pueden elaborar factores de crecimiento, son ejemplo, la astroglia produce mitógeno para células microgliales y viceversa, y estos son probablemente completados por otras fuentes del SNC (Frei et al., 1986) -Inmunobiología e inmunopatología del Sistema Nervioso Central. Un asunto que ha intrigado los investigadores durante décadas, es si existe comunicación entre el sistema inmune y el SNC; qué saben cada uno del otro (Besedovky et al., 1983). Existen varias evidencias que sugieren una relación recíproca. 1. En 1964 Reif y Allen (Reif y Allen, 1985) describieron un antígeno THY-1 en ratones, el cual es común al cerebro y a los órganos linfoides. Actualmente se sabe que existen varios determinantes antigénicos compartidos entre las células hematopoyéticas y las células nerviosas normales o neoplásicas. 2. El daño experimental del hipotálamo tiene efectos transitorios sobre la respuesta inmune, incluyendo disminución de la actividad de los macrófagos supresores y de los linfocitos asesinos. (Cross et al.,1984) la cual sugiere una capacidad para la modulación neural de la función inmune.( Roszman y Brooks, 1985). 3. Los órganos linfoides (ganglios linfáticos, bazo, timo) y la médula osea roja están inervados por fibras de los nervios linfáticos y las células inmunes tienen receptores para los neurotransmisores químicos y las hormonas.(Besedovsky et al., 1983). 4. La exposición a antígenos extraños trae consigo un aumento en la frecuencia de impulsos de las neuronas ventromediales del hipotálamo.(Besedovksy et al., 1983). -Inflamación del SNC La leptomeningitis bacteriana en neonatos se caracteriza por la secreción de un fluido rico en fibrina y leucocitos polimorfonucleares hacia los compartimentos ventriculares y leptomeningeos. Esta es a menudo letal debido a la inflamación del cerebro y la compresión consecuente del tallo encefálico, antes de que los cambios inflamatorios agudos hayan progresado más. Otras enfermedades inflamatorias del SNC se caracterizan por proliferación e hipertrofia de los elementos gliales - específicamente la microglia y astroglia - además de manguitos perivasculares, con una salida mínima de fluidos y proteínas. Estas características son típicas de un grupo importante de encefalomielitis virales. El libre acceso de los constituyentes sanguíneos hacia el tejido nervioso esta impedido por la barrera hemato-encefálica del endotelio capilar. El SNC carece de células dendríticas especializadas en la presentación de antígenos y expresión intrínseca de moléculas del Complejo Mayor de Histocompatibilidad, especialmente de clase 2 que sobre todo es baja.(Schawendemann et al., 1983), Andersson et al., 1992a ), no obstante, estas características no parecen ser un impedimento para el desarrollo de procesos inflamatorios en el neuroaxis. No hay sistema linfático dentro del tejido nervioso, pero las células y los antígenos en el SNC drenan a la circulación y a los ganglios linfáticos cervicales.(Cserr et al., 1992). Los signos cardinales de la inflamación del SNC son: manguitos perivasculares, gliosis, satelitosis neuronal y neuronofagia. Existe un compartimiento perivascular, ya sea real o potencial alrededor de todas las arterias, arteriolas, vénulas y venas del SNC, este compartimiento perivascular se encuentra entre las paredes de los vasos sanguíneos y los límites de la glia, contiene LCR y normalmente se ve como una continuidad del espacio subaracnoideo. En los animales domésticos los capilares de la médula espinal y de algunas áreas del tallo encefálico también poseen espacio perivascular (el cual puede contener escasas fibras de colágeno), mientras que en otras áreas del cerebro el espacio vascular alrededor de los capilares esta obliterado. También hay evidencias de que las células piales rodean los vasos sanguíneos en el espacio subaracnoideo. Se sugiere que en la cara arterial, esta vaina pial se extienda hacia el interior del cerebro produciendo un espacio periarterial separado. Esta capa pial es fenestrada, lo cual permite la comunicación entre los espacios periarterial y perivascular. Una característica típica de inflamación del SNC es el manguito perivascular, que lo constituyen las acumulaciones de leucocitos de uno o múltiples tipos en el espacio perivascular. La composición del manguito puede variar durante el curso de la enfermedad y está influenciado por el uso de algunos medicamentos. La mayoría de las células de los manguitos perivasculares son de origen hematógeno, pero no son las únicas constituyentes. Los pericitos vasculares del SNC y las células de la adventicia son capaces de redondearse para formar células mononucleares con probable actividad fagocítica y presentadora de antígenos. Puede llegarse a algunas generalizaciones con respecto a la composición de los manguitos perivasculares. En las enfermedades bacterianas predominan las células polimorfonucleares con un componente menor de células mononucleares. En las infecciones fungosas las proporciones se invierten, a menudo con macrófagos epitelioides hinchados que pueden alojar el agente causal. Las infecciones virales se manifiestan por una población rica en linfocitos con células plasmáticas y monocitos. Algunas infecciones por arbovirus incitan una respuesta celular polimorfonuclear, como la Encefalomielitis Equina Oriental, que es un buen ejemplo. La gliosis es el aumento de la prominencia de las células gliales como resultado del hinchamiento citoplasmático, la adquisición de más procesos celulares, de la proliferación celular o de todas estas. La gliosis puede ser isomórfica, en la cual las células gliales siguen el patrón normal de disposición en el tejido (como puede verse en el cerebro envejecido), o anisomórfica en la cual la acumulación es desordenada. La última es mucho más común y son un indicio importante para el microscopista en la detección de áreas discretas de tejido que aparecen inapropiadamente hipercelulares. Muy a menudo lo que se designa como gliosis contiene tantas células circulatorias como endógenas (Morshead, 1990). -Agentes infecciosos e inflamación del SNC. No se conocen ejemplos de enfermedad neurológica primaria entre los animales domésticos, ni encefalolimielitis causada por mycoplasma, aunque en algunas infecciones causadas por mycoplasma se produce meningitis como parte de la poliserositis producida por estos agentes. Los virus constituyen el agente infeccioso neuropatógeno más estudiado. Muchas encefalitis se presentan como parte de una infección más generalizada, pero el compromiso del SNC es comúnmente la causa de morbilidad y mortalidad. Los virus neuropatógenos incluyen el DNA y RNA virus convencionales, los retrovirus que son algo más atípicos y se incorporan en los genomas del hospedero y (usan el término menos rigurosamente) el grupo no clasificado scrapie. La familia de los retrovirus contiene tres subfamilias. Miembros de las tres Oncovirus, Lentivirus y Espumavirus son subfamilias que pueden invadir el SNC (Georgsson et al., 1991), pero solo las oncovirus y los lentivirus se asocian con enfermedades neurológicas de aparición espontánea o frecuentemente, estos agentes son neuropatógenos en el hombre y en un amplio rango de animales. -Meningitis y Abscesos cerebrales. Por definición la leptomeningitis es la inflamación de la piamadre y la aracnoides, mientras que la paquimeningitis es la inflamación de la duramadre. Son registrados casos esporádicos en animales adultos (Andersson et al., 1992)a algunas veces causados por gérmenes aerobios (Andersson et al., 1992)b . La mayoría de los casos de leptomeningitis canina son estériles como respuesta a corticoides y se piensa que son mediados por el sistema inmune. Otras bacterias que producen cambios anatomopatológicos similares pero solo en ciertas especies son: - En cerdos Hemofilus swis en la enfermedad de Glässer y Salmonella dublin (Sedgwick et al., 1991). -Aspectos regulatorios para productos biotecnológicos. La fabricación y control de los medicamentos ha sido desde sus inicios una de las grandes preocupaciones del hombre y por ello las administraciones públicas, sintieron la necesidad de legislar normativas que velasen por la calidad, eficacia e inocuidad de los mismos. En cualquier país la implementación de las Buenas Práctica de Laboratorio (BPL) constituye un requisito indispensable para garantizar la integridad de los estudios y la comercialización de los medicamentos, tanto nacional como internacionalmente (Pérez y Ferrándiz, 1998). Las evaluaciones preclínicas constituyen una de las fases críticas en la ruta de desarrollo de un medicamento (Hernández et al., 1998). Otros aspectos a considerar son los preservantes, adyuvantes y excipientes del formulado. Se conoce que el modelo animal adecuado no está siempre disponible y que la respuesta en tales modelos no siempre es predictiva de la respuesta humana (Díaz y Agramonte, 1998). Los adyuvantes son compuestos de naturaleza química diversa, capaces de potenciar las respuestas inmunitarias. Se han utilizado con este fin desde hace más de seis décadas, pero en la actualidad ha crecido el interés por su utilización, toda vez que las nuevas tendencias en vaccinología son las de desarrollar vacunas cada vez más específicas, pero que generalmente son menos inmunogénicas por lo que necesitan de adyuvantes para incrementar su inmunogenicidad. Sin embargo, el mayor inconveniente en la utilización de estos, está relacionado con los efectos adversos que producen, por esta razón existe una gran resistencia por parte de las entidades regulatorias para aceptar nuevos adyuvantes usados en vacunas humanas y animales. Cuba está alcanzando un desarrollo acelerado en materia de vacunas y esto también implica la búsqueda incesante de nuevos candidatos a adyuvantes, de los cuales no basta con demostrar sus propiedades inmunopotenciadoras, sino también es imprescindible una evaluación toxicológica integral, en aras de lograr su aceptación (Batista y Pascual, 1998). Como resultado de esta revisión bibliográfica podemos plantear que existe abundante información donde se precisan aspecto de mucha utilidad con relación a los tópicos que aborda en la tesis planteada, tales como: epidemiología y aspectos clínicos de la enfermedad meningocócica, modelos animales y su aplicación en el desarrollo de la biotecnología, utilización de los biomodelos en la investigación de vacunas y sueros, papel de hierro en la microbiología, vacunas, SNC, inmunobiología e inmunopatología del SNC, histoquímica e inmunohistoquímica, meningitis y absceso cerebral y aspectos regulatorios para productos biotecnologícos, todo lo que nos permitió profundizar en el conocimiento de estos tema. PARTE EXPERIMENTAL Capítulo II MATERIALES Y MÉTODOS GENERALES: Ver Diagrama General. (Reverso página anterior) II-1. Animales: Todos los animales eran de la categoría convencional procedentes del Centro Nacional para la Producción de Animales de Laboratorio (CENPALAB) y fueron recibidos junto con los correspondientes certificados de calidad higiénico sanitaria y genética. - Ratones de la línea Balb/cj de 18 -22 g, con 6-10 semanas de edad, de ambos sexos, alojados en cajas T2 en grupos de cinco animales. - Ratas recién nacidas de la línea Sprague Dawley, con edad comprendida entre 3 y 5 días de nacidas de ambos sexos, con sus correspondientes madres, alojados en cajas T2 en grupos de ocho animales con su correspondiente madre. - Ratas convencionales de la línea Sprague Dawley, con edad de 12 semanas y peso de 150 200 g, de ambos sexos, alojados en cajas T3 en grupos de cinco animales. II-2. Procedimientos de tenencia y manejo de los animales. Se mantuvo un régimen de iluminación de 12 h luz y 12 horas de oscuridad. La temperatura ambiental fue de 222ºC y la humedad relativa de 6010. El alimento fue suministrado por el CENPALAB, del tipo roedores, con sus correspondientes certificado de calidad bromatológica, e higiénico sanitaria, tanto estos como el agua acidulada con ácido clorhídrico a un pH de 2.7-2.9. fue suministrada “al libitum” El encamado consistió en bagazo del desmehollado de la caña de azúcar, previamente autoclaviado a 121 o C y 1.5 Atmósferas de presión Los cambios se realizaron semanalmente. Tanto los animales fueron sometidos a un período de adaptación nunca menor de 15 días en los locales donde posteriormente serían realizados los ensayos, según las características de cada especie. II-3. Etica. Todos los protocolos del ensayo, incluyendo los métodos de eutanasia, estuvieron sometidos a la consideración, análisis y aprobación de la comisión de ética en el ámbito institucional observando lo establecido por las regulaciones de seguridad biológica. II-4. Métodos de inoculación. Los animales fueron inoculados por vía intraperitoneal (I. P) tomando los ratones y las ratas recién nacidas por sus extremidades posteriores, quedando suspendidos con la cabeza hacia abajo, procediendo a realizar la punción a un lado de la línea alba en la pared abdominal. Esta se realizó con aguja de calibre 23 G X 1 pulgadas de longitud y jeringuillas desechables esteriles de 1 mL de capacidad, mientras que para las inoculaciones intramusculares de la vacuna se utilizaron dosis de 0.05 mL aplicadas en la cara interna de una de las extremidades posteriores. II-5. Factores estimulantes de la virulencia (F. E .V). Dextrana Férrica: Hierro - Dextrana (ENSUMEFA) Lote No. 4490 de 1990. Diluido hasta la concentración deseada en Solución Salina Fisiológica. Las mezclas fueron realizadas momentos antes de su inyección Para hacer los ratones susceptibles a Nm también se utilizó en el momento de la inoculación de las bacterias, una mezcla de dextrana férrica (Hierro dextrana, ENSUMEFA, 2 mg de hierro/mL, 0.25 mL) y mucina (Mucina gástrica de cerdo, SIGMA, 8%, 0.4 mL) Lote 55H0130 . Se utilizó la vía intraperitoneal (IP). II-6. Inoculación de gérmenes y FEV. Los inóculos de los gérmenes se administraron por vía intraperitoneal (IP) con jeringuillas de cristal de 1 mL y agujas 23G x 1. II-7. Características químicas de la vacuna VA-MENGOC-BC. La vacuna VA-MENGOC-BCestá compuesta por: - Proteínas de Neisseria Meningitidis del serogrupo B. - Polisacáridos de Neisseria Meningitidis del serogrupo C. Cada dosis de 0.5 mL contiene: Proteínas purificadas de la membrana externa 50 g. Polisacárido purificado 50 g. Gel de Hidróxido de Aluminio 2 mg. Tiomersal 0.05 mg. Cloruro de Sodio 4.25 mg. Fosfatos 0.05 mg Agua para inyección c.s.p 5 mL. II-8. Características de la Inmunoglobulina Antimeningocócica BC. Se empleó una Inmunoglobulina Antimeningocócica específica hiperinmune, elaborada por el Instituto Finlay (Ciudad de La Habana, Cuba) cuyo objetivo es comprobar de manera indirecta la eficacia de VA-MENGOC-BC. Con las siguientes características. Fuente: plasma de donantes voluntarios inmunizados con vacuna cubana antimeningocócica BC ( VAMENGOC.BC®). Composición: 50 mg de Inmunoglobulina Antimeningocócica BC por mL estabilizada con albúmina humana. Título bactericida medio: 1/264. Actividad específica media: 21 678 U/mL. Pureza electroforética: Más del 90% de IgG monomérica. Controles de VIH y del VHB: negativos. Controles de esterilidad y seguridad: satisfactorios. Pirogenicidad: apirogénica II-9. Germen y Preparación de los inóculos. Se empleó la cepa 385 de NmB (perteneciente al cepario del Instituto Finlay), realizando un cultivo en placa (a partir de la cepa liofilizada en Sacarosa Gelatina) e incubada a 37ºC y atmósfera de 5 CO2, durante 18-24 horas. Las colonias formadas fueron inoculadas en 100mL de caldo Mueller-Hinton ajustado a una densidad óptica de 0.2 y mantenidas en zaranda orbital a 170rpm, a 37ºC durante 6 horas. Transcurrido este tiempo se realizó tinción de Gram y centrifugación del cultivo a 7000 rpm, durante 10 min a una temperatura de 20 ºC. El sobrenadante fue eliminado y el pellet se resuspendió el pellet en 25 mL de PBS + Gelatina al 1 y procediendose a centrifugar en las mismas condiciones. Repetiendo este paso, pero resuspendiendo esta vez en la mitad del volumen, ajustando finalmente la suspención bacteriana a una concentración de 1x1010 UFC/mL, apartir de la cual se realizaron diluciones seriadas hasta obtener las concentracione deseadas (Sotolongo, 1995). II-10. Observación clínica. Los animales fueron mantenidos en observación después de terminada la inoculación, teniendo en cuenta que las muertes sucedidas durante las primeras 4 horas se consideraron debidas a la técnica operatoria. Después se observaron cada doce horas por espacio de una semana, registrando el número de muertes y síntomas en general y en particular erizamiento del pelo, postración, inapetencia, agrupamiento, diarreas y conjuntivitis. II-11.Obtención de sangre y líquido cefalorraquídeo de animales inoculados con el germen. La evolución de la bacteriemia se determinó por extracción intracardíaca de sangre. El líquido cefalorraquídeo (LCR) fue obtenido por punción en la Cisterna Magna, con un equipo de inyección hipocraneal No 27. Los frotis o extensiones fueron realizados en láminas porta objeto coloreados con coloración de Gram. II-12. Estudios anatomopatológicos . Para los estudios anatomopatológicos se aplicó la técnica descrita para el sacrificio y la necropsia por Castillo (1985), las alteraciones macroscópicas fueron recogidas en protocolos establecidos al efecto mientras que para los estudios histopatológicos se fijaron los fragmentos seleccionados de órganos y tejidos en formalina neutra al 10 %. Se procesaron por la técnica de inclusión y cortes en parafina, y se colorearon con Hematoxilina Eosina Periodic acid of Shiff (P.A.S) y Van Giesson. El cráneo fue separado inmediatamente después de producirse la muerte por un corte transversal en el ámbito cervical y se despojó de la piel y el maxilar inferior, con una tijera de microcirugía, se realizaron incisiones de los huesos craneales. Removimos ligeramente la tapa ósea formada sin eliminarla (para conservar las meninges). Se realizaron cortes en planos longitudinales. La identificación de las muestras fue codificada para realizar las evaluaciones histopatológicas a doble ciegas. II-13. Procesamiento estadístico. Cálculo de la protección Se realizó el cálculo de la protección teniendo en cuenta la siguiente fórmula: P=100(1-Mt/Mc ) Mt : mortalidad de los tratados Mc : mortalidad de los controles Para el análisis de los resultados se utilizó el programa Statistica. La comparación de los tiempos de sobrevivencia se llevó acabo mediante el método Long-rank (todos contra todos con niveles de significación de 0.01,0.05 y 0.1). Estadísticamente se aplicó un análisis de Varianzas ANOVA, de clasificación triple sin interacción para explicar las variaciones de los niveles de anticuerpos según sexo, vía de inoculación y estado (placebo y vacunado) y prueba t de Students para la comparación de medias entre grupos y pruebas de comparación de proporciones para la bacteremia. II-14. Determinación de la dosis letal 50: Para cada ensayo de DL50 se formaron aleatoriamente 6 grupos de 10 animales cada uno. Estos fueron tratados con FEV (Dextrana Férrica 1mg/mL 0.5mL, IP) y diluciones seriadas de las suspensiones de bacterianas en concentraciones desde 1x1010 hasta 1x10 5 UFC/mL (0.2mL/Aniama, vía IP). En todos los casos se confirmaron las concentraciones bacterianas mediante conteo de viables en plancas. Se registraron las muertes ocurridas hasta 72 h post-inoculación y se determinaron las mortalidades acumuladas para cada dosis de germen. Con los puntos de dosis - mortalidad acumulada se buscó la curva de mejor ajuste al modelo de Emax sigmoidea 100dosis n [ Mort(%) ] para obtener la DL50 directamente a partir de la ecuación de DL50 n dosis n regresión (Holford, 1991). II-15. Procedimiento de las técnicas de ELISA.. Se cuantificaron las densidades ópticas (DO) en los grupos tratados con respecto al control positivo (Placebo). La determinación de anticuerpos se realizó según metodología previamente descrita por Sotolongo (1995).En esta se usaron placas de poliestireno de 96 pocillos de fondo plano (Maxisorp, Nunc, E.U.), las cuales se recubrieron con 100 l de vesículas de la membrana externa (VME) 20g/ml en tampón Na 2CO3-NaHCO 3, 0.1 M pH 9.6 y se incubaron a 4º C durante toda la noche. Después de lavar con Agua-Tween 20 (Sigma St. Louis, MO, E.U.), al 0.05% (esta solución se usó para todos los lavados), se incubaron las muestras problemas y los controles (100l) (Oxoid, UK) al 3 % en TFS durante 1 h a 37º C. Se lavó 3 veces, se añadieron 100l/pozo del conjugado IgG – fosfatasa alcalina (Sigma St. Louis, MO, E.U.) diluido 1:20. Se lavó 3 veces y se reveló añadiendo 100 l/pozo de H2O2 0.15 % más O-fenilendiamina (Sigma St. Louis, MO, E.U.). Se detuvo la reacción con 50 l/pozo de H2SO 4 2 M y se midió la absorbancia a 405nm en un lector Titertek Multiskan. II-16. Metodología de trabajo. Todas las operaciones fueron ejecutadas ajustándose a las Buenas Prácticas de Laboratorio (BPL) establecidas en los Planes Normalizados de Operaciones (PNO) elaborados por la Dirección de Aseguramiento de la Calidad del Instituto Finlay. II-17. Recolección y conservación de muestras de suero. Los sueros colectados durante la necropsia de los diferentes grupos en estudio fueron guardados a -20para la posterior evaluación. II-18. Prueba de vacunación: Animales de 4-5 semanas de edad y 15 - 17g de peso vivo fueron separados al azar en dos grupos. Uno de ellos fue vacunado con VA- MENGOC- BC®, 0.5 mL., IP y el otro no recibió ningún tratamiento. Al cabo de 21 día, fueron seleccionados aquellos de 18 - 22g de peso vivo. Formando cuatro grupos de ratones vacunados y cuatro sin vacunar. En cada caso dos grupos fueron para seguir la mortalidad (Ensayo de mortalidad), uno para la bacteriemia (Ensayo de bacteremia) y uno para titular los niveles de anticuerpos contra NmB en el momento del reto. Ensayo de mortalidad: Se inocularon 7.4 x 107 UFC/mL (aproximadamente 100 LD50) a un grupo vacunado y a otro sin vacunar; a otros dos similares, 0.5 mL de P.B.S, vía IP (controles). Título de anticuerpos: El día del reto se desangraron 15 animales vacunados y 15 controles por incisión de la arteria subclavia derecha, determinando los títulos de anticuerpos contra NmB mediante técnica de ELISA. II-19. Estudios citológico y bacteriológico del LCR. En las muestras del LCR obtenidas a diferentes intervalos de tiempo se les realizaron extensiones a las que se les practicó tinción de Gram y May Grunwal Giemsa, para los estudios bacteriológicos se realizaron frotis y cultivos en medios específicos para el crecimiento del meningococo. Capítulo III ESTABLECIMIENTO, CARACTERIZACIÓN Y PERFECCIONAMIENTO DE LOS BIOMODELOS RATÓN Y RATA PARA REPRODUCIR LA INFECCION CAUSADA POR N. MENINGITIDIS. Introducción Existen experiencias con los biomodelos para reproducir las infecciones causadas por bacterias que afectan el SNC. Así, han sido utilizados los ratones en las infecciones experimentales causadas por H. infuenzae (Sifontes et al., 1996), (Hugosson et al., 1996,) y Pneumococcus spp (Giebink et al., 1993), de igual modo las ratas recién nacidas han sido empleadas (Nurminen et al., 1992), (Fusco et al., 1998.) Por otra parte, en el caso de Nm los ratones y las ratas recién nacidas han servido de biomodelos para probar infecciones con este germen (Idanpaan - Heikkila et al., 1995), (Stojiljkovic et al., 1995), (Buchanan et al., 1998), (Rubinstain et al., 1998), (Saunder et al., 1999). No obstante demostrar que estos biomodelos pueden utilizarse en las pruebas preclínicas de eficacia de la vacuna VA- MENGOC –BC®, constituía una necesidad a la que había que dar respuesta. Objetivos - Reproducir la infección causada por NmB en el ratón utilizando como Factor Estimulante de la Virulencia la dextrana férrica, caracterizar el cuadro clínico y anatomopatológico en esta especie. - Caracterizar el cuadro clínico y anatomopatológico de la infección en ratas recién nacidas de la línea Sprague Dawley. - Evaluar la utilidad del biomodelo ratón para conocer la eficacia de la terapia inmunológica (vacunas, inmunogenicidad protectogénica activa), frente a NmB. Materiales y Métodos: 1- Animales, condiciones de tenencia, manejo, ética, método de inoculación, la utilización de los Factores estimuladores de la virulencia FEV preparación e inoculación de los gérmenes y la determinación de LD50,se realizaron según se expresa en el Capítulo II Materiales y Métodos Generales, Acápites II (1,2,3,4,5,6,9 y 14). Ver Diagrama Capítulo III. (Reverso página anterior). 2- Formación de grupos experimentales: Se utilizaron un total de 130 ratones en los tres ensayos, para estos se formaron grupos entre 10 y 15 animales, para los estudios de mortalidad 40 (machos y hembras), protección 40, para bacteremia 20 y 30 en el ensayo de títulos de anticuerpos, utilizando siempre el mismo número de animales controles y de sujetos experimentales para comprobar la cinética de la mortalidad, la clínica, bacteremia, presencia de anticuerpos frente a NmB en sangre, estudio anatomopatológico y la protección conferida por VA-MENGOC-BC. Las camadas de ratas recién nacidas a utilizar, fueron separadas de sus madres, se mezclaron y distribuyeron aleatoriamente en cajas correspondientes a los diferentes grupos. Inmediatamente se incorporaron a las cajas que alojaban a las ratas madres, 22 ratas recién nacidas en el ensayo de bacteremia, 20 para la observación del LCR y 76 para los estudios histopatológicos en proporción de 8 a 10 crías por rata. En ese momento las crías se pusieron en contacto con el encamado de sus respectivas cajas para que adquirieran su olor y fueran reconocidas por la rata adulta como hijas suyas, evitando fenómenos de canibalismo (Saukkonen, 1987), (Saukkonen et al., 1989). Los grupos de ratas recién nacidas inoculadas se mantuvieron separados de las no inoculados previendo infecciones cruzadas por aereosoles (Laitenen, 1988). 3- Estudios clínicos: En el caso de los ratones, durante el tiempo que duró el experimento (7 días) se evaluó la aparición de síntomas clínicos en los grupos inoculados y controles, así como el consumo de alimento, agua y la ganancia de peso vivo. Se realizaron evaluaciones del peso vivo antes de la inoculación y cinco días después. Los demás parámetros fueron registrados cada 24 h. Los animales recibieron dextrana férrica en dosis de 1 mg/mL y NmB 1x107 UFC/mL con el fin de determinar curso el clínico de la infección en los biomodelos y la cinética de mortalidad. Los controles fueron inoculados con PBS para otros aspectos del estudio clínico ver Materiales y Métodos Generales, Capítulo II, Acápite II (10). 4- Estudios anatomopatológicos: Ver Capítulo II Materiales y Métodos Generales, acápite II (12). Las ratas y ratones fueron sacrificados mediante una sobredosis de pentobarbital sódico. Se tomaron muestras de encéfalo de los grupos controles y de animales inoculados 2, 3, 5, 6, 7 y 9 días post-inoculación con el fin de conocer la evolución del proceso lesional. Las observaciones macroscópicas fueron realizadas en este momento. Los cráneos fueron sumergidos en formol neutro al 10 % para su fijación. A la 24 h se diluyó la solución fijadora hasta un 5%. Después las muestras fueron sumergidas en formalina descalcificadora durante un período de 48 a 72 h. Los fragmentos se incluyeron en parafina, se cortaron y posteriormente fueron teñidos con hematoxilina y eosina. En el caso de las ratas recién nacidas los cráneos fueron introducidos en formalina descalcificadora por 24 h. Se realizaron de 4 ó 5 cortes coronales a cada cráneo incluyendo en cada sección cavidad cranial, ojos, encéfalo, cerebro y puente. Se realizó inclusión en parafina, cortes histológicos de 4 - 5 micras y tinción con hematoxilina - eosina y Gram. Los ojos fueron fijados en Bouin. En el caso de los ratones el corte del encéfalo se realizó en forma longitudinal y los demás aspectos fueron similares a los realizados en las ratas recién nacidas. El procedimiento de observación microscópica se realizó de igual forma en animales no inoculados (controles) y en los inoculados para evaluar la apariencia de las meninges en estado fisiológico en los primeros y las posibles lesiones en los infectados con NmB, teniendo en cuenta que, como se definió, la meningitis se expresa como la presencia de células inflamatorias en las meninges o en el espacio subaracnoideo (Moxon 1974). El análisis microscópico fue realizado por dos especialistas independientemente y “a ciegas” (muestras codificadas cuya identificación desconocían los observadores). 5 - Obtención de sangre y LCR, metodología de trabajo, recolección y conservación de muestras de suero, estudio de líquido cefalorraquídeo y citología de este, las prueba de vacunación, técnica de ELISA, métodos estadísticos. Ver Capítulo II Materiales y Métodos Generales acápite II(11,15,17 y 19). 6- Para el ensayo de la bacteremia se elige la dosis de 7.4 x 105 UFC/mL, tanto en las ratas recién nacidas (González y Leinonen, 1991), como en los ratones (Pedroso, 1998). Resultados y Discusión A- El ratón como biomodelo para la infección causada por N. meningitidis. serogrupo. Partiendo de las experiencias obtenidas por (Brodeur, 1986), (Sifontes et al., 1999) para los FEV, donde demostraron los efectos beneficiosos de la referida sustancia con el objetivo de facilitar la infección por NmB, decidimos utilizar la dextrana férrica con este objetivo. El rango en que encontramos la DL50 de NmB fue de 5x10 6UFC/mL, que es similar al reportado (Brodeur et al., 1986) para Nm cuando se usa dextrana férrica como FEV. La infección en los ratones se caracterizó por la presencia de piloerección, agrupamiento entre 1 y 2 h postinoculación entre 6 y 8 h presentaron diarreas mucosas de volumen variable, disminución en el consumo de agua y alimentos así como decremento de peso con respecto a los controles. Fue prácticamente constante la presentación de conjuntivitis serosa con tendencia a tornarse purulenta al transcurrir entre 24 y 36 h; llegando a obstaculizarse la visión por producirse una adherencia palpebral considerablemente firme entre 48 y 56 h. Se observó marcado debilitamiento, terminando con postración en los casos fatales. En la agonía los animales mostraron disnea grave. Aquellos que se recuperaron, lo hicieron en breve tiempo. Entre 4 y 6% de los animales sobrevivieron presentaron un estado de caquexia y tras un curso crónico murieron, no se observaron diferencias en el comportamiento clínico entre los animales machos y hembras. (Figura III/A/1) Ver ANEXOS Capítulo III. La cinética de la mortalidad mostró una marcada agudeza de la enfermedad. Los porcientos más altos de mortalidad se obtuvieron dentro de las primeras 24 h posteriores a la inoculación del germen, un porciento mucho menor de animales murieron al segundo día y este fue aun menor al tercer día. En este sentido debemos tener en cuenta que la sepsis por Nm se produce parcialmente, debido a un exceso de citoquinas que aparecen inmediatamente después de la interacción establecida entre los macrófagos y las endotoxinas bacterianas (Quakyi et al., 1997). (Gráfico III/A/1). La única lesión macroscópica en SNC que pudo detectarse fue la congestión de los vasos meníngeos. En el 25% de los casos trabajados encontramos histopatológicamente una meningitis focal, leve, de tipo neutrofílica al principio y luego con predominio mononuclear. La piamadre, sobre todo, se presentó engrosada focalmente por el infiltrado inflamatorio antes mencionado. Los vasos meníngeos se observaron congestivos y apareció un infiltrado netrofílico escaso al nivel de los plexos coroideos.(Figuras desde la III/A/2 a la III/A/4) en ANEXO Capítulo. III). Las lesiones observadas en los animales inoculados avalan la reproducción de una meningitis (leptomeningitis) por NmB en los ratones. A pesar de que las lesiones fueron focales y relativamente leves evidenciaron los efectos patógenos del germen a este nivel. Esto permitió además, interpretar algunos aspectos de la patogenia de la enfermedad; sobre todo si tenemos en cuenta las afectaciones observadas en los plexos coroideos que pudieran sugerir el acceso de los microorganismos al SNC por esta vía según plantea Williams et al., (1990a, 1990b) quienes utilizaron las vías naturales e intracisternal para infectar cerdos con Streptococus suis tipo 2, mientras que para Nm Hugosson et al., (1996), demostró la acción específica que contra células dianas tienen esta bacteria mediante los gliocoesfingolípidos que la une a los granulocitos, células epiteliales, tonsilas de la nasofaringe, y los plexos coroideos. No menos interesante puede considerarse la presencia de las proteínas de la membrana externas de Nm Opc considerada un factor de virulencia que facilita su adherencia a las células y la posterior invasión del germen, mientras que la Opa le confiere la propiedad de tropismo hacia determinados tejidos (Virji et al., 1992,1993),(Sacchi et al., 1995). Asimismo, el innato perfil de citoquinas antiinflamatorias del huésped puede contribuir al desarrollo fatal de la infección meningocóccica (Westerndrop, 1997). Cuando se aplicaron los FEV se encontraron ratones bacteriémicos incluso 144 h post inoculación en contra posición a lo sucedido cuando no se usan, donde resultó difícil el aislamiento del germen a partir de la sangre de los ratones infectados. En la Tabla III/A/1 se puede ver los resultados del reto en animales vacunados con VAMENGOC- BC® y sin vacunar. Al inocular 100 DL50 aproximadamente, como dosis de confrontación, la vacuna protegió al 60 % de los animales, mientras que murieron todos los controles (P<0.01). Lo anterior pudiera tener explicación en el hecho de que los oligosacáridos de Nm son responsables de la inmunomodulación y la toxicidad. Pero cuando los oligosacáridos están íntimamente asociados a las proteínas de la membrana externa, como sucede en el caso de VA-MENGOC-BC, se producen una reducción de la toxicidad y un aumento de la inmunomodulación (Quakyi et al., 1997). En el comportamiento de los ratones al cabo de 12 h de retados pudo comprobarse que tanto los vacunados como los controles mostraron bacteremia. Transcurridas 24 h, los primeros ya la habían vencido; mientras que aún después de 48 h los no vacunados permanecían bacteriémicos (Tabla III/A/2), coincidiendo de este modo con los resultados de Westerink y Giardina, (1992). En cuanto al tiempo de eliminación de la bacteremia sucedió de igual modo, coincidiendo con lo planteado por Westerink et al., (1994,1995) cuando utilizó los ratones Balb/c como modelo de infección por meningococo utilizando dextrana férrica como FEV,. Así, obtuvo 100% de sobrevivencia y una reducción significativa de la bacteremia en 24 h cuando los animales fueron protegidos con un anticuerpo antiidiotipo 6F9 del polisacárido capsular de NmC. Los títulos de anticuerpos contra NmB en el suero de los animales infectados mostraron diferencias altamente significativas (P<0.01) a favor de los vacunados (Tabla III/A/3). Similares resultados han sido obtenidos por Infante et al., (1998) al usar VA -MENGOCBC® con diferentes dosis y esquemas de vacunación en el biomodelo ratón y por Westerink et al., (1995) cuando inmunizó ratones Balb/c con complejos péptido protosoma de NmC produciendo un significativo aumento de anticuerpos y evidente protección frente a retos letales con el serogrupo C. Los ensayos efectuados en animales previamente vacunados con VA-MENGOC-BC® y sus controles (no vacunados), todos los cuales fueron confrontados con altas dosis (100 LD50 ) de NmB demostraron el alto nivel de protección conferido por esta vacuna. Los controles murieron en su totalidad con la dosis de reto, mientras que los animales vacunados mostraron una sobrevivencia significativamente mayor, pudiendo estar relacionado esto con la presencia de anticuerpos contra los lipopolisacáridos y las proteínas de la membrana externa de Nm presentes en esta vacuna. Esto coincide con el criterio de que estas sustancias evocan la producción de anticuerpos protectores contra la bacteremia producida por gérmenes Gram (-) en ratas leucopénicas según reportaron Bhattachajee et al., (1996). Los vacunados eliminaron más rápidamente la bacteremia que los controles y a la vez presentaban títulos de anticuerpos contra NmB en el momento del reto muy superiores a los controles (Tabla III/A/2). Los títulos de anticuerpos demostrados en los animales no inmunizados (valor medio de, 872) están por debajo del valor mínimo establecido (1050 U.) para ser considerados positivos por las Especificaciones de la vacuna antimeningocócica VA-MENGOC-BC® (F01-001 versión 1, 1998). No obstante, pudieran reflejar algún tipo de reacción cruzada demostrada por Artenstein et al., (1975) entre NmB y E. coli K1 cuya presencia no es descartable en ratones convencionales. Similares resultados fueron obtenidos al ser empleados ratones Balb/c vacunados con una vacuna conjugada que contenía polisacárido de NmC y toxoide tetánico que actúa como transportador según Rubinstein et al., (1998). B- Las ratas recién nacidas como biomodelo para reproducir la enfermedad causada por NmB. En los ensayos conducidos en las ratas recién nacidas los síntomas no fueron evidentes. No obstante después de la infección pudimos apreciar menor movilidad, dificultad para la lactación y en períodos más avanzados del proceso, la respiración se hace lenta, lo que precede a la muerte. La bacteremia de las ratas inoculadas con una carga bacteriana de 106 UFC/animal (Tabla III/B/1) se prolongó por 144 horas post-inoculación, resultando todos los animales bacteriémicos. Con cargas bacterianas superiores, como 5x107UFC/mL, no fue posible estudiar la bacteremia más allá de la 72 h por producirse considerables niveles de mortalidad. Es decir, la bacteremia se prolonga hasta la muerte cuando se usan dosis del germen altamente letales, y al menos, hasta la 144 h post-infección con dosis menores. Esto guarda relación directa con las posibilidades de la respuesta inmune de los animales, la cual trata de mantener un desplazamiento del equilibrio germen-hospedero a favor del último. Sin embargo cuando las concentraciones del inóculo son tan altas que vencen las posibilidades del sistema inmune para combatir y controlar los daños que resultan de la acción del microorganismo, causan la muerte del individuo. Se han reportado resultados similares (Moxon 1974) en el que solo se describe bacteremia hasta la 72 h con un inóculo de 106UFC/mL, mientras en nuestro caso esta se prolongó hasta las 144 h post-inoculación. Lo anterior pudiera tener relación con las características de la cepa utilizada en el ensayo, si tenemos en cuenta que tanto el ácido siálico como los lipooligosacáridos de NmB son responsables de la expresión de la virulencia del meningococo en ratas recién nacidas (Vogel et al., 1996). En líquido cefalorraquídeo (LCR) se observaron, a las 72 horas post-inoculación múltiples formaciones redondeadas Gram (-), identificadas como NmB mediante los estudios bacteriológicos (Frotis y cultivo). La DL50 se estableció teniendo en consideración el comportamiento de la mortalidad en relación con las dosis utilizadas para la infección con NmB. Utilizando una carga de 1x106 UFC/animal se alcanzó un 32 %, mientras que con 1x107 UFC/mL el nivel de mortalidad fue de un 50 % y con 1x108UFC/mL la mortalidad fue total (100%) al cuarto día postinoculación. Se estimó por ello una DL50 de 5.1 X 106UFC/mL con un inóculo de 0.1 mL establecida sobre la base del comportamiento de la mortalidad en las ratas infectadas. El estudio citológico del líquido cefalorraquídeo se realizó en las ratas inoculadas con 1x106 UFC y trajo como resultado que en el 30 % (6 de 20) de los frotis analizados se encontraron signos evidentes de alteraciones a este nivel, como fueron abundantes células blancas con predominio de neutrófilos. . A las 72 horas de la inoculación los frotis mostraron que las células inflamatorias predominantes en LCR eran polimorfonucleares neutrófilos y macrófagos, en los cuales según McNeil et al (1994) la Nm mantiene su virulencia durante un período prolongado de tiempo, aunque además se apreciaron abundantes linfocitos. Se observaron formaciones identificadas como N. meningitidis a través de la coloración de Gram, que luego fueron aisladas. El estudio anatomopatológico macroscópico en ratas inoculadas con NmB reveló que el 100 % de los casos investigados mostraron congestión de los vasos meníngeos, mientras que se apreciaron en 10 (27.5%) hemorragias focales. No se observaron lesiones de valor diagnóstico en otros órganos, ni tejidos de la economía animal. En la histopatología de las ratas inoculadas, de un total de 40 cráneos provenientes de ratas infectadas con 1x106UFC/animal de NmB, mostraron lesiones indicativas de inflamación meníngea 25 de ellos para un 62.5 % de positividad. Sin embargo de un total de 36 cráneos pertenecientes a ratas inoculadas con NmB con una carga de 1x107 y 5x107UFC/mL, microscópicamente resultaron con lesiones evidentes de meningitis 27 de ellos, para un 75 %. El grado de severidad de dichas alteraciones varió, en las tres concentraciones, según el tiempo transcurrido desde el momento de provocada la infección y el sacrificio o la muerte, como puede apreciarse, la presencia de las lesiones en las meninges es alto, en contraste con la mortalidad ocurrida en los inóculos con las menores concentraciones de NmB. Estos muestreos siempre se realizaron en combinación con estudios bacteriémicos previos en las ratas inoculadas y controles. Los animales que mostraron alteraciones meníngeas resultaron anteriormente bacterémicos. Antes de 48 h post-inoculación no se observaron alteraciones que indicaran la ocurrencia de meningitis. A las 48 h postinoculación las meninges solo mostraron congestión, mientras que a la 72 h, ya se observaron discretas lesiones que incluyeron el engrosamiento de la piamadre con infiltrado de células predominantemente polimorfonucleares neutrófilos. Ya a los 6-8 días se observaron lesiones severas en meninges mostrando congestión y hemorragias de los vasos meníngeos en la corteza y los plexos coroideos, engrosamiento de la aracnoides fundamentalmente basado en células mononucleares, coexistiendo con abundantes neutrófilos, gliosis focal astrocítica y microglial, sobre todo hacia la zona frontal, cerca de las meninges. También se encontró infiltrado inflamatorio neutrofílica alrededor de la Vaina de Schawan del nervio olfatorio. Además, se observó la presencia de macrófagos fagocitando unas partículas grandes redondeado Gram (-). A los 9 días postinoculación se apreció que las lesiones anteriores persistieron, solo que las células inflamatorias predominantes fueron mononucleares (linfocitos, macrófagos y células plasmáticas), con tendencia a desaparecer los polimorfonucleares neutrófilos, mientras que los procesos gliales se tornaron difusos y generalmente por debajo de las leptomeninges. Basado en los análisis realizados se logró conformar una escala según el grado de severidad de la meningitis (Tabla III/B/2). El alto porciento de positividad respecto a las lesiones histopatológicas de meningitis que ascendió a un 62.5%, la presencia de células inflamatorias y de gérmenes en el LCR (30 % de los casos) es similar a los reportados por Granoff y Nankervis, (1977), para gérmenes productores de meningitis tales como H. influenzae, constituyendo una franca evidencia de la utilidad de este biomodelo. Esto a su vez guardó relación estrecha con los estados bacteriémicos presentados por las ratas cuando se utilizaron inóculos de 1x106UFC/mL de NmB, coincidiendo así con lo planteado por Saukkonen y Leinonen, (1986), González y Leinonen, (1991), aunque los porcientos alcanzados por nosotros fueron superiores a los logrados por (González y Leinonen, 1991). El cuadro lesional observado en nuestros ensayos resultó similar al descrito por estos autores para el biomodelo, solo que ellos reportan la aparición de las células plasmáticas después del décimo día post inoculación, así como también con los casos letales descritos en humanos reportados por Tunkel et al, (1990) y por Infante et al., (1997) en ratas SPF infectadas con NmB y NmC. (Figuras desde la III/B/1 hasta la III/B/5 Ver ANEXOS Capítulo III). El mecanismo por el cual las bacterias circulantes ingresan al espacio subdural todavía no se ha determinado. Existen varias teorías que tratan de explicar el proceso patogénico mediante el cual estas bacterias patógenas penetran en el espacio subdural. Entre estas teorías está el ingreso a través de los sinusoides venosos durales, los vasos sanguíneos leptomeníngeos (Moxon et al., 1974), (Salit y Tomalty, 1986), (Nakajima et al., 1991), (Seimiya y Ohshima, 1992), (Green y Smith, 1992), los plexos coroideos (Daum et al., 1978), (Quagliarello y Scheld, 1992), necrosis de la vellosidades aracnoideas y sitios de trauma meníngeo directo. Somos partidarios de que los plexos coroideos pudieran ser la puertas de entrada de NmB al conducto cerebroespinal, teniendo en cuenta las lesiones iniciales observadas y caracterizadas por la presencia de células inflamatorias tales como neutrofilos y monocitos marginados en las paredes del plexo, así como la presencia de las células de Kolmer a este nivel. Esto indica que este es el sitio por donde circulan y pudieran penetrar dichos gérmenes, coincidiendo con las observaciones realizadas por Sifontes, (1996) cuando estudió la meningitis experimental producida por H. influenzae. Algunos estudios en ratón han confirmado que las Células Kolmer (macrófagos residentes en LCR) son de origen monocítico. Estas además demostraron que partículas inoculadas de forma intravenosa pueden ser fagocitadas por células mononucleares y así ser transportadas hasta el conducto del LCR. Estas células se observan con más frecuencia entre el parénquima y la superficie de los plexos coroideos confirmando que el principal sitio de ingreso de los monocitos en el canal del LCR son dichas estructuras (Carpenter et al., 1970). Dicha migración probablemente se mantiene durante toda la vida para sustituir o mantener los macrófagos residentes en todos los tejidos (Perry y Gordon, 1988), además se ha demostrado que la infección a través de la cisterna cerebral en ratas recién nacidas con 1020ng de endotoxinas del meningococo produce marcada pleocitosis (Kartalija et al., 1995). Por todo lo anterior pensamos que estas bacterias llegan a dicho canal asociadas con monocitos migrantes hacia él para mantener las poblaciones de macrófagos residentes, es decir, los microorganismos circulantes entran en una forma de asociación celular por un mecanismo análogo al de algunas virosis (Rubin et al., 1985). Lo anterior es corroborado por la presencia en las leptomeninges de grandes macrófagos repletos de contenido finamente granular, Gram negativo, principalmente al nivel de las aracnoides y libres en el líquido cefalorraquídeo (Peluso et al., 1985), lo cual pudiera corresponderse con las bacterias inoculadas y posteriormente fagocitadas. Además, se ha demostrado que algunas bacterias que frecuentemente producen meningitis. (Haemophilus influenzae b, Streptococcus tipo b y Listeria Monocitogenes) pueden sobrevivir en el interior de los fagocitos (Geoffroy et al., 1987). Otros sitios que pudieran sugerirse como probables vías de ingreso al canal cerebroespinal, tales como las vellosidades aracnoideas y la región rostral de los bulbos olfatorios, pensamos que más bien sean lugares de acumulación de células inflamatorias coincidiendo con (William y Blakemore, 1990a ). Mientras tanto los referidos acúmulos en las vainas de los nervios olfatorios creemos ocurran por la evacuación del líquido cefalorraquídeo a través de estas, proceso que se incrementa cuando aumenta la presión intracraneal, como sucede en las meningitis (William y Blakemore, 1990 b). CONCLUSIONES DEL CAPÍTULO 1- Se logró reproducir la infección por NmB en la línea isogénica de ratones Balb/c, inoculando como FEV la dextrana férrica. 2- Se estableció la caracterización clínico y patológico de la meningitis por NmB en ratón y ratas recién nacidas como un proceso de presentación aguda, mientras que las lesiones anatomopatológicas a nivel del SNC, sugieren el compromiso del referido sistema. . 3- Se demostró que VA-MENGOC-BC® es altamente eficaz para proteger los animales retados con NmB. CAPITULO IV EFICACIA DE VA-MENGOC-BC Y DE LA INMUNOGLOBULINA ANTIMENINGOCOCCICA BC EN LOS BIOMODELOS RATA Y RATON. Introducción Luego de establecer los biomodelos rata y ratón para la enfermedad producida por NmB en estas especies se imponía la necesidad de comprobar su eficacia para demostrar el potencial protector de VAMENGOC-BC® frente a cepas homólogas y heterólogas, así como conocer el alcance de la protección conferida por la Inmunoglobulina Antimeningocócica BC en los referidos biomodelos frente a estas cepas. Los estudios epidemiológicos revelan que el mayor número de infecciones en niños por debajo de 4 años de edad se debe a H. influenzae, N. meningitidis y Streptoccus pneumoniae (Goldblatt, 1988). Diversos modelos experimentales han sido usados para demostrar la eficacia de productos inmunoterapéuticos o los mecanismos de protección de los organismos vivos frente a Nm. Tal es el caso de los ratones Knockout para conocer los efectos del IFN gamma sobre esta infección (Buchanan et al., 1998). Por otra parte el modelo ratón fue utilizado en la comprobación de la eficacia de vacunas contra Nm elaboradas a base de la membrana externa de este germen mostrando una fuerte inmunogenicidad cuando fueron administrados por la vía intranasal (Hanaberg et al., 1998). En tanto Stojiljkovic et al., (1995) han empleado las ratas recién nacidas para confirmar la importancia de los lipooligosacáridos en la virulencia de Nm. También se han realizado estudios en ratones Balb/c para demostrar la biodistribución de VA-MENGOC-BC® observando la cinética de la radioactividad en órganos como bazo, timo y músculos (Pérez et al., 1997). Objetivos: - Establecer la protección conferida por la Inmunoglobulina Antimeningocóccica BC en ratas y ratones frente a la infección con NmB. (protección pasiva con la Inmunoglobulina Antimeningocócica humana) - Verificar la protección conferida por VA-MENGOC-BC® y la Inmunoglobulina Antimeningocócica BC en ratones frente a la infección con cepas de NmB aisladas en América Latina. (Conocer la amplitud del efecto protector frente diferentes serotipos y subtipos de NmB. - Comprobar la eficacia de VA-MENGOC-BC® y de la Inmunoglobulina Antimeningocócica BC en ratones frente a retos con Nm A, B y C y verificar su efecto según la vía de administración. MATERIALES Y MÉTODOS Ver Diagrama Capítulo IV. (Reverso página anterior.) 1- Todo lo relacionado con animales, tenencia y manejo de estos, ética, método de inoculación, factores estimulantes de la virulencia, puede encontrarse en él: Capítulo II, Materiales y Métodos Generales, Acápite II (9,10,11,12,13). 2- Gérmenes y preparación de inóculos se encuentran en el Capítulo II, Materiales y Métodos Generales, Acápite II (9) Para los ensayos de eficacia de VA-MENGOC-BC® y de la Inmunoglobulina Antimeningocócica BC en ratones frente a diferentes cepas de NmB aisladas en América Latina, ensayos (B1 y B2 ), las cepas de NmB utilizadas en los retos fueron aisladas de casos clínicos y pertenecientes a los serotipos siguientes: Argentina B no tipable: P1.10; Chile no tipable, Colombia, B4:P1:15; Cuba B4:P1.15; Brasil B4: P1.15. Para esta última se cálculo la DL50, pero no se llevó a cabo el reto en animales vacunados con VA-MENGOC-BC®, pero sí frente a la Inmunoglobulina Antimeningocóccica BC. 3- Inoculación de gérmenes y FEV. Determinación de la DL50-. Composición de la Inmunoglobulina Antimeningocócica BC y de VA-MENGOC-BC® y para otros datos ver el Capítulo II, Materiales y Métodos Generales, Acápite II (6,7,9,14). Ensayo A Comprobación de la eficacia de la inmunoglobulina antimeningocócica BC en ratas recién nacidas y ratones Balb/c. En este estudio se emplearon 84 ratas recién nacidas. Con ellas se llevaron acabo tres experimentos, 32 animales en el ensayo de mortalidad, 32 en la bacteremia con la aplicación dela Inmunoglobulina Antimtningocócica y 20 animales para determinar la mortalidad con la Inmunoglobulina Antimeningocóccica BC, con el objetivo de medir su eficacia. Esta fue aplicada 2 h antes de la confrontación (mortalidad y bacteremia), empleando una carga bacteriana de NmB de 1 x 106UFC/mL, 2 h después de la 7 infección, con una carga de 1 x 10 UFC/mL de NmB por vía intraperitoneal. En el caso de los ratones se utilizaron un total de 100 animales, 40 para determinar la dosis de selección expresada en mg y en diferentes tiempos posteriores al reto para aplicarla en los siguientes experimentos y 60 en la comprobación de la eficacia de la dosis única y repetida. Se emplearon 10mg y 5mg, respectivamente de la Inmunoglobulina antimeningocócica BC durante 30 y 15 minutos como tratamiento preinfección, en los estudios de dosis única, utilizando una dosis de confrontación de 1 x 108UFC/mL. Para las aplicaciones de las dosis repetidas de 5 mg los intervalos fueron: 30, 45 minutos, 30, 45,min. 1h, y 30, 45, 1h y 17, mientras que para 5 aplicaciones e igual dosis los tiempos fueron 30, 45 minutos, 1, 2 y 17 horas, post - confrontación. Eficacia de VA-MENGOC-BC® y de la Inmunoglobulina Antimeningocóccica BC en ratones frente a diferentes cepas de N. meningitidis serogrupo B aisladas en América Latina. Ensayo B1 Eficacia de VA-MENGOC-BC® en ratones. Para conocer la eficacia de la vacuna VA-MENGOC-BCen ratones, luego de la aplicación del esquema (Ver Tabla IV/M/2), se llevaron a cabo retos con las cepas latinoamericanas (Ver Tabla IV/M/1) con las siguientes cantidades de ratones: 26 para la cepa argentina, 30 para la chilena, 30 en la colombiana y 31 en la cubana. Los animales fueron separados en 3 grupos, uno de ellos recibió una dosis de VAMENGOC-BC®, otro 2 dosis (45 días entre una y otra) y el control no fue vacunado. Ver Tabla IV/M/2 En la determinación de la bacteremia el grupo 5 solo recibió Inmunoglobulina antimeningocóccica BC después de la inoculación con la cepa D (ver tabla IV/M/1). La Inmunoglobulina Antimeningocócica BC fue aplicada en 0.2 mL de volumen por vía intraperitoneal 30 minutos antes del reto ó 30 minutos después de este. Para otros datos ver Capítulo II, Materiales y Métodos Generales, Acápite II (1,2,3,4,5,6,8,9,10,14,16.17). Análisis estadístico Los ensayos para determinar DL50 se realizaron paralelamente a los retos que se utilizaron para medir la protección con Inmunoglobulina antimeningocóccica BC. Las diluciones se prepararon en el rango comprendido entre 1x10 6 y 1x10 10UFC /mL. Para otros datos: Ver capítulo II, Materiales y Métodos Generales, Acápite II (14). Retos Los animales recibieron el reto con las cepas ya mencionadas 21 días después de la segunda dosis y 66 días después de la primera, con un inoculo que en términos de UFC/mL oscilaron entre 1x106-8 UFC/mL (1x106/mL en el modelo rata recién nacidas y 1x108 /mL para el modelo ratón en un volumen de 0.2 ml aplicado por vía intraperitoneal. Las muertes fueron registradas en el intervalo de 4-6 h durante 3 días. Los retos así como los ensayos para determinar el valor de la DL50 de cada inoculo empleado siempre fueron realizados simultáneamente con el objetivo de obtener el valor más adecuado en términos de la LD50 y conocer la virulencia y utilidad comparativa de las diferentes cepas usadas. Cálculo de la protección: Ver Capítulo II, Materiales y Métodos Generales, Acápite II (13). B2- Eficacia de la Inmunoglobulina Antimeningocócica BC en ratones retados con cepas de N. meningitidis serogrupo B aisladas en América Latina. En este se utilizaron las mismas cepas que en el anterior y se aplicó la Inmunoglobulina Antimeningocócica BC por la vía intraperitoneal 30 min. antes o después del reto. Ver Tabla IV/M/1. C- Evaluación de la eficacia de VA-MENGOC-BCen ratón Balb/cj retados frente a los serogrupos A, B y C. Vacunación y reto Se utilizaron un total de 180 ratones distribuidos en grupos de 90 animales. Los grupos compuestos de 60 ratones recibieron 1, 2 ó 3 dosis de VA-MENGOC-BC y en cada subgrupo 20 ratones, los cuales fueron posteriormente retados con las cepas de los serogrupos A, B y C antes mencionada en cantidad de 10 animales a los 15 y 21 días después de la última vacunación. En cada serogrupo se realizó el estudio de mortalidad para conocer la DL50 aproximada de cada cepa empleando 6 concentraciones en los cuales se usaron 10 animales por dosis Los ratones fueron inmunizados con 1, 2 ó 3 dosis de VA-MENGOC-BC®, 0.5 mL, IP. El intervalo entre dosis fue de 21 (1ra-2da) y 15 días (2da-3ra). El reto se realizó después de los 15 y 21 días de aplicada la última dosis correspondiente. Se mantuvieron animales controles no inmunizados, de igual peso y edad. Protección con la Inmunoglobulina Antimeningocócica. Para los estudios de protección con la inmunoglobulina se utilizaron 90 ratones, 30 para cada serogrupo incluyendo 10 en cada vía (I/P y I/V) y 10 animales controles por cada serogrupo. La Inmunoglobulina Antimeningocócica BC fue aplicada 30min, 2 y 6h después de la inoculación de 1x107UFC/mL de Nm A, B y C. Se administraron en cada caso 5 mg de la inmunoglobulina por vía intraperitoneal o intravenosa en un volumen de 0.1 mL, la primera elegida por permitir la administración de grandes volúmenes y la segunda por constituir una de las vías de elección para la aplicación del producto en humanos. Cepas utilizadas en los retos Se realizaron retos con NmA (Nigeria A4 P1 9), NmB (Colombia B4 P1 15) y NmC (ATCC). Se suspendieron las bacterias en PBS y se inocularon 0.2 mL, IP, de las diluciones equivalentes, a dosis superiores o iguales a la DL50. RESULTADOS Y DISCUSIÓN A- Comprobación de la eficacia de la Inmunoglobulina Antimeningocócica BC en ratas recién nacidas y ratones Balb/c. - Ratas recién nacidas: En los estudios de mortalidad y bacteriemia, al aplicar la Inmunoglobulina Antimeningocócica BC 2 h antes de inocular una carga bacteriana de 1x106 UFC/mL, no se produjeron muertes, mientras en el grupo no inmunizado, murió una proporción (37.5%) significativamente mayor (p0.004) (Tabla IV/A/1). Al realizar el estudio de la bacteremia se apreció que en los animales inmunizados pasivamente 2 horas antes del reto no se llegó a observar bacteremia. En tanto que en los controles no inmunizados, ésta se mantuvo durante los tres días de chequeo, (Tabla IV/A/2). Resultados similares fueron obtenidos por Sacchi et al., (1995) cuando utilizó ratas recién nacidas tratadas con sueros procedentes de individuos vacunados con proteína clase 5 de NmB. El reto y tratamiento posterior con la Inmunoglobulina Antimeningocócica BC (Tabla IV/A/3) corroboró su valor terapéutico, protegiendo todos los animales tratados, lo que difiere significativamente de los no tratados, cuando se aplicó 1x107UFC/mL en los que la mortalidad alcanzó el 50%. Esto puede atribuirse a los estados de shock que sobrevienen tras la bacteremia causada por microorganismos Gram negativos, los cuales pueden estar mediados en parte por el complemento, activado extensamente por las endotoxinas (Gerard y Gerard, 1991). Estos resultados evidenciaron la efectividad del biomodelo en los ensayos de inmunización pasiva pre y post infección con NmB aplicando la Inmunoglobulina, donde probablemente ciertos anticuerpos dirigidos contra la superficie bacteriana puedan bloquear los requerimientos funcionales del microorganismo, como pueden ser las uniones con los compuestos quelantes del hierro o la captación de otros nutrientes. Los resultados demuestran al mismo tiempo el alto poder protector de este medio terapéutico. Similares resultados han sido obtenidos al usar anticuerpos monoclonales contra proteínas de la membrana externa del germen en cuestión (Saukkonen, et al., 1988), (González y Leinonen, 1991), así como a (Nassif et al., 1992) cuando empleo el TNF durante la bacteremia letal en ratas recién nacidas retadas con Nm. Todo esto demuestra indirectamente una vez más la elevada eficacia de la vacuna antimeningocócica VA-MENGOC-BC, ya que la Inmunoglobulina Antimeningocócica BC es obtenida a partir de humanos hiperinmunizados con esta vacuna. - Ratones: Con respecto a la dosis de confrontación pudo comprobarse que 1x108UFC/mL resulta excesivamente alta, causando 100% de mortalidad en el grupo retado. Por el contrario, con 3x106UFC/mL, aun cuando se logra 100% de protección en los retados, esta no es significativa ya que muere solo el 10% de los controles no inmunizados. Empleando una dosis logarítmicamente intermedia (5x107UFC/mL), se logra una protección del 40% que si resulta estadísticamente significativa (p0.05) con 10 animales por grupo experimental y la mortalidad de 100% en los controles. Respecto a la efectividad de la Inmunoglobulina Antimeningocócica BC en dos momentos y dos dosis diferentes aplicándolas antes del reto, cuando se administran dosis de 10 mg y 5 mg de Inmunoglobulina Antimeningocócica, 30 y 15 min respectivamente antes de la dosis de confrontación (1x108 UFC/mL) se observó (Tabla IV/A/4) que al aplicar 10 mg de Inmunoglobulina Antimeningocócica BC (0,2 mL) 30 min. antes de la dosis de reto, la sobrervivencia fue del 70%. Sin embargo, al reducir la dosis a la mitad (5 mg) esta aumentó al 80%. Con esta misma dosis de Inmunoglobulina Antimeningocóccica BC, reduciendo a la mitad el tiempo antes de la inoculación, aumentó la sobrevivencia hasta el 100%. Esto resultados aunque las diferencias estadísticas a favor 5mg y 15 son pequeñas indica que la Inmunoglobulina Antimeningocóccica BC ensayada tiene altos efectos protectores, contra la cepa de NmB inoculada en una carga bacteriana de 1x108UFC/mL, la cual fue capaz de desencadenar la infección con una mortalidad del 100% en los controles no protegidos. En este sentido no debemos olvidar la posibilidad que presentan microorganismos como N. meningitidis, E. coli K1, y Streptococus del grupo B que resisten la acción del complemento al ser este inactivado por la unión con los factores H e I (Lynn y Wolenbooch, 1992). Debemos destacar que cuando se actúa más próximo a la invasión bacteriana, se logra mejor efecto y se puede reducir la dosis de Inmunoglobulina Antimeningocócica BC a la mitad; de aquí que los resultados permitieran tomar la dosis de 5 mg para ensayo de dosis única y repetidas. Sin embargo nosotros decidimos utilizar esta dosis, pero aplicarla en vez de a los 15min. a los 30, teniendo en cuenta el criterio de que a los 15 min. es muy poco tiempo para que se implanten los gérmenes y produzcan el efecto sobre el biomodelo, resultando poco similar a lo que podría suceder en el humano y además por razones de tipo práctica. En relación con la efectividad por dosis repetidas de Inmunoglobulina Antimeningocócica post-infección, (Tabla IV/A/5) se puede observar que al analizar la estadísticamente mediante la prueba t de Student paramétrica, la sobrevivencia de los ratones fallecidos no manifestó diferencias significativas entre los grupos que recibieron una y dos dosis. Sin embargo, hubo un incremento en el tiempo medio de sobrevivencia de 11,7 horas (30 min y 1 hora) y 9,6 (30 y 45 min) respecto a una sola dosis (30 min). Al analizar la supervivencia se aprecia que la administración más temprana (30 y 45 min) provocó que ésta aumentara hasta el 70%, de donde se infiere que dos dosis surten mejor efecto que una o dos aplicaciones más tardíamente. Al aplicar cuatro dosis comenzando tempranamente después de la invasión (30, 45 min, 1 y 17 horas), la sobrevivencia alcanzó el 90% de los casos, con el 100% de protección al aplicar cinco dosis en el esquema de 30, 45 min, 1, 2 y 17 horas. Esto hace pensar en la posibilidad de que los anticuerpos suministrados mediante terapia pasiva paulatina y constante se combinan con los antígenos (gérmenes), que en este caso abundan en el interior del peritoneo, por haber sido esta la vía de aplicación del inóculo, así como en el interior del aparato circulatorio y otros sitios de la economía, facilitado este proceso por el efecto que producen los FEV al aumentar la permeabilidad del lecho capilar existente en el peritoneo (Sifontes et al., 1996). Así se evita la acción deletérea del germen y sus toxinas, sobre los distintos órganos y tejidos del animal, por lo que resultan más eficaces que en dosis única o menos fraccionadas, además queda demostrado su efecto dosis de pendiente. Por otra parte se considera que la forma de aplicación de la Inmunoglobulina Antimeningocócica BC posterior al proceso de confrontación con el germen, resulta la forma más adecuada. Esta mimetiza el proceso que sucede en el humano afectado de meningitis meningocócica, ya que el tratamiento se aplica generalmente a las personas a las cuales la enfermedad les ha sido diagnosticada. Esto no niega en modo alguno las posibilidades de utilización del producto de forma preventiva en personas con contacto cercano a enfermos, o en peligro de contraer la enfermedad ante la ocurrencia de brotes y en personas consideradas de alto riesgo. Con relación al tiempo de sobrevivencia (entendido este como la cantidad de horas vividas por los animales tratados que murieron antes de finalizar el ensayo), podemos plantear que este aumentó en la misma medida que la cantidad de aplicaciones. Este experimento pone de manifiesto la alta efectividad de la Inmunoglobulina Antimeningocócica BC humana en el ratón, coincidiendo con los ensayos realizados por Danve et al., (1993) cuando inmunizó pasivamente ratones con sueros obtenidos de animales vacunados con proteínas unidas a transferrina en la membrana externa de Nm, los cuales resistieron posteriormente un reto con 100 DL50 de NmB. Todo lo anterior pone en evidencia la factibilidad del modelo animal para valorar la Inmunoglobulina Antimeningocócica BC de origen humano y darnos una idea de la protección que puede conferir al hombre, lo que ha sido confirmado posteriormente por los trabajos de Galguera et al., (1991). B- Eficacia de VA-MENGOC-BC® y de la Inmunoglobulina Antimeningocóccica BC en ratones frente a diferentes cepas de N. meningitidis serogrupo B aisladas en América Latina. B1- Eficacia de VA-MENGOC-BCen ratones. Los ratones fueron susceptibles a los 5 aislamientos de NmB que se evaluaron, cuando se aplicó un inóculo equivalente a 1x108 UFC/mL en un volumen de 0.2ml. No obstante la virulencia fue variable en un rango de DL50 (Figura IV/B 1/1). Cuando los ratones recibieron un reto equivalente a 37.3 DL50 de la cepa Argentina (B nt P 1-10), la aplicación de una dosis de la vacuna fue capaz de proporcionar un 33.3% de protección, mientras que con el esquema de inmunización de dos dosis esta se elevó al 70% (Tabla IV/B1/1). Con un reto equivalente a 32 DL50 de la cepa chilena (B: no tipable) una dosis de VAMENGOC-BC® protegió al 10% de los animales y 70 % para dos dosis (Tabla IV/B1/2). Los inóculos más fuertes evaluados fueron los de la cepa colombiana B4P1.15 equivalente a 505 DL50 . En este caso una dosis de VA-MENGOC-BC® no fue capaz de proteger frente al reto en los animales vacunados con una dosis. Por otra parte en los que recibieron dos dosis la protección fue de 20 % (Tabla IV/B1/3). No obstante el tiempo de supervivencia de los animales vacunados fue significativamente mayor que el de los controles correspondientes (Tabla IV/B1/4). Los ratones que recibieron una dosis de la vacuna y fueron retados con una dosis equivalente a 7.8 DL50 de la cepa cubana (B4: P1. 15) tuvieron una protección (35.7%) y con dos dosis (85.9) aunque, la mortalidad de los controles no fue del 100% pero sí significativa con respecto a la de los vacunados (Tabla IV/B1/5). Los ratones tratados con hierro pueden infectarse con NmB y desarrollar una infección aguda y fatal, aunque son requeridas altas dosis de microorganismos. El curso clínico en humanos también es agudo y si no se aplica tratamiento, la proporción de muertes puede alcanzar el 100% (Saukkonen y Leinonen, 1986). En el hombre es posible causar la enfermedad bacterémica con una dosis comprendida entre 1 y 224 UFC (Brandtzaeg, 1994). En sentido general se obtuvo una significativa protección frente a las cepas evaluadas las cuales tuvieron gran variación en su virulencia demostrada mediante el cálculo de la LD50 realizado en paralelo a los estudios de reto (Tabla IV/B/6). Del mismo modo, podemos considerar que con la cepa colombiana se obtuvo un buen nivel de protección, ya que aunque en esta no hubo diferencias significativas con relación con la mortalidad de los protegidos con respecto a los controles, sí se observó diferencia en términos de tiempo de sobrevivencia post-infección comparados con estos últimos. Debemos además tener en cuenta que la dosis de reto era extremadamente alta con un 60% de mortalidad en los ratones inmunizados antes de las 16 h post-inoculación, aspecto que recuerda lo que sucede en los humanos afectados por la forma fulminante la enfermedad meningocócica conocida como meningococémica. Similares resultados fueron obtenidos por (Jamison y Prescott, 1987), cuando estudió preparados vacunales hechos a partir de vesículas de la membrana externa de Nm inoculadas en el modelo de ratón. Del mismo modo sucedió cuando (Quakyi et al., 1997) vacunó ratones lipooligosacaridos o inmunodeprimidos con ciclofosfamida e inoculados con vesículas de la membrana externa de Nm. Por otra parte nuestros resultados son similares a los obtenidos por Claassen et al., (1996) cuando protegió ratones con una vacuna antimeningocóccica elaborada a partir de vesículas de la membrana externa por ingeniería genética, observando protección frente a 5 cepas de Nm aisladas de casos clínicos. Las ventajas de aplicar un esquema de dos dosis en lugar de una quedaron demostradas frente a las cepas evaluadas, lo cual coincide con lo sucedido en el mismo esquema practicado en humanos y en animales cuando (Haneberg et al., 1998) utilizó una vacuna obtenida a partir de vesículas de la membrana externa de NmB aplicada por la vía intranasal. La DL50 es el índice de virulencia usado en estos experimentos, sin embargo, a veces este no permite realizar una correcta comparación entre diferentes cepas. Una dosis de 2-3 DL50 de una cepa puede causar 100% de mortalidad y esta podría ser superior a la DL100; por el contrario la 2-3 LD50 de otra cepa podría no ser letal para todos los animales en el experimento. Por tanto dos cepas pudieran tener grados diferentes de virulencia, incluso cuando sus DL50 sean muy similares si sus curvas de dosis mortalidad no son paralelas. En este caso la cepa más virulenta es aquella cuya curva sea más perpendicular. Por esta razón los resultados obtenidos con cada cepa no son del todo comparables entre sí. Los ratones mostraron un alto grado de inmunidad si tenemos en cuenta la protección obtenida frente a la gran cantidad de gérmenes inoculados en el reto y la agudeza del curso clínico. La destrucción de la bacteria es causada por los mecanismos de defensa de los ratones, las cuales liberan las endotoxinas que pueden empeorar la infección, ocultando de esta forma la protección conferida por la vacuna y la de los otros sistemas defensivos del organismos como son los mecanismos destructivos oxígenodependientes (Anión superóxido, peróxido de hidrógeno, ácido nítrico) y las proteínas catiónicas (defensinas), catepsina y la azurocidina (Chan et al., 1992). Sin embargo en la infección de curso más largo es facilitada la acción del sistema inmune frente a la infección. La vacuna contra la leptospirosis es un ejemplo donde altos niveles de protección pueden observarse incluso frente a retos con inóculos en el orden de 10 000 DL50. Esto pudiera deberse al largo período de incubación de la infección en los animales no inmunizados de 8-11 días(Oliva et al., 1994). Por otra parte pudo comprobarse que con mayor o menor eficacia en VA-MENGOC-BC fue capaz de conferir protección frente a las diferentes cepas de NmB procedentes de casos de Latinoamérica. B2- Eficacia de la Inmunoglobulina Antimeningocócica BC en ratones retados con cepas de N. meningitidis serogrupo B aisladas en América Latina. En el grupo 1 retado con la cepa argentina B: nt: P1. 10 el subgrupo Control (A-1) tuvo un 62.5% de animales que murieron antes de la 16 h post-inoculación alcanzando un 100% de mortalidad a las 24 h. Todos los ratones que recibieron Inmunoglobulina 30min antes del reto (A-2) sobrevivieron, mientras que en los animales que recibieron el tratamiento 30min después solo hubo un 37.5% (A-3) de sobrevivencia. Estos grupos presentaron diferencias estadísticamente significativas con respecto al control A-1, tanto desde el punto de vista de la mortalidad como del tiempo de sobrevivencia (P<0.05). Tabla IV/B2/1. Como pudo observarse en el grupo 1 cepa Argentina, hubo una buena protección. Todos los animales tratados previamente con la Inmunoglobulina sobrevivieron frente a un 100% de mortalidad en los controles. Cuando se empleó la terapia pasiva posterior al reto, también existieron diferencias significativas con relación al control. En el caso del grupo 2 con la cepa Brasileña B4:P1.15 el subgrupo control B-1 presentó un 80% de mortalidad, y el 60% de las muertes ocurrieron antes de las 24 horas posteriores al reto. Por otra parte, los que recibieron la Inmunoglobulina tuvieron un período de mortalidad que se extendió por espacio de 40 h, en los subgrupos B-2, B-3. La mortalidad registrada alcanzó el 80 y 100 % respectivamente. Cuando comparamos los promedios de tiempo de sobrevivencia de cada grupo observamos que existieron diferencias estadísticas (P<0.1) entre los controles y los animales tratados con la Inmunboglobulina. (Tabla IV/B2/2). Los resultados del grupo 2 subgrupo B-2, B-3 sugieren el efecto protector conferidos por la Inmunoglobulina Antimeningocócica BC frente a la cepa brasileña, puesto que se produce un alargamiento de la vida en los animales tratados, lo cual desde el punto de vista práctico, proporciona un margen de seguridad, al facilitar el tiempo necesario para que actúen otros agentes terapéuticos sobre el microorganismo. Para el grupo 3 con la cepa Colombiana B4:P1.15, la mortalidad de los controles del subgrupo C-1 fue de 100% durante las 16 horas posteriores al reto. En los grupos tratados C-2 y C-3, el 50% de las muertes se produjeron durante un período de 40 h, por lo cual hubo diferencias estadísticamente significativas entre estos y los controles tanto en los parámetros de protección como en el tiempo de sobrevivencia (P< 0.05) (Tabla IV/B2 /3). Por otra parte, la eficacia de la Inmunoglobulina fue demostrada frente a la cepa colombiana (incluso frente a un reto extremadamente cruento): 44.5 DL50. En este caso la protección demostrada ascendió en el subgrupo C-2 al 40% y en el C-3 a un 30% de sobrevivencia. En el caso de la cepa CH Chilena (no tipable) correspondiente al grupo 4 se observó que el subgrupo CH-1 tuvo un 100 % de mortalidad, muriendo el 60 % durante las primeras 16 h, con un promedio de sobrevivencia de 18,1 h; mientras que en el subgrupo CH-2 hubo una mortalidad del 80% con una media de 26,5 horas. En el subgrupo CH-3 todos los ratones murieron con un promedio de tiempo de sobrevivencia de 25,8 h; comprobándose de esta manera que el tiempo de sobrevivencia de los animales tratados fue estadísticamente diferente (P< 0.05). a la de los del grupo control Tabla IV/B2/4. En el grupo 4 a pesar de la virulencia sustancial mostrada por la cepa chilena (no tipable), la Inmunoglobulina mostró sus efectos benéficos al prolongar la vida de los animales. En los humanos este tiempo pudiera facilitar el espacio necesario para que diferentes drogas pudieran actuar sobre el paciente enfermo. El factor de prolongación vital constituye por sí solo un índice de protección. Para el grupo 5 de la cepa D cubana (B4:P1 .15) la Inmunoglobulina Antimeningocócica BC solo fue aplicada después de la infección, cuando se realizaron los retos con 20 DL50 la mortalidad del subgrupo D-2 alcanzó el 40% mientras que el control presentó 100% de mortalidad grupo D-1 con diferencias significativas a favor de los tratados, de igual forma que el tiempo de sobrevivenvia (Tabla IV/B2/5). Basado en el cálculo hecho por Brandtzaeg, (1994), cuando un paciente de meningitis meningocócica recibe atención médica, el nivel de la bacteremia y la endotoxemia son bajos encontrando menos de 1UFC/mL (rango 1-240 UFC/mL), en sangre y en el plasma 3-260 pg/mL de lipopolisacárido. Esto pudiera darnos una idea de la magnitud de la dosis de reto en los ratones 20 DL50 (1x108UFC/mL), lo cual representa más de 10 millones de veces la cantidad de germen necesario para producir la enfermedad en el humano. Lo anterior fue observado en 48 pacientes estudiados que tuvieron la enfermedad meningocócica y un período de admisión hospitalaria significativamente mayor que otros con septicemia fatal que evolucionó en un período de 12-22 h. (Brandtzaeg, 1994). Estos resultados evidenciaron que cuando la Inmunoglobulina Antimeningocócica BC se aplicó 30 min después de la inoculación, el promedio de sobrevivencia frente a las cepas utilizadas varió desde 25.8 a 42 h. Lo anterior demostró que el uso de la Inmunoglobulina Antimeningocócica BC específica podría proporcionar una ventaja en las primeras horas de la terapia. Los autores consideran que una sola dosis de Inmunoglobulina Antimeningocócica BC en ratones es capaz de conferir protección, constituyendo una buena evidencia para el uso de estos como biomodelo en los estudios de eficacia de la Inmunoglobulina Antimeningocócica BC para humanos. Estos modelos brindan la posibilidad de conocer los efectos benéficos de la terapia pasiva a base de la Inmunoglobulina obtenida a partir de humanos inmunizados con VA-MENCOGBC® lo cual tiene importancia en la evaluación de toda una amplia gama de cepas circulantes en el mundo. La Inmunoglobulina Antimeningocócica BC fue capaz de prevenir la infección entre un 20 y un 100% de los casos en el biomodelo ratón, dependiendo esto de la virulencia de las cepas y las dosis de reto que guardaron correspondencia con la mortalidad y el tiempo de sobrevivencia en todos los grupos estudiados, coincidiendo con los resultados obtenidos por Bhattacharjee et al., (1996) cuando trató ratones con suero post-inmune obtenido por inoculación de NmB en ratas. Los efectos protectores de la Inmunoglobulina Antimeningocócica BC han sido estudiados en ratones frente a virosis y se conoce que el uso de esta en etapas iniciales es mejor que después de las 48 h de haber comenzado la enfermedad (Kohl et al., 1981). En ratas recién nacidas la infección intraperitoneal y el tratamiento con dos anticuerpos monoclonales contra NmB demostraron un mejor efecto cuando fueron administrados al inicio de la infección (Mckendall, 1985). Los resultados obtenidos aquí demostraron que la Inmunoglobulina Antimeningocócica BC, incluso cuando las dosis de reto fueron muy altas, tuvo un efecto protector en ratones Balb/c frente a las diferentes cepas latinoamericanas ensayadas. De esta manera, podría considerarse efectiva la referida terapia frente la enfermedad meningocócica causada por NmB. Además quedó demostrado que el biomodelo de ratón Balb/c es adecuado para la evaluación de la Inmunoglobulina Antimeningocóccica BC. Como ha podido apreciarse este producto fue capaz, en sentido general, de conferir protección frente a las diferentes cepas, si bien en algunas no evitó la mortalidad, pero aumentó el tiempo de sobrevivencia, factor de gran importancia que permite realizar intervenciones con otros agentes terapéuticos en la práctica clínica humana. C- Evaluación de la eficacia de VA-MENGOC-BCen ratón Balb/cj retados frente a N. meningitidis de los serogrupos A, B y C. La protección conferida por VA-MENGOC-BC® frente a NmA en ratones a los 15 días post-vacunación fue considerable sobre todo a partir de la segunda dosis, llegando a un 83.33% con la aplicación de tres dosis, en tanto que el tiempo de sobrevivencia fue directamente proporcional al número de dosis a partir de la segunda (Tabla IV/C/1), mientras que a los 21 días post-vacunación la protección ascendió al 100% en todas las dosis con una confrontación equivalente a la DL50 (Tabla IV/C/2). La efectividad de VA-MENGOC-BC® frente a NmB en ratones tratados con mucina dextrana férrica como estimulantes de la virulencia, como puede apreciarse en la DL50 (Tabla IV/C/3), fue estimada en 33.3x105UFC/mL. No obstante cuando se inocularon 3DL50, la protección fue considerable con respecto a los controles no inmunizados, llegando a 87.5% para una dosis y 100% para dos y tres dosis respectivamente (Tabla IV/C/4) en tanto que a los 21 días la protección se mantuvo a altos niveles que estuvieron entre 75 y 100% respectivamente (Tabla IV/C/5). La efectividad de VA-MENGOC-BC® frente a NmC en ratones fue considerablemente menor que las obtenidas con los otros serogrupos a los 15 días post-vacunación. No obstante estas estuvieron entre 33.3 y 55.6 % (Tabla IV/C/6), mientras a los 21 días fueron de 100% para 1 y 2 dosis y de un 60% para tres aplicaciones de la vacuna(Tabla IV/C/7). Con relación a la efectividad de la Inmunoglobulina Antimeningocócica BC frente a Nm A, B y C en ratones, la protección para los tres serogrupos fue significativa sobre todo cuando la terapia pasiva fue aplicada por la vía intravenosa con niveles que estuvieron entre 44.4, 90 y 100%. Sin embargo por la vía intraperitoneal, los niveles de protección se mantuvieron en 11.1,70 y 50% para los tres respectivos serogrupos (Tabla IV/C/8). Si tenemos en cuenta que VA-MENGOC-BC no solo es la vacuna más efectiva de la segunda generación a partir de proteínas de membrana externa (OMP), sino que por lo genuino de su diseño y formulación, pudiera considerarse la primera licenciada de la segunda generación de candidatos vacunales, y esta vez contra diferentes serogrupos y serotipos de Nm. El desarrollo de ensayos inmunológicos que correlacionan la protección verdadera producida por estas vacunas constituye uno de los objetivos estratégicos en el plan de investigaciones priorizadas por el programa para inmunización y vacunas de la Organización Mundial de la Salud. Esta además propone la utilización de las ratas infantes en los estudios de la respuesta inmune frente a sueros obtenidos a partir de vacunados con preparados que contienen proteínas de la membrana externa de NmB, comprobando su actividad protectora y comparándola con la actividad bactericida de dichos sueros (SAGE, 1999). Se ha demostrado por (Pettersson et al., 1990), (Danve et al., 1993) que las proteínas de Nm reguladas por hierro pueden inducir la producción de anticuerpos bactericidas capaces de proteger a los ratones frente a infecciones experimentales. En tanto el uso de anticuerpos dirigidos contra Por A demostró protección pasiva contra el meningococo en infecciones experimentales, (Frasch et al., 1986), (Saukkonen et al., 1987), con Por B y con Opc (Fernández et al., 1992) y LPS (Verheul, 1994). Por otra parte se plantea por (Poolman, 1996) la necesidad de focalizar la atención en los antígenos con demostrada habilidad para inducir anticuerpos bactericidas. Nosotros hemos obtenido resultados satisfactorios en la utilización del biomodelo ratón, tanto en estudios de protección activa con VA-MENGOC-BC® como de protección pasiva con la Inmunoglobulina Antimeningocócica BC a través de retos efectuados con NmB. No obstante, recientes estudios demuestran la existencia de inmunidad cruzada entre proteínas de membrana externa de Nm que incluyen la protección frente a las infecciones experimentales en el ratón ( Martin et al., 1997), (Manning et al., 1998). En los últimos ensayos llevados a cabo en nuestro laboratorio hemos encontrado resultados que apoyan estos planteamientos al lograrse adecuados niveles de protección en ratones previamente inmunizados con VA-MENGOC-BC® y confrontados después con cepas de los serogrupos A y C, así como la protección conferida con la Inmunoglobulina Antimeningocócica BC frente a estas cepas, Westerink y Giardina, (1992), Ala`Aldeen et al., (1996) obtuvieron una fuerte inmunogenicidad basada en la presencia de anticuerpos Men-E2p en el suero de humanos y animales frente a cepas homólogas y heterólogas de Nm. Por otro parte, Gómez et al., (1996) demostraron la presencia de inmunidad cruzada en ratones frente a la proteína de 37 Kda (Fbp) de Nm y por (Ferron et al., 1992) con proteínas unidas a transferrina en Nm. Resultados similares fueron obtenidos por Colino y Outschoorn, (1998) cuando inmunizaron ratones adultos con polisacáridos de NmB, mientras que Chrislodoulides y Heckels, (1994) encontraron respuesta de anticuerpos bactericida frente a cepas homólogas y heterólogos de NmB. Los polisacáridos capsulares pertenecientes a determinados serogrupos conjugados con proteínas resultaron mejores inmunógenos y tuvieron una larga persistencia de anticuerpos específicos contra los aislamientos pertenecientes a estos serogrupos (Costantino et al., 1992), (Anderson et al., 1994), (Zakirov et al., 1995). Lo anterior pudiera estar sucediendo en el caso de VA-MENGOC-BC® y la Inmunoglobulina Antimeningocócica BC, por tanto justificaría en buena medida la respuesta obtenida frente al serogrupo C, toda vez que esta vacuna contiene las proteínas de la membrana externa de NmB y el polisacárido del referido serogrupo. Resultados similares fueron obtenidos por Bhattacharjee et al., (1996) cuando utilizó una vacuna obtenida a partir de lipopolisacárido de E. coli J5 y proteínas de la membrana externa de NmB constatando su protección frente retos letales con diferentes gérmenes Gram negativos en ratas (Young et al., 1991). Todo lo anterior nos permite plantear que nuestra vacuna VA-MENGOC-BC® está en mejores condiciones que cualquier otra para enfrentar el reto de una prueba de campo en un país donde las cepas circulantes de Nm fueran diferentes a las del serogrupo B. CONCLUSIONES DEL CAPITULO - Se demostró la eficacia de la Inmunoglobulina Antimeningocócica BC en ratones retados con cepas de NmB, demostrándose el carácter protectogénico de la Inmunoglobulina específica de origen humano y estableciéndose un excelente correlato de protección. Obtuvimos por primera vez mediante sueroterapia con la Inmunoglobulina antimeningocóccica BC protección frente a retos con NmA y NmC. La Inmunoglobulina por la vía intravenosa resultó más eficaz que la vía intraperitoneal frente a los retos con Nm de los serogrupos A, B y C. - Sé logró comprobar la eficacia de VA-MENGOC-BC y la Inmunoglobulina Antimeningocócica BC en ratones frente a diferentes cepas de NmB procedentes de casos clínicos de América Latina. Obtuvimos por primera vez protección en ratones vacunados con VA-MENGOC-BC, al ser retados con NmA y NmC. Capítulo V INOCUIDAD DEL PRODUCTO VA-MENGOC-BC EN BIOMODELOS DE RATA Y RATÓN. Introducción. El empleo de los modelos animales para comprobar la eficacia de la vacuna contra Nm ha sido estudiado, se hace necesario entonces determinar los posibles efectos tóxicos de esta y de ese modo establecer la adecuada relación riesgo - beneficio que garantice la seguridad de la misma y cumplir con ello parte de los requisitos preclínicos para su uso en humanos. La inoculación de vacunas por la vía intramuscular constituye una práctica común en medicina humana y veterinaria. Los antígenos inyectados por esta vía inducen procesos tisulares. En muchos casos las vacunas requieren de sustancias capaces de potenciar y prolongar su efecto protector por largo tiempo, dichas sustancias reciben el nombre de adyuvantes. Algunos de estos, los llamados adyuvantes de depósito, generan procesos granulomatosos al nivel de los tejidos donde son inyectados (Infante et al., 1995). La mayoría de las vacunas utilizadas actualmente usan compuestos de aluminio como adyuvantes. El fosfato y el hidróxido de este metal han resultado los mejores. A menudo la cantidad de adyuvante requerido para obtener una eficacia adecuada es tan alta que los principales efectos adversos de la vacuna dependen del adyuvante. Lesiones locales severas, como la tumefacción, el rubor y el dolor están frecuentemente asociadas a la dosis del adyuvante. El metabolismo del aluminio aún no se conoce bien, pero se sabe que es excretado por los riñones y que su toxicidad es de tipo acumulativo (Gupta y Relyveld, 1991). En el caso de la vacuna VA-MENGOC-BC se conocen estudios realizados a los procesos lesionales inducidos por este producto en los tejidos de ratones que fueron inoculados por vía IP (Infante et al., 1989), y en conejos usando la vía intramuscular con adyuvantes oleosos (Mebus y de Mello, 1981). No obstante, la importancia que tiene la relación entre la respuesta tisular, los posibles daños en el ámbito de órganos y tejidos, justifica la profundización en el conocimiento de estos aspectos. Junto a los avances de la Vaccinología sigue prevaleciendo la evaluación del posible efecto que puedan provocar vacunas como VA-MENGOC-BCa escala sistémica, debido a que se han sentado pautas, en cierta medida rígidas y difíciles de enfrentar en el mundo desarrollado, para la introducción de productos en el mercado. Esto constituye un reto y una necesidad imperiosa para que nuestro país perfeccione la metodología ya existente, evaluando los efectos “in vivo” que pueda acarrear la referida vacuna en los biomodelos. OBJETIVOS: - Determinar los posibles efectos adversos desde el punto de vista clínico veterinario y de las lesiones anatomopatológicas causadas por la administración de la vacuna VA- MENGOC-BC en ratas Sprague Dawley. - Determinar la tolerancia local provocada por la referida vacuna en las ratas Sprague Dawley y ratones Balb/c. Caracterizar histopatológicamente el granuloma inducido. - Estimar la asociación existente entre los títulos de anticuerpos contra VA-MENGOCBC® en los ratones inoculados y la duración de los granulomas generados a nivel del músculo. MATERIALES Y METODOS Ver Diagrama Capítulo V. (Reverso página anterior). A- Evaluación toxicológica de VA-MENGOC-BCen ratas Sprague Dawley. 1.Animales experimentales. (Ver Capítulo II, Materiales y Métodos Generales, Acápite II1) La atención de las ratas se realizó dé acuerdo con las normas institucionales establecidas para el cuidado y uso de animales de laboratorio, según la Guía para el cuidado y empleo de los animales de laboratorio de la Council on Animal Care, (1980). Estas fueron cuarentenados y observados en detalle por 7 días, posteriormente se distribuyeron por sexos en cajas tipo Macrolón 2154F001, de tamaño suficiente para permitir el libre movimiento dé las mismas. Recibieron una dieta adecuada de alimento pelletizado para roedores, fórmula CM 01000 certificado por CENPALAB y agua fresca acidulada con HCl a un pH de 2.5-2.8 “ad libitum”. Además, este centro suministró la cama de bagazo de caña, la cual se encontraba libre de sustancias nocivas, siendo la misma cambiada en días alternos. 2-Distribución de los animales experimentales. Al concluir la cuarentena y verificarse el estado de salud de los animales, estos fueron seleccionados para el experimento, distribuyéndose aleatoriamente a razón de cinco animales por cajas, dos cajas por cada grupo (una por sexo), mediante un sistema automatizado suministrado por el Instituto de Farmacia y Alimentos (IFAL) de la Universidad de la Habana. El peso corporal inicial de los animales estuvo en el rango de 272,3 ± 54,5 g para las ratas machos y 192,3 ± 38,46 g para las ratas hembras. 3-Sistema de identificación. La identificación de los animales se realizó individualmente mediante tatuaje de ambas orejas usando tatuador para roedores EBECO (T 21R3104341), asignándose un único número a cada animal. Por otra parte se colocó una tarjeta a cada caja con las siguientes informaciones: - Especie, sexo, fecha de recepción, número de cuarto, número de animales por cajas, código de estudio, número aprobado del protocolo por el Comité Bioético del centro y patrocinador. 4-Control de la sustancia de estudio. 4.1-Sustancia de estudio. (Ver Capítulo II, Materiales y Métodos Generales, Acápite II-7) La sustancia ensayada fue la vacuna VA-MENGOC-BC, envasada en viales de 10 dosis, cantidad suficiente para el estudio y con la calidad requerida, cuyo certificado de calidad fue emitido por el Departamento Técnico y Control Analítico, así como por el Centro Estatal para el Control de la Calidad de Medicamentos (CECMED) del Ministerio de Salud Pública. Esta vacuna satisfizo todos los Standard Nacionales y requerimientos establecidos por el Registro de Drogas Cubanas. Una vez que la vacuna fue aceptada por la entidad de aseguramiento de la calidad, la misma se almacenó hasta ser usada, a la temperatura indicada (2-8 0C), en frascos de vidrio transparente con tapa de goma y retapa de aluminio. 4.2 – Placebo (Sustancia control positivo) Gel de Hidróxido de Aluminio 2 mg Tiomersal 0.05 mg Hidrógeno fosfato disódico 0.03 mg Dihidrógeno fosfato de sodio dihidratado 0.02 mg Cloruro de sodio 4.25 mg Agua para inyección 0.5 ml Condiciones de Almacenamiento: Mantenidas a temperatura de 2-80C. La materia prima y los materiales empleados en la elaboración de la sustancia utilizada como control fue analizado, a fin de garantizar la calidad de los mismos, Estas fueron avaladas por el patrocinador (Control No .084). 5-Preparación de la dosis de la sustancia de estudio. Los viales de 10 dosis se mantuvieron cinco minutos fuera del refrigerador, previos a la inoculación, siendo sometidos a agitación suave para lograr su adecuada homogeneización. Las cantidades empleadas en el experimento se obtuvieron con jeringuillas plásticas desechables de 1ml y agujas hipodérmicas calibre 23Gx1y1½. 6-Procedimiento a seguir para el tratamiento y rango de dosis. Atendiendo a que no existe una definición reguladora para llevar a cabo un estudio de toxicidad en productos biológicos semejantes a las vacunas y considerando que la sustancia ensayada posee un programa de administración discontinua con intervalos de 6-8 semanas entre la primera y la segunda administración, se elaboró un esquema de inmunización que permitió seguir una evaluación del riesgo derivado de la misma, acorde con la administración propuesta para humanos. Los animales fueron inoculados por vía intramuscular en las extremidades, estableciéndose dos variantes: una variante propuesta para uso clínico y la otra propuesta como dosis subaguda. En la primera variante se emplearon tres niveles de dosis, la dosis mínima que es equivalente a la dosis empleada en humanos, con un volumen de 0.5 ml (Grupo II) en una inoculación; la dosis media fue de 1 ml (Grupo III) y la dosis máxima, 1.5 ml (Grupo IV). Estas dos últimas dosis fueron equivalentes en dos y tres veces la dosis de inmunización, aplicándose en dos y tres sitios de inoculación, respectivamente, repitiendo este procedimiento a los 42 días de la primera dosis (dosis de refuerzo). Por su parte, los animales empleados como control (Placebo) recibieron una dosis de 0.5 ml (Grupo I). La segunda variante de inoculación fue en dosis única, el volumen fue de 1.5 ml (Grupo V) y se aplicó en tres puntos diferentes, solo en una ocasión. En la tabla V/M/1 se muestran las dosis empleadas para cada grupo en estudio y el esquema seguido. Tabla V/M/1: Sustancias del ensayo y dosis. Grupo Volumen (ml/Animal) Dosis de cada principio No. de inoculaciones activo. (g/ Animal) I 0.5 1 - II 0.5 1 50 III 1 2 100 IV 1.5 3 150 V* 1.5 3 150 *Grupo V: no recibió inmunización secundaria. 7-Observaciones clínicas Los animales fueron observados detalladamente luego de ser administrada la sustancia en estudio y la sustancia control, realizando un programa de examen del comportamiento físico general de cada animal, para lo cual se tuvo en cuenta las siguientes manifestaciones: - Movimientos voluntarios, involuntarios, salivación, lagrimeo, ataxia, anorexia, incoordinación, características de la piel, el pelo y las mucosas visibles. 8- Determinación de la letalidad causada por VA -MENGOC –BC. La letalidad se determinó diariamente, representando el tiempo de vida del animal desde el momento en que se administró la vacuna VA-MENGOC-BC hasta la posible muerte del animal. Esta depende del efecto tóxico que pudiera causar la vacuna sobre los animales para cada dosis en estudio. 9-Estudio Hematológico. Las determinaciones hematológicas se llevó a cabo en las dos etapas del experimento 30 días para el grupo V y 57 para los grupos del I al IV, de los cuales se obtuvieron 5 ml de sangre por animal mediante extracción a través de la vena cava al realizarse la necropsia. La sangre se distribuyó en alícuotas de 0.5 ml para las diferentes determinaciones. Para la determinación de los valores de hematocrito se siguió el PNO 51.33 BO. 008.CENATOX Las células rojas y leucocitos fueron determinados en la primera y segunda dilución acorde con la técnica descrita en el sistema de operaciones manuales. Los valores son expresados en unidades de 106 cél/l y 103 cél/ l respectivamente. El Conteo diferencial de los leucocitos se desarrolló según PNO 51.33 BO. 006: CENATOX 10-Estudio bioquímico de la sangre. Para las determinaciones bioquímicas, la sangre fue colectada en viales plásticos y se centrifugó a 1200 r.p.m. después de la formación y retracción del coágulo. El suero obtenido fue trasvasado a viales plásticos y guardado a –20 0C. La actividad enzimática de la fosfatasa alcalina, TGO, TGP y colinesterasa se determinó con un analizador automático Hitachi Modelo 705 (Hitachi, Boehringer Mennhein), del Departamento Técnico de Especificaciones de la Calidad del Hospital Carlos J. Finlay. Además, se determinaron otros indicadores tales como: bilirrubina, creatinina, proteínas totales, triglicéridos, urea, nitrógeno, glucosa y calcio. 11-Estudios serológico (Ver Capítulo II, Materiales y Métodos Generales, Acápite II-16) 12-Estudios anatomopatológicos.( Ver Capítulo II, Materiales y Métodos Generales, Acápite II-12) El análisis anatomopatológico se realizaron aplicando el PNO 61.34 BO. 005: CENATOX comenzó el día 30 para el grupo V y el día 57 para los grupos I – IV, con eutanasia de todos los animales después de su narcosis con una sobredosis de Pentobarbital Sódico, seguida del desangrado y necropsia, teniendo en cuenta las recomendaciones para la eutanasia de los animales de experimentación (Council on Animal Care, 1980). Se realizó el estudio anatomopatológico general con énfasis en el cerebro, médula espinal, mesenterio y nódulos linfático submaxilares y otros órganos como hígado, timo, corazón, pulmones, bazo, riñones y punto de inoculación, los cuales fueron pesados en balanzas Sartorios Modelo LC6200S y LC 1200 S. En la toma de muestras los órganos fueron colectados en frascos de boca ancha de color ámbar, los cuales estaban adecuadamente rotulados con el número de estudio, número del animal y grupo de estudio para su posterior procesamiento y análisis histopatológico. Los órganos fueron fijados en solución buffer de formalina al 10% para las primeras 24 horas, luego se transfirieron a una solución menos concentrada al 4% para su conservación. Posteriormente los fragmentos referidos fueron procesados en el Histoquine Modelo Leyca 2000 (procesador de tejidos) e incluidos en parafina distribuyéndose en bloques de la siguiente manera: - Bloque No 1: Corazón, hígado, bazo, riñón y ganglios. - Bloque No 2: S.N.C. (Cerebro y cordón espinal), glándulas salivares, ganglios linfáticos y pulmones. - Bloque No.3: Piel y músculo (Area de inoculación), aparato digestivo (Intestino delgado e intestino grueso). Luego los bloques fueron cortados en secciones de 4-6 micras de espesor, mediante un micrótomo Modelo Leyca 1400. El número de secciones se realizó acorde con las recomendaciones del Procedimiento Operativo de Toxicología, Patología, Histopatología. Centro Nacional para la Producción de Animales de Laboratorio y el Centro de Toxicología Experimental(CETEX) (POT/PAT/ HO5. Métodos Histológicos Empleados. Las muestras de tejidos fueron procesadas, empleando 209 láminas correspondientes a 60 animales que pertenecían a los 5 grupos de estudio y el placebo correspondiente. Las mismas fueron sometidas a diferentes métodos histológicos, los cuales permitieron verificar el efecto de la vacuna en las diferentes dosis de tratamiento con respecto al grupo control. Para ello se empleó la coloración de Hematoxilina-Eosina como método de rutina. Además, en las secciones que contenían músculo, se realizó la coloración de Van Giesson (Para la tinción de fibras colágenas) y la técnica histoquímica del PAS (Acido Periódico de Schiff) para determinar la presencia del polisacárido a nivel del sitio de inoculación. Las observaciones se realizaron en microscopio convencional Leyca y microscopio Olympus (modelos DMLB y CH-2 respectivamente), estudiándose detalladamente las lesiones encontradas en los tejidos para cada animal y en cada grupo de estudio. Los datos primarios se registraron en modelos habilitados al efecto y posteriormente en tablas para facilitar su interpretación. (Ver Materiales y Métodos Generales, Acápite II- 12). 13-Análisis estadístico. A todos los grupos de valores pertenecientes a las variables de naturaleza continua y medición escalar o proporcional se les verificó la homogeneidad de las varianzas mediante la prueba de Bartlett y se realizó el análisis de la normalidad. Teniendo en cuenta estos resultados se aplicaron pruebas paramétricas o no paramétricas para comparar las distribuciones de los diferentes grupos experimentales. En todos los casos se consideraron significativos los valores de p<0.05 y se usó para los análisis estadísticos el paquete STATISTICA for Windows, Release 4.5, StatSoft, Inc. 1993. Las variables hematológicos y bioquímicas y el peso corporal se compararon mediante análisis de varianza (ANOVA) de dos factores (sexo y tratamiento) y la prueba de diferencias significativas mínimas (LSD). El test de Kruskal Wallis y el de comparaciones múltiples para distribuciones libres fueron empleado para procesar las muestras del estudio de inmunogenicidad. B- Evaluación anatomopatológica de VA-MENGOC-BC en ratones Balb/cj utilizando la vía intramuscular. Se utilizaron en este experimento 99 ratones machos de la línea Balb/cj de peso inicial comprendido entre 17 y 22 g, mantenidos en las condiciones de tenencia y alimentación convencionales. Todos los animales fueron vacunados con VA-MENGOC-BC por vía intramuscular a nivel del músculo bíceps femoral de la extremidad derecha, con dosis única de 0,05mL. Para otros datos referentes a los animales, inoculaciones, etc (ver Capítulo II, Materiales y Métodos Generales Acápite II - 1, 2, 3,4, 5,6). Los datos referentes a la inoculación, la vacuna y el método de ELISA utilizado (ver Capítulo, Materiales y Métodos Generales II Acápite, II –16). Los animales fueron sacrificados luego de ser desangrados en grupos de tres con la siguiente periodicidad:12 y 24 horas y los días siguientes: 2, 4, 6, 8, 12,16,20, 24,28,32,36,40,44,48,5,60,67,75,85,92,110,117,125,135,143,158,169,204,241,263 y 286. Las necropsias se realizaron mediante los métodos de rutina, haciendo énfasis en el punto de inoculación de la vacuna y en los ganglios regionales que drenan la linfa correspondiente a la zona relacionada con el referido punto. Se tomaron muestras para el estudio histopatológico de esta zona, las cuales fueron fijadas en formalina neutra al 10%, procesadas por la técnica convencional y coloreadas con Hemotoxilina-Eosina, VanGiesson y Aluminón al 2%. Las investigaciones serológicas para la determinación de los niveles de anticuerpos contra NmB se realizaron coincidiendo con los sacrificios y se aplicó la técnica de ELISA con este objetivo. RESULTADOS Y DISCUSION A- Evaluación toxicológica de VA-MENGOC-BCen ratas Sprague Dawley. En los resultados de este experimento se evidencia el comportamiento de los diferentes parámetros analizados en las ratas Sprague Dawley de ambos sexos, de acuerdo con el esquema de inmunización seguido en las dos variantes analizadas: Una variante para uso clínico en tres niveles de dosis (dosis mínima, dosis media y dosis máxima) y la variante propuesta como dosis subaguda. En las figuras V/A/1 y V/A/ 2 se presentan los resultados del peso corporal en las ratas Sprague Dawley hembras y machos respectivamente. Los valores se corresponden con lo reportado por (Alemán et al., 1998) para esta especie y edad. Los resultados obtenidos respecto a la letalidad causada por VA-MENGOC-BC® arrojaron un 100% de sobrevida en los grupos tratados con la vacuna durante el experimento. Por otra parte no se observaron síntomas clínicos en la conducta de los animales que pudieran ser atribuidos a la vacunación. Se observó un incremento progresivo del peso corporal en cada grupo en estudio hasta concluir el experimento a los 30 días para el grupo V, V/A/3 y a los 57 días para el resto de los grupos, cuyo comportamiento se corresponde con lo planteado para esta especie por (Alemán et al., 1998) figura V/A/3. Estos resultados confirman las evidencias obtenidas en Ratas SPF (Infante et al., 1997) descritos en estudios anteriores al usarse la vacuna VA-MENGOC-BC con respecto a los aspectos clínicos y de la mortalidad. Estudio hematológico: Primeramente debemos aclarar que todos los parámetros hematológicos se mantuvieron en el rango de valores considerados normales para esta especie, línea, sexo y edad. En las (Tablas V/A/1 y V/A/2) se muestran las medias y desviaciones estándares, los valores de la hemoglobina y el hematocrito para ambos sexos. Con relación a la hemoglobina se obtuvieron diferencias estadísticamente significativas con respecto al grupo control, no obstante todos los grupos tuvieron medias comprendidas entre los intervalos normales. Para el hematocrito se observaron algunas diferencias significativas con respecto al control (Placebo) en las ratas hembras para la dosis media (Grupo III) y dosis máxima (Grupo IV), mientras que en las ratas machos no se observaron diferencias con respecto al control, pero sí entre el grupo IV y el grupo V, dosis máxima y dosis subaguda respectivamente. Los resultados obtenidos en el conteo de leucocitos totales (Tabla V/A/1 y V/A/2) no muestran diferencias significativas entre los grupos, obteniéndose un aumento de los leucocitos sanguíneos para la dosis máxima (Grupo IV ratas hembras) con respecto al resto de los grupos. El resultado se relacionó con el conteo diferencial que se realizó para los mismos, resultando superiores el porciento de los linfocitos en sangre de todos los grupos tratados y el Placebo, no así para los polimorfonucleares neutrófilos y monocitos. Química sanguínea: Los valores promedio en cuanto a la química sanguínea para ambos sexos se presentan en las (Tablas V/A/3 y V/A/4), observándose estadísticamente significativas para el caso de las ratas hembras en cuanto a: diferencias - La TGO entre los grupos I y II, así como entre los grupos III y IV; sin embargo estas diferencias no se observan en las ratas machos. - La TGP difiere significativamente en las hembras al compararse los grupos II y III con el control positivo y también se observaron estas diferencias en los grupos II y IV. Por su parte las ratas machos mostraron estas diferencias entre los grupos IV y V. - Los resultados obtenidos en cuanto a la fosfatasa alcalina no arrojaron diferencias entre los grupos tratados con respecto al control, pero sí entre el grupo IV y V de las hembras. Por su parte, los machos no presentaron diferencias en cuanto a estas variables. - Los valores obtenidos en el balance Urea-Nitrógeno muestran diferencias significativas entre los grupos IV y V de las hembras y no entre los grupos tratados con respecto al control. Cuyas diferencias sí fueron observadas en las ratas machos en el grupo V con respecto al control y entre los grupos II y III. - Creatinina: En las hembras fueron encontradas diferencias entre los grupos II y III, II y IV, grupos III y I, así como entre los grupos IV y V. En los machos solo se encontraron diferencias entre los grupos IV y V. - Calcio: Se observaron diferencias significativas entre los grupos IV y V en ambos sexos. - Proteínas totales: En las ratas hembras se observaron diferencias significativas entre los grupos II y III con respecto al control, encontrándose diferencias entre los grupos II y IV, entre los grupos III y IV. En los machos no hay diferencias. El resto de los parámetros, bilirrubina directa e indirecta triglicéridos y glucosa no mostraron diferencias significativas en ninguno de los grupos. Los resultados en cuanto a la hematología y química sanguínea denotan algunas diferencias significativas entre los grupos tratados con respecto al control y entre los tratados entre sí, principalmente entre las ratas hembras. Estas diferencias no parecen ser dosis dependientes, debido a que los valores no están distribuidos de forma similar para todos los grupos. En contraste, el tamaño de muestra es pequeño, lo cual pudo influir en los resultados, sin embargo estos se encuentran dentro del rango normal establecido por la Guía para el cuidado y uso de animales de laboratorio propuesto por la (Council on Animal Care, 1980),(Alemán et al., 1998). Estudio de inmunogenicidad: Los componentes proteicos de la vacuna Antimeningocócica VA-MENGOC-BC®, desencadenaron una respuesta humoral en los grupos vacunados (p0.0005). Las muestras obtenidas de los machos del grupo I no se tuvieron en cuenta en los análisis debido que la cantidad de muestra no fue representativa para realizar el estudio. Las ratas hembras mostraron una respuesta superior que los machos del grupo III (p 0.05) y IV ( p 0.02). Por otra parte se obtuvo el mayor título de anticuerpos en las ratas hembras del grupo IV al ser comparadas estas con el grupo V (p0.04). El promedio de los valores de ambos sexos se muestra en la figura V/A/4. La respuesta inmune después de la vacunación fue evaluada por el ensayo de ELISA, encontrándose aumento significativo de la respuesta de IgG después de la vacunación, demostrando la buena inducción de anticuerpos séricos que provoca esta vacuna. Los resultados obtenidos en el título de anticuerpos específicos contra la proteína de la membrana externa de NmB mostraron altos niveles para cada variante de vacunación analizada con respecto al control. Este último no presentó reactividad frente a los antígenos de la bacteria, como era de esperar, debido a que los adyuvantes tienen la característica de no desencadenar por sí solos la respuesta inmune (Roitt, 1994). Los mejores resultados fueron obtenidos en las hembras del grupo III (dosis media) con un título de anticuerpos de 12952 u/mL. Se puede inferir que esto pudiera deberse a factores hormonales reportados para esta especie o a la respuesta inmunológica del animal contra los antígenos de la bacteria cuya dosis supera a la usada en humano. Esto puede acarrear una mayor respuesta frente a los antígenos con el correspondiente aumento de la respuesta humoral, mientras que no ocurre igual en el caso de la dosis máxima (Grupo IV) en que a pesar de poseer mayor dosis no se logra una mayor respuesta. Esto pudiera estar influenciado por un exceso de antígenos, cuya propiedad tal vez este relacionada con el efecto de zona planteado por Roitt,(1994). Este autor postula que al encontrarse los anticuerpos y/o los antígenos en exceso ocurre una sobresaturación que trae consigo resultados muy por debajo de los resultados que pudieran esperarse, o a un fenómeno de tolerancia inmunológica previamente descritos en estos casos. Las diferencias observadas entre las hembras y los machos del grupo IV (dosis máxima) y las del V (dosis subaguda) en relación con los diferentes parámetros analizados pudieran ser esperadas debido a que el esquema de inmunización y el tiempo de sacrificio fueron diferentes. Se conoce la necesidad de una inmunización secundaria para lograr una alta producción de anticuerpos, fenómeno denominado memoria inmunológica (Abbas, 1995). Esto se debe a que, en una segunda inmunización, los linfocitos tienen la capacidad de reconocer los antígenos de forma rápida trayendo consigo un reforzamiento de los mecanismos fisiológicos de la reactividad inmunológica, con el consiguiente aumento de los anticuerpos circulantes. La evaluación de la memoria es un área de transcendental importancia para la vaccinología. Los procedimientos empleados para su evaluación pueden resumirse en: efecto de refuerzo (“booster”), tras un nuevo desafío antigénico algún tiempo después de la inmunización, estimulación de IgG en las inmunizaciones parenterales, mayor concentración y afinidad de los anticuerpos producidos (Roitt, 1994). Estudio anatomopatológico: Las observaciones macroscópicas no arrojaron ninguna variación relacionada con la textura y color de los órganos en los grupos de tratamiento, ni en el grupo control. En la necropsia se observó una esplenitis moderada en un animal correspondiente al grupo III. Los estudios histológicos no mostraron alteraciones en la citoarquitectura de los tejidos estudiados que podían ser atribuibles a procesos de tipo tóxico (figuras desde la V/A/1 hasta la V/A/4 ver ANEXOS Capítulo, V) La (Tabla V/A/5 y VA/6) muestran el peso de los órganos. Se observaron algunas diferencias significativas entre las ratas hembras con relación a los pesos de órganos tales como: hígado, pulmón, riñón, timo y bazo para los grupos de tratamiento. Estas diferencias no se observaron en las ratas machos. Estos resultados demuestran la gran variabilidad que puede existir entre animales de una misma especie con la misma edad y sexos diferentes, analizadas con el mismo rango de dosis. A pesar de las diferencias anteriormente citadas, estos valores estuvieron dentro del rango normal reportado para estas ratas por (Alemán et al., 1998). En las (Tablas V/A/7 y V/A/8) se muestra la correlación entre los pesos de los órganos linfoide, y los resultados hematológicos e histológicos de estos órganos a fin de determinar la posible existencia de algún efecto inmunotóxico de la vacuna, los cuales no fueron observados. Desde el punto de vista histopatológico, fueron encontrados algunos cambios, siendo considerados incidentales en este tipo de estudio (Tabla V/A/9), destacándose las urolitiasis renales y las esplenitis. La urolitiasis tubular renal observada en todos los grupos está descrita como una lesión de frecuente presentación en ratas jóvenes, fundamentalmente en las hembras. Su presencia se relaciona con el contenido de calcio y fósforo en la dieta (Magnusson y Ramsay, 1971), (Casey et al., 1978), (Clapp, 1980). La hemosiderosis del bazo tiene igualmente poca connotación, ya que está relacionada con la función hemocaterética de este órgano y con los niveles de hierro (Richter, 1974), (Ward y Reznik, 1980). Las formaciones granulomatosas macrofágicas han sido reportadas como consecuencia de los adyuvantes empleados, Hidróxido de Aluminio, Glucanos, Adyuvante Completo de Freud (ACF), Adyuvante Incompleto de Freud (AIF), Aceite mineral entre otros (Azuma, 1993), (Infante et al., 1995), (Cox y Coulter, 1997), cuyo modo de actuar se basa en la generación de un depósito en el sitio de la inoculación, permitiendo la lenta liberación del antígeno, con lo que se prolonga el tiempo de interacción de estos es decir, las células presentadoras de antígenos (CPA) y los linfocitos. Es necesario que la CPA capten el antígeno y este sea tansportado por las referidas células, hasta los ganglios linfáticos regionales, donde ocurren entre otros eventos la presentación y proliferación de los linfocitos T y B (Moncada y Palmer, 1990),(Nathan, 1992), (Rivero et al., 1993). Similar fenómeno es de esperar que ocurra en el humano, donde la separación entre las dosis es de 6-8 semanas. Por otra parte los compuestos de aluminio activan la vía clásica del complemento provocando una respuesta inflamatoria local, importante en el desarrollo de las células B de memoria e inducen a los linfocitos T a desarrollar una respuesta de tipo Th2. Los granulomas macrofágicos han sido descritos en estudios toxicológicos llevados a cabo en otras vacunas que contienen hidróxido de aluminio como adyuvantes. La presencia de esta sustancia dentro del granuloma ha sido demostrada en otro trabajo mediante una coloración especial para detectar aluminio. Se encontraron estructuras de tipo granulomatosas macrofágicas en los músculos a nivel del sitio de inoculación. Estas estuvieron bien delimitadas por el tejido conjuntivo que las rodeaba, formando un halo que circunscribe el proceso, compuesto por fibroblastos y fibrocitos, (identificados por la tinción para fibras colágenas Van Giesson). A través de la técnica de PAS, los granulomas resultaron positivos, indicando la presencia de polisácaridos en su interior. Además estas lesiones estuvieron caracterizadas por acumulaciones de histiocitios, macrófagos (que contenían en su interior abundantes depósitos de una sustancia granular basófila) y otras células inflamatorias, incluyendo un gran número de neutrófilos, también se observó la movilización de los endotelios capilares en los vasos sanguíneos de la zona adyacente. Las formaciones de granulomas en el sitio de la inyección se observaron en todos los grupos, no así en todos los animales de cada uno de estos. Estas diferencias individuales pudieran estar influenciadas por dificultades en las tomas de las muestras de los músculos. La relativa larga duración de los granulomas en el músculo puede estar relacionada con la adsorción de las PME de esta vacuna en el hidróxido de aluminio. Como es sabido uno de los mecanismos de acción de los adyuvantes es servir de depósito (Rosenqvit et al 1991). B- Evaluación anatomopatológica de VA-MENGOC-BC en ratones Balb/cj utilizando la vía intramuscular. En los animales investigados macroscópicamente no se encontraron alteraciones en órganos y tejidos de los ratones excepto un proceso inflamatorio agudo en la zona del punto de inoculación a las 12 horas post-inoculación. Este caracterizado por la presencia de hemorragias puntiformes con un diámetro aproximado de 0,5 mm. Posteriormente, apareció una coloración grisácea de aspecto opaco, algo difusa en el ámbito de los planos de clivaje muscular. A partir del sexto día se constató un aumento de tamaño del ganglio linfático poplíteo, correspondiente a la extremidad inoculada y de los ganglios inguinales profundos. Estos aumentos se mantuvieron hasta el día 36. A partir del día 24, la coloración se tornó amarillo-grisácea, con disminución paulatina en el diámetro de la lesión y tendencia a ser más larga que ancha. A los 158 días la lesión fue poco perceptible, apareciendo una pequeña área gris blanquecina de 0.5 a 1 mm de diámetro, que persistieron hasta los 286 días post-inoculación. Aunque se observó una considerable disminución de tamaño, estas lesiones son similares a las descritas en otras especies inoculadas con biológicos que contenían adyuvantes de depósito (Infante et al., 1989). En otros tejidos de la economía animal no se observaron lesiones de valor diagnóstico. Cronológicamente, desde el punto de vista histopatológico, a las 12 horas post-inoculación se encontraron amplias zonas de hemorragias, con necrosis de Zenker en los músculos del sitio de inoculación, áreas de infiltración a partir de los polimorfonucleares neutrofilos y acumulaciones de sustancias granulares basófilas. (Figura V/B/1 y V/B/2). Cerca de los pequeños vasos de esta zona abundan los neutrófilos distribuidos concéntricamente, hay movilización del endotelio capilar y edema entre las fibras musculares, todo lo cual se corresponde con los procesos inflamatorios agudos. Transcurridos dos días, al proceso se suma un infiltrado de escasos macrófagos, haciéndose menos destacadas las hemorragias y el edema intersticial. Al cuarto día hay amplias zonas de miocitólisis, donde se aprecian numerosas fibras musculares con aspecto hialino, pérdida de las estrías transversas, retracción y fragmentación parcial del sarcolema y amplias áreas edematosas en el intersticio, con abundantes macrófagos llenos de aluminio. El tejido conjuntivo fibroso mostró una marcada tendencia a circunscribir el proceso inflamatorio, formando un halo con vasos neoformados y fibroblastos. Coexistió en este estado un escaso número de neutrófilos y, cerca de las vénulas, abundantes linfocitos pequeños que en su mayoría forman manguitos perivasculares. Al nivel de los ganglios linfáticos correspondientes a la zona de inoculación, al sexto día se encontraron discretas hiperplasias de los folículos paracorticales (figura V/B/3) y algunas células plasmáticas en los senos, que se hicieron más numerosas hacia el hilio A partir de este día las lesiones en el músculo mantuvieron las características antes descritas, aunque hubo un aumento gradual del tejido conjuntivo (demostrado mediante la coloración de VanGiesson,) hasta el día 75, y una tendencia del granuloma a hacerse más macrofágico y menos piógeno. En los ganglios linfáticos se incrementaron las hiperplasias paracorticales hacia el decimosegundo día; al igual que el número de células plasmáticas a nivel de los senos del hilio. Todas las características se mantuvieron, salvo discretas variaciones respecto a la deposición del tejido conjuntivo. Hasta el día 85, los granulomas se hicieron muy discretos, conformados por macrófagos y escasos neutrófilos, siendo gruesas las cápsulas de tejido conjuntivo. Esto se mantuvo de forma reparativa hasta el día 286, en que se culminaron las observaciones (Figuras de la V/B/4 hasta la V/B/6). Estos hallazgos pueden estar en relación con lo plateado por Roitt, (1994), quien consideró que en los casos en que el antígeno persiste localmente en determinado tejido no linfoides, como es el músculo, su presencia estimula las reacciones macrofágicas, seguidas por las segregaciones de linfocitos y liberaciones de linfocinas lo que conduce a la formación de granulomas. Por otra parte, gracias a la destrucción del exceso de antígeno y a la presentación por los macrófagos del resto en forma fuertemente inmunogénica, se puede inhibir la tolerancia inmunológica y reforzar de este modo la respuesta inmune, haciendo del pequeño granuloma macrofágico un evento de significativa importancia en el desarrollo del proceso de inmunización. Sin embargo en las reacciones inflamatorias iniciadas por el sistema inmunitario, el control último es ejercido por el propio antígeno, del mismo modo en que éste es controlado por la respuesta inmunitaria. Por este motivo, las acumulaciones celulares en los lugares de reacciones autoinmunitarias (en las que el antígeno no puede quedar finalmente erradicado) son completamente diferentes de las de los lugares donde el estímulo antigénico queda rápidamente eliminado (Pround y Kaplan, 1988), (Male, 1991). Por otra parte las lesiones descritas en nuestro experimento fueron similares a las encontradas por McKercher et al., (1971) cuando estudiaron formaciones de origen similar en otras especies, con la única diferencia de que solo completaron los estudios hasta los 90 días post-inoculación. Los resultados del estudio de anticuerpos en sangre (Gráfico V/B/1) muestran que a partir del decimosexto día comienzan a incrementarse para alcanzar un máximo a los 67 días. A partir de entonces decrecen progresivamente hasta desaparecer luego de los 160 días aproximadamente. Datos similares fueron reportados por Echeverrg et al., (1995) y Rappuoli and Gianozzi, (1994), cuando estudiaron vacunas conjugadas de polisacáridos y proteínas de la membrana externa de Nm en humanos. Si tenemos en consideración los niveles de anticuerpos en el suero de los ratones vacunados y su asociación a la presencia de los procesos granulomatosos macrofágicos a nivel muscular, encontramos una estrecha relación entre la presencia e intensidad de éstos y la aparición y mantenimiento de la respuesta humoral, que en el casos de VA-MENGOCBC® es potenciada por la base celular de la respuesta inmune. CONCLUSIONES DEL CAPITULO - La vacuna VA-MENGOC-BC® resultó inocua. - Se demostró que existe tolerancia a la vacuna VA-MENGOC-BC®. La formación de granulomas macrofágicos encontrados en las ratas Sprague Dawley y los ratones Balb/cj en el sitio de la inyección son propias de los adyuvantes de depósitos, las cuales evolucionaron hacia un proceso de reparación. - Se demostró que la presencia del granuloma se asocia a altos títulos de anticuerpos. DISCUSIÓN GENERAL Haciendo un análisis de los resultados obtenidos observamos una franca resistencia natural de los ratones adultos jóvenes utilizados por nosotros frente a NmB si tenemos en cuenta que la enfermedad producida por este germen en el humano no se presenta de forma natural en el ratón. Ello ha sido señalado con anterioridad por varios investigadores (Huet y Suire, 1981) Para vencer este escollo Lausund y Louik, (1994) han recurrido al uso de ratones recién nacidos por ser estos animales inmunológicamente incompetentes. Nosotros decidimos no emplear esta variante al menos para comprobar la eficacia de VA-MENGOC-BC, teniendo en cuenta los objetivos propuestos, entre los que se destacan la necesidad de comprobar los efectos protectores de preparados vacunales, para los cuales es imprescindible disponer de un animal con un sistema inmune maduro, capaz de responder a los antígenos inoculados. En este sentido fue necesario acudir a la utilización de los FEV, los cuales facilitaron la infección del modelo animal ratón, de la categoría adulto joven. Luego de realizar la comprobación de varios de estos factores y sus correspondientes mezclas en experimentos previos, decidimos utilizar primero la Dextrana Férrica y luego mezcla de esta y la mucina gástrica de cerdo, dada su posibilidad de disminuir la LD50 de Nm lo cual favorece la similitud entre el proceso patológico desencadenado por este germen en el referido modelo animal y la enfermedad humana, al realizar una comparación con otros FEV. Resultados similares han sido obtenidos con H. influenzae tipo b por (Sifontes et al., 1999). Lo anterior pudiera tener su explicación en el conocido papel que desempeñan las cantidades del hierro en el desarrollo de las infecciones producidas por muchos gérmenes patógenos. Además hay que tener en cuenta su disponibilidad restringida en los líquidos tisulares por lo que los microorganismos patógenos tienen que enfrentar el problema de adquirir cantidades suficientes de hierro para su multiplicación “in vivo”. No obstante, no debemos olvidar que el hierro también regula la expresión de varios genes metabólicos y de virulencia (Griffiths, 1991) que desempeñan un papel fundamental en el desarrollo de la infección. Los fluidos corporales en estado de normalidad contienen proteínas fijadoras del hierro con una elevada afinidad por este metal como son las transferrinas y la lactoferrinas. Estos fijadores aseguran que no haya hierro libre, útil para el crecimiento y posterior invasión de las bacterias. Patógenos como Neisseria spp. y Haemophilus influenzae que expresan proteínas de membrana que actúan como receptores de alta afinidad para la transferrina y lactoferrina, adquiriendo el hierro por interacción directa entre las proteínas fijadoras y los receptores situados en la superficie celular bacteriana (Holland et al., 1992), pero esta utilización por parte de los gérmenes solo se produce en el humano y algunos primates (Kolsto y Rod, 1990). Como estimulador de la virulencia, la dextrana con hierro resultó un efectivo artificio, toda vez que al emplearlo junto con dosis de 1x107UFC/mL de NmB en el ratón la misma favoreció niveles de mortalidad estadísticamente significativos en comparación con los controles inoculados con los gérmenes solamente. Con esto quedó demostrado el efecto beneficioso que sobre la virulencia ejercen los compuestos del hierro al burlar la llamada “inmunidad nutricional”, mecanismo de defensa inespecífico que poseen los organismos animales y el humano para mantener bajos niveles de hierro sérico, dada la unión de este a las proteínas séricas, disminuyendo su absorción a escala intestinal y aumentando el acumulo de esta sustancia en el hígado. Por su parte la mucina gástrica de cerdo, además de aportar hierra al medio peritoneal, produce determinada irritación al nivel de los vasos sanguíneos facilitando de ese modo la penetración de las bacterias hacia el torrente circulatorio a la vez que crea una especie de trama que le sirve de sostén a los gérmenes propiciando su sobrevivencia. Todo esto hace del ratón adulto joven un buen modelo para reproducir aspectos tales como shock séptico y la forma meningococémica letal de la enfermedad la cual responde mayormente a las endotoxinas del germen que resultan de interés para comprobar la eficacia de preparados vacunales y sueros como la vacuna VAMENGOC- BC y la Inmunoglubulina Antimeningocócica BC, los LPS presentes en el proteoliposoma de la vacuna generan una potente respuesta inmune específica que provoca la inducción de anticuerpo neutralizantes, antiLPS presentes tanto en la Inmunoglobulina como en la vacuna. Otros antígenos como las proteínas de la membrana externa de este germen generan una importante respuesta humoral y celular que contrarresta la patogenicidad del este. La utilización de los combinados (Mucina y dextrana) como F.E.V para comprobar la eficacia de VA-MENGOC-BC y de la Inmunoglobulina Antimeningocócica, han demostrado la ventaja de disminuir considerablemente las dosis utilizadas para las confrontaciones, pues como es conocido, esta enfermedad se produce en el hombre con la presencia de pequeñas cantidades de las bacterias que llegan al SNC a través de la nasofaringe generalmente, por lo cual mientras menor sea la concentración del inóculo más similitud existirá entre el modelo animal y la enfermedad en cuestión. El cuadro clínico caracterizado por trastornos de carácter general tales como piloerección, fiebre, diarreas, conjuntivitis, ataxia, movimientos de pedaleo y cuyo desenlace final se acompañaba de una disnea grave en los ratones inoculados con NmB y los FEV, estos son síntomas comunes a los procesos septicémicos que se producen en esta especie cuando son infectados por diferentes gérmenes patógenos y en especial organismos responsables de daños en SNC según refieren Hudson, (1976) y McErlean, (1968). Debemos señalar que aunque este modelo no desarrolla un cuadro anatomopatológico florido como el observado en el modelo de ratas infantes donde es conocido su estatus inmunológico caracterizado por la inmadurez en sus mecanismos de la inmunidad, que hacen posible la presentación del referido proceso lesional. No obstante, sí se presentan alteraciones en el SNC como son los procesos congestivos graves al nivel de los vasos meníngeos, los infiltrados inflamatorios focales en los cuales durante las primeras etapas predominan los polimorfonucleares neutrófilos, posteriormente células redondas y las plexitis, lugar de posible acceso de los microorganismos al SNC de acuerdo a lo referido por (Wiliams y Blakemore, 1990). El biomodelo de rata recién nacida resultó útil para la realización de los estudios en el campo de la fisiopatología de los procesos causados por Nm si tenemos en cuenta el cuadro lesional desarrollado a nivel del SNC de estos animales, que estuvo caracterizado por una grave meningitis. Este tiene una gran similitud con la observada en la enfermedad producida por el referido germen cuando afecta a los humanos (Cordy, 1982). Por otra parte este modelo nos permitió comprobar la eficacia de la Inmunoglobulina Antimeningocócica BC en aplicaciones previas o posteriores al reto con NmB. En las ratas recién nacidas los gérmenes alcanzan la sangre y producen una considerable bacteremia sin necedad de utilizar FEV aprovechando las posibilidades que brinda el estado de inmadurez inmunológica de estos animales en sus primeros días de nacidos. Esto permite el paso a la cavidad cerebroespinal de los agentes patógenos, ya que esta zona puede considerarse un espacio abierto según Sanford, (1987). Así, las partículas suspendidas en el LCR están sujetas a sus movimientos a menos que las mismas se adhieran a la superficie aracnoidea o pial, o sean atrapadas por las redes de fibrina (Quagliarello y Scheld, 1992). Después de su entrada en la cavidad cerebroespinal los organismos se dispersan en ella y generan la entrada de células inflamatorias. Mientras este proceso continúa dichas células son transportadas hacia las vellocidades aracnoideas craneales y espinales donde se acumulan, dando lugar a los focos inflamatorios. Esto podría explicar los acúmulos presentes en estos sitios comparado con otras partes del neuroeje (Williams y Blakemore, 1990). Analizando lo explicado y tratando de fundamentar las observaciones realizadas en nuestros experimentos creemos que la distribución de las lesiones en casos clínicos de meningitis generalizadas agudas en los biomodelos, ejemplo: la acumulación de células inflamatorias son resultado del flujo del líquido cerebroespinal en nervios, bulbos olfatorios y vellocidades aracnoideas, parece guardar relación con el punto de entrada de los microorganismos en la cavidad cerebroespinal, concordando con Williams y Blakemore, (1990). Además creemos que el nivel y duración de la bacteriemia son las cuestiones que ejercen mayor influencia en el desarrollo de meningitis generalizadas causadas por bacterias como Neisseria meningitidis grupo B, Haemophilus influenzae b, Escherichia coli Kl, Streptococcus B, coincidiendo con Ferrieri et al., (1980), ya que los cuadros anatomopatológicos más intensos coinciden con los periodos más largos de la bacteremia y los inóculos de mayor concentración. Establecidos los modelos en ratas recién nacidas y ratón, con F.E.V. decidimos comprobar la eficacia de la Inmunoglobulina Antimeningocócica BC frente a retos con NmB. Pudimos apreciar que las ratas recién nacidas cuando fueron retadas con 1x106UFC/mL y posteriormente inoculados con la inmunoglobulina como un método de terapia pasiva, presentaron niveles significativos de protección en comparación con los controles no inmunizados. De igual modo se comportaron los estados de la bacteremia, la cual en los inmunizados desapareció antes de las 24 horas, mientras que en los controles permaneció más de 72 horas. Toda la anterior evidencia demuestra lo útil del modelo con los fines propuestos al permitirnos obtener un valor predictivo de referencia respecto a la protección que este medio terapéutico es capaz de conferir. Los ratones con F.E.V. mostraron resultados similares a las ratas recién nacidas, pero superiores en cuanto a los niveles de protección sobre todo cuando la inmunoglobulina fue aplicada en dosis repetida. Llegaron a obtener 100 % de protección, lo que evidenció las ventajas de una dosificación fraccionada sobre las aplicaciones únicas o con menor frecuencia, mientras que en los controles no inmunizados morían en su totalidad, quedando demostrados de esta forma los efectos beneficiosos de la inmunoglobulina antimeningocóccica BC, tanto cuando se aplicó previa al reto, como posteriormente. En este último caso se simula lo que sucede en sentido general en los humanos que enferman con el meningococo y reciben el correspondiente tratamiento con dosis repetidas de este producto con el cual se obtienen magníficos resultados (Galgura et al., 1991). Por otra parte la protección de la vacuna VA-MENGOC-BC- BC se puso de manifiesto cuando se utilizó el modelo de ratón con la aplicación de los F.E.V para realizar confrontaciones frente a diferentes cepas procedentes de Latinoamérica, evidenciando que a pesar de las elevadas concentraciones de gérmenes presentes en los retos a que fueron sometidos estos animales, se observaron diferencias estadísticamente significativas a favor de los animales inmunizados, en comparación con los controles no protegidos, tanto en el parámetro de tiempo de sobrevivencia como en los niveles de mortalidad. Resultados similares han sido obtenidos para N. meningitidis y H. inflenzae por Hamel et al., (1987) y Olyhoek et al., (1991). El modelo de ratón con FEV evidenció su eficacia también en las pruebas de terapia pasiva con Inmunoglobulina Antimeningocócica BC, frente a retos con las cepas antes mencionadas procedentes de América Latina (Argentina, Colombia, Cuba, Brasil y Chile) del serogrupo B, mostrando altos niveles de protección o un aumento de la sobrevivencia de los ratones protegidos respecto a los controles, este último aspecto guarda similitud con lo acaecido en los humanos que padecen la enfermedad meningocóccica y son tratados con este producto según lo expresado por Galguera et al., (1991). Estos resultados revisten una enorme importancia tanto desde el aspecto referente a la posible eficacia de VA-MENGOC-BC® y de a inmunoglobulina frente diferentes cepas de países afectados de la enfermedad meningocócica en los que mediante estos ensayos es posible un acertado correlato de protección y además las posibilidades de facilitar la comercialización del producto partiendo de la base de la eficacia obtenida con los modelos murinos retados con las referidas cepas. Como se ha podido apreciar se demostraron variaciones entre las DL50 de las diferentes cepas incluso de una misma cepa en diferentes momentos o con diferentes animales, esto coincide plenamente con lo planteado por Sandoval et al., (2000) respecto a que las DL50 a pesar de ser muy utilizada en la toxicología ha sido discutido su objetividad sobre la base de que los valores resultados obtenidos varían ampliamente entre especies animales y razas, con el sexo, la dieta la edad y otros factores exógenos. No obstante, utilizamos estos valores para demostrar las diferencias con respecto a la virulencia de las cepas, cuando usamos un inoculo igual para todas en lo referido a la concentración bacteriana (1x108UFC/mL). Nosotros hemos obtenido resultados satisfactorios en la utilización del biomodelo ratón, tanto en estudios de protección activa con VA-MENGOC-BC® como de protección pasiva con la Inmunoglobulina Antimeningocócica BC a través de retos efectuados con NmB. No obstante, recientes estudios demuestran la existencia de inmunidad cruzada entre proteínas de membrana externa de Nm que incluyen la protección frente a las enfermedades experimentales en el ratón (Fernández et al., 1992). Lo anterior motivó nuestro interés por conocer el espectro de protección de VAMENGOC-BCpara lo cual utilizamos el modelo de ratón con FEV en experimentos de protección mediante reto frente a cepas pertenecientes a los serogrupos A y C obteniendo resultados satisfactorios, tanto en la vacunoterapia como la sueroterapia (datos no publicados). Hay que tener en cuenta que VA-MENGOC-BCno solo es la vacuna más efectiva frente a NmB de las de segunda generación elaborada a partir de proteínas de membrana externa del serogrupo B (OMP) y el polisacárido perteneciente al serogrupo C, sino que por lo genuino de su diseño y formulación, pudiera considerarse la primera vacuna licenciada de los candidatos vacunales, y que además protege frente a los retos con diferentes serogrupos y serotipos de Nm. Vemos, además, que esta mostró eficacia frente a cepas de los serogrupos A y C, resultando interesante la protección cruzada frente un antígeno que no está presente en el preparado vacunal, constituyendo un aspecto a tener en cuenta, pues demuestra las posibilidades potenciales de la vacuna para actuar en epidemia ocurrida a partir del serogrupo A de Nm que a pesar de no estar presente en la formulación de VAMENGOC-BC ofrece la posibilidad de proteger frente a cepas de este serogrupo. No menos importantes resultaron los niveles de protección frente a NmC, aspecto ignorado hasta el momento en este biomodelo y donde muchos sostenían la idea de que en roedores no era posible obtener niveles de protección adecuados frente al serogrupo C debido a que esta especie de roedores no respondía bien a los estímulos antigénicos de origen polisacarídico, (Verheul et al., 1994). Sin embargo, en nuestros resultados se demostró una protección consistente tanto de la vacuna como de la inmunoglobulina frente a los retos con este germen. Los polisacáridos capsulares pertenecientes al serogrupo C, conjugados con proteínas resultan mejores inmunógenos que cuando se utilizan sin conjugar, además se obtuvo una larga persistencia de anticuerpos específicos contra los aislamientos pertenecientes a este serogrupo según Costantino et al., (1992 y Anderson et al., (1994). Lo anterior pudiera estar sucediendo en el caso de VA-MENGOC-BC y la Inmunoglobulina Antimeningocócica BC. Por tanto se justificaría en buena medida la respuesta obtenida frente al serogrupo C, toda vez que esta vacuna contiene las proteínas de la membrana externa de NmB y el polisacárido del referido serogrupo. Los resultados obtenidos mediante ELISA (Datos no publicados) demostraron que aún cuando la vacuna no contiene antígenos proteicos procedentes del serogrupo C, esta es capaz de inducir la formación de anticuerpos contra las proteínas de este serogrupo, sobre todo cuando se realizaron más de una inoculación. Frente al serogrupo A sucedió algo similar; aunque la respuesta fue mucho más marcada y evidente desde la primera inmunización. La presencia de anticuerpos bactericidas contra Nm representa una alta correlación con la respuesta inmune y la protección tanto en humanos como en animales (Goldberg y Bernstein, 1988). Mientras que la existencia del reconocimiento a las bandas de proteínas situadas entre 30 y 100 kDa en los sueros de animales vacunados y enfrentados a los antígenos correspondientes mediante la técnica de Western-blott verifican la existencia de una fuerte reacción antígeno-anticuerpo (Martínez et al., 1998) que pudiera respaldar los resultados obtenidos en estos ratones y que además coinciden con la existencia de antígenos de 22 kDa y de las transferrinas Tbp1 y Tbp2.según Bhatnagar y Frasch, (1990). Estan también incluidas las proteínas de alto peso molecular (65-95 kDa) y otras como las de 70 kDa presentes en varios gérmenes del género Neisseria (Martin et al., 1997) las cuales estimulan la respuesta inmune de amplio espectro para varias cepas de Nm (Goldberg y Bernstein, 1988), (Núñez 1997) sin dejar de tener en cuenta los aspectos inherentes a la respuesta celular demostrada para VA-MENGOC-BC(Lastre et al., 1995). Las pruebas de anticuerpos bactericidas y el ELISA son procedimientos “in vitro” que contribuyen a avalar o no los resultados obtenidos “in vivo”. Estas pruebas evidencian de forma parcial lo que ocurre “in vivo”, pues sus resultados son solo una parte del complejo engranaje que se desencadena cuando ocurre una estimulación antigénica. De ahí la importancia de considerar los biomodelos animales como una herramienta esencial e imprescindible para la evaluación de cualquier candidato vacunal. Después de conocer la eficacia de la vacuna VA-MENGOCV-BC y dando cumplimiento a las regulaciones establecidas, realizamos desde el punto vista toxicológico ensayos utilizando las ratas Sprague Dawley y el ratón Balb/c. Ante la necesidad de comprobar la inocuidad de la vacuna VA-MENGOC-BC® se acometieron ensayos en ratas y ratones que arrojaron los siguientes resultados. Las ratas partenecientes presentaron diferencias entre el grupo IV (dosis máxima) y el grupo V (dosis subaguda) en cuanto a título de anticuerpos, superiores en el grupo IV para las hembras, lo que permite considerar que tal superioridad tal vez pudiera estar relacionada con que el grupo V tenía un esquema de inmunización diferente en cuanto a número de dosis, pues estas solo recibieron una dosis según lo propuesto en el protocolo, con relación al grupo de dosis máxima, ya que estos no recibieron la segunda inmunización, en tanto que el grupo IV fue sacrificado a los 57 días del estudio. Por otra parte las diferencias pudieran estar dadas, por lo reportado respecto a la necesidad de una inmunización secundaria para lograr una alta producción de anticuerpos, propiedad conocida como memoria inmunológica (Abbas, 1995), debido a que los linfocitos tienen la capacidad de reconocer los antígenos de forma rápida trayendo consigo un reforzamiento de los mecanismos fisiológico de la reactividad inmunológica con el consiguiente aumento de los anticuerpos circulantes. La evaluación de la memoria es un área de transcendental importancia para la vaccinología. Los procedimientos empleados para su evaluación puede resumirse en: efecto de refuerzo (“booster”), tras un nuevo desafío antigénico, algún tiempo después de la anterior inmunización; estimulación de IgG en las inmunizaciones parenterales, mayor concentración y afinidad de los anticuerpos producidos (Roitt et al., 1994). La no-existencia de alteraciones entre algunos parámetros hematológicos, el peso de órganos con importantes funciones dentro del sistema inmune como fueron el bazo y el timo, los cuales responden ante la acción de una amplia gama de productos inmunotóxico con las correspondientes alteraciones según Hogan y Hoel (1989), y finalmente los estudios histológicos que no demostraron procesos patológicos en general y en particular al referido sistema, nos hacen pensar con mucha certidumbre que el producto evaluado no presenta dificultades para el sistema inmune. Debemos comentar además que no se encontraron cambios fuera de los niveles considerados como normales para esta línea por Alemán et al., (1991,1998) respecto a los parámetros químicos y hematológicos, mientras que la falta de existencia de las alteraciones entre la hematología, el peso de los órganos linfoides y su correspondiente histología, permiten plantear la inocuidad de la vacuna. Desde el punto de vista anatomopatológico las formaciones granulomatosas macrofágicas observadas en las ratas inoculadas con la vacuna y los animales controles que recibieron hidróxido de aluminio, han sido reportadas como consecuencia de la utilización de los adyuvantes empleados en diferentes vacunas (Hidróxido de Aluminio, Glucanos, ACF, AIF, Aceite mineral entre otros) (Azuma, 1993). El modo de actuar es la generación de un depósito en el sitio de la inoculación, el cual permite la lenta liberación del antígeno y prolonga el tiempo de interacción de este con las células presentadoras de antígenos (CPA) y los linfocitos. De aquí la necesidad de que las CPA capten el antígeno y que el mismo sea acarreado por las referidas células, hasta los ganglios linfáticos regionales, donde ocurren entre otros eventos la presentación y proliferación de los linfocitos T y B (Nathan, 1992), (Rivero et al., 1993). Por otra parte los compuestos de aluminio activan la vía clásica del complemento provocando una respuesta inflamatoria local, importante en el desarrollo de las células B de memorias e inducen respuesta Th2. Estas han sido descritas en estudios toxicológicos llevados a cabo en otras vacunas que contienen hidróxido de aluminio como adyuvantes, cuya presencia dentro del granuloma se demostró en trabajos previos mediante una coloración especial para detectar aluminio. Las lesiones consideradas incidentales observadas tanto en controles como en los animales de los grupos experimentales, no fueron considerados con valor diagnóstico, entre ellas los depósitos de hemosiderina a nivel del parénquima esplénico y las urolitiasis, se consideran normales en esta especie y edad. Las lesiones observadas en el punto de inoculación por nosotros, también han sido descritas en otras especies de animales en que se han empleado vacunas preparadas con adyuvantes oleosos (Anónima, 1988). En nuestro estudio, el adyuvante empleado fue el gel de AL(OH)3. Consideramos que éste, con su efecto de depósito, provoca las alteraciones granulomatosas anteriormente mencionadas las cuales tienden a desaparecer después de los 8-9 meses según los estudio conducidos por nosotros en ratones. Estas formaciones de granulomas también se han observado en tejidos humanos y de animales con otras vacunas absorbidas en gel AL(OH)3 (Gupta et al., 1993). Aunque en pequeño número y vinculado principalmente al macho, las lesiones granulomatosas se demostraron en la vacuna de tétanos-difteria adsorbida con este adyuvante cuando se llevaron a cabo inoculaciones intramuscularmente en la especie ratón (Posada 1995). No obstante, al resultar una incógnita para nosotros el comportamiento del comportamiento de este adyuvante en la vacuna, nos propusimos realizar un ensayo en ratones inoculados con VA- MENGOC- BC por la vía intramuscular relacionando la presencia del granuloma con los niveles de anticuerpos específicos antimeningocócicos, que evolucionaron de la siguiente forma: Hacia el decimosexto día, comienzan a incrementarse los títulos de anticuerpos en los ratones inoculados por vía intramuscular, se produce una hiperplasia paracortical en el ámbito del ganglio linfático regional correspondiente a la formación granulomatosa. Sin embargo, hacia los días 82-85 comienzan a disminuir los niveles más altos de anticuerpos. El granuloma aunque persistió se hizo más discreto con presencia de macrófagos y escasos neutrófilos, para desaparecer finalmente al ser sustituido por tejido conjuntivo. Por todo lo demostrado consideramos que la existencia del proceso histopatológico y su evolución, caracterizada a través de una reparación con sustitución paulatina del granuloma macrofágico por el referido tejido conjuntivo, constituye un evento de singular importancia en la respuesta inmune. Quedo comprobado, además la relación existente entre la respuesta celular representada por el referido granuloma y la humoral dada por los títulos de anticuerpos específicos demostrados a través de la técnica de ELISA en los animales estudiados. Luego de lograr la reproducción experimental en murinos (rata y ratón) y conocer los aspectos que cada uno de ellos son capaces de reproducir más fielmente a la infección producida por NmB, se demostró la eficacia de la vacuna VA-MENGOC-BC® y de Inmunoglobulina Antimeningocócica BC frente diferentes cepas, serogrupos, serotipo subtipo e inmunotipos de Nm, partiendo de estas premisas comprobamos la inocuidad de la referida vacuna en rata y ratones, todo lo cual demostró que lo planteado en nuestra hipótesis. CONCLUSIONES FINALES - Quedaron establecidos los biomodelos murinos, para reproducir la infección causada por NmB. - Se determinó que la vacuna VA-MENGOC-BC® fue eficaz para contrarrestar la infección en los modelos murinos. - Se comprobó que la vacuna VA-MENGOC-BC® es inocua. Todo lo anterior nos permite defender como tesis nuestro planteamiento de la hipótesis. RECOMENDACIONES - Incluir estos modelos dentro de los ensayos de control de calidad de VA-MENGOCBC® y realizarlos con la frecuencia que se considere necesaria, después de efectuar los ensayos de reproducivilidad, estandarización y validación, constituyendo este un parámetro más de consistencia para la referida vacuna, así como instrumento de evaluación de nuevos candidatos vacunales y combinaciones en el futuro. - Realizar todos los ensayos de reproducivilidad y estandarización de todos los ensayos para que formen parte de los métodos propuestos para comprobar la eficacia de VAMENGOC-BC® e incluyendo la Inmunoglobulina Antimeningocóccica BC, sancionados por la Dirección de Control de la Calidad. - Realizar los estudios inmunológicos que fundamenten las reacciones cruzadas de protección de VA-MENGOC-BC® frente a cepas no incluidas en esta vacuna y que pudieran ser las bases de la predicción de un más amplio espectro protector. - Aplicar las experiencias obtenidas en los biomodelos por Nm en otros gérmenes bacterianos productores de meningitis en los aspectos de reproducción de los procesos patológicos causados por estos. - Transferir la tecnología para comprobar la eficacia de VA-MENGOC-BC® y de la Inmunoglobulina Antimeningocóccica BC a otras vacunas y sueros de características similares. - Completar los estudios a nivel del punto de inoculación de VA-MENGOC-BC® aplicando técnicas de inmunohistoquímica y citometría de flujo, perfil de citoquinas en tejidos. Varias de estas recomendaciones en el momento de defender estas tesis ya han sido o están siendo implementados. Referencias Bibliográficas Referata 1. Abbas KA. Inmunidad frente a los microorganismo. En: Abbas KA, Lichtman HA, Pober JS. Inmunología Celular y Molecular. 2 da edición. Madrid: McGraw Hill Interamericana; 1995:369-376. 2. Alemán CL, Más RM, Rodeiro I, Noa M, Henández C, Menéndez R, Gámez R: Refeence database of the main physilogical parameters in Spreague Dawley rats from 6 to 32 Months. Lab. Animals. 1998; 32:457-466. 3. Azuma I. Potentiation of host defence machanisms against infection by a cytokine inducen and acyl-MDP-Lys (48) in mice and human. Med. Res. Rev. 1993; 14(4): 401414. 4. Brandtzaeg P. Clinical aspects of Neisseria meningitidis infection. (editores )Proc IX Intern Pathog Neisseria Conf. Winchester. England: (Editorial); 1994:234-5. 5. Bhatnagar NB, Frasch YCA. Expression of Neisseria meningitidis iron-regulate outer membrane protein including a 70-Kilodalton transferrin receptor, and their potential for use as vaccines. Infect. Immun. 1990; 58: 2875-2881. 6. Brandtzaeg P. Clinical aspects of Neisseria meningitidis infection. In: Evans JS, Yost SE, Maiden MCJ, Feaver IM. Proceedings of the IX International Pathogenicc Neisseria Conference, September 26-30, Winchester. England: SCC, Pasteur Merieux Company; 1994: 234. 7. Brodeur BR, Tsang PS. Mouse models of infection from Neisseria meningitidis B, 2b and Haemophylus influenzae type b diseases. Can J. Microbiol. 1986; 32:33-37. 8. Buchanan RM, Arulanandam BP, Metzger DW. IL-12 enhances antibody responses to T-independent polysaccharide vaccines in the absence of T and NK cells. J Immunol. 1998; 161(10): 5525-33. 9. Casey HW, Ayers KM, Robinson FR. The urinary sistem. In: Benirschk K, Garner FM, Jones TC, Eds. Pathology of laboratory animals. Vol. 1. chap. 3. New York: SpringerVerlag; 1978:115-173. 10.Castillo F. El ratón como animal de Laboratorio. La Habana: Ed. Ciencias Médicas; 1985. 11.Clapp MJL. Effect of diet on some paraeters measured in toxicological studies in the aging rat. Lab. Anim. 1980; 14:253-261. 12.Cordy DR. Pathomorphology and pathogenesis of bacterial meningoventriculitis of neonatal ungulates. Vet. Pathol. 1984; 21: 587-591. 13.Council on Animal Care.Laboratory Mice In: Canadian Council on Animal Care. Guide to the care and use of experimental animals. Vol. 1. Canada: Canadian Council on Animal Care; 1980:79-106 Council on Animal Care.Laboratory Mice In: Canadian Council on Animal Care. Guide to the care and use of experimental animals. Vol. 1. Canada: Canadian Council on Animal Care; 1980:79-106 14.Danve B, Lissolo L, Mignon M, Dumas P, Colombani PP, Colombani AB, at al. Tranferrin - binding protein isolated from Neisseria meningitidis protective and bactericida antibodies. Laboratory Animals Vaccine. 1993; 11: 1214- 1220. 15.Deng SQ, Yee RB and Zhang HC. Tree new- serogroups of Neisseria meningitidis. J. Biol.Scand. . 1998; 1:307-315. 16.Echeverrg M, Malberty JA, et al. Respuesta inmune humoral a las proteínas de una vacuna antimeningocóccica BC® ensayo realizado en Antioquia, Colombia. Bol. Oficina Sanit. Panam. 1995; 118 (4) 295-301. 17.Favorova LA, Teleshevskaya EA, Trochina LD, Ershova GA, Salnicova GP. Epidemiology and Prophilaxis of Meningococcal Infection. Sov. Med. 1995; 11: 113118. 18.Fernández de Cossio M F, Dhlin M, Selander B, Cruz S, Del Valle J, Burrebaeck CA. Human monoclonal antibodies against an epitope on the class 5c outer membrane protein common to many patogenic strain of Nesseria meningitidis . J. Infect .Dis. 1992; 166: 1322-1328. 19.Ferrieri, P.; Burke, B.; Nelson, J. Production of bacteremia and meningitis in infant rats with group B Streptococcal serotypes. Infect. Immun. 1980; 27: 1023-1032. 20.Frasch CE, Tsai CM. Mocca LF. Outer membrane proteins of Neisseria Meningitidis: Structure and importance in meningococcal disease. Clin. Invest. Med. 1986; 9:101-107. 21.Frasch CE. Immunization against Neisseria meningitidis. In: Easmon C and Jelzaszewies J, eds. Medical Microbiology. London: Academic Press; 1993:115-144. 22.Frasch CE. Serogroup and serotype classification of bacterial pathogens. In: Methods en Enzymology. New York: Academic Press Inc; 1994: 159-174 23.Galguera DM, Sierra GG, Campa HC. Aplicación terapéutica de Gammaglobulina Hiperinmune contra la enfermedad meningocóccica B en niños. (editores). In: Achtman M, Kohl P, Marchal C, Morelli G, Seiler A, Thiesen B. Proceedings of the Seventh International Pathogenic Neisseria conference, Neisseria 1990, September 9-14; West Berlin, Germany. Berlin- New York: Walter de Gruyter Co; 1991: 679-683. 24.Gerard NP, Gerard C. The chemotactic receptor for C5 a anaphylataxi. Nature 198; 23440 1991. 25.González S, Leinonen M. Therapeutic efficacy of monoclonal antibodies to class2 protein against meningococcal infant rat. New York: Walter de Grister; 1991. 26.Granoff DM, Nankervis GA Circulating capsular antigen in infant rats infected with Haemophilus influenzae type b. J. Infect. Dis. 1977; 136: 292-296. 27.Griffiths, E. Enviromental regulation of bacterial virulence-implications for vaccine design and production. (UK): Elsevier Science Publisch. LTD; 1991: 309-315 28.Hamel J, Brodeur BR, Larose Y, Tsang PS, Belmaaza B, Montplaisir S. A monoclonal antibodies directed against a serotype-specific, outer-membrane protein of Haemophilus influenzae type b. J. Med. Microbiol. 1987; 23: 163-170.. 29.Holland J, Langford RP, Towner JK, Williams P. Evidence for “in vivo” expression of Transferrin- Binding Proteins in Haemophilus influenzae type b. Infect. Immun. 1992; 60: 2986-2991. 30.Hudson MD. Bacterial meningitis: a case study and review, J. Am. Anim Hosp Assoc. 1976; 12: 88-91. 31.Huet M, Andrée Suire. Un modele animal pour l´acyivité des vaccins polysidiques méningococciques. J Biol Standarization. 1981; 67-74. 32.Infante JF, Pérez P, Sifontes S, Izquierdo L, Fariñas M, Sierra G, Muñoz E, Malberty M, Caro E, Oliva R. Determinación del esquema de inmunización con VA-MENGOC-BC para pruebas de reto en ratones. VacciMonitor. 1998; 7(8):7-11 . 33.Infante JF, Sierra GV, Campa HC, Acosta A, Azahares E, Pérez V, Muñoz E, et al. Las Ratas SPF como modelo experimental para la enfermedad meningocóccica y la evaluación de la efectividad de VA-MENGOC-BC. II. Estudio microbiológico e inmunológico. VacciMonitor. 1997; 6(7):2-6. 34.Infante JF, Muñoz E, Medina R, Oliva R: Inocuidad inespecífica realizada al Beta (1-3) glucano (EVIMUNK) de fabricación cubana. Rev. Salud Anim. 1995; 17: 45-54. 35.Kolsto Anne-Brit, Rod TO. Iron and Meningococcal disease. Serum Lactoferrin in Meningococcal disease patiens and controls. In: Achtman M, Kohl P, Marchal C, Morelli G, Seiler A, Thiesen B. Proceedings of the Seventh International Pathogenic Neisseria conference, Neisseria 1990, September 9-14; West Berlin, Germany. Berlin- New York: Walter de Gruyter Co; 1991: 413. 36.Laitinen K. Experimental meningococcal meningitis in the infant rat and protective efficacy of antibodies. Helsinki: National Public Health Institute (NPHI); 1988. 37.Lastre M, Batista A, Sierra G, Pérez O. Cuantificación de células formadoras de IgG totales y específicas por ELISPOT en Balb/c durante la respuesta primaria a VAMENGOC-BC. Vacccimonitor. 1995; 4(7):2-3. 38.Lausund P, Louik M. Infection of new born mice with Neisseria meningitidis. In Conde-Glez CJ, Morse S, Rise P, Sparling F, Calderon E, editors. Pathobiology and Immunology of Neisseriaceae. Proceeding of the 8th International Patuologenic Neisseria Conference1992 Oct 4-9Cuernavaca. México. CuernavacaINSP1994: 190. 39.Lee JW. Keynotes Adrres. In, Brounf, aproaches to stabilization of vaccine potency. Rev.Biol. Stand. 1998; 87:3-5. 40.Lynn WA, Wolenbooch DT. Lipopolysaccharide antagonists. Immunol. Today. 1992; (7): 271-7 41.Magnusson G, Ramsay C H. Urolithiasis in the rat. Lab. Anim. 1971, 5; 158-162. 42.Martínez M. Estudio de Waster Blott en sueros de ratones Balb/c inoculados con VAMENGOC-BC. En: Instituto Finlay, Ed. Libro Resumen. Forum de Ciencia y Técnica. La Habana: Instituto Finlay; 1998. 43.Martin D, Cadieux N, Hamel J, Brodeur BR. Highly: Conserved Neisseria meningitidis surface protein with diferens protection against . Experimental infection .J Exp .Med. 1997; 185(7):1173- 1183. 44.McErlean BA. Salmonella dublin meningitis in piglets. Vet. Rec. 1968; 82: 257-258. 45.Mc Neil G, Virji M, Moxon ER. Interactions of Neisseria meningitidis with human monocytes. Microb Pathog. 1994; 16(2):153-63. 46.Moxon ER, Ostrow PT. Haemophilus influenzae meningitis in infant rats: Role of bacteremia in pathogenesis of age- dependent inflamatory responses in cerebrospinal fluid. J. Infect. Dis. 1974; 135: 303- 307. 47.Nathan C. Nitric oxide as a secretory product of mammalian cells. FASEB J. 1992; 6: 3051-3064. 48.Olyhoek AJ M, Sarkari J, Bopp M, Morelli G, Achtman M. Clonig and expression in Escherichia coli of Opc, the gene for an unusual class 5 outer membrane protein from Neisseria meningitidis. Microbiol. Pathog. 1991; 11: 249-257. 49.Pérez -Martín O, Lastre- González M, Díaz- Orellana M, Zayas- Vignier C, Caso R, Hernández I, Sierra- González G. : Biodistribution of the Cuban anti-meningococcal vaccine, VA-MENGOC-BC, in Balb/c mice. Arch. Med. Res. Spring 1997; 28(1): 37-40. 50.Pettersson A, Kuipers A.J, Pelzer M,Verhangen EPM, et al. Monoclonal antibodies against the 70- kilodalton iron- regulated protein of Neisseria meningitidis are bactericidal and strains-specific. Infect. Immun. 1990; 58: 3036-3041. 51.Poolman JT. Clinical trial with outer membrane protein vaccine and Por A recombinant vaccine. 10th International Pathogenic Neisseria Conference. Sept. 8-13 1979; Baltimore Maryland. 1996. 52.Quagliarello V, Scheld WM: Bacterial meningitis: pathogenesis, pathophysiology, and progress. N Engl J Med. 1992; 327: 864-872. 53.Quakyi EK, Hochstein HD, Tsai CM: Modulation of the biological activities of meningococcal endotoxins by association with outer membrane proteins is not inevitably linked to toxicity. Infect Immun. 1997; 65(5): 1972-9. 54.Quakyi EK, Carter PH, Tsai CM, Marti GE. Immunization with meningococcal membrane-bound lipooligosaccharide accelerates granulocyte recovery and enhances lymphocyte proliferation in myelosuppressed mice. Pathobiology. 1997; 65 (1):26-28. 55.Richter GW. Effects of cyclic starvation feeding and splenoctoy on the develoment of hemosiderosis in rat livers. AM. J. Pathol. 1974; 74, 481-506. 56.Rivero- Moreno V, Beddell C and Moncada S. Nitric oxide synthase Structural studies using anti-peptide antibodies. Eur. J. Biochem. 1993; 215: 801-808. 57.Roitt L, Brostaff J, Mole D. Inmunología. Reaccciones inmunítorias mediadas por células, formación de granulomas. Barcelona: Ed. Ciencias Técnicas S.A; 1994. 58.Rubinstein LJ, Garcia-Ojeda PA, Michon F, Jennings HJ, Stein KE: Murine immune responses to Neisseria meningitidis group C capsular polysaccharide and a thymusdependent toxoid conjugate vaccine. Infect Immun. 1998; 66(11): 5450-6. 59.Sacchi CT, Gorla MC, de Lemos AP, Brandileone MC: Considerations on the use of Neisseria meningitidis class 5 proteins as meningococcal BC vaccine components. Vaccine. 1995; 13 (1): 112-8. 60.SAGE. Plan of activities for 1996, including full report of the research an development subgroup session of the meeting of the scientific advisory group of Vaccine Reserch an development (VRD) elicit Global program for vaccines an immuniza. experts June 1999; 12-14. 61. Sandoval C M, Anadon B J M, Delgado J M, Capó M M: Estudio comparativo de toxicidad aguda in vivo e in vitro. Extrapolación al hombre. Animales de Experimentación. 5 (1); 2000, 14-16. 62.Sanford SE. Gross and histopathological findings in unusual lesions caused by Streptococcus suis pig: II- central nervous system lesions, Can. J Vet. Res. 1987; 51: 486-489. 63.a Saukkonen K, Abdillahi H, Poolman JT, Leinonen M. Protective efficacy of monoclonal antibodies to class 1 and class 3 outer membrane proteins of Neisseria meningitidis B:15:P1:.16 in infant rat infection model: new prospects for vaccine development. Microbiol. Pathogen. 1987; 3:261. 64. Saukkonen K, Leinonen M. Infant rat model for meningococcal group B infection. In: (Editores), Proceeding of 5TH Internat Path Neisseria Conf. Noordwijkerhout, the Netherland, 1986. Netherland: (editorial); 1986:14-18. 65.b Saukkonen K, Abdillahi H, Poolman JT, Leinonen M. Protective efficacy of monoclonal antibodies to class 1 and class3 outer membrane proteins of Neisseria meningitidis B: 15: P1: P1.16 in infant rat infection model : new prospects for vaccine development. Microbiol Pathogen. 1987; 3: 261 66.Saukkonenn K, Leinonen M, Abdillahi H, Poolman J T: Comparative evaluation of potential components of group B meningococcal vaccine by pasive protection in the infat rat. J. Infect. Dis. 1988; 158:209. 67.Saunders NB, Shoemaker DR, Brandt BL, Moran EE, Larsen T, Zollinger WD: Inmunogenicity of intranasally administered meningococcal native outer membrane vesicles in mice. Infect Immun. 1999; 67(1): 113-9. 68.Sierra GV, Campa HC and Volcárcel M. Vaccine against group B Neisseria meningitidis: protection trial and mass vaccination. Results in Cuba. NIPH Ann. 1991; 14(2):195-207. 69.Sifontes S, Infante JF, Marrero O, Fariñas M, Muñoz E, López Y. Virulence enhancement agents for Haemophilus influenzae type B infection in mice: Laboratory Animal Science. 1999; 49(1): 95-98. 70.Sifontes S, Infante JF, Capó V, Marrero O, Azahares E, Muñoz E, Sierra G, Campa C. Experimental Infection by Haemophilus influenzae type b in Inbred mice. Arc. Med. Res. 1996; 27(2):133-138. 71.Sotologo. F Neisseria meningitidis: Aspectos Habana: Ed. Finlay; 1996. Serie Monográfica. bacteriológicos e inmunológicos. La 72.Statistica for Windows, Release 4.5, StatSoft, Inc. 1993. 73.Stojiljkovic I, Hwa V, Larson J, Lin L, So M, Nassif X. Cloning and characterization of the Neisseria meningitidis rfaC gene encoding alpha-1,5 heptosyltransferase I. FEMS Microbiol Lett. 1995; 151(1): 41-9. 74.Tunkel AR, Wispelwey B, Scheld WM. Bacterial meningitis: recent advances in pathophysiology and theatment. Ann. Intern. Med. 1990; 112(8):610-623. 75.Verheul AF, Kuipers AJ, Braat AK, Dekker HA, Peeters CC, Snippe H, Poolman JT. Development characterization and biological properties of meningococcal immunotype L 3,7 (8), 9 specific monoclonal antibodies. Clin . Diag . Lab . Immunol. 1994; 1: 729-736. 76.Virji MK, Ferguson DJ, Achtman M, Sarkari J, Moxon ER. Expression of the Opc protein correlates with invasion of epithelial and endothelial cells by Neisseria meningitidis. Mol microbiol. 1992; 6 (19): 2785-95. 77.Vogel U, Hammerschmmid S, Frosch M. Sialic acids of both the capsule and the sialylated lipooligosaccharide of Neisseria meningitis serogroup B are prerequisites for virulence of meningococci in the infant rat. Med Microbiol Immunol (Berl). 1996; 185 (2): 81-7. 78.Ward JM, Reznik-Schuller H. Morphologic and histocheical characteristics of pigments in aging F 344 Rats. Vet. Pathol. 1980; 17, 678-685. 79.Westendorp RG, Langermans JA, Huizinga TW, Elouali AH, Verweij CL, Boomsma DI, Vanderbrougke JP. Genotic influence on cytokine production and fatal menigococcal disease. Lancet. 1997; 349 (9046): 170-3. 80.Westerink MA, Campagnari AA, Giardina P, Apicella MA: Antiidiotype antibodies as surrogates for polysaccharide vaccines. Ann N Y Acad Sci. 1994; 151(730): 209-16. 81.Westerink MA, Giardina PC, Apicella MA, Kieber-Emmons T. Peptide mimicry of the meningococcal group C capsular polysacchride. Proc Natl Acad Sci USA. 1995; 92(9): 4021-5. 82.Westerink MA, Giardina PC. Anti- idiotype induced protection against Neisseria meningitidis serogroup C bacteremia. Microb Pathog. 1992; 12(1):19-26. 83.a Wiliams AE, Blakemore WF. Pathogenesis of meningitis caused by Streptococus sevis type 2. Journ. Infect. Dis. 1990; 162: 474-481. 84.b Williams AE, Blakemore WF. Pathology of streptococcal meningitis following intravenous, intracisternal and natural routes of infection. Neurophath. And Applied Neurobio. 1990; 16: 345-356. Tabla III/A/1: Estudio de protección en ratones Balb/c vacunados con VAMENGOC-BC ® Tratamiento Inóculo N VA-MENGOC-BC® P.B.S NmB* P.B.S 10 10 10 10 Sin vacunar Mortalidad (%) 0 40a 0 100b NmB Se usa Dextrana-Fe como FEV *: 100 x LD 50 ab: P=0.0002 0.5mL, IP, 21 días antes del reto. Tabla III/A/2: Bacteremia en ratones Balb/cj vacunados con VA-MENGOC-BC® Tiempo post-inoculación (h) Grupo 12 24 36 48 72 Vacunados* + + Sin vacunar + + + + + * VA-MENGOC-BC, 0.5 mL, IP, 21 días previos al reto Dosis de reto: 7.4 x 105 UFC de NmB. Tabla III/A/3: Títulos de anticuerpos contra NmB en ratones Balb/cj Tratamiento N Media SD P Vacunados* 15 5149 2022 0.0003 Sin vacunar 15 872 1561 VA-MENGOC-BC® 0.5mL, IP, 21 días previos al muestreo. Tabla III/B/1: Bacteremia en ratas recién nacidas infectadas con NmB No. de animales Inócul Vía o Bacteremia (horas) 24 48 72 96 120 12 1x10 I.P + + + + + 7 10 1x10 I.P + + + * Nota: Se muestrearon en cada tiempo 2 animales. *: Mortalidad del 100% 6 144 + Tabla III/B/2: Estudio histopatológico secuencial del SNC de ratas recién nacidas inoculadas con NmB. Días inoculación 3 días 6 días 9 días post- Lesiones histopatológicas Discreto engrosamiento de la piamadre, congestión de los vasos meníngeos, infiltración de escasas células inflamatorias con predominio de polimorfonucleares neutrófilos Congestión de los vasos meníngeos, el engrosamiento de la piamadre basado en células inflamatorias redondas (linfocitos y macrófagos) y persisten discretos focos de supuración sobre todo al nivel de la aracnoides y focos de gliosis astrocítica en la zona situada por debajo de la piamadre Congestión de los vasos sanguíneos, engrosamiento al nivel de la aracnoides a base de (linfocitos, células plasmáticas y grandes macrófagos fagocitando un material granular); focos de gliosis astrocítica y microglial en la zona situada por debajo de la leptomeninge. Infiltrado mononuclear al nivel de la vaina del nervio olfatorio. Grados I III II y III Tabla IV/M/1. Cepas utilizadas en los ensayos para comprobar la eficacia de VA-MENGOC-BC y de la Inmunoglobulina Antimeningocócica BC y dosis de la confrontación a que fueron sometidos los animales. Grupo Cepa I II III IV V A B:nt P1.10 (Argentina) B B4: P1.15 (Brasilera) C B4: P1.15 (Colombiana) CH No tipable (Chilena) D B4: P1.15 (Cubana) Diseño Experimental. N Subgrupo Para la IgG 8 A1: Controles 8 A2:Ig30 min antes 8 A3:Ig30 min después 10 B1: Controles 10 B2:Ig30 min antes 10 B3:Ig30 min después 10 C1: Controles 10 C2:Ig30 min antes 10 C3:Ig30 min después 10 CH1: Controles 10 CH2:Ig30 min antes 10 CH3:Ig30 min después 10 D1: Controles 10 D3:Ig30 min después Dosis (DL50) 20 20 20 25 25 25 44.5 44.5 44.5 36 36 36 20 20 Tabla IV/M/2 Esquema de inmunización para el ensayo de eficacia de VA-MENGOCBC. Grupo NI 1d 2d día 0 vacunación día 45 vacunación vacunación Tabla IV/A/1. Resultados del reto realizado con NmB en ratas recién nacidas (mortalidad) para evaluar la eficacia de la Inmunoglobulina Antimeningocócica aplicada previamente. No. Animales inmunizados Animales sin inmunizados ab: P= 0.003 Carga (UFC) 1x10 6 1x10 6 16 16 Vía Mort. (%) I.P. I.P. Oa 37.5b Tabla IV/A/2 Resultados del reto realizado con NmB en ratas infantes (bacteremia) evaluando la eficacia de la Inmunoglobulina Antimeningocócica aplicada previamente. No. Carga (UFC) Vía 24 + 6 Bacteremia (horas) 48 + Animales inmunizados 16 1x10 I.P 6 Animales 16 1x10 I.P sin inmunizar Nota: Se extrajo sangre a dos animales por grupo en cada tiempo. 72 + Tabla IV/A/3 Reto con NmB en ratas recién nacidas y posterior tratamiento con Inmunoglobulina Antimeningocócica (mortalidad) No. Ratas tratadas Ratas no tratadas ab: P= 0.004 10 10 Carga (UFC) 1x10 7 1x10 7 Vía Mort. (%) I.P. I.P. Oa 50b Tabla IV/A/4 Efectividad de la Inmunoglobulina Antimeningocócica en ratones con dos dosis diferentes, inyectada 30 y 15 min antes de la prueba de confrontación con la cepa B4:P1.15 de Nm en dosis de 1 x 10 8 UFC Dosis Ig Mortalidad Supervivencia Grupo * (mg) n % n % Control Sin 10 100 0 0a tratamiento 30 min 10 3 30 7 70b 30 min 5 2 20 8 80c 15 min 5 0 0,0 10 100d ab, ac, ad: P<0.001, bc: P=0.303, bd: P=0.030, cd: P=0.068 Tabla IV/A/5 Efectividad de la Inmunoglobulina Antimeningocóccica repetida en ratones. en dosis Tratamiento Secuencia de administración N sobrevivencia (%)** Controles 1d 2d 3d 4d Sin tratamiento 30 min 30 min, 1h 30, 45 min y 1h 30, 45 min, 1 y 17 h 30, 45 min, 1, 2 y 17 h 10 10 10 10 10 0a 60b 60b 70b 90bc Tiempo medio de** sobrevivencia (h) 19.7 h 55,6 h 60,3 h 62,6 h 66,7 h 10 100c 72 h 5d * tiempo posterior a la inoculación de las bacterias. ** superíndices diferentes indican diferencias estadísticas significativas (P 0,05) Tabla IV/B1/1 Protección y Mortalidad en ratones retados con la cepa aislada en Argentina. Grup o Inóculo (LD50) N Mortalida d (%) Protecció n (%) Valores de P NI 37.3 8 100a - ab: 0.036 1d 37.3 9 66.7b 33.3 ac: 0.001 2d 37.3 9 30c 70 bc: 0.060 NI: no inoculados 1d: una dosis 2d: dos dosis Tabla IV/B1/2 Protección y Mortalidad en ratones retados con la cepa aislada en Chile Grup o Inóculo (LD50) N Mortalid ad (%) Protecció n (%) Valores de P NI 32 10 100a - ab: 0.0005 1d 32 10 90b 10 cd: 0.003 2d 32 10 30c 70 NI: no inoculados 1d: una dosis 2d: dos dosis Tabla IV/B1/3 Protección y Mortalidad en ratones retados con la cepa aislada en Colombia. Grupo Inóculo (LD50) N Mortalid ad (%) Protecció n (%) NI 505 10 100a - 1d 505 10 100a 0 2d 505 10 80b 20 Valores de P ab: 0.068 Tabla IV/B1/4 Tiempo medio de vida en ratones retados con la cepa aislada en Colombia N* Grupo SD (h) Valores de P NI 10 19.58a 4.80 ab: 0.0165 1d 10 23.86b 3.35 ac: 0.0264 2d 8 25.65c 7.48 bc: 0.2533 Tabla IV/B1/5 Protección y Mortalidad en ratones retados con la cepa aislada en Cuba. Grupo Inóculo (LD 50) N Mortalid ad (%) Protecció n (%) Valores de P NI 7.8 9 88.8 a - ab: 0.0532 1d 7.8 14 57.1 b 35.7 ac: 0.0008 2d 7.8 8 12.5c 85.9 bc: 0.0202 Tabla IV/B2/1. Mortalidad y tiempo de sobrevivencia de animales tratados con Inmunoglubulina Antimeningocócica BC y retados con la cepa A. (Grupo 1) Subgrupo Tratamiento N Mortalidad (%) Tiempo de sobrevivencia (h) 18.1d 72e 28f Control 8 100a Ig 30 min antes 8 0b Ig 30 min 8 37.5c después Dosis de Reto: 20 DL50 ab: P<0.001, ac: P=0.004, bc: P=0.027). ANOVA (d-e-f: P<0.01). Test de Duncan (d-e: P< 0.01, d-f : P<0.05, e-f: P<0.01). A-1 A-2 A-3 Tabla IV/B2/2 Mortalidad y tiempo de sobrevivencia de animales tratados con Inmunoglubulina Antimeningocócica BC y retados con la cepa B. (Grupo 2) Subgrupo Tratamiento N Mortalidad (%) Tiempo de sobrevivencia (h) 29.5d 42.0e 42.0f Control 10 80a Ig 30 min antes 10 80a Ig 30 min 10 100b después Dosis de reto: 25 DL50, ab: P=0.068; de: P<0.05, df: P<0.05 B-1 B-2 B-3 Tabla IV/B2/3 Mortalidad y tiempo de sobrevivencia de animales tratados con Inmunoglubulina Antimeningocócica BC y retados con la cepa C. (Grupo 3) Subgrupo Tratamiento N Mortalidad Tiempo de (%) sobrevivencia (h) C-1 Control 10 100a 16.5d C-2 Ig 30 min antes 10 60b 35.0c C-3 Ig 30 min después 10 70c 27.3f Dosis de Reto: 44.5 DL50, ab: P= 0.013, ac: P= 0.030, bc: P= 0.320). Tabla IV/B2/4.(Grupo 4) Mortalidad y tiempo de sobrevivencia de animales tratados con Inmunoglubulina Antimeningocócica BC y retados con la cepa CH. Subgrup Tratamiento N Mortalidad Tiempo de o (%) sobrevivencia (h) CH-1 Control 10 100a 18.1c CH-2 Ig 30 min antes 8 80b 26.5d CH-3 Ig 30 min después 10 100a 25.8e Dosis de Reto: 36 DL50. ab: P=0.069, cd: P<0.05, ce: P<0.05, de: P>0.05 Tabla IV/B2/5. (Grupo 5) Mortalidad y tiempo de sobrevivencia de animales tratados con Inmunoglubulina Antimeningocócica BC y retados con la cepa D. Subgrup Tratamiento N Mortalidad Tiempo de o (%) Sobrevivencia (h) a D-3 Control 10 100 20.3c b D-4 Ig 30 min 10 40 25.8d después ab: P>0.05, : cd: P<0.05 Tabla IV/B/6 . Análisis resumen de la protección de VA-MENGOC-BC ® de la Inmunoglobulina Antimeningocócica respecto a la virulencia de las diferentes cepas LatinoamericanasPaís Argentina Chile Colombia Cuba Brasil Vacuna VA MENGOC-BC 1era 2da 33.3 10 0 35.7 No probada Inmunoglobulina Antimeningocócica BC Antes Después 70 0 70 20 20 40 85.9 No probada No probada 0 37.5 0 30 100 0 Sobre vida Valor DL50 + + + + + 3.88 x 10 7 3.16 x 10 7 1.98 x 10 6 1.28 x 10 8 6.24 x 10 6 Gráfico III/A/1: Cinética de mortalidad en los ratones infectados con Nmb en dosis de 10UFC/L % mortaldad Valores de P: ab< 00001 bc< 0.0001 100 80 60 40 20 0 Mortalidad 0-24 24-48 Tiempo (h) 48 Tabla IV/C/1 Protección cruzada de VA-MENGOC-BC® frente a NmA, 15 días después de la última vacunación Grupo Mortalidad Protección Tiempo de sobrevida (h) (%) (%) Contro 60 a 41.9 l 1 dosis 70 b -16.66 33.7 2 dosis 40 c 33.33 56.9 3 dosis 10 d 83.33 66.5 Dosis de reto: 1x107 UFC/mL, ab: P=0.320, ac: P=0.186, ad: P=0.01 Tabla IV/C/2 Protección cruzada de VA-MENGOC-BC® frente a NmA, 21 días después de la última vacunación Grupo Mortalidad Protección (%) Tiempo de sobrevida (%) (h) a Control 50 48 b 1 dosis 0 100 72 b 2 dosis 0 100 72 3 dosis 0b 100 72 7 Dosis de reto:1x 10 UFC/mL, ab: P= 0.0049 Tabla IV/C/3 Comportamiento de la mortalidad en el grupo control en tres de las 6 concentraciones utilizadas para calcular la DL50 de NmB Concentración N Muertos Mortalidad UFC % 5 10 10 3 30 6 10 10 8 80 7 10 10 9 90 DL50=3.33x10 5 UFC/mL, 0.2 mL, IP (límites 95%: 1.91-5.80x105) Tabla IV/C/4 Efectividad de VA-MENGOC-BC® frente a NmB a los 15 días de la última vacunación. Grupo Control 1 dosis 2 dosis 3 dosis Mortalidad (%) 80 a 10 b 0c 0c Protección (%) 87.5 100 100 Tiempo de sobrevida (h) 30.9 66.5 72 72 Dosis de reto: 1x10 6 UFC/mL (3xDL50), ab: P=0.001, ac: P<0.001, bc: P=0.153 Tabla IV/C/5 Efectividad de VA-MENGOC-BC® frente a NmB, a los 21 días de la última vacunación Grupo Mortalidad Protección (%) Tiempo de (%) sobrevida (h) Control 80 a 30.9 1 dosis 0b 100 72 2 dosis 20 c 75 56.2 3 dosis 10 d 87.5 66.2 6 Dosis de reto: 1x10 UFC/mL (3xDL50), ab: P=0.068, ac: P=0.004, ad: P=0.001 Tabla IV/C/6 Efectividad de VA-MENGOC-BC frente a NmC a los 15 días de la última dosis Grupo Mortalidad Protección (%) Tiempo de sobrevida (%) (h) Control 90 a 22.4 1 dosis 40 b 55.6 2 dosis 60 c 33.3 3 dosis 40 b 55.6 ab: P=0.01, ac: P=0.061, bc: P=0.186 51.2 39.6 50.6 Tabla IV/C/7 Grupo Control 1 dosis 2 dosis 3 dosis Efectividad de VA-MENGOC-BC® frente a NmC, 21 días después de la última dosis. Mortalidad Protección (%) Tiempo de sobrevida (%) (h) 50 a 44.5 0b 100 72.0 0b 100 72.0 20 c 60 63.3 ab: P<0.005, bc. P=0.068, ac: P=0.080 Tabla IV/C/8 Protección y tiempo de sobrevida en ratones retados con Nm A, B o C y tratados con inmunoglobulina por vía intraperitoneal (IP) o intravenosa (IV). Serogrup Tratamien Mortalidad Protección (%) Tiempo de sobrevida o to * (%) Sobrevivencia (h)* A Control 90ª 23.4ª IP 80ab 11.1 31.2ab IV 50b 44.4 45.0b B Control 100ª 18.0a b IP 30 70 57.0b IV 10b 90 68.9b C Control 100ª 39.8a b IP 50 50 45.4a IV 0c 100 72.0b * Superíndices distintos indican diferencias significativas para P<0.05 Mortalidad (%) Curvas dosis mortalida d (Regresió n lineal) DL50 Brasil 6.24x106 Chile 3.16x107 Colombia 1.98x106 Cuba 1.28x108 Arg. :3.88x107 Log (dosis) Gráfico No. IV/B/1 Curva DosisMortalidad (regresión lineal) Tabla V/A/ 1: Valores hematológicos (Media y D.S)de las variables analizadas en las ratas SPRD hembras VARIABLES Grupo I Grupo II Grupo III Grupo IV Grupo V (*) a,d a,d b,c Hb (g/ dl ) 11.7 12.9 0.55 13.9 1.20 11.2 0.34 12.1 1.01 0.40b,c Hto ( % ) 38.8 0.83 c 40.8 1.64 42.0 2.82 a,d 37.6 1.67a,c 38.7 3.83 3 Leuco (10 /l) 7.4 4.81 6.97 4.02 6.2 3.29 11.2 7.37 8.0 2.81 Bas ( % ) 0.4 0.54 0.0 0.0 0.2 0.44 0.3 0.48 Eos ( % ) 1.8 1.09 d 0.6 0.89 1.2 0.44d 0.0 a,c 0.4 0.51 Neut ( % ) 15.4 5.27 19.0 13.83 24.0 9.92 19.0 11.55 17.6 4.67 Linf ( % ) 82.2 5.35 79.8 13.75 73.6 10.08 80.2 10.84 80.8 5.20 Mono ( % ) 0.4 0.54 0.6 1.34 1.2 0.44 0.6 0.54 0.9 1.10 Retic. ( % ) 1.15 0.25 1.16 0.40 1.15 0.25 1.02 0.37 0.87 0.14 (*) a los 30 días Diferencia significativa (p 0.05). a: grupo I; b: grupo II; c: grupo III; d: grupo IV; e: grupo V. Tabla V/A/2. Valores hematológicos (Media ±. D.S) en las variables analizadas de las ratas SPRD machos. Variables Grupo I Grupo II Grupo III Grupo IV Grupo V (*) d b.e d Hb (g/ dl) 11.6 0.75 13.2 1.18 12.4 1.40 10.1 1.95 13.5 1.14 e d Hto ( % ) 41.0 2.07 42.4 3.13 39.8 7.25 36.4 6.26 44.4 2.45 3 Leuco (10 /l) 11.2 9.33 9.1 1.88 10.0 1.27 5.8 3.96 5.6 1.14 Bas ( % ) 0.4 0.54 0.2 0.44 0.2 0.44 0.2 0.44 0.1 0.31 Eos ( % ) 1.2 0.83 1.4 0.89 0.2 0.44 0.4 0.54 0.3 0.48 Neut ( % ) 10.4 5.41 12 5.70 15 3.31 22.8 10.42 18.1 6.20 Linf ( % ) 87.6 5.98 85.4 6.52 83.4 4.82 76 11.9 80.5 6.22 Mono ( % ) 0.6 0.89 1 0.70 1.2 1.64 0.6 0.89 1 0.65 Retic. ( % ) 1.14 0.80 0.9 0.53 1.16 0.31 1.12 0.38 0.76 0.32 (*) a los 30 días. Diferencia significativa (p 0.05). a: grupo I; b: grupo II; c: grupo III; d: grupo IV; e: grupo V. Tabla V/A/ 3: Valores de la química sanguínea (Media ±. D.S) en ratas hembras SPRD. Variables Grupo I Grupo II Grupo III Grupo IV Grupo V (*) Colinesterasa (U/l) TGO (U/l) TGP (U/l) Fosfatasa Alc Bilirrubina indirecta Bilirrubina directa Proteínas totales (mg/dl) Urea - nitrógeno Creatinina. (mg/dl) Triglicéridos (mmol/l) Glucosa (mmol/l) Calcio (mg/dl) 175.7 48.8 c 226.8 65.2 b,c 56.6 7.6 39.0 15.5 1.26 1.2 0.16 0.30 70.4 6.95 b,c 262.6 99.5 165.0 25.5 a,d 37.8 5.6 60.4 17.6 0.76 0.4 0.0 56.0 3.54 a,d 292.0 83.3 a,d 113.8 19.8 a 38.3 4.9 54.8 26.6 1.22 0.7 0.02 0.044 57.6 4.85 a,d 245.6 108.2 c 297.2128.9 b 117.4 93.6 47.2 22.1 0.36 0.8 0.0 74.5 7.94 b,c 957.6 672.8 200.0 56.3 52.1 9.0 d 92.8 35.2 0.41 0.3 0.0 61.5 3.57 6.88 7.82 b,c 70.4 15.2 1.19 0.21 6.08 0.88 a,d 48.0 4.69 0.96 0.66 6.36 1.16 a,d 47.0 5.33 0.69 0.13 8.06 2.22 b,c,e 70.8 13.55 1.22 0.37 6.12 0.47 d 48.0 5.67 1.30 0.46 5.8 3.07 2.98 0.43 7.3 1.46 2.45 0.31 6.5 0.39 2.68 0.20 5.7 2.95 e 2.71 0.06 7.0 0.94 d 2.39 0.22 e d (*) a los 30 días Diferencia significativa (p 0.05). a: grupo I; b: grupo II; c: grupo III; d: grupo IV; e: grupo V. A/4: Valores químicos sanguíneos (Media ± D.S). en ratas machos SPRD. Variables Colinesterasa (U/l) TGO (U/l) TGP (U/l) Fosfatasa Alc. Bilirrubina indirecta Bilirrubina directa Proteínas totales (mg/dl) Urea - nitrógeno Creatinina. (mg/dl) Triglicérido (mmol/l) Glucosa (mmol/l) Calcio (mg/dl) Grupo I Grupo II Grupo III Grupo IV Grupo V (*) 262.6 58.90 258.4 65.22 188 51.69 1028.5871.08 240.6 64.62 143.6 33.20 224.6 57.33 212.3 39.25 63 6.08 109.2 19.38 54.2 8.25 119.2 39.42 66.6 16.9 97.8 21.99 e 43.2 4.15 86.5 21.07 0.8 1.47 0.26 0.58 240.6 45.97 173.2 46.73 50.2 7.04 101.2 23.01 0.86 0.94 0.56 1.25 0.16 0.30 0.06 0.13 63.0 3.33 0.0 61.1 2.67 0.0 59.7 4.99 0.02 0.04 61.6 2.60 0.0 61.5 4.31 7.2 0.38 d 63.6 22.64 7.0 0.47 c 51.4 8.23 5.9 0.47 b 48.2 4.38 6.4 0.45 a 57.8 7.85e 6.8 1.39 48.4 6.39d 0.96 0.33 1.15 0.23 1.03 0.44 1.09 0.31 0.93 0.37 6.0 3.74 2.58 0.05 9.1 1.84 2.71 0.07 7.2 2.33 2.60 0.19 5.5 1.42 e 2.57 8.64 6.6 0.42 d 2.35 0.08 d (*) a los 30 días Diferencia significativa (p 0.05). a: grupo I; b: grupo II; c: grupo III; d: grupo IV; e: grupo 14000 12625 12952 12000 11038 hembras machos U/ml 10000 7886 8000 7267 6812 6363 6000 4000 2000 195 0 1 64 n.a. 2 3 4 5 Grupos Figura V/A/4 títulos de anticuerpos frente N. meningitidis de los diferentes grupos de ratas Tabla V/A/5. Pesos de los órganos (Media ± D.S) de las ratas hembras SPRD. Organos Grupo II Grupo III Grupo IV Grupo I Grupo V (*) Corazón 0.77 0.07 0.80 0.07 0.79 0.07 0.83 0.06 0.79 0.06 d d a,b Pulmón izquierdo 0.31 0.02 0.31 0.04 0.37 0.02 0.43 0.09 0.38 0.14 d d d ‡ d Pulmón derecho 0.59 0.04 0.59 0.10 0.61 0.09 0.79 0.69 0.62 0.08 d d b,c Timo 0.32 0.09 0.32 0.05 0.28 0.07 0.43 0.06 0.34 0.06 d c,e d Hígado 7.56 0.80 7.84 1.04 7.55 0.64 8.76 0.64 7.96 0.62 c b Bazo 0.53 0.06 0.48 0.04 0.60 0.05 0.59 0.12 0.49 0.06 Riñón izquierdo 0.70 0.04 0.73 0.06 0.68 0.04d 0.80 0.08 c 0.67 0.14 Riñón derecho 0.71 0.04 0.75 0.07 0.66 0.05 0.79 0.14 0.72 0.06 (*) a los 30 días Diferencia significativa (p 0.05). a: grupo I; b: grupo II; c: grupo III; d: grupo IV; e: grupo V. Tabla V/A/6 Pesos de los órganos en gramos (Media ± D.S) de las ratas machos SPRD. Organos Grupo I Grupo II Grupo III Grupo IV Corazón 1.11 0.11 1.14 0.06 1.12 1.14 1.12 0.08 Pulmón izquierdo 0.46 0.11 0.39 0.03 0.40 0.03 0.45 0.03 Pulmón derecho 0.67 0.14 0.73 0.03 0.80 0.08 0.83 0.12 Timo 0.34 0.05 0.43 0.09 0.38 0.06 0.34 0.05 Hígado 10.53 0.67 11.36 1.08 11.22 1.04 11.01 1.91 Bazo 0.69 0.12 0.68 0.06 0.77 0.05 0.68 0.06 Riñón izquierdo 1.12 0.09 1.15 0.05 1.16 0.07 1.19 0.12 Riñón derecho 1.13 0.08 1.18 0.06 1.18 0.09 1.21 0.12 Grupo V (*) 1.12 0.11 0.41 0.09 0.71 0.08 0.43 0.09 10.43 1.58 0.66 0.06 1.15 0.12 1.17 0.12 (*) a los 30 días . Diferencia significativa (p 0.05). a: grupo I; b: grupo II; c: grupo III; d: grupo IV; e: grupo V. Tabla V/A/7: Relación entre el peso de los órganos linfoide, la histología y algunos parámetros hematológicos (Media ± D:S) en ratas hembras SPRD. Variables Grupo I Grupo II Grupo III Grupo IV Grupo V () Bazo (g) 0.53 0.06 0.48 0.04 0.60 0.05 0.59 0.12 0.49 0.06 Timo (g) 0.32 0.09 0.32 0.05 0.28 0.07 0.43 0.06 0.34 0.06 Histología Normal Normal Normal Normal Normal Hb (g/dl) 11.7 0.40 12.9 0.55 13.9 1.20 11.2 0.34 12.1 1.01 Hto ( % ) 38.8 0.83 40.8 1.64 42.0 2.82 37.6 1.67 38.7 3.83 Leuco ( % ) 7.4 4.81 6.97 4.02 6.2 3.29 11.2 7.37 8.0 2.81 () a los 30 días Tabla VA/8. Relación entre el peso de los órganos linfoide, la histología y algunos parámetros hematológicos(Media ± D:S) en ratas machos SPRD. Variables Bazo(g) Timo (g) Histología Hb (g/dl) Hto ( % ) Leuco ( % ) Grupo I Grupo II Grupo III Grupo IV 0.69 0.12 0.34 0.05 Normal 11.6 0.75 41.0 2.07 11.2 9.33 0.68 0.06 0.43 0.09 Normal 13.2 1.18 42.4 3.13 9.1 1.88 0.77 0.05 0.38 0.06 Normal 12.4 1.40 39.8 7.25 10.0 1.27 0.68 0.06 0.34 0.05 Normal 10.1 1.95 36.4 6.26 5.8 3.96 Grupo V () 0660.06 0.43 0.09 Normal 13.5 1.14 44.4 2.45 5.6 1.14 () a los 30 días Tabla V/A/9: Alteraciones histopatológicas observadas en los animales tratados con diferentes dosis de VA- MENGOC- BC a en ambos sexos. TIPO DE GRUPO I GRUPO II GRUPO III GRUPO IV GRUPO V (*) LESIONES H M T H M H M T H M T H M T T Urolitiasis renal 5/5 2/5 7 4/5 2/5 6 5/5 4/5 9 4/5 1/5 5 8/1 3/ 11 tubular 70 60 90 % 50 % 1 55 % (moderada) % % Hemosiderosis 1/5 0/5 1 1/5 0/5 1 2/5 2/5 4 2/5 1/5 3 3/1 1/ 4 del Bazo (mild) 10 10 40 % 30 % 1 20 % % % Granuloma 3/5 0/5 3 3/5 0/5 3 3/5 3/5 6 2/5 3/5 5 4/1 4/ 8 macrofagico a 30 30 60 % 50 % 1 40 % nivel del % % músculo (*) a los 30 días, H: Hembras, M: Machos, T: Total Gramos 300 250 200 150 100 Grupo I Grupo II Grupo III Grupo IV 50 0 1 2 3 4 5 6 7 8 Semanas Figura V/A/1Comportamiento del peso corporal correspondiente a los grupos I, II, III y IV de las ratas SPRD hembras G ramos 400 350 300 250 200 150 G rupo I 100 G rupo II G rupo III G rupo IV 50 0 1 2 3 4 5 6 7 8 Semanas Figura V/A/2. Comportamiento del peso corporal correspondiente a los grupos I, II, III y IV de las ratas SPRD machos Gramos 400 350 300 250 200 Figura . V/A/3 Comportamiento del peso corporal en los grupos IV y V en las ratas 150 SPRD hembras 100 50 Grupo IV Grupo V 0 1 2 3 4 5 Semanas 6 7 8 Gráfico V/B/1. Resultados del estudio serológico en ratones Balb/c inoculados con VA-MENGOC-BC por vía intramuscular. Figura 1. Aspecto clínico de los ratones Balb/cj luego de ser infectados con Neisseria meningitides serogrupo B, previa aplicaciòn de FEV (Dextrana - Fc), inóculo bacteriano 108 UFC/ml 12h Post infecciòn Figura 2. Inflamaciòn de la meninges a nivel del lóbulo frontal de un ratòn Balb/cj inoculado con Neisseria meningitidis grupo B, 3 días post-inoculaciòn. Figura 3. Leptomeninges y focos de gliosis a nivel de la corteza cerebral de un ratón Balb/cj a los 4 días de haber sido infectado con Neisseria meningitidis serogrupo B. H-E x 100. Figura 4. Extenso infiltrado a predominio de polimorfonucleares neutrófilosa a nivel de la meninge aracnoides en un ratón Balb/cj infectado por Neisseria meningitidis serogrupo B 3 dias póst-infección. H.E. x 200. . Figura 1.Comportamiento del peso corporal correspondiente a los grupos I, II, III y IV de las ratas SPRD hembras gramos 300 250 200 150 100 Grupo I Grupo II Grupo III Grupo IV 50 0 1 2 3 4 semanas 5 6 7 8 Figura 2. Comportamiento del peso corporal correspondiente a los grupos I, II, III y IV de las ratas SPRD machos gramos 400grammes 300 350 250 300 200 250 150200 150 100 Grupo I 100 Group IV Grupo II Group V Grupo III Grupo IV 50 50 0 0 1 1 2 2 3 3 4 4 weeks semanas 5 5 6 6 7 7 8 8 . gramos 400 350 300 250 200 150 100 Grupo IV 50 Grupo V 0 1 2 3 4 5 6 7 semanas Figura 3. Comportamiento del peso corporal en los grupos IV y V en las ratas SPRD hembras 8 . gramos 400 350 300 250 200 150 100 Grupo IV 50 Grupo V 0 1 2 3 4 5 6 7 8 semanas Figure 4.Comportamiento del peso corporal en los grupos IV y V en ratas machos SPRD. 14000 12625 12952 12000 11038 hembras 10000 m achos U/ml 7886 7267 8000 6812 6363 6000 4000 2000 195 0 1 64 n.a. 2 3 4 5 Grupos Figura 5 Concentración de IgG anti-VME en muestras de suero de ratas SPRD de ambos sexos. n.a. no representativa g r a mme s 4 0 0 3 5 0 3 0 0 2 5 0 2 0 0 1 5 0 G r upo I V G r upo V 1 0 0 5 0 0 1 2 3 4 5 6 7 8 we e ks Figure 4. Average body weight corresponding to different groups of male SPRD rats. 14000 12625 12952 12000 11038 Female Male 10000 U/ml 7886 7267 8000 6812 6363 6000 4000 2000 0 195 64 1 n.a. 2 3 4 5 Groups Figure 5. Mean concentration of IgG anti-OMP in SPRD rat serum samples. n.a. not available Figura 5. Infiltrado inflamatorio mixto a nivel de las meninges del cerebelo de un ratón Balb/cj infectado por Neisseria meningitidis serogrupo B, 38 horas post-inoculación. H.E. x 120. Figura 11. Focos de gliosis astroglia situado por debajo de las meninges en una rata recién nacida de la línea Sprague Dawley, infectada con Neisseria meningitidis serogrupo B, 4 días post-infección. H.E. x 100. Figura 13. Hiperplasia de los folículos linfoides subcapsulares correspondientes a ganglio linfatico poplíteo del ratón Balb/cj donde se practicó la inoculación con VA-MENGOCBC, 30 días posteriores a la vacunación. Figura 14. Pequeña formación granulomatosa macrofágica en el músculo de un ratón Balb/cj inoculado con VA-MENGOC-BC, 286 días después de la aplicación por vía intramuscular. H.E. x 220. Figura III / A / 6. Infiltrado inflamatorio mixto a nivel de las meninges del cerebelo de un ratón Balb/cj infectado por Neisseria meningitidis serogrupo B, 38 horas póst-inoculación. H.E. x 120. Figura III / B / 5. Focos de gliosis astroglia situado por debajo de las meninges en una rata recién nacida de la línea Sprague Dawley, infectada con Neisseria meningitidis serogrupo B, 4 días post-infección. H.E. x 100. Figura 7. Engrosamiento de las meninges en ratas recién nacidas Sprague Dawley, infectada con Neisseria meningitidis serogrupo B, 5 días post-inoculación. H.E. x 80. Figura 8. Meningitis focal en ratas recién nacidas de la línea Spreague Dawley, infectada con Neisseria meningitidis serogtupo B, 4 días post-inoculación. H.E. x 80 Figura 9. Infiltrado a base de polimorfonucleares neutrófilos en el punto de inoculación, 24 horas después de la inoculación pro vía intramuscular con VA-MENGOC-BC en ratón Balb/cj. H.E. x 80. Figura 10. Infiltración de los músculos por macrófagos y neutrófilos. Necrosis de Zenker a nivel del punto de inoculación con VA-MENGOC-BC en ratón Balb/cj 48 horas de la aplicación por vía intramuscular. H.E. x 160. Figura 11. Abundantes células plasmáticas a nivel del ganglio linfático poplíteo correspondiente a la extremidad donde se realizó la inoculación con VA-MENGOC-BC. H.E. x 120. Figura 13. Aspecto de liquido cefalorraquídeo en un frotis de una rata recién nacida de la línea Spreague Dawley infectada con Neisseria meningitidis serogrupo B, dos días postinfección. Obsérvese la abundancia de células inflamatorias a predominio de neutrófilos. Romanosky-Giemsa x400.