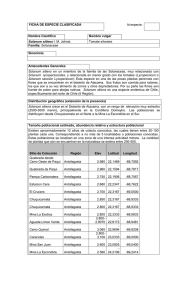
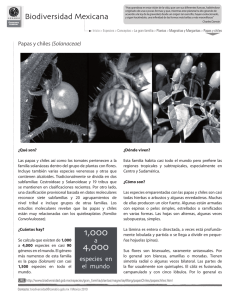
Consideraciones taxonómicas, germinación de semillas y
Anuncio

UNIVERSIDAD NACIONAL DE TUCUMÁN FACULTAD DE AGRONOMÍA Y ZOOTECNIA MAGÍSTER EN DESARROLLO DE ZONAS ARIDAS Y SEMIARIDAS Consideraciones taxonómicas, germinación de semillas y emergencia de plántulas del Complejo Solanum nigrum en la Provincia de Tucumán Ing. Agr. Nora Josefina Mansilla de Andrada San Miguel de Tucumán 12 República Argentina 2009 Consideraciones taxonómicas, germinación de semillas y emergencia de plántulas del Complejo Solanum nigrum en la Provincia de Tucumán 13 Integrantes de la Comisión de Supervisión Director Dr. Ing. Agr. Salvador Chaila Comisión de Supervisión M.Sc. Ing. Agr. María Teresa Sobrero M.Sc. Ing. Agr. Jorge Perera 14 PREFACIO En la producción agrícola siempre se han destacado las Solanáceas por ser un grupo de plantas que brindan enormes beneficios pero también ocasionan grandes daños. Son plantas cosmopolitas de importancia económica positiva por la presencia de especies alimenticias, medicinales, industriales y ornamentales y negativa por la gran cantidad de Solanáceas que se comportan como malezas y que, además, suelen ser tóxicas para el hombre y los animales. La familia Solanáceas, con aproximadamente 96 géneros y unas 2.300 especies, está ampliamente distribuida en las regiones templadas y tropicales de todo el mundo; aunque su mayor diversidad se encuentra en el Hemisferio Sur, particularmente en América Central y Sud América. Las malezas constituyen una población vegetal espontánea que como componentes del agroecosistema deben mantener un equilibrio con el cultivo. El manejo que se realice sobre estos problemas de la producción agrícola apuntará a evitar la disminución de rendimientos con controles de las malezas en el momento oportuno y tratando siempre de conservar el equilibrio del ecosistema. Se ha considerado para este trabajo de tesis a malezas Solanáceas que integran un grupo de especies estrechamente relacionadas, de compleja identificación y ampliamente distribuidas en las regiones de clima cálido. Debido a ello se las ha llamado Complejo Solanum nigrum. Numerosos cultivos presentan interferencias producidas por especies del género Solanum. La interferencia competitiva aparece en cultivos hortícolas, en especial en la producción de verduras de hojas, zanahoria, remolacha, frutilla, papa y en el cultivo de plantas ornamentales de la región. En el Sur de la provincia de Tucumán su presencia fue detectada en tabaco y soja; en la zona Este de la provincia aparece en los cultivos de melón, sandía, maíz, poroto negro y soja. Si bien la densidad no alcanza niveles críticos, aparecen pequeños rodales de hasta 15 plantas/m2. El control no debe efectuarse con 2,4-D porque el género Solanum presenta estadios tolerantes al herbicida hormonal. En citricultura su presencia no reviste peligrosidad ni competencia crítica, aparece en las tazas de las plantas conviniendo efectuar limpieza mecánica y aplicaciones dirigidas de herbicidas en manchoneos. En Tucumán, en caña de azúcar su aparición se limita a la región centro-este y noreste. En este cultivo Solanum nigrum puede alcanzar densidades que obligan a la aplicación de herbicidas relativamente enérgicos como MCPA, Dicamba, Metsulfuron metil y Fluroxipir. En zonas semiáridas de la provincia de Tucumán el empleo del riego determina la aparición de las especies del Complejo Solanum nigrum, especialmente en cultivos hortícolas; pero, su presencia está relacionada a condiciones específicas de manejo. No se registran antecedentes del uso de herbicidas para su control en esta región. La composición cualitativa y cuantitativa de la flora perjudicial de una determinada localidad refleja el sistema de cultivo en uso (Voll et al., 2005). Solanum predomina en suelos con laboreos de rastra y de escarda. Los cambios producidos en la especies presentes pueden variar entre 15 localidades y se refleja en la introducción de nuevas especies de malezas o el aumento de las ya existentes (Debaeke y Sebillotte, 1988). Es necesario relacionar el comportamiento de las especies perjudiciales con las prácticas culturales ejecutadas, principalmente la preparación de suelo, rotación de cultivos, controles químicos y operaciones de cosecha (Fernández Quintanilla, 1988). Esos comportamientos deben estar relacionados con las causas que producen las alteraciones, tales como las condiciones internas de las semillas, el medio ambiente y los factores de manejo (Voll et al., 2005). Para Arévalo (1977) y Chaila (1997) las pérdidas por competencia de malezas con el cultivo dependen: a) Del grado de tecnología agrícola y desarrollo económico alcanzado por los países; b) De la densidad crítica de malezas; c) De factores ecológicos críticos del cultivo; d) De los cultivares; e) De la especie de maleza; f) Del estadio de crecimiento del cultivo; g) Del estadio de crecimiento de la maleza. Kogan (1993) sostiene que el conocimiento de los efectos perjudiciales que las malezas ocasionan es de vital importancia, ya que de ellos resulta la reducción de los rendimientos y consecuentemente la dificultad para producir económicamente. En cambio en nuestra región no existen publicaciones que aclaren los conceptos competitivos específicos de Solanum nigrum con cultivos de importancia. Este trabajo consta de dos partes: Consideraciones taxonómicas Germinación de semillas y emergencia de plántulas del Complejo Solanum nigrum en la provincia de Tucumán. Los capítulos en que se ha dividido son: El primer capítulo trata aspectos botánicos, aspectos agronómicos y la relación con el agroecosistema. Luego se analizan los antecedentes sobre los aspectos taxonómicos de las plantas que integrarían el Complejo Solanum nigrum en la zona agrícola de la provincia de Tucumán. El segundo considera el estudio de la totalidad de los Aspectos Taxonómicos del Complejo Solanum nigrum en la región con el propósito de identificar las especies que lo componen. Se aplican métodos clásicos de identificación que comprenden a: - Caracteres morfológicos - Caracteres anatómicos - Caracteres cariológicos El tercer capítulo considera una etapa de la biología de estas malezas, la germinación y la emergencia de las plántulas. El cuarto estudia aspectos de importancia sobre el Complejo, positivos y negativos. El último enumera conclusiones y recomendaciones. 16 CONSIDERACIONES TAXONÓMICAS, GERMINACIÓN DE SEMILLAS Y EMERGENCIA DE PLÁNTULAS DEL COMPLEJO SOLANUM NIGRUM EN LA PROVINCIA DE TUCUMÁN RESUMEN Las Solanáceas es un grupo de plantas cosmopolitas que brindan enormes beneficios pero que también ocasionan grandes daños. En la Familia Solanáceas, Solanum es el género más grande y complejo con más de 1.500 especies. Una de las especies más estudiadas en el mundo es Solanum nigrum L. conocida como black nightshade en algunos países del Hemisferio Norte, Australia y África, y como yerba mora en los de habla hispana. Se la considera como una de las 76 peores malezas del mundo de importancia en 67 países y en 37 cultivos. Solanum nigrum está estrechamente relacionada a otras especies del género Solanum constituyendo un grupo confuso, con un gran número de especies, de aparente uniformidad, gran plasticidad y variabilidad genética llamado Complejo Solanum nigrum (Solanum nigrum Complex). La composición de especies del Complejo varía en las diferentes áreas de estudio. Con respecto al área agrícola de Tucumán, un grupo de especies estrechamente relacionadas con Solanum nigrum se encontraría constituyendo un complejo semejante al de otras regiones. Dado que de estas especies, en el ámbito regional o provincial no existen publicaciones sobre su taxonomía y tampoco referencias de la importancia como malezas y su biología, los objetivos propuestos en esta tesis fueron seleccionar los caracteres taxonómicos de las especies que componen el Complejo Solanum nigrum en Tucumán con el propósito de identificación con métodos clásicos. Se evaluaron caracteres morfológicos, anatómicos y cariológicos y se realizó el análisis de la germinación de las especies, en forma comparativa. Los caracteres diferenciales de importancia taxonómica que permitieron la identificación de las especies que componen el Complejo son 17 morfológicos, de los cuales los más relevantes son diez (10) carpológicos, cinco (5) florales y dos (2) vegetativos. Como carácter anatómico de diferenciación se observó la presencia de corpúsculos pétreos. Mediante la caracterización citológica se demostró que las poblaciones del Complejo Solanum nigrum estudiadas en Tucumán son diploides, con un número de cromosómico 2n= 24, que concuerda con el número básico x= 12 citado para el género Solanum. Lo que representa una contribución al conocimiento de la citogenética de las malezas del Complejo Solanum nigrum en esta región. Otro aporte que confirma la hipótesis planteada es la identificación de Solanum nigrum, Solanum lorentzii y Solanum nigrum – Intermedia como parte de un Complejo de especies de malezas Solanáceas. Con respecto a la evaluación de los parámetros de germinación y época emergencia de las especies del Complejo Solanum nigrum, en el área agrícola de Tucumán, las diferencias son escasas o nula y la época de emergencia es semejante, lo que no modifica la importancia como malezas. Estos resultados no confirman la hipótesis planteada. 17 TAXONOMIC CONSIDERATIONS, SEED GERMINATION AND SEEDLING EMERGENCE OF THE SOLANUM NIGRUM COMPLEX IN THE PROVINCE OF TUCUMÁN. SUMMARY The Solanaceae constitutes a cosmopolitan group of plants offering important benefits and also causing severe economic damages. The Genus Solanum within the Solanaceae Family is the largest and more complex Genus having over 1500 Species. Solanum nigrum L. is one of the most studied species being known as black nightshade in some countries of the Northern Hemisphere, in Australia and Africa. The common name in the Spanish speaking countries is yerba mora. This plant is considered to be one of the 76 world worst weeds and it is important in 67 countries and 37 crops. Solanum nigrum L. is closely related to other Species of the Genus Solanum constituting a confused group known as Solanum nigrum Complex with a great number of Species apparently uniform and with remarkable plasticity and genetic variability. The Species complex composition varies in the different studied areas. In the agricultural zones of the Province of Tucumán exists a group closely related to Solanum nigrum L. which constitutes a complex probably similar to those ones from some other regions. No papers or studies related to the taxonomy of the complex or its biology and importance as weeds in the Province of Tucumán or in the Argentine NW could be found. The purpose of this contribution is the selection of taxonomic characters of the Species constituting the Solanum nigrum complex in the Province of Tucumán. The task comprises classical practices for morphological, anatomical and cariological character identification. Comparative analysis of germination is also intended. The differential characters of taxonomic importance allowing a correct identification are the 17 morphological ones, the most relevant being ten (10) carpological, five (5) floral and two (2) vegetative. An anatomic character useful for differentiation was the presence of stone cells. It was demonstrated, by means of cytological characterization, that the populations of the Solanum nigrum complex studied in Tucumán are diploid, with a chromosome number 2n=24, matching the basic number x=12 cited for the Genus Solanum. This is a contribution to the knowledge of the cytogenetic of the Solanum nigrum complex weeds in this region. Another contribution confirming the initial hypothesis is the identification of Solanum nigrum, Solanum lorentzii and Solanum nigrum – Intermediate as a part of a weed species complex belonging to the Solanaceae family. The evaluation of germination parameters and emergency moments in the agricultural area of Tucumán Province showed similar values. This fact does not have incidence in their importance as weeds. These results do not confirm the initial hypothesis. 18 ÍNDICE PREFACIO RESUMEN SUMMERY ÍNDICE INTRODUCCIÓN - Hipótesis - Objetivos CAPÍTULO 1 ANTECEDENTES DEL Complejo Solanum nigrum 1.1.- La Maleza 1.1.1.- Aspectos Botánicos 1.1.2.- Aspectos Agronómicos 1.1.3.- Malezas y el Agroecosistema 1.2.- Las Solanáceas 1.2.1.- Aspectos Taxonómicos del Género Solanum 1.2.2.- Aspectos Taxonómicos del Complejo Solanum nigrum 1.2.3.- Las especies del Género Solanum en la República Argentina 1.2.3.1.- Solanum nigrum L. 1.2.3.2.- Solanum americanum Mill 1.2.3.3.- Solanum lorentzii Bitter 1.2.3.4.- Solanum bermejense Bitter 1.2.3.5.- Solanum sublobatum Willd. 1.2.4.- Las especies del Género Solanum en Tucumán ANEXO del CAPÍTULO 1 Nombres de Solanum nigrum L. Descripción botánica de Cabrera, A. L. (1983) del género Solanum Solanum nigrum L Solanum americanum Mill Solanum lorentzii Bitter Solanum sublobatum Willd. Solanum bermejense Bitter Registro de Malezas de la Familia Solanáceas. 1991 - 2001 CAPÍTULO 2 ASPECTOS TAXONÓMICOS DEL Complejo Solanum nigrum 2.1. MATERIALES 2.2. MÉTODOS 2.3. CARACTERES MORFOLÓGICOS 2.3.1. Aspecto Vegetativo 2.3.2. Aspecto Reproductivo 2.4. CARACTERES ANATÓMICOS 2.5. CARACTERIZACIÓN CITOLÓGICA iv v vi vii 1 2 3 4 4 4 4 4 6 7 7 8 8 9 9 9 10 11 12 13 14 19 22 27 34 37 39 39 39 40 40 42 45 46 19 Tabla 2-1: Lista de caracteres 2.6. RESULTADOS y DISCUSIÓN Caracteres Morfológicos Caracteres Vegetativos Caracteres Reproductivos Caracteres Anatómicos Caracteres Citológicos 2.7. ANEXO del CAPÍTULO 2 2.7.1. Altura Bifurcación Del Tallo (AB) 2.7.2.- Longitud (LL), Ancho (AL) y Longitud del Peciolo (LP) de la Hoja 2.7.3.- Pedúnculo (LPD) 2.7.4.- Longitud de Sépalos (en la flor) (KL) 2.7.5.- Longitud del Cáliz (en la flor) (KL) 2.7.6. -Longitud de los Dientes (KLD) (en la flor) 2.7.7.- Longitud de Estambres (ALF)y Ancho del Filamento (AAF) 2.7.8.- Longitud (SL) de las Semillas 2.7.9.- Ancho de las Semillas (SA) CAPÍTULO 3 GERMINACIÓN Y EMERGENCIA DE PLÁNTULAS 3.1. INTRODUCCIÓN 3.2. ANTECEDENTES 3.2.1. La longevidad de las semillas 3.2.2. Las condiciones necesarias para la germinación 3.2.3. Ruptura de la dormancia y época de emergencia 3.2.4. Diferentes situaciones que afectan el crecimiento y la producción de frutos y semillas 3.2.5. Objetivos 3.3. MATERIALES y MÉTODOS 3.3.1. Parámetros 3.3.2. Observación y Recolección del material; Identificación de las especies. 3.3.3. Obtención de semillas pertenecientes a las diferentes especies. 3.3.4. Métodos utilizados en la extracción de las semillas 3.3.4.1. Utilización inmediata 3.3.4.2. Secado de semillas a temperatura ambiente 3.3.4.3. Extracción de semillas de frutos secados a temperatura ambiente 3.3.4.4. Extracción de semillas de frutos conservados en frío hasta 6 meses 3.3.5. Métodos utilizados en la siembra de las semillas de las diferentes especies. Ensayos de germinación 3.3.5.1. En laboratorio 3.3.5.1.1. Siembra inmediata 3.3.5.1.2. Siembra de semillas conservadas 47 50 50 50 61 90 92 93 93 94 96 97 98 99 100 101 102 103 103 103 104 104 104 105 105 106 106 107 107 108 108 108 108 108 108 109 109 110 20 3.3.5.1.2.1. En cajas de Petri a temperatura ambiente, con luz natural 3.3.5.1.2.2. En Germinador 3.3.5.1.3. En estufa, en oscuridad 3.3.5.1.4. Germinación de una especie intermedia. 3.3.5.1.4.1. En tierra 3.3.5.1.4.2. En agua 3.3.5.2. En invernáculo y a la intemperie 3.3.5.2.1. Siembra 3.3.5.2.2. Trasplante 3.3.6. Emergencia en función de la profundidad 3.3.7. Emergencia a campo y en laboratorio 3.3.8. plantación de estacas 3.4. RESULTADOS Y DISCUSIÓN SIEMBRA INMEDIATA En cajas de Petri En recipientes con tierra (macetas y bandejas) En medio líquido Registro fotográfico en medio líquido SIEMBRA DE SEMILLAS CONSERVADAS. Grupos. 1 er grupo: Sin almacenaje 2º grupo: Almacenaje seco Subgrupos A: siembra después de 10 días del lavado. B: siembra después de 15 días del lavado. C: siembra después de 30 días del lavado. D: siembra luego de 90 días del lavado. E: siembra luego de 180 días del lavado. 3er grupo. Almacenaje seco con frío A: siembra después de 10 días de frío. B: siembra después de 15 días de frío. C: siembra después de 30 días de frío. D: siembra luego de 90 días de frío. E: siembra luego de 180 días de frío. 4º grupo. Almacenaje de bayas enteras. (Almacenaje húmedo). SIEMBRAS EN GERMINADOR, con temperaturas constantes y luz alternada Primer grupo A. siembra con semillas frescas (s. inmediata) a 25 º C B. siembra con semillas frescas (s. inmediata) a 30 º C Segundo grupo C. siembra con semillas conservadas a tº ambiente a 25 ºC. D. siembra con semillas conservadas a tº ambiente a 30 ºC. Tercer grupo E. siembra de semillas conservadas en frío a 25ºC. 110 110 111 111 111 111 112 112 112 113 113 113 114 114 114 115 115 117 119 119 119 119 119 119 119 119 119 121 121 121 121 121 121 122 123 123 123 124 124 124 125 125 21 F. siembra de semillas conservadas en frío a 30ºC. Registros fotográficos Otras pruebas de germinación. Ensayos en Oscuridad Germinación de una especie intermedia Siembras en Invernáculo / Intemperie A. Germinación B. Emergencia: Comparación del comportamiento y del desarrollo de las especies del Complejo Solanum nigrum en invernáculo y en el campo. Trasplantes. Emergencia en Función de la Profundidad Parámetros de Germinación Observaciones finales CAPÍTULO 4 OTROS ASPECTOS DE IMPORTANCIA SOBRE EL Complejo Solanum nigrum 4.1. INTRODUCCIÓN 4.2. ANTECEDENTES 4.2.1. Medicinal 4.2.2. Tóxica Importancia para el hombre Importancia agronómica 4.2.3. Forrajera 4.2.4. Alimentación 4.2.5. Reservorio de plagas y/o portadores de resistencias a. Reservorio de plagas b. Portadores de resistencias 4.2.6. Otros usos 4.2.7. Encuestas 4.2.8. Distribución 4.3. ANEXO CAPÍTULO 4 Resultado de la encuesta CAPÍTULO 5 CONCLUSIONES GENERALES BIBLIOGRAFÍA 126 128 130 130 130 131 131 132 133 134 135 136 137 137 137 137 139 139 139 142 143 143 143 150 151 151 152 155 165 173 22 INTRODUCCIÓN Desde los orígenes de la agricultura la productividad de los cultivos se ha visto reducida por la acción de las plagas, enfermedades y malezas. Para tratar de disminuir esas pérdidas, los agricultores han buscado formas y medios de control cada vez más sofisticados. Hoy en día los conocimientos científicos ofrecen nuevas pautas para el control de estos agentes, compatibilizando eficacia, economía y protección ambiental. Las malezas compiten con el cultivo por luz, agua y nutrientes del sustrato. Según Marzocca (1993) los daños que causan las malezas han sido estimados entre un 20 y 30 % del valor de las cosechas, aunque en algunos cultivos, especialmente los intensivos (frutales, hortícolas) estos valores suelen ser ampliamente superados. En la producción agrícola siempre se han destacado las Solanáceas por ser un grupo de plantas que brindan enormes beneficios, pero que también ocasionan grandes daños. En la República Argentina existen 35 géneros de Solanáceas con aproximadamente 400 especies, indígenas o naturalizadas. La mayoría de éstas son del género Solanum (Cabrera, 1983). En Tucumán las Solanáceas están presentes en la totalidad de los departamentos, en sus 10 formaciones vegetales y por su adaptación a distintos hábitats, crecen en diversos niveles altitudinales que oscilan entre los 250 y los 4.300 m.s.m. (Figueroa Romero et al., 2000). En el área agrícola de Tucumán, además de las malezas Solanáceas nativas, se registran especies exóticas que con los años han ido en aumento y que posiblemente no estaban correctamente identificadas. Debido a que estas malezas Solanáceas se encuentran estrechamente relacionadas y su identificación es confusa, se las ha llamado Complejo Solanum nigrum. Las especies del Complejo Solanum nigrum cumplen dentro del ecosistema funciones de malezas y de plantas útiles. Como malezas aparecen en áreas de riego en toda la Región Árida y Semiárida del NOA y producen efectos competitivos en cultivos hortícolas y en otros. Pese a que su presencia no es significativa en abundancia y cobertura producen un importante daño en el crecimiento por utilizar el agua destinada al desarrollo del cultivo. En estas regiones por efecto del riego la maleza produce una abundante cabellera radicular sobre la zona irrigada. Entre los recursos utilizados por los cultivos en la región mencionada, el más importante es el agua, pues el riego permite el manejo de estos y una producción adecuada. Las malezas que interfieran en el consumo del cultivo serán las que se deban eliminar primero. Para interiorizarse sobre las posibilidades de reinfestación en un área dada es necesario conocer los porcentajes de germinación, de emergencia y sobre todo las expectativas de supervivencia de simientes no germinadas o dormantes de las especies más importantes del banco de semillas (Voll et al., 2005). Entre el 5-7 % del total de las semillas germinan y emergen del suelo para competir con el cultivo. Esos valores son variables en función de diversos factores. La periodicidad de emergencia anual se relaciona, primeramente, con la ocurrencia de lluvias, y con el período de aumento de las temperaturas (Stoller y Wax, 1974). 23 Holt y Boose (2000), Chaila (2001), concluyeron que la especie, después de arribar al nuevo hábitat, se reproduce favoreciendo el establecimiento y adaptándose a las condiciones del ambiente. De esta forma favorece la integración con los componentes del sistema natural o agrícola. Todo el proceso, dentro del cual también está el establecimiento, es la invasión de las nuevas especies por un nuevo o distinto hábitat. La dispersión y la diseminación en el nuevo ambiente completarán el mecanismo de invasión. Radosevich y Holt (1984) afirman que la comunidad de malezas en vías de establecerse se va modificando gradualmente a lo largo de un gradiente ambiental, y la vegetación que llega a equilibrarse es la suma de las distribuciones individuales de especies. En un hábitat inestable (Chaila et al., 1997) la habilidad para producir descendientes con alto poder germinativo o protegerlos mediante tácticas adecuadas, es el factor más importante para asegurar la perpetuación en el tiempo. La emergencia de las plantas indeseables a partir del banco de semillas del suelo está influenciada por factores de tipo biológico, físico y ambiental; tales como el estado de dormición de las semillas, humedad, temperatura, luz, fertilidad el suelo, tipo, momento y frecuencia de las labores culturales (Mulugeta y Stoltenberg, 1997). Numerosos y diversos estudios se han realizado sobre el Complejo Solanum nigrum en todo el planeta. Las especies que los componen son diferentes según las zonas y en Argentina no se han encontrado antecedentes sobre el tema. Las malezas constituyen una población vegetal espontánea que como componentes del agroecosistema deben mantener un equilibrio con el cultivo. El manejo que se realice sobre estos problemas de la producción agrícola apunta a evitar la disminución de rendimientos con controles de las malezas, en el momento oportuno, tratando siempre de mantener el equilibrio del ecosistema. Para ello, es necesario tener una identificación precisa y conocer los aspectos más relevantes de su biología. Sobre la base de las anteriores consideraciones se plantea la siguiente hipótesis: Solanum nigrum integra un complejo en el área agrícola de Tucumán con características ecobiológicas que diferencian sus efectos como malezas. Y sobre esta hipótesis se presentan estos planteos o Las especies del Complejo Solanum nigrum tienen importancia en las zonas áridas y semiáridas de la región. o Están presentes en cultivos de la región. o Existen elementos básicos de diferenciación de especies en el complejo. o Hay características morfológicas similares en las especies del complejo. o Algunas de las especies tiene propiedades medicinales. o Tienen cualidades comprobables como forrajeras. o Algunas de las especies son reconocidas popularmente. Por lo que, los objetivos que proponen son: 24 Seleccionar caracteres taxonómicos de las especies que componen el Complejo Solanum nigrum en Tucumán con el propósito de identificación. Analizar comparativamente la germinación del Complejo Solanum nigrum. Estudiar la distribución geográfica de las especies del género que integran el Complejo, mediante encuestas y consultas a personal calificado. Lograr la confección de mapas de distribución del Complejo Solanum nigrum. Establecer los parámetros demográficos de Solanum nigrum para áreas áridas y semiáridas. CAPÍTULO 1 Antecedentes del Complejo Solanum nigrum 1.1.- La Maleza 1.1.1.- Aspectos Botánicos Las malezas son plantas que crecen siempre en forma predominante en situaciones marcadamente alteradas por el hombre, que resultan no deseadas por él en un momento y lugar determinado y producen un daño económico. Son una forma especial de vegetación altamente exitosa en ambientes agrícolas, ambientes perturbados por el hombre, donde crecen sin haber sido sembradas (Marzocca, 1993; Leguizamón, 2003). Para Voll et al., (2005) son plantas que crecen en donde no son deseadas. 1.1.2.- Aspectos Agronómicos Las Solanáceas son plantas que brindan enormes beneficios pero que también ocasionan grandes daños. Son especies cosmopolitas de importancia económica positiva por la presencia de alimenticias, medicinales, industriales y ornamentales y negativa por la gran cantidad de Solanáceas que se comportan como malezas y que, además, suelen ser tóxicas para el hombre y los animales (Cabrera, 1983; Kissman y Groth, 1995). Las malezas compiten con el cultivo por luz, agua y nutrientes del sustrato. El resultado final de la competencia de las malezas es una reducción en el rendimiento, o en la calidad de la cosecha, dos de los más graves problemas de la agricultura. Los perjuicios que causan las malezas han sido estimados entre un 20 y 30% del valor de las cosechas (Marzocca, 1993). Estos valores suelen ser superados en algunos cultivos, principalmente los intensivos (frutales, hortícolas). Para tratar de disminuir esas pérdidas, el hombre constantemente ha buscado diversas formas y medios de control, que en muchos casos han llegado a perjudicar en gran medida al agroecosistema. Hoy en día los conocimientos científicos ofrecen nuevas pautas para el control de las malezas, compatibilizando eficacia, economía y protección ambiental. En general en el banco de semillas de malezas se observa un decrecimiento exponencial del número de semillas viables, de un año a otro, una media de 32 % en suelo cultivado y de 12 % para suelos no cultivados, incluyendo todas las especies. La tasa de decrecimiento varía con el manejo del suelo, o sea, las poblaciones declinan más rápidamente (2-3 %) en suelos cultivados que en suelos no cultivados utilizando siembra convencional (Roberts y Feast, 1972). 25 La supervivencia de un banco de semillas de malezas está en función de las condiciones ecológicas en las que las mismas se encuentran, y de las alteraciones ambientales introducidas en un sistema de producción agrícola (Voll et al., 2005). 1.1.3.- Malezas y el Agroecosistema Altieri (1997) define el agroecosistema como una unidad compuesta por el complejo total de organismos de un área agrícola junto con el ambiente físico externo, en la cual se originan intercambios de flujos de energía e interacciones de ciclos de materiales a través de sus componentes. Las malezas forman parte del agroecosistema y pueden ubicarse dentro de las pioneras de la sucesión secundaria debido a la recurrencia y periodicidad de las perturbaciones. El impacto más crítico de las malezas es el efecto negativo sobre las plantas cultivadas ejercido a través de la competencia por recursos limitados y las alelopatías. Por ejemplo, los trastornos en la recolección y el acondicionamiento de los granos y la disminución de la calidad del forraje constituyen perjuicios adicionales en muchos sistemas (Leguizamón, 2003). Navas (1991) dice que para lograr un manejo sustentable de malezas el enfoque debe cambiar hacia el estudio de las interacciones entre organismos, que permitan conocer las relaciones biológicas, ecológicas y económicas que rigen el funcionamiento de los sistemas productivos. Para Sarandón (2005) un manejo sustentable de malezas (MSM) debe estar basado en una concepción holística y sistémica que permita ver a este componente de manera integral, percibiendo su interrelación con los otros componentes como así también las salidas y entradas de los agroecosistemas que afectan dicho componente. En el marco de los conceptos de agricultura sustentable, el uso racional de los herbicidas es aquel que se efectúa dentro de las oportunidades agrícolas permitidas por el cultivo, en las concentraciones y dosis adecuadas, sin dejar residuos y que garantiza un control muy bueno, satisfactorio o excelente de las malezas, logrando una mejor y mayor producción del cultivo. Se entiende que los rindes logrados no alterarán de ninguna forma el equilibrio del agroecosistema y al mismo tiempo se generará un ciclo de oportunidades laborales favoreciendo el bienestar de la sociedad (Chaila et al., 2006). Existe un replanteo en el manejo de malezas que va más allá de la propuesta productivista de aplicar productos de bajo impacto ambiental o disminuir al máximo el uso de productos de síntesis bajando la posibilidad de aparición de resistencia a herbicidas. Ese replanteo está sujeto a la consideración de otro rol para las especies que son malezas al ser un integrante destacado del agroecosistema (Altieri, 1997). De allí surgirán nuevas estrategias que estarán relacionadas con el comportamiento bioecológico y no solamente con el control. El MSM (Manejo Sustentable de Malezas) tiene como objetivo lograr sistemas donde el control de malezas se realice sólo cuando sea estrictamente necesario considerando a largo plazo todos los efectos ambientales y económicos además de tener presente la repercusión o impacto sobre otros componentes del sistema (Sarandón, 2005; Chaila et al., 2006). 26 Los métodos alternativos comprenden los enfoques de manejo de malezas basados en conocimientos de la semilla y de la planta perjudicial sus flujos de emergencia, su viabilidad, dispersión, estado del banco de semillas, demografía, índices estructurales y dinámica de sus poblaciones, diseminación, invasión, establecimiento, senescencia, controles manuales, mecánicos y MIM (Chaila et al., 2006). El MIM (Manejo Integrado de Malezas) está correlacionado con los programas de manejo del cultivo en especial con los programas MIP (Manejo Integrado de Plagas) y está relacionado con todas las estrategias que apunten a reducir los niveles de infestación de las malezas (cultural, biológico, mecánico o química). En sus concepciones el MIM deberá ser económica, ambiental y socialmente aceptable involucrando el empleo de tácticas biológicas y físicas en forma integrada, justificando el uso de herbicidas solamente por encima de determinado umbral y solamente cuando su empleo sea adecuado desde un punto de vista ambiental, económico y productivo. (Swanton y Weise, 1991; Lyon et al., 1996; Sarandón, 2005; Buhler, 1995; Buhler, 1996, Chaila et al., 2006). 1.2.- Las Solanáceas La familia Solanáceas con aproximadamente 96 géneros y unas 2300 especies, está ampliamente distribuida en las regiones templadas y tropicales de todo el mundo aunque su mayor diversidad se encuentra en el Hemisferio Sur, particularmente en América Central y Sud América. Otros centros de origen se localizan en Australia y África, con pocas especies en Europa y Asia. (Cabrera, 1983; Edmonds y Chweya, 1997; Figueroa Romero et al., 2000). Varias especies de esta familia tienen importancia económica, por ser la fuente de plantas alimenticias como la papa (Solanum tuberosum L.), el tomate (Lycopersicon esculentum Mill.), la berenjena (Solanum melongena L.), pimientos y ajíes (Capsicum spp). Otras Solanáceas interesan desde el punto de vista farmacológico por la producción de alcaloides como el estramonio (Datura stramonium L.), la belladona (Atropa belladona) y el tabaco (Nicotiana tabacum L.). Algunas especies de los géneros Petunia, Brunfelsia y Nierembergia, son a menudo cultivadas por lo atractivo de sus flores y numerosas son tóxicas para animales como los “chuschos” (Nierembergia linariefolia Graham var. linariefolia), el “yuyo de San Vicente” (Nicandra physalodes (L.) Gaertn ) y la “hediondilla” (Cestrum parqui L´Hér.) entre otras. En la familia Solanáceas, Solanum es el género más grande y complejo; lo componen más de 1500 especies, y como ya se mencionara, muchas de ellas económicamente importantes en todo el planeta (Edmonds y Chweya, 1997). En la República Argentina existen 35 géneros de la Familia Solanáceas con aproximadamente 400 especies, indígenas o naturalizadas; la mayoría de las especies son del género Solanum (Cabrera, 1983). Para Barboza et al., (1999) en Argentina son 33 los géneros de Solanáceas y 328 las especies. En el género Solanum 58 es el número de las especies argentinas. (Ver descripción botánica del género en Anexo Cap. 1 Pág. 15). Figueroa Romero et al., (2000), en una sinopsis de los géneros y especies en Tucumán, afirman que la familia Solanáceas es una de las más importantes de la flora de la provincia, tanto por el número de géneros como por las especies presentes. Estas Solanáceas están ampliamente 27 representadas y adaptadas en los 17 Departamentos de Tucumán, en sus 10 formaciones vegetales y a distintos hábitats; crecen en niveles altitudinales que oscilan entre los 250 hasta los 4.300 m.s.m. Mediante el empleo de datos exomorfológicos y fitogeográficos se han reconocido en Tucumán 21 géneros de Solanáceas, con aproximadamente 100 especies. El mayor número de géneros está representado en el Bosque Chaqueño (17 géneros de un total de 21, un 81%) y la mayor cantidad de especies pertenecen al género Solanum (42 especies). 1.2.1. Aspectos Taxonómicos del Género Solanum Las publicaciones de Edmonds y Chweya, (1997) se refieren a la complejidad taxonómica de las especies asociadas a la sección Solanum. Varios estudios clásicos, experimentales y numéricos han demostrado que la complejidad es atribuible a varias causas. Estas causas son: Históricas: desde 1732 hasta la fecha la sección Solanum ha sido estudiada, clasificada y reclasificada en especies y subespecies y aun hoy no hay un acuerdo. Plasticidad fenotípica: estas especies presentan variación particularmente en sus caracteres vegetativos, (hábito, ramas aladas, tamaño y forma de las hojas) y en algunos caracteres reproductivos (número, tamaño y coloración de los frutos). Estos dependen de factores como la intensidad de la luz y de la temperatura, por consiguiente, es difícil determinar cuales son los caracteres genéticamente fijos y cuales dependen del ambiente. Poliploidía: Las especies pertenecientes a la sección constituyen series poliploides (di, tetra, exa y hasta octoploides). Hibridación Natural: son predominantemente autopolinizadas, pero pueden ocurrir cruzamientos e hibridaciones inter e intraespecíficas especialmente en diploides de flores pequeñas. Posiblemente, esta hibridación natural está más extendida de lo que generalmente se supone. Variación Discordante: se define como un patrón de variación en el cual las magnitudes relativas en que se diferencian las poblaciones no coinciden con el conjunto de los diferentes atributos. De las 42 especies del género Solanum encontradas en la provincia de Tucumán, algunas han sido citadas como malezas en trabajos de Arévalo (1975), de Devani et al., (1993) y Roncaglia et al.,(2000), pero no siempre ha sido precisa la identificación. 1.2.2. Aspectos Taxonómicos del Complejo Solanum nigrum Ogg et al., (1989); Roberts y Lockett (1978); Orzoleck (1999) entre otros, han llamado Complejo Solanum nigrum (Solanum nigrum Complex) a varias especies de malezas del género Solanum. El Complejo Solanum nigrum está constituido por un grupo de especies estrechamente relacionadas, ampliamente distribuidas en las regiones de clima cálido de todo el planeta, desde el nivel del mar hasta las altas montañas (Edmonds y Chweya, 1997). Según Ogg et al., (1981, 1989) y Orzolek, (1999), el Complejo Solanum nigrum está formado en América del Norte por tres especies: Solanum americanum Mill. (american black nightshade) que 28 es común en muchas áreas del sur y costeras de EEUU, Solanum nigrum L. (black nightshade) en los estados más occidentales y Solanum ptycanthum Dun. (eastern black nightshade) en el este de las Montañas Rocallosas. Además, se considera en este complejo a una cuarta especie, Solanum sarrachoides Sendtn. (hairy nightshade) que se la encuentra en toda América del Norte. En África y Asia, otras son las especies estrechamente relacionadas con Solanum nigrum, que también presentan una identificación confusa (Edmonds y Chweya, 1997). Solanum nigrum, Solanum americanum y Solanum sarrachoides se mencionan en un complejo de 13 especies de Solanum. Desde el punto de vista taxonómico, el complejo Solanum nigrum es confuso debido al gran número de especies, a la aparente uniformidad de muchas de ellas, y a su plasticidad y variabilidad genética (Edmonds y Chweya, 1997; Orzolek, 1999). Este grupo de plantas ha sido objeto de numerosos estudios a escala mundial como malezas y en algunas regiones, como recursos alimenticios y medicinales, existiendo una gran confusión sobre la precisa identificación del mismo (Holm et al., 1977; Edmonds y Chweya, 1997). En el Hemisferio Sur se encuentran dos de las cuatro especies del Complejo Solanum nigrum citadas en el Hemisferio Norte, en África y en Asia: Solanum nigrum L. y Solanum americanum Mill. (Lorenzi, 2000). Las especies del Complejo Solanum nigrum son malezas que invaden una gran cantidad de cultivos y ocasionan daños directos o indirectos, que en ciertos casos pueden llegar a ser muy severos (Holm et al., 1977; Keeley y Tullen, 1989, 1991; Kleifeld et al., 1991; Kogan, 1992; McGiffen y Masiunas, 1992; Kissman y Groth, 1995; Tan y Weaver, 1996; Orzolek, 1999). También como malezas han sido citadas por estos autores en caña de azúcar, textiles, granos y hortícolas. En estos últimos es difícil el control por pertenecer a la misma familia botánica de la papa, del tomate, berenjena y pimiento. Entre los numerosos problemas que ocasionan, Holm et al., (1977) destacan los siguientes: ramas y hojas producen atascamiento de cosechadoras; las bayas maduras manchan los granos, los capullos de algodón y la lana de las ovejas; el control es difícil en cultivos de hoja ancha; son muy visitadas por hormigas, vaquitas y otros insectos, agentes transmisores de enfermedades; son hospederas de nematodos, bacterias, hongos y virus que causan importantes enfermedades en diversos cultivos y son plantas tóxicas para el hombre y animales. Solanum nigrum L., la especie que da el nombre a este complejo, es la más estudiada en el mundo. En algunos países del Hemisferio Norte, Australia y África se la conoce como “black nightshade” y como “yerba mora” en los de habla hispana. ( Ver Tabla Nº1-1. Nombres de Solanum nigrum L., en Anexo Capítulo 1, pág 14). Es originaria de Europa, crece desde los 54º de Lat. N hasta los 45º de Lat. S y está considerada como una de las 76 peores malezas del mundo, mencionada como muy importante en 67 países y en 37 cultivos (Holm et al., 1977). Además, es común en jardines, campos, áreas sin cultivar, desmontes, tanto en ambientes húmedos como en áreas con bajas precipitaciones. 1.2.3.- Las especies del Género Solanum en la República Argentina 29 1.2.3.1.- Solanum nigrum L. En la República Argentina Solanum nigrum L. es conocida como “yerba mora”. Ha sido citada como maleza de caña de azúcar (Holm et al., 1977; Arévalo, 1975), pero sin especificar datos sobre su incidencia en el rendimiento de este cultivo o en otros de importancia económica. Morton (1976) la incluye en la Revisión de las especies del género Solanum para la República Argentina a pesar de que es una especie exótica de origen europeo, aparentemente introducida en el Este de América del Norte. En las observaciones de su descripción, el autor aclara que: ---“numerosos ejemplares argentinos han sido identificados como Solanum nigrum, pero no he visto ninguno que represente verdaderamente a esta especie. Sin embargo, no sería raro que aparezca como maleza o fuera introducida, y por esta razón se la incluye en la clave.”--(Ver Anexo Cap.1 Pág. 16-20. Fig 1-1 a 1-3). 1.2.3.2.- Solanum americanum Mill Conocida como “maria-pretinha”, “erva-moura”, es descripta como una maleza frecuente en Brasil. Produce daños en áreas de cultivos anuales y perennes, en huertos, cafetales, y también aparece en jardines y en terrenos baldíos (Lorenzi, 2000). Solanum nigrum var. americanum es uno de los cinco sinónimos que Lorenzi (2000) cita para Solanum americanum. En la República Argentina Morton (1976) y Barboza et al., (1999) se refieren a Solanum americanum como una de las especies excluidas de la flora argentina. También Morton expresa que puede estar presente por haber sido introducida como maleza. En las observaciones, este autor aclara que: ---“Muchos de los ejemplares tradicionalmente identificados como S. nigrum de los trópicos americanos probablemente se refieren a S. americanum, el verdadero S. nigrum que se limita quizás a Europa, o que pudo introducirse desde otra parte. No he visto ningún espécimen de Argentina pero este puede aparecer como maleza.”--(Ver Anexo Cap.1, pág 21-23; Fig. Nº 1-4 y 1-5). 1.2.3.3.- Solanum lorentzii Bitter Solanum lorentzii, “yerba mora”, es una especie nativa del Noroeste Argentino (Cabrera, 1983; Barboza et al., 1999). Se la encuentra desde Jujuy hasta La Rioja. Figueroa Romero et al., 2000 la registran como parte de la flora en 10 Departamentos de Tucumán. Es considerada maleza de cultivos de granos en el Norte de Argentina por Roncaglia et al. (2000). Cuenta con las descripciones botánicas de Morton (1976), Cabrera (1983) y referencias de Barboza et al. (1999). No se encontraron otras referencias en las búsquedas bibliográficas. 30 (Ver en Anexo Capítulo 1, Pág. 24 a 28; Fig. Nº 1-6 y 1-7). 1.2.3.4.- Solanum bermejense Bitter Solanum bermejense es otra especie nativa de Argentina que Cabrera (1983), describe y también la llama “yerba mora”. Ha sido registrada en las provincias de Salta y Jujuy y en Bolivia entre los 1000 y 2000 m s n m (Barboza et al., 1999). Para esta especie sólo se cuenta con la descripción de Cabrera (1983), el material del herbario del Instituto Miguel Lillo (LIL) y la referencia del Catálogo del Instituto de Botánica Darwinion (IBODA) (Barboza et al., 1999). No se encontraron otras referencias para Argentina y Sud América. (Ver en Anexo Capítulo 1, Pág. 28-29). 1.2.3.5.- Solanum sublobatum Willd. En la búsqueda de información sobre las especies del Complejo Solanum nigrum se encontraron las opiniones de autores como Cabrera (1965), C. V. Morton (1972), Ragonese y Milano (1984) y Marzocca (1994 y 1997) quienes dicen que es una planta común en los alrededores de la ciudad de Buenos Aires y que ha sido clasificada erróneamente como Solanum nigrum L.. La localidad típica de Solanum sublobatum es Buenos Aires. En 1972, C. V. Morton, taxónomo del Departamento de Botánica del Smithsonian Institutions, Washington, al efectuar una revisión del material del género Solanum en herbarios de nuestro país, estableció que Solanum nigrum L. no se encuentra en Argentina y que los ejemplares atribuidos a esta especie corresponden a Solanum sublobatum Willd.. Ragonese y Milano, 1984, la incluyen en la recopilación de plantas tóxicas para animales domésticos donde consideran que los antecedentes tóxicos existentes en nuestro país atribuidos a la “yerba mora”, Solanum nigrum L., deben ser referidos a la especie del epígrafe (Solanum sublobatum), o a otras especies de Solanum, conocidas con el nombre vernáculo de “yerba mora”. Marzocca describe a esta planta como una de las malezas del Cono Sur de América (1994) y en su Vademécum de Malezas Medicinales (1997) como una especie fácilmente confundible con Solanum gracilis (Solanum gracilius) y con sus sinónimos, y afirma que constituye un complejo sistemático confuso y de difícil dilucidación. Por último agrega que frecuentemente se la identifica como Solanum nigrum. Su hábitat: ruderal; en baldíos y diversos cultivos (trigo, arroz, soja, y otras leguminosas de grano, algodón, café), montes frutales y pasturas; prefiere los suelos arenosos, pedregosos, húmedos y sombreados fértiles. Es una de las primeras colonizadoras de terrenos de reciente desmonte, y su presencia en pasturas es inconveniente pues se trata de una especie sumamente tóxica para el ganado que, aunque no es consumida normalmente por éste cuando tiene bastante forraje a su disposición, resulta peligrosa en época de escasez. 31 Cuando invade los cultivos de soja es perjudicial principalmente en época de cosecha, pues sus frutos maduros pueden manchar de morado o violáceo los granos depreciándolos comercialmente. Otros de los sinónimos encontrados son Solanum chenopodioides Lam.; Solanum gracile Dun. (Ver en Anexo Capítulo 1, Fig. 1-10 a 1-15). Como Solanum bermejense y Solanum lorentzii es una especie nativa de Argentina pero su distribución es muy distante de donde crecen estas; se la encuentra de 0 a 500 m s m. No se encontraron citas para el Noroeste o para Tucumán. 1.2.4.- Las especies del Género Solanum en Tucumán De las 1.500 especies del género Solanum encontradas en el mundo se han mencionado alrededor de 150 en Argentina y 42 en la Provincia de Tucumán. De estas 42, seis (6) se han citado como malezas en trabajos de Arévalo (1975) y Devani et al. (1993), pero la identificación no siempre ha sido precisa. Arévalo (1975) registra a Solanum nigrum como maleza perenne de caña de azúcar, mientras que Devani et al. (1993) como una maleza anual de soja pero con su nombre común, “yerba mora”. Relevamientos realizados durante diez años en los cultivos más importantes en la provincia de Tucumán informan sobre 12 malezas del género Solanum (de Andrada et al., 1995; Mansilla et al., 1998; Roncaglia et al., 1991, 1993 a, 1993 b, 1995, 1998, 2000). Las especies del género Solanum registradas e identificadas en estos relevamientos son: (Anexo Cap. 1. Tabla Nº 1-4) Solanum adventitium Polg.; Solanum atriplicifolium Gillies; Solanum chacoense Bitter; Solanum chenopodioides Lam.; Solanum conditum C.V. Morton; Solanum hieronymii Kuntze; Solanum lorentzii Bitter; Solanum nitidibaccatum Bitter; Solanum physalidicalyx Bitter; Solanum pygmaeum Cav; Solanum pilcomayense Morong; Solanum sisymbriifolium Lam. Se destaca la ausencia de Solanum nigrum, y algunos ejemplares podrían ser de otras especies, como Solanum bermejense o Solanum americanum que fueron observados sólo en el Herbario del Instituto Miguel Lillo, Tucumán. En general, en este grupo de Solanáceas, que forman el Complejo Solanum nigrum, hay un polimorfismo notable. Al respecto, según Cabrera (1983), en la Flora de la Provincia de Jujuy, las descripciones de Solanum lorentzii Bitter y Solanum bermejense Bitter no son lo suficientemente claras para establecer una diferencia consistente; se basan en la forma de las hojas y en el número de corpúsculos pétreos del fruto. Sin embargo, las características morfológicas son frecuentemente influenciadas por las condiciones ambientales (Edmonds y Chweya, 1997; Botta, 1979), lo que complica aún más las identificaciones. (Tabla Nº 1-5.) Tabla Nº 1-5. Caracteres diferenciales de dos especies de Solanum. (Cabrera, 1983) 32 Especie Solanum lorentzii Solanum bermejense Forma de la lámina ovado u ovado – lanceolada lámina lanceolada Corpúsculos Pétreos Corpúsculos 2 - 6 Corpúsculos numerosos Son escasas las publicaciones en el ámbito regional o provincial sobre taxonomía de las especies de Solanáceas aquí mencionadas. Tampoco se encontraron referencias sobre la importancia como malezas y su control o sobre la biología de éstas. Anexo capítulo1: 1.2.5.- Distribución de Solanum nigrum en el mundo (Holm et al., 1977). 1.2.7.- Distribución de Solanum lorentzii en Tucumán (Figueroa Romero et al., 2000). 33 Tabla Nº 1-1. Nombres de Solanum nigrum L. en los países que se encuentra como maleza (Holm et al., 1977). 34 ALEMANIA: schwarzer Nachtschatten ARGENTINA: yerba mora AUSTRALIA: blackberry nightshade BANGLADESH: gurki BIRMANIA: baung-laung-nyo BRASIL: erva moura CAMERÚN: black nightshade CHILE: hierba mora, llague COLOMBIA: yerba mora COSTA RICA: yerba mora DINAMARCA: black nightshade EGIPTO: enab el-deib EL SALVADOR: yerba mora ESPAÑA: tomatito de moro, tomatitos ESTADOS UNIDOS: black nightshade FIDJI: black nightshade FILIPINAS: kamakamatisan, kunti, malasili FINLANDIA: mustakoiso FRANCIA: crève-chien HAWAII: black nightshade HONDURAS: hierba mora INDIA: makhoi (Rajasthani), nunununia (Oriya) INDONESIA: anti (Indonesian), leuda, leuntja, leuntja pait (Sundanese), ranti (Javanese) INGLATERRA: black nightshade IRAN: taj rizi IRAQ: innaib el-theeb ITALIA: ballerina, erba morella, solano nero JAMAICA: black nightshade, branched calalu, guma JAPÓN: inuhōzuki 137 MALASIA: terong meranti MARRUECOS: morelle noire MAURICIO: brède martin MÉJICO: hierba mora, trompillo NATAL: black nightshade NORUEGA: sort sǿtvider NUEVA ZEALANDIA: black nightshade PAÍSES BAJOS: zwarte nachtschade PAKISTÁN: kanper makoo PANAMÁ: pintamora PERÚ: yerba mora RHODESIA: i xabaxaba (Ndebele), musaka (Shona), mutsungutsugu (Ndeo) SUD ÁFRICA: black nightshade, galbessie, nagskaduuse, nastagal, nastergal SUECIA: nattskatta TAILANDIA: toem tok, ya-tomtok TAIWAN: lung-kwei TRINIDAD: agouma TÚNEZ: morelle noire TURQUÍA: kopek uzumii VENEZUELA: hierba mora, yocoyoco YUGOSLAVIA: pomocnica 138 Descripción botánica de Cabrera, A. L. (1983) del género Solanum Solanum L. Linnaeus, Spec.1:184, 1753 Flores hermafroditas, actinomormorfas. Cáliz acampanado, pentadentado o pentapartido, más raramente casi entero, con 5 o 10 dientes muy cortos. Corola rotácea o acampanada, con tubo muy corto y limbo pentalobado ( a veces con lóbulos casi inconspicuos) de prefloración plegada. Estambres 5, insertos en la garganta de la corola, con las anteras libres pero generalmente conniventes, oblongas o atenuadas hacia el ápice, dehiscentes por un poro apical más tarde continuando hacia la base de la antera. Disco poco conspicuo. Ovario bilocular, multiovulado. Estilo simple, con estigma algo engrosado, obtuso. Fruto baya carnosa acompañada por el cáliz que, algunas veces es acrescente. Semillas numerosas, comprimidas orbiculares o semireniformes, a veces acompañadas por corpúsculos pétreos. Embrión curvo. Hierbas anuales o perennes, subfrútices, arbustos o árboles, a veces volubles o apoyantes, aculeados o innermes. Hojas alternas o acopladas, sin estípulas (a veces con hojas nuevas en la axila que semejan estípulas). Inflorescencias cimosas. Flores pequeñas o medianas, de colores diversos. Especie tipo: Solanum nigrum L. Género tropical y subtropical con cerca de 2000 especies. Algunas tienen gran importancia económica, como la papa (Solanum tuberosum), la berenjena (Solanum melongena) y otras. 139 Solanum nigrum L. C. V. Morton, (1976). Traducción de la descripción. Plantas decumbentes o erectas, no ramificadas desde la base, las plantas jóvenes con o sin las hojas basales en rosetas, el diámetro de las ramas en la base es de 3,3 – 4,3 mm, carece de crestas suberosas; limbo de las hojas ovadas, de 4,6 – 6,2 cm de long., con un promedio de 0,60 – 0,75 veces, tan largo como ancho, los limbos de 4,9 – 8,5 veces tan largos como los pecíolos, cuneados en la base, con un promedio de 1,8 – 4,4 dientes en cada lado, estos de 1,1 – 2,8 mm de profundidad, los pelos de las hojas de 3 – 19 por 7 mm2, de 0,1 – 0,2 mm de long., pelos glandulares largos, presentes o ausentes, pelos glandulares cortos y simples presentes; inflorescencias extra axilares, simples, racimosas o subumbeladas-racimosas, con un promedio de 3,8 – 5,6 flores por inflorescencia; pedúnculos en un ángulo de 68-79º, de 12 – 16,5 mm de long., pedicelos en un ángulo de 75 –108º, de 9 – 11,4 mm de long., pubescentes o casi glabros, no recurvados; cáliz regular o irregular de 2,25 – 2,45 mm de long. en las flores, senos de 0,65 – 0,75 mm de profundidad, sin mancha morada (púrpura), sin un área luminosa entre los lóbulos; corola blanca, la estrella central amarilla o amarillaverde, los lóbulos con promedio de 6,8 – 6,9 mm de long., los senos de 4,3 – 4,7 mm de profundidad, lóbulos con 2,6 – 3,1 mm de ancho; filamentos glabros o pubescentes en toda su longitud; anteras amarillas de 2,1 - 2,4 mm de long; polen de 10,8 – 11,2 µ de diámetro; estilo de 3,1 – 3,4 mm de long., casi igual que las anteras, pubescentes casi ½ del largo; cáliz 2, 8 – 3,4 mm en el fruto, reflexo; bayas de textura blanda, verdes o negras, globosas, de 7,7 - 8,8 mm de diámetro, brillante u opaco; semillas de 34 – 40 por fruto de 2 – 2,1 mm de diámetro, color suave; corpúsculos pétreos ausentes. Tipo.--- De Europa pero aun no determinado Rango. --- Europa, introducido en el este de Norte América y otros sitios. Obs. --- Numerosos ejemplares argentinos han sido identificados como S. nigrum pero no he visto ninguno que represente verdaderamente a esta especie. Sin embargo, es probable que ésta se encuentre como maleza o sean introducidas y razón por la se incluye en la clave. Cabrera, 1979 cita a Solanum nigrum en la Flora de la Provincia de Jujuy sólo como la especie tipo. Barboza et al., (1999) incluyen a esta especie en el Catálogo de las Plantas Vasculares de la República Argentina, pero se aclara que no está incluida dentro de la flora nativa. Solanum nigrum L. Ref.: Morton, 1976; Cabrera, 1979. Nota: Especie excluida de la flora argentina Solanum nigrum L. var. americanum (Mill.) O.E. Schulz =Solanum americanum Mill. Solanum nigrum L. var. frutescens auct. non A. Br. =Solanum sublobatum Willd. Solanum nigrum L. var. frutescens auct. non A. Br. = Solanum lorentzii Bitter. Solanum nigrum L. var. humile auct. non Bernh. = Solanum pygmaeum Cav. var. pygmaeum Solanum nigrum L. var. humile auct. non Bernh. = Solanum tripartitum Dunal [Solanum nigrum L. var. miniatum Bernh.] Nota: Taxón dudoso. Solanum nigrum L. var. pilcomayense (Morong) Chodat = Solanum pilcomayense Morong var. pilcomayense Solanum nigrum L. var. virginicum L. = [Solanum americanum Mill.] 140 Solanum nigrum L. subsp. chacoense Hassl. = Solanum pilcomayense Morong var. pilcomayense Solanum nigrum L. subsp. chacoense Hassl. var. genuinum Hassl., = Solanum pilcomayense Morong var. pilcomayense Solanum nigrum L. subsp. chacoense Hassl. var. genuinum Hassl. f. floribundum Hassl. = Solanum pilcomayense Morong var. pilcomayense. Solanum nigrum L. subsp. chacoense Hassl. var. subhastatum Hassl. f. longepetiolatum Hassl. = Solanum pilcomayense Morong var. pilcomayense 141 Solanum nigrum L. A. planta – B. Hoja - C. Corola estrellada – D. Flor madura – E. Infrutescencia - F. Fruto. Ilustración: Luciana Andrada. 2009 142 Fig. Nº 1-2. Solanum nigrum L. detalles de la planta. (Holm et al., 1977). 143 Fig. Nº 1-3. Mapa de distribución mundial de Solanum nigrum L. (Holm et al., 1977). 144 Solanum americanum Miller C. V. Morton, (1976). Traducción de la descripción. Plantas erectas, no ramificadas desde la base, las plantas jóvenes con hojas basales en rosetas, el diámetro de las ramas en la base es de 5,3 mm, éstas carecen de crestas suberosas; hojas ovadas o deltoides-ovadas, de 8,1 – 8,7 cm de long. por 0,61 – 0,66 de ancho, los limbos de 5,4 – 5,6 veces tan largos como los pecíolos, cuneados en la base, con 4,3 – 5 dientes en cada lado, estos de 4,1 – 5,5 mm de profundidad, los pelos de las hojas de 12 – 34 por 7 mm2, de 0,2 – 0,4 mm de long., pelos glandulares largos ausentes, pelos glandulares cortos y simples presentes; inflorescencias extra axilares, simples, subumbeladas, con un promedio de 3,2 – 3,7 flores por inflorescencia, pedúnculos en un ángulo de 99º, de 16 – 22 mm de long., pedicelos en un ángulo de 71 –79º, de 8,2 – 8,9 mm de long., escasamente pubescentes no recurved; cáliz regular o irregular de 2,5 mm de long. en las flores, senos de 0,89 – 1,15 mm de profundidad, sin mancha morada (púrpura) o un área luminosa entre los lóbulos; corola blanca con púrpura, la estrella central verde, los lóbulos de 5,1 – 6,5 mm de long. con promedio estrechoen la amplitud de 2 – 2,5 mm de ancho, los senos de 3,5 – 4,1 mm de profundidad; filamentos pubescentes en la base o en toda su longitud ; anteras amarillas de 1,4 – 1,7 mm de long; polen de 7,5 – 8,3 µ de diámetro; estilo de 2,4 – 2,6 mm de long. casi igual que las anteras, pubescentes en la base ⅓ o ½; cáliz 2, 45 – 3,3 mm en el fruto, reflexo o ligeramente reflexo; bayas de texturas blandas, negras, globosas, de 8,3 – 9 mm de diámetro, brillante; semillas de 63 – 66 por fruto de 1,43 – 1,7 mm de diámetro, color suave; corpúsculos pétreos en promedio de 7,7 – 7,9 por fruto. Tipo.--- Virginia (holotipo presumiblemente en Herb. Miller, BM, no lo vi). Rango. --- Incierto, pero probablemente este de los EEUU, West Indians y norte de Sud América. Obs. --- Muchos de los ejemplares tradicionalmente identificados como S. nigrum de América tropical, son probablemente S. americanum. El verdadero S. nigrum L. está quizás limitado a Europa, excepto que se haya introducido en otro lugar. No he visto ejemplares de Argentina, pero esta especie puede encontrarse como maleza. En la República Argentina la revisión de las Solanáceas Argentinas de Morton (1976) y Barboza et al., (1999) en el Catálogo de las Plantas Vasculares de la República Argentina II, excluyen a Solanum americanum de la flora Argentina. Esta última es la información más reciente y sólo brinda los sinónimos de esta especie. [Solanum americanum Mill.] Sinónimos: Solanum adventitium Polg., Solanum nigrum L. var. americanum (Mill.) O.E. Schulz, Solanum nigrum L. var. virginicum L., Solanum nodiflorum Jacq., Solanum oleraceum Dunal Ref.: Morton, 1976; D'Arcy, 1993. Nota: Especie excluida de la flora argentina. 145 Descripción http://www.jamart.com.br/plantas Planta herbácea anual, se reproduce por semillas. Hojas alternas, pecioladas, ovaladas o romboidales, de hasta 7cm de longitud de color verde. Inflorescencia axilar, con un pedúnculo de cuyo ápice sale una umbela ostentando hasta 6 pequeñas flores. Flores con cáliz de coloración verde con 5 lóbulos agudos; el cáliz persiste hasta la maduración de los frutos. Corola con 5 lóbulos lanceolados, blancos o con leve tonalidad púrpura. El fruto es un solanídio globoso de color verde cuando inmaduro y negro brillante cuando maduro. Cada fruto encierra de 50 a 100 semillas. La planta acepta bien condiciones de climas tropical, subtropical o templado, mas para un buen desarrollo exige suelo fértil, rico en nitrógeno y con buena humedad. En la parte meridional de Brasil se la encuentra de primavera a otoño y recibe los siguientes nombres populares: "erva-moura", maria-preta, maria-pretinha, pimenta, pimenta-de-galinha, pimenta-de-rato, pimenta-de-cachorro, erva-de-bicho erva-mocó, aguarágua, aguaraquiá, guaraquinha e caraxixú. Las diferencias entre Solanum americanum y Solanum nigrum se muestran en la Tabla Nº1-2, aunque en la práctica esas diferencias no son consistentes. Los autores han encontrado en Brasil muchas plantas con inflorescencia en umbelas imperfectas, las diferencias de fracciones de milímetro de largo de las anteras no son relevantes y la coloración de las hojas depende del estado nutricional de la planta. La separación más aceptable sería la geográfica, pero es rota por la dispersión. Tabla Nº 1-2. Caracteres diferenciales de SOLAM y SOLNI Solanum americanum Inflorescencia en umbelas con hasta 6 flores, cuyos pedicelos salen todos de un mismo punto (lo que en la práctica no siempre acontece). Anteras nunca con más de 2mm de longitud. Hojas de coloración verde poco intensa. Frutos con 50 a 100 semillas. Solanum nigrum Inflorescencia en umbelas con 5 a 10 flores, en umbelas imperfectas con algunos pedicelos saliendo abajo del punto apical del pedúnculo. Anteras con hasta 2,6mm de longitud. Hojas de coloración verde-oscura. Frutos con 15-60 semillas. solanìdio: fruto carnoso indehiscente, bacóide 146 Fig.Nº 1-4. SOLAM – Flores y frutos Fig. Nº 1-5 Detalles de SOLAM 147 Solanum lorentzii Bitter Especie nativa de la República Argentina, cuenta con las descripciones botánicas de Morton (1976), Cabrera (1983) y referencias de Barboza et al. (1999). Roncaglia et al. (2000) realizaron una descripción como maleza frecuente del Norte Argentino y Figueroa Romero et al. (2000) dan la distribución de ésta en la provincia de Tucumán. C. V. Morton, (1976). Traducción de la descripción Arbusto erecto, mayormente de 1 – 1,5 m de altura (raro solo o,6 m); ramas cortamente estrigilosas, no glandulares, las ramas más viejas con crestas evidentes, internodios elongados, 6 – 10,5 cm de long. pecíolos de 10 –15 mm de long. estrigilosos todo el margen del limbo con alas decurrentes; hojas ovadas a lanceoladas, de 7 – 15 cm de long. por 3 – 7 cm de ancho, atenuados en el ápice y en la base, enteros, herbáceos, muy finamente pubescente en ambos lados, no glandular; inflorescencias extra axilares, laterales nunca opuestas a las hojas, generalmente de la mitad a dos tercios arriba del entrenudo, con una o generalmente con dos bifurcaciones; pedúnculo elongado, ascendente, 2 – 6,5 cm de long., flores por inflorescencia numerosas (más de 10 y apiñadas); pedicelos cortos, 5 – 9 mm de long. en las flores, llegando a 12 mm en los frutos, con pubescencia densa no glandular-estrigilosa; cáliz campanulado, de 2 - 2,5 mm de long., 5-dentado, dientes iguales, no connados, deltoides, de 1,5 mm de long. y 1,5 mm de ancho en la base, agudos o no acuminados, con pelos muy finamente estrigilosos no glandulares en la parte externa; corola rotácea, púrpura o blanca, 12 – 16 mm de ancho, finamente pubescente por fuera, 5-lobadas llegan cerca de la mitad, la parte basal del tubo en general cilíndrica, los lóbulos triangulares, de 5 – 7 mm de long.; filamentos igual, 1,25 mm de long., insertos 1 mm encima de la base del tubo de la corola, connados en la base por 0,7 mm, con pelos mas bien largos en el interior; anteras libres, angostamente oblongas, de 3,8 – 4,7 mm de long. 1 – 1,1 mm de ancho, el poro terminal introrso, de 0,6 mm long.; ovario glabro; estilo de 6,5 – 7 mm de long.; densamente piloso por dos tercios hacia la parte superior; bayas esféricas, de 5 mm de diámetro; corpúsculos pétreos 2 apicales, o 6 (2 apicales y 4 en pares en los lados opuestos del fruto). Tipo.--- Chacarita de los padres, Catamarca, Nov. 21 – 24, 1872, LORENTZ et HIERONYMUS 440 (CORD, lectotipo; B, destruido, F foto 2782) Rango. --- Argentina (Jujuy, Salta, Catamarca, Tucumán, Santiago del Estero, La Rioja). Obs. --- La información argentina de S. cymosum RUIZ et PAVON se basa en esta especie. De acuerdo con BITTER, que vio el isotipo, S. cymosum es bastante diferente, y pertenece a la sección Dulcamara, y es probablemente un sinónimo de S. corymbosum JACQ.. Por el contrario, S. lorentzii es un típico miembro de la sección Morella con inflorescencias laterales; de hecho, algunos de los ejemplares citados arriba fueron identificados originalmente como S. nigrum L. lo cual sin embargo, no es del todo exacto. BITTER que siempre insiste en el número y posición de los corpúsculos pétreos en las bayas, ha distinguido el típico S. lorentzii (de Jujuy y Catamarca) porque tiene sólo 2 corpúsculos pétreos apicales, de la var. tucumanicum, que tiene 6, dos apicales y 4 en dos pares en cada lado de la baya. Este carácter necesita ser investigado en el campo. Todos los ejemplares que 148 vi tienen 6 corpúsculos en el fruto, incluyendo el único que vi de Catamarca. Soy por consiguiente reacio de reconocer var. tucumanicum esta vez. Morton clasifica a Solanum lorentzii en dos variedades: - Solanum lorentzii Bitter var. tucumanicum Bitter = Solanum lorentzii Bitter Ref.: Morton, 1976; Cabrera, 1983. - Solanum lorentzii Bitter var. montigenum C.V. Morton = Solanum montigenum (C.V. Morton) Cabrera Ref.: Cabrera, 1983. Difiere de la variedad típica en sus dientes irregulares en el borde de las hojas. Tipo. --- En el U. S. National Herbarium. Nº 1.548.937, colectado en Estancia Santa Rosa, Dpto. Chicligasta; Provincia de Tucumán, Argentina, Enero 8, 1927, a 3300 m.s.m., VENTURI 4760, Isotipos: AA; LP; SI Rango. --- Conocido sólo en Argentina (Jujuy, Catamarca, Tucumán). Según Cabrera (1983) se la encuentra en el Noroeste de Argentina, desde Jujuy a La Rioja. Vegeta preferentemente en bosques y selvas, pero también aparece en los matorrales de la prepuna. Florece en primavera y verano. En esta publicación, cuando se describe a Solanum bermejense Bitter se encuentra la siguiente observación de Cabrera: Obs. Especie próxima a Solanum lorentzii, Bitter del que difiere por las hojas lanceoladas. El número de Corpúsculos pétreos de los frutos parece ser mayor. Tal vez se trate sólo de una variedad. Se transcribe a continuación la descripción de la Flora de Jujuy, Argentina. Solanum lorentzii Bitter, Fedde Repert. 11: 2, 1912 Arbusto erecto o apoyante de 1-2 m de altura, con ramas delgadas, más o menos densamente pubescentes, con pelos tendidos multicelulares, hojosas hasta el ápice. Entrenudos de 3 - 10 cm de long. Hojas alternas, pecioladas, con pecíolo de 1-2 cm de long., pubescente, algo alado por la decurrencia de la lámina. Lámina ovada u ovado – lanceolada, atenuada en el ápice y cuneada en la base, entera, laxamente pubescente en el haz y densamente en el envés, de 4 12 cm de long. Por 2 – 5 cm de lat. Inflorescencias extra- axilares, generalmente bifurcadas, 8 – 18 – floras; pedúnculo delgado, pubescente, de 2 – 6 cm de long.;pedicelos pubescentes de 5 – 10 mm. Cáliz acampanado, pubescente, de 2 – 2,5 mm de long., pentadentado, con dientes triangulares. Corola blanca de 12 – 15 mm de diámetro, profundamente pentalobada, con lóbulos triangulares, agudos, de 5 – 7 mm de long., menudamente pubescentes exteriormente. Anteras con filamentos de 1,2 – 1,5 mm de long. Y tecas oblongas, de alrededor de 4 mm de long. Estilo piloso en sus dos tercios inferiores. Bayas globosas, glabras, de 5 mm de diám. Con semillas comprimidas, redondeadas, de cerca de 1 mm de long. y 2 – 6 corpúsculos pétreos. 149 Solanum lorentzii Bitter: A. Planta – B. Hoja - C. Flor madura – D. Corola estrellada – E. Infrutescencia – F Fruto. Ilustró Luciana Andrada Mansilla (2009). 150 Barboza et al. (1999) brinda los siguientes datos en el Catálogo de las Plantas. Vasculares de la República Argentina II, del Instituto Darwinion: Solanum lorentzii Bitter Sinónimos: Solanum cymosum auct. non Ruiz & Pav., Solanum filiforme Ruiz & Pav. var. lanceolatum Kuntze, Solanum filiforme Ruiz & Pav. var. subdentatum Kuntze, Solanum lorentzii Bitter var. tucumanicum Bitter, Solanum nigrum L. var. frutescens auct. non A. Br., Solanum polystrichostylum Bitter var. lorentzii (Bitter) Edmonds Ref.: Edmonds, 1972; Morton, 1976; Cabrera, 1983. Hábito: Arbusto. Status: Nativa. Elevación: 500-2500 m. Prov.: CA, JU, LR, SA, SE, TU. Paises limítrofes: BO, PA. Ejemplar examinado: Venturi 5064 (SI). Y las dos variedades mencionadas anteriormente: - Solanum lorentzii Bitter var. montigenum C.V. Morton = Solanum montigenum (C.V. Morton) Cabrera. Ref.: Cabrera, 1983. - Solanum lorentzii Bitter var. tucumanicum Bitter = Solanum lorentzii Bitter Ref.: Morton, 1976; Cabrera, 1983. 151 Para Figueroa Romero et al., (2000) la distribución de Solanum lorentzii en la provincia de Tucumán es la siguiente: Departamentos Burruyacu, Capital, Cruz Alta, Chicligasta, Famaillá, Leales, Lules, Tafí del Valle, Tafí Viejo, Trancas y Yerba Buena que se observan en la Fig. Nº1-7.. Fig. Nº 1-7. Distribución de Solanum lorentzii Bitter en la provincia de Tucumán. Figueroa Romero et al., (2000). 152 Solanum lorentzii Bitter Especie nativa de la República Argentina, cuenta con las descripciones botánicas de Morton (1976), Cabrera (1983) y referencias de Barboza et al. (1999). Roncaglia et al. (2000) realizaron una descripción como maleza frecuente del Norte Argentino y Figueroa Romero et al. (2000) dan la distribución de ésta en la provincia de Tucumán. C. V. Morton, (1976). Traducción de la descripción Arbusto erecto, mayormente de 1 – 1,5 m de altura (raro solo o,6 m); ramas cortamente estrigilosas, no glandulares, las ramas más viejas con crestas evidentes, internodios elongados, 6 – 10,5 cm de long. pecíolos de 10 –15 mm de long. estrigilosos todo el margen del limbo con alas decurrentes; hojas ovadas a lanceoladas, de 7 – 15 cm de long. por 3 – 7 cm de ancho, atenuados en el ápice y en la base, enteros, herbáceos, muy finamente pubescente en ambos lados, no glandular; inflorescencias extra axilares, laterales nunca opuestas a las hojas, generalmente de la mitad a dos tercios arriba del entrenudo, con una o generalmente con dos bifurcaciones; pedúnculo elongado, ascendente, 2 – 6,5 cm de long., flores por inflorescencia numerosas (más de 10 y apiñadas); pedicelos cortos, 5 – 9 mm de long. en las flores, llegando a 12 mm en los frutos, con pubescencia densa no glandular-estrigilosa; cáliz campanulado, de 2 - 2,5 mm de long., 5-dentado, dientes iguales, no connados, deltoides, de 1,5 mm de long. y 1,5 mm de ancho en la base, agudos o no acuminados, con pelos muy finamente estrigilosos no glandulares en la parte externa; corola rotácea, púrpura o blanca, 12 – 16 mm de ancho, finamente pubescente por fuera, 5-lobadas llegan cerca de la mitad, la parte basal del tubo en general cilíndrica, los lóbulos triangulares, de 5 – 7 mm de long.; filamentos igual, 1,25 mm de long., insertos 1 mm encima de la base del tubo de la corola, connados en la base por 0,7 mm, con pelos mas bien largos en el interior; anteras libres, angostamente oblongas, de 3,8 – 4,7 mm de long. 1 – 1,1 mm de ancho, el poro terminal introrso, de 0,6 mm long.; ovario glabro; estilo de 6,5 – 7 mm de long.; densamente piloso por dos tercios hacia la parte superior; bayas esféricas, de 5 mm de diámetro; corpúsculos pétreos 2 apicales, o 6 (2 apicales y 4 en pares en los lados opuestos del fruto). Tipo.--- Chacarita de los padres, Catamarca, Nov. 21 – 24, 1872, LORENTZ et HIERONYMUS 440 (CORD, lectotipo; B, destruido, F foto 2782) Rango. --- Argentina (Jujuy, Salta, Catamarca, Tucumán, Santiago del Estero, La Rioja). Obs. --- La información argentina de S. cymosum RUIZ et PAVON se basa en esta especie. De acuerdo con BITTER, que vio el isotipo, S. cymosum es bastante diferente, y pertenece a la sección Dulcamara, y es probablemente un sinónimo de S. corymbosum JACQ.. Por el contrario, S. lorentzii es un típico miembro de la sección Morella con inflorescencias laterales; de hecho, algunos de los ejemplares citados arriba fueron identificados originalmente como S. nigrum L. lo cual sin embargo, no es del todo exacto. BITTER que siempre insiste en el número y posición de los corpúsculos pétreos en las bayas, ha distinguido el típico S. lorentzii (de Jujuy y Catamarca) porque tiene sólo 2 corpúsculos pétreos apicales, de la var. tucumanicum, que tiene 6, dos apicales y 4 en dos pares en cada lado de la baya. Este carácter necesita ser investigado en el campo. Todos los ejemplares que 153 vi tienen 6 corpúsculos en el fruto, incluyendo el único que vi de Catamarca. Soy por consiguiente reacio de reconocer var. tucumanicum esta vez. Morton clasifica a Solanum lorentzii en dos variedades: - Solanum lorentzii Bitter var. tucumanicum Bitter = Solanum lorentzii Bitter Ref.: Morton, 1976; Cabrera, 1983. - Solanum lorentzii Bitter var. montigenum C.V. Morton = Solanum montigenum (C.V. Morton) Cabrera Ref.: Cabrera, 1983. Difiere de la variedad típica en sus dientes irregulares en el borde de las hojas. Tipo. --- En el U. S. National Herbarium. Nº 1.548.937, colectado en Estancia Santa Rosa, Dpto. Chicligasta; Provincia de Tucumán, Argentina, Enero 8, 1927, a 3300 m.s.m., VENTURI 4760, Isotipos: AA; LP; SI Rango. --- Conocido sólo en Argentina (Jujuy, Catamarca, Tucumán). Según Cabrera (1983) se la encuentra en el Noroeste de Argentina, desde Jujuy a La Rioja. Vegeta preferentemente en bosques y selvas, pero también aparece en los matorrales de la prepuna. Florece en primavera y verano. En esta publicación, cuando se describe a Solanum bermejense Bitter se encuentra la siguiente observación de Cabrera: Obs. Especie próxima a Solanum lorentzii, Bitter del que difiere por las hojas lanceoladas. El número de Corpúsculos pétreos de los frutos parece ser mayor. Tal vez se trate sólo de una variedad. Se transcribe a continuación la descripción de la Flora de Jujuy, Argentina. Solanum lorentzii Bitter, Fedde Repert. 11: 2, 1912 Arbusto erecto o apoyante de 1-2 m de altura, con ramas delgadas, más o menos densamente pubescentes, con pelos tendidos multicelulares, hojosas hasta el ápice. Entrenudos de 3 - 10 cm de long. Hojas alternas, pecioladas, con pecíolo de 1-2 cm de long., pubescente, algo alado por la decurrencia de la lámina. Lámina ovada u ovado – lanceolada, atenuada en el ápice y cuneada en la base, entera, laxamente pubescente en el haz y densamente en el envés, de 4 12 cm de long. Por 2 – 5 cm de lat. Inflorescencias extra- axilares, generalmente bifurcadas, 8 – 18 – floras; pedúnculo delgado, pubescente, de 2 – 6 cm de long.;pedicelos pubescentes de 5 – 10 mm. Cáliz acampanado, pubescente, de 2 – 2,5 mm de long., pentadentado, con dientes triangulares. Corola blanca de 12 – 15 mm de diámetro, profundamente pentalobada, con lóbulos triangulares, agudos, de 5 – 7 mm de long., menudamente pubescentes exteriormente. Anteras con filamentos de 1,2 – 1,5 mm de long. Y tecas oblongas, de alrededor de 4 mm de long. Estilo piloso en sus dos tercios inferiores. Bayas globosas, glabras, de 5 mm de diám. Con semillas comprimidas, redondeadas, de cerca de 1 mm de long. y 2 – 6 corpúsculos pétreos. 154 Solanum lorentzii Bitter: A. Planta – B. Hoja - C. Flor madura – D. Corola estrellada – E. Infrutescencia – F Fruto. Ilustró Luciana Andrada Mansilla (2009). 155 Barboza et al. (1999) brinda los siguientes datos en el Catálogo de las Plantas. Vasculares de la República Argentina II, del Instituto Darwinion: Solanum lorentzii Bitter Sinónimos: Solanum cymosum auct. non Ruiz & Pav., Solanum filiforme Ruiz & Pav. var. lanceolatum Kuntze, Solanum filiforme Ruiz & Pav. var. subdentatum Kuntze, Solanum lorentzii Bitter var. tucumanicum Bitter, Solanum nigrum L. var. frutescens auct. non A. Br., Solanum polystrichostylum Bitter var. lorentzii (Bitter) Edmonds Ref.: Edmonds, 1972; Morton, 1976; Cabrera, 1983. Hábito: Arbusto. Status: Nativa. Elevación: 500-2500 m. Prov.: CA, JU, LR, SA, SE, TU. Paises limítrofes: BO, PA. Ejemplar examinado: Venturi 5064 (SI). Y las dos variedades mencionadas anteriormente: - Solanum lorentzii Bitter var. montigenum C.V. Morton = Solanum montigenum (C.V. Morton) Cabrera. Ref.: Cabrera, 1983. - Solanum lorentzii Bitter var. tucumanicum Bitter = Solanum lorentzii Bitter Ref.: Morton, 1976; Cabrera, 1983. 156 Para Figueroa Romero et al., (2000) la distribución de Solanum lorentzii en la provincia de Tucumán es la siguiente: Departamentos Burruyacu, Capital, Cruz Alta, Chicligasta, Famaillá, Leales, Lules, Tafí del Valle, Tafí Viejo, Trancas y Yerba Buena que se observan en la Fig. Nº1-7.. Fig. Nº 1-7. Distribución de Solanum lorentzii Bitter en la provincia de Tucumán. Figueroa Romero et al., (2000). Solanum lorentzii Bitter Especie nativa de la República Argentina, cuenta con las descripciones botánicas de Morton (1976), Cabrera (1983) y referencias de Barboza et al. (1999). Roncaglia et al. (2000) realizaron una descripción como maleza frecuente del Norte Argentino y Figueroa Romero et al. (2000) dan la distribución de ésta en la provincia de Tucumán. C. V. Morton, (1976). Traducción de la descripción Arbusto erecto, mayormente de 1 – 1,5 m de altura (raro solo o,6 m); ramas cortamente estrigilosas, no glandulares, las ramas más viejas con crestas evidentes, internodios elongados, 157 6 – 10,5 cm de long. pecíolos de 10 –15 mm de long. estrigilosos todo el margen del limbo con alas decurrentes; hojas ovadas a lanceoladas, de 7 – 15 cm de long. por 3 – 7 cm de ancho, atenuados en el ápice y en la base, enteros, herbáceos, muy finamente pubescente en ambos lados, no glandular; inflorescencias extra axilares, laterales nunca opuestas a las hojas, generalmente de la mitad a dos tercios arriba del entrenudo, con una o generalmente con dos bifurcaciones; pedúnculo elongado, ascendente, 2 – 6,5 cm de long., flores por inflorescencia numerosas (más de 10 y apiñadas); pedicelos cortos, 5 – 9 mm de long. en las flores, llegando a 12 mm en los frutos, con pubescencia densa no glandular-estrigilosa; cáliz campanulado, de 2 - 2,5 mm de long., 5-dentado, dientes iguales, no connados, deltoides, de 1,5 mm de long. y 1,5 mm de ancho en la base, agudos o no acuminados, con pelos muy finamente estrigilosos no glandulares en la parte externa; corola rotácea, púrpura o blanca, 12 – 16 mm de ancho, finamente pubescente por fuera, 5-lobadas llegan cerca de la mitad, la parte basal del tubo en general cilíndrica, los lóbulos triangulares, de 5 – 7 mm de long.; filamentos igual, 1,25 mm de long., insertos 1 mm encima de la base del tubo de la corola, connados en la base por 0,7 mm, con pelos mas bien largos en el interior; anteras libres, angostamente oblongas, de 3,8 – 4,7 mm de long. 1 – 1,1 mm de ancho, el poro terminal introrso, de 0,6 mm long.; ovario glabro; estilo de 6,5 – 7 mm de long.; densamente piloso por dos tercios hacia la parte superior; bayas esféricas, de 5 mm de diámetro; corpúsculos pétreos 2 apicales, o 6 (2 apicales y 4 en pares en los lados opuestos del fruto). Tipo.--- Chacarita de los padres, Catamarca, Nov. 21 – 24, 1872, LORENTZ et HIERONYMUS 440 (CORD, lectotipo; B, destruido, F foto 2782) Rango. --- Argentina (Jujuy, Salta, Catamarca, Tucumán, Santiago del Estero, La Rioja). Obs. --- La información argentina de S. cymosum RUIZ et PAVON se basa en esta especie. De acuerdo con BITTER, que vio el isotipo, S. cymosum es bastante diferente, y pertenece a la sección Dulcamara, y es probablemente un sinónimo de S. corymbosum JACQ.. Por el contrario, S. lorentzii es un típico miembro de la sección Morella con inflorescencias laterales; de hecho, algunos de los ejemplares citados arriba fueron identificados originalmente como S. nigrum L. lo cual sin embargo, no es del todo exacto. BITTER que siempre insiste en el número y posición de los corpúsculos pétreos en las bayas, ha distinguido el típico S. lorentzii (de Jujuy y Catamarca) porque tiene sólo 2 corpúsculos pétreos apicales, de la var. tucumanicum, que tiene 6, dos apicales y 4 en dos pares en cada lado de la baya. Este carácter necesita ser investigado en el campo. Todos los ejemplares que vi tienen 6 corpúsculos en el fruto, incluyendo el único que vi de Catamarca. Soy por consiguiente reacio de reconocer var. tucumanicum esta vez. Morton clasifica a Solanum lorentzii en dos variedades: - Solanum lorentzii Bitter var. tucumanicum Bitter = Solanum lorentzii Bitter Ref.: Morton, 1976; Cabrera, 1983. - Solanum lorentzii Bitter var. montigenum C.V. Morton = Solanum montigenum (C.V. Morton) Cabrera Ref.: Cabrera, 1983. Difiere de la variedad típica en sus dientes irregulares en el borde de las hojas. Tipo. --- En el U. S. National Herbarium. Nº 1.548.937, colectado en Estancia Santa Rosa, Dpto. Chicligasta; Provincia de Tucumán, Argentina, Enero 8, 1927, a 3300 m.s.m., VENTURI 4760, Isotipos: AA; LP; SI 158 Rango. --- Conocido sólo en Argentina (Jujuy, Catamarca, Tucumán). Según Cabrera (1983) se la encuentra en el Noroeste de Argentina, desde Jujuy a La Rioja. Vegeta preferentemente en bosques y selvas, pero también aparece en los matorrales de la prepuna. Florece en primavera y verano. En esta publicación, cuando se describe a Solanum bermejense Bitter se encuentra la siguiente observación de Cabrera: Obs. Especie próxima a Solanum lorentzii, Bitter del que difiere por las hojas lanceoladas. El número de Corpúsculos pétreos de los frutos parece ser mayor. Tal vez se trate sólo de una variedad. Se transcribe a continuación la descripción de la Flora de Jujuy, Argentina. Solanum lorentzii Bitter, Fedde Repert. 11: 2, 1912 Arbusto erecto o apoyante de 1-2 m de altura, con ramas delgadas, más o menos densamente pubescentes, con pelos tendidos multicelulares, hojosas hasta el ápice. Entrenudos de 3 - 10 cm de long. Hojas alternas, pecioladas, con pecíolo de 1-2 cm de long., pubescente, algo alado por la decurrencia de la lámina. Lámina ovada u ovado – lanceolada, atenuada en el ápice y cuneada en la base, entera, laxamente pubescente en el haz y densamente en el envés, de 4 12 cm de long. Por 2 – 5 cm de lat. Inflorescencias extra- axilares, generalmente bifurcadas, 8 – 18 – floras; pedúnculo delgado, pubescente, de 2 – 6 cm de long.;pedicelos pubescentes de 5 – 10 mm. Cáliz acampanado, pubescente, de 2 – 2,5 mm de long., pentadentado, con dientes triangulares. Corola blanca de 12 – 15 mm de diámetro, profundamente pentalobada, con lóbulos triangulares, agudos, de 5 – 7 mm de long., menudamente pubescentes exteriormente. Anteras con filamentos de 1,2 – 1,5 mm de long. Y tecas oblongas, de alrededor de 4 mm de long. Estilo piloso en sus dos tercios inferiores. Bayas globosas, glabras, de 5 mm de diám. Con semillas comprimidas, redondeadas, de cerca de 1 mm de long. y 2 – 6 corpúsculos pétreos. 159 Solanum lorentzii Bitter: A. Planta – B. Hoja - C. Flor madura – D. Corola estrellada – E. Infrutescencia – F Fruto. Ilustró Luciana Andrada Mansilla (2009). 160 Barboza et al. (1999) brinda los siguientes datos en el Catálogo de las Plantas. Vasculares de la República Argentina II, del Instituto Darwinion: Solanum lorentzii Bitter Sinónimos: Solanum cymosum auct. non Ruiz & Pav., Solanum filiforme Ruiz & Pav. var. lanceolatum Kuntze, Solanum filiforme Ruiz & Pav. var. subdentatum Kuntze, Solanum lorentzii Bitter var. tucumanicum Bitter, Solanum nigrum L. var. frutescens auct. non A. Br., Solanum polystrichostylum Bitter var. lorentzii (Bitter) Edmonds Ref.: Edmonds, 1972; Morton, 1976; Cabrera, 1983. Hábito: Arbusto. Status: Nativa. Elevación: 500-2500 m. Prov.: CA, JU, LR, SA, SE, TU. Paises limítrofes: BO, PA. Ejemplar examinado: Venturi 5064 (SI). Y las dos variedades mencionadas anteriormente: - Solanum lorentzii Bitter var. montigenum C.V. Morton = Solanum montigenum (C.V. Morton) Cabrera. Ref.: Cabrera, 1983. - Solanum lorentzii Bitter var. tucumanicum Bitter = Solanum lorentzii Bitter Ref.: Morton, 1976; Cabrera, 1983. 161 Para Figueroa Romero et al., (2000) la distribución de Solanum lorentzii en la provincia de Tucumán es la siguiente: Departamentos Burruyacu, Capital, Cruz Alta, Chicligasta, Famaillá, Leales, Lules, Tafí del Valle, Tafí Viejo, Trancas y Yerba Buena que se observan en la Fig. Nº1-7.. Fig. Nº 1-7. Distribución de Solanum lorentzii Bitter en la provincia de Tucumán. Figueroa Romero et al., (2000). 162 Solanum sublobatum Willd C. V. Morton, (1976). Traducción de la descripción Hierba de 30 – 70 cm de altura, muy ramificadas, el diámetro de las ramas en la base es por encima de 4 mm, de sección circular algo aplastadas(terete) o ligeramente angulosa-alada (wing-angled), con la pubescencia en dos líneas, las ramas jóvenes pilosas, los pelos de 0,5 mm de long. o más, curvos - ascendentes, más bien flácidos, pluricelulares, no glandulares, la punta angosta y puntiaguda; pecíolos 8 – 11 mm de long, densamente pilosos; hojas ovadas a ovadas – lanceoladas, de 4,5 cm de long. o más y 1,6 de ancho, obtusas o redondeadas en el ápice, angostamente cuneadas en la base y pecíolos decurrentes, enteras o a menudo con un par de pequeños lóbulos subbasales o a veces con 2 pares, escasamente pilosas en el haz, con una pubescencia muy fina, densa y persistente en el envés tanto en la superficie como en las nervaduras , la nervadura primaria con 5 pares; inflorescencias opuestas o sub opuestas a las hojas más superiores, el pedúnculo tiene de 10 – 15 mm de long. en antesis, con una muy fina pubescencia curvada no glandular, a menudo bifurcada, la bifurcación inferior a menudo uniflora, en la superior 2-4-flores, o a veces una inflorescencia simple, umbelada, con 3–4 (raramente 5) flores por inflorescencia, pedicelos de 4 mm de long. en las flores, con una muy fina pubescencia curvada no glandular; cáliz de 1,5 - 2 mm de long. ligeramente no glandularestrigilosa en la parte externa, lóbulos cercanos a la mitad, lóbulos deltoides de 0,75 – 1 mm de longitud , 0,6 mm de ancho en la base, obtusos o redondeados; corola blanca rotácea de 4 – 5 mm de long., 5 mm de ancho, externamente muy finamente estrigilosa, los lóbulos de 3 mm de long. y 1 mm de ancho en la base; filamentos iguales, insertos 0,75 mm encima de la base del tubo de la corola, las partes libres escasas, de 0,75 mm de long, pilosas adentro; anteras oblongas, de 2 – 2,5 mm de long 0,75 de ancho; estilo recto, escaso de 4 mm de long. piloso hasta encima de la mitad, el ápice no piloso; pedúnculo del fruto de 2 cm de long. pedicelos frutales (fruiting) de 10 mm de long. que se balancea o gira; cáliz no acrescente en el fruto de 1,5 – 6 mm de ancho; bayas globosas de 6 mm de diámetro, no brillante; carecen de corpúsculos pétreos. Tipo.--- Buenos Aires, Argentina (holotipo Herb. Willd, B. Field photograph 2912). No dan el recolector pero con certeza es COMMERSON; el tipo es muy semejante al tipo de S. chenopodifolium y es sin duda, una parte del mismo material original. Rango. --- Conocido sólo en el sur de Brasil, Uruguay y Argentina (Entre Ríos, Buenos Aires, Córdoba). Muestras examinadas en las tres provincias mencionadas. Obs. --- no hay observaciones. La información más reciente es la del Catálogo de las Plantas. Vasculares de la República Argentina II, (Barboza et al, 1999) del Instituto Darwinion que brinda los siguientes datos: Solanum sublobatum Willd. Sinónimos: Solanum aguaraquiya Piso ex Sendtn., Solanum chenopodiifolium Dunal, Solanum gracile Dunal var. microphyllum Dunal, Solanum isabellii Dunal, 163 Solanum nigrum L. var. frutescens auct. non A. Br., Solanum pterocaulon Dunal var. aguaraquiya (Piso ex Sendtn.) Dunal, Solanum subspatulatum Sendtn., Solanum vile Bitter Ref.: Edmonds, 1972; Morton, 1976; Cabrera, 1979. Hábito: Hierba. Status: Nativa. Elevación: 0-500 m. Prov.: BA, CO, ER. Paises limítrofes: BR, UR. Ejemplar examinado: Rodríguez 128 (SI). Tabla Nº 1-3. Descripciones y Distribución de Solanum sublobatum Willd. en Argentina. Ragonese y Milano (1984) Marzocca (1997) Subfrútice de 0,50 a 0,90 m de altura. Hierba anual, glabra o levemente Hojas ovado – lanceolada, base cuneada, pubescente (pelos simples), de 0,20 – generalmente con uno o dos gruesos 0,60 m de alt. con tallos ascendentes y dientes a cada lado, próximos a la base, erguidos, ramosos, inermes, y hojas a veces enteras. Flores en número de 3 – alternas, ovadas u ovado-lanceoladas, 5; corola blanca en cimas umbeliformes. de 3-5 cm de largo por 1-2 cm de Bayas negruzcas. ancho, acuminadas o subagudas, enteras o sinuado-lobadas o dentadas y de base atenuada o redondeada, pecioladas (pecíolo de 1 – 3 cm de largo); flores blancas, de unos 8-10 mm de diámetro, en racimos umbeliformes 3-10-floros; pedúnculos péndulos, de unos 2-3 cm de largo; lóbulos corolinos dirigidos hacia atrás; bayas esféricas, negro azuladas a la madurez, de 8-10 mm de diámetro, amarillas hasta pardo-oscuras. Distribución Sudeste de Brasil, Uruguay y la República Argentina (Córdoba, San Luis, Santa Fe, Buenos Aires, Entre Ríos). Distribución Cosmopolita. En América: Chile, Bolivia, Paraguay, Uruguay, Brasil, Argentina (Misiones, Corrientes, Chaco, Formosa, Entre Ríos, Buenos Aires, La Pampa, Río Negro). Marzocca (1994) da esta especie el nombre común de “hierba mora”, “yerba mora”. Además enumera los nombres con que se la conoce en otros países de Sudamérica: Chile: llague; tomatillo Bolivia: guapurucillo; ñusco 164 Paraguay: ará chichú; ara sichú; mata gallina; yerba gallina Brasil: aguarágua; aguaraquiya-açu; erva-moura; maria preta Fig. Nº 1-9. Solanum sublobatum Willd. “yerba mora”. Ragonese y Milano (1984). 165 Fig. Nº 1-10. Mapa de distribución de Solanum sublobatum Willd. “yerba mora”. en la República Argentina. Ragonese y Milano (1984). 166 Las imágenes son presentadas con los nombres que aparecen en documentos publicados en Internet. Fig. Nº 1-11. Solanum chenopodioides Lam Fig. Nº 1-12. Solanum sublobatum : = S. gracile Dun. fleurs et fruits = S. sublobatum Willd. ex Roemer & Schultes Belles Fleurs de France 2 Friche. Dolus-d'Oléron (Charente Maritime) 03-07-06 167 Fig. Nº 1-13. Solanum sublobatum= Solanum chenopodioides Hierba mora (tall nightshade) Vía Rural - Argentina 168 03-07-06 Fig. Nº 1-14. Detalle del haz las hojas de Solanum sublobatum Willd. Vía Rural – Argentina. 04-07-06 169 Fig.Nº 1-15. Detalle del envés de las hojas de Solanum sublobatum Willd. Vía Rural – Argentina. 04-07-06 Solanum bermejense Bitter Son escasas las referencias sobre esta planta. Es una especie nativa descripta por Cabrera (1983) para Bermejo, Departamento de Tarija, en el límite con Salta. Parece ser frecuente en el norte de Salta y en el nordeste de Jujuy. Vegeta en selvas. También para la República Argentina la cita más actual es la del Catálogo de las Plantas. Vasculares de la República Argentina II, (Barboza et al., 1999) que hace referencia a Cabrera (1983). No se encontraron otros datos, esquemas ni registros fotográficos actuales de esta planta. Descripción de A. Cabrera (1983). Solanum bermejense Bitter, Fedde Repert. 12: 87, 1913. Arbusto ramoso de 1 – 1,5 m de altura, con ramas delgadas costadas, pubescentes, con pelos pluricelulares, hojosas hasta el ápice. Entrenudos de 3 - 8 cm de long. Hojas alternas o, las superiores, geminadas, pecioladas, con pecíolo pubescente de 1,5 - 5 cm de long. y lámina herbácea lanceolada, aguda en el ápice y atenuada en la base, entera o, más raramente con uno o dos gruesos dientes cerca de su base, casi glabra en el haz y laxamente pubescente en el envés, con pelos pluricelulares, de 4 - 15 cm de long. por 1 – 3,5 cm de lat. Inflorescencias extra - axilares, bifurcadas, 5 – 15 – flora. Pedúnculo delgado, pubescente, de 1 – 2,5 mm de long., pedicelos de 7 – 12 mm. Cáliz acampanado, pubescente, 1 – 2 mm de alto cortamente pentadentado, con dientes triangulares anchos. Corola blanca de unos 15 mm de diámetro, profundamente pentapartida, con segmentos lanceolados, agudos, corta y densamente pubescentes exteriormente. Anteras con filamentos de 1 mm de long. y tecas oblongas, de alrededor de 4,5 mm de long. Estilo densamente pubescente en sus dos tercios inferiores. Bayas globosas, glabras, de 5 mm de diámetro. Semillas numerosas. Corpúsculos pétreos numerosos. Obs. Especie próxima a Solanum lorentzii, Bitter, del que difiere por las hojas lanceoladas. El número de Corpúsculos pétreos de los frutos parece ser mayor. Tal vez se trate sólo de una variedad. Barboza et al., (1999) en el Catálogo de las Plantas. Vasculares de la República Argentina II, del Instituto Darwinion brinda los siguientes datos: Solanum bermejense Bitter Ref.: Cabrera, 1983. Hábito: Arbusto. Status: Nativa. Elevación: 1000-2000 m. Prov.: JU, SA. Paises limítrofes: BO. Ejemplar examinado: Cabrera et al. 26412 (LP, SI). 170 La ficha que se presenta a continuación tiene la única referencia fotográfica para Argentina. Instituto de Botánica Darwinion Academia Nacional de Ciencias Exactas, Físicas y Naturales Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) Especie: Solanum bermejense Bitter Categoría de Tipo: Isotipo Colector: Fiebrig, K. A. G. 2131 Localidad: Bolivia. Tarija: Bermejo. Coordenadas: Altitud: 1400 m s.m. Fecha de colección: 17/11/1903 Observaciones: Det.: Bitter, F. A. G., 1913 Imágen: sol-ber-001.jpg 171 Fig. Nº 1-8 Ejemplar de herbario de Solanum bermejense Bitter 172 Registro de Malezas de la Familia Solanáceas 1991 - 2001 Este listado comprende a los trabajos realizados en las provincias del NOA, en diferentes cultivos entre los años 1991 y 2001. Tabla Nº 1-4. Registro de Malezas Nombre Científico Año Provincia Datura ferox 2001 Tucumán Departamento Trancas Cultivo ají picante Physalis viscosa Solanum atriplicifolium Solanum conditum Solanum hyeronimi Solanum lorentzii Tucumán Salta 2000 Santiago del Estero Nicandra physalodes 1999 Tucumán Lules Huerta Nicandra physalodes 1998 Tucumán Lules Huerta Datura ferox Nicandra physalodes Nicotiana glauca Nicotiana longiflora Physalis angulata 1997 Salta Physalis pubescens Physalis viscosa Solanum atriplicifolium Solanum chacoense Solanum conditum Solanum hyeronimi Solanum sisymbriifolium Nicandra physalodes Physalis pubescens Solanum atriplicifolium Solanum physalidicalyx Metán - Anta soja Catamarca Santa Rosa Tucumán 1995 Salta Datura ferox Nicandra physalodes Physalis pubescens soja; poroto; algodón 1995 Tucumán La Cocha; Graneros; J. B. Alberdi; Cruz Alta; Burruyacu granos Metán; Anta Burruyacu; Cruz Alta; Leales soja Nicotiana longiflora 173 Nierembergia hippomanica 1995 Tucumán Datura ferox Nicandra physalodes Tucumán Physalis angulata 1993 Physalis pubescens Solanum atriplicifolium Salta Solanum chacoense Solanum sisymbriifolium Lules La Cocha; Graneros; J. B. Alberdi Metán; Anta Huerta soja; maíz; poroto; sorgo Cestrum parqui Datura ferox Nicandra physalodes Catamarca Santa Rosa Solanum adventitium Solanum atriplicifolium Solanum chacoense 1993 Solanum chacoense Solanum chenopodioides Tucumán La Cocha; Graneros; J. Solanum nitidibaccatum B. Alberdi Solanum pigmaeum Solanum pilcomayense Solanum sisymbriifolium soja Cestrum parqui Tucumán La Cocha; Graneros; J. Datura ferox 1991 B. Alberdi Lycium cestroides Nicotiana longiflora Catamarca Santa Rosa Nierembergia gracilis Solanum sisymbriifolium trigo Datura ferox Nicandra physalodes Nicotiana longiflora Catamarca Physalis angulata Physalis pubescens 1998 Tucumán Physalis viscosa Solanum atriplicifolium Solanum chacoense Salta Solanum lorentzii Solanum physalidicalyx Solanum sisymbriifolium soja Santa Rosa La Cocha; Graneros; J. B. Alberdi Metán; Anta 174 Resultan de estos relevamientos en la zona NOA 22 géneros de la Familia Solanáceas y 12 de estas son del género Solanum. No se identificó entre éstas a Solanum nigrum. 175 CAPÍTULO 2 ASPECTOS TAXONÓMICOS DEL Complejo Solanum nigrum Introducción La Familia Solanáceas ha sido extensamente estudiada, agrupada y reagrupada, ordenada y reordenada, pero por sus caracteres variables y complejos no es un taxón en el que se pueda llegar a tener datos concluyentes. La Taxonomía agrupa plantas sobre la base de similitudes y diferencias, que son expresiones de parentesco filogenético actual. Establece y ordena a los grupos de plantas emparentadas entre sí poseedoras de características comunes (Marzocca, 1985). Para analizar todos los caracteres botánicos de este Complejo de plantas se han utilizado los métodos taxonómicos clásicos (Subils, 1977; Botta, 1979). En las provincias del Noroeste Argentino se han registrado a lo largo de una década aproximadamente la presencia de malezas Solanáceas en diferentes cultivos (de Andrada et al., 1995; Mansilla et al., 1998; Roncaglia et al., 1991, 1993 a, 1993 b, 1995, 1998, 2000). En estos relevamientos no aparece Solanum nigrum debido a que es posible que estas plantas no estuvieron correctamente identificadas. Por ello, el objetivo de esta parte del trabajo es seleccionar los caracteres taxonómicos que permitan identificar a las especies de malezas que componen el Complejo Solanum nigrum en el área agrícola de la Provincia de Tucumán. 2.1. MATERIALES Los ejemplares utilizados en este estudio provienen de las regiones agroecológicas del área agrícola de la provincia de Tucumán. En la mayor parte de las tareas se trabajó con materiales frescos. 2.2. MÉTODOS Para determinar los aspectos taxonómicos propuestos se utilizaron métodos clásicos que evalúan: a.- caracteres morfológicos, b.- caracteres anatómicos y c.- caracteres cariológicos (Subils, 1977; Botta, 1979). Se consideraron en total 64 caracteres morfológicos para evaluar a las especies del Complejo Solanum nigrum. En el aspecto vegetativo se utilizaron 16 caracteres (7 caracteres para ramas y 9 para hojas), en el reproductivo 48 caracteres (9 en las inflorescencias, 24 en las flores, 9 en los frutos y 6 en las semillas) considerándose dimensiones (en cm y mm), formas, colores y presencia o ausencia de detalles (pubescencia, dientes, crestas, etc.). Además, se tuvo en cuenta 2 caracteres citológicos y 1 cariológico (Tabla Nº 2-1) . Además de los caracteres propuestos en el proyecto se agregaron otros por considerarlos relevantes. 176 En el análisis estadístico se utilizó el programa SSPS y Microsoft Excel 2.3. CARACTERES MORFOLÓGICOS Se utilizó material vegetal fresco y material herborizado para estudiar los caracteres morfológicos. En algunos casos se analizó la planta completa; en otros se eligieron partes de plantas bien desarrolladas (no en los brotes). Los materiales frescos fueron observados en la totalidad del ciclo de vida de la planta. La observación se realizó con lupa de mano, lupa binocular (Marca Carl Zeiss 100x), microscopio Modelo Axiolab de polarización (Marca Carl Zeiss 100x); la disección con agujas, pinzas, hojas de afeitar y espátula de arrastre y en las mediciones se utilizaron ruleta, reglas (comunes y de precisión) y compás de dos puntas. Se emplearon entre 50-100 ejemplares de cada especie. 1. Número: Se dio un número de identificación para cada ejemplar estudiado que no cambia si este desaparece (se seca, pudre, lo destruyen los insectos o lo machetean). 2. Localización y Fecha: Se registró la ubicación geográfica, fecha de recolección o de observación y datos del estado fenológico de cada planta. Referencias: V: vegetativo F: floración Fr: fructificación 3. Ciclo: Se observó el desarrollo de las plantas en el campo y en las llevadas a invernáculo, desde la germinación o trasplante hasta su muerte. Referencias: A: anual P: perenne SP: semiperenne 2.3.1. ASPECTO VEGETATIVO Se consideraron 16 caracteres vegetativos de las plantas jóvenes hasta el inicio de la floración. 4. Hábito: Se observó el hábito de crecimiento de las diferentes especies: hierba, arbusto, subarbusto y si son erguidas o decumbentes. Referencias 1. hierba 2. arbusto 3. subarbusto 4. erguida 5. decumbente 177 Tallo. (7 caracteres) 5. Altura: Se midió la altura de 110 plantas en metros (m) desde el ras del suelo hasta el borde superior de la copa. SOLNI: de un total de 68 ejemplares medidos, 7 eran partes de plantas. SOLLO: se midieron 42 plantas completas SOLNI-I: en las plantas con caracteres intermedios se repitió el número correspondiente a SOLNI. 6. Ramificación: Para describir el tipo de ramificación se observó la forma en que las plantas distribuyen sus ramas. 1. Presentan un tallo único que se bifurca en dos ramas. 2. Se ramifican desde la base del tallo. 7. Altura de la bifurcación de las ramas: Utilizando una ruleta se midió la altura en que el tallo se bifurca en metros (m). 8. Diámetro del tallo: Con compás de dos puntas y regla se midió el diámetro del tallo principal, por debajo de la bifurcación de las ramas, en centímetros (cm). 9. Observación de detalles del tallo: a) Estrías: Se contó el número de crestas de las estrías presentes en el tallo. 1. sin estrías 2. 3-4 crestas 3. más crestas Se observó la amplitud entre éstas. 1. espacios estrechos entre las crestas 2. espacios amplios entre las crestas b) Pubescencia: La observación de la pubescencia se hizo en forma directa y con lupa binocular. 1. glabra 2. pubescente 3. muy pubescente El tipo de pubescencia sólo se observó con lupa binocular. 1. pelos simples 2. pelos glandulosos 3. pelos de otro tipo 10. Hojas: (9 caracteres) Por las características especiales que posee la familia Solanáceas (Concaulescencia y recaulescencia) se observaron las hojas bien desarrolladas que se encontraban lejos del ápice de la planta. 178 Algunos datos fueron tomados de la planta completa y otros, de las partes cortadas y herborizadas, al azar. Se observó: - la disposición (filotaxis) de las hojas en el tallo: 1. alternas 2. opuestas - el tipo de hoja: 1. simple 2. compuesta - la forma de la lámina: 1. oval terminada en punta 2. oval – romboidal 3. lámina decurrente 4. lámina no decurrente 5. otra - la forma de la base de la lámina: 1. base simétrica 2. base asimétrica 3. otra Se anotaron las dimensiones de las hojas en cm: - Largo de la lámina - Ancho de la lámina - longitud pecíolo En el borde de la lámina se observaron los diferentes tipos: 1. liso 2. algo ondulado, casi liso 3. liso o ligeramente dentado 4. liso o ligeramente ondulado 5. presencia de dientes notables 2.3.2. ASPECTO REPRODUCTIVO Se consideraron 48 caracteres en plantas con flores y frutos desde el inicio de floración hasta el fin de su ciclo. Inflorescencias (9 caracteres) 11. Posición: Se observa y registra a dónde nacen las inflorescencias y su posición en la rama, con respecto a una hoja: 1- axilares 2- extraaxilares 179 12. Tipo: Se describió el tipo de inflorescencia 1- cimosa subumbelada sin bifurcación 2- cimosa subumbelada con bifurcación 3- cimosa bifurcada en planta no bifurcada 13. Número de inflorescencias: Se contó el número de inflorescencias y el número de flores por plantas en ejemplares adultos. 14. Número de flores: Se contó el número de flores de cada inflorescencia. 15. Pedúnculo: Se midió la longitud del pedúnculo de la inflorescencia (cm) en muestras de 50 inflorescencias en SOLNI y SOLLO y 5 en la forma intermedia. 16. Pedicelo: Se midió la longitud de los pedicelos en las ramas de las inflorescencias bifurcadas (mm). 17. Bifurcación: Se registró si las inflorescencias son: 1. bifurcadas 2. no bifurcadas 3. otras 18. Ramas de la bifurcación: Se midió la longitud de las ramas de la bifurcación de la inflorescencia (mm) identificándolas como rama izquierda (I) y (D) rama derecha. Se contaron las flores de cada una de las ramas. Flores (24 caracteres) 19. Cáliz: Tipo: Se describe el tipo de cáliz. Número: Se contó el número de piezas del cáliz. Forma: Se describió la forma del cáliz y la disposición de los sépalos: 1: reflexo 2: normal (no reflexo) Longitud en mm: La longitud de cada sépalo se midió en mm. Dientes: Se contó el número de dientes del cáliz, se observó la forma de éstos dientes, y se midió su longitud en mm. 20. Corola: Tipo: Se describió el tipo de corola. 180 Forma: Se observó la forma de la corola. Número de pétalos: Se contó el número de piezas de la corola. Color: Se anotó el color de las flores. 1: blanca 2: blanca con el centro morado Diámetro: Con un compás de dos puntas se midió el diámetro (mm). Tamaño: Con un compás de dos puntas se midió la longitud de los pétalos (mm). Lóbulos de los pétalos: Se describió la forma de los lóbulos 1: terminados en punta 2: romos 3: otros Se observó la profundidad de las escotaduras: 1. sin escotaduras 2. escotaduras poco profundas 3. escotaduras profundas Con compás de dos puntas y regla de precisión se midió la longitud de los lóbulos (mm). 21. Androceo: Número de piezas: Se detalló el número de estambres. Tipo: Se observó el tipo de androceo. Tamaño: Con compás de dos puntas y regla de precisión se midieron la totalidad de las partes de los estambres. Filamentos: Se midió ancho y largo de los filamentos (mm). Anteras: Se midió ancho y largo de estas (mm). Pubescencia: Se observó la presencia de pelos. 1. glabra 2. pubescente 3. muy pubescente Se observó el tipo de pubescencia 1. pelos simples 2. pelos glandulosos 3. otro tipo 22. Gineceo: 181 Se observó la posición del gineceo, forma del ovario, características del estilo y pubescencia. 23. Fruto (9 caracteres) Tipo de fruto: Se describió el tipo de fruto. Número de infrutescencias por planta: Se contó el número de infrutescencias. Número de frutos por infrutescencia: Se contó el número de frutos por infrutescencia. Número de frutos por planta: Se calculó el número de frutos de la totalidad de inflorescencias en una planta. Diámetro en mm: Se midió el diámetro de los frutos (mm) considerando el mayor debido a que no son perfectamente esféricos. Color de los frutos: Se describió el color de las bayas. 1: negro brillante 2: morado opaco Color de la pulpa: Se observó el color de la pulpa y del líquido. 1: morado 2: verde Descripción del cáliz en el fruto maduro: Se describió la forma del cáliz persistente en el fruto y la disposición de los sépalos. 1: reflexo 2: normal (no reflexo) Longitud de los sépalos en el fruto maduro, medido en mm 24. Semilla (6 caracteres) Número de semillas por fruto: Se contaron las semillas que contienen por fruto. Forma: Se describió la forma de las semillas. Tamaño: (Ancho-Largo) Con regla de precisión se midieron el ancho y largo de las semillas (en mm). Peso: Con tamices de diferentes tamaños se hizo separación de muestras de semillas y se pesaron las diferentes fracciones. Además, se pesaron semillas de 20, 30, 50 y 100 frutos y muestras de 50 semillas en balanza granataria (Marca: Sartorius 220 P – Sensibilidad 0,1) y en balanza de precisión (Marca: Mettler, Tipo H6). 182 Color: Se describió el color de las semillas extraídas de frutos maduros. 1: Marrón claro 2: Marrón oscuro 2.4. CARACTERES ANATÓMICOS En frutos completamente maduros de las especies del Complejo Solanum nigrum se registró: a.- la presencia (1) y la ausencia (2) de corpúsculos pétreos en el fruto. b.- el número de corpúsculos pétreos por fruto. c.- la disposición de éstos en la pulpa del fruto (cortes longitudinales y transversales). Para determinar la naturaleza de los corpúsculos pétreos, extraídos y lavados con agua de red, se utilizó safranina La caracterización de los corpúsculos pétreos se hizo con la colaboración de la Cátedra de Geología Estructural de la Facultad de Ciencias Naturales de la U.N.T.. 183 2.5. CARACTERIZACIÓN CITOLÓGICA: Con la colaboración de la Cátedra de Genética de la Facultad de Agronomía y Zootecnia se realizó el análisis de los cromosomas mitóticos y de los meióticos para caracterizar citológicamente a estas especies. 1: Solanum nigrum Los preparados microscópicos para la observación de los cromosomas somáticos se realizaron en meristemas de raíz y de hojas jóvenes. Se pretrataron los mismos en una solución saturada de para-di-clorobenceno durante 2 horas 15 minutos, luego se fijaron en Etanol – acético 3:1. Los preparados microscópicos se obtuvieron hidrolizando el material en HCl 1 N a 58ºC durante 5 minutos, y luego se aplastó entre porta y cubreobjetos. se utilizó como colorante hematoxilina al 2% y citrato férrico al 1 % como mordiente. Para la observación de los cromosomas en meiosis se fijaron anteras jóvenes en una solución de Newcomer. Se observaron las distintas fases de la meiosis en las células madre del grano de polen (CMP), utilizándose la técnica de coloración empleada para los cromosomas somáticos. (Andrada et al., 2003). 2: Solanum lorentzii Para el análisis de los cromosomas en meiosis de Solanum lorentzii se fijaron flores en una solución de Newcomer y en la realización de los preparados microscópicos se utilizó la técnica de coloración con hematoxilina al 2%, empleando citrato férrico al 1% como mordiente. 3: Intermedia Con el objeto de dilucidar aspectos relacionados con la identidad de estas especies se realizó un estudio de la morfología de sus inflorescencias y un análisis de sus respectivos complementos cromosómicos, para lo cual se compararon los cromosomas mitóticos y meióticos de formas intermedias con los de Solanum nigrum y de Solanum lorentzii. (Mansilla de Andrada et al., 2004). Los caracteres propuestos para el análisis y su codificación se detallan en la Tabla Nº 2-1. 184 Tabla 2-1: Lista de Caracteres, con sus respectivos Estados y su Codificación. 1- Ciclo (C) Caracteres 2- Estado fenológico (EF) 3- Porte (P) y Hábito de crecimiento(HC) 4- Altura (A) 5- Ramificación (R) 6- Altura de bifurcación del tallo(AB) 7- Diámetro del tallo (D) 8- Estrías (E) Número y amplitud entre crestas 9- Pubescencia (Pb) 10- Tipo de pelos (TP) 11- Filotaxis (F) 12- Tipo de hoja (TH) 13- Forma de la lámina (FL) 14- Decurrencia (DR) 15- Forma de la base de la lámina (FBL) 16- Borde de la lámina (BL) 17- Longitud de la lámina (LL) 18- Ancho de la lámina (AL) 19- Longitud del pecíolo (LP) 20- Posición de la inflorescencia en la rama con respecto a las hojas (PIR) 21- Tipo de inflorescencia (TI) Estados Anual Perenne Vegetativo Floración Fructificación Hierba Arbusto Subarbusto Erguida Decumbente En m Tallo único Ramificado desde la base En m En cm Sin estrías 3-4 crestas Más crestas Espacios estrechos entre crestas Espacios amplios entre crestas Glabra Pubescente Muy pubescente Simples Glandulosos Otro tipo Alterna Opuesta Simple Compuesta Oval terminada en punta Oval punta roma Oval romboidal Lámina decurrente Lámina no decurrente Base simétrica Base asimétrica Otra Liso Algo ondulado, casi liso Liso o ligeramente dentado Liso o ligeramente ondulado Presencia de dientes notables Sin dientes notables En cm En cm En cm Axilar Extraxilar Cimosa no bifurcada Cimosa bifurcada Codificación 1 2 1 2 3 1 2 3 4 5 1 2 1 2 3 4 5 1 2 3 1 2 3 1 2 1 2 1 2 3 1 2 1 2 3 1 2 3 4 5 6 1 2 1 2 185 22- Posición de la inflorescencia (PI) 23- Número de inflorescencias por planta (IP) 24- Número de flores por planta (FP) 25- Número de flores por inflorescencia (FI) 26- Longitud del pedúnculo (LPD) 27- Longitud del pedicelo (LPC) 28- Longitud de las ramas de bifurcación de la inflorescencia (LBI) 29- Tipo de cáliz (KT) 30- Forma del cáliz (KF) 31- Número de los sépalos (KN) 32- Número de dientes del cáliz (KND) 33- Longitud del cáliz (KL) 34- Forma de los dientes del cáliz (KFD) 35- Longitud de los dientes (KLD) 36- Tipo de corola (CT) 37- Forma de la corola (CF) 38- Número de piezas de la corola (CN) 39- Color de la Corola (CC) 40- Diámetro de la Corola (CD) 41- Tamaño de los pétalos Long.– Ancho (CL) 42- Forma lóbulos de los pétalos (CFP) 43- Profundidad de las escotaduras (CE) 44- Tipo de androceo (AT) 45- Número de piezas del androceo (AN) 46- Longitud de los estambres (AL) 47- Ancho del filamento (AAF) 48- Largo del filamento (ALF) 49- Pubescencia del androceo (AP) 50- Tipo de pubescencia (APT) 51- Posición del gineceo (GT) 52- Longitud del gineceo (GL) 53- Tipo de fruto (FT) Cimosa bifurcada en planta no bifurcada Erecta Nutante 3 1 2 En cm En mm En mm Gamosépalo Dialisépalo Reflexo No reflexo En mm Deltoidea, punta roma Deltoidea, punta roma aguda En mm Gamopétala Dialipétala Rotácea Otra Blanca Blanca con el centro morado Otro En mm En mm Terminados en punta Romos Otros Sin escotaduras Escotaduras poco profundas Escotaduras profundas Anteras conniventes Anteras no conniventes En mm En mm En mm Glabra Pubescente Muy pubescente Pelos simples Pelos glandulosos Otro tipo Súpero Ínfero No sobresale del androceo Sobresale del androceo Baya Otro tipo 1 2 1 2 1 2 1 2 1 2 1 2 3 1 2 3 1 2 3 1 2 1 2 3 1 2 3 1 2 1 2 1 2 186 54- Número de infrutescencias por planta (FIP) 55- Número de frutos por infrutrescencia (FNI) 56- Número de frutos por planta (FNP) 57- Diámetro de los frutos (FD) 58- Color de los frutos (FC) 59- Color de la pulpa y del jugo(FCP) 60- Forma del cáliz en el fruto maduro (FFK) 61- Longitud de los sépalos (FKL) 62- Número de semillas por fruto (SN) 63- Forma de la semilla (SF) 64- Ancho de la semilla (SA) 65- Largo de la semilla (SL) 66- Color de semillas en el fruto maduro (SC) 67- Peso de las semillas (SP) 68- Corpúsculos pétreos (CP) 69- Número de corpúsculos pétreos por fruto (NCP) En mm Negro brillante Morado opaco Morado Verde Reflexo No reflexo En mm 1 2 1 2 1 2 Reniforme Redondeada Otra En mm En mm Marrón claro Marrón oscuro En balanza de precision 1 2 3 Ausentes Presentes 1 2 1 2 70- Número cromosómico (NCr) REFERENCIAS: CARACTERES MORFOLÓGICOS 1 a 19 CARACTERES VEGETATIVOS 20 a 67 CARACTERES REPRODUCTIVOS CARACTERES ANATÓMICOS 68 a 69 CARACTERES CITOLÓGICOS 70 cada uno consta de: N º de orden del carácter Nombre del carácter taxonómico Codificación (C). 187 2.6. RESULTADOS Y DISCUSIÓN Con los métodos clásicos de identificación (Subils, 1977; Botta, 1979), en cada ejemplar de Solanum, se reconocieron 64 caracteres morfológicos, 1 anatómico y 1 carácter citológico. Entre los morfológicos se obtienen 16 caracteres vegetativos y 48 reproductivos que se analizaron con el propósito de lograr la diferenciación de las especies del Complejo. Todos los caracteres sirven para identificar estas especies pero sólo algunos para diferenciarlas. Estos últimos se remarcarán a medida que se muestren los resultados. Para nombrar a las especies de este Complejo se utiliza la codificación internacional propuesta por la Weed Science Society of America (1984), SOLNI= Solanum nigrum, SOLLO= Solanum lorentzii, SOLNI-I forma intermedia de la dos especies anteriores. Se analizaron las variables cualitativas y cuantitativas. Los resultados obtenidos para los aspectos taxonómicos de este Complejo de Solanáceas se los comparó con las referencias de todos los autores consultados a lo largo de la investigación. Debido a que el objetivo es descriptivo, los datos de las variables cuantitativas fueron analizados con los procedimientos estadísticos propuestos por la bibliografía. Se hicieron: cálculo de los principales indicadores estadísticos (promedio y error estándar de la media principalmente); la representación gráfica de las mismas (barra de error de la media) y el análisis de la varianza de una vía de clasificación, cuando se comparaban los tres grupos de las especies del Complejo Solanum nigrum. Para los procedimientos estadísticos de utilizó el paquete estadístico SSPS y la planilla de cálculo Excel. CARACTERES MORFOLÓGICOS CARACTERES VEGETATIVOS 1- Ciclo (C): Diferentes autores citan a Solanum nigrum como especie de ciclo anual (Roberts y Lockett, 1978; Ogg et al., 1989; Devani et al., 1993; Edmonds y Chweya, 1997); otros la consideran anual a bienal Holm et al. (1977) y para Arévalo (1975) es perenne. Solanum lorentzii es citada sólo como especie perenne (Morton, 1976; Cabrera, 1983; Roncaglia et al., 2000). Del seguimiento realizado a lo largo del ciclo de vida de las especies de Solanum del Complejo Solanum nigrum se comprobó que: SOLNI se comporta generalmente como anual y en algunos casos es perenne. SOLLO siempre se comporta como perenne. SOLNI-I En las plantas donde se observó la forma intermedia corresponde decir lo mismo que SOLNI. 188 En la provincia de Tucumán las observaciones coinciden con las referencias bibliográficas. Se debe aclarar que cuando Solanum nigrum extiende su ciclo y se considera perenne, no produce flores ni frutos, es decir, sólo vegeta. 2- Estado fenológico (E F): El Complejo Solanum nigrum ha sido ampliamente estudiado en el hemisferio Norte por Roberts y Lockett (1978), Ogg et al. (1989), Keeley y Thullen (1989, 1991) y Edmonds y Chweya (1997). Estos autores han demostrado que en condiciones de laboratorio e invernáculo los resultados sobre germinación, emergencia y etapa reproductiva del Complejo Solanum nigrum, en general, son semejantes a lo que ocurre en el campo. También sostienen que predominan diferentes especies de este Complejo según las regiones. En el hemisferio Sur no se encontraron referencias sobre la biología de Solanum nigrum y las especies relacionadas. El seguimiento realizado del ciclo de vida de las especies de Solanum del Complejo Solanum nigrum en el área de estudio, se muestra en Tabla Nº 3-33 del Cap. 3. 3- Porte (P) y Hábito de crecimiento (HC): Del porte de Solanum nigrum la bibliografía dice que pueden ser hierbas o arbustos (Holm et al., 1977; Edmonds y Chweya, 1997); del hábito de crecimiento, erguidas o decumbentes (Morton, 1976; Goodhue, 1998). Para Morton (1976) Solanum lorentzii es un arbusto erecto, mientras que para Cabrera (1983) es erecto o apoyante. Estas plantas en la zona estudiada, son generalmente hierbas erguidas, pudiendo volcar en zonas con mucha humedad o cuando la densidad de plantas es muy elevada, pero esto no es lo normal. Las plantas rebrotan cuando el crecimiento es alterado y toman aspecto de arbusto. 4- Altura (A): Las alturas registradas en 50 – 100 plantas completas, en estado reproductivo se muestran en la Tabla Nº 2-2, el análisis estadístico en las Tablas Nº 2-3, 2-4 y están representadas en la Figura N°2-1. Estos datos coinciden con los de la bibliografía consultada en Solanum nigrum Holm et al, (1977) es de 0,3 – 0,9 m y para Goodhue (1998) 0,5 m. En Solanum lorentzii con las medidas de las descripciones de Morton (1976) 1- 1,5 m y de Cabrera (1983) 1-2 m. Tabla Nº 2-2. Altura de las plantas (m) del Complejo Solanum nigrum Altura (m) m M 0,11 1,50 SOLNI 0,49 0,82 SOLNI-I 0,56 2,20 SOLLO m: altura menor – M: altura mayor Promedios 0,66 0,66 1,25 189 Figura N° 2-1: Altura de las especies del Complejo Solanum nigrum Tabla N° 2-3: Análisis de las alturas del Complejo Solanum nigrum Altura de Planta Inter-grupos Intra-grupos Total SC 8,870 14,156 23,026 gl 2 103 105 CM 4,435 ,137 F 32,271 Sig. ,000 Tabla N° 2-4: Pruebas estadísticas para el análisis de altura de planta en las distintas especies del Complejo Solanum nigrum Su bcon juntos Homo geneos seg ún Altura de Plan ta a,b HSD de Tukey ESPECIE SOLNI SOLNI-I SOLLO Sig. N 59 6 41 Subconjunto para alf a = .05 1 2 ,6586 ,6650 1,2532 ,999 1,000 a. Usa el tamaño muestral de la media armónica = 14,423. b. Los tamaños de los grupos no son iguales. Se utilizará la media armónica de los tamaños de los grupos. Los niv eles de error de tipo I no están garantizados. La altura coincide con las referencias bibliográficas y la altura mayor corresponde a Solanum lorentzii. 190 En general la altura depende de los factores ambientales (humedad y temperatura) y de la altitud. En zonas áridas, la altura es menor, lo que coincide con lo que ocurre en Tucumán en laderas con exposición al oeste. 5- Ramificación (R): Se observaron ejemplares no ramificados y ramificados con aspecto de arbustos. Los porcentajes se presentan en la Tabla Nº 2-5 y los números corresponden a lo propuesto en Materiales y Métodos. Tabla N º 2-5. Ramificación de las plantas del Complejo S. nigrum (%) SOLNI SOLNI-I SOLLO No Ramificados 1 48 10 38 Porcentajes % 81,35 100 92,58 Ramificados 2 11 0 1 Porcentajes % 18,64 0 2,43 Las plantas observadas casi siempre presentan un tallo único que se bifurca en dos ramas a veces se ramifican desde la base con aspecto de arbustos. En Holm et al. (1977) y en Morton (1976) se describen las dos posibilidades, pero se debe destacar que las plantas ramificadas desde la base, aparentemente son las que fueron macheteadas o rotas por alguna maquinaria o producto químico, y rebrotaron. No se observaron plantas que rebrotaran cuando fueron comidas por animales. 6- Altura de la bifurcación del tallo (AB): La variación de la altura de la bifurcación en metros (m) se muestra en Tabla Nº 2-6. Tabla N º 2-6. Alturas y promedios de la bifurcación del tallo en el Complejo Solanum nigrum SOLNI SOLNI-I SOLLO Altura (m) Promedios m M 0,115 0,93 0,40 0,28 0,48 0,39 0,46 1,24 0,90 m: altura menor – M: altura mayor No se encontraron datos a cerca de la altura donde el tallo se bifurca. En los tomados sólo en plantas completas se observó que la altura de la bifurcación varía, dependiendo del lugar a donde se desarrolla. Si ocurre debajo de otras plantas, el tallo se alarga buscando luz y la bifurcación ocurre a mayor altura; pero en otras condiciones, hay diferencias notables en este detalle. Como era de esperar, la bifurcación ocurre a mayor altura en Solanum lorentzii. El análisis estadístico de los datos se encuentra en Anexo Capítulo 2, Pág. 93. (Fig. Nº 2-2 y Tablas Nº 2-7 y 2-8). 7 - Diámetro (D): 191 Sobre el diámetro del tallo medido debajo de la bifurcación de las ramas tampoco se encontraron referencias en la bibliografía. Posiblemente porque si bien hay diferencias en las especies observadas, estas dimensiones no son importantes. El diámetro del tallo medido debajo de la bifurcación, se observa en Tabla Nº 2-9. El análisis estadístico de los datos en Fig. Nº 2-3 y Tablas Nº 2-9 y 2-10. Tabla N º 2-9. Diámetro del tallo en cm del Complejo Solanum nigrum Diámetro (cm) Promedios m M 0,30 1,40 0,5726 SOLNI 0,50 0,70 0,4817 SOLNI-I 0,20 2,50 0,8207 SOLLO m: diámetro menor – M: diámetro mayor IC-95% del Diametro del Tallo (mm) 1,2 1,0 ,8 ,6 ,4 ,2 N= 6 43 41 SOLNI -I SOLNI SOLLO Figura N°2-3. Diámetro de tallo de las especies del Complejo Solanum nigrum Tabla N° 2-9. Análisis de diámetro de tallo del Complejo Solanum nigrum Diametro del Tallo (mm) ESPECIE SOLNI SOLLO N 43 41 Media ,5726 ,8207 DE ,25128 ,54886 EE ,03832 ,08572 Tabla N° 2-10. Pruebas estadísticas para el análisis del diámetro de los tallos en las distintas especies del Complejo Solanum nigrum 192 Prueba de Lev ene para la igualdad de v arianzas Diametro del Tallo (mm) Se han asumido v arianzas iguales No se han asumido v arianzas iguales F 12,882 Sig. ,001 Prueba T para la igualdad de medias t Sig. (bilateral) gl -2,685 82 ,009 -2,643 55,476 ,011 193 8- Estrías (E): Todos los ejemplares observados presentaban pocas estrías (3 - 4 crestas) (Tabla N º 2-11). Son escasamente estriados, con espacios amplios entre ellos. En esta característica se coincide con los diferentes autores sobre la presencia de estrías o crestas en los tallos en Solanum nigrum y Solanum lorentzii, (Morton, 1976; Holm et al., 1977; Edmonds y Chweya, 1997), por lo que este carácter sirve para la identificación pero no se usará para hacer una diferenciación. Tabla N º 2-11. Porcentaje de número de estrías del tallo en el Complejo Solanum nigrum SOLNI SOLNI-I SOLLO Sin estrías 1 0% 0% 0% 3 – 4 crestas 2 100 % 100 % 100 % más crestas 3 0% 0% 0% 9- Pubescencia (Pb): Este carácter es propio de la familia Solanáceas en la que se observan pelos de formas variadas. En la observación directa y con lupa de mano del Complejo Solanum nigrum se notó que la pilosidad es muy tenue. En general la pubescencia es rala, aumentando la densidad en zonas de mayor aridez, sin llegar a considerarse “muy pubescente” como se informa en trabajos de Dieckmann (1912), Morton (1976) y Cabrera (1983). En nuestra región se ha observado este detalle y está relacionado con las condiciones ambientales. Los porcentajes se muestran en la Tabla N º 2-12. Esta característica no se usará para diferenciar a las especies. Tabla N º 2-12. Porcentaje de presencia de pubescencia en el Complejo Solanum nigrum SOLNI SOLNI-I SOLLO glabra 1 0% 0% 0% pubescente 2 100 % 100 % 100 % muy pubescente 3 0% 0% 0% 10- Tipo de pelos (TP): La lupa binocular muestra en todos los órganos del Complejo Solanum nigrum una pubescencia de pelos simples, cortos, a veces muy ralos, detalles que lógicamente no son visibles a simple vista. Los datos del tipo de pubescencia tampoco son importantes para hacer una diferenciación. Tabla Nº 2-13. Tabla N º 2-13. Porcentaje de Tipo de pubescencia en el Complejo Solanum nigrum SOLNI SOLNI-I SOLLO pelos simples 1 100 % 100 % 100 % pelos glandulosos 2 0% 0% 0% pelos de otro tipo 3 0% 0% 0% 194 En la Fig. Nº 2-4 se muestran las crestas y los pelos del tallo de Solanum nigrum. Crestas o estrías Pelos simples Fig. Nº 2-4. Pelos simples y estrías en el tallo de SOLNI 195 HOJAS : Se observó en coincidencia con lo determinado por Morton (1976) y Cabrera (1983) que las hojas, que son los órganos siempre visibles en las plantas, son simples, de disposición alterna, de forma oval y lámina decurrente; la nervadura principal es muy marcada; el color es verde oscuro en el haz y un poco más claro en el envés. (Figuras 2-5 a 2-8). La base de la lámina es levemente asimétrica. Esta característica observada en todas las hojas de las especies del Complejo no está citada en la bibliografía. Fig. 2-9. El gran dimorfismo foliar se debe a los distintos bordes que presentan las hojas. Ogg et al. (1989) afirman que es un carácter estacional, pero en esta región se lo ha observado en todas las estaciones del año. El dimorfismo foliar es un poco más marcado en SOLNI como se observa en la Fig. 2-5 y Fig. 2-6. Debido al fenómeno de concaulescencia y recaulescencia, que ocurre en la Familia Solanáceas, las medidas se tomaron en las hojas maduras de plantas vivas. Los materiales conservados en los herbarios generalmente son partes de plantas. En éstos, se observó una gran variación de formas: lineares a muy anchas, con borde entero o con bordes lobulados, con 1-5 lóbulos por lado o con dientes muy notables y con idéntica distribución. Se evaluó una gran variedad de dimensiones y se registraron formas variadas. La variación en las mediciones de las hojas depende fundamentalmente de las condiciones ambientales y no es un carácter que permita la diferenciación de especies como expresan Cabrera (1983) y Long (2002). Los datos que se obtuvieron en la observación y mediciones de las hojas (11 a 19) no muestran diferencias que sirvan para caracterizar o diferenciar a las especies del Complejo Solanum nigrum. Sólo se realizó el análisis estadístico de las dimensiones de las hojas. Las observaciones realizadas en plantas con hojas bien desarrolladas, lejos del ápice, mostraron las siguientes características: 11- Filotaxis (F): la disposición siempre es alterna. Tabla N º 2-14 Tabla N º 2- 14. Porcentaje de la filotaxis en el Complejo Solanum nigrum SOLNI SOLNI-I SOLLO Alternas 1 100 % 100 % 100 % Opuestas 2 0% 0% 0% 12- Tipo (TH): siempre es simple. Tabla N º 2-15. Tabla N º 2-15. Porcentaje del Tipo de hojas del Complejo Solanum nigrum SOLNI Simple 1 100 % Compuesta 2 0% 196 100 % 100 % SOLNI-I SOLLO 0% 0% 13- Forma de la lámina (FL) – 14- Decurrencia (DR) - 15- Forma de la base de la lámina (FBL): En una misma planta aparecen hojas con diferentes formas (dimorfismo foliar). Por ello, no pueden ser expresados los porcentajes de las formas. Estas son ovales terminadas en punta u ovales romboidales, siempre con la base de la lámina levemente asimétrica y con lámina decurrente. Tablas Nº 2-16 y 2- 17, Figuras 2-10 a 2-13. Tabla N º 2- 16. Forma de las hojas y Decurrencia en el Complejo Solanum nigrum SOLNI SOLNI-I SOLLO oval terminada en punta 1 oval –romboidal lámina decurrente 3 100 % 100 % 100 % 2 lámina no decurrente 4 0% 0% 0% Otra 5 Tabla N º 2-17. Porcentaje de la Forma de la base de la lámina en el Complejo Solanum nigrum SOLNI SOLNI-I SOLLO base simétrica 1 0% 0% 0% base asimétrica 2 100 % 100 % 100 % Otra 3 0% 0% 0% 16- Borde de la lámina (BL): Los tipos de bordes que se observaron son variados y en muchos casos todos ocurren en una sola planta. Tampoco se pueden calcular los porcentajes de la forma de los bordes. (Figuras Nº 2-5 a 2-8) . En estas fotografías se muestran detalles de las diferentes formas de las hojas que tiene cada planta. Dimensiones de las Hojas: Las medidas del largo, el ancho de lámina y del pecíolo se expresan en cm. Algunos datos son de la planta completa, otros de las partes cortadas y herborizadas. Los gráficos y análisis correspondiente a estos datos se encuentra en Anexo Capítulo 2, Pág. 94 Figuras 2-14 a 2-16 y Tablas Nº 19 y 20. 17- Longitud de la lámina (LL) 18- Ancho de la lámina (AL) 19- Longitud del pecíolo (LP) (cm) se observan en la siguiente Tabla. Tabla N º 2- 18. Dimensiones de las hojas en cm en el Complejo Solanum nigrum Largo de la hoja Ancho de la hoja SOLNI- SOLNI-I m M 2,9 9,3 1,7 4,2 promedios 5,60 2,82 m 3,3 0,7 SOLLO M 10,5 4,9 promedios 5,41 2,14 197 Largo del pecíolo m:menor – M: mayor 0,7 3,1 1,43 0,4 2,7 1,02 198 Dimorfismo foliar en plantas del Complejo Solanum nigrum Figura Nº 2-5. Aspecto vegetativo de SOLNI. Figura Nº 2-7. Aspecto vegetativo de SOLLO. Figura Nº 2-6. Detalles de la forma y el borde de las hojas en SOLNI. Figura Nº 2-8. Detalle de la forma y el borde de las hojas. 199 Figura 2-9. Base de la lámina levemente asimétrica y lámina decurrente. La misma planta de SOLNI muestra gran dimorfismo foliar en un corto espacio de tiempo. Fig.2-10 a 2-13. Fig. 2-10. Planta joven de SOLNI Fig. 2-11. La misma planta adulta Fig. 2-12 y 2-13. Detalles de las hojas 200 Inflorescencia CARACTERES REPRODUCTIVOS Una de las características más destacadas son los detalles fácilmente observables que presentan las inflorescencias. 20- Posición de las inflorescencias en las ramas con respecto a las hojas (PIR): Los ejemplares observados pertenecen a la Sección Morella como lo describió Morton (1976) donde la posición típica es la extraaxilar. También esta característica la describieron Holm et al. (1977), Ogg y Rogers (1989) y Edmonds y Chweya (1997) en Solanum nigrum y Cabrera, (1983) en Solanum lorentzii. La posición no se utiliza como característica diferencial entre las especies, pero este detalle es una característica constante en este Complejo de malezas. Tabla Nº 2-21. Tabla Nº 2-21. Porcentaje de la posición de las inflorescencias en el Complejo Solanum nigrum SOLNI SOLNI-I SOLLO 1.- axilar 0% 0% 0% 2.- extraaxilar 100 % 100 % 100 % 21- Tipo de inflorescencia (TI): En las plantas del Complejo Solanum nigrum las inflorescencias son de dos tipos: cimosa subumbelada sin bifurcación y cimosa subumbelada bifurcada. En SOLNI, la inflorescencia es del tipo cimosa subumbelada; en SOLLO es cimosa subumbelada con dos ramas de bifurcación (Fig Nº 2-17- A; 2-18 y 2-19). Es un carácter constante que permite diferenciar a las especies. En otras plantas registradas como SOLNI I se observaron los dos tipos de inflorescencias en un mismo ejemplar. A esta última se la ha llamado “especie intermedia” (Fig Nº 2-17- B). Se debe destacar que en SOLNI-I, por todos los caracteres observados corresponde a SOLNI. Además presentan una cantidad baja y variable de inflorescencias cimosas subumbeladas bifurcadas de SOLLO; el número de estas inflorescencias varía de 1 a 10 por planta llamada “intermedia”. Tabla Nº 2-22. Esta particularidad de inflorescencias que crecen de esta manera no se describe en ningún texto de taxonomía. 201 SOLNI-I SOLNI SOLLO A B Figura.Nº 2-17. Esquemas de las inflorescencias del Complejo Solanum nigrum 202 Detalles de las inflorescencias en plantas del Complejo Solanum nigrum Cimosa extraaxilar, sin bifurcación, erecta Figura Nº 2-18. Inflorescencias de SOLNI Cimosa, bifurcada Figura Nº 2-19. Inflorescencias de SOLLO 203 SOLLO nutante SOLNI erecta Figura Nº 2-20. Posición de las inflorescencias Tabla Nº 2-22. Porcentaje del tipo de inflorescencias en el Complejo Solanum nigrum Tipo de Inflorescencia Porcentaje % cimosa no bifurcada (1) 100% SOLNI 18% SOLNI-I cimosa bifurcada en planta no bifurcada (3) cimosa bifurcada (2) 100% SOLLO 22- Posición de las inflorescencias (PI): La posición de las inflorescencias es de dos tipos: son erectas o son nutantes (colgantes). En SOLNI son generalmente erectas; en SOLLO son generalmente nutantes. (Fig. Nº 2-20, Pág 62; Fig. Nº 2-25, Pag.67). Esta característica no es citada en los trabajos de taxonomía. 23- Número de inflorescencias (IP) y 24- Número de flores por planta (FP): La floración y maduración de los frutos es escalonada, contínua, por lo que resulta difícil determinar con exactitud estos valores. Los datos de la Tabla Nº 2-23 muestran que: El número de inflorescencias en plantas adultas es mayor en SOLNI El número de flores por planta es mayor en SOLNI Con respecto a estos números no se encontraron referencias para comparar o discutir Tabla N º 2-23. Número de inflorescencias y Número de flores por planta en el Complejo Solanum nigrum SOLNI Promedios SOLLO Promedios m M m M Nº inflorescencias 12 415 71,71 10 45 19,87 Nº flores/frutos 55 1509 345,6 120 491 261,1 m: Número menor – M: Número mayor 25- Número de flores por inflorescencia (FI) 204 En el Número de flores por inflorescencia, los registros obtenidos son semejantes a todas las citas bibliográficas mencionadas. Se debe tener en cuenta que en Solanum nigrum, la mayoría son trabajos realizados en el hemisferio Norte y que los de Solanum lorentzii en la República Argentina, no están muy detallados pero los números se parecen a los descriptos por Morton (1976). El número de flores por inflorescencia es mayor en SOLLO. En la Tabla N º 2-24 se indica el número de flores por inflorescencia más frecuente y entre paréntesis los que siguen en cantidad. En general, se contaron las flores y los frutos por lo que el número de flores sería mayor aún, pues algunas se caen y otras no llegan a desarrollarse. Tabla N º 2-24. Número de flores por inflorescencias en el Complejo Solanum nigrum SOLNI m M Nº flores/frutos / 2 8 inflorescencias m: Número menor – M: Número mayor El Nº más frecuente 5 ( 4 y 6) SOLLO m 5 M 34 El Nº más frecuente 8 – 9 (10, 14 y 19) 26- Pedúnculo (LPD): Sobre la longitud del Pedúnculo y de los Pedicelos hay coincidencias con los autores consultados por lo que no se discuten los resultados. La longitud del pedúnculo de la inflorescencia en cm, muestra una mínima diferencia (de 0,33 cm). Corresponde a SOLLO el pedúnculo de mayor longitud. Estos datos no se usarán para diferenciar a las especies. Tabla N º 2-25. Longitud del pedúnculo en cm en el Complejo Solanum nigrum Largo del pedúnculo Promedios m M 0,8 2,0 1,16 SOLNI 1,9 2,5 2,18 SOLNI-I 0,4 2,9 1,49 SOLLO m: menor – M: mayor El análisis estadístico de los datos se encuentra en Anexo Capítulo 2, Pág. 96. (Fig. Nº 2-21 y Tablas Nº 2-26 y 2-27). 27- Pedicelo (LPC) (Longitud) en mm. Los datos obtenidos no se utilizarán para la diferenciación. 28- Longitud de las ramas de bifurcación de la inflorescencia (LBI) 205 Las ramas tienen diferentes longitudes. La diferencia entre la rama 1 y la rama 2 es mayor en SOLLO. En la forma intermedia si bien sus pedúnculos son más largos casi no hay diferencia en la longitud de las ramas. Las diferencias se observan sólo en los promedios como se muestra en Tabla N º 2-28. Tabla Nº 2-28. Longitud de las ramas de bifurcación de las inflorescencias en el Complejo Solanum nigrum Largo de la Rama 1 (mm) m M 3 10 SOLLO 3 3,3 SOLNI-I m: menor – M: mayor Promedio 5,79 3,08 Largo de la Rama 2 (mm) m M 3 10 3,1 3,6 Promedio 6,6 3,38 Los valores de longitudes de pedúnculo, pedicelo y ramas de la bifurcación son semejantes a los descriptos por Morton (1976). No se usarán para hacer una diferenciación. Más que medir o referirse a la longitud de estas partes hay que hacer hincapié en el tipo de inflorescencia (si se ramifica o no), en el número de flores por inflorescencia y en el tamaño de las flores. FLORES Cáliz El cáliz de las especies de este Complejo responde a las características de la familia Solanáceas y del género Solanum en cuanto a tipo, forma, número de piezas y dimensiones de las mismas (Cabrera, 1983). 29- Tipo (KT): Si bien las piezas son de pequeñas dimensiones y con diferencias mínimas en sus medidas se debe destacar que el cáliz es irregular. Frecuentemente se unen dos dientes de éste y tres quedan más o menos libres, pero siempre es gamosépalo en la base. Esta característica no se encontró en ningún texto. El cáliz observado en la flor es campanulado, 5-dentado, normal (no retroflexo) y pubescente. La pubescencia la componen sólo pelos simples y ralos, no visibles a simple vista. En el análisis del carácter Nº 60 se describirá el cáliz en el fruto. 30- Forma del cáliz (KF): Tabla N º 2-29. Forma del cáliz en las flores del Complejo Solanum nigrum Tipos SOLNI 2 SOLNI-I 2 SOLLO 2 1- Reflexo 2- No reflexo Porcentaje 100 % 100 % 100 % Se observó que en las flores, las piezas del cáliz siempre tienen la misma forma, no reflexo. 206 Esta característica no se usará para hacer una diferenciación. 31- Número de sépalos (KN) y 32- Número de dientes del cáliz (KND): Se contaron 5 piezas del cáliz en todos los ejemplares. El número de dientes coincide con el número de sépalos que es 5. Estas son características de la Familia Solanáceas. No se utilizarán para hacer una diferenciación. 33- Longitud del cáliz (KL): Se encontró una diferencia mínima en la longitud de los sépalos. Se debe destacar que el cáliz es más pequeño en SOLNI, las flores también son más pequeñas. Tabla Nº 2-30. Tabla Nº 2-30. Longitud de los sépalos en las flores del Complejo Solanum nigrum (mm) Largo del cáliz Promedio m M 1,5 3,0 2,08 * SOLNI 1,8 4,0 2,52 SOLLO * 2,08 mm (incluye a SOLNI-I) m: menor – M: mayor 34- Forma de los dientes del cáliz (KFD): En todos los dientes la forma es deltoidea, con punta roma. 35- Longitud de los dientes (KLD) (en mm): Tabla Nº 2-31. Tabla Nº 2-31. Longitud de los dientes del cáliz en el Complejo Solanum nigrum Largo de los dientes Promedio m M 0,65 0,75 0,73* SOLNI 1,1 1,7 1,45 SOLLO * 0,73 mm (incluye SOLNI-I) m: menor – M: mayor Los datos de número, forma y longitud de las piezas del cáliz no se utilizarán para hacer una diferenciación. El análisis estadístico de los siguientes datos se encuentra en Anexo Capítulo 2: Longitud de sépalos (KLS) (en la flor): Pág. 97. (Fig. Nº 2-22 y Tablas Nº 2-32 y 2-33). Longitud del cáliz (KL) (en la flor): Pág. 98. (Fig. Nº 2-23 y Tablas Nº 2-34 y 2-35). Longitud de los dientes (KLD) (en la flor): Pág. 99. (Fig. Nº 2-24 y Tablas Nº 2-36 y 2-37). Corola La corola en el Complejo también responde, como el cáliz, a las características de la familia y del género Solanum en cuanto a tipo, forma, número de piezas y dimensiones de las mismas (Morton, 1976; Cabrera, 1986). 207 36- Tipo (CT): Gamopétala siempre. 37- Forma de la corola (CF): Rotácea, estrellada. Se observó que cuando las flores se van desarrollando los pétalos se doblan hacia atrás. Fig. Nº 2-27, 2-28 y 2-32. 38- Número de piezas (CN): 5 (cinco) pétalos en todos los ejemplares. 39- Color (CC): En las especies del Complejo Solanum nigrum, el color de los pétalos es blanco. Tabla Nº 232; Fig. 2-25 a 2-32. En Solanum nigrum a veces se dice que las flores son blancas con el centro amarillo como un sol (Morton, 1976). La coloración amarilla corresponde a las anteras del androceo. Las flores de Solanum nigrum tienen pétalos blancos. En algunas observaciones se registró color de flores lilas. Esto se debe a que Solanum lorentzii tiene un tinte morado en el centro de la corola más marcado lo que las hace parecer de color lila, pero observando con atención las flores son blancas (Fig. Nº 2-30). Tabla N º 2-32. Color de las flores en el Complejo Solanum nigrum Color Porcentaje Blanca 100 % SOLNI Blanca 100 % SOLNI-I 100 % SOLLO Blanca con el centro morado 208 Solanum nigrum L. Detalle de las flores Figura Nº 2-25. Inflorescencia erecta Figura Nº 2-27. Corola rotácea y detalle del cáliz en la flor. Figura Nº 2-26. Corola rotácea, estrellada Figura Nº 2-28. Pétalos en la flor madura. 209 Solanum lorentzii Bitter Detalle de las flores Figura Nº 2-29. Inflorescencia cimosa bifurcada y detalle del cáliz en la flor Figura Nº 2-30. Corola rotácea con el centro morado 210 Centro morado Figura Nº 2-31. Corola rotácea, estrellada Figura Nº 2-32. Pétalos en la flor madura 211 De estos cuatro caracteres (36 a 39), el que permite obtener una diferencia de las especies a simple vista es el color de la flor. Como ya se dijo el centro morado en las flores de SOLLO las hace parecer de color lila, pero observando con atención las flores son blancas. 40- Diámetro (CD): (en mm) La corola rotácea permite medir fácilmente el diámetro de las flores. Los valores se presentan en Tabla Nº 2-33. En SOLNI los promedios de Morton (1976) son ligeramente mayores, de 6,8 a 6,9, pero se debe considerar que este carácter está influenciado por el ambiente. Los promedios en SOLLO de Morton (1976), de 12 a 16 mm y Cabrera (1983), de 12 a 15 mm son semejantes a los valores obtenidos en esta tesis. Tabla N º 2-33. Diámetro de la corola en el Complejo Solanum nigrum Diámetro (en mm) Promedio m M 3 9 4,9 * SOLNI – SOLNI-I 12 22 15,5 SOLLO * 4,9 mm (incluye a SOLNI-I) m: menor – M: mayor Si bien las flores son pequeñas, hay diferencias en las medidas de cada especie: SOLLO es de mayor tamaño que SOLNI. Los tamaños pueden observarse en forma directa. El resultado en este carácter sirve para la diferenciación pero complementado con la observación de número de flores y la bifurcación de las inflorescencias. 41- Tamaño de los pétalos (CL): (Longitud –ancho en mm) Los registros se muestran numérica y gráficamente y confirman el mayor tamaño en las flores de SOLLO. Tabla N º 2-34. El análisis estadístico de los datos se encuentra en Páginas 70 y 71, Figuras Nº 2-33 a 2-35 y Tablas Nº 2-35 y 2-36. Tabla N º 2-34. Dimensiones de pétalos en el Complejo Solanum nigrum Longitud Promedio Ancho Promedio m M m M 5 3,94 1 1,5 1,24 * SOLNI – SOLNI-I 2 5 10 6,7 2,9 5 3,67 SOLLO * promedio de 50 datos: 1,24 mm (incluye a SOLNI-I) 212 4,5 16 4,0 IC -95% del Ancho de los Pétalos 18 14 12 10 8 6 3,5 3,0 2,5 2,0 1,5 4 1,0 2 N= ,5 100 100 SOLN I SOLLO Figura N° 2-33. Diámetro de corola de diferentes especies del Complejo Solanum nigrum N= 30 31 SOLN I SOLLO Figura N° 2-34. Ancho de pétalos de diferentes especies del Complejo Solanum nigrum 213 Tabla N° 2-35. Análisis de diámetro de la corola, largo y ancho de los pétalos del Complejo Solanum nigrum Diametro de la Corola (mm) Longitud de los Petalos (mm) IC-95% de la Longitud de los Petalos (mm) Ancho de los Petalos ESPECIE SOLNI SOLLO SOLNI SOLLO 8 SOLNI SOLLO 7 N 100 100 30 Media 4,9050 15,5100 3,9467 DE 1,37909 2,49239 1,06568 EE ,13791 ,24924 ,19457 31 6,7000 1,81016 ,32511 30 31 1,1933 3,6774 ,19286 ,63651 ,03521 ,11432 Tabla N° 2-36. Pruebas estadísticas para el análisis del diámetro de la corola largo y ancho de6 los pétalos del Complejo Solanum nigrum 5 4 3 N= 30 31 SO LN I SO LLO Fig. N° 2-35. Largo de pétalos de diferentes especies del Complejo Solanum nigrum 214 Prueba de muestras independientes Prueba de Lev ene para la igualdad de v arianzas Diametro de la Corola (mm) Longitud de los Petalos (mm) Ancho de los Petalos Varianzas iguales Varianzas desiguales Varianzas iguales Varianzas desiguales Varianzas iguales Varianzas desiguales F 32,741 Sig. ,000 16,190 ,000 58,117 ,000 Prueba T para la igualdad de medias t -37,230 -37,230 -7,208 198 154,425 59 Sig. (bilateral) ,000 ,000 ,000 -7,267 48,854 ,000 -20,481 -20,767 59 35,630 ,000 ,000 gl 215 42- Forma de los lóbulos de los pétalos (CFP): Los lóbulos, en todos los ejemplares observados terminan en punta (triangulares) o son algo romos (Morton (1976); Cabrera (1983)). Este dimorfismo ocurre no sólo en flores de una misma planta, sino en una misma inflorescencia. Tabla N º 2-37. Tabla N º 2-37. Forma de lóbulos en el Complejo Solanum nigrum Forma Porcentaje 1 terminados en punta 50 % 50 % 2 romos 0% 3 otros 43- Profundidad de las escotaduras (CE): Las flores presentan diferencias en la profundidad de las escotaduras, pero siempre están soldados en la base. La mayor cantidad de flores presenta escotaduras profundas (más de un tercio de longitud de pétalos); el resto escotaduras poco profundas (menos de la mitad de la longitud de éstos). No se observaron flores sin escotaduras. Este dimorfismo también fue advertido en una misma planta y hasta en una misma inflorescencia. (Fig. Nº 2-36 y 2-37). Las características de forma y profundidad coinciden con las descripciones de Morton (1976) y de Cabrera (1983). Tabla Nº 2-38. Profundidad de escotaduras de corola en Complejo Solanum nigrum (%) 1 2 3 Profundidad de las escotaduras Porcentaje sin escotaduras 0% escotaduras poco profundas 30 % escotaduras profundas 70 % Figura Nº 2-36 Escotaduras de corola de SOLNI Figura Nº 2-37 Escotaduras de corola de SOLLO 216 ANDROCEO El tipo, número de piezas y dimensiones del androceo de las especies de este Complejo, responden a las características de la familia Solanáceas y del género Solanum (Cabrera, 1983). Estambres 44- Tipo (AT): Las anteras grandes y conniventes se muestran como un centro de color amarillo, de estructura compacta. Fig. Nº 2-38 y 2-39. (Pág 74). 45- Número de piezas (AN): Todos los ejemplares observados tienen 5 estambres. 46- Longitud de las anteras (AL): (en mm) Las mediciones y promedios del largo de las anteras en las especies del Complejo se exponen en la Tabla Nº 2-39. Estos números muestran que las anteras de SOLNI son de menor tamaño que SOLLO. Fig. 2-38 y 2-39. Tabla N º 2-39. Largo y ancho de anteras en el Complejo Solanum nigrum Longitud Promedio Ancho Promedio m M m M 1,5 4 2,86 * 0,5 0,8 0,64 * SOLNI – SOLNI-I 3,8 5 4,25 1 1,1 1,05 SOLLO * 2,86 mm y 0,64 mm incluyen a SOLNI-I m: menor – M: mayor 47- Ancho de los filamentos (AAF): (en mm) Se determinó que los valores son mayores en SOLLO como también sus flores son más grandes. Tabla Nº 2-40. Ancho de filamentos en el Complejo Solanum nigrum SOLNI – SOLNI-I SOLLO Ancho m M 0,4 0,7 0,6 1,2 * 0,515 mm (incluye a SOLNI-I) Promedio 0,515 * 0,871 m: menor – M: mayor 48- Longitud de los filamentos (ALF): (mm) El largo de los filamentos es mayor en SOLLO. Tabla Nº 2-41. Largo de filamentos en Complejo Solanum nigrum SOLNI – SOLNI-I SOLLO * 1,9 mm (incluye a SOLNI-I) Ancho m M 1,5 2 4 5 Promedio 1,9 * 4,83 m: menor – M: mayor 217 La longitud y ancho de los filamentos registrados en 46, 47 y 48 están relacionados con el tamaño de las flores. A mayor tamaño de las flores, mayores valores, sirven para confirmar la identificación pero no para realizar una diferenciación. Los gráficos y análisis estadísticos se muestran en Anexo Capítulo 2 (Pág. 100). Fig. Nº 2-40 y Nº 2-41; Tablas Nº 2-42 y Nº 2-43. 49- Pubescencia del androceo (AP): El androceo, como los otros órganos de la planta tiene una pubescencia rala. Tabla Nº 2-44. Pubescencia del androceo en el Complejo Solanum nigrum SOLNI SOLNI-I SOLLO 1 glabra 0% 0% 0% 2 3 pubescencia densa pubescencia rala 0% 100 % 0% 100 % 0% 100 % 50- Tipo de pubescencia (APT): Los pelos son simples. Tabla Nº 2-45. Tipo de pubescencia del androceo en Complejo Solanum nigrum 1 2 3 Pelos simples Pelos glandulosos Otro tipo 100 % 0% 0% SOLNI 100 % 0% 0% SOLNI-I 100 % 0% 0% SOLLO De este grupo de caracteres hay que destacar que todas las especies del Complejo presentan 5 (cinco) estambres, con gruesas anteras conniventes, amarillas, lo que le da a las flores el aspecto de tener un sol en el centro. La pubescencia es rala con pelos simples. 218 Fig. Nº 2-38. Anteras conniventes de SOLNI Fig. Nº 2-39. Anteras conniventes de SOLLO Pistilo que sobresale las Anteras en ambas especies 219 GINECEO La posición del gineceo de las especies del Complejo responde a las características de la familia Solanáceas aunque sus dimensiones pueden variar en las diferentes especies del género Solanum (Cabrera, 1983). Los resultados obtenidos concuerdan con este autor y se detallan a continuación: 51- Posición del ovario (GT): La posición del gineceo es súpero. (Tabla N º 2-46). Tabla Nº 2-46. Posición del gineceo en el Complejo Solanum nigrum 1 2 Súpero Ínfero 100 % 0% SOLNI 100 % 0% SOLNI-I 100 % 0% SOLLO 52- Longitud del gineceo (GL): En todos los ejemplares se observó que el gineceo sobresale por entre las anteras: aproximadamente ⅓ (un tercio) de su longitud en SOLLO y menos de ⅓ (un tercio) en SOLNI. Fig. Nº 2-38 y 2-39. (Página 74). Tabla N º 2-47. Longitud del gineceo en Complejo Solanum nigrum 1 2 No sobresale del androceo Sobresale del androceo 0% 100 % SOLNI 0% 100 % SOLNI-I 0% 100 % SOLLO 220 FRUTO Esta característica se considera que es la más importante de las reproductivas estudiadas. El Tipo de fruto es una baya de forma globosa ligeramente comprimida en el polo apical, fruto típico del género Solanum (Cabrera, 1983; Dottori,1995). Los trabajos de Roberts y Locket, 1978; Ogg y Rogers, 1989 y Edmods y Chweya, 1997 sobre la biología del Complejo Solanum nigrum se refieren solamente a la maduración de los frutos, la que se produce en forma escalonada; por lo que en plenitud de fructificación se contó la totalidad de las partes reproductivas. Esto incluye pimpollos, flores bien desarrolladas, frutos verdes y frutos maduros lo que daría un número aproximado del total de frutos. Sobre Número de infrutescencias por planta, Número de frutos por planta ni del Número de frutos por infrutescencia en plenitud de fructificación, no se han encontrado referencias bibliográficas. Con respecto al Diámetro se han encontrado valores con escasa diferencia a los citados por Morton, 1976; Cabrera, 1983; Ogg y Rogers, 1989; Edmonds y Chweya, 1997. En la observación de plantas en el campo, los frutos de Solanum lorentzii parecían de mayor tamaño; en cambio en las mediciones los resultados mostraron lo contrario. Este carácter también está relacionado con los factores ambientales. Sobre el Color de las bayas, sólo hay referencias de los frutos de Solanum nigrum (Morton, 1976; Ogg y Rogers, 1989; Edmods y Chweya, 1997); tampoco ningún autor describe a la baya de Solanum lorentzii. Para este estudio esta es una de las características más importantes por su constancia y fácil visualización. También se debe destacar que en ningún texto se describió el color de la pulpa y del jugo de los frutos y éstos también son caracteres constantes y fáciles de visualizar al hacer la identificación. El cáliz en el fruto maduro es otro carácter importante por su constancia, en la bibliografía consultada tiene algunas diferencias y las descripciones se mezclan posiblemente por la difícil identificación de este grupo de plantas. La longitud de los sépalos y cuánto cubren éstos a las bayas son coincidentes con la bibliografía, pero no se consideran en esta tesis porque no es un carácter diferencial de importancia. Se detallan a continuación los caracteres carpológicos observados. 53- Tipo (FT): Es un fruto carnoso, una baya globosa ovoide o elipsoide generalmente comprimida en el polo apical, con cáliz persistente. Este cáliz no crece con el fruto, sólo lo acompaña y adquiere diferentes formas. 221 54- Número de infrutescencias por planta (FIP): (en plenitud de fructificación) Tabla Nº 2-48. Número y promedios de infrutescencias por planta en el Complejo Solanum nigrum SOLNI Promedios SOLLO Promedios m M m M Nº infrutescencias/planta 16 415 71,71 10 45 19,87 Nº flores/frutos* 80 2075 -80 360 - * se calculan estos valores considerando 5 frutos por infrutescencia en SOLNI y 8 en SOLLO. m: menor – M: mayor El menor número de infrutescencias observado en plantas adultas de SOLNI es 16 y el mayor es 415, mientras que en SOLLO el menor es 10 y 45 el mayor. El número de infrutescencias en adultas es notablemente mayor en SOLNI y por lo tanto también el número de frutos por planta. (Fig. Nº 2-42 a 2-44 y Tablas Nº 2-51 y 2-52. Pág. 78 y 79). La maduración de los frutos se produce en forma escalonada, por lo que en plenitud de fructificación se han contado las inflorescencias y las infrutescencias. 55- Número de frutos por infrutescencias (FNI): El menor Nº de frutos por infrutescencia observado es 2 y el mayor es 8 en SOLNI, mientras que en SOLLO va de 5 hasta 34. Es mucho mayor la cantidad de frutos en SOLLO. El número de frutos más frecuente en SOLNI es 5 y en menor cantidad 4 y 6, mientras que en SOLLO los valores más frecuentes son en orden decreciente 8, 9, 10, 14 y 19. (Tabla Nº 2-49). Las infrutescencias encontradas con un solo fruto son las que sus flores abortaron y un solo fruto se desarrolló, quedando las marcas donde había flores. No se consideraron para este análisis. Fig. Nº 2-44. Tabla N º 2-49. Número de frutos por infrutescencias en el Complejo Solanum nigrum SOLNI Nº más frecuente SOLLO Nº más frecuente m M m M Nº flores/frutos / infrutescencias 2 8 5 (4 y 6) 5 34 8–9 (10, 14 y 19) m: número menor – M: número mayor 56- Número de frutos por planta (FNP): De acuerdo con estos registros el número de frutos por planta es mayor en SOLNI. Fig. Nº 243. Tabla N º 2-50. Tabla N º 2-50. Número de frutos por planta en el Complejo Solanum nigrum SOLNI Promedio SOLLO Promedio m M m M 222 Nº flores/frutos 55 1509 345,6 120 491 m: número menor – M: número mayor 261,1 A continuación se muestran los gráficos y las pruebas estadísticas de los caracteres N º 54, 55 y 56 223 140 IC-95% del Nº Inflor / Planta 120 100 80 60 40 20 0 N= 23 20 SO LNI SO LLO Figura N° 2-42. Número de infrutescencias por planta (FIP) de las especies del Complejo Solanum nigrum IC-95% del Nº Fruto / Planta 600 500 400 300 200 100 N= 23 20 SO LNI SO LLO Figura N° 2-43. Número de frutos por planta (FNP) de las especies del Complejo Solanum nigrum 16 IC-95% del Nº Fruto / Infloresc 14 12 10 8 6 4 2 N= 164 SOLNI 275 SOLLO Figura N° 2-44. Número de frutos por infrutescencias (FNI) de las especies del Complejo Solanum nigrum 224 Tabla N° 2-51. Análisis del Nº de infrutescencias por planta y Nº de frutos por planta del Complejo Solanum nigrum Nº Inf lor / Planta Nº Fruto / Planta ESPECIE SOLNI SOLLO SOLNI SOLLO N 23 20 23 20 Media 80,78 23,40 336,83 306,05 DE 95,532 9,133 354,224 129,867 EE 19,920 2,042 73,861 29,039 Tabla N° 2-52. Pruebas estadísticas para el análisis del Nº de infrutescencias por planta y Nº de frutos por planta en las distintas especies del Complejo Solanum nigrum Prueba de Lev ene para la igualdad de v arianzas Nº Inf lor / Planta Nº Fruto / Planta Se han asumido v arianzas iguales No se han asumido v arianzas iguales Se han asumido v arianzas iguales No se han asumido v arianzas iguales F Sig. 10,768 4,503 Prueba T para la igualdad de medias t ,002 ,040 Sig. (bilateral) gl 2,671 41 ,011 2,866 22,462 ,009 ,367 41 ,715 ,388 28,537 ,701 57- Diámetro (FD): en mm El diámetro varía entre 3 mm y 10 mm en SOLNI y 3 mm y 9,5 mm en SOLLO. El valor más frecuente es 7 mm para ambas especies con porcentajes diferentes como se observa en Tabla Nº 2-53. Tabla N º 2-53. Diámetro de frutos en el Complejo Solanum nigrum SOLNI SOLLO más frecuente 7 (151) 7 (40) % Menor 21,01 3 (5) 15,30 3 (11) % Mayor 0,727 10 (1) 4,23 9,5 (2) % 0,145 0,769 En la Tabla N° 2-54, se muestra la cantidad de frutos que tienen un determinado diámetro y el porcentaje que representa en la muestra analizada. Con colores se indican los valores más frecuentes para SOLNI Y SOLLO que muestran una semejanza en los tamaños y en el porcentaje de estos tamaños. 225 Tabla N° 2-54. Relación entre diámetro y cantidad de frutos en el Complejo Solanum nigrum SOLNI Diámetro 3,0 3,5 4,0 4,5 5,0 5,5 6,0 6,2 6,5 6,6 7,0 7,1 7,2 7,5 8,0 8,5 9,0 9,5 10,0 Total Frutos Cantidad de frutos 5 1 18 8 68 38 119 1 78 1 151 1 1 79 84 18 13 2 1 687 SOLLO % 0,727 0,145 2,62 1,164 9,898 5,531 17,321 0,145 11,353 0,145 21,979 0,145 0,145 11,499 12,227 2,62 1,892 0,291 0,145 Diámetro 3,0 3,5 4,0 4,5 5,0 5,5 6,0 6,2 6,5 6,9 7,0 7,1 7,3 7,5 8,0 8,5 9,0 9,5 10,0 Cantidad de frutos 11 0 13 4 39 7 38 0 18 2 40 1 1 27 24 14 19 2 0 260 % 4,23 0 5 1,538 15 2,692 14,6 0 6,923 0,769 15,384 0,384 0,384 10,384 9,23 5,384 7,307 0,769 0 Ref. Tabla Nº2-54: se indica con los colores el orden del valor más frecuente:1 º amarillo; 2º verde; 3º celeste El análisis estadístico de los datos se encuentra a continuación (Pág. 81). Fig. Nº 2-45 y Tablas Nº 2-55 y 2-56. 226 6,9 IC-95% del Diametro del Fruto (mm) 6,8 6,7 6,6 6,5 6,4 6,3 6,2 6,1 N= 630 262 SOLNI SOLLO 59 SOLNI -I Figura N° 2-45. Diámetro de frutos de las diferentes especies del Complejo Solanum nigrum Tabla N° 2-55. Diámetro de frutos de especies del Complejo Solanum nigrum Diametro del Frut o (mm) SC 2,116 1550,313 1552,428 Inter-grupos Intra-grupos Total gl CM 1,058 1,635 2 948 950 F ,647 Sig. ,524 Tabla N° 2-56. Pruebas estadísticas para el análisis del diámetro en las especies del Complejo Solanum nigrum Subconjuntos Homogéneos según Diametro del Fruto (mm) a,b HSD de Tukey ESPECIE SOLNI-I SOLLO SOLNI Sig. N 59 262 630 Subconjunto para alf a = .05 1 6,5254 6,5695 6,6589 ,669 a. Usa el tamaño muestral de la media armónica = 134,209. b. Los tamaños de los grupos no son iguales. Se utilizará la media armónica de los tamaños de los grupos. Los niv eles de error de tipo I no están garantizados. 227 58- Color (FC): Ogg y Rogers (1989) describen el color del fruto en SOLNI, negro brillante, Edmonds y Chweya (1997) varios colores (negro, marrón, rojo, naranja, y verde). De SOLLO no hay descripciones. Tampoco se describe el color de la pulpa y del jugo. Los colores observados en los frutos y los colores de la pulpa y del jugo de las bayas son características constantes. En SOLNI y en la forma intermedia, SOLNI-I, el color es negro brillante. En cambio en SOLLO es morado opaco. Tabla Nº 2-57 y Figuras 2-46 a 2-48. Tabla Nº 2-57. Color de frutos en el Complejo Solanum nigrum Color Porcentaje 100 % SOLNI – SOLNI-I 1. negro brillante 2. morado opaco 100 % SOLLO FRUTOS E INFRUTESCENCIAS SOLLO SOLNI Fig. Nº 2-46. Infrutescencias del Complejo Solanum nigrum 228 SOLNI SOLLO Figura Nº 2-47. Color de bayas y forma del cáliz en el fruto maduro SOLNI-I Figura Nº 2-48 Color de bayas y forma del cáliz en fruto maduro SOLNI-I 59- Color de la pulpa ( y del jugo) (FCP): El color de la pulpa y del jugo es morado en SOLNI y en SOLNI-I y en SOLLO es verde. Como se observa en Tabla Nº 2-58 y en las Fig. 2-49 a 2-51. Tabla Nº 2-58. Color de pulpa en Complejo Solanum nigrum Color de la pulpa Porcentaje 100 % SOLNI – SOLNI-I morado verde 100 % SOLLO 229 SOLLO SOLNI Figura Nº 2-49. Color de pulpa en bayas maduras de SOLNI y SOLLO CP Figura Nº 2-50. Color de la pulpa en la baya de SOLNI Figura Nº 2-51. Color de la pulpa y corpúsculos pétreos en la baya de SOLLO 60- Forma del cáliz en el fruto maduro (FFK): El cáliz es persistente. De forma acampanada, gamosépalo, con 5 sépalos, regulares o irregulares. A la madurez se desgarran de manera irregular y quedan, generalmente, 2 sépalos de un lado y 3 del otro. 230 En los frutos maduros se observaron dos tipos: con sépalos reflexos (vueltos hacia atrás) y con sépalos de aspecto y posición normales (extendidos o no reflexos). Tabla N º 2-59. Tabla N º 2-59. Forma del cáliz en fruto maduro en Complejo Solanum nigrum Tipos Porcentaje 100 % SOLNI – SOLNI-I reflexo 2 normal (no reflexo) 100 % SOLLO En las Solanáceas el cáliz generalmente es persistente o acrescente. En las especies observadas en esta tesis es persistente, es decir, que sólo acompaña al fruto en su madurez. En SOLNI cuando el fruto está verde, el cáliz se encuentra adherido a la baya. Al madurar, el cáliz se desprende y se dobla hacia atrás (cáliz retroflexo). El cáliz cubre ¼ (un cuarto) de la baya. Esta característica también se ha observado en la forma intermedia de SOLNI. (Figura Nº 2-52). En SOLLO cuando el fruto está verde el cáliz se encuentra adherido a la baya. Al madurar, el cáliz no se desprende, no se dobla hacia atrás, queda extendido. El cáliz cubre 1/8 (un octavo) de la baya. (Figura Nº2-53). En Morton (1976), las medidas del cáliz en el fruto de SOLNI van de 2,8-3,4 mm de long., pero en SOLLO las medidas se refieren a la flor, 2,0-2,5 mm de long., que son aproximadamente los valores registrados. Lo importante es la forma que toma el cáliz en el fruto maduro. Forma del cáliz en el Complejo Solanum nigrum Figuras Nº 2-52. Forma del cáliz en frutos verdes y en maduros de SOLNI 231 Figuras Nº 2-53 Forma del cáliz en frutos verdes y en maduros de SOLLO 61- Longitud de los sépalos (FKL): (mm) Las mediciones se hicieron en frutos completamente maduros. Los valores se muestran en la siguiente tabla. Tabla Nº 2-60. Tabla Nº 2-60. Longitud de sépalos en Complejo Solanum nigrum Longitud (mm) Promedios m M 3,0 2,087 SOLNI - SOLNI I 1,5 1,8 4,0 2,520 SOLLO m: menor – M: mayor Estos datos no sirven para diferenciar las especies sino para confirmar su identificación. Lo importante es la forma que toma el cáliz en el fruto maduro. De los detalles obtenidos de las piezas del cáliz en el fruto sólo tiene importancia la forma en el fruto maduro como carácter de diferenciación de las especies del Complejo, por su fácil observación. 232 SEMILLA La mayoría de los autores coinciden en que estas plantas sólo se reproducen por semillas y que producen una gran cantidad de éstas. Por ejemplo, Solanum nigrum, puede llegar a tener 178.000 semillas por planta (Holm et al., 1977). Según Morton (1976) y Ogg y Rogers (1989) es menor la cantidad de semillas por fruto. No hay datos sobre número de semillas de Solanum lorentzii. La forma y el tamaño observados coinciden con las descripciones. Si bien el peso no es un carácter taxonómico, se considera importante por que los datos obtenidos muestran la diferencia de tamaño de estas pequeñas semillas. Se coincide con Ogg y Rogers (1989) y Edmonds y Chweya (1997) en el color de las semillas de Solanum nigrum y no hay referencias de las semillas de Solanum lorentzii. 62- Número de semillas por fruto (SN): El resultado de la cuenta se muestra en Tabla Nº 2-61. El análisis estadístico de los datos se encuentra en Pág. 87. Fig. Nº 2-54 y Tablas Nº 2-62 y 2-63. Tabla Nº 2-61. Semillas por fruto en el Complejo Solanum nigrum SOLNI SOLNI-I SOLLO Nº menor/fruto Nº mayor/fruto Promedios 14 93 58,64 56 91 74,66 10 128 50,95 Además, se encontraron frutos con un número muy bajo de semillas, 5 en SOLNI y 2 en SOLLO, pero esos datos no se tuvieron en cuenta. 233 90 IC-95% del Nº Semillas / Fruto 80 70 60 50 40 N= 89 114 SOLLO SOLNI 18 SOLNI -I Figura N° 2-54. Número de semillas de las diferentes especies del Complejo Solanum nigrum Tabla N° 2-62. Análisis del número de semillas (sem/fruto) del Complejo Solanum nigrum Nº Semillas / Fruto Inter-grupos Intra-grupos Total SC 9171,998 166650,075 175822,072 gl 2 218 220 CM 4585,999 764,450 F 5,999 Sig. ,003 Tabla N° 2-63. Pruebas estadísticas para el análisis del número de semillas (sem/fruto) en las distintas especies del Complejo Solanum nigrum Nº Semillas / Fruto a,b HSD de Tukey ESPECIE SOLLO SOLNI SOLNI-I Sig. N 89 114 18 Subconjunto para alf a = .05 1 2 50,9551 58,6404 74,6667 ,432 1,000 a. Usa el tamaño muestral de la media armónica = 39, 702. b. Los tamaños de los grupos no son iguales. Se utilizará la media armónica de los tamaños de los grupos. Los niv eles de error de tipo I no están garant izados. 63- Forma (SF): Son reniformes, aplanadas y aplastadas. Son semejantes en estas especies. Tamaño: 64- Ancho de la semilla (SA), 65- Largo de la semilla (SL) (en mm) 234 Son semillas muy pequeñas. Las de SOLLO son de menor tamaño que las de SOLNI y de la forma intermedia. Los valores encontrados de longitud, ancho, valor más frecuente (VMF) y promedios se presentan en la Tabla Nº 2-64. Tabla Nº 2-64. Dimensiones de las semillas en el Complejo Solanum nigrum SOLNI SOLNI-I SOLLO Largo (mm) m M VMF Prom. 0,9 2,0 1,5 1,354 1,1 1,6 1,2 1,258 0,5 2,0 1,0 1,023 m: menor – M: mayor Ancho (mm) m M VMF Prom. 0,8 1,9 1,0 1,032 0,9 1,2 1,1 1,06 0,4 1,1 0,6 0,747 MF: valor más frecuente Prom.: promedio El análisis estadístico de los datos se encuentra en Anexo Capítulo 2, Pág. 101-102. Fig. Nº 2-55 y 2-56. Tablas Nº 2-65 a 2-68. 66- Color (SC): El color de las semillas extraídas de frutos maduros es marrón, con diferentes tonalidades en las especies: marrón claro en SOLNI y marrón oscuro en SOLLO. Figura Nº 2-57. SOLNI SOLLO Figura Nº 2-57. Color de las semillas del Complejo Solanum nigrum 67- Peso (SP): El peso no es un carácter taxonómico, pero es importante por que los datos obtenidos muestran la diferencia de tamaño de las semillas. De los pesos obtenidos en balanza granataria y en balanza de precisión, se muestran estos últimos por ser más representativos. (Tabla Nº 2-69). Tabla Nº 2-69. Peso de semillas del Complejo Solanum nigrum PESO MENOR PESO MAYOR SOLNI SOLLO SOLNI-I 0,0108 0,0035 0,0124 0,0123 0,0046 0,0157 235 0,2291 0,0812 0,2871 TOTAL 0,0115 0,0041 0,0144 PROMEDIO UNA SEMILLA 0,00023 0,00008 0,00029 Muestra: de 50 semillas -- Balanza de precisión El análisis estadístico y la representación gráfica de esta variable cuantitativa se muestra en la Figura Nº 2-58; Tablas 2-70 y 2-71. , 016 IC-95% del Peso 50 Semilla (gr) , 014 , 012 , 010 , 008 , 006 , 004 , 002 N= 20 20 SO LLO SO LNI 20 SO LNI-I Figura N° 2-58. Peso de 50 semillas de diferentes especies del Complejo Solanum nigrum. Tabla N° 2-70. Peso de 50 semillas (gr) del Complejo Solanum nigrum. Peso 50 Semilla (gr) Inter-grupos Intra-grupos Total SC ,001 ,000 ,001 gl 2 57 59 CM ,001 ,000 F 1297,120 Sig. ,000 Tabla N° 2-71. Pruebas estadísticas para el análisis del peso de 50 semillas (gr) en las distintas especies del Complejo Solanum nigrum. 236 Subconjuntos Homogéneos Peso 50 Semil la (gr) a HSD de Tukey ESPE SOLLO SOLNI SOLNI-I Sig. N 20 20 20 Subconjunto para alf a = .05 1 2 3 ,004060 ,011455 ,014355 1,000 1,000 1,000 a. Usa el tamaño muestral de la media armónica = 20,000. 237 CARACTERES ANATÓMICOS CORPÚSCULOS PÉTREOS (CP) Los corpúsculos pétreos (grain stones, esclereidas o células pétreas) son grupos de células que en el fruto se encuentran en el pericarpio y cuyas paredes están engrosadas con lignina. Algunas especies de Solanáceas contienen muchos, otras unos cuantos o bien carecen de ellos. 68- Presencia / ausencia de corpúsculos pétreos: En Solanum nigrum y en la forma intermedia no se observaron corpúsculos pétreos, mientras que sí se los encuentra en Solanum lorentzii. Este importante detalle coincide con la bibliografía consultada (Morton, 1976; Cabrera, 1983). La información recopilada de diferentes autores se muestra en Tabla Nº 2-72. Tabla Nº 2-72. Presencia / ausencia de corpúsculos pétreos según los autores Especie Morton Ogg Cabrera SOLAM 7,7-7,9 0-5 --------------SOLNI Ausente Ausente --------------SOLLO 2ó6 -------------2-6 SOLBE ------------ -------------- Numerosos Edmods y Chweya Presente o Ausente Presente o Ausente ------------------------------------------- Por lo expuesto en tabla 2-72 se descarta que las plantas observadas pudieran ser Solanum americanum. La observación realizada en numerosos ejemplares se muestra en Tabla Nº 2-73. Tabla N º 2-73. Corpúsculos pétreos en frutos del Complejo Solanum nigrum SOLNI – SOLNI-I SOLLO 1.- Presentes 2.- Ausentes 0% 100 % 100% 0% 69- Número de corpúsculos pétreos por fruto (NCP): En Solanum lorentzii, Morton (1976) y Cabrera (1983) citan un número fijo de corpúsculos pétreos. En los registros de estas bayas se encontró que el número de este carácter varía hasta en los frutos de una misma inflorescencia y que no tiene relación con el número de semillas por baya. (Tabla N º 2-74). Tabla N º 2-74. Corpúsculos pétreos por fruto en Complejo Solanum nigrum Número por fruto VMF 0 0 SOLNI – SOLNI-I 1 - 15 7 (8 – 9) SOLLO VMF: valor más frecuente 238 Disposición en la pulpa del fruto: En Solanum lorentzii, en los cortes longitudinales del fruto se observó una disposición constante: 2 corpúsculos apicales y dos en los lados opuestos de la mitad de la baya. Generalmente el número es mucho mayor de cuatro; se encuentran en la pulpa mezclados con las semillas. No coincide con la disposición descripta por Morton (1976). Color de los corpúsculos: Nada se encontró sobre este carácter. El color de los corpúsculos es blanco brillante en la pulpa de los frutos. (Figura 2-59). En cambio cuando son extraídos y lavados tienen un color marfil o crema y son opacos. (Figura 2-60). Naturaleza de los corpúsculos: Sobre la muestra de los corpúsculos, enviada para su análisis, se informó que son concreciones amorfas de naturaleza orgánica, sin determinación química. La naturaleza orgánica se confirmó coloreándola con safranina. Figura 2-60. Corpúsculos pétreos de Solanum lorentzii Figura Nº 2-59. Semillas, pulpa y corpúsculos pétreos Figura Nº 2-60. Corpúsculos pétreos sin colorear y coloreados con safranina Fotografías tomadas con lupa binocular (Marca Carl Zeiss 100 x) 239 Semilla Corpúsculos pétreos Figura Nº 2-61. SOLLO: semilla y corpúsculos pétreos. Fotografía tomada con Microscopio modelo Axiolab de polarización Carl Zeiss 100x CARACTERES CITOLÓGICOS 70- Número cromosómico El número cromosómico de Solanum nigrum, citado en la bibliografía europea (Edmonds y Chweya, 1997) y norteamericana (Ogg y Rogers, 1989), es 72. Allí se menciona también la poliploidía de estas especies. Los resultados muestran que las poblaciones del Complejo Solanum nigrum estudiadas en Tucumán son diploides, con un número de cromosómico 2n= 24, que concuerda con el número básico x= 12 citado para el género. 240 ANEXO CAPÍTULO 2 CARACTERES VEGETATIVOS 1.- ALTURA BIFURCACION DEL TALLO (AB) 1,2 IC-95% de la Altura de Bifurcacion del Tallo (m) 1,0 ,8 ,6 ,4 ,2 N= 36 6 S OLNI S OLNI-I 32 S OLLO Figura N° 2-2. Altura de la bifurcación del tallo de las diferentes especies del Complejo Solanum nigrum. Tabla N° 2-7. Análisis de la altura de la bifurcación de tallos del Complejo Solanum nigrum. Altura de Bif urcacion del Tallo (m) Inter-grupos Intra-grupos Total SC 4,527 3,426 7,953 gl 2 71 73 CM 2,263 ,048 F 46,906 Sig. ,000 Tabla N° 2-8. Pruebas estadísticas para el análisis de la altura de bifurcación del tallo en las distintas especies. Subconjuntos Homogéneos - Altura de Bi furcacion del Tallo (m) a,b HSD de Tukey ESPECIE SOLNI-I SOLNI SOLLO Sig. N 6 36 32 Subconjunto para alf a = .05 1 2 ,3850 ,4009 ,8978 ,981 1,000 a. Usa el tamaño muestral de la media armónica = 13,292. b. Los tamaños de los grupos no son iguales. Se utilizará la media armónica de los tamaños de los grupos. Los niv eles de error de tipo I no están garantizados. 241 2. - LONGITUD (LL), ANCHO (AL) y LONGITUD DEL PECIOLO (LP) DE LA HOJA 6,0 IC-95% de la Longitud de Hoja (cm) 5,8 5,6 5,4 5,2 5,0 N= 102 SOLNI 130 SOLLO Figura N° 2- 14. Longitud de las hojas para las diferentes especies. 3,0 IC-95% del Ancho de Hoja (cm) 2,8 2,6 2,4 2,2 2,0 1,8 N= 102 SOLNI 130 SOLLO Figura N° 2- 15. Ancho de las hojas para las diferentes especies. 1,6 1,5 IC-95% del PEC 1,4 1,3 1,2 1,1 1,0 ,9 ,8 N= 102 SOLNI 130 SOLLO Figura N° 2- 16. Longitud del pecíolo para las diferentes especies. 242 Tabla N° 2- 19. Análisis de longitud, ancho y pecíolo de las hojas en las diferentes especies del Complejo Solanum nigrum. Longitud de Hoja (cm) Ancho de Hoja (cm) PEC ESPECIE SOLNI SOLLO SOLNI SOLLO SOLNI SOLLO N 102 130 102 130 102 130 Media 5,6078 5,4177 2,8216 2,1469 1,4314 1,0292 DE 1,08561 1,49087 ,54202 ,68655 ,52407 ,54736 EE ,10749 ,13076 ,05367 ,06021 ,05189 ,04801 Tabla N° 2- 20. Pruebas estadísticas para el análisis de longitud, ancho y pecíolo de las hojas en las distintas especies. Prueba de Lev ene para la igualdad de v arianzas Longitud de Hoja (cm) Ancho de Hoja (cm) PEC Se han asumido v arianzas iguales No se han asumido v arianzas iguales Se han asumido v arianzas iguales No se han asumido v arianzas iguales Se han asumido v arianzas iguales No se han asumido v arianzas iguales F Sig. 3,907 3,425 ,534 ,049 ,066 ,466 Prueba T para la igualdad de medias t Sig. (bilateral) gl 1,082 230 ,280 1,123 228,805 ,262 8,132 230 ,000 8,364 229,988 ,000 5,659 230 ,000 5,689 221,079 ,000 243 CARACTERES REPRODUCTIVOS 3.- PEDÚNCULO (LPD) 2,0 IC-95% del Pedúnculo (cm) 1,8 1,6 1,4 1,2 1,0 N= 47 30 SOLNI SOLLO Figura N° 2-21. Longitud del pedúnculo de la inflorescencia para las diferentes especies del Complejo Solanum nigrum. Tabla N° 2-26. Análisis de longitud del pedúnculo en las diferentes especies. Pedúnculo ESPECIE SOLNI SOLLO N 47 30 Media 1,1615 1,4867 DE ,25949 ,77537 EE ,03785 ,14156 Tabla N° 2-27. Pruebas estadísticas para el análisis de longitud pedúnculo en las distintas especies. Prueba de Lev ene para la igualdad de v arianzas Pedúnculo Se han asumido v arianzas iguales No se han asumido v arianzas iguales F 47,271 Sig. Prueba T para la igualdad de medias t ,000 Sig. (bilateral) gl -2,659 75 ,010 -2,219 33,188 ,033 244 4.- LONGITUD DE SEPALOS (en la flor) (KL) 3,2 IC-95% de la Longitud Sepalos (mm) 3,0 2,8 2,6 2,4 2,2 2,0 1,8 N= 100 147 SOLNI SOLLO 10 SOLNI-I Figura N° 2-22. Longitud de los sépalos para las diferentes especies del Complejo Solanum nigrum. Tabla N° 2-32. Análisis de longitud de los sépalos en las diferentes especies del Complejo Solanum nigrum. Longitud Sepalos (mm) Inter-grupos Intra-grupos Total SC 18,543 38,142 56,685 gl 2 254 256 CM 9,272 ,150 F 61,743 Sig. ,000 Tabla N° 2-33. Pruebas estadísticas para el análisis de longitud de los sépalos en las distintas especies. Subconjuntos Homogéneos según Longitud Sepal os (mm) a,b HSD de Tukey ESPECIE SOLNI SOLLO SOLNI-I Sig. N 100 147 10 Subconjunto para alf a = .05 1 2 3 2,0090 2,5204 2,8700 1,000 1,000 1,000 a. Usa el tamaño muestral de la media armónica = 25,684. b. Los tamaños de los grupos no son iguales. Se ut ilizará la media armónica de los tamaños de los grupos. Los niv eles de error de tipo I no están garantizados. 245 5.- LONGITUD DEL CÁLIZ (en la flor) (KL) 2,7 IC-95% de la Longitud del Caliz (mm) 2,6 2,5 2,4 2,3 2,2 2,1 2,0 1,9 N= 110 147 SOLNI SOLLO Figura N° 2- 23. Longitud del cáliz para las diferentes especies del Complejo Solanum nigrum. Tabla N° 2-34. Análisis de long. del cáliz en las diferentes especies del Complejo Solanum nigrum. Longitud del Caliz (mm) ESPECIE SOLNI SOLLO N 110 147 Media 2,0873 2,5204 DE ,36299 ,45720 ES ,03461 ,03771 Tabla N° 2-35. Pruebas estadísticas para el análisis de longitud del cáliz en las distintas especies. Prueba de Lev ene para la igualdad de varianzas Longitud del Caliz (mm) Se han asumido varianzas iguales No se han asumido varianzas iguales F 8,813 Sig. ,003 Prueba T para la igualdad de medias t gl Sig. (bilateral) -8,189 255 ,000 -8,462 254,080 ,000 246 6. -LONGITUD DE LOS DIENTES (KLD) (en la flor) IC-95% de la Longitud de los Dientes (mm) 1,6 1,4 1,2 1,0 ,8 ,6 N= 100 SOLNI 99 SOLLO Figura N° 2-24. Longitud de los dientes del cáliz en la flor para las diferentes especies del Complejo Solanum nigrum. Tabla N° 2-36. Análisis de longitud de los dientes en las diferentes especies del Complejo Solanum nigrum. Longitud de los Dientes (mm) ESPECIE SOLNI SOLLO N 100 99 Media ,7288 1,4545 DE ,05885 ,25245 EE ,00588 ,02537 Tabla N° 2-37. Pruebas estadísticas para el análisis de longitud de los dientes en las distintas especies. Prueba de Lev ene para la igualdad de v arianzas Longitud de los Dient es (mm) Se han asumido v arianzas iguales No se han asumido v arianzas iguales F 199,528 Sig. ,000 Prueba T para la igualdad de medias t gl Sig. (bilatera l) -27,991 197 ,000 -27,865 108,517 ,000 247 7.- LONGITUD DE ESTAMBRES (ALF)y ANCHO DEL FILAMENTO (AAF) 1,0 5,0 4,5 3,5 3,0 2,5 IC-95% del Ancho del Filamento 4,0 ,9 ,8 ,7 ,6 ,5 2,0 ,4 1,5 N= 50 N= 51 SOLNI 60 60 SOLNI SOLLO SOLLO Figuras N° 2-40 (izq.) y Nº 2-41 (der): Longitud de los estambres y ancho de los filamentos para las diferentes especies del Complejo Solanum nigrum. Tabla N° 2-42. Análisis de longitud de los estambres y ancho de los filamentos en las diferentes especies del Complejo Solanum nigrum. Longitud del Estambre (mm) Ancho del Filamento ESPECIE SOLNI SOLLO SOLNI SOLLO N 50 51 60 60 Media 2,2440 4,2520 ,5150 ,8717 DE ,63957 ,37536 ,07988 ,15304 EE ,09045 ,05256 ,01031 ,01976 Tabla N° 2-43. Pruebas estadísticas para el análisis de longitud de los estambres y ancho de los filamentos en las distintas especies. Prueba de muestras i ndependientes Prueba de Lev ene para la igualdad de v arianzas Longitud del Estambre (mm) Ancho del Filamento Se han asumido v arianzas iguales No se han asumido v arianzas iguales Se han asumido v arianzas iguales No se han asumido v arianzas iguales F 1,496 22,375 Sig. ,224 ,000 Prueba T para la igualdad de medias t Sig. (bilateral) gl -19,288 99 ,000 -19,194 78,868 ,000 -16,003 118 ,000 -16,003 88,929 ,000 248 8.- LONGITUD (SL) DE LAS SEMILLAS 1,5 IC-95% de la Longitud Semilla (mm) 1,4 1,3 1,2 1,1 1,0 ,9 N= 225 50 SOLLO SOLNI-I 225 SOLNI Figura N° 2-55. Longitud de las semillas para las diferentes especies del Complejo Solanum nigrum. Tabla N° 2-65. Análisis de la longitud de las semillas en las diferentes especies. Longitud Semilla (mm) Inter-grupos Intra-grupos Total SC 12,547 20,385 32,932 gl 2 497 499 CM 6,274 ,041 F 152,961 Sig. ,000 Tabla N° 2-66. Pruebas estadísticas para el análisis de la longitud de las semillas en las distintas especies. Subconjuntos Homogeneos según Longitud Semi lla (mm) a,b HSD de Tukey ESP SOLLO SOLNI-I SOLNI Sig. N 225 50 225 Subconjunto para alf a = .05 1 2 3 1,0236 1,2580 1,3547 1,000 1,000 1,000 a. Usa el tamaño muestral de la media armónica = 103,846. b. Los tamaños de los grupos no son iguales. Se ut ilizará la media armónica de los tamaños de los grupos. Los niv eles de error de tipo I no están garantizados. 249 9.- ANCHO DE LAS SEMILLAS (SA) 1,2 IC-95% del Ancho Semilla (mm) 1,1 1,0 ,9 ,8 ,7 N= 225 225 SOLLO SOLNI 50 SOLNI-I Figura N° 2-56. Ancho de las semillas para las diferentes especies del Complejo Solanum nigrum. Tabla N° 2-67. Análisis del ancho de semillas en las diferentes especies. Ancho Semilla (mm) SC 10,500 11,603 22,103 Inter-grupos Intra-grupos Total gl 2 497 499 CM 5,250 ,023 F 224,871 Sig. ,000 Tabla N° 2-68. Pruebas estadísticas para el análisis del ancho de semillas en las distintas especies. Subconjuntos Homogéneos segúnAncho Semilla (mm) a,b HSD de Tukey ESP SOLLO SOLNI SOLNI-I Sig. N 225 225 50 Subconjunto para alf a = .05 1 2 ,7476 1,0320 1,0660 1,000 ,245 a. Usa el tamaño muestral de la media armónica = 103,846. b. Los tamaños de los grupos no son iguales. Se utilizará la media armónica de los tamaños de los grupos. Los niv eles de error de tipo I no están garantizados. 250 CAPÍTULO 3 GERMINACIÓN DE SEMILLAS Y EMERGENCIA DE PLÁNTULAS 3.1. INTRODUCCIÓN En la lucha contra las malezas, para lograr eficiencia y proteger el medio ambiente, es necesario conocer el hábito de crecimiento y ciclo de vida de las mismas, para evitar que lleguen a producir semillas y puedan difundirse (Kogan, 1992). Una maleza no constituye una clase botánica particular sino que es una población vegetal espontánea, que exhibe características propias para un sistema, en un determinado lugar y tiempo. Las malezas son una forma especial de vegetación altamente exitosa en ambientes agrícolas. Son poblaciones vegetales que crecen en ambientes perturbados por el hombre sin haber sido sembradas y que causan numerosos problemas en los cultivos siendo el impacto más crítico el efecto negativo ejercido a través de la competencia por recursos limitados y las alelopatías (Leguizamón, 2003). En la tierra habitan aproximadamente 250.000 fanerógamas, menos de 250 se consideran importantes en amplios sectores del planeta y de estas, unas 70 especies se consideran "las peores malezas del mundo". Estas malezas están distribuidas en 30 familias y en 5 de ellas, Poáceas, Solanáceas, Convolvuláceas, Euforbiáceas y Fabáceas, se encuentran las especies que suministran el 75 % del alimento mundial. Esta observación implica que los cultivos y las malezas comparten características taxonómicas y orígenes evolutivos comunes (Holm et al., 1977; Leguizamón, 2003). El bajo número de especies consideradas maleza en relación con el total, demuestra la tremenda presión selectiva ocurrida desde tiempos remotos y los hechos que suceden en la fase regenerativa de las plantas (dispersión de las semillas, dormición, germinación y establecimiento de las plántulas), son particularmente importantes para las malezas en ambientes agrícolas. Otros autores como Lambelet-Haueter (1985, 1986); Barralis y Chadoeuf (1987); Mayor y Dessaint (1998), aseguran que el banco de semillas es el mayor indicador de la influencia de las prácticas agronómicas sobre las malezas, más que la vegetación existente por encima del suelo. Para Grundy y Mead (2000) el banco de semillas del suelo constituye la principal fuente de infestación de malezas que tienen los cultivos. Según Allmaras y Dowdy (1985), aparte de la longevidad específica, las semillas incorporadas al suelo permanecen más tiempo viables que las que quedan en la superficie, en especial si el suelo es biológicamente activo, lo que ocurre cuando su superficie se mantiene con restos de los cultivos precedentes. El conocimiento del banco de semillas, cuanti y cualitativamente, es de interés para la evaluación del impacto de diferentes técnicas agronómicas (tratamientos herbicidas, laboreos mecánicos, mulching, empleo del fuego, etc.) y para el estudio de la dinámica de las poblaciones de malas hierbas. El conocimiento de la composición del banco de semillas permite el análisis de la infestación en un momento y situación dada (Dorado et al., 1997; Zanin et al., 1989; Chaila, 2001). 251 Sahoo et al. (1994) demostraron que la dinámica de población de las semillas de malezas está afectada por el tipo de especie que se cultiva y los sistemas de labranza empleados. De esa forma, de acuerdo al sistema de cultivo, la presencia de las especies será diferente tanto en número como en viabilidad. Mayor y Dessaint (1998) dicen que en un sistema de producción integrada con manejo orgánico del cultivo, la presencia de grandes cantidades de semillas de especies infestantes en el banco del suelo, influirá sobre los rendimientos, haciéndolos decrecer dramáticamente afectando las ganancias en forma considerable. 3.2. ANTECEDENTES Son numerosos los antecedentes sobre estudios de la biología de las especies del Complejo Solanum nigrum en Norte América (EEUU y Canadá), Europa, Asia, África y Australia (Holm et al., 1977; Roberts y Lockett, 1978; Ogg et al., 1981, 1989; Keeley y Thullen, 1989, 1991; McGiffen y Masiunas, 1992; del Monte y Tarquis, 1997; Edmonds y Chweya, 1997). La mayoría de los autores coinciden en que estas plantas sólo se reproducen por semillas y producen una gran cantidad de éstas. Holm et al. (1977) manifiestan que Solanum nigrum puede llegar a producir 178.000 semillas por planta. Los estudios que se han realizado para comprender algunos de los aspectos de la biología de las especies del Complejo Solanum nigrum son: 3.2.1. La longevidad de las semillas: Las experiencias realizadas en el Hemisferio Norte sobre la emergencia de las especies del Complejo Solanum nigrum, demostraron que las semillas de estas malezas pueden permanecer viables en el suelo por períodos prolongados. Esto depende de la situación en y del sustrato (profundidad a que se encuentran enterradas, suelo alterado o no). Holm et al. (1977) señalaron que las semillas de Solanum nigrum tienen una gran pérdida de viabilidad a través de los años. Estas, cuando se almacenaron a temperatura ambiente por dos años, alcanzaron un 27 % de germinación y luego de 8 y 9 años las semillas sólo germinaron 2 y 0 %, respectivamente. 3.2.2. Las condiciones necesarias para la germinación: Varios autores determinaron los factores que interactúan en el proceso de germinación de semillas frescas o almacenadas del Complejo Solanum nigrum. Estos son: Temperatura: constante o alternas en diferentes rangos; Luz: constante, intermitente, oscuridad y Humedad: en situaciones de almacenaje seco o húmedo (Ogg et al., 1989; Roberts y Lockett, 1978; del Monte y Tarquis, 1997). Algunos autores coincidieron en que la germinación se produce entre 20 y 40º C, con una temperatura óptima de 30º C y que con temperaturas por encima de los 45º C las semillas mueren (Ogg et al., 1989; Holm et al., 1977; Roberts y Lockett, 1978). 3.2.3. Además, se refieren a la necesidad y los tratamientos de ruptura de la dormancia, la época de emergencia y las condiciones que la favorecen: En general las semillas de Solanum nigrum deben sufrir algún tipo de tratamiento poscosecha para que germinen. Se ha determinado que un 99 % lo hace en primavera, luego de la exposición a 252 temperaturas invernales (Holm et al., 1977) y que están en condiciones de germinar entre 2 y 8 semanas después de la antesis, con un promedio de 4 semanas y llegan a un porcentaje de germinación del 100 % a los 32 días (Ogg et al., 1989; Roberts y Lockett, 1978). Los experimentos sobre ruptura de dormancia y de emergencia demostraron que en condiciones de laboratorio e invernáculo, los resultados, en general, son semejantes al patrón de la emergencia en el campo de especies de malezas de ciclo anual (Ogg et al., 1989; Roberts y Lockett, 1978; Edmonds y Chweya, 1997). 3.2.4. Diferentes situaciones que afectan la emergencia, el crecimiento y la producción de frutos y semillas: En general, en el hemisferio norte las especies del Complejo Solanum nigrum comienzan a emerger en primavera (marzo) cuando la temperatura máxima diaria del ambiente se aproxima a 20º C y la temperatura del suelo, a una profundidad de 5 cm, alcanza los 17º C. Las experiencias de Ogg et al. (1989) y Roberts y Lockett (1978) en suelos no disturbados, en el campo y en macetas en invernaderos, determinaron que son muy sensibles a las bajas temperaturas (heladas) y que la emergencia de Solanum nigrum se inicia a principios de mayo (primavera), declina en agosto, cesando en septiembre (otoño). En condiciones de invernáculo, entre los 40 y 55 días de realizada la siembra comienza la floración y a los 70 días inicia la maduración de los frutos. En condiciones de campo ocurre a los 60 días de la siembra (Roberts y Lockett, 1978; Keeley y Thullen, 1989, 1991). Con respecto a profundidad en que se encuentran las semillas en el suelo se afirma, que en general, las plántulas no emergerán si están enterradas más de 5 cm. Todos estos experimentos se realizaron en el hemisferio Norte donde el Complejo Solanum nigrum ha sido ampliamente estudiado mientras que en el hemisferio Sur no se encontraron referencias sobre la biología de Solanum nigrum y las especies relacionadas. 3.2.5. Por lo expuesto, los objetivos son: Conocer el ciclo de vida de las especies que forman el Complejo Solanum nigrum. Determinar cuales son las condiciones que favorecen la germinación y la emergencia de las plántulas del Complejo Solanum nigrum. Analizar comparativamente la germinación del Complejo Solanum nigrum. Comparar el comportamiento de estas malezas en condiciones naturales y en condiciones artificiales. Cotejar si la época de emergencia de las especies que forman un complejo propio del área agrícola de la provincia de Tucumán influye en su importancia como malezas. 253 3.3. MATERIALES y MÉTODOS Se siguió el método de Roberts y Lockett (1978) para estudiar la germinación de las semillas y la emergencia de las plántulas. Se experimentaron los factores que favorecen la germinación en las especies del Complejo Solanum nigrum, en semillas frescas y en semillas almacenadas, la emergencia estacional de las plántulas, la emergencia en función de la profundidad, la pérdida de viabilidad y la dormancia. En la caracterización de la germinación de los ensayos se utilizaron los parámetros de: - Porcentaje de germinación: es la fracción de semillas germinadas del número total de semillas. PG = (Ng x 100)/Nt PG = Porcentaje de germinación Ng = Nº de semillas germinadas Nt = Total de semillas sembradas - Germinación relativa: la fracción (x100) de semillas germinadas en un tiempo dado como porcentaje del total de semillas germinadas. Para las experiencias realizadas se consideraron las semillas que germinaron en 30 días. - Velocidad de germinación: tiempo para la germinación de un determinado número de semillas. G1 G2 G IVG = ..... n N1 N 2 Nn IVG = ÍNDICE DE VELOCIDAD DE GERMINACIÓN G = NÚMERO DE PLÁNTULAS NORMALES EN LOS DIFERENTES RECUENTOS N = NÚMERO DE DÍAS DESDE LA SIEMBRA AL CONTEO RESPECTIVO - Tasa de germinación: la inversa del tiempo de germinación de un porcentaje dado de semillas. TG = ( N1 .T1 ) ( N 2T2 ) ... ( N nTn ) Nt TG = Nº de semillas germinadas en “n” días Tn = Tiempo en días Nt = Nº de semillas germinadas totales - Demografía experimental: Los parámetros demográficos son: (expresados en ‰) TBM (tasa bruta de mortalidad): representa las plantas muertas por año. TBN (tasa bruta de natalidad): es la diferencia entre (1000 –TBM). Sv (Supervivencia): son las plantas que sobreviven dentro del año, obtenida de la diferencia entre vivas y muertas. PSv (Superv. probable): PSv expresa los individuos que sobrevivirán sobre 1000. 254 Para el PI (Potencial de Infestación) se empleó un modelo propuesto por Chaila (2001) que relaciona la viabilidad (test de tetrazolium a la mínima tinción), poder germinativo (en cámaras de laboratorio), las semillas ingresantes al banco y la densidad de la especie. El modelo se expresa: ( SemViab. P.Germ. N .sem.ingres.) PI dens 10000 PI= Potencial de Infestación SemViabl= Semillas viables P.Germ= poder germinativo N.Sem.ingres.= semillas ingresantes al banco Dens x 10000= densidad de la especie 3.3.2. Observación y recolección del material; Identificación de las especies La observación se hizo en el campo, en diferentes regiones agroecológicas (llanura chacopampeana, pedemonte y selvas de transición) durante las cuatro estaciones del año en predios con cultivo (hortícolas, cítricos, soja y caña de azúcar) y sin cultivo (banquinas, montes naturales, laderas de cerros). Se recolectó material de Solanáceas en estos predios eligiéndose plantas completas en estado reproductivo (con flores y frutos). Los ejemplares que estaban en estado vegetativo se extrajeron con pilón de tierra y se llevaron a invernáculo para continuar la observación del desarrollo completo de las plantas, luego cosechar frutos y por último separar las semillas. Identificación de las especies: En gabinete se identificaron con los métodos clásicos que evalúan caracteres morfológicos, caracteres anatómicos y caracteres cariológicos (Subils, 1977; Botta, 1979). Las plantas se herborizaron y se realizó un registro fotográfico de los caracteres de diferenciación más relevantes. De éstos, los de observación directa sirvieron en recolecciones posteriores para una rápida identificación a campo. 3.3.3. Obtención de semillas pertenecientes a las diferentes especies: La recolección de frutos se hizo en forma escalonada a lo largo del período de fructificación desde el año 2000 al presente. Para la extracción de las semillas se seleccionaron y recolectaron sólo frutos maduros. Se registró el origen de cada uno de los ejemplares, el estado fenológico y la fecha de recolección. Además, se tomó nota de detalles que pudieran perderse en el herborizado de las plantas, como el color de las bayas, la forma del cáliz y las características de éste en frutos verdes y maduros. 255 Para separar las semillas se formaron grupos de 50 bayas. Los frutos fueron aplastados suavemente en un mortero de porcelana. Se observó el color del jugo. Se agregó agua, se mezcló y luego se filtró con una malla para separar las semillas de la pulpa. Las semillas fueron lavadas con agua corriente durante treinta minutos dejándolas secar a temperatura ambiente sobre papel de filtro siguiendo el método de Roberts y Lockett (1978). Además, se probaron 15, 60 minutos y 3 horas. Una vez completado el secado se separaron grupos de 50 semillas para realizar siembras. Se aprovechó este material para observar además del color de las semillas, la forma y una aproximación del largo y el ancho de las mismas. 3.3.4. Métodos utilizados en la extracción de las semillas 3.3.4.1. Utilización inmediata de semillas: Las semillas se sacaron de las bayas arrastrándolas con una espátula para sembrarlas inmediatamente, sin lavarlas. 3.3.4.2. Secado de semillas a temperatura ambiente: Las semillas se extrajeron de las bayas maduras aplastándolas suavemente en un mortero de porcelana. Se agregó agua a esta pulpa, se mezcló y luego se separaron las semillas de la pulpa mediante un tamiz que permite sólo el paso de las semillas. Estas se colocaron en un filtro de malla fina y se lavaron con agua corriente durante 15, 30, 60 minutos y 3 horas. Se dejaron secar a temperatura ambiente sobre papeles absorbentes. 3.3.4.3. Extracción de semillas de frutos secados a temperatura ambiente: Los frutos se dejaron secar en bandejas a temperatura ambiente. Después de 2, 3 y 12 meses se hidrataron y se aplastaron en un mortero de porcelana. Usando un tamiz se separaron las semillas de la pulpa hidratada, se lavaron 30 minutos con agua corriente. Luego se las dejó secar sobre papeles de filtro, a temperatura ambiente durante 15 días aproximadamente. Las semillas se almacenaron en sobres de papel hasta el momento de su utilización. 3.3.4.4. Extracción de semillas de frutos conservados en frío hasta seis meses: Inmediatamente después de la cosecha los frutos fueron puestos en recipientes cerrados y conservados en heladera durante 10, 15, 30, 90 y 180 días. El almacenaje de 90 días simula el período invernal. Para la extracción y secado de las semillas se siguieron los pasos indicados en 3.3.4.3. 256 3.3.5. Métodos utilizados en la siembra de las semillas de las diferentes especies. Ensayos de germinación: (lugar – recipiente) Los ensayos de germinación de las especies del Complejo Solanum nigrum se realizaron en laboratorio y en invernáculo empleándose distintos tipos de recipientes: cajas de Petri, recipientes de plástico transparentes con tapa, macetas, cubeteras y recipientes de vidrio (vasos de precipitación y frascos). 3.3.5.1. En el laboratorio. Se utilizaron cajas de Petri y también cajas de plástico transparentes con tapa, con algodón y papel de filtro, vasitos de plástico con tierra y sin tapa, vasos de vidrio o de plástico transparente con agua. Las experiencias se llevaron a cabo en laboratorio, a temperatura ambiente y en germinador con temperatura controlada. En cada experiencia se utilizaron semillas de las especies identificadas como Solanum nigrum y Solanum lorentzii 3.3.5.1.1. Siembra inmediata: A. En cajas de Petri: Cincuenta semillas frescas, lavadas y sin lavar, fueron sembradas en cajas de Petri conteniendo como sustrato algodón y papel de filtro. Se efectuaron cuatro repeticiones en Solanum nigrum y cuatro en Solanum lorentzii B. En recipientes con tierra (macetas y bandejas): Una siembra casi superficial se efectuó en vasitos de plástico rellenados con tierra. También en este experimento fueron 4 repeticiones de 50 semillas en cada vaso y de las dos especies. Se colocaron en laboratorio a temperatura ambiente próximos a una ventana. Se regó cada vez que fue necesario empleando agua de red. C. En medio líquido: Para esta experiencia se colocaron cincuenta semillas frescas de cada especie en vasos de vidrio o de plástico transparente, con agua de red y con agua destilada. Se mantuvo el nivel de agua en los recipientes. Las condiciones fueron temperatura ambiente y luz natural (períodos de alternancia de luz y oscuridad). Se controló periódicamente la temperatura del ambiente. Las observaciones se hicieron durante 12 y en algunos casos hasta 18 meses. La variación de la temperatura y humedad relativa se muestra en la Tabla Nº 31. 257 Tabla Nº 3-1 Registros de Temperaturas. Sección Agrometeorología. EEAOC. Tucumán Lules-Período: ENERO 2002 - DICIEMBRE 2004 2002-2004 Tª MAX Tª MINTª MEDH.R.MAX H.R.MINH.R.MED 37,1 16,1 25,3 100 43 79 ENERO FEBRERO SIN DATOS 35,2 13,8 23,8 100 59 89 MARZO 31,3 8,9 18,8 100 38 88 ABRIL 29,5 8,5 17,4 100 60 89 MAYO 23,3 2,7 12,0 100 44 83 JUNIO JULIO SIN DATOS 35,4 1,8 17,1 97 18 69 AGOSTO 4,7 19,7 100 28 63 SEPTIEMBRE 34,6 39,0 8,2 23,3 100 27 72 OCTUBRE 12,4 23,6 NOVIEMBRE 36,6 SIN DATOS 35,8 12,1 23,4 DICIEMBRE 3.3.5.1.2. Siembra de semillas conservadas: Las experiencias se realizaron en condiciones de laboratorio durante 12 a 18 meses, con temperatura del ambiente y luz natural y en germinador con temperatura controlada y alternancia de luz y oscuridad, se registró diariamente, durante un mes, en cada temperatura regulada en el aparato. 3.3.5.1.2.1. En cajas de Petri a temperatura ambiente Se sembraron 50 semillas de Solanum nigrum y de Solanum lorentzii por recipiente, con cuatro repeticiones en cada caso. Se utilizó algodón, dos hojas de papel de filtro y se mantuvo la humedad. Se formaron grupos y subgrupos de semillas para efectuar las siembras: 1er grupo: siembra con semillas frescas (sin almacenaje o siembra inmediata). Se incluyeron semillas frescas para poder comparar con los otros grupos. 2º grupo: semillas almacenadas a temperatura ambiente y en ambiente seco. Una porción de semillas se lavó con agua y se dejó secar, la otra se conservó sin lavar. (almacenaje seco) 3er grupo: semillas almacenadas en heladera en recipientes de plástico durante 10, 15, 30, 90 y 180 días. (almacenaje seco con frío de 4ºC). 4º grupo: semillas húmedas. Se almacenaron las bayas enteras durante 30, 60, 90 y 180 días. (almacenaje húmedo) 258 3.3.5.1.2.2. En Germinador En estas experiencias las semillas utilizadas tenían diferentes condiciones de conservación (cosecha reciente, temperatura ambiente, frío de 4 ºC). En cada experiencia se sembraron 50 semillas de Solanum nigrum y 50 de Solanum lorentzii. Se efectuaron cuatro repeticiones de cada una. Se utilizaron cajas de Petri con algodón y papel de filtro. Los recipientes se mantuvieron tapados humedeciéndolos periódicamente con agua de red. Las pruebas se hicieron con diferentes temperaturas (18, 20, 25, 30 y 35º C) y con luz natural. Se observó diariamente durante 1 mes. 3.3.5.1.3. En estufa, en oscuridad: Grupos de 50 semillas lavadas y sembradas en cajas de Petri, se mantuvieron en oscuridad, en estufa a 27º C durante 1, 2 y 3 meses. Las siembras se hicieron sobre algodón cubierto con papel de filtro y las cajas envueltas con papel negro. Se mantuvo la humedad con agua de red. En los registros la iluminación fue con luz de lámpara fluorescente. 3.3.5.1.4. Germinación de una especie intermedia. Como se dijo en el capítulo 2, el tipo de inflorescencia es un carácter constante que permite diferenciar a las especies. En algunas de estas plantas se había observado un detalle en las inflorescencias que no era el normal. Por ello, se recolectaron frutos maduros de plantas de Solanum nigrum (SOLNI) y de otras plantas de Solanum nigrum (SOLNI-I), que presentan un tipo diferente de inflorescencia. Esta característica en la inflorescencia, se había observado en varias muestras, pero al inicio del trabajo no se le dio importancia y por eso, en ese momento no se separaron las semillas. 3.3.5.1.4.1. En tierra En el año 2003, se procesaron y se sembraron en macetas con tierra. Se utilizaron semillas de inflorescencias cimosas (Solanum nigrum), y las de inflorescencias bifurcadas encontradas en la misma planta. Se hicieron 5 repeticiones con 10 semillas por maceta a 1 cm de profundidad que fueron observadas y registrada la emergencia de las plántulas diariamente durante 3 meses. Muestra 1: semillas de una inflorescencia bifurcada conservada durante 8 meses a temperatura ambiente. 259 Muestra 2: semillas de una inflorescencia bifurcada que se cosecharon, lavaron inmediatamente de separadas de la baya y luego se sembraron. Muestra 3: semillas de una inflorescencia cimosa de la misma planta que la Muestra 2 3.3.5.1.4.2. En agua También se probó la germinación de las semillas de las inflorescencias bifurcadas en agua de red y en agua destilada. 3.3.5.2. En invernáculo y a la intemperie. Los recipientes utilizados en estas siembras fueron macetas y cubeteras con tierra. 3.3.5.2.1. Siembra: La siembra fue casi superficial efectuándose riegos periódicos con agua de red, dependiendo de la temperatura y humedad ambiente. En cada maceta llenada con tierra, se hicieron 4 repeticiones de 50 semillas en cada una. La mitad de las macetas se mantuvo en invernáculo, y la otra, a la intemperie. En otros recipientes, con tierra y con semillas del mismo origen se probó el humedecimiento con agua destilada. Las siembras en macetas se observaron durante 3 meses (octubre a enero) y en las bandejas 6 meses (junio a diciembre). Macetas: Siembra: 50 semillas sembradas al voleo en cada maceta. Detalles: 4 macetas a la intemperie 1-2 SOLNI – 1-2 SOLLO. Riego diario. 4 macetas en el invernáculo. 3-4 SOLNI – 3-4 SOLLO. Riego cuando están secas. Cubeteras: Siembra: En cada hueco de las cubeteras se sembraron 10 semillas, (48 huecos). Detalles : Total 480 semillas de SOLNI y la misma cantidad de SOLLO. 3.3.5.2.2. Trasplante: El propósito del trasplante fue disponer de plantas donde se completaran las observaciones de los caracteres taxonómicos y obtener semillas para los ensayos en forma rápida y permanente. De una población numerosa de Solanum sp. se seleccionaron y trasplantaron en macetas plantas con dos o tres hojas verdaderas. Los lugares donde se recolectó material se presentan en la Tabla Nº 3-2. Tabla Nº 3-2. Sitios de recolección de ejemplares del Complejo Solanum nigrum Lugar Finca El Manantial - FAZ Huerta Cultivo Fecha 09/10/01 Especie SOLNI 260 Finca El Manantial - FAZ Quinta Agronómica - FAZ Quinta Agronómica - FAZ Yerba Buena Yerba Buena Durazneros Parcelas de Botánica Sin cultivo (Ficus seco) Sin cultivo Limoneros 09/10/01 20/09/01 13/08/01 13/09/01 22/09/01 SOLLO SOLNI SOLNI SOLLO SOLLO A un grupo de macetas se las mantuvo en el exterior (terraza) y a otro en invernáculo, a temperatura ambiente, observándose la evolución de las plantas durante seis meses. Se llevó un registro de las que vegetaron, de las que florecieron y de las que llegaron a producir frutos. Se completaron observaciones de aspectos taxonómicos de las especies del complejo. Estos datos se compararon con las plantas que siguieron creciendo en los predios de donde se extrajeron las plantas para trasplante. Los frutos maduros se cosecharon y procesaron para realizar las pruebas de germinación. En general en este período de seis meses el tiempo fue muy húmedo y caluroso, no se controló temperatura. Ver los promedios de los registros meteorológicos en Cap. 3 - Tabla 3-1 - Pág. 8. 3.3.6. Emergencia en función de la profundidad: En invernáculo y en laboratorio se sembraron en macetas llenadas con tierra a 0; 2,5; 5 y 10 cm de profundidad. La tierra se recolectó de la huerta de Finca el Manantial (FAZ) y de la Quinta Agronómica (FAZ) de los lugares donde había grupos de estas malezas. Se hicieron 4 repeticiones con 50 semillas en cada uno. Se regó con agua de red. 3.3.7. Emergencia a campo y en laboratorio. Se observó la emergencia de las especies del Complejo Solanum nigrum a lo largo de un año. 3.3.8. Plantación de estacas En el mes de setiembre se hicieron plantaciones de estacas de Solanum nigrum y de Solanum lorentzii. Se enterraron 20 estacas de cada especie en macetas con tierra, (cinco por maceta). Se mantuvo a la intemperie con riegos periódicos observándolas semanalmente durante dos meses. 261 3.4. RESULTADOS Y DISCUSIÓN Siembra Inmediata En las muestras sembradas inmediatamente después de cosechar, en diferentes sustratos, sin procesar y procesadas (separadas en mortero y luego lavadas y secadas) los resultados fueron los siguientes: En cajas de Petri: Tabla Nº 3-3. Registro de germinación en siembra inmediata del Complejo Solanum nigrum Repeticiones Fechas de observación 03/06/03 (siembra) 12/06/03 24/06/03 07/07/03 04/06/04 1 SOLNI 2 3 4 1 0 0 0 0 0 0 0 0 0 7 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 SOLLO 2 3 0 0 0 0 0 0 0 0 0 0 4 0 0 0 0 0 De las 4 repeticiones de Solanum nigrum sólo en una caja de Petri, a casi un año de la siembra, de las 50 germinaron 7 semillas. El papel de filtro estaba cubierto de hongos y apenas húmedo. (Tabla Nº 3-3; Fig. Nº 3-1). Fig. Nº 3-1. SOLNI: Semillas germinadas luego de un año de sembradas Las semillas que germinaron fueron las del grupo que se habían lavado. La germinación se produjo al año de sembradas. No hubo germinación en las sin lavado. No germinaron las semillas de Solanum lorentzii. - Análisis del porcentaje de germinación: TOTAL: 200 semillas SOLNI: PG = (7x100)/200 = 3,5% SOLLO: 0 % En los trabajos consultados no se encontraron experiencias sobre germinación en siembra directa de semillas del Complejo Solanum nigrum. 262 En recipientes con tierra (macetas y bandejas): En semillas sembradas inmediatamente después de separadas, con y sin lavado, no hubo germinación. Tampoco se encontraron antecedentes de siembras inmediatas en tierra. Como el resultado es 0 % de germinación, no se analizan los parámetros. En medio líquido: Un elevado porcentaje de semillas colocadas en recipientes de vidrio con agua de red germinaron en algunos casos en forma escalonada, en otros varios meses más tarde (hasta 8 meses). Tabla Nº 3-4 Solanum nigrum - Tabla Nº 3-6 Solanum lorentzii. No hubo germinación cuando se usó agua destilada. Se analizaron los siguientes parámetros: Porcentaje de germinación (PG), Porcentaje de germinación relativa (PGR) y Tasa de germinación (TG). No se analiza la velocidad de Germinación (VG) porque en el Modelo de Maguire (1962), la temperatura debe ser constante. (Tabla Nº 3-5 y Tabla Nº 3-7). Tabla Nº 3-4. Germinación de Solanum nigrum en medio líquido, en siembra inmediata. 1.- SOLNI (La Cruz) Fechas de observación Días 12/2002 (siembra) 02/2003 60 03/2003 90 04/2003 120 052003 150 062003 180 07/2003 210 08/2003 240 ------------11/2003 330 12/2003 360 Total Nº 5 7 3 0 0 2 6 10 9 42 2.- SOLNI (El Manantial) Fechas de observación Días 11/2002 (siembra) 12/2002 ----02/2003 90 03/2003 120 04/2003 150 05/2003 180 06/2003 210 07/2003 240 08/2003 270 09/2003 300 10/2003 330 11/2003 360 12/2003 390 -----------08/2004 600 Total Nº 0 0 0 7 4 5 0 0 21 5 2 3 --5 47 3.- SOLNI (La Cruz) Fechas de observación Días 11/2003 (siembra) 12/2003 01/2004 02/2004 03/2004 04/2004 05/2004 180 06/2004 210 -------------11/2004 360 12/2004 390 Total Nº 0 0 0 0 0 5 10 3 3 21 Tabla Nº 3-5. Parámetros de germinación de Solanum nigrum en medio líquido, en siembra inmediata. Muestra 1 2 3 PG % 84 94 70 PGR % 0 0 0 VG TG ------------- 230,71 333,19 250,00 263 Tabla Nº 3-6. Germinación de Solanum lorentzii en medio líquido, en siembra inmediata. 1.- SOLLO (Yerba Buena) Fechas de Días Nº observación 08/2003 (siembra) 09/2003 30 6 10/2003 60 8 11/2003 90 3 12/2003 120 5 01/2004 150 0 02/2004 180 0 03/2004 210 0 --------------10/2004 420 10 11/2004 450 9 12/2004 480 4 Total 45 2.- SOLLO (El Manantial) Fechas de observación Días Nº 3.- SOLLO (La Cruz) Fechas de observación Días 10/2002 (siembra) ---------02/2003 03/2003 04/2003 05/2003 06/2003 07/2003 08/2003 ----------11/2003 12/2003 Total 11/2002 (siembra) --------02/2003 03/2003 04/2003 05/2003 06/2003 07/2003 08/2003 09/2003 10/2003 11/2003 Total ----120 150 180 210 240 270 300 ---390 420 Tabla Nº 3-7. Parámetros de germinación de Solanum inmediata. Muestra PG PGR % % 1 90 12 2 68 0 3 90 0 2 3 0 7 0 5 5 10 2 34 ----90 120 150 180 210 240 270 300 330 360 Nº 0 8 7 0 0 0 14 6 0 6 45 lorentzii en medio líquido, en siembra VG TG ------------- 260,00 263,51 216,66 Los registros se hicieron durante 12 hasta 18 meses. Los resultados de Siembra Inmediata (A, B y C), demuestran que las semillas cosechadas y sembradas inmediatamente no germinan, son dormantes, y que necesitan sufrir algún tipo de tratamiento poscosecha para que lo hagan. En esto se coincide con Roberts y Lockett (1978) y Ogg y Rogers (1989). Con los métodos utilizados los porcentajes de germinación fueron nulos o bajos salvo en la siembra que se utilizó sólo agua de red. La mayor cantidad de semillas germinadas fue de Solanum lorentzii. Para el Porcentaje de Germinación Relativa (PGR), 30 días es el tiempo de registro de germinación. Las que germinaron lo hicieron después de ese tiempo. No se analiza la Velocidad de Germinación (VG) porque en el modelo de Maguire (1962), la temperatura debe ser constante. La tasa de germinación (TG) de las especies es de mediana a baja, 216,66 – 263,51. lo que indica que las posibilidades de germinar de las especies es baja en el ambiente estudiado. Estos valores están vinculados a un porcentaje de germinación relativa bajo, es decir que en un tiempo dado (30 días iniciales) germinarán muy pocas o ninguna de las semillas del 264 Complejo. Del total de las semillas analizadas en los tratamientos en medio líquido el porcentaje de germinación es alto superando al 80 % (Tabla Nº 3-5 y Nº 3-7). Sobre este método (germinación en medio líquido), no se encontró referencia alguna. Figuras 3-2 a 3-5 germinación de SOLLO y Figuras 3-6, 3-7 y 3-8 germinación de SOLNI en medio líquido. GERMINACIÓN EN MEDIO LÍQUIDO Registro fotográfico Solanum lorentzii (SOLLO) Figuras Nº 3-2 – 3-3. Germinación de SOLL0 Figuras Nº 3-4 – 3-5. Germinación de SOLLO en medio líquido 265 266 Solanum nigrum (SOLNI) Fig.Nº 3-6 - 3-7 Germinación en agua de SOLNI Figura Nº 3-8. Germinación en agua de SOLNI 267 SIEMBRA DE SEMILLAS CONSERVADAS En los grupos de semillas conservadas los porcentajes de germinación fueron bajos. Se observaron durante 1, 3, 6 y 12 meses (y en algunos casos 18 meses). En algunas cajas se produjeron germinaciones luego de un año de la siembra y en medio de los hongos que cubrían el papel de filtro. 1er grupo. Sin almacenaje o siembra inmediata: en esta experiencia no se produjo germinación. Las semillas sin ningún tratamiento de conservación se sembraron inmediatamente después de la extracción. 2º grupo. Almacenaje seco: se hicieron 5 subgrupos de las semillas almacenadas a temperatura ambiente durante diferentes tiempos (10, 15, 30, 90 y 180 días) luego de lavadas. Los resultados de germinación de los subgrupos D y E se muestran en tablas. Se omitieron las tablas de los subgrupos A, B y C porque no hubo germinación. Subgrupos A: siembra después de 10 días del lavado. B: siembra después de 15 días del lavado. C: siembra después de 30 días del lavado. D: siembra luego de 90 días del lavado. E: siembra luego de 180 días del lavado. Se analiza el subgrupo“D: siembra luego de 90 días del lavado”. Se registraron las semillas germinadas durante 10 meses. Los valores obtenidos y el análisis de los parámetros de germinación en Tabla Nº 3-8. Tabla Nº 3-8. Análisis de los parámetros de germinación de semillas del Complejo Solanum nigrum conservadas a temperatura ambiente, 90 días. Repeticiones P G P G R % % 1 2 2 2 6 4 SOLNI 3 8 0 4 2 0 1 0 0 2 6 0 SOLLO 3 0 0 4 2 0 VG TG ------------------------------------------------- 30 50 75 150 0 180 0 240 Se analiza “E: siembra luego de 180 días del lavado. Se trabajó de la misma manera, con 4 repeticiones en cada especie. Se observó durante 10 meses. Los resultados y el Análisis de los parámetros se presentan en Tabla Nº 3-9. 268 Tabla Nº 3-9. Análisis de los parámetros de germinación de semillas del Complejo Solanum nigrum conservadas a temperatura ambiente, 180 días. Repeticiones P G P G R % % 1 6 0 2 2 0 SOLNI 3 2 0 4 6 4 1 8 0 2 10 2 SOLLO 3 10 0 4 14 2 VG TG ------90 ------- 120 ------90 ------50 ------90 ------- 108 ------- 132 ------- 167,14 Los registros totales de germinación (RTG), obtenidos de los cinco subgrupos en un año de observación aproximadamente, se reúnen en la TablaNº 3-10. Tabla Nº 3-10. RTG de semillas del Complejo Solanum nigrum conservadas a temperatura ambiente, durante 15 meses. Jun. Jul. Ago. Set. A 0603 0 0 0 0 B 0603 0 0 0 0 C 0803 D E E 0 2 0 0 Oct. Nov. Dic. Ene. Feb. Mar. Abr. May Jun . 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0903 3 0 2 0 3 0 0 1 1 0 0 0 0 2 0 1 0 0 0204 2 2 0 0 5 7 1 4 Continúan observaciones de E 0 6 0 0 Referencia de Colores: Gris: fechas de siembra - Rojo: SOLNI - Verde: SOLLO Se coincide con Roberts y Lockett (1978) y Ogg y Rogers (1989) sobre esta necesidad de un período de reposo que tienen las especies del Complejo Solanum nigrum. Estos resultados mostraron que luego de un reposo de 3 meses o más, recién comienzan a germinar aunque con resultados diferentes según las especies. En las condiciones de almacenaje seco y a temperatura ambiente, el porcentaje de germinación mayor (14 %) corresponde a Solanum lorentzii (SOLLO) cuando tuvo un periodo de 180 días de reposo luego de ser lavados. El porcentaje en Solanum nigrum (SOLNI) es más bajo y con porcentajes semejantes en ambos períodos de conservación, (90 y 180 días). Tanto para Solanum nigrum como para Solanum lorentzii los porcentajes de germinación son bajos y demoran un gran número de días en germinar. No se analizó la Velocidad de germinación (VG) porque en el modelo de Maguire (1962) la temperatura debe ser constante. 269 En este caso los valores de tasa de germinación son bajos y muy bajos debido a que los porcentajes semillas germinadas y los porcentajes de germinación relativa son bajos y muy bajos respectivamente. En este grupo, no se analizaron los parámetros de germinación en los subgrupos A, B, C, porque no hubo germinación. 3er grupo. Almacenaje seco con frío: Luego de lavadas y secas, las semillas se guardaron en recipientes cerrados durante 10, 15, 30, 90 y 180 días. Se conservaron en heladera a 4ºC. Se hicieron subgrupos de acuerdo con el tiempo de almacenado en heladera. Subgrupos: A: siembra después de 10 días de frío. B: siembra después de 15 días de frío. C: siembra después de 30 días de frío. D: siembra luego de 90 días de frío. E: siembra luego de 180 días de frío. “A, B y C” no se analizan porque la germinación fue 0 %. D: Siembra luego de 90 días de frío: Los valores obtenidos en 6 meses y el Análisis de los parámetros se observan en Tabla Nº 3-11. Tabla Nº 3-11. Análisis de los parámetros de germinación de semillas del Complejo Solanum nigrum conservadas 90 días a 4ºC Repeticiones P G P G R % % 1 0 0 2 2 0 SOLNI 3 8 0 4 0 0 1 4 0 2 2 0 SOLLO 3 10 0 4 0 0 VG TG ------------------------------------------ 0 90 75 0 90 120 90 0 E: siembra luego de 180 días de frío: son 4 repeticiones de 50 semillas en cada especie. El Análisis de los parámetros de la germinación observados durante 10 meses se presentan en Tabla Nº 3-12. Tabla Nº 3-12. Análisis de los parámetros de germinación de semillas del Complejo Solanum nigrum conservadas 180 días a 4ºC. Repeticiones P G P G R VG TG % % 1 22 22 ------- 21,90 2 0 0 -----0 SOLNI 3 4 4 -----21 270 4 1 2 3 4 SOLLO 4 4 8 24 4 4 4 6 10 4 ------ 15,5 -----21 ------ 37,5 ------ 184,83 ------ 15,5 En Tabla Nº 3-13 se muestra el registro total de la germinación (RTG) de la experiencia con semillas conservadas en frío a 4ºC. Tabla Nº 3-13. RTG de semillas del Complejo Solanum nigrum conservadas en frío, durante 15 meses. Jun. Jul. Ago. Set. A 0602 0 0 0 0 B 0602 0 0 0 0 C 0702 D E E 0 0 0 0 0 0 0902 Oct. Nov. Dic. Ene. Feb. Mar. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 7 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0203 Continúan observaciones de E 0 0 0 0 0 0 0 0 0 0 0 7 0 0 0 0 15 Abr. May. Jun . 0 0 0 0 0 0 0 0 0 0 12 0 0 0 1 0 0 Referencia de Colores: Gris: fechas de siembra - Rojo: SOLNI - Verde: SOLLO En las experiencias de conservación con bajas temperaturas se confirma lo expresado por Roberts y Lockett (1978) y Ogg y Rogers (1989). Estos resultados muestran que con un almacenaje en frío de 3 meses o más, recién se inicia la germinación. El área agrícola de Tucumán presenta temperaturas frías en invierno que favorecen a este período de reposo con temperaturas bajas pero que no son extremadamente frías como ocurre en el hemisferio norte. En condiciones de conservación en frío el porcentaje mayor de germinadas, 24 %, correspondió a Solanum lorentzii (SOLLO) cuando tuvo 180 días de frío. Y lo mismo ocurre en Solanum nigrum (SOLNI) que con 180 días en frío de 4 ºC la germinación alcanzó un porcentaje de 22 %. En estos experimentos la velocidad de germinación no se analiza. La tasa de germinación en estas condiciones es baja debido a que todos los porcentajes de germinación para el tratamiento son bajos. 4º grupo. Almacenaje de bayas enteras. (Almacenaje húmedo). Las semillas permanecieron dentro de las bayas durante 30, 60, 90 y 180 días. Una parte de la muestra se había conseró a temperatura ambiente y otra en heladera. Las que estuvieron almacenadas a temperatura ambiente durante 30 días se procesaron y se colocaron en recipientes con algodón y papel de filtro que se humedeció periódicamente con agua. 271 Hubo una pronta invasión de hongos. No se obtuvieron resultados de germinación. En las muestras de 60 días no se pudo trabajar por que al ser hidratadas y aplastadas se desintegraba el contenido. Más adelante, al abrir los restantes grupos de bayas se comprobó la pulverización total del interior. No se hicieron pruebas de germinación. En estas especies, no sirve la conservación de semillas en la humedad de las bayas. En la Tabla Nº 3-14 se comparan los porcentajes de germinación obtenidos de semillas conservadas a temperatura ambiente, en frío y en almacenaje húmedo. Tabla Nº 3-14. Comparación del Porcentaje de la germinación de semillas conservadas del Complejo Solanum nigrum Tiempo de conservación Temperatura ambiente Frío (4ºC) Almacenaje húmedo SOLNI 4,5 2,5 0 90 días SOLLO 2 4 0 180 días SOLNI SOLLO 4 10,5 7,5 10 0 0 De esta comparación surge que además del reposo que demandan las semillas para recién comenzar la actividad, necesitan un período de frío que es semejante al del invierno en la zona agrícola de Tucumán. Las semillas almacenadas en las bayas no sobreviven por la putrefacción que ocurre a lo largo de los diferentes tiempos de conservación. SIEMBRAS EN GERMINADOR con temperaturas constantes y luz alternada Para el análisis de los resultados se consideraron tres grupos. En cada grupo se analizaron las dos temperaturas en las que hubo germinación y emergencia de plántulas: 25 y 30º C. En cada caso la observación se hizo durante un mes. Los grupos analizados son: Primer grupo: A. siembra con semillas frescas (s. inmediata) a 25 º C. B. siembra con semillas frescas (s. inmediata) a 30 º C. Segundo grupo: B: siembra con semillas conservadas a tº ambiente a 25 º C. siembra con semillas conservadas a tº ambiente a 30 º C. 272 Tercer grupo: C: siembra de semillas conservadas en frío a 25ºC. siembra de semillas conservadas en frío a 30 º C. Primer grupo: A: siembra con semillas frescas (siembra inmediata) En la siembra a 25 ºC, no se analizan los parámetros porque hay 0 % de germinación. En siembra en germinador a 30º C de semillas frescas, con 4 repeticiones en cada especie se obtienen estos resultados: Tablas Nº 3-15 y 3-16 muestran la velocidad de germinación y Tabla Nº 3-17 el análisis de los parámetros. Tabla Nº 3-15 Velocidad de germinación de Solanum nigrum (30ºC) (Calculada utilizando el modelo de Maguire) Repeticiones 1 2 3 4 Días dr de 15 16 16 17 24 16 17 20 Plántulas Germinación IVG N° % 8 2 6 3 16 4 12 6 0,469 0,125 0,360 0,158 dr: días empleados para empezar a germinar de :días empleados para la germinación total de las semillas Tabla Nº 3-16 Velocidad de germinación de Solanum lorentzii (30ºC) (Calculada utilizando el modelo de Maguire) Repeticiones 1 2 3 4 Días dr de 15 15 17 8 20 15 17 16 Plántulas Germinación IVG N° % 3 1 2 3 6 2 4 6 0,216 0,066 0,360 0,250 dr: días empleados para empezar a germinar de :días empleados para la germinación total de las semillas Tabla Nº 3-17. Análisis de los parámetros de germinación de semillas del Complejo Solanum nigrum en germinador, siembra inmediata a 30ºC. Repeticiones P G P G R % % 1 16 16 2 4 4 SOLNI 3 12 12 4 6 6 1 8 6 2 10 2 SOLLO 3 10 4 4 14 6 Segundo grupo: VG TG 0,469 17,5 0,125 16 0,360 15,66 0,158 19 0,216 17 0,066 15 0,360 17 0,250 8 273 C. siembra de semillas conservadas a temperatura ambiente Se analiza la germinación de semillas conservadas a temperatura ambiente, sembradas en germinador a 25 º C. Tablas Nº 3-18 y 3-19. Sólo germinó Solanum lorentzii. Tabla Nº 3-18. Velocidad de germinación de Solanum lorentzii (25ºC) (Calculada utilizando el modelo de Maguire) Repeticiones Días dr de - 19 19 19 19 14 14 1 2 3 4 Plántulas Germinación IVG N° % 0 1 9 2 0 2 18 4 0 0,052 0,473 0,142 dr: días empleados para empezar a germinar de :días empleados para la germinación total de las semillas Tabla Nº 3-19. Análisis de los parámetros de germinación de semillas del Complejo Solanum nigrum , 25ºC. Conservación a temperatura ambiente. Repeticiones P G P G R VG % % 1 0 0 0 2 0 0 0 SOLNI 3 0 0 0 4 0 0 0 1 0 0 0 2 2 2 0,052 SOLLO 3 18 18 0,473 4 4 4 0,142 TG 0 0 0 0 0 19 19 14 Se analiza la germinación de semillas conservadas a temperatura ambiente, sembradas en germinador a 30 º C, Tablas Nº 3-20 y 3-21. Sólo germinó Solanum lorentzii. Tabla Nº 3-20. Velocidad de germinación de Solanum lorentzii (30ºC) (Calculada utilizando el modelo de Maguire) Repeticiones 1 2 3 4 Días dr de 13 13 0 13 25 15 0 25 Plántulas Germinación IVG N° % 3 3 0 8 6 4 0 16 0,156 0,209 0 0,43 dr: días empleados para empezar a germinar de :días empleados para la germinación total de las semillas Tabla Nº 3-21. Análisis de los parámetros de germinación de semillas conservadas a temperatura ambiente, germinador a 30ºC Repeticiones P G P G R % % VG TG 274 1 2 3 4 1 2 3 4 SOLNI SOLLO 0 0 0 0 6 4 0 16 0 0 0 0 6 4 0 16 0 0 0 0 0 0 0 0 0,156 21 0,209 28 0 0 0,43 20,5 Tercer grupo: E. siembra de semillas conservadas en frío a 25 ºC Se analiza la germinación de semillas conservadas en frío, sembradas en germinador a 25 ºC. Tablas Nº 3-22 y 3-23. Sólo germinó Solanum lorentzii, SOLNI no se analiza. Tabla Nº 3-22. Velocidad de germinación de Solanum lorentzii (25ºC) (Calculada utilizando el modelo de Maguire) Repeticiones 1 2 3 4 Días dr de 10 10 10 10 11 10 10 10 Plántulas Germinación IVG N° % 3 1 2 1 6 2 4 2 0,281 0,100 0,200 0,100 dr: días empleados para empezar a germinar de :días empleados para la germinación total de las semillas Tabla Nº 3-23. Análisis de los parámetros de germinación de semillas conservadas en frío (germinador a 25ºC) Repeticiones P G P G R % % 1 0 0 2 2 2 SOLNI 3 0 0 4 0 0 1 6 0 2 2 2 SOLLO 3 4 0 4 2 2 VG TG 0 0 11 11 0 0 0 0 0,281 7,33 0,100 10 0,200 10 0,100 10 Se analiza la germinación de semillas conservadas en frío, sembradas en germinador a 30 º C, Tablas Nº 3-24 y 3-25. No se analizan los parámetros de SOLNI. Tabla Nº 3-24. Velocidad de germinación de Solanum lorentzii (30ºC) (Calculada utilizando el modelo de Maguire) Días 275 Repeticiones dr de Plántulas Germinación IVG N° % 13 25 8 16 0,535 1 2 13 13 1 2 0,076 3 8 13 10 20 1,201 4 8 8 10 20 1,250 dr: días empleados para empezar a germinar de :días empleados para la germinación total de las semillas Tabla Nº 3-25. Análisis de los parámetros de germinación de semillas conservadas en frío (germinador a 30ºC) Repeticiones P G P G R % % 1 0 0 2 0 0 SOLNI 3 0 0 4 0 0 1 16 16 2 2 2 SOLLO 3 20 20 4 20 20 VG TG 0 0 0 0 0 0 0 0 0,535 16,12 0,075 13 1,201 20 1,250 20 En Tabla Nº 3-26 se comparan los porcentajes de germinación de semillas sin conservación, conservadas a temperatura ambiente y en frío, sembradas en germinador con diferentes temperaturas. Tabla Nº 3-26. Comparación del Porcentaje de la germinación de semillas del Complejo Solanum nigrum en germinador Temperatura Conservación Siembra Inmediata Temperatura ambiente Frío (4ºC) 30 ºC 25 ºC SOLNI 0 0 0,5 SOLLO 0 6 3,5 SOLNI 9,5 0 0 SOLLO 4,5 6,5 14,5 En condiciones de temperatura controlada, con excepción de siembra inmediata, el mayor porcentaje de germinación (PG) fue el de Solanum lorentzii. El rango de 25-30 º C es el de mejor germinación. Por debajo y por encima de estas temperaturas no se produce la germinación. La temperatura óptima para ambas especies es 30 º C. Estos valores obtenidos por Mansilla y Chaila (2004) se ajustan con los determinados por algunos autores. Ogg et al., (1989); Holm et al., (1977); Roberts y Lockett, (1978); del Monte y Tarquis, (1997) coinciden en que la germinación se produce entre 20 y 40 º C, con una temperatura óptima de 30 º C. Los valores de germinación relativa (PGR) son los mismos que los porcentajes de germinación (PG) pues en todos los casos los registros se hicieron durante un mes. 276 En el análisis de la velocidad de germinación (VG), se utilizó el índice de Maguire (1962) pues la temperatura era constante. Lo obtenido muestra que a 30ºC germinan más rápido, especialmente en la siembra inmediata. La tasa de germinación es muy baja en todos los casos para las dos especies estudiadas y la velocidad de germinación obtenida es importante y de consideración (10 plantas en 8 días ) en el caso de SOLLO y muy lenta para SOLNI (ninguna en 30 días). Registros fotográficos: En las figuras 3-9 a 3-14 se muestran imágenes de la germinación de las especies del Complejo Solanum nigrum. Los cotiledones de ambas tienen forma oval – lanceolada, con la base atenuada. Levemente pilosos. El largo de cada hojita es aproximadamente 5 mm. En las plántulas se observó que SOLNI tiene las primeras hojitas algo redondeadas, mientras que SOLLO las tiene mas bien lineal lanceoladas. No ha sido posible con estos caracteres diferenciar con seguridad a las especies del Complejo Solanum nigrum al estado de plántulas y también resulta difícil identificarlas cuando se encuentran en estado vegetativo. Recién cuando se inicia la floración es posible hacerlo 277 Germinación de Solanum nigrum Fig.Nº 3-9. Plántulas y frutos de SOLNI Fig.Nº 3-10. Esquema y plántula de SOLNI 278 Fig. Nº 3-11. Germinación y emergencia de SOLNI 279 Germinación de Solanum lorentzii Fig.Nº 3-12. Germinación de SOLLO Fig.Nº 3-13 . Esquema y cotiledones de SOLLO Fig.Nº 3-14.Germinación de SOLLO 280 ENSAYOS EN OSCURIDAD Otras pruebas de germinación No hubo germinación en estas condiciones. No se analiza. En los trabajos consultados no se encontraron referencias sobre experimentos de germinación en oscuridad. GERMINACIÓN DE UNA ESPECIE INTERMEDIA. En macetas con tierra: El análisis de la germinación de semillas de Solanum nigrum Intermedio (SOLNI-I), que presentan dos tipos de inflorescencias, se muestran en las Tablas 3-27, 3-28 y 3-29. Se exponen los registros de los 15 primeros días. Tabla Nº 3-27. Análisis de los parámetros de germinación de semillas de SOLNI-I conservadas a Tº ambiente Repeticiones P G P G R % % 1 20 20 2 20 20 SOLNI-I 3 0 0 4 20 20 5 10 10 VG TG ------- 8 ---- 8 ---- 0 ---- 8 ---- 8 Tabla Nº 3-28. Análisis de los parámetros de germinación de semillas sin conservación de SOLNI-I Repeticiones PG PGR VG TG % % 1 0 0 ----0 2 40 40 ----- 11,75 SOLNI-I 3 20 20 ----- 12 4 0 0 ----0 5 0 0 ----0 Tabla Nº 3-29. Análisis de los parámetros de germinación de semillas de una inflorescencia cimosa de SOLNI-I, siembra inmediata Repeticiones P G P G R % % 1 50 50 2 60 60 SOLNI-I 3 0 0 (cimosa) 4 0 0 5 20 20 VG TG ---- 11 ---- 10,83 ---0 ---0 ---- 15 El porcentaje de germinación más elevado en estas plantas con inflorescencias particulares fue el de las semillas de la inflorescencia cimosa. 281 Se debe destacar que ninguna de las dos formas de inflorescencias, normal (cimosa) y la intermedia (bifurcada), necesitaron un período de reposo para germinar como ocurrió en las otras experiencias. En estas experiencias la temperatura del ambiente se registró a diario por lo tanto no se analiza la velocidad de germinación. En agua: Se produjeron germinaciones escalonadas a lo largo de los seis meses que estuvieron en agua . Tabla Nº 3-30. Porcentajes de la germinación de semillas de forma Intermedia del Complejo Solanum nigrum, en medio líquido. SOLNI – I (Capital) 19/11/2003 Siembra Junio /20 04 Octubre / 2004 Diciembre / 2004 Total PG=(Nx100)/30 PGR=(Nx100)/30 TG Nº 7 3 5 15 50 % 0 - No se encontró referencia alguna sobre la presencia de esta particularidad en el tipo de inflorescencia que se presenta en SOLNI. Se destaca la capacidad de germinar sin un periodo de reposo, al poco tiempo de cosechadas, en algunas siembras en solo siete días. No se produjo germinación en agua destilada. SIEMBRAS EN INVERNÁCULO / INTEMPERIE Varios autores habían demostrado que en condiciones de laboratorio e invernáculo los resultados de germinación y emergencia del Complejo Solanum nigrum, en general, eran semejantes al patrón de la emergencia en el campo de otras malezas de ciclo anual y que según las regiones, predominan diferentes especies en este complejo (Ogg et al., 1989; Roberts y Lockett, 1978; Edmonds y Chweya, 1997). En Tucumán, se debe destacar que hay marcadas diferencias entre lo observado a campo y en laboratorio con respecto a la germinación de las semillas y a la emergencia de las plántulas del Complejo Solanum nigrum. A- Germinación a.- En el campo se observó una situación de germinación continua, en forma escalonada y con un máximo en el número de plantas en primavera lo que coincide con los autores mencionados. El mayor número de plantas observadas en el campo correspondió a Solanum nigrum. 282 b.- En laboratorio la germinación no fue continua ni escalonada, y se observaron porcentajes muy bajos de ésta. En Solanum nigrum, en todas las condiciones de conservación de las semillas y con las diferentes temperaturas a que fueron sometidas las muestras, los porcentajes fueron 2 a 9,5 % mientras que en Solanum lorentzii los valores observados fueron un poco mayor, entre el 2 y 14 %. Las plántulas en el laboratorio son débiles, viven un corto período donde se pudren o se secan. Esto no ocurre en el campo. B- Emergencia: Se observó la emergencia en el campo de las especies de Solanum a partir de la segunda quincena del mes de julio. El número de las que nace en el campo es mucho mayor que las que germinan en medios artificiales, pero pocas son las plantas que completan su ciclo. Las fechas de los distintos estados fenológicos se han comparado con los trabajos de Ogg et al. (1989) y de Roberts y Lockett (1978) y se observa que en el hemisferio norte hay un patrón semejante al de nuestra región que se repite todos los años. En nuestro medio, Solanum nigrum germina y emerge casi un mes antes que Solanum lorentzii. También florece antes y ambas llegan a plenitud de fructificación en la misma época del año, en verano. Los registros del seguimiento realizado a lo largo del ciclo de vida de las especies de Solanum se observan en la Tabla Nº 3-31 donde se comparan los estados fenológicos de las dos especies. Tabla Nº 3-31. Estados fenológicos del Complejo Solanum nigrum Especie SOLNI SOLLO Jul. germina Ago. Set. Oct. Nov. Dic. Ene. Feb. Mar. florece fructifica muere germina florece fructifica Declina la producción de frutos, sigue vegetando De los experimentos realizados se observó que: No germinaron las semillas de las macetas llenadas con tierra (en invernáculo e intemperie). No germinaron en cubeteras mantenidas en el invernáculo, hubo una rápida invasión de hongos. No germinaron en ningún recipiente humedecido con agua destilada Sólo germinaron las sembradas en bandejas mantenidas a la intemperie, riego con agua de red. En las dos especies la germinación fue pobre, con plántulas muy débiles que murieron en pocos días. 283 Se observaron diariamente durante aproximadamente 6 meses y de estas experiencias sólo se muestra el resultado de la siembra en bandejas en Tabla Nº 3-32; en las otras no hubo germinación. Tabla Nº 3-32. Siembra en bandejas del Complejo Solanum nigrum mantenidas a la intemperie. SOLNI SOLLO Repeticiones 1 2 1 2 Total 1 0 0 2 PG= (Nx100)/480 0,2 % 0% 0% 0,4% PGR= (Nx100)/480 0,2 % 0% 0% 0,4% Comparación del comportamiento y del desarrollo de las especies del Complejo Solanum nigrum en invernáculo y en el campo: Al iniciarse la floración las especies marcadas se identificaron como Solanum nigrum y Solanum lorentzii. Las plantas que se trasplantaron y se llevaron a un invernáculo en Agosto-Septiembre sufrieron un rápido debilitamiento, perdieron hojas, produjeron flores, pocos frutos y luego murieron (aproximadamente 2 o 3 meses después del trasplante). Por planta produjeron 1 a 3 inflorescencias de 4 en 5 frutos cada una. Las que continuaron en el predio siguieron creciendo hasta completar su ciclo de vida con abundante floración y fructificación. Solanum nigrum lo completó en enero-febrero y Solanum lorentzii en varios meses más (más de un año en algunos casos). Este comportamiento confirma los ciclos de vida que citan: anual para Solanum nigrum (Holm et al,. 1977) y perenne en Solanum lorentzii según Morton (1976), Cabrera (1983) y Roncaglia et al., (2000). En el campo se observó que Solanum nigrum completó su ciclo al finalizar el verano, las plantas no tenían hojas, sólo frutos maduros; mientras que Solanum lorentzii luego de perder las hojas, algunas plantas murieron y otras se recuperaron y continuaron vegetando. En general las plantas de Solanum lorentzii produjeron menos frutos en la temporada siguiente y de un tamaño menor. Un 90 % menos. Cuando fue trasplantada Solanum lorentzii se mantuvo en mejores condiciones que Solanum nigrum. En ambas situaciones, invernáculo y campo, ocurrieron ataques de insectos y de hongos. Tanto en Solanum nigrum como en Solanum lorentzii las especies de insectos fueron las mismas. En los brotes se observaron pulgones de color negro identificados como Myzus persicae. 284 En plantas más desarrolladas se observó la aparición de un ácaro identificado como Tetranichus telarium. El ataque de éste fue el responsable de la muerte de la mayoría de las plantas en el invernáculo y de la desfoliación de las que continuaban en el campo. Si bien se hacen referencias sobre estas plantas como hospederas de insectos, (Edmonds y Chweya, 1997) no se han encontrado referencias bibliográficas sobre cuáles son las especies insectos que suelen encontrarse en estas malezas. El material que se trajo de zonas del este y norte de Tucumán no sirvió para trasplante por su pronta marchitez a pesar de ser extraídas con pilón. Se debe destacar, que en las temporadas siguientes no se encontraron plantas del Complejo en los sitios donde fueron registradas y/o recolectadas. EMERGENCIA EN FUNCIÓN DE LA PROFUNDIDAD Los valores obtenidos de la germinación en distintas profundidades confirman los antecedentes que muestran en sus experiencias Ogg et al. (1989), Holm et al. (1977) y Roberts y Lockett, (1978). Sólo germinaron con bajos porcentajes a 0 y 2,5 cm, a una profundidad mayor no se produjo la germinación. Se presentan los datos de las semillas sembradas casi en la superficie y a 2,5 cm. (Tabla Nº 333 y Tabla Nº 3-34). No se analiza la velocidad de germinación. Tabla Nº 3-33. Germinación del Complejo Solanum nigrum sembradas en superficie. (0 cm) Repeticiones SOLNI SOLLO 1 2 3 4 1 2 3 4 Total P G P G R % % 4 8 2 6 12 2 5 10 0 2 4 0 11 22 6 11 22 4 13 26 2 12 24 14 VG TG --------------------------------- 33,50 47,16 35 42 36,54 34,54 39,92 23 Tabla Nº 3-34. Germinación del Complejo Solanum nigrum sembradas a 2,5 cm. Repeticiones SOLNI 1 2 3 4 Total P G P G R % % 2 4 0 5 10 0 2 4 0 2 4 0 VG TG ----- 51 ----- 39,2 ----- 35 ----- 42 285 SOLLO 1 2 3 4 9 1 4 2 18 2 8 4 4 0 0 0 ----- 40,11 ----- 42,0 ----- 40,25 ----- 42 PARÁMETROS DE GERMINACION Los resultados de los parámetros que se utilizaron en la caracterización de la germinación de los ensayos, se fueron indicando a medida que se analizaba cada grupo de experiencias. (Porcentaje, porcentaje de germinación relativa, velocidad y tasa de germinación). (Mansilla de Andrada y Chaila, 2007). Las plántulas emergidas en un área determinada sirven para estudiar los flujos de emergencia en el tiempo y efectuar todos los estudios concernientes a la demografía de malezas. Dentro de los estudios demográficos es importante tener los datos de las especies sobrevivientes en un sistema agrícola. Estas últimas son las especies que se establecieron en el proceso de competencia y llegan a ser plantas adultas que son las encargadas de las estrategias reproductivas. En demografía experimental de Solanum, se trabajó en cañaverales infestados de la Localidad de Los Ralos (Cruz Alta – Tucumán) durante 2005 con el cv TUC 77-42. (Mansilla de Andrada y Chaila, 2008). El diseño experimental fue en bloques totalmente al azar con tres repeticiones, en parcelas de 480 m2, determinadas en áreas de infestación media a alta de la maleza (de 5 a 12 plantas por m2), dispuestos en un sector de cañaveral de 5000 m2. Para este estudio demográfico se siguió la metodología propuesta por Chaila (1998). Se contaron las plantas totales al 15 de noviembre y 15 de diciembre para establecer por diferencia plantas vivas y muertas. Las sobrevivientes fueron las registradas al 15 de enero. Se realizaron análisis estadísticos no paramétricos con las pruebas correspondientes. En la demografía experimental, la media teórica del género establece la posible variación en número de plantas vivas y muertas; siendo de 8.457,94 pl.ha -1 para las sobrevivientes con una densidad de 0,84 pl.m-2 . En Los Ralos, en cambio, los valores medios se encuentran por debajo de las medias teóricas para sobrevivientes en 6.284,65 pl.ha-1 con una densidad de 0,62 pl.m-2 . Se empleó un test no paramétrico para determinar las diferencias con respecto a la misma tasa. La significancia del estadístico de Kruskal-Wallis es mayor que el 5 %, esto indica que los tratamientos tienen igual localización dentro de una serie originada a partir de la variable ordenada, es decir, no existen diferencias significativas entre tratamientos (tasas de igual denominación). Por eso el género es determinante de las tasas demográficas y el hábitat influye sobre las mismas, pero ante igual situación de manejo del agroecosistema no habrá variaciones o diferencias entre tasas. Se encontró una TBM (tasa bruta de mortalidad) de 750,00‰; una TBN (tasa bruta de natalidad) de 250,00‰ y una Sv (Supervivencia) de 63,10‰. (Fig Nº 3-15). 286 El potencial de infestación (PI) de la especie es bajo, llegando a 218,62 m2.pl-1, al igual que la media teórica del género y expresa que una planta puede infestar 161,36 m2.pl-1 con su próxima descendencia (Mansilla de Andrada y Chaila, 2007). En la determinación del potencial de infestación la comparación de las dos especies arroja los siguientes resultados: SOLNI: Mayor cantidad de semillas ingresantes al banco. Individuos con capacidad reproductiva alta y capacidad reproductiva real baja e igual que SOLLO. El Potencial de infestación aporta valores intermedios y superiores a SOLLO. SOLLO: Menor cantidad de semillas ingresantes al banco que SOLNI. Individuos con capacidad reproductiva baja. Capacidad reproductiva real baja e igual que SOLNI. El Potencial de infestación arroja valores bajos e inferiores a SOLNI. SOLNI posee los valores más altos obtenidos en este análisis, tanto para cantidad de semillas y viabilidad como para semillas ingresantes, para individuos con capacidad reproductiva, para real capacidad reproductiva y para potencial de infestación. Esta especie infestará mayor superficie a partir de la producción de semillas de sus frutos. SOLLO posee los valores más bajos del análisis excepto poder germinativo y densidad. Su capacidad de infestación es menor pero por ello no deja de ser importante como competidora de cultivos. Sv TBN TBM Fig. Nº 3-15. TASAS DEMOGRÁFICAS DE SOLNI Observaciones finales: Se agregan estas observaciones por considerarlas interesantes. Ninguna de ellas se menciona en la bibliografía consultada. - En la plantación de estacas no se obtuvo resultado alguno lo que confirma lo expresado por Holm et al. (1977), Ogg et al. (1981) y Edmonds y Chweya, (1997), que SOLNI sólo se reproduce por semillas. - En los recipientes retirados del germinador y conservados un año en el laboratorio tapados y a temperatura ambiente, se produjeron germinaciones en medio de los hongos que invadieron el papel de filtro. 287 - En un grupo de semillas retiradas del germinador y colocadas en una maceta con tierra, la germinación fue de un 90 %. Las plantas que emergieron y siguieron desarrollándose no llegaron a florecer. Fueron atacadas por hongos y luego murieron. - El trasplante de las que nacieron en el germinador no dio resultado, al poco tiempo las plantitas murieron. 288 CAPÍTULO 4 OTROS ASPECTOS DE IMPORTANCIA SOBRE EL Complejo Solanum nigrum 4.1. INTRODUCCIÓN Este grupo de plantas ha sido objeto de numerosos estudios en el mundo como malezas y en algunas regiones, como recursos alimenticios y medicinales, existiendo una gran confusión sobre la identidad del mismo (Holm et al. 1977; Edmonds y Chweya, 1997). Por ejemplo en algunas regiones de África en algunos trabajos se comenta sobre el consumo de hojas de Solanum nigrum cuando en realidad se trata de Solanum villosum o S. scabrum (Guarino, 1977). Se debe destacar que en la mayor parte de los trabajos en el mundo se han hecho en Solanum nigrum y ninguno en Solanum lorentzii. Por ello, fue necesario en primer lugar precisar la identificación de las especies del Complejo, con el propósito de reconocerlas y estudiar su biología, para saber a ciencia cierta si realmente son un problema o si podrían considerarse como un recurso para el desarrollo de las zonas áridas y semiáridas de nuestra región. En este capítulo los diferentes matices encontrados se han tratado en el siguiente orden: medicinales, tóxicas, alimentación, reservorio de plagas y/o portadores de resistencias y por último su distribución. El objetivo de este capítulo es brindar datos teóricos obtenidos en la extensa búsqueda bibliográfica y otros de experimentación propia sobre otros aspectos del Complejo Solanum nigrum. 4.2. ANTECEDENTES 4.2.1. Medicinal Si bien es cierto que el hombre, desde el momento mismo de su aparición sobre la tierra, ha hecho uso de las plantas para proporcionarse alivio a los malestares que lo acosan y ese uso ha llegado hasta nuestros días, no es menos cierto que todavía se desconocen los tóxicos que se ocultan en ellas. En El Dioscórides renovado, entre las plantas medicinales de España, se describe a Solanum nigrum como “una hierba medicinal conocida desde antiguo y que ha sido utilizada en muchas regiones” (Font Quer, 1962). Se dice que el jugo de sus bayas evaporado combate el dolor de muelas, la cocción de las hojas se aplica a los pruritos vulvares y hemorroidales, las hojas en emplasto para sanar llagas, para reducir el ardor de estómago, para diferentes dolores (de cabeza, auditivo, el provocado por la artritis y contusiones), contra la crisis hepática, para regularizar el flujo menstrual... y así, unas cuantas aplicaciones más. En general, se usaba por vía externa, ya que su ingestión podía resultar tóxica y se sabía que tenía propiedades como sedante, narcótica, emoliente y analgésica. 289 Esta hierba humilde, que a menudo crece entre estiércol, rastrojos y orillas de caminos, si se la conoce y se utiliza correctamente, resulta ser toda una fuente de curas y panacea de muchos males. Etimológicamente Solanum significa “que mitiga el dolor”. (Enciclopedia Universal Ilustrada. Europeo – Americana. 1927). En la actualidad, su uso como planta medicinal está registrado en todo el planeta. * En Colombia y Méjico los frutos de la “yerba mora” se consideran narcóticos y venenosos. Las hojas y los frutos son estupefacientes y desinfectantes por lo que en medicina popular se emplean en decocción o triturados para curar algunas afecciones de la piel, (infecciones, eczemas, sarpullido, granos de etiología incierta, pústulas, etc.). En general su uso es externo en el tratamiento de los desórdenes cutáneos, pero como su acción es cambiante, es considerado un remedio algo peligroso salvo en dosis muy pequeñas. También los campesinos usan con frecuencia los emplastos de hojas y frutos frescos para curar la gangrena y el ántrax. (Uninca, 2003; Semarnap, 2003). * En Japón, usan la planta como emoliente, laxativa y antiespasmódica. Se dice que la decocción de las hojas cura el cáncer de estómago y la úlcera duodenal, es también desinfectante, cura el prurito y las hemorroides. El fruto es reconocido como venenoso (Uninca, 2003). * Esta planta es ampliamente utilizada en países como China para tratar la impotencia sexual masculina (Wikipedia, 2003). * También la infusión de hojas es un agente sudorífico fuerte y en Bohemia, (República Checa), las hojas se ponen en las cunas de los niños para promover el sueño (Grieve, 2004). * Los Árabes aplican las hojas frescas y machacadas a las quemaduras y úlceras para aliviar el dolor y bajar la inflamación. Su jugo es usado para la tiña (ringworm), gota y dolor de oídos y mezclado con vinagre para hacer gárgaras y enjuague bucal. (Grieve, 2004; (Wikipedia, 2004). * En zonas húmedas de la India se estudiaron las propiedades tóxicas de la “yerba mora” para eliminar las larvas de los mosquitos (Wikipedia, 2003; Botanical, 2006). * Lorenzi y Abreu (2002) incluyen a esta planta dañina de la agricultura de todo Brasil entre las plantas medicinales y con los mismos usos. * Alonso (2004) informa que entre las plantas medicinales empleadas por los nativos del noroeste de la República Argentina, “la infusión de hojas de yerba mora era utilizada por los chamanes para el reumatismo”. * En el Vademécum de malezas medicinales de la Argentina, Marzocca (1997) sugiere todos estos usos pero aclara el carácter de planta tóxica. * Edmonds y Chweya (1997) expresan que muchas de las especies de la Sección Solanum son ampliamente utilizadas como medicinales en todo el mundo. El uso como medicinal se registra desde tiempos remotos y varias especies, especialmente Solanum nigrum son mencionadas y a menudo ilustradas en todos los antiguos herbarios. Relatan a cerca de estos 290 usos principalmente en países de Asia, África y Europa y que en el nuevo mundo hay unas pocas citas en Norte América, Hawai y en Ecuador en Sud América. En la zona en estudio no se ha encontrado información sobre usos como planta medicinal de las especies del Complejo Solanum nigrum. 4.2.2. Tóxica Importancia para el hombre La mayoría de las especies asociadas con Solanum de la sección Solanum tienen fama de venenosas, tanto para el hombre como para los animales. Mucha información sobre los efectos tóxicos ya se encontraba en la literatura antigua y en la actualidad, en casi en todos los manuales de plantas venenosas en todas las regiones del mundo se incluye a Solanum nigrum. Son tóxicas en forma variable tanto para el hombre, como para el ganado vacuno, cerdos, cabras, patos y gallinas (Edmonds y Chweya, 1997). La toxicidad ha sido atribuida a los alcaloides que poseen. Se han aislado dos alcaloides de este grupo de plantas: solanina, un glicoalcaloide tóxico y solanidina un alcaloide esteroide, también tóxico. Se ha comprobado que la toxicidad de las especies varía con el ambiente, el tipo de suelo, con la porción de planta ingerida o el grado de madurez de la misma (Edmonds y Chweya, 1997; Otero Aira, 2001; World Wise Garden Seeds, 2004). En las especies de Solanum, en general, las bayas verdes son más tóxicas que las bayas maduras; estas más tóxicas que las hojas y las hojas más tóxicas que los tallos o las raíces. El principio venenoso está generalmente asociado con todas las partes verdes de las plantas. Las bayas de Solanum nigrum aun las maduras, pueden ser peligrosas para los niños, pero no para los adultos (World Wise Garden Seeds, 2004). La solanina, causa dolor de cabeza, insomnio, agitación, sed, vértigos, cara congestionada, náuseas, cólicos, diarreas y vómito, sudores profusos, mareos, arritmia, trismus, temblor de todo el cuerpo, somnolencia, dificultad para hablar y moverse, respiración dificultosa y en mayores dosis, delirio, convulsiones, contracturas tetánicas, enfriamiento, coma y finalmente la muerte por fallo cardíaco (Edmonds y Chweya, 1997; Uninca, 2003). Es importante destacar que con la cocción se pierde la toxicidad porque ocurre la destrucción de la solanina. Esto explica por qué esta especie considerada tóxica se puede usar como medicinal o como comestible, como se verá más adelante. Importancia agronómica Las plantas tóxicas se encuentran distribuidas por todo el mundo, pero su concentración es mayor en aquellas regiones áridas y semiáridas sometidas a sobrepastoreo. En estas regiones son frecuentes fenómenos meteorológicos como escasa precipitación y bajas temperaturas (Arbiza Aguirre, 1986). Gallo (1987) define como “Planta tóxica” a la que ingerida por un animal le produce trastornos de los distintos aparatos, que pueden llevarle inclusive a la muerte. Las plantas tóxicas para el ganado han constituido desde el inicio de los estudios veterinarios en Argentina, una preocupación constante y una amenaza a nuestra ganadería, causando grandes pérdidas anuales. 291 En Argentina no hay estadísticas, sobre las pérdidas que experimenta la ganadería, a consecuencia de las intoxicaciones alimentarias del ganado, atribuibles al consumo de plantas tóxicas. La información con que se cuenta en nuestro país sobre esta cuestión es escasa y algo similar ocurre con los países del cono Sur de América. Gallo ha realizado diferentes clasificaciones de plantas tóxicas: 1. según la parte vegetal, que ingerida, resulta tóxica a. semillas y frutos b. hojas, tallos verdes y corteza c. durante un período del desarrollo bajo condiciones ambientales especiales (diferentes partes de la planta como cotiledones, floración o fructificación, etc.) 2. según los principios activos que contienen a. Heterósidos b. Alcaloides c. Resinas d. Sustancias de acción fotodinámica e. Nitratos f. Principios anticoagulantes g. Aceites h. Ácido tánico i. Látex j. Euphorbona k. Toxoalbúmina l. Ácido oxálico m. Etc. ............. 292 En el grupo de los Heterósidos (glucósidos), que son sustancias que una vez hidrolizadas forman azúcares y agliconas se encuentran las saponinas, la solanina y los alcaloides. En muchas Solanáceas están presentes Alcaloides y Saponinas, como en Solanum eleagnifolium, “meloncillo del campo, revienta caballo” y en Solanum nigrum, “yerba mora”. Según la clasificación propuesta por Gallo (1987), la Solanina sería una saponina y también un alcaloide. Fig. Nº 4-1. Estructura química de la Solanina La importancia agronómica de las plantas que contienen alcaloides reside en su potencial poder tóxico para el ganado que las ingiere y no existe una información muy extensa en nuestro país sobre este tema (Troiani, 2006). El problema de intoxicación en animales es la contaminación de forraje por mezcla de este con las Solanáceas que principalmente sucede en aquellos que provienen de otras zonas, con hambre y acostumbrados a otras especies de pasturas (Arbiza Aguirre, 1986). Pero, si Solanum nigrum se encuentra mezclado en el forraje ensilado, la toxicidad de la solanina disminuye aparentemente por una hidrólisis ácida producida en la fermentación (Edmonds y Chweya, 1997). En el ganado vacuno, caballos, ovejas y cabras, los alcaloides y saponinas que posee la “yerba mora”en las partes verdes de la planta, pueden producir pulso y respiración rápida, parálisis, pupilas dilatadas, vómitos, diarreas y trastornos intestinales, membranas mucosas pálidas, hocicos secos, ansiedad seguida de depresión, somnolencia, baja temperatura, debilidad, falta de coordinación, convulsiones, colapso y posiblemente la muerte. En general se puede afirmar que las especies de Solanum producen disminución de la producción de leche y, además, alteran el gusto y el olor de la misma. (Edmonds y Chweya, 1997; Caribbean area, 2003; Botanical-online, 2006). Este grupo de Solanáceas, además, contiene altos niveles de nitratos. La intoxicación aguda por nitratos puede llevar a la muerte. La intoxicación crónica produce disminución del rendimiento de leche, aborto, deterioro de la vitamina A y del yodo, músculos temblorosos y andar tambaleante, pulso rápido, orina frecuente, respiración esforzada, seguida de colapso y coma, con o sin convulsiones. Sin embargo, el nivel al cual la toxicidad de los nitratos puede causar deterioro en la producción y en la salud de los animales es incierta. Se piensa que los efectos observados luego de la ingestión de la yerba mora, puede deberse a la solanina o a los nitratos, o a ambos. Como ya se dijo, la cantidad de las sustancias tóxicas disminuyen con la madurez de los órganos de las plantas (Edmonds y Chweya, 1997, Mansilla y Díaz, 2006). 155 Para Portolano (1990) las plantas más conocidas capaces de causar intoxicación a los animales, o de transmitir características particulares a sus producciones, no son plantas apetecidas por el ganado. Este fenómeno está vinculado a las leyes sobre la conservación de las especies. Los animales pueden circunstancialmente ingerir plantas tóxicas condicionados por situaciones extremas de falta de recursos alimenticios y la gravedad de las intoxicaciones está relacionada con la mayor o menor rapidez con que las plantas ingeridas cedan al organismo sus principios tóxicos. (Arbiza Aguirre, 1986). Las características principales de las plantas tóxicas para Arbiza Aguirre (1986) son: Gran potencialidad genética, que le permite reproducir formas y ecotipos fácilmente adaptables a diferentes ecosistemas. Multiplicación rápida y producción de numerosas generaciones de semillas. Resistencia a condiciones adversas. Diseminación a grandes distancias por medio de diferentes agentes de dispersión. Latencia diferencial y a veces prolongada de las semillas. La mayoría de las plantas tóxicas permanecen verdes en los meses de sequía. Gran capacidad competitiva. Salvo la última de las características, las especies del Complejo Solanum nigrum responden a todas las otras. Si se consideran los antecedentes de las Solanáceas como plantas medicinales y como plantas tóxicas se puede decir que no existen plantas tóxicas sino un uso indebido de las plantas. Citando a Paracelso se puede decir que: "Un tóxico puede ser un remedio y un remedio puede ser un tóxico. Todo depende de la dosis". 4.2.3. Forrajera A pesar de lo que se ha considerado en el punto anterior, las especies del Complejo Solanum nigrum pueden usarse como forraje para el ganado y cabras y también pueden comerlas los animales salvajes (Edmonds y Chweya, 1997). En general la bibliografía consultada no hace referencia sobre este tema. Para determinar la aptitud como forrajeras se efectuó el Análisis Proximal de las especies de Solanáceas recolectadas en la zona de estudio. Este análisis se realizó con la colaboración de personal técnico de la cátedra de Forrajicultura y Cerealicultura de la Facultad de Agronomía y Zootecnia de la Universidad Nacional de Tucumán. Se recolectaron plantas enteras en plenitud de floración, se cortaron la totalidad de las hojas, descartándose tallos, ramas, flores y frutos. Las hojas se secaron en estufa a menos de 60 º C y se utilizaron 100 gr de hojas secas y molidas, de cada especie. Tabla Nº 4-1. Análisis del valor nutritivo del Complejo Solanum nigrum en plenitud de floración MUESTRA SOLNI % P.B. 24,50 % E.E. 2,47 % % % % % % % F.C. CZA E.NoN2 F.D.N. F.D.A. L.D.A. DIG 18,82 15,02 39,19 30,17 20,15 6,60 59,90 156 MUESTRA SOLLO % P.B. 18,29 % E.E. 1,21 % % % % % % % F.C. CZA E.NoN2 F.D.N. F.D.A. L.D.A. DIG 20,76 14,30 45,43 33,29 28,30 9,13 61,84 Tabla Nº 4-2. Referencias del Análisis Análisis Proximal P.B. E.E. F.C. CZA Proteína bruta Extracto Etereo. (Lípidos) Fibra Cruda o Fibra bruta Ceniza total (mineral) Análisis químico E.NoN2 F.D.N. F.D.A. L.D.A. DIG. Hidratos de Carbono Fibra Detergente Neutro Fibra Detergente Ácida Lignina Detergente Ácida Digestibilidad De los valores obtenidos se destaca que la proteína bruta (P.B.) presenta un excelente nivel. La fibra cruda (F.C.) que comprende celulosa, hemicelulosa y lignina, la porción menos digerible de la planta, tiene valores medios a bajos en este estado fenológico. Esto significa que tiene una buena aptitud forrajera. También son buenos los valores de la ceniza total (CZA) y de los hidratos de Carbono (E.NoN2). Por medio de las cenizas los animales toman los minerales, y los hidratos de carbono son los jugos celulares citoplásmicos fácilmente digeribles. La digestibilidad (DIG) es muy buena a excelente. Con estos resultados se puede afirmar que en este estado fenológico, plenitud de floración, las especies analizadas tienen una aptitud forrajera buena. Debería, además, determinarse su palatabilidad. Esto confirma lo citado por World Wise Garden Seeds, (2004) que a medida que maduran las plantas pierden la toxicidad. 4.2.4. Alimentación Solanum nigrum y las especies relacionadas a ésta, han sido usadas por el hombre como fuente de alimentación por muchos siglos pues en plantas adultas, tanto las hojas como sus frutos son comestibles (Edmonds y Chweya, 1997). Las partes vegetativas son muy nutritivas por el contenido de proteínas, fibra, minerales y vitaminas. Las bayas contienen calcio, hierro, caroteno y vitaminas B y C y las semillas también contienen vitamina C y caroteno (Edmonds y Chweya, 1997). Los brotes tiernos se comen como quelite (ensalada), sus hojas como verdura de hoja en reemplazo de la espinaca. Los brotes y las hojas también son utilizadas para preparar puchero (Semarnap, 2003). Con los frutos se preparan jaleas, mermeladas y confituras, que son utilizadas en el relleno de pasteles. Con las bayas también se preparan conservas o se comen crudas y pueden usarse como tintura y colorante de comidas (Holm et al., 1977; Edmonds y Chweya, 1997). En algunas regiones de África y de Asia se cultivan en un pequeño pedazo de tierra o simplemente en macetas para proporcionar una buena cosecha de hojas y de bayas. No hay datos estadísticos sobre estos cultivos (superficie, rendimientos, etc.) (Edmonds y Chweya, 1997). En nuestro país no hay información sobre estas especies como fuente de alimentación. 157 4.2.5. Reservorio de plagas y/o portadores de resistencias a. Reservorio de plagas Muchas especies de insectos y ácaros actúan como vectores o agentes transmisores de patógenos que producen enfermedades de las plantas. Los patógenos transmitidos de esta manera son en su mayor parte virus y micoplasmas, en menos proporción hongos y bacterias. Algunos insectos actúan como agentes de propagación de patógenos por el mero hecho de posarse en tejidos enfermos y después en plantas sanas, llevando a los agentes patógenos adheridos a sus apéndices. En el estudio del Complejo Solanum nigrum en el área agrícola de la provincia de Tucumán se observó que éstas son hospederos alternativos de insectos por lo que estas malezas son plagas potenciales para los cultivos. (Mansilla de Andrada y Salvatore, 2004). Los relevamientos se hicieron en localidades de las distintas regiones agroecológicas de Tucumán: La Cruz (Dpto. Burruyacú), Lolita (Dpto. Cruz Alta), Campo Experimental Finca el Manantial (Dpto. Lules), La Rinconada (Dpto. Yerba Buena). Las especies del Complejo Solanum nigrum se encontraban como plantas acompañantes en los cultivos de soja, caña de azúcar, trigo, cítricos y hortícolas. Para la toma de muestras, se consideraron las malezas que se encontraban dentro y fuera de los cultivos. Se realizaron observaciones sobre los insectos que estaban alimentándose en las mismas. Las muestras fueron tomadas al azar, una vez cada 15 días durante tres años (2000 – 2003). Entre los insectos recolectados se encontraron especies fitófagas y predadores. Además, se encontraron ácaros. Éstos totalizaron 6 especies correspondientes a tres órdenes: 50% Hemípteros, 20% Coleópteros y 30% Arácnidos. Se observó, además, la presencia persistente de hormigas y abejas. Del conjunto de insectos se destacan las chinches pentatómidos. Tabla Nº 4-2. Lista de las especies recolectadas Nombre Común Chinche de los cuernos Chinche Chinche verde Nombre Científico Dichelops furcatus (F) Edessa meditabunda (F) Nezara viridula L. Cochinilla algodonosa o Icerya purchasi Mask acanalada de Australia Pulgón verde del duraznero Siete de oro Vaquita de San José Myzus persicae Sulzer Astylus atromaculatus Blanch Cycloneda sanguínea L Arañuela roja Tetranychus telarius L Orden Hemíptero Hemíptero Hemíptero Homóptero Homóptero Coleóptero Coleóptero Clase Arácnida 158 En los Hemípteros se destacan las: Chinches pentatómidos: Son llamadas “plagas potenciales” por el daño que causan al inyectar toxinas en el momento de succionar la savia en hojas, tallos y fundamentalmente en vainas provocando abortos de flores, vaneos y manchado de semillas. Los principales cultivos que atacan son: tomate, papa, lechuga, pimiento, berenjena, acelga, soja, maíz, trigo, poroto, alfalfa, tabaco y frutales. (Giganti et al., 1997; Bayón, N. D., 2001; Futuros y Opciones, 2004, 2005; Olicer cereales, 2005; Todoagro, 2005; Massaro y Gamundi, 2006). Se debe destacar que Nezara viridula se encuentra con facilidad en las plantas espontáneas de donde pasan a los cultivos (Bayón, 2001). El grupo de chinches observado en el Complejo Solanum nigrum se compone de: Dichelops furcatus (F.) (chinche de los cuernos), Edessa meditabunda (F.) (alquiche chico) y Nezara viridula L. (chinche verde) (Mansilla de Andrada y Salvatore, 2004). (Fig. Nº 4-1 a 4-3). Entre los Homópteros se encontraron: Cochinillas Icerya purchasi Mask es la cochinilla que se encuentra en los cítricos de la región NOA en densidades muy bajas. Teniendo en cuenta que alguna vez fue plaga importante, hoy se la considera plaga en potencia (Nasca et al., 1981). (Fig. Nº 4-6). Áfidos Los pulgones provocan daños directos e indirectos a las plantas que se traducen en importantes disminuciones del rendimiento. El primer tipo de daño se genera por la incorporación de saliva tóxica y la extracción de grandes cantidades de savia, lo que provoca clorosis, manchas y muerte de hojas. El segundo tipo de daño se observa cuando los áfidos presentes en el cultivo son transmisores de virus (Infoagro, 2004; Saini, 2006). Myzus persicae Sulzer (pulgón verde del duraznero) ataca a numerosos frutales (duraznero, ciruelo, cítricos). Durante los meses del verano abandona sus hospederos primarios leñosos por hospederos secundarios herbáceos, como acelga, berenjena, batata, lechuga, papa, pimentón, remolacha, tabaco, tomate, trébol, zapallo, etc. (Ortego, 1990; BayerCropScience, 2004; Imwinkelried et al., 2004; Infoagro, 2004). Causa daños directos e indirectos. Entre los directos produce estrés hídrico, marchitamiento, enrollamientos de las hojas, reducción del crecimiento. En nectarinas es capaz de atacar al fruto produciendo áreas deprimidas y descoloridas. Los indirectos producen la transmisión de virus. Entre éstos, el virus Y de la papa (PVY) que se encuentra con frecuencia en regiones templadas y subtropicales. Malezas como Solanum nigrum L., Solanum dulcamara L., Portulaca oleracea L. y Senecio vulgaris L. no presentan síntomas pero actúan como reservorios naturales del virus Y de la papa por lo tanto, Myzus persicae es considerada la especie vectora más importante de PVY (Infoagro, 2004). 159 En nuestra región Myzus persicae está presente en todas las plantas del Complejo Solanum nigrum. Su presencia se manifiesta en un enrollamiento y gran deformación de las hojas y el aborto de flores. (Fig. Nº 4-5). En los Coleópteros se recolectaron insectos perjudiciales y benéficos. Astylus atromaculatus Blanch (siete de oro) en nuestra región tiene importancia en el cultivo de soja. También se considera plaga en maíz, girasol, y sorgo. (Fig. Nº 4-4). En primavera las larvas de Astylus destruyen semillas de los cultivos de verano y sus poblaciones se incrementan en lotes con mayor presencia de rastrojos. Los adultos entre enero y marzo consumen polen y sólo provocan daños de consideración en el sorgo (Frana, 1998; Saini, 2006). Cycloneda sanguinea Linneo (vaquita de San José) es un coleóptero benéfico. Es el coleóptero predador más abundante, enemigo natural de los pulgones. Son entomófagos predadores que ejercen fuerte presión para mantener las poblaciones perjudiciales por debajo de los umbrales de daño (Saini, 2006). (Fig. Nº 4-7). Ácaros Tetranychus telarius L. o Tetranychus urticae Koch afecta a más de 150 especies de interés económico, hortícolas, cultivos extensivos, frutales y ornamentales (melón, pimiento, berenjena, tomate, poroto, frutilla, cítricos). Los daños se producen en hojas, frutos, ramas, flores ocasionando un retraso del crecimiento, marchitamiento de hojas y manchas de frutos. (Agroinformación, 2002; Saini, 2006). (Fig. Nº 4-8). Las referencias bibliográficas sobre estas plagas demuestran la importancia de este aspecto debido a que todos los cultivos citados en éstas, también se hacen en Tucumán. Las especies del Complejo Solanum nigrum que sirven como los hospederos alternativos de plagas en varios cultivos de importancia comercial podrían ser usados como potenciales agentes de control (Edmonds Chweya, 1997). Por lo expuesto se hace necesario conocer la relación entre los insectos y estas malezas como hospederos alternativos, para poder realizar la planificación del control de plagas o bien mantenerlas como reservorios de insectos benéficos. En Tucumán se carece de suficiente información al respecto por lo que se debería continuar con el estudio de estas malezas asociadas a insectos o ácaros. 160 Especies recolectadas en el Complejo Solanum nigrum Orden Hemíptero - “chinches” Fig. Nº 4-2. Dichelops furcatus (F) “chinche de los cuernos” Fig.Nº 4-3. Nezara viridula L “chinche verde” www.viarural.com.ar http//bp1.blogger.com Fig Nº 4-4. Edessa meditabunda (F.) Extranet granos.fyo.com Orden Homóptero 161 Fig.Nº 4-5. Astylus atromaculatus Blanch “siete de oro” www.viarural.com.ar 162 Fig. Nº 4-6. Myzus persicae Sulzer “Pulgón verde del duraznero” 163 www.agroinformación.com/plaga-enfermedad Fig. Nº 4-7. Icerya purchasi Mask “cochinilla algodonosa o acanalada de Australia” Orden Coleóptero Fig. Nº 4-8. Cycloneda sanguinea L.“vaquita de San José” www.inta.com.ar Saini D. E. 164 Clase Arácnida Fig. Nº 4-9. Tetranychus telarius L “arañuela roja” www.agroinformación.com/plaga-enfermedad b. Portadores de resistencias Edmonds y Chweya (1997) señalan que las especies del Complejo Solanum nigrum por ellos consideradas son fuentes de varios rasgos de resistencia que podrían ser útiles para mejorar a varios cultivos económicamente importantes. Éstos incluyen: 165 - resistencia a las atrazinas - al virus del mosaico de tabaco - al tizón tardío de la papa - a la Phomopsis, o putrefacción de frutos Una gran cantidad de información se ha encontrado sobre estos temas, pero en nuestra región aun nada se ha investigado. 4.2.6. Otros usos Las tribus del sudoeste de Norte América usaban las bayas para alimentarse, y también para curtir cueros (Godhue, 1998). En China, antiguamente se usaba como cosmético (para quitar los granos) y como insecticida, para espantar moscas (Wikipedia. 2004). En países de África se empleaban como tinturas (Wikipedia. 2004; Guarino, 1997). En Méjico el follaje se usa como ceremonial y religioso, contra la envidia (Semarnap, 2003). En Argentina el único uso encontrado es como planta medicinal (Marzocca, 1997). 4.2.7. Encuestas Con el fin de obtener datos sobre el conocimiento y usos de las especies del Complejo Solanum nigrum en diferentes áreas agroecológicas de Tucumán, se realizó una encuesta. La encuesta posee cinco preguntas y dos dibujos (Fig.4-10). La distribución de las encuestas se hizo en cuatro escuelas agrotécnicas ubicadas en los departamentos Lules, Río Chico y Burruyacu de la Provincia de Tucumán. La edad de los alumnos encuestados variaba entre 14 y 16 años con un porcentaje aproximadamente igual de varones y de mujeres. El análisis de los datos obtenidos se procesó con el programa SPSS. Los resultados y análisis estadístico se muestran en Anexo Capítulo 4. 166 Encuesta sobre Solanum Nombre: .......................................... Localidad: ............................................ Fecha: .................................. 1.- ¿Conoce la planta de la figura? 2.- ¿Cómo la llama? 3.- ¿Para qué se la usa? 4.- ¿Es venenosa? 5.- ¿Es una maleza? Fig. Nº 4-10. Encuesta Con el análisis de las respuestas, se puede afirmar que no conocen esta planta, o la confunden con otras plantas útiles. Se debe destacar que son personas muy jóvenes las que contestaron la encuesta. 4.2.8. Distribución Con la búsqueda de información se ha logrado tener una idea de la distribución de estas especies en el mundo, en Argentina y en Tucumán. Los mapas de distribución se presentan en Anexo Capítulo 1 acompañando a la descripción de las especies y en Capítulo 4. En el mundo: Mapa de distribución mundial de Solanum nigrum L. de Holm et al., (1977). Fig. Nº 1-3. En Argentina: distribución de Solanum sublobatum Willd. “yerba mora”. Ragonese y Milano (1984). Fig. Nº 1-9. En Tucumán: Distribución de Solanum lorentzii Bitter en la provincia de Tucumán. Figueroa Romero et al., (2000). Fig. Nº 1-7. El mapa logrado a partir de las encuestas para las especies del complejo, en Anexo Capítulo 4, Fig. Nº 4-11. El mapa logrado en este trabajo. Fig. Nº4-19. 167 Fig.Nº 4-19. Distribución del Complejo Solanum nigrum en Tucumán 168 ENCUESTA SOBRE SOLANUM La encuesta realizada consta de cinco preguntas. Se distribuyó a grupos de alumnos de cuatro escuelas agrotécnicas de la Provincia de Tucumán. Además, a modo de orientarlos, se proporcionaba un esquema de la planta consultada. (Fig.Nº 4-10, Cap. 4.) La edad de los alumnos encuestados varía entre 14 y 16 años. Las escuelas se encuentran en diferentes zonas agroecológicas de la provincia. Fig. Nº 4-11, Tabla Nº 4-3 Fig. Nº 4-11. Distribución de las escuelas encuestadas en la Provincia de Tucumán. 169 Tabla Nº 4-3. Distribución por Departamentos de las escuelas encuestadas en Tucumán. Escuela 20 de Junio Departamento Localidad Lules Lules Los Sarmientos Los Ríos Los Sarmientos Río Chico La Tipa Aguilares Mal Paso El Chañar La Ramada Alderete Mariano Ramos Burruyacu Macomita Mariño La Cruz Taquello Soldado Cajal Burruyacu Las Cejas Banda del Río Salí Se realizó el análisis por sexo de los encuestados en las diferentes escuelas. 60 50 40 30 Percent 20 10 0 F M NS/NC Sexo Figura Nº 4-12: Distribución por sexo de los encuestados La figura Nº 4-12 muestra la distribución por sexo del total de alumnos que respondieron la encuesta. De un total de 151 encuestados, 72 son mujeres y 77 varones. Aproximadamente es un porcentaje igual de varones y de mujeres. 170 También se analizó el porcentaje de la distribución por sexo en las escuelas. 40 30 Nº de encuestados 20 Sexo 10 F M 0 NS/NC 20 DE JUNIO MARIANO RAMOS LOS SARMIENTOS SOLDADO CAJAL Escuela Figura Nº 4-13: Distribución del sexo de los encuestados en las diferentes escuelas Tabla Nº 4-4 . Distribución del sexo de los encuestados en las diferentes escuelas. Escuela * Sexo Crosstabulation Count Escuela Total 20 DE JUNIO LOS SARMIEN TOS MARIANO RAMOS SOLDADO CAJAL Sexo F 32 M 28 NS/NC Total 19 23 1 43 13 14 1 28 8 12 72 77 60 20 2 151 Tabla Nº 4-5 . Porcentaje de la distribución del sexo de los encuestados en las diferentes escuelas. Escuela * Sexo Crosstabulation % within Escuela Escuela 20 DE JUNIO LOS SARMIEN TOS MARIANO RAMOS SOLDADO Sexo F 53.3% M 46.7% NS/NC Total 44.2% 53.5% 2.3% 100.0% 46.4% 50.0% 3.6% 100.0% 40.0% 60.0% 100.0% 100.0% 171 CAJAL Total 47.7% 51.0% 1.3% 100.0% Pregunta Nº 1: ¿Conoce la planta de la figura? La primera pregunta es sobre el conocimiento que tiene el alumno que debe reconocer la planta en una figura. De acuerdo con las respuestas un 60% de los alumnos conocen la planta de la figura. (Figura Nº 4-14). 70 60 50 40 30 P o r 20 c e n t 10 a j e 0 NO SI P re g u n ta 1 Figura Nº 4-14: Pregunta Nº 1: ¿Conoce la planta de la figura? Tabla Nº 4-6. Análisis estadístico de la Pregunta Nº 1 ¿Conoce la planta de la figura? Pregunta 1 Frequency Percent Valid NO SI Total 58 93 151 38.4 61.6 100.0 Valid Percent 38.4 61.6 100.0 Cumulativ e Percent 38.4 100.0 Tabla Nº 4-7 . Porcentaje de las respuestas en las diferentes escuelas Pregunta 1 * Escuela Crosstabulation Pregunta 1 NO SI Total Count % within Escuela Count % within Escuela Count % within Escuela Escuela 20 DE LOS MARIANO SOLDADO JUNIO SARMIEN RAMOS CAJAL TOS 14 34 10 23.3% 79.1% 35.7% Total 58 38.4% 46 76.7% 9 20.9% 18 64.3% 20 100.0% 93 61.6% 60 100.0% 43 100.0% 28 100.0% 20 100.0% 151 100.0% 172 Pregunta Nº 2: ¿Cómo la llama? La pregunta Nº 2 está referida al nombre común que le dan a esta planta. Las respuestas son muy variadas y casi un 50% no contesta lo que se ve claramente en la Figura Nº 413. Tablas Nº 4-8 y 4-9. 50 40 30 P o r c e n t a j e 20 10 0 AL AR CH CO CO MA NC NO PO TA T A TO T O TO YU GODÁN ER CH I CO LVA S CO TLA ( LA CMATMATMATYO N ÓNDA NRY N I LBA E CEI A I B E o S E O LA BO ) O ( E CHEEDEEJGRO ALV r oj o RR AR IA Y DÍ N ) P re g u n t a 2 Figura Nº 4-15. Porcentajes de los diferentes nombres que dan a la planta Tabla Nº 4-8 Análisis estadístico de las respuestas a: ¿Cómo la llama? Pregunta 2 Frequenc y Valid ALGODÓN 1 ARÁNDANO 21 CHERRY 1 COCHINILLA 1 COCOBA 1 MALVA o 1 SALVIA NC 69 NO SE 9 POCOTE 1 TALA (CEIBO) 1 TALA CAIBO 1 (rojo) TOMATE 3 TOMATE 14 CHERRY TOMATE DE 7 JARDÍN YUYO 20 NEGRO Total 151 Percent .7 13.9 .7 .7 .7 .7 Valid Cumulativ Percent e Percent .7 .7 13.9 14.6 .7 15.2 .7 15.9 .7 16.6 .7 17.2 45.7 6.0 .7 .7 .7 45.7 6.0 .7 .7 .7 62.9 68.9 69.5 70.2 70.9 2.0 9.3 2.0 9.3 72.8 82.1 4.6 4.6 86.8 13.2 13.2 100.0 100.0 100.0 173 N C: no contesta Tabla Nº 4-9. Análisis estadístico de las respuestas por escuela a: ¿Cómo la llama? Pregunta 2 * Escuela Crosstabulation Escuela Total Pregunta 2 ALGODÓN Count % within Escuela ARÁNDANO Count % within Escuela CHERRY Count % within Escuela COCHINILL Count A % within Escuela COCOBA Count % within Escuela MALVA o Count SALVIA % within Escuela NC Count % within Escuela NO SE Count % within Escuela POCOTE Count % within Escuela TALA Count (CEIBO) % within Escuela TALA Count CAIBO (rojo) % within Escuela TOMATE Count % within Escuela TOMATE Count CHERRY % within Escuela TOMATE Count DE JARDÍN % within Escuela YUYO Count NEGRO % within 20 DE JUNIO LOS MARIANO SOLDADO SARMIENTO RAMOS CAJAL 1 2.3% 6 14.0% 15 53.6% 1 .7% 21 13.9% 1 2.3% 1 .7% 1 1 1.7% .7% 25 41.7% 34 79.1% 1 3.6% 1 .7% 1 1 3.6% .7% 10 35.7% 69 45.7% 9 15.0% 9 6.0% 1 3.6% 1 .7% 1 1 1.7% .7% 1 1 1.7% .7% 2 3.3% 1 2.3% 3 2.0% 14 14 23.3% 9.3% 7 7 11.7% 4.6% 20 20 100.0% 13.2% 174 Escuela Count 60 % within 100.0% Escuela Total 43 100.0% 28 100.0% 20 100.0% 151 100.0% Pregunta Nº 3: ¿Para qué se la usa? En esta pregunta se intenta conocer los usos que se dan a Solanum nigrum. 50 40 30 20 P o r c e 10 n t a j e 0 C CC CZ I-C I-F I M -C M NC NS S SU VU P re g u n ta 3 Figura Nº 4-16. Porcentajes de los diferentes usos que dan a la planta. Tabla Nº 4-10. Referencias de las iniciales utilizadas para indicar los diferentes usos Referencias: C CC CZ FF I-C I-F I M NS S SU VU COMESTIBLE-CONSUMO CULTIVO COBERTURA COMERCIALIZAR FRUTA FRESCA INDUSTRIAL-CONSUMO INDUSTRIAL-FRUTAL INDUSTRIAL MEDICINAL NO SABE SOMBRA SIN USO VARIAS UTILIDADES 175 Pregunta 3 Tabla Nº 4-11. Análisis estadístico de los diferentes usos que dan a la planta. Valid C CC CZ I-C I-F I M-C M NC NS S SU VU Total Frequency Percent 4 25 1 1 5 3 5 2 1 69 16 2 16 1 151 2.6 16.6 .7 .7 3.3 2.0 3.3 1.3 .7 45.7 10.6 1.3 10.6 .7 100.0 Valid Cumulativ Percent e Percent 2.6 2.6 16.6 19.2 .7 19.9 .7 20.5 3.3 23.8 2.0 25.8 3.3 29.1 1.3 30.5 .7 31.1 45.7 76.8 10.6 87.4 1.3 88.7 10.6 99.3 .7 100.0 100.0 Tabla Nº 4-12. Porcentaje de los usos que dan a la planta en las diferentes escuelas. Pregunta 3 * Escuela Crosstabulation Pregunta 3 C CC CZ I I-C I-F M M-C Count % within Escuela Count % within Escuela Count % within Escuela Count % within Escuela Count % within Escuela Count % within Escuela Count % within Escuela Count % within Escuela Count % within Escuela Escuela 20 DE LOS MARIANO SOLDADO JUNIO SARMIEN RAMOS CAJAL TOS 2 2 3.3% 4.7% 19 31.7% 5 11.6% 1 3.6% 1 1.7% 4 2.6% 25 16.6% 1 .7% 1 2.3% 2 3.3% Total 1 3.6% 1 .7% 5 17.9% 5 3.3% 4 14.3% 5 3.3% 3 10.7% 3 2.0% 1 3.6% 1 .7% 2 1.3% NC NS S SU VU Total Count % within Escuela Count % within Escuela Count % within Escuela Count % within Escuela Count % within Escuela Count % within Escuela 20 33.3% 34 79.1% 12 20.0% 10 35.7% 5 25.0% 69 45.7% 3 10.7% 1 5.0% 16 10.6% 2 3.3% 2 1.3% 2 3.3% 14 70.0% 16 10.6% 1 2.3% 60 100.0% 43 100.0% 1 .7% 28 100.0% 20 100.0% 151 100.0% Pregunta Nº 4: ¿ Es venenosa? De acuerdo a las respuestas sobre si es venenosa los alumnos no contestan o dicen que “no lo es” en un alto porcentaje. Sólo un 15% dicen que es venenosa. 50 40 30 20 P o r c e 10 n t a j e 0 NC NO N O S E S e d ic e q u e lo e sS I P re g u n ta 4 Figura Nº 4-17. Porcentajes de la pregunta 4 ¿ es venenosa? Tabla Nº 4-13 Análisis estadístico de los porcentajes de la pregunta ¿ es venenosa? Frequency Percent Valid Cumulativ Percent e Percent ii Valid NC NO NO SE SI Total 61 52 2 36 151 40.4 34.4 1.3 23.8 100.0 40.4 34.4 1.3 23.8 100.0 40.4 74.8 76.2 100.0 Tabla Nº 4-14. Porcentajes de la pregunta 4 ¿ es venenosa? en las diferentes escuelas. Pregunta 4 * Escuela Crosstabulation Pregunta 4 SI NC NO NO SE SI Se dice que lo es Total Count % within Escuela Count % within Escuela Count % within Escuela Count % within Escuela Count % within Escuela Count % within Escuela Count % within Escuela Escuela 20 DE LOS MARIANO SOLDADO JUNIO SARMIEN RAMOS CAJAL TOS 1 5.0% 14 23.3% 35 81.4% 10 35.7% 39 65.0% 8 18.6% 17 60.7% 3 15.0% Total 1 .7% 62 41.1% 64 42.4% 1 1.7% 1 .7% 5 8.3% 1 3.6% 16 80.0% 22 14.6% 1 1 1.7% .7% 60 100.0% 43 100.0% 28 100.0% 20 100.0% 151 100.0% Pregunta Nº 5: ¿Es una maleza? Un 30% de los alumnos dice que es una maleza, una gran proporción dice que no o no contesta. 50 40 30 20 P o r c e 10 n t a j e 0 NC NO N O SE SI P re g u n ta 5 iii Figura Nº 4-18. Porcentajes de la pregunta ¿ es maleza? Tabla Nº 4-15 Análisis estadístico de los porcentajes de la pregunta ¿es maleza? en las diferentes escuelas. Pregunta 5 * Escuela Crosstabulation Pregunta 5 NC NO NO SE SI Total Count % within Escuela Count % within Escuela Count % within Escuela Count % within Escuela Count % within Escuela Escuela 20 DE LOS MARIANO SOLDADO JUNIO SARMIEN RAMOS CAJAL TOS 15 35 10 1 25.0% 81.4% 35.7% 5.0% 17 60.7% 1 5.0% Total 61 40.4% 28 46.7% 6 14.0% 52 34.4% 1 1.7% 1 2.3% 16 26.7% 1 2.3% 1 3.6% 18 90.0% 36 23.8% 60 100.0% 43 100.0% 28 100.0% 20 100.0% 151 100.0% 2 1.3% Con el análisis de estas respuestas, se puede afirmar que no conocen esta planta, o que la confunden con otras plantas útiles. Se debe destacar que son personas muy jóvenes las que contestaron la encuesta. iv CAPÍTULO 5 CONCLUSIONES I- CONCLUSIONES SOBRE LOS ASPECTOS TAXONÓMICOS: En los caracteres diferenciales de importancia taxonómica, ordenados según su importancia se encontraron 17 morfológicos, de los cuales los más relevantes son los carpológicos, diez (10), cinco (5) florales, y por último dos (2) vegetativos los que resultaron de interés para diferenciar las especies. Algunos son de observación directa, caracteres macroscópicos y otros, caracteres microscópicos, necesitan instrumental de laboratorio para ser notados. Se destaca sólo un carácter anatómico de diferenciación. La caracterización citológica es simplemente una contribución a la citogenética de las malezas que componen el Complejo Solanum nigrum. Ordenados según su importancia los CARACTERES MORFOLÓGICOS comprenden a: Caracteres Reproductivos: * CARACTERES CARPOLÓGICOS 1. – Número de infrutescencias por plantas 2. – Número de frutos por planta 3. – Número de frutos por infrutescencia 4. – Diámetro de los frutos 5. – Color de los frutos 6. – Color de la pulpa 7. – Forma del cáliz en el fruto maduro 8. – Número de semillas 9. – Dimensión de las semillas (largo – ancho – peso) 10. – Color de las semillas * CARACTERES FLORALES 1. - Tipo de inflorescencias 2. - Posición de las inflorescencias (erecta – nutante) 3. – Número de inflorescencias por planta 4. – Número de flores por planta 5. – Número de flores por inflorescencias Los Caracteres Vegetativos tienen menos relevancia pero pueden servir junto a los otros para la diferenciación en el campo. 1. - Altura de la planta 2. – Altura de la bifurcación del tallo CARACTERES ANATÓMICOS Un carácter anatómico se ha encontrado, importante por su constancia: 1. - Presencia / ausencia de corpúsculos pétreos en el fruto 2. – Disposición de los corpúsculos pétreos e la pulpa de la baya CARACTERES CITOLÓGICOS v En los caracteres citológicos, si bien no se encontraron diferencias en el número cromosómico de las especies del Complejo Solanum nigrum, con este estudio se confirmó su concordancia con el número básico citado para el género y la poliploidía frecuente en estas especies de Solanáceas. Es importante destacar que algunos caracteres diferenciales en el Complejo Solanum nigrum son citados por primera vez en esta área de estudio. Las características reunidas en el siguiente esquema son las que han permitido identificar a las especies. Algunos son sólo datos teóricos y otros de experimentación propia. Solanum nigrum L., Solanum lorentzii Bitter y una especie que se la llamó Solanum nigrum – Intermedia, son las especies que componen este complejo, propio de la zona agrícola de Tucumán. Solanum nigrum L Solanum americanum Mill. Nº de frutos por infrutescencia 2-6 Color del fruto: negro brillante Diámetro: 7 mm Corpúsculos pétreos: ausentes Cáliz: reflexo en fruto maduro Flor: corola blanca con estrella central amarilla Nº de flores por inflorescencia 4-6 Tipo de inflorescencia: cimosa, simple, subumbelada Status: exótica Elevación: 0-3048 m sobre el mar Distribución: Cosmopolita Nº de frutos por infrutescencia 3-4 Color del fruto: negro brillante Diámetro: 8-9 mm Corpúsculos pétreos: 7,7 – 8 (prom) Cáliz: reflexo o ligeramente reflexo Flor: corola blanca con púrpura, con estrella central verde Nº de flores por inflorescencia 3-4 Tipo de inflorescencia: cimosa, simple, subumbelada Status: exótica Elevación: 0-2926 m sobre el mar Distribución: Incierta, pero probablemente este de los EEUU, West Indians y norte de Sud América. Solanum lorentzii Bitter Solanum bermejense Bitter Nº de frutos por infrutescencia 8-9 Color del fruto: morado opaco Diámetro: 9,5 mm Corpúsculos pétreos: 6 o más Cáliz: adherido al fruto maduro Flor: corola blanca con el centro morado y la estrella central amar. Nº de flores por inflorescencia 8-19 Tipo de inflorescencia: cimosa, bifurcada en 2 ramas Status: nativa Elevación: 500-2500 m Distribución: CA, JU, LR, SA, SE, TU Solanum sublobatum Willd. Nº de frutos por infrutescencia 3-5 Color del fruto: negro opaco Diámetro: 6 mm Corpúsculos pétreos: carece Cáliz: no acrecente Flor: corola blanca con estrella central amarilla Nº de flores por inflorescencia 2-4 Tipo de inflorescencia: cimosa, simple, subumbelada Status: nativa Elevación: 0-500 m sobre el mar Distribución: Sudeste de Brasil, Uruguay y la República Argentina (Córdoba, San Luis, Santa Fe, Buenos Aires, Entre Ríos). Nº de frutos por infrutescencia 3-4 Color del fruto: negro brillante Diámetro: 8-9 mm Corpúsculos pétreos: numerosos Cáliz: reflexo o ligeramente reflexo Flor: corola blanca con púrpura, con estrella central verde Nº de flores por inflorescencia 5-15 Tipo de inflorescencia: cimosa, bifurcada en 2 ramas Status: nativa Elevación: 1000- 2000 m Distribución: JU, SA Lo expuesto confirma la hipótesis. Solanum nigrum integra un complejo de malezas en el área agrícola de Tucumán. vi II-CONCLUSIONES SOBRE GERMINACIONY EMERGENCIA Sobre Identificación del Material de trabajo: Después de realizar exhaustivos análisis de las plantas que componen el Complejo Solanum nigrum, se determinó que en el campo el mayor número corresponde a Solanum nigrum. Sobre Reproducción: Las especies del Complejo Solanum nigrum se reproducen sólo por semillas. Sobre Germinación del Complejo, lo experimentado muestra que: Sólo germinan las semillas que se encuentran casi en la superficie. Sólo germinan las que se lavaron con agua corriente durante no menos de 30 minutos. Mejor lo hacen las que estuvieron un tiempo en la heladera (3-4 º C); necesitan un período de reposo y frío. La germinación en medio líquido es el mejor sustrato de todos los experimentados. Son semillas dormantes y longevas. La temperatura óptima de germinación es de 30 º C, luego de un período de reposo de aproximadamente tres meses. Las temperaturas óptimas se producen en el período primavero-estival, asegurándose con esto un flujo de emergencia continuo desde el banco de semillas del suelo durante el ciclo. Las temperaturas elevadas matan las semillas. El porcentaje de germinación a campo es bajo pero se compensa con la gran producción de semillas por planta lo que asegura una gran población de plántulas en los picos de emergencia. Con los métodos utilizados los porcentajes de germinación fueron nulos o bajos en SOLNI y en SOLLO salvo en la siembra que se utilizó sólo agua de red. La excepción se produjo en la forma intermedia (SOLNI-I) que sin tratamiento y sin reposo las semillas germinaron en macetas con tierra, en cajas de petri y en recipientes con agua de red. Sobre Emergencia en nuestras condiciones, con inviernos moderados se comprobó que: La emergencia de las plántulas se produce a fines del invierno y principios de primavera. Durante el flujo de emergencia se producen picos de abundancia de germinación semejantes en las especies del Complejo. Además, se observó que: La plenitud de floración y la aparición de los primeros frutos ocurre durante la primavera. vii La maduración generalizada de frutos ocurre a lo largo del verano, declinando con la llegada del otoño. Solanum nigrum generalmente muere, Solanum lorentzii continua viviendo, pero sólo vegeta. En años subsiguientes no se vuelve a encontrar especies del Complejo en los lugares donde se las observó, sin importar la cantidad de plantas que tuviera esa población. Es importante destacar que: El primer botón floral aparece en el campo entre los 40 y 50 días de producida la emergencia. Cuando la germinación ocurre más tarde, la floración se produce en la misma época que las que emergieron antes, en plantas que alcanzan una altura notablemente menor. Con las observaciones realizadas a campo, en nuestro medio, Solanum nigrum germina y emerge casi un mes antes que Solanum lorentzii. Solanum nigrum florece antes y ambas especies llegan a plenitud de fructificación en la misma época del año, en verano. Sobre el comportamiento en laboratorio / invernáculo/campo Las condiciones de laboratorio, y de invernáculo no son propicias para la germinación y el desarrollo de las especies del Complejo Solanum nigrum. Las plántulas que emergen en estos medios artificiales son débiles y en pocos días mueren. Las que sobreviven raramente llegan al estado reproductivo. La emergencia no es continua. El número de las que nace en el campo es mucho mayor, la emergencia es continua. Si bien pocas son las que extienden su desarrollo esto es muy importante por la gran cantidad de frutos y de semillas que produce cada planta. Con los métodos utilizados en laboratorio los Porcentajes de Germinación fueron nulos o bajos salvo en la siembra que se utilizó sólo agua de red. La mayor cantidad de semillas germinadas fue la de Solanum lorentzii. Para el Porcentaje de Germinación Relativa (PGR), 30 días es el tiempo de registro de germinación. Las que germinaron generalmente lo hicieron después de ese tiempo. La Velocidad de Germinación (VG) sólo se analizó en las siembras en germinador. Lo obtenido muestra que a 30ºC germinan más rápido, especialmente en la siembra inmediata. Con el IVG del modelo Maguire, cuanto mayor sea el valor obtenido, mayores serán la velocidad de germinación y el vigor de la simiente. En general la velocidad de estas especies es variable y más bien baja. La Tasa de germinación es muy baja en todos los casos para las dos especies estudiadas En el Potencial de infestación SOLNI posee los valores más altos obtenidos tanto para cantidad de semillas y viabilidad como para semillas ingresantes, para individuos con capacidad reproductiva, para real capacidad reproductiva y para potencial de infestación. Esta especie infestará mayor superficie a partir de la producción de semillas de sus frutos. SOLLO posee los viii valores más bajos del análisis excepto poder germinativo y densidad. Su capacidad de infestación es menor pero por ello no deja de ser importante como competidora de cultivos. En Demografía experimental la tasa de supervivencia baja determinó que la especie puede mantener sus relaciones de interferencia con moderadas estrategias de competencia, buscando ocupar lentamente un territorio. Esto explica su presencia en manchones y a espacios breves alrededor de la planta original. La TBM es alta y superior a la media de la especie; la TBN es intermedia e inferior a la media y la probabilidad de sobrevivir es muy baja. Un grupo de especies estrechamente relacionadas a Solanum nigrum, forman un complejo propio del área agrícola de la provincia de Tucumán y las diferencias en relación con la germinación y emergencia, no influyen en su importancia como malezas. Estas características ecobiológicas no diferencian sus efectos como malezas. Lo expuesto sobre germinación y emergencia no confirma la hipótesis. ix III- CONCLUSIONES SOBRE OTROS ASPECTOS DE IMPORTANCIA SOBRE EL Complejo Solanum nigrum De los antecedentes bibliográficos se rescata que: Hay menciones importantes sobre el uso medicinal pero en nuestra región hay pocos antecedentes y poco conocimiento sobre la especie. Se destaca su importancia agronómica como planta tóxica para el ganado, cerdos, patos y gallinas, lo mismo que para el hombre, debido a su alto contenido en alcaloides (solanina y solanidina). Desde el punto de vista de la alimentación humana los frutos y las hojas son comestibles, los brotes tiernos se comen como verdura, sus hojas como espinaca, y de muchas otras formas. Con los frutos se preparan jaleas, mermeladas, confituras, conservas, y se utilizan las bayas para extraer tinturas y colorantes. En algunos lugares de África y Asia se cultivan estas especies consideradas malezas, para cosechar sus hojas y sus bayas pero no existen datos relevantes sobre la extensión cultivada. Se ha estudiado que las partes vegetativas son muy nutritivas por el contenido en proteínas, fibras, minerales y vitaminas y que las bayas contienen calcio, hierro, caroteno, y vitaminas B y C. Las semillas contienen vitaminas C y caroteno. En Argentina no existen datos de estas especies como fuente de alimentación. Al experimentar sobre las propiedades como forrajeras se encontró que la proteína bruta presenta un excelente nivel. La fibra cruda, (celulosa, hemicelulosa y lignina, la porción menos digerible de la planta) tiene valores medios a bajos en el estado fenológico de floración a fructificación. Los valores de ceniza total y de los hidratos de carbono son buenos. Por medio de las cenizas los animales toman los minerales, y los hidratos de carbono son los jugos celulares citoplasmáticos, fácilmente digeribles. El resultado es una digestibilidad buena a excelente. Con los resultados obtenidos se puede afirmar que este estado fenológico (plenitud de floración a fructificación) las especies analizadas tienen una aptitud forrajera buena. Los datos de palatabilidad completarían el panorama necesario para establecer la verdadera importancia forrajera de estas especies. Entre los insectos recolectados se encontraron especies fitófagas y predadores. Además se encontraron ácaros. Estos totalizaron seis especies correspondientes a tres órdenes: 50 % Hemípteros, 20 % Coleópteros y 30 % Arácnidos. Se observó, además, las presencia persistente de hormigas y abejas. Son plantas hospederas de áfidos que producen daños de consideración en los cultivos hortícolas y frutales del medio. x Las referencias bibliográficas sobre estas plagas demuestran la importancia de este aspecto debido a que todos los cultivos citados en éstas, también se hacen en Tucumán. Las especies del Complejo Solanum nigrum que sirven como los hospederos alternativos de plagas en varios cultivos de importancia comercial podrían ser usados como potenciales agentes de control. Por lo expuesto se hace necesario conocer la relación entre los insectos y estas malezas como hospederos alternativos, para poder realizar la planificación del control de plagas o bien mantenerlas como reservorios de insectos benéficos. En Tucumán se carece de suficiente información al respecto por lo que se debería continuar con el estudio de estas malezas asociadas a insectos o ácaros. La encuesta realizada en escuelas agrotécnicas en diferentes áreas agroecológicas de Tucumán sirvió para de obtener datos sobre el conocimiento y usos de las especies del Complejo Solanum nigrum. Con el análisis de las respuestas, se puede afirmar que no conocen esta planta, o la confunden con otras plantas útiles. Se debe destacar que fueron personas muy jóvenes las que contestaron la encuesta. Si se consideran los antecedentes de las Solanáceas como plantas medicinales, como alimento para el hombre y los animales (forrajes) y además como plantas tóxicas se puede decir que no existen plantas tóxicas sino un uso indebido de las plantas. Citando a Paracelso podemos decir que: "Un tóxico puede ser un remedio y un remedio puede ser un tóxico. Todo depende de la dosis". RECOMENDACIONES De acuerdo a lo expuesto, estas plantas, si bien se han considerado principalmente como malezas muy perjudiciales, presentan varios aspectos positivos. No son tan malas y tampoco tan buenas..... Holm et al., (1977) habían incluido a Solanum nigrum entre las peores malezas del mundo pero no es así, pues se ha comprobado principalmente que no tiene un carácter invasor. Es una maleza importante. Si el pequeño productor la conoce puede arrancarla antes de que llegue al estado reproductivo. Se puede decir lo mismo para Solanum lorentzii. Sobre esta especie no se encontraron datos. Estas plantas podrían usarse como una alternativa forrajera, para ello es necesario conocer a las especies y el momento en que éstas carecen de toxicidad. Se ha comprobado la buena aptitud forrajera del Complejo, y esto debería aprovecharse. Con la información obtenida de la bibliografía consultada se puede afirmar que el conocimiento de estas plantas podría resultar de interés a los pobladores rurales proporcionando una fuente de ingreso no habitual o ser utilizadas como fuente de medicinas no tradicionales. xi Por último se ve la necesidad de enseñar a los productores caracteres simples que permitan la identificación de las plantas del Complejo Solanum nigrum para su aprovechamiento, para su control o para prevenciones, según sean sus necesidades. xii BIBLIOGRAFÍA Agroinformación. 2002. Ficha de la plaga: Araña http//www.agroinformacion.com [consultada el 010705]. roja. Consultado en: Allmaras, R. R. y R. H. Dowdy. 1985. Conservation tillage systems and their adoption in the United States. Soil Tillage Research. 5: 197-222 Alonso, J. 2004. Plantas Medicinales empleadas por los Nativos del Noroeste. Consultado en: http//www.aglia.com/sitio/index. [consultada el 200304] Altieri, M. 1997. Ecología y Manejo de Malezas. En: Agroecología. Bases científicas para una agricultura sustentable. CLADES. Pp 391-420. Andrada, A. B.; Mansilla de Andrada, N. Pastoriza, A. 2003. Caracterización citológica en Solanum nigrum L. Editado por Agrosur. Universidad Austral de Chile. Facultad de Ciencias Agrarias. Chile. ISNN 0304 - 8802. Volumen 31 N º 1. Enero - Junio 2003. 77 - 81. Arbiza Aguirre, S. I. 1986. Producción de caprinos. Apéndice 2. Plantas tóxicas para cabras en México. A.G.T. Editor S. A. Primera Edición. 420 – 441pág. Arévalo, R. A. 1975. Especies de Malezas de la Caña de Azúcar de la República Argentina. Revista Agronómica del Noroeste Argentino. XXII(*1-2) 95-103. Arévalo, R. A. 1977. Competencia de malezas y Períodos Críticos. Primer Curso Básico de Control de Malezas. Facultad de Agronomía y Zootecnia. UNT. p 6. Barboza, G., Romanutti, A. Arroyo, S. C. Leuenberger, A. A. Cocucci, y M. R. Figueroa Romero. 1999. Solanaceae en Zuluaga, F. O. y O. Morrone (eds.), Catálogo de las Plantas Vasculares de la República Argentina II. Fabaceae, Zygophyllaceae (Dicotiledoneae): 1059 – 1114. Missouri Botanical Garden Press. Barralis, G. y R. Chadoeuf. 1987. Potencial semencier des terres arables. Weed Research. 27: 417-424. BayerCropScience. 2004. PULGÓN VERDE DEL DURAZNERO (Myzus persicae). Consultado en: http//www.bayercropscience.com.ar/arg/pages/actualidades [consultada el 250505]. Bayón, N. D. 2001. Producción de Legumbres: Caracterización del Sector: Poroto C.F.I. Consultado en: http//www.cfired.org.ar/Legumbres [consultada el 210605]. Botanical. Hierba Mora [Consultado en Botanical on-line.com/alcaloideshierbamora/htm#] consultada el 190806. Botta, S. M. 1979. Las especies Argentinas del género Aloysia (Verbenáceas). Darwiniana Vol. 22, N º 1-3, 67- 108. Buhler, D. D. 1995. Influence of tillage systems on weed populations dynamics and management in corn and soybean in the central USA. Crop Sci. 35: 1247-1258. Buhler, D. D. 1996. Development of alternative weed management strategies. J. Prod. Agric. 9(4): 501-505. Cabrera, A. L. 1983. Flora de la Provincia de Jujuy. (Argentina). (Cletráceas a Solanáceas). Tomo XIII. Parte VIII. Colección Científica del INTA. Buenos Aires. 508 Pág. xiii Caribbean area. Plantas tóxicas. Consultado http//www.pr.nrcs.usda.gov/pm/ptindex.htm.[consultada el 160803]. en: Chaila, S. 1997. Matoecología de la Caña de Azúcar. Facultad de Agronomía y Zootecnia. UNT. Publicación Especial Nº 32. 130 pp. Chaila, S.; F.R. Piscitelli; H. González Navarro; M.H. de la Vega y N. Ortiz. 1997. Efecto de tratamientos mecánicos y químicos sobre la mortalidad y supervivencia de Sicyos polyacanthus Cogn en caña de azúcar. ASAPROVE. 2: 202-209. Chaila, S. 2001. Bioecología, demografía y estrategias de manejo de Sicyos polyacanthus Cogn. Tesis doctoral. Facultad de Agronomía y Zootecnia. Universidad Nacional de Tucumán. 414 p. Chaila, S.; M. T. Sobrero; R. Fuentes; y M. del C. Ochoa. 2006. Sustentabilidad y efecto ambiental y agronómico de los herbicidas aplicados al suelo. Universidad Nacional de Santiago del Estero. 150 p. De Andrada, N.; H. Robinet; O. Arce; B. Díaz; S. Guillén; E. Gallo; N. Mansilla. 1995. "Relevamiento y determinación de la distribución de malezas en la zona sojera del Noreste de Tucumán. República Argentina". XII Congreso Latinoamericano de Malezas. Asociación Latinoamericana de Malezas (ALAM) – Instituto Nacional de Investigación Agropecuaria (INIA). Serie Técnica N ° 56:50-60. Montevideo - Uruguay. Del Monte, J. P. y A. M. Tarquis. 1997. The role of temperature in the seed germination of two species of Solanum nigrum complex. Journal of experimental botany. v. 48 (317) p 2087-2093. Debaeke, P. y M. Sebillotte. 1988. Modélisation de l’evolution a long terme de la flore adventice: I.Constructiond’un modéle descriptif de l’evolution quatitative du stock de semences a l’horizon travaillé. Agronomie, Paris, 8(5) :393-403. Devani, M. R., O. R. Ricci y G. Salas. 1993. Malezas anuales en soja y su control en Tucumán. Avance Agroindustrial, octubre 1993. 10-13. Dieckmann, J. G. 1912. Contribución al Estudio de las Solanáceas Argentinas. Tesis. Facultad de Ciencias Exactas, Físicas y Naturales. Universidad Nacional de Buenos Aires. 193 Pág. Dorado, J., J. P. Del Monte y C. López Fando. 1997. Influencia de los sistemas de manejo en la evolución del banco de semillas del suelo en cultivos cerealistas. Anales VI ° Congreso de la Sociedad Española de Malherbología. pp. 121-126. Dottori, N. 1995. Desarrollo y estructura de fruto y semilla en Solanum sect. Cyphomandropsis (Solanaceae) de Argentina. Kurtziana. 24:83-104. Edmonds, J. M. and J. A. Chweya. 1997. Black nightshades. Solanum nigrum L. and related species. Promoting the conservation and use of underutilized and neglected crops. 15. Institute of Plant Genetics and Crop Plant Research, Gatersleben/International Plant Genetic Resources Institute, Rome, Italy. 115 Pág. Enciclopedia Universal Ilustrada. Europeo – Americana. 1927. Espasa Calpe S. A. Madrid Barcelona. Tomo 57 Sola – Subn. 9-12. Especies con usos no maderables en bosques de Encino, Pino y Pino-Encino en los estados de Chihuahua, Durango, Jalisco, Michoacán, Guerrero y Oaxaca. xiv Consultado en:http//www.semarnap.gob.mx/pfnm/index.html. [consultada el 160803]. Fernández Quintanilla, C. 1988. Studying the population dynamics of weeds. Weed Research. 25(6) :443-447. Figueroa Romero, M. R.; E. Bulacio y N. B. Muruaga. 2000. Sinopsis de la Familia Solanaceae en Tucumán (Argentina). Lilloa 40 (1): 15 – 64. Font Quer, P. 1962. Plantas Medicinales. El Dioscórides Renovado. Editorial Labor. Barcelona. España. 1033 Pág. Frana, J. 1998. Información técnica Nº 228. Relevamiento de artrópodos presentes en un cultivo de maíz en siembra convencional y con riego suplementario. Consultado en: http//www.inta.gov.ar/rafaela [consultada el 140206]. Futuros y Opciones. 2004. Plagas del cultivo de la soja. Consultado en: http//www.futurosyopciones.com./granos/producción/especiales/soja [consultada el 210605]. Futuros y Opciones. 2005. Chinches de los cuernos (Dichelops furcatus). Consultado en: http//www.fyo.com/granos/produccion. [consultada el 210605]. Gallo, G. G. 1987. Plantas Tóxicas para el ganado en el cono sur de América. Editorial Hemisferio Sur. 2ª edición. 213 Pág. Giganti, H., G. Dapoto y R. Gonzalez Junyent. 1997. Insectos y Ácaros Asociados a los Principales Cultivos Hortícolas del Alto Valle de Río Negro y Neuquén, Argentina. Horticultura Argentina 16 (40-41):29-36. Consultado en: http//www.redagraria.com/profesionales/ingenieros/giganti_humberto_e.html. [consultada el 210605]. Goodhue R. 1998. Nightshade (Solanum spp.) Biology Outline & Bibliography. Consultado en: http//www.iastate.edu/~weeds/WeedBiolLibrary/solanumbiblio.htm. [consultada el 170308]. Grieve, M. 2004. A Modern Herbal. Nightshade, Black. Consultado en: http//www.botanical.com. [consultada el 080404]. Grundi, A. C. y A. Mead. 2000. Modeling weed emergente as a function of meteorological records. Weed Science. 48: 594-603. Guarino, L. 1977. Global Facilitation Unit (GDU). Traditional African Vegetables. Consultada en http://www.underutilized-species.org/species/herb.asp. Consultada el 120408 Heap, J. W. and R. J. Carter. 1999. The biology of Australian Weeds. 35 Solanum eleagnifolium Cav. Plant Protection Quarterly. Vol.14 (1):2-12. Holm, L. F., D. L. Plucknet, J. V. Pancho and J. P. Herberger. 1977. Solanum nigrum L., Solanaceae, Nightshade Family. P. 430-435. The World’s Worst Weeds. Distribution and Biology. Univ. Press of Hawaii, Honolulu. 609 Pág. Holt, J. S. y A. B. Boose. 2000. Potential for spread of Abutilon thephrasti in California. Weed Science. 48:43-52. Imwinkelried, J. M., F.D. Fava y E.V. Trumper. 2004. Pulgones que atacan al cultivo del trigo. Boletín Nº 7. Consultado en: http//www.inta.gov.ar/manfredi [consultada el 210605]. xv Infoagro. 2004. El virus PVY en pimiento. Consultado en: http//www.infoagro.com [consultada el 130705]. Keeley, P. E. and R. J. Thullen. 1989. Growth and Competition of Black Nightshade (Solanum nigrum) and Palmer Amaranth (Amaranthus palmeri) with cotton (Gossypium hirsutum). Weed Science. Volume 37:326-334. Keeley, P. E. and R. J. Thullen. 1991. Biology and Control of Black Nightshade (Solanum nigrum) in cotton (Gossypium hirsutum). Weed Technology. Volume 5:713-722. Kissman, K. G. y D. Groth. 1995. Plantas Infestantes e Nocivas. Tomo III. 1ª Edición. BASF Brasileira S.A. - Indústrias Químicas. São Paulo. Brasil. 683 Pág. Kleifeld, Y., T. Blunfeld and G. Herzlinger. 1991. BAY SMY 1550 for Selective Control of Black Nightshade (Solanum nigrum) Other Weeds in Tomato (Lycopersicon esculentum) Fields. Weed Technology. Volume 5:592-597. Kogan, M. 1992. Malezas. Ecofisiología y estrategias de Control. Facultad de Agronomía. Pontificia Universidad Católica de Chile. Santiago. Chile. 402 Pág. Kogan, M. 1993. Manejo de malezas en plantaciones frutales. Pontificia Universidad Católica de Chile. Colección en Agricultura. 277 pp Lambelet-Haueter, C. 1985. Comparaisons entre flore réelle et flore potentielle en grandes cultures de la région genevoise. Candollea. 40 : 99-107. Lambelet-Haueter, C. 1986. Analyse de la flore potentielle, en relation avec la flore réelle, en grandes cultures de la région genevoise. Candollea. 41 : 299-323. Leguizamón E. S. 2003. Las Malezas y el Agroecosistema. Cátedra de Malezas. Departamento de Producción Vegetal. Facultad de Ciencias Agrarias. U.N.R. (2123). Zavalla. Santa Fe. [Consultado en: http://www.e-campo.com]. (consultado el 050707) Long, A. M. 2002. Senecio quequensis, nuevo sinónimo de Senecio bergii (Asteraceae, Senecioneae). Hickenia 3. (33): 125-127. Lorenzi, H. 2000. Manual de Identificação e Controle de Plantas Daninhas. Plantio direto e convencional. Instituto Plantarum de Estudos da Flora Ltda. 5ª Edição. Nova Odessa, São Paulo. Brazil. 371 pág. Lorenzi, H. 2000. Plantas Daninhas do Brasil. Terrestres, aquáticas, parásitas e tóxicas. Instituto Plantarum de Estudos da Flora Ltda. 3ª Edição. Nova Odessa, São Paulo. Brazil. 640 Pág. Lorenzi H. y F. J. Abreu Matos. 2002. Plantas Medicinais no Brasil. Nativas e Exóticas. Nova Odessa. Instituto Plantarum de Flora Ltade. 544 Pág. Lyon, D. J.; S. D. Millar y G. A. Wicks. 1996. The future of herbicide in weed control systems of the great plains. J. Prod. Agric. 9(2):209-215. Maguire, J. D. 1962. Speed of germination aid in selection and evaluation for seedling emergency and vigor. Crop Sci., 2(1):176-177. xvi Mansilla, N.; L. Villagrán; N. De Marco; O. Arce y B. Díaz. 1998. "Caracterización morfológica de Solanáceas que se comportan como malezas en campos de soja del noroeste argentino". Resumen N ° 51, XV Jornadas de la Sociedad de Biología de Tucumán. Tafí del Valle. Tucumán. Mansilla de Andrada, N. Chaila S..2004. Requerimientos térmicos para la germinación de Solanum nigrum L. (Solanáceas). XXI Jornadas Científicas de la Sociedad de Biología de Tucumán. Tafí del Valle. Tucumán. Octubre de 2004. Edición Digital. Pág. 38-39 Mansilla de Andrada, N.; Andrada, A. B.; Pastoriza, A..2004. Aspectos relacionados con la identidad de especies del Género Solanum. XXI Jornadas Científicas de la Sociedad de Biología de Tucumán. Tafí del Valle. Tucumán. Octubre de Edición Digital. Pág. 390-391. Mansilla de Andrada, N. J. y Salvatore A. R.. 2004. Presencia de Insectos en dos especies del Género Solanum en diferentes localidades de la Provincia de Tucumán. XXI Jornadas Científicas de la Sociedad de Biología de Tucumán. Tafí del Valle. Tucumán. Edición Digital. Pág. 477. Mansilla de Andrada, Nora J. y Díaz, Bibiana E. L.. 2006. Reconocimiento de Solanum nigrum L. (Solanáceas) una especie tóxica para el ganado. XXIII Jornadas Científicas Asociación de Biología de Tucumán. Tafí del Valle. Tucumán. Mansilla de Andrada N. J. y Chaila S. 2007. Cálculo de la velocidad de germinación relativa(“RVG”) de Solanum nigrum L. y Solanum lorentzii Bitter. Pág. 128. XXIV Jornadas Científicas de la Asociación de Biología de Tucumán. Tafí del Valle. Tucumán. Mansilla de Andrada N. y Chaila S. 2007. Potencial de infestación de Solanum nigrum L. y Solanum lorentzii Bitter. Pág. 157. XXIV Jornadas Científicas de la Asociación de Biología de Tucumán. Tafí del Valle. Tucumán. Mansilla de Andrada, Nora J. y Salvador Chaila. 2008. Demografía experimental de la matoespecie Solanum nigrum L. en cañaverales de la localidad de Los Ralos (Tucumán – Argentina). Resumen Nº 36. XXV Jornadas Científicas de la Asociación de Biología de Tucumán. Tafí del Valle. Tucumán. Marzocca, A. 1985. Nociones Básicas de Taxonomía. San José. Costa Rica. IICA. 263 Pág. Marzocca, A. 1993. Manual de Malezas. (4ª ed.). Edit. Hemisferio Sur. Buenos Aires. (Argentina). 684 Pág. Marzocca, A. 1994. Guía descriptiva de malezas del cono sur. Buenos Aires. INTA. 304 Pág. Marzocca, A. 1997.Vademécum de Malezas medicinales de la Argentina. Indígenas y Exóticas. Orientación Gráfica Editora. Buenos Aires. Argentina. 363 Pág. Massaro, R. y J.C. Gamundi. Detectan un aumento de los ataques de chinches en soja. INTA Oliveros. INTA INFORMA Nº 379. Consultado en: http//www.inta.gov.ar/ [consultada el 250306]. Mayor, J. P. y F. Dessaint .1998. Influence of weed management strategies on soil seed bank diversity. Weed Research. 38: 95-105 McGiffen, M. E. Jr. and J. B. Masiunas. 1992. Prediction of Black and Eastern Black Nightshade (Solanum nigrum and Solanum ptycanthum) Growth Using Degree-Days. Weed Science. Volume 40:86-89. xvii Mcgiffen, M. E. Jr., J. B. Masiunas and J. D. Hesketh. 1992. Competition for Light Between Tomatoes and Nightshades (Solanum nigrum or Solanum ptycanthum ). Weed Science. Volume 40:220226. Moran Lemir, A. H. 1997. Comportamiento germinativo de Ipomoea nil (L.) Roth en función de la temperatura y del stress hídrico. Actas del XIII Congreso Latinoamericano de Malezas. Buenos Aires. Argentina. 66-74. Morton, C. V. 1976. A revision of the Argentine Species of Solanum. Academia Nacional de Ciencias. Córdoba. República Argentina. 260 Pág. Mulugeta, D. y D. E. Stoltenberg. 1997. Seed bank characterization and emergence of a weed community in a moldboard plow system. Weed Science. 45: 54-60. Nasca, A. J., A. L. Terán, R. V. Fernández y A. J. Pasqualini. 1981. Animales perjudiciales y benéficos a los cítricos en el noroeste argentino. CIRPON. Centro de investigaciones sobre regulaciones de poblaciones de organismos nocivos. 362 Pág. Navas, M. L. 1991. Using plant populations biology in weed research a strategy to improve weed management. Weed Res. 31: 171-179 Ogg, A. G. Jr., B. S. Rogers and E. E. Schilling. 1981. Characterization of Black Nightshade (Solanum nigrum L.) and related species in the Unites States. Weed Science. Volume 29, Issue 1:27-32. Ogg, A. G. Jr and B. S. Rogers. 1989. Taxonomy, Distribution Biology, and Control of Black Nightshade (Solanum nigrum L.) and related species in the Unites States and Canada. Rev. Weed Science. 4:25-58. Olicer cereales. 2005. Soja. Síntomas sospechosos. Chinches. Consultado en: http//www.olicer.com.ar [consultada el 210605]. Ortego, J. 1990. Bioecología de los áfidos (Homoptera aphidoidea) de Malargüe, Mendoza, Argentina y su relación con la epidemiología del virus "Y" de la papa (raza necrótica). Consultado en: http//www.inta.gov.ar/ramacaida/index.html [consultada el 250505]. Orzolek, M. D. 1999. Weed of the Month: Nightshade (Solanum nigrum complex). Department of Horticulture. The Vegetable Gazette. Vol. 3 N º 2. (Documento de Internet consultado el 021999). Otero Aira, L. 2001. Las Plantas Alucinógenas. Editorial Paidotribo. Barcelona. 4ª Edición. 224 Pág. Portolano, N. 1990. Flora Tóxica y medicinal, en Explotación de ganado ovino y caprino. Ediciones Mundi-Prensa. España. 49 - 56. Radosevich, S. R. y J. S. Holt. 1984. Weed Ecology: Implications for vegetation management. J. Wiley and Sons. N.Y. Ragonese, A. E. y V. A. Milano. 1984. Vegetales y substancias de la flora argentina, en L. R. Parodi. Enciclopedia Argentina de Agricultura y Jardinería. 2(8-2). Buenos Aires. Ed. ACME S.A.C.I.. 2ª. Edición. 413 pág Roberts, H. A. and P. M. Lockett. 1978. Seed dormancy and field emergence in Solanum nigrum L. Weed Research. Vol. 18. 231-241. xviii Roberts, H. A. y P. M. Feast. 1972. Fate of seeds of some annual weeds in different depths of cultivated and undisturbed soil. Weed Research. 12: 316-324. Roncaglia, R. V. R.; N. D. M. de Roncaglia; B. Díaz; N. de Andrada; G. Würschmidt; E. Gallo; L. F. de Krapovickas. 1991. "Inventario de Malezas en Trigo (Triticum aestivum L.) en cultivos del Sur de la Provincia de Tucumán y Dpto. Santa Rosa (Catamarca). República Argentina". ASAM - XII Reunión Argentina sobre la Maleza y su Control. Cultivos de Cereales. Tomo 2: 153 -156. Mar del Plata, Argentina. Roncaglia, R. V. R.; N. D. M. de Roncaglia; B. Díaz; N. de Andrada; E. Gallo; L. F. de Krapovickas; O. Arce. 1993a. Malezas frecuentes en el noroeste argentino. Manual de Malezas. Facultad de Agronomía y Zootecnia. U.N.T.- DowElanco. Tucumán. Argentina. 79 Pág. Roncaglia, R. V. R.; N. D. M. de Roncaglia; E. Gallo; G. Würschmidt; B. Díaz; N. de Andrada; L.F. de Krapovikcas. 1993 b. "Relevamiento y Determinación de Malezas en Campos de Soja (Glycine max (L.) Merr.) en el Sur de la Provincia de Tucumán y Noroeste de la Provincia de Catamarca". Miscelánea N ° 87. Facultad de Agronomía y Zootecnia. U.N.T.. Tucumán. 17 Pág. Roncaglia, R.; N. De Marco de Roncaglia; O. Arce; B. Díaz; N. M. de Andrada; L. V. de Krapovickas. 1995. "Claves prácticas para el reconocimiento de géneros y especies de malezas con caracteres morfológicos semejantes en cultivos de granos de verano del noroeste argentino". XII Congreso Latinoamericano de Malezas. Asociación Latinoamericana de Malezas (ALAM) – Instituto Nacional de Investigación Agropecuaria (INIA). Serie Técnica N ° 56: 61-80. Montevideo - Uruguay. Roncaglia, R. V.; O. Arce; N. de M. Roncaglia; N. de Andrada; L. de Krapovickas y B. Díaz. 1998. "Relevamiento de malezas en campos de soja en el este de Salta, Argentina." XIII Congreso Latino Americano de Malezas. (A.L.A.M.). Cereales y Cultivos Industriales. Tomo II. 159 168. Buenos Aires. Argentina Roncaglia, R.,V.,R., N. De Marco de Roncaglia; O. Arce; B. Díaz; N. Mansilla de Andrada; L. de Krapovickas. 2000. Malezas Frecuentes en el Norte Argentino. F.A.Z.-U.N.TDowAgroSciences. 63 Pág. Sahoo, U. K.; R. S. Tripathi; H. N. Pandey y J. Misra. 1994. Population dynamics of buried weed seeds as influenced by shifting and terrace cultivation in the humid subtropics of India. Weed Research. 34: 157-165. Saini, E. D. 2006. Insectos y ácaros perjudiciales al cultivo de soja y sus enemigos naturales. Publicación del Instituto de Microbiología y Zoología Agrícola Nº 4 . CICVyA / INTA. Castelar. Buenos Aires. 80 Pág. Sarandón, S. J. 2005. Agroecología. El camino hacia una agricultura sustentable. Ediciones Científicas Americanas. Bs. As.557 p Stoller, E. W. y L. M. Wax. 1974. Dormancy changes and fate of some annual weed seeds in soil. Weed Science. 22(1): 151-155. Subils, R. 1977. Las especies de Euphorbia de la República Argentina. Museo Botánico. Facultad de Ciencias Exactas, Físicas y Naturales. Córdoba. Argentina. Kurtziana 10:83-248. xix Swanton, C. J. y S. F. Weise. 1991. Integrated weed management in Ontario the rationale and approach. Weed Technol. 5:657-663. Tan, C. S. and S. E. Weaver. 1996. Water use patterns of eastern black nightshade Solanum ptycanthum) and hairy nightshade (Solanum sarrachoides) in response to shading an water stress. Horticulture Section. Canadian Journal of Plant Science. 261-265. Todoagro. 2005. Sistema de alerta. Consultado http//www.todoagro.com.ar/seccion/Producción Vegetal. [consultada el 210605]. en: Troiani, R. M. de. 2006. Metabolitos Secundarios. Alcaloides. Universidad Nacional de La Pampa. Facultad de Agronomía. Química III. [Documento de Internet consultado el. 270106] Uninca. Región de la Sabana Occidente - Colombia. Plantas medicinales de la región– Solanáceas. Consultado en: http//www.uninca.edu.co/tesis/FTWeb/Solanaceas.html#Solanum Nigrum. [consultada el 160803]. Voll, E.; D. L. Pizza Gazziero; A. M. Brighenti; F. S. Adegas; C. De Almeida Gaudéncio y C. E. Voll. 2005. A dinámica das plantas danihas e práticas de manejo. EMBRAPA. Brasil. 85 pp. Weed Science Society of America. 1984. Composite List of Weeds. Weed Science, 32 (Supp 1.2.): 1137. Wikipedia. Leaves, shoots and seeds used in Chinese medicine, Perry, (1980). Consultado en: http//www.wikipedia.org/wiki/Solanum_nigrum [consultada el 160803]. Wikipedia - Solanum nigrum. Consultado en: http//es,wikipedia.org/wiki/solanum-nigrum. [consultada el 281204]. World Wise Garden Seeds. Black nightshade is a very valuable plant. Consultado en: http//store4.yimg.com/l/worldwise-gardenseeds_1736_5577377. [consultada el 080404]. Zanin, G.; A. Berti y M. C. Zuin. 1989. Estimation du stock semencier d’un soil labouré ou en semis direct. Weed Research. 29 : 407-417. xx