Copia digital - Biblioteca Virtual de la Real Academia
Anuncio

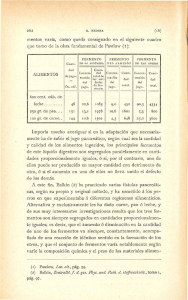
LA FARMACIA MODERNA» U Á R I O Página. F . BUSTINZA.—Glucósidos cianogenéticos y fermentosque catalizan su hidrólisis Combate biológico dé los insectos dañinos Extractos de publicaciones Legislación Q. E S C O L A R . — E s indispensable una Asociación de titulares Academia Nacional de Farmacia J . VÁZQUEZ.—Análisis de la leche ( c o n t i n u a c i ó n ) . — Noticias 591 609 618 323 529 550 353 559 MADRID Año XLVII 10 Enero 1936 LA F A R M A C I A M O D E R N A REVISTA C I E N T I F I C O - P R O F E S I O N A L Fundada en 1890 por L. Siboni y A . Bellogín. Se publica los d í a s 10 y 25 de cada mes. Director: A . CHALMETA España Precio de la suscripción Portugal y América latina anual Otros países 15 pías. 20 » 25 » Número suelto, 1 pta. » atrasado, 2 » R e d a c c i ó n y Administración: Guzmán el Bueno, 53 pral. izqd. Teléf. 48209 Advertencias á los nuevos suscriptores. Todas las suscripciones terminarán en 3 1 de Diciembre, pudiendo optar el suscriptor por abonar únicamente la fracción correspondiente al tiempo que falte para terminar el año natural ó suscribirse por el año entero, remitiéndole todos los números del mismo, incluso los atrasados. Advertencias á nuestros colaboradores. Se ruega á los colaboradores de trabajos científicos indiquen en la primera página si desean corregir personalmente las pruebas y el número de separatas que quieren se les remita, entendiendo que se les otorga gratuitamente 2 5 ejemplares. Las figuras y gráficos que acompañen á los trabajos deben estar trazados en tinta china sobre fondo blanco. Conviene dejar en lápiz las letras y leyendas para ponerlas á escala. De los conceptos emitidos en todos los trabajos responden los autores. No se devuelven los originales. A nuestros suscriptores. Toda consulta de asuntos profesionales será resuelta gratuitamente, remitiendo un sello de 0,30 pesetas, para el franqueo de la correspondencia. G l u c ó s i d o s c i a n o g e n é t i c o s y fermentos que catalizan su hidrólisis por Florencio B u s t i n z a Catedrático de Agricultura. Farmacéutico. El ácido cianhídrico ó ácido prúsico (CNH) es un líquido cuyo punto de ebullición es alrededor de los 26°, ó sea que emite fácilmente vapores que son de extraordinaria toxicidad. Hay plantas que desprenden C N H en determinadas fases de su desarrollo, y son mu«has las especies que contienen glucósidos cianogenéticos, los cuales por hidrólisis con los ácidos diluidos ó por la acción en presencia del agua, de ciertos fermentos solubles desprenden ácido cianhídrico. Los vapores de ácido cianhídrico pueden caracterizarse utilizando el Reactivo de Guignard, que se prepara sumergiendo tiritas rectangulares de papel de filtro en una disolución de ácido pícrico al 1 por 100; cuando están bien empapadas de dicho ácido se desecan al abrigo de la luz entre papeles de filtro y ya secas se sumergen en una disolución de carbonato sódico al 10 por 100, para sacarlas al poco rato y desecarlas también al abrigo de la luz entre papel de filtro. E l C N H da una coloración roja m á s ó menos i n tensa al papel de Guignard por formarse la sal sódica del ácido isopurpúrico (1). El glucósido cianogenético más interesante y mejor estudiado es la amigdalina, aislada en 1830 por ROBIQUET y BOUTRON, á partir de las almendras amargas. La amigdalina es un glucósido de un disacárido (que según los trabajos de HAWORTH y HuDSON^en la genciobiosa) y por la ac(1) El CNH puede también diagnosticarse mediante el papel de sulfato de cobre y de guayaco, para cuya preparación se sumergen tiritas de papel de filtro en una disolución de sulfato de cobre al 1 por 1.000 y luego se desecan y en el momento de emplearlas se tocan en determinados puntos con una disolución de tintura alcohólica de resina de guayaco recientemente preparada. Las zonas de papel, impregnadas de la tintura de guyaco, se colorean de azul intenso en presencia de indicios de CNH. Hay que tener presente que esta reacción no es especifica del CNH, pues la dan también el ozono, el cloro, los vapores de bromo ó de iodo, los vapores nitrosos y los de amoniaco. 592 v. BÜSTINZA.-GLUCÓSIDOS OIANOGBNÉTICOS Y FERMENTOS ción de los ácidos diluidos se hidroliza engendrando una molécula de aldehido benzoico, una de C N H , y dos moléculas de glucosa. COH C6H5-GH \)(Cl2H21O10) Amisfdalina +2HoO = 4-CNHH-2CG Hjo 06 Ald. benzoico En el año 1837 LIEBIG y WOHLER observaron que la amigdalina es hidrolizada en presencia del agua, por un fermento soluble, la emulsina contenido en las almendras amargas y muy especialmente en las almendras dulces. La vidanina es también un glucósido cianogenético extraído en 1906 por BERTRAND (1) de las semillas de la Vicia angustifolia y se diferencia de la amignalina en que la biosa que integra su molécula es la vicianosa. La vicianina por la acción de un fermento soluble presente en la Vicia angustifolia y en presencia del agua se desdobla: en una molécula de aldehido benzóico, una de C N H , y una de vicianosa. COH C0 H5 — C H \ 0 (CH H18 Og) Vicianina - f H2 O = + CNH + C n H 0O ni ^ Esta vicianosa por la acción de la emulsina se desdobla en una molécula de glucosa y en otra de 1-arabinosa. Cu H20 O]0 - f Ha O = CG H12 06 + C5 H10 0'5 Vicianosa Avabinosa Hay glucósidos cianogenéticos que dan por hidrólisis una sola molécula de glucosa, además de la molécula de aldehido benzóico y del C N H , y son: la prunasina que se extrae de la corteza del Prunus Padus, la prulaurasina aislada por HÉRISSEY en 1906 de las hojas del laurel-cerezo y la sambunigrina descubierta por BOURQUELOT y DANJOU en 1906 en las hojas de Sambucus nigra (2). Estos tres glucósidos son estereoisómeros y se derivan del nitrilo del ácido mandélico, ó ácido fenilglicólico, y como hay tres ácidos fenilglicólicos, levógiro uno, dextrogiro otro, é inactivo el tercero, resulta (1) G. BERTRAND «La vicianine, nouveau glucoside cyanhydriquc contenu dans les graines des Vesces», C B. Ac. des Sciences de París, 1906, 143, p. 832. (2) E. HOURQUELOT et DANJOU «Préparation du g-lucoside cyanbydrique du sureau á l'état cristaliisé». Jour. de Pharm. et de Chim. 1905, 22, 219. F, BUSTINZA. —GLUCOSIDOS CIANOGENÉTICOS Y FERMENTOS 593 que la. prunasina se deriva del ácido levógiro, la sambunigrina del dextrogiro y la prulaurasina del ácido racémico. Los tres glucósidos dan por hidrólisis: una molécula de aldehido benzoico, una de C N H y una de glucosa. CgHg — CH = Ac. mandélico ó fenilg'licólico ^COOH COH C6H5-CH +H20 = + CNH + C(!H1206 X 0 (C6 H n ü5) Glucósidos del nitrilo mandélico Hay glucósidos cianogenéticos que no producen en su hidrólisis aldehido benzoico y son: la clurrina, la gynocardina, la Unamarina, la faseolunaüna, la manihotoxina, la lotusina y la Mptagina. La durrina fué descubierta por DUNSTAN y HENRY en 1902 (1) en las plántulas del Sorghum vulgare y por la acción de los ácidos diluidos ó de la emulsina se hidroliza engendrando: una molécula de p-oxibenzoaldehido, una de C N H y una de glucosa. COH CH +H20 \ ) (C6Hn Og) OH Durrina + CNH + C6HJ2O0 OH p. oxibenzoaldehido La gynocardina fué descubierta por POWER y GORNALL en 1904 (2) en las semillas de la chaulmoogra o Taraktogenos Kurzü, de la familia de las bixáceas, planta que vive en los bosques de Birmania (3). L a gynocardina por la acción de un fermento contenido en dichas semillas, la Gynocardasa, (1) W. R. DUNSTAN and Th. A. HENRY «Cyanogenesis in plants». Part. I I . The great millet, «Sorghum vulgare», Philos. Trans. Roy. Soc. of London. Serie A. 1902, 199, 399. (2) POWER and LEES «Gynocardin, á new Cyanog-enetic glucoside». Journal Chem. Soc. of London. 1905, 87, 349. (3) De las semillas de chaulmoogra se extrae un aceite utilizado en el tratamiento de la lepra. Hoy se recomienda para este fin, mejor que el aceite, las sales sódicas ó los esteres etílicos de los ácidos grasos cíclicos, gynocárdico y chaulmoógrico, contenidos en dicho aceite. 594 F. BUSTINZA. —GLUCÓSIDOS CIANOGBNÉTICOS Y FERMENTOS en presencia del agua se hidroliza y engendra: C N H , glucosa y un cuerpo no bien determinado. C5H4(OH)3-CH N ) (CeH,, 05) Gynocardina " C5H5(OH)3 = C +H20 ^OCCeHn 05) - Ó CNH + C6Hl2 06 + C6H804 El cuerpo C6 H8 O4 que se engendra en la hidrólisis de la gynocardina, puede ser un trioxialdehido de la fórmula C5 H4 (OH)3 COH ó una trioxicetona de la fórmula C5 Hg (OH)3 CO. La Unamarina descubierta en 1887 por JORISSEN y HAIRS en las plántulas del Linum usitatissimum, por la acción de un fermento llamado linasa, se hidroliza en: una molécula de CNH, otra de acetona y otra de glucosa. (CH3)2- C +H20 ^ 0 (CeH^Os) Linamarina = CNH + C6H12 06 + C H 3 - C O ~ C H 3 La faseolunatina contenida en las semillas del Fhaseolus lunatus (1) y la Manihotoxina de los rizomas de la tapioca amarga, son glucósidos iguales que la linamarina y, por lo tanto, dan también por hidrólisis: C N H , glucosa y acetona. DUNSTAN y HENRY en 1900 (2) descubrieron en el Lotus arábicus un glucósido cianogenético la Lotusina el cual por hidrólisis proporciona: dos moléculas de glucosa, una de C N H y un pigmento amarillo del grupo de las flavonas llamado la Lotoflavina. GORTER, en el año 1920, ha descubierto otro glucósido cianogenético la Hiptagina (3) en la corteza de la raíz del Hiptage madablota de la familia malpigiáceas. Se trata de un glucósido muy complejo que por la acción de los álcalis da C N H y otros cuerpos. Conocidos ya los glucósidos cianogenéticos, pasemos al estudio de su hidrólisis por la acción de fermentos solubles específicos. (1) W. R. DUNSTAN and TH. A. HENRY «Cyanogenesis in plants». Part. I I I . On phaseolunatin, the cyanogenetic glucoside of Phaseolus lunatus. Proc. Roy. Soc. of London. 1903, 72, 285. (2) W. R. DUNSTAN and Th. A. HENRY. «The nature and origin of the poison of Lotus arabicus», Phüosoph. Trans. Roy. Soc. of Londcn. Serie B, 1900, 194, 515. (3) K. GORTER. «L'hiptagine», Bull. Jard. Bot. de Buitenzorg. 1920. 595 F. BUSTINZA.—GLUCOSIDOS CIANOGHNBTICOS Y FERMENTOS Si machacamos almendras amargas en presencia del agua, percibiremos un olor especial debido á la formación de aldehido benzoico y de ácido cianhídrico. Si la pasta obtenida la colocamos en un pequeño Erlenmeyer que cerraremos con un tapón bien limpio y del cual cuelga un papel de picrato sódico (Erlenmeyer A de la figura 1), veremos que éste se pondrá rojo al cabo de pocos minutos, signo revelador del desprendimiento de C N H . Pape/ </e f/crafo Soc/ico A//n amarocis con HXQ Amijdai/n¿> A//77. Ju/ces ton H z O con H t 0 Amij-\-alm. dulces^0 £rnu/. ck ¿//n. c/u/ces ¿erv/eü fígua Figura 1.—El picrato sódico se pondrá rojo en A y en D, y permanecerá amarillo en B, C, E y F. Tanto el aldehido benzoico como el C N H no existen preformados en las almendras amargas, aparecen al ser éstas trituradas y proceden del desdoblamiento de la amigdalina en ellas contenidas. Ahora bien, para que tenga lugar la hidrólisis de este glucósido es indispensable el concurso del agua, pero ello no es suficiente, pues si colocamos en el matraz B figura 1 amigdalina (1) con agua, por ejemplo, 0,1 gr. de amigdalina y 26 c. c. de agua, veremos que no hay hidrólisis, pues el papel de picrato sódico permanece amarillo. Para que el agua hidrolice á la amigdalina, es indispensable el concurso, la cooperación de un fermento soluble que actúe de vector del agua y permita que ésta hidrolice al glucósido. Este fermento, llamado eraulsina, se encuentra en las almendras amargas localizado en células diferentes de las en que está localizada la amigdalina (2) y por eso la almendra amarga entera no huele á aldehido benzóico ni desprende C N H , pero al triturarla y ponerse en contacto en presencia del agua las células (1) Un método sencillo para obtener amigdalina es el siguiente: Se toman 12 almendrasamargas, se las sumerge en agua hirviendo, al cabo de 10 ó 15 minutos se las priva de su tegumento, y se reducen á pasta en un mortero; se introduce ésta en un Erlenmeyer, se agregan 50 c. c. de etanol y se calienta sobre baño maría. El extracto alcohólico se evapora á sequedad y el producto obtenido contiene amigdalina impura. (2) La amigdalina está localizada en las almendras amargas en el parénquima cotiledonar y la emulsina, tanto en las amargas como en las almendras dulces, en el periciclo de la raicilla y del tallito y también en el periciclo y en el endodermo de los hacecillos cotiledonares. 596 F. BUSTINZA.- GLUCÓSIDOS CIANOGBNÉT1C0S Y FERMENTOS que contienen la emulsina y las que contienen la amigdalina, entonces este glucósido se hidroliza y se desdobla en una molécula de aldehido benzóico, una G N H y dos de glucosa, con arreglo á la ecuación que en otro lugar hemos formulado. Esta reaccióu nos explica los casos de envenenamiento observados, especialmente en niños, á consecuencia de haber comido unas pocas almendras amargas (6 ó 6 bastan). A l ser trituradas en la boca, la emulsina conduce al agua de la saliva sobre la amigdalina y se produce el C N H ; que si bien como ácido es débil y no tiene acción corrosiva sobre las mucosas, sin embargo, al penetrar en el torrente circulatorio, ejerce una acción tóxica considerable. La emulsina es un fermento muy difundido en el reino vegetal, habiendo sido diagnosticado en numerosas plantas, tanto criptógamas como fanerógamas, y abundando muy especialmente en las almendras dulces. Antes de diagnosticar la emulsina en las almendras dulces, vamos á demostrar que en ellas no hay amigdalina, y para ello trituraremos un par de almendras dulces (desprovistas de su tegumento) con un poco de agua, y colocaremos la pasta ó la papilla así obtenida en el Erlenmeyer C de la figura 1, el cual lo taponaremos después de colocar en el borde de su boca un papel de picrato sódico, y veremos que éste, ni aun después de muchas horas, cambia de color, prueba de que no hay desprendimiento de C N H , ó sea que en las almendras dulces no hay amigdalina. La presencia de la emulsiua en las almendras dulces la demostraremos sin m á s que repetir la experiencia anterior, pero triturando las almendras dulces con agua en la que habremos disuelto previamente un poco de amigdalina (por ejemplo, 0,1 gr. del glucósido en 25 c. c. de agua). (Erlenmeyer D de la figura 1) y veremos que al cabo de pocos minutos el papel de picrato está rojo, prueba de que la amigdalina se ha hidrolizado por un fermento soluble, por la emulsina. Vamos ahora á demostrar que la substancia contenida en las almendras dulces que cataliza la hidrólisis de la amigdalina es un fermento. Para ello trituraremos una ó dos almendras con agua hasta formar una papilla que trasladaremos á un Erlenmeyer que sumergiremos durante unos minutos (5 á 10) en un baño de agua hirviente con objeto de destruir al agente capaz de hidrolizar á la amigdalina, se retira el Erlenmeyer del baño de agua y se agrega á la emulsión de almendras así tratada unos c. c. de una disolución de a m i g d a l i n a (por e j e m p l o , 0,1 gr. de glucósido en 10 c. c. de agua), se coloca en la boca del matracito el papel de picrato y se tapona. ¿Qué observaremos? Pues sencillamente que el papel no acusará desprendimiento de C N H , y sacaremos, por lo tanto, de esta experiencia la consecuencia de que la substancia contenida en las almendras dulces y que es capaz de provocar en presencia del agua la hidrólisis de la amigdalina, es una substancia del tipo de los fermentos solubles, pues es F. BUSTINZA. GLUCÓSIDOS CIANOGBNÉTICOS Y FERMENTOS 597 muy frágil y se desnaturaliza y pierde totalmente su actividad por la acción del calor de baño de agua hirviente. Las almendras dulces constituyen un excelente material para la preparación de la emulsina (1). Veamos cómo debemos proceder para obtener este interesante fermento. Tomemos 100 gr. de almendras dulces que sumergiremos durante medio minuto en agua hirviente para facilitar la separación de los tegumentos; practicada esta operación, se reducen en un mortero las almendras á pasta que colocaremos en un Erlenmeyer con 200 c. c. de éter sulfúrico para extraer la mayor cantidad posible de grasa, tapemos el matraz y agitémosle de vez en cuando, y al cabo de un par de horas se decanta el extracto etéreo y se vuelve á agregar al Erlenmeyer nueva cantidad de éter, se tapona y se agita; á las dos horas se filtra, y el residuo no disuelto en el éter se extiende sobre papel de filtro y se deseca en un local donde no haya ningún mechero encendido ni ninguna llama, dada la facilidad con que se inflama el éter. El residuo, una vez seco, se trata con '200 c. c. de agua destilada saturada de cloroformo y se introduce en un matraz que se t a p o n a r á , y se deja en maceración durante 24 á 48 horas agitando de vez en cuando. Se filtra, y á la disolución acuosa obtenida se agregan 5 ó 6 c. c. de ácido acético al 10 por 100 para precipitar las proteínas, se agita, se espera á que el precipitado se forme bien, se decanta y se filtra sobre filtro mojado, se comprueba si el líquido filtrado precipita por adición de unas gotas de acético al 10 por 100, y si no precipita se le agrega tres ó cuatro volúmenes de alcohol etílico de 96° con objeto de cambiar el medio de dispersión y flocular á la emulsina, se agita, se deja en reposo, se decanta la mayor parte del líquido hidro-alcohólico, se centrifuga, y el precipitado se lava varias veces con alcohol, se recoge en un vidrio de reloj y se coloca en un desecador de vacío sobre sulfúrico concentrado. La emulsina perfectamente seca se conserva bien. Vamos á comprobar ahora que el producto obtenido es emulsina. Para ello dispersaremos 0,05 gr. del producto en 10 c. c. de agua y agreguemos 2 c. c. de esta disolución á 10 c. c. de una disolución de amigdalina al 2 por 100, contenida en un pequeño Erlenmeyer A, de cuya boca colgaremos un papel de picrato sódico, y tapémosle con un corcho que ajuste bien. En otro Erlenmeyer idéntico B, pongamos 10 c. c. de la disolución de amigdalina y 2 c. c. de la disolución del producto (previamente hervida durante un minuto), cerremos con el tapón de corcho después de colocado el papel de picrato sódico y metamos ahora ambos matracitos en una estufa á 45° (2) (1) Puede también prepararse la emulsina partiendo de las almendras amargas, pero la técnica á seguir en este caso es más complicada debido á la presencia de la amigdalina, (2) A 45° la emulsina hidroliza más rápidamente á la amigdalina que á la temperatura ordinaria. 598 F. BÜSTINZA. — GLUCÓSIDOS CIANOGENÉT1COS Y FERMENTOS y veremos al cabo de pocos minutos que el papel de picrato del matracito A, se pone rojo, y que el del B no cambia de coloración ni al cabo de muchas horas, lo cual nos demuestra que efectivamente el producto obtenido es un fermento soluble que hidroliza á la amigdalina y que pierde su actividad por ebullición, y como el único fermento que cataliza dicha hidrólisis es la emulsina, resulta que el producto obtenido contiene emulsina. En el matraz A se puede comprobar que se ha formado aldehido benzóico, por su olor particular y la glucosa, comprobando que el liquido tiene acción reductora sobre el licor de Fehling. Teniendo en cuenta el interesante trabajo del Dr. F. CHODAT (1) «sobre la importancia de los puntos isoeléctricos en la preparación y en la actividad de los fermentos», recomiendo (aunque no es absolutamente preciso), que la dispersión de la emulsina (y la trituración de los órganos vegetales en los cuales tratemos de investigar la emulsina), se efectúe con una disolución tampón; cuyo p H sea de 6,3, pues la concentración de hidrogeniones que corresponde á ese p H , es la óptima para la hidrólisis de la amigdalina por la emulsina. Utilizo como disolución tampón de p H = 5,3, la preparada mezclando 0,25 c. c. de una disolución de fosfato secundario y 9,75 c. c. de fosfato primario. L a disolución de fosfato secundario se prepara pesando con precisión 11,876 gr. de fosfato disódico Sorensen y disolviéndolo en agua destilada neutra en cantidad suficiente hasta obtener un litro de disolución, y la disolución de fosfato primario se obtiene pesando con precisión 9,078 gr. del fosfato monopotásico Sorensen y disolviéndolo en agua destilada neutra obtener un litro de disolución. Estas dos disoluciones se alteran con facilidad y conviene renovarlas cada mes. Para evitar esta alteración acostumbro á cubrir cada una de estas dos disoluciones con una capa de toluol. La emulsina es un fermento complejo que ha motivado numerosos trabajos, destacando los de ARMSTRONG y colaboradores, en Inglaterra, y los de BOUEQUELOT y colaboradores, en Francia. La emulsina, como ya hemos indicado, cataliza la hidrólisis de la amigdalina en dos moléculas de glucosa, una de C N f í y una de aldehido benzóico, y también cataliza la hidrólisis de los tres glucósidos del nitrilo mandélico, prunasina, prulaurasina y sambunigrina, los cuales se desdoblan por su acción en una molécula de glucosa, una de C N H y una de aldehido benzóico. El jugo de Helix pomatia, actuando sobre la amigdalina, separa una molécula de glucosa y la convierte en el glucósido del nitrilo mandélico, sobre el cual dicho jugo no tiene acción y CALDWELL demostró que el Saccharomyces Ludwigii, actuando s ó b r e l a amigdalina separa también una mo(1) C. B. Soc. de Fhysique et d'H. Naturelle de Genéoe. Yol. 44, nüm, 1, p. 35-40 (127). F. BU8TINZA. — GLUCÓSIDOS CIANOGBNÉTICOS Y FERMENTOS 599 lécula de glucosa transformándola en el glucósido del nitrilo mandélico. Estos hechos demuestran la existencia en el jugo de Helix pomatia y en el Sach. Ludwigii de un agente, un fermento soluble (porque por la acción de calor se inactiva), llamado amigdalinasa capaz de separar una molécula de glucosa de la maléenla de la amigdalina. De la corteza del cirolero se puede extraer un fermento, la prmasa, que no actúa sobre la amigdalina, pero si sobre las tres formas del glucósido del nitrilo mandélico. ARMSTRONG y colaboradores deducen (1) de todo esto que la emulsina está formada por esos dos fermentos: la amigdalinasa y la prunasa, actuando primero la amigdalinasa sobre la amigdalina de la qne separa una molécula de glucosa y luego la prunasa, actuando sobre el glucósido del nitrilo mandélico, lo desdobla en una molécula de glucosa, una de C N H y una de aldehido benzoico. BOURQUELOT y discípulos sostienen en cambio que no puede considerarse a la emulsina como formada por la unión de esos dos fermentos, sino que se trata de un fermento complejo. La emulsina actúa sobre gran número de glucósidos naturales, así, actuando sobre la salicina (2) en presencia del agua la desdobla en glucosa y saligenina. 0-C6Hu05 l -CHoOH CH.ÜH + CG H12 06 -f H , 0 Salicina Saligenina A 10 c. c. de una disolución al 1 por 100 de salicina agreguemos 2 c, c. de una disolución (le emulsina al 0,6 por 100, coloquemos el tubo en el que practicamos la experiencia en una estufa de 45°, y á la media hora comprobemos que la salicina se ha desdoblado, pues podremos diagnosticar en el líquido la saligenina por la coloración p u r p ú r e a que da con unas gotas de disolución de cloruro férrico y la glucosa por su acción reductora sobre el licor de Fehling. 4. (1) H . E. ARMSTRONG, E. F. ARMSTRONG and E. HORTON Studies on Enzyme Action, X I I . The enzymes of emulsin. Proc. Boy. Soc. Vol. 80,1908, p. 321-331. Los mismos autores Studies on Enzyme action, X V I , The enzymes of emulsin. Proc. Roy. Soc. Vol. 85,1912. I . Primase the correlate of prunasin, p. 359-352. I I . Distribution of -enzymes inplants, p. 363-369. (2) La salicina se encuentra en la corteza de diversas especies de Salix y Populus, y también en los botones florales de la Spirea TJlmaria. 600 F. BUSTINZA. — GLUCÓSIDOS CIANOGBNÉTICOS Y FERMENTOS La emulsina en presencia del agua hidroliza arreglo á la siguiente ecuación: 0 - C 6 H u 05 a la arhutina (1) con OH + H20 + C6 OH Arbutina 06 OH Hidroquinona Actuando sobre la coniferina (2) la desdobla en glucosa y en el alcohol coniferilico. C H = : O H ~ CH90H CH = CH - CH, OH + C0Hl2O(! + H2 O - OCH, O-CeH^Os Coniferina - OCH. OH Ale. Coniferilico Actuando sobre la siringina ó metoxiconiferina (3) la desdobla en glucosa y siringenina. CH = C H - C H ) O H CH = CH - CH90H CH,0 - - OCH, 0 ~ C Q R n 05 Siringina + H20 = CHgO + C6 H ^ 06 OCH, OH Siringenina Actuando sobre la esculina (glucósido derivado de la cumarina) la desdobla en glucosa y esculetina (4). (1) La arbutina es un glucósido que se encuentra en las hojas del Ardostaphylos uva-ursi, Pyrola, Vaccinium, Pyrus communis, etc. (2) La coniferina se halla en los tallos de abetos, de alerces y de gran número de coniferas, en los espárragos, en la raíz de Scorzonera hispánica, etc. (3) La siringina se encuentra en las hojas de Ligustrum, Jasminum y de diversas oleáceas. (4) La esculina se encuentra en la corteza del Castaño de las Indias y sus disoluciones tienen fluorescencia azulada, y para su compi-obación se cortan pequeños trozos de corteza joven de Aesculus hippocastanum, se toman unos 5 gr. y se hierven en una cápsula con P.fiUSTlNZA.—GLUCÓSIDOS ClANO&KNIíTICOS Y PBRMEKTOS CH=CH I -O - co -f H 2 0 = C6 Hu 05 - O CH-CH v I O - CO + C6 Hí2 O0 HO OH Esculina 601 OH Esculetina Actuando sobre la geina la transforma en d-glucosa, 1-arabinosa y eugenol (1). Vi ci añosa. O - C0 H10 0^ - O — C5 H9 04 OH - OCH, — OCHc + 2 H2 O = CH2 — CH = CH2 Geina + C6 H ^ Oc -f- C5 HJO O5 CH2 - CH = CH<! Euffenol BKIDEL ha descubierto que la emulsina actuando sobre la MonotropiHna (2), glucósido cuyo a z ú c a r es un disacárido, la primaverosa, y cuya aglucona es el éster metílico del ácido salicílico, la desdoblan en: salicilato de metilo, glucosa y xilosa. COOCH3 H10 O4 Glucosa —O — + 2H20 C5 H9 04 Xilosa Primaverosa - COOCH3 + C(iH12 O(i + C5H10 O5 - OH Salicilato de metilo Monotropina. 50 c. c. de agua destilada durante unos 15 minutos, se filtra y se vierte la disolución filtrada sobre gran cantidad de agua (uno á dos litóos) contenida en un recipiente de vidrio de gran altura y paredes planas. Observaremos una bella fluorescencia azulada que se intensifica por la adición de un álcali y que se debilita por la adición de un ácido. (1) La geina se encuentra en las raices del Geum urbanum, en las que existe un fermento llamado geasa que desdobla á la geina en eugenol y vicianosa, J. CHBYMOL. Die chemische zusammensetzung der Wurzel von Geum urbanum L . , Schivez. Apoth. Ztg. 1928, 66, 283. (2) La Mo7iotropitina es un glucósido descubierto por BRIDBL en 1923 en la Monotropa hypopitys y, posteriormente, en muchas otras plantas, Etude biochimique sur la composition du Monotropa hypopitys L . Obtentión d'un nouveau glucoside ¿1 salycilate de méthyle, la monotropine. C. B . Ac. Se. de París, 1923, 177, p. 642. 602 v. BUSTINÍÜA.—(tLUCÓSIDOS CIANOGKNKTICOS Y FERMENTOS Según BRIDEL (1) la emulsina también actúa sobre los dos glucósidos (isómeros) la primaverina y la primulaverina de la raíz de la Prímula officinalis, á los cuales desdobla en presencia del agua en una molécula de COOCH, COOCH, O - C o H10 0 4 - 0 - C 5 H 9 04 O — Cfi Hin Od — O — CR H0 04 CH,0 OCH3 Primaverina Primulaverina glucosa, otra de xilosa y una molécula del derivado metoxílico del salicilato de metilo. La emulsina actúa también sobre otros muchos glucósidos naturales, y ¿por qué?, pues, sencillamente, porque la emulsina es una ¡S.-glucosidasa y , por lo tanto, es un fermento especifico de todos los ¡S-glucósidos (2). BOUEQÜELOT utiliza el fenómeno de hidrólisis de los glucósidos por la emulsina para investigar dichas sustancias en las plantas. Su método (3) se basa en el hecho de que los glucósidos hidrolizados por la emulsina son levógiros y que, en su desdoblamiento, se produce glucosa que desvia el plano de polarización hacia la derecha. T é c n i c a de Bourquelot para l a i n v e s t i g a c i ó n de g l u c ó s i d o s : Se toman 200 gramos del órgano fresco, en el cual se trata de investigar la existencia de un glucósido; se corta en fragmentos que se vierten inmediatamente sobre alcohol en ebullición con objeto de destruir rápidamente todos los fermentos que pueden haber en dicho órgano. L a disolución alcohólica se destila bajo presión reducida, y el residuo obtenido se disuelve en agua que contenga un poco de timol (como antiséptico), y se completa á 200 c. c , se determina el poder rotatorio de esta disolución, luego se agrega la emulsina, se deja que ésta actúe á la temperatura ordinaria y al cabo de algunos días se procede á determinar nuevamente el poder rotatorio del líquido, y si observamos desviación hacia la derecha, es evidente que la emulsina ha hidrolizado á un glucósido. Esta técnica de BOURQUELOT ha sido muy fecunda y ha permitido á sus discípulos, entre los que destaca BRIDEL, el descubrimiento, en diferentes plantas, de gran número de glucósidos. (1) M. BRIDEL. Le primevérose, les primevérosides et la primevérosidase. C. R. Ac. Se. 1925, 180, p. 1421, Sur la présence dans l'émulsine des amandes de deux nouveaux ferments, la primevérosidase et la pimevérase. C. B . Ac. Se. 1925, 181, p. 523. (2) Precisamente por ser una p-glucosidasa, la emulsina cataliza la hidrólisis de la lactosa que es un ^-glucósido (glucosa-galactosa). (3) Les principes actifs de quelques plantes employées en médecine populaire; leur recherche par la méthode biochimique, Bull. de la Soc. de Chim. Biol, 1921, 3, p. 71, V, BÜSTtN^A. —GLUCÓSIDOS CtANOGBN¿TICOS Y ÍBRMíJNTOg 603 A partir de 1912, BOURQUELOT y discípulos han efectuado una serie de trabajos de alto valor científico acerca de la reversibilidad de la acción de la emulsina. Primero, demostraron que este fermento puede ejercer su acción hidrolítica sobre los glucósidos aun en disolución alcohólica de elevada concentración alcohólica, y después observaron que el fermento era capaz de catalizar la unión de la glucosa con el alcohol empleado, engendrando el ¡3-glucósido correspondiente. Son numerosos los glucósidos que, desde esa fecha hasta el momento actual, se han obtenido por síntesis bioquímica, utilizando la emulsina como catalizador (1). De todo lo que llevo expuesto se deduce claramente que la emulsina es un fermento de xetraordinario interés científico. Su caracterización en las plantas se efectúa con la misma sencillez con que la hemos diagnosticado en las almendras dulces (matraz D de la fig. 1), solamente recomiendo, para el mejor éxito en el diagnóstico, dispersar el órgano (en que tratamos de averiguar la existencia de este fermento) en una disolución tampón de p H = 5,3 á base de fosfatos y también colocar el Erlenmeyer en una estufa á 46°. Respecto á la existencia de la emulsina entre los fermentos del tubo digestivo, resulta que las célulos del intestino contienen este fermento; en cambio, está demostrada su ausencia en la sangre, y por esto la inyección intravenosa de amigdalina se soporta bien, y , en cambio, la ingestión de dicho glucósido provoca accidentes mortales. Del ilustre Prof. ROGER (2) tomo los siguientes datos: «4 conejos á los que se ha inyectado 1 gr. de amigdalina en 3 ó 4 inyecciones intravenosas han sobrevivido». «27 conejos han recibido 0,26 gr. á 1 gr. de amigdalina en inyección intraperitoneal; de ellos, en 11 la primera inyección ha sido mortal^ en 5 ha sido mortal la segunda inyección, en 5 ha sido mortal la tercera, en 1 la cuarta inyección y en 3 la quinta, y solamente 2 han sobrevivido después de haber recibido uno de ello 10 inyecciones y el otro 13 de 0,26 gramos de glucósido, o sea que existe una sensibilidad y una resistencia particular que no está ligada á las dosis inyectadas». Desde luego, los conejos fallecidos en las experiencias de ROGER mueren por intoxicación típicamente cianhídrica, lo que prueba que el glucósido ha sido hidrolizado por la emulsina que ha salido del intestino á la cavidad peritoneal. (1) E. BOURQUELOT et M. BRIDBL.—Z)e V action hydrolisante et de V action synthétisante de V émulsine dans V alcool méthylique. Obtention du méthylglucoside. J. de Pharmacie et de Chimie. 6, 56-62. (1912). E. BOURQUELOT et E. VERDÓN.—La réversibüité des actions fermentaires. Emulsine et méthylglucoside. J. de Ph. et de Gh„ 1913. 7, 377-383. (2) G. H. ROGER, Questions actuelles de Biologie médicale, Ma&son et Cié., París, 1924, Págs. 158-159. 604 e. BÜST1N55A. —GLUCÓSIDOS ClAÍSTOGBNÉTICOg Y FERMENTOS Los glucósidos cianogenéticos se hallan localizados en células diferentes de las en que están localizados los fermentos capaces de transformarlos. En 1890, GuiGNARD demostró (1) que en el laurel-cerezo la prulaurasina existe en pequeña proporción en las células del parénquima foliar y la emulsina en la vaina endodérmica que rodea á los hacecillos liberoleñosos y en algunas células no escleriflcadas del periciclo. Coloquemos en un tubo de ensayo (a, flg. 2) una hoja de laurel-cerezo entera, y en & otra hoja previamente machacada en un mortero, pongamos ahora en la boca de cada tuvo un papel de picrato sódico y cerremos ambos con un tapón de corcho bien limpio. A l cabo de pocos minutos veré- Fg. 2.—a, hoja entera; b, hoja machacada de laurel-cerezo; R. G., reactivo Guignard. mos que el papel del tubo h está rojo, y en cambio el a está amarillo, y permanece asi durante muchas horas y días. Estas experiencias nos demuestran que en el laurel-cerezo hay un glucósido cianogenético y un fermento capaz de hidrolizarlo, pero ambos cuerpos están localizados en células diferentes, y únicamente cuando se ponen en contacto al machacar la hoja, es cuando se acusa la hidrólisis de dicho glucósido por el enrojecimiento del picrato sódico, que nos revela el desprendimiento de C N H . MARCEL MIRANDE presentó á la Academia de Ciencias de París (2) un método para diagnosticar las plantas cianogenéticas. Se coloca, como indica la figura 3, un poco de cloroformo en un tubo de Roux de los empleados en Bacteriología, y encima, y sostenida por el estrangulamiento del tubo, coloquemos una hoja joven, entera y fresca del laurel-cerezo, se cuelga del borde un papel de picrato, se tapa con un corcho limpio y que ajuste bien y se incuba la experiencia á 37°. Veremos al cabo de unos pocos mi(1) L . GUGNARD: Sur la localisation de principes qui fournissant de V acide cyanliydrique. Jourrual de Botauique, 1890. (2) M. MIRANDE: Influence exercée par certaines vapeurs sur la cyanogénese végétale. Procédé rapide pour la recherclie des plantes á acide cyanhydrique. C. R. Ac. des Se. de París. 1909, 149, p. 140-142. f.feÜSTINZA.-—CILÚCOSiDOSCÍANOGBNÉTlCOS Y FBRMElNTOS 605 ñutos (3 á 10) que el papel amarillo se va enrojeciendo, señal evidente del desprendimiento de C N H . ¿Cómo ha actuado el cloroformo? Pues sencillamentCj los vapores de cloroformo han anestesiado á las hojas, y si las examinamos al microscopio, veremos que sus células han experimentado la plasmolisis, estando en ellas el saco protoplásmico m á s ó menos separado de la membrana celu- Quijnorcf //o/a cíe /cn/re/-cerezo H,Oa5J CCUH Fig. 3 lósica, debido á que han perdido parte del jugo celular, saliendo así de ciertas células agua con prulaurasina y de otras agua con emulsina, y al ponerse en contacto ambas substancias, el glucósido se hidroliza y se produce C N H , que se desprende de la hoja y asciende á la pared superior del tubo, enrojeciendo al picrato sódico (1). Esta técnica sencilla y elegante puede aplicarse para el diagnóstico de glucósidos cianogenéticos en los órganos frescos vegetales, especialmente en las hojas, pudiéndose practicar la incubación en un bolsillo del chaleco ó del pantalón. Entre las muchas plantas que pueden utilizarse para practicar la experiencia anterior, recomiendo, además de las hojas de laurel-cerezo, las de Photinia serrulata, Samiucus nigra, Aquilegia vulgaris, Arum maculatum, Crataegus oxyacaniha, Lotus corniculatus, Ehamnus frángula, Ribes nigrum, Ribes i'ubrum, Mhes grossularia y diversas Vicias. También pueden diagnosticarse las plantas cianogenéticas sometiéndolas á bajas temperaturas (2) y disponiéndola experiencia como indica la figura 4. (1) Las intoxicaciones por el laurel-cerezo han sido frecuentes, ya sea por el empleo del agua de laurel-cerezo con elevada proporción de CNH ó por el uso culinario, siendo wuy sensibles los niños y habiéndose registrado numerosos casos de intoxicación por esta planta en el ganado. (2) L . GUIGNARD: Infiuence de l'anestJiésie et du gel sur le dédouhlement de certains glucosides ches les plantes. C. R. Ac. Se. de París. 1909, 149, p. 91-93. 606 F. BÜSTÍNÍÍA.—GLUCÓSIDOS OIANOGBNÉTICOS Y FERMENTOS Se rodea el tubo que contiene la hoja que se desea ensayar (en el case de la figura, es el laurel-cerezo) con la mezcla criógena de hielo y sal, so ffojo ofe cerezo Fig. 4. cuelga de la parte superior del tubo el papel de picrato sódico y se tapa bien con un corcho limpio. Las células se defienden del descenso de temperatura á que se les somete concentrando su jugo celular, y para ello dejan salir de su interior á los espacios intercelulares, agua, pero al mismo tiempo que sale el agua, sale también de ciertas células algo del glucósido cianogenético y de otras aguas y emulsina, y al ponerse en contacto se hidrolizael glucósido y se desprende el G N H , que será puesto en evidencia por el enrojecimiento del reactivo de Guignard, Hemos visto cómo la emulsina cataliza la hidrólisis de gran número de glucósidos, pero no es capaz de hidrolizar á todos, pues no ejerce acción sobre los glucósidos de la digital, sobre los glucósidos antraquinónicos ni sobre los dépsidos, ni tampoco es activa sobre ciertos glucósidos cianogenéticos tales como: la lotusina y la Unamarina ó faseolunatina. DUNSTAN y HENRY descubrieron en el Lotus arábicus la lotusina, glucósido que se halla también en el Lotus corniculatus en proporción variable, faltando en el Lotus corniculatus var. majo?* que para algunos botánicos es una especie diferente. La lotusina es un glucósido derivado de la maltosa y de la 5, 7, 2', 4', tetraoxiflavona y por la acción del fermento lotasa, presente en todos aquellos órganos donde se halla dicho glucósido, se desdobla en una molécula de maltosa, otra de CNEL y una de un pigmento amarillo, la lotoflavina. Núcleo de la ñavona F. BUSTINZA. —GLUCÓSIDOS CIANOGBNÉTICOS Y FORMBNTOS 607 OH - CH - O CN HO0 Lotusina OH OH H22 On + CNH + Lotoflavina La lotusina por hidrólisis con los ácidos diluidos engendra una molécula de C N H , una de lotoflavina y dos de glucosa. La linamarina y la faseolunatina, son un mismo glucósido y se hallan en las plántulas y en las semillas del lino y en las semillas del Phaseolus lunatus. La emulsina, no ejerce acción sobre este glucósido que solamente es hidrolizado por el fermento llamado linasa ó faseolunatasa (1). La presencia de linamarina en las semillas de lino, explica la toxicidad de ciertas tortas de lino, las cuales, en general, contienen pequeña cantidad de glucósido, pero éste puede alcanzar tan elevada dosis que represente de 40 á 45 centígrados de O N H por 100 gr. de torta, habiendo dado cuenta en 1927 el Prof. Moussu de Alfort (2) de accidentes mortales en el ganado á consecuencia de la ingestión de tortas de lino, ricas en C N H . La presencia de linamarina y de la linasa en la torta de lino, se pone en evidencia de la siguiente forma: se hace una pasta con una pequeña cantidad de torta y agua tibia á 350-380 que se colocará en un pequeño Erlenmeyer (fig. 6), se cuelga el papel de picrato, se tapa con un corcho que ajuste bien y se coloca en la estufa á 38° ó 40°. Si el papel de picrato no cambia de color, la torta no es peligrosa, pues ello indica que no se ha desprendido C N H , y si hay desprendimiento de este cuerpo, el papel se pondrá m á s ó menos rojo y con más ó menos rapidez, según la proporción de gluFig. 5.—A, torta de lino con agua. cósido presente en la torta. He aquí, pues, un procedi(1) H . E. ARMSTRONG and. J. VARGAS EYRB Studies on Enzyme action. X V I I I . Enzymes of the emulsin type. I I I . Linase and other enzymes i n Linaceae. Proceedings, 1912, 85 p. 370-378. (2) HUGUIRR: Í e s intoxications alimentaires des animaux de la feime. La vie Agricole et rurale, 26 de Agosto de 1928, p, 138. 608 F, BUSTINZA.—GLUCÓSIDOS CIANOGBNBTICOS Y FERMENTOS miento sencillo que permitirá separar de la alimentación del ganado un producto tóxico y que e v i t a r á pérdidas irreparables al ganadero. A las tortas tóxicas se les puede hacer perder su toxicidad calentándolas á 100°, temperatura á la cual el fermento hidrolizante de la linamarina se destruye, y ya libre de linasa, la torta no es peligrosa, porque la linamarina que pueda contener no es hidrolizada por la emulsina de la mucosa intestinal. La faseolunatina, cuerpo idéntico á la linamarina, se halla en el Phaseolus lunatus L . = Fhaseolus Capensis Thoub (habichuela asesina, habichuela de Lima, habichuela de Java), en una proporción que puede oscilar de 0.003 a 3,6 por 1.000 aunque frecuentemente no pasa de 0.002 por 1.000, y existiendo algunas variedades blancas carentes de dicho glucósido. En la excelente obra de BECKER-DILLINGEN: «Handbuch der Hubsenfruchterbaues and Futterbaues», 1929, p á g . 238, se indica que una proporción de C N H por bajo de 2 por 1.000 en estas semillas no las hace tóxicas, porque la mayor parte del C N H se desprende al cocer las habichuelas y, lo que si debe prohibirse terminantemente, es el utilizar las aguas de cocción en las cuales puede haber C N H . Hace algunos años llegó á Galicia, procedente de Amsterdam, un cargamento de habichuelas para pienso del ganado, y se dieron casos de intoxicación que motivaron el envío de muestras al Instituto Nacional de Higiene, de Madrid, donde diagnosticaron que dichas habichuelas eran cianogenéticas. Una muestra de estas habichuelas nos fué facilitada por el Dr. GARMENDÍA, Jefe de la Sección de análisis de alimentos de dicho Instituto, y las he utilizado con éxito en las diferentes ocasiones que he efectuado esta práctica ante mis alumnos. Combate biológico de los insectos dañinos a) A l lado del combate con medios químicos de los animales perjudiciales para las plantas, tiene gran importancia otro método llamado biológico. Los primeros y mayores resultados fueron alcanzados con estos métodos en los Estados Unidos y su dominio colonial (California, Hawai, Filipinas), donde los parásitos producían los mayores estragos en las grandes explotaciones agronómicas y en las plantaciones, pero donde también se reunían las mejores condiciones para la eficacia del método. Después de los Estados Unidos, están muy interesados en el combate biológico el Imperio británico, Francia, Holanda y otros con sus grandes posesiones coloniales. Pero también se pueden anotar hermosos resultados en algunos países europeos, especialmente en los países mediterráneos. Menor importancia tiene el método, por el momento, para Alemania; aunque en la Biologischen Reichsanstalt se prosigue atentamente su desarrollo, fomentado por investigaciones y experiencias, porque debe sostenerse siempre que Alemania debe obtener otra vez colonias entiempo no lejano. En los Estados Unidos se encuentra la organización central en el Burean of Entomology, de Washington, y en Inglaterra en el Imperial Burean of Entomology (Farriham House Laboratory). Las ideas directivas y el m é t o d o de combate biológico.—Su eficacia y límites han sido expuestos detenidamente por diferentes autores ingleses y americanos. Un extenso resumen da TEAPPMAN (B. R. A.) en su libro, y MORSTATT (B. R. A.) una relación de los resultados en las islas de Hawai, el país clásico de los métodos biológicos (B. R. A.) A continuación indicamos la bibliografía sobre esta materia (2): W. R. THOMPSON.-The principies of Biological control. Annals ofapplied Biology, 1930, 17, p. 306. W . R. THOMPSON.---The Biological control of insect and plant pests. A report on the arganisation and progrese of thé toork of Farriham Hause Laboratory, 1930. H . S, SMITH.—On sorae phases of insect control by the Biological method. Journal of economical Entomology, t. 12, p. 288. (1) Del Chemiker-Zeitung, 1935, 59, p. 724. (2) N . de la R. Una magnifica recopilación de todo lo publicado encontrarán los lectores españoles en la obra de R. GARCÍA MBRCBT «LOS parásitos de los insectos perjudiciales». 610 COMBATE BIOLÓGICO DE LOS INSECTOS DAÑINOS J. G. MYERS.—The principlesof Biological control. Tropical Agriculture, 1929, 6, p. 163. MORSTATT. —Die Erfolge der biol. Bekampfüng in E.&w&n.Anzeiger f. Schadlingskunde, 1927, 3, p. 46. TRAPPMANN.—Schadlingsbekampfung. 68 ff. ¿Qué se debe comprender ahora por combate biológico de los parásitos? Un autor inglés, MYERS, dice: «En cierto sentido, es biológico todo método de combate de una plaga de parásitos, con medios diferentes de los directamente físicos ó químicos; tal es, por ejemplo, también, el muy prometiente cultivo de variedades de plantas inmunes». Pero, en seguida, añade lo siguiente: «Se debe restringir el concepto al empleo de alguna clase de organismos para la limitación ó destrucción de otra especie». L a multitud de posibilidades que aún contiene esta respuesta restringida, se muestran en el esquema siguiente: I Combate de los animales dañinos. A) Por otros animales. 1. Nematodes por nematodes rapaces. 2. Moluscos por vertebrados. 3. Insectos y otros artrópodos por ácaros, otros insectos, pájaros y otros vertebrados. 4. Vertebrados por otros vertebrados. B) Por organismos vegetales. l i Insectos y otros artrópodos por bacterias, hongos parásitos, algas y fanerógamas. 2. Combate de plantas dañinas por insectos, ácaros, hongos parásitos y bacterias. De todas estas posibilidades, en algunos se trata solamente de proposiciones, de modo que hasta ahora solamente tiene importancia práctica y real el combate de insectos y otros artrópodos por insectos. Esto es el combate biológico de los parásitos en el sentido estricto, y en este dominio han sido alcanzados sorprendentes resultados con el empleo de principios biológicos. Todos los otros métodos están aún en estado de experimentación ó no se les concede valor alguno. Con la importación de pájaros y mamíferos para combatir las plagas de parásitos, se han hecho tan malas experiencias, que en general se debiera prescindir de ellas. En cuanto se refiere á las enfermedades producidas por bacterias y hongos, es ciertamente conocido, cómo obran en la Naturaleza sobre ciertos insectos parásitos, exterminándolos en determinadas condiciones, y se han descrito también los grandes resultados obtenidos con su cría artificial. Pero estos resultados no han podido ser confirmados en experimentos posteriores, y las perspectivas del método son juzgadas muy escépticas por autoridades reconocidas. Así, B. PAILLOT escribe (1916): COMBATE BIOLÓGICO DE LOS IKSBCTOS DAÑINOS 611 «La producción de epidemias artificiales, que sean comparables en intensidad y extensión á las naturales, parece casi imposible en el estado actual de nuestros conocimientos». Y PETOH llega á la afirmación: «Hoy, después de 30 años de experiencia, no se conoce n i un solo caso de un combate eficaz de ningún insecto perjudicial por medio de hongos parásitos». Por consiguiente, si en el combate de los parásitos insectos, que son con mucho los causantes de los peores daños, se está pendiente á su vez de la intervención de sus parásitos y de insectos rapaces, se hace necesaria también una exacta investigación de las condiciones para poder distinguir lo que puede dar resultado y es realizable y lo que no lo es. Ha resultado que todos los grandes éxitos se han obtenido solamente con parásitos importados, mientras que fracasaron la mayoría de ensayos realizados con especies indígenas. Los resultados con los parásitos importados se comprenden por la siguiente consideración: El combate biológico de los parásitos se establece automáticamente en la Naturaleza cuando por cualquier factor (variaciones del clima, condiciones atmosféricas anormales ú otras) tiene lugar un incremento extraordinario de los perjudiciales; al aumento de los animales parasitables sigue en seguida, en condiciones favorables, un aumento de sus parásitos, que por destrucción de los perjudiciales, sus huevos y larvas reducen ampliamente su número. De esta manera, se establece en la Biozonose (comunidad de vida de los organismos animales y vegetales) un nuevo estado de equilibrio m á s ó menos aproximado al anterior. Pero si las condiciones son desfavorables para los parásitos, entonces tampoco tendrá ninguna acción decisiva una ayuda artificial (trasplante del parásito indígena criado), porque tan sólo puede producir un efecto momentáneo. Las condiciones son completamente diferentes cuando el dañino extranjero se introduce sin su parásito adaptado, porque según las experiencias realizadas hasta ahora, son muy poco atacados por los parásitos indígenas. La autoregulación es entonces imposible, y se presenta una procreación desenfrenada del dañino. Si entonces se logra incorporarles los parásitos que tenían en su antigua patria y éstos encuentran en el nuevo país condiciones apropiadas para su incremento, pronto se obtienen resultados importantes. De las experiencias practicadas de Hawai, pueden deducirse (según Myers), las mejores condiciones para el éxito de los métodos biológicos, las cuales se indican á continuación. 1. Los dañinos deben ser trasplantados sin parásitos. 2. La fauna indígena debe ser pobre en formas y de carácter especial, 612 COMBATE BIOLÓGICO DH LOS INSECTOS DAÑINOS para que el dañino importado tenga pocas probabilidades de encontrar allí nuevos enemigos. 3. El clima debe ser cálido, uniforme, para que los parásitos introducidos puedan acrecentarse sin trastornos estacionales. 4. L a agricultura local debe producir solamente algunos pocos productos en gran escala, con lo cual hasta las pequeñas mejoras producen un gran efecto. Quizá en ningún país de la tierra se encuentran estas condiciones tan favorablemente reunidas como en Hawai, siendo por el contrario las m á s inadecuadas las regiones continentales con una fauna muy multiforme y clima moderado con inviernos fríos. Aquí sólo pueden emplearse los métodos biológicos, en el caso de que se críen los parásitos en el laboratorio y se implanten continuamente nuevas cantidades contra los dañinos, para asegurarles su superioridad duradera. Así se procede en los lugares elevados de California, en la lucha contra las cochinillas del naranjo, realizadas con especies de coccinélidos Cryptolaemus monirouzieri. Los costes son, naturalmente, mayores que si se logra la aclimatación de los parásitos (como en la franja del litoral de California), pero, sin embargo, son aún menores que los necesarios para combatirlos con medios químicos. • Es natural aplicar también estos principios contra los parásitos indígenas, y ya se han emprendido ensayos en grande en Louisiana contra las orugas, gusanos de los frutos en California y el taladrador de la caña de azúcar. Este desarrollo más reciente de los métodos biológicos, como ya se ha mencionado, no puede presentar en ninguna parte un resultado eficaz. También en el porvenir se podrá esperar el mejor resultado de los parásitos introducidos. De gran importancia para el método biológico es el Hiperparasitismo, es decir, el fenómeno de que los parásitos mismos sean atacados á su vez por parásitos que impidan su acción favorable. Si se descuida eliminar los hiperparásitos se originan perturbaciones que la mayoría de las veces no pueden corregirse. Aprovisionamiento de parásitos.—Como las indicaciones que se encuentran en la literatura, sobre los parásitos de los dañinos, son generalmente muy incompletos y frecuentemente desacertadas, rara vez será posible, ante un nuevo caso, tomar una decisión^sin realizar previamente investigaciones cuidadosas en el mismo lugar,|Del pirausta del maíz europeo se conocía en 1919 un sólo parásito, hoy se conocen veinte. Dado el gran número d e ' d a ñ i n o s , deben implantarse también los p a r á sitos en la mayor cantidad posible. Para este aprovisionamiento pueden seguirse dos caminos: la crianza de un pequeño núcleo, procedente de la COMBATE BIOLÓGICO DE LOS INSECTOS DAÑINOS 613 patria del parásito ó su recolección en masa. Ambos métodos, tienen sus ventajas é inconvenientes. La crianza es más barata en algunos casos, cuando se tiene una organización adecuada, suministra un material libre de hiperparásitos y de otros insectos indeseables, y este material puede ser implantado en el momento más favorable; por otra parte, es necesario un personal adiestrado y una instalación especial y en algunos casos, resulta difícil la crianza de los animales parasitados. En la recolección en masa, pueden reunirse grandes cantidades con personal inexperto y equipo sencillo, en tiempo mucho más breve que en la crianza, y no necesita ningún otro control; pero los gastos son mayores, existe el peligro de la implantación de hiperparásitos y frecuentemente no es posible llevar el material en el momento conveniente para la implantación. Los perjuicios por insectos importados.—Sobre esto, así como sobre los gastos para su combate en los Estados Unidos, hace el Director del Farnham House Laboratory (Inglaterra), W . R. THOMPSON, la siguiente declaración: «De los 183 insectos perjudiciales m á s dañinos, existentes en Norteamérica, casi la mitad fueron importados de países extranjeros y de ellos una gran parte de Europa. La mosca de Hesia sola, destruye cada año una parte de la cosecha de trigo por valor de 800.000 libras. JíA Pyrausta nubüalis causa tantos daños, que para su combate se emplean anualmente medios oficiales por valor de 50.000-150.000 libras, y en e laño 1927 se gastaron 2 millones de libras para detener su amplia difusión. También para el Canadá ha llegado á ser un gran peligro: en 1925 fué destruida completamente la cosecha de maíz en un confín de m á s de 400 millas cuadradas. Los gastos del Gobierno americano para combatir el taladro del maíz el coleóptero japonés, el del suelo mejicano y la polilla oriental del melocotón han alcanzado en total hasta el año 1930., una suma que sobrepasa 12 millones de libros. En un periódico americano, el Saturday Evening Post, se calculó en 1928 que los daños producidos por los insectos implantados y los gastos oficiales y privados para combatirlos ascendía á una suma total de 1,6 millares de millones de dólares, de los cuales correspondían 1.000 millones á la pérdida de la cosecha. Resultados con el empleo de los m é t o d o s b i o l ó g i c o s . — A continuación se citan algunos de sus empleos m á s eficaces. Para el combate del Icerya purchasi, cochinilla muy dañina de las plantaciones de citrus en California que había sido importado de Australia, fueron introducidos los Noviue cardinalis (de KOÉBELE). 614 COMBATE BIOLÓGICO DE LOS INSECTOS DAÑINOS Los 100 Novius puestos en libertad, se habían convertido al cabo de un año en 10.000 y al año y medio casi habían exterminado á las cochinillas. El Novius cardinalis fué implantado más tarde también en Italia, Dalmatia, Africa del Norte y el Cabo, con completo éxito contra las Icerya (1). Contra la cochinilla del olivo de California se emplearon en 1893 con los mejores resultados la RMzobius ventrális y Cryptolaemus montrouzieri. Una serie de fracasos en otros dañinos, disminuyeron las excesivas esperanzas de los plantadores de California, pero el método biológico sufrió un nuevo impulso en 1918 por el éxito alcanzado con el Aphicus Lounsburgi HOWARD contra algunas especies de cochinillas de Citrus. Un ejemplo que demuestra la posibilidad extraordinaria d é l o s métodos biológicos es la salvación de la industria del azúcar en las Islas de Hawai ante la catástrofe amenazante que produjo la importación, en 1898, del dañino de la c a ñ a de azúcar, el PerMusiella saccharicida, lo cual se logró mediante la introducción de su parásito. Mientras anteriormente los plantadores tenían un déficit de cosecha de 3 millones de dólares anuales, á partir de 1915 se consiguió la reducción del dañino hasta una proporción no peligrosa. Finalmente citaremos los resultados obtenidos c o n t r a í a cochinilla de la morera Diaspis pentágona en Italia, Dalmatia y el Sur del Tirol por el afelínido Prospaltella herlesei (2), importado de América. Son ejemplos de casos inversos—combate de las plantas dañinas por los insectos, el control del Lantana camasa en las Islas de Hawai y del prickly pear (opuntie) en Australia. Los lantanas que fueron introducidos como arbustos de adorno y en el transcurso de los años habían inutilizado grandes superficies de dehesa pudieron ser disminuidos en una proporción no peligrosa por introducción de insectos dañinos principalmente para las flores y semillas. luos pricldy pear son vencidos cada vez más por los ataques de una gran cantidad de insectos y ácaros importados de América. Los peligros de l a c o n t a m i n a c i ó n con dañinos.—Por el aumento del tráfico mundial y el mejoramiento de los medios de transporte, se han allanado los obstáculos, en otra época inaccesibles, para la peregrinación de los insectos dañinos y ha sido aumentada considerablemente la posibilidad de su implantación. Tampoco es suficiente el control en la entrada para impedir completamente su introducción, como resulta recientemente de la implantación en los Estados Unidos de varios de los perjudiciales peores. (1) N de la R.—También se implantó en España en la región de Levante. (2) N. de la E.—Este parásito procede del Japón, donde se pudo comprobar que las abundantes plantaciones de moreras no sufrían daños de consideración, á pesar de la existencia de la cochinilla, debido á la presencia de este parásito, que la destruye. Su introducción en Europa se debe al Profesor italiano BERLESE. COMBATE BlOLÓaiCO DE LOS INSECTOS DAÑINOS 615 De otra parte, en determinadas condiciones, se obtienen resultados con los métodos biológicos, que no hubieran podido conseguirse con los métodos químicos ó solamente con gastos muchísimo mayores. En los casos más propicios se alcanzan con los métodos biológicos una destrucción duradera de una plaga de insectos, mientras que el combate químico debe ser siempre repetido. Así es comprensible que todos los Institutos entomológicos importantes hayan implantado en su programa de trabajo los métodos de combate biológicos y que hayan sido construidas organizaciones de protección en las regiones amenazadas. El U. S. Burean of Entomólogy de Washington, sostiene en todas las partes del mundo estaciones en las cuales se coleccionan y estudian por especialistas los parásitos de los perjudiciales importados. Los medios de que se dispone desde hace años, solamente el laboratorio europeo (1) del Pyrausta del maíz importa alrededor de 6.000 libras, anuales, y casi otro tanto se gastó en una serie do investigaciones. L a suma total que el Gobierno americano emplea cada año para trabajos sobre el control de los dañinos en el dominio biológico, es superior á 100.000 libras. California posee una notable organización para el combate práctico de un insecto perjudicial para las plantas, en la estación de crianza para los ya mencionados Cryptolaemus montrouzieri, empleados para combatir los «mealy bug» (cochinilla del naranjo). Las estaciones de crianza son impulsadas en parte por el Estado de California, en parte por los plantadores, pero también en parte como puro negocio (1 cent, por parásito). Según las declaraciones de H . S. SMITH, el valor material de las estaciones alcanzó en 1924 á 45,000 dólares. E l área total atacada tenía una magnitud de unas 8.000 hectáreas, y el año 1924 se implantaron allí m á s de cuatro millones de insectos. El coste de un solo rociado con medios químicos, de solamente la tercera parte del á r e a atacada, hubiera importado unos 200.000 dólares, y además, el combate con medios químicos de salpicado ó ahumado no conduce al fin, porque la cochinilla de la naranja es muy resistente contra ellos. En el Canadá, el Entomologicál service de Belleville (Ontario) sostiene un Instituto de investigación de parásitos que se ocupa principalmente del problema del Pirausta del maíz, pero sin duda se extenderá el dominio de su trabajo. 4 Tuvo un éxito completo la introducción de parásitos de Inglaterra contra un enemigo muy peligroso de las posesiones de alerce la avispa de alerce. El Instituto Cawthron, en Nueva Zelanda, y en especial el Servicio federal entomológico, en Australia,, han dedicado la mayor parte de su acti(1) N . de la R.—Laboratorio establecido por los norteamericanos en el Mediodía de Francia con el fin de encontrar, criar y utilizar los parásitos de este insecto. 616 COMBA'TB BIOLÓGICO D f i L O S I N S E C T O S DAÑINOS vidad al combate biológico de los perjudiciales, y en el mismo sentido se realizan esfuerzos en India, Sud-Africa y otras partes del Imperio británico. Observaciones finales.—El procedimiento biológico, arma reciente en el combate contra los perjudiciales de plantas, se ha mostrado, según acabamos de ver, como un medio muy valioso cuando se emplea juiciosamente. Pero para evitar desilusiones será necesario, como dice un investigador inglés, «distinguir muy cuidadosamente entre lo que puede ser emprendido desde el punto de vista del resultado práctico y lo que debe quedar como objeto de cuidadosos ensayos». BEEVE COMPLEMENTO DE LAS NOTAS ANTERIORES POR EL PR. DR. A . HASE, BERLÍN-DAHLEM. Involuntariamente se p r e g u n t a r á el lector qué se ha hecho en Alemania en el dominio del combate biológico de los perjudiciales. Antes de la guerra no se hizo prácticamente nada en este sentido. Después de la guerra,, cuando también en Alemania tomaba un gran vuelo la Entomología aplicada, se ha ocupado cuidadosamente de esta cuestión. No escaseaban, en verdad, las voces que sobreestimando sin crítica los éxitos americanos, consideraban el procedimiento biológico como el «único posible», y casi no querían saber nada de los combates químicos. Pero pronto se vió que en nuestro país había que vencer dificultades mucho mayores de todas clases (en parte relativas al problema mismo y otras del dominio económico), que las que se había supuesto primeramente. Sin embargo, el Biol. Reichsanstalt, Berlín-DaMem, ha realizado al aire libre primeramente en ensayos previos de crianza, y también algunos ensayos para establecer normas al aire, después en combinación con el / . Zool. Inst. d. Forstl. Hochschule Eberswalde, especialmente con el Trichogramma evanescens WESTW, y el Trich, minutum RILEY; finalmente con dos icneumónidos del gusano de la harina (Ephestia Mehniella). Además fueron criados en el laboratorio un icneumónido (Encarsis formosa) apropiado para la destrucción de la llamada mosca blanca (Trialeurodes) en invernaderos, y otro (Aphelinus mali), para la destrucción de los piojos de la flor del manzano. Finalmente fueron criados los icneumónidos, que pueden servir para la destrucción de polillas de cera y de pupas de moscas—para comprobar fundamentalmente los procedimientos de crianza—, cría de varios años consecutivos en el laboratorio, y también para sus correspondientes ensayos directivos al aire libre. Todos los ensayos de laboratorio aportaron la prueba de que la Entomología práctica alemana está en condiciones de dominar las dificultades técnicas, pero no debe achacársele la duradera carencia en medios para la conducción de los experimentos en grande. Los ensayos más amplios con la avispa Trichogramma para la destrucción de la polilla del pino, han sido realizados por el Zool. Instituí der Forstlichen Hochschule COHÍBATE BIOLÓGICO DE LOS INSECTOS DAÑINOS 61*1 Hann. Münden, después que el Lahoratorium, fur physiologische Zoologie an der Biol. Beichsanst. había criado exclusivamente para este fin en números redondos, ocho millones de estos icneumónido. Desgraciadamente todos los esfuerzos para llevar á cabo otras crianzas y ensayos más extensos, han fracasado otra vez por falta de medios. En Austria se sostienen crianzas de Aphelinus en diferentes instituciones escolares agrícolas. Finalmente diremos aún, que por la B. R. A . sé han realizado con éxito ensayos para eliminar la plaga de las chinches por la araña comedora de chinches Thanatos. Los resultados son alentadores; desgraciadamente no puede disponerse por ahora de medios para criar en grande esta útil a r a ñ a , natural de Grecia. Pero los ensayos realizados por la B. R. A. han demostrado la posibilidad de utilizar estos procedimientos. (Destrucción de un perjudicial por a r a ñ a s rapaces). En resumen, debe destacarse que puede y debe juzgarse meditando muy cuidadosamente sobre el valor de estos procedimientos de combate para nuestras condiciones climáticas, oconológicas y de fauna. Es posible que volvamos á ocuparnos aún de estas cosas, partiendo de consideraciones económicas determinadas. Quizá nos obligue el peligro cada vez más amenazante del escarabajo de la patata, á dedicar nuestra atención al posible combate ecológico de este dañino en grande. Extractos de publicaciones. Efecto de las sales amónicas sobre el líquido de Fehling.-—R. BAÍ/ME1UO\J.—Bull. Ph. Sud-Est., 1934, 38, p. 382.—Cuando se añaden sales amónicas al líquido de Pehling, se forma un tartrato doble de cobre y amoníaco que no se puede reducir por la glucosa. Una solución de glucosa á 0,5 por 100 conteniendo 110 gr. de cloruro amónico por 1.000 no produce ninguna reducción del reactivo, lo cual es debido a una cantidad insuficiente de sosa. Cuando se emplea el líquido de Fehling para determinar los azúcares reductores en las orinas, debe tener suficiente cantidad de sosa para neutralizar su acidez, para descomponer las sales amónicas y para formar el tartrato de cobre y de sodio reductible. Para precaverse contra los errores, la concentración en sosa del reactivo será elevada y se empleará una cantidad por lo menos igual a la del volumen de la orina 'con que se opere. Cuando sea necesario, se a ñ a d i r á sosa. Reacción luminosa para i d e n t i f i c a r el agua o x i g e n a d a . — i M . des Biol. Pharm. 1935, p. 360.—•Alcalinizando ligeramente el agua oxigenada y mezclándola con una solución de dimetilacridilium ligeramente alcalina, se produce una luminiscencia verde azulada. Se aumenta la sensibilidad de esta reacción añadiendo tetraóxido de osmio que actúa como catalizador. Conservación del complemento del conejillo de Indias.--P. MOUNIEE.— Bull. Biol. Ph., 1935, p. 285.—El autor examina la conservación de estos sueros con la solución siguiente: Acido bórico Cloruro sódico Fluoruro sódico Agua destilada c. s. p 4 gr. 9 » 6 » 100 c. c. Colocar este líquido en un matraz, añadir 20 c. c. de agua destilada y hervir hasta retorno al peso primitivo. Dejar enfriar completamente después de haber colocado en la boca del matraz una cápsula previamente flameada. Por otra parte, extraer sangre de uno o varios conejillos de Indias por medio de una punción i n t r a c a r d í a c a , colocarla en tubos de centrífuga estériles, tapar con capuchón de caucho, colocar media hora á la estufa, desprender el coágulo de las paredes y dejar 3-4 horas en un lugar fresco. Después de este tiempo, centrifugar algunos minutos y extraer el suero con una pipeta, mezclarle á partes iguales con el líquido conservador anterior, y meter inmediatamente en ampollas de 1-2 c. c. esterilizadas. En estas condiciones el suero conserva sus propiedades aléxicas casi íntegramente durante más de seis meses. Las ampollas deben conservarse en un lugar fresco, pero sin que sea necesario colocarlas en la nevera.