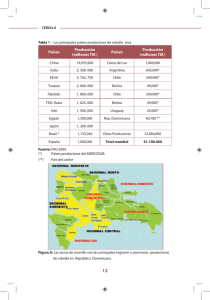
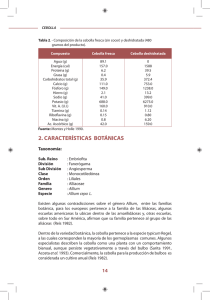
V o lu m en 3 / N o . 1 / En ero -Ju n io 2 0 0 9 R evista C o lo m b ian
Anuncio

Editorial 5 Sección de frutales Climatic change effects on the supply and prices of bulb onion. Case of Duitama municipality, Boyacá (Colombia) Biotechnologies applicable to the development of some Caricaceae species cultivated in the Andean Region: advances and problems 9 Sección de hortalizas Evaluación de dos sistemas de tutorado para el cultivo de la arveja voluble (Pisum sativum L.) en condiciones de la Sabana de Bogotá Evaluation of two training systems for indeterminate pea (Pisum sativum L.) in conditions of the Bogota Plateau Andrés Fernando Forero, Gustavo Adolfo Ligarreto Melhoramento genético de cebola para as condições tropicais e subtropicais do Brasil 18 Mejoramiento genético y producción de “semilla” de ajo (Allium sativum L.). Posibilidades de adaptación a diferentes ambientes Effect of planting density on growth, production and quality of five accessions of hot pepper (Capsicum spp.) in the Colombian Western Amazon Camila Castellanos, Jaime A. Barrera, María Soledad Hernández, Luz Marina Melgarejo, Marcela Carrillo, Ligia Rodríguez, Orlando Martínez Garlic (Allium sativum L.) genetic improvement and seed production. Possibilities of adaptation to variable environments José Luis Burba Raíces y tubérculos tropicales olvidados o subutilizados en Brasil Tropical root and tuber crops forgotten or underexploited in Brazil Marcos Vinícius Bohrer Monteiro Siqueira, Elizabeth Ann Veasey Onion and garlic crops in Colombia: current state and prospects 45 Evaluación de fuentes de fertilización edáfica y el efecto sobre la microbiota presente en un cultivo de cebolla cabezona (Allium cepa L.) 110 Caracterização fisiologica de mudas de Jatropha curcas L. produzidas em diferentes níveis de irradiancia Physiological characterization of Jatropha curcas L. plants grown under different levels of irradiance 56 Fábio Santos Matos, Cleílton Vasconcelos Moreira, Robson Fernando Missio, Luíz Antônio dos Santos Dias 126 Politica editorial e instrucciones a los autores 135 Volumen 3 / No. 1 / Enero-Junio 2009 Evaluation of fertilization sources and the effect on microbiota present in bulb onion (Allium cepa L.) culture Carlos Alberto Luengas–Gómez, Diana Paola Mora–Castro, Dorisana Guerrero–Flórez 95 Sección de otras especies 28 Los cultivos de cebolla y ajo en Colombia: estado del arte y perspectivas Hernán Pinzón R. 81 Efecto de la densidad de plantación sobre crecimiento, producción y calidad en cinco accesiones de aji (Capsicum spp.) cultivadas en la Amazonia occidental colombiana Onion breeding for the tropical and subtropical conditions in Brazil Daniela Lopes Leite, Valter Rodrigues Oliveira, Carlos Antonio Fernandes Santos, Nivaldo Duarte Costa, Maria Esther de Noronha Fonseca, Leonardo Silva Boiteux, Paulo Eduardo de Melo, Ailton Reis, Bernardo Ueno, Mirian Josefina Baptista 69 Juan Carlos Barrientos F., Germán A. Leyva V., Juan Camilo Morales B. Revista Colombiana de Ciencias Hortícolas Biotecnologías aplicables al desarrollo de algunas especies de Caricáceas cultivadas en la región Andina: avances y problemas Miguel Jordán, Diego Vélez, Rosa Armijos Efectos del cambio climático sobre la oferta y precios de cebolla de bulbo. Caso del municipio Duitama, Boyacá (Colombia) SOCIEDAD COLOMBIANA DE CIENCIAS HORTÍCOLAS – SCCH BOGOTÁ – COLOMBIA PRESIDENTE Diego Miranda VICEPRESIDENTE María Soledad Hernández EDITOR COMITÉ CIENTÍFICO Fánor Casierra–Posada Miguel A. Altieri University of California, Berkely (Estados Unidos) Galdino Andrade Filho COEDITOR Gerhard Fischer Universidade Estadual de Londrina (Brasil) Raúl Cabrera Texas A+M University (Estados Unidos) Ana Maria Castagnino Universidad Nacional del Centro de la Provincia de Buenos Aires (Argentina) Daniel H. Díaz Laboratorios Agroenzimas (México) Georg Ebert Kali + Salz Gruppe (Alemania) José Miguel Guzmán Universidad de Almería (España) Miguel Jordán Universidad Mayor, Santiago (Chile) Jairo Antonio Osorio COMITÉ EDITORIAL Marco Cabezas Bernardo Chaves Miguel Espitia Rebecca Lee Nelson Rodríguez Edison Valencia Kris Wyckhuys Corporación Colombiana de Investigación Agropecuaria (Colombia) Marcelo Francisco Pompelli Universidade Federal de Pernambuco (Brasil) Ricardo Ramírez University of Guelph (Canadá) Philip A. Stansly University of Florida (USA) © 2009 Revista Colombiana de Ciencias Hortícolas Sociedad Colombiana de Ciencias Hortícolas ISSN: 2011-2173 INFORMACIÓN, CORRESPONDENCIA Y CANJE Sociedad Colombiana de Ciencias Hortícolas Facultad de Agronomía Universidad Nacional de Colombia A.A. 14490, Bogotá, Colombia Teléfono: 571 - 316 5000 Ext. 19041/19043 Tel/fax: 571 - 316 5498 [email protected], [email protected] CORRECTOR DE ESTILO EN INGLÉS Stanislav Magnitskiy DISEÑO GRÁFICO Y ARMADA ELECTRÓNICA Formato Comunicación Diseño Ltda. [email protected] IMPRESIÓN Offset Gráfico Editores S.A. Bogotá, Colombia Junio de 2009 La presente publicación es de carácter científico (Artículo 4, resolución 1508, Ministerio de Cultura, Octubre 3 de 2000). A nivel internacional la Revista Colombiana de Ciencias Hortícolas esta integrada en las bases de datos de CAB Abstracts y AGRIS (FAO). Foto de ajo en la portada por cortesía de: Cátedra de Dibujo y Técnica Fotográfica, Facultad de Ciencias Agrarias, Universidad Nacional de Cuyo, Mendoza (Argentina) JUNTA DIRECTIVA DE LA SOCIEDAD COLOMBIANA DE CIENCIAS HORTÍCOLAS (SCCH) PRESIDENTE VICE-PRESIDENTE SECRETARIO EJECUTIVO TESORERO Diego Miranda Universidad Nacional de Colombia, Bogotá [email protected] María Soledad Hernández Instituto Sinchi, Bogotá [email protected] Gerhard Fischer Universidad Nacional de Colombia, Bogotá [email protected] Wilson de Jesús Piedrahita Universidad Nacional de Colombia, Bogotá [email protected] VOCALES Alfredo Jarma Universidad de Córdoba, Montería [email protected] Hernán Pinzón Asesor Hortícola, Bogotá [email protected] Stanislav Magnitskiy Universidad Nacional de Colombia, Bogotá [email protected] FISCAL EDITOR REVISTA Luís Enrique Flórez Universidad de Bogotá Jorge Tadeo Lozano [email protected] Fánor Casierra – Posada Universidad Pedagógica y Tecnológica de Colombia, Tunja [email protected] Marlon Hans Rodríguez Sociedad Colombiana de Ciencias Hortícolas [email protected] FÁNOR CASIERRA–POSADA Director Comité Editorial Revista Colombiana de Ciencias Hortícolas EDITORIAL Nos encontramos ad portas de la celebración del 3er Congreso Colombiano de Horticultura y del Simposio Internacional de Cebolla y Ajo en el Trópico, eventos oportunos para hacer una entrega más de nuestra Revista Colombiana de Ciencias Hortícolas, con énfasis en estas especies hortícolas. El objetivo del Simposio será la divulgación de resultados de investigación obtenidos con estas aliáceas en condiciones tropicales, incluyendo las áreas de genética y mejoramiento, fisiología, sistemas de producción, problemas fitosanitarios, tecnología de poscosecha, procesamiento, así como aspectos económicos y de mercado. De igual manera, el Simposio será un buen escenario para que destacados investigadores e interesados en estos cultivos intercambien opiniones de carácter técnico y comercial. Es la primera vez que incluimos dos trabajos en portugués como muestra de nuestra apertura a otros idiomas aparte del español, el portugués y el inglés. Además, por ejemplo esto resalta la cooperación entre investigadores brasileños y colombianos en temas importantes del desarrollo hortícola de los dos países. El primer trabajo presentado en este número para la sección de frutales se titula “Biotecnologías aplicables al desarrollo de algunas especies de Caricáceae cultivadas en la región andina”, un tema de gran actualidad, útil para el desarrollo frutícola nacional y mundial, destacando algunos hallazgos científicos relevantes y su aplicación a la fruticultura. Hemos optado por incluir prioritariamente en este número dos géneros representativos de la familia Alliaceae en la sección de hortalizas las cuales han ganado reconocimiento mundial no solo por sus cualidades culinarias, sino también por sus propiedades antisépticas, lo que ha hecho que muchos hayan sido utilizados en la medicina tradicional. Los compuestos organosulfurados de las aliáceas tienen propiedades antioxidantes, antibióticas, anticarcinogénicas, antiteratogénicas y estimulantes del sistema inmune y protectoras de la función hepática, de ahí su gran importancia. Por otro lado, sus sistemas productivos se constituyen en un importante generador de ingresos para los productores de la región y un destacado aporte al producto interno bruto agrícola de éstas zonas. Destacamos en los artículos los principales avances en las áreas de recursos genéticos, ecofisiología, manejo integrado del cultivo, cosecha, poscosecha y comercio nacional e internacional. Vol. 3 - No.1 - 2009 De igual manera se incluyen temas novedosos relacionados con algunos recursos genéticos de cultivos ancestrales, olvidados o subutilizados, pero que nuevamente cobran importancia. También sobre otras especies consideradas como promisorias (Capsicum, Jatropha), sus aspectos ecofisiológicos y de manejo integrado del cultivo que servirán de apoyo y orientación a los productores de las diferentes regiones del país. El artículo de Jatropha curcas de Brasil es de especial importancia, teniendo en cuenta que se trata de una interesante alternativa para la producción de biodiesel. Finalmente, queremos agradecer a todos los científicos nacionales e internacionales que participan en nuestro Tercer Congreso Colombiano de Horticultura y en el Primer Simposo Internacional de Cebolla y Ajo en el Trópico, porque sus artículos aseguran la calidad y actualidad de nuestro principal órgano de divulgación científica. REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS Volumen 3 - No.1 - 2009 CONTENIDO PÁG. 5 Editorial Sección de frutales Biotecnologías aplicables al desarrollo de algunas especies de Caricáceas cultivadas en la región Andina: avances y problemas Biotechnologies applicable to the development of some Caricaceae species cultivated in the Andean Region: advances and problems Miguel Jordán, Diego Vélez, Rosa Armijos................................................................................................................................................... 9 Sección de hortalizas Melhoramento genético de cebola para as condições tropicais e subtropicais do Brasil Onion breeding for the tropical and subtropical conditions in Brazil Daniela Lopes Leite, Valter Rodrigues Oliveira, Carlos Antonio Fernandes Santos, Nivaldo Duarte Costa, Maria Esther de Noronha Fonseca, Leonardo Silva Boiteux, Paulo Eduardo de Melo, Ailton Reis, Bernardo Ueno, Mirian Josefina Baptista.................................................................................................................................................. 18 Mejoramiento genético y producción de “semilla” de ajo (Allium sativum L.). Posibilidades de adaptación a diferentes ambientes Garlic (Allium sativum L.) genetic improvement and seed production. Possibilities of adaptation to variable environments José Luis Burba........................................................................................................................................................................................ 28 Los cultivos de cebolla y ajo en Colombia: estado del arte y perspectivas Onion and garlic crops in Colombia: current state and prospects Hernán Pinzón R........................................................................................................................................................................................ 45 Evaluación de fuentes de fertilización edáfica y el efecto sobre la microbiota presente en un cultivo de cebolla cabezona (Allium cepa L.) Evaluation of fertilization sources and the effect on microbiota present in bulb onion (Allium cepa L.) culture Carlos Alberto Luengas–Gómez, Diana Paola Mora–Castro, Dorisana Guerrero–Flórez............................................................................. 56 Efectos del cambio climático sobre la oferta y precios de cebolla de bulbo. Caso del municipio Duitama, Boyacá (Colombia) Climatic change effects on the supply and prices of bulb onion. Case of Duitama municipality, Boyacá (Colombia) Juan Carlos Barrientos F., Germán A. Leyva V., Juan Camilo Morales B................................................................................................... 69 Evaluación de dos sistemas de tutorado para el cultivo de la arveja voluble (Pisum sativum L.) en condiciones de la Sabana de Bogotá Evaluation of two training systems for indeterminate pea (Pisum sativum L.) in conditions of the Bogota Plateau Andrés Fernando Forero, Gustavo Adolfo Ligarreto................................................................................................................................... 81 Efecto de la densidad de plantación sobre crecimiento, producción y calidad en cinco accesiones de aji (Capsicum spp.) cultivadas en la Amazonia occidental colombiana Effect of planting density on growth, production and quality of five accessions of hot pepper (Capsicum spp.) in the Colombian Western Amazon Camila Castellanos, Jaime A. Barrera, María Soledad Hernández, Luz Marina Melgarejo, Marcela Carrillo, Ligia Rodríguez, Orlando Martínez............................................................................................................................................................. 95 Sección de otras especies Raíces y tubérculos tropicales olvidados o subutilizados en Brasil Tropical root and tuber crops forgotten or underexploited in Brazil Marcos Vinícius Bohrer Monteiro Siqueira, Elizabeth Ann Veasey........................................................................................................ 110 Caracterização fisiologica de mudas de Jatropha curcas L. produzidas em diferentes níveis de irradiancia Physiological characterization of Jatropha curcas L. plants grown under different levels of irradiance Fábio Santos Matos, Cleílton Vasconcelos Moreira, Robson Fernando Missio, Luíz Antônio dos Santos Dias...................................... 126 Politica editorial e instrucciones a los autores 135 REV. COLOMB. CIENC. HORTÍC. Biotecnologías aplicables al desarrollo de algunas especies de Caricáceas cultivadas en la región Andina: avances y problemas Biotechnologies applicable to the development of some Caricaceae species cultivated in the Andean Region: advances and problems Miguel Jordán1, 3 DIEGO Vélez2 ROSA Armijos2 Frutos maduros de Vasconcellea cundimarcensis. Foto: M. Jordan RESUMEN Diferentes especies de la Familia Caricaceae corresponden a cultivos importantes de la región Andina, teniendo usos como frutales, producción de compuestos enzimáticos con acción proteolítica, como fuentes de germoplasma y potencial génico de interés posible de usar en programas de mejoramiento. Entre ellas se destacan Carica papaya y otro grupo de cultivos clasificados dentro del género Vasconcellea; entre ellos V. cundinamarcensis [sin. C. pubescens] (papayo de montaña); y V. stipulata; ambas portadoras de genes que confieren resistencia a “papaya ringspot virus”, así como para tolerancia de frío, con características organolépticas y como patrones de injerto de otro cultivo como es V. x heilbornii nm. pentagona (babaco). Según cada cultivo, se evidencian diferentes potenciales regenerativos, ya sea a nivel sexual y asexual, pero también grandes limitaciones que afectan su desarrollo y su regeneración. Algunas de las varias limitaciones específicas a mencionar, según especie, corresponden a problemas de germinación por causa de dormancia influenciada por la testa, variabilidad en el grado de viabilidad según proveniencia y lento desarrollo de postemergencia (V. stipulata). En otras situaciones, la reproducción asexual es obligatoria, tratándose de híbridos interespecíficos estériles, como es el caso de V. x heilbornii, respuesta que se ve parcialmente limitada. Fundamentalmente la presencia de virosis es el mayor problema que afecta los cultivos comerciales de C. papaya; conjuntamente la existencia de las diferentes formas sexuales limita la productividad de V. cundinamarcensis. El objeto del presente trabajo es señalar algunos resultados de estudios conducidos en nuestros laboratorios, así como enfocar algunas estrategias biotecnológicas que han sido exploradas y en desarrollo, útiles en la superación de algunas de dichas problemáticas. 1 2 Instituto de Biotecnología, Universidad Mayor, Santiago de Chile. Instituto de Ecología, Universidad Técnica Particular de Loja, Loja, Ecuador. 3 Autor para correspondencia. [email protected] REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 3 - No.1 - pp. 9-17, 2009 10 JORDAN/VÉLEZ/ARMIJOS Palabras clave adicionales: Vasconcellea, Carica papaya, papaya de montaña, organogénesis, embriogénesis somática, regeneración in vivo & in vitro, germoplasma. ABSTRACT Different species of Caricaceae family are intensively cultivated in the Andean Region consumed as fruit, used as sources of proteolytic enzymatic compounds and as germplasm sources, and have genetic potential of possible interest for improvement programs. Among these species stay Carica papaya and another group of species classified within the Vasconcellea genus, such as V. cundinamarcensis [syn. C. pubescens] (mountain papaya) and V. stipulata. Both species carry genes conferring resistance to “papaya ringspot virus” as well as cold tolerance and also have specific organoleptic characteristics. They can be used as graft stocks for other cultures such as V. x heilbornii nm. pentagona (babaco). Different regenerative potentials are evidenced, according to each species at the sexual or asexual level as well as important limitations affecting their development and regeneration. Some of the specific limitations to be mentioned are: germination problems due to dormancy influenced by the testa, variability in the degree of viability according to provenance, slow post-emergence development and associated aspects, with significant losses to fungal attack (V. stipulata). Asexual reproduction is otherwise mandatory, for sterile interspecific hybrids such as V. x heilbornii, whose response is partially limited. The presence of virosis is the main problem affecting commercial cultures of C. papaya; the existence of different sexual forms affects the productivity of V. cundinamarcensis. The objective of this work is to mention several results of studies carried out in our laboratories, focusing on various biotechnological strategies already explored or being developed, which can be used to overcome some regeneration problems. Additional key words: Vasconcellea, Carica papaya, mountain papaya, organogenesis, somatic embryogenesis, in vivo & in vitro regeneration, germplasm. Fecha de recepción: 28-04-2009 Aprobado para publicación: 01-06-2009 Lista de abreviaciones: AIA: ácido indolacético; AIB: ácido indol-3-butírico; AG3: ácido giberélico; ANA: ácido α-naftalenacético; BAP: 6-bencilaminopurina; TDZ- 1-fenil-3-(1,2,3,-tidiazol-5-il)urea (tidiazuron). INTRODUCCIÓN La Región Andina, centro de origen de las numerosas especies e híbridos pertenecientes a la Familia Caricaceae, y que evidencian considerable variabilidad genética, constituyen una fuente importante de recursos genéticos para mejoramiento vegetal y productos tanto agrícolas, medicinales o industriales. Diferentes especies que proveen de características de resistencia a virosis pueden participar en cruzamientos interespecífi- REV. COLOMB. CIENC. HORTIC. cos generando híbridos, aportar genes de tolerancia para el frío favoreciendo la extensión de cultivos a zonas por hoy limitantes o para otorgar características organolépticas. La recombinación genética, los aspectos de selección y generación de genotipos y la multiplicación, en sus formas, agámica y sexual, vía in vivo o in vitro son en su conjunto todas relevantes. Diferentes potencialidades biotecnológicas han sido exploradas en las B I OT E C N O LO G Í A S A P L I C A B L E S A L D E S A R R O L LO D E A L G U N A S E S P E C I E S D E C aric á ceas diferentes especies de Caricáceas y han conducido al conocimiento del potencial morfogénico y regenerativo existente. En el presente trabajo se intenta recopilar algunos antecedentes sobre el potencial de respuestas inductivas y regenerativas inducidas, expresables en células, tejidos y órganos de algunas Caricáceas, biotecnologías de posible utilidad en estudios atingentes a estas especies. MATERIAL Y MéTODOS A fin de evaluar el potencial morfogénico y regenerativo in vivo e in vitro según la especie, se evaluaron respuestas bajo diferentes condiciones a partir de células y protoplastos derivados, de varios tipos de tejidos y órganos. Se ensayaron anteras a fin de conocer el potencial androgénico existente en microsporas. A partir de suspensiones celulares se evaluó la capacidad de iniciar embriogénesis somática; embriogénesis somática adventicia y producción de semillas sintéticas mediante encapsulación, y en protoplastos la posibilidad de fusiones interespecíficas. A partir de hojas se evaluó la inducción de nuevos brotes o embriogénesis somática, así como desde hipocótilos, ápices caulinares, yemas axilares–secciones nodales y secciones de tallo. Respuestas fisiológicas y adaptativas fueron referidas a aspectos de viabilidad, germinación y control de la hiperhidricidad y caracterización parcial de germoplasma. La metodología empleada se encuentra resumida en Jordan et al. (1986); Jordan (1992); Jordan y Velozo (1996); Jordan y Piwanski (1999) y Vélez et al. (2009). RESULTADOS Y DISCUSIóN Respuestas morfogénicas y regenerativas del material estudiado, parcial caracterización del germoplasma (V. cundinamarcensis), condiciones de cultivo y de requerimientos hormonales se resumen en las tablas 1 y 2. De los resultados se desprende que una gran parte de los diferentes tipos de explantes ensayados expresan respuestas morfogénicas dependientes de las potencialidades inherentes a la especie y del órgano–tejido examinado, aun en presencia de niveles de reguladores de crecimiento y condiciones de cultivo muy semejantes. Así, por ejemplo, la inducción de embriones somáticos en callo de diferente origen V. cundinamarcensis es fácilmente inducible (Jordan, 1992), mientras que en V. heilbornii nm. pentagona este tejido prospera difícilmente, aun en presencia de addenda orgánica, evidenciando pardeamiento sin generar embriones; respuesta en parte semejante a nuestros resultados con C. papaya. Por el contrario, explantes de lámina foliar de V. cundinamarcensis no son inducibles a expresar la formación de brotes de–novo, mientras sí se inician en hojas adultas de babaco (Jordan y Piwanski, 1997, 1999). V. cundimamarcensis expresa amplia respuesta mor­ fogénica en sus tejidos. Diferentes tipos y niveles hormonales (combinaciones de ANA con BA; ANA con Z; ANA con K; AIA con BA y AIA con Z) inician todos la formación de embriones en callo derivados de temas axilares de plantas adultas o de hipocótilos (Jordan, 1986). Dichos embriones son inducibles, además de generar embriones adventicios, con lo cual se facilita su multiplicación masiva (Jordan y Velozo, 1996). Un encapsulamiento de estos embriones (para uso como semilla sintética) también parece factible (Jordan, 2005). Dadas las variadas formas sexuales (por ejemplo en V. cundinamarcensis poco reconocibles en material juvenil), estrategias como la inducción homozigosis de doble haploides mediante el proceso de androgénesis en plantas altamente productivas aparece como una estrategia de selección y producción de germoplasma en programas de mejoramiento genético. Si bien las respuestas androgénicas en Caricáceas han sido poco descritas, en estudios preliminares el potencial en V. cundimarcensis, alcanza a aprox. > 5% de inducción de embrioides derivados de polen y hasta un 91% de callo embriogénico, probablemente también de carácter haploide (Jordan, 1996). Vol. 3 - No.1 - 2009 11 12 JORDAN/VÉLEZ/ARMIJOS El potencial de cruzamientos interespecíficos entre especies de la familia Caricaceae está ampliamente establecido, y con ello su potencial de recombinación (con resultados tan notables como en babaco). Al respecto, Kyndt et al. (2005 a, b) indica que la hibridación es común entre especies del genero Vasconcellea, encontrándose signos de introgresión de V. cundinamarcensis en V. stipulata en condiciones naturales como controladas (De Zerpa, 1980); dicha variabilidad es notable en V. stipulata en material colectado de diferentes procedencias (Horovitz y Jiménez, 1967). Al respecto, cabe notar que en las poblaciones de cultivo de V. cundinamarcensis, originarias de las regiones de altura de la Region Andina, distribuida desde Panamá a Bolivia en zonas temperadas entre 1.500-3.000 m (Badillo, 1971; Muñoz, 1965), adaptadas a la zona costera en Chile central (28° y 36° S.), éstas parecen ser más bien homogéneas; estudios isoenzimáticos denotan extrema similitud en muestras en más de 1.000 km de extensión. Así, considerando el potencial de cruzamiento interespecies existente, trabajos experimentales de fusión a nivel de protoplastos han sido sin embargo bastante limitados. Al respecto, algunos experimentos de fusiones de protoplastos entre V. cundinamarcensis y C. papaya con perspectivas de lograr híbridos o recombinantes con caracteristicas de interés posibles de usar en programas genéticos (inserción de resistencia a PRV, características de tolerancia al frío, generación de recombinantes como patrones de injerto), sólo inician las primeras fases de desarrollo embriogénico in vitro y no desarrollan estructuras viables (Jordan et al., 1986). La imcompatibilidad génica al recombinar el genoma de ambas procedencias es probablemente la causa de los escasos resultados disponibles. Sin embargo, establecidos los mecanismos morfogénicos y de regeneración celular y tisular y con el advenimiento de metodologías que permiten la inserción de genes foráneos entre una especie y otra permite la fijación y expresión de características particulares deseables. Entre éstas, ayudan marcadores moleculares ligados a genes de resistencia a PRV presentes en V. cundinamarcensis (Dillon et al., 2006). Finalmente, mediante ingeniería genética, insertando el gene codificante de la proteina viral del PRV, ha sido posible obtener existosamente plantas transgénicas de C. papaya resistentes en variedades comerciales, intensidad de respuesta que depende de la dosificación presente de dicho gen en las nuevas plantas transformadas (Fitch et al., 1992; Tennant et al., 1994, 2001.; Gonzalves y Ferreira, 2003). Otros muchos mecanismos de regulación son necesarios en la estabilización de respuestas ante la modificación genética y consecución final de un producto transgénico comercial. Entre las Caricaeas, las respuestas regenerativas in vitro, según el caso, parecen ser más satisfactorias que en muchas especies vegetales de carácter recalcitrante, característica de no menor relevancia cuando a nivel del material celular transformado los procesos morfogénicos deben encaminarse hacia la obtención in vitro de una planta viable completa. Tabla 1.Resumen de respuestas regenerativas, potencial regenerativo y parcial caracterización del germoplasma en algunas Caricáceas*. Especie–explante Microsporas uninucleadas Cultivos celulares REV. COLOMB. CIENC. HORTIC. Vasconcellea cundimarcensis Androgénesis, embriones somáticos derivados de polen–callo Embriones somáticos, plántulas, embriogénesis; embriog. adventicia, encapsulación V. heilbornii nm. pentagona V. stipulata Carica papaya Formación de callo Formación de callo (No aplica) Formación de callo B I OT E C N O LO G Í A S A P L I C A B L E S A L D E S A R R O L LO D E A L G U N A S E S P E C I E S D E C aric á ceas Tabla 1. Continuación Especie–explante Vasconcellea cundimarcensis Fusión de protoplastos Con Carica papaya; primeros estados de embriogénesis de células fusionadas Lámina foliar Sin respuesta morfogénica Elemento bulboso en porción basal de lámina foliar Hipocótilo Embriogénesis somática Ápices caulinares Yemas axilares– secciones nodales Estacas de tallo Semillas Embriones Brotación de las yemas axilares preexistentes; formación nuevos brotes múltiples, callo y embriogénesis indirecta Raíces; plantas Incremento en el porcentaje de germinación in vitro en presencia de ANA y Kin, medio MS Caracterización de germoplasma Formas sexuales en 1. Determinación de plantas masculinas extractos de fenoles y productoras HPLC de hojas y flores femeninas plantas ambos sexos 2. Electroforesis SDS poliacrilamida hojas, flores, pecíolos en plantas de ambos sexos Ecotipos–variación Electroforesis en gel somaclonal de isoenzimas (AAT) y otras enzimas indicadoras V. heilbornii nm. pentagona V. stipulata Carica papaya Con Vasconcellea. cundinamarcensis; primeros estados de embriogénesis de células fusionadas Formación de callo, brotes de novo o embriones somáticos, hoja adulta Inducción de nuevos brotes; plántulas (No aplica) A partir de subcultivo de los nuevos bro­ tes generados del elemento bulboso que evidencian inducción de raíces, regeneración de plántulas Formación de raíces y plantas escasa Raíces; plantas (No aplica) (No aplica) Iniciación de brotes de hojas derivadas de ejes embrionarios (Estructura visible poco desarrollada según ontogenia foliar) Regeneración de plántulas, control de la hiperhidricidad Formación de callo Formación de callo de ápices de plántulas en desarrollo Brotación de las yemas axilares preexistentes; nuevos brotes, callo Escarificación promueve porcentaje y reduce tiempo de germinación Reducción tiempo de latencia * Resultados experimentales realizados por autores exclusivamente. Vol. 3 - No.1 - 2009 13 14 JORDAN/VÉLEZ/ARMIJOS Tabla 2.Resumen de mejores condiciones inductoras de respuestas morfogénicas y regenerativas en varias Caricáceas Respuesta morfogénica Inducción – tratamiento Medio nutritivo – reguladores Addenda – otros Resultado Generación de embrioides de polen 6,5% Androgenesis (V. cundinamarcensis) Cultivo de anteras y microsporas uninucleadas de plantas femeninas Sales NN; microelementos de MS, sacarosa 60 g L-1. AIA 1,0 mg L-1 BAP 1,0 mg L-1 (papel Whatman N° 1 reemplaza agar) De preferencia cultivo bajo luz fluorescente tenue, ciclos de 18 horas; aprox. 48 µmol m-2 s-1 Embriogenesis somática [hipocótilo] (V. cundinamarcensis; C. papaya) Formación de callo inicial, posteriormente subcultivo, estático o agitación cte. Varios embriones somáticos, plántulas, embriogénesis; embriog. adventicia, encapsulación Para el caso de C. papaya se requiere oscuridad. Medios: MS y NN En V. cundinamarcensis formación de callo y embriones somáticos, tb. embriones adventicios, abundantes y plántulas usando pretratamientos con 2,4-D Fusión de protoplastos (V. cundinamarcensis x C. papaya) Células de C. cundinamarcensi s derivadas de hoja juvenil; de C. papaya obtenidas de callo formado de hipocótilo Preplasmólisis con manitol 13%, 60 min; digestión por 39 h en celulasa (2%) , Macerozyma R-10 (0,5%) y manitol 13%, en oscuridad, 30°C, sales MS Desinfección hoja en etanol 70%, 2 min Protoplastos viables de ambos tejidos, densidad aprox. 44 x 104 en C. cundinamarcensis. Primeros estados de división y embriogénesis de productos de fusión aunque sin posterior desarrollo Formación de embriones somáticos–brotes en lámina foliar (V. heilbornii nm. pentagona) Inducción de brotes y embriones somáticos en secciones de lamina foliar; respuestas de acuerdo a uso y niveles de reguladores Fases de cultivo de hoja y subcultivo de callo al inicio con TDZ y AIA, luego con AG3. Callo en placa, estático desarrolla embriones somáticos. En babaco formación de brotes solo con alta concentración de TDZ Inducción de embriones somáticos (indirecto) sólo en presencia glutamina, hidrolizado de caseína y cisteína Varias fases de subcultivo; con respuesta embriogénica menor que observada en V. cundinamarcensis, en la cual no se generan por contrario brotes a partir de explantes foliares Formación de brotes [elemento bulboso] en lámina foliar (V. heilbornii nm. pentagona) Inducción de brotes sólo del elemento bulboso, ligado vascularmente y cultivado con sección de hoja Medio MS con TDZ solo o combinado incluyendo hidrolizado de caseína y sulfato de adenina. Para rizogénesis de brotes en subcultivo AIB 2,0 mg L-1 Presencia de maltosa (2%) y TDZ 3,0 mg L-1 imprescindibles para la inducción de respuesta y síntesis de clorofila en los nuevos brotes Obtenidos los brotes el enraizamiento en subcultivo ocurre en un 50% aprox. REV. COLOMB. CIENC. HORTIC. B I OT E C N O LO G Í A S A P L I C A B L E S A L D E S A R R O L LO D E A L G U N A S E S P E C I E S D E C aric á ceas Tabla 2. Continuación. Respuesta morfogénica Inducción – tratamiento Medio nutritivo – reguladores Addenda – otros Resultado Ápices caulinares [regenerados in vitro]. (V. heilbornii nm. pentagona) Formación de callo A partir de nuevos brotes in vitro, formación de callo, inducción de raíces, plántulas Yemas axilares (V. heilbornii nm. pentagona; V. cundinamarcensis; C. papaya; V. stipulata) Inducción de callo morfogénico en porción proximal del ápice en varias especies conducente a embriones somáticos en subcultivo En babaco, inducción raíces en presencia de ANA 0,01 mg L-1 y BAP 0,8 mg L-1. Nutrientes NN. Para C. papaya, V. cundinamarcenesis y V. stipulata (secciones nodales), brotación de yemas axilares; con formación de nuevos brotes, brotes múltiples, callo y embriogénesis indirecta Babaco requiere incluir junto a reguladores carbón activado 0,6 g L-1; tb. AdS, hidrolizado caseína y cisteína a fin de evitar pardeamiento. Medios WPM y MS En babaco, inducción de raíces en la base con producción de plantas en forma errática Estacas de tallo (V. heilbornii nm. pentagona) Tratamiento directo de inmersión y posterior mantención de estacas en agua conteniendo AIB Formación de raíces en presencia de AIB, 50 mg L-1, 7 min Addenda no requiere; solo condiciones de invernadero. Respuesta a luz Todas las estacas con 3-4 raíces en la porción proximal Semillas (V. stipulata) Escarificación de la esclerotesta Peróxido de hidrógeno (no aplica) Incremento de porcentaje de germinación de 0,5% a aprox. 53% en 40 días Embriones (V. stipulata) Ruptura de la dormancia, desarrollo rápido eje embrionario Ajuste concentración de sales MS y NN más AG3 (no aplica) Germinación más temprana, control de hiperhidricidad de ejes embrionarios en desarrollo; reducción al 0-13% MS: Murashige y Skoog, 1962; NN: Nitsch y Nitsch, 1969; WPM: Lloyd y McCown, 1981. CONCLUSIONES Con base en la vasta literatura existente, varias especies adscritas a la familia de la literatura existente, sometidas a condiciones inductivas específicas in vivo e in vitro, parecen expresar un po- tencial embriogénico y regenerativo satisfactorio a nivel celular, tisular o de órganos. Lo anterior es ventajoso en programas de mejoramiento o de propagación masiva para fines productivos. Particularmente relevante, considerando estrategias como las de transgenia, es la posibilidad de poder ser regeneradas plantas viables llevando íntegra- Vol. 3 - No.1 - 2009 15 16 JORDAN/VÉLEZ/ARMIJOS mente el gen–carácter de interés a partir de células o tejidos virtualmente transformados. En segundo lugar, la posibilidad de poder seleccionar material de interés y la regeneración de plantas a partir del nivel celular hace posible y facilita también la selección temprana de características particulares, naturales o inducidas detectables a ese nivel y, en tercer lugar, la expresión de la androgénesis, derivando en la regeneración de plantas di-haploides (doble haploides o de carácter homocigoto) derivadas del polen, respuesta inducible y obtenible a corto plazo, en una sola generación, permite igualmente la selección de material con características no posibles de ser obtenidas en programas de mejoramiento convencionales. La obtención de plantas en forma directa a partir del polen descarta la fase de generación de heterocigotos predominantes en las primeras fases de cruzamientos convencionales. La resultante del empleo conjunto de las biotecnologías mencionadas implica poder incrementar en el ámbito de las Caricaeas la variabilidad de germoplasma por hoy disponible y hacer uso de éste en futuros programas de selección, recombinación o de mejoramiento genético en las especies adscritas a esta familia de plantas. REFERENCIAS BIBLIOGRÁFICAS Badillo, V.M. 1971. Monografía de la familia Caricaceae. Fac. Agr. Maracay, Venezuela. censis and C. papaya protoplasts. Gartenbauwissenschaft 51, 175-178. De Zerpa, D.M. 1980. Comportamiento meiótico de la descendencia híbrida producida al transferir el carácter bisexual de Carica pubescens a Carica stipulata. Rev. Fac. Agron. (Maracay) 11, 5-47. Jordan, M. 1992. Capítulo II 12: Micropropagation of Papaya (Carica sp). pp. 418-459. En: Bajaj, Y.P.S. (ed.) Biotechnology in Agriculture and Forestry Vol. 18, Springer-Verlag, Berlin-Heidelberg. Dillon, S.; C. Ramage; S. Ashmore y R.A. Drew. 2006. Development of a codominant CAPS marker linked to PRSV-P resistance in highland papaya. Theor. Appl. Genet. 113, 1159-1169. Jordan, M. 1996. In vitro regeneration of some little known Andean fruit crops. p. 34. Proc. 1st. Conference on Fruit Production in the Tropics and Subtropics. Humboldt-Universität zu Berlin, Berlin. Fitch, M.; R. Manshardt; D. Gonsalves; J. Slightom y J. Sanford. 1992. Virus resistant papaya derived from tissues bombarded with the coat protein gene of papaya ringspot virus. BioTechnol. 10, 1466-1472. Jordan, M. y J. Velozo. 1996. Improvement of somatic embryogenesis in highland-papaya cell suspensions. Plant Cell Tissue Organ Cult. 44, 189-194. Gonsalves, D. y S. Ferreira. 2003. Transgenic papaya: A case for managing risks of papaya ringspot virus in Hawaii. Online. Plant Health Progress doi:10.1094/ PHP-2003-1113-03-RV. Horovitz S. y H. Jiménez. 1967. Cruzamientos interespecíficos e intergenéricos en Caricáceas y sus implicaciones fitotécnicas. Agronomía Tropical (Maracay) 17, 323-343. Jordan, M.; G. Ciudad; M.L. Rojas y F. Valverde. 1986. Isolation, culture and fusion of Carica candamar- REV. COLOMB. CIENC. HORTIC. Jordan, M. y D. Piwanski. 1997. Regeneration of babaco [Carica pentagona (Heilborn) Badillo] using leaf explants and shoot-tip culture. Rev. Intl. Bot. Exp. 61, 109-115. Jordan, M. y D. Piwanski. 1999. Regeneration of babaco (Carica pentagona) through somatic embryogenesis. Φyton 64, 101-105. Jordan, M. 2005. Capítulo 4: Aplicaciones de la técnica de cultivo de tejidos. pp.73-98. En: Prieto, H., M. Jordan; L.P. Barrueto; M.C. Rocha y D. Durzan. Biotecnologia vegetal. Colección Libros INIA, Nº 15. B I OT E C N O LO G Í A S A P L I C A B L E S A L D E S A R R O L LO D E A L G U N A S E S P E C I E S D E C aric á ceas I­ nstituto de Investigaciones Agropecuarias, Ministerio de Agricultura, Prograf Impresores, Santiago. Nitsch J.P. y C. Nitsch. 1969. Haploid plants from pollen grains. Science 163, 85-87. Kyndt, T.; E. Romeijn-Peeters; B. Van Droogenbroeck; J. Romero-Motochi; G. Gheysen y P. Goetghebeur. 2005a. Species relationships in the genus Vasconcellea (Caricaceae) based on molecular and morphological evidence. Amer. J. Bot. 92, 1033–1044. Muñoz, M. 1965. Síntomas de deficiencias nutricionales en plantas de papayo (Carica candamarcensis Hook). Trabajo de grado. Facultad de Agronomía, Universidad de Chile, Santiago. Kyndt, T.; B. Van Droogenbroeck; E. Romeijn-Peeters; J. Romero-Motochi; X. Scheldeman; P. Goetghebeur; P. van Damme y G. Gheysen. 2005b. Molecular phylogeny and evolution of Caricaceae based on rDNA internal transcribed spacer (ITS) and chloroplast sequence data. Mol. Phylogenet. Evol. 37, 442-459. Lloyd, G. y B.H. McCown. 1981. Woody Plant MediumA mineral nutrient formulation for microculture of woody plant species. HortScience 16, 453. Murashige, T. y F. Skoog. 1962. A revised medium for rapid growth at bio-assays with tobacco tissue cultures. Physiol. Plant. 15, 473-497. Tennant, P., C. Gonsalves; K. Ling; M. Fitch; R. Manshardt; J. Slightom y D. Gonsalves. 1994. Differential protection against papaya ringspot virus isolates in coat protein gene transgenic papaya and classically cross-protected papaya. Phytopathol. 84, 1359-1366. Tennant, P.; G. Fermin; M. Fitch; R. Manshardt; J. Slightom y D. Gonsalves. 2001. Papaya ringspot virus resistance of transgenic Rainbow and SunUp is affected by gene dosage, plant development, and coat protein homology. Eur. J. Plant Pathol. 107, 645-653. Vélez D.; R. Armijos y M. Jordan. 2009. Germinación y desarrollo in vitro de embriones de Vasconcellea stipulata Badillo: tratamientos pre-germinativos y control de hiperhidricidad (en preparación). Vol. 3 - No.1 - 2009 17 Melhoramento genético de cebola para as condições tropicais e subtropicais do Brasil Onion breeding for the tropical and subtropical conditions in Brazil Daniela Lopes Leite1,4 Valter Rodrigues Oliveira2 Carlos Antonio Fernandes Santos3 Nivaldo Duarte Costa3 Maria Esther de Noronha Fonseca2 Leonardo Silva Boiteux2 Paulo Eduardo de Melo2 Ailton Reis2 Bernardo Ueno1 Mirian Josefina Baptista2 Bulbos de cebola da cultivar BRS Cascata. Foto: C. Ruas Schimulfening Resumo O agronegócio cebola é importante para o Brasil, onde se cultivam cerca de 62.000 há ano -1 nas regiões Sul, Sudeste, Nordeste e Centro-Oeste. Há demanda por cultivares melhor adaptadas às diferentes condições edafoclimáticas; para cultivo em sistemas convencionais e agroecológicos; com bulbos de sabor pungente para mercado interno, suave/doce e do tipo “cascuda bronzeada” para mercados interno e de exportação. O trabalho objetiva desenvolver cultivares de polinização livre e/ou híbridas de cebola adaptadas às diferentes condições edafoclimáticas brasileiras e resistentes às principais doenças que ocorrem na cultura. As atividades de pesquisa vem sendo conduzidas por uma rede de Pesquisa e Desenvolvimento envolvendo três Unidades da Embrapa e instituições parceiras/colaboradoras, públicas e privadas, além de produtores rurais. Estão sendo usados métodos de melhoramento com aplicação de seleção recorrente e com desenvolvimento de linhas A (macho-estéril), B e C (macho-férteis) e produção e testes de híbridos experimentais, na criação de novas cultivares. Técnicas de biologia molecular estão sendo aplicadas na identificação de mantenedoras da machoesterilidade, na caracterização da variabilidade genética de germoplasma e no mapeamento de caracteres de qualidade e resistência a doenças. Os impactos potenciais decorrentes da incorporação de cultivares mais atrativas e resistentes a doenças e com padrão genético superior são, a médio prazo, maior competitividade da cebola brasileira; estabilização e potencial redução da importação de sementes; possibilidade de exportação de cebola; diminuição no uso de agrotóxicos pela incorporação de cultivares resistentes a doenças com diminuição do custo de produção e dos impactos ambientais. 1 Embrapa Clima Temperado, Pelotas, Brasil. Embrapa Hortaliças, Brasília, Brasil. 3 Embrapa Semi-Árido, Petrolina, Brasil. 4 Autor para correspondência. [email protected] 2 REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 3 - No.1 - pp. 18-27, 2009 Melhoramento genético de cebola Palavras chave adicionais: Allium cepa L., cultivares, resistência a doenças, pungência, biologia molecular. Abstract The onion culture is an important agribusiness in Brazil, where it is cultivated around 62,000 ha year-1 in the South, Southeast, Northeast and the Midwest. There is a demand for improved cultivars adapted to different soil and climatic conditions; for cultivation in conventional and agroecological systems, with bulbs of pungent flavor to the internal market, soft / sweet and “dark bronze and thick scales” types for domestic and export markets. The work aims to develop open-pollinated and / or hybrid cultivars of onion adapted to the different Brazilian soil and climatic conditions and resistant to major diseases occurring in the culture. The research activities have being conducted by a network of Research and Development involving three Embrapa units and public and private partner/collaborator institutions, and rural producers. The breeding methods being used for the new cultivar development are the application of recurrent selection and development of A lines (malesterile), B and C (male-fertile) and production and testing of experimental hybrids. Techniques of molecular biology have being applied in the identification of male sterility maintainer lines, in the characterization of germplasm genetic variability and mapping of quality traits and resistance to diseases. The potential impacts from the incorporation of more attractive and disease resistant cultivars with superior genetic pattern, are in the medium term, higher competitiveness of Brazilian onion, and stabilization and potential reduction of seed importation; possibility of onion exportation, decrease of pesticide use by disease resistant cultivars with reduction in production costs and environmental impacts. Additional key words: Allium cepa L., cultivars, disease resistance, pungency, molecular biology. Fecha de recepción: 22-04-2009 Aprobado para publicación: 01-06-2009 Introdução O início do cultivo de cebola amarela no Brasil ocorreu com a chegada de imigrantes açorianos que colonizaram a região de Rio Grande, no Rio Grande do Sul, durante o século XVIII e início do século XIX (Melo et al., 1988; França e Candeia, 1997). Das cebolas introduzidas da Europa, desenvolveram-se, por seleção natural e pela ação de agricultores, diversas populações que são agrupadas em dois tipos de acordo com a cultivar de origem: ‘Baia Periforme’, que engloba as populações derivadas de uma cebola portuguesa conhecida como Garrafal e ‘Pêra’, possivelmente populações derivadas de genótipos egípcios introduzidos na Ilha dos Açores e posteriormente trazidas para o Brasil. Um terceiro tipo, possivelmente resultante do cruzamento entre populações do tipo ‘Baia Periforme’ e ‘Pêra’ e denominado ‘Crioula’ surgiu na região do Alto Vale do Itajaí, em Santa Catarina (Costa, 1997). Como características, estes três tipos possuem elevada tolerância a moléstias e, sobretudo, boa conservação pós colheita. Seleção nesses tipos, especialmente em ‘Baia Periforme’, ocorreu no Rio Grande do Sul e em outros estados brasileiros a partir da criação dos programas de melhoramento genético de cebola no Centro de Pesquisa da Região Sul em Rio Grande (RS), da Fundação Estadual de Pesquisa Agropecuária (Fepagro), e no Insti- Vol. 3 - No.1 - 2009 19 20 LEITE/OLIVEIRA/SANTOS/COSTA/FONSECA/BOITEUX/DE MELO/REIS/UENO/BAPTISTA tuto Agronômico de Campinas, e no Instituto de Genética da ESALQ/USP, em Piracicaba (SP), em 1940. Diversas cultivares de cebola foram disponibilizadas pela Fepagro com destaque para ‘Jubileu’, ‘Norte 14’, ‘Petroline’ e ‘Madrugada’. Em São Paulo, a seleção para dias curtos resultou na disponibilização das cultivares Baia Periforme Piracicaba do Cedo, Pira Ouro e Pirana, entre outras. Em 1972, com a criação da Empresa Pernambucana de Pesquisa Agropecuária, cebola do tipo ‘Baia Periforme’ foi levada de São Paulo para Belém do São Francisco (PE) e selecionada para latitudes 8 - 9º, ou seja, para bulbificação em dias ainda mais curtos e resistência a condições de calor constante (Wanderley et al., 1973; Candeia e Costa, 2000). Foram disponibilizadas cultivares até a presente data, com destaque para ‘Pêra IPA 4’, ‘Composto IPA 6’, ‘Belém IPA 9’, ‘Franciscana IPA 10’, ‘Vale Ouro IPA 11’ e ‘Brisa IPA 12’. Ainda nas décadas de 1970 e 1980, outros programas de melhoramento genético públicos e privados foram criados para atender às demandas das regiões produtoras, destacando-se a criação do programa da Empresa de Pesquisa Agropecuária e Extensão Rural de Santa Catarina S.A. (Epagri), em Ituporanga (SC), em 1976. Como cultivares de destaque desenvolvidas pela Epagri citam-se ‘Bola Precoce’, ‘Juporanga’, ‘Crioula Alto Vale’ e ‘Superprecoce’. Na Embrapa, os trabalhos de melhoramento genético de cebola foram iniciados em 1977 de forma descentralizada, atendendo as demandas regionais. A cebola, fortemente dependente de fotoperíodo e temperatura para bulbificação e com forte interação com outros fatores ambientais e práticas culturais, requer programas de melhoramento para latitudes ou regiões específicas, sendo a melhor cultivar aquela obtida na própria região de cultivo (Jones e Mann, 1963; Pike, 1986). Atualmente, a Embrapa Clima Temperado, na região Sul, a Embrapa Hortaliças, na região Centro-Oeste, e a Embrapa Semi-Árido na região Nordeste, desenvolvem programas de melhoramento de cebola. Estes programas visam disponibilizar cultivares adaptadas às mais REV. COLOMB. CIENC. HORTIC. variadas condições edafoclimáticas, sistemas de cultivo e preferências regionais, já tendo sido disponibilizadas oito cultivares. A Embrapa Clima Temperado disponibilizou as cultivares Aurora em 1988, Primavera em 1992 e BRS Cascata em 2002. Enquanto a ‘BRS Cascata’ é do grupo das cebolas tardias com bulbos de coloração pinhão-bronzeada e de excelente retenção de escamas, o que condiciona excelente conservação pós-colheita (Leite et al., 2002), as cultivares Aurora e Primavera são do grupo das precoces. Estas cultivares foram importantes para o Rio Grande do Sul, substituindo a população Baia Periforme, que era extremamente desuniforme. Além de adaptadas à região Sul, comportam-se bem nas condições de inverno das regiões Sudeste e Centro-Oeste. A Embrapa Hortaliças disponibilizou as cultivares Conquista, em 1988, São Paulo, em 1991, Alfa Tropical, em 1997 e Beta Cristal, em 1998. ‘Conquista’ é do grupo ‘Baia Periforme’ e possui resistência a míldio na fase de florescimento, ‘São Paulo’ é do grupo das claras precoces, ‘Beta Cristal’ é branca do tipo indústria e ‘Alfa Tropical’ é cebola de verão. A ‘Alfa Tropical’ continua sendo a de maior destaque, pois é a única cultivar de cebola disponível no Brasil para cultivo nas condições de verão das regiões Sudeste e CentroOeste, possibilitando abastecer o mercado no período de entressafra. Além das regiões Sudeste e Centro-Oeste, esta cultivar tem se mostrado como opção para o cultivo no segundo semestre do ano no Vale do Rio São Francisco. Pelas suas características de resistência a doenças foliares e rusticidade, ‘Alfa Tropical’ também vem sendo amplamente utilizada em sistemas agroecológicos de cultivo. O programa de melhoramento de cebola da Embrapa Semi-Árido, iniciado recentemente, já disponibilizou a cultivar BRS Alfa São Francisco para as condições de dias curtos do Vale do Rio São Francisco (Costa et al., 2005). Esta cultivar foi licenciada recentemente para empresas de se- Melhoramento genético de cebola mentes, e por isso ainda não há estimativas do seu impacto no mercado. Por ser derivada da ‘Alfa Tropical’ possui potencial para ser recomendada também para as condições de verão das regiões Sudeste e Centro do Brasil. A pesquisa em melhoramento genético de cebola no Brasil, principalmente os programas públicos, já disponibilizou cerca de 50 cultivares, com ganhos em produtividade, diversidade, adaptação a estresses bióticos e abióticos e possibilitado a modernização dos sistemas de cultivo, tendo contribuído de forma efetiva para o desenvolvimento e sustentação da cebolicultura no Brasil. As cultivares disponíveis no Brasil visam atender as exigências do consumidor brasileiro, que prefere bulbos de tamanho médio (50-90 mm de diâmetro), de formato globular, de película externa de cor bronzeada uniforme, e escamas internas de cor branca (Melo e Boiteux, 2001). A produção de cebola no Brasil continua baseada em cultivares de polinização livre (cerca de 75% da área plantada) com seleções de ‘Baia Periforme’ e ‘Crioula’ dominando o mercado. Possuem, entre outras qualidades, tolerância a doenças, conservação pós-colheita boa e variação ampla em formato, tamanho, cor, número e espessura de películas de bulbos. Cebolas do grupo ‘Crioula’ são adaptadas principalmente à região Sul e são responsáveis pelo desenvolvimento alcançado pela cultura da cebola em Santa Catarina (Leite, 2007). O agronegócio cebola é importante para o Brasil, com previsão, em 2009, de uma área média de plantio dessa cultura de 62.351 ha, com produção de 1.364.098 t distribuídas nas regiões Sul, Sudeste, Nordeste e Centro-Oeste (IBGE, 2009). Estima-se que 70% da cebolicultura brasileira seja proveniente da agricultura familiar, principalmente nas regiões Sul e Nordeste, envolvendo cerca de 60.000 famílias que têm a cebolicultura como atividade principal (Epagri, 2000). O uso de cultivares superiores e de técnicas modernas de produção como irrigação, alta densidade populacional, semeadura direta, mecanização da produção, adubação balanceada, etc. associadas ao uso de sementes de melhor padrão genético vêm favorecendo aumentos gradativos e constantes no rendimento. A adoção de cultivares híbridas associada ao uso de alta tecnologia de produção tem sido fator de aumentos de produtividades, especialmente nas regiões Sudeste e Centro-Oeste e em parte do Nordeste nos últimos anos. Para competir no mercado globalizado de sementes, o país precisará desenvolver bons híbridos com níveis mais altos possíveis de resistência às doenças mais importantes, usando como base populações dos tipos ‘Baia Periforme’ e ‘Crioula’. Cultivares para nichos de mercado específicos, como o de produtos “orgânicos” e produtos “nutracêuticos” e de “cebolas doces”, têm sido buscadas. Como alimento funcional, a cebola é rica em três grupos de compostos com beneficios à saúde humana: flavonóides, tiosulfinatos e frutanas (Bertolucci et al., 2002). Flavonóides são compostos polifenólicos com propriedades antioxidantes (Hollman e Katan, 1997). A cebola é a principal fonte de quercetina na dieta humana, contribuindo com cerca de 30% dos flavonóides consumidos (Hertog et al., 1992). A quantidade de quercetina na cebola varia com a cor e o tipo de bulbo e cultivar, sendo distribuída, principalmente, nas camadas externas (Lombard et al., 2005). Ferramentas de biologia molecular vêm sendo empregadas nos programas de melhoramento da Embrapa e têm sido úteis no conhecimento da distribuição da variabilidade genética e na identificação de mantenedoras da macho-esterilidade. Recentemente foi demonstrado por Imai et al., (2002) que o efeito de lacrimejação da cebola é controlado por uma enzima, denominada “sintase do fator de lacrimejação” - SFL. Esta descoberta abre a possibilidade do desenvolvimento de cultivares sem lacrimejação, porém preservando Vol. 3 - No.1 - 2009 21 22 LEITE/OLIVEIRA/SANTOS/COSTA/FONSECA/BOITEUX/DE MELO/REIS/UENO/BAPTISTA compostos sulfurados de valor nutracêutico, controlados por outro sistema enzimático. Em termos de diversidade e filogenia, o fato de o gene SFL existir como uma cópia abre a possibilidade de utilizá-lo como ferramenta na caracterização e na estimativa de distância genética entre espécies e de acessos dentro das espécies do gênero Allium. Entre os fatores que afetam o rendimento da cebolicultura brasileira estão os danos causados por doenças: antracnose (C. gloeosporioides), manchapúrpura (A. porri) e raiz rosada (P. terrestris) nas regiões Sudeste, Centro-Oeste e Nordeste; míldio (P. destructor) e mofo cinzento (B. squamosa) na região Sul, cujas ocorrências estão relacionadas às condições ambientais do local de cultivo e o nível de resistência das cultivares plantadas (Carneiro e Amorim, 1999). A antracnose foliar é a mais importante doença da cebola em regiões tropicais, afetando a cultura desde a fase de canteiro até o armazenamento dos bulbos, freqüentemente, em proporções epidemicamente alarmantes quando as condições climáticas favorecem (Galván et al., 1997). O controle genético por meio de cultivares com níveis mais altos de resistência a doenças, associado ao controle cultural tem sido buscado como forma de minimização do uso de agrotóxicos e riscos de contaminação de produto e ambiente (Galván et al., 1997). Com base no exposto, este trabalho visa ampliar as informações genéticas básicas já levantadas (Batista et al., 2007; Valêncio et al., 2004; Santos et al., 2007; Santos et al., 2008), bem como identificar marcadores moleculares associados a caracteres de interesse, possibilitando em futuro próximo, o melhoramento assistido por marcadores moleculares. Populações estão sendo melhoradas para sistemas agroecológicos e convencionais, visando a disponibilização contínua de boas cultivares para o segmento de cebolas pungentes, de cebolas suaves/doces, de produtos orgânicos e de alimentos funcionais. REV. COLOMB. CIENC. HORTIC. Materiais e métodos Estão sendo realizadas atividades de pré-melhoramento, que visam auxiliar a identificação de genótipos e genes de interesse dos programas de melhoramento de cebola da Embrapa, através da manutenção e a caracterização de germoplasma mais promissores disponíveis nas coleções de trabalho de cebola da Embrapa e por introdução de novos materiais. Contempla também a caracterização de progênies e/ou populações e/ou híbridos dos programas de melhoramento da Embrapa. A caracterização está sendo realizada com base em caracteres fenótípicos (UPOV, 1999) e agronômicos, marcadores moleculares, através de AFLP (Vos et al., 1995) e de RAPD (Williams, 1990), teores de sólidos solúveis, quercetina (Lombard et al., 2002). e matéria seca, pungência (Schwimmer e Weston,1961), e resistência a doenças. E serão iniciadas atividades de caracterização de germoplasma de cebola para o conteúdo de frutanas (Simonovska, 2000) e para a resistência ao déficit hídrico. A dissimilaridade genética entre os acessos está sendo estimada por métodos estatísticos multivariados (Cruz e Regazzi, 1994). As avaliações quanto a resistência ao míldio (Peronospora destructor), mancha púrpura (Alternaria porri), mofo cinzento (Botrytis squamosa), antracnose (C. gloeosporioides) e raiz rosada (Pyrenochaeta terrestris) estão sendo conduzidas em ensaios de campo e telados, com base na severidade dos sintomas das doenças. Os genótipos mais resistentes são utilizados para continuidade do programa de melhoramento. Um banco de seqüências gênicas está sendo construído visando o isolamento/caracterização de genes potencialmente associados a resistência à C. gloeosporioides e genes das vias metabólicas de compostos organosulfurados, flavonóides e açúcares, que poderão funcionar como marcadores moleculares para fenótipos de interesse, visando a obtenção de cultivares combinando alto potencial de rendimento e resistência a doenças Melhoramento genético de cebola e/ou elevada qualidade nutracêutica e/ou melhor qualidade sensorial, além de elevada capacidade de conservação pós-colheita e com adaptação para cultivo em diferentes latitudes. Com base em marcador para o gene da “sintase do fator de lacrimejação” de modo a verificar a possibilidade de amplificar alelos deste gene em outras espécies do gênero Allium via uma estratégia de PCR heterólogo usando “primers” desenhados para o gene isolado em cebola e analisar a diversidade das seqüências gênicas obtidas, amplificaram-se os segmentos análogos ao gene SFL de cebola a partir de DNA de cinco espécies do gênero Allium: A. cepa; A. chinense; A. fistulosum; A. sativum e A. ampeloprasum, com “primers” (LF 300 e LF 600) sintetizados a partir de informações do cDNA do gene SFL (Imai et al., 2002). Os amplicons foram seqüenciados usando o protocolo BigDye® version 3 e as seqüências analisadas usando o programa LaserGene (DNAStar, Madison, WI). Linhas A (macho-estéreis) e B (macho-férteis, mantenedoras da macho-esterilidade) estão sendo identificadas, em populações e cultivares de cebola para produção de cultivares híbridas e/ou de linhas B para produção de cultivares de polinização livre. Técnicas de biologia molecular disponíveis para caracterização de fatores citoplasmáticos associados a macho-esterilidade estão sendo aplicadas, de forma a acelerar a identificação de linhas B (Havey, 1995; Sato, 1998; Szklarczyc et al. 2002; Engelke et al. 2003). Plantas macho-estéreis identificadas visualmente são pareadas com as macho-férteis (citoplasma N) identificadas com os “primers”, para realização de cruzamentos-testes para identificação de plantas mantenedoras da macho-esterilidade. Está sendo realizada endogamia nos pares de linhas A e B identificados, os quais serão avaliados em esquema dialélico para identificação de linhas com boa capacidade combinatória. Está sendo aplicada seleção recorrente fenotípica e com base em famílias meio-irmãs ou S1 em populações de cebola do programa de melhoramento genético em sistema convencional e em sistema de base ecológica. São considerados a nível de campo: produtividade, ciclo de maturação, uniformidade de estalamento, diâmetro do pseudocaule, resistência a doenças, tamanho e formato de bulbo, número, espessura e aderência da película, ausência de florescimento. Após a cura e armazenamento em galpões ventilados durante pelo menos 30 dias, são feitas avaliações pós-colheitas dos bulbos, considerando-se: capacidade de conservação, pungência, cor, número, espessura e retenção de película seca, e resistência ao brotamento. Após seleção, bulbos são armazenados em câmara fria em condições naturais (região Sul) ou em câmaras frias (regiões Sudeste, Centro-Oeste e Nordeste) até o plantio. Anualmente são produzidas sementes genéticas das cultivares de cebola desenvolvidas pela Embrapa. Havendo o desenvolvimento de novas cultivar(es), serão adotados procedimentos para obtenção do Registro e Proteção das mesmas, atendendo às determinações da Embrapa e seguindo as instruções do Serviço Nacional de Proteção de Cultivares do Ministério da Agricultura, Pecuária e Abastecimento para os ensaios de Distingüibilidade, Homogeneidade e Estabilidade de cultivares de cebola. Estão sendo realizadas atividades de avaliação (unidades de observação e/ou validação) de populações elites e/ou híbridos experimentais de cebola nas regiões produtoras de cebola do Brasil (Sul e/ou Sudeste e/ou Centro-Oeste e/ou Nordeste), de forma a subsidiar a escolha de potenciais cultivares. Resultados e discussão A caracterização da diversidade genética baseada em 22 acessos de cebola do banco ativo de germoplasma de cebola da Embrapa Clima Temperado, por meio de características da película seca (espessura e aderência) e números de pontos Vol. 3 - No.1 - 2009 23 24 LEITE/OLIVEIRA/SANTOS/COSTA/FONSECA/BOITEUX/DE MELO/REIS/UENO/BAPTISTA vegetativos de bulbos, permitiu a separação dos acessos em três grupos, cada um representando uma das três principais populações de cebola brasileiras: Baia Periforme, Crioula e Pêra. Por meio da caracterização da diversidade genética em um grupo de 64 acessos de cebola do banco ativo de germoplasma da Embrapa Hortaliças, foi possível separar os acessos em 12 grupos com a utilização de descritores morfológicos, agronômicos e bioquímicos. Os caracteres que mais contribuíram para a diferenciação foram o teor de açúcares totais, comprimento de bulbo, número de dias para a colheita, porcentagem de bulbos com diâmetro entre 70 e 90 mm e massa média de bulbos, sendo responsável por 58% de toda a variabilidade genética presente entre os acessos. Um grupo de 14 genótipos, incluindo cultivares comerciais, avaliados quanto ao conteúdo de quercetina dos bulbos, através de leituras espectrofotométricas, apresentaram-se altamente variáveis quanto a este flavonóide. Os valores de quercetina variaram de 212,81 mg kg-1 (‘Super Precoce’) até 606,99 mg kg-1 (‘Crioula Roxa’). Em vista das diferenças de concentração de quercetina em cebola ser influenciada pelo genótipo, pretende-se por meio de seleção desenvolver materiais com quantidades superiores de quercetina e, consequentemente, maiores propriedades nutracêuticas. A técnica de RAPD mostrou-se eficiente na estimativa da variabilidade genética de seis cultivares de cebola recomendadas para cultivo na região Sul do Brasil, separando as plantas individuais de cada cultivar em um grupo e a formação de três grupos maiores (‘Baia Periforme’, ‘Crioula’ e ‘Pêra’), correspondendo às principais populações de base das cultivares. Em um outro estudo, com um grupo de 24 cultivares e populações de cebola, o emprego da técnica de RAPD, com a utilização de 53 “primers”, resultou em 102 marcadores, que permitiu o agrupamento dos genótipos de acordo com os grupos morfoagronômicos. REV. COLOMB. CIENC. HORTIC. Com base no marcador AFLP avaliou-se a divergência genética entre populações de cebola potenciais e comerciais para o Nordeste, com a obtenção de fenograma que manteve as relações conhecidas do pedigree dos genótipos avaliados. Houve a formação de três grupos: o primeiro com as populações experimentais de cebola suave as cultivares IPA 10, IPA 11, Brisa e Alfa São Francisco; o segundo com o híbrido Granex 429 e TEG 502 PRR; e o terceiro com as populações experimentais de cebola cascuda bronzeada. Com base no emprego de marcador para o gene codificador da “sintase” do “fator de lacrimejação”, observaram-se amplicons polimórficos entre espécies e entre acessos de A.cepa e A. fistulosum, indicando haver variabilidade natural de pungência entre acessos de cebola. A identificação de marcadores moleculares associados ao fenótipo de cebola com ausência de lacrimejação poderá facilitar a seleção de cebola isentas do desagradável fator “choro”. Com base na caracterização molecular de citoplasma, segundo sistema de marcador descrito por Havey (1995), foram avaliadas 27 populações elites da Embrapa Hortaliças. Dezesseis populações e a cultivar Conquista apresentaram exclusivamente citoplasma N, sete populações apresentaram exclusivamente citoplasma S e apenas três populações apresentaram mistura de ambos citoplasmas. Outras 65 populações foram caracterizadas molecularmente, incluindo os “primers” descritos por Engelke et al., (2003) e Sato (1998), obtendo-se 38 acessos apresentando apenas citoplasma S, 18 exibiram citoplasma N e T e nove apenas citoplasma N. Para a região Nordeste, na Embrapa Semi-Árido, as identificações citoplasmáticas realizadas na cultivar BRS São Francisco e em uma população experimental, apresentaram respectivamente, citoplasma N e T e apenas citoplasma S. Isto indica que para que sejam desenvolvidos híbridos nesta população, se faz necessária a introgressão de citoplasma N. Enquanto para a Região Sul, as caracterizações citoplasmáticas na Embrapa Clima Tem- Melhoramento genético de cebola perado têm sido realizadas principalmente nas cultivares Bola Precoce e Crioula apresentando respectivamente 51,0 e 43,0% das plantas com citoplasma normal, e onde o citoplasma estéril predominante é do tipo T. Os estudos de caracterização da resistência de genótipos de cebola à Coletotrichum. gloeosporioides e Alternaria porri possibilitaram identificar genótipos promissores como fontes de resistência à estas doenças. Dos 65 acessos de cebola da coleção de trabalho da Embrapa Hortaliças avaliados quanto a resistência a C. gloesporiodes, destacaram-se com níveis mais altos de resistência: ‘Pêra IPA 4’, ‘Roxa IPA 3’, ‘Vale Ouro IPA 11’ e a população ‘Alfa Tropical antracnose’. Para A. porri, dos 47 acessos testados por inoculação artificial da doença, destacaram-se com os níveis mais altos de resistência os genótipos ‘Boreal’, ‘Primero’, ‘XP 8010’, ‘Brisa IPA 12’, ‘White Creole’, ‘Crioula Alto Vale’, ‘BRS Cascata’ e ‘Aurora’. Estas populações são, portanto, as mais promissoras para seleção visando o desenvolvimento de cultivares de cebola com maiores níveis de resistência à antracnose e mancha-púrpura. Em um ensaio de campo para caracterização de 10 genótipos de cebola quanto a avaliação agronômica e resistência as doenças mofo cinzento e mancha-púrpura, em sistema de cultivo de base ecológica, destacaram-se as cultivares Petroline e Primavera, em produtividade, tamanho de bulbos e menor susceptibilidade às doenças, as quais seguirão sendo trabalhadas no melhoramento para este sistema de cultivo. Visando a criação de novas cultivares, foram autofecundadas 131 plantas de várias populações de interesse e foram geradas 36 populações base pelos programas de melhoramento da Embrapa, incluindo amarelas baias, cascudas bronzeadas e claras suaves/doces e foram incorporadas mais 37 populações do extinto programa de melhoramento de cebola da ESALQ-USP. Vinte e seis populações foram submetidas à pelo menos um ciclo de seleção massal ou recorrente e estão sendo submetidas a novos ciclos de seleção. Dos 297 pareamentos de plantas macho-estéreis e macho-férteis avaliados foram identificados 26 pares de linhagens A e B. Adicionalmente, 42 pares de linhas A e B, altamente endogâmicos e isogênicos recebidos da ESALQ-USP, também estão sendo avaliados quanto a capacidade combinatória e os primeiros híbridos experimentais produzidos e testados a partir deste ano. Conclusões Para que se consiga realizar melhoramento genético em cebola, assim como em outras espécies, é fundamental a caracterização e preservação de toda a variabilidade genética presente nas cultivares antigas e “landraces”. Os programas de melhoramento genético de cebola de instituições públicas de pesquisa brasileiras têm uma longa tradição de trabalho, sendo responsáveis pelo lançamento da maioria das cultivares em cultivo no país. É importante que estes órgãos continuem recebendo apoio e investimentos para o desenvolvimento e incremento das pesquisas com cebola, com vistas à criação de cultivares cada vez mais competitivas no mercado. Agradecimentos Os autores agradecem a Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul – Fapergs, pelo apoio financeiro. Vol. 3 - No.1 - 2009 25 26 LEITE/OLIVEIRA/SANTOS/COSTA/FONSECA/BOITEUX/DE MELO/REIS/UENO/BAPTISTA REFERENCIAS BIBLIOGRÁFICAS Batista, P.P; R.S.N. de Lima; C.A.F. Santos; M.A.R. Rodrigues e J.C. Alves da S.F. 2007. Divergência genética entre populações de cebola potenciais e comerciais para o Nordeste, com base em marcador AFLP. In: Anais eletrônicos. Congresso Brasileiro de Olericultura 47. Porto Seguro. Campinas: ABH, 2007, http://www.abh.org.br/anais/anais.htm; con­sulta: setembro 2007. Bertolucci, S.K.V.; R.C. Pinheiro; J.E.B.P. Pinto e R.J. de Souza A. 2002. Qualidade e valor nutracêutico da cebola. Informe Agropecuário 23(218), 88-92. Candeia, J.A. e N.D. Costa. 2000. A cebolicultura Nordestina e a necessidade de pesquisa no contexto atual. pp. 15-17. In: Pereira, W.; J.V. VieraA, J.V. e J.L. de Mendonca (Org.). Relatório do Workshop sobre cebolicultura no Brasil. Embrapa Hortaliças. Série Documentos 25. Brasilia. Carneiro, L.C. e J.L. Amorim. 1999. Influência da temperatura e do molhamento foliar no monociclo do mal-de-sete-voltas da cebola. Fitopatologia Brasileira 24(3), 422-427. Costa, C.P., da. 1997. Germoplasma de cebola brasileiro e seu uso no melhoramento. p. 2. In: Anais Seminario Nacional de Cebola 9, 1997. Centro de Pesquisa Agropecuária de Clima Temperado, Pelotas, Brasil. Costa, N.D.; C.A.F. Santos; M.A. de Queiroz; H.M. de Araújo; V.R. Oliveira; J.L. de Mendonca e J.A. Candeia. 2005. Alfa São Francisco: variedade de cebola para cultivo no verão. p. 420. In: Anais Congresso Brasileiro de Olericultura 45, Horticultura Brasileira 23, Fortaleza, Brasil. Galván, G.A.; W.A. Wiestma; S. Putrasemedja; A.H. Permandi e C. Kik. 1997. Screening for resistance to antracnose (C. gloeosporioides Penz.) in Allium cepa and its wild relatives. Euphytica 95(2), 173-178. Havey, M.J. 1995. Identification of cytoplasms using the polymerase chain reaction to aid in the extraction of maintainer lines from open-pollinated populations of onion. Theor. Appl. Genet. 90, 263-268. Hertog, M.G.L.; P.C.H. Hollman e D.P. Venema. 1992. Optimalization of a quantitative HPLC determination of potentially anticarcinogenic flavonoids in vegetables and fruits. J. Agr. Food Chem. 40, 15911598. Hollman, P.C. e M.B. Katan. 1997. Absorption, metabolism and health effects on dietary flavonoids in man. Biomed. Pharmacoterapy 51(8), 305-310. IBGE. 2009. Levantamento sistemático da produção agrícola – cebola: produção e área plantada, Brasil e Unidades da Federação. Rio de Janeiro, In: http:// www.sidra.ibge.gov.br/bda/prevsaf; consulta: julho 2009. Imai, S.; N. Tsuge; M. Tomotake; Y. Nagatome; H. Sawad; T. Nagata e H. Kumaga. 2002. Plant biochemistry: an onion enzyme that makes the eyes water. Nature 419, 685. Jones, H.A. e L.K. Mann. 1963. Onion and their allies. Leonard Hill, London. Cruz, C.D. e A.J. Regazzi. 1994. Modelos biométricos aplicados ao melhoramento genético. UFV, Vicosa, Brasil. Leite, D.L.; A. Garcia e A.M. Santos. 2002. BRS Cascata a new cultivar released by Temperate Climate Agricultural Research Center, Embrapa, Brazil. Abstracts National Allium Research Conference. Washington State University, Pasco, WA. Engelke, T.; D. Terefe e T.A. Tatlioglu. 2003. PCR-based marker system monitoring CMS-(S), CMS-(T) and (N)-cytoplasm in the onion (Allium cepa L.). Theor. Appl. Genet. 107, 162-167. Leite, T.L. 2007. Melhoramento genético de cebola. pp. 79-113. In: Barbieri, R.L. (ed.). Cebola: ciência, arte e história. 2. ed. Embrapa Informação Tecnológica, Brasília. EPAGRI. 2000. Sistema de produção para cebola. Sistemas de Producao 16. Epagri, Florianópolis, Brasil. Lombard, K.; E. Geoffriau e E. Peffley. 2002. Flavonoid quantificationin onion by spectrophotometric and high performance liquid chromatography analysis. HortScience, 37(4), 682-685. França, J.G.E. de e J.A. Candeia. 1997. Development of short-day yellow onion for tropical environments of the Brazilian Northeast. Acta Hort. 433, 285287. REV. COLOMB. CIENC. HORTIC. Lombard, K.; E. Peffley; E. Geoffriau; L. Thompson e A. Herring. 2005. Quercetin in onion (Allium cepa L.) Melhoramento genético de cebola after heat-treatment simulating home preparation. J. Food Comp. Analysis 18(6), 571-581. Melo, P.C.T. e L.S. Boiteux. 2001. Análise retrospectiva do melhoramento genético de cebola (Allium cepa L.) no Brasil e potencial aplicação de novas estratégias biotecnológicas. In: Anais Congresso Brasileiro de Melhoramiento de Plantas 1, 2001. Goiania, Brasil (CD). Melo, P.C.T; A. Ribeiro e M.G.C. Churata-Masca. 1988. Sistemas de produção, cultivares de cebola e o seu desenvolvimento para as condições brasileiras. pp. 27-61. In: Anais Seminario Nacional de Cebola 2, 1988. Piedrade, Brasil. Pike, L.M. 1986. Onion breeding. pp. 357-394. In: Basset, M.J. (ed.). Breeding vegetable crops. AVI Publishing Company, Connecticut, CT. Santos, C.A.F.; D.L. Leite; N.D. Costa; V.R. Oliveira; I.C.N. dos Santos e M.A. Rodrigues. 2008. Identificação dos citoplasmas “S”, “T” e “N” via PCR em populações de cebola no Vale do São Francisco. Hortic. Bras. 26(3), 306-309. Santos, M. do D.M. dos; A.G.R. Buzar; M.E.N. Fonseca; V.R. Oliveira; A.C. Torres e L.S. Boiteux. 2007. Relação genética entre acessos de cebola adaptados para cultivo no Brasil estimada via marcadores RAPD. In: Anais Congresso Brasileiro de Olericultura 47, Porto Seguro, http://www.abh.org.br/anais/anais. htm; consulta: setembro 2007. Sato, Y. 1998. PCR amplification of CMS-specific mitochondrial nucleotide sequences to identify cytoplasmic genotypes of onion (Allium cepa L.). Theor. Appl. Genet. 96, 367-370. Schwimmer, S. e W.J. Weston. 1961. Enzymatic development of pyruvic acid in onion as a measure of pungency. J. Agr. Food Chem. 9(4), 301-304. Simonovska, B. 2000. Determination of inulin in foods. J. AOAC Intl. 83(3), 675-678. Szklarczyk, M.; M. Simlat; B. Jagosz e G. Ba. 2002. The use of cytoplasmic markers in onion hybrid breeding. Cell. Mol. Biol. Lett. 7, 625-634. UPOV. 1999. Guidelines for conduct of test for distinctness, uniformity and stability of Allium cepa and Allium ascalonicum. UPOV, Genebra. Valencio, A.G.B.; A.H. Araújo; L.S. Boiteux; M.E.N. Fonseca; V.R. Oliveira e P.P. Silva. 2004. Estabelecimento de uma coleção de clones genômicos e de cDNA de cebola (Allium cepa L.) contendo seqüências similares a genes de resistência a doenças. p. 39. In: Anais Congresso Brasileiro de Genética 50, 2004. Gramado, Brasil. Vos, P.; S. Hogers; M. Bleeker; M. Reijans; T. van de Lee; M. Hornes; A. Fritjers; J. Pot; J. Peleman; M. Kuiper e M. Zabeau. 1995. AFLP: a new technique for DNA fingerprinting. Nucleic Acids Res. 23, 4407-4414. Wanderley, L.J. da G.; M.A. de Queiroz; P.C.T. de Melo; J.P. de M. Souto; M.A.C. dos Santos; H.M. Silva e D.T. de Lima. 1973. Melhoramento e produção de semente de cebola no Nordeste. Sudene/Brascan Nordeste/IPA. Recife, Brasil. Williams, J.G.K.; A.R. Kubelik; K.J. Livak; J.A. Rafalski e S.V. Tigey. 1990. DNA polymorphism amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 18, 6531-6535. Vol. 3 - No.1 - 2009 27 Mejoramiento genético y producción de “semilla” de ajo (Allium sativum L.). Posibilidades de adaptación a diferentes ambientes Garlic (Allium sativum L.) genetic improvement and seed production. Possibilities of adaptation to variable environments José Luis Burba1 Plantulas de tomate. Ajo INTA’. Foto:‘CASTAÑO Álvarez-Herrera. Foto: INTA, Argentina. RESUMEN Algunos ecotipos de ajo son capaces de florecer y producir semilla botánica, sin embargo la selección clonal sigue siendo la herramienta más válida para aprovechar la abundante variabilidad genética que tiene esta especie y sus variedades botánicas (var. sativum; var. ophioscorodon; var. pekinense). Estas variaciones y la fuerte interacción que tiene esta especie con el ambiente (por su reñida relación con el termo–fotoperiodo) hacen que en el mundo se confundan ecotipos, biotipos, tipos comerciales y cultivares. La selección clonal puede ser masal o individual. La masal es más efectiva desde el punto de vista de la cantidad de “semilla” alcanzada, pero más demorada para encontrar las mejores respuestas. La individual alcanza más rápido los objetivos propuestos, pero es más lenta para alcanzar grandes volúmenes de “semillas”. En el transcurso de seis o siete años puede lograrse una nueva variedad de ajo, capaz de superar en ensayos comparativos a los testigos locales. Este nuevo material, luego de inscribirse formalmente en un registro, pasa a la etapa de producción de “semilla”. Para formalizar un programa de producción de “semilla” de ajo debe contarse con legislación y normas específicas que protejan el patrimonio genético y sanitario de un país. Multiplicaciones seriadas año tras año generan diferentes categorías de “semillas fiscalizadas” (básicas, registradas, certificadas). En el mercado se conocen “semillas mejoradas”, (provenientes de planes de selección), y “semillas saneadas”, (provenientes de sistemas de control sanitario de virosis y otras patologías). Éstas pueden ser inicialmente multiplicadas in vitro y luego por multiplicación convencional, sin embargo existen alternativas intermedias que mejoran los tiempos de disponibilidad y el negocio, como son las multiplicaciones a través de bulbillos aéreos. Conocer a qué grupo ecofisiológico pertenecen las cultivares es imprescindible para predecir la adaptación y el comportamiento en diferentes ambientes. 1 Coordinador Nacional Proyecto Ajo/INTA, Mendoza, Argentina. [email protected] REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 3 - No.1 - pp. 28-44, 2009 M ejoramiento genético y producci ó n de “ semilla” de ajo Palabras clave adicionales: selección clonal, ecotipos, tipos comerciales, cultivares, grupos ecofisiológicos, semilla certificada. Abstract Some ecotypes of garlic (Allium sativum L.) are capable of blooming and producing botanical seeds, nevertheless the clonal selection remains to be the most valuable tool to make use of the abundant genetic diversity that have this species and its botanical varieties (var. sativum, var. ophioscorodon, var. pekinense). These changes and the strong interaction that this crop presents with the environment, such as the one related to thermoand photoperiod, makes that the ecotypos, biotypes, varieties and cultivars are worldwide mistaken. The clonal selection can be mass or individual. The mass selection is more effective from the point of view of the quantity of acquired “seed”, but more delayed in terms of the best responses. The individual selection reaches more rapidly the proposed goals, but it is slower to reach large volumes of “seeds”. In the course of six or seven years, it can be achieved a new variety of garlic, capable of overcoming in comparative essays the local control plants. After formal registration in a record, this new material enters the stage of “seed” production. In order to formalize a program of garlic “seed” production, one must be provided with legislation and specific norms that should protect the genetic and sanitary patrimony of the country. Serial multiplications year after year generate different categories of “fiscal seeds” (basic, registered, and certified). On the market these are known as “improved seeds” (proceeded from selection plans) and “healthy seeds” (proceeded from the systems of sanitary control of viruses and other pathogens). These can be initially multiplied in vitro and then via conventional multiplication, nevertheless there exist intermediate alternatives that improve the period of availability and commercialization, such as the multiplications through aerial bulbils. Knowing the ecophysiological group to which the culture belongs is essential to predict its adaptation and behaviour in different environments. Additional key words: clonal selection, ecotypes, commercial types, cultivars, ecophysiological groups, seed quality certification. Fecha de recepción: 03-04-2009 Aprobado para publicación: 01-06-2009 IntroduCCIÓN El mejoramiento genético del ajo (Allium sativum L.) ha sido motivo de encuentros y desencuentros e historias curiosas. Durante mucho tiempo de la edad moderna y contemporánea nadie creyó en la posibilidad de mejora genética del ajo bajo el argumento que una especie agámica estricta como el ajo no tenía fuentes de variabilidad. Cuando en la década del cincuenta un investigador ruso anunció haber encontrado en un clon la capacidad de dar semilla botánica, otros tantos se encargaron de demostrarlo, ya que nadie lograba que sus plantas produjeran semillas siguiendo su técnica (Burba, 2008). Tuvieron que pasar más de 30 años para que un investigador japonés encontrara “nuevamente” plantas con semilla que casualmente eran del mismo ecotipo ruso ya reportado. Aún hoy no existe en el mercado un cultivar comercial obtenido por hibridación, por lo que la técnica tradicional de selección clonal sigue siendo la principal herramienta. La carencia de reproducción sexual en esta especie y, en consecuencia, la falta de recombinación meiótica limitan la variabilidad natural sólo a la acumulación de mutaciones somáticas, sin que puedan descartarse otras fuentes secundarias de Vol. 3 - No.1 - 2009 29 30 Burba variabilidad de menor frecuencia, como pueden ser las recombinaciones mitóticas o la presencia de tramposones (Lopez Frasca et al., 1997). plasma) hayan acumulado mutaciones, muchas de ellas, favorables, las cuales, pueden ser aprovechadas. Miles de años de acumular pequeños pero significativos cambios (no muy perceptibles por parte del agricultor), acompañados de selección negativa (se venden los bulbos grandes y se guardan para semilla los bulbos chicos), contribuyeron para que el ajo en el mundo llegara a mediados del siglo pasado a su piso más bajo. La selección y la liberación de virus fueron las herramientas para superar esas barreras comerciales. Estas variaciones y la fuerte interacción que tiene esta especie con el ambiente (por su reñida relación con el termo–fotoperíodo) hacen que en el mundo se confundan ecotipos, biotipos, tipos comerciales y cultivares. Por otra parte, hay confusiones en las denominaciones populares, por lo que se requiere corregir esta situación. Hoy existen cultivares con muy bajas y muy altas concentraciones de allicina y de inulina; muy suaves y muy picantes; con y sin aptitud como saborizante de panes; con muy altas y muy bajas concentraciones de selenio; muy precoces y muy tardíos; con y sin fuentes de tolerancia a enfermedades causadas por hongos y virus; con pocas o muchas propiedades antiplaquetarias en sangre; aptos para el pelado, deshidratado o elaboración de fármacos. Sólo para dar un ejemplo, al mismo tipo comercial de ajo en España se los llama “morados”, en Argentina “colorados” y en Chile “rosados”. Pero los “rosados” en Argentina son ajos subtropicales muy diferentes a los chilenos, y los “morados” de ese país son ajos asiáticos de bajos requerimientos de frío que nada tienen que ver con los “colorados” (Burba, 1997a). El mejoramiento genético convencional tiene como objetivos: •El aumento del rendimiento comercial. Si bien en la actualidad algunos ecotipos de ajo son capaces de florecer y producir semilla botánica, la selección clonal sigue siendo la herramienta más válida para aprovechar la abundante variabilidad genética que tiene esta especie (Allium sativum) y sus variedades botánicas (var. sativum; var. ophioscorodon y var. pekinense). Allium sativum var. sativum son los ecotipos más comunes de ajos “rosados”, “violetas”, “blancos” y “colorados”, mientras que A. sativum var pekinense corresponde a los ajos “morados”, mal llamados “chinos”. A. sativum var ophioscorodon son los ajos “castaños”, mal llamados “rusos” o “polacos” (Burba, 2008). La falta de selección sistemática durante siglos hizo que las actuales poblaciones clonales (en manos de los agricultores o de bancos de germo- REV. COLOMB. CIENC. HORTIC. •La ampliación del periodo de oferta (ya sea a través de la precocidad de los materiales o la prolongación de la vida útil pos cosecha). •La concentración de principios organoazufrados (para mejorar las propiedades nutracéuticas o las aptitudes industriales). •El mejor maridaje con las preparaciones culinarias. La estrategia de mejoramiento genético del ajo en Argentina (segundo exportador mundial) apunta a la obtención de un producto diferenciado, con sólidas bases científicas y tecnológicas en los parámetros de diferenciación, estudiando y aprovechando la aún amplia variabilidad natural existente. M ejoramiento genético y producci ó n de “ semilla” de ajo Métodos convencionales de mejoramiento El sistema reproductivo de una especie condiciona la aplicación de los métodos tradicionales de mejoramiento, ya que aquel determina la variabilidad genética original; las posibilidades de generarla; el manejo durante el proceso de selección y la estructura genética de los posibles tipos de cultivares. En muchos de los clones de ajo las flores son total o parcialmente sustituidas por bulbillos, no alcanzando por tanto a formar semillas; en otros no hay ni siquiera formación de escapo floral. Esta menor variabilidad natural y la imposibilidad de generarla por cruzamientos artificiales hacen que el ajo sea una de las especies cultivadas con mayores limitantes en la aplicación de los métodos convencionales de mejoramiento. Las posibilidades se reducen a la aplicación de selecciones sobre los materiales existentes y por tanto el éxito dependerá más que nunca de la variabilidad genética que se encuentre en las colecciones de trabajo. De ahí la importancia de apoyar la formación de bancos de germoplasma. Complementariamente a estas ideas, es conveniente señalar que la variabilidad somaclonal constituye, en esta especie, una interesante estrategia para la generación de variabilidad genética seleccionable (López Frasca et al., 1997). Se comprobó que plantas de ajo regeneradas a partir del cultivo in vitro de tejidos (callos) mostraron marcada variabilidad para características fenotípicas como altura de planta, número de hojas, peso y forma de los “dientes” (bulbillos), características morfológicas de las “cabezas” (bulbos), coloración de hojas envolventes y presencia de bulbillos aéreos, a partir de las cuales fue posible la selección de somaclones sobresalientes que posibilitaron la creación de nuevos cultivares. Para evaluar las posibilidades de aprovechar la variación fenotípica existente en una población clo- nal es necesario determinar cuánto de ella es de origen genético y cuánto ambiental. Los parámetros genéticos que expresan esta relación son dos: • Heredabilidad en sentido amplio (h2) o grado de determinación genética: h2= VG/(VG+VE) VG = variancia genética VE = variancia ambiental •Coeficiente de variación genética (CVG): (CVG) % = (√ VG √ X)* 100 X= media √VG = desvío de la variancia genética Además, resulta útil reunir información acerca de los grados de asociación de los distintos caracteres, lo que se estima a través de correlaciones fenotípicas (rP) y genotípicas (rG). Diversos autores han estimado la variabilidad genética de caracteres de interés comercial, sus correlaciones y su contribución como componentes del rendimiento (López Frasca et al., 1997). En general, los resultados son coincidentes en estimar mayor heredabilidad en peso de bulbo, peso de bulbillos, número de bulbillos y altura de planta. En cuanto a las correlaciones genéticas, se encontraron valores altos y positivos entre diámetro y peso de bulbo, mientras que la contribución directa de diámetro y peso de bulbillos al rendimiento tuvo una respuesta variable de acuerdo con el ciclo de evaluación. En Argentina, en colecciones de ajo tipo “rosado” (GE II), cultivado en la provincia mediterránea de Córdoba (centro del país), y en colecciones de los tipos “blanco” (GE III) y “colorado” (GE IVa), cultivados en la provincia de Mendoza (oeste del país), se encontró una apreciable variación genética, lo que confirma que estos tipos por su origen y estructura deben ser consideradas como poblaciones o “variedades criollas”. Vol. 3 - No.1 - 2009 31 32 Burba En el ajo tipo “rosado”, la heredabilidad en peso, perímetro y número de bulbillos por bulbo, al igual que número de hojas fértiles, indicó efectos ambientales preponderantes, aunque la mayor variabilidad aprovechable por selección (variabilidad genética) fue estimada en número y peso de bulbillos. También se determinó que el número y peso promedio de bulbillos por bulbo tuvieron efectos directos sobre el rendimiento y fueron los componentes de principal importancia. Los resultados en ajo tipo clonal “blanco” confirmaron mayores posibilidad de selección en el carácter, peso y número de bulbillos y escasa variabilidad en número de hojas fértiles, en consecuencia poca oportunidad de selección. Considerando el alto grado de asociación a nivel fenotípico y genotípico que se presentó entre peso y diámetro y los valores heredabilidad, se previó un mayor avance en la selección de diámetro a través de la selección indirecta por peso. mayor a 2 gramos). Durante el cultivo se pueden eliminar plantas fuera de tipo y a partir de la cosecha se vuelven a repetir los pasos descritos. De esta manera, en 4 ó 5 años se puede contar con muy buen material, bastante homogéneo y adaptado a las condiciones locales. La selección individual (que por lo general la realizan los organismos oficiales) alcanza más rápido los objetivos propuestos, pero es más lenta para obtener grandes cantidades de “semillas” disponibles. A partir de una población local se estudia la frecuencia de caracteres deseables (formas, colores, calibres, número de dientes, etc.) y se define el ideotipo. Se eligen todos aquellos bulbos que respondan al ideotipo y cada bulbo constituye una familia. Los dientes de cada bulbo se plantan en una parcela individual. El número de familias no debería ser inferior a cien. La existencia de variación genética para número de bulbillos se confirmó por la respuesta a la selección, que les permitió obtener clones con distribuciones muy diferentes. En ensayos entre clones experimentales de ajo “colorado” se verificó una mayor variación en peso y una alta, positiva y significativa (P≤0,01) correlación fenotípica entre diámetro y peso, que osciló entre 0,939 y 0,715 (López Frasca et al., 1997). Durante el cultivo se analiza el comportamiento de cada familia y se eliminan todas aquellas que muestran anormalidades indeseables (síntoma de virosis, plantas dobles, etc.). Luego de la cosecha se analiza para cada familia el rendimiento (diámetro, peso y PER: peso específico relativo), y con una presión de selección aproximada al 50% se eliminan todas aquellas familias con mal o regular comportamiento (Burba, J.L., 1997b). La metodología clásica de selección clonal puede ser masal o individual. La selección masal (que la puede realizar cualquier agricultor esmerado) es más efectiva desde el punto de vista de los volúmenes de “semilla” alcanzados, pero más lenta para encontrar las mejores respuestas (Burba, 1997b). Repitiendo este proceso, en el transcurso de 6 ó 7 años puede lograrse una nueva variedad de ajo, capaz de superar en ensayos comparativos a los testigos locales. Este nuevo material, luego de inscribirse formalmente en un registro, pasa a la etapa de producción de “semilla”. Se seleccionan importantes cantidades de bulbos “ideales” (buena forma, sanidad y compacidad), se calibran y eliminan los de diámetro más pequeño (los menores de 45 o 50 mm). Los bulbos se desgranan, los dientes se clasifican y se plantan solamente los de mayor peso (por ejemplo, Producción de semilla REV. COLOMB. CIENC. HORTIC. Para formalizar un programa de producción de “semilla” de ajo, debe contarse con legislación y normas específicas que protejan el patrimonio genético y sanitario de un país (Burba, 1990). M ejoramiento genético y producci ó n de “ semilla” de ajo Como se mencionó,, el ajo carece en la práctica de semilla verdadera, y por tanto se utilizan los “dientes” de los bulbos como órgano de multiplicación. Estos “dientes” acarrean por lo general serios problemas de plagas (nemátodos y eriófidos) y enfermedades (hongos, fitoplasmas y virus) éstas por tanto se transmiten de generación a generación, lo que obliga a organizar un sistema especializado de producción de “semilla”. Para el común de la gente, existen varios “tipos” de “semillas”, sin embargo no siempre son claras las diferencias entre ellas. Hay una gran confusión en las denominaciones debido a lo complejo del sistema de producción. Es importante entonces conocer realmente qué alcance y utilidad tiene cada una de ellas y su denominación correcta (Burba, 1993). Se utilizan, entre otros, los siguientes términos aplicados a la “semilla”, los que no siempre están bien utilizados: y aunque los resultados de los análisis sanitarios sean aceptables, estas "semillas" tienen escasa pureza genética. La “semilla fiscalizada” es la que controla el estado a través de los organismos especializados, la cual tiene distintas categorías según su nivel de pureza y calidad: básica, registrada y certificada. Por lo general, solamente esta última es utilizada por los productores de ajo para consumo, mientras que las categorías anteriores son utilizadas por los “semilleros”. Las básicas y registradas son las denominadas “semillas saneadas” o “semillas libres de virus”, aunque estos términos merecen mayor explicación. Las categorías básicas y registradas provienen de un proceso de laboratorio, en el cual se eliminan total o parcialmente los virus que posee. Es exagerado hablar de “semillas libres de virus” ya que es muy difícil asegurar que no contienen ninguna partícula de viral, por eso suelen llamarse “semillas saneadas” o “semillas de sanidad controlada”. •Común o identificada. •Fiscalizada (básica, registrada o certificada). •Saneada o libre de virus. •Original. Existe una fuerte tendencia por parte del productor o empacador a utilizar (o vender) los descartes del ajo de consumo como “semilla”, sin embargo las diferencias entre ambos productos son grandes: el punto de cosecha, la pureza genética, la sanidad, el tamaño de los “dientes” y la temperatura de conservación, sólo para mencionar algunas. El ajo de consumo podría transformarse en “semilla común” si es sometido a ciertos análisis de laboratorio y estos arrojan resultados favorables. Por tanto, su propietario, bajo su más absoluta responsabilidad, podrá utilizarla o comercializarla como “semilla identificada”. Por lo general, Las “semillas saneadas” en los laboratorios se re contaminan en el campo en las sucesivas multiplicaciones, y con el paso del tiempo vuelven a adquirir los virus (transmitidos por pulgones y eriófidos), aunque en proporciones menores que las “semillas” crónicamente enfermas que nunca pasaron por este proceso. Por esta razón, sólo se controla de virus a las básicas y registradas y no a las certificadas. Por lo anterior, se considera que existen en el mercado dos tipos de "semillas" certificadas: •Aquellas que provienen de un sistema de saneamiento de virus (aunque ya estén parcialmente contaminadas). •Las que no provienen de un sistema de saneamiento, aunque sí de selección y control de otro tipo de problemas (como nemátodos, eriófidos y algunos hongos). Vol. 3 - No.1 - 2009 33 34 Burba Las primeras siempre tienen un mayor potencial de rendimiento. La semilla (saneada o no) puede ser de variedades “impuras” (llamadas poblaciones clonales), es decir, son mezclas de varios tipos aunque se parezcan mucho, o de variedades “puras” (llamadas mono o policlonales). Estas últimas provienen de un programa de selección genética. La “semilla madre” que se entrega a los “semilleros” para su multiplicación se denomina comúnmente “semilla original”. También se confunden los establecimientos “criaderos” de los establecimientos “semilleros”. Los primeros son los que producen nuevas variedades a través de los programas de mejoramiento genético (y producen “semilla original”), mientras que los segundos son los responsables de multiplicar ésta. Si bien existe un mercado potencial para la exportación de "semillas de ajo", esta situación es muy delicada debido a la gran especificidad de adaptación que tienen las variedades. Sólo países del hemisferio sur con condiciones similares podrán importar "semilla". La venta y posterior adaptación transhemisférica, si bien no es imposible, es un negocio de mediano a largo plazo. Las condiciones de los valles aislados y frescos tienen las mejores posibilidades de producir "semilla" de alta calidad. Si bien es deseable que existan zonas especializadas en la producción de semillas de ajo (un poco más frías que las zonas de producción de ajo para consumo), suele ser difícil su organización y protección del sistema semillero. Cuando la situación anterior no es posible, se aconseja que cada productor o grupo de productores destinen un área especial de aproximadamente el 15% de lo que cultiva con ajos para consumo y produzca su propia semilla de tercera o cuarta multiplicación a partir de semilla básica adquirida en los establecimientos especializados. REV. COLOMB. CIENC. HORTIC. Por ejemplo, si un productor cultiva cinco hectáreas para consumo necesitará destinar unos 6.000 m2 para producir su semilla de tercera multiplicación, unos 800 m 2 para su semilla de segunda multiplicación, y 100 m 2 para su semilla de primera, por lo que se verá obligado a adquirir una pequeña cantidad de semilla básica. Partiendo de la premisa que la “semilla” es la principal responsable de los altos rendimientos comerciales, un programa “semillero” será obligatorio para cualquier zona productora. Ella sola explica más del 70% del rendimiento potencial a alcanzar. Manejo de semilla básica Con el avance de la tecnología de producción de semilla de ajo, ya nadie duda sobre la importancia de la calidad de la misma, principal componente de los rendimientos comerciales del cultivo. El atributo calidad en semilla de ajo implica dos conceptos: calidad genética, es decir, alta pureza varietal, y calidad sanitaria, es decir, libre de patógenos sistémicos (Burba, 1990; Moriconi et al., 1991). La semilla de alta pureza varietal (cultivares inscriptos por criaderos y producidos por semilleros inscritos) garantiza fundamentalmente un gran potencial de rendimiento comercial y gran uniformidad, no sólo en las características agronómicas (periodo de dormición, simultaneidad de emisión de vara floral, homogeneidad de punto de cosecha, etc.), sino características comerciales (grandes calibres, formatos uniformes, compacidad, etc.). La semilla de alta sanidad (producida por laboratorios y viveros prestigiosos) garantiza estar libre tanto de patógenos como el causante de la “podredumbre blanca” (Sclerotium cepivorum y Sclerotium rolfsii), o de la “tristeza” (fitoplasma), M ejoramiento genético y producci ó n de “ semilla” de ajo como de nematodos (Ditylenchus dipsaci), o saneados de virosis de alta peligrosidad (OYDV, LYSV, entre otros). Esta semilla se obtiene en diferentes etapas (tabla 1), en las que, durante varios años, participan criaderos, laboratorios, viveros y multiplicadores (Burba, 2009). La semilla básica puede salir del laboratorio en forma de micro plantas o microbulbillos (obtenidos in vitro), o minibulbillos rusticados, los que deben continuar las sucesivas etapas de multiplicación en jaulas con mallas a pruebas de pulgones (jaulas antiáfidos), ya que son estos insectos los vectores de los principales virus. Los propágulos (micro plantas, micro o minibulbillos) provenientes del laboratorio deben cultivarse en condiciones agroclimáticas similares a las requeridas por la variedad (cultivar), es decir, capaces de cumplir con los requerimientos termo y fotoperiódicos de ésta, pero en ambientes aislados como son las jaulas antiáfidos de alta seguridad.Una vez cumplida la primera multiplicación en jaulas fijas antiáfidos, es conveniente, desde el punto de vista económico, realizar la o las etapas siguientes en estructuras más económicas, instaladas directamente en suelo agrícola, denominadas túneles antiáfidos. Los bulbillos aéreos de ciertas variedades (morados, colorados o castaños) pueden multiplicarse en canteros en forma de almácigos (microtúneles antiáfidos), bajo cobertura de una malla antiáfidos o manta térmica. Posibilidades de adaptación Se entiende por adaptación la capacidad que tienen los organismos para modificar su comportamiento ante una nueva situación y se acepta que ellos tienen mecanismos capaces de lograrlo (Burba, 1997a). El ajo, originario de zonas con inviernos rigurosos y fotoperiodo primaveral largo (alrededor de 40° N), se ha dispersado adaptándose hasta en las regiones tropicales, sin embargo el “precio” de esta adaptación se evidencia por la modificación de la pungencia, conservación poscosecha y calidad estética (gran número de dientes asociado a deformaciones o escaso número de dientes asociado a bajos rendimientos). Actualmente existe tecnología suficientemente probada para contribuir a que este proceso de adaptación tenga éxito en el corto o mediano plazo. El esquema agroecologista (más conservador y tradicional) consiste en introducir materiales, estudiar el manejo agronómico y efectuar selección clonal Tabla 1. Categorías y subcategorías de ajo semilla de la clase fiscalizada. Ámbito de uso Categoría Básica Criaderos Viveros o semilleros Registrada Productores de ajo de consumo Certificada Subcategoría Denominación Pre inicial M0 Inicial M1 Prefundación M2 Fundación M3 Registrada 1 M4 Registrada 2 M5 Registrada 3 M6 Certificada M7 en adelante Fuente: Burba et al., 2009. Vol. 3 - No.1 - 2009 35 36 Burba para lograr nuevos ecotipos. Éstos, alcanzados en el mediano a largo plazo, podrán obtener altos rendimientos, aunque no aseguren siempre alta rentabilidad. El esquema productivista (más audaz) posee las mismas dos etapas anteriores, pero reemplaza a la tercera por la utilización de tecnología más o menos sofisticada que modifique el comportamiento del cultivo en alguna de sus etapas. Los resultados, logrados en el corto o mediano plazo, quizás sacrifiquen rendimiento en campo, pero ofrecen la posibilidad de mejorar la rentabilidad por la oportunidad de la oferta. La pérdida de rendimiento está generalmente asociada con una mayor precocidad. Ambos esquemas combinados podrán ser herramientas valiosas para resolver los problemas de adaptación en una región determinada. Estrategias de adaptación En este proceso se utilizan dos vías: manejo del genotipo y manejo fisiológico. El conocimiento profundo de ambos aspectos facilitará las acciones. Poblaciones clonales, variedades y cultivares Prácticamente todos los cultivos de ajo del mundo están realizados con base en poblaciones clonales, producto de mezclas mecánicas y ausencia de programas de mejoramiento genético. El término poblaciones clonales encuentra resistencia para su adopción, sin embargo refleja mejor la realidad. Denominaciones como “biotipos”, “ecotipos”, “variedades”, “genotipos”, “tipos clonales”, “tipos comerciales” sólo confunden la interpretación de resultados. Gran parte de la bibliografía existente, con resultados aparentemente erráticos, es el producto de haber utilizado poblaciones clonales no descritas o poco conocidas. Las verdaderas “variedades cultivadas” o cultivares (genotipos), producto de planes sistemáticos de mejoramiento genético, podrán ser mono o policlonales, dependiendo de si REV. COLOMB. CIENC. HORTIC. se utilizó como modalidad la selección individual, la selección masal, o ambas. El conocimiento de los requerimientos ecofisiológicos de éstos permitirá tomar decisiones sobre las oportunidades de adaptación de dichos cultivares (Burba, 1997a). Relación genotipo x ambiente En una especie tan sensible como el ajo al termo y fotoperiodo (en ese orden), cuyo periodo de cultivo (C) y el de dormición (D) son fuertemente influenciados por el ambiente, debe (al menos teóricamente) cumplirse la ecuación: C + D = K, donde K = 365 días Cuando se introducen cultivares de otras regiones (en o fuera del hemisferio en que nos encontramos) y esta sumatoria no se cumple, podemos decir que los cultivares no se adaptan y por tanto requieren de la modificación de las condiciones de manejo. Si la sumatoria C + D < 365, como es el caso de cultivares subtropicales cuando son llevados a regiones más frías y de fotoperiodos primaverales más largos, resta un periodo del año (desde la cosecha hasta el momento óptimo de plantación), en el cual probablemente la “semilla” sufra deterioros y comprometa el rendimiento. Si la sumatoria C + D > 365, como es el caso de cultivares de mucho requerimiento de frío y fotoperiodo largo cuando se llevan a regiones subtropicales, le sobra una fracción de tiempo que obligaría a plantar siempre tarde y por tanto con menores rendimientos. La adecuación de la época de plantación y el manejo del periodo de dormición (mediante el almacenamiento de la “semilla” en una relación tiempo–temperatura ideal para cada caso) serán las posibles soluciones para la adaptación del cultivar introducido. Por otra parte, y teniendo en cuenta que el requerimiento fotoperiódico puede verse modificado por la cantidad de frío inductor recibido, podemos adaptar cultivares provenientes de regiones más frías a regiones subtropicales de altura. Los M ejoramiento genético y producci ó n de “ semilla” de ajo índices latialtimétricos de adaptación pueden ayudar a tomar decisiones. Así, cultivares de buen comportamiento a 35º de latitud a 700 msnm pueden tener un comportamiento similar a 20º de latitud, pero a 2.500 msnm. Manejo del periodo de dormición como estrategia Relación temperatura y tiempo de almacenaje Son numerosas las experiencias realizadas aplicando bajas temperaturas a la “semilla” en la búsqueda de diferentes respuestas. La mayoría de los resultados alcanzados demuestran que, mientras más bajas sean las temperaturas y más largos los tiempos de almacenaje, los bulbos producidos disminuyen su calidad comercial o su productividad total, sin embargo hay resultados favorables que podemos aprovechar en un sistema “semillero”. Las respuestas directas o indirectas que podemos encontrar con el uso de las denominadas “frigoplantas” seguramente están condicionadas por el genotipo, las condiciones de almacenamiento, el estado fisiológico del órgano que recibe frío y las condiciones de campo a que serán sometidas. Efectos sobre la dormición y brotación El balance entre promotores e inhibidores y las proporciones relativas entre ellos controlarían la entrada y salida de dormición, y el efecto del frío en preplantación serviría de “gatillo” para modificar este balance en favor de los promotores (auxinas, giberelinas, citocininas) y en contra de los inhibidores (ABA o compuestos relacionados). A este último proceso se lo denomina frigoinducción (“choque de frío”, “golpe de frío” o “vernalización”). A la semilla tratada se la denomina frigoinducida y a las plantas logradas se las llama frigoplantas (Burba, 2009). El ajo requiere para bulbificar o “encabezar” dos condiciones: tomar cierta cantidad de frío y cre- cer con días largos. Existen variedades que tienen escasos requerimientos de frío y longitud del día (como es el caso de los “rosados”, GE II), y otras que tienen altos requerimientos, como es el caso de los ajos “colorados” (GE IVa). Mientras más frío toma, los requerimientos de largo del día son menores. El aporte de frío artificial a la “semilla” que lo recibe “a cuenta” del frío invernal. La frigoinducción se utiliza con diversos objetivos: •Lograr precocidad en la cosecha de las mismas variedades utilizadas en la región. •Adaptar ajos de zonas frías a zonas tropicales. •Escalonar la cosecha. Este tratamiento preplantación con bajas temperaturas a la semilla de ajo es una práctica antigua, utilizada para aumentar la velocidad de emergencia y acortar el ciclo vegetativo de las plantas en búsqueda del objetivo comercial de lograr primicias, aunque esto siempre está asociado con la caída de rendimientos y la aparición de disturbios fisiológicos como el “rebrote”. La base teórica se estudió a fines de la década del cincuenta, sin embargo, pasaron más de 30 años para que se promoviera como alternativa práctica y más de 50 años para que fuese utilizada como técnica de rutina. A inicios de la primera década del siglo XXI, todavía se presenta como una tecnología de uso restringido, ya que a la enorme cantidad de factores agronómicos que interactúan se le agrega el mal manejo de las cámaras frigoríficas. En muchas regiones productoras de ajo en el mundo se trata con frío la “semilla” (proceso denominado vulgarmente “vernalización”), simplemente para darle mayor precocidad a las variedades típicas de la zona. Esa precocidad casi siempre está acompañada por menores rendimientos y aumento de deformaciones. Vol. 3 - No.1 - 2009 37 38 Burba En regiones tropicales de Brasil, donde sólo es factible cultivar ajos de poca calidad (por lo general pequeños, de muchos “dientes” y de escasa conservación), se utiliza el almacenamiento frigorífico de la “semilla” de variedades de mayor calidad, provenientes de regiones más frías (bulbos medianos a grandes, pocos “dientes”, pero más grandes y de buena conservación). En algunas zonas de Australia, Egipto o Corea, se dispone de suficiente experiencia para brindarle a la “semilla” diferentes “dosis” de frío, que producen bulbificación en distintos momentos, y de esa manera permiten escalonar la cosecha, disponiendo de programas de computación que permiten aportar el frío “exacto” en función del clima de una determinada campaña agrícola. En Argentina, los primeros antecedentes datan de mediados de la década del setenta en Córdoba sobre ajos del tipo “rosado”, sin embargo es una práctica que no se difundió. Consistía en tratamientos entre 5ºC durante 60 días, lo que permitía acortar el ciclo del cultivo de 210 días a 150 días. Por otra parte, frigoinducir y plantar temprano no arroja los mismos resultados que realizar el tratamiento y plantar tarde. Asimismo, no todas las variedades de ajo reaccionan ante el estímulo de la misma manera. Mientras más frío toma “se hace menos exigente al largo del día” (los requerimientos de largo del día son menores) y bulbifica o “encabeza” antes. Esta es una razón para aportarle frío artificial a la “semilla”, que lo recibe “a cuenta” del frío invernal. La “semilla” almacenada en frío produce varios resultados, dependiendo de la temperatura, del tiempo de almacenamiento, de la variedad, del estado de dormición que tenga el brote, de la fecha de plantación, etc. Estos efectos no siempre son deseables y ese es el “precio” que se paga cuando se modifican artificialmente a las plantas. Como ya quedó expresado, se debe distinguir al “choque” de frío (que por lo general es el tratamiento a temperaturas inferiores a los 10ºC) de las temperaturas de almacenamiento de “semillas”, que ocurre entre 14ºC y 18ºC. Estas últimas tienen ventajas como la mayor sanidad, el mejor estado de hidratación o la uniformidad de la brotación y generalmente no poseen desventajas. Entre los efectos deseables se pueden mencionar la brotación rápida (ya que el frío en “semilla” rompe el estado de reposo), la formación prematura del bulbo (ya que haber recibido frío la hace menos dependiente del largo del día) y, como consecuencia de esto, la cosecha anticipada. Entre los indeseables se encuentran los bajos rendimientos y la manifestación de deformaciones (“rebrotes”), que comprometen seriamente la comercialización. Estos efectos negativos son tanto más graves cuanto más bajas sean las temperaturas de almacenamiento y más largo el período del mismo. Acortarle la vida al cultivo de ajo para obtener primicias mediante alguna técnica implica por lo general pérdidas de rendimiento, aunque el mejor precio del mercado en ese momento puede compensarla ampliamente. La frigoinducción permite al diente semilla “tomar frío a cuenta del invierno”, sin embargo, y debido a las diferencias térmicas en dicha estación entre año y año, hace que muchas veces los resultados sean erráticos. Existe para cada región una y sólo una combinación exitosa entre variedad, tiempo y temperatura de almacenamiento, momento en que se realiza el tratamiento y época de plantación (figura 1). A pesar de ello, las cambiantes condiciones meteorológicas entre año y año modifican también esta combinación. Por esta razón, sólo en algunas regiones y después de muchas pruebas se consigue la combinación perfecta. REV. COLOMB. CIENC. HORTIC. M ejoramiento genético y producci ó n de “ semilla” de ajo Fuente: Burba, 2009. Figura 1.Interrelaciones de los factores que afectan la producción de frigoplantas. Como ya es conocido, los grandes grupos varietales de ajo tienen diferentes requerimientos de frío para bulbificar, por esta razón hoy existen variedades adaptadas a las diversas condiciones agroecológicas, desde regiones tropicales a nivel del mar hasta regiones frías a más 2.000 msnm. La tabla 2 muestra de manera sintética la relación entre los grandes grupos varietales y sus requerimientos de frío. Los grupos varietales En el mundo moderno se reconocen ajos de todo tipo, sin embargo la selección natural que la especie ha sufrido, las denominaciones populares en las diferentes culturas, los intentos de los investigadores por agruparlos y los planes de mejoramiento genético han complicado el panorama y su interpretación. Los ajos pueden ser agrupados por sus características botánicas, fisiológicas o comerciales. Los intentos por hacerlo no fueron muy exitosos, complicándose aún más la situación debido a las barreras idiomáticas. En la práctica comercial los ajos se denominan en algunos casos según el color de los bulbos, o el color de los “dientes” o bulbillos en otros. Este panorama es más complejo cuando de país en país las tonalidades reciben diferentes denominaciones. En español, los mismos tipos comerciales pueden denominarse rosa, rosado, rojo, violeta, morado o colorado. En portugués se denominan roxo y roxao. En italiano, rosso y rosa, y en francés, rose, rosé, rouge y violet. Los grupos ecofisiológicos de Argentina, distinto para Francia, China o Japón, no serán motivo de este análisis, sin embargo, las mencionaremos, ya que dichas clasificaciones guardan fuerte relación con otros tipos de agrupamientos, generando muchas veces mayor confusión. Vol. 3 - No.1 - 2009 39 40 Burba Tabla 2.Relación entre cultivares y requerimientos de frío. Tipo comercial Grupo ecofisiológico Cultivares Requerimientos de frío Rosados II Alpa Suquía Muy bajos Morados II Morado INTA Bajos Violetas III Lican INTA Medios Blancos Norteño INTA Nieve INTA Unión Perla INTA Plata INTA Tempranillo Gostoso INTA Fuego INTA Sureño INTA Rubi INTA III Colorados IV a Castaños IV b Medios Altos Castaño INTA Muy altos Fuente: Burba, 2009. Variedades botánicas A la especie Allium sativum L., denominada ajo común, ajo doméstico o ajo de huerta, se le conocen principalmente tres subespecies (botánicamente llamadas variedades), que son pekinense, sativum y ophioscorodon (tabla 3). Según algunas escuelas populares, algunas de estas variedades estarían “especializadas” en producir bulbillos aéreos y otras en bulbillos subterráneos, aunque en rigor de verdad, según las condiciones ambientales de cultivo o las temperaturas del almacenamiento de la “semilla”, las variedades pueden producir los dos tipos de propágulos, tal es el caso de ajos “blancos”, que por lo general no emiten vara floral, pero sí lo hacen en regiones muy frías, y por consiguiente la denominación hardneck (de cuello duro o con vara floral) o softneck (de cuello blando o sin vara floral) es también relativa. Ajos de cuellos duros y de cuellos blandos En algunos ambientes de los Estados Unidos, a la var. sativum se la denomina popularmente como softneck, ya que interpretan que florece parcialmente o no lo hace, y por tanto presentan el “cuello blando fácil de trenzar” en el momento de la cosecha. Englobaría a todos aquellos ecotipos de abundante número de hojas fértiles y relativamente muchos “dientes”. Tabla 3.Equivalencia probable de las denominaciones vulgares de grupos varietales de ajo de “cuello duro” (hardneck). Variedad botánica Según Burba Según England pekinense asiáticos (morados) asiáticos ophioscorodon rusos (castaños) continental sativum mediterráneos (colorados) rocambole Fuente: Burba, 2008. REV. COLOMB. CIENC. HORTIC. M ejoramiento genético y producci ó n de “ semilla” de ajo Denominan variedades de “cuello duro” (hardneck) a las que interpretan que siempre tienen tallo floral emergente y por tanto en el momento de la cosecha mantienen dicho cuello duro. Se consideran como var. sativum: los “violetas” (del tipo Violeta de Cadours o Germidour de Francia); los “blancos” (del tipo California Early de Estados Unidos), y también los “colorados”, sean tanto ecotipos de “cuello duro” como de “cuello blando”. Ajos rosados y morados Una misión científica china, radicada durante muchos años en la Estación Experimental de Caacupé (Paraguay), habría sido la responsable de la introducción de muchas variedades subtropicales (Fen Sang No. 1 y No. 2, entre otros). Éstos pertenecen al Grupo Ecofisiológico II de Argentina. Una población denominada “nacional” en Paraguay sería la misma denominada “rosado paraguayo” en Argentina. Existen en Brasil numerosas poblaciones “asiáticas” o “subtropicales”, como Amarante, Contestado y Lavinia; en Francia, estos grupos estarían representados por Rouge d’Afrique y Mallorí; en Japón, por Ishuwase y Okinawa, mientras en Italia, por Rosa di Napoli. Estos muestran plantas muy vigorosas, de cosecha temprana, bulbos grandes con muchos dientes pigmentados con estrías moradas, globoso piriforme, vara floral no emergente (queda incluida en el falso tallo pero puede presentar algunos bulbillos laterales grandes sobre el escapo), de poca conservación. En Argentina, de la población denominada “paraguayo”, se seleccionó Alpa Suquía y, probablemente, la variedad Rubí 1 de Colombia derive también de aquellas. Los “ajos morados”, mal llamados “ajos chinos”, ya que en rigor son muchos y variados los procedentes de ese país, son poblaciones de plantas muy vigorosas, de cosecha temprana, bulbos grandes, pigmentados con estrías moradas, globosos achatados, de escasa conservación, poco ajustados en el cuello y “dientes” de color castaño claro. En Argentina, los ajos “asiáticos” o “morados” (como la cultivar Morado INTA, similar a Xi’an de China), y los “rosados” (del tipo “rosado paraguayo”) pertenecerían a la variedad botánica pekinense, aunque no hay coincidencia entre los autores. En Francia, Blanc d’Egypte (del que se seleccionó Ramsès), y un grupo de asiáticos precoces (de los que seleccionó Sprint), junto con los conocidos en Estados Unidos como Beijing, Sahntung Purple o Asian Tempest, pertenecerían también a esta variedad botánica. Ajos blancos y violetas Son ajos medianos a grandes, chatos, de color externo blanco o blanco amarillento, eventualmente con tintes violáceos, mediana cantidad de “dientes” (entre 8 y 14), de color lechoso o con leves tintes rosados o parduscos, distribuidos en varias hojas fértiles (entre tres y cinco), de cosecha temprana o semitemprana y en general de buena capacidad de conservación. En Estados Unidos distinguen tres subtipos: artichoke, silverskin e italian. Los del subtipo artichoke presentan bulbos que muestran las puntas de bulbillos abiertos en la parte superior, similar a un alcaucil, del que provendría su nombre. La aparición de síntomas moderados de “rebrote”, prolongando las hojas envolventes adicionales de los “dientes”, le da un aspecto que también podría asemejarse a las brácteas del alcaucil. En Argentina se pueden distinguir variantes como “blancos nativos”, “blancos americanos” y “violetas”. Son características de los “blancos nativos” las cultivares argentinas Blanco Mendoza, Nieve INTA, Norteño INTA, Plata INTA y Unión y las variedades francesas derivados de Blanc de la Drome (Messidrome y Thermidrome); los seleccionados de Blanc de Beaumont o de Lomagne (Jolimont y Corail) y los derivadas de Blanco de Ronda (Novatop) o Aguilar de la Frontera. Entre los “blancos americanos” son típicas las cultivares California Early y Vigor Supreme (EsVol. 3 - No.1 - 2009 41 42 Burba tados Unidos), Perla INTA (Argentina), y entre los “violetas” Germidour (Francia) y Lican INTA (Argentina). Los del subtipo silverskin presentan bulbos medianos, globosos piriformes, de color blanco níveo (o “plateados”), con muchas hojas fértiles (5 a 8), con numerosos “dientes” (18 a 24), de color blanco con tintes rosados, de cosecha muy tardía y largo periodo de conservación. Los típicos representantes de este subtipo serían California Late de Estados Unidos y las francesas Rosé du Var (de la que se seleccionaron Jardinor, Cristo, Moulinen y Moulinor); Rose d’Auvergne (de la que se seleccionó Perle de Auvergne); Ail du Nord, d’Arleux, d’Arrase (de las que se seleccionaron Gayant, Artop y Atenor); Rose d’Italie (de la que seleccionó Rovigo 24); Rose de Brignoles (de la que seleccionó Brignoles 14); Rose de Corse (de la que se seleccionaron Fructidor y Printanor). También pertenecerían a este grupo Bianco Piacentino de Italia y Blanco de Chinchon de España. Ajos colorados y castaños Son ajos medianos a chicos, globosos a globosos achatados, de color externo blanco o blanco amarillento, eventualmente con tintes violáceos, con pocos “dientes” (entre 5 y 14), de colores rosados, violáceos o parduscos, distribuidos sólo en dos hojas fértiles, de cosecha semitardía o tardía y en general de muy buena capacidad de conservación. En Estados Unidos, la variedad ophioscorodon es denominada popularmente ophios, topsetting (con bulbillos aéreos) y serpent garlic (por la forma retorcida del tallo floral). Serían materiales del tipo hardneck, debido a que interpretan que siempre tiene tallo floral emergente y por tanto en el momento de la cosecha mantienen el “cuello duro”. El subtipo rocambole se corresponde con los conocidos mundialmente como red garlic o purple garlic en Estados Unidos, ajos rojos o morados en REV. COLOMB. CIENC. HORTIC. España, rosados en Chile o colorados en Argentina. Pertenecen al Grupo fisiológico I de Francia, Grupo IV de Argentina, Grupo 1 de Japón o Grupo 2B de China, con bulbos con envolturas blancas y “dientes” pigmentados entre rosa claro y morado oscuro. La variedad denominada popularmente ophios, del subtipo rocambole, se trataría en realidad de ajos de “cuello duro”, pero de la var. sativum y no de la variedad ophioscorodon, que, como se manifestó, se trataría de los tradicionales ajos rojos, y que Argentina posee inscritas varias cultivares como Colorado Mendoza, Fuego INTA, Gostoso INTA, Rubí INTA y Sureño INTA. Pertenecerían también a este subtipo Spanish Roja de Estados Unidos, algunas poblaciones francesas como Rose de Lautrec (de la que se seleccionaron Iberose y Goulurose; Violet de Kabyle (de la que se seleccionó Morasur); Rose d’Espagne (de la que se seleccionaron Morasol, Moraluz y Moratop); Cazador y Chonan (Brasil); Yamagata (Japón), Rosso di Sulmona (Italia), de la que seleccionó en Francia Sultop y Rosado Platina (Chile). Estudios realizados sobre el comportamiento del Grupo fisiológico IV de Argentina, respecto a sus hábitos de floración, indican la existencia de subgrupos como “rusos”, “mediterráneos” (“criollos” y “españoles”) y “asiáticos”, que en principio corresponderían respectivamente a los denominados Continental, Rocambole y asiáticos de la clasificación de Engeland. Existen también diferencias de apreciación, ya que, para este autor, estos grupos tienen escasa conservación, mientras que para las condiciones locales los ajos “colorados” se consideran como ajos “nobles” por su presentación y “de guarda” por su capacidad de almacenamiento en el tiempo. Por otra parte, el término Rocambole (debido probablemente a la forma curvada del tallo floral cuando crece) se aplicó muchas veces a otras especies del género Allium. Rocambole o de España (ajo murciano o español), a veces denominada “cebolla de Egipto”, es Allium scordoprasum L. M ejoramiento genético y producci ó n de “ semilla” de ajo Los ajos “colorados criollos” se caracterizan por poseer mayores requerimientos de frío y fotoperiodo largo en primavera; son de bulbos medianos a grandes, globosos achatados, con disco plano o levemente hundido y emisión relativamente temprana de vara floral. Mientras, por su parte, los “colorados españoles” poseen menores requerimientos de frío y fotoperiodo largo en primavera; son de bulbos medianos a chicos, globosos piriformes, con disco cóncavo y emisión tardía de vara floral. El subtipo continental está considerado como el más próximo a los ajos silvestres nativos del antiguo Turkestán (Turkmenistán, Kirguizistán Kazajstán, Tajikistán y Uzbekistán). En Argentina son denominados popularmente como “ajos rusos”, introducidos y cultivados en regiones muy frías y de los que se conoce sólo una cultivar monoclonal inscrita en los registros oficiales, denominada Castaño INTA. Los “ajos rusos” tienen aroma y sabor muy similar a los ajos silvestres, y quienes son entendidos en apreciar flavors manifiestan que nadie conoce el verdadero sabor del ajo hasta no probar o degustar los ophios. Para los gourmets, el hecho que posean pocos “dientes” grandes y pesados, de sabor semisalvaje (intermedio entre crudo y cocido, pungente pero de aroma muy suave), y de fácil remoción del bulbo lo convierten en un ajo de gran perfomance culinaria. Estas variedades se caracterizan por presentar plantas de hojas verde claro, de largos entre- nudos en el falso tallo y posición opuesta muy marcada, asemejando una hoja de palmera. El tallo floral es emergente, grueso y fuertemente lignificado, rematando en una umbela con gran espata, llena de flores estériles y bulbillos aéreos. Para algunos autores, la vara floral de este sub tipo aparece más tardíamente que en los Rocambole y poseen muchos bulbillos aéreos, pero de tamaño pequeño, sin embargo se citan datos que se contradicen con aquellos. Los bulbos agrupan pocos y grandes “dientes” asentados sobre dos hojas fértiles. Existirían entre los “ajos rusos” variantes denominadas “porcelana” (de 4 a 6 “dientes” y “púrpuras” (de 8 a 12 “dientes”), refiriéndose al color de envoltura del bulbo. El color de los “dientes” es de base marrón o castaña, aunque pueden mostrar tintes rojizos. Si bien se les atribuye escasa conservación, en principio ésta se debe a una extrema sensibilidad al ataque de eriófidos (Aceria tulipae), principal responsable del manchado y deshidratación de la hoja reservante. Cuando este problema sanitario es controlado su conservación es excelente. En síntesis, para la clasificación argentina de ajos de “cuello duro”, los cultivares Fuego, Gostoso y Sureño pertenecerían a la var. sativum del tipo comercial “colorado”, mientras que Morado sería del tipo “asiático”. Por su parte, Castaño pertenecería a la var. ophioscorodon y al tipo comercial “ruso”. La tabla 4 resume esta situación. Vol. 3 - No.1 - 2009 43 44 Burba Tabla 4.Interpretación sintética del agrupamiento de cultivares de ajos argentinos de la especie Allium sativum L. Variedad Tipos comerciales Grupo ecofisiológico Cultivar típico En argentina (*) softneck sativum “blancos” (artichoke) III Nieve INTA “violetas” III Lican INTA “plateados” (silverskin) hardneck “colorados” (rocambole) pekinense (asiáticos) ophioscorodon IV a Fuego INTA “morados” II Morado INTA “rosados” II Alpa Suquía IV b Castaño INTA “castaños” (continental) (*) No se descartan otros Fuente: Burba, 2008. REFERENCIAS BIBLIOGRÁFICAS Burba, J.L. 1990. Producción de “semillas” de sanidad controlada en hortalizas de propagación agámica. pp. 245-250. En: Actas Curso-Taller en Tecnología de Producción de Semillas Hortícolas para Pequeños Agricultores. FAO–INTA, Mendoza, Argentina. Burba, J.L. 1993. Producción de semilla de ajo. En: Crnko, J. (ed.). Manual de Producción de Semillas. INTA. Centro Regional Cuyo, Mendoza, Argentina. 1993. 136 p. Burba, J.L. 1997a. Panorama mundial y nacional de poblaciones y cultivares de ajo. Posibilidades de adaptación. Cultivares y Producción de Semilla (INTA, Argentina) 2, 11-31. Burba, J.L. 1997b. Obtención de nuevas cultivares de ajo. Cultivares y Producción de Semilla (INTA, Argentina) 2, 49-53. Burba, J.L. 2008. Los grupos varietales del ajo (Allium sativum L.). Contribución para su entendimiento. Horticultura Argentina 27(62), 20-27. REV. COLOMB. CIENC. HORTIC. Burba, J.L. 2009. Manejo de semilla de ajo frigoinducida. Documentos Proyecto Ajo/INTA 088. Estación Experimental Agropecuaria INTA La Consulta, Mendoza, Argentina. Burba, J.L.; R. Ocañas; G.E. Lanzavecha y M. Paganini. 2009. Manejo de semilla básica de ajo en condiciones controladas. Documentos Proyecto Ajo/INTA 087. Estación Experimental Agropecuaria INTA La Consulta, Mendoza, Argentina. López Frasca, A.; V. Silvestri y C. Rigoni. 1997. Métodos convencionales del mejoramiento genético del ajo. Cultivares y Producción de Semilla (INTA, Argentina) 2, 32-48. Moriconi, D.N.; J.L: Burba y J. Izquierdo. 1991. Manual de intercambio y propagación de germoplasma de ajo a través de microbulbillos. FAO/RLAC-UN Córdoba, Santiago. Los cultivos de cebolla y ajo en Colombia: estado del arte y perspectivas Onion and garlic crops in Colombia: current state and prospects Hernán Pinzón R.1 Cultivo de cebolla de rama en Aquitania, Boyacá. Foto: H. Pinzón R. RESUMEN Los alliums comestibles que se cultivan en Colombia comprenden varias especies, pero sin duda las de mayor importancia económica son las cebollas de bulbo (incluyendo la ocañera), de rama y el ajo. Sus cultivos tradicionalmente se han localizado en valles de climas medios y fríos moderados de las regiones andinas y cercanos a los centros de consumo. El consumo de estas hortalizas se ha incrementado por sus cualidades condimentarias, asociada a sus beneficios de carácter medicinal. Su importancia socioeconómica es grande por la cantidad de mano de obra, directa e indirecta, asociada a los cultivos y por el capital invertido. Importantes problemas, especialmente de índole fitopatológica (hongos, bacterias y nemátodos en todas estas especies) y de semillas (en ajo, cebolla de rama y ocañera) han causado incremento de los costos de producción, baja en los rendimientos y por tanto disminución del área sembrada y desplazamiento de cultivos a otras zonas buscando suelos no infectados. En el ajo, lo anterior ha causado prácticamente su desaparición como cultivo y en la actualidad se importa casi el 90% de lo que se consume. El ICA y luego otras entidades de investigación y docencia, públicas y privadas, se han vinculado en la búsqueda de la solución a la problemática enunciada mediante proyectos de investigación. Los resultados hasta el momento son de carácter preliminar y desconocidos por la gran mayoría de productores. Falta una coordinación para integrar esfuerzos, los cuales han nacido y se han desarrollado aisladamente. Palabras clave adicionales: fitopatología, semillas, investigación, hongos, nemátodos. 1 Asesor externo hortícola, Bogotá (Colombia). [email protected] REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 3 - No.1 - pp. 45-55, 2009 46 Pinzón R. ABSTRACT The edible Allium plants grown in Colombia include several species, but without doubt the highest economic importance have bulb onions (including ocañera) and bunching onions and garlic. Their crops have traditionally been located in moderate and cold climate valleys of the Andean regions and close to centers of consumption. The consumption of these vegetables has increased by their condimentary qualities associated with the benefits of medicinal characteristics. Its economic importance is high due to the large amount of labor directly associated with cultivation and the capital invested. Major problems, especially phytopathological (fungi, bacteria and nematodes in all these species) and that of seeds (garlic, green onions and ocañera) have caused increased costs of production, low yields and, therefore, decrease the planted area of crops and cause movement to other areas seeking no infection lands. In garlic, these have caused virtually disappearance of it as a crop and now it is imported at almost 90% of what is consumed. ICA and then other research and teaching institutions, public and private, have been linked with the search to solve the problems listed by research projects. The results so far are preliminary and unknown by the vast majority of producers. There is a lack of coordination efforts to integrate these that have been born and developed in isolation. Additional key words: plant pathology, seeds, research, fungi, nematodes. Fecha de recepción: 18-03-2009 Aprobado para publicación: 01-06-2009 IntroduCCIÓN El botánico Hanelt (1990) dividió el género Allium en cinco subgéneros, dos de los cuales contienen las plantas comercialmente importantes (Brewster, 1990). La tabla 1 resume este esquema taxonómico y nos sirve para señalar las especies cultivadas en Colombia. En Colombia, las más cultivadas y consumidas por su aroma, sabor y propiedades medicinales, a las cuales nos vamos a referir, son, en orden de importancia, la cebolla de bulbo, la cebolla de rama y el ajo. Tabla 1.Grupos del género Allium en los que se incluyen los cultivos más importantes. Subgénero Rhizirideum Sección Rhizirideum Schoenoprasum Cepa Allium Fuente: Hanelt, 1990. REV. COLOMB. CIENC. HORTIC. Allium Especies A. tuberosum A. schoenoprasum A. cepa A. fistulosum A. chinense A. sativum A. ampeloprasum Nombres vulgares Cebollino chino Cebollín Cebolla de bulbo Cebolla de rama Rakkyo Ajo Puerro Los cultivos de cebolla y ajo en C olombia : E stado del arte y perspectivas Los tipos de Allium cepa cultivados en Colombia se distribuyen en dos grupos hortícolas: el grupo de la cebolla común y el grupo de los agregados (Hanelt, 1990; Jones y Mann, 1963; Brewster, 1991). Al primer grupo corresponden las cebollas que más se consumen y son aquellas producto de semillas sexuales, importadas en su totalidad, y en el segundo grupo están las cebollas llamadas “ocañeras”, producto de asexuales (bulbos). La denominada cebolla de rama (A. fistulosum), conocida mundialmente como cebolla japonesa, se cultiva para obtener pseudotallos largos y blancos. El ajo tradicional de nuestro país corresponde a los llamados “ajos rosados” (Pinzón, 1999). DESCRIPCIóN La cebolla de bulbo común tradicionalmente ha correspondido al tipo Yellow Granex, con bulbos grandes, aplanados y con piel de un color amarillo muy agradable; presenta problemas en almacenamiento debido a que su piel es muy fina (Acosta et al., 1993; Galmarini, 1990). En los últimos años, las evaluaciones de materiales (semillas) importados apuntan a seleccionar uno o varios híbridos tipo Sweet (menos picantes) y con ventajas para almacenar por un periodo mayor a dos meses (Pinzón, 1996). El sistema de producción de estas cebollas se realiza a partir de un semillero y su cultivo se realiza en muchos climas, especialmente en el frío, 2.600 msnm (Instituto Colombiano Agropecuario, 1983; Cásseres, 1980). Para 2007, el Ministerio de Agricultura y Desarrollo Rural reporta 11.500 ha cosechadas, una producción de 233.286 t y un rendimiento promedio de 19,7 t ha-1. Los departamentos más productores son Boyacá (6.149 ha) y Cundinamarca (3.100 ha) (Agronet, 2009). El otro tipo de cebolla de bulbo que se produce en Colombia corresponde a la llamada “ocañera”, que se caracteriza porque su sistema de propagación se hace vegetativamente a partir de bulbos, luego la planta genera de 1 a 10 bulbos, pero prevalecen plantas de tres a siete bulbos de color rojo pálido a rosado claro y con fuerte sabor picante, apetecido especialmente en la Costa Atlántica (Vergel et al., 2003; Vergel et al., 2004). Su cultivo, localizado en zonas entre 1.200 y 1.900 msnm, prevalece en la Provincia de Ocaña (Norte de Santander) y para 2007 se cosecharon 4.300 ha, con una producción de 79.387 t. La cebolla de rama es apetecida por su pseudotallo largo, jugoso y picante. Su sistema de producción se caracteriza por el abundante aporque (colocación de tierra a la base de la planta), lo cual favorece un abundante macollamiento (número de pseudotallos). Normalmente, a este cultivo se le pueden hacer varios cortes o cosechas (a partir de una sola siembra), el primer corte se hace entre los cinco y los seis meses y los posteriores cada dos a tres meses. Los rendimientos mínimos por corte están cerca de las 30 t ha-1, con un máximo de 75 t y un promedio de 40 t (Pinzón, 2004). Según el Ministerio de Agricultura y Desarrollo Rural, durante 2007 se cosecharon 6.900 ha y una producción de 184.000 t, la región con mayor área sembrada es Aquitania (Boyacá) con 2.425 ha. Este cultivo se adapta a casi todos los climas: la región de Aquitania es considerada zona de páramo (entre 3.000 y 3.400 msnm) y las demás regiones productoras están situadas en altitudes más bajas, con rangos entre 2.700 y 1.500 msnm (Raymond, 1990). En Colombia, todos los materiales cultivados de ajo pertenecen al grupo de los rosados, y su cultivo se ha desarrollado en las subregiones del Altiplano Cundiboyacense, Altiplano de Nariño y Montaña Santandereana. En 2007, según el Ministerio de Agricultura y Desarrollo Rural, se sembraron 375 ha con una producción de 4.300 t y un rendimiento de 11,5 t ha-1, las zonas más productoras son Santander (234 ha), Cundinamarca (70 ha) y Boyacá (35 ha). A pesar de la disminución del área sembrada, su consumo en el país ha tenido un notable incremento: de 0,29 kg/persona–año en 1992 a 0,50 kg/persona–año en 2001, frente a un consumo mundial de 0,80 kg/persona–año. Vol. 3 - No.1 - 2009 47 48 Pinzón R. Su participación en el gasto mensual de los colombianos es alto y por encima de muchas hortalizas consideradas de importancia. La baja en el área cultivada a nivel nacional ha causado un estímulo a las importaciones y más del 90% del ajo que se consume proviene de China (Pinzón, 2008; Calderón, 1999; Departamento Administrativo Nacional de Estadística, 2001). PROBLEMÁTICA Los factores tecnológicos de la producción de estas especies en Colombia tienen características muy similares en las diferentes zonas de producción: baja sostenibilidad y competitividad, relacionadas principalmente con una compleja problemática sanitaria y de deterioro de los recursos ambientales, que resultan en bajos rendimientos y obligan al uso intensivo de agroquímicos (Pinzón, 1996). Esta situación ha desplazado el cultivo a muchas zonas (caso de Boyacá), elevando exageradamente los costos de producción, agudizando los procesos de deterioro ambiental y acentuando los riesgos para la salud humana. En ajo, esta situación produjo una reducción dramática del área sembrada (hasta 400 hectáreas en la actualidad), y como consecuencia las importaciones (principalmente de China) del ajo que se consume aumentaron hasta un 90%. Una descripción puntual de estos problemas es la siguiente: Materiales de propagación: con excepción de la cebolla de bulbo común, los Alliums nombrados anteriormente son propagados vegetativamente. En Colombia no existen empresas o entidades creadas para la producción de esta clase de semillas y no existe una normatización por parte del Estado, por ello su mercado es caótico, teniendo como consecuencia un insumo de muy baja calidad sanitaria y agronómica (Pinzón, 2002, 2003). Los bulbos, pseudotallos y dientes, al no ser producidos adecuada y técnicamente, pueden ser portadores sistemáticos de nemáto- REV. COLOMB. CIENC. HORTIC. dos y algunas estructuras de reproducción de hongos, por tanto, al utilizarlos como semillas causan un efecto multiplicador de infección e infestación en los terrenos (López y Ávila, 1996; Rico y Ávila, 1989; Gaviola y Oliva, 1985; López y Heredia, 1997) Enfermedades: son varios los patógenos que están afectando negativamente a los cultivos de cebollas y ajo, pero sin duda los que están causando mayor impacto socio–económico son los que afectan al sistema radicular y bulbos. El nemátodo Ditylenchus dipsaci ha tomado mucha importancia en nuestro medio debido a su gran poder de diseminación a través de los dientes, gajos o bulbos utilizados como semillas y a su poder de infectar la planta a partir del suelo contaminado. Los adelantos en investigación demuestran la asociación de nemátodos con bacterias y nemátodos con hongos (Hio, 2009; Tovar et al., 1997). El hongo Sclerotium cepivorum, que causa la enfermedad llamada “pudrición blanca”, es otro de los patógenos que está causando grandes pérdidas económicas, especialmente a los cultivos de cebolla común y ajo. En nuestro medio se constató una disminución en los rendimientos de los cultivos de ajo entre un 45% y 61%, por ataque de virus igualmente se determinó una reducción del 17% en altura de las plantas y un 39% en el peso de los dientes (Pinzón, 2008; Angarita, 1998; Ávila, 1996). Estos dos problemas: semillas de mala calidad sanitaria y enfermedades han conformado un círculo vicioso e infeccioso que ha provocado desplazamientos de cultivos buscando zonas “nuevas” y, por tanto, han ampliado el radio de infección. En la mayor zona de producción de cebolla de rama, causó un descenso del número de “cortes” o cosechas de 18 a 3, a partir de una sola siembra (Herrera et al., 2006a) y, como se comentó antes, provocó un descenso vertiginoso del área nacional sembrada de ajo, trayendo como consecuencia el incremento en las importaciones. Los cultivos de cebolla y ajo en C olombia : E stado del arte y perspectivas Poscosecha: la comercialización de nuestras cebollas y ajos en su gran mayoría se realiza sin o con un mínimo valor agregado. En el caso de las cebollas comunes, al momento de la cosecha sólo se eliminan pseudotallos y raíces y los bulbos no se curan, ni se someten a procesos de selección y clasificación. Es de anotar que la producción de esta cebolla es típicamente estacional, lo cual genera variaciones en los precios, restándole competitividad al cultivo, por lo cual es importante generar tecnología para su almacenamiento (Báez y Díaz, 2005; Herrera et al., 2006a). La cebolla de rama cosechada sólo tiene un proceso de presentación y transporte en las llamadas “ruedas”. Por su parte, el ajo cosechado todavía conserva la tradicional y antitécnica presentación en “manojos” y tampoco se le hace selección ni clasificación. Otros problemas: aparte de los problemas de tipo tecnológico que se presentan a nivel de cultivo, se encuentran otros de igual o mayor importancia, entre los cuales se destaca la deficiente comercialización. Colombia cuenta con centrales mayoristas de mercadeo creadas para, supuestamente, servir de apoyo al agricultor en la comercialización de sus productos, sin embargo, estos centros han caído en manos de unos pocos comercializadores que pagan valores arbitrarios por las cosechas, disminuyendo sensiblemente los ingresos de los productores (Alonso y Pinzón, 2002). RESPUESTAS DESDE LA INVESTIGACIÓN La investigación en los Alliums ha sido paralela al desarrollo de la investigación hortícola en Colombia: ésta se inició en 1959 en la División de Investigación Agropecuaria (DIA) y continuó con la creación del Instituto Colombiano Agropecuario (ICA) en 1963. En ese Instituto se creó el Programa de Investigación en Hortalizas y Frutales y en 1977 se estructuraron como programas independientes. En 1979, el ICA elaboró un diagnóstico del sector agropecuario, en el cual presentó la problemática tecnológica, basada en el limitante y en la oferta tecnológica para solucionarla. Elaborado el diagnóstico, estructuró un Plan Nacional de Investigación (Plania) en el cual indicó y jerarquizó las áreas de investigación, con base en la importancia de los problemas a resolver y en la oferta tecnológica disponible. La investigación propuesta en el Plania respecto a hortalizas comprendió 18 especies y propuso 38 proyectos en las disciplinas de mejoramiento, fisiología, entomología, fitopatología, procesos y suelos. En 1989, el Programa de Hortalizas pasó a ser una Sección Nacional constituida por un Grupo Multidisciplinario. A partir de 1993, las actividades de investigación agropecuaria pasaron a la Corporación Colombiana de Investigación Agropecuaria (Corpoica), que en sus primeros años realizó un esfuerzo tendiente a priorizar las especies, sus problemas y las posibles soluciones tecnológicas (Pinzón, 1997; Rondón y Becerra, 1996). Teniendo en cuenta que el país adolece de una gran “central de datos” que recopile el historial investigativo agropecuario nacional, la siguiente constituye una aproximación a los principales logros alcanzados en la investigación en cebollas y ajo durante los últimos años: Cebolla de bulbo Las cebollas en general aparecieron como especies prioritarias en la mayoría de estas entidades y programas de investigación. En sus primeros años, el ICA privilegió los estudios de fertilización e incorporó los de fitopatología y entomología, luego acogió la política de evaluación de materiales (semillas) importados en especies hortícolas de propagación sexual, como los de cebolla de bubo. A Colombia llegan semillas provenientes principalmente de Estados Unidos (las de mayor demanda), Europa, Israel y Japón, y últimamente las evaluaciones han estado dirigidas a seleccionar híbridos de tipo Sweet (cebollas menos picantes) y con un tiempo razonable de almacenamiento (López y Ávila, 1996). Vol. 3 - No.1 - 2009 49 50 Pinzón R. El manejo de enfermedades se ha abordado teniendo en cuenta los factores principales que inciden en ellas, como el manejo del agua para riego y la fertilización adecuada. En un proyecto financiado por el Ministerio de Agricultura y Desarrollo Rural y Asohofrucol, actualmente se trabaja en los estudios preliminares para implementar sistemas de fertirriego en la zona de producción de Sogamoso (Boyacá), con miras a fomentar el ahorro en el consumo de agua y a fertilizar de acuerdo con los requerimientos de la planta y el suelo. En el transcurso de los años, se han efectuado diagnósticos y trabajos tendientes a la identificación y manejo de los patógenos que afectan al follaje y especialmente al bulbo; sin embargo, al tema no se le ha otorgado la importancia que debería tener y los problemas prevalecen. Nuestras condiciones ambientales son variables y por esa razón hay dificultad para curar los bulbos en campo después de su cosecha, por tanto, a los diversos ensayos de secado natural que utilizan cubiertas de protección se agregó un estudio incluido en un proyecto sobre “Producción y tecnologías de curado y almacenamiento de cebolla de bulbo”, financiado por Sena y Asohofrucol en 2005, el cual utiliza prototipos de secamiento con aire forzado, en los que el aire es calentado mediante quemadores de combustible (coque, ACPM) y se hace pasar un flujo de aire a través de las cebollas, retirando de esta manera la humedad de los dos o tres catáfilos exteriores de los bulbos; la temperatura del aire para el secado puede variar entre 35°C y 45°C, de esta condición depende el tiempo de secado, que es aproximadamente 24 h (Pinzón et al., 2003; Suárez y Cadena, 2005). Cebolla de rama Con la finalidad de preservar los materiales existentes y conocer su variabilidad, el ICA, en 1980, inició las colecciones de cebolla de rama, en 1984 ya se tenía un Banco de 34 clones y en la actualidad se cuenta con 60. El análisis de esa colección arrojó diferencias significativas para las variables REV. COLOMB. CIENC. HORTIC. cuantitativas y cualitativas, especialmente para rendimiento, número y peso de tallos; el análisis multivariado definió seis grupos para doce variables analizadas, las cuales explican el 64% de la variabilidad total encontrada y se halló que la interacción genotipo–ambiente fue significativa para caracteres asociados con rendimiento (Jaramillo et al., 1986; Rozo y Tinjacá, 1995). A partir de 1985, el ICA llevó los mejores materiales de la Colección para Aquitania, con el propósito de evaluarlos y seleccionar los de mejor adaptación, comportamiento agronómico y sanitario y mejor calidad (pungencia, color, grosor). En 1995, se presentó una carencia de recursos económicos en Corpoica y hubo que suspender esas evaluaciones, por esta razón un agricultor multiplicó por su cuenta el clon que presentaba las mejores perspectivas, le puso nombre y lo comercializó con buena aceptación en su comienzo, pero, desafortunadamente, no pudo sostener la pureza del material y éste se perdió. Posteriormente y con intermitencias se prosiguió la tarea de las evaluaciones en Aquitania y en la actualidad hay en marcha un proyecto financiado por el Ministerio de Agricultura y Desarrollo Rural, destinado a obtener variedades para esa región (Herrera et al., 2006a). En 2005, Corpoica comenzó a trabajar en un proyecto, financiado en parte por Corpoboyacá, destinado a desarrollar procesos de innovación y tecnologías de producción más limpias, a través de un programa integral para prevenir y mitigar impactos ambientales negativos generados por el cultivo de cebolla en Aquitania. Los aspectos más relevantes de este programa fueron (Herrera et al., 2006): •La obtención del protocolo de producción de plantas in vitro de cebolla de rama. Se cumplieron las etapas de selección de plantas donantes, desinfección de esas plantas, el proceso de introducción in vitro de ápices y el proceso de multiplicación in vitro de plántulas El proyecto se encuentra suspendido. Los cultivos de cebolla y ajo en C olombia : E stado del arte y perspectivas •Investigar y producir biabono a través de un proceso de compostaje, utilizando inóculos micro­bianos y a partir de residuos de elodea, cebolla y gallinaza; este proceso se ha continuado en manos de una cooperativa privada, Asoparcela. • Con el concurso de la Corporación PBA, el fortalecimiento de la capacidad asociativa de los productores con la creación y consolidación de Asoparcela, entidad que agrupa en la actualidad cerca de 50 socios productores de cebolla. •Mediante reconocimientos en campo e identificación en laboratorio de las enfermedades más limitantes, se obtuvo información valiosa sobre la presencia de Ditylenchus dipsaci, Pseudomonas sp., Peronospora destructor, Stemphylium allí y Cladosporium allí. Corpoica en 2008, con financiación del Ministerio de Agricultura y Asohofrucol, desarrolló un proyecto de carácter sanitario para el estudio sobre la interacción del nemátodo Ditylenchus dipsaci (Kuhn) y la bacteria Burkholderia cepacia (Hio, 2009) y su relación con la pudrición radical en cebolla de rama. Entre sus resultados más importantes se destacan los siguientes: descripción de los cambios morfológicos de plantas de cebolla, ocasionados por los microorganismos patógenos involucrados en el proceso de la enfermedad; descripción de síntomas de pudrición radical de plantas de cebolla en campo; aislamiento de bacterias a partir de material vegetal y suelo; extracción de nemátodos; sinergismo entre los microorganismos relacionados con la pudrición radical e incidencia de la enfermedad (pudrición radical) en condiciones de campo (Hio, 2009; Corporación Colombiana de Investigación Agropecuaria, 1997). Ajo En 1982, el ICA inició y completó un banco de germoplasma de ajo consistente en 20 clones provenientes de diferentes regiones del país. Entre 1990 y 1991, a esta colección se le hicieron estudios de caracterización para conocer su variabilidad genética disponible; el análisis de las variables canónicas y del dendograma indicó una diversidad genética que permitió su clasificación en cuatro grupos genéticamente uniformes y se definieron algunos genotipos aptos para incluirlos en un programa de selección de clones por su precocidad, uniformidad del bulbo y porte de la planta (Ligarreto, 1991). En 1994, Corpoica estructuró un proyecto destinado a obtener la primera variedad nacional mejorada de ajo. Ésta fue obtenida en un proceso investigativo de cinco años a través de la caracterización, evaluación y selección de diez clones promisorios, cinco de ellos de origen argentino, los cuales fueron enviados al ICA en 1992 por la Red de Cooperación Técnica en producción de cultivos que auspicia la FAO; los otros cinco eran los mejores materiales pertenecientes al Banco de Germoplasma del ICA. Como producto de este trabajo, surgió la variedad denominada Rubí-1 y en 2004 a Corpoica le fue concedido el Certificado de Obtentor (Pinzón, 1999; Ballesteros y Gómez, 2004). Después de estar un tiempo en el mercado, esta variedad fue retirada provisionalmente mientras se accedía a un protocolo de multiplicación in vitro con el propósito de obtener semillas limpias. Colciencias, en 2004, financió a Corpoica un proyecto para desarrollar procesos de innovación tecnológica para la producción de ajo Rubí-1 basado en el preescalamiento de semillas limpias de alta calidad, el secamiento y almacenamiento y mediante la técnica de cultivo in vitro de meristemos (Parra, 2006). Este proyecto actualmente se encuentra en ejecución. En la actualidad, el Centro de Investigaciones y Asesorías Agroindustriales (CIAA) de la Universidad Jorge Tadeo Lozano está desarrollando dos proyectos financiados por el Ministerio de Agricultura y Asohofrucol destinados a la evaluación participativa de tecnologías de manejo integrado de plagas para los principales limitantes fitopa- Vol. 3 - No.1 - 2009 51 52 Pinzón R. togénicos Sclerotium cepivorum y nemátodos y el mejoramiento de la competitividad del ajo en Colombia mediante el ajuste participativo de tecnologías disponibles para la obtención y comercialización de semillas garantizadas. PERSPECTIVAS Las perspectivas de este grupo de especies hortícolas nacen de su importancia: de acuerdo con el Plan Hortícola Nacional (PHN) la tendencia histórica muestra que la cebolla de bulbo domina la producción hortícola, seguida de la cebolla de rama, y estas mismas especies dominan el área de producción, después de la arveja. De hecho, entre los 15 productos de mayor preferencia para los colombianos en su canasta alimenticia diaria se encuentran el tomate, la cebolla cabezona y la zanahoria. El tomate es la hortaliza más consumida con 38% de preferencia y una cantidad diaria de aproximadamente 45 g y la cebolla cabezona es la siguiente con 30% de preferencia y una cantidad diaria de aproximadamente 31,8 g. Por otro lado, las importaciones colombianas de cebollas y ajo, entre frescas y secas, alcanzaron en 2006 un valor de 18 millones de dólares, cifra que constituye un 52% del valor total de las importaciones nacionales de hortalizas (Corporación Colombiana Internacional, 2007; Jaramillo, 2006). Como se expuso anteriormente, es de anotar que ha habido respuesta de algunas entidades de investigación a la problemática puntual del sector de los Alliums comestibles, si bien no en la proporción que se requiere. Sin entrar en zonas críticas de manejo político, como las de comercialización y precios, no es claro el panorama que se visualiza: por un lado, las áreas de siembra disminuyen y las zonas infectadas por enfermedades crecen amenazando zonas nuevas y, por otro lado, los recursos de financiación para investigación compiten con otros sectores agrícolas. REV. COLOMB. CIENC. HORTIC. Recordemos, además, que los Alliums comestibles en Colombia constituyen un sector de alto impacto socio–económico que demanda de casi tres millones de jornales anuales, aporta más de 500.000 t de productos frescos para el consumo y favorece otros sectores sensibles de la economía como el transporte y la producción de insumos para el cultivo (Ministerio de Agricultura y Desarrollo Rural, 2005). Es claro que dos grandes problemas se destacan actualmente y merecen la máxima atención, desde el punto de vista de la investigación, ellos son las enfermedades y la calidad sanitaria de las semillas de propagación asexual. Como se aprecia en la breve reseña de proyectos desarrollados y en curso, el aspecto fitopatológico no ha sido el más importante y requiere la máxima atención y, por otro lado, tanto a los proyectos sobre evaluación y selección de materiales destinados a la obtención de nuevas variedades como a los estudios para obtener “semillas limpias”, acudiendo a la herramienta de la biotecnología, se les debe garantizar su continuidad. Sin embargo, es necesario resaltar algunos aspectos que pueden ayudar en la búsqueda de soluciones a la problemática global de estas especies: •La falta de capacidad asociativa de los productores dificulta tener interlocutores para la elaboración y presentación de proyectos que los beneficien y apoyen cuando se están desarrollando. •El sector privado difícilmente aporta recursos para, por lo menos, apoyar económicamente en un porcentaje a los proyectos que demanda la comunidad de productores. •La investigación, en nuestro medio, necesita priorizar regularmente sus objetivos y alcances y necesita planes sostenidos a mediano y largo plazo. Los cultivos de cebolla y ajo en C olombia : E stado del arte y perspectivas •Si bien las individualidades están presentes en todas las actividades, es necesario privilegiar y fortalecer el sentido de equipo de trabajo. Los grupos de investigación–extensión de carácter multidisciplinario son importantes en este proceso. La perspectiva en Colombia, desde el punto de vista relacionado con la agroindustria de los Alliums comestibles es todavía muy reducida, debido a que, en general, a pesar de la estacionalidad de la oferta, se encuentran productos frescos todo el año. Sin embargo, los nuevos requerimientos en los mercados, ocasionados por los cambios en la sociedad relacionados con la participación de la mujer en el mercado laboral, menos tiempo en el hogar, personas independientes y la practicidad de los alimentos preparados, están empezando a generar cambios, que seguramente se acelerarán a corto plazo. Las estadísticas comerciales mundiales muestran claramente el gran incremento que se ha experimentado en el comercio mundial de procesados, esta situación es una tendencia mundial y, por tanto, Colombia deberá producir y comercializar de acuerdo con los requerimientos de la demanda. En el mercado local, ya se están ofertando diversas presentaciones de Alliums (vinagreta, deshidratados, cremas, concentrados). Sin embargo, es difícil conseguir información sobre el estado actual de esa agroindustria, porque todavía su producción es muy pequeña (Báez y Díaz, 2005; Corporación Colombiana Internacional, 1994; Pinzón et al., 2006). REFERENCIAS BIBLIOGRÁFICAS Acosta, A.; J. Gaviola y C. Galvamarini. 1993. Cebolla. Manual de producción de semillas hortícolas. EEA La Consulta. INTA, Argentina. Agronet. 2009. Producción nacional por producto. En: Análisis y estadísticas, 1997. http://www.agronet. gov.co; consulta: marzo de 2009. Alonso, A. y M.S. Pinzón. 2002. Situación de la comercialización del ajo en la Central Mayorista de Abastos en Bogotá y su inserción en la cadena hortícola. Trabajo de grado. Universidad de Cundinamarca, Facultad de Ciencias Agropecuarias, Ubaté, Colombia. Ballesteros, C.A. y L.E. Gómez. 2004. Estudio de la adaptabilidad del ajo Rubí-1 en la Región del Sumapaz. Trabajo de grado. Facultad de Ciencias Agropecuarias, Universidad de Cundinamarca, Fusagasugá, Colombia. Brewster, J.L. 1990. Physiology of crop growth and bulbing. pp. 53-88. En: Rabinowicht, H.D. y J.L. Brewster (eds.). Onions and allied crops. Vol.1. CRC Press, Boca Raton, FL. Brewster, J. 2001. Las cebollas y otros alliums. Editorial Acribia, Zaragoza, España. Angarita, A. 1998. Distribución, incidencia y severidad de las principales enfermedades de cebolla de rama en la cuenca del lago de Tota. Trabajo de maestría en Ciencias Agrarias. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. Calderón, P.N. 1999. Sondeo del mercado del ajo en Colombia. Corpoica, Mosquera, Colombia. Ávila de M., C. 1996. Principales enfermedades del ajo. pp. 73-86. En: López, A. (ed). El cultivo del ajo y las cebollas en Colombia. Produmedios, Bogotá. Casseres, E. 1980. Producción de hortalizas. Instituto Interamericano de Ciencias Agrícolas, San José de Costa Rica. Báez, A. y E. Diaz. 2005. Evaluación de dos sistemas de almacenamiento de cebolla de bulbo (Allium cepa) curada utilizando condiciones de refrigeración y aire ligeramente calentado. Trabajo de grado. Facultad de Ingeniería Agrícola, Universidad Nacional de Colombia, Bogotá. 79 p. Corporación Colombiana Internacional. 1994. Análisis Internacional del sector hortifrutícola para Colombia. Serie CCI Informes técnicos, No. 1. Bogotá. Caicedo, L. 1972. Curso de horticultura. Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Palmira. Corporación Colombiana Internacional. 2007. Plán Hortícola Nacional. CCI, Bogotá. Vol. 3 - No.1 - 2009 53 54 Pinzón R. Corporación Colombiana de Investigación Agropecuaria. 1997. Control biológico de fitopatógenos. Seminario internacional. Produmedios, Bogotá. Departamento Administrativo Nacional de Estadística. 2001. Primer censo del cultivo de cebolla larga en la Región de la laguna de Tota. DANE, Bogotá. Galmarini, C. 1990. Caracterización de cultivares argentinos de cebolla (Allium cepa) de acuerdo a sus requerimientos de vernalización. Tesis de maestría. Instituto Agronómico Mediterráneo de Zaragoza, España. Gaviola, J. y N. Oliva. 1985. Efectos de la densidad de plantación de bulbos de cebolla (Allium cepa) sobre el rendimiento y la calidad de la semilla. pp. 1-39. En: Resúmenes XI Seminario Panamericano de Semillas. Cali, Colombia. Hanelt, P. 1990. Taxonomy, evolution and history. pp. 1-26. En: Rabinowicht, H.D. y J.L. Brewster (eds.). Onions and allied crops. Vol.1. CRC Press, Boca Raton, FL. Hio, J.C. 2009. Estudio sobre la interacción del nematodo Ditylenchus dipsaci (Kuhn) y la bacteria Burkholderia cepacia (Palleroni and Holmes 1981) y su relación con la pudrición radical en cebolla de rama Allium fistulosum L., en el municipio de Aquitania (Boyacá). Tesis de Maestría en Biología Aplicada. Facultad de Ciencias, Universidad Militar Nueva Granada, Bogotá. Herrera, C.; G. Sánchez y V. Peña. 2006a. Avances de resultados de investigación en cebolla de rama en Aquitania, Boyacá. Produmedios, Bogotá. Herrera, C.; M. Laitón y G. Sánchez. 2006b. Cebolla de bulbo. Guía para la implementación de buenas prácticas agrícolas. Produmedios, Bogotá. Instituto Colombiano Agropecuario. 1983. Hortalizas. Manual de asistencia técnica. División de Comunicación del ICA, Bogotá. Jaramillo, J.; Y. Palacios y O. Martínez. 1986. Descripción cuantitativa de la colección colombiana de cebolla de rama. ICA, Mosquera, Colombia. Jaramillo, J.E. 2006. Situación de la producción de las hortalizas en Colombia y sus perspectivas. pp. 137149. En: Memorias Primer Congreso Colombiano de Horticultura. Sociedad Colombiana de Ciencias Hortícolas, Bogotá. Jones, H. y Mann, L. 1963. Onions and their allies. Leonard Hill, London. pp. 36-38. Ligarreto, G.A. 1991. Caracterización morfológica y electroforética de cultivares de ajo. Trabajo de grado. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. REV. COLOMB. CIENC. HORTIC. López, A. y C. Ávila. 1996. Manejo fitosanitario del ajo y las cebollas. Produmedios, Bogotá. López, E. y J. Heredia. 1997. Evaluación de diferentes densidades de siembra en la propagación vegetativa de la cebolla de rama (Allium fistulosum) en condiciones de la Sabana de Bogotá. Trabajo de grado. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. Ministerio de Agricultura y Desarrollo Rural, IICA. 2005. La competitividad de las cadenas agroproductivas en Colombia: análisis de su estructura y dinámica. (1991-2004). Bogotá. Ministerio de Agricultura y Desarrollo Rural. 2006. Acuerdo de competitividad de la Cadena de Hortalizas. Bogotá. Parra, M. 2006. Desarrollo de tecnologías para la producción de semilla prebásica indexada de ajo (Allium sativum, L.) var. Rubí-1 y criollo en el Laboratorio de Micropropagación de Plantas de Corpoica C.I. Tibaitatá del Municipio de Mosquera-Cundinamarca. Trabajo de grado. Facultad de Ciencias Agrarias y del Ambiente, Universidad Francisco de Paula Santander, Colombia. Pinzón, H. 1996. Botánica, morfología y fisiología. pp. 9-16. En: López, A. (ed.). El cultivo del ajo y las cebollas en Colombia. Produmedios, Bogotá. Pinzón, H. 1997. Consideraciones para la investigación hortícola en Colombia. Corpoica, Mosquera, Colombia. Pinzón, H. 1999. Rubí-1 primera variedad colombiana de ajo. Produmedios, Bogotá. Pinzón, H. 2002. Producción de Alliaceas. pp 11-13. En: Memorias Taller de Hortalizas, Productividad y Mercadeo. Produmedios, Bogotá. Pinzón, H. 2003. Producción de semillas de algunas alliáceas. pp. 33-36. En: Memorias II Taller de Hortalizas, Productividad y Mercadeo. Produmedios, Bogotá. Pinzón, H. 2004. La cebolla de rama (Allium fistulosum) y su cultivo. Produmedios, Bogotá. Pinzón, H. 2008. El cultivo del ajo. Manejo empresarial del campo. Produmedios, Bogotá. Pinzón, H.; J. Ospina y A. Baez. 2006. Curado y almacenamiento de cebolla de bulbo. Editorial Litolaser, Bogotá. Pinzón, H.; J. Torres; P. Prada y L. Barrera. 2003. Informe final proyecto: Validación participativa y transferencia de tecnología para la producción sostenible de cebolla de bulbo, cebolla de rama y ajo en Cundinamarca y Boyacá. Corpoica, Bogotá. Los cultivos de cebolla y ajo en C olombia : E stado del arte y perspectivas Raymond, P. 1990. El lago de Tota ahogado en cebolla. Serie investigación y desarrollo No. 1. Facultad de Ciencias Económicas y Administrativas, Pontificia Universidad Javeriana. Editorial Presencia, Bogotá. Suarez, L. y D. Cadena. 2005. Estudio del secado en cebolla de bulbo (Allium cepa). Trabajo de grado. Departamento de Ingeniería Agrícola, Universidad Nacional de Colombia, Bogotá. Rico, L. y M. Ávila 1989. Estudios preliminares sobre el efecto de un virus en ajo. p. 18. En: Memorias XXIX Congreso de Ascolfi, Cali. Tovar, D.R.; A.M. Yara y L.P. Zuleta. 1997. Estudio preliminar para control biológico de Sclerotium cepivorum en cebolla de bulbo y Ditylenchus dipsaci en cebolla de rama. Trabajo de grado. Facultad de Ciencias de la Salud, Universidad Colegio Mayor de Cundinamarca, Bogotá. Rondón, S. y N. Becerra. 1996. Efecto de las coberturas plásticas en dos sistemas de siembra sobre el crecimiento y desarrollo de la cebolla de bulbo (Allium cepa) en la Sabana de Bogotá. Trabajo de grado. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. Rozo, I. y G. Tinjacá. 1995. Caracterización morfológica de cultivares de la colección colombiana de cebolla de rama Trabajo de grado. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. Vergel, L.; J. Jaramillo y P. Conde. 2003. Producción de cebolla de bulbo ocañera. Manual técnico. Litografía La Bastilla, Bucaramanga, Colombia. Vergel, L.; C. Terán y C. Gómez. 2004. Manejo de riego en cebolla ocañera. Manual técnico. Litografía La Bastilla, Bucaramanga, Colombia. Vol. 3 - No.1 - 2009 55 Evaluación de fuentes de fertilización edáfica y el efecto sobre la microbiota presente en un cultivo de cebolla cabezona (Allium cepa L.) Evaluation of fertilization sources and the effect on microbiota present in bulb onion (Allium cepa L.) culture Carlos Alberto Luengas–Gómez1,3 Diana Paola Mora–Castro2 Dorisana Guerrero–Flórez1 Cultivo dede Plantulas cebolla tomate. cabezona en Pasca, Cundinamarca. Foto: Álvarez-Herrera. Foto: D. Guerrero–Flórez RESUMEN En Pasca (Cundinamarca, Colombia), municipio ubicado a 2.090 msnm, temperatura y precipitaciones anuales promedio de 14°C y 1.800 mm, la cebolla cabezona está distribuida ampliamente. En un lote de este municipio se estudió el híbrido Yellow Granex PRR, utilizando las siguientes fuentes y épocas de suministro para cada tratamiento; orgánico: compost, roca fosfórica, KCl, MgO y azotobacter, incorporados en trasplante; orgánico–mineral: CO(NH2)2, (NH4)2HPO4, MgSO4, KCl, Ca(NO3)2, Compost, adicionados en trasplante y 30 días después transplante (ddt); convencional se usaron las mismas fuentes, intercambiando el compost por micronutrientes, agregados en trasplante, 30 y 75 ddt. Se tomaron datos durante dos ciclos de cultivo consecutivos (abril 2008 a enero 2009), a cada unidad experimental se le realizaron análisis químicos de suelo y microbiológicos, al iniciar y finalizar. Se evaluaron unidades formadoras de colonia (ufc): bacterias, actinomicetos, hongos, solubilizadoras de fósforo, fijadoras de nitrógeno; se midieron variables: altura, peso fresco y seco de planta y bulbo, registros de cosecha. Los resultados mostraron respuestas favorables en rendimiento para el tratamiento orgánico en dosis media de 26,35 t ha-1, seguido del orgánico–mineral en dosis baja 22,65 t ha-1 y el convencional en dosis media 19,17 t ha-1, se atribuye a las altas reservas minerales que posee este suelo, debido a aportes continuos de materia orgánica y cales en ciclos anteriores, justificado en todos los aná- 1 Universidad de Ciencias Aplicadas y Ambientales (UDCA), Bogotá (Colombia). Universidad de Cundinamarca, Fusagasugá (Colombia). 3 Autor para correspondencia. [email protected] 2 REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 3 - No.1 - pp. 56-109, 2009 E valuaci ó n de fuentes de fertilizaci ó n ed á fica y el efecto sobre la microbiota lisis de suelo realizados. Se observó en el tratamiento convencional en dosis media aumento en las poblaciones evaluadas con excepción de los hongos, en el que el orgánico en dosis media reportó poblaciones superiores. En cambio, el tratamiento orgánico en dosis alta sólo aumentó para actinomicetos y fijadoras de nitrógeno, la abundancia se explica porque hubo condiciones favorables como: pH neutro, suelo cercano a capacidad de campo y alto contenido nutricional. Palabras clave adicionales: nutrición, actividad microbiológica, producción. ABSTRACT In Pasca (Cundinamarca, Colombia), municipality located at 2,090 msnm, with annual temperature14°C and precipitation average 1,800 mm, onion bulb is widely distributed. The experimental trial was done with hybrid Yellow Granex PRR using the following sources and times for each treatment: organic that included compost, phosphoric rock, KCl, MgO and azotobacter, built-in transplant; organo-mineral: CO(NH2)2, (NH4)2HPO4, MgSO4, KCl, Ca(NO3)2, compost added at transplant and 30 days after transplanting (dat); conventional fertilizer that used the same sources, interchanging compost by micronutrients aggregated at transplanting, at 30, and 75 dat. The data were taken during two consecutive culture cycles (April 2008 to January 2009) and soil microbiological and chemical analyses were realized on each experimental unit; when initiating and finishing form units of colony (ufc) were evaluated: bacteria, actinomycota, fungi, phosphorus solubilizers, nitrogen fixers. The variables measured were: height, fresh and dry weight of plant and bulb, registries of harvest. The results showed favorable response in yield for the organic treatment in average dose 26.35 t ha-1, followed by the organic-mineral in low dose 22.65 t ha-1 and conventional one in average dose 19.17 t ha-1, it attributes to the high levels of mineral reserves that this soil has, due to the continuous contributions of organic matter and lime to it in previous cycles, justified in all realized soil analyses. An increase in the populations evaluated except that of fungi was observed in the conventional treatment in average dose, where the organic one in average dose report superior populations. However, the organic treatment in high dose only increased for actinomycota and nitrogen fixers. The abundance is explained because there were favorable conditions, such as neutral pH, soil moisture near field capacity and high nutrient contents. Additional keywords: nutrition, production, microbiological activity. Fecha de recepción: 20-03-2009 Aprobado para publicación: 01-06-2009 IntroduCCIÓN La actividad de la agricultura que provee al suelo y planta los nutrientes faltantes, denominada fertilización, y el material aplicado lleva el nombre de fertilizante. Lo ideal es devolver al suelo todo lo extraído de él; por lo menos, el agricultor debe procurar devolver no sólo la máxima cantidad de nutrientes perdidos, sino dar alimento a los microorganismos del suelo y, al mismo tiempo, mantener el suelo en buenas condiciones fí- sicas. La mayoría de nutrientes del suelo tienen origen en el material parental del cual se ha formado, la otra parte se incorpora por intercambio de gases, flujo de agua freática, fijación biológica de nitrógeno y mediante la fertilización. Los microorganismos son componentes importantes del suelo, constituyen su parte viva y son responsables de la dinámica de transformación Vol. 3 - No.1 - 2009 57 58 Luengas-Gómez/Mora-Castro/Guerrero-Flórez y desarrollo. La diversidad de microorganismos encontrados en una fracción de suelo cumple funciones determinantes en transformación de los componentes orgánicos e inorgánicos que se incorporan. La humificación de la materia orgánica es proceso netamente microbiológico. La microflora del suelo está compuesta por bacterias, actinomicetos, hongos, algas, virus y protozoarios (Abril, 2002). El suelo utilizado en el proyecto está descrito como Humic Dystrudepts, caracterizado por un paisaje montañoso, clima frío húmedo, relieve ligeramente plano a ligeramente ondulado, pendientes 3% a 7%, suelos profundos, moderadamente bien drenados, presentan reacción fuerte a fuertemente ácida, saturación de aluminio alta y fertilidad moderada a baja (Borrero, 2000). Actualmente estas tierras del Municipio de Pasca (Cundinamarca) se encuentran dedicadas a cultivos transitorios y semipermanentes, hortalizas como la cebolla cabezona, pertenece a la familia Liliaceae, posee hojas tubulares y alargadas que dan origen al bulbo, formado por hojas modificadas (catafilas); excelente captadora de radiación solar, a su vez es ineficiente para competir con malezas. Presenta raíces cortas que salen de la parte inferior del tallo, por lo que las prácticas de riego y fertilización son muy importantes (ICA, 1989). El cultivo tiene un periodo vegetativo aproximadamente de 120 días. A nivel nacional, la producción de este cultivo se concentra en Boyacá como el mayor productor y que registra mejores rendimientos por área (23,36 t ha-1), tercero Cundinamarca con 16,88 t ha-1 (Melo et al., 2006). Para las zonas de Boyacá, Cundinamarca y Nariño, el híbrido Yellow Granex tiene amplia aceptación por su tolerancia a raíz rosada y bulbos de tamaño grande forma gruesa y aplanada, sabor suave, atractivas capas exteriores. Debido a los costos elevados de los fertilizantes sintéticos y su impacto sobre los microorganismos benéficos para las plantas, el objetivo de este trabajo fue la evaluación de diferentes fuentes y dosis de fertilización en el crecimiento y rendimiento del cultivo de cebolla para la zona de Pasca. Figura 1.Estadios fenológicos de la cebolla cabezona, según Dogliotti y Galvan (2008). REV. COLOMB. CIENC. HORTIC. E valuaci ó n de fuentes de fertilizaci ó n ed á fica y el efecto sobre la microbiota MATERIALES Y MéTODOS El lote donde se llevó a cabo el ensayo experimental de 2.000 m2 está localizado en el municipio de Pasca (Cundinamarca), con coordenadas geográficas de 4°18’ 45,3” N y 74°19’ 05,6” W, una altitud de 2.090 msnm, una temperatura promedio de 14,0°C y una precipitacion anual de 1.800 mm. Según el historial, hace cuatro años se cultiva el hibrido Yellow Granex (PRR) sin hacer rotación alguna, tiempo en el que se ha aplicado cal agrícola, fertilizantes químicos y altas cantidades de gallinaza para cada ciclo del cultivo, sin realizar análisis de suelo, base para efectuar un correcto plan nutricional. El proyecto fue evaluado mediante un diseño completamente aleatorizado, utilizando las siguientes fuentes y épocas de suministro para cada tratamiento; orgánico: compost, roca fosfórica, KCl, MgO y Azotobacter incorporados en trasplante; en la inoculación de Azotobacter, se utilizó cepa del mismo suelo, donde se realizó el adecuado aislamiento y multiplicación en laboratorios de la Universidad de Cundinamarca. Para orgánico–mineral se usó: CO(NH2)2, (NH4)2HPO4, MgSO4, KCl, CaNO3, Compost, adicionados en trasplante y a los 30 ddt, el convencional contó con las mismas fuentes del anterior, intercambiando compost por menores, agregados en trasplante, 30 ddt y 75 ddt; con dosis según extracción de la especie: 30, 40 y 50 t ha-1 reportada por Bertsch (2003), para dos ciclos de cultivo. Se obtuvieron nueve tratamientos con tres repeticiones, para un total de 27 unidades experimentales (tabla 1), evaluadas entre los meses de abril de 2008 a enero de 2009, obteniendo cosecha a mediados de agosto y finales de enero. Se realizaron tres análisis químicos de suelos antes del trasplante, para determinar algún tipo de Tabla 1. Tratamientos establecidos. No. Tratamiento Abreviación 1 Orgánico alto Org. alto 2 Orgánico medio Org. medio 3 Orgánico bajo Org. bajo 4 Orgánico–mineral alto Org. min. alto 5 Orgánico–mineral medio Org. min. medio 6 Orgánico–mineral bajo Org. min. bajo 7 Convencional alto Conv. alto 8 Convencional medio Conv. medio 9 Convencional bajo Conv. bajo efecto en el suelo, posteriormente al primer y segundo ciclo de cultivo, se tomó una muestra por cada tratamiento para un número de 54 análisis. En total se efectuaron 57 muestras de suelo. Las variables fisiológicas evaluadas se tomaron quincenalmente en cada ciclo de cultivo y algunas sólo en las cosechas, en cuanto a las primeras se tienen altura (Al), peso fresco planta (PFP), peso fresco bulbo (PFB), peso seco planta (PSP) y peso seco bulbo (PSB). En cosecha se tomaron datos de peso fresco bulbo primera (PFBP), peso fresco bulbo segunda (PFBS), peso seco bulbo primera (PSBP), peso seco bulbo segunda (PSBS) y en rendimiento primera calidad (RP) y rendimiento segunda calidad (RS). En cuanto a variables de suelo, se tuvieron en cuenta las arrojadas por los análisis de suelos, textura (Bouyucus), pH (Potenciómetro relación 1:1), fósforo (Bray II), carbono orgánico (Walkley Black modificado), bases intercambiables, CIC (extracción acetato de amonio 1N neutro), aluminio y acidez intercambiable (extracción con KCl 1N, titulación), densidad aparente por método del terrón parafinado. El análisis microbiológico incluyó grupos de hongos, bacterias, actinomicetos, solubilizadoras de fósforo y fijadoras de nitrógeno cuantificando poblaciones unidades formadoras de colonia (ufc). Vol. 3 - No.1 - 2009 59 60 Luengas-Gómez/Mora-Castro/Guerrero-Flórez La cuantificación de las ufc realizada mediante la técnica de conteo microbiano, para conocer el número de microorganismos viables en el suelo, realizando dilución y vaciado en placa de agar (Ferrera–Cerrato et al., 2007). El análisis estadístico se realizó mediante el programa estadístico SSPS 15. RESULTADOS Y DISCUSIóN Parámetros de crecimiento y producción Según la tabla 2, no se obtuvieron respuestas a las aplicaciones de fertilizantes en las variables peso seco planta y rendimiento segunda, las demás variables analizadas presentaron respuesta significativa y en ocasiones altamente significativa a diferentes dosis y tipos de mezclas. Se observó el caso del tratamiento Org. alto, las menores respuestas para variables fisiológicas (Al, PFP, PFB, PSP, PSB), posiblemente un efecto adverso generado por empleo de compostaje (estiércoles avícolas), que presenta altos niveles de calcio, lo que genera antagonismo con cationes, principalmente amonio, magnesio y potasio; derivando una menor disponibilidad y por consiguiente menor aprovechamiento por la planta (Jaramillo, 2001). De otra forma, el fósforo, fundamental en desarrollo y crecimiento de plantas, puede formar compuestos menos solubles y baja disponibilidad en suelo en presencia de altos niveles de calcio (Jaramillo, 2001). Lo anterior representó que tratamientos orgánicos presentaran menores respuestas fisiológicas frente a las mezclas y la fertilización convencional, se observó en PFP que los tratamientos que presentaron mayor diferencia significativa entre sí fueron Org. min. bajo y Conv. bajo, siendo este último el que mostró una mayor tasa de acumulación de biomasa durante los ciclos, que podría ser por una buena eficiencia fotosintética, provocando un incremento de la biomasa, atribuido REV. COLOMB. CIENC. HORTIC. a disponibilidad de nitrógeno, ya que en déficit las plantas tienden a disminuir peso, número de hojas y área foliar (Taiz y Zeiger, 2006). Al mismo tiempo, PFB muestra alta diferencia significativa entre los tratamientos Org. alto y medio, este último mostrando promedio superior al primero, se podría decir que existió mayor transporte de savia elaborada y agua al bulbo, produciendo un incremento en su biomasa, marcando la diferencia con los tratamientos (Taiz y Zeiger, 2006). Los datos obtenidos se hicieron bajo el modelo de un diseño completamente aleatorizado, sin ninguna estratificación. Sin embargo, la acumulación de fotosintatos no reflejó la condición anterior, como se observa en la variable PSP, en la que no existe ninguna diferencia significativa entre tratamientos, lo cual se explica desde la función de parte aérea, el transporte de nutrientes hacia los órganos de interés, el bulbo de la cebolla, disminuyendo considerablemente peso al eliminarse el contenido de agua (Denisen, 1991). Por otro lado, el PSP no siempre refleja acumulación de materia seca en el órgano de interés agronómico (bulbo). Al respecto se encontró mayor diferencia entre tratamientos Org. min. bajo y Conv. medio, siendo superior el segundo, indicando que estos bulbos quizás tienen contenido nutricional mucho menor al tratamiento convencional, lo cual puede ser por una mayor absorción de minerales. Por encontrarse estos elementos en mayor disponibilidad, es importante el manejo de la nutrición mineral, práctica que se hace más necesaria debido a sus raíces superficiales y a alta densidad de siembra usada en cebolla (Añez y Tavira, 1986). Se esperaría que las respuestas en cosecha tuvieran comportamiento similar al presentado durante el ciclo del cultivo, sin embargo se encontró que las fertilizaciones Org. medio y Org. min. bajo presentaron los valores más altos. Los tratamientos Org. medio con un peso promedio por 45,05 ab 45,67 ab 44,84 ab 44,49 ab 45,25 ab 45,38 ab 46,50 b 44,15 ab Org. medio Org. bajo Org. min. alto Org. min. medio Org. min. alto Conv. alto Conv. medio Conv. bajo 44,55 c 43,19 bc 37,34 abc 31,98 a 37,19 abc 33,66 ab 43,93 b 42,76 bc 30,63 a Peso freso planta (g) 18,08 ab 21,26 b 13,16 ab 11,81 a 11,72 a 15,58 ab 17,81 ab 14,93 ab 11,90 a Peso fresco bulbo (g) 4,97 a 4,88 a 3,67 a 3,46 a 3,32 a 3,24 a 4,70 a 4,10 a 3,15 a Peso seco planta (g) 3,52 ab 4,48 b 2,37 ab 2,23 ab 2,03 a 2,42 ab 3,89 ab 2,51 ab 1,89 a Peso seco bulbo (g) 101,19 a 126,99 ab 107,02 a 133,88 ab 105,21 a 114,58 a 110,47 a 158,63 b 118,68 a Peso fresco primera (g) Promedios con letras distintas en la misma columna indican diferencia significativa según la prueba de Tukey (P≤0,05). 42,68 a Altura (cm) Org. alto Tratamiento Tabla 2.Efecto de la fertilización en la altura, el peso de la planta y el rendimiento del bulbo. 12,74 ab 12,41 ab 13,33 ab 16,00 ab 10,39 a 10,32 a 11,77 ab 17,04 b 10,55 a Peso seco primera (g) 44,50 ab 48,18 ab 47,35 ab 53,83 bc 45,70 ab 42,93 a 46,63 ab 61,58 c 47,52 ab Peso fresco segunda (g) 3,77 a 5,34 ab 6,23 ab 5,90 ab 3,56 a 4,61 ab 6,15 ab 6,71 b 4,53 ab Peso seco segunda (g) 90,42 a 97,38 a 88,21 a 136,04 b 83,07 a 97,79 a 97,96 a 168,33 b 91,75 a 27,38 a 44,96 a 31,23 a 47,75 a 32,61 a 36,84 a 31,40 a 26,88 a 30,79 a Rendimiento Rendimiento primera (kg) segunda (kg) E valuaci ó n de fuentes de fertilizaci ó n ed á fica y el efecto sobre la microbiota Vol. 3 - No.1 - 2009 61 62 Luengas-Gómez/Mora-Castro/Guerrero-Flórez ciclo de 158,63 g y el Org. min. bajo con 133,88 g presentaron un grado de significancia (P≤0,05), lo cual encuentra coherencia con lo que reporta García (2001), quien menciona que la incorporación de abonos orgánicos bajo un correcto plan de fertilización y nutrición vegetal tiene respuestas favorables en cuanto a peso de los bulbos. El comportamiento de PSBP es similar frente al PFBP, para el cual los tratamientos con diferencia significativa (P≤0,05) son Org. medio y Org. min. bajo, mostrando respectivamente 17,04 g y 15,99 g. En cuanto a esto, Socorro (1994) reporta que el empleo de Azotobacter como parte del plan de fertilización es benéfico para obtener bulbos de primera de mayor peso. En lo correspondiente a PFBS, los tratamientos que manifestaron mayor peso y mostraron diferencia significativa (P≤0,05) fueron Org. medio y Org. min. bajo, alcanzando 61,58 g y 53,83 g, respectivamente, atribuido a un efecto positivo de la adición de compost, ya que según Gardezi et al. (1995) esta estimula la actividad biológica, haciendo que la planta se nutra mejor. Igualmente, en PSBS se halló diferencia significativa entre los mismos tratamientos, relacionándose concretamente con la variable PFBS, en la que el tratamiento orgánico en dosis media cuenta con mayor calidad, es decir, al tener mayor contenido de materia orgánica es probable que cuente con un contenido nutricional superior a los demás tratamientos (Agamennoi et al., 1998). Para RP se observó diferencia significativa de los tratamientos Org. medio con rendimiento 22,72 t ha-1 y Org. min. bajo con rendimiento 18,35 t ha-1, el cual se encuentra en un rango mayor con respecto al rendimiento nacional de 21,6 t ha-1 referenciado por Melo et al. (2006). Es de notar que sólo se ha hecho el cálculo para la producción de bulbos de primera. Para el RS no se presentaron diferencias significativas, como se había mencionado anteriormente REV. COLOMB. CIENC. HORTIC. entre los tratamientos, indicando homogeneidad en producción de bulbos segunda, lo que representó 23,67% del total de la producción. La explicación a los resultados obtenidos en cosecha es atribuible a la frecuencia de aplicación que presentaban los diferentes tratamientos, los orgánicos fueron aplicados al inicio del experimento, algo que generó un efecto contrario en las evaluaciones iniciales de las variables fisiológicas, marcando respuestas negativas en el desarrollo del cultivo, sin embargo, hacia final del ciclo la no aplicación de ningún tipo de fertilizante hizo que no se presentaran antagonismos, evidenciados en la primera parte del ciclo. Lo anterior puede corroborarse con la respuesta del tratamiento que tuvo la dosis más baja de mezcla orgánico–mineral, en la que sólo se realizaron dos aplicaciones de fertilizante y, contrario a lo encontrado en la fertilización convencional, en la cual al realizarse tres aplicaciones se obtuvieron respuestas inferiores a las esperadas, probablemente a baja disponibilidad de nutrientes generada por exceso de iones en el suelo que incrementa la conductividad eléctrica, generando estrés a la planta (Jaramillo, 2001). Parámetros edáficos En lo referente al suelo, se debe mencionar que las condiciones actuales del suelo distan notablemente de lo esperado en un Humic dystrudept que presentan una reacción fuerte–fuertemente ácida, saturación de aluminio alta y fertilidad moderada a baja, los valores encontrados se aprecian en la tabla 3. El pH del suelo está cercano a la neutralidad, la acidez intercambiable es mínima, generada por altos niveles de carbono orgánico y la fertilidad de este suelo se puede considerar alta. Lo anterior se presenta por el manejo dado, durante cuatro años, realizando aplicaciones de cales agrícolas de 2,5 t ha-1 año-1 y de gallinazas en dosis superiores a las 8 t ha-1 por ciclo. Probablemente la 63 E valuaci ó n de fuentes de fertilizaci ó n ed á fica y el efecto sobre la microbiota Tabla 3. Promedio de valores de las propiedades químicas del suelo. ppm cmol(+) kg-1 % pH P CO Ca Mg K Na Aci CICE CIC 6,96 946,20 3,87 39,87 2,80 3,23 0,30 0,10 46,16 32,65 clasificación actual ya no lo coloca en un Humic dystrudept, sino en otra ubicación taxonómica. Como se observa en la tabla 4, el pH presentó una correlación inversa con las variables fisiológicas (Al, PFP, PSP, PFB, PSB) evaluadas durante el ciclo de cultivo, ocasionando un efecto adverso en el desarrollo vegetativo, mientras que para las variables trabajadas en la cosecha tiene una correlación directamente proporcional con el PFBP, PFBS, PSBP y PSBS, corroborando de esta manera un mejor comportamiento para los tratamientos. En el rango de pH de 5,5 a 6,5, la solubilidad es la ideal, por debajo de 5,5 el Al puede ser tóxico y en el caso del P es menos disponible, mientras por encima de 6,5 los elementos menores no son disponibles (Pansu y Gautheyrou, 2003). Para el segundo ciclo, se cambió la fuente de urea por sulfato de amonio, buscando generar acidez en el suelo y, por ende, mayor disponibilidad de los nutrientes menores para la planta, lo cual se evidenció al finalizar la segunda etapa, en la que se obtuvo un pH entre 6,5 a 6,8, que sigue considerándose neutro. Esto se puede explicar teniendo en cuenta los procesos químicos que se realizan en el suelo y su influencia sobre la nutrición vegetal, como: disolución, oxidación, hidrolización y reducción (Malagón, 1995). La disolución de nutrientes puede realizarse en un medio básico o en uno ácido. Al acidificarse la solución del suelo, se disuelven los fosfatos cálcicos, pero tienden a formar los óxidos e hidróxidos de aluminio y de hierro; por otro lado, al dis- minuir la acidez de un suelo, se solubilizan parte de los fosfatos de hierro y aluminio. Por medio de la oxidación, se transforman los sulfuros en sulfatos, y de esta manera, el azufre se hace asimilable para las plantas, elemento de interés en el cultivo de cebolla cabezona (Fassbender, 1980; Zapata, 2006). El fósforo es el elemento que interviene en todos los procesos fisiológicos, por tanto, tiene alta importancia en la mejora de la calidad de los productos agrícolas (Wild y Jones, 1992). En el análisis químico se considera que un buen contenido de P se encuentra por encima de las 25 ppm, sin embargo para este caso osciló entre las 800 y 1.100 ppm, mostrando una alta riqueza en este elemento. Es importante recordar que la disponibilidad de este elemento depende del pH de la solución del suelo, sin embargo, para el ensayo, al encontrarse entre 6 y 7, se tiene una condición favorable para la absorción por las plantas (Hanke, 2000). El P no mostró diferencia significativa (P>0,05) en ninguna de las variables evaluadas, aunque los tratamientos que incluyeron en su fertilización el compost y la roca fosfórica presentaron mayores cantidades de fósforo. Cabe resaltar que, según Hanke (2000), el abonamiento con materia orgánica tiene generalmente el efecto de un mejor suministro de los fosfatos a los cultivos, ya que la descomposición de ésta, y con ayuda de los microorganismos, mejora la disposición de fósforo para las plantas. Vol. 3 - No.1 - 2009 REV. COLOMB. CIENC. HORTIC. 0,22 0,07 0,62** 0,06 0,58** 0,38** -0,10 0,64** 0,06 0,67** 0,20 0,68** 0,25 -0,19 0,70** -0,42** -0,37** -0,24 CO Ca Mg K Na AcI CICE CIC 0,49** -0,01 0,41** 0,54** -0,17 0,47** -0,12 0,03 -0,16 -0,30* 0,42** 0,12 0,27 0,46** 0,68** 0,61** 1 PFBP 0,10 -0,12 -0,21 -0,11 0,17 0,46** -0,33* -0,07 0,27* 0,24 0,25 0,46** -0,33* -0,35** -0,37** 0,43** -0,09 0,21 0,22 0,24 0,43** 0,09 0,07 -0,31* 0,11 0,14 P 0,24 -0,55** -0,51** -0,58** -0,29* pH 0,21 -0,21 0,24 -0,20 0,19 -0,21 0,27* -0,26 0,31* -0,21 -0,44** -0,43** PSBS -0,44** -0,42** -0,20 RS -0,43** -0,43** RP -0,55** -0,53** -0,61** -0,51** -0,22 1 PSBP -0,37** -0,38** -0,27* 0,98** PFBS 0,36** -0,50** -0,48** 0,80** 1 0,71** 0,26 1 PSB -0,40** -0,45** -0,27 0,78** 0,69** PSP PSP PFBP 0,66** 0,52** PFB PFB PSB 1 0,83** PFP PFP 1 Al Al -0,03 -0,28* 0,02 -0,30* 0,40** 0,33* 0,29* 0,63** 1 PSBP 0,35** -0,31* 0,08 -0,29* 0,19 -0,29* 0,22 -0,24 -0,43** -0,19 -0,28* -0,26 -0,11 -0,32* 0,51** 0,15 0,13 0,67** 0,61** 1 PFBS 1 -0,01 RP 0,26 -0,21 -0,02 -0,22 -0,32* -0,27* -0,17 -0,10 -0,31* -0,24 0,26 0,01 0,02 0,30* 0,09 0,23 -0,02 0,1 0,48** -0,19 0,17 0,09 1 PSBS 0,06 -0,23 0,29* -0,09 -0,20 -0,01 -0,23 0,07 -0,24 0,06 1 RS 0,31* -0,40** -0,08 -0,17 -0,64** 0,03 -0,36** -0,07 0,02 1 pH 0,19 0,31* 0,06 0,07 0,15 0,42** 0,29* 0,32* 1 P 0,12 0,25 -0,06 -0,19 0,12 0,23 0,25 1 CO Ca -0,09 0,99** -0,31* 0,24 0,59** 0,11 1 Tabla 4. Análisis de correlación de las variables de respuestas fisiológicas y algunas propiedades químicas del suelo. -0,01 0,16 0,04 -0,03 0,11 1 Mg -0,30* 0,66** -0,10 0,15 1 K -0,26 0,26 -0,16 1 Na 1 1 CICE 0,40** -0,12 -0,29* AcI 1 CIC 64 Luengas-Gómez/Mora-Castro/Guerrero-Flórez E valuaci ó n de fuentes de fertilizaci ó n ed á fica y el efecto sobre la microbiota Tabla 5. Efecto de la fertilización sobre algunos grupos microbiológicos (en ufc/g de suelo seco a la dilución de 10). Tratamiento Fijadoras de N Solubilizadores de P Hongos Actinomicetos Bacterias Org. alto 87,25 abc 57,25 a 75,00 a 35,16 a 99,25 a Org. medio 95,08 abc 47,83 a 55,91 a 57,58 a 80,75 a Org. bajo 53,66 a 93,25 a 34,08 a 91,83 a Org. min. alto 105,83 c 90,91 abc 47,50 a 88,66 a 39,41 a 90,41 a Org. min. medio 66,83 abc 53,91 a 58,08 a 45,50 a 103,83 a Org. min. bajo 63,91 ab 56,91 a 49,83 a 43,25 a 73,75 a Conv. alto 84,91 abc 47,75 a 93,58 a 19,66 a 100,08 a Conv. medio 99,41 b 49,00 a 84,16 a 51,25 a 98,50 a Conv. bajo 55,75 a 48,00 a 109,91 a 37,08 a 84,83 a Promedios con letras distintas en la misma columna indican diferencia significativa según la prueba de Tukey (P≤0,05). Otro factor importante que atañe a este elemento es el tipo de arcillas del suelo. Para el ensayo se encontró que, al ser de un origen ándico, es posible la presencia de alófanas, las cuales pueden llegar a fijar este elemento hasta un 85%, no obstante al poseer tan altas cantidades aún se tendría una buena cantidad disponible a las plantas. Revisando la correlación, se encuentra este elemento vinculado de manera significativa e inversa con la producción de cebolla de segunda, lo que se le podría atribuir a la ley de los rendimientos decrecientes, en la que se indica que, a medida que aumenta el insumo (fertilizante), se llega a un punto en que la producción disminuye (Fremont, 1961; Zapata, 2006). La necesidad del cultivo en calcio (Ca) es baja, hasta en suelos ácidos no se restringe la disponibilidad de iones de Ca para abastecer las exigencias de la cebolla, además varios fertilizantes lo contienen como componente secundario. Al lote donde se trabajó, por varios años se le ha incorporado cantidades altas de cal. Sin haber realizado algún análisis de suelo, se encontró un porcentaje de saturación mayor al 80%. Se encontró correlación directa entre los niveles de calcio en el suelo con la producción. Por otra parte, altos contenidos de este elemento inhiben la absorción de Fe, B, Zn, Cu y Mg, porque estos elementos pierden su solubilidad con un pH mayor de seis (Zapata, 2006). El contenido de magnesio (Mg) se encontró por debajo de los niveles ideales, localizándose en un rango de 6 a 8% de saturación, lo cual se debe a la interacción con el calcio, que, como ya se mencionó, inhibe otros elementos, por su alta presencia en el suelo. Se encontró además correlacionado de manera altamente significativa y positiva con la altura de la planta, lo cual se manifiesta, posiblemente, porque el Mg forma parte de la molécula esencial de las plantas (clorofila) y se fomenta el crecimiento y desarrollo vegetativo de éstas (Wild y Jones, 1992). El potasio (K) tiene una relación altamente significativa y positiva con las siguientes variables: altura, peso fresco y seco de planta y bulbo, de manera un poco menos significativa con la producción de bulbo de primera; ya que este elemento se encontró por encima del óptimo de saturación en el suelo, lo que reporta incluso protección contra ataque de hongos, porque este elemento forma fibras más fuertes, haciendo a las plantas más resistente contra este tipo de ataques, además crea resistencia contra la sequia, ya que disminuye la traspiración (Hanke, 2000). Vol. 3 - No.1 - 2009 65 66 Luengas-Gómez/Mora-Castro/Guerrero-Flórez Dogliotti y Galvan (2008) determinan que para el uso entre cloruros o los sulfatos se debe tener presente el cultivo, ya que plantas como la papa, el tabaco, frutales y hortalizas prefieren los sulfatos, mientras que plantas halófitas, como la remolacha, la caña de azúcar y forrajes, prefieren los cloruros. El sodio (Na) no se considera un elemento esencial, pero puede reemplazar algunas funciones del K. Se observa una alta correlación de Pearson entre el el peso fresco de la planta y el bulbo. Parámetros microbiológicos Las bacterias se encontraron en abundancia en todos los tratamientos y no mostraron diferencias entre los tratamientos, ni entre los ciclos. En cuanto a la correlación de Pearson, se hallaron relacionadas con un nivel de significancia de 0,05 de forma directa con las siguientes variables: altura, peso fresco de planta y bulbo, además de manera inversa se correlacionaron con el pH. Esto último se puede dar debido a que las bacterias prefieren pH cercanos a la neutralidad; en el experimento, el pH, a pesar de encontrarse en un rango de neutralidad, pasó de 7,2 a 6,5, por lo que se diría que son susceptibles a estas variaciones, ya que se incrementaron en el segundo ciclo, pasando en promedio de 83 ufc a 100 ufc. Durante el estudio, los hongos no mostraron diferencia significativa para el número de sus poblaciones, sin embargo los tratamientos que evidenciaron una mayor riqueza en estos microorganismos fueron los convencionales en sus tres dosis, estando inclusive por encima de las bacterias en el tratamiento Conv. bajo (hongos 33% y bacterias 25%). Los solubilizadores de fósforo presentaron mejores respuestas en cuanto a los tratamientos con mejor rendimiento, fueron aquellos que coinciden con la mejor presencia de este tipo de microorganismos. El porcentaje promedio que se manejó de estas poblaciones en los diferentes tratamientos fue de 15%, el cual es considerado bueno, teniendo en cuenta la intensa actividad microbiológica que aportan al suelo. En cuanto a las fijadoras de nitrógeno, la correlación de Pearson se determinó relacionada con todas las variables fisiológicas en forma altamente significativa y directa, indicando que, a mayor proporción de ufc de estos microorganismos en el suelo, la planta presenta una mejor nutrición que conlleva a un óptimo desarrollo vegetativo y reproductivo. CONCLUSIONES En lo referente a los actinomicetos, es importante reconocer que los tratamientos Org. medio y Org. min. bajo nuevamente mostraron mayor cantidad de estos microorganismos, mostrando su intrínseca relación con el desarrollo y productividad de las plantas. El suelo del estudio demostró mediante los análisis químicos que el manejo convencional de éstos ha generado una modificación de las condiciones naturales, elevando las cantidades de los elementos por encima de los niveles críticos, por lo cual posiblemente no se requieran aplicaciones de fertilizantes de forma continua. Con este grupo de microorganismos se encontró una correlación inversa (P≤0,05) con el carbono orgánico y el magnesio, lo que puede darse porque algunos actinomicetos contribuyen a la precipitación de CaCO3 en el suelo, alterando de esta manera el pH (Damian, 1992). Los suelos enriquecidos por aplicaciones de materia orgánica y enmiendas con cales generan niveles de elementos por encima de niveles críticos, en los que la respuesta en la producción se atribuye más a una mejora en la condición química del suelo que a un aporte de nutrientes a través de la fertilización. REV. COLOMB. CIENC. HORTIC. E valuaci ó n de fuentes de fertilizaci ó n ed á fica y el efecto sobre la microbiota El análisis de suelo forma parte del manejo de la fertilización y debe considerarse como una buena herramienta para la toma de decisiones. Es posible que el aumento de los microorganismos se haya debido a condiciones climáticas de la zona en el tiempo de evaluación y no a los tratamientos empleados, ya que se observaron poblaciones de éstos inclusive en tratamientos como el convencional, por lo que se cree que es más un efecto ambiental que del experimento. La microbiota edáfica es un factor bastante relevante a considerar cuando se planea el establecimiento del cultivo. Ya que el establecimiento de una flora microbiana permite una mayor asimilación de los nutrientes por las plantas y, por tanto, la cantidad de fertilizantes a utilizar puede llegar a reducirse. Es necesario realizar más investigación sobre los grupos de microorganismos, ya que por su capacidad de producir sustancias antagonistas y benéficas (Gutiérrez, 2000) se podría explotar en mayor proporción su potencial en agricultura y en la industria. REFERENCIAS BIBLIOGRÁFICAS Abril, A. 2002. La microbiología del suelo: su relación con la agricultura sustentable. pp. 129-150. En: Sarandon, S.J. (ed.). Agroecología. El camino hacía una agricultura sustentable. Ediciones Científicas Americanas, La Plata, Argentina. Agamennoi, R.; J. Rivas; R. Delhey y A. Azpilicueta. 1998. Abonos verdes de verano en la producción de cebolla. Resúmenes XXI Congreso Argentino de Horticultura. San Pedro, Buenos Aires. Añez, B. y E. Tavira. 1986. Aplicación de N, P y K a diferentes poblaciones de plantas de cebolla. Turrialba 36(2), 163-169. Borrero, S. 2000. Estudio general de suelos y zonificación de tierras. IGAC, Bogotá. Burbano, H. 1989 El suelo: una visión sobre sus componentes biorgánicos. Universidad de Nariño, Pasto, Colombia. Damian, C.A. 1992. Actinomicetos y su efecto antagónico sobre Phytophtora cinnamomi Rands (tristeza del aguacate) bajo la incorporación de dos mejoradores orgánicos del suelo y un fungicida. Tesis. Departamento de Parasitología Agrícola, Universidad Autónoma de Chapingo, México. Denisen, E.L. 1991. Fundamentos de la horticultura. 2a. ed. Editorial Limusa, México. Dogliotti, S. y G. Galvan. 2008. Bases fisiológicas del crecimiento y desarrollo de cultivo de cebolla (Allium cepa L.). p. 19. En: Facultad de Agronomía, Universidad de la Republica, Uruguay, http://www.fagro. edu.uy/~cultivos/hortalizas/Repartido_Fisiologia_Cebolla.pdf; consulta: marzo de 2009. Fassbender, H.W. 1980. Quimica de los suelos. IICA, San José, Costa Rica. Ferrera–Cerrato, R. y A. Alarcon. 2007. Microbiologia agricola. Hongos, bacterias, micro y macrofauna, control biológico y planta-microorganismo. Trillas, México. Fremont, P. 1961. Economía agrícola. Aguilar, Madrid. García, J. 1963. Fertilización agrícola. Ediciones Agrociencia, Zaragoza, España. García, M. 2001. Evaluación de la respuesta de una sucesión de cultivos hortícolas a diferentes abonos orgánico. Universidad de la Republica, Montevideo. Gardezi, A.K. 1995. Endomycorrhiza, rock phosphate, and organic matter effects on growth of Erythrina americana. Nitrogen Fixing Tree Res. 13, 48-50. Gutiérrez, L.M.T. 2000. Potencial herbicida de metabolitos aislados de especies selectas de actinomicetos. Actinomyces MIV2B (31) como una fuente novedosa del antibiótico maduramicina. Tesis de Doctorado. UNAM, México. Hanke, F. 2000. La nutrición de la planta y su problemática en la agricultura. Edición Talleres Gráficos Fundación Universitaria Juan de Castellanos, Tunja, Colombia. Vol. 3 - No.1 - 2009 67 68 Luengas-Gómez/Mora-Castro/Guerrero-Flórez Instituto Colombiano Agropecuario. 1989. Análisis de suelos, plantas y aguas para riego. ICA, Bogotá. Scharrer, K.1960. Química agrícola. Editorial Hispano Americana, México. Jaramillo, D. 2001. Introducción a la ciencia del suelo. Escuela de Geociencias, Universidad Nacional de Colombia, Medellín, Colombia. Socorro, M.; N. Bouza; O. Morales; P. Meneses; R. Cabello y B.Dibut. 1994. Empleo del Azotobacter chroococcum en la fertilización nitrogenada del arroz. p. 113. En: Resúmenes XVII Reunión Latinoamericana de Rhizobiología. La Habana. Malagón, D.; C. Pulido; R. Llinas y C. Chamorro. 1995. Suelos de Colombia, origen, evolución, clasificación, distribución y uso. Canal Ramírez Antares, Bogotá. Melo, L.I.; M.M. Melo y L.F. Rodríguez. 2006. Competitividad del sistema agroalimentario de la cebolla de bulbo con enfoque de cadena productiva en el distrito de riego del alto Chicamocha (Boyacá). Agron. Colomb. 24(2), 378-389. Pansu, M. y J. Gautheyrou. 2003. Handbook of soil analysis. Mineralogical, organic and inorganic methods. Springer-Verlag, Berlin. REV. COLOMB. CIENC. HORTIC. Wild, A. y L.H.P. Jones. 1992. Nutrición mineral de las plantas cultivadas. pp. 73-119. En: Wild. A. (ed.). Condiciones del suelo y desarrollo de las plantas según Russel. Ediciones Mundi-Prensa, Madrid. Taiz, L. y E. Zeiger. 2006. Plant physiology. 4th ed. Sinauer Associates Publ., Massachussetts, MA. Zapata, H. 2006. Química de los procesos. Pedogenéticos. Universidad Nacional de Colombia, Medellín, Colombia. Efectos del cambio climático sobre la oferta y precios de cebolla de bulbo. Caso del municipio Duitama, Boyacá (Colombia) Climatic change effects on the supply and prices of bulb onion. Case of Duitama municipality, Boyacá (Colombia) Juan Carlos Barrientos F.1, 2 Germán A. Leyva V.1 Juan Camilo Morales B.1 Riego de cultivo de cebolla en Duitama, Boyacá. Foto: Usochicamocha RESUMEN La cebolla (Allium cepa L.) es la segunda hortaliza más consumida en Colombia. La producción y los precios de mercado de esta hortaliza están determinados por varios factores, entre ellos el clima. Últimamente, el clima ha experimentado cambios en su comportamiento. Estos cambios están afectando, entre otros, la productividad agraria. En el caso de la cebolla, se presume que el cambio climático está afectando tanto la producción, como la oferta y los precios de la cebolla de bulbo. En ese sentido, el objetivo de esta investigación es determinar el modo en que el cambio climático está afectando la producción y precios de la cebolla de bulbo, basado en un estudio de caso. Se analizaron datos climáticos de la zona de estudio, distrito de riego Alto Chicamocha en Duitama, Boyacá, y datos de producción y precios del departamento y del país. Se realizaron entrevistas con productores y un experto de la zona en cuestión. Con esta información se ha llegado a los siguientes resultados y conclusiones: el cambio climático es un fenómeno presente en la zona de estudio, caracterizado, entre otros, por la disminución del brillo solar y de la humedad relativa, incremento de la precipitación, evaporación y temperatura; sus efectos han determinado cambios en el manejo del cultivo, como incremento de dosis y frecuencia de aplicación de fungicidas principalmente, pero no han incidido de manera determinante en la cantidad de producción (oferta local); los precios de la cebolla son afectados por la importación y las expectativas (de mayor ganancia) de los productores más que por el cambio climático. Palabras clave adicionales: clima, brillo solar, temperatura, oferta, precios. 1 2 Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá (Colombia). Autor para correspondencia. [email protected] REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 3 - No.1 - pp. 69 - 80, 2009 70 Barrientos/Leyva/Morales ABSTRACT Onion (Allium cepa L.) is the second vegetable most consumed in Colombia. The production and the market prices of this vegetable are determined by many factors, among them, the climate. Lately, the climate has undergone changes in behavior. These changes are affecting, among others, the agrarian productivity. In onion case, it is presumed that the climatic change is affecting so much the production as the supply and the prices of the bulb onion. In that sense, the objective of this research is, based on a case study, to determine the way in that the climatic change is affecting the production and prices of bulb onion. Climatic data of the study zone, irrigation district of Alto Chicamocha in Duitama, Boyacá, have been analyzed, as well as production and prices data of the department and the country. Interviews with producers and the expert of the zone have been realized. With this information the research has reached the following results and conclusions: the climatic change is a present phenomenon in the zone of study, characterized, among others, by the diminution of solar brightness and relative humidity, an increase of precipitation, evaporation and temperature; their effects have determined changes in crop management, such as increase of dose and frequency of application of fungicides mainly, but these have not affected in a determent form the amount of production (local supply); the onion prices are affected by the import and the expectations (of greater gain) of the producers more than by the climatic change. Additional keywords: climate, solar intensity, temperature, supply, prices. Fecha de recepción: 11-05-2009 Aprobado para publicación: 01-06-2009 IntroduCCIÓN Oferta de cebolla en Colombia La cebolla (Allium cepa L.) es la segunda hortaliza más consumida en Colombia. Según la CCI (2003), el consumo per cápita nacional de cebolla (junca o larga y de bulbo o cabezona) es de 6 kg/persona-año, mostrándose un incremento aproximado del 2,7% entre 1997 y 2001 (Agronet, 2009). La cebolla que se vende en Colombia proviene principalmente de la oferta nacional, aunque en los últimos años la oferta de cebolla importada ha incrementado sus volúmenes, de 11.028 t en 1997 a 71.826 t en 2008 (Agronet, 2009). La producción nacional, que en 1997 era de 897.199 t, ha tenido un descenso considerable, tal que para 2008 fue de 568.457 t. Este descenso se debió principalmente a la disminución de la producción anual de cebolla junca, la que bajó de 666.604 t en 1997 a 291.305 t en 2008, mientras que la producción anual de cebolla de bulbo se mantuvo oscilando entre 200.000 (1998) y 300.000 t (2007) aproximadamente, con una REV. COLOMB. CIENC. HORTIC. clara tendencia de incremento de cerca del 3% anual. La superficie dedicada a la producción de cebolla en Colombia en los últimos 12 años ha oscilado alrededor de 20.000 ha, según las estadísticas 26.714 ha en 1997, 16.286 ha en 2002 y 31.384 ha en 2008 (Agronet, 2009). Del mismo modo que la producción, la superficie nacional de cultivo de cebolla junca ha disminuido de 15.110 ha en 1997 a 14.868 ha en 2008; mientras que el cultivo de cebolla de bulbo se ha incrementado paulatinamente, de 11.604 ha en 1997 a 16.516 ha en 2008 (Agronet, 2009). Por otra parte, el rendimiento de cebolla de bulbo en los últimos 12 años se ha mantenido alrededor de 20 t ha1; mientras que el de cebolla junca ha bajado de 44,1 t ha-1 en 1997 a 19,5 t ha-1 en 2008 (Agronet, 2009). Actualmente, son siete los departamentos donde se cultiva cebolla de bulbo, de los cuales, para el año 2008, Boyacá aportó con 49% de la producción nacional, Norte de Santander con 23% y Cundinamarca 20%. La superficie de cultivo en Boyacá osciló notablemente, por ejem- E fectos del cambio clim á tico sobre la oferta y precios de cebolla de bulbo plo en 1998 fue de 3.686 ha, en 2001 de 4.005 ha, en 2002 de 5.553 ha, en 2007 de 6.149 ha y en 2008 de 4.262 ha. Por el contrario, en Norte de Santander la superficie de cultivo ha tenido un incremento sostenido de aproximadamente un 14% anual. Entretanto, también Cundinamarca incrementó su superficie de cultivo, pero recién a partir de 2005, en aproximadamente un 44% anual. Boyacá, aunque con oscilaciones de casi 5 t ha-1, ha venido incrementando sus rendimientos, por ejemplo de 24 t ha-1 en 1998 a 28 t ha-1 el 2006 y a 32 t ha-1 el 2008. Por el contrario, Norte de Santander, con un promedio aproximado de 21 t ha-1, y Cundinamarca, con un promedio aproximado de 17 t ha-1, han experimentado en los últimos dos años descensos notables (6 a 10 t ha-1) en sus rendimientos (Agronet, 2009). Factores determinantes de la oferta de cebolla Los principales factores que determinan la oferta de cebolla de bulbo, como en otros cultivos, son, entre otros, los precios del producto, los costos de los factores de producción, efectos climáticos y fitosanitarios y la tecnología de producción. Precios y costos de producción de la cebolla Los precios del producto incentivan o desaniman a los productores, a su vez, la cantidad de oferta en el mercado determina la subida o bajada de precios. En el caso de la cebolla de bulbo, los precios promedio en Corabastos (Bogotá) correspondientes a los años 1997 (511 $/kg) hasta 2002 (792 $/kg) muestran un comportamiento más o menos regular de bajadas y subidas, atribuible al efecto del teorema de la telaraña (Ezekiel, 1938), después del cual se advierten oscilaciones leves hasta 2007 y luego un incremento notorio entre 2007 (831 $/kg) y 2008 (1063 $/kg). Según datos del Banco Agrario de Colombia (2008), el costo de producción desde semillero de una hectárea de cebolla de bulbo (cabezona) en Boyacá asciende a cerca de diez millones de pesos, de los cuales aproximadamente un 15% corresponde a aplicación de riego, un 10% a semilla (Agrocadenas, 2005, reporta un 23%), un 10% a fertilizantes, un 3% a control de plagas y enfermedades, un 1,5% a fungicidas y un 0,5% a fertilización. El gasto en mano de obra abarca cerca del 50% de los costos totales de producción, de los cuales el riego, la cosecha y siembra (trasplante) son las labores de mayor demanda de trabajo. Factores fitosanitarios Si bien el costo del control de enfermedades y de fungicidas es porcentualmente bajo (3-5%), su importancia en el proceso de producción de cebolla es mayor, ya que de ello depende, en gran medida, el éxito o fracaso del negocio. Dos enfermedades son especialmente restrictivas para el cultivo de cebolla: el tizón (Peronospora destructor) y la alternaria (Alternaria porri) (Agrios, 2005; Maroto et al., 1995). Su control depende tanto del manejo del cultivo como de las condiciones climáticas de la zona y periodo del año. Cambio climático y producción de cebolla El fenómeno del cambio climático, expresado a través de cambios y variaciones en las temperaturas, precipitaciones y concentraciones de CO2 en la atmósfera, está afectando, entre otros, la productividad agraria, especialmente de las zonas tropicales del planeta (Kurukulasuriya y Rosenthal, 2003). En el caso de la producción de cebolla, los cambios en la forma, la cantidad, la frecuencia e intensidad de las precipitaciones han determinado un nuevo régimen de humedad del suelo y del aire que, sumado a los cambios en las variaciones diarias, mensuales y anuales de la temperatura, ha creado un nuevo ambiente, muchas veces más favorable, para la proliferación de enfermedades (Brewster, 1997). Planteamiento del problema La producción de cebolla depende, entre otros, de los factores climáticos. La precipitación determina en el cultivo de cebolla de bulbo la cantidad y frecuencia de riego o drenaje. La luminosidad (horas luz) influye en la calidad y tamaño del Vol. 3 - No.1 - 2009 71 72 Barrientos/Leyva/Morales bulbo. La humedad relativa y la temperatura, así como sus oscilaciones, determinan las condiciones favorables o desfavorables para enfermedades que atacan a la especie en estudio. Este escenario hace suponer que también en Colombia la producción de cebolla, y por ende la oferta y precios del producto, está siendo afectada por el cambio climático. Para confirmar o rechazar el supuesto mencionado, se ha tomado como objeto de estudio una zona productora de cebolla en Boyacá, la zona del distrito de riego Alto Chicamocha. Objetivo general Determinar los efectos que produce el cambio climático en los cultivos de cebolla de bulbo en la región del Alto Chicamocha (Boyacá). Objetivos específicos • Determinar las características del cambio climático en la zona. Zona de estudio El distrito de Riego y Drenaje de Gran Escala Alto Chicamocha y Firavitoba abarca los municipios de Paipa, Duitama, Santa Rosa, Tibasosa, Nobsa, Sogamoso y Firavitoba. Tiene una superficie bruta de cubrimiento de 11.300 ha, un área de drenaje de 9.258 ha y un área de infraestructura de riego de 7.335 ha. Sus usuarios beneficiarios son 6.200, de los cuales el 76,7% tiene parcelas agrarias menores a 1 ha, 18,5% tiene parcelas entre 1 y 5 ha, 3,23% tiene parcelas entre 5 y 10 ha y sólo 1,37% tiene parcelas mayores a 10 ha. El área dedicada a producción agrícola es de 2.500 ha y a producción pecuaria, principalmente lechería, 6.758 ha. Según Usochicamocha (2008), en la zona se han destinado 2.300 ha a la producción de cebolla de bulbo, 12 ha para brócoli, 10 ha para coliflor, 70 ha para arveja y 35 ha para repollo. Para el presente estudio se ha considerado solamente el municipio Duitama y en éste la zona productora de cebolla de bulbo. • Determinar las manifestaciones del cambio climático y sus efectos en la producción de cebolla de bulbo en el distrito de riego Alto Chicamocha. Tipo de información y su obtención • Determinar las medidas adoptadas por los productores para contrarrestar los efectos del cambio climático en la producción de cebolla de bulbo. • Información mensual de los parámetros climáticos desde 1990 hasta 2008: brillo solar, evaporación, humedad relativa, precipitación, precipitación en 24 h, número de días con lluvia, temperatura media, temperatura máxima y temperatura mínima. Esta información, procedente de la estación Surbatá–Bonza del municipio Duitama, se obtuvo del Ideam (2009). • Determinar los efectos de la producción en la oferta (producción nacional e importación) y precios de la cebolla de bulbo. MÉTODO Esta investigación es, en una proporción mayor, de tipo exploratoria, sobre todo cuando se trata de la información primaria. Sin embargo, la información secundaria permite hacer análisis descriptivos y en algunos casos causales. REV. COLOMB. CIENC. HORTIC. La información requerida para este trabajo fue la siguiente: • Información de oferta de cebolla en Colombia desde 1997 hasta 2008: superficie de producción, producción anual, rendimiento e importación anual por países (Agronet, 2009). • Información de precios promedio mensual de mayoristas de cebolla de bulbo desde 1997 hasta 2008 (Corabastos, 2009; CCI, 2009). E fectos del cambio clim á tico sobre la oferta y precios de cebolla de bulbo • Información primaria sobre el efecto del cambio climático en la producción de cebolla. Esta información se obtuvo de seis productores de cebolla “representativos” del municipio Duitama a través de entrevistas. Las entrevistas, con una duración aproximada de 25 a 40 min, se realizaron el 25 de junio de 2009, con base en un cuestionario de 20 preguntas abiertas. El cuestionario contenía, entre otros, los temas: presencia–ausencia del cambio climático en la zona, manifestación del cambio climático, efectos del cambio climático en la producción de cebolla, medidas adoptadas para contrarrestar el efecto del cambio climático, factores que influyen en la fijación de precios de la cebolla, efectos del cambio climático en el precio de la cebolla y algunas características de las fincas productoras de cebolla de bulbo de la zona de estudio. Paralelamente, se entrevistó al jefe del Departamento de Planeación y Desarrollo Productivo del Distrito de Riego para tratar temas relativos a la producción de cebolla: superficie, producción, rendimientos, problemas más importantes, etc. La entrevista, con una duración de aproximadamente 3 h, se llevó a cabo mediante un cuestionario no estructurado con preguntas abiertas. • Análisis de la importación de la cebolla. Análisis de la información • Análisis de los efectos del cambio climático en los precios de la cebolla. Información mensual y anual de los parámetros climáticos • Descripción de las características climáticas de la zona de estudio con base en datos anuales acumulados o promedio. • Determinación de la regularidad o irregularidad de ocurrencia de los eventos climáticos. Información de oferta de cebolla en Colombia • Análisis de la producción nacional. • Relación de la producción con los parámetros de clima. • Comparación de la oferta nacional e importación de cebolla con los precios nacionales. Información de precios promedio mensual de mayoristas de cebolla de bulbo • Análisis comparativo de los precios por meses. • Análisis comparativo de los precios por años. • Relación de los precios con la oferta nacional e importación de cebolla de bulbo. • Relación de los precios con algunos factores climáticos. Información primaria sobre el efecto del cambio climático en la producción de cebolla • Análisis de los efectos del cambio climático en la producción de cebolla. • Análisis de las medidas adoptadas por los productores para contrarrestar el efecto del cambio climático en la producción de cebolla. RESULTADOS Y DISCUSIÓN Características del cambio climático en la zona De los datos climáticos presentados en la tabla 1, lo que más se destaca, caracterizando a la zona, es lo siguiente: • En promedio, hay 5 h diarias de brillo solar. Esto podría ser restrictivo para variedades de cebolla de bulbo que requieren de más horas luz para desarrollo foliar o llenado de bulbo. Vol. 3 - No.1 - 2009 73 REV. COLOMB. CIENC. HORTIC. 1.715 1.831 1.726 1.838 1.702 1.782 1.784 1.967 1.782 1.740 1.884 1.973 1.885 1.793 1.968 1.811 1.611 1.578 1.697 1.793 (h año-1) 48 68 83 75 111 83 103 116 87 98 130 109 129 155 95 106 118 134 98 102 Rango* 1.171 1.272 1.261 1.218 1.169 1.201 1.208 1.399 1.389 1.247 1.225 1.398 1.408 1.347 1.223 1.311 1.256 1.216 1.240 1.271 (mm año-1) 47 30 52 31 40 43 40 44 65 63 47 43 44 56 25 63 26 54 57 46 Rango* Evaporación 80 79 80 84 78 78 80 75 77 80 79 69 74 81 86 83 81 76 77 79 Promedio anual Precipitación 11 18 10 11 25 15 8 9 13 7 7 11 12 14 7 5 17 9 7 11 872 760 556 961 895 887 918 669 964 1.091 856 611 719 1.053 906 904 1.003 934 882 865 198 150 88 180 153 127 102 98 196 130 104 104 132 225 175 161 139 124 142 144 Rango* (mm año-1) Rango* Humedad relativa (%) Fuente: Elaborado con base en datos del Ideam, 2009. * Diferencia entre el mes del año con el valor más alto y el mes del año con el valor más bajo. 1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 Promedio Años Brillo solar 49 23 32 32 32 27 32 36 54 32 33 25 31 42 56 37 33 39 36 36 (mm) 43 18 30 26 31 18 23 29 54 25 28 20 28 40 51 29 26 31 29 30 Rango* Precipitación máxima en 24 h 166 157 139 188 195 201 202 147 199 206 184 158 152 173 158 172 180 166 171 174 Anual 14 17 15 20 22 21 14 17 29 15 15 17 15 22 21 16 15 17 18 18 Rango* No. de días con lluvia/año Tabla 1. Principales datos climáticos anuales de la estación Surbatá–Bonza del municipio Duitama, Boyacá (1990-2008). 14 14 14 14 14 14 14 14 15 14 14 14 14 14 15 15 15 15 15 14 Anual 1 2 2 2 2 2 2 1 3 1 1 2 2 1 1 1 3 2 1 2 Rango* Temperatura media (°C) 24 26 27 24 25 28 24 25 25 28 24 26 25 29 29 25 24 26 27 26 Anual 2 4 5 2 4 7 3 3 2 6 2 3 3 6 6 3 4 2 4 5 Rango* Temperatura máxima (°C) -1 -3 -2 -3 -1 -4 -1 -2 0 1 -1 -3 0 1 -4 1 0 -8 0 -2 Anual 5 7 7 8 6 10 7 7 6 5 6 8 6 5 10 6 6 13 5 7 Rango* Temperatura mínima (°C) 74 Barrientos/Leyva/Morales E fectos del cambio clim á tico sobre la oferta y precios de cebolla de bulbo • La evaporación es, en promedio, mayor a la precipitación en aproximadamente 400 mm. Esto significa, de manera general, que para la producción vegetal se necesita un aporte complementario de agua al suelo. • Cerca de la mitad del tiempo está lloviendo, lo que favorece el mantenimiento de la humedad del ambiente. • La humedad del ambiente es relativamente elevada. Esta es una condición favorable para la proliferación de enfermedades en el cultivo de la cebolla. • Las precipitaciones en 24 h están alrededor de 36 mm. Esto podría significar, en algunos casos, encharcamiento de las parcelas de cebolla creándose un ambiente favorable para el desarrollo de enfermedades del cultivo. • Si bien la temperatura media se ha mantenido entre 14°C y 15°C, el rango entre temperaturas máximas y mínimas es mayor a 25°C. Esto puede ir en detrimento del cultivo de cebolla, ya que las temperaturas bajas reducen o detienen el crecimiento, pudiendo dañar también los tejidos de la planta. Por otra parte, las temperaturas mayores a 23°C favorecen el desarrollo de la planta, pero también de enfermedades del cultivo. 75 Mediante los “rangos”, presentados en la tabla 1, se desea hacer notar el comportamiento regular o irregular de los parámetros climáticos durante el periodo de estudio. Teóricamente, si los eventos climáticos se repitieran y ocurrieran con regularidad, los rangos entre los valores máximos y mínimos mensuales del año serían similares en todos los casos. Sin embargo, la tendencia de los valores hallados es de incremento en la diferencia entre los extremos, es decir, los eventos climáticos se estarían haciendo más variables y extremos (tabla 1). Llaman la atención especialmente el brillo solar, la humedad relativa y las temperaturas máximas. Este fenómeno de mayor diferencia entre extremos dificultaría aún más la precisión en la planificación de la producción vegetal. La tabla 2 muestra que, según las tendencias lineales, a largo plazo: • Hay un incremento en la temperatura media, máxima y mínima. Esto podría favorecer al desarrollo del cultivo de cebolla, pero también a la proliferación de enfermedades. • A consecuencia del incremento de temperatura, hay un incremento de la evaporación, lo que hace pensar en una mayor necesidad de riego para el cultivo. Sin embargo, también hay un Tabla 2. Tendencia lineal de los principales parámetros climáticos de la estación Surbatá–Bonza del municipio Duitama, Boyacá (1990-2008). Ítem Promedio Brillo solar Evaporación Humedad relativa (h día-1) (mm año-1) Promedio anual (%) PrecipiPrecipitación No. tación máxima en de días 24 horas (mm año-1) (mm) con lluvia/ año Temperatura media anual Temperatura máxima anual Temperatura mínima anual (°C) (°C) (°C) 5 1271 79 865 36 174 14 26 -2 Tendencia de valores entre 1990 y 2008 (%) - 2,3 +5 -1 + 16 + 17 - 2,4 + 7,8 +5 + 21 Tendencia de los rangos (%)* + 73 + 19 + 40 +1 + 13 0 - 25 + 42 + 14 (1990-2008) Fuente: Elaborado con base en datos del Ideam, 2009. * Con base en los datos de la Tabla 1. Vol. 3 - No.1 - 2009 76 Barrientos/Leyva/Morales incremento en la precipitación, y en mayor proporción que la evaporación, por lo que la necesidad de riego tendería a reducirse. • Como efecto del incremento de la precipitación, las horas luz tienden a disminuir. Esto podría incidir negativamente en la ganancia de peso y calidad del bulbo de la cebolla. • Hay un incremento en la intensidad de las lluvias y una disminución en su frecuencia, es decir, las lluvias serán más cortas, más fuertes y menos frecuentes, lo que podría llegar a ser menos favorable para la producción vegetal. Manifestaciones del cambio climático y sus efectos en la producción de cebolla de bulbo en el distrito de riego Alto Chicamocha Los productores entrevistados y sus parcelas de producción presentan características que se resumen a continuación: Los productores entrevistados tienen basta experiencia en el cultivo de cebolla de bulbo. En atención a la superficie de cultivo, se diferencian en pequeños, medianos y grandes productores. Los rendimientos que obtienen en esta zona están entre los más altos del país, favorecidos, entre otros, por la disponibilidad de riego. La mayoría siembra cebolla dos veces al año, pero también hay agricultores que han escalonado las siembras, llegando a valores de hasta seis por año. Algo que llama la atención es la frecuencia de riego. Según el técnico entrevistado, los riegos se realizan rutinariamente (1-3 horas por riego) y en abundancia, siendo común ver parcelas casi encharcadas después de un riego. Dado que la mayoría de las propiedades agrícolas son pequeñas (<1ha), los productores de cebolla optan por arrendar o cultivar en compañía, así las parcelas de cebolla son itinerantes. Otro motivo para la itinerancia de las parcelas de cultivo de cebolla, según uno de los entrevistados, es la infección de los suelos (después de cuatro años seguidos de cultivo de cebolla) con el “tizón” (Peronospora destructor) y la “al- REV. COLOMB. CIENC. HORTIC. ternaria” (Alternaria porri) y la disponibilidad de riego. Es así que la mayoría de los productores de la zona de estudio proceden de otros municipios como: Samacá, Sáchica, Villa de Leyva, Sutamarchan, Tinjacá, Choachí y Cáqueza. Todos los entrevistados manifestaron haber percibido cambios en el clima. Los cambios a los que hicieron referencia son los siguientes: • El ambiente está más seco. • La temperatura es mayor, hace más calor. • Hay mayor presencia de niebla, por tanto más frío por las mañanas y menor brillo solar. • Las lluvias son más irregulares. Las épocas de lluvia, que eran bien marcadas (abril–mayo; octubre–noviembre), ya no tienen la regularidad de antes. • Los cambios de temperatura y lluvias son más bruscos e impredecibles. La percepción de los productores entrevistados sobre el cambio climático coincide, en su mayoría, con las tendencias obtenidas de los datos del clima de la zona (tablas 1 y 2). Aunque la superficie de cultivo no ha crecido considerablemente, Boyacá sigue siendo el mayor productor nacional de cebolla de bulbo (tabla 4). A su vez, el distrito de riego Alto Chicamocha concentra la mayor superficie de producción en el departamento, aproximadamente 2.500 ha (Usochicamocha, 2008); adicionalmente, los mayores rendimientos se presentan en esta zona, aproximadamente 40 t ha-1 (obtenido de tabla 3). Si bien el clima tiene un efecto determinante en la producción agrícola, sus variables no han influido significativamente en el comportamiento de la producción, como lo muestra la tabla 5, excepto el de evaporación. Los factores climáticos determinantes del cultivo están siendo controla- 77 E fectos del cambio clim á tico sobre la oferta y precios de cebolla de bulbo Tabla 3. Características de los productores y de las unidades de producción de cebolla de bulbo en la zona de estudio. Experiencia en el cultivo Cantidad de semilla Superficie parcela de cebolla Rendimiento Número de siembras Frecuencia de riego (años) (lb/fanegada) (fanegadas) (bultos/ fanegada)1 al año (No./semana) 5–20 3–4 1–70 500–560 2–6 2 1–3 Tenencia de tierra Otros cultivos que maneja Arriendo, en compañía Papa, lechuga, arveja, fríjol, maíz, remolacha, nabo y avena Fuente: Entrevista a productores de cebolla de bulbo del Distrito de Riego Alto Chicamocha (junio, 2009). 1Bulto = 50 kg; fanegada = 6.400 m²; 2En siembras escalonadas. Tabla 4. Producción anual, superficie de cultivo y rendimiento de cebolla de bulbo en Boyacá y Colombia (1997-2008). Variable 1997 Producción Boyacá (t) 1998 95.894 Producción Colombia (t) 1999 2000 2001 88.391 127.656 142.436 104.793 2002 2003 2004 2005 2006 2007 2008 95.500 110.889 117.272 113.803 133.351 151.727 136.770 230.595 203.410 257.099 276.300 224.945 216.558 232.817 251.752 253.728 294.790 312.673 277.152 Superficie cosechada Boyacá (ha) 3.921 3.686 4.913 5.553 4.005 4.094 4.734 4.290 4.328 4.798 6.149 4.262 Superficie cosechada Colombia (ha) 11.604 10.298 11.766 12.424 10.330 10.250 11.058 11.642 11.838 13.870 15.858 16.516 Rendimiento Boyacá (t ha-1) 24,5 24 26 25,7 26,2 23,3 23,4 27,6 26,3 27,8 24,7 32,1 Rendimiento Colombia (t ha-1) 19,9 19,8 21,9 22,2 21,8 21,1 21,1 21,6 21,4 21,6 19,7 16,8 Fuente: Elaborado con base en datos de Agronet, 2009. Tabla 5. Correlación entre los parámetros climáticos de la estación Surbatá/Bonza del municipio Duitama (Boyacá) y la producción anual del departamento Boyacá (1997–2008). Precipitación No. de días Temperatura máx. anual Temperatura mín. anual Brillo solar Evaporación (h año-1) (mm año-1) promedio anual (%) (mm año-1) (mm) con lluvia/ año (°C) (°C) (°C) R² 0,3725 0,8171 0,0666 0,1574 0,0384 0,052 0,0757 0,0011 0,1198 Relación* directa inversa directa directa inversa directa directa directa inversa Coeficiente Precipitación máx. en 24 h Temperatura media anual Humedad relativa Fuente: Elaborado con base a datos del Ideam, 2009 y Agronet, 2009. * Relación directamente proporcional o relación inversamente proporcional. Vol. 3 - No.1 - 2009 78 Barrientos/Leyva/Morales dos a través del manejo del cultivo: semilla, épocas de siembra, riego, control de plagas, enfermedades y malezas. Según los productores entrevistados, los factores que más han incidido en el incremento o disminución de cultivos de cebolla en la zona han sido los precios y las enfermedades (tizón y alternaria). Según las fuentes primarias, a finales de 2007 los precios llegaron hasta 5.000 $/bulto, así como a finales de 2008 llegaron a 140.000 $/bulto. La caída de los precios se dio por la abundante producción (clima favorable y expectativas de los productores) y por la importación de cebolla. El ataque de las enfermedades proliferó en los últimos dos años. Este incremento se atribuye a dos factores: incremento de fuentes de inóculo (más cultivos con enfermedad) y condiciones ambientales favorables (mayor humedad por efecto del riego y precipitaciones y mayores temperaturas). Lógicamente, el incremento de la frecuencia de aplicaciones y dosis de fungicidas ha incrementado los costos de producción, tanto por producto comprado como por mano de obra utilizada. A diferencia de los datos provenientes del Banco Agrario (2008), el técnico entrevistado sostenía que los costos de producción de cebolla ascendía a 18.000.000 $/ha, de los cuales alrededor del 30% correspondían a controles fitosanitarios. Algo que llamó la atención fue que la mayoría de los productores no han intentado mejorar el manejo del riego, las densidades de siembra y el monitoreo de enfermedades. Sin embargo, los productores más grandes ya lo están haciendo, por ejemplo, a través de aplicación de riegos por demanda del cultivo y no por rutina, disminución de densidades de siembra a 3 libras/fanegada y monitoreos del ataque de enfermedades antes de fumigaciones rutinarias. Medidas adoptadas por los productores para contrarrestar los efectos del cambio climático en la producción de cebolla de bulbo Efectos de la producción en la oferta (producción nacional e importación) y precios de la cebolla de bulbo Frente a los problemas de precios bajos y ataque de enfermedades que se han presentado en los últimos años, los productores de cebolla que han permanecido en el negocio han adoptado, casi todos, las siguientes medidas: Si bien la producción nacional cubre cerca del 80% de la oferta nacional de cebolla de bulbo (año 2008), la importación está cobrando importancia (tabla 6). Mientras la producción nacional tiene una tendencia de crecimiento de 31%, la de importación es de 355%. Por el contrario, las exportaciones de cebolla tienen una tendencia de disminución del 87%. La producción de Boyacá, que aporta aproximadamente el 50% a la producción colombiana, tiene gran influencia en la oferta nacional de cebolla. • Tratar de utilizar semilla certificada. Antes se utilizaban hasta 15 libras por hectárea, ahora aproximadamente 8 libras/hectárea. • Escalonar las épocas de siembra. • Incrementar la frecuencia de fumigado contra enfermedades. Antes cada 10-12 días, ahora cada 3-7 días. Otros productores dicen que ahora se fumiga de 9 a 15 veces por ciclo. • Incrementar las dosis de fungicidas (casi el doble de lo que se utilizaba antes). • Reducir la superficie de cultivo. En algunos casos se redujo hasta un 50%. REV. COLOMB. CIENC. HORTIC. Los precios nominales de la cebolla de bulbo han tenido una tendencia al incremento en una proporción del 64% en los últimos 12 años (tabla 6). Relacionando precios y producción (oferta nacional), se ha encontrado una baja correlación (<0,2) entre estas dos variables, de lo que podría deducirse una baja influencia de solamente la producción sobre los precios y viceversa (Corabastos, 2009; Agronet, 2009). Sin embargo, se advierte una creciente (aproximadamente 5% 79 E fectos del cambio clim á tico sobre la oferta y precios de cebolla de bulbo Tabla 6.Oferta nacional y precios mayoristas (Corabastos, Bogotá) de cebolla de bulbo en Colombia (1997-2008). Variable Producción nacional (t) 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 230.595 203.410 257.099 276.300 224.945 216.558 232.817 251.752 253.728 294.790 312.673 277.152 Importación (t) 11.028 14.154 21.005 38.333 30.936 35.620 34.538 44.436 43.990 51.449 44.713 71.826 Exportación (t) -3.090 -53 -3.657 -20.396 -2.024 -619 -340 -910 -990 -1.517 -1.626 -1.328 Oferta para Colombia (t) Precios mayoristas ($/kg) 238.533 217.511 274.447 294.237 253.857 251.559 267.015 295.278 296.728 344.722 355.760 347.650 511 829 478 854 474 792 812 852 780 861 831 1063 Fuente: Elaborado con base en datos de Agronet, 2009 y Corabastos, 2009. anual) importación de cebolla proveniente de Ecuador (77% el año 2008), Perú (19% el año 2008), Holanda (2% el año 2008) y Chile (<1% el año 2008) (Agronet, 2009). El comportamiento de la importación de cebolla tiene una correlación del 0,5 con el comportamiento de los precios en las centrales mayoristas como Corabastos de Bogotá, lo que hace pensar que la importación está influyendo en la formación de los precios de cebolla mucho más que la producción local y nacional. Así lo manifestaron también los productores entrevistados, quienes aducían a la importación la caída de los precios y la disminución de la superficie de cultivo de la cebolla. CONCLUSIONES • El cambio climático es un fenómeno que ya se ha hecho presente en la zona de estudio. Se caracteriza principalmente por el incremento de la temperatura media, de la evaporación, de la precipitación (menor frecuencia y mayor intensidad) y por una disminución del brillo solar y de la humedad relativa. Además, los rangos entre máximos y mínimos de la mayoría de los parámetros climáticos tienen una tendencia a ampliarse. • El cambio climático, según los entrevistados, ha incidido en los cultivos de cebolla de la zona de estudio, pero no ha ocasionado grandes cambios en la cantidad de producción. • Los efectos más notorios de los cambios en el clima se han manifestado, según los productores, con incrementos del ataque de enfermedades (tizón y alternaria) y mayor necesidad de riego para el cultivo. Siendo éste el efecto más visible, no significa, sin embargo, que problemas como ataque de plagas y presión de malezas al cultivo no estén asociados al cambio climático. • Las medidas adoptadas por los productores frente al fenómeno del cambio climático, de una manera general, fueron el incremento de la frecuencia y dosis de las fumigaciones contra enfermedades y el incremento de la frecuencia de riegos. Estas medidas no son suficientes, y en muchos casos no son las más apropiadas, teniendo en cuenta que hay medidas de manejo integral del cultivo que podrían hacer más eficiente el uso de recursos (entre otros, agua, fungicidas y mano de obra) durante el proceso de producción. • Los productores grandes, a diferencia de los medianos y pequeños, son los que están adoptando otras medidas, como el manejo de las densidades y del riego, así como el monitoreo de enfermedades. • El cambio climático, a través de una mayor o menor producción, no afecta el precio de la cebolla de manera significativa, su efecto se manifiesta más en los costos de producción y en la ganancia de los productores. Las importaciones y las expectativas (de una mejor ganancia) Vol. 3 - No.1 - 2009 80 Barrientos/Leyva/Morales de los productores han resultado ser los factores que más determinan los cambios de precios de la cebolla de bulbo. • Este estudio exploratorio da pie para profundizar en temas específicos como: influencia del cambio climático en el incremento de problemas fitosanitarios del cultivo de cebolla, respuesta estratégica de los productores de cebolla al cambio climático e introducción de nuevas estrategias de transferencia de tecnología. AGRADECIMIENTOS Los autores expresan su agradecimiento al Instituto Colombiano Agropecuario (ICA), seccional Boyacá, al Instituto de Hidrología, Meteorología y Estudios Ambientales (Ideam), estación Surbatá–Bonza, Duitama (Boyacá) y a los productores de cebolla entrevistados por la información brindada. Un especial agradecimiento al Distrito de Riego y Drenaje de Gran Escala Alto Chicamocha y Firavitoba (Usochicamocha), que por medio de los señores Horacio Pachón y Hector Centeno nos facilitó el recorrido por la zona de estudio y el contacto con los productores de cebolla. REFERENCIAS BIBLIOGRÁFICAS Agrios, G. 2005. Plant pathology. 5th ed. Elsevier Academic Press, Burlington, MA. home.php?men=226&con=70&idHm=2&opc=9 9; consulta: mayo de 2009. Agronet. 2009a. Encuesta Nacional Agropecuaria 20012008. En: Agronet, http://www.agronet.gov.co/ agronetweb/Boletines/tabid/75/Default.aspx; consulta: mayo de 2009. Corporación de Abastos de Bogotá S.A. (Coraboastos, Mercado mayorista). 2009. Información anual de precios de mercado mayorista. Calidad extra Cebolla cabezona blanca. Bogotá. Agronet. 2009b. Área, rendimiento y producción. Cebolla cabezona. En: Agronet, http://www.agronet.gov.co; consulta: junio de 2009. Distrito de Riego y Drenaje de Gran Escala Alto Chicamocha y Firavitoba (Usochicamocha) 2008. Presentación de la institución (ppt). Duitama, Boyacá. Agronet. 2009c. Importaciones de Cebolla fresca. En: Agronet, http://www.agronet.gov.co; consulta: junio de 2009. Ezekiel, M. 1938. The cobweb theorem. Quarterly Journal of Economics 52: 255-280. Cambridge. Traducción al castellano: El teorema de la telaraña; Gottfried Haberler (ed.) 1944. Ensayos sobre el ciclo económico. pp. 437-457. Fondo de Cultura Económica, México. Banco Nacional Agrario de Colombia. 2008. Cultivo cebolla cabezona – Boyacá (Allium cepa L.). Costo de producción por cosecha/hectárea. En: http:// www.bancoagrario.gov.co/webapp/PaintServle t?node=003007&treeManagerId=1&treeId=1; consulta: junio de 2009. Brewster, J. 1997. Onions and garlic. pp. 581-619. En: Wien, H. (ed.). The physiology of vegetable crops. CABI, Wallingford, UK. CCI. 2003. Monitoreo de mercados: Mercado nacional de frutas y hortalizas No. 5. junio de 2003. Corporacion Colombia Internacional, Bogotá. CCI. 2009. Encuesta Nacional Agropecuaria 2001-2008. En: CCI, http://www.cci.org.co/cci/cci_x/scripts/ REV. COLOMB. CIENC. HORTIC. Instituto de Hidrología, Meteorología y Estudios Ambientales (Ideam). 2009. Sistema de información nacional ambiental. Bogotá. Kurukulasuriya, P. y S. Rosenthal. 2003. Climate change and agriculture: A review of impacts and adaptations. Paper No. 91 in Climate Change Series, Ariculture and Rural. Development Department and Environment Department, World Bank, Washington, DC. Maroto, J.; B. Pascual y V. Borrego. 1995. Enfermedades de las hortalizas. Ediciones Mundi-Prensa, Madrid. pp. 361-400. Evaluación de dos sistemas de tutorado para el cultivo de la arveja voluble (Pisum sativum L.) en condiciones de la Sabana de Bogotá Evaluation of two training systems for indeterminate pea (Pisum sativum L.) culture in conditions of the Bogota Plateau Andrés Fernando Forero1 Gustavo Adolfo Ligarreto2,3 Tutorado en arco para el cultivo de la arveja. Foto: A.F. Forero RESUMEN Se evaluaron dos sistemas de tutorado; el tutorado vertical y el de arco, que consiste en una propuesta de innovación de los sistemas tradicionales, en el que se recurrió a nuevos materiales y disposiciones que no afecten las densidades siembra que se manejan en la actualidad y permitan el ingreso de maquinaria para el desarrollo de labores durante el ciclo del cultivo. El ensayo comprendió dos partes: la determinación de la estructura adecuada para el tutorado en arco y la evaluación de la viabilidad técnica y económica de este sistema de cultivo de la arveja voluble y su comparación con el tutorado tradicional. Para la estructura del tutorado en arco se utilizó guadua, fibra de polipropileno y mallas y se evaluaron: distancia entre guaduas, enganche entre guaduas y el tamaño del hueco interno de la malla. Los resultados en la primera parte del ensayo permitieron definir que el sistema de tutorado en arco debe tener una distancia entre guaduas de dos metros, un solo enganche entre guaduas y una malla de 15 x 17 cm2 de huecos interiores. En la segunda parte del ensayo, el tutorado en arco presentó un rendimiento menor que el tutorado vertical, costos de instalación más altos y una vida útil más larga. Palabras clave adicionales: sistema de siembra, manejo fitosanitario, rendimiento, calidad de vainas. 1 Programa Agricultura Sostenible, Centro de Investigaciones y Asesorías Agroindustriales (CIAA), Universidad de Bogotá Jorge Tadeo Lozano, Chía (Colombia). 2 Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá (Colombia). 3 Autor para correspondencia. [email protected] REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 3 - No.1 - pp. 81 - 94, 2009 82 Forero/Ligarreto ABSTRACT Two training systems were evaluated. The trained pea system is an innovation to the traditional systems of training. Vertical training and the new training system is built with new materials have no affect on plant density and allow a better access to machinery. The study encompassed two parts: determination of the appropriate structure for arch training, and evaluation of the economic and technical viability of the proposed system in indeterminate pea compared to the traditional training. The arch training was built with bamboo, polypropylene fiber and meshes. The following variables were evaluated: bamboo distance, hook type between bamboo and size of the mesh. The largest yields were obtained with a pole bamboo distance of two meters, a single hook between bamboo and mesh of 15 x 17 cm2. In the second part of the trial vertical training showed a lower yield, higher costs of installation and a longer useful life. Additional key words: planting system, phytosanitory managing, yield, pod quality. Fecha de recepción: 22-01-2009 Aprobado para publicación: 01-06-2009 IntroduCCIÓN La arveja ha sido cultivada como una fuente importante de alimento humano durante muchos siglos y es reconocida por su gran valor nutricional, ya que representa una alta fuente de proteína y carbohidratos, destacándose como fuente de sacarosa y aminoácidos, incluyendo lisina. También es un alimento de contenidos significativos de minerales y vitaminas, por esta razón ocupa un sitio de preferencia en el ámbito alimenticio de los consumidores de las principales ciudades de Colombia. FAO (2009) reporta que la arveja en verde se siembra en una basta área geográfica como cultivo comercial; países como China, India, Estados Unidos, Francia, Inglaterra, Egipto y Marruecos se conocen como los mayores productores de arveja comestible durante 2006 y 2007, mientras Warkentin et al. (1995) reportan al este de Canadá los cultivos de arveja con mayor calidad. En Suramérica, Perú es el mayor productor de arveja verde con 86.415 t anuales para 2006 y 86,500 toneladas para 2007, seguido de Colombia con producción de 33.800 t en 2007. REV. COLOMB. CIENC. HORTIC. La arveja se produce en 11 de los 32 departamentos de Colombia, pero la mayor producción está concentrada en Cundinamarca y Boyacá, con 35% y 33% respectivamente (CCI, 2000). La producción nacional se destina a dos usos diferentes: la verde en vaina para el consumo directo y la seca como semilla, para renovar el ciclo productivo, principalmente por los altos costos que implica su transformación para la obtención del grano seco (SIPSA, 2001). El cultivo de la arveja en Colombia es considerado rentable, pero implica una inversión inicial para su producción, un alto riesgo de pérdidas por las constantes fluctuaciones de los precios en el mercado, susceptibilidad a problemas fitosanitarios y un alto grado exigencia del consumidor, que rechaza cualquier tipo de daño causado por enfermedades o plagas, representando para el productor un gran reto, que se ve inclinado hacia la utilización de agroquímicos como respuesta efectiva para resolver estas situaciones, pero en la mayoría de los casos el uso inadecuado de químicos es una constante, acarreando consigo graves implicaciones para el productor, el consumidor y el medio ambiente. E valuaci ó n de dos sistemas de tutorado para el cultivo de la arveja voluble La arveja es la hortaliza de mayor área cosechada en Colombia con 25.658 ha, es decir, el 26,2% del área hortícola cosechada total en 2004. Sin embargo, su peso en la producción es sustancialmente menor. Esto se reflejó en los bajos rendimientos obtenidos: 1,4 t ha-1 en 2000 (CCI, 2000) y 1,24 t ha-1 en 2000 (MADR, 2006). 1997). Se ha reportado en las regiones templadas de Europa, Norte América, Australia, Nueva Zelanda y en áreas subtropicales de África, Centro y Sur América y Haití (Kraft y Pfleger, 2001), mientras en Colombia, Buriticá (1999) reporta la enfermedad en los departamentos de Cundinamarca, Boyacá y Nariño. Existen diversas variedades de arveja como la Piquinegra, la Parda, la Pajarito, la Guatecana y la Bogotana (CCI, 2000), pero el material Santa Isabel ocupa casi la totalidad del área sembrada, ya que satisface los requerimientos del mercado. Esta homogeneidad de la variedad representa una desventaja desde el punto de vista fitosanitario, haciendo que el manejo de enfermedades producidas por patógenos sea cada vez más costosa y difícil (Fenalce, 2002). Otra de las enfermedades limitantes del cultivo es el mildeo velloso, causado por el hongo Peronospora viciae f. sp. pisi (Sydow) Boherema & Verhoeven. la enfermedad ocurre en todas las partes del mundo donde se cultiva la arveja (Kraft y Pfleger, 2001) y en Colombia, Buriticá (1999) la ha reportado en los departamentos de Antioquia, Boyacá, Cundinamarca y Nariño, en Cundinamarca ha llegado a causar perdidas totales de cultivo (Tamayo, 2000). Otras enfermedades reportadas con gran impacto son la antracnosis, causada por Colletotrichum pisi Pat.; el moho gris, causado por Botryotinia fuckeliana (de Bary) Whetzel y la marchitez, causada por Fusarium oxysporum f. sp. pisi (van Hall) Snyd. & Hans. En la actualidad no se ofrecen materiales de siembra que satisfagan las necesidades del productor y del mercado y con adaptación a Cundinamarca y Boyacá, como departamentos de mayor área de siembra, ya que según Cousin (1997) son muchos los caracteres que tienen que ser mejorados para satisfacer las necesidades de productores y la industria de alimentos. Una de esas necesidades para los productores es la alta susceptibilidad a las enfermedades que afectan a la planta, siendo las manchas foliares las más limitantes, principalmente el tizón, añublo de la arveja causado por el complejo Ascochyta spp. (Alarcón, 1998), que reduce los rendimientos entre un 20% y un 50%, demeritando así la calidad de la vaina y los granos cosechados (Tamayo, 2000). Entre los factores que más afectan la incidencia de las enfermedades al cultivo están las altas densidades de siembra, las condiciones medioambientales de alta humedad relativa, la falta de rotación de cultivos y la poca disponibilidad de variedades mejoradas. Aún no se tiene conocimiento sobre la evaluación de los sistemas de tutorado en la incidencia de las enfermedades en arveja. Las enfermedades causadas por Ascohyta spp. se presentan en todos los lugares del mundo donde se siembra la arveja (Aragona y Porta-Plugia, De acuerdo con anteriores observaciones, es viable el mejoramiento de algunos de los factores internos de la planta, como lo es la resistencia varietal, y externos para aumentar la productividad hasta alcanzar el máximo genético de la planta. El tutorado es uno de los aspectos externos más importantes que se tiene en cuenta al adoptar un sistema de siembra, ya que está relacionado con el rendimiento y la buena calidad del producto final. El hábito voluble de la arveja exige un sistema de tutorado que permita la aireación de las plantas y su apropiado manejo, facilitando las labores como aspersiones, control de arvenses, fertilización y cosecha. Para Almanza y Fischer (1993), el tutorado tiene como finalidad cambiar el hábito de crecimiento de la planta, optimizar la distribución de la luz para aumentar la rata fotosintética y mejorar la calidad de los frutos. Arévalo y Ortega (1995) reportan que con la utilización de tutores en el cultivo de arveja se obVol. 3 - No.1 - 2009 83 84 Forero/Ligarreto tiene un mayor rendimiento por hectárea y una mejor calidad de frutos. El mayor rendimiento se debe a que los tutores permiten aprovechar mejor el espacio aéreo, disponiendo de mayor área de terreno para sembrar más plantas. La mejor calidad de los frutos se debe a la mayor iluminación que recibe el cultivo, favoreciendo un mejor llenado de las vainas; además, la posición vertical de las plantas contribuye a un control más eficiente de plagas, enfermedades y daños por pájaros. En Colombia existen diversos sistemas de siembra de arveja en monocultivo, entre los que Castro (1995) destaca los siguientes: Con tutor vertical En las variedades hortícolas de enrame es muy conveniente preparar un dispositivo capaz de soportar los tallos trepadores, ayudando con ello a que éstos se desarrollen y produzcan una mayor cosecha. Se requieren de 30 a 35 kg de semilla por ha. Se siembra a distancias entre surcos de 1,0 y 1,2 m, 10 cm entre sitios, una semilla por sitio o cada 20 cm dos semillas por sitios, o cada 30 cm tres semillas por sitio. Para tutorar se usan varas de madera de 2,0 a 2,5 m cada cuatro a cinco m, alambre calibre 16 para colgar allí las plantas en amarre individual con hilaza agrícola cuando la planta tiene de 10 a 20 cm de altura. Con tutor horizontal Se requieren 30 a 35 kg de semilla por ha. Se siembra a distancias entre surcos de 1,0 y 1,2 m, 10 cm entre sitios, una semilla por sitio o cada 20 cm dos semillas por sitio, o cada 30 cm tres semillas por sitio. Se utilizan varas de 2,5 m colocadas a una distancia de 4 m, se disminuye la distancia porque no hay utilización de alambre. A medida que va creciendo la planta se van pasando hiladas horizontales cada 15 a 20 cm durante la etapa de crecimiento (siete a diez REV. COLOMB. CIENC. HORTIC. hiladas) con polipropileno encanastándola, alternando una por cada lado. Entre los dos sistemas de tutorado mencionados, el vertical se utiliza en más del 90% de los cultivos por su facilidad en realizar la labor y por la estabilidad y resistencia del tutor, ya que en el encanaste con frecuencia hay volcamiento de plantas causado por el peso del follaje del cultivo. La desventaja de estos tutorados es que impide la mecanización del cultivo. Una propuesta de innovación en el sistema tradicional es el tutorado en arco sin afectar la densidad de siembra y facilitando el ingreso de maquinaria para el desarrollo de labores durante el ciclo del cultivo. El sistema de tutorado en arco es una metodología concertada entre los investigadores del proyecto de leguminosas de la Universidad Nacional de Colombia y profesionales de las entidades Agrointegral Andina S.A. y Syngenta y el agricultor de la Sabana de Bogotá Miguel García, que pretende facilitar el manejo fitosanitario conseguido con el tutor vertical y horizontal al lograr realizar de forma mecanizada las aplicaciones fitosanitarias en zonas donde la mano de obra es escasa, con la opción de tener una optima calidad y sanidad de las vainas, al igual que mayor peso; lograr aumentar la eficiencia fotosintética de la planta; implementar el sistema en la época de invierno; incrementar lar eficiencia en el tiempo de realización de las aplicaciones y tener coberturas homogéneas. MATERIALES Y MÉTODOS El trabajo experimental se llevó a cabo en la finca San Francisco, ubicada en la vereda Laguna Larga, entre las localidades de Madrid y Facatativá (Cundinamarca). El manejo fitosanitario y la fertilización se realizaron de acuerdo con las prácticas corrientes que realiza el agricultor. El material vegetal utilizado fue la variedad Santa Isabel por ser la más cultivada a nivel nacional, y principalmente en la Sabana de Bogotá, ésta E valuaci ó n de dos sistemas de tutorado para el cultivo de la arveja voluble tiene crecimiento indeterminado y alcanza alturas mayores a 1,60 m. El desarrollo experimental comprendió dos partes. La primera parte fue evaluar la estructura adecuada para realizar el tutorado en arco y la segunda fue el análisis de viabilidad técnica y económica del cultivo de la arveja mediante los dos sistemas de tutorado en estudio: arco y vertical. El tutorado en arco se instaló a los 60 dds, para ello se usaron guaduas de 1,70 m de longitud y cuatro cm de ancho. Con estas dimensiones, la guadua, por su flexibilidad, logró formar un arco con 0,4 m de altura y 1,20 m de ancho (figura 1a), la distancia entre surcos fue de 1,80 m. El diseño experimental utilizado en esta primera fase fue el de bloques, completamente aleatorio en arreglo factorial con 27 tratamientos y tres repeticio- nes, en las que el factor principal correspondió a la distancia entre guaduas, el factor secundario a enganches entre guaduas y el tercer factor a la referencia de la malla; a continuación se relacionan los diferentes niveles de cada factor: Distancia entre guaduas: G1: 2 m; G2: 4 m; G3: 6 m. Enganche entre guaduas: E1: un punto; E2: dos puntos; E3: tres puntos. En la figura 1b, se muestra la ubicación de los enganches entre guaduas, se utilizó fibra de 100% polipropileno referencia 18 x 5 monofilamentada, que presentó alta flexibilidad y resistencia. Tipos de malla: M1: malla con 4 x 4 cm2 de cuadros interiores; M2: malla con 7,5 x 7,5 cm2 entre cuadros; M3: malla con 15 x 17 cm2 de cuadros interiores; la malla 3 tenía 80 cm de ancho, las otras dos cubrieron en su totalidad al arco (figura 1c). Figura 1a.Estructura del tutorado en arco, utilizando guadua. Figura 1b. Posición de los enganches en los arcos de guadua. Vol. 3 - No.1 - 2009 85 86 Forero/Ligarreto Di sta ia nc en tr u eg ad ua s Figura 1c.Estructura del sistema de tutorado en arco Cada unidad experimental estuvo conformada por 21,6 m2. Las mediciones se realizaron durante todo el ciclo del cultivo, el cual fue de 23 semanas. El rendimiento se obtuvo al cosechar las vainas de cada unidad experimental. Posteriormente, se procedió a determinar los datos de los componentes de rendimiento, los cuales son: número de vainas por planta que se tomó en cinco plantas por tratamiento, numero de granos por vaina evaluado sobre 10 vainas cosechadas, peso de 20 vainas en verde y el peso de 100 granos. sencia de lesiones visibles; 3=2%, 5=5%, 7=10% y 9=25% o más del área de las vainas afectadas, con base en las escalas utilizadas para ascochyta, reportadas por Van Shoonhoven y Pastor-Corrales (1991). La incidencia de la enfermedad se midió como el número de vainas afectadas en cinco plantas por parcela con al menos una cantidad de tejido afectado del 2%, expresando luego estas unidades o vainas como porcentaje de la población total de las unidades cosechadas. De manera simultánea al experimento de tutorado en arco, se realizó un cultivo de 500 m2 en tutorado vertical. Para el efecto, se utilizaron distancias entre surcos de 1,2 m, varas de 2,5 m de largo cada 4,6 m, alambre calibre 16 y a los 60 dds se colgaron del tutor las plantas en amarre individual con hilaza agrícola, con recolgado a los 105 y 125 dds. La comparación entre los dos sistemas de tutorado no tuvo un diseño estadístico, se realizó extrapolando las áreas de los dos sistemas de tutorado a una hectárea de producción, para el caso del sistema en arco el tratamiento seleccionado fue distancias entre guaduas de 2 m y un enganche que, a su vez, fueron los mejores niveles de cada factor evaluado, con un tamaño de hueco de la malla de 15 x 17 cm2 por ser la más económica. RESULTADOS Y DISCUSIÓN Para cuantificar la enfermedad, se utilizó una escala de severidad basada en la percepción visual. Se establecieron niveles de 1 a 9, donde 1 es au- REV. COLOMB. CIENC. HORTIC. Evaluación del sistema de tutorado en arco El ciclo del cultivo en las condiciones de la Sábana de Bogotá con el tutorado en arco fue de 21 semanas y se lograron realizar dos pases de cosecha. Del análisis estadístico se reporta en la tabla 1 que hay diferencias significativas en el número de granos por vaina y en el peso de las vainas cuando se varió la distancia entre las guaduas, para este mismo factor las diferencias fueron altamente significativas cuando se cuantificó el número de vainas por planta, también se establecieron diferencias significativas para las variables peso de 100 granos y peso de las vainas en estado verde al utilizar diferente número de enganches entre guaduas, resultados concordantes con lo reportado por Lehman y Lambert, citados por Castro (1995), que concluyeron que, al estudiar el efecto de las distancias de siembra sobre el ren- E valuaci ó n de dos sistemas de tutorado para el cultivo de la arveja voluble Tabla 1.Resumen del análisis de varianza para los componentes del rendimiento de arveja. Modalidad de tutorado en arco. Cuadrados medios Fuente de variación GL Peso de granos (100) No. de granos por vaina Peso de vainas verdes (20) No. vainas verdes por planta G 2 0,40 2,65* 34,72* 7,73** E 2 5,20* 0,70 18,39* 1,29 M 2 0,70 0,61 11,62 0,60 G*E 4 3,54* 0,96 1,18 0,96 G*M 4 3,26 1,13 4,32 2,00 E*M 4 0.84 0,55 1,58 3,57 E*M*G 8 1,00 0,15 5,96 2,39 1,83 15,47 1,61 18,47 Coeficiente de variación (%) G, Distancia entre guaduas; E, Enganche entre guaduas; M, Referencia de malla. * Diferencias significativas P≤0,05; ** Diferencia altamente significativa P≤0,01. dimiento y sus componentes en diferentes leguminosas de grano, el componente más afectado es el número de vainas por planta y, en menor grado, el número de granos por vaina y el peso de los granos. La distancia entre guaduas de dos metros generó mayor rendimiento en vainas por planta y afectó favorablemente a las variables número de granos por vaina y peso de las vainas, también asociadas con el rendimiento. El comportamiento se dio porque las plantas estuvieron a mayor altura del suelo, respecto a las otras distancias de cuatro y seis metros entre guaduas, originando mayor aireación entre las plantas, lo cual favorece el desarrollo fisiológico y reproductivo. Al haber mayor fotosíntesis y translocación de asimilados a las estructuras reproductivas, también hay menor afectación por enfermedades, lo cual se refleja de manera directa en el mayor peso de los granos y en el peso de las vainas (tabla 2). En cuanto a la estructura del tutorado, es importante considerar que con la fibra de polipropileno, que tiene una fuerza de tensión de 79 kg (Ciplas, 2005) y se sostiene mediante los arcos de guadua, a medida que es mayor la distancia entre guaduas, la fibra realiza mayor fuerza de tensión, lo que provocó que a las distancias mayores a 2 m no cumpliera su objetivo de forma satisfactoria y se cediera, permitiendo que las vainas rozaran con el suelo. Para la fuente de variación enganche entre guaduas se observaron diferencias significativas en el enganche entre un punto y dos puntos, mientras que entre el enganche a un punto y tres puntos las diferencias fueron altamente significativas. Tabla 2.Resultados de los componentes de rendimiento de la arveja en el tutorado en arco variando la distancia entre guaduas. Distancia entre guaduas (G) No. de vainas por planta Peso de vainas verde (20) No. de granos por vaina 2m 7,04 a 135,34 a 4,84 a 4m 6,18 b 134,81 a 4,68 ab 6m 5,74 c 133,42 b 4,60 b Promedios con letras distintas indican diferencia significativa según la prueba de Duncan (P≤0,05). Vol. 3 - No.1 - 2009 87 88 Forero/Ligarreto La media más alta se presentó en el enganche entre guaduas a un punto (tabla 3), tal vez porque la planta tiende a permanecer más erecta que en dos y tres enganches, lo que favorece la traslocación de asimilados hacia las estructuras reproductivas. La referencia de la malla no presentó diferencias significativas en ninguno de los componentes del rendimiento, por esta razón la más conveniente fue la referencia 15 x 17 cm2 de cuadros interiores, ya que fue la de menor costo. El tutorado en arco, por ser un trabajo experimental inicial, no posee un paquete tecnológico de cultivo. Muchas de las labores realizadas fueron basadas en prácticas que se realizan con el tutorado vertical y otras prácticas se implementaron con el fin de dar solución a limitantes de la producción. Análisis técnico y económico de los sistemas de tutorado vertical y en arco Durante el ciclo del cultivo no se presentaron diferencias en el manejo de plagas con los sistemas de tutorado, en los primeros estados del cultivo se hizo necesario el control químico de trozadores del tallo (Spodoptera sp. y Agrotis ipsilon), mientras para el resto del ciclo no se presentó daño por insectos plaga. El manejo de malezas fue una de las labores dispendiosa de realizar bajo el tutorado en arco, debido a que los 40 cm de altura que posee el arco dificultan el acceso del personal para realizar la labor manual, mientras que en el tutorado tradicional la labor se realiza de manera fácil y con mayor rapidez. Una vez instalado el sistema de tutora- do en arco, se requiere el doble de jornales para la realización de la labor de control de las malezas respecto al sistema de tutorado vertical. Las enfermedades con mayor incidencia y severidad fueron el mildeo velloso y el tizón de la arveja o pecoseo, y presentaron diferencias en cada sistema de tutorado. El mildeo velloso, causado por Peronospora viciae f. sp. pisi (Sydow) Boherema & Verhoeven, presentó mayor incidencia bajo el tutorado vertical, mientras que el tizón o “pecoseo” de la arveja, causado por el complejo Ascochyta pisi, Micosphaerella pinodes y Phoma medicaginis (micosphaerella medicaginis)(Kraft y Pfleger, 2001), presentó la mayor incidencia en el tutorado en arco, comportamiento que muestra una de las diferencias a resaltar entre los dos sistemas (figuras 2 y 3). El mildeo velloso recubre el envés de las hojas de un micelio grisáceo o gris–marrón, en el haz, la infección se manifiesta con manchas de color amarillo–verdoso, también infecciones sistémicas provocan clorosis, deformación de los brotes y enanismo. Ocurre en condiciones frías (15°C), el bajo número de horas luz en días consecutivos es un factor determinante para el desarrollo de la enfermedad, la humedad ambiental y el mantenimiento de una película de agua en la superficie de las hojas favorece la germinación de oosporas y desarrollo de los esporangios con la consiguiente propagación de la enfermedad (Kraft y Pfleger, 2001). Para el tutorado vertical, la enfermedad mildeo velloso alcanzó los mayores valores de incidencia en las vainas en los tres pases de cosecha con valores de 8%, 21% y 30% de vainas afectadas respectivamente en primero, segundo y tercer pase, Tabla 3.Resultados de los componentes de rendimiento en arveja tutorada en arco variando el número de enganches entre guaduas. Enganche entre guaduas (E) Peso de granos (100) Peso de vaina verde (20) Un punto 52,01 a 135,25 a Dos puntos 51,40 b 134,48 ab Tres puntos 51,30 b 133,81b Promedios con letras distintas indican diferencia significativa según la prueba de Duncan (P≤0,05). REV. COLOMB. CIENC. HORTIC. E valuaci ó n de dos sistemas de tutorado para el cultivo de la arveja voluble Figura 2.Incidencia de mildeo velloso sobre vainas cosechadas. Figura 3.Incidencia del tizón o pecoseo de la arveja sobre vainas cosechadas. Vol. 3 - No.1 - 2009 89 90 Forero/Ligarreto factor que tiene alto impacto en la reducción de la productividad del cultivo de arveja (figura 2). Por el contrario, en el sistema en arco la incidencia del mildeo velloso fue baja con promedio de porcentaje del 6% y 10% en primero y segundo pase respectivamente, la variación de estos resultados pudo ser causada porque la densidad de las plantas fue mayor en el tutor vertical y se presenta sombreamiento de las plantas de un surco con las de los surcos vecinos, principalmente en los dos primeros tercios de la planta, también porque hay alta humedad ambiental favorable al establecimiento y daño del patógeno, mientras en el sistema de arco no hay sombreamiento entre plantas de un surco con los surcos vecinos, aun cuando las plantas en su tercio inferior que da sobre la malla también presentan alta humedad y tienen poca aireación. El pecoseo prevalece en condiciones de lluvia y alta humedad relativa, los cultivos sin tutor y con deficiente control de las malezas son más susceptibles. Los síntomas y signos se hacen más evidentes después de floración, en el tercio inferior de la planta. En las hojas se forman lesiones circulares de 2 a 10 mm de color café. En las vainas se forman lesiones deprimidas que dañan la presentación y pueden llegar a causar daño en la semilla (Tamayo, 2000). Contrario al caso anterior, el pecoseo fue limitante en la producción bajo el tutorado en arco, lo cual disminuyó la cantidad de vainas verdes de óptima calidad para su comercialización. La incidencia de la enfermedad sobre las vainas cosechadas en el cultivo con tutor vertical fue inferior al 5%, permitiendo obtener una cosecha de muy buena calidad por este factor, en el sistema en arco se alcanzo el 22% y 42% de vainas con daño de tizón en primer y segundo pase respectivamente, deteriorando drásticamente el producto para la venta (figura 3). La alta incidencia del daño fue causada porque en el costado de las plantas que cae sobre el tutor no recibe aplicaciones de control químico para la enfermedad, REV. COLOMB. CIENC. HORTIC. excepto lo que la planta logre trasportar a esos tejidos, lo cual, unido a factores de alta población de las malezas y alta densidad del follaje de la planta, origina la humedad ambiental favorable al establecimiento y desarrollo del patógeno, afectando drásticamente la producción y calidad del producto. Factores que tuvieron mejor control en el sistema de tutorado vertical. La incidencia de ambas enfermedades aumentó en cada pase de cosecha, debido a que no se realizaron aplicaciones fitosanitarias a partir de 130 dds. El ciclo del cultivo en las condiciones de la Sábana de Bogotá con el tutorado vertical fue de 23 semanas y se logró realizar tres pases de cosecha, uno más que al tutorado en arco, aunque este último sólo representó el 9% del rendimiento total en el tutorado vertical. Las diferencias grandes de producción entre los dos sistemas se presentaron en el segundo pase, el cual representó el 58% de la producción total en el tutorado vertical y el 62% de la misma en el tutorado en arco. El rendimiento del tutorado vertical superó en un 36% al tutorado en arco (figura 4). Por tanto, la propuesta más viable para la Sabana de Bogotá es el sistema de tutorado vertical, ya que con él se obtienen rendimientos de 9,7 t ha-1 con tres pases de cosecha, mientras que bajo el sistema de tutorado en arco los rendimientos son de 6.2 ton ha con dos pases de cosecha (figura 4). El mayor rendimiento del cultivo en el sistema de tutorado vertical se debe a dos aspectos. El primer aspecto a considerar es el programa tecnológico que se ha desarrollado para las condiciones de la Sabana de Bogotá, el cual consiste en la adecuada realización de la preparación del terreno, fertilización, control de malezas y aspersiones para el control de enfermedades e insectos plagas del cultivo, proveniente de varios años de experiencia y ensayos realizados por los agricultores, los investigadores y los profesionales que asisten a los cultivos. El segundo aspecto es la eficiencia fotosintética de la planta en el tutorado vertical, al lograr las plantas recibir mayor iluminación E valuaci ó n de dos sistemas de tutorado para el cultivo de la arveja voluble Figura 4.Rendimiento del tutorado vertical vs. el tutorado en arco en condiciones de la Sabana de Bogotá. en todo su canopy y aprovechar mejor el espacio aéreo, favoreciendo una mejor distribución de las vainas a lo largo del tallo principal y las ramas laterales, un mejor llenado de las vainas y una vainas con menor daño por Ascochyta como la enfermedad que más limita la producción del cultivo. La cosecha fue la labor más limitante del tutorado en arco, porque hay que levantar las plantas para recoger las vainas que se han enredado en la malla, actividad dispendiosa porque las plantas son de hábito indeterminado y se conectan y amarran por zarcillos, lo cual dificulta la labor y, en consecuencia, se empleó más tiempo que la cosecha en el tutorado vertical. En el tutorado vertical, el operario realiza la labor de cosecha de pie y sin agacharse, mientras que en el tutorado en arco la labor se tenía que comenzar a hacer a una altura de 40 cm del piso, lo que no permitió que rindiera por las constantes inclinadas de los recolectores. En el primer pase de cosecha con el tutorado en arco las plantas fueron maltratadas por intentar recoger las vainas que se encontra- ban ocultas, en el segundo pase de cosecha también se presentó esta situación, pero los recolectores manejaron con más cuidado la labor. Los materiales que se emplearon para el tutorado en arco se encuentran en la capacidad de cumplir más de cuatro ciclos de cultivo, puesto que la guadua es de alta resistencia, mientras que el tutorado vertical conformado por varas de eucalipto es reutilizado para máximo tres cosechas, lapso de tiempo en el cual se descompone. Al considerar los costos de instalación del tutorado vertical en 100% y compararlo con el tutorado en arco, que superó en 151% los costos del tutorado vertical, resulta una opción no viable para los productores (tablas 4 y 5). La mano de obra es uno de los ítems en los que se descompensa el tutorado en arco, ya que se emplea mayor número de jornales y ante todo mayor tiempo para la instalación. La cantidad de semilla que se emplea en el tutorado vertical es mayor (tabla 4) que la que se emplea bajo el tutorado en arco (tabla 5) por las distancias de siembra entre los surcos. Vol. 3 - No.1 - 2009 91 92 Forero/Ligarreto Tabla 4. Costos de instalación (en pesos colombianos) del cultivo de arveja bajo el tutorado vertical por hectárea (año 2006). Insumo Unidad Semilla kg Varas Unidad Alambre kg Hilaza Mano de obra Cantidad Valor unitario Valor total 28 6.000 168.000 1450 430 623.500 250 2.800 700.000 Cono 38 8.000 304.000 Jornal 32 30.000 960.000 Total 2.755.500 Tabla 5. Costos de instalación (en pesos colombianos) del cultivo de arveja bajo el tutorado en arco por hectárea (año 2006). Insumo Unidad Semilla kg Guadua Unidad Fibra Rollo Malla m Mano de obra Jornal Cantidad Total Valor unitario Valor total 3 6.000 18.000 2750 1.200 3.300.000 3 43.639 130.917 5560 400 2.224.000 42 30.000 1.260.000 6.932.917 CONCLUSIONES Las mayores limitantes bajo el sistema de tutorado en arco fueron el control de malezas luego de haber instalado el tutorado y la labor de cosecha, lo que aumentó la cantidad de mano de obra para realizar estas actividades con respecto al tutorado tradicional. La propuesta de tutorado en arco puede llevarse a cabo en Colombia, pero para ello se necesitaría innovar más en su estructura, para llegar a los rendimientos que se están obteniendo con el sistema de tutorado vertical en la Sabana de Bo- REV. COLOMB. CIENC. HORTIC. gotá, ya que ella representa una alternativa que puede reducir los costos de manejo en el sistema de producción al permitir el acceso del tractor en el cultivo para aplicaciones fitosanitarias. Las investigación se debe encaminar a mejorar el sistema de tutorado vertical mediante la investigación de materiales de siembra con mayor rendimiento para la Sabana de Bogotá, el perfeccionamiento de las labores de instalación del tutorado y con el correcto uso de los agroinsumos para el manejo de las enfermedades e insectos plagas. E valuaci ó n de dos sistemas de tutorado para el cultivo de la arveja voluble AGRADECIMIENTOS Los autores expresan agradecimientos a los ingenieros Rodrigo Prada y Emilio García por la colaboración y aportes en la ejecución de este trabajo; a Miguel García, por generar la idea del sistema de tutorado en arco con el propósito de dar soluciones prácticas a las problemáticas fitosanitarias del cultivo de arveja y por la colaboración en la elaboración de este trabajo; a Agrointegral Andina S.A. y Syngenta, entidades interesadas en estimular la innovación en los sistemas de producción y por su aporte financiero para la realización de la investigación; al ingeniero José Manuel García y al administrador Néstor Hernández por sus sugerencias técnicas en el sistema de producción de arveja. REFERENCIAS BIBLIOGRÁFICAS Alarcón, P. 1998. Incidencia y severidad del tizón o añublo (Ascochyta spp.) en dos variedades de arveja (Pisum sativum var. Arvense (L.) Poir) con fertilización convencional y biofertilizantes en el municipio de Pasca (Cundinamarca).Tesis de maestría en Ciencias Agrarias. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. Almanza, P. y G. Fischer. 1993. Nuevas tecnologías en el cultivo de la uchuva. Agro-Desarrollo 4(1-2), 292304. Aragona, M. y A. Porta-Plugia. 1997. Improvement of grins y legumes. General part: Diseases. Field Crops Res. 53(1), 17-30. Arévalo, C. y V. Ortega. 1995. Uso de tutores en el cultivo de la arveja. Instituto Nacional de Investigación Agraria, Perú. Buriticá, P. 1999. Directorio de patógenos y enfermedades de las plantas de importancia económica en Colombia. Produmedios, Bogotá. Castro, M. 1995. Evaluación de arveja voluble (Pisum sativum L.) en diferentes densidades y sistemas de siembra en la región de Simijaca (Cundinamarca). Trabajo de grado. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. Ciplas S.A. 2005. Our products. En: http://www.ciplas. com/; consulta: enero de 2009. Corporación Colombia Internacional (CCI). 2000. Manual del exportador de fritas, hortalizas y tubérculos en Colombia. En: http://www.cci.org.co/ Manual%20Exportador/desempeno_prod/princ_ prod_mapas/princ_prod11.htm; consulta: diciembre de 2008. Cousin, R. 1997. Peas (Pisum sativum L.). Field Crops Res. 53, 111-130. FAO. 2009. Faostat. Organización de las Naciones Unidas para la agricultura y la alimentación. En: http:// faostat.fao.org/; consulta: 23 de abril de 2009. Federación Nacional de Cultivadores de Cereales y Leguminosas (Fenalce). 2002. Conocimiento Técnico: Amarillamiento en arveja. En: http://www. fenalceorg.c_Cuotas.htm; consulta: diciembre de 2008. Vol. 3 - No.1 - 2009 93 94 Forero/Ligarreto Kraft, J. y F. Pfleger. 2001. Compendium of pea diseases. APS Press, St. Paul, MN. Ministerio de Agricultura y Desarrollo Rural (MADR). 2006. Acuerdo de competitividad de la cadena de hortalizas. Observatorio agrocadenas Colombia. 19 p. En: http://www.agrocadenas.gov.co/home.htm; consulta: diciembre de 2008. SIPSA. 2001. Relaciones de precios y consumo de las arvejas seca y verde en vaina. Boletín mensual No. 43. Agosto-Septiembre de 2001. En: http//:www. cci.org.co/publicaciones/Sipsa/sipsa43.doc; consulta: diciembre de 2008. REV. COLOMB. CIENC. HORTIC. Tamayo, P. J. 2000. Enfermedades del cultivo de la arveja en Colombia: Guía de reconocimiento y control. Boletín técnico, Corpoica. Produmedios, Bogotá. Van Shoonhoven, A. y M. Pastor-Corrales (eds.). 1991. Sistema estándar para la evaluación de germoplasma de fríjol. Centro Internacional de Agricultura Tropical (CIAT), Cali, Colombia. Warkentin, T.D.; K.Y. Rashid y A. Xue. 1995. Fungicidal control de ascochyta blight of field pea. Can. J. Plant Sci. 75, 67-71. Efecto de la densidad de plantación sobre crecimiento, producción y calidad en cinco accesiones de aji (Capsicum spp.) cultivadas en la Amazonia occidental colombiana Effect of planting density on growth, production and quality of five accessions of hot pepper (Capsicum spp.) in the Colombian Western Amazon Camila Castellanos1 Jaime A. Barrera2 María Soledad Hernández2,6 Luz Marina Melgarejo3 Marcela Carrillo2 Ligia Rodríguez4 Orlando Martínez5 Ají amazónico en fructificación. Foto: J.A. Barrera RESUMEN El género Capsicum, perteneciente a la familia Solanaceae, presenta condiciones favorables para un buen desarrollo en la Amazonia colombiana. De la colección de germoplasma de Capsicum Amazónico del Instituto Sinchi se seleccionaron cinco accesiones por su alto grado de pungencia con el objeto de evaluar su crecimiento y el efecto sobre los patrones de interceptación de la radiación fotosintéticamente activa (RFA), producción y calidad de los frutos bajo tres distancias de siembra. Para este propósito se estableció un experimento en condiciones de campo en la ciudad de Florencia, Caquetá. Durante las nueve semanas del estudio, los índices de crecimiento en general mostraron una producción de biomasa de manera eficiente y sostenida en todas las accesiones. No se presentaron diferencias significativas en producción y en la radiación interceptada bajo diferentes distancias de siembra, pero sí entre las accesiones, siendo la CS 285 la que logra mayor porcentaje de interceptación de RFA, asociado este resultado a la arquitectura que exhiben las accesiones. Para las cinco accesiones la TAN exhibió su máximo valor entre los 27 y 41 días después de transplante. 1 Departamento de Agronomía, Facultad de Agronomia, Universidad Nacional de Colombia, Bogotá (Colombia). Instituto Amazónico de Investigaciones Científicas (Sinchi), Bogotá (Colombia). 3 Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, Bogotá (Colombia). 4 Facultad de Ciencias Naturales, Universidad de Bogotá Jorge Tadeo Lozano, Bogotá (Colombia). 5 Facultad de Ciencias, Universidad de los Andes, Bogotá (Colombia). 6 Autor para correspondencia. [email protected] 2 REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 3 - No. 1 - pp. 95-109, 2009 96 Castellanos/Barrera/Hernández/Melgarejo/Carrillo/Rodríguez/Martínez Palabras clave adicionales: RFA interceptada, índice de área foliar, tasa de asimilación neta, tasa de crecimiento relativo. ABSTRACT The genus Capsicum belonging to Solanaceae family enjoys favorable conditions for successful development in the Colombian Amazon. Five accessions of the Germplasm collection of Amazonian Capsicum of the Institute SINCHI were selected for their high pungency to study the growth, development and the effect on the patterns of interception of photosynthetic active radiation (PAR), production and quality of fruits under three planting distances (50, 60 and 70 cm between plants). For this purpose, an experiment was conducted in field conditions in Florencia, Caqueta. Growth rates, in general, showed an efficient and sustained production of biomass in all accessions in response to the strengthening of their photosynthetic apparatus during the growth. There were no significant differences among the plants in production and intercepted radiation under different distances, but among the accessions it was the CS 285 that achieves the highest percentage PAR interception; this result is associated with the architecture that have the accessions. The five accessions showed highest NAR rate between 27 and 41 days after transplanting. Additional keywords: intercepted PAR, leaf area index, net assimilation rate, relative growth rate. Fecha de recepción: 14-01-2009 Aprobado para publicación: 01-06-2009 IntroduCCIÓN Bolivia, Perú, el Sur de México y Colombia son países considerados como el lugar de origen del género Capsicum –uno de los más importantes de la familia de las Solanaceae–, que comprende cerca de 25 especies silvestres y cinco domesticadas, incluyendo todas la variedades de ají y pimentón. Las cinco especies domesticadas y cultivadas comercialmente son C. annuum, C. frutescens, C. baccatum, C. pubescens y C. chinense (IBPGR, 1983). El Instituto Amazónico de Investigaciones Científicas, Sinchi, tiene a su cuidado un banco de germoplasma de Capsicum ex situ con una colección de 472 materiales de la región amazónica (Méndez et al., 2004) con diferente contenido de pungencia, característica propia de los ajíes. Según Villachica (1996), la Amazonia presenta REV. COLOMB. CIENC. HORTIC. condiciones favorables para la producción continua de ají durante el año, lo que podría servir para abastecer el mercado de los países con invierno prolongado. Los factores que influyen la respuesta fisiológica frente a la densidad de plantas, viendo ésta como un factor cambiante, se pueden comparar mediante la influencia de la radiación solar durante la competencia de las plantas por recursos disponibles (Rodríguez et al., 2004). La respuesta de las plantas frente a la densidad de siembra o por competencia con otras es comparativa frente al efecto de la radiación solar. Para comprender cómo afecta la luz a la tasa fotosintética, hay que considerar que las longitudes de onda comprendidas entre 400 y 700 nm son la porción del espectro que las plantas usan para la fotosínte- E F E C TO D E L A D E N S I DA D D E P L A N TA C I Ó N S O B R E C R E C I M I E N TO, P R O D U C C I Ó N y calidad sis, las cuales se conocen como radiación fotosintéticamente activa (RFA). Esta radiación ha sido establecida como uno de los factores determinantes para calcular los rendimientos potenciales de los cultivos, ya sean densos o sembrados en arreglos especiales (Castillo et al., 1997). La disponibilidad de la radiación solar necesaria para los procesos biológicos que ocurren en un cultivo está determinada en primer término por el macroclima de la región (latitud y día del año), por las condiciones de nubosidad que determinan las cantidades de radiación directa y difusa y por las propiedades del follaje, como el tamaño y la disposición de las hojas, estructura de la planta y distancia de siembra (Jaramillo-Robledo, 2005). Una correcta y apropiada distribución de radiación solar, entre y dentro de los doseles, dará como resultado un trabajo más homogéneo del índice de área foliar (IAF), mejor aprovechamiento de la luz, aumento en la eficiencia fotosintética, menos respiración de mantenimiento y, por tanto, mayores rendimientos agronómicos (Lee et al., 2000). El incremento en la densidad de siembra es uno de los principales manejos que se hacen para mejorar la captura de radiación solar por los cultivos (Idinoba et al., 2002). El rendimiento y la producción de las plantas cultivadas puede medirse mediante el empleo de índices de crecimiento, los cuales indican la eficacia de las plantas para aprovechar los factores ambientales del sitio donde crecen y la forma en que las plantas distribuyen sus asimilados (Méndez et al., 2004). Las diferentes estrategias de manejo durante el ciclo de vida de un cultivo pueden producir efectos en el crecimiento y desarrollo de las plantas; en algunos casos, como el del pimentón (C. annuum L.), hay variaciones que pueden ser inherentes a los diversos genotipos o pueden ser modificadas por factores, como la distancia de siembra entre las plantas (Viloria et al., 1998). En evaluaciones anteriores de material del Instituto Sinchi, Méndez et al. (2003) observaron que el aborto de frutos se encuentra influenciado por la alta densidad de siembra, generando una alta competencia por los nutrientes y luz entre plantas y por fotoasimilados por parte de los frutos. También se ha encontrado que las distancias de siembradesempeñan un papel determinante en la intercepción de radiación solar por el cultivo para convertir esta energía en biomasa, sin dejar de lado otros factores importantes, como el cultivar elegido y sus características de crecimiento, al igual que las características climáticas del lugar donde se va a desarrollar el cultivo (Carrillo et al., 2003). En esta investigación se determinó el efecto que la densidad de siembra ejerce sobre el crecimiento y desarrollo de cinco accesiones de ají amazónico (Capsicum sp.) colección Sinchi cultivadas bajo condiciones de la Amazonia occidental y compara su capacidad individual de interceptación de radiación (RFA). MATERIALES Y MÉTODOS El trabajo se realizó en la granja experimental Santo Domingo de la Universidad de la Amazonia a 7 km de Florencia vía Morelia (Departamento de Caquetá, Colombia), caracterizada con una temperatura promedio anual de 26ºC, humedad relativa entre 70 y 80% y precipitación de 2.500 a 3.300 mm año-1. En el ensayo se empleó un diseño de parcelas divididas distribuidas completamente al azar con estructura factorial de tres densidades de siembra y cinco accesiones en tres repeticiones, para un total de 15 tratamientos, que se describen en la tabla 1. Las accesiones fueron seleccionadas por su alto nivel de pungencia (tabla 2). Las plántulas de 15 cm de altura se sembraron en suelo, en camas de 0,9 m de ancho por 18 m de largo con una polisombra de 30% a 1,5 m de altura del suelo. Para el mantenimiento nutricional en cada parcela se empleó abonamiento orgánico con Bocachi, en aplicación al suelo alrededor de la planta en dosis de 500 g por planta cada dos meses. Vol. 3 - No.1 - 2009 97 Castellanos/Barrera/Hernández/Melgarejo/Carrillo/Rodríguez/Martínez Distancia entre plantas (cm) Accesión (CS) Tratamiento Distancia entre plantas (cm) Accesión (CS) Tratamiento Distancia entre plantas (cm) Accesión (CS) Tabla 1. Tratamientos realizados en el ensayo experimental. Tratamiento 98 T1 50 009 T6 60 009 T11 70 009 T2 50 210 T7 60 210 T12 70 210 T3 50 219 T8 60 219 T13 70 219 T4 50 285 T9 60 285 T14 70 285 T5 50 469 T10 60 469 T15 70 469 CS, Colección Sinchi. Tabla 2.Nivel de pungencia reportado para las accesiones del ensayo experimental. Accesión Especie Pungencia USH CS 210 C. chinense 53.068 CS 469 C. chinense 19.000 CS 219 C. annuum 17.500 CS 285 C. frutescens 16.000 CS 009 C. chinense 12.100 USH, Units Scoville Heats (Unidades de pungencia). Dos semanas después de sembradas las plantas se iniciaron evaluaciones semanales, durante un lapso de nueve semanas, tiempo en el cual se alcanzó el ciclo de cosecha. Se registraron las variables de altura de la planta y área del dosel, diámetro basal del tallo con el uso de un calibrador, peso seco de la planta completa mediante secado en horno de circulación de aire a 75ºC por un periodo de 72 h, área foliar utilizando un planímetro digital marca LI-COR®. La radiación fotosintéticamente activa (RFA) se midió en la segunda floración y fructificación de las plantas con un Ceptómetro AccuPAR® Modelo LP-80. Estos datos se tomaron sobre y bajo el dosel de las plantas, ubicando la barra de tal forma que abarcara dos de las cuatro plantas de cada unidad experimental. Los índices de crecimiento se calcularon utilizando las fórmulas propuesta por Gardner et al. (1985) (tabla 3). En la cosecha, 48 d después de trasplante, se colectaron los frutos para determinar el número de frutos y su peso fresco. Tabla 3. Fórmulas para calcular los índices de crecimiento (Gardner et al., 1990). Índice de crecimiento Ecuación Unidades Índice de área foliar (IAF) sin unidad Tasa de asimilación neta (TAN) g cm -2 día-1 Duración de área foliar (DAF) días Tasa de crecimiento relativo (TCR) g g-1 día-1 Tasa de crecimiento del cultivo (TCC) g m-2 día-1 AF2=Área foliar final; AF1 =Área foliar inicial; AS=Área del dosel; P2=Peso final; P1=Peso inicial; t2=Tiempo final; t1=Tiempo inicial; IAF2=Índice de área foliar final; IAF1=Índice de área foliar inicial. REV. COLOMB. CIENC. HORTIC. E F E C TO D E L A D E N S I DA D D E P L A N TA C I Ó N S O B R E C R E C I M I E N TO, P R O D U C C I Ó N y calidad La calidad de los frutos se midió de acuerdo con el contenido de capsaicinoides y con base en la metodología reportada por Barrera et al. (2008), tomando una muestra de 6,25 g de frutos dispuestos en 50 mL de alcohol al 96%, sometidos a extracción durante 5 h, al cabo de las cuales se filtra el líquido y se pone en un Rotavapor Heidolph® LABOROTA 4000-Efficient a 90 rpm y 75°C hasta obtener 10 mL de extracto. Posteriormente se envasó en frasco ámbar rotulado y se realizó la cuantificación del contenido de capsaicinoides por el sistema CLAE (HPLC) con columna Zorbax Eclipse XDB-C18 a 40ºC. La fase móvil utilizada fue una mezcla de acetonitrilo, ácido acético y agua en la proporción 40:59:1 (v: v) con un flujo de 1,0 mL min-1. Se utilizó como patrón 8-metil-n-vanilil-6-nonenamida (Sigma, St. Louis, MO). Los datos se expresaron en USH (Unidades Scoville de calor), usando el valor de Todd reportado por Sinan et al. (2005). La información obtenida se sometió a análisis de varianza y las diferencias entre los tratamientos se establecieron mediante la prueba de diferencia significativa mínima (LSD) con un alfa de 0,05. Los valores de área foliar y peso seco se procesaron mediante análisis de regresión exponencial. Se utilizó el paquete estadístico Statitistix 8.0. RESULTADOS Y DISCUSIÓN Análisis de crecimiento En la figura 1 se observa el rápido aumento del área foliar entre los días 41 y 69 después de trasplante ddt, coincidiendo con el comienzo de la etapa de fructificación, en la que la planta debe obtener fotoasimilados para empezar el llenado del fruto. Los promedios más altos se dieron para las accesiones CS 219 (C. annuum) y CS 210 (C. chinense). Probablemente, los factores como la RFA movieron asimilación neta de la planta, de esta manera la planta asignó materia seca hacia las hojas entre los 40 ddt y 70 ddt. El peso seco (figura 2) aumenta rápidamente durante estos intervalos de tiempo, debido a la presencia de los nuevos frutos. Los modelos que explican el comportamiento del área foliar y el peso seco de las accesiones se aprecian en la tabla 4. Los resultados (figura 1 y 2) muestran que la fase de crecimiento rápido se presenta después de los 41 ddt para las accesiones CS 009, CS 210 y CS 469 (C. chinense), de los 27 ddt para la accesión CS 285 (C. frutescens) y de los 34 ddt para la accesión CS 219 (C. annuum). Tabla 4.Ecuaciones de regresión para los valores de área foliar y peso seco de las cinco accesiones de ají Capsicum spp. Accesión CS 009 CS210 CS 219 CS 285 CS 469 Variable Ecuación r2 Área foliar y = 0,3438 x 2 – 13,431 x + 191,46 0,9148 Peso seco y = 0,00625 x 2 – 0,2546 x + 3,4455 0,8762 Área foliar y = 0,2536 x 2 – 5,4358 x + 63,446 0,8214 Peso seco y = 0,00565 x 2 – 0,1246 x + 1,6179 0,8484 Área foliar y = 0,4004 x 2 + 12,985 x + 145,18 0,7969 Peso seco y = 27,505 x 2 + 4,1600 x + 1,7404 0,8718 Área foliar y = 0,2016 x 2 – 1,2705 x – 1,732 0,8399 Peso seco y = 0,0032 x 2 – 0,0602 x + 1,7064 0,8240 Área foliar y = 0,293 x 2 – 7,7746 + 82,623 0,9319 Peso seco y = 0,005 x 2 – 0,1409 x + 1,7764 0,9000 Vol. 3 - No.1 - 2009 99 100 Castellanos/Barrera/Hernández/Melgarejo/Carrillo/Rodríguez/Martínez Figura 1.Área foliar en cinco accesiones de ají Capsicum sp. de la colección Sinchi establecidas en Florencia, Caquetá. REV. COLOMB. CIENC. HORTIC. E F E C TO D E L A D E N S I DA D D E P L A N TA C I Ó N S O B R E C R E C I M I E N TO, P R O D U C C I Ó N y calidad Figura 2. Peso seco en cinco accesiones de ají Capsicum spp. de la colección Sinchi establecidas en Florencia, Caquetá. Vol. 3 - No.1 - 2009 101 102 Castellanos/Barrera/Hernández/Melgarejo/Carrillo/Rodríguez/Martínez Índices de crecimiento Los índices de crecimiento de cinco accesiones de ají amazónico presentaron una tendencia ascendente entre los 13 ddt y los 69 ddt (tabla 5) que está acorde con el comportamiento de los índices de crecimiento de un cultivo anual que crece bajo libre exposicion En condiciones de cultivo, de manera particular a partir de los 27 ddt, el comportamiento de la TCR sigue una tendencia acorde con lo observado para muchas especies de crecimiento determinado, teniendo los valores altos que disminuyen con la edad; esta disminución se debe al aumento gradual de tejidos no asimilatorios (Jarma et al., 1999). La tasa de acumulación de materia seca (MS) por unidad de área foliar y tiempo se define como la TAN, que es una medida de la eficiencia media fotosintética de las hojas (Mansab et al., 2003). Para las cinco accesiones, la TAN exhibe su máximo valor entre los 27 ddt y los 41 ddt. Hunt (2003) indica que la TAN exhibe en general un nivel constante relativo, modificado por fluctuaciones en el ambiente durante el periodo de crecimiento. Páez et al. (2000) encontraron en una Solanacea como el tomate var. Río Grande una TAN mayor en los primeros estados de desarrollo de plantas expuestas a radiación total, condición que se hizo contraria en los estados más avanzados de crecimiento. Los resultados anteriores sugieren que, en términos generales, en las etapas vegetativas de las especies anuales la planta produce biomasa de manera eficiente y sostenida en respuesta a la capacidad fotosintética. Esto explicaría la tendencia similar que muestran los índices de crecimiento IAF, TAN y TCR de los materiales de ají en etapas tempranas de crecimiento. Jarma et al. (2006) reportan que esta eficiencia, ante la diversidad de estímulos ambientales y genéticos, se vuelve diferencial en las fases tardías, cuando la planta ha alcanzado su madurez fisiológica. Tabla 5.Índices de crecimiento obtenidos para cinco accesiones de ají amazónico en Florencia, Caquetá. Accesión CS 009 CS 210 CS 219 CS 285 CS 469 REV. COLOMB. CIENC. HORTIC. ddt 13 27 41 55 69 13 27 41 55 62 13 27 41 55 69 13 27 41 55 69 13 27 41 55 69 IAF 0,5041 0,9895 0,6967 0,9520 0,4816 0,3038 0,6682 1,7452 1,9887 6,6095 0,7172 0,9244 1,0669 2,0962 5,2270 0,2886 0,6782 1,0638 0,9377 0,5500 0,3062 0,4469 0,8418 0,9898 0,4554 TAN 0,0019 0,0000 0,0020 0,0006 0,00162 0,00329 0,00051 0,00482 0,0062 -0,0009 0,0019 0,0032 0,0047 -0,0002 0,0004 0,0004 0,0021 0,0003 0,0010 0,0002 DAF 6,8825 5,1844 5,8311 3,6290 4,7254 9,9624 17,0251 43,1604 6,033 7,681 13,473 35,502 3,875 7,435 7,482 6,041 2,903 5,316 7,683 5,114 TCR -0,0004 0,0533 0,1015 0,0380 0,0647 0,1365 0,0212 0,1957 0,0878 -0,0252 0,0948 0,1729 0,0614 -0,0023 0,0277 0,0209 - E F E C TO D E L A D E N S I DA D D E P L A N TA C I Ó N S O B R E C R E C I M I E N TO, P R O D U C C I Ó N y calidad Efecto de la densidad de siembra sobre la RAF, producción y calidad de frutos En las densidades de siembra evaluadas (figura 3) se observa la evolución de la altura durante las nueve semanas de ciclo de cultivo para las cinco accesiones. Contrario a lo reportado, a mayor densidad de siembra, esto es, a distancia de 50 cm entre plantas, se observaron los valores más bajos de altura para las accesiones CS 009, CS 210, CS 219 y CS 295. Experiencias previas indican que la posibilidad de usar pequeñas distancias de siembra en plantas del género Capsicum es un poco limitada, ya que las altas densidades traen como respuesta un incremento en la altura (Decoteau y Hatt Graham, 1994), como lo observado para la accesión CS 469 (C. chinense). Generalmente, los incrementos en la altura de la planta, en respuesta al aumento en la densidad de población, son indicativos de una mayor competencia por luz entre plantas, debido a menor disponibilidad de radiación dentro del dosel (Castillo et al., 1997). Las diferencias significativas se encontraron a la densidad de siembra más baja en las accesiones CS 009 y CS 285 donde se registraron los mayores valores de altura y no hubo diferencias en las densidades medias y altas (50 cm y 60 cm entre plantas). En el caso de la accesión CS 210, se presentan diferencias significativas en la densidad media, siendo ésta en la que se alcanzó la mayor altura. La accesión CS 219 presentó diferencias significativas entre las tres distancias, obteniendo la mayor altura a la menor densidad de siembra. La accesión CS 469 presenta diferencias significativas para la menor densidad de siembra, en la que se alcanzó el menor valor de altura (figura 3). La interacción genotipo y ambiente determinó la respuesta obtenida en cuatro de las cinco accesiones porque las plantas de las especies chinense, frutescens y annuum difieren en cuando a la arquitectura. De acuerdo con Nuez y Costa (1996) y los estudios morfo- lógicos adelantados por Melgarejo et al. (2004), las plantas de ají varían ampliamente en forma y tamaño, encontrando desde herbáceas hasta arbustos entre 25 cm y 190 cm de altura; son plantas semiarbustivas perennes, pero de cultivo anual, y alcanzan entre los 0,3 y 1,5 m de altura, dependiendo principalmente de la variedad, de las condiciones climáticas y de la fertilización. El diámetro basal del tallo fue mayor en las plantas sembradas a menor densidad con excepción de la accesión CS 469 (C. chinense). Díaz et al. (1999) obtuvieron una disminución de diámetro del tallo a mayor altura en las plantas de pimentón. Sin embargo, el comportamiento del diámetro en la accesión CS 469 no muestra diferencias significativas entre ninguna de las distancias de siembra. La accesión CS 219 (C. annuum) presentó diferencias significativas entre todas las distancias, mientras que en las accesiones CS 009 y CS 285 existieron diferencias significativas a la distancia 70 cm. Finalmente, en la accesión CS 210, aunque el mayor valor es alcanzado en la distancia de 70 cm, la diferencia significativa sólo se presenta entre esta distancia y la de 50 cm (figura 4). En general, una mayor exposición a radiación solar favorecerá el crecimiento de la planta, lo cual se observó en casi todas las accesiones. Producción y calidad de frutos Se presentaron diferencias significativas para la variable número de frutos (figura 5) entre la accesión CS 285 (C. frutescens) y las demás accesiones. La accesión CS 219 (C. annuum) sólo presentó diferencias significativas con la accesión CS 285. Entre las accesiones de C. chinense, CS 009 y CS210 no se presentaron diferencias significativas, pero entre éstas y la accesión CS 469 (C. chinense) se las presentaron. Mendez et al. (2003) evaluando accesiones de ají amazónico en condiciones de Leticia, Amazonas, encontraron respuestas diferentes en producción de frutos entre materiales de C. chinense y C. baccatum. Vol. 3 - No.1 - 2009 103 104 Castellanos/Barrera/Hernández/Melgarejo/Carrillo/Rodríguez/Martínez Figura 3. Comportamiento de la altura de la planta en cinco accesiones de ají Capsicum spp. con tres distancias de siembra. Promedios con letras distintas en cada semana indican diferencia significativa según la prueba de LSD (P≤0,05). Las barras sobre las columnas indican desviación estándar. REV. COLOMB. CIENC. HORTIC. E F E C TO D E L A D E N S I DA D D E P L A N TA C I Ó N S O B R E C R E C I M I E N TO, P R O D U C C I Ó N y calidad Figura 4. Comportamiento del diámetro basal del tallo en cinco accesiones de ají Capsicum spp. con tres distancias de siembra. Promedios con letras distintas en cada semana indican diferencia significativa según la prueba de LSD (P≤0,05). Las barras sobre las columnas indican desviación estándar. Vol. 3 - No.1 - 2009 105 106 Castellanos/Barrera/Hernández/Melgarejo/Carrillo/Rodríguez/Martínez Figura 5. Peso y número de frutos cosechados de cinco accesiones de ají Capsicum spp. con tres distancias de siembra. Las accesiones con letras distintas sobre las columnas indican diferencia significativa según la prueba de LSD (P≤0,05). Entre distancias de siembra no se registraron diferencias significativas en las accesiones ni en la interacción accesión por distancia (P>0,05). Según Perez et al. (2003), en condiciones de alta competencia, las plantas responden fenotípicamente, sintetizando mayor proporción de asimilados para contrarrestar el efecto de competencia por RFA. De acuerdo con Arjona et al. (1992) y Azcón-Bieto y Talón (1993), las diferencias en el peso de los frutos puede atribuirse a la composición genética de los diferentes materiales evaluados porque la variedad tiene una gran influencia sobre la velocidad de crecimiento, el tamaño final y la forma del fruto, además del importante efecto ambiental. La accesión con mayor contenido de pungencia es la accesión CS 210 (C. chinense), seguida de la CS 219 (C. annuum) y la CS 285 (C. frutescens). En la tabla 2 se muestra que la pungencia en USH de estas accesiones estaba en su orden de mayor a menor (CS 210-CS 469-CS 219- CS285-CS 009). Los resultados de la figura 6 muestran que indistintamente de la densidad de siembra la accesión CS 210 mantuvo el mayor nivel de pungencia, pero la CS 469 que contenía el segundo mayor nivel se ubicó en el cuarto lugar. Fujikawi et al. (1980) y Calva et al. (2000) anotan que los capsaicionoides son producidos en las glándulas localizadas cerca de la placenta en las paredes del fruto, producto de la ruta del áciREV. COLOMB. CIENC. HORTIC. do cinámico. Ésta empieza a acumularse a partir de los 8 d a 10 d después de la antesis, aumentando a medida que transcurre la maduración de los frutos para llegar a un máximo cuando éstos se tornan completamente maduros (rojos, naranjas o amarillos) y cayendo drásticamente en la senescencia (Iwai et al., 1979). Barrera et al. (2008) mencionan que al inicio de la senescencia hay una importante pérdida de capsaicionides de los frutos producto de la actividad de las peroxidasas. También mencionan que para accesiones pungentes de ají amazónico los niveles óptimos de síntesis de capsaicina y dihidrocapsaicina se dan durante la etapa que corresponde a la máxima coloración típica de cada una de esas accesiones y coincide con el momento de la recolección. Es probable que la CS 469 haya sido colectada en una etapa temprana de desarrollo porque el cambio de color verde a morado y rojo se da en etapas tempranas de desarrollo. Radiación solar interceptada Las accesiones CS 009, CS 219 y CS 285 mostraron un mayor porcentaje de interceptación (figura 7) en la distancia de siembra de 70 cm, seguida de la de 60 cm y finalmente la de 50 cm. Midmore et al. (1988) encontraron que una gran cobertura del follaje con la consecuente interceptación de radiación fotosintéticamente activa fue característica en las altas densidades. No se presentaron diferencias entre las condiciones de E F E C TO D E L A D E N S I DA D D E P L A N TA C I Ó N S O B R E C R E C I M I E N TO, P R O D U C C I Ó N y calidad Figura 6. Contenido de capsaicinoides en unidades USH de cinco accesiones de ají Capsicum spp. con tres distancias de siembra. Las accesiones con letras distintas sobre las columnas indican diferencia significativa según la prueba de LSD (P≤0,05). cielo despejado y nublado, al igual que lo encontrado por Ortega-Farias et al. (2004) en tomate. La radiación interceptada por el dosel mostró diferencias significativas entre distancias de siembra y entre accesiones. La accesión CS 210 presentó los valores significativos más bajos (P>0,05) de porcentaje de radiación absorbida, producto del escaso desarrollo foliar de la planta, y con un tipo de ramificación antocladial con entrenudos largos, muy común en hábito de crecimiento voluble (Nuez y Costa, 1996). Esta disposición reduce la capacidad de interceptar RFA en un nivel superior al 40%, que fue el común para las demás accesiones. El mayor porcentaje de interceptación (tabla 6) lo obtuvo la accesión CS 285 (C. frutescens), el cual fue significativo (P≤0,05) en los dos estados fenológicos evaluados. Sin embargo, esta accesión presentó los valores más altos de número de frutos y el menor peso de frutos. Ésta accesión se caracteriza por agrupar la mayor capacidad de producción en el estrato superior debido al tamaño reducido de sus frutos, lo cual permite inferir que es un material de alta eficiencia en el uso de la radiación solar, concentrando la mayor cantidad de frutos en este estrato, en el que será mejor la relación fuente–demanda. Figura 7. Peso y número de frutos cosechados de cinco accesiones de ají Capsicum spp. con tres distancias de siembra. Las accesiones con letras distintas sobre las columnas indican diferencia significativa según la prueba de LSD (P≤0,05). Las barras sobre las columnas indican desviación estándar. Vol. 3 - No.1 - 2009 107 108 Castellanos/Barrera/Hernández/Melgarejo/Carrillo/Rodríguez/Martínez Tabla 6. Porcentaje de intercepción de interceptación de radiación entre cinco accesiones de ají amazónico en Florencia,Caquetá. Promedios con letras distintas en la misma columna indican diferencia significativa según la prueba de LSD (P≤0,05). Accesión Floración Promedio Fructificación Promedio CS 009 47,270 b 45,976 b CS 210 24,497 c 26,409 c CS 219 44,515 b 41,225 b CS 285 57,522 a 57,352 a CS 469 48,805 b 45,549 b Conclusiones Los ajíes amazónicos en sus etapas vegetativas presentaron un comportamiento típico de las especies anuales en las que la planta produce biomasa de manera eficiente y sostenida en res- puesta a la capacidad fotosintética. Este periodo va hasta los 33 ddt en promedio, momento a partir del cual se da la curva de crecimiento exponencial, aunque en algunas accesiones se dio de manera más tardía. La interacción entre el genotipo y el ambiente determinó la respuesta poco común obtenida en cuatro de las cinco accesiones porque la arquitectura de la planta en las especies chinense, frutescens y annuum es diferente. La producción y calidad de los frutos no se vio afectada por las densidades de siembra, lo cual es una respuesta de aclimatación de las especies del género Capsicum a las condiciones ambientales de disipación de radiación directa. Larcher (2003) menciona que la luz directa pasa a hojas bajeras, por lo que éstas absorben gran cantidad de luz difusa, y hace suponer adaptación a bajos puntos de compensación de luz, por lo que habría una ventaja en condiciones de alta densidad (Cabezas y Corchuelo, 2005). REFERENCIAS BIBLIOGRÁFICAS Arjona, H.; M. Montalvo y M. Soto. 1992. Evaluación del comportamiento agronómico de tres híbridos y dos cultivares de pepino cohombro (Cucumis sativus L) bajo condiciones de invernadero en la sabana de Bogotá. Revista Comalfi 19, 11-14. Carrillo, J.C.; F. Jiménez; J. Ruiz; G. Díaz; P. Sánchez; C. Perales y A. Arellanes. 2003. Evaluación de densidades de siembra en Tomate (Lycopersicon esculentum Mill.) en invernadero. Agron. Mesoam. 14(1), 85-88. Azcón-Bieto, J. y M. Talón. 1993. Fisiología y bioquímica vegetal. McGraw-Hill Interamericana de España, Madrid. Castillo, R.E.; P.J. Arcila.; R.A. Jaramillo y J. Sanabria. 1997. Interceptación de la radiación fotosintéticamente activa y su relación con el área foliar de Coffea arabica. Cenicafe 48(3), 182-194. Barrera, K.; M.S. Hernández; L.M. Melgarejo; O. Martínez y J.P. Fernández-Trujillo. 2008. Physiological behavior and quality traits during fruit growth and ripening of four Amazonic hot pepper accessions. J. Sci. Food Agric. 88, 847-857. Cabezas, M. y G. Corchuelo. 2005. Estimación de la interceptación de la radiación solar en papa criolla (Solanum phureja Juz. et Buk.) en tres localidades colombianas. Agron. Colomb. 23(1), 62-73. Calva, G.; A. Ochoa.; L.E. Rios.; J. Sánchez y V. Martínez. 2000. Catabolismo de capsaicina En: Cultivos de células de Capsicum. Cinvestav. San Pedro Zacatenco, México D.F. REV. COLOMB. CIENC. HORTIC. Decoteau, D.R. y H.A. Hatt Graham. 1994. Plant spatial arrangement affects growth, yield, and pod distribution of cayenne peppers. HortScience 29(3), 149-151. Díaz, L.; A. Vitoria y L. Arteaga. 1999. Crecimiento vegetativo del Pimentón en función de la densidad de plantas y edad del cultivo. Bioagro 11(2), 69-73. Fujikawi, H.; T. Suzuki; S. Osaka y K. Iwai. 1980. Enzymatic formation of capsaicionoid from vanillylamine and iso-type fatty acids by cells free extracts of Capsicum annuum var Karayatsubusa. Agric. Biol. Chem. 44, 2907-2912. E F E C TO D E L A D E N S I DA D D E P L A N TA C I Ó N S O B R E C R E C I M I E N TO, P R O D U C C I Ó N y calidad Gardner, F.P.; R.B. Pearce y R.L. Mitchell. 1985. Physiology of crop plants. Iowa State University, Ames, IA. Hunt, R. 2003. Plant growth analysis: individual plants. pp. 579-588. En: Thomas, B.; D.J. Murphy y D. Murray (eds.). Encyclopedia of Applied Plant Sciences. Academic Press, London. IBPGR. 1983. Genetic resources of Capsicum. A global plan action. International Board of Plant Genetic Resources AGPG/ IBPGR/82/12. Roma. Idinoba, M.E.; P.A. Idinoba y A.S. Gbadegesin. 2002. Radiation interception and its efficiency for dry matter production in the three crop species in the transitional humid zone of Nigeria. Agronomie 22(7), 273-281. Iwai, K.; T. Suzuki y H. Fujikawe. 1979. Formation and accumulation or pungent principle of Hot Pepper fruits. Capsaicin and its analogues, in Capsicum annuum var. annuum cv. Karayatsubusa at different growth stages after flowering. Agric. Biol. Chem. 6, 2493-2498 M.C. Duque y J. Tohme. 2004. Caracterización de accesiones del banco de germoplasma de la región amazónica colombiana. pp. 13-26. En: Melgarejo, L.M.; M.S. Hernández; J.A. Barrera y X. Bardales, (eds.). Caracterización y usos potenciales del banco de germoplasma de ají amazónico. Instituto Amazónico de Investigaciones Científicas, Sinchi, Universidad Nacional de Colombia, Gráficas Ducal Ltda., Bogotá. Méndez, M.A.; G.A. Ligarreto; M.S. Hernández; L.M. Melgarejo y V. González. 2003. Evaluación agronómica de 4 accesiones de ají amazónico del banco de germoplasma ex situ del Instituto Sinchi. Agron. Colomb. 21(3), 165-174. Méndez, M.A.; G.A. Ligarreto; M.S. Hernández y L.M. Melgarejo. 2004. Evaluación del crecimiento y determinación de índices de cosecha en frutos de cuatro materiales de Ají (Capsicum sp.) cultivados en la Amazonía Colombiana. Agron. Colomb. 22(1), 7-17. Nuez, F. y J. Costa. 1996. El cultivo de pimientos, chiles y ajíes. Ediciones Mundi-Prensa, Madrid. Jaramillo-Robledo, A. 2005. La redistribución de la radiación solar y la lluvia dentro de plantaciones de café (Coffea arabica L.). Rev. Acad. Colom. Cienc. 29(112), 371-382. Ortega-Farias, S.; R. Calderón; N. Martelli y R. Antonioletti. 2004. Evaluación de un modelo para estimar la radiación neta sobre un cultivo de tomate industrial. Agric. Técnic. 64, 41-49. Jarma, A.; C. Buitrago y S. Gutiérrez. 1999. Respuesta del crecimiento de la habichuela (Phaseolus vulgaris L. var. Blue Lake) a tres niveles de radiación incidente. Revista Comalfi 26(1-3), 62-73. Páez, A.; V. Paz y J. López. 2000. Crecimiento y respuestas fisiológicas de plantas de tomate cv. ‘Río Grande’. Rev. Fac. Agron. (LUZ) 17(2), 173-184. Jarma, A.; T. Rengifo y H. Aramendia. 2006. Fisiología de estevia (Stevia rebaudiana) en función de la radiación en el Caribe colombiano. II. Análisis de crecimiento. Agron. Colomb. 24(1), 38-47 Larcher, W. 2003. Physiological plant ecology. 4th ed. Springer-Verlag, Berlin. Lee, D.W.; S.F. Oberbauer y P. Johnson. 2000. Effects of irradiance and spectral quality on leaf structure and function in seedlings of two southeast asian Hopea (Dipterocarpaceae) species. Amer. J. Bot. 87(4), 447-455. Mansab A.; D. Jeffers y P. Henderlong. 2003. Interrelationship between leaf area, light interception and growth rate in a soybean-wheat system. Asian J. Plant Sci. 2181, 605-612. Melgarejo, L.M; F. Rodríguez; M. Giraldo; G. Cardona; M. Celis; J.C. Arias; M. García; L. Quintero; M. Cudris; S. Toquita; I. Monroy; M.E. Rodríguez; Perez, L.; G. Corchuelo y E. Ñustez. 2003. Influencia del espaciamiento entre plantas sobre la morfología y el crecimiento de la papa (Solanum tuberosum L. cv. Parda pastusa) bajo dos ambientes contrastantes. Agron. Colomb. 21(3), 210-219. Rodríguez, P.L.; R.G. Corchuelo y C.E. Ñústez. 2004. Influencia del espaciamiento entre plantas sobre la morfología y el crecimiento de la papa (Solanum tuberosum L. cv. Parda pastusa) bajo dos ambientes contrastantes. Agron. Colomb. 21(3), 210-219. Sinan, E.; O. Yemis; C. Kadakal y N. Artik. 2005. Determination of capsaicinoid profile of different chilli peppers grown in Turkey. J. Sci. Food Agr. 85,1435-1438. Villachica, H. 1996. Frutales y hortalizas promisorias de la Amazonia. Tratado de Cooperación Amazónica, Lima. Viloria, A.; L. Arteaga y H.A. Rodríguez. 1998. Efecto de la distancia de siembra en las estructuras de la planta del pimentón. Agron. Tropic. 48(4), 413-423. Vol. 3 - No.1 - 2009 109 Raíces y tubérculos tropicales olvidados o subutilizados en Brasil Tropical root and tuber crops forgotten or underexploited in Brazil Marcos Vinícius Bohrer Monteiro Siqueira1 Elizabeth Ann Veasey1 Tubérculosdedetomate. Plantulas Dioscorea bulbifera L. Foto: Álvarez-Herrera. Foto: D.M. Silva RESUMEN La agricultura tradicional en Brasil básicamente es mantenida para fines de subsistencia en comunidades locales, sin el uso de tecnologías modernas, usufructuando la mano de obra familiar. Una amplia gama de especies son usadas por esos agricultores que mantienen gran diversidad de variedades de cada especie, sea dentro o entre quintales y chacras, presentando también un intenso sistema de intercambio de esos genotipos entre familiares y vecinos. De esa gran diversidad de cultivos, se destacan muchas tuberosas que son parte de la dieta de la mayoría de los brasileros, siendo la yuca, los ñames y la papa dulce (camote) las más cultivadas. A pesar de toda la importancia cultural y alimentaria, esas tuberosas demarcan un “olvido” por parte de grupos de investigación, extensionistas y sociedad en general, visto que son escasas las investigaciones asociadas a ellas, si comparamos con culturas de ciclo corto, en las que predominan la fuerza del agro negocio y el apoyo gubernamental. Cultivos “huérfanos” o “despreciados” son algunas de las connotaciones peyorativas que reciben esas especies de propagación vegetativa. Por la importancia como materia prima en programas de mejoramiento, por la urgencia de evitar la erosión genética, estudios actuales con énfasis en marcadores moleculares trajeron, especialmente en los últimos años, informaciones importantes en relación con la diversidad genética mantenida por agricultores tradicionales y cómo esta diversidad se encuentra estructurada. Por la importancia que posee en Brasil, la caracterización y el análisis genético de la yuca, ñame y papa dulce (camote) harán el cuerpo central de esta revisión. 1 Escola Superior de Agricultura “Luiz de Queiroz”, Departamento de Genética, Universidade de São Paulo, Brasil. [email protected] REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 3 - No.1 - pp. 110-125, 2009 R aíces y tubérculos tropicales olvidados o subutilizados en B rasil Palabras clave adicionales: agricultura tradicional, cultivos despreciados, tubérculos, yuca, ñame, papa dulce. ABSTRACT The traditional agriculture in Brazil basically is maintained for the needs of existence of local communities, without the use of modern technologies and employing the family labour. A wide scale of species is used by the farmers who support high diversity of varieties of each species, within or outside of the small farms, presenting also an intense system of exchanging of these genotypes among family members and neighbours. Of this high diversity of cultures, stand out tuberous root crops that are a part of the diet of the majority of Brazilians, being yucca, yam and sweet potato (camote) the most cultivated. In spite of all cultural and food importance, these tuberous crops demonstrate a “negligence” by the part of investigation groups, extensionists and society in general, taking into account that the investigations associated with these are scarce, if we compare these crops with the ones of short cycle culture, where predominate the strength of the agricultural business and governmental support. “Orphan” or “despised” cultures are some of the poor connotations that receive these species in part of vegetative propagation. Due to the importance of raw material in improvement programs, for the urgency of avoiding the genetic erosion, current studies with emphasis on molecular markers brought, especially in the last years, important information in relation to the genetic diversity supported by traditional farmers and the structure of this diversity. For the importance that these possess in Brazil, the characterization and genetic analysis of the yucca, yam and sweet potato (camote) will occupy the central volume of this review. Additional keywords: traditional agriculture, underused crops, tubers, yucca, yam, sweet potato. Fecha de recepción: 26-04-2009 Aprobado para publicación: 01-06-2009 IntroduCCIÓN La chacra, unidad básica evolutiva de la agricultura tradicional, donde se desarrolla la conservación in situ (on farm) de las etnovariedades, o también denominadas variedades locales, y también donde existe ampliación de la diversidad genética, la cual beneficia a los agricultores locales (Martins, 2001), es un patrimonio agroecológico que se ha perdido a lo largo de los años. La migración de los pueblos a las grandes ciudades, la erosión de los suelos, el aumento del turismo, el uso abusivo del fuego para generar pastizales y el monocultivo, sobre todo de la soya, y la prohibición de la formación de claros en áreas de preservación ambiental ha llevado a muchas chacras al exterminio (Peroni y Hanazaki, 2002; Klink y Machado, 2005; Jackson et al., 2007). Uno de los muchos beneficios que la agricultura tradicional ha proveído a la humanidad es la conservación in situ de germoplasma de muchas especies de importancia económica, manteniendo y amplificando en el tiempo la variabilidad genética (Martins, 1994; Peroni y Martins, 2000; Sambatti et al., 2001). Entre las dos principales estrategias de conservación de germoplasma (in situ y ex situ), la menos exigente económicamente es la primera. Además de ser más barata, la conservación in situ, viabilizada por las chacras de agricultores tradicionales, es de fundamental importancia no sólo Vol. 3 - No.1 - 2009 111 112 Siqueira/Veasey desde el punto de vista genético, sino también para la seguridad alimentaria y preservación del patrimonio cultural de muchas comunidades. Así, la conservación de los recursos genéticos in situ llevada cabo por los agricultores tradicionales tiene un papel fundamental, tomando en cuenta la estrechez de la base genética de las especies cultivadas que se viene manifestando en los últimos años, consecuencia del modelo de agricultura de la Revolución Verde y las limitaciones de la conservación ex situ, extremadamente onerosa y de difícil mantenimiento (Clement et al., 1982; Amorozo, 2000, 2008). La conservación in situ (en la chacra) se refiere a la selección y transformación genética continuas, en vez de preservación estática. Los cultivos así seleccionados se mantienen adaptados a las condiciones del ambiente local, de manejo y a todos los problemas bióticos del cultivo (plagas, enfermedades y hierbas dañinas). La conservación in situ requiere que las unidades de producción agrícola y los productores sean los depositarios tanto de la información genética como del conocimiento cultural de cómo los cultivos son cuidados y manejados. Por tanto, el principio de la conservación in situ propone que cada unidad de producción agrícola tenga su propio programa de mejoramiento y preservación. En realidad, los productores deben ser capaces de seleccionar y preservar sus propias etnovariedades adaptadas localmente, donde eso fuese posible. Como las características de una región establecen criterios importantes de selección, puede haber cierta centralización de programas de selección para una determinada área, definida ecológica y geográficamente, toda vez que el intercambio constante entre material genético de cultivos agrícolas ocurra entre los productores de aquella región. Esas unidades agrícolas son en gran parte áreas pequeñas y medianas donde es practicada de forma plena o con algunas adaptaciones modernas la agricultura tradicional de tipo tumba y quema (Altieri y Montecinos, 1993). La agricultura de tipo tumba y quema está caracterizada generalmente por el uso de un ciclo REV. COLOMB. CIENC. HORTIC. de corte de la vegetación en estado de sucesión secundaria, seguido por la quema de esta vegetación cuando seca, plantío, cultivo, abandono del área después de la cosecha y el retorno al área después de algunos años (Éden, 1988). Segundo Diegues y Arruda (2001), varias culturas tradicionales existen en el Brasil y se debe a ellas que tenemos una gran amplitud de cultivos. Varias comunidades rurales encontradas en el país son resultado de un intenso mestizaje entre pueblos blancos, negros e indios (Arruda, 1999). La introducción de varias especies y variedades de hortalizas, además de diversificar la alimentación de los primeros colonizadores, sirvió de material básico para el mejoramiento genético, en la búsqueda de una mejor adaptación de estas especies a las diferentes condiciones edafoclimáticas encontradas en el Brasil. Otras introducciones se volvieron indispensables a la culinaria regional en algunas regiones brasileras. Otra contribución es indirecta y poco honrosa para los descubridores portugueses, pero ocurrió de hecho y hoy resultó en el enriquecimiento aún mayor del mestizaje del pueblo brasilero, fue el flujo de ñames traídos por los esclavos africanos (Madeira et al., 2008). La yuca y la papa dulce (camote), cultivadas por pueblos indígenas de América del Sur, siempre fueron la base de sus dietas alimenticias y a partir de varios sistemas de cultivo tradicional han sido creadas variedades regionales a partir del conocimiento local, muchas veces centenario y poco comprendido (Brush, 2005). Fue a través de las rutas migratorias de pueblos indígenas y de colonizadores europeos que muchas tuberosas fueron incorporadas y ampliamente cultivadas hasta hoy. Actualmente, la diversidad genética de ciertas especies de hortalizas es mantenida por esos agricultores locales de forma que satisfacen sus necesidades, de acuerdo con el contexto social en el que están incluidos (Bellon, 1996; Veasey et al., 2007). En Brasil, al igual que en otros países en desarrollo, especies como la yuca (Manihot esculenta), los ñames (Dioscorea spp.) y la papa dulce (Ipo- R aíces y tubérculos tropicales olvidados o subutilizados en B rasil mea batatas), tienen una profunda importancia en la agricultura familiar, siendo utilizadas básicamente como fuente de carbohidrato, sobre todo en la región donde residen comunidades de bajo poder adquisitivo. Sus características van más allá de las altas cualidades nutritivas; esas especies también presentan importantes propiedades medicinales, muchas de ellas desconocidas entre nosotros. En Brasil, en algunos estados como Paraíba y Pernambuco, el ñame es un importante recurso alimenticio, difundido por todos los municipios. Ya la papa dulce o camote gana destaque en regiones del sur y sureste, con alguna importancia en el noreste (Silva et al., 2002). La yuca, en cambio, está difundida por todo el país, con una cadena productiva sui generis. Sin embargo, el gran porcentaje del material cultivado en territorio brasilero para ambas especies es de origen tradicional, con una gran diversidad de variedades. Lo que se encuentra en el mercado, en muchos casos, son cultivos poco o nada mejorados, desmereciendo la aceptación del consumidor. Son pocas las instituciones actualmente involucradas en investigaciones relacionadas con el cultivo de la yuca, ñames y papa dulce (camote), siendo urgente el incremento de estudios e investigaciones profundas, para que se comprendan los factores que interfieren en la evolución de esas especies y cómo la diversidad genética de éstas se encuentra distribuida actualmente. La literatura existente sobre esas “hortalizas subutilizadas” es escasa y se encuentra alejada de la realidad, obligando al investigador e interesado por estos cultivos a un intenso ejercicio de análisis de lo que es información correcta y errónea. Partiendo de la realidad, son raros los proyectos de investigación siendo realizados en el país, y con el propósito de rescate y valorización en el que se busque el mejoramiento y conservación de esas especies, el Departamento de Genética de la Escuela Superior de Agricultura “Luiz de Queiróz” en la Universidad de São Paulo (ESALQ/USP), en los últimos años, a partir de su germoplasma de raíces y tubérculos, ha buscado contribuir con nuevas informaciones sobre la caracterización morfológica y molecular de especies de propagación vegetativa. Se postula que la conservación genética de variedades tradicionales es parte de un conjunto de políticas cruciales en el mantenimiento de la biodiversidad global, podremos afirmar que algunas variedades locales desaparecerán más temprano o más tarde, trayendo perjuicio a la población, sobre todo desde el punto de vista alimenticio. La urgencia de más estudios en especies como la yuca, los ñames y la papa dulce o camote se hace evidente por los siguientes factores: agricultores y mejoradores que cultivan tuberosas enfrentan una serie de dificultades generadas por la falta de informaciones técnicas sobre el manejo de estos cultivos, de cara a la escasez de investigación en esta área. Acciones conjuntas entre instituciones académicas, de investigación y extensión son fundamentales para revertir este proceso, de modo que se pueda crear profesionales capacitados, generando y difundiendo tecnologías para el mejoramiento de estos mismos cultivos. La rápida erosión genética de esos materiales, sobre todo por el aumento de los monocultivos y áreas de pastizales, y consecuentemente el abandono de las tierras por pequeños agricultores, lleva a la necesidad de rescate y preservación del conocimiento tradicional y paralelamente a la orientación de programas de conservación de germoplasmas in situ (Jarvis et al., 2000; Altieri, 2002). Eso nos lleva a creer que la yuca, los ñames y la papa dulce, a semejanza de otros cultivos difundidos en chacras tradicionales, merecen una mayor atención del dominio público. Muchos investigadores afirman que apenas las plantas de mayor dinámica fisiológica son las que prevalecerán a los cambios climáticos severos, como es el caso de esas hortalizas, a veces señaladas como “alternativas” o “no convencionales”. Se sabe que la mayoría de los sistemas productivos de esas tuberosas despreciadas se encuentra en pequeñas propiedades y su manejo es técnicamente poco desarrollado. Áreas de mayor explotación también se encuentran desarticuladas de cara a los nuevos recursos, perdiendo Vol. 3 - No.1 - 2009 113 114 Siqueira/Veasey en productividad tanto por la falta de acceso a los tubérculos semillas y tallos de buena calidad, como por el mal uso de los recursos, en los cuales se incluyen el suelo, el agua y la mano de obra no calificada. Se suman a estos factores la falta de apoyo de los órganos de gobierno para el incentivo de la pequeña y mediana explotación y la inexistencia de una cadena de exportación para responder a la exigencia de un mercado internacional competitivo y exigente. En relación con los programas de mejoramiento, un largo camino se ha de recorrer, ya que ellos están reducidos a pocas instituciones. La yuca ha recibido mayor atención por los mejoradores, con varias instituciones involucradas con programas de mejoramiento en Brasil, como la Empresa Brasileira de Pesquisa Agropecuaria (Embrapa), el Instituto Agronómico (IAC) y la Empresa de Pesquisa Agropecuaria y Extensión Rural de Santa Catarina (Epagri), entre otras (Fukuda, 1999). Si los abordajes de pre y poscosecha fuesen diferentes, los (pocos) problemas fitopatológicos en esos cultivos serían reducidos, tornándose más fácil las perspectivas de exportación en el Brasil. La micropropagación, así como otras herramientas que la biotecnología ofrece, puede beneficiar a esos cultivos (Fregene et al., 2002). enorme peso cultural, toda vez que compone la dieta y el folklore de poblaciones tradicionales en diversos países. En este contexto, al hacer la pregunta “?por qué continúan siendo esos cultivos marginalizados en Brasil¿” parece ser fácil la respuesta; difícil sin duda será trazar directrices en el sentido de mejoramiento y conservación de esas especies en el extenso territorio brasilero. Desde el punto de vista agronómico, biológico y experimental, la yuca es una especie muy interesante, pues consiguió, incluso después de un intenso proceso de domesticación, mantener la rusticidad, sin perder la capacidad de reproducirse sexualmente. Si consideramos el proceso de domesticación como continuo, la domesticación de la yuca continúa ocurriendo, como ocurre en las chacras de la agricultura tradicional autóctona de todo el mundo (Cury, 1998). La agricultura tradicional, realizada por poblaciones autóctonas de los países tropicales, es de suma importancia para la evolución de la especie. Yuca La yuca (Manihot esculenta Crantz), Euphorbiaceae, es una de las principales plantas de interés económico del planeta, que ejerce importante papel en la alimentación de millones de personas, principalmente en los países tropicales, siendo también enormemente utilizada en la industria para fabricación de diversos productos. Además de la importancia económica, la yuca posee REV. COLOMB. CIENC. HORTIC. Brasil, según Allem (2002), abriga a la mayoría de las especies (cerca de 55, lo que representa 80% de la diversidad). México viene en seguida, con 17 especies, y 15% de la diversidad. Se estima en 7.000 el número de variedades encontradas en todo el mundo (Mühlen et al., 2000). Las dos especies de Manihot de mayor importancia económica son la M. esculenta, utilizada para alimentación, con raíces tuberosas que poseen alta capacidad de acumulación de almidón, y M. glaziovii, utilizada en Africa para la producción de latex (Cury, 1993). En relación con M. esculenta, existen dos tipos de yuca, clasificadas de acuerdo con la concentración de glucósidos cianogénicos, las yucas bravas, denominadas por los agricultores apenas como yuca, o yuca brava, y las yucas dulces, conocidas por macaxeira (Valle et al., 2004). Las yucas bravas son las más apropiadas para la fabricación de harina, y las yucas dulces son utilizadas en incontables recetas culinarias, sin embargo, ambas tienen otras innumerables aplicaciones (Cereda, 2003). Cury (1993) estableció principios básicos para el establecimiento del modelo de la dinámica evolutiva de la yuca (Martins, 2001). El modelo utiliza la yuca como unidad evolutiva y lleva R aíces y tubérculos tropicales olvidados o subutilizados en B rasil en consideración el hecho de que la especie se propaga vegetativamente, sin dejar de poseer un sistema reproductivo sexual activo, capaz de generar nuevos recombinantes, que aparecen en las chacras a través de la germinación espontánea de semillas. De modo general, las chacras de yuca presentan elevada diversidad intra e inter variedades, sea en las chacras, en una misma comunidad, o entre comunidades (Peroni y Martins, 2000; Pereira, 2008), y varios trabajos refuerzan la tesis de que la Amazonía es uno de los principales centros de origen y diversidad de la especie en el planeta (Olsen y Schaal, 1999; Léotard, 2003; Olsen, 2004). Las diversas etnovariedades se diferencian en múltiples aspectos de interés ecológico y agronómico, como adaptación a condiciones adversas. Para elucidar cuestiones relativas al origen y filogenia, los marcadores moleculares han sido importantes para la caracterización de la diversidad genética de la yuca, que también puede ser medida por indicadores morfológicos y agronómicos. Sambatti et al. (2000) estudiaron la diversidad fenotípica de etnovariedades provenientes de cuatro chacras autóctonas situadas en el municipio de Ubatuba (litoral del Estado de São Paulo), por medio de patrones isoenzimáticos y morfológicos. Concluyeron que hubo diversidad fenotípica, y que la diversidad isoenzimática se concentra básicamente en las chacras, lo que sugiere un gran flujo genético entre los agricultores estudiados. La diversidad de caracteres morfológicos tuvo una distribución similar a la encontrada para caracteres isoenzimáticos. De acuerdo con los resultados obtenidos, se cree que los caracteres morfológicos son objeto de selección por los agricultores, los cuales los utilizan como forma de cuantificar sus cultivos. Sambatti et al. (2001), evaluando la percepción de los agricultores en relación con la identificación de cultivos, compararon los nombres dados por ellos a los individuos encontrados con sus fenotipos isoenzimáticos y concluyeron que ellos tienen un sistema poco específico y generalizado para el reconocimiento de las variedades o morfotipos y tienden a subestimar la diversidad genética de la yuca en sus chacras. En el Valle de Ribeira, Peroni (1998) concluyó, evaluando caracteres morfológicos e isoenzimáticos, que cada variedad está contenida en genotipos distintos, sin embargo, con gran semejanza morfológica, componiendo una población heterogénea. Cada etnovariedad puede entonces ser considerada una familia de genotipos. En ese estudio, él testificó que los agricultores diferencian correctamente las etnovariedades a través de la morfología. Entre tanto, en relación con los resultados obtenidos por Sambatti et al. (2001), los agricultores también subestiman la variabilidad en sus chacras, ya que no diferencian genotipos diferentes que presentan gran semejanza morfológica de las etnovariedades, dando el mismo nombre a genotipos diferentes. Faraldo et al. (2000) evaluaron 141 etnovariedades de varios locales de Brasil a través de la técnica de electroforesis de isoenzimas, estudiando 11 sistemas enzimáticos. Los resultados revelaron índices elevados de heterozigosidad, observándose una mayor concentración de variabilidad en las chacras de la misma región y mayor variabilidad en las regiones que entre regiones. La diversidad intra específica está directamente relacionada con la estructura sociocultural del local. El grupo colectado en las chacras amazónicas presenta mayor diversidad genética. Colombo et al. (2000) estudiaron la estructuración genética de etnovariedades de yuca provenientes de diversos locales de las Américas, utilizando marcadores RAPD (Random Amplified Polymorphic DNA). Encontraron baja estructuración genética entre los materiales evaluados, lo que fue explicado por los autores a través de la práctica común de intercambio de tallo. Analizando los genotipos en función de los datos climáticos de las localidades de origen, encontraron una estructuración en función de la temperatura y de la precipitación de esos locales. Vol. 3 - No.1 - 2009 115 116 Siqueira/Veasey Pereira (2008), evaluando la diversidad y estructura genética en chacras de Amazonas a partir de marcadores microsatélites, concluyó que las chacras son extremadamente complejas desde el punto de vista genético, presentando altos valores de riqueza alélica, polimorfismo y heterozigosidad, sublineando que la diversidad está estructurada básicamente en cada chacra y entre las variedades de las diferentes chacras, consecuencia del intercambio de etnovariedades entre agricultores. Siqueira et al. (2009a), evaluando etnovariedades de diferentes regiones de Brasil, también con marcadores microsatélites, obtuvieron conclusiones semejantes, realzando que la migración de poblaciones puede responder a la mayor o menor proximidad genética entre los diferentes agrupamientos obtenidos. Estudios como esos tienen como objetivo contribuir con una mayor comprensión sobre cómo la diversidad de la yuca se encuentra distribuida en diferentes regiones geográficas. Sus resultados se enfocan en promover la conservación in situ/on farm de un importante recurso genético a través de los agricultores tradicionales. Ñame Dioscorea es el mayor género de la familia Dioscoreaceae, con cerca de 600 especies (Pedralli et al., 2002). Aunque sea elevado el número de especies de Dioscorea, seis de esas especies son consideradas más importantes en la alimentación humana: D. cayenensis Lam., D. alata L., D. bulbifera L., D. esculenta (Lour.) Burkill, D. rotundata Poir. y D. trifida L. De estas, apenas D. trifida es originaria del continente americano, teniendo como probable centro de origen a América Central, aunque Brasil también es considerado centro de origen de esta especie. Relatos revelan que, cuando colonizadores llegaron a América, los indios se alimentaban de la raíz “cará”, nombre que tiene origen en la lengua tupi–guaraní (Abramo, 1990). Durante la expansión marítima y colonización de nuevos territorios en el siglo REV. COLOMB. CIENC. HORTIC. XVI, portugueses y españoles tuvieron un papel importante en la dispersión de las otras especies cultivadas de Dioscorea, originarias de Africa tropical (complejo D. cayenensis/D. rotundata y D. bulbifera), India Central (D. alata y D. esculenta) y región indo–malaya (D. bulbifera) (Montaldo, 1991; Monteiro y Peressin, 2002). El ñame, planta poliploide y de propagación vegetativa, constituye la alimentación básica para más de 100 millones de personas en todo el mundo, sobre todo en los trópicos húmedos y sub–húmedos (Mignouna et al., 2003b). A pesar de ser producido por todo el continente africano, Nigeria es conocida como el mayor productor de ñame del mundo, siendo este país responsable por 76% de la producción mundial, lo que representa aproximadamente 18,3 millones de toneladas. Datos totales apuntan para una producción africana de 28,8 millones de toneladas (Ike y Inoni, 2006). La producción brasilera, en 1999, fue estimada en 225.000 t en una área cultivada de 24.500 ha (Monteiro y Peressin, 2002). La mayor producción de ñame en Brasil ocurre en el Nordeste, especialmente en los estados de Paraíba, Pernambuco, Alagoas, Bahía, Piauí, seguidos de otros de menor importancia. Los agricultores cultivan básicamente la especie D. cayenensis, conocida vulgarmente como “roxo-da-costa”, aunque sean utilizados también algunos clones de D. alata, como el “cará São Tomé” y el ñame “corneta”. Vale resaltar que ocurre en menor escala el cultivo de ñame “mimoso” o “inhambu” (D. trifida) y de ñame “hígado” (D. bulbifera) (Carvalho y Carvalho, 1999). Las especies de Dioscorea son también cultivadas por sus propiedades medicinales (sapogeninas esteroidales, utilizadas en la producción de cortisona y hormonas sintéticas), en el siguiente orden de importancia: D. bulbifera, D. cayenensis, D. dumetorum (Kunth) Pax, D. alata, D. trifida, D laxiflora Griseb. y D. microbotrya Griseb. (Pedralli et al., 2002). Entre muchas de las propiedades R aíces y tubérculos tropicales olvidados o subutilizados en B rasil medicinales del ñame, se destaca la prevención de enfermedades como la malaria, fiebre amarilla y dengue. La medicina tradicional utiliza este rizoma para erupciones de la piel, creyéndose en la limpieza de impurezas. Los tubérculos de Dioscorea son ricos en carbohidratos y pobres en lípidos. Presentan, además de eso, importantes propiedades nutricionales, siendo ricos en proteínas, vitaminas C, B1 (tiamina) y B2, B5 (niacina), riboflavinas, provitamina A, β-caroteno, provitamina D, almidón, ácido ascórbico, Ca, Fe y P, fitoesteroides, entre otras (Seagri, 2002). Su almidón es parecido al del maíz en sabor, textura y color. La harina puede ser adicionada a la de trigo para la fabricación de panes o puede ser utilizada en diversos platos, dulces o salados (Abramo, 1990). Los ñames tienen gran importancia en la agricultura tradicional en Brasil (Peroni y Hanazaki, 2002; Bressan, 2005), así como en muchas pequeñas comunidades rurales de otros países (CBDC, 2001), distribuyéndose por los trópicos húmedos y semihúmedos (Lebot et al., 2005). La especie con mayor representación en la literatura científica y ampliamente consumida en Brasil es la D. alata, especie que fue domesticada en varios países, con gran aceptación tanto en el mercado local como en grandes superficies comerciales. Algunos trabajos con la finalidad de preservación en D. alata han sido realizados in vitro con éxito, porque su conservación en bancos de germoplasma, además de volverse dispendiosa, tiene los inconvenientes de los riesgos de ataques patogénicos y de erosión genética (Rodríguez et al., 2003). A pesar de toda la importancia alimenticia y medicinal, aún es un cultivo despreciado y mal comprendido. Martin y Rhodes (1977) relataron que la clasificación intraespecífica de D. alata es problemática y las relaciones genéticas entre cultivos son difíciles de explicar. Años después Lebot et al. (1998) usaron marcadores isoenzimáticos para estudios de la diversidad genética en 269 cultivares de D. alata originarios del sur del Pacífico, Asia, África, Caribe y América del Sur, concluyendo que muchos cultivos exhibían variaciones diversas, muy probablemente debido al proceso de selección humana. Según los autores, la existencia de una variación genética significativa se debe también a las recombinaciones genéticas verificadas en los patrones isoenzimáticos. En Brasil, los marcadores isoenzimáticos fueron usados para el estudio de la diversidad genética entre etnovariedades de ñames del Valle de Ribeira (Bressan, 2005), constatándose alta variabilidad genética mantenida por los agricultores de esta región, siendo que esa variabilidad no se encuentra estructurada en el espacio. Ya los marcadores RAPD fueron utilizados para evaluar la variabilidad intraespecífica en accesos originarios de Jamaica (Asemota et al., 1996) y en otras especies de Dioscorea de Africa, Asia y Polinesia (Ramser et al., 1996), caracterizando accesos importantes de Dioscorea como entre las especies D. cayenensis/D. rotundata (Dansi et al., 2000). A partir de estos marcadores, Munõz (2003) consiguió determinar qué genotipos de D. alata, de la colección de germoplasma del IIAP (Instituto de Investigación Agropecuaria de Panamá), se separaban claramente de los genotipos de la colección de germoplasma del Catie (Centro Agronómico Tropical de Investigación y Enseñanza). El marcador AFLP (Amplified Fragment Length Polymorphism) fue también utilizado para evaluar la diversidad genética en D. alata y su relación con otras nueve especies comestibles de Dioscorea (Malapa et al., 2005) y en estudios sobre la domesticación del género Dioscorea (Scarcelli et al., 2006). Egesi et al. (2006), utilizando marcadores AFLP, consiguieron demostrar que, a partir de 53 accesos de D. alata, en África occidental y central, cada grupo formado era una mezcla de los accesos de origen geográfico diferente, indicando que la geografía no tuvo un papel central en la diferenciación de la especie. Vol. 3 - No.1 - 2009 117 118 Siqueira/Veasey Otro marcador que ha sido utilizado para especies de Dioscorea son los microsatélites (Hochu et al., 2006; Bousalem et al., 2006). Fundamentalmente, los microsatélites han sido utilizados para estudios de los patrones de segregación de esos marcadores y caracterización de accesos de varias especies del género Dioscorea (Terauchi y Konuma, 1994; Mignouna et al., 2003a; Mignouna et al., 2003b; Scarcelli et al., 2005). Con el objetivo de caracterizar especies cultivadas del género Dioscorea de frecuentes en áreas de agricultura tradicional de Brasil, enfatizando traer subsidios para la conservación on farm, se pueden citar nuevos estudios con D. trifida (Borges et al., 2007, 2008), D. cayenensis (Recchia et al., 2007) y D. bulbifera (Kreyci et al., 2008). Siqueira et al. (2009b) evaluaron la diversidad genética de etnovariedades de ñame colectadas en chacras de agricultura tradicional y de variedades comerciales obtenidas en ferias y mercados por medio de marcadores microsatélites, verificando alto polimorfismo entre los locus y gran variabilidad genética para el conjunto de accesos de D. alata. Con todo, en la investigación no observaron separación entre las etnovariedades y las variedades comerciales. Papa Dulce (camote) La papa dulce o camote (Ipomoea batatas (L.) Lam.) es una planta de clima tropical o subtropical, también cultivada en regiones templadas. Es de fácil cultivo, rústica, de amplia adaptación, alta tolerancia a la sequía y bajo costo de producción. Los riesgos ofrecidos por el cultivo son mínimos si se compara a otras hortalizas. Es cultivada en toda la faja tropical del globo terrestre, donde tiene usos múltiples, tanto en la alimentación humana como animal, o como materia prima para la industria. Es la séptima más importante fuente de alimento en todo el mundo, junto con el trigo, el arroz, el maíz, la papa, la cebada y la yuca (CIP, 2009). REV. COLOMB. CIENC. HORTIC. Actualmente, la papa dulce puede ser considerada como un cultivo asiático, ya que gran parte de la producción mundial es producida y consumida en Asia. Los países en desarrollo producen y consumen más de 95% del total mundial. Los mayores productores son China, Indonesia, India, Japón y Filipinas, siendo China el productor del 90% del total mundial (CIP, 2009). En Brasil, ella es cultivada en todo el territorio. Los mayores productores son los estados de Rio Grande do Sul, Santa Catarina, Bahía, Paraná y Rio Grande do Norte. Brasil produjo cerca de 500 mil toneladas en 2005 (IBGE, 2007) y las estimaciones apuntaban que ese número no sería muy diferente en los años siguientes. El valor nutritivo del camote es alto, comparable al de la papa. Las raíces son buena fuente de energía (azúcares y otros carbohidratos), calcio, hierro y otros minerales y vitaminas, principalmente vitamina C. Algunos cultivos son ricos en vitamina A, particularmente las de pulpa anaranjada (Steinbauer y Kushman, 1971; Edmond y Ammerman, 1971; Kay, 1973; Folquer, 1978). La composición química varía con el cultivo, condiciones climáticas, época y periodo de cosecha, duración y condiciones de almacenamiento y tratos culturales (Edmond y Ammerman, 1971; Miranda et al., 1984; Cereda et al., 1984; Fischer y Karnatz, 1992). La papa dulce y las especies relacionadas con ella son poco estudiadas, comparativamente a otros cultivos, del punto de vista de la citogenética, taxonomía, morfología, fisiología, distribución geográfica, sistemas de incompatibilidad, cruzamientos compatibles y otros. Naturalmente, estos tópicos son diferentes en especies silvestres y tipos cultivados, e importantes sobre varios aspectos, inclusive para elucidar y comprender el origen genético y la evolución de la especie. La existencia de innumerables especies silvestres del género Ipomoea y la gran difusión y diversidad de tipos existentes en toda América tropical, supe- R aíces y tubérculos tropicales olvidados o subutilizados en B rasil rior a la encontrada en Oceanía (Yen, 1976) hace suponer que la domesticación de la papa dulce o camote sucedió en el continente americano (Folquer, 1978; Austin, 1977; FAO, 1981). Uno de los trabajos más importantes sobre la evolución de I. batatas fue dirigido por Nishiyama (1982). Este autor tuvo sus trabajos de investigación basados en la compatibilidad sexual, síntesis de poliploides, análisis citogenéticos, comparaciones de genomas y características morfológicas y tuvo como principal premisa que la papa dulce se originó de I. trifida hexaploide por diferenciación génica, especialmente para la formación de raíces tuberosas. Las informaciones generadas por Nishiyama (1982) y sus colegas trajeron, en la época, un gran debate y mucha controversia. Entre tanto, sin entrar en mérito de la cuestión del origen genético, todos los investigadores son unánimes en reconocer el gran valor de las investigaciones conducidas por los científicos japoneses, cuyo trabajo es extremadamente importante, no sólo para elucidar y comprender el origen genético y la evolución de la papa dulce, sino sobre todo por la posibilidad que se abre con la utilización de los híbridos hexaploides (2n = 90), sintetizados a partir de otras especies del género Ipomoea, en programas de mejoramiento genético. Lo que está en discusión aún hoy es el local exacto del centro de origen y domesticación: si es en el noroeste de América del Sur (Perú, Colombia y Ecuador) o el sur de México o América Central (principalmente Guatemala). Austin (1988), basado en análisis numéricos de características morfológicas en papa dulce o camote en especies silvestres de Ipomoea, había postulado que la papa dulce se habría originado en la región entre la Península de Yucatán, en México, y el Río Orinoco, en Venezuela. Estudios más recientes, basados en marcadores moleculares, indican a América Central como el centro primario de diversidad y el más probable centro de origen de la especie (Zhang et al., 2000; Gichuki et al., 2003). Perú y Ecuador deberían ser considerados como el centro secundario de la diversidad de la papa dulce (Zhang et al., 2000). Brasil debería también ser considerado como un centro secundario de diversidad (Austin, 1988), una vez que varias especies de Ipomoea, de la sección papas, se encuentran en este país. Concordando con esta hipótesis, altos niveles de diversidad morfológica y molecular fueron observados entre etnovariedades del Valle de Ribeira, Estado de São Paulo, Brasil (Veasey et al., 2007, 2008). Usando ocho marcadores microsatélites, Veasey et al. (2008) verificaron gran polimorfismo, siendo que la mayor variabilidad genética se encontraba en las chacras. Los autores concluyeron que el gran intercambio de material entre agricultores, así como otros factores antrópicos, fueron determinantes para esos resultados. Oliveira et al. (2002), Daros et al. (2002) y Fabri (2009) también observaron elevada diversidad morfológica, isoenzimática y molecular entre variedades de papa dulce originarias de diferentes regiones de Brasil. Entre 1940 y finales de los años sesenta, algunas instituciones de investigación mantienen colecciones de germoplasma e iniciaron programas de mejoramiento de papa dulce en Brasil. Entre ellas, se destacan el Instituto Agronómico (IAC), el antiguo Instituto de Ecología y Experimentación Agrícola, después Ipeacs–Itaguaí/ RJ, la Escola de Agronomia de Cruz das Almas (en el Estado de Bahía), y la Escola de Agronomia de Viçosa, hoy UFV (Estado de Minas Gerais), entre otras. En abril de 1980, se inició en Embrapa–Centro Nacional de Pesquisas en Hortalizas (CNPH), en Brasilia, D.F. un programa de mejoramiento genético de papa dulce (Miranda, 1983). Hoy, algunas instituciones de investigación en Brasil mantienen colecciones de germoplasma de papa dulce, existiendo un banco activo de germoplasma (BAG), mantenido por la Embrapa–CNPH, en Brasilia, otro en Embrapa–CNPMF, en Cruz das Almas, uno en el IAC, en Campinas, y uno en ESALQ/USP. Vol. 3 - No.1 - 2009 119 120 Siqueira/Veasey Notas finales Grandes áreas de preservación del territorio brasilero son ocupadas por poblaciones indígenas o de comunidades rurales “tradicionales” –caiçaras, ribeirinhos, seringueiros, quilombolas, caipiras– para las cuales la conservación de fauna y flora es una garantía de su perennidad. El proceso de expansión de las fronteras agrícolas y extractivistas de la sociedad brasilera, a su vez, reproduce y se pauta por un modelo de ocupación del espacio y de uso de los recursos naturales, generador de degradación ambiental y de enormes costos sociales, en particular para las sociedades rurales (Arruda, 1999). El mantenimiento de la diversidad genética de los agroecosistemas tradicionales contribuye para la adaptabilidad de las poblaciones humanas, ya que provee garantías de sobrevivencia para el agricultor. Al mismo tiempo, a pesar del importante papel de las comunidades rurales para la conservación de la biodiversidad, innumerables factores se han constituido en riesgos para ésta, tales como presiones del mercado, éxodo rural y la globalización, que mina a las culturas locales (incluso los hábitos alimenticios, basados en el consumo de una pequeña variedad de alimentos) en pro de una supremacía cultural reduccionista, importada de los países desarrollados. Nos parece claro que, a partir de los datos presentados, además de generadora de diversidad, la agricultura tradicional es también mantenedora de esas etnovariedades de yuca, ñame y papa dulce o camote. En el momento en que la acción humana deja de ejercer influencia directa, esas variedades locales permanecen en la naturaleza por poco tiempo o desaparecen en la competición con otras plantas silvestres. Este es un aspecto que diferencia a las plantas domesticadas– cultivadas de las plantas silvestres o salvajes y demuestra la estrecha relación entre las plantas cultivadas y el ser humano. Para rescatar las etnovariedades de yuca, ñame y papa dulce y ampliar su uso en la agricultura, se torna necesario pensar en estrategias construidas a partir de un diagnóstico de las causas que vienen provocando el abandono de esas variedades locales. Tal diagnóstico puede ser iniciado a través de una lectura del espacio rural, y en especial de la situación socio–económica y ambiental en la cual la agricultura familiar está inserta. La compleja y alta diversidad genética de las tuberosas “despreciadas” revisadas aquí, y continuamente estudiadas en ESALQ/USP, realzan per se la importancia de cómo se encuentra el patrimonio hortícola nacional. No obstante, nuevos estudios se hacen necesarios para que las políticas de conservación in situ y on farm se puedan concretizar. AGRADECIMIENTOS A Jorge Luis Ferrer Uribe, Eleonora Z. Blaco y Paulo Tavares de Melo por la ayuda en la revisión del manuscrito y valiosas sugerencias tanto en los aspectos de redacción como de contenido. REV. COLOMB. CIENC. HORTIC. R aíces y tubérculos tropicales olvidados o subutilizados en B rasil REFERENCIAS BIBLIOGRÁFICAS Allem, A.C. 2002. The origins and taxonomy of cassava. pp. 1-16. En: Hillocks, R.J.; J.M. Tresh y A.C. Bellotti (eds.). Cassava: biology, production and utilization. CABI Publishing, Oxon, UK. Altieri, M. 2002. Agroecology: the science of natural resource management for poor farmers in marginal environments Agr. Ecosyst. Environ. 93, 1-24. Altieri, M.A. y C. Montecinos. 1993. Conserving crop genetic resources in Latin America through farmers’ participation. pp. 45-64. En: Potter, C.S.; J.I. Cohen y D. Janczewski (eds.). Perspectives on Biodiversity: Case Studies of Genetic Resource Conservation and Development. Washington D.C. Abramo, M.A. 1990. Taioba, cará e inhame: o grande potencial inexplorado. Editora Ícone, São Paulo, Brasil. Amorozo, M.C.M. 2000. Management and conservation of Manihot esculenta Crantz germplasm by traditional farmers in Santo Antonio do Leverger, Mato Grosso State, Brazil. Etnoecologica 4, 69-83. Amorozo, M.C.M. 2008. Maintenance and management of agrobiodiversity in small-scale agriculture. Functional Ecosystems Communities 2, 11-20. Arruda, R. 1999. “Populações tradicionais” e a proteção dos recursos naturais em unidades de conservação. Ambiente e Sociedade 2(5), 79-92. Asemota, H.N.; J. Ramser; C. Lopéz-Peralta; K. Weising y G. Kahl. 1996. Genetic variation and cultivar identification of Jamaican yam germplasm by random amplified polymorphic DNA analysis. Euphytica 92, 341-351. Austin, D.F. 1977. Hybrid polyploids in Ipomoea section batatas (Convolvulaceae). J. Heredity 68, 259-260. Austin, D.F. 1988. The taxonomy, evolution and genetic diversity of sweetpotatoes and related wild species. pp. 27-60. En: Gregory, P. (ed.). Exploration, maintenance, and utilization of sweetpotato genetic resources. International Potato Center, Lima. crossatélites em Dioscorea trifida. En: Anais Simpósio Internacional De Iniciação Científica Da USP, Ribeirão Preto, Brasil. Borges, A.; G.H. Recchia; L.C. Ming y E.A. Veasey. 2008. Role of traditional farmers in maintaining the genetic diversity assessed with microsatellites in Dioscorea trifida. En: Proc. Harlan II. An International Symposium. Biodiversity in Agriculture: Domestication, Evolution & Sustainability, Davis, CA. Bousalem, M.; G. Arnau; I. Hochu; R. Arnolin; V. Viader; S. Santoni y J. David. 2006. Microsatellite segregation analysis and cytogenetic evidence for tetrasomic inheritance in the American yam Dioscorea trifida and a new basic chromosome number in the Dioscoreae. Theor. Appl. Genet. 113, 439–451. Bressan, E.A. 2005. Diversidade isoenzimática e morfológica de inhame (Dioscorea spp.) coletados em roças de agricultura tradicional do Vale do Ribeira – SP. Tese (Mestrado em Ecologia de Agroecossistemas) – Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, Brasil. Brush, S.B. 2005. Protecting traditional agricultural knowledge. Washington Univ. J. Law Policy 17, 59-109. Carvalho, P.C.L. y R.L. Carvalho. 1999. Coleção de genótipos silvestres e cultivados de Dioscorea. En: Queiróz, M.A. de; C.O. Goedert y S.R.R. Ramos (eds.). Recursos Genéticos e Melhoramento de Plantas para o Nordeste brasileiro. Versão 1.0. Embrapa Semi-Árido, Petrolina-PE; Embrapa Recursos Genéticos e Biotecnologia, Brasília-DF, nov. 1999. http://www.cpatsa.embrapa.br; consulta: 22 de enero de 2007. CBDC. 2001. Community Biodiversity Development and Conservation Programme - Bohol Project. Crop Genetic Resources Enhancement of Yam (Dioscorea alata) in Four Towns in Bohol, Philippines. Technical Report No. 7. Southeast Asia Regional Institute for Community Education, Quezon City, Philippines. Bellon, M.R. 1996. The dynamics of crop infraspecific diversity: a conceptual framework at the farmer level. Econ. Bot. 50, 26-39. Cereda, M.P. 2003. Tecnologia, usos e potencialidades de tuberosas amiláceas Latino Americanas. Fundação Cargill, Série Culturas de Tuberosas Amiláceas Latino Americanas. Vol 3. São Paulo, Brasil. Borges, A; G.H. Recchia y E.A. Veasey. 2007. Otimização de protocolo para amplificação de marcadores mi- Cereda, M.P.; G. Wosiack y F.A.D. Conceição. 1984. Avaliação físico-química e reológica de vinte e seis cul- Vol. 3 - No.1 - 2009 121 122 Siqueira/Veasey tivares de batata-doce (Ipomoea batatas (L.) Lam.). Hortic. Bras. 2(1), 6-12. CIP - International Potato Center. 2009. Sweet potato production and consumption: major global impact. En: http://www.all-about-sweet-potatoes. com/production-consumption.html; consulta: 17 de junio de 2009. Clement, C.R.; W.E. Kerr y T. Ken-Hiri. 1982. Erosão cultural na Amazônia uma melhor adaptação às condições ecológicas da região. Ciência e Cultura 34, 1150-1153. Colombo, C.; G. Second y A. Charrier. 2000. Diversity within of american cassava germplasm based on RAPD markers. Genet. Mol. Biol. 23, 189-199. Cury, R. 1993. Dinâmica evolutiva e caracterização de germoplasma de mandioca (Manihot esculenta Crantz) na agricultura autóctone do sul do Estado de São Paulo. Piracicaba, 103 p. Dissertação (Mestrado) – Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Brasil. Cury, R. 1998. Distribuição da diversidade genética e correlações de caracteres em etnovariedades de mandioca (Manihot esculenta Crantz) provenientes da agricultura tradicional do Brasil. Piracicaba, 163 p. Tese (Doutrorado) – Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Brasil. Dansi, A.; H.D. Mignouna; J. Zoundjihékpon; A. Sangare; N. Ahoussou y R. Asiedu. 2000. Identification of some Benin Republic’s Guinea yam (Dioscorea cayenensis/rotundata complex) cultivars using Randomly Amplified Polymorphic DNA. Genet. Resources Crop Evolution 47, 619-625. Daros, M.; A.T. Amaral Júnior; T.N.S. Pereira; N.R. Leal; S.P. Freitas y T. Sediyama. 2002. Caracterização morfológica de acessos de batata-doce. Hortic. Bras. 20, 43-47. Diegues, A.C.S. y R.S. Arruda. 2001. Saberes tradicionais e biodiversidade no Brasil. Ministério do Meio Ambiente, Brasília. Eden, M.J. 1988. Crop diversity in tropical swidden cultivation: comparative data from Colombia and Papua New Guinea. Agr. Ecosyst. Environ. 20, 127-136. Edmond, J.B. y G.R. Ammerman. 1971. Sweet Potato: Production, Processing, marketing. AVI Publishing, Westport, CT. REV. COLOMB. CIENC. HORTIC. Egesi, C.N.; R. Asiedu; G. Ude; S. Ogunyemi y J.K. Egunjobi. 2006. AFLP marker diversity in water yam (Dioscorea alata L.) Plant Genet. Resources Characterization Utilization 4(3), 181-187. Fabri, E.G. 2009. Diversidade genética entre acessos de batata-doce (Ipomoea batatas L. Lam.) avaliada através de marcadores microssatélites e descritores morfoagronômicos. Tese (Doutorado) – Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, Brasil. Faraldo, M.I.F.; M.R. Silva; A. Ando y P.S. Martins. 2000. Variabilidade genética de etnovariedades de mandioca em regiões geográficas do Brasil. Scientia Agrícola 57, 499-505. FAO - Food and Agriculture Organization of the United Nations. 1981. Genetic resources of sweet potato. IBPGR report, Roma. Fischer, G. y A. Karnatz. 1992. Effect of soil temperature on growth and carbohydrates of two sweet potato (Ipomoea batatas) varieties from Papua-New Guinea. Plant Res. Dev. 35, 64-72. Folquer, F. 1978. La batata (camote). Estudio de la planta y su producción comercial. Editorial Hemisfério Sur, Buenos Aires. Fregene, M.; J. Tohme; W. Roca; P. Chavarriaga; R. Escobar y H. Ceballos. 2002. Biotecnología para la yuca. pp. 377-405. En: Ospina, B. y H. Ceballos (eds.). La Yuca en el Tercer Milenio. Sistemas Modernos de Producción, Procesamiento, Utilización y Comercialización. CIAT, CLAYUCA, Ministerio de Agricultura y Desarrollo y FENAVI, Cali, Colombia. Fukuda, W.M.G. 1999. Melhoramento da mandioca. pp. 409-428. En: Borém, A. (ed.). Melhorameto de espécies cultivadas. Viçosa, UFV, Brasil. Gichuki, S.T.; M. Berenyi; D. Zhang; M. Hermann; J. Schmidt; J. Glössl y K. Burg. 2003. Genetic diversity in sweetpotato [Ipomoea batatas (L.) Lam.] in relationship to geographic sources as assessed with RAPD markers. Genet. Resources Crop Evolution 50, 429-437. 429-437. Hochu, I.; S. Santoni y M. Bousalem. 2006. Isolation, characterization and cross-species amplification of microsatellite DNA loci in the tropical American yam Dioscorea trifida. Mol. Ecol. Notes 6, 137-140. IBGE, 2007. Brazilian Institute of Geography and Statistics. En: www.ibge.com.br R aíces y tubérculos tropicales olvidados o subutilizados en B rasil Ike, P.C. y O.E. Inoni. 2006. Determinants of yam production and economic efficiency among smallholder farmers in southeastern Nigeria. J. Central Eur. Agr. 7, 337-342. Jackson, L.E.; U. Pascual y T. Hodgkin. 2007. Utilizing and conserving agrobiodiversity in agricultural landscapes. Agr. Ecosyst. Environ. 121, 196-210. Jarvis, D. I.; L. Myer; H. Klemick; L. Guarino; M. Smale; A.H.D. Brown; M. Sadiki; B. Sthapit y T. Hodgkin. 2000. A training guide for in situ conservation on-farm: version 1. International Plant Genetic Resources Institute, Rome, Italy. Kay, D.E. 1973. Root crops. The Tropical Products Institute, London. Klink, C.A. y R. Machado. 2005. Conservation of the Brazilian Cerrado. Conservation Biol. 19, 707-713. Kreyci P.F.; O.M. Iamamoto; M.V.B.M. Siqueira y E.A. Veasey. 2008. Otimização de protocolo de amplificação de marcadores microssatélites em Dioscorea bulbifera para análises genéticas. En: Anais 16. Simpósio Internacional de Iniciação Científica da USP, Piracicaba. Martin, F.W. y A. Jones. 1972. The species of Ipomoea closely related to the sweet potato. Econ. Bot. 26(3), 201-215. Martin, F.W. y A.M. Rhodes. 1977. Intra-specific classification of Dioscorea alata. Trop. Agr. 54, 1-13. Martins, P.S. 1994. Biodiversity and agriculture: patterns of domestication of Brazilian native plants species. Anais da Academia Brasileira de Ciências 66, 219-224. Martins, P.S. 2001. Dinâmica evolutiva em roças de caboclos amazônicos. pp. 369-384. En: Vieira, I.C.G.; J.M.C. Silva; D.C. Oren y M.A. D’incao (eds.). Diversidade Biológica e Cultural na Amazônia. Museu Paraense Emílio Goeldi, Belém, Brasil. Mignouna, H.D.; M.M. Abang y S.A. Fagbemi. 2003a. A comparative assessment of molecular marker assays (AFLP, RAPD and SSR) for white yam (Dioscorea rotundata) germplasm characterization. Ann. Appl. Biol. 142, 269-276. Mignouna, H.D; M.M. Abang y R. Asiedu. 2003b. Harnessing modern biotechnology for tropical tuber crop improvement: Yam (Dioscorea spp.) molecular breeding. Afr. J. Biotech. 2, 478-485. Lebot, V.; R. Malapa; T. Molisale y J.L. Marchand. 2005. Physico-chemical characterisation of yam (Dioscorea alata L.) tubers from Vanuatu. Genet. Resources Crop Evolution 00, 1-10. Miranda, J.E.C. 1983. Programa de melhoramento da batata-doce (Ipomoea batatas (L.) Lam.) no CNPHortaliças. p. 135. En: Resumo Congresso Brasileiro de Olericultura, Rio de Janeiro, SOB, Brasil. Lebot, V.; B. Trilles; J.L. Noyer y J. Modesto. 1998. Genetic relationships between Dioscorea alata L. cultivars. Genet. Resources Crop Evolution 45, 499-509. Miranda, J.E.C. y W.R. Maluf. 1984. Projeto de melhoramento genético da batata-doce. EMBRAPA-CNPH, Brasília (dados não publicados). Léotard, G. 2003. Phylogéographie et origine de la domestication du manioc (Manihot esculenta Crantz, Euphorbiaceae): les apports d’un échantillon élargi à l’écotone nord de l’Amazonie. DEA Biologie de l’Evolution et Ecologie, Université Montpellier II, Francia. Montaldo, A. 1991. Cultivo de raíces y tubérculos tropicales. Instituto Interamericano de Ciências Agrícolas de la OEA, São José. Madeira, N.R.; F.J.B. Reifschneider y L.B. Giordano. 2008. Contribuição portuguesa à produção e ao consumo de hortaliças no Brasil: uma revisão histórica. Hortic. Bras. 26, 428-432. Malapa, R.; G. Arnau; J.L. Noyer y V. Lebot. 2005. Genetic diversity of the greater yam (Dioscorea alata L.) and relatedness to D. nummularia Lam. and D. transversa Br. as revealed with AFLP markers. Genet. Res. Crop Evol. 52, 919-929. Martin, F.W. y A. Jones. 1971. Flowering and fertility in six generations of open pollinated sweet potatoes. Amer. Soc. Hort. Sci. 96(4), 493-495. Monteiro, D.A. e V.A. Peressin. 2002. Cultura do inhame. pp. 511-518. En: Cereda, M.P. (coord.). Agricultura: tuberosas amiláceas Latino Americanas. Fundação Cargill, São Paulo, Brasil. Mühlen, G.S.; P.S. Martins y A. Ando. 2000. Variabilidade genética de etnovariedades de mandioca, avaliada por marcadores de DNA. Scientia Agric. 57, 319-328. Muñoz, Y.C.G. 2003. Caracterización morfológica y molecular de genotipos de Dioscorea alata y D. trifida del Instituto de Investigación Agropecuaria de Panamá, IDIAP y CATIE, Costa Rica. Tese (Mestrado em Educación para el Desarrollo y la Conservación). Centro Agronómico Tropical de Investigación y Enseñanza-CATIE, Costa Rica. Vol. 3 - No.1 - 2009 123 124 Siqueira/Veasey Nishiyama, I. 1982. Autohexaploid evolution of the sweet potato. pp. 263-274. En: Villareal, R.L. e T.O. Grigs (eds.). Sweet potato: porceedings of the first international symposium. Shanhua, Tainan, Taiwan. Recchia, G.H.; A. Borges y E.A. Veasey. 2007. Protocolo de amplificação de marcadores microssatélites em Dioscorea cayenensis Lam. para análises genéticas. In: Anais 15. Simpósio Internacional de Iniciação Científica da USP. Ribeirão, Brasil. Oliveira, A.C.B.; M.A.N. Sediyama; T. Sediyama; F.L. Finger y C.D. Cruz. 2002. Variabilidade genética em batata-doce com base em marcadores isoenzimáticos. Hortic. Bras. 20, 576-582. Rodríguez, A.; M. García; M. Borges; S. Meneses; N. Aguilera; J. Vázquez; Z. Infante y M. Fonseca. 2003. In vitro conservation of Dioscorea alata L. germplasm by slow growth. Plant Genet. Resources Newsletter 133, 8-12. Olsen, K.M. 2004. SNPs, SSRs and inferences on cassava’s origin. Plant Mol. Biol. 56, 517-526. Olsen, K.M. y B.A. Schaal. 1999. Evidence on the origin of cassava: phylogeography of Manihot esculenta. Proceedings of the National Academy of Sciences 96, 5586-5591. Pedralli, G.; C.A.S. Carmo; C. Marney y M. Puiatti. 2002. Uso de nomes populares para as espécies de Araceae e Dioscoreaceae no Brasil. Hortic. Bras. 20, 530-532. Pereira, K.J.C. 2008. Agricultura tradicional e manejo da agrobiodiversidade na Amazônia Central: um estudo de caso nos roçados de mandioca nas Reservas de Desenvolvimento Sustentável Amanã e Mamirauá, Amazonas, Brasil. Tese (Doutorado) – Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, Brasil. Peroni, N. 1998. Taxonomia folk e diversidade intraespecífica de mandioca (Manihot esculenta Crantz) em roças de agricultura tradicional em áreas de mata atlântica no estado de São Paulo. Dissertação (Mestrado) – Escola Superior de Agricultura “Luíz de Queiróz”, Universidade de São Paulo, Piracicaba, Brasil. Peroni, N. y N. Hanazaki. 2002. Current and lost diversity of cultivated varieties, especially cassava, under swidden cultivation systems in the Brazilian Atlantic Forest. Agr. Ecosys. Environ. 92, 171-183. Peroni, N. y P.S. Martins. 2000. Influência da dinâmica agrícola itinerante na geração de diversidade de etnovariedades cultivadas vegetativamente. Interciencia 25, 22-29. Ramser, J.; C. López-Peralta; R. Wetzel; K. Weising y G. Kahl. 1996. Genomic variation and relationships in aerial yam (Dioscorea bulbifera L.) detected by random amplified polymorphic DNA. Genome 39, 17-25. REV. COLOMB. CIENC. HORTIC. Sambatti, J.B.M.; P.S. Martins y A. Ando. 2000. Distribuição da diversidade isoenzimática e morfológica da mandioca na agricultura autóctone de Ubatuba. Scientia Agric. 57, 75-80. Sambatti, J.B.M.; P.S. Martins y A. Ando. 2001. Folk taxonomy and evolutionary dynamics of cassava: a case study in Ubatuba, Brazil. Econ. Bot. 55, 93-105. Silva, J.B.C.; C.A. Lopes y J.S. Magalhães. 2002. Cultura da batata-doce. pp. 449-503. En: Cereda, M.P. (coord.). Agricultura: tuberosas amiláceas Latino Americanas, Vol. 2, Cap. 22. Fundação Cargill, São Paulo, Brasil. Scarcelli, N.; O. Daïnou; C. Agbangla; S. Tostain y J.L. Pham. 2005. Segregation patterns of isozyme loci and microsatellite markers show the diploidy of African yam Dioscorea rotundata (2n=40). Theor. Appl. Genet. 111, 226-232. Scarcelli, N.; Tostain, S.; C. Mariac; C. Agbangla; O. Da; J. Berthaud y J. Pham. 2006. Genetic nature of yams (Dioscorea sp.) domesticated by farmers in Benin (West Africa). Genet. Resources Crop Evolution 53, 121-130. Siqueira, M.V.B.M.; J.R.Q. Silva; E.A. Bressan; A. Borges; P. Kayo y E.A. Veasey. 2009a. Genetic characterization of cassava (Manihot esculenta) landraces in Brazil assessed with simple sequence repeats. Genet. Mol. Biol. 32, 104-110 Siqueira, M.V.B.M.; O.M. Iamamoto y E.A. Veasey. 2009b. Genetic characterization of landraces and commercial varieties of yam (Dioscorea alata) in Brazil with microsatellites markers. p. 51. En: Anais II Simpósio Brasileiro de Genética Molecular de Plantas, 1. Búzios, Brasil. Steinbauer, C.E. y L.J. Kushman. 1971. Sweet-potato culture and diseases. Agriculture Handbook No. 388. USDA, Washington, D.C. R aíces y tubérculos tropicales olvidados o subutilizados en B rasil Terauchi, R. y A. Konuma. 1994. Microsatellite polymorphism in Dioscorea tokoro, a wild yam species. Genome 37, 794-801. zilian sweetpotato (Ipomoea batatas (L.) Lam., Solanales, Convolvulaceae) landraces assessed with microsatellite markers. Genet. Mol. Biol. 31, 725-733. Valle, T.L.; Carvalho, C.R.L.; M.T.B. Ramos; G.S. Mühlen y O.V. Villela. 2004. Conteúdo cianogênico em progênies de mandioca originadas do cruzamento de variedades mansas e bravas. Bragantia 63, 221-226. Yen, D.E. 1976. Sweet potato – Ipomoea batatas (Convolvulaceae). pp. 42-45. En: Simmonos, N.W. (ed.). Evolution of crop plantas. Longman, London. Veasey, E.A.; Silva, J.R.Q.; M.S. Silva; A. Borges; E.A. Bressan y N. Peroni. 2007. Phenology and morphological diversity of sweet potato (Ipomoea batatas) landraces of the Vale do Ribeira. Scientia Agric. 64(4), 416-427. Veasey, E. A.; A. Borges; M.S. Rosa; J.R. Queiroz; A.A. Bressan y N. Peroni. 2008. Genetic diversity in Bra- Wedderburn, M.M. 1967. A study of hybridization involving the sweet potato and related species. Euphytica 16, 69-75. Zhang, D.P.; J. Cervantes; Z. Huamán; E. Carey y M. Ghislain. 2000. Assessing genetic diversity of sweet potato (Ipomoea batatas (L.) Lam.) cultivars from tropical America using AFLP. Genet. Resources Crop Evolution 47, 659-665. Vol. 3 - No.1 - 2009 125 Caracterização fisiologica de mudas de Jatropha curcas L. produzidas em diferentes níveis de irradiância Physiological characterization of Jatropha curcas L. plants grown under different levels of irradiance Fábio Santos Matos1,3 Cleílton Vasconcelos Moreira2 Robson Fernando Missio1 Luíz Antônio dos Santos Dias1 Plantulas Dos frutosde detomate. Jatropha curcas L. Foto: R.F. Missio Álvarez-Herrera. Resumo O incremento dos níveis dos gases de efeito estufa, notadamente dióxido de carbono (CO2) na atmosfera terrestre, tem intensificado a busca por combustíveis renováveis, tipo biodiesel, visando reduzir o consumo de combustíveis fósseis, responsáveis por 40% das emissões de CO2. Diversas são as vantagens do uso do biodiesel em comparação ao diesel de petróleo, dentre elas destaca-se a sustentabilidade do ecossistema. Neste contexto, a espécie Jatropha curcas é uma importante alternativa para produção de biodiesel. Neste trabalho, avaliou-se o efeito da irradiância sobre as características fisiológicas de folhas de J. curcas. O experimento foi conduzido em Viçosa (20°45’S, 42°54’W; 650 m de altitude), Minas Gerais, Brasil. Sementes de J. curcas foram semeadas em vasos plásticos de 12 L, contendo uma mistura de 30% de areia e 70% de solo. Metade das mudas foram cultivadas sob telado com 40% da radiação solar, e a outra metade a pleno sol. A irradiância de saturação foi semelhante entre os dois sistemas de cultivo. Entretanto, as plantas cultivadas a pleno sol apresentaram maiores fotossíntese, irradiância de compensação e acúmulo de massa seca. Estes resultados sugerem uma relativa aclimatação da maquinaria fotossintética de J. curcas à variação da irradiância. Palavras-chave: biodiesel, energia renovável, sustentabilidade ambiental. 1 Departamento de Fitotecnia (DFT), Universidade Federal de Viçosa (UFV), Viçosa, MG, Brasil. Universidade Federal de Lavras (UFLA), Lavras, MG, Brasil. 3 Autor para correspondência. [email protected] 2 REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 3 - No.1 - pp. 126-134, 2009 C aracteriza ç ã o fisiologica de mudas de J a t r o p h a c u r c a s L . ABSTRACT The increase in carbon dioxide levels in the atmospheres has intensified the search for renewable fuel sources in order to further reduce the consumption of fossil fuel. There are several advantages of employing biodiesel as compared to conventional diesel, but the main benefit of using biodiesel is the sustainability of the ecosystem. In this context, the species Jatropha curcas is an important alternative for production of biodiesel. This study aimed at assessing the effects of irradiation on the physiological characteristics of leaves of that species. The experiment was conducted in Viçosa (20º45’S, 42°54’W, 650 m altitude) Minas Gerais state, Brazil. Seeds were sown in 12 L pots, containing a mixture 30% sand and 70% soil. A half of plants was grown under black shade 40% of solar radiation treatment (T1), and the other half was exposed to full irradiation, constituting treatment T2. Despite the saturation irradiance was similar between treatments, the leaves at full sun exhibited higher photosynthesis, compensation irradiance and increased accumulation of dry mass. These results suggest a high acclimation of the photosynthetic machinery of J. curcas to the variation of irradiance. Additional keywords: biofuel, renewable energy, environmental sustainability. Fecha de recepción: 03-02-2009 Aprobado para publicación: 01-06-2009 Introdução O incremento dos níveis dos gases de efeito estufa (GEE), notadamente dióxido de carbono (CO2) na atmosfera terrestre, tem intensificado a busca por combustíveis renováveis, tipo biodiesel, visando reduzir o consumo de combustíveis fósseis. O combustível renovável denominado biodiesel consiste de metil ésteres de ácidos graxos, usualmente derivado de óleo vegetal ou animal. Diversas são as vantagens da utilização do biodiesel quando comparado com o diesel convencional, no entanto, a principal delas é a sustentabilidade da produção, isto porque a energia contida na cadeia de triacilglicerol é derivada da luz solar. Quando comparado com o diesel convencional, o biodiesel reduz a emissão de GEE em 40% (Hill et al., 2006). A cultura do pinhão manso ( Jatropha curcas L.) é uma importante alternativa para produção de biodiesel. No atual contexto político, com o programa nacional de produção e uso do biodiesel (PNPB), esta cultura torna-se extremamente importante por produzir óleo com características semelhantes ao diesel. No entanto, os plantios têm sido estabelecidos com genótipos silvestres, com praticamente nenhum grau de melhoramento. O pinhão manso pertence à família das euforbiáceas, a mesma da mamona (Ricinus communis). É um arbusto, de crescimento rápido, cuja altura pode atingir de 2 a 3 m, mas pode alcançar até 5 m em condições especiais. A planta adulta tem diâmetro do tronco de aproximadamente 20 cm, com raízes curtas e pouco ramificadas, caule liso, de lenho mole e medula desenvolvida, mas pouco resistente a ventos e floema com longos canais que se estendem até as raízes, nos quais circulam o látex. O tronco ou fuste é dividido desde a base, em compridos ramos, com numerosas cicatrizes provocadas pela queda das folhas na estação seca, as quais ressurgem logo após as primeiras chuvas (Arruda et al., 2004). O óleo é extraído dos grãos de pinhão manso, os quais são submetidos a cozimento prévio e prensagem subseqüente em prensas tipo “expeller”. Posteriormente, o óleo é filtrado, centrifugado e Vol. 3 - No.1 - 2009 127 128 MATOS/MOREIRA/MISSIO/DIAS clarificado, resultando em um produto livre de impurezas (Ministério da Indústria e do Comércio, 1985). A torta resultante da prensagem dos grãos contém ainda, aproximadamente 8% de óleo, o qual é re-extraído com solventes orgânicos, geralmente hexano, sendo o farelo residual ensacado para aproveitamento como fertilizante natural, em virtude dos teores elevados de nitrogênio, fósforo, potássio e material orgânico. Desintoxicada, a torta pode também ser transformada em ração, como tem sido feita com a torta de Ricinus communis. O óleo do pinhão manso tem 83,9% do poder calorífico do óleo diesel, e a sua utilização pode gerar a mesma potência (Fundação CETEC, 1983). Modificações nos níveis de luminosidade para os quais uma espécie está aclimatada pode acarretar diferentes respostas fisiológicas em sua características bioquímicas, anatômicas e de crescimento (Atroch et al., 2001). Em geral, as plantas desenvolvem “folhas de sol” e “de sombra”, quando aclimatadas a diferentes níveis de luminosidade. Área foliar específica, espessura da cutícula, densidade estomática, cloroplastos com menos ou mais grana e menos ou mais tilacóides por granum, são algumas das características que variam em resposta à irradiância (Fahl et al., 1994). Obviamente, não apenas a irradiância interceptada, mas também a temperatura foliar deve variar ao longo do dossel, com possíveis reflexos sobre o comportamento estomático e a fotossíntese. A atividade fotossintética é um parâmetro relacionado com o ganho de biomassa da planta. A análise de crescimento tem sido usada na tentativa de explicar diferenças no crescimento, de ordem genética ou resultante de modificações do ambiente (Peixoto, 1998) e constitui uma ferramenta muito eficiente para a identificação de materiais promissores (Benincasa, 2003), além de identificar características que, no crescimento inicial, indiquem possibilidade de aumento no rendimento da planta adulta, favorecendo os trabalhos de melhoramento na busca por materiais mais produtivos. O fundamento da análi- REV. COLOMB. CIENC. HORTIC. se de crescimento baseia-se no fato de que, em média, 90% da matéria orgânica acumulada ao longo do crescimento da planta resultam da atividade fotossintética e o restante, da absorção mineral do solo (Benincasa, 2003). Em experimentos com culturas de ciclos longos, em que a mensuração da produtividade não seja possível devido ao longo período para desenvolvimento dos frutos, a mensuração da fotossíntese associada com a análise de crescimento da planta pode prever uma produtividade potencial da cultura. Não se conhece o grau de tolerância e a capacidade de aclimatação da maquinaria fotossintética do pinhão manso a fatores ambientais, como o sombreamento imposto pelo consórcio com outras culturas, que poderiam ser limitantes para a introdução desta espécie fora de seu habitat natural. Folhas submetidas ao sombreamento com irradiância inferior ao ponto de saturação luminoso possuem menor atividade fotossintética devido principalmente a menor energia fotoquímica disponível. O pinhão manso por ter evoluído em ambientes sob alta luminosidade, deve possuir relativamente baixa aclimatação de sua maquinaria fotossintética para ajustar-se à disponibilidade de luz. Tendo em vista a necessidade de buscar informações dessa natureza para a produção de mudas para o plantio em campo, bem como o melhor entendimento dos atributos para tolerar o sombreamento, possibilitando sua exploração comercial, avaliou-se neste trabalho o efeito do sombreamento sobre as características fisiológicas de folhas em mudas de J. curcas. Material e Métodos Material vegetal e condições de cultivo O experimento foi conduzido em Viçosa (20°45’S, 42°54’W, 650 m de altitude, Minas Gerais, Brasil, em delineamento inteiramente casualizado. Sementes de J. curcas L. foram semeadas em vasos plásticos de 12 L, contendo uma mistura de 30% de areia e 70% de solo. Após a análise da composição química da mistura, foi realizada a adu- C aracteriza ç ã o fisiologica de mudas de J a t r o p h a c u r c a s L . bação e a correção do pH, de acordo com análise de solo e recomendações técnicas para a cultura (Dias, 2000). A umidade do substrato foi mantida próxima à capacidade de campo, via irrigações periódicas. Metade das plantas foi cultivada sob telado (Sombrite®) com 40% da radiação solar, constituíndo um tratamento (T1) e a outra metade a pleno sol constituindo outro tratamento (T2). As análises fisiológicas foram realizadas em folhas totalmente expandidas de plantas com 80 dias após a germinação. Trocas gasosas e parâmetros de fluorescência A taxa de assimilação líquida do carbono (A), a condutância estomática (gs) e a razão entre as concentrações interna e ambiente de CO2 (Ci/Ca) foram medidas entre às 09:00-11:00 h, em sistema aberto, sob concentração de CO2 ambiente, com um analisador de gases a infravermelho portátil (LICOR 6400, Li-COR, Lincoln, NE). Curvas de resposta da taxa de assimilação liquida de carbono (A) à irradiância (curva A/RFA) foram obtidas em laboratório, em folhas de ramos coletados no início da manhã, utilizando-se de um analisador de gases a infravermelho (LCpro+, Analytical Development Company, Hoddesdon, UK), variando-se a RFA de 0 a 1.400 µmol (fótons) m2 s-1, a 25ºC, e concentração de CO constante 2 de 380 µmol mol-1. Antes de se aplicar a RFA às folhas, foi fornecida uma concentração inicial de CO2 igual a 50 µmol mol-1, por 3 min, para induzir a abertura dos estômatos. O rendimento quântico aparente (Φa) foi estimado por meio da regressão linear da porção inicial da curva [0 ≤ RFA ≤ 100 µmol (fótons) m-2 s-1]. Os parâmetros de fluorescência da clorofila (CHL) a foram determinados naqueles horários, bem como na antemanhã, nas mesmas folhas utilizadas para as medições das trocas gasosas, por meio do fluorômetro com pulso modulado (FMS2, Hansatech, Norfolk, UK). Após serem adaptados ao escuro, por 30 min, tecidos foliares foram inicialmente expostos a pulsos de luz vermelho-distante [1-2 µmol (fótons) m-2 s-1] para a determinação da fluorescência inicial (F0). Em seguida, pulsos de luz saturante, com irradiância de 6.000 µmol (fótons) m-2 s-1 e duração de 1 s, foram aplicados para determinar a fluorescência máxima emitida (Fm). Nas amostras adaptadas ao escuro a eficiência máxima do FSII foi estimada pela razão entre as fluorescências variável e máxima, sendo a fluorescência variável obtida pela subtração entre a fluorescência máxima e mínima. Fv/Fm = (Fm-F0)/Fm. Para a obtenção da fluorescência em steady-state (Fs), foi utilizada uma irradiância actínica artificial de 500 e 1000 µmol (fótons) m-2 s-1, respectivamente para plantas cultivadas à sombra ou a pleno sol. Essa irradiância, aplicada por 300 s, foi equivalente à irradiância média interceptada pelas folhas à sombra e ao sol. Subseqüentemente, pulsos de luz saturante [6.000 µmol (fótons) m-2 s-1; 1 s] foram aplicados para medir-se a fluorescência máxima das amostras sob luz (Fm’). Pigmentos fotossintéticos e nitrogênio Clorofilas e carotenóides totais (Car) foram quantificados espectrofotometricamente, utilizando quatro discos foliares (1,4 cm de diâmetro). Amostras foliares foram coletadas e determinadas as diferenças de N orgânico e N nítrico, conforme descrito por Cataldo et al. (1974, 1975). A soma das duas frações representa o N-total. Massa seca Após realizadas todas as análises não destrutivas, iniciou-se a separação da parte aérea e do sistema radicular, sendo ambos colocados em estufa à 80ºC por 72 h para quantificação da massa seca da parte aérea e sistema radicular. Delineamento experimental e análise estatística O experimento, em delineamento inteiramente casualizado, foi analisado considerando dois tratamentos, cinco repetições e parcelas de 1 muda por vaso. Vol. 3 - No.1 - 2009 129 130 MATOS/MOREIRA/MISSIO/DIAS As análises de variância foram processadas e as diferenças entre médias dos tratamentos inferidas do próprio teste F, em nível de 5% de probabilidade, das análises de variância. CO2 ambientes, foram, em média, 48,0%, 44,9% e 63,6% maior nas folhas de T2, quando comparada com folhas de T1, respectivamente (Tabela 2). A razão Fv/Fm foi superior a 0,82 em todos os tratamentos, indicando ausência de fotoinibição da fotossíntese (dados não mostrados). Resultados Nitrogênio total e pigmentos Trocas gasosas e parâmetros de fluorescência A concentração foliar de nitrogênio total, em base de massa, foi, em média, 20% maior nas folhas de T1 quando comparada com T2. A área foliar unitária foi maior (~59,2%) nas folhas de T1 em relação às de T2. A concentração de clorofilas totais, em base de massa, foi significativamente maior nas folhas de T1 em relação às de T2. A concentração de carotenóides totais (em base de massa) bem como a razão entre as clorofilas a/b foi superior nas folhas de T2 em relação à T1, enquanto a razão entre clorofilas e carotenóides foi, em média, 60,8% maior nas folhas de T1 em relação às de T2. As razões clorofilas totais/N e clorofilas/Car foram relativamente maiores nas folhas de T1 em relação às de T2, fatos explicáveis, principalmente, em função da maior concentração de clorofilas totais em T1. A razão clorofila a/b não respondeu estatisticamente aos diferentes tratamentos (tabela 2). A irradiância de compensação (Ic) foi, em média, 35,6% maior nas plantas a pleno sol, quando comparada com as plantas sombreadas, variando de 10,61 a 16,47 µmol (fótons) m-2 s-1 (tabela 1). No entanto, a irradiância de saturação (Is) foi semelhante, independemente dos tratamentos (tabela 2), ficando em torno de 460 µmol (fótons) m-2 s-1. A respiração foi, em média, 39,7% maior nas folhas de T2, quando comparada com folhas de T1. A taxa de assimilação líquida do carbono saturada pela luz (ARFA) foi semelhante, ao compararem-se as folhas de T1 e T2, enquanto o rendimento quântico aparente (Φa) foi, em média, 32,9% maior em T2 quando comparada com a de T1. A taxa de assimilação líquida do carbono (A), a condutância estomática (gs) e a razão entre a concentração interna e ambiente de CO2 (Ci/Ca) quando medidas em sistema aberto, sob luz e concentração de Tabela 1.Irradiância de compensação [Ic, µmol (fótons) m -2 s -1], irradiância de saturação [Is, µmol (fótons) m -2 s -1], rendimento quântico aparente [Φa, mol (CO2) mol -1 (fótons)], respiração [Rd µmol m -2 s -1] e taxa de assimilação líquida de carbono saturada pela luz [ARFA, µmol (CO2) m -2 s -1] obtidas a partir de curvas A/RFA de folhas de Jatropha curcas L. Parâmetros Tratamentos* T1 T2 Ic 10,61 ± 1,71 A 16,47 ± 3,9 A Is 458,5 ± 1,07 A 466,6 ± 4,31A Φa A 0,073 ± 0,009 A Rd 0,44± 0,020 A 0,73± 0,03 A ARFA 0,30 A 13,35 ± 0,75 A 0,049 ± 0,004 14,18 ± *Valores representam a média ± erro-padrão (n=7). Médias seguidas pela mesma letra na linha não diferem entre si, a 5% de probabilidade, pelo teste F. T1: plantas à sombra (40% de radiação solar); T2: plantas a pleno sol. REV. COLOMB. CIENC. HORTIC. C aracteriza ç ã o fisiologica de mudas de J a t r o p h a c u r c a s L . Tabela 2. Concentrações foliares de N-total, clorofilas (Cl) totais (a+ b) e carotenóides totais (Car), e razões Cl/N, Cl a/ Cl b (Cl a/b) e Cl/Car de folhas de Jatropha curcas L. Tratamentos Parâmetros T1 T2 A, µmol (CO2) m-2 s-1 4,96 ± 0,26B 9,54 ± 0,90A gs, mmol (H2O) m-2 s-1 0,138 ± 0,008A 0,076 ± 0,011B 0,77 ± 0,09A 0,28 ± 0,06 B 24,81 ± 0,41A 19,84 ± 0,38B Cl (a+b), g kg-1 MS 6,12 ± 0,32A 3,18 ± 0,18 B Car, g kg-1 MS 0,94 ± 0,12 A 1,19 ± 0,13A Cl/N, mmol mol-1 6,40 ± 0,20A 2,69 ± 0,18B Cl a/b 1,77 ± 0,25A 2,44 ± 0,43A Cl/Car 7,27 ± 1,36A 2,85 ± 0,35 B Ci /Ca N total, g kg-1 MS Valores representam a média ± erro-padrão (n=7). Médias seguidas pela mesma letra na linha não diferem entre si, a 5% de probabilidade, pelo teste F. T1, plantas à sombra (40% de radiação solar); T2, plantas a pleno sol. Análise de crescimento Discussão A massa seca da parte aérea e sistema radicular foram, em média, 33,6% e 64,0% maiores nas folhas de T2 em relação às de T1, respectivamente, resultando em uma razão parte aérea/sistema radicular 46,5% maior no tratamento T2, quando comparado com T1. No entanto, o diâmetro do caule e altura das plantas, foram maiores no tratamento T1 (tabela 3). Aclimatação a baixas irradiâncias Este trabalho é o primeiro a explorar as características fisiológicas de mudas de Jatropha curcas L. De modo geral, a área foliar unitária aumentou nas folhas mais sombreadas. O incremento da superfície foliar pode ser atribuído tanto ao aumento do número de células, devido ao aumento Tabela 3.Área foliar, massa seca da parte aérea e sistema radicular, diâmetro do caule e altura da planta de Jatropha curcas L. distribuídas em dois tratamentos, correspondendo aos tratamentos à sombra (T1) e a pleno sol (T 2). Parâmetros Tratamentos T1 T2 Área foliar (cm) 397 ± 22,0 A 162 ± 12,3 B Massa seca parte aérea (g) 4,24 ± 0,29 B 6,29 ± 0,63 A Massa seca raiz (g) 0,63 ± 0,038B 1,78 ± 0,23 A Razão parte aérea/ raiz 6,77 ± 0,23 A 3,62 ± 0,28B Diâmetro do caule (cm) 1,58 ± 0,058 A 1,12 ± 0,020 B Altura da planta (cm) 14,78 ± 1,14A 13,52 ± 0,83A Valores representam a média ± erro-padrão (n=7). Médias seguidas pela mesma letra na linha não diferem entre si, a 5% de probabilidade, pelo teste F. T1, plantas à sombra (40% de radiação solar); T2, plantas a pleno sol. Vol. 3 - No.1 - 2009 131 132 MATOS/MOREIRA/MISSIO/DIAS das divisões celulares, quanto ao maior volume celular (Fahn, 1990). Nesse sentido, nas folhas das plantas cultivadas à sombra, a expansão celular pode ter sobrepujado as divisões celulares que acarretam aumento do número de células. Apesar do fato de o número de divisões celulares ser controlado geneticamente (Nadeau e Sack, 2003), o ambiente pode exercer forte pressão e alterar esse padrão. Dessa forma, folhas sombreadas, freqüentemente, apresentam incremento do número de divisões celulares, como forma de aumentar a área foliar, conforme descrito para Populus sp. (Ceulemans et al., 1995). tica nas plantas deste tratamento, uma vez que a atividade fotossintética mensurada em sistema aberto nestas folhas foi muito inferior a atividade fotossintética de T2. Fisiologicamente, a baixa Ic deve permitir a obtenção de um balanço positivo de carbono sob irradiâncias extremamente baixas. Os valores reduzidos de Ic observados nas folhas mais sombreadas permitiu a obtenção de um balanço positivo de carbono sob menor irradiância. Isso esteve provavelmente associado com uma menor taxa respiratória, maior concentração de clorofilas totais em base de massa e maior razão Cl/N. A maior razão Cl/N nas folhas de sombra indica maior alocação de N para produção de pigmentos fotossintéticos, em detrimento de compostos diretamente envolvidos na fixação do CO2 (Evans, 1989; Ellsworth e Reich, 1993). No entanto, não ocorreu redução da razão Cl/a/b – uma das principais respostas observadas em plantas (folhas) desenvolvidas à sombra (Walters, 2005). Contudo, a razão Cl/a/b não foi afetada pelas variações da irradiância. Essa razão é um indicador da proporção de complexos coletores de luz associados ao FSII (CCL-II), em relação a outros complexos contendo clorofilas (Murchie & Horton, 1997). As menores concentrações de carotenóides, bem como, menor razão clorofila/carotenóide nas folhas de T1 resultam em menor fotoproteção nas folhas deste tratamento. Isso poderia explicar, em parte, o porque de mudas de pinhão manso, quando transferidas de viveiros sombreados para o campo exibirem, usualmente, sintomas típicos de escaldadura (danos fotooxidativos). Aclimatação a altas irradiâncias A menor massa seca das plantas de T1 está possivelmente associada a menor atividade fotossintéREV. COLOMB. CIENC. HORTIC. Os resultados sugerem que o pinhão manso possui relativamente alta aclimatação da maquinaria fotossintética à disponibilidade de luz, particularmente a Ic, que deve estar intimamente relacionada com a aclimatação de J. curcas a baixas irradiâncias, e em boa extensão, pode explicar um possível sucesso do cultivo da espécie em condição de consórcio com outras culturas. A maior massa seca nas plantas de T2 esteve associado a maior atividade fotossintética nas folhas deste tratamento, enquanto o menor diâmetro do caule das plantas de T2 pode ser resultado da menor alocação de assimilados para o caule. A menor razão Ci/Ca nas folhas à pleno sol, está intimamente relacionada a maior taxa de assimilação líquida do carbono (A) nestas folhas, uma vez que não houve restrição estomática. No entanto, a maior disponibilidade de energia luminosa parece explicar a maior atividade fotossintética das folhas de T2. As plantas podem apresentar diversos meios para se protegerem contra irradiâncias potencialmente fotoinibitórias. Neste trabalho, a redução da concentração de clorofilas, nas folhas de T2 em relação às folhas de T1, deve ter auxiliado na redução da absortância foliar e, associada com os menores valores da eficiência de captura de energia de excitação pelos centros de reação abertos do PSII, reduzido a quantidade total de energia efetivamente absorvida pelos fotossistemas. Decréscimos na concentração de clorofilas totais em folhas submetidas a maiores níveis de irradiância podem estar associados a processos fotoxidativos (e.g., Krause, 1988), ou estar relacionado com alterações na organização dos fotossistemas, de modo a servir como um mecanismo fotoprotetor (e.g., Elvira et al., 1998; Ottander et al., 1995). A segunda hipótese é mais provável, uma vez que não houve qualquer indício de dano oxidativo (razão Fv/Fm ≥ 0,82), ape- C aracteriza ç ã o fisiologica de mudas de J a t r o p h a c u r c a s L . sar da grande diferença da irradiância interceptada entre as folhas dos tratamentos. Conforme esperado, houve um decréscimo na razão Cl/Car nas folhas sob maiores níveis de luz, de modo a permitir-lhes um aumento na capacidade de dissipação da energia de excitação, principalmente via ciclo das xantofilas (Demmig-Adams e Adams, 1996a; Ma et al., 2003). Com efeito, a elevação na concentração de carotenóides e/ou uma redução da concentração de clorofilas pode auxiliar as plantas a minimizarem a fotooxidação (Corcuera et al., 2005). Desta forma, as mudas desenvolvidas a pleno sol, apresentam características fisiologicamente desejáveis como maiores taxas fotossintética, ca- pacidade de dissipação do excesso de energia luminosa e maior rendimento quântico, bem como maior acúmulo de massa seca, o que corrobora com a origem da espécie em ambiente de elevada irradiância, supostamente regiões semi-áridas. Conclusões Tomados em conjunto, estes resultados explicam, em boa extensão, um possível sucesso do cultivo da espécie à sombra ou a pleno sol, ainda que tenha evoluído em ambientes a pleno sol. As mudas de J. curcas L. demonstraram relativa variação das características fisiológicas quando submetidas a diferentes irradiâncias. Referências BIBLIOGRAFICAS Arruda, F.P. de; N.E. de M. Beltrão; A.P. de Andrade; W.E. Pereira e L.S. Severino. 2004. Cultivo do Pinhão manso (Jatropha curcas L.) como alternativa para o semi-árido nordestino. Revista Brasileira de Oleaginosas e Fibrosas 8(1), 789-799. Corcuera, L.; F. Morales; A. Abadía e E. Gil-Pelegrín. 2005. Seasonal changes in photosynthesis and photoprotection in a Quercus ilex subsp. ballota woodland located in its upper altitudinal extreme in the Iberian Peninsula. Tree Physiol. 25, 599-608. Atroch, E.M.A.C.; A.M. Soares; A.A. Alvarenga e E.M. Castro. 2001. Crescimento, teor de clorofilas, distribuição de biomassa e características anatômicas de plantas jovens de Bauhinia forticata Link submetidas a diferentes condições de sombreamento. Ciência e Agrotecnologia 25, 853-862. Demmig-Adams, B. e W.W. Adams. 1996. Photoprotection and other responses of plants to high light stress. Annu. Rev. Plant Physiol. Plant Mol. Biol. 43, 599-626. Benincasa, M.M.P. 2003. Análise de crescimento de plantas: noções básicas. Funep, Jaboticabal, Brasil. Cataldo, D.A.; L.E. Schrader e V.L. Youngs. 1974. Analysis by digestion and colorimetric assay of total nitrogen in plant tissues high in nitrate. Crop Sci. 14, 854-856. Cataldo, D.A.; M. Haroon; L.E. Schrader e V.I. Youngs. 1975. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Comm. Soil Sci. Plant Anal. 6, 71-80. Ceulemans, R.; L.V. Praet e X.N. Jiang. 1999. Effects of CO2 enrichment, leaf position and clone on stomatal index and epidermal cell density in poplar (Populus). New Phytologist 131, 99-107. Dias, L.A.S. 2000. Calagem e adubação. pp. 18-20. In: Cultivo de pinhão manso Jatropha curcas L. para produção de óleo combustível. Universidade Federal de Viçosa, Viçosa, Brasil. Ellsworth, D.S. y P.B. Reich. 1993. Canopy structure and vertical patterns of photosynthesis and related leaf traits in a deciduous forest. Oecologia 96, 169-178. Elvira, R.; S. Alonso; J. Castilho e B.S. Gimeno. 1998. On the response of pigments and antioxidants of Pinus hapelensis to mediterranean climate factors and long-term ozone exposure. New Phytologist 138, 419-432. Evans, J.R. 1989. Photosynthesis and nitrogen relationship in leaves of C3 plants. Oecologia 78, 9-19. Fahl, J.I.; M.L.C. Carelli; J. Vega e A.C. Magalhães. 1994. Nitrogen and irradiance levels affecting net pho- Vol. 3 - No.1 - 2009 133 134 MATOS/MOREIRA/MISSIO/DIAS tosynthesis and growth of young coffee plants (Coffea arabica L.). J. Hort. Sci. 69, 161-169. Fahn, A. 1990. Plant anatomy. 2nd ed. Butterworth Heinemann, Oxford, UK. Fundação Centro Tecnológico de Minas Gerais (CETEC). 1983. Produção de biocombustíveis líquidos a partir de óleos vegetais. Relatório final. Belo Horizonte, Brasil. Hill, J.; E. Nelson; D. Tilman; S. Polasky e D. Tiffany. 2006. Environmental, conomic, and energetic costs and benefits of biodiesel and ethanol biofuels. Proc. Natl. Acad. Sci. USA 103, 11206-11210. Krause, G.H. 1988. Photoinhibition of photosynthesis - an evaluation of damaging and protective mechanisms. Physiol. Plant. 74, 566-574. Ma, Y-Z.; N.E. Holt; X.-P. Li; K.K. Niyogi e G.R. Fleming. 2003. Evidence for direct carotenoid involvement in the regulation of photosynthetic light harvesting. Proc. Natl. Acad. Sci. USA 100, 43774382. REV. COLOMB. CIENC. HORTIC. Ministério da indústria e do comércio. Secretária de tecnologia industrial. 1985. Produção de combustíveis líquidos a partir de óleos vegetais. STI/CIT, Brasília. Murchie, E.H. e P. Horton. 1997. Acclimation of photosynthesis to irradiance and spectral quality in British plant species: Chlorophyll content, photosynthetic capacity and habitat preference. Plant Cell Environ. 20, 438-448. Nadeau, J.A. e F.D. Sack. 2003. Control of stomata distribution on the Arabidopsis leaf surface. Sci. 296, 1700. Ottander, C.; D. Campbell e G. Oquist. 1995. Seasonal changes in photosystem-Ii organization and pigment composition in Pinus-Sylvestris. Planta 197, 176-183. Peixoto, C.P. 1998. Análise de crescimento e rendimento de três cultivares de soja em três épocas de semeadura e três densidades de plantas. Tese (Doutorado em Fitotecnia). Escola Superior de Agricultura Luiz de Queiroz,Universidade de São Paulo, Piracicaba, Brasil. Walters, R.G. 2005. Towards an understanding of photosynthetic acclimation. J. Exp. Bot. 56, 435-447. 135 POLÍTICA EDITORIAL La Revista Colombiana de Ciencias Hortícolas es el órgano de divulgación de la Sociedad Colombiana de Ciencias Hortícolas (SCCH – www.soccolhort.com), adscrita a la ISHS (Internacional Society for Horticultural Science – www.ishs.org), con frecuencia semestral en el territorio nacional. Constituye una publicación abierta a la discusión y difusión de trabajos técnico-científicos en el área de las ciencias agrícolas, con énfasis en horticultura (frutales, hortalizas, ornamentales, hierbas aromáticas, viveros) y disciplinas afines, propuestos por autores nacionales e internacionales. Busca divulgar trabajos inéditos desarrollados por investigadores de diversas universidades y centros de investigación del país y difundir y someter a discusión los avances científicos que se producen, con el fin de contribuir a la consolidación de una comunidad académica congregada en torno a las disciplinas afines a la horticultura. Los artículos deben presentarse según las normas establecidas en las instrucciones para los autores. El Comité Editorial se reserva el derecho de aceptar o no las contribuciones que se hagan a la revista. Los manuscritos se enviarán a dos pares evaluadores expertos en el área específica del conocimiento, quienes decidirán si el artículo tiene el nivel científico necesario para publicarse. Si se llega a presentar alguna diferencia de conceptos en cuanto a la aprobación para publicación, el artículo será enviado a un tercer par, quien entregará la decisión final. Si el artículo se acepta para publicación, el autor debe realizar las correcciones sugeridas en el tiempo indicado por el editor. La Revista Colombiana de Ciencias Hortícolas autoriza la reproducción y citación del contenido de la publicación, siempre y cuando se registre de forma explícita el nombre de la revista, nombre del autor (es), año, volumen, número y página del artículo consultado. Los artículos publicados en la Revista serán enviados en medio magnético a las diferentes bases de datos y sistemas de indexación (Publindex), para la divulgación de su contenido y se podrán consultar mediante Internet. Las ideas y afirmaciones emitidas por los autores son responsabilidad de éstos y no expresan necesariamente las opiniones y políticas del Comité Editorial de la SCCH, ni de las instituciones a las cuales están vinculados quienes las proponen. INSTRUCCIONES PARA LOS AUTORES Los artículos puestos a consideración del Comité Editorial de la Revista Colombiana de Ciencias Hortícolas deben ser inéditos; en consecuencia, aquellos manuscritos que hayan sido publicados en otras revistas o publicaciones técnico-científicas no serán aceptados. La Revista Colombiana de Ciencias Hortícolas acepta artículos originales de los siguientes tipos, en concordancia con lo requerido por el programa Publindex de Colciencias para la categoría B: • Artículo de investigación científica y tecnológica: documento que presenta de manera detallada los resultados originales de proyectos de investigación. La estructura generalmente utilizada tiene cuatro partes esenciales: introducción, metodología (materiales y métodos), resultados y discusión, y conclusiones. • Artículo de reflexión: documento que presenta resultados de investigación desde una perspectiva analítica, interpretativa y crítica del autor, sobre un tema específico y recurriendo a fuentes originales. Vol. 3 - No.1 - 2009 136 • Artículo de revisión: documento resultado de una investigación en la que se analizan, sistematizan e integran los resultados de investigaciones, publicados o no, sobre un campo en ciencia o tecnología, con el fin de dar cuenta de los avances y tendencia de desarrollo; se caracteriza por presentar un soporte bibliográfico cuidadoso no menor a 50 referencias. Extensión y remisión del artículo y de la ilustración La extensión del artículo de investigación no debe exceder las 5.200 palabras (incluido literatura y las tablas), excepto el artículo de revisión, que puede contener hasta 6.500 palabras; tamaño carta, escritas a doble espaciado, letra o fuente Times New Roman con tamaño de 12 puntos, márgenes de 3 cm en la parte superior, 2 cm en la inferior y 2,5 cm en las márgenes laterales derecha e izquierda. Las tablas y figuras (es decir, los gráficos, dibujos, esquemas, diagramas de flujo, fotos y mapas) deben presentarse en hojas independientes y con numeración consecutiva (Tabla 1 ... Tabla n. Figura 1 ... Figura n; etc.). Los textos y tablas deben presentarse en el procesador de palabra MS-Word®; las tablas y los diagramas de frecuencia (barras y torta) originales deben suministrarse en el archivo del manuscrito y también en su original de MS-Excel®; otras figuras, como fotografías sobre papel y dibujos, se pueden enviar en originales o escanearlas y remitirlas en el formato digital de compresión JPG (o JPEG), preferiblemente con una resolución de 600 x 600 dpi (mínimo 300 dpi). Como norma general, las tablas y figuras sólo se aceptan en blanco y negro. El artículo con sus archivos en Word, Excel y JPG debe ser enviado al correo electrónico: [email protected], al editor profesor Dr. Fánor Casierra, [email protected] o coeditor profesor Dr. Gerhard Fischer, [email protected]. Título y autores El título es en minúscula y no debe exceder las 15 palabras. Es obligatoria su respectiva traducción al idioma inglés. Cuando incluya nombres científicos de vegetales o animales, éstos se deben escribir con letra cursiva (itálica) y en minúsculas, y sólo con mayúsculas la primera letra del género y el clasificador. Debajo del título en inglés se escribe el nombre(s) y apellido(s) de los autores, sin sus respectivos títulos académicos ni cargos laborales, en una línea horizontal y de acuerdo con su contribución en la investigación o preparación del artículo. En la parte inferior de la primera página, es decir, en aquélla que contiene el título del artículo, el nombre y la ciudad de ubicación de la entidad a la cual prestan sus servicios o del patrocinador para la realización del trabajo y el autor para correspondencia con su correo electrónico (e-mail). Resumen, abstract y palabras clave adicionales El resumen debe escribirse en español, portugués o inglés, correspondiente al idioma del artículo. Los artículos en español o portugués deben contener un abstract en inglés, un artículo en inglés y un resumen en español. Ambos textos deben describir en forma breve el problema, los métodos utilizados, su justificación y los resultados obtenidos más relevantes, y no deben exceder de 250 palabras escritas en un único párrafo. Es obligatorio acompañar el resumen con máximo seis palabras clave, que no hayan sido usadas en el título, y de su traducción a inglés (additional key words), las que tampoco deben figurar en el título en inglés. Introducción Texto que debe contener la situación actual del problema, su definición y la revisión de los trabajos previos relacionados con él; además, los objetivos y la justificación de la investigación. Es obligatorio acompañar los nombres vulgares con el nombre(s) científico(s) y la abreviatura(s) del clasificador en la primera mención en el artículo. Materiales y métodos En este apartado se deben describir de forma clara, concisa y secuencial los materiales (vegetales, animales, implementos agrícolas o de laboratorio) utilizados en desarrollo del trabajo, además de los procedimientos o protocolos seguidos y el diseño escogido para el tratamiento estadístico de los datos. Resultados y discusión Los resultados deben presentarse de manera lógica, objetiva y secuencial mediante textos, tablas y figuras; estos dos últimos apoyos deben ser fáciles de leer y deben poderse interpretar de manera autónoma, aunque deben citarse siempre en el texto. Las gráficas serán bidimensionales y a una sola tinta, con porcentajes de negro para las variaciones de las columnas; las líneas de las curvas deben ser de color negro, punteadas o continuas (- - - - ó --------), usando convenciones como: l, , s, ∆, etc. REV. COLOMB. CIENC. HORTIC. 137 Las tablas se deben elaborar con pocas columnas y renglones. La discusión de resultados debe ser completa y exhaustiva, contrastando los resultados obtenidos con la literatura más actual sobre el tema. En esta sección se relacionan los hallazgos más concluyentes de la investigación. Conclusiones En este apartado se relacionan los hallazgos más concluyentes de la investigación, es decir, aquellos que constituyan un aporte significativo para el avance del campo temático explorado, además de un direccionamiento sobre futuras investigaciones. Agradecimientos Si se considera necesario, se agradecen aquellas contribuciones importantes en la concepción, financiación o realización de la investigación: especialistas, firmas comerciales, entidades oficiales o privadas, asociaciones de profesionales y operarios. Literatura citada Para las citas bibliográficas que sustentan las afirmaciones en el texto, se utilizará el sistema [autor(es), año] de forma uniforme; cuando la publicación citada tenga tres o más autores, se debe mencionar el apellido del primer autor acompañado de la expresión latina et al., equivalente a “y otros”, en cursivas y con el año (ej: García et al., 2003). La lista completa con las referencias bibliográficas mencionadas se debe incluir al final del artículo. Los apellidos y nombres de todos los autores deben escribirse en redondas, en el orden alfabético de sus apellidos; cuando se citan varias publicaciones del mismo(s) autor(es) deben listarse en orden cronológico. Se prevén algunos casos: • Para libros: Autor (es), año. Título del libro, edición, casa editora y ciudad de su sede, páginas consultadas (pp. # - #). Ejemplo: Agrios, G. 1996. Fitopatología. Segunda edición. Editorial Limusa, México D.F. • Para capítulos de libros: Autor (es), año. Título del capítulo, páginas consultadas (pp. # - #). En: Apellidos y nombres de los compiladores o editores (eds.)., título del libro, edición, casa editora y ciudad de su sede. Ejemplo: Bernal, H. 1996. Capítulo 6: Evapotranspiración. pp. 112-125. En: Agrios, G. (ed.). Fitopatología. 2a ed. Editorial Limusa, México D.F. • Para revistas: Autor (es), año. Título del artículo, nombre abreviado de la revista volumen(número), páginapágina. Ejemplo: García, S.; W. Clinton; L. Arreaza y R. Thibaud. 2004. Inhibitory effect of flowering and early fruit growth on leaf photosynthesis in mango. Tree Physiol. 24(3), 387-399. • Para citas de internet: Autor (es), año. Título del artículo. En: Nombre de la publicación electrónica del website, portal o página y su URL, fecha de consulta. Ejemplo: Arafat, Y. 1996. Siembra de olivos en el desierto palestino. En: Agricultura Tropical, http://agrotropical.edunet.es; consulta: noviembre de 2003. Vol. 3 - No.1 - 2009 139 Suscripción Revista Colombiana de Ciencias Hortícolas Sociedad Colombiana de Ciencias Hortícolas - Facultad de Agronomía Universidad Nacional de Colombia A.A. 14490, Bogotá-Colombia Teléfono: 571 - 316 5000 Ext. 19041/19043 Tel/fax: 571 - 316 5498 [email protected], [email protected] Nombre / Name Documento de identidad / Identification document Dirección / Address Ciudad / City Teléfono / Phone Departamento / Department Zona postal / Zip code País / Country E-mail Profesión / Profession Institución / Employer Dirección de envío / Mailing address Forma de pago suscriptores nacionales Pago en efectivo o en cheque a nombre de la Sociedad Colombiana de Ciencias Hortícolas o por consignación nacional en el Banco de Bogotá, cuenta corriente 033811761, y envío posterior de copia de recibo y formato de suscripción diligenciado al fax (57+1) 316 5498. Valor para el año 2009 (dos números, incluyendo costos postales para el envío): Colombia: $40.000 (pesos colombianos); América Latina y el Caribe: US$ 30; USA y Canadá: US$ 50; otras regiones y países: US$ 65. Último ejemplar recibido / Last issue mailed. Año / Year Volumen Número Fecha de envío / Mailing date Vol. 3 - No.1 - 2009 Editorial 5 Sección de frutales Climatic change effects on the supply and prices of bulb onion. Case of Duitama municipality, Boyacá (Colombia) Biotechnologies applicable to the development of some Caricaceae species cultivated in the Andean Region: advances and problems 9 Sección de hortalizas Evaluación de dos sistemas de tutorado para el cultivo de la arveja voluble (Pisum sativum L.) en condiciones de la Sabana de Bogotá Evaluation of two training systems for indeterminate pea (Pisum sativum L.) in conditions of the Bogota Plateau Andrés Fernando Forero, Gustavo Adolfo Ligarreto Melhoramento genético de cebola para as condições tropicais e subtropicais do Brasil 18 Mejoramiento genético y producción de “semilla” de ajo (Allium sativum L.). Posibilidades de adaptación a diferentes ambientes Effect of planting density on growth, production and quality of five accessions of hot pepper (Capsicum spp.) in the Colombian Western Amazon Camila Castellanos, Jaime A. Barrera, María Soledad Hernández, Luz Marina Melgarejo, Marcela Carrillo, Ligia Rodríguez, Orlando Martínez Garlic (Allium sativum L.) genetic improvement and seed production. Possibilities of adaptation to variable environments José Luis Burba Raíces y tubérculos tropicales olvidados o subutilizados en Brasil Tropical root and tuber crops forgotten or underexploited in Brazil Marcos Vinícius Bohrer Monteiro Siqueira, Elizabeth Ann Veasey Onion and garlic crops in Colombia: current state and prospects 45 Evaluación de fuentes de fertilización edáfica y el efecto sobre la microbiota presente en un cultivo de cebolla cabezona (Allium cepa L.) 110 Caracterização fisiologica de mudas de Jatropha curcas L. produzidas em diferentes níveis de irradiancia Physiological characterization of Jatropha curcas L. plants grown under different levels of irradiance 56 Fábio Santos Matos, Cleílton Vasconcelos Moreira, Robson Fernando Missio, Luíz Antônio dos Santos Dias 126 Politica editorial e instrucciones a los autores 135 Volumen 3 / No. 1 / Enero-Junio 2009 Evaluation of fertilization sources and the effect on microbiota present in bulb onion (Allium cepa L.) culture Carlos Alberto Luengas–Gómez, Diana Paola Mora–Castro, Dorisana Guerrero–Flórez 95 Sección de otras especies 28 Los cultivos de cebolla y ajo en Colombia: estado del arte y perspectivas Hernán Pinzón R. 81 Efecto de la densidad de plantación sobre crecimiento, producción y calidad en cinco accesiones de aji (Capsicum spp.) cultivadas en la Amazonia occidental colombiana Onion breeding for the tropical and subtropical conditions in Brazil Daniela Lopes Leite, Valter Rodrigues Oliveira, Carlos Antonio Fernandes Santos, Nivaldo Duarte Costa, Maria Esther de Noronha Fonseca, Leonardo Silva Boiteux, Paulo Eduardo de Melo, Ailton Reis, Bernardo Ueno, Mirian Josefina Baptista 69 Juan Carlos Barrientos F., Germán A. Leyva V., Juan Camilo Morales B. Revista Colombiana de Ciencias Hortícolas Biotecnologías aplicables al desarrollo de algunas especies de Caricáceas cultivadas en la región Andina: avances y problemas Miguel Jordán, Diego Vélez, Rosa Armijos Efectos del cambio climático sobre la oferta y precios de cebolla de bulbo. Caso del municipio Duitama, Boyacá (Colombia)