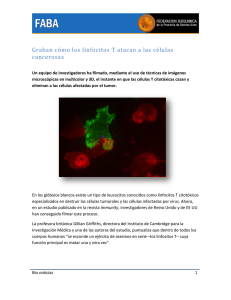
La historia del linfocito en el siglo XX
Anuncio

TESIS DOCTORAL LA HISTORIA DEL LINFOCITO EN EL SIGLO XX Teresa Rinaldi Catalá Departamento Terapéutica Médico-Quirúrgica 2015 TESIS DOCTORAL LA HISTORIA DEL LINFOCITO EN EL SIGLO XX Teresa Rinaldi Catalá Departamento Terapéutica Médico-Quirúrgica Conformidad de los Directores Fdo.: Diego Peral Pacheco Fdo.: Francisca Lourdes Márquez Pérez 2015 “Lucha por tus sueños o los demás te impondrán los suyos” Paulo Coelho Agradecimientos Quiero dejar constancia de la gratitud que siento a todas las personas que han estado presentes en esta etapa de mi vida. En primer lugar, quiero dar las gracias a Dña. Francisca Lourdes Márquez Pérez y D. Diego Peral Pacheco, mis directores de tesis, por su confianza hacia mí en unos difíciles momentos y su apoyo y constante ayuda en la elaboración de la tesis y, sobre todo por su amistad. A mi compañero y amigo D. Félix Conde Martín por su apoyo incondicional y su infatigable ayuda. A Dña. Ana Hidalgo Romero por su empujón y estímulo inicial. A mis familiares. A mi marido por su paciencia y comprensión, a mi hijo por su ayuda en la traducción y apoyo informático, y a mi padre, que aunque ya hace años que no está, supo contagiarme su amor a la lectura. A todas las personas que aunque no aparecen con nombres y apellidos, de alguna forma han hecho posible la realización de esta tesis. ÍNDICE GENERAL RESUMEN……………………………………………………………………………………………… 1 SUMMARY…………………………………………………………………………………………….. 3 1. INTRODUCCIÓN………………………………………………………………………….. 1.1. 7 Notas en la historia temprana del microscopio. La teoría celular y los primeros estudios inmunológicos………………………… 2. OBJETIVOS……………………………………………………………………………………. 9 19 3. MATERIAL Y MÉTODO……………………………………………….………………. 23 4. RESULTADOS……………………………………………………………………………….. 27 4.1. Resultados generales de la historia del linfocito en el siglo XX……………………………………………………..…………………….... 27 4.2. Los primeros treinta años (1900-­‐1930)……………………………………… 28 4.3. La cuarta década del siglo XX…………………...………………………………... 92 4.4. La quinta década (1941-­‐1950)……………………………………………….….. 146 4.5. La sexta década del siglo XX (1951-­‐1960)………………………………….. 204 4.6. La séptima década (los años sesenta)………………………………………… 261 4.7. La octava década del siglo XX (los años setenta)…………………….…… 372 4.8. Las dos últimas décadas del siglo XX………………………………………….. 471 5. DISCUSIÓN…………………………………………………………………………………….. 569 5.1. El linfocito y las tres primeras décadas (1900-­‐1930)………………..…. 569 5.2. La cuarta década del siglo XX (1931-­‐1940)……………………………….... 581 5.3. La quinta década (1941-­‐1950)…………………………………………………... 591 5.4. La sexta década del siglo XX (1951-­‐1960)…………………………………... 600 5.5. La séptima década del siglo XX (1961-­‐1970)…………………………….... 609 5.6. La octava década (1971-­‐1980)……………………………………………….….. 624 5.7. Las dos últimas décadas del siglo XX (1981-­‐2000)……………………... 642 6. CONCLUSIONES……………………………………………………………………………. 665 7. BIBLIOGRAFÍA……………………………………………………………………………… 669 INDICE DE TABLAS Tabla I. Clasificación de tumores y condiciones semejantes a tumores del tejido linfático………………………………………………. 122 Tabla II. Distribución de las muestras por edad y sexo……………………. 343 Tabla III. Enfermedades infecciosas asociadas con linfocitosis atípica periférica……………………………………………………………… Tabla IV. Efecto de la timectomía en la capacidad de transferir memoria………………………………………………………………………….. 377 Tabla V. Reactividad entre suero y linfocitos en el carcinoma de piel…………………………………………………………….. 391 Tabla VI. Movilidad electroforética…………………………………………………. 407 Tabla VII. Talasemia y linfocitos………………………………………………………. 458 Tabla VIII. Tumores y grasas…………………………………………………………….. 487 350 Tabla IX. Anticuerpos monoclonales. ……………………………………………… 503 Tabla X. Frecuencia de las mutaciones del gen RAS en tumores linfoides. …………………………………………………………… 518 Tabla XI. Marcadores fenotípicos de células B…………………………………. 529 ÍNDICE DE GRÁFICOS GRÁFICO 1: Documentos por años 1900/2000 ………………………………….. 27 GRÁFICO 2: Documentos por país/territorio 1900/1910……………………. 28 GRÁFICO 3: Documentos por país/territorio 1931/1940……………………. 93 GRÁFICO 4: Documentos por años 1941/1950 ………………………………….. 147 GRÁFICO 5: Documentos por años 1961/1970 ………………………………….. 262 GRÁFICO 6: Documentos por años 1971/1980 …………………………………... 374 GRÁFICO 7: Documentos por años 1981/2000 …………………………………... 473 GRÁFICO 8: Documentos por materia…………………………………………………. 473 ÍNDICE DE FIGURAS Figura 1. Bordes desiguales en linfocitos……………………………………… 31 Figura 2. Linfocito en reposo. ……………………………………………………... 35 Figura 3. Célula en profase temprana …………………………………………… 35 Figura 4. Célula en profase. …………………………………………………………. 35 Figura 5. Un estadio más avanzado………………………………………………. 35 Figura 6. Dibujo de una etapa posterior. ……………………………………… 35 Figura 7. División completa. ……………………………………………………….. 35 Figura 8. Pequeñas células tímicas. …………………………………………….. 39 Figura 9. Cambios en órganos linfoides después de aplicar pequeñas dosis de rayos X……………………………………………. 58 Figura 10. Migración de linfocitos. ……………………………………………….. 61 Figura 11. Material reticular. ……………………………………………………….. 63 Figura 12. Criterios de Madurez de linfocitos. ……………………………….. 99 Figura 13. Valores del numero de linfocitos según la evolución de la enfermedad. ………………………………………………………… 104 Figura 14. Células plasmáticas………………………………………………………. 109 Figura 15. Porcentaje celular según altitud. …………………………………… 114 Figura 16. Células sanguíneas de 115 conejos jóvenes según los datos recogidos. …………………………………………….. 129 Figura 17. Clasmatocitos………………………………………………………………… 137 Figura 18. Nódulo linfoide solitario de colon…………………………………… 148 Figura 19. Sección vertical del epitelio de una glándula folicular de la lengua de un hombre adulto. ………………………………….. 148 Figura 20. Títulos de anticuerpo y número de linfocitos en el linfoide eferente comparado con el peso del ganglio linfático ………. 152 Figura 21. Diversos tipos de linfocitos. ……………………………………………. 159 Figura 22. Linfocito intraepitelial……………………………………………………. 166 Figura 23. Tipos celulares de ganglios linfáticos………………………………. 175 Figura 24. ARN y título de anticuerpos. ………………………………….. 187 Figura 25. Tipos celulares en la infección por virus X……………………….. 192 Figura 26. Valor del número de linfocitos después de la administración de una suspensión de linfocitos concentrados…………………. 209 Figura 27. Contenido de linfocitos en médula ósea de animales normales y de experimentación……………………………………… 212 Figura 28. Grandes y pequeños linfocitos ……………………………………….. 220 Figura 29. Aparato de reinfusión continua intravenosa de linfa y linfocitos. …………………………………………………………………… 233 Figura 30. Contenido tímico total de sustancia estimulante de linfocitosis (LSS) ………………………………………………………. 237 Figura 31. Porcentaje de neutrófilos, linfocitos y monocitos que fagocitan tinta china ………………………………………………………. 240 Figura 32. Célula cráter. ………………………………………………………………… 243 Figura 33. Emperipolesis………………………………………………………………… 248 Figura 34. Valores medios de linfocitos/hora. …………………………………. 253 Figura 35. Autorradiografía de una suspensión de células del timo de rata. …………………………………………………………….. 265 Figura 36. Linfocitos con inclusiones……………………………………………….. 279 Figura 37. Linfocito transformado de un cultivo de tres días con PHA. 289 Figura 38. Células plasmáticas………………………………………………………… 292 Figura 39. Ratones timectomizados………………………………………………… 298 Figura 40. Ratón timectomizado e injertado. …………………………………… 299 Figura 41. Fenómeno de la adherencia. …………………………………………… 305 Figura 42. La especificidad celular en la reacción al homoinjerto……… 315 Figura 43. Timo y bolsa de Fabricio de un pollo recién nacido………….. 319 Figura 44. Transferencia pasiva. …………………………………………………….. 324 Figura 45. Seguimiento del estado de 59 casos de trasplante renal. …. 335 Figura 46. Movilidad electroforética. ……………………………………………… 340 Figura 47. Volumen. ………………………………………………………………………. 341 Figura 48. Unión de AO al complejo DNP. ………………………………………… 369 Figura 49. Epitelio de las vellosidades del yeyuno humano………………. 383 Figura 50. Nº de linfocitos intraepiteliales por 100 células epiteliales de las vellosidades. ……………………………………….. 384 Figura 51. Linfocito unido a célula tumoral. …………………………………… 393 Figura 52. EMG y miastenia. Electromiograma realizado a ratones dos semanas después de la inmunización………………………… 396 Figura 53. Análisis inmunoelectroforético en la enfermedad de Chagas. 402 Figura 54. Influencia del ácido linoleico en la interacción antígeno-­‐ anticuerpo……………………………………………………………………….. 412 Figura 55. Células formadoras de placas. ………………………………………….. 418 Figura 56. Linfocitos B y T en asma bronquial…………………………………… 426 Figura 57. Reactividad de linfocitos in vitro……………………………………… 434 Figura 58. Pequeños linfocitos al ME………………………………………………… 438 Figura 59. Organización ultraestructural del linfocito……………………….. 440 Figura 60. Malnutrición y células T. ………………………………………………… 447 Figura 61. Células E-­‐rosetas. …………………………………………………………… 451 Figura 62. Linfocitos y trasplantes. …………………………………………………. 454 Figura 63. Diabetes de inicio juvenil y células T. ……………………………… 465 Figura 64. Esclerosis múltiple y ERFC. …………………………………………….. 469 Figura 65. Amígdala teñida con Anti-­‐IgD. ………………………………………… 477 Figura 66. Inmunofluorescencia a actina. ………………………………………… 481 Figura 67. Trasplante y células T. …………………………………………………….. 491 Figura 68. Anticuerpos monoclonales. ……………………………………………… 507 Figura 69. Cinética de la respuesta ISC inducida por antígenos del VIH. 514 Figura 70. Expresión del gen del receptor cannabinoide en tejido linfoide humano. …………………………………………………………… 540 ÍNDICE DE ACRÓNIMOS Y ABREVIATURAS − Å: Unidades angström − AAVS: Liga Contra la Vivisección Americana − ACT: Terapia celular adoptiva. − ACTH: Hormona adrenocorticotropa. − AchR: Receptor de Acetilcolina. − ADN: Ácido desoxiribonucleico. − ADNP: Fosfato de ADN. − AFCP: Células Formadoras de Anticuerpos. − AGL: Ácidos grasos libres. − AHTS: Suero antilinfocito T humano. − AK: Adenilato Kinasa − AL: Ácido Linoléico. − ALG: Globulina antilinfocito. − ALL: Leucemia linfoblástica aguda. − AMM: Asociación Médica Mundial. − AO: Naranja de Acridina. − ARN: Ácido ribonucleico. − ARNr: ARN ribosomal. − ARNm: ARN mensajero. − ARNP: Fosfato de ARN. − AS: Ácido Esteárico. − ATG: Globulina anti-­‐timocito. − ATP: Adenosintrifosfato. − A-­‐9-­‐THC: Tetrahidrocannabinol. − BM: Banco Mundial. − CBMC: Células mononucleares de sangre del cordón umbilical. − CEE: Comunidad Económica Europea. − CFC: Clorofluorocarbonados. − CGG: Triplete Citosina-­‐Guanina-­‐Guanina. − CGH: Hibridación Genómica Comparada. − CMB: Total de linfocitos en sangre. − CMI: Inmunidad mediada por células. − CNIO: Centro Nacional de Investigaciones Oncológicas. − Con A: Concavalina A. − CT: Conducto torácico. − DGLA: Ácido dihomo-­‐gamma-­‐linoléico. − DH: Hipersensibilidad retardada. − DNCB: Dinitroclorobenceno. − DNP: Deoxyribonucleoproteínas. − DZ: Dicigotos. − EAC: Anticuerpos de eritrocitos. − EBV: Virus de Epstein-­‐Barr. − EDTA: Ácido etildiaminotetraacético. − EEUU: Estados Unidos. − EM: Esclerosis Múltiple. − EMG: Electromiograma. − EPM: Movilidad electroforética. − Epon: Resina Epoxy. − ERFC: Células Formadoras de Rosetas Tempranas. − FEDER: Federación Española de Enfermedades Raras. − FISH: Hibridación Fluorescente in Situ. − FJRC: Carcinoma Flexner Jobling de la Rata. − FLA: Frente de Liberación Animal. − FMI: Fondo Monetario Internacional. − FRC: Cáncer relacionado con las grasas. − GATT: Acuerdo General sobre Comercio y Aranceles. − GDP: Guanosin-­‐difosfato. − GLA: Ácido gamma linoléico. − GTP: Guanosin-­‐trifosfato − HER-­‐2: Receptor del factor de crecimiento humano. − HLA: Antígeno Leucocitario Humano. − HLV: Virus Linfotrópico Humano. − HP: Helicobacter Pylori. − HRBC: Eritrocitos de caballo. − HSA: Albúmina de Suero Humano. − IARC: Agencia Internacional para la Investigación del Cáncer. − IFN: Interferón − IgG, IgA, IgM: Inmunoglobulinas G, A y M. − IL12: Interleucina 12. − IL16: Interleucina 16. − INIG: Grupo Internacional de Inmunología Nutricional. − ISCS: Células secretoras de Inmunoglobulinas. − JOD: Diabetes de inicio juvenil. − LAV: Virus asociado a linfadenopatía. − LDF: Factor depresor de linfocitos. − LPS: Lipopolisacáridos. − LR: Reactividad de linfocitos. − LSF: Factor Estimulante de Linfocitosis. − LSS: Sustancia Estimulante de Linfocitosis. − MEM: Movilidad electroforética de macrófagos. − MG: Miastenia gravis. − MGG: May-­‐Grümwald-­‐Giemsa. − MHA: Antígeno del complejo mayor de histocompatibilidad. − MI: Mononucleosis Infecciosa. − M/L: Razón monocito/linfocito. − MO: Médula Ósea. − MZ: Homocigotos. − NCI: Instituto Nacional contra el Cáncer. − NIH: Instituto Nacional de la Salud. − ONU: Organización de Naciones Unidas. − OMS: Organización Mundial de la Salud. − PAS: Ácido Peryódico de Schiff. − PCR: Reacción en Cadena de la Polimerasa. − PFC: Células formadoras de placas. − PGK: Fosfogliceratokinasa. − PHA: Fitohemaglutinina. − PK: Piruvatokinasa. − PPD: Derivado Proteico Purificado. − PSA: Agregados submembranosos polarizados. − PUFA: Ácidos grasos poliinsaturados. − PWM: Hierba carmín. − RAHTG: Globulina de conejo anti-­‐timocitos humanos. − RFC: Células formadoras de rosetas. − RN: Recién Nacido. − SDS-­‐PAGE: Electroforesis en geles de poliacrilamida. − SFC: Síndrome de Fatiga Crónica. − SIDA: Síndrome de Inmunodeficiencia Adquirido. − SKY: Cariotipo espectral multicolor. − SRBC: Eritrocitos de oveja. − SXF: Síndrome del X Frágil. − TCR: Receptor de células T. − TDL: Linfocitos del conducto torácico. − TEC: Número total de eosinófilos. − Th: Células T colaboradoras − Tio-­‐TEPA: N-­‐trietilenetiofosforamide. − Ts: Células T supresoras. − TVP: Trombosis venosa profunda. − UV: Radiación Ultravioleta. − VIH: Virus de la Inmunodeficiencia Humana. − WBC: Valores medios totales de leucocitos en sangre. RESUMEN Introducción. El linfocito como tal surge a principios del siglo XX, gracias a autores como Pappenheimer. Los trabajos que marcan su historia y sus implicaciones a nivel fisiológico y clínico, son objeto de estudio. Objetivos. Estudiar los acontecimientos que rodearon al descubrimiento del linfocito, indagar las fases del conocimiento sobre la célula linfocítica y constatar la valoración clínica (actual) sobre el linfocito. Material y métodos. Una vez verificada la viabilidad del proyecto, la búsqueda inicial de artículos se hizo con la palabra lymphocyt* en Pubmed y Scopus, la de libros se hizo en Google Books, que se completa con la de temas o autores concretos en Pubmed, Scopus ó Dialnet, cuando el desarrollo del tema lo requiere. Resultados. Los estudios en relación al linfocito han ido paralelos al avance técnico. En un principio interesa la morfología linfoide, su respuesta ante diversos estímulos y su papel en la inmunidad tomando como modelo la tuberculosis y el cáncer. El microscopio electrónico permitirá conocer su ultraestructura, y técnicas como la electroforesis y la autorradiografía, facilitarán numerosas investigaciones que ayudarán a una mejor comprensión de la enfermedad. Los cultivos de células linfoides o los anticuerpos monoclonales son algunos de los logros conseguidos. Conclusiones. Los trabajos sobre el linfocito han ayudado a comprender mecanismos fisiológicos y patológicos, a avanzar en la investigación y el abordaje de enfermedades. El linfocito es aún hoy tema de investigación. 1 SUMMARY Introduction. The lymphocyte as such emerged in the early twentieth century, thanks to authors like Pappenheimer. Jobs that marked its history and its implications to physiological and clinical level, are studied. Objectives. Study the events surrounding the discovery of lymphocyte, investigate the phases of knowledge on lymphocyte cell and show the (current) clinical assessment of the lymphocyte. Material and methods. After verifying the feasibility of the project, the initial search was done with the word lymphocyt * in PubMed and Scopus, the book was made in Google Books, complete with specific themes or authors in Pubmed, Scopus or Dialnet when the subject requires development Results. Studies in relation to lymphocyte have gone parallel to technical advance. Initially interested lymphoid morphology, their response to various stimuli and their role in immunity taking as model tuberculosis and cancer. The electron microscope reveal its ultrastructure, and techniques such as electrophoresis and autoradiography, numerous investigations that help facilitate a better understanding of the disease. Lymphoid cell cultures or monoclonal antibodies are some of the achievements. Conclusions. Works about the lymphocyte have helped to understand physiological and pathological mechanisms, to advance research and addressing diseases. Lymphocytes are still the subject of research. 3 1. INTRODUCCIÓN INTRODUCCIÓN Durante los siglos XVI y XVII crece el interés por el conocimiento de la sangre, gracias a los estudios anatómicos de Vesalio, Harvey y de Malpighi, descubridor del flujo sanguíneo capilar. La invención del microscopio da paso a la identificación de hematíes, plaquetas y componentes leucocitarios, iniciándose así en la historia una nueva era, que cambiará el concepto de enfermedad existente hasta entonces y abrirá numerosos caminos en el mundo científico (2)(11). Los leucocitos junto con eritrocitos y plaquetas, son los elementos formes de la sangre. La valoración porcentual de las diferentes subpoblaciones de leucocitos (identificadas fácilmente en las extensiones de sangre teñidas con colorantes apropiados) es conocida como fórmula leucocitaria. Actualmente la fórmula leucocitaria junto al resto de valores sanguíneos (como hemoglobina, hematocrito y volumen corpuscular) constituye el exámen básico de sangre o hemograma. El concepto de hemograma, ha ido evolucionando paralelamente al progreso de la tecnología, en un lento proceso de más de cien años. Hasta la llegada de la automatización en los años 70 del siglo XX, el hemograma fue sinónimo de fórmula leucocitaria, tal como lo definió Schilling en 1933 (1)(2). La historia de la fórmula leucocitaria empieza con el descubrimiento de los métodos de tinción de los tejidos, pero en realidad comienza muchos años antes, siendo necesario el desarrollo de los dos pilares básicos de la hematología: el microscopio y los métodos de tinción celular. Los verdaderos inicios de esta historia se remontan al siglo XVII, cuando tiene lugar el gran auge del comercio textil y con él las técnicas de tinción, y cuando el naturalista holandés Leeuvenhoek observa por primera vez células de la sangre, concretamente eritrocitos, en un rudimentario microscopio. Fueron necesarios cien años más para que el anatomista inglés Wiliam Hewson en 1770 (3,4), pudiera ver los leucocitos y las plaquetas. Pero todavía va a ser necesario otro siglo para que en el año 1891, Paul Ehrlich (5), bacteriólogo alemán especializado en colorantes de tinción celular, analice en detalle la morfología de las células sanguíneas y 7 INTRODUCCIÓN describa la existencia de dos grandes subpoblaciones de leucocitos: granulares y no granulares. Podemos considerar este hecho como el inicio de lo que más tarde se llamaría fórmula leucocitaria. A partir de entonces, hay un rápido avance en el perfeccionamiento de los procedimientos de tinción de las células sanguíneas, y en pocos años se consolidan métodos de tinción universales como el de May-­‐Grunwald-­‐Giemsa, reconocido como el procedimiento de referencia para el estudio morfológico de la sangre y médula ósea (6)(7). Esta generalización de los métodos de tinción va a facilitar en gran medida el estudio de la sangre y su patología, y será la morfología el núcleo principal de una nueva especialidad médica, la hematología. En el desarrollo de esta etapa van a intervenir personajes como Gustav Giemsa, Artur Papperheimer, Wihelm Turk y Alexander Maximov, entre otros. Entre ellos, no podemos olvidar a Arneth (8)(9), quien en el año 1904 estableció una clasificación de granulocitos neutrófilos que va a llamar hemograma y que divide a los granulocitos según sus lobulaciones, colocando los más inmaduros, con menos lobulaciones a la izquierda, y a la derecha los más maduros y lobulados. Esta fórmula leucocitaria se conoce también como hemograma de Arneth, por ser la primera vez que se utiliza el término hemograma. Schilling en 1933, establece un procedimiento de recuento individualizado de las diferentes subpoblaciones leucocitarias, que es conocido como hemograma de Schilling: fórmula leucocitaria realizada con conteo de cien elementos (10). A partir de los años 70, la historia del hemograma y de la fórmula leucocitaria se ve marcadamente influenciada por el gran avance de la automatización, pero hemograma sigue siendo sinónimo de fórmula leucocitaria, tal y como lo definió Schilling. Hoy día, sin embargo, la fórmula leucocitaria es una parte inherente del hemograma. Un hemograma ya no es sinónimo de fórmula leucocitaria, sino que recoge toda la información sobre el estado global de las células de la sangre que aporta un sistema automatizado. El hemograma forma parte del estudio inicial de todo paciente, siendo 8 INTRODUCCIÓN imprescindible en las hemopatías y en el seguimiento evolutivo de muchas enfermedades (1) (10). El reconocimiento de la célula que hoy día se conoce como linfocitos va unido al descubrimiento y perfeccionamiento del microscopio, al desarrollo de la inmunología, y al gran avance que en estos últimos cuarenta años ha tenido el campo de la biología molecular. Para conocer cómo ha ido evolucionando el conocimiento sobre el llamado linfocito, es necesario conocer el marco científico y social en el que surge. 1.1. NOTAS EN LA HISTORIA TEMPRANA DEL MICROSCOPIO. LA TEORÍA CELULAR Y LOS PRIMEROS ESTUDIOS INMUNOLÓGICOS. El microscopio parece surgir en el siglo XVI, siendo muy difícil atribuir su invención a una sola persona (11). Varios siglos antes, esferas de vidrio llenas de agua fueron usadas para aumentar los cortes en piedras preciosas y facilitar la visión de letras pequeñas (12). En el año 1589 un italiano, Gianbattista Della Porta, describió la posibilidad de construir un instrumento para observar seres diminutos, y unos años después, en 1595, Zacharías Janssen construyó un microscopio rudimentario, que Galileo Galilei empezó a utilizar como venía haciendo con éxito con el telescopio y desarrolló un “occhiolino” o microscopio compuesto por una lente convexa y una cóncava (11) (13). El interés por las cosas pequeñas tiene especial desarrollo a finales del siglo XVI y principios del XVII. En 1665, el inglés Robert Hooke creó el primer microscopio compuesto, utilizando dos sistemas de lentes, lentes oculares y lentes objetivos, y publicó el primer tratado de microscopía en el que se describe la célula (3)(13). El holandés Antoni Van Leeuvenhoek fabricó sus propios microscopios, que lo llevaron al descubrimiento de los glóbulos rojos. 9 INTRODUCCIÓN Son conocidos sus estudios pioneros sobre protozoos y ciclos vitales de los insectos, y es considerado por muchos el padre del microscopio simple (14). Ya en el siglo XVII, la microscopía tuvo un rápido avance, entre otros, debido a Marcello Malpighi (15), quien gracias al microscopio puso en evidencia la existencia de los capilares. Malphighi inició el estudio de la estructura de los órganos como hígado, bazo y riñón. Sus descripciones fueron tan exactas, que su nombre ha quedado firmemente ligado a la histología de estos tejidos. También estudió la estructura del gusano de seda, plantas (reconoció su diferenciación sexual y el significado de los anillos de crecimiento), hueso, dientes, cerebro y pulmón. A finales del siglo XIX Pasteur y Koch sustentaron científicamente la etiopatogenia infecciosa de la enfermedad. Rudolf Virchow amplió el campo de interés de la microscopía hacia la anatomía patológica. Así se inician las ciencias de la microbiología, histología y patología celular (18)(27). Comenzaron a aparecer publicaciones de trabajos científicos que se basaban en la utilización del microscopio. Como ejemplo, una de la Revista Médica de la Academia de Medicina y Ciencias Naturales de septiembre de 1873 (16), es la trascripción de la lección oral dictada por el doctor Nicolás Osorio (Colombia) sobre la leucocythemia, la cual se iniciaba en los siguientes términos: “Hace algunos días que un enfermo en el hospital, por su estado caquéctico y color especial, llamó la atención del señor doctor Rengifo, y él manifestó que debíamos examinar la sangre de dicho enfermo, pues probablemente tenía lecocithemia, enfermedad que tiene algunos puntos de contacto con la melania. El microscopio me ofreció un dato precioso para diagnosticar durante la vida la melania palúdica, y en el caso que vamos a ocuparnos, no vacilé en diagnosticar la leucocithemia cuando encontré en la sangre enorme cantidad de glóbulos blancos y solo uno que otro glóbulo rojo”. Virchow (1821-­‐1902) fue pionero en el concepto moderno del proceso patológico al presentar su teoría celular, explicando que las enfermedades 10 INTRODUCCIÓN surgen no en los órganos o tejidos, sino de forma primaria en células individuales. A el se debe el término “omnis cellula ex cellula” (cada célula procede de otra célula ya existente) (17)(18). Los primeros estudios de Virchow giraron en torno a la inflamación de las venas, lo que le llevó a investigar la patología de la serie blanca y a describir por primera vez la leucemia e introducir el término de leucocitosis. Pero Virchow no es el primero en utilizar material histológico como base para el estudio de la patología, cinco años antes, Paget había publicado sus “Lecturas de Patología Quirúrgica”, donde mencionaba abundantes observaciones al microscopio (19)(20). De sus numerosas publicaciones, Virchow (17) es recordado por su “Patología Celular” publicada en 1858. El libro lo constituyen una serie de 20 lecturas entregadas en el recién formado Instituto de Patología de la Universidad de Berlín. Comienza comparando la estructura de las células animales con la de células vegetales, con base en el trabajo de Schwann. Habla sobre los elementos celulares, comenzando por las células rojas de la sangre, el efecto de la lisis que libera “hematina”, que en su forma cristalina es llamada “hematoidina”, reconoce la similitud de las células blancas sanguíneas con las células del pus, discute sobre leucocitosis y leucemia, y describe la diátesis hemorrágica encontrada en casos de leucemia, la cual se asocia a un aumento del bazo y de ganglios linfáticos. Distingue en los ganglios linfáticos, el córtex de la médula y reconoce los folículos, los cuales dice estar especialmente desarrollados en los agrandamientos patológicos de los ganglios. Reconoció que Placas de Peyer, amígdalas, timo y corpúsculos de Malpighio del bazo son parte del sistema linfático. Louis-­‐Antoine Ranvier (21)(22) es considerado como uno de los histopatólogos más destacados de la segunda mitad del siglo XIX. Su trabajo más importante de investigación se centró en el sistema nervioso. Ranvier se adentró también en el estudio de la estructura de los ganglios linfáticos realizando estudios sobre células linfoides in vitro. Mediante canulación del 11 INTRODUCCIÓN conducto torácico estudió las células que obtenía de perros y conejos, describiendo sus características morfológicas y su movimiento ameboide. Demostró que los linfocitos son móviles a 37ºC, pero no por debajo de 20ºC ni por encima de 40ºC, y que el movimiento sólo sucede cuando las células tienen superficie sobre las que apoyarse, pero no cuando se encuentran en suspensión. Describió la diapédesis de los linfocitos, es decir, su habilidad para atravesar las paredes de vasos sanguíneos, observando que muchas células se añadían a la linfa cuando ésta pasa a través del ganglio, hecho que también había descrito Heyfelder en 1852 y Erns Wilhelm von Brucke en 1854. En 1897 Ranvier fundó junto con Balbiani, los “Archivos de Anatomía Microscópica”, la primera revista francesa consagrada a los estudios microscópicos. El “Manual de Histología Patológica” (22)(21) publicado en París en 1869, cuyos autores son Cornil y Ranvier, lleva a cabo un estudio de la Anatomía Patológica General. En su parte primera, se exponen las generalidades sobre la histología normal, y hace referencia a la teoría celular y la constitución de las células. Continúa con el significado de célula según distintos autores; y con una descripción de las células embrionarias, su tamaño y propiedades. Las observaciones de Virchow y Bennet sobre la leucocitemia, son la base para el estudio de los linfoadenomas: “Los ganglios linfáticos, corpúsculos del bazo y folículos del intestino se caracterizan por tejido conjuntivo reticular, donde las mallas se encuentran llenas de células linfáticas”. Se afirma que estas células son pequeñas, redondas y con un solo núcleo. Hace también referencia a la transformación sufrida por el timo con los linfoadenomas: “…con la edad sufre una atrofia casi completa, pero con la leucocitemia toma su forma y adquiere un volumen considerable. Estamos convencidos que se trata de una neoformación, no de ganglios peritraqueales hipertróficos”. Y termina este estudio sobre los linfoadenomas haciendo referencia a su pronóstico: “…sucumben a la progresión, generalmente rápida del mal”. A lo largo de la historia del hombre, y hasta prácticamente el siglo XX, las principales causas de muerte estuvieron vinculadas a las enfermedades 12 INTRODUCCIÓN infecciosas. Así en el siglo XVII, las enfermedades infecciosas suponían el 90% de las causas de muerte, porcentaje que se reduce al 25% en el siglo XX. Fueron las enfermedades infecciosas las que dieron lugar al inicio de la inmunología debido al interés que suscitó el estudio de sus causas, medios de transmisión y métodos de prevención. Y, precisamente una de esas enfermedades, la viruela dio origen a muchos conceptos vigentes en inmunología. Gracias al trabajo llevado a cabo por la O.M.S desde 1966 a 1980, la viruela fue declarada oficialmente erradicada en 1980, gracias a la colaboración de países de todo el mundo. Se considera a James Pylarino (23) como el primer inmunólogo (1659-­‐ 1718). En 1701 Pylarino inocula con el virus de la viruela a los tres hijos de un amigo, con la intención de provocar una enfermedad suave, y evitar un ataque severo de la enfermedad. Gracias a Pylarino y su colaborador Timoni, la variolación fue practicada extensamente en el Imperio Otomano siendo conocida como el “método griego”. Un paso más en el campo de la inmunización, lo dio Edward Jenner a finales del siglo XVIII (24)(25). Sus estudios se centraron en la relación existente entre dos virus, el virus cowpox del ganado vacuno y el virus smallpox de la viruela humana. En 1798, Jenner publica todas sus investigaciones sobre la viruela en un libro que lleva por título “Preguntas sobre las causas y efectos de la vacuna”, donde describe 23 casos que apoyan su teoría. En los dos siguientes años, 1799 y 1800, publica el resultado de otros trabajos, los cuales confirman la original teoría de que la enfermedad de las vacas protege de la viruela (26). Gracias a los trabajos de Pasteur en Francia y de Koch en Alemania, la historia reconocería el trabajo científico de Jenner. En 1885 Pasteur vacunó por primera vez a un humano, Joseph Meister, un niño que había sido mordido por un perro rabioso. Joseph creció y se convirtió en el custodio del Instituto Pasteur. 13 INTRODUCCIÓN Koch es bien conocido por su trabajo con el bacilo de la tuberculosis (bacilo de Koch) (27)(28). Cuando Koch empezó su estudio, ya había evidencia de que la tuberculosis era una enfermedad infecciosa y transmisible en animales de experimentación; pero el organismo causal no se había identificado. Koch establece el ciclo de vida del bacilo y muestra su asociación con la enfermedad y desde sus inicios asegura que la enfermedad humana y animal estaban causadas por el mismo organismo. El nombre de Koch está inevitablemente asociado a los postulados que llevan su nombre (29)(30). Estos postulados dictaban los criterios necesarios para poder hablar de agente causal: • El organismo debe encontrarse en cada uno de los casos de enfermedad. • Debe ser posible aislarlo mediante cultivo. • La reintroducción del microorganismo debe reproducir la enfermedad. Si estos criterios se cumplen debe existir una relación causal entre microorganismo y enfermedad. Elie Metchnikoff, describe la fagocitosis de bacterias por células sanguíneas abogando por la Teoría de la Inmunidad Celular (31)(32)(33). Metchnikoff es el primero en clasificar las células llamadas “macrófagos” (grandes comedores) y “micrófagos” (un tipo de células fagocíticas pequeñas, el leucocito polimorfonuclear). Demostró que ambos tipos celulares juegan un papel importante en la resistencia del huésped contra la infección. Reconoce la relación existente entre las células fagocíticas mononucleares y el bazo, médula ósea y ganglios linfáticos, y fuera de estos órganos, en el tejido conectivo, introduciendo el término “sistema macrofágico”. Metchnikoff definió la inmunidad a enfermedades infecciosas como sigue: “La inmunidad contra las enfermedades infecciosas debe ser entendida como el grupo de fenómenos en virtud del cual un organismo es capaz de resistir el ataque del microorganismo productor de esta enfermedad. La inmunidad puede ser de 14 INTRODUCCIÓN nacimiento o adquirida. Esta última puede ser el resultado de la intervención humana directa como ocurre con la práctica de la vacunación”. Esta definición permanece vigente después de más de cien años (34). Paul Ehrlich logró eliminar los gérmenes causantes de enfermedad sin lesionar el organismo, mediante la inyección de un producto en sangre (balas mágicas), que supuso el primer tratamiento eficaz de la sífilis, y que plasmaba el principio fundamental de la quimioterapia: la toxicidad selectiva. El trabajo de Ehrlich sobre la formación de anticuerpos, apoyaba la teoría humoral de Behring y Kitasato (35)(36)(37). En 1908, Ehrlich compartió con Metchnikoff el Premio Novel, por el reconocimiento de su trabajo sobre el sistema inmune A finales del siglo XIX y primeros años el siglo XX, existían dos grandes tendencias científicas: los que creían que los mecanismos de protección residían en el suero (Teoría de la Inmunidad Humoral), y otros que suponían que la protección frente a patógenos dependía fundamentalmente de células (Teoría de la Inmunidad Celular). Estos dos grupos, en principio, antagónicos y excluyentes, posteriormente reconocieron la existencia de una respuesta inmune tanto humoral como celular y, al demostrarse que la opsonización facilitaba la fagocitosis, que ambas eran altamente interdependientes (31, 35). A principios del siglo XX, la microbiología se había constituido como una disciplina autónoma. Una de sus hijas fue la inmunología, que pronto se sistematizó como una disciplina científica, autónoma y relativamente independiente. Para ello se sucedieron una serie de hallazgos experimentales, como el descubrimiento de la fagocitosis, la toxina diftérica, el poder de los sueros inmunes, descubrimiento del complemento, el intento de conciliación de las dos concepciones de inmunidad, y por último el descubrimiento de la anafilaxia, de cuyo estudio Bordet fue uno de los pioneros. 15 2. OBJETIVOS OBJETIVOS 1. Estudiar los acontecimientos científicos y sociales que rodearon al descubrimiento del linfocito. 2. Averiguar las distintas fases del conocimiento en que se han ido desarrollando los nuevos hallazgos sobre la célula linfocítica. 3. Explorar los distintos campos y patologías donde el linfocito ha estado presente. 4. Constatar la valoración clínica (actual) sobre el linfocito. 19 3. MATERIAL Y MÉTODO MATERIAL Y MÉTODO Se realiza en principio una búsqueda bibliográfica en las bases de datos de Pubmed, Scopus, Web of Science y Teseo con las palabras history y lymphocyt*, a fin de comprobar la no existencia de trabajos similares y verificar la viabilidad del proyecto. La búsqueda inicial de artículos se hizo con la palabra lymphocyt* en las bases de datos Pubmed y Scopus, y para la búsqueda de libros de autores concretos en Google Books. Se completa esta búsqueda con la de temas o autores en Pubmed, Scopus o Dialnet, cuando el desarrollo del tema y/o artículo así lo requieran. Desarrollo de la búsqueda bibliográfica: el trabajo se estructuró con la búsqueda año a año, y el resultado se agrupó en décadas. Conforme avanzábamos en las décadas el número de documentos aumentaba y se hizo necesario seleccionar los artículos a desarrollar. Esta selección se hizo en base a criterios tales como la representatividad del artículo, la originalidad del tema tratado o bien su trascendencia histórica. 23 4. RESULTADOS RESULTADOS 4.1. RESULTADOS GENERALES DE LA HISTORIA DEL LINFOCITO EN EL SIGLO XX. Creemos que es de interés conocer la trascendencia numérica de las publicaciones realizadas a lo largo del siglo XX que guardan relación con el linfocito. La búsqueda en Scopus con la palabra lymphocyt* abarcando todo el siglo XX nos da un total de 558.754 documentos. Observamos en la gráfica como estas publicaciones son muy escasas hasta mediados de la década de los años sesenta, momento a partir del cual la curva adquiere una gran pendiente ascendente. Los cuatro países con mayor número de publicaciones en orden descendente son Estados Unidos, Japón, Reino Unido y Alemania. GRÁFICO 1 27 RESULTADOS 4.2. LOS PRIMEROS TREINTA AÑOS (1900-­‐1930) Si acotamos la búsqueda a las tres primeras décadas encontramos 70 documentos, lo que supone un 0,012 % de todos los documentos publicados en el siglo XX. A diferencia de la tendencia global que sitúa a Estados Unidos como primer país en número de publicaciones, durante estas tres primeras décadas el primer puesto lo ocupa Alemania. Los documentos encontrados en la primera década (1900-­‐1910) son escasos, representando tan solo el 0,0026% del total, sin embargo destacan por su gran interés científico e histórico, al ser trabajos pioneros y punto de partida de posteriores estudios. Se aprecia un claro dominio de la lengua germana, como podemos apreciar en la gráfica acompañante, destacando Pappenheimer como el autor más prolífico. GRÁFICO 2 28 RESULTADOS En la segunda década (1911-­‐1920), al igual que en la primera, los documentos encontrados suponen tan solo el 0,0026% del total. Continúa la supremacía de la lengua germana en el mundo científico, ya que es Alemania, seguida de Austria y Dinamarca los países que más trabajos publican. En la tercera década, aunque el porcentaje de publicaciones con respecto al total registrado en el siglo XX sigue siendo insignificante (0,0071%), la importancia científica e histórica de los trabajos recogidos es indudable. Continúa el predominio de Alemania, pero seguido muy de cerca por Estados Unidos, lo que hace que la lengua inglesa se equipare en volumen de publicaciones a la alemana. Los primeros escritos donde se hace referencia y se estudia la célula linfocitaria, se realizan en lengua germana. Así, recién estrenado el siglo, en 1901, aparece en los Archivos de Anatomía Patológica, Fisiología y Medicina Clínica (mundialmente conocidos como Archivos de Virchow) un trabajo del Dr. A. Pappenheimer (38). En él Pappenheimer reconoce la importancia de los linfocitos, entendiendo el término linfocito como un espacio común para un grupo de leucocitos con una estructura determinada. El linfocito es ahora un término específico, morfológica y funcionalmente, con propiedades tintoriales características. Hace referencia a las diferentes formas morfológicas de los linfocitos, del conocimiento que sobre los mismos existía entonces, de su origen y destino: “… son los constituyentes celulares en los centros germinales de los llamados órganos linfoides. En el tejido conectivo de ganglios linfáticos, intestino, glándulas, bazo y médula ósea, por división mitótica, los grandes linfocitos da lugar a leucocitos, esplenocitos, mielocitos o células megaloblásticas”. En La Revista Europea de Patología, Archivos de Virchow, tienen lugar varias publicaciones en los primeros años de la centuria. En 1902, se recoge por el Dr. Johann Almkvist (39), un trabajo “Sobre la capacidad de emigración de los linfocitos”, donde se realiza una descripción del linfocito, y se señala su capacidad de fagocitosis. Del mismo año y en la misma revista, se encuentra 29 RESULTADOS otro trabajo “Sobre la granulación en los linfocitos” por el Dr. Leonor Michaelis y Dr. Alfred Wolf (40). Un tercer estudio “Sobre células plasmáticas y linfocitos”, tiene por autor a Schlesinger (41). En este último, se recogen las investigaciones realizadas en relación a la morfología de las células plasmáticas por Unna y Marschalko. Mientras la célula de Unna presenta una forma aplanada, ramificada y dotada de movilidad, la célula de Marschalko es de morfología redondeada u oval, con núcleo excéntrico, cromatina en grumos gruesos y halo perinuclear. Las observaciones de Schlesinger difieren de las de Pappenheimer, ya que éste mantiene la no existencia de diferencias en los dos tipos de células, y que las formas diferentes dependen del tejido; si es un tejido seco estaríamos ante la célula de Unna, y si es suculento, las células serían de Marschalko. Según Pappenheimer, las células de Unna y las de Marschalko, son diferente tipos celulares, pero únicamente como diferentes formas de expresión de la misma célula. Autores como H.C. Ward, del Laboratorio del Hospital de Connecticut para la demencia (42) centran su atención, en 1904, en el estudio de “los bordes desiguales de los pequeños linfocitos”. Ehrlich y Lazarus ya habían observado el contorno raído, dentado y desigual de los linfocitos más grandes. Ward utiliza en su trabajo sangre secada en portaobjetos y teñida con los métodos de Jenner y Wright. Reconoce dos variedades de linfocitos, grandes y pequeños, indicando la presencia de formas intermedias que dificultan el reconocimiento de los dos grupos, y realiza la siguiente descripción: “ el pequeño linfocito es ligeramente más grande que un corpúsculo rojo (8-­‐10 micras), y consiste en un núcleo grande, redondo, a veces mostrando mellas, rodeado por una capa muy delgada de citoplasma. La parte central se tiñe más intensamente que el citoplasma, que es más variable en la tinción y a veces presenta gránulos, con una estructura mucho menos definida que el núcleo”. Describe la presencia de procesos citoplasmáticos finos y gruesos, llamados seudópodos, observando que son más numerosos donde se ha aplicado más presión en la extensión sobre el portaobjetos, y da una serie de explicaciones a estos hallazgos: 30 RESULTADOS 1. “…son un artefacto, resultado de la retracción producida por los reactivos de fijación o por la presión ejercida en la extensión”. 2. “…son debidas al estado moribundo de los linfocitos”. 3. “en la sangre fresca también se observa el borde desigual, visto en extensiones secas y teñidas”. Ward intenta buscar un significado fisiológico a este hallazgo, y se pregunta por la posibilidad de que jueguen un papel importante en el proceso de diapédesis y tengan relación con la gran acumulación de linfocitos que tiene lugar en los procesos inflamatorios. Figura 1. Bordes desiguales en linfocitos. Diferentes formas de presentación: apariencia de espinas, como brotes o como prolongaciones muy cortas. Tomado de: Ward, 1904 (42) . Basado en los trabajos de Horbaczewski, el primero en reconocer que la pilocarpina da lugar a una leucocitosis, y los de Ruzicka, que en un largo (15) Dock W. THE LURE OF trabajo llegó al mismo resultado pero no MEDICAL pudo formular ninguna conclusión HISTORY: MALPIGHI AND LEEUWENHOEK: The Early definitiva, W. H. Harvey, en 1906 lleva a cabo estudios sobre Linfocitosis Microscopists. Cal West Med 1928 Nov;29(5):333-334. experimental” (43) con el objetivo de determinar la clase y origen de la leucocitosis. Realizó su trabajo en conejos, gatos Médica y perros, el (16) Revista de lautilizando Academia de Medicina y Ciencias hemocitómetro de Thoma-­‐Zeiss, y la tinción de Jenner. Dividió Naturales. los leucocitos en El microscopio en Colombia. 1873 septiembre 1873;4:35-37. 31 (17) Virchow R. CELLULAR PATHOLOGY. London: J.E.ADLARD; 1859. (18) Virchow R. Archiv für Pathologische Anatomie und RESULTADOS dos grupos, linfocitos y polimorfonucleares, llegando a las siguientes conclusiones: 1. Inmediatamente después de la inyección de pilocarpina, muscarina y cloruro de bario aparece una leucocitosis. 2. Esta leucocitosis es una linfocitosis verdadera. 3. Que es de origen puramente mecánico, debido a contracción del bazo y glándulas linfáticas. 4. Que puede ser inhibida si la atropina precede a la incorporación de cualquier medicina que estimula el tejido muscular. 5. Que la estimulación del nervio esplénico puede producir una linfocitosis similar. Peyton Rous, instructor de patología en la Universidad de Michigan, publica en 1908 dos estudios sobre el comportamiento de los linfocitos. En el primero de ellos (44), investiga el papel de “Algunos factores mecánicos en la producción linfocitaria”. Basa su trabajo en observaciones sobre la fluctuación del contenido de la sangre en linfocitos, y en las investigaciones realizadas con anterioridad por varios autores como Delamere, quien aseguraba: “los linfocitos son los invitados ocasionales de la linfa”. “Se suponen formados en las glándulas de linfa y tejido linfoadenoide, y se considera al conducto torácico como el centro principal de la circulación linfática”. Por su parte Parodi, Crescenzi y Selinoff (45)(46) llegaron a la conclusión de que en perros, la fístula del conducto torácico, causaba una disminución en la cantidad de linfocitos circulantes. Rous (44) estudió el efecto que ocasionaba en perros la realización de una fístula torácica. En todos los animales se observó una disminución gradual de la cantidad de linfa y una concentración celular que varió poco en las primeras horas, pero mostró una tendencia a la disminución progresiva. Las conclusiones a las que llega Rous, son que la linfa del conducto torácico proporciona a la sangre una cantidad de linfocitos mayor que la que se supone en circulación, y que la cantidad de linfocitos suministrado por el conducto torácico del perro sano, permanece prácticamente constante de hora a hora, si las condiciones fisiológicas no cambian. Observa también que la 32 RESULTADOS actividad muscular da lugar a un aumento de la salida de linfocitos por el conducto torácico, que puede ser duplicada o triplicada, con un ejercicio continuado. Rous asegura que ello se debe a la existencia de un fondo de reserva de linfocitos, que rápidamente es cedido a la sangre en ciertas condiciones fisiológicas. Unos meses más tarde, Rous publica un segundo trabajo, también realizado en los Laboratorios de la Universidad de Michigan (47), donde analiza “El efecto de la pilocarpina en la salida de linfocitos a través del conducto torácico”. En 1891, Horbaczewski ya había observado que la pilocarpina aumentaba los glóbulos blancos. Walstein en 1893 empleó pilorcapina como tratamiento en algunas enfermedades infecciosas, teniendo como resultado una linfocitosis absoluta. Este descubrimiento originó muchas teorías, pero en realidad fue poco estudiado. Los perros estudiados por Rous eran anestesiados con morfina para la introducción de una cánula directamente en el conducto torácico, a través de la cual se realizaban los recuentos celulares. Comprobó que el flujo de linfa era influido por factores como la pilocarpina, que producía un aumento, pero también por la respiración disneica inducida por la pilocarpina. La contracción del músculo liso es otro de los factores a tener en cuenta y, según Harvey (43) esta contracción de las glándulas linfáticas y bazo, es el único factor responsable de la linfocitosis. Del análisis de sus resultados, Rous (47) asegura que la inyección intravenosa de pilocarpina en el perro, ocasiona un aumento rápido y considerable en la salida de linfocitos a través del conducto torácico, y que el aumento en la producción celular explica gran parte del cuadro, si no todo, ya que la respiración acelerada y el flujo linfático derivada de ella tienen un papel secundario en la salida de células a la linfa. Dice: “deben existir más factores de los supuestos, no siendo la linfocitosis sólo debida a la migración directa de células desde el bazo, médula ósea o glándulas linfáticas”. 33 RESULTADOS Parte de los estudios durante estos años, están destinados al conocimiento del comportamiento de la célula linfocitaria bajo diversos estímulos. En este sentido realizan sus investigaciones Ross y Macalister en Liverpool (48). En publicaciones previas (49)(50), los autores describen como el plasma de los pacientes de cáncer excita movimientos ameboides en los leucocitos de personas sanas, y el hecho de que los leucocitos pueden ser excitados mediante productos químicos como el azul de metileno y la atropina. En el trabajo, “La flagelación de linfocitos con excitantes artificiales y cancerosos” se presta atención al comportamiento de los linfocitos en personas sanas bajo la presencia de excitantes, ya sean cancerosos o químicos, pero concluyen que no han podido aislar una célula flagelada, ya que sus filamentos se rompen al iniciar su manipulación (48). En 1912, Ross lleva a cabo un estudio comparando la división inducida en los linfocitos por auxetics con el método de Jelly y las figuras mitóticas observadas en secciones de tejido por métodos antiguos (51). Llega a la conclusión de que la división en las células blancas de la sangre no parece depender de alteraciones en la tensión superficial: “Las jaleas se han preparado con diferentes concentraciones de sapolin, glicolato de Na, sodium-­‐tauro-­‐colato y ácido oléico, y ninguno de ellos tiene una acción visible, excepto si la concentración es fuerte, que tiene como resultado una rotura celular. No excitan movimientos ameboides y no provocan un aumento en la división celular”. Afirma, que en el estudio de la citología sanguínea, las especulaciones son algo común, que a veces se aceptan los hechos sin cuestionarlos ni comprobarlos. Nos muestra varias figuras en las etapas de la división del linfocito, comenzando por la célula en reposo ilustra todo el proceso, la profase temprana, y poco a poco hasta la metafase, anafase, telofase y división completa. 34 RESULTADOS Figura 2. Linfocito en reposo. El llamado núcleo, puede verse con la superficie tachonada de gránulos de cromatina. Dentro del núcleo se distingue el nucleolo en forma de anillo. Fig 2 Figura 3. Célula en profase temprana. Ligeramente ovalada y con el nucleolo dividido en dos partes que divergen a los polos. Fig 3 Figura 4. Célula en profase. El núcleo de forma alargada y con los nucleolos en posición de centrosomas. Fig 4 Figura 5. Un estadio más avanzado. Se observa la placa ecuatorial. Fig 5 Figura 6. Dibujo de una etapa posterior. Los núcleos se forman a partir del huso nuclear, por constricción de su centro. Fig 6 Figura 7. División completa. Los cromosomas se han revertido completamente en gránulos, que cubren la superficie del llamado núcleo. El nuevo centrosoma aparece como el nucleolo. Fig. 7 Tomado de: Ross, 1912 (51). Durante varios años (1901-­‐1916), Leo Loeb publica varios trabajos sobre trasplantes y crecimiento de tumores e inmunidad (52)(53)(54). En una de sus primeras publicaciones, en 1902, expone los estudios realizados sobre “Las células linfáticas de la sangre y los procesos inflamatorios del limulus” (55). 35 RESULTADOS Los estudios realizados por P.Geddes (56) y más tarde por Schäfer (57), sobre las células sanguíneas de invertebrados como las estrellas de mar o erizos, sirvieron a Loeb de punto de partida para continuar con esta línea de investigación, ahora en el limulus. El limulus o cangrejo en herradura, es una especie más próxima a los arácnidos que a los cangrejos. Su sangre contiene hemocianina que se colorea de azul con la exposición al aire, y poseen unas células sanguíneas llamadas amebocitos, semejantes a los leucocitos de los vertebrados, que liberan una sustancia coagulante al detectar endotoxinas bacterianas. Las observaciones (56) sobre la coalescencia de las células sanguíneas de invertebrados, sugieren la existencia de una cierta similitud entre el coágulo de los invertebrados y la formación del trombo en los mamíferos, ya que en muchos casos el trombo se inicia como una aglutinación de las plaquetas de la sangre. Estas analogías suponen el estímulo necesario para que Loeb inicie sus investigaciones. Realiza así una serie de experimentos en los que cuerpos extraños como hilos de seda, piezas de gasa, agar o clara de huevo coagulada por calor, son introducidos en el cuerpo del limulus. Seguidamente, en diferentes tiempos, de pocas horas a 11 días, se eliminan las piezas del animal donde se ha introducido el cuerpo extraño y se observan al microscopio. El resultado del estudio del comportamiento de las células del limulus sobre los cuerpos extraños es idéntico en todos los casos, y difiere de la observada en los mamíferos. En un mamífero el implante de un pedazo de gasa o agar, da lugar a una serie de cambios, apreciables a las 7 horas por la presencia de polimorfonucleares que vagan por el agar, y más tarde por la organización del agar a partir del tejido conectivo que lo infiltra a modo de ramificaciones, además de la presencia de células gigantes de distintos aspectos alrededor de los diversos tipos de cuerpos extraños. La reacción en el limulus es muy distinta a ésta, según Loeb: “nunca encontré migración de las células de la sangre del limulus en agar o clara de huevo coagulada, en comparación con la observada en el conejo de indias, ni tampoco hubo formaciones ramificadas. Cinco días después, se puede ver un tejido conectivo bien preservado en contacto 36 RESULTADOS directo con el cuerpo extraño, pero las células no emigraron a su interior”. La reacción usual consistió en la formación de un coágulo alrededor del cuerpo extraño, que finalmente, se necrosa parcialmente, con desaparición de núcleos y células. Loeb (55) habla también de la capacidad del limulus, al igual que otros muchos invertebrados, de regenerar parte de su cuerpo: “... cerca de la hipodermis necrótica no se pudo observar regeneración, aunque el limulus tiene el poder de regenerar las extremidades”, sugiriendo vagamente la posible relación de este hecho con la forma distinta de reacción a los cuerpos extraños: “Las células de mamíferos suelen reaccionar muy fuertemente a los cuerpos extraños, y carecen del poder de regenerar extremidades perdidas, en cambio en el limulus, la reacción es distinta y más débil, pero tienen ese poder de regeneración”. Este comentario que Loeb realiza sin darle mayor importancia pudiera ser lo más interesante de todo este trabajo, y puede convertirse en una inagotable fuente de las mas variadas dudas, preguntas y pensamientos. ¿Quizás esta forma de comportamiento celular esté relacionada con el poder de regeneración? ¿Subyacen en ella diferencias inmunológicas que hacen posible la regeneración en algunos animales? Si entendemos la regeneración como la reactivación del desarrollo para restaurar tejidos perdidos, y sabemos que puede ser a nivel celular, de tejidos o de órganos, y que se ha observado, entre otros animales, en salamandras, estrellas de mar y cangrejos. ¿Tiene la regeneración un origen ancestral genético, que los mamíferos han perdido y los invertebrados no? ¿O serían estas células del limulus células madre totipotentes, a diferencia de las células de mamíferos mucho más diferenciadas? ¿Puede el coágulo servir de soporte para que células madre puedan realizar su función regenerativa? ¿De qué tipo celular estamos hablando cuando nos referimos a células del limulus, son macrófagos, células linfoides, son blastos no definidos? Preguntas y preguntas no contestadas por ahora. Loeb inició sus estudios en el verano de 1901, y en el escrito deja un comentario donde dice: “el final de la temporada no permitió la realización de 37 RESULTADOS todos los experimentos que se habían planeado”. En la actualidad, siglo XXI, sabemos poco más al respecto. Conocemos la importancia del sistema inmunológico para que la salamandra, uno de los animales más estudiados en este sentido, pueda regenerar sus partes afectadas. Los estudios realizados por James Godwin (58) ponen de manifiesto que si no están presentes los macrófagos en las primeras etapas de la recuperación, la regeneración en la salamandra no se produce. Pappenheimer en 1910, siendo patólogo del Hospital de Bellevue en Nueva York, lleva a cabo el “Estudio de la Histología Normal y Patológica del timo” (59) publicado en lengua inglesa, donde realiza un detallado trabajo sobre la histogénesis del timo y sus elementos histológicos: las pequeñas células tímicas, células reticulares y los cuerpos de Hassall. Describe la involución normal del timo y la histología patológica del mismo. Dentro del apartado dedicado a la histogénesis del timo, hace referencia a los estudios de Beard (60), quien sugiere que el linfocito surge en el timo in situ, de repetidas divisiones de células epiteliales, considerando al timo como fuente principal de suministro para los linfocitos fetales. Sin embargo, existen discrepancias al respecto, como la de Ver Eecke (61), quien asegura que los linfocitos surgen en el tejido mesodermal e invaden el epitelial; correspondiendo dichas estructuras epiteliales a los cuerpos de Hassall en los vertebrados superiores. Menciona los trabajos de Bell (62), quien en 1905 también contribuyó al estudio de la histogénesis de las células tímicas en embriones de cerdo, y asegura que las pequeñas células tímicas, aunque de origen epitelial, son verdaderos linfocitos, que en una etapa posterior emigran al tejido circundante. Pappenheimer describe al linfocito. Asegura que las pequeñas células, correspondientes a las supuestas células linfoides: “… tienen un núcleo esférico bien marcado, con abundantes gránulos de cromatina y por lo general un distintivo nucleolo colocado excéntricamente. De citoplasma escaso, que se tiñe claramente con eosina y con frecuencia muestra conexiones citoplasmáticas con 38 RESULTADOS las células más grandes”. Añade que el citoplasma puede no ser distinguible, y que la intensidad de la tinción nuclear varía en células diferentes y con métodos diferentes: “En la mayoría, la sustancia nuclear tiñe intensa y difusamente, de modo que un nucleolo o la membrana nuclear no pueden ser distinguidas”. Figura 8. Pequeñas células tímicas. Escaso citoplasma eosinófilo. (nº3) Tomado de: Pappenheimer, 1910 (59) 39 RESULTADOS Otro de los autores que Pappenheimer cita en el texto, Hammar (63), asegura que las pequeñas células tímicas tienen movimientos ameboides. Hammar también estudió la sensibilidad de las pequeñas células tímicas a la acción de los rayos X: “… una exposición de unas horas es suficiente para causar una desintegración casi completa de estas células; las células grandes reticulares son más resistentes a la Rontgenización”. Cuando Pappenheimer (59) trata a considerar la histología patológica, hace referencia al “estado linfático” o aumento de la glándula tímica. “Dicho estado linfático se asocia en ocasiones a enfermedades endocrinas como enfermedad de Basedow, acromegalia y enfermedad de Addison, llegando a la conclusión de que la característica histológica del timo asociado a enfermedades de glándulas endocrinas son una activa proliferación de los elementos tímicos y ausencia de regresión del mismo”. Según Pappenheimer (59) el timo es un órgano epitelial, compuesto por dos tipos de células epiteliales. Las primeras, designadas como “epitelio reticular” sirven de soporte estructural y son activamente fagocíticas. Los cuerpos de Hassall estarían formados por complejos de células reticulares hipertróficas. Las segundas, las pequeñas células tímicas, son también de origen epitelial, y no son linfocitos. “El timo histogenética e histopatológicamente no tiene relación con las glándulas linfáticas”, asegura Pappenheimer. En este artículo se aprecia claramente los esfuerzos realizados en esta época de la historia por numerosos autores, como los citados por Pappenheimer (entre ellos Beard, Ver Eecke, Bell y Hammar), para determinar el origen del linfocito y el papel desempeñado por el timo. No están definidos los órganos linfoides y está en juego el hecho de que el timo entre a formar parte de los mismos o no, y quizás que uno o varios nombres permanezcan para siempre en la historia. Para ello es fundamental el conocimiento histológico del mismo y el conocimiento del origen de sus dos tipos celulares, células de aspecto epitelial y células de aspecto linfoide. Si es la misma célula con diferente morfología o si son células de distinta etiología es de vital importancia en este momento. 40 RESULTADOS Pappenheimer, identifica los pequeños linfocitos y los describe, pero considera al timo no relacionado con las glándulas linfáticas. Durante muchas décadas, el timo seguirá siendo motivo de estudio. La importancia del timo en el sistema inmune fue descubierta por Jacques Miller (64)(65) en los años sesenta, pero no será hasta la década de los años setenta cuando se empiece a definir y entender su papel. Dorothy Norris, del Instituto Lister en Londres, escribe unas notas en 1914, “Sobre las bases de las fábricas de gas de alquitrán de hulla, que se cree puede ser la causa de predisposición al cáncer en los trabajadores de estas fábricas, en particular su acción sobre los linfocitos, junto a un método para su inactivación” (66). Es conocido el hecho de que el alquitrán de hulla y el hollín causan dermatitis crónicas, verrugas y úlceras, dando lugar a epiteliomas en la piel de los trabajadores en contacto con estas sustancias. En 1910, H.C. Ross y J.W. Cropper (67) propusieron la hipótesis de trabajo que conectaba la aparición de carcinoma y la predisposición al mismo, con sustancias químicas conocidas como auxetics y sustancias cinéticas. Los auxetics (la mayor parte de ellas son cuerpos de aminoácidos) inducen la división de las células blancas de la sangre, tales como linfocitos. Las cinéticas (incluyen la mayoría de alcaloides) dan lugar a movimientos ameboides en las células. Las pruebas las realizaban sometiendo células sanguíneas a estas sustancias auxetics o cinéticas, en películas de gelatina que las contenían. Si estaban presentes auxetics, aparecían figuras de mitosis, si eran sustancias cinéticas, tenían lugar exagerados movimientos ameboides. Ross y Cropper (67) determinaron la fracción activa responsable de la predisposición al cáncer, que corresponde a la que destila entre 260º y 320ºC, es decir la correspondiente al antraceno. Norris (66) que centra su investigación en el papel de las sustancias auxetics, utiliza en sus estudios la metodología empleada por Ross y Cropper, e intenta aislar el agente o agentes responsables del daño producido en los trabajadores de estas fábricas. Como resultado de estos estudios: 41 RESULTADOS 1. Se discuten los métodos para la separación de los constituyentes básicos que estimulan la división celular, de las fábricas de gas de alquitrán de hulla. 2. Se determina que las bases capaces de ejercer esta acción se encuentran en el antraceno del alquitrán de hulla. 3. Dos de estas bases son identificadas como picratos, pero no son satisfactoriamente aisladas. 4. Se describe un método para la inactivación de estas bases de alquitrán. Además de la aportación del trabajo de Norris, al conocimiento del linfocito, la trascendencia de éste y otros trabajos similares en la historia, es doble. En primer lugar, el cáncer es abordado como el efecto de una causa y empieza a ser contemplado y entendido con una nueva perspectiva. La humanidad empieza a dejar atrás la sensación de impotencia que durante siglos ha estado asociada a la palabra cáncer. Un muy breve repaso histórico nos puede dar una idea de su importancia. La descripción más antigua que se conoce de un proceso canceroso tuvo lugar en Egipto (1600 años a C) en el papiro de Edwin Smith (68), un documento médico copia de textos más antiguos. En él se asegura refiriéndose a un caso de cáncer “no existir tratamiento para tal condición”. Mucho más tarde, en el siglo XVI, se inicia la cirugía para el tratamiento de tumores, como lo indican Vesalio, Ambrose Pare, Servet y Johanes Scultetus y a partir de 1715, los oficios de barbero y cirujano se separan, desarrollándose la cirugía experimental y la patología quirúrgica. No podemos dejar de mencionar a Virchow (17), fundador de las bases microscópicas para el estudio patológico del cáncer y quien, en el siglo XIX, más se acerca a una causa etiológica de cáncer con su “teoría de la irritación crónica”. Gracias al trabajo de autores como Ross, Cropper (67) y Norris (66) el cáncer empieza a dejar de ser una enfermedad tabú y maldita, castigo de dioses, y pasa a ser algo con entidad, con una causa palpable, que puede ser evitado y tratado. En segundo lugar, es de suma importancia la atención prestada al estudio de las sustancias cancerígenas. Se inicia un largo camino, 42 RESULTADOS del que todavía sólo hemos visto los primeros metros. El número de sustancias cancerígenas ha ido creciendo desde entonces y, actualmente la Agencia Internacional para la Investigación del Cáncer (IARC) (69) de la OMS, es uno de los organismos de mayor prestigio y autoridad sobre compuestos cancerígenos. Todavía en algunos casos, no se ha conseguido especificar el compuesto responsable; es el caso de la auramina, cromo, cadmio y níquel, entre otros. Respecto al poder cancerígeno del hollín y alquitrán está totalmente demostrado, y se suponen debidos al contenido de hidrocarburos aromáticos policíclicos. Llama la atención el caso del tabaco, no sometido a ningún tipo de regulación, como lo hacen la ropa, los aparatos eléctricos o los alimentos. De las 4000 sustancias descritas en el cigarrillo, al menos 40 de ellas están consideradas cancerígenas para el hombre, entre algunas cabe citar el amoniaco, arsénico, formaldehído, cadmio y alquitrán, considerado el vehículo de todos los demás venenos, sin olvidar la nicotina, sustancia responsable de la adicción. En el año 2003 se prohibió el uso de la creosota, una mezclas de sustancias obtenidas a partir del carbón de hulla y utilizado como conservante de la madera, debido a su potencial cancerígeno, mayor de lo que se pensaba. ¿Cuándo va a ocupar el tabaco el lugar que le corresponde como producto con un alto potencial oncogénico? Pappenheimer que continúa sus estudios en años sucesivos, trabajando en el Departamento de Patología del Colegio de Médicos y Cirujanos de la Universidad de Columbia en Nueva York, publicó en 1914 sobre “los efectos de la extirpación temprana del timo en ratas albinas” (70). Recoge la influencia de la timectomía en órganos como testículo, tiroides y bazo, donde no observa modificaciones. En la sangre, observa un aumento de polimorfonucleares en las primeras semanas después de la intervención, pero los resultado en el recuento linfocitario son contradictorios. Observa que: 1. Las ratas blancas pueden sobrevivir a la extirpación completa del timo al menos 131 días, incluso si se realiza en las primeras semanas antes de la vida. 43 RESULTADOS 2. La timectomía no produce retraso en el crecimiento del cuerpo, ni cambios cuantitativos en el sistema esquelético o dientes. 3. El porcentaje relativo de linfocitos en sangre está disminuido en las primeras semanas después de la intervención, pero no determina cuanto tiempo persiste esta alteración. Junto con Pappenheimer, seguramente es Murphy, el autor que en estos primeros años del siglo XX más se dedica al estudio del linfocito, como podemos apreciar por sus numerosas publicaciones, procedentes la mayor parte de ellas de los laboratorios del hospital de Instituto Rockefeller para la Investigación Médica. En 1914, publica junto con Ellis, los resultados de sus experimentos sobre “El papel del tejido linfático en la resistencia a la tuberculosis experimental en ratones” (71). El papel de la célula linfoide había sido poco estudiado con relación a la tuberculosis, ya que la infiltración de células redondas en el tubérculo se había considerado resultado de la destrucción del tejido y carente de importancia. Sin embargo, algunos médicos clínicos habían señalado la importancia pronóstica de los linfocitos en sangre circulante. Así en la tuberculosis miliar, había una caída de linfocitos en sangre a prácticamente cero, mientras que tenía lugar un incremento en la tuberculosis en estado inicial o en la tuberculosis curada, y en la forma subaguda, los tubérculos contenían grandes cantidades de células linfoides. Los linfocitos son considerados por Murphy factores de resistencia, como lo indican los estudios con rayos X en animales, que provocan una depleción linfoide, facilitando que un tejido injertado, perteneciente a una especie foránea crezca fácilmente. Considera muy significativo, que en dichos animales haya una ausencia total de la infiltración de células redondas sobre los bordes del injerto, siempre presente en animales no radiados (72,73)(74). Sobre el efecto de los rayos X, Heineke (75) demostró la acción destructiva inmediata y específica de los mismos en el sistema linfoide, y que utilizados en pequeñas dosis tenían poco o ningún efecto sobre las otras células del cuerpo. 44 RESULTADOS Murphy (71) al igual que Heineke, encontró que dosis adecuadas y repetidas de rayos X, producen una atrofia gradual del tejido linfoide, sin ningún efecto apreciable sobre los otros tejidos o la salud general del animal. Murphy (71) considera la infección tuberculosa en ratones radiados un excelente método para valorar el papel del linfocito en diversas condiciones. Se realizan varios experimentos con la idea de probar la resistencia de los animales a la tuberculosis cuando se varía la cantidad de tejido linfoide. Los animales tratados con rayos X fueron los primeros en morir a causa de la enfermedad, seguido de los esplenectomizados poco antes de la inoculación (eran considerados con reducción del tejido linfoide). Los animales inoculados 8-­‐10 días después de la esplenectomía, con una proliferación activa de células linfoides en esta etapa, tiene la misma resistencia que los animales normales. Los esplenectomizados tres o más semanas antes de la inoculación, sobreviven a los animales normales en unos días, debido probablemente a un aumento de actividad de agentes defensivos. Los trabajos de Lewis y Margot, publicados en 1914 (76) tuvieron resultados similares. Observaron que las ratas y ratones afectados experimentalmente por tuberculosis desarrollan grandes bazos, y que los animales esplenectomizados vivían más tiempo, después de la inoculación a la tuberculosis, que los animales normales. Ante este resultado difícil de explicar, señalan que las inoculaciones nunca se realizaban antes de 2 semanas después de la esplenectomía y que durante este periodo de tiempo, el tejido linfoide del cuerpo sufría una hipertrofia, mostrando muchos animales un aumento de linfocitos circulantes con respecto a animales normales. La pregunta que se plantea Murphy, motivo de discusión en este trabajo, es, si estos tratamientos causan variaciones en otros factores distintos a los linfocitos, que podrían desempeñar un papel en la resistencia a la tuberculosis. Los resultados de estos experimentos sugieren fuertemente que los linfocitos juegan un importante papel en la resistencia del animal a la tuberculosis. 45 RESULTADOS Murphy y Morton, presentan en 1915 procedente del Instituto Rockefeller, un trabajo titulado “El linfocito, un factor en la resistencia natural e inducida al cáncer trasplantado” (77). Descubren una gran semejanza entre una serie de fenómenos que tienen lugar alrededor de un injerto de tejido de otra especie y un injerto canceroso homólogo en un animal con una inmunidad natural o inducida a cáncer trasplantado. Un hallazgo constante es una reacción local linfoide que aparece tempranamente en el proceso, y dura hasta la destrucción completa del injerto. Hablan de dos tipos de inmunidad: la natural, existente normalmente en un determinado porcentaje de animales inoculados y la inducida, obtenida por un tratamiento previo de los animales con inoculaciones de tejidos homólogos vivos. Se estudia la circulación de linfocitos en estos dos tipos de inmunidad. Con el fin de determinar la importancia de la reacción linfoide en el proceso de inmunidad, se destruye la mayor parte de las células linfoides en ratones con uno de los dos tipos de inmunidad. Se utilizaron rayos X, resultando una destrucción completa de la inmunidad y un crecimiento más rápido de los cánceres en estos animales que en animales normales. Demuestran que, después de la inoculación de cáncer en ratones con inmunidad natural o inducida, tiene lugar un marcado aumento de linfocitos en la circulación. Cuando esta reacción linfoide es impedida, por una destrucción previa del tejido linfoide con rayos X, el estado inmune es destruido. De la observación de los resultados, Murphy y Morton (77) concluyen que: “el linfocito es un factor necesario para la inmunidad contra el cáncer”. En 1917 Pappenheimer (78), en sus “Estudios Experimentales sobre linfocitos”, recoge observaciones sobre la acción del suero inmune en linfocitos y pequeñas células tímicas. Hace referencia a los trabajos de Metchnikoff y de Besredka quien encontró que las propiedades tóxicas de la leucotoxina eran destruidas por calor a 55ºC durante 30 minutos. Pappenheimer, preparó suero citotóxico inyectando a conejos suspensiones de células lavadas de timo de rata o de linfocitos de amígdalas. Asegura que es fácil producir suero con actividad leucotóxica tanto in vitro como in vivo, y que la especificidad de estos 46 RESULTADOS sueros leucotóxicos, lo son para el tipo particular de célula usada para la formación del mismo. Sin embargo, reconoce que la especificidad de estos sueros para un tipo particular de célula, utilizada como antígeno, es cuestionable, ya que los métodos utilizados podrían ocultar posibles errores inevitables. “… incluso un cuidadoso lavado de un órgano o suspensión celular no garantiza la total ausencia de sangre. No es de extrañar que el suero sea moderadamente hemolítico y aglutinante” afirma Pappenheimer, y finalmente añade: “Las pruebas presentadas indican que las pequeñas células tímicas están biológicamente relacionadas, si no son idénticas, a los linfocitos de las glándulas linfáticas”. En 1917 se recoge una segunda publicación de Pappenheimer: “Reacción de los linfocitos bajo varias condiciones experimentales” (79). Asegura que el hambre, la inanición crónica, las infecciones o determinadas lesiones como aquellas causadas por rayos X, causan una destrucción repentina o gradual de las pequeñas células tímicas. Reconoce la falta de conocimiento que existe sobre el tema: “Si se compara con los estudios existentes sobre el comportamiento biológico de los glóbulos rojos, nuestro conocimiento de las células tímicas y, la influencia de las condiciones ambientales en ellos es pobre”. Utiliza una técnica sistemática para ver el comportamiento de las células del timo, bajo una amplia gama de condiciones experimentales: • El efecto de soluciones hipotónicas-­‐hipertónicas. • Las variaciones en la concentración del ión hidrógeno. • El efecto de la temperatura. • La asfixia. • La acción protectora del suero y otros coloides como la gelatina, albúmina de huevo, goma arábica y emulsión de colesterol. • El efecto de ciertas sustancias fotodinámicas como hematoporfirina y clorofila. • El efecto de la exposición a rayos X sobre la capacidad tintorial del linfocito. 47 RESULTADOS • La comparación de los linfocitos en animales jóvenes y viejos. • El efecto de la inanición sobre la capacidad tintorial de los linfocitos. • El efecto tóxico de sueros heterólogos y de sueros citotóxicos específicos. Hace una valoración de la difusión del colorante trypa azul en el núcleo, como criterio de lesión celular, y cuantifica así el efecto de todos estos factores sobre las células tímicas. Consideramos interesante hacer mención al pensamiento científico que predomina en este momento histórico, y que inevitablemente va a influir en el trabajo de todos estos investigadores de primeros años del siglo XX. Podemos definir el método científico como una serie ordenada de procedimientos de que hace uso la investigación científica para observar la extensión de nuestros conocimientos. Fueron los antiguos griegos (80) los que dieron forma a lo que hoy se conoce como ciencia teórica racional. Tales de Mileto (500 a C) fue el primero en negarse a aceptar las explicaciones sobrenaturales, religiosas o mitológicas de los fenómenos naturales, proclamando que: “todo proceso tiene una causa natural”. El método científico (81) ha sido objeto de intensos debate a lo largo de la historia, entre los principales pensadores que han contribuido a darle forma, hay que destacar, entre otros, a Aristóteles con su razonamiento deductivo, a Galileo Galiei, quien presentó su tratado con demostraciones matemáticas, algo innovador en términos de método científico, Isaac Newton que describió las cuatro reglas del razonamiento, sentando las bases del método inductivo, y Claude Bernard (82), quien a mediados del siglo XIX en su “Introducción al estudio de la medicina experimental” discute sobre las bases que hace que una teoría sea buena y un científico un verdadero descubridor. Es importante saber que ningún método es infalible para el conocimiento y que todos constituyen una propuesta racional para llegar a su obtención. El primer paso en el método científico empírico es la observación cuidadosa de un fenómeno y la descripción de los hechos que tratan de explicar las hipótesis. 48 RESULTADOS Solamente las ideas que pueden ser comprobadas experimentalmente están dentro del ámbito de la ciencia. El proceso experimental comienza siempre con una pregunta. En el caso que nos ocupa podría ser: ¿Qué tipos celulares hemos encontrado? ¿Cual de ellos se corresponde con el linfocito? ¿Cómo se comporta ante diversos estímulos? ¿Cómo podemos reconocerlo? ¿Qué función tiene? ¿Cuál es su vida media? ¿Dónde se destruyen los linfocitos? Peguntas sencillas y básicas que inician un proceso experimental observacional en principio, a veces seguido de sencillas hipótesis de trabajo. Esta es la base de trabajo de los autores aquí presentados. Pappenheimer que basado primero en la observación, continúa con estudios experimentales, lo mismo que Harvey con su linfocitosis experimental, o que Murphy, Rous, Ross y Macalister, o Norris, por citar algunos. Formulan una hipótesis de trabajo, que una vez analizados los resultados será aceptada, rechazada, modificada o incluso podrá dar lugar a nuevas hipótesis. El método científico se basa en dos pilares fundamentales: 1. Reproducibilidad. La mayoría de los trabajos, se basan en experiencias previas, derivan de otros anteriores, que serán ratificados, refutados o ampliados. Para que esto sea posible se busca la publicidad de los resultados obtenidos, generalmente en forma de artículo científico. 2. Refutabilidad. Todo estudio científico tiene que ser susceptible de ser rechazado o aceptado. Mediante la discusión, el trabajo es objeto de crítica interna por el propio autor y comparado con los resultados de otros autores (81, 82). Una de las características fundamentales del siglo XX, es la preocupación por el método científico. A finales del siglo XIX, el modelo de Newton entra en crisis, y se replantea una vez más la cuestión del método en la ciencia, apareciendo un cierto desorden en la búsqueda de siempre: teorizar sobre la realidad (83). 49 RESULTADOS En 1917, Taylor y Murphy (84) realiza nuevos estudios para determinar “El papel del tejido linfoide en la resistencia a la tuberculosis experimental en el ratón”. Ahora lleva a cabo sus estudios en ratones, con la idea de establecer una relación entre el cáncer y la tuberculosis, ya que a priori, no existen razones suficientes para tal suposición. Existe la creencia del antagonismo específico entre las dos enfermedades, pero hay que considerar a este respecto la distinta edad de presentación de las dos afecciones, y tener en cuenta que ambas pueden ocurrir simultáneamente en el mismo individuo, por lo que esta idea del antagonismo específico tiene poca base científica. La presente comunicación trata de la influencia de una linfocitosis pronunciada inducida por otros medios que no sea la esplenectomía, sobre la resistencia del ratón a la infección tuberculosa. Ya es sabido que los ratones inmunizados y luego inoculados con cáncer desarrollan una reacción linfoide marcada en sangre que dura varias semanas. Las indicaciones de que la célula linfoide desempeña un papel en la resistencia a la tuberculosis son numerosas. Los recuentos de linfocitos en tuberculosos muestran variaciones interesantes. Un número normal de células linfoides en un animal adulto puede variar entre 1500 y 2500 por ml. En la tuberculosis miliar hay una disminución notoria, y el número de linfocitos varía directamente con el grado de resistencia a la infección tuberculosa. Como resultado de este experimento se obtienen las siguientes conclusiones: 1. Los ratones sometidos a rayos X tienen reducido su tejido linfoide en gran medida y son altamente susceptibles a la infección tuberculosa. 2. Cuando se induce una marcada linfocitosis en ratones, y posteriormente son inoculados con cáncer, la resistencia a la infección tuberculosa es mucho mayor que en ratones normales. 50 RESULTADOS 3. Esta mayor resistencia puede disminuir, incluso virar a un estado de mayor susceptibilidad a la infección por agotamiento de los linfocitos debido a rayos X. En 1918, Loeb publica los resultados de un estudio sobre “múltiples trasplantes de la glándula tiroides y la reacción linfocítica” (85). Hace referencia al trabajo de Hesselberg sobre trasplantes sucesivos de tiroides en el mismo huésped. Loeb observa lo siguiente: • Los primeros trasplantes, difieren según sea el animal del que se ha obtenido. Los animales relacionados parecen tener mejores resultados. • En el trasplante de tiroides múltiple, los dos lóbulos del tiroides derivados del mismo donante se comportan de igual forma y difieren de los tiroides derivados de otros donantes y trasplantados en el mismo hospedador. • La fuerza de la reacción linfocítica varía, pudiendo ser muy marcada en unos tiroides y casi ausente en otros. Considera que la reacción linfocitaria es una respuesta localizada a las toxinas, que se forman independientemente de las sustancias inmunes. • Que en ciertas condiciones, tiene lugar la formación de cuerpos inmunes, cuyo efecto puede ser añadido al de las toxinas. • Que es probable que el componente celular que directa o indirectamente da lugar a la formación original de toxinas, también pueda actuar para provocar la formación de sustancias inmunes. En definitiva, Loeb (85) reconoce que los factores experimentales son complejos y los resultados no son decisivos, pero muestran que tanto el donante como el receptor son importantes para el resultado del trasplante, así como la relación que exista entre ellos. En 1918, Murphy (86) continúa sus experimentos iniciados en 1915 sobre la importancia del linfocito en el cáncer trasplantado. Esta vez junto a Taylor 51 RESULTADOS estudia el efecto de los rayos X sobre la inmunidad inducida artificialmente. Ahora, los ratones son inmunizados por una inyección de sangre homóloga bajo la piel de la espalda, y diez días más tarde, se inocula con tumor (Adenocarcinoma Bashford nº 63) en la ingle izquierda de cada animal. Se utiliza como control una serie de ratones no inmunizados pero inoculados con tumor al mismo tiempo que los experimentales. Después de tres semanas, los animales inmunes se dividen en dos grupos, siendo sometido uno de los grupos a pequeñas dosis repetidas de rayos X. Una semana más tarde, ambos grupos fueron inoculados de nuevo en la ingle derecha, con el mismo tumor. También se inocularon los ratones normales. La dosis de rayos X era la usada para destruir la mayor parte de tejido linfoide (determinada en experimentos anteriores), sin afectar a la salud general del animal. Se obtuvieron los siguientes resultados: 1. Entre los animales inmunizados artificialmente, que no recibieron ningún tratamiento de rayos X, desarrollan tumor un 21%. 2. El segundo grupo de ratones, que fue radiado antes de la segunda inoculación, mostró un grado menor de inmunidad, desarrollando tumores el 52,4%. 3. De los animales que formaban el grupo control, el 90,9% desarrolló tumor Por tanto, los ratones inmunizados artificialmente contra un cáncer trasplantable, inoculados y demostrados inmunes, pueden ser convertidos en susceptibles al mismo tumor por la exposición a rayos X. Los animales inmunes no radiados, demostraron en gran medida su resistencia a una segunda inoculación del tumor en cuestión. Estos experimentos indican que los linfocitos son un factor potente de inmunidad al cáncer en el ratón. Las observaciones realizadas por Murphy (77)(86) son apoyadas por las de Loeb (85) sobre el papel del linfocito con respecto a los injertos de tejido homólogo. 52 RESULTADOS Sobre la importancia del linfocito en la inmunidad contra el cáncer también trabaja Sittenfield, quien en 1918 publica los resultados de sus experimentos realizados en el Departamento de Patología de la Universidad de Columbia (87). Sittenfield, inocula en ratas, a las que previamente había inducido inmunidad, el carcinoma Flexner Jobling de la rata (F.J.R.C). Sus resultados contrarios a los observados por Murphy y Morton, parecían concluir que el grado de linfocitosis tenía muy poco que ver con la inmunidad al F.J.R.C, y decidió realizar nuevos estudios y repetir el experimento que Murphy había realizado en ratones, ya que estaba en juego un factor importante implicado en la inmunidad al cáncer. Inoculó F.J.R.C en 89 ratas, y posteriormente extirpó el tumor completamente en varios intervalos de tiempo que van de los 10 a los 28 días. Mientras el tumor inoculado crecía, una dosis estimulante de rayos X fue administrada, inoculando de nuevo al animal con el propio tumor en la otra axila. Se hicieron recuentos antes y después de la radiación, pero se observó el crecimiento del segundo tumor. En 47 ratas se realizó la extirpación incompleta del tumor deliberadamente y se administró la misma dosis de radiación, se inoculó por segunda vez en la otra axila, observándose su crecimiento en ésta y la recurrencia en un 48% de los casos en el lugar de la intervención. Estos resultados parecen concluir que el grado de linfocitosis tenía un escaso papel en la inmunidad al F.J.R.C. Cuando el experimento se repitió en ratones, con el tumor suministrado por el Dr. Murphy, los resultados obtenidos eran muy similares a los aportados por Murphy y Morton (77) en su comunicación. No obstante, Woglom (conocido en la sociedad científica de la época por sus trabajos sobre cáncer e inmunidad) (88)(89), en relación con este tema, divulgó lo siguiente: “El cuadro histológico en los nodos linfáticos en ratas inmunes no se diferencia del de ratas normales. El comportamiento en el crecimiento del tumor en ratas blancas inoculadas con F.J.R.C no fue afectado lo más mínimo por la alta tasa de linfocitos, y por tanto el agente que causa algún grado de inmunidad en ratones y falla totalmente en ratas debe ser insignificante”. 53 RESULTADOS Sittenfield (87) basa su trabajo en las experiencias previas de otros autores, que intenta ratificar y ampliar utilizando ratas. La reproducibilidad y la refutabilidad, los dos pilares básicos del método científico, se aprecian aquí claramente. Finalmente, el trabajo, vista la crítica tanto interna como externa por otros autores, será rechazado o aceptado. Sin embargo, el mayor enemigo, en el avance científico a lo largo de toda la historia, ha estado representado por el propio mundo científico. En palabras de López Piñero (88): “Las novedades médicas, químicas o biológicas, encontraron una barrera que dependía casi exclusivamente de un simple proceso de inercia social…ceder ante las novedades les hubiera significado, en efecto, o un esfuerzo de asimilación que difícilmente podrían realizar desde su situación y su ambiente, o verse desplazados de sus cargos y sus puestos por los miembros de una nueva generación que había sabido ponerse al día. Por ello, la lucha en torno a las nuevas ideas, la polémica entre “antiguos” y “modernos” fue en gran parte un encuentro entre generaciones”. En este mismo año (1918), se publica otro de los trabajos de Murphy y Sturm (89), bajo el título: “Efecto del calor seco sobre el recuento sanguíneo en animales. Estudio sobre actividad linfoide”. Muchos métodos se habían probado para producir un aumento intenso y duradero de linfocitos y tejido linfoide: drogas y tintes, luz ultravioleta y luz solar. Excepto el empleo de la luz, todos los intentos habían sido insatisfactorios. Un método prometedor era el empleo de calor seco intenso. La fuente de calor en estos experimentos era un bulbo eléctrico de cristal, variando el método de aplicación según el tamaño del animal y su capacidad para soportar altas temperaturas. Para ratones la temperatura debería oscilar entre 55-­‐65ºC, nunca sobrepasando éste límite. Para animales más grandes como ratas y conejos, más susceptibles a altas temperaturas, 60ºC era una temperatura ya no soportada, pero el tiempo de exposición podía ser mayor en estos animales. Después de someter a los ratones durante 5 minutos a la fuente de calor, se observó una caída tanto de polimorfonucleares como linfocitos. El leucocito 54 RESULTADOS polimorfonuclear se recuperó despacio, mientras los linfocitos aumentan rápidamente antes de la segunda semana después de la calefacción. Son linfocitos sanos y normales, y no se diferencian aparentemente de aquellos del animal normal. Los linfocitos circulantes después del calentamiento presentan un número elevado de figuras de división amitótica (división de núcleo sin que se hagan patentes los cromosomas). Las observaciones fueron hechas sobre ratones, ratas y conejos de indias. Los animales muestran después del calor seco una caída repentina en el recuento total de glóbulos blancos. Después tiene lugar una recuperación lenta por parte de polimorfonucleares, de varias semanas para llegar a valores normales. Los linfocitos se elevan rápidamente después de la caída inicial, y siguen elevándose 2-­‐3 semanas, a menudo con valores de 200-­‐300% sobre cifras normales. Murphy y Sturm (89) manifiestan la imposibilidad de asegurar de dónde proceden estos linfocitos, dando como probable un origen en los centros linfoides estimulados. Añaden la no existencia de datos suficientes en la literatura, para determinar variación estacional, efecto de la altitud o variaciones climáticas, para establecer una relación entre estos resultados y los observados en el hombre. Y señalan la utilidad de este método en futuras investigaciones para producir un aumento marcado y duradero de linfocitos circulantes, dando así una oportunidad para el estudio de la función de estas células. El “Papel del calor como inductor de linfocitosis”, es uno de los estudios de Waro Nakahara en 1919 (90), quien trabajó con Murphy varios años en los Laboratorios del Instituto Rockefeller, publicando juntos numerosos artículos. Basado en las observaciones de Murphy y Sturm (89) sobre el efecto del calor seco, Nakahara utiliza el mismo material empleado por éstos. Los ratones, que eran de la misma población y de la misma edad, se sometieron a calor seco durante 5 minutos a una temperatura entre 55 y 65ºC, siendo sacrificados en 55 RESULTADOS grupos, a intervalos de 2, 4, 6, 8, 12 y 14 días, a fin de estudiar los ganglios linfáticos, bazo y otros órganos, los cuales eran fijados en el líquido de Bouin, de Zenker o de Flemming, antes de su estudio histológico. Una de las observaciones más sorprendentes era el paralelismo encontrado en los cambios celulares en el bazo y ganglios linfáticos después del tratamiento con calor. Lo primero que ocurre es una degeneración generalizada de los elementos celulares por acción del calor (fase destructiva), seguida a las 48 horas, de una regeneración caracterizada por un aumento de figuras mitóticas (fase de compensación). Sobre estas bases, Nakahara (90) concluye: “... parece evidente que la linfocitosis marcada inducida por medio de calor se debe, al menos en parte, a la actividad mejorada de centros germinales de bazo y ganglios linfáticos, en respuesta a la acción destructiva sobre las células linfoides”. Otro estudio realizado en el Instituto Rockefeller, y publicado en 1919, tiene por título “El efecto estimulante de los rayos X sobre el linfocito” (91). Taylor, Witherbee y Murphy, han constatado la acción destructiva de los rayos X en linfocitos circulantes (92). “Dosis grandes eran destructivas, en cambio, pequeñas dosis servían de estímulo”, afirmaba Murphy (86). Con la introducción de los nuevos tubos Coolidge, las dificultades encontradas con los viejos tubos de rayos X eran eliminadas, y existía la oportunidad de comprobar esta observación y ampliarla. Se utilizaron nueve conejos del mismo tamaño, y dosis de rayos X de baja penetración bajo unas determinadas condiciones de intensidad, distancia al objeto y tiempo de exposición, que fue fijado en 20 minutos. En siete de los nueve animales, se observó un aumento de linfocitos circulantes. De éstos, en cinco el aumento fue marcado, y en los otros dos definido, pero no tan grande. De los animales que no mostraron ningún estímulo, uno presentó una fluctuación marcada en el conteo antes y después de la radiación y el otro, poco o ningún cambio. Los autores manifiestan (91):“Los resultados obtenidos en esta pequeña serie de animales, no puede ser aceptada como prueba concluyente, pero su interés principal radica en su paralelismo con los estudios histológicos”. 56 RESULTADOS Los cambios que se producen en los órganos linfoides después de la aplicación de pequeñas dosis de radiación es estudiado por Nakahara, que publica sus resultados en 1919 (93). Emplea conejos normales del mismo color y aproximadamente de la misma edad, tratados con las mismas condiciones y dosis de radiación que utilizaron Thomas, Taylor y Witherbee (intensidad, distancia y tiempo de 20 minutos) (91). Los animales fueron sacrificados a distintos intervalos de tiempo para el estudio histológico de bazo y ganglios linfáticos. Los resultados obtenidos, bien por aplicación de calor o rayos X son indistinguibles, ya que los cambios en los linfocitos son idénticos en ambos casos, parece pues probable que el fenómeno de la linfocitosis sea de la misma naturaleza, independientemente si el agente usado es calor o radiación. En el bazo, se observó una mejora de la actividad proliferativa celular que alcanzó su máximo a los cuatro días después del tratamiento, persistiendo hasta los 14 días. En los ganglios linfáticos, el cambio estimulante es manifestado claramente antes, a los 2 días, y es más extenso que en bazo. El aumento de linfocitos se hace más pronunciado a la semana del tratamiento, y ello se relaciona con los hallazgos histológicos. Las conclusiones a las que llega Nakahara (93) son: 1. Las pequeñas dosis de rayos X aplicadas a conejos no tienen ningún efecto destructivo apreciable sobre tejido linfoide. 2. Los indicadores del estímulo en tejido linfoide aparecen inmediatamente, se hacen más pronunciados a los dos días en las glándulas de linfa y a los cuatro en el bazo, y persisten en un grado más leve hasta el día catorce. 3. Estos hechos sugieren que la linfocitosis inducida por pequeñas dosis de rayos X, es debido a un efecto estimulante primario sobre el tejido linfoide del animal. 57 RESULTADOS Figura 9. Cambios en órganos linfoides después de aplicar pequeñas dosis de rayos X. Nódulo esplénico 4 días después del tratamiento. (M representa las figuras mitóticas) Tomado de: Nakahara, 1919 (93). También el sol es estudiado como fuente de radiación capaz de influir en las células sanguíneas. En este sentido debe ser considerado el trabajo de Taylor en 1919 (94). Taylor, inicia su estudio en la primavera de 1916 realizando una serie de recuentos sanguíneos sobre varios individuos, antes y después de ser expuestos al sol durante los meses siguientes de verano en Massachusetts. La observación se hizo en personas de ambos sexos, entre 20 y 65 años, que mostraban pruebas de dermatitis solar crónica (curtido o bronceado) en la mayor parte de su cuerpo, no sólo en cara y manos, y se admitió que el bronceado era debido a los cambios actínicos y no al calor. De los resultados registrados, parece que los rayos del espectro solar causan cambios similares que los debidos a rayos X, y que la forma de acción es igualmente desconocida en ambos agentes. En conexión con este estudio, 58 RESULTADOS Taylor nombra los realizados por Chamberlain y Vedder en 1911(95) sobre la población americana que vive en Filipinas, y conocer así el efecto que el clima tropical tiene sobre la sangre. Los resultados de sus conteos muestran un porcentaje más alto de linfocitos que lo considerado normal en individuos de zonas templadas. Otro estudio realizado por Wickline en 1908 (96) sobre 104 soldados, a 3, 12 y 19 meses después de la llegada a Filipinas, mostró un aumento definido y progresivo de células mononucleares, siendo el porcentaje de los pequeños linfocitos de 21,8%, 26,6% y 33,3% respectivamente. Wickline considera que este efecto sobre la sangre parece estar relacionado con la luz solar, pero es imposible excluir en estas observaciones el efecto del infrarrojo en olas de calor En el estudio de Taylor (94), la dermatitis crónica solar fue acompañada en 25 de los 38 individuos estudiados, de un apreciable aumento en el porcentaje y número absoluto de linfocitos circulantes. En 8 se encontró una clara disminución y en 5 no había ningún cambio apreciable después de la exposición prolongada a los rayos del sol. De los 13 sujetos que no aumentaron sus linfocitos en sangre, 6 no presentaban bronceado, 3 eran de piel tan oscura que determinar la existencia de bronceado era imposible, y 5 tenían un alto número de linfocitos en el conteo del primer análisis. Se sugiere pues, que las personas de piel blanca que viven en Filipinas aumentan su número de linfocitos circulantes después de un periodo prolongado de residencia en zona tropical. Basado en el paralelismo existente entre el bronceado y los cambios en la sangre, Taylor dice: “... parece probable que una linfocitosis, similar a la observada en animales como respuesta a pequeñas dosis de rayos X, es debida al efecto de los rayos ultravioletas del sol”. Una vez más, en 1920, Pappenheimer (97) publica sus estudios referentes a “el efecto de las inyecciones intravenosas de dicloroetilsulfide en conejos, con especial referencia a su acción leucotóxica”. Establece la dosis mortal del compuesto en conejos, describe el cuadro clínico y las lesiones patológicas. 59 RESULTADOS Determina que administrado por vía intravenosa es venenoso para el sistema hematopoyético, dando lugar a lesiones severas en médula ósea y ocasionado una disminución importante del número de leucocitos circulantes, siendo las células granulares de la médula ósea más sensibles que los eritrocitos y las células linfoides. En 1920, Lewis y Webster realizan sus estudios sobre “la migración de los linfocitos en cultivos plasmáticos de ganglios linfáticos humanos” (98). Los cultivos se obtienen de ganglios linfáticos normales y patológicos, procedentes de las intervenciones quirúrgicas del Hospital Johns Hopkins. Se registran los patrones de migración de linfocitos, tasa de migración, su aspecto y estructura. Observaron que los linfocitos eran casi siempre las primeras células en emigrar fuera del coágulo de plasma: “…a veces hacían círculos complejos, migraban, descansaban, para volver a migrar, pero no siempre seguían la misma dirección. Los núcleos sufrieron muchos cambios de forma, se veían torcidos, volteados y alargados durante la migración”. En cuanto a la forma, el linfocito que descansaba era más o menos esférico, y con la imagen característica de una cáscara delgada de citoplasma con un núcleo relativamente grande. En movimiento, tenían una forma peculiar, alargada, con el núcleo siempre en el extremo anterior. El grado de elongación variaba, pero el núcleo permanecía siempre en el borde anterior, con una pequeña cantidad de citoplasma ameboide. No se observaron mitosis ni se apreciaron diferencias notables en cultivos de ganglios normales y ganglios patológicos. 60 RESULTADOS Figura 10. Migración de linfocitos. Nº1. Cultivo de 48 horas de un nódulo sarcomatoso metastático en homoplasma. La migración de un linfocito a intervalos de ½ minuto. Se muestra el cambio de la forma de migración a reposo y vuelta a migración. Nº2. Cultivo de 20 horas a partir de un nodo normal. Migración de un linfocito a intervalos de 1 minuto durante 1 hora. Nº3. Cultivo de 24 horas de linfadenitis en homoplasma. Migración de un linfocito en el mismo campo dibujado a intervalos de 1 minuto. Nº4. Cultivo de 1 ½ hora de un nodo carcinomatoso metastático en homoplasma. Migración de un linfocito dibujado a intervalos de 1 minuto. Nº5. Cultivo de 4 días de un nódulo tuberculoso en autoplasma. 4 linfocitos en migración en el mismo campo, a intervalos de 1 minuto. Tomado de: Lewis y Webster, 1920 (98). Unos meses después Lewis y Webster publican sobre las células gigantes que aparecen en estos cultivos procedentes de ganglios linfáticos humanos (99). Reconocen varios tipos celulares que emigran fuera del coágulo: “Hay células pequeñas como los linfocitos, células grandes, células gigantes fusiformes y triangulares, polimorfonucleares, leucocitos y eosinófilos”. Se discute el posible origen de la célula gigante y su estructura: “Son células con múltiples núcleos y gran acumulación de gotas de grasa, de movimientos lentos, y se asemejan a las 61 RESULTADOS células que se encuentran en los ganglios tuberculosos. No existe seguridad de donde proceden ni cómo se desarrollan”. Con el título “el material reticular de las células sanguíneas en desarrollo”, publica Cowdry (100) en 1920, procedente del Laboratorio de Anatomía del Colegio Médico Unión de Pekín. Su objetivo es determinar la apariencia del material reticular de las células sanguíneas normales, para que sirva de guía en futuros estudios experimentales, usándolo como indicador de actividad celular. Utiliza médula ósea de conejo de indias para sus estudios, aplicando el método osmótico ácido de Kopsch, el método de nitrato de plata de Cajal y los métodos de fijación recomendados por Bensley. Sobre los resultados en linfocitos, Cowdry dice: “no he tenido éxito en la búsqueda de cualquier rastro de la presencia de un aparato reticular en células vivas de la serie linfocítica. En la médula ósea es sólo posible distinguir el linfoblasto del eritroblasto por la ausencia de hemoglobina”. Cowdry, encuentra sin embargo, que los linfocitos que habitan otros tejidos, como el tiroideo, muestran una red ennegrecida y definida en un lado del núcleo, cuando son tratados con el método de Cajal. Parece ser que fue Sinigaglia en 1910, el primero en describir una estructura en forma de red, alrededor del núcleo de las células rojas de anfibios. En 1912, von Berenberg-­‐Gossler (101) hizo mención a una red similar, pero colocada excéntricamente, en células de embriones de pollo, Fañanás observó que cuando aparece la hemoglobina de los glóbulos rojos, la sustancia reticular sufre cambios regresivos, y finalmente desaparece por completo, y Deineka (102) no tuvo dificultad para identificar una red en el tejido conectivo con el método de tinción de plata. Cowdry (100) menciona una serie de propiedades características de este material, entre ellas: • Que puede ser teñido de negro con ácido ósmico y nitrato de plata. • Que está dispuesto en una red de hebras anastomosadas, cuya forma, en cierta medida, es característica de los distintos tipos de células. 62 • RESULTADOS Esta red, se rompe en la división celular, distribuyéndose por igual en las células hijas. Por conveniencia, a esta sustancia que aparece en las células formando un retículo y que cambia lentamente de forma, la denominaron material reticular. Las observaciones realizadas por los mencionados autores, indican la existencia de “algún material” en el citoplasma de glóbulos rojos y linfocitos en desarrollo, que reaccionan de un modo más o menos característico a dos métodos diferentes de tinción. Este material encontrado también en otros tipos de células (nerviosas, glandulares y epiteliales), debe estar ampliamente distribuido en el protoplasma vivo y su presencia en las tres categorías de células sanguíneas, así como en células activas de casi todas las clases, ponen en evidencia que es un componente citoplasmático muy importante. Figura 11. Material reticular. 1-12. Células de la médula ósea de un conejo de indias macho, adulto, fijadas en 2% de ácido ósmico durante 15 días para la demostración del aparato reticular después del método de Kopsch. 13-21. Células de la médula ósea de un conejo de indias macho, preparado por el método de nitrato de plata de Cajal. Tomado de: Cowdry, 1920 (100). 63 RESULTADOS El término de fibra reticular, en el sentido que Cowdry lo emplea, se empieza a utilizar en 1892, gracias a Siegfried (103). La teoría reticular defendida por Golgi a finales del siglo XIX hacía referencia a la presencia en el sistema nervioso de una red difusa constituida por las anastomosis de las células nerviosas. Cajal, utilizando la tinción de plata de Golgi, describió la terminación libre y perfectamente delimitada de los axones, estableciendo que las células nerviosas son unidades independientes, nunca se anastomosan, abogando así por la teoría de la neurona y rechazando la teoría reticular de Golgi. Cuando hoy día hablamos de sustancia reticular o fibras reticulares, nos referimos a un tipo de fibras del tejido conectivo compuestas fundamentalmente por colágeno tipo III. Estas fibras, constituyen una fina red que actúa como una malla de soporte de los tejidos más blandos, como pueden ser hígado, médula ósea o tejidos linfáticos, y se tiñen con métodos de plata. Actualmente su mención se limita a las fibras compuestas por colágeno tipo III. Sin embargo, durante años, hubo confusión con el término, que se utilizaba para describir dos estructuras: a. Estructuras fibrosas argirófilas presentes en las membranas basales. b. Fibras histológicamente similares presentes durante el desarrollo del tejido conectivo. En 1920 nuevos estudios sobre la acción de los rayos X son llevados a cabo por Nakahara y Murphy (104) y publicados con el título: “Efecto de pequeñas dosis de Rayos X de baja penetración, en el tejido linfoide del ratón”. Ya que los ratones eran los animales más utilizados, los autores consideraron importante duplicar el estudio histológico en este animal. Estudiaron la repercusión del estímulo en los centros germinales de bazo y glándulas linfáticas, valorando principalmente la tasa de mitosis. Observaron que, dentro de los cuatro días siguientes a la administración de la radiación, aparecen un gran número de figuras mitóticas en el tejido linfoide de bazo y ganglios linfáticos indicativas de un aumento de la proliferación celular del tejido. Los autores manifiestan: “…la uniformidad de los cambios mostrados en los experimentos, tanto en el grado como en el período que aparecen, no pueden ser considerados una mera coincidencia”. La conclusión a la que llegan Nakahara y Murphy (104) es que 64 RESULTADOS las pequeñas dosis de rayos X son capaces de estimular la proliferación del tejido linfoide en ratones: “…con este conocimiento podemos decir que el aumento cuantitativo de elementos linfoides en el cuerpo es principalmente debido a hiperactividad del tejido linfoblástico de los órganos linfoides”. El linfocito en la resistencia natural e inducida al cáncer trasplantado, es un motivo reiterado de estudios. En 1920, Nakahara y Murphy (105) publican los resultados de las observaciones que llevan a cabo en la comparación histológica del tejido linfoide entre ratones con inmunidad natural y ratones susceptibles. Los órganos linfoides de ratones adultos jóvenes, son examinados histológicamente tres semanas después de ser inoculados con la cepa tumoral Adenocarcinoma Bashford nº 63. Se estudió el bazo y ganglios linfáticos de 105 ratones, 29 de los cuales eran ratones inmunes inoculados con cáncer. El punto a destacar en este trabajo, es la diferencia en la naturaleza de los cambios en los órganos linfoides de uno y otro grupo. En los ratones inmunes, hay en mayor o menor medida cambios que indican una hiperplasia linfoide, mientras que en los ratones susceptibles tiende a surgir una depleción linfoide. En el primer caso, la imagen histológica es muy parecida a la descrita en animales con estimulación linfoide artificial. Nakahara y Murphy (105) mantienen: “…estos resultados están totalmente de acuerdo con los de experimentos anteriores, que señalan el importante papel de los linfocitos, en la resistencia inducida artificialmente para el cáncer trasplantado”. Murphy, Nakahara y Sturm (Diciembre de 1920) continúan sus estudios sobre actividad linfoide. “La relación entre el tiempo y alcance de la estimulación linfoide inducida por agentes físicos y el grado de resistencia al cáncer en ratones” es ahora el motivo del trabajo (106). Se intenta determinar si existe una relación cuantitativa entre el grado de inmunidad y la estimulación. Los rayos X dan lugar a una estimulación de corta duración, el calentamiento provoca una estimulación aguda que persiste durante más tiempo. La inoculación al cáncer realizada en ratones inmediatamente después de la exposición a la radiación, origina poca resistencia, mientras que si la inoculación es hecha a la altura de la fase de estimulación, tiene lugar un claro 65 RESULTADOS aumento de la inmunidad. Los ratones inoculados inmediatamente después del tratamiento térmico, presentan una inmunidad pronunciada, pero no tan marcada como la que aparece cuando la inoculación se realiza a la altura de la estimulación. Todos estos resultados indican que el grado de inmunidad depende de la cantidad de estimulación linfoide existente. Un nuevo experimento para valorar el efecto de los rayos X es llevado a cabo por Nakahara (107) en 1920, cuyo objetivo es determinar la influencia del cáncer inoculado en la estimulación linfoide inducida por pequeñas dosis de rayos X. Los resultados de este estudio están de acuerdo con los resultados de los experimentos anteriores. Las conclusiones que se desprenden son las siguientes: 1. Si la inoculación del cáncer se realiza inmediatamente después de una dosis estimulante de radiación, interfiere con la reacción linfoide y la estimulación resultante es poca o ninguna. No tiene lugar la estimulación linfoide tal como ocurriría si no se hiciera la inoculación. 2. Si la inoculación del cáncer se realiza al mismo tiempo que la estimulación, tiene lugar un aumento de la reacción linfoide. 3. Si la inoculación se realiza 7 días después de la radiación, permitiendo que la estimulación se desarrolle antes de la inoculación, existe en la mayoría de los casos lo que podría llamarse una segunda estimulación del tejido linfoide. Nakahara (107) asegura que: “Los recuentos de sangre de los animales después de la inoculación muestran que sólo los animales que presentan un aumento de linfocitos llegan a ser resistentes al cáncer” Los rayos X son empleados como una herramienta de ayuda en la exploración del sistema linfoide. Ya en 1913, se registra un trabajo de Murphy y Ellis (71) con rayos X en animales, que provoca una depleción del tejido linfoide y facilita el crecimiento del tejido injertado. Murphy es, sin duda, el autor que más 66 RESULTADOS trabajos publica en este campo, observa una atrofia gradual del tejido linfoide por la acción de rayos X y estudia la resistencia a la tuberculosis de los animales así tratados (71,84,106). En un estudio sobre cáncer trasplantado (año 1915), los rayos X se utilizan para destruir completamente la inmunidad y facilitar el crecimiento de los implantes cancerosos (77). Sin embargo, no se conoce todavía el papel del linfocito en el organismo, ni los mecanismos fisiológicos y las células que participan en la inmunidad y la tolerancia a tejidos extraños, pero ya se diseñan experimentos con rayos X. Hace apenas dos décadas del descubrimiento de los Rayos X por Röntgen, en 1895 (108), y este interés por estos sorprendentes rayos, capaces de atravesar elementos sólidos y producir fotografías de huesos, podemos considerarlo una consecuencia lógica del proceso normal en la exploración de una nueva herramienta con infinitas posibilidades. A partir de su descubrimiento, los rayos X se convierten en uno de los elementos más revolucionarios para las distintas ramas de la ciencia, desde la astronomía hasta la radiografías en medicina. En palabras del propio Röntgen: “…casi inmediatamente, la posibilidad de aplicaciones prácticas atrajo al público, y muy especialmente a la profesión médica. Se tenía un método de gran utilidad para el diagnóstico de fracturas complicadas, o para localizar cuerpos extraños en el organismo”. Sólo un año después de su descubrimiento, ya aparecen publicaciones sobre su utilidad, como por ejemplo, para la extracción de cuerpos extraños de cobre en el vítreo (109), sobre sus aplicaciones en cirugía (110), o localización de cuerpos extraños en el esófago (111), y también se registran los primeros efectos nocivos, como la aparición de dermatitis causada por rayos Roentgen (112). A las pocas semanas de su descubrimiento, se publicaron los primeros informes de los efectos nocivos por la exposición a rayos X. El físico americano Thomson fue el primero en acusar una acción directa entre la exposición a rayos X y efectos nocivos, exponiendo su propio dedo índice media hora diaria durante varios días, y William Rollins propuso medidas de protección, tales como la utilización de vidrios opacos, protección con plomo de las 67 RESULTADOS habitaciones, y la sola radiación del área de interés (113). Se observó que las radiaciones de aquellos “rayos mágicos” actuaban sobre las células destruyéndolas, pero incluso a este inconveniente se le dio utilidad, como lo indican numerosas publicaciones: “El valor terapéutico de los rayos X en medicina” (114), “Diagnóstico de cálculos renales y su tratamiento mediante rayos X” (115), “Tratamiento del epitelioma de los párpados por rayos X” (116), “cuatro úlceras Rodent tratadas con rayos X”(117), por citar algunas de ellas. En 1906 aparece una publicación que refleja su utilización en el tratamiento de las leucemias (118): “El efecto de los rayos X sobre los leucocitos de la sangre y médula ósea en leucemias”. Se recoge que la acción de los rayos X se manifiesta clínicamente por una reducción en el número de leucocitos en la sangre, y se mantiene que la influencia de los rayos X alterando la proporción de células sanguíneas, puede actuar inhibiendo la excesiva producción, ya que las principales células presentes en la leucemia, los grandes mielocitos, disminuyen en número. En opinión de Buchanan: “…es evidente que la acción de los rayos X en la leucemia puede actuar de dos formas, inhibiendo la sobreproducción en médula ósea y destruyendo leucocitos” Un año después del descubrimiento de Röntgen, en 1896 Becquerel descubre la radiactividad natural (119), y en 1898 los esposos Curie descubren el polonio y el radio. En 1936, casi cuatro décadas después del descubrimiento de los rayos X y del radio, la sociedad alemana Röntgen erigió un monumento en memoria de todos los que habían muerto como consecuencia de la exposición a rayos X o radio. Marie Curie (113) reconoció que “el radio era peligroso en manos no entrenadas”, y defendió la formación específica para los trabajadores con sustancias radiactivas. Otro de los interrogantes no resueltos es, ¿a dónde van los linfocitos en su viaje por el torrente sanguíneo? ¿Qué ocurre con ellos? ¿Se desintegran dentro de los vasos sanguíneos? La cuestión sobre “el destino de los linfocitos” es abordada por Bunting y Huston en 1921, del Laboratorio de Patología de la Universidad de Wisconsin (120). Es evidente la desintegración de linfocitos en 68 RESULTADOS extensiones de sangre de individuos normales o enfermos, y parece incuestionable que algunos se fragmentan en el torrente sanguíneo. ¿Es éste el destino de la gran mayoría de linfocitos? Bunting y Huston realizan una serie de experimentos en conejos, para determinar el número de linfocitos que circulan libremente en la vena marginal de la oreja derecha, para ello aíslan una sección completa de la vena yugular derecha, entre ligaduras dobles aplicadas cuidadosamente, y hacen el recuento celular 6 y 24 horas después. A pesar de las imperfecciones del método, como la reducción de oxígeno que provoca en las células, hay una mayor desaparición de polimorfonucleares que de linfocitos, que puede ser explicado por su emigración hacia el campo operatorio. Los linfocitos no muestran ninguna tendencia a la desintegración, observándose una clara diferenciación de núcleo y borde protoplásmico como en las extensiones de sangre circulante. Bunting y Huston (120) expresan su creencia de que los experimentos, aunque defectuosos y abiertos a la crítica, indican que no es la desintegración de las células la que ocasiona la tasa de su desaparición en la sangre, cuando el suministro se interrumpe: “…ellos deben abandonar el torrente sanguíneo” manifiestan. Como el movimiento ameboide de los linfocitos es aceptado, no hay objeciones a priori a la proposición de que salen de la corriente sanguínea en condiciones normales. La cuestión ahora es la siguiente: “¿a dónde se dirigen?”. Y su respuesta: “No lo sabemos, pero deben destruirse, ya que sin destrucción, necesariamente habría una acumulación cada vez mayor en los órganos, como puede verse en las leucemias linfocitarias”. Los autores llegan a las siguientes conclusiones: 1. Aunque el número de linfocitos circulantes permanece constante, desde el conducto torácico se produce una entrada continua de los mismos a sangre. 2. Este exceso de linfocitos, no se destruye en la circulación sanguínea, migran desde los vasos a la superficie de las membranas mucosas, sobre todo del tracto gastrointestinal, y es al parecer en la mucosa 69 RESULTADOS intestinal, dentro de la luz intestinal, donde normalmente tiene lugar la función de los linfocitos. Gerald B. Webb (121) en 1922 estudia la relación entre linfocitos e infección tuberculosa y centra sus investigaciones en la importancia del índice linfocitario. No es un hecho nuevo que una linfocitosis acompañe a la tuberculosis, ni que cuanto mayor sea esta linfocitosis mejor será el pronóstico, pero estas observaciones no habían recibido el reconocimiento que requerían dada su importancia clínica. Existía un índice linfocitario que asignaba los siguientes porcentajes: • En adultos normales, 43,6% • En pacientes curados de tuberculosis, 41,7%, • En casos con mejoría, 38,5%, • En casos estacionarios, 34,4%, y • Con progresión desfavorable, 27,5%. Hay que reseñar que estos porcentajes incluían todas las células mononucleares basófilas y estaban determinados a una altitud de 6000 pies (1828 metros). El porcentaje para un adulto a nivel del mar se sitúa en torno al 34%. El aumento de linfocitos como efecto de la altitud, fue establecido por Webb y Williams en 1909 (122) y posteriormente confirmado por autores como Staübli, Staines y James (123). Así mismo existían comunicaciones que establecían una estrecha relación entre el porcentaje de linfocitos, la condición del paciente y la actividad de la enfermedad (124). Webb hace referencia a los trabajos de Murphy y Taylor (84) con ratones radiados y la susceptibilidad a tuberculosis (ya comentado con anterioridad). Albert Calmette (125), en 1920, en “L´infectión Bacillaire et la tuberculose” habla de tres distintas fórmulas leucocitarias en los diversos tipos o etapas de la enfermedad. Así habría una “fórmula de resistencia”, en aquellos casos de fibrosis benigna y evolución lenta, que sería una leucocitosis moderada de no más de 10.000 por cm, con una linfocitosis marcada. La “fórmula de defensa”, 70 RESULTADOS en casos de infección masiva o especial virulencia, consiste en una hiperleucocitosis y abundante polinucleosis con disminución constante de eosinófilos y linfocitos. Por último, la “fórmula del desastre”, en casos de progresión a caseificación grave y formación de cavidades, caracterizada por una hiperleucocitosis de 16.000 a 20.000 con polimorfonucleares cercanos al 90%, y una gran disminución del número de linfocitos, en ocasiones degenerados, mientras los eosinófilos están ausentes o fragmentados. Webb (121), durante diez años, realiza un seguimiento a pacientes tuberculosos, a los que somete a recuentos sanguíneos de rutina. Observa así el importante valor del porcentaje linfocitario como una medida simple y precisa de la condición y el pronóstico del paciente. Con el fin de asegurar los resultados, analiza más de 200 determinaciones sucesivas en 400 pacientes, en todas las etapas de la enfermedad. Establece un índice linfocitario medio, que para adultos normales es del 43%, en pacientes curados o estacionarios del 45%, en casos que no mejoran del 24%, y del 0,16% si la evolución es mortal. Observa que si este índice se mantiene por debajo de 20, la evolución es muy desfavorable, en cambio un aumento gradual es muy esperanzador. Establece también los valores del índice en niños, teniendo en cuenta que durante los dos primeros años de vida es mayor, disminuyendo lentamente hasta los 10-­‐12 años. Queda la duda, si realmente es el linfocito u otro tipo célula semejante morfológicamente, el elemento implicado en la resistencia a la tuberculosis. Bergel (126), considera a todos los mononucleares presente en la sangre linfocitos, afirmando que el aspecto diferente entre grandes fagocitos mononucleares y linfocitos es simplemente una cuestión de funcionalidad. Por otro lado, parece establecido (127), que las grandes células mononucleares tienen su origen en el endotelio vascular y una capacidad funcional para ingerir partículas extrañas, propiedades que no son compartidas por las células que derivan del tejido linfoide. Aunque hay casos de tuberculosis muy avanzada que muestran una disminución en el porcentaje de leucocitos endoteliales, en realidad se produce un aumento absoluto debido a la leucocitosis presente. 71 RESULTADOS Webb (121) teniendo en cuenta todas estas diserciones, concluye que todos los hallazgos parecen indicar que: “son los linfocitos y no los leucocitos endoteliales, los factores más importantes para medir la resistencia a la tuberculosis en seres humanos”. Para poder entender el significado y la importancia de una enfermedad como la tuberculosis en la historia de la humanidad, necesariamente hemos de retroceder algunos años. Como consunción, tisis, mal del rey, peste blanca o plaga blanca ha sido nombrada a través de la historia (128). Se han constatado indicios de su presencia en huesos del neolítico, aunque es muy difícil conocer su magnitud con anterioridad al siglo XIX (129) (130). Según Hipócrates (siglo V a. C): “... un tísico viene de otro tísico, y prende más fácilmente en ciertos temperamentos como pituitosos, flemáticos e imberbes rubios de ojos brillantes, carnes blandas y omóplatos sobresalientes”. En la Europa de la Edad Media, acorde con la concepción teocentrista, se introducen como remedio a la tuberculosis terapias alternativas de carácter sobrenatural, siendo la más popular el “toque del rey”. El rey es considerado un delegado de Dios con propiedades mágico-­‐curativas, así Felipe el Hermoso, Roberto II el Piadoso o San Luis de Francia, tocaban las escrófulas pronunciando las palabras “el Rey te toca, Dios te cura”. A Santa Águeda, protectora de los enfermos del pecho, se encomiendan los tísicos cuando no pueden acudir al rey. Una enfermedad, que ni un rey ni una santa pueden curar es considerada un castigo divino, algo que provoca terror y vergüenza. Paracelso en el siglo XVI, propone una actitud indiferente hacia la tisis, por ser una enfermedad incurable. En el siglo XVIII, Benjamín Marten (131), autor de un tratado sobre la tuberculosis titulado “A new theory of consumptions more specially of phthisis or consumptions of the lungs” propone la novedosa teoría de que la tuberculosis es debida a algún tipo de “animacula” o ser vivo capaz de vivir en nuestro organismo. Teoría que es rápidamente rechazada por absurda. Ciento cincuenta años después (año 1882) Koch demuestra esto de forma inequívoca (129)(132). 72 RESULTADOS La epidemia de tuberculosis en Europa, que probablemente se inició a comienzo del siglo XVII y continuó durante más de doscientos años, se conoció como “gran plaga blanca”, teniendo su máxima extensión desde finales del siglo XVIII a finales del siglo XIX. En estos años tuberculosis era sinónimo de muerte inevitable. La revolución industrial que favorece la extensión de la enfermedad, debido a las malas condiciones de vida de los trabajadores en las grandes ciudades, el hacinamiento y las jornadas de trabajo abusivas, va a favorecer al mismo tiempo el inicio del desarrollo de la medicina científica. La tuberculosis se marginaliza a medida que se evidencia su carácter contagioso, y anticipa el fenómeno que a mayor escala se producirá un siglo después con el SIDA (128)(133). Hay que esperar a los últimos años del siglo XIX para que se produzcan los avances científicos que van a marcar un antes y un después en la historia de la enfermedad. En 1869, Villemin demostró que la tuberculosis era una enfermedad infecciosa, y pocos años después Koch descubrió el bacilo causante. Con el descubrimiento de los rayos X en 1895 (134), es ahora posible el diagnóstico y seguimiento de la enfermedad, pero todavía está lejos el poder curarla, ya que no es hasta mediados del siglo XX cuando comienza a emplearse un tratamiento eficaz. En los primeros años del siglo XX se asiste a un esfuerzo comunitario global por saber más sobre cómo abordar la enfermedad, los tratamientos siguen siendo muy diversos e ineficaces, flebotomías, eméticos, purgantes y dietas se encuentran entre ellos (135). Surgen sanatorios especializados, lo que por vez primera supone un obstáculo a la cadena de transmisión del microorganismo, y esto junto a la mejora de las condiciones de vida, hace que la tuberculosis inicie un lento descenso antes de la aparición de los primeros fármacos eficaces (136). En 1944, en plena guerra mundial, Schatz y Waksman descubren la estreptomicina, y en 1952 se desarrolla la Isoniacida, el primer antibiótico específico que conseguirá que la tuberculosis se convierta en una enfermedad curable. Pero quizás, donde mejor podemos apreciar el cortejo anímico que acompañaba a la enfermedad y su significado social es en el arte. Dada su 73 RESULTADOS extensión, la tuberculosis acabó convirtiéndose en un tópico o subgénero recurrente en arte, literatura, pintura y cine durante estos doscientos años de máximo apogeo (137)(138). “La Miseria” de Cristóbal Rojas, pintada en 1886 es una muestra de ello. El autor aquejado de tuberculosis, refleja en su obra el aspecto social de la enfermedad y su relación con las condiciones de vida. La tuberculosis también aparece en óperas como la Bohéme y la Traviata. Moliére afectado de tisis muere en el escenario representando una de sus obras (el enfermo imaginario). En la literatura, “la montaña mágica” de Thomas Mann o “el pabellón de reposo” de Camilo José Cela (año 1943) hacen referencia a los sanatorios para tuberculosos. Este breve recordatorio sobre el pasado histórico de la tuberculosis, nos ayuda a entender la importancia histórica actual de los estudios que relacionan tuberculosis y linfocito de los autores aquí contemplados como Murphy (año 1914), Taylor y Murphy (año 1917) y Webb (año 1922), y comprender no sólo el valor científico de su trabajo en ese momento, sino que constituyen también una pequeña muestra del esfuerzo comunitario global en la lucha contra una terrible enfermedad. El aceite de oliva es reiteradamente estudiado por Nakahara como agente capaz de modificar la actividad linfoide. En 1922, publica sobre la inmunidad al cáncer trasplantado inducida por la inyección de aceite de oliva (139). En 1923, estudia la supresión de la inmunidad inducida al cáncer trasplantado por grandes dosis de aceite de oliva (140). En 1924, de nuevo, trabaja en la resistencia al cáncer espontáneo en ratones, inducida por inyecciones de ácido oleico (141). Basado en los estudios de Clark (142), quien unos años antes observa que la inyección subcutánea de aceite de oliva ejerce, una acción quimiotáctica sobre los linfocitos, Nakahara (139) considera interesante determinar si el aceite puede dar lugar a una estimulación linfoide general, y si es así, el efecto que ejerce sobre la resistencia al cáncer. Se utilizaron 25 ratones blancos que fueron divididos en 5 grupos de 5 ratones cada uno. A cada grupo se les 74 RESULTADOS administró una inyección intraperitoneal a dosis distintas (0,1 cc, 0,2 cc, 0,3 cc, 0,5 cc y 0,7 cc). Un animal de cada grupo fue sacrificado para estudio histológico a las 24, 48 horas, 3, 4 y 5 días después de la inyección. La reacción más pronunciada en órganos linfoides se obtuvo a la dosis de 0,2cc de aceite de oliva. Se repitió el experimento en 12 ratones, a los que se inyectó 0,2 cc de aceite de oliva, siendo sacrificados a las 48 horas, 4, 7 y 10 días después. Se observó que en el período de 4-­‐7 días todos mostraron un aumento del número de figuras mitóticas en órganos linfoides. Como control, un grupo de ratones normales fue inyectado con vaselina líquida intraperitoneal, sin que diese lugar a ningún aumento en el número de mitosis. Una vez determinada la dosis idónea de aceite, se pasó a estudiar el grado de resistencia de los ratones al cáncer mediante la inoculación en la ingle izquierda de fragmentos de Adenocarcinoma Bashford nº 63. Se tuvo en cuenta el recuento sanguíneo, la estimulación del tejido linfoide valorado según el número de mitosis y la reacción local en el injerto celular, tanto en los animales resistentes como susceptibles. Los resultados mostraron que el recuento de células blancas linfoides a los 10 días de la inoculación en los animales resistentes era superior al encontrado en animales susceptibles, que la estimulación linfoide era mayor en animales resistentes y que la reacción local en el animal resistente estaba fundamentalmente formada por linfocitos. Nakahara (139) expone la situación con estas palabras: “… por tanto podemos decir que la resistencia al cáncer trasplantado puede ser inducida por varios agentes: agentes biológicos como el tejido homólogo, agentes físicos como los rayos X y el calor y agentes químicos como el aceite de oliva”. “No se conoce la naturaleza común de estos factores, pero las manifestaciones asociadas con fenómenos de resistencia son las mismas con independencia del medio utilizado para inducir tal estado”. Estas manifestaciones comunes son la existencia de un periodo de latencia después del tratamiento, una reacción local celular sobre el cáncer injertado, el aumento de linfocitos circulantes y una marcada actividad proliferativa de órganos linfoides. 75 RESULTADOS El trabajo de Nakahara muestra: a. Que es posible lograr ratones resistentes a cáncer inoculado, mediante inyecciones de una adecuada cantidad de aceite de oliva, b. Que existe un período de latencia y el grado de máxima resistencia aparece a los 10 días, c. Que el estado de resistencia estudiado histológicamente, es precedido por la proliferación de las células linfoides de los centros germinales, y después de la inoculación del cáncer, se asocia con una infiltración linfoide sobre los injertos, así como con una segunda estimulación de los centros germinales y un aumento del número de linfocitos circulantes. El segundo estudio de Nakahara (140) tiene por título “La supresión de la inmunidad inducida al cáncer trasplantado por grandes dosis de aceite de oliva”. Si se administran grandes dosis (0,7 a 1 cc) de aceite de oliva a ratones potencialmente inmunizados, inmediatamente antes de inocular el cáncer (Adenocarcinoma Bashford nº 63), se observa un aumento en el número de los tumores que crecen en relación con los controles y en relación con los ratones inmunizados a los que no se les administró aceite. Este efecto depresor del aceite de oliva es operativo siete días después de la inyección. En cambio, si la administración de grandes dosis de aceite se realiza en ratones inmunizados, después de la inoculación del cáncer, es imposible, al parecer, suprimir los mecanismos de resistencia después ser activados por inoculación de cáncer. Como ya sabemos, las reacciones celulares que acompañan a la inmunidad, son una marcada infiltración linfocitaria en todo el injerto inoculado, una estimulación de la división celular en centros germinales de órganos linfoides y la aparición en sangre de linfocitosis. El estudio histológico de los ratones una vez sacrificados, nos muestra que el aceite de oliva en grandes dosis suprime la proliferación de células linfoides, disminuyendo el tamaño de centros germinales en bazo y ganglios linfáticos. 76 RESULTADOS Como consecuencia de los resultados obtenidos, Nakahara (140) mantiene que: • El estado de inmunidad potencial al cáncer trasplantado, inducido en ratones mediante inyección de sangre homóloga, puede ser invertido a un estado susceptible, por una inyección intraperitoneal de aceite de oliva antes de la inoculación del cáncer. • La supresión de la resistencia va acompañada del fallo de ciertas reacciones celulares conocidas durante el establecimiento de la inmunidad. Una vez demostrado que la resistencia al cáncer injertado se puede aumentar en gran medida con una inyección previa de una pequeña cantidad de aceite de oliva o ácidos grasos insaturados, el interés de Nakahara se centra en el estudio de “la acción de los ácidos grasos en la resistencia de los animales a sus propias neoplasias espontáneas” (141)(143). Se utilizó ácido oleico, debido a que la gama de dosis eficaz es más ancha que la de otros ácidos grasos insaturados. Los ratones fueron tomados de la población tumoral mantenida en el Instituto Rockefeller (cepa Lathrop) y todos fueron ratones hembras con tumores de la glándula mamaria. Se hizo un intento de determinar en primer lugar, el efecto del ácido oleico en las recurrencias después de la eliminación quirúrgica del cáncer de ratón espontáneo y, en segundo lugar, el efecto sobre la incidencia del cáncer tomado de injertos autoplásticos. Las observaciones realizadas indican: 1) Una reducción en la incidencia de recidivas locales en el lugar de la intervención. 2) Una disminución de la incidencia de tumores primarios en otros lugares. 3) Disminución de las metástasis. 4) Disminución de crecimiento de injertos autoplásticos. 77 RESULTADOS 5) Una actividad aumentada del tejido conectivo y del número de linfocitos en los ratones tratados. 6) Un aumento apreciable de la longevidad media postoperatoria. Nakahara (143) asegura: “Los resultados muestran que la resistencia de los ratones a las enfermedades neoplásicas espontáneas se puede aumentar mediante la administración de ácido oleico”. No sabemos en realidad los motivos que llevaron a Nakahara a investigar sobre las propiedades del aceite de oliva. En su primer trabajo de 1922 (139) hace referencia a Ramond (año 1904), quien encuentra que el aceite de oliva inyectado en el peritoneo es absorbido por las células blancas, especialmente por linfocitos, a Clark (año 1917), que observa la acción quimiotáctica ejercida por el aceite de oliva sobre los linfocitos, y a Bergel (año 1919), que advierte que el exudado después de un inyección intraperitoneal de aceite de oliva está formado fundamentalmente por células de tipo linfoide. Aparte de estos antecedentes, y una vez consultada la literatura al respecto, no se encuentran publicaciones anteriores que estudien al aceite de oliva en el sentido que lo hizo Nakahara. Aparecen publicaciones donde figura el aceite pero contemplado desde una perspectiva muy distinta. Así, encontramos en 1892 “observaciones sobre los cálculos biliares y su tratamiento por grandes cantidades de aceite de oliva” (144), en 1894 “notas sobre la esterilización de Catgut (cuerda de tripa) en aceite de oliva hirviendo” (145), en 1895 “curaciones de cólico hepático por ingestión de aceite de oliva” (146)(147), en 1918 “uso del aceite de oliva en la sutura de vasos sanguíneos” (148) o “el valor de una mezcla de Cyllín, yodoformo y aceite de oliva en la sepsis intratorácica” (149). Los estudios de Nakahara sobre actividad linfoide relacionando al aceite de oliva con la inmunidad al cáncer son estudios pioneros en la historia. El aceite de oliva es tratado como un agente químico con poder de intervención en los mecanismos inmunológicos. 78 RESULTADOS Publicaciones posteriores, como ésta de 1936: “las propiedades bactericidas y fotoquímicas del aceite de hígado y un ozónido del aceite de oliva” (150), o la de 1945 “efecto del consumo del aceite de oliva en la absorción y depósito de colesterol” (151), siguen teniendo un enfoque muy distinto al dado por Nakahara. Si la búsqueda se realiza con ácido oleico, encontramos un trabajo anterior a 1924, del año 1913, sobre “las propiedades hemolíticas de los ácidos grasos y su relación con la hemólisis tóxica y la anemia perniciosa” (152), que tampoco tiene el espíritu de los trabajos de Nakahara. Pero sí encontramos trabajos de índole muy parecida en fechas posteriores, de lo que deducimos que los estudios de Nakahara pudieron servir de inspiración o modelo a estos autores. Así, en 1959, treinta y cinco años después, tenemos “efectos promotores de tumores en la piel del ratón; comparación entre ácido oleico, mono-­‐oleato de sorbitán (Span 80) y, mono-­‐oleato de sorbitán polioxietileno (Tween 80)”(153), y en 1989, sesenta y cinco años más tarde, “efecto de la inhibición del crecimiento por ácido oleico, ácido linoleico y sus ésteres de metilo en tumores trasplantados en ratones” (154), publicación cuyos resultados indican que ambos ácidos, oleico y linoleico, prolongan significativamente la vida de ratones portadores de carcinoma, en comparación con los ratones de control no tratados, y se observa que los ácidos grasos libres inhiben selectivamente el crecimiento de células tumorales. En 1922 Ferdinand Lee publica los resultados de sus experimentos sobre “Cambios en el número de pequeños linfocitos de la sangre que siguen a la ligadura del conducto torácico” (155). Lee utiliza gatos machos adultos, en los que se determina, en un primer momento, el hemograma y la citología a partir de la sangre obtenida de la oreja, para en un segundo paso, llevar a cabo la ligadura del conducto torácico y del plexo linfático periaórtico. Cinco horas después de la operación, la sangre fue examinada de nuevo, haciendo recuentos durante los 7 días siguientes y después a días alternos. Al final de la tercera semana los animales son 79 RESULTADOS sacrificados para el estudio histológico de diversos órganos. Una vez establecidas las normas para la identificación de las células (linfocitos pequeños y grandes) en los recuentos diferenciales con la tinción de Wright, Lee realiza las siguientes observaciones experimentales: “… el número de pequeños linfocitos cae aproximadamente un 56% inmediatamente después de la ligadura, y vuelve gradualmente al nivel pre-­‐ligadura, sobre el día 19 a 21”. Muchos investigadores habían estudiado el conducto torácico respecto al tipo y numero de células que se encuentran en la circulación. Se observó que la ligadura no impedía por completo la entrada de mononucleares a la sangre, pero las inyecciones de color no mostraron conexiones entre los sistemas sanguíneos y linfáticos, demostrándose posteriormente que pueden llegar a establecerse caminos colaterales después de la ligadura del conducto torácico. Parece que el retorno de los linfocitos a niveles normales aproximadamente a la 3ª semana post-­‐ligadura, es debido al re-­‐establecimiento de la circulación colateral del conducto torácico, aunque no existen pruebas absolutas de ello, sin embargo, es definitivo que el conducto torácico es una importante vía para la entrada de los pequeños linfocitos a la sangre circulante, y que es la vía a través de la cual, al menos el 50% de los linfocitos pequeños llegan a la circulación en el gato. En 1923, Alexis Carrel y Albert Ebeling (156) trabajando en el Instituto Rockefeller, intentan determinar si los linfocitos y células mononucleares, producen sustancias libres que influyan en la proliferación de fibroblastos, cuando se encuentran en un medio que contiene suero como única fuente de nitrógeno. Proceden a determinar la acción del suero en la multiplicación de los linfocitos, demostrando que son capaces de vivir y multiplicarse en un medio que contiene solo suero de la sangre, y que éste no es suficiente en el caso de los fibroblastos. Observan que la presencia de linfocitos y grandes células mononucleadas en la vecindad de un cultivo de fibroblastos en suero, aumenta notablemente la actividad y duración de la vida de los fibroblastos. En vista de estos resultados los autores manifiestan: “Es probable que los linfocitos puedan sintetizar a partir de compuestos de nitrógeno contenidos en el suero, 80 RESULTADOS sustancias que los fibroblastos y células epiteliales requieren para su multiplicación”. Un original trabajo es publicado en 1924 por Jordan y Speidel (157), procedentes del Departamento de Histología y Embriología de la Universidad de Virginia, Charlottesville, sobre “el comportamiento de los leucocitos durante la regeneración y metamorfosis inducida por tiroides, en la larva de la rana, con un análisis de factores de crecimiento”. Carrel (156) había obtenido resultados que indicaban que los leucocitos tienen una acción promotora de crecimiento, ya que emiten sustancias estimulantes del crecimiento de células adyacentes. Los tejidos de la larva de la rana es un excelente material para el estudio de las células sanguíneas en relación a los cambios progresivos y regresivos que tienen lugar en el renacuajo. Los cambios regresivos pueden ser fácilmente inducidos por la administración simultánea de tiroides y la eliminación de parte de la cola, que es rápidamente seguida por la regeneración de la parte eliminada. El tratamiento del animal con extracto de tiroides no sólo inhibe el crecimiento en la cola, sino que inicia el proceso degenerativo. Cuando se realiza el examen microscópico de los tejidos en la zona de regeneración, se observa que los linfocitos están presentes en el proceso de regeneración de la parte perdida de la cola. Sin embargo, no lo son en suficiente número para justificar la conclusión de que son de vital importancia en el crecimiento de la nueva cola. Se observan los siguientes resultados: ! En los cambios coincidentes regresivos y progresivos inducidos en la cola del renacuajo de rana mediante administración de tiroides y simultáneamente eliminación de parte de la cola, se observa una tasa de crecimiento mayor en animales no tratados con tiroides, seguida por los animales tratados 2 días después de la eliminación de la cola, algo menos en animales tratados simultáneamente a la intervención, y menos en los tratados con tiroides 2 días antes de la retirada de la cola. 81 RESULTADOS ! Los linfocitos aparentemente no son lo suficientemente numerosos como para ejercer una importante función promotora del crecimiento. Por lo que Jordan y Speidel dicen: “… si presentan alguna función promotora del crecimiento, debe ser pequeña en comparación a su correspondiente actividad en las extremidades de los animales tratados con tiroides”. Harold A. Abramson centra su interés en la electroforesis de las células sanguíneas. Fruto de su trabajo en el Departamento de Cirugía de la Universidad de Columbia en Nueva York (año 1924)(158) y en el Instituto de Física, Química y Electroquímica de Kaiserwilhelen, en Alemania (año 1927) (159), son algunas de sus publicaciones. El primero de sus trabajos estudia “la influencia de una fuerza electromotriz de bajo voltaje en la electroforesis de linfocitos de diferentes edades” (158). Por experiencias anteriores es sabido que los linfocitos que se mantienen a 37ºC fuera del cuerpo, exhiben movimientos espontáneos que aumentan con el tiempo. La velocidad media espontánea de estos pequeños linfocitos se calculó en 4 µ por minuto, llegando a 15 µ por minuto después de cinco horas. Entre 1922 y 1924 (160) tienen lugar una serie de experimentos a fin de determinar la influencia de una fuerza electromotriz en los movimientos de las células blancas de la sangre, comparando esta fuerza electromotriz con las que pasan entre los tejidos dañados y los tejidos normales, deduciéndose que tal vez uno de los factores que atrae a las células blancas de la sangre al punto de la lesión pueda ser “esta corriente de lesión”. Con la finalidad de realizar estudios de este tipo, Abramson construye una cámara de electroforesis de porcelana sin esmaltar, donde coloca una pequeña gota de sangre desfibrinada y centrifugada entre dos electrodos. En estos experimentos, los linfocitos estudiados fueron los que se movieron en una línea casi recta a través del campo, en dirección hacia el ánodo. No se observaron linfocitos que se dirigieran hacia el cátodo. La sangre empleada en el estudio fue extraída de la vena del brazo de un hombre adulto supuestamente sano, y después de ser desfibrinada y centrifugada se mantuvo en refrigeración hasta ser utilizada. 82 RESULTADOS La pregunta que se plantea Abramson es, si en los tejidos de animales vivos, la fuerza electromotriz que pasa entre tejidos sanos y lesionados es del mismo orden de magnitud que la experimental de 0,8 voltios por centímetro. Y la explicación que da del fenómeno es la siguiente: “Cuando los tejidos se dañan, los cambios físico-­‐químicos que tienen lugar hacen que el área lesionada sea electropositiva, por tanto puede ser considerada similar al ánodo, y la dirección en la migración de los linfocitos podría corresponder con la sugerencia de que la fuerza electromotriz es parcialmente responsable de la reacción inflamatoria celular”. En un segundo trabajo sobre “la electroforesis de las células sanguíneas en el caballo y su relación con la migración leucocitaria” (159), Abramson menciona que la relación entre el potencial electrocinético de los leucocitos y las diferencias existentes entre zonas de tejido lesionado y no lesionado, no está resuelta, ni cualitativamente ni cuantitativamente, y quizás la solución se encuentre en la migración del leucocito como respuesta a la lesión tisular. Hace un extenso e interesante recuerdo histórico sobre el estudio del comportamiento eléctrico de las células blancas sanguíneas. Dineur (año 1893) parece haber sido el primero en el estudio de la carga eléctrica de las células blancas de la sangre. Observó el movimiento de los leucocitos en la cavidad peritoneal de la rana en estado normal e inflamada, registrando lo ocurrido cuando tenía lugar el paso de una corriente eléctrica. Lillie (año 1903) utilizó una suspensión isotónica de azúcar-­‐NaCl, y estudió el comportamiento de las células y su migración hacia los polos del campo electromagnético. Schwyzer y Feringa (161) coinciden en la creencia de que la diferencia de potencial entre tejido sano y lesionado es la causante de la migración leucocitaria al punto de la lesión. Abramson inició sus estudios sobre linfocitos humanos suspendidos en suero, encontrando que éstos estaban cargados negativamente, y estimó la velocidad media de cataforesis (CV) en 0,3µ por voltio por centímetro por segundo. Mantenía que debería fluir una corriente continua entre el tejido sano y el lesionado, y que la energía procedía de los cambios metabólicos incidentales a 83 RESULTADOS la lesión. Presenta datos relativos a las velocidades de linfocitos y polimorfonucleares en el plasma y suero de caballo, y sus observaciones las extiende a geles de distinta viscosidad. Los puntos fundamentales de trabajo de Abramson (159) son los siguientes: • La velocidad de migración de células de la sangre por cataforesis en plasma y suero es proporcional a la caída de potencial aplicada. • Se describe la velocidad de cataforesis de glóbulos rojos, leucocitos, polimorfos, pequeños linfocitos y leucocitos grandes para suero y plasma. • La relación entre el potencial electrocinético de células blancas de la sangre y la diferencia de potencial existente en tejidos lesionados están relacionados cuantitativamente. • Esta correlación sugiere que la migración de leucocitos a un punto de la lesión, es dependiente de fuerzas electromagnéticas en juego en los tejidos. En 1928, se registra una comunicación de Philip Silver, en Montreal, Canadá (162), con el propósito de presentar dos casos de Linfocitoma Maligno, y abogar por una mayor uniformidad en la nomenclatura de los tumores linfoides. En los dos casos se hace una descripción detallada de clínica, estudio histológico, tratamiento y seguimiento de los pacientes. Silver hace un breve repaso de los tumores linfoides, dentro de los cuales incluye las hiperplasias inflamatorias simples, neoplasias de rápido crecimiento y una variedad de tipos intermedios, y plasma su opinión al respecto: “los tumores linfoides son el resultado de un sobrecrecimiento de células linfoides atípicas. Son enfermedades de rara aparición. También son de etiología desconocida el linfoadenoma, leucemia y pseudoleucemia, que producen en mayor o menor medida un aumento del tejido linfoide, y dificultan su diagnóstico diferencial”. Se queja de la existencia de una falta de uniformidad en la nomenclatura de los tumores linfoides, que cree debido a la falta de conocimiento en su etiología. Expone la clasificación de Ewing, basada en tres elementos capaces de originar tumores 84 RESULTADOS linfoides: linfocitos, células reticulares de folículo y pulpa y células endoteliales de pulpa y senos cavernosos. Según esta histogénesis, los tumores linfoides se clasificarían en: 1-­‐ Los derivados de linfocitos o linfocitomas, que incluye linfoma simple, linfoma tuberculoso, leucemia linfática, pseudoleucemia y linfocitoma maligno. 2-­‐ Los que derivan de células reticulares como el granuloma maligno, leucemia mieloide, sarcoma Hodgkin y sarcoma de células grandes y, 3-­‐ Los derivados de células endoteliales (hiperplasia endotelial de tuberculosis y endotelioma), que pueden ser de origen hiperplásico o neoplásico. En 1928 W.R. Earle (163) publica los resultados de un estudio sobre el efecto que la luz tiene en la sangre y tejidos celulares, concretamente sobre “la acción de la luz en las células blancas sanguíneas in vitro”. Trabajó con sangre de conejo, gato y cerdo. La sangre una vez extraída, se centrifugaba aislándose la capa leucocitaria, que se lava con solución de tyrode (PH 7,4) y se cultiva en un incubador a oscuras a 37,5ºC. En el trabajo se presenta un esquema de la naturaleza y secuencia de los cambios degenerativos observados, cuando las células se someten a examen microscópico con distintas fuentes de luz, todas incandescentes. Fueron eliminados aquellos factores que pudieran causar degeneración, como el aceite de inmersión o la temperatura alcanzada por el microscopio. Después de un corto tiempo de exposición a la luz del microscopio, se señalaron en el neutrófilo movimientos ameboides, que pronto cesaron y las células pasaron a ser esféricas. El núcleo se mantuvo turgente durante más tiempo en este proceso de degeneración. Las células gradualmente se fueron hinchando, en ocasiones hasta 3 veces el diámetro normal. El tipo de degeneración mostrada por el eosinófilo y basófilo era esencialmente igual a la del neutrófilo. Los monocitos sufrieron una serie de cambios casi idénticos a 85 RESULTADOS los que tuvieron lugar en las células anteriores, y a medida que aumentaba el movimiento browniano de las vacuolas celulares, la célula se redondeaba gradualmente, su contorno se hizo más visible, y la viscosidad del citoplasma disminuyó rápidamente. El pequeño e intermedio linfocito era seguido más fácilmente en el curso de este proceso, debido a su morfología característica, que los grandes linfocitos, más difíciles de identificar. El primer cambio visible en el linfocito fue un marcado aumento en el tamaño celular, de 1 a 2 veces su diámetro original. Hubo pocos cambios en el tamaño nuclear, pero el citoplasma pasó a ser francamente edematoso. Los datos obtenidos (163) indican que la luz ultravioleta, visible o infrarrojos ya sea la luz del día o de la lámpara del microscopio, fue el agente más probable causante de la degeneración. Con el fin de poner a prueba la posibilidad de que la luz fuese la causante, se prepararon una serie de 20 cultivos. Se inocularon con luz de día muy tenue y se colocaron en la incubadora, en oscuridad, lo más rápidamente posible. Toda manipulación posterior se hizo con luz muy tenue. La degeneración observada en estos cultivos coincide en todos los aspectos con la observada en cultivos anteriores. La exposición continuada a la luz no es esencial, ya que una exposición de muy corto tiempo (3 minutos) produce poco o ningún efecto en este tiempo, pero es suficiente para causar la muerte de las células en unas pocas horas. Se incorporó un filtro de calor especial que eliminaba gran parte de los rayos infrarrojos, y se observó que en estas condiciones la degeneración era aparentemente la misma que sin filtro de calor. El paso siguiente fue determinar las longitudes de onda responsables de la degeneración. El rango espectral de la luz fue dividido en intervalos de longitud de onda mediante la utilización de filtros de luz Wratten y Corning. Se utilizaron filtros azules, verdes y rojos, y se determinó el espectro de cada uno de ellos. Se demostró 86 RESULTADOS que la luz transmitida por cualquiera de estos filtros era causa de degeneración, y que los periodos de tiempo necesarios para la degeneración eran variables en diferentes cultivos para cualquier filtro. Al considerar la relación entre la degeneración de células in vitro y la degeneración de células in vivo bajo la acción de la luz, Earle dice:”… es difícil de establecer paralelismo, ya que las condiciones en los dos casos son muy distintas e imposibles de comparar”. Los puntos más importantes del trabajo de Earle (163) son: 1. En general, en el proceso de degeneración, hubo un período de latencia, durante el cual no se observó cambio alguno en las células, seguido de un período de estimulación en el que tiene lugar una aceleración enorme del movimiento celular. Esta segunda etapa se ha observado en todas las células, excepto en linfocitos, en los que puede ocurrir en un ligero grado. En una etapa terminal, la célula puede morir por hinchazón o coagulación. 2. El factor causante de esta degeneración es la exposición del cultivo a la luz, por ejemplo, durante el examen microscópico. 3. Por reducción de la parte infrarroja del espectro, se indicó que el efecto no se debió a una coagulación por calor. 4. Esta degeneración también se producía en completa ausencia de longitudes de onda ultravioleta. 5. Esta degeneración fue causada por la luz dentro de cada una de las tres zonas de longitud de onda siguientes: a-­‐ 430-­‐550 nm, infrarrojos, b-­‐ 475-­‐ 630 nm; 690 nm de infrarrojos, c-­‐ 600 nm de infrarrojos. 6. La degeneración de los leucocitos también tenía lugar en las gotas de sangre entera, lo que indica que no era dependiente de factores adicionales como la centrifugación, disminución de temperatura o presencia de solución salina anormal. 7. Se observó, sin embargo, que los leucocitos en cultivos de sangre entera, requerían un tiempo considerablemente más largo para su degeneración en virtud de la acción de la luz. Esto se considera como posiblemente 87 RESULTADOS debido a un daño celular durante la centrifugación y subsiguiente manipulación. En 1930 merecen ser recordados dos autores, Claude E. Forkner (164) y Louise Pearce (165). Forkner (164), que desarrolla su trabajo en el Instituto de Patología de Freiburgo, Alemania, estudia sobre el origen de los monocitos en ciertos ganglios linfáticos, y su relación genética con otras células del tejido conectivo. La finalidad del trabajo de Forkner es determinar la posición de los monocitos en los ganglios linfáticos y cuales son sus células progenitoras. Forkner, en una comunicación anterior (166), había mostrado que los ganglios linfáticos de diferentes partes del cuerpo exhiben marcadas diferencias citológicas, y que el monocito puede ser encontrado como constituyente normal en todos los ganglios linfáticos del conejo, excepto en el mesenterio. El origen del monocito ha sido una cuestión muy debatida. Existían diversas teorías, según las cuales, los monocitos surgen de mieloblastos, linfocitos, histiocitos, endotelio o células del sistema reticuloendotelial. Sin embargo, ninguno de los experimentos realizados pudo demostrar su origen. Para realizar el estudio se utilizaron conejos, cobayas y ratas. La tinción empleada fue trypan azul, y se indican los protocolos seguidos en el manejo de los animales. Se describen y hacen observaciones de los tipos celulares encontrados: a) Células primitivas no diferenciadas, también llamadas mesenquimales. Son indiferenciadas y poseen la capacidad potencial para la formación de diversos tejidos mesenquimales. Son pálidas, de núcleo redondo u oval y moderado citoplasma sin gránulos. Se pueden encontrar en cualquiera de los órganos que forman la sangre. b) Linfocitos. Aparecen como pequeños, intermedios y grandes. Núcleos redondos, ovalados o en herradura. Cromatina característica en gránulos 88 RESULTADOS gruesos y citoplasma claro, hialino. Poseen un tipo característico de motilidad, moviéndose con el núcleo en la parte delantera de la célula. c) Monocitos. Otros términos aplicados son grandes mononucleares, células tradicionales de Ehrlich, leucocitos endoteliales e histiocitos. Son células grandes en general, con núcleo preferentemente en forma de riñón o herradura y cromatina de estructura delicada. d) Pre-­‐monocito. Nuevo término que se propone con el fin de designar una de las etapas en el desarrollo del monocito, intermedia entre el monoblasto y los monocitos maduros. Se caracteriza por su presencia constante en los ganglios linfáticos de los conejos, excepto en el tejido linfoide del mesenterio intestinal. e) Monoblasto. El precursor del monocito. Se considera derivado de una célula indiferenciada primitiva y precursora del pre-­‐monocito. f) Macrófagos mesenquimales. La terminología empleada para designarla es confusa: macrófago, clasmocito, célula adventicia, célula errante en reposo, célula reticular, célula reticuloendotelial, leucocito endotelial o histiocito, entre otras. El escritor revisa críticamente la literatura sobre el tema y propone retomar el término descriptivo “macrófago”, que fue aplicado por primera vez a estas células por Metchnikoff. Son células de gran tamaño, núcleo redondo u ovalado con una delicada red de cromatina y cuyo citoplasma contiene pocos o muchos cuerpos irregulares en formas y tamaños, que representan restos celulares (debris) fagocitados. El trabajo de Forkner (164), nos muestra el sitio de origen y el desarrollo de monocitos en virtud de condiciones fisiológicas: 1. Demuestra que se forman en ciertos órganos hematopoyéticos específicos, y que esto es igual para todos los demás elementos estructurales de la sangre. 2. Encuentra que estas células derivan de células primitivas indiferenciadas y pueden diferenciarse claramente las diversas etapas de desarrollo. 89 RESULTADOS 3. No se niega que los monocitos en condiciones anómalas puedan ser anormalmente producidos en lugares no habituales, así pueden surgir en diversos órganos y tejidos del cuerpo, y esto es igualmente cierto para linfocitos y granulocitos. 4. En cuanto a la relación de los monocitos con el sistema reticuloendotelial, la única conclusión que puede hacerse de manera segura es que no se tiñen como lo hace el sistema reticuloendotelial con el método habitualmente utilizado. Louise Pearce y Albert E. Casey (165), investigadores del Instituto Rockefeller, publican sus estudios en 1930 sobre “citología en la sangre del conejo. Observaciones en linfocitos y monocitos en grupos de conejos normales”. El trabajo forma parte de un serie de estudios que recogen los resultados de los exámenes repetidos en la sangre de cinco grupos de conejos, respecto a las determinaciones de eritrocitos, hemoglobina, el total de células blancas, neutrófilos, basófilos, eosinófilos y recuentos celulares, y que fueron publicados de forma sucesiva, (167)(168). El período estudiado fue en total de 20 meses, desde el 24 de octubre de 1927 a 24 de julio de 1929. Un total de 45 conejos, en 5 grupos de 9 conejos, se sometieron a análisis semanales sanguíneos, y los resultados fueron analizados cada semana en base a los valores medios de grupo. Los resultados son expuestos en orden cronológico: 1. En el grupo I, se examinaron a intervalos semanales desde el 24 de octubre de 1927 a 24 de junio de 1928. Los valores referentes a los linfocitos se incrementaron al final de este período, observándose una caída pronunciada a finales de febrero y primera quincena de marzo. Los monocitos se mantuvieron por encima del valor normal. 2. En el grupo II, desde el 29 de marzo a 19 junio de 1928, las curvas que representan los valores numéricos de linfocitos y monocitos ponen de 90 RESULTADOS manifiesto con claridad valores inferiores en ambos tipos de células, en comparación con el grupo I. 3. Grupo III, de 20 de septiembre a 22 de noviembre de 1928. Se observan valores tanto en linfocitos como en monocitos del mismo orden que en el grupo II. El total de células blancas describe una tendencia a la baja. 4. Grupo IV. Desde el 27 de noviembre a 19 de junio de 1929. El nivel general en los valores medios de linfocitos es comparable a los resultados en los grupos II y III. En diciembre, la tendencia de la media fue a disminuir, encontrándose un bajo nivel en enero, febrero y marzo. En abril tuvo lugar un aumento brusco, que fue seguido de una disminución. La curva de monocitos es comparable a la de los grupos II y III. A partir de niveles muy bajos en diciembre y primeros de enero, la curva comienza a subir en febrero, marzo y abril. 5. Grupo V. De 29 de diciembre de 1928 a 21 de junio de 1929. Los linfocitos comienzan a subir en febrero, y a partir de entonces prevalecen a un nivel relativamente alto. La curva de monocitos en los grupos IV y V es muy similar. En general, la tendencia de linfocitos y monocitos a niveles más altos, fue menos pronunciada en monocitos que en linfocitos. El período de mayor irregularidad en los valores medios de linfocitos fue a finales de los meses de invierno y primavera en los dos años. Con los monocitos, las fluctuaciones tienen lugar en otoño de 1927, primavera y principios de verano de 1928 y el final del invierno, primavera y principios de verano de 1929. Los resultados (165) basados en el estudio de las tendencias de los valores medios del grupo, obtenidos a partir de las observaciones semanales consecutivas no mostraron evidencia de una relación numérica constante entre linfocitos y monocitos. 91 RESULTADOS 4.3. LA CUARTA DÉCADA DEL SIGLO XX La década de los años treinta, desde 1931 a 1940, constituye uno de los puntos de inflexión fundamentales en la historia de Europa y América. En ella tienen lugar la crisis del liberalismo y la democracia, el crecimiento del fascismo, la irrupción de las masas y la expansión del estado (219)(220). El crecimiento desmesurado de los años 20 desemboca en una crisis profunda que provoca la quiebra de los bancos y la falta de capitales para la industria, y todo de forma brusca. El miedo detiene la inversión, el paro aumenta, el consumo desciende, los precios caen y se descapitaliza la banca. La llamada Gran Depresión se originó en EEUU a partir de la caída de la bolsa en Octubre de 1929, y rápidamente se extendió a casi todos los países del mundo, prolongándose hasta principios de la década siguiente, la década de los cuarenta (169). En 1931 la crisis financiera es definitiva y muy profunda, pero comienzan las medidas para salir de ella. El ministro de economía de Gran Bretaña, John Keynes renuncia al patrón oro y el Estado se hace intervencionista en la economía aumentando el gasto público. El llamado Keynesianismo (170) es la política que ayudará a la recuperación de Europa y gran parte del mundo. Será una recuperación lenta, y los efectos de la crisis llegarán hasta el año 1939. En 1932, el Partido Nacional Socialista consigue la victoria en las elecciones generales de Alemania, y en 1933, Adolf Hitler, canciller de Alemania, obtienen plenos poderes en el país. Con la invasión de Polonia por parte de la Alemania nazi, el Reino Unido y Francia le declaran la guerra al Tercer Reich, iniciándose la Segunda Guerra Mundial. En lo relacionado con la ciencia, a finales de los años 30 se incorporan nuevos avances técnicos como la electroforesis, la cromatografía en papel, la ultracentrifugación y el microscopio electrónico. En 1931 Ernst Ruska y Max 92 RESULTADOS Knoll construyeron el primer microscopio electrónico (171) y en 1935, W.M. Stanley aisló el primer virus. Heidelberger y Forrest E. Kendall en 1936 purifican anticuerpos a partir de sueros por disociación de precipitados. El principal descubrimiento de Heidelberger (172)(173), que los anticuerpos no son sino proteínas, fue de gran importancia en el desarrollo de los estudios inmunológicos. La búsqueda en Scopus con la palabra lymphocyt* da como resultado un total de 70 documentos, un volumen de publicaciones igual al registrado durante las tres décadas anteriores, lo que supone un porcentaje del 0,0125 % con respecto al total (558.754 documentos en el siglo XX). Se observa en la gráfica acompañante, como Estados Unidos encabeza la lista en número de publicaciones seguido por Alemania. GRÁFICO 3 93 RESULTADOS En 1931, Bruce K. Wiseman, publica dos trabajos fruto de sus experimentos en los Laboratorios del Instituto Rockefeller para la investigación médica. El primero de ellos (enero de 1931) tiene por título “La inducción de linfocitosis e hiperplasia linfoide mediante la administración parenteral de proteínas” (174). Justifica su trabajo alegando que los métodos de estimulación del tejido linfoide mediante sustancias químicas han sido insatisfactorios en sus resultados, que los efectos de las drogas, dietas y medios físicos han sido ya revisados, pero que, sin embargo, la efectividad de la proteína como sustancia estimulante del aumento de linfocitos en sangre, cuando se inyecta por vía intravenosa durante períodos de tiempo prolongados, no se ha observado hasta ahora. Refiere la experiencia de Ehrich (175), quien inyectando estafilococos muertos vía intravenosa, obtiene una linfocitosis que alcanza su máximo entre los días 10 y 14, para luego volver a la normalidad aun con la persistencia de las inoculaciones. Observó una hiperplasia del bazo y ganglios linfáticos, pero no advirtió una clara diferenciación entre monocitos y linfocitos. Las observaciones que Wiseman recoge fueron incidentales a un estudio del efecto del extracto de embriones de pollo en las células del epiplon de conejo. Se utilizaron conejos adultos que fueron sometidos a observación durante 4-­‐13 semanas, a fin de asegurar en lo posible la ausencia de infección, antes de ser empleados experimentalmente. Se controlaba en ellos la temperatura, y se hacían numerosos recuentos sanguíneos. El estudio histológico post-­‐morten era la rutina habitual. A 12 animales se les administró la proteína por tres rutas diferentes (intraperitoneal, intravenosa y subcutánea), uno recibió suero normal de caballo y otro una solución salina normal vía intravenosa. Se realizó esplenectomía a dos animales, a uno de ellos se le había administrado albúmina de huevo y el otro era un control al que no se había inyectado nada. En el examen de los resultados (174) se observó que las tres formas de administración de proteínas provocaban hiperplasia del bazo, acompañado de una linfocitosis periférica que oscilaba de un 23% a 139%. La mayor respuesta 94 RESULTADOS parece obtenerse por vía intravenosa, y probablemente es la albúmina de huevo la sustancia más potente (el autor considera necesario hacer un estudio en una serie mayor para comprobar este punto). Los resultados muestran un aumento casi específico de linfocitos, que se hizo evidente por primera vez entre los días 7 y 10, observando que la respuesta linfocitaria no era dependiente del bazo y que las inyecciones de soluciones salinas normales no provocan cambios sanguíneos fuera de los límites normales para conejos. Los cambios cualitativos de los linfocitos fueron muy marcados, con un aumento del linfocito intermedio-­‐grande, células con citoplasma basófilo y gran número de mitocondrias. No hubo cambios definidos en células rojas, hemoglobina, eosinófilos y leucocitos basófilos. El bazo y ganglios linfáticos mostraron una histología acorde con la hiperplasia de estos tejidos. En los nodos de los animales inyectados tuvo lugar un aumento del tamaño de los nódulos corticales y del número de figuras mitóticas dentro de los centros germinales. Muchos de los nódulos fueron confluentes, dando como resultado un aspecto de hiperplasia difusa del tejido linfoide, en contraste con la clara arquitectura cortico-­‐medular del nodo normal original del mismo animal. En el bazo, los folículos agrandados de Malpighi se podían ver a través de la cápsula esplénica, muchos medían más de 1 cm de diámetro, y microscópicamente se observó una gran hiperplasia con muchas figuras mitóticas. No hubo evidencia de proliferación del timo, de lo que se desprende, que el timo no participó de la hiperplasia general del tejido linfático, y favorece la duda sobre la identidad de las pequeñas células tímicas como verdaderos linfocitos sanguíneos. El examen histológico de los restantes órganos no mostró cambios significativos. La explicación del autor (174) sobre estos hechos es que la proteína puede actuar bien como una sustancia madurativa o como un quimiotáctico, y dado que el número de linfocitos no aumenta hasta el día 7, es difícil creer el efecto quimiotáctico, y piensa que la reducción o neutralización de la proteína extraña a través de las barreras naturales del cuerpo, pudiera ser una de las funciones de los linfocitos, anunciando futuros estudios en este campo. 95 RESULTADOS En mayo del mismo año 1931, Wiseman escribe sobre los “criterios de madurez o edad de los linfocitos en sangre periférica” (176). Este autor considera de gran importancia poder determinar en el análisis sanguíneo de una enfermedad el estado de madurez de sus elementos celulares: “Observando las variaciones en ciertas características citoplasmáticas y nucleares, se puede estimar la edad de estas células, obteniéndose así una valiosa información”. El índice de variación nuclear en linfocitos, estudiado por Arneth, no tuvo el mismo reconocimiento ni aceptación que las observaciones realizadas en el leucocito polimorfonuclear, existiendo algunas razones por las que la búsqueda de criterios reconocibles en la maduración del linfocito no ha tenido éxito. En primer lugar, porque entre las células mononucleares llamadas linfocitos, algunos, sino todos, surgen como linfocitos inmaduros (linfoblastos), y maduran de forma ordenada, degenerando sin desdiferenciación. Otra dificultad radica en el carácter físico del linfocito, en el que aparentemente no hay elaboración de estructuras específicas en el citoplasma. Así por ejemplo, el desarrollo de gránulos citoplasmáticos en el granulocito, y de hemoglobina en las células rojas sanguíneas, han servido como criterios objetivos sobre los cuales se ha basado la identificación de las fases madurativas. La falta de datos comparables ha sido un obstáculo para el reconocimiento de la edad de los linfocitos, por lo que resulta evidente la necesidad de buscar criterios de carácter diferente. Una tercera dificultad es la que se encuentra probablemente en la fisiología de los tejidos linfoides, siendo muy probable que muchos de los linfocitos realicen su función dentro del tejido en que surgen. Podemos esperar que linfocitos funcionalmente maduros se entremezclen con células de diversos grados de inmadurez en los ganglios linfáticos. Y, por último la incertidumbre que existe en relación a la estructura exacta del tejido linfoide, contribuye a la complejidad del problema. En este sentido, hay que decir que, aunque la mayoría de autores reconocen el centro germinal de Flemming como el foco de la linfogénesis, en cambio otros lo interpretan como zonas reactivas no partícipes en la génesis del linfocito. 96 RESULTADOS Una vez hechas todas estas consideraciones, Wiseman (176) establece posibles criterios de edad celular: • Basofilia. En 1898, Pappenheimer había señalado que el citoplasma de mielocitos jóvenes era muy basófilo; desde entonces, la asociación de la basofilia con la juventud de los granulocitos ha sido señalada por muchos investigadores. El mismo hecho se ha observado en las células rojas de la sangre; el “reticulado” de los pequeños glóbulos rojos, no es más que una precipitación de sustancia basófila, y los recuentos de células reticuladas son un ejemplo de la utilización de la presencia de basofilia como índice de inmadurez. Una cuidadosa aplicación de este criterio en linfocitos no ha tenido lugar. • Mitocondrias. Las mitocondrias están presentes en grandes cantidades durante la maduración de células sanguíneas jóvenes. Cowdry (100) acepta que la senescencia se asocia con una disminución de mitocondrias en el citoplasma de las células en general. • Tamaño celular. Los hematólogos han utilizado con gran frecuencia el tamaño como criterio de edad de los linfocitos. Sin embargo, la incertidumbre no está resuelta, ya que unos consideran al linfocito de gran tamaño, y otros al pequeño linfocito como el más joven, mientras un tercer grupo sostiene que el tamaño no es criterio de edad en absoluto. Es significativo que estas opiniones se relacionan la mayoría de las veces con la técnica de fijación y tinción utilizada. • Otras características. Motilidad, vacuolas, contenido del núcleo en cromatina, proporción entre núcleo y citoplasma y forma del núcleo, son a veces utilizados como criterios de madurez. Cuando se evalúan todos estos parámetros (176), parece ser que, por analogía con otras células, la estimación del número de mitocondrias y el grado de basofilia, son criterios válidos para calcular la edad del linfocito. De ellos, la basofilia es el más fácil y preciso de determinar. Para su estudio se sigue el método de Wrigth y Giemsa. Después de acostumbrar el ojo a los 97 RESULTADOS distintos tipos de tonos azules del citoplasma, se pueden dividir arbitrariamente las células en tres tipos: muy basófila (tipo Y), moderadamente basófila (tipo M), y poco o nada basófila (tipo O). En sangre normal, tanto humana como de conejo, la composición porcentual de basófilos es relativamente definida y estable. La fórmula (Y-­‐M-­‐O) que expresa los grados de basofilia en sangre humana es 5-­‐ 49-­‐ 46 y en conejos 4,7-­‐ 48-­‐ 47,3. Sin embargo, el número de ejemplares estudiados no son suficientes para aceptar la fórmula como representativa de la fisiología normal, pero sí es válida para este trabajo. El estudio en sangre obtenida de hiperplasias fisiológicas o patológicas del tejido linfoide, debería reflejar un aumento de los elementos inmaduros, por ello se llevó a cabo el examen sanguíneo de animales en los que la hiperplasia linfoide se había inducido artificialmente mediante inyecciones parenterales de proteína extraña. Tuvo lugar un aumento de los ganglios linfáticos y bazo, y aumentaron también las formas basófilas en sangre, concluyéndose que la basofília es una medida de la falta de madurez del linfocito. El estudio del contenido en mitocondrias se realizó con tinción supravital, en los mismos grupos anteriores y con la misma gota de sangre empleada y el contenido de mitocondrias se determinó bien por recuento, o por estimación aproximada. Se clasificó de la siguiente forma: clase Y, muchas mitocondias (> 20), clase M, valor intermedio (5-­‐20 mitocondrias), clase O (0-­‐5 mitocondrias). Se observó que cuando la tendencia de la linfocitosis es hacia una mayor basofília citoplasmática, también hay el correspondiente cambio hacia un mayor contenido mitocondrial. El tamaño celular, está muy influido por la técnica empleada que puede distorsionar la célula por factores mecánicos (la extensión) o factores químicos (fijador que contrae la célula). Se observó que formas inmaduras pueden ocurrir sin cambios notables en el tamaño, pero que en algunos casos, el aumento de las formas jóvenes se asoció también con un aumento de células de tamaño grande, y se estudió al linfocito de tamaño grande para determinar su edad valorando su grado de basofilia y cantidad de mitocondrias. Los 98 RESULTADOS resultados indicaron que la mayoría tienen una marcada basofilia y contienen gran número de mitocondrias. En general, en los linfocitos grandes, a mayor cantidad de célula ocupada por el núcleo más basófilo es el citoplasma. Figura 12. Criterios de Madurez de linfocitos. 1- joven linfocito grande de la sangre periférica del conejo. Observar el carácter granular y el grado extremo de basofilia del citoplasma (célula tipo Y). 2- un pequeño linfocito mostrando las mismas características. 3- linfocito intermedio (célula tipo M). 5- gran linfocito de sangre periférica de conejo (célula tipo O). 8- célula roja sanguínea. Tomado de: Wiseman, 1931 (176) En cuanto a la relación entre linfocitosis e inmadurez celular, el postulado de Arneth de que el número total de granulocitos en sangre no es necesariamente un criterio de actividad de la médula ósea, ha sido ampliamente confirmado. Si aceptamos que a mayor basofilia mayor inmadurez, es necesario comparar el porcentaje de basófilos con el número total de células circulantes, ya que el total de linfocitos circulantes, no es un criterio de un porcentaje aumentado de células jóvenes. La expresión de Arneth “giro a la izquierda”, tuvo su inicio de la observación de que en condiciones patológicas se produce un aumento del porcentaje de granulocitos que muestran un menor número de lobulaciones en el núcleo, es decir, un aumento de células más jóvenes. En un sentido más amplio “desviación o giro a la izquierda”, viene a expresar un cambio hacia las 99 RESULTADOS etapas más jóvenes en la maduración de algunos otros elementos de la sangre. Si se produce un aumento de elementos más viejos la expresión utilizada es “giro a la derecha”. En definitiva, el estudio de la sangre de conejos con tejido linfático normal e hiperactivo, revela en este último un aumento de linfocitos con citoplasma fuertemente basófilo y aumento en el número de mitocondrias. Según Wiseman (176): “… esto indica que el grado de basofilia citoplasmática y el contenido mitocondrial puede servir como criterio del grado de madurez de los linfocitos, y que la basofilia es el más evidente y fiable indicador de la juventud de las células. El número total de linfocitos circulantes en un momento dado, no es necesariamente un índice de actividad linfoide”. Louis Pearce y Albert E. Casey, continúan con sus investigaciones iniciadas en años anteriores, sobre la citología sanguínea del conejo, presentando tres de sus trabajos procedentes del Instituto Rockefeller. El primero de ellos, en febrero de 1931, sobre la “relación de las células sanguíneas en grupos de conejos normales con respecto al tiempo” (177), comienza con un breve resumen de los resultados de sus trabajos anteriores (I, II, III, IV y V): “… se informó de observaciones repetidas en las células de la sangre periférica en conejos normales… examinadas durante largos períodos de tiempo, se demostró claramente que el contenido en células y en hemoglobina fluctúa en mayor o menor grado. Estos resultados se obtuvieron en condiciones experimentales uniformes. El análisis demostró además que las variaciones en los niveles de ciertas clases de células fueron acompañadas por variaciones similares u opuestas en otras”. Con la finalidad de determinar la importancia de estas tendencias y fluctuaciones en los diversos tipos celulares Casey realizó un análisis estadístico que recoge en este documento (177) . Se utilizaron 45 conejos normales distribuidos en 5 grupos de la siguiente forma: los grupos 1, 2, 3 y 4 estaban formados cada uno por 10 conejos, el grupo 5 por 5 conejos. Eran conejos adultos, de aproximadamente 8 meses de edad, todos aparentemente libres de enfermedad. Se realizaron 100 RESULTADOS determinaciones del hemograma y de las diferentes células blancas, a intervalos semanales desde octubre de 1927 a junio de 1929. Los coeficientes de correlación promedios para todos los cinco grupos fueron utilizados como la base, y se calculó un coeficiente de correlación múltiple, en el que el total de las células blancas de la sangre fue la variable dependiente, y los neutrófilos, eosinófilos, basófilos y linfocitos, las variables independientes. El coeficiente de correlación múltiple encontrado fue =1 (R = 1,00), lo que significa que no se había cometido ningún error en el cálculo. Ya que R = 1,00 cuando las células blancas de la sangre son la variable dependiente, se explica por el hecho de que el valor de las células blancas es igual a la suma de los valores para las distintas clases de células blancas. Hay que recordar que este estudio estadístico de las tendencias y variaciones en sangre periférica de conejos normales, se llevó a cabo con el propósito de determinar la existencia de asociación o falta de asociación entre las fluctuaciones observadas en los niveles celulares y el contenido de hemoglobina. Es aceptable que el número medio de cualquiera de los constituyentes de la sangre pueda variar con el tiempo. Por ejemplo, el nivel numérico de dos clases de células puede tener oscilaciones bruscas de corta duración, en que los dos niveles fluctúan en la misma u opuesta dirección, o bien no tienen relación aparente entre sí. A este cambio en breve período de tiempo hemos llamado “fluctuación u oscilación” para diferenciarlo de “tendencia” que se utiliza cuando el cambio es progresivo, en meses. Las determinaciones efectuadas se muestran en una serie de tablas. En la tabla I se muestra las correlaciones simples de orden cero sobre las determinaciones semanales en los cinco grupos de conejos normales. La tabla II representa un intento de medir las relaciones entre las células de los diversos grupos de animales, respecto a fluctuaciones a corto plazo. En la tabla III encontramos un resumen del análisis del coeficiente de correlación combinado (método Z aplicado a cinco grupos de animales normales). 101 RESULTADOS Encontramos que las células blancas de la sangre están fuertemente relacionadas con neutrófilos y linfocitos, tanto en lo que respecta a tendencia general y oscilación. La relación también existe con los movimientos en monocitos, sangre roja y basófilos, en el orden nombrado. Los niveles de células rojas varían de semana a semana, y de mes en mes en todos los grupos de animales. Estas variaciones están acompañadas de cerca por variaciones en basófilos, linfocitos, hemoglobina y monocitos, en este orden. Uno de los puntos más sorprendentes es el hecho de que el contenido en hemoglobina, no siempre ha seguido el de los valores de células rojas de la sangre. Casey (177) establece como relaciones significativas las siguientes: 1. La cantidad de glóbulos rojos, la cantidad de hemoglobina por mm3 y número de linfocitos. Hubo una relación inversa entre la cantidad de hemoglobina y el número de monocitos. 2. De las células blancas sanguíneas con las variaciones en células rojas y contenido de hemoglobina. Las variaciones numéricas en el total de células blancas, se asociaba con variaciones similares en el grupo de neutrófilos, linfocitos, monocitos, basófilos y eosinófilos. 3. Glóbulos rojos, linfocitos y basófilos como un grupo, eosinófilos y monocitos como otro grupo, y el contenido de hemoglobina y neutrófilos como un tercer grupo, describen un cambio definitivo desde un máximo a un bajo valor numérico, durante los dos años del período de observación. Respecto a los linfocitos, Casey encontró que: “… no tenían relación significativa con los otros elementos de la sangre, en lo que se refiere a fluctuaciones a corto plazo, excepto, tal vez, con basófilos y células rojas de la sangre. Sin embargo había una relación negativa con neutrófilos y eosinófilos”. Respecto a la tendencia: “los linfocitos estuvieron relacionados positiva y significativamente con las células rojas de la sangre, neutrófilos y basófilos, pero en absoluto con la hemoglobina y monocitos. La única diferencia significativa en las relaciones 102 RESULTADOS intercelulares de los linfocitos tanto en tendencia como en oscilación ocurrió con los neutrófilos”. En marzo de 1931. Pearce y Casey escriben sus “observaciones en conejos inoculados con una neoplasia maligna trasplantable” (167). El objeto de las observaciones en conejos normales, realizadas con anterioridad, no era sólo obtener información sobre el carácter del cuadro sanguíneo con respecto a condiciones ambientales, sino además proporcionar un fondo para observaciones similares en experimentos con diversos agentes patógenos como Treponema Pallidum, virus y tumores malignos trasplantables. En el presente trabajo se exponen los resultados de numerosas observaciones en sangre periférica, de conejos inoculados con un tumor maligno trasplantable. Se estudió un total de 78 animales, conejos adultos de 6-­‐8 meses al tiempo de la inoculación. El curso de la enfermedad fue seguida muy de cerca en cada animal, con exámenes clínicos frecuentes, y el experimento se dio por finalizado 8-­‐9 semanas después de la inoculación, cuando los animales que sobrevivieron fueron sacrificados. Los resultados han sido considerados con respecto al carácter del proceso tumoral determinado en la autopsia y se presentan en una serie de gráficos y tablas contenidos en el texto, donde se recogen las desviaciones medias porcentuales, llamadas valores estándar, de los elementos de la sangre estudiados en semanas sucesivas: células rojas, hemoglobina, total de células blancas, neutrófilos, basófilos, eosinófilos, linfocitos y monocitos Los conejos quedan clasificados en cuatro grupos según su evolución: los que murieron a causa de la enfermedad maligna (muertes), los que continúan el experimento, los que probablemente se han recuperado (recuperaciones probables) y los conejos recuperados en los que no se encontró tumor (recuperaciones). Respecto a las células rojas de la sangre y hemoglobina, los valores de los casos con probables recuperaciones y recuperaciones, están por encima de la línea base a lo largo de la observación, mientras que el de muertes y muertes probables caen por debajo del valor normal. La curva de las recuperaciones 103 RESULTADOS muestra una tendencia hacia el mantenimiento de los valores, más alta después de la inoculación. Figura 13. Valores del numero de linfocitos según la evolución de la enfermedad. D: muerte; PD: probable muerte; PR: probable recuperación; R: recuperación. Las curvas indican que los valores medios de linfocitos en estos cuatro grupos de conejos antes de la inoculación fueron variables. Tomado de Pearce y Casey, 1931 (167). Antes de la inoculación el conteo total de células blancas de todos los cuatro grupos de animales variaron de 5-­‐25% por debajo del valor estándar de 9.560 células por mm3. Después de la inoculación, la tendencia en todos los grupos fue hacia valores mayores. Esta tendencia fue más marcada en el caso de los grupos de fatal desenlace, en los que se encontraron los valores más altos durante las últimas tres semanas. En el caso de recuperaciones probables, los resultados mostraron una tendencia general hacia el aumento de células blancas, pero no a niveles tan altos como en los grupos de fatal desenlace. En animales recuperados los glóbulos blancos totales mostraron un ligero aumento después de la inoculación, que asciende aproximadamente al 15% por encima del nivel de pre-­‐inoculación. 104 RESULTADOS En las observaciones relativas a los linfocitos, los niveles medios en los cuatro grupos de conejos, antes de la inoculación fueron variables, con límites que se extienden entre el valor estándar de 3.050 células por mm3, en el caso de muertes probables, a aproximadamente un 20% por debajo de esta cifra en el caso del grupo recuperado. Después de la inoculación, se observó una caída en la media del número de linfocitos en dos grupos, en los que murieron y en las probables recuperaciones, mientras que en los otros grupos, se encontró poco o ningún cambio. Desde este momento, los linfocitos del grupo de desenlace fatal, mostraron una disminución numérica constante. En el caso de las muertes probables, no hubo cambio significativo hasta la sexta semana después de la inoculación; en este momento, hay una disminución moderada del número de linfocitos, que persistió hasta el final del período de observación. Los resultados en las recuperaciones probables y las recuperaciones son menos definidos y hubo una clara tendencia para el aumento sobre el nivel de pre-­‐inoculación, según manifiestan los autores (167): “al parecer, el número medio de linfocitos en sangre periférica, tiende a ser algo menor en los conejos que sucumben a la enfermedad neoplásica. Por otro lado, los valores medios de linfocitos, tienden a ser ligeramente mayores en conejos en los que el proceso es leve y en aquellos que se recuperan”. En definitiva, cuando el proceso tumoral marcó gravedad, el número de monocitos, neutrófilos y el total de células blancas de la sangre, eosinófilos y basófilos aumentó, en cambio los linfocitos disminuyeron, y no hubo cambios destacados en la sangre de los conejos que se recuperaron del tumor, cuando los resultados se consideran en todo el periodo de observación. Un tercer artículo obra de Casey y Pearce es publicado en junio de 1931, sobre la citología sanguínea del conejo: “La sangre de conejos normales como índice de resistencia a neoplasias trasplantables” (178). Los datos se obtuvieron a partir de dos grupos de conejos inoculados con el tumor de conejo Brown-­‐Pearce, un epitelioma que surgió espontáneamente en la piel del escroto de un conejo 10 años antes, y se ha mantenido pasándolo de 105 RESULTADOS animal a animal por inoculación intratesticular en unas 100 generaciones. Se estableció un primer grupo de 78 conejos que se dividió en 9 subgrupos para facilitar su estudio y seguimiento. Una segunda serie de conejos (13 conejos) fue utilizada como testigo, a fin de comprobar la precisión de los métodos desarrollados en el primer grupo. La técnica hematológica empleada incluía determinaciones del total de células rojas y blancas con pipetas estandarizadas, hemoglobina por el método Newcomer y conteo diferencial con rojo neutro supravital. El experimento se mantuvo durante dos meses, se sacrificaron los animales y se realizó la autopsia con especial referencia al carácter y la distribución de cualquier tumor. Con el fin de dar un valor numérico a la malignidad tumoral en cada animal, se clasificaron en: recuperaciones probables (33% de malignidad), muertes probables (67% de malignidad), muertes (100% de malignidad) y recuperaciones (0% de malignidad). Si se toma en consideración simplemente la presencia del tumor, el número de posibles focos tumorales se fijó arbitrariamente en 20, y la distribución de animales sobre esta base, es la siguiente: recuperaciones (0 focos), recuperaciones probables (1-­‐4 focos), muertes probables (4-­‐12 focos), y muertes (4-­‐20 focos). El método de análisis empleado por los autores (178), determinante de la relación de las células sanguíneas en el curso de la enfermedad maligna, ha sido construir una tabla de correspondencias simple para cada tipo de célula estudiada. Se calcularon los coeficientes de correlación simples para cada uno de los 9 grupos, entre los valores anteriores a la inoculación, y la estimación numérica usando los dos métodos de la estimación de la malignidad del tumor. En el trabajo de Casey y Pearce se ilustra la relación de varias clases de células blancas con el curso de la enfermedad. Los conejos más resistentes, tienen valores de linfocitos pre-­‐inoculación de aproximadamente 2000/ml. Hubo una tendencia a incluir los animales con valores inferiores a 2000/ml en la categoría de susceptibles. También se refleja la media de cada tipo de célula blanca por cada 100 células, es decir los valores relativos antes de la 106 RESULTADOS inoculación. Los conejos susceptibles, tenían valores de neutrófilos, basófilos y eosinófilos relativamente bajos antes de la inoculación, mientras que los más resistentes tenían valores altos. La tendencia de los linfocitos contrasta fuertemente con la de eosinófilos, manteniendo una relación inversa con éstos. Los autores (178) trataron con este estudio dar un esbozo de las relaciones en las células sanguíneas antes de la inoculación tumoral, con el curso de la enfermedad inducida por una neoplasia maligna trasplantable, y predecir, en la generalidad de los casos, el resultado final de la enfermedad a partir de un estudio citológico de sangre previo a la inoculación tumoral. Las conclusiones a las que llegan, son las siguientes: 4-­‐ Los valores medios de los recuentos pre-­‐inoculación en los animales más resistente a la enfermedad se encontraban dentro de valores normales, tanto en glóbulos rojos, hemoglobina y células blancas, en cambio los linfocitos presentaron valores bajos. 5-­‐ Los animales más susceptibles fueron los que tenían antes de la inoculación, valores de células rojas superiores a 5.500.000 o inferiores a 5.000.000 por ml, hemoglobina superior al 70% o inferior al 60%, valores de células blancas inferior a 6.000 o superior a 8.500 por ml, un número de eosinófilos bajo y un recuento de monocitos o linfocitos alto. Y aseguran: “a partir de los resultados parece posible poder predecir, con una precisión del 80-­‐90%, la resistencia individual o la susceptibilidad de los conejos al tumor, según los recuentos de las células sanguíneas antes de ser inoculados”. Franklin R. Miller, del Instituto Rockfeller, muestra interés por la histogénesis de las células plasmáticas, publicando en 1931: “El desarrollo inducido y la histogénesis de las células plasmáticas” (179). El conocimiento existente en este momento sobre las células plasmáticas no era muy extenso, indicaba que procedían del tejido conectivo y que su número aumentaba en varias circunstancias patológicas. Miller, inspirado en el estudio 107 RESULTADOS realizado unos años antes por Sabin y Doan (180) que informaron de la presencia de células plasmáticas en el epiplon de conejo tras la inyección intraperitoneal de proteína tuberculosa, pretendía determinar la eficacia de dicha proteína como sustancia estimulante de células plasmáticas, y estudiar así su histogénesis: “Si la célula plasmática es aceptada como entidad celular definida, debe tener un ciclo de maduración, al igual que los elementos mieloides y linfoides”, aseguraba Miller. El nombre de célula plasmática fue introducido por Waldeyer en 1875 (181), pero en su trabajo no queda claro si describe un tipo específico de célula. Un tipo celular definido fue reconocido por Cajal (año 1890) en el condiloma de la sífilis; las llamó células cianófilas. Unna en 1891 encontró unas células en las lesiones de la tuberculosis dérmica, a las que llamó células plasmáticas. Las describió con un citoplasma basófilo, esponjoso y granular, el llamado granuloplasma. De núcleo central o excéntrico, y de variable contenido en cromatina. El único criterio válido era la presencia constante de granuloplasma basófilo, y como resultado de ello, en la categoría de célula plasmática se incluyeron muchas células con citoplasma basófilo que probablemente derivaban de otras cepas celulares. Marshalkó (179), seleccionó ciertas características morfológicas como criterios esenciales de célula plasmática: “debe tener citoplasma basófilo con un área claro en el centro, ser redonda u ovalada, y el núcleo colocado excéntricamente, con la cromatina condensada cerca de la membrana nuclear y dispuesta radialmente”. Las células con estos criterios son conocidas hoy día como células plasmáticas maduras o células de Marshalkó. La discusión sobre el origen de estas células, se mueve entre los que creen que son derivadas de los linfocitos, ya sean hematógenos ó histógenos, y los que piensan que su origen es el tejido mesenquimal primitivo. Miller utilizó para estimular la producción de células plasmáticas tres tuberculoproteínas, dos solubles en agua y una soluble en álcali, y empleó 18 animales, 17 de ellos recibieron inyecciones intraperitoneales de una proteína, 108 RESULTADOS 2 de ellos inyecciones intravenosas y 3 por vía subcutánea. El animal restante, sólo inyecciones intravenosas. Al finalizar el experimento, los animales son sacrificados y estudiados por autopsia. En la autopsia, el epiplon generalmente mostró una reacción hiperplásica; estaba engrosado, con manchas lechosas y crecimiento de vasos y tejido fibroso. El omento era el sitio donde mayor proliferación de células plasmáticas había, pero también estaban presentes en hígado, bazo, ciego, y ganglios linfáticos dispersos. Algunos omentos estaban tan estimulados, que las células plasmáticas era el elemento predominante, y donde había un aumento de plasmáticas, los linfocitos también se incrementaron. Los animales que recibieron la mayor cantidad de proteína y por tiempo más prolongado, mostraron la mayor proliferación de plasmáticas, y la mayor variedad de etapas en el ciclo de la vida de éstas. La mayoría de células plasmáticas, tanto en tejidos estimulados como normales eran del tipo Marshalkó, sin embargo siempre se veían células con características de inmadurez que, a medida que maduraban, se acercaban a los criterios de Marshalkó de células plasmáticas. Figura 14. Células plasmáticas 5- sección de epiplón de conejo mostrando un foco de células plasmáticas maduras típicas. 6- sección de epiplón de conejo mostrando células plasmáticas en varias fases de maduración. Tomado de Miller, 1931, (179) 109 RESULTADOS Miller (179) observó que mediante la técnica supravital, las células plasmáticas se diferencian de los linfocitos por su citoplasma gris más oscuro, núcleos excéntricos, variaciones en el tamaño y distribución de mitocondrias, y añade: “Las plasmáticas se encuentran donde se desarrollan los linfocitos, y el efecto estimulante de las proteínas tuberculosas hace que se formen muchos pequeños linfocitos, nuevos folículos linfáticos, así como células plasmáticas”. Y advierte también sus diferencias con los pequeños linfocitos: “Las células plasmáticas jóvenes difieren mucho de los pequeños linfocitos en su morfología. Los núcleos de las plasmáticas son más densos en cromatina, más grandes y por lo general contienen nucleolos”. Sin embargo Miller reconoce que el carácter diferencial de las jóvenes células linfocitarias y células plasmáticas está definido, pero no es muy marcado. La importancia del trabajo de Miller podemos concretarla en los siguientes puntos: • Descubre un nuevo método para inducir la producción de gran número de células plasmáticas. • Establece el origen de las células plasmáticas en un tejido conectivo primitivo celular y describe su trasformación desde la célula precursora: “experimenta un aumento progresivo de la basofilia del citoplasma, el núcleo se sitúa excéntricamente, y se produce una condensación de la cromatina cerca de la membrana nuclear. Cuando el núcleo asume la posición excéntrica, aparece un área clara en el centro del citoplasma”. • Asegura que la célula plasmática es una entidad definida, y presenta un ciclo de maduración: “… las diferencias entre formas jóvenes, maduras y seniles eran claramente reconocibles cuando trascurrían algunos días después de la última inyección del agente estimulante” En 1931, el germano Oelkers, presenta su trabajo sobre “Enzimas proteolíticas de linfocitos” (182). La existencia de comunicaciones anteriores, entre ellas las de Willstätter y Bamann, donde se alude a la existencia de proteasas en 110 RESULTADOS linfocitos, y la de Husfeldt que estudia enzimas tales como peptidasa, triptasa y catepsina, de extractos en glicerol de linfocitos humanos de sangre normal y de pacientes con leucemia mieloide, animan a Oelkers a iniciar su estudio. Oelkers (182), estudia los fermentos que producen los linfocitos en dos pacientes leucémicos. En el primer paciente, al que denomina H, observó una cifra 6.400 leucocitos por ml, con un 75 % de linfocitos tras tres semanas de terapia con rayos X. Un segundo paciente sin tratamiento previo, presentó una cifra de 174.000 células blancas de la sangre con un 99 % de linfocitos. Para la detección de los fermentos en linfocitos se empleó una técnica a partir de sangre desfibrinada, y se siguió un protocolo estándar que determinaba con precisión los volúmenes, sustratos utilizados, pH idóneo y el medio de suspensión empleado (glicerol), para cada uno de los dos pacientes y según la enzima a estudiar. Los resultados obtenidos por Oelkers indican la existencia en los linfocitos de los dos pacientes con leucemia linfocítica, valores significativos de erepsina, débiles de tripsina y catepsina, así como la presencia de lipasa. Nuestro conocimiento sobre las enzimas se remonta al siglo XIX. En 1836, Theodore Schwann, médico y bioquímico, aisló partiendo del jugo gástrico un fermento capaz de desintegrar y disolver la albúmina, la enzima peptidasa. En 1878, Wilhelm Kühne, profesor de fisiología en Heidelberg, dio el nombre de enzimas a los biocatalizadores que tenían actividad también en cultivos fuera de la célula viva, y en 1907 Eduard Buchner (183) recibió el Novel de Química por sus investigaciones bioquímicas y el descubrimiento de la fermentación libre de células. Las enzimas proteolíticas o proteasas son un grupo de enzimas que descomponen las proteínas en unidades más pequeñas; tripsina y quimotripsina son dos ejemplos de proteasas de origen digestivo. En el grupo también se incluyen la bromelaína de la piña, la papaína existente en la papaya y las proteasas fungales (184). 111 RESULTADOS Al realizar la búsqueda en un intento por conocer más sobre el estado científico y las publicaciones existentes sobre enzimas proteolíticas, encontramos, en 1901, una primera comunicación: “la acción de enzimas proteolíticas sobre toxinas bacterianas” (185), y algunas otras que llaman nuestra atención como por ejemplo: “presencia de una enzima proteolítica en el suero normal del buey” en 1903 (186), o “las enzimas proteolíticas de la leche humana” en 1908 (187). Cuando la búsqueda incluye enzimas proteolíticas y linfocitos en fechas anteriores al estudio de Oelkers, los trabajos son escasos: “estudio de los fermentos proteolíticos de los grandes linfocitos en un caso de leucemia aguda” de 1908 (188), y “estudios de fermentación. La proteolisis leucocitaria” en 1912 escrita por Jobling JW (189), donde el autor demuestra que la acción proteolítica de los leucocitos no es una escisión simple debida a un único fermento, sino que se debe a la acción de dos proteasas, una que actúa en un medio alcalino y otra en medio ácido. También nos llama la atención la existencia, entre las comunicaciones encontradas, de un número no despreciable de ellas que tratan sobre enzimas y cáncer: “investigación sobre la acción enzimática dirigida al estudio de la bioquímica del cáncer. Actuación de la pro-­‐lipasa pancreática” de 1932 (190); “enzimas y cáncer. La ß-­‐glicofosfatasa en el eritrocito”de 1935 (191), y “estudios y tratamiento del cáncer con enzimas proteolíticas” de 1935 (192). Hecho que no debe extrañarnos, ya que el uso de las enzimas digestivas para el tratamiento de las enfermedades es muy antiguo, la utilización de papaya (papaína) era algo muy común en el tratamiento de las úlceras malignas en la civilización Maya, y a principios del siglo XX, el jugo pancreático se empezó a usar para tratamiento del cáncer. La enzimoterapia se utilizó para mejorar las digestiones, eliminar virus, estimular las defensas, acelerar la curación de heridas y disminuir la inflamación. No es de extrañar que Oelkers pudiera verse influido por esta tendencia general hacia las bondades de las enzimas y decidiese estudiarlas en el linfocito. Sea cual fuere la intención de Oelkers con su estudio, lo que está fuera de duda es que es uno de los primeros de esta 112 RESULTADOS índole, no apareciendo trabajos similares hasta mitad de los años setenta, en 1975. “El ratio polimorfonuclear/linfocito existente a una altitud de 5.750 pies” es publicado por Artur Dighton Stammers, del Departamento de Fisiología de la Universidad de Johannesburg, en 1933 (193). Los registros que aparecen en su escrito se refieren al conteo diferencial realizado durante un período de cuatro años a 171 estudiantes de medicina, de edades comprendidas entre 18 y 24 años. Stammers considera un motivo interesante de estudio la variación de los valores de polimorfonucleares y linfocitos con la altitud. Los datos registrados en algunos libros de texto, como por ejemplo los de Bainbridge y Menzies, Bucknester, Burhhan, Fleco y Hill, de 68,2% de polimorfos y 25,8% de linfocitos, probablemente corresponden a observaciones hechas a nivel del mar, y las referencias en la literatura de determinaciones realizadas a altitudes superiores al nivel del mar son muy escasas. Sin embargo, es un hecho bien establecido, que la radiación ultravioleta ejerce un profundo efecto sobre estos valores diferenciales. Clark (194) afirmaba que la radiación ultravioleta produce una marcada linfocitosis, conclusión a la que también llegó Hardy aplicando rayos ultravioletas de más de 300 nm a conejos. La ciudad de Johannesburgo, situada a una latitud 26.11 S, longitud 28.03 E, y a una altitud de 5.750 pies (1.752,6 metros) sobre el nivel del mar, recibe un 73% del total de horas de sol, en comparación con el 40% de Suiza y el 29% de Londres. Por otra parte, está bien establecido que la luz solar en lugares de gran altitud, es más rica en ultravioletas que a nivel del mar, y además, cuanto más cerca del Ecuador se encuentra un lugar, existe mayor tendencia a que la luz sea de longitudes de onda más corta. Por tanto, es de esperar que una linfocitosis relativa se encuentre en la sangre de los residentes en un distrito como Johannesburgo. 113 RESULTADOS La técnica empleada por Stammers (193) con resultados satisfactorios era muy sencilla: “El dedo se pincha justo por encima de la uña, sin presión, y la sangre se pone en contacto con un portaobjetos libre de grasa. Con un cubreobjetos aplicado en ángulo de 30º se extiende la sangre de forma uniforme a lo largo de la superficie del primer portaobjetos. La película así formada se deja secar y se colorea con tinción de Leishman”. El conteo se realizó sobre un total de 300 células por cada cubreobjeto, cifra óptima para evitar errores e igual de fiable en cuanto a exactitud que utilizando conteos más extensos de 400 a 700 células. Una comparación de los datos obtenidos por Stammers y los aportados por otros autores a nivel del mar, quedan reflejados en la siguiente figura donde se aprecia un aumento claro en el número de linfocitos. Se observa una caída de polimorfos aproximadamente del 14%, y un aumento de linfocitos en la misma proporción, en comparación con los valores medios registrados como normales a nivel del mar (68.2% y 25.8% respectivamente). Figura 15. Porcentaje celular según altitud. Áreas sombreadas de oscuro: polimorfonucleares. Áreas sombreadas de claro: linfocitos Blanco: Otras células. A: media de las medidas a nivel del mar. B: medidas de la presente investigación. Tomado de Stammers, 1933, (193). Una vez que finalizó su trabajo, Stammers tuvo la oportunidad de examinar la tesis de Sachs sobre el sistema nervioso autónomo y la demencia precoz, en la 114 RESULTADOS que figuraban los resultados de los valores diferenciales de 81 nativos africanos del Hospital Mental de Pretoria (latitud 25.5 S, longitud 28.11 E), situado a una altitud de 4.392 pies (1.338,7 metros) sobre el nivel del mar, y comprobó que las cifras dada por Sachs sobre linfocitos y polimorfos eran muy similares a las que él había obtenido: “…se verá que hay una estrecha concordancia entre sus cifras y las mías”. El interés del trabajo de Stammers queda patente en la medida que el recuento diferencial de leucocitos se utiliza como garantía de pruebas para fines diagnósticos, lo que concede al reconocimiento de estas variaciones dependientes de la altitud suma importancia. Otro de los autores con una trayectoria de varios años de investigación y varios artículos publicados es J.M.Yoffey. Sus estudios de 1929 sobre la formación de la sangre en peces, darían lugar a un nuevo trabajo acerca del tejido linfoide de mamíferos y su relación con el resto del sistema hematopoyético (195)(196). Yoffey profesor de anatomía de la Universidad de Gales del Sur y Monmouth, Cardiff, publica en 1933: “Estudio cuantitativo de la producción linfocitaria” (197). Considera una suerte que los linfocitos se presten a un estudio cuantitativo más fácilmente que cualquiera de los otros elementos de la sangre: “Es posible intervenir el conducto torácico inmediatamente antes de que drene la linfa en la sangre, y estimar así el contenido en linfocitos”. Yoffey utilizó para su estudio a 12 perros de peso entre 4 y 12 Kg, que después de 24 horas de ayuno se alimentaron con 1/4 Kg de manteca de cerdo. Los animales fueron anestesiados con cloralosa a razón de 0,1 gramo por Kg de peso y se localizó el lugar más idóneo para acceder al conducto torácico teniendo en cuente su distribución anatómica irregular. La principal complicación a temer es la coagulación de la linfa, teniendo a veces que abandonar por coagulación persistente, y no pudiendo emplear anticoagulantes como el extracto de sanguijuela por ser un poderoso linfagogo. 115 RESULTADOS Se registró la cantidad de linfa recogida por hora, tomando tres gotas de la cánula al final de cada hora para recuento linfocitario. El recuento resulta ser sorprendentemente constante a lo largo de las horas, pudiendo así calcular la producción de linfocitos. Por ejemplo, si al final de una hora, a linfa contenía 10.000 linfocitos por ml, y si el flujo total para esa hora era de 25 cc, la salida total de linfocitos para esa hora sería: 25 x 10.000 x 1.000 = 250.000.000, que para un período de 24 horas representaría una producción de linfocitos de 6.000.000.000. El siguiente paso fue el estudio de los componentes celulares de la linfa del conducto torácico. Las células encontradas eran predominantemente linfocitos pequeños constituyentes del 95% de las células totales, el restante 5% eran células mucho más grandes (linfocitos grandes y macrófagos). Las técnicas de tinción supravital permiten diferenciar bastante bien los dos tipos celulares, lo que es de importancia para determinar la posibilidad de una circulación continua de linfocitos, de la sangre a los ganglios, y de la linfa a la sangre, siendo el número de linfocitos en sangre el resultado de un equilibrio entre estos dos procesos, salida y entrada de la sangre. La linfocitosis se divide así en dos grupos, lo que Yoffey denomina linfocitosis activa o verdadera, por entrada en sangre de un mayor número de linfocitos, y linfocitosis pasiva o de retención, debido a una interferencia en la salida de linfocitos. “¿Cual es el destino final de los linfocitos sanguíneos?”, se pregunta Yoffey (197). La cuestión fundamental es en qué parte del cuerpo acaban estos linfocitos y con qué propósito. Para intentar dar respuesta, Yoffey sigue en su estudio de los linfocitos a los que define como células jóvenes, analiza los linfocitos del tejido conectivo, a los que llama linfocitos histiógenos, y considera que el único tejido del cuerpo que regularmente fabrica grandes cantidades de células es la médula ósea, pero considera su interpretación histológica extremadamente difícil. Acaba reconociendo que los factores que controlan la producción de linfocitos permanecen en la oscuridad. 116 RESULTADOS A continuación expone las dos teorías sobre la formación de la sangre: la monofilética, que considera que la célula madre de eritrocitos y granulocitos es la misma, y la teoría polifilética, que cree que aunque la célula madre se parece mucho, existen pequeñas diferencias significativas entre ellas. Refleja también sus observaciones sobre el control de la producción linfocitaria y comenta el posible papel de la linfa y linfocitos en el transporte de grasa y la función de barrera que ejerce el tejido linfoide en los casos de inflamación o depósitos malignos: “clínicos y patólogos han asumido que el tejido linfoide existe para este propósito, que sirve como una barrera que evita por un tiempo, la propagación de la materia perjudicial…pero si es cierto que actúa como barrera, no es menos cierto que los vasos linfáticos facilitan enormemente la propagación”. Reconoce la necesidad de entender la función de este tejido linfoide “silencioso” que existe en personas normales, antes de poder empezar a apreciar el significado de los cambios patológicos, y añade: “la función que definitivamente podemos asignar a los tejidos linfoides es la formación de linfocitos. Por tanto, es el conocimiento de la vida de los linfocitos lo que debe constituir el punto de partida de cualquier investigación”. Zeyland, presenta en 1933 una comunicación sobre el estudio del índice monocito/linfocito en la tuberculosis usando el método supravital (198). El método de tinción supravital ha sido utilizado por numerosos investigadores para el análisis de la sangre, entre ellos, Arnold, Unna, Marcus, Forkner (164), Bettmann, Rosin, Wiseman (176), Achard, Ramond y Schilling. Este método se difundió rápidamente en América, pero tuvo poca aceptación en Europa, figurando entre algunos de sus defensores del Continente Europeo, Masugi y Seemann de la Escuela Aschoffschen, Sanctis Monaldi de los Laboratorios Calmettes, Mareggiani y Rezzesi en Italia y Lorentz en Holanda. Se describe y explica la técnica empleada por Lorentz, y también el método original de Sabin (199). Zeyland, lleva a cabo sus estudio en cobayas, registrando los valores medios de 11 cobayas normales y 20 tuberculosos y realiza un registro sistemático en 16 117 RESULTADOS conejos con resultados similares, observando una disminución progresiva en el recuento leucocitario y un aumento del índice monocito/linfocito (M/L) relacionado con la evolución de la enfermedad. Los resultados obtenidos no son considerados por el propio autor lo suficientemente fiables como para realizar un diagnóstico a partir de ellos, estando en discrepancia con la teoría de la escuela americana sobre el diagnóstico basado en el índice M/L. Realiza también diferentes estudios en seres humanos (198), principalmente niños, estableciendo tres grupos según los resultados de dicho índice. El grupo I, de casos con índice M/L considerado normal, engloba 2 casos de tuberculosis crónica pulmonar, 2 complicados con anemia y 5 casos de tuberculosis miliar terminal, y asegura: “el relativo bajo índice M/L también aparece en la tuberculosis experimental poco antes de la muerte del animal”. El grupo II correspondiente a la tuberculosis activa, se corresponde con un alto índice M/L, y el grupo III engloba casos con alto índice M/L debidos a tuberculosis progresiva, pero también debidos a peritonitis, neumonía lobar o neumonía crónica, por lo que en este último grupo dice: “…el alto índice M/L no siempre sería indicador de actividad tuberculosa”. Del estudio realizado en 545 muestras de sangre tomadas de personas, principalmente niños, y animales, Zeyland (198) obtiene las siguientes conclusiones: 1-­‐ La tinción supravital de Sabin es insustituible en el diagnóstico diferencial de laboratorio para linfocito, monocito, célula monocitoide y clasmatocito. 2-­‐ Como resultado de la infección tuberculosa experimental en conejos y conejillos de indias, aumenta el número de monocitos y disminuye el de linfocitos en sangre periférica, en otras palabras aumenta el índice M/L. 3-­‐ Un alto índice M/L se presenta con mayor frecuencia en la tuberculosis progresiva de la edad infantil, y representa un pobre pronóstico para estos niños. 118 RESULTADOS Otro curioso trabajo es realizado por Leonhard Löffler, de la Universidad de Clarité–Berlín en el año 1934, que tiene como motivo de estudio “el comportamiento de los linfocitos y la reacción tisular en el grupo experimental del edema congestivo” (200). Löffler en sus experimentos intenta determinar el papel que juegan los linfocitos en los tejidos del cuerpo. Utiliza ratas a las que sacrifica tres días después de provocarles un fuerte edema en toda la extremidad inferior mediante la ligadura de la vena femoral, y realiza el estudio microscópico de todos los tejidos de esta extremidad, incluyendo el edema subcutáneo del área de rodilla y muslo. En estas zonas, observó un aumento significativo de los pequeños linfocitos en comparación con las zonas sanas no edematosas. En un segundo experimento, las ratas son sacrificadas a los seis días de la ligadura, e igualmente en este caso se puede encontrar una pronunciada proliferación de linfocitos sobre la zona del edema (200). Realiza un total de nueve experimentos, sacrificando los animales a intervalos de tiempo cada vez más largos, de tal forma que en el experimento número 9 las ratas se sacrifican a los 42 días, y observa que al aumentar los días de vida de los animales, aumentan también los signos de degeneración en los linfocitos como son la presencia de núcleo borroso, limitación del protoplasma y rotura del citoplasma entre los más significativos, además tiene lugar una disminución de la proliferación linfocitaria, y se observa un aumento en el número de histiocitos. A los 42 días, en el tejido subcutáneo de pierna, rodilla y muslo se detecta un edema residual, y cuando se compara con el lado sano, se aprecia una pequeña proliferación de linfocitos, con predominio de histiocitos y un desarrollo del tejido conectivo Löffler (200) resume los acontecimientos observados en el tejido edematoso cuando dice: “en el edema provocado experimentalmente tiene lugar una intensa proliferación de linfocitos, en respuesta a la probable modificación metabólica de los tejidos. Bajo el estímulo del edema también se desarrollan otras respuestas 119 RESULTADOS tisulares como la proliferación de histiocitos, y con su resolución aparece tejido conectivo”. Helmuth Richter, escribe en su lengua materna, el alemán, un análisis sobre las posibles causas de linfocitosis (año 1934), titulado: “Sobre la linfocitosis familiar” (201). Richter presenta las posibles causas de linfocitosis, haciendo referencia a la presencia de ésta como algo normal o patológico en una serie de estados y enfermedades que agrupa en: a. Linfocitosis asociada a neutropenia, como en el tifus o la gripe. b. Asociada a severa anemia como en la leucemia linfocítica. c. Asociada a enfermedades endocrinas, acromegalia y mixedema. d. Ocasionalmente junto con anemia no regenerativa. e. Asociada a tuberculosis y lues. f. En respuesta a ciertas vacunas. g. Combinada con monocitosis y enfermedades protozoarias. h. En el escorbuto, la aquilia gástrica simple, y i. Convalecencia de enfermedades infecciosas Asume la existencia de una linfocitosis constitucional, que observó en dos casos con una clara influencia familiar: “La linfocitosis asociada a granulopenia, que ocurre en la constitución hiperplásica y asténica, puede tener un probable origen familiar”, aclara. También considera que cuando la linfocitosis ocurre en enfermedades endocrinas, desórdenes hormonales y enfermedades degenerativas de la sangre puede observarse la existencia de un origen familiar. Según Richter, el propósito de esta comunicación es simplemente: “dar a conocer la existencia de una linfocitosis constitucional y su posible relación con los cambios en el cuadro hemático para el conocimiento de algunas enfermedades”. 120 RESULTADOS En la Revista Americana de Patología de 1934 se recoge una publicación sobre “tumores y condiciones similares a tumores de linfocitos, mielocitos, eritrocitos y células reticulares”, de autor G.R Callender (202). El documento expone una clasificación basada en los 380 casos recogidos durante 7 años en el Registro de Tumores Linfáticos de la Asociación Americana de Patólogos y Bacteriólogos. Todos los casos de este registro se revisaban cada año para elaborar el informe anual, y aquellos casos con dificultades diagnósticas y con suficiente material disponible, se remitían a la comisión de Consultoría y, a menudo, a otro patólogo. El resultado de estos estudios es esta clasificación de tumores. Callender (202) reconoce que la nomenclatura de estas enfermedades es extraordinariamente variable en textos y artículos, y observa que: “… el grupo de condiciones derivadas del linfocito es el mejor definido y mejor entendido de todos los grupos aquí contemplados”. Considera, con respecto al término linfoblastoma, que puede ser aplicado a la totalidad del grupo, pero los diferentes miembros deben ser renombrados ya que el término no define suficientemente a la entidad. Dentro del grupo derivado de los linfocitos, existe un considerable paso de una condición a otra, tanto de forma espontánea como resultado del efecto de la irradiación. En esta clasificación, la extensión del proceso a otros nodos o tejidos linfáticos no se considera metástasis. Respecto al linfocito, la clasificación que se muestra es la siguiente: 121 RESULTADOS Tabla I. Clasificación de tumores y condiciones semejantes a tumores del tejido linfático. Tomado de: Callender, 1934 (202). En el grupo I se encuentran los procesos reactivos, no contemplados en el registro. El grupo II y III comprende las proliferaciones de tipo neoplásico: linfocitoma leucémico, agudo y crónico, y linfocitoma aleucémico, difuso o nodular. El grupo IV corresponde a los tumores malignos, linfosarcomas ya sean leucémicos o aleucémicos. Se definen cada uno de estos grupos y se describen sus hallazgos microscópicos, dando algunos criterios para el diagnóstico diferencial en la explicación de la clasificación. Así, por ejemplo, se habla de leucemia definitiva, si existen más de 25.000 células blancas en sangre con un predominio de linfocitos, observándose al microscopio un aumento de los nódulos primarios debido a proliferación de linfocitos y una distorsión de la estructura de los nodos linfáticos. 122 RESULTADOS Curiosamente la Enfermedad de Hodgkin queda incluida dentro del grupo de las células reticulares, y Callender (202) describe detalladamente dos variedades, una forma localizada, que se extiende por continuidad linfática local, y una forma generalizada, que se extiende a todos los grupos de ganglios linfáticos, hígado, bazo o ambos, y hace referencia al Sarcoma de Hodgkin, cuando dice: “ocasionalmente en el curso de una Enfermedad de Hodgkin focal o generalizada, se produce un proceso neoplásico metastático, un verdadero sarcoma, siendo la célula neoplásica de estirpe reticular”. Nos muestra asimismo su espíritu de trabajo y el objetivo de dicha clasificación, a la que no considera perfecta ni definitiva: “… esta clasificación se presta a la crítica y estimula el debate, quedando abierta a la aportación de casos de apoyo que puedan refutar o modificar las opiniones expresadas en el presente documento”, algo no demasiado habitual en el mundo científico, digno de elogio y respeto. Como consecuencia de las observaciones realizadas en su anterior trabajo (197), Yoffey estudia las consecuencias de la anestesia en el perro, publicando su trabajo en 1935: “disminución de los linfocitos en la sangre del perro anestesiado con cloralosa” (203). Los datos presentados en este trabajo constituyen el punto de partida para una consideración de los factores involucrados en el mantenimiento del equilibrio de la linfocitosis. Entiende por “balance de linfocitos” el mecanismo mediante el cual los linfocitos de la sangre se mantienen a un nivel bastante constante, y se les impide someterse a fluctuaciones demasiado extremas. A los perros bajo anestesia intravenosa con cloralosa se les realiza conteo inmediato (A) de glóbulos blancos total y diferencial, y en algunos animales recuento de glóbulos rojos. Transcurridas 2 ¼ y 4 ¼ horas se realizan nuevos conteos sanguíneos (B). Los datos completos se expresan en tablas, observándose que la caída de linfocitos en sangre es un fenómeno casi constante, siendo habitual el aumento de neutrófilos, pero no de forma tan regular. Se realizaron un total de 13 experimentos, y se observó una caída de linfocitos en todos ellos, excepto en uno (4ES64); en éste hubo un ligero aparente aumento (50 por ml), que no tenía ningún significado por ser un cambio muy 123 RESULTADOS pequeño. Al revisar los resultados de experimentos anteriores, parece claro que en el perro bajo las condiciones experimentales descritas, una caída en los linfocitos de la sangre es prácticamente un cambio constante. Yoffey asegura: “es imposible formarse una opinión, en cuanto a la importancia de este cambio, ó incluso de cómo el cambio se ha llevado a cabo. La cuestión de si es debida a un aumento en el número de linfocitos que salen de la sangre ó una disminución de los entran, solamente puede ser resuelta mediante el estudio experimental de estos dos procesos, y la investigación de cada uno por separado” que Yoffey presentará en un nuevo documento. Yoffey, continúa sus investigaciones sobre producción linfocitaria, publicando de nuevo en 1936 (204). Mientras realizaba el estudio cuantitativo de la producción de linfocitos en 1933, Yoffey observó que el número de linfocitos en la linfa del conducto torácico del perro mostraba una considerable fluctuación, sugiriendo la posibilidad de variaciones entre un animal y otro, lo que dio lugar a un estudio para comparar la producción linfocitaria en diferentes animales, siendo necesario para ello, el conocimiento de la producción total de linfocitos. Los resultados de los 21 experimentos realizados, los resume en una tabla, y con la finalidad de facilitar su comparación, se muestran calculados por unidad de tiempo (1 hora) y unidad de peso corporal (10 Kg) Cuando se estudia la producción horaria de linfocitos en cada perro, se observa una gran variación entre animales, e inmediatamente surge la pregunta: “¿esta variabilidad es o no permanente?” Yoffey (204) considera que para su resolución es necesario conocer el número total de linfocitos en la circulación, y puesto que los linfocitos en sangre se mantienen relativamente constantes durante un período, el mismo número de linfocitos deben entrar y salir diariamente en la sangre. Determina el factor de reemplazo diario ó DRF, relación entre el número de linfocitos que entran diariamente a sangre a través del conducto torácico y los linfocitos en sangre al inicio del experimento, calculando así la producción media diaria de linfocitos del conducto torácico de un perro de aproximadamente 10 Kg, en 5.078 millones. Lo que significa que 124 RESULTADOS los linfocitos de la sangre son normalmente reemplazados 2.06 veces diarias. Por lo tanto, los linfocitos permanecen en sangre durante 11,65 horas. Otra de las preguntas que Yoffey trata de contestar es: “¿son los linfocitos del conducto torácico células recién formadas?” Parte del supuesto de que los linfocitos del conducto torácico han sido recientemente formados en los tejidos linfoides y entran por vez primera en el sistema linfático. Pero debe ser considerada la posibilidad alternativa: “…que no sean células recién formadas, sino que exista una continua circulación de linfocitos de la sangre a ganglios linfáticos y viceversa”. Sobre el tema existen opiniones dispares. David y Carlson en 1909 observaron que el contenido en linfocitos de la linfa eferente se incrementó en gran medida por el masaje del tejido linfoide. Haynes y Field en 1931 encontraron que en el perro, la linfa obtenida de la parte frontal del tobillo, contenía un promedio de solo 33 células/ ml, y concluyen: “el hecho de que la linfa del tobillo que a pasado a través de donde no hay nódulos linfáticos, prácticamente no contiene células, indicaría que normalmente las células entran en la linfa desde los ganglios linfáticos, y no por diapédesis desde la sangre de los espacios de los tejidos”. En cambio Drinker y Field (205) llegan a conclusiones diametralmente opuestas, y dicen: “en nuestra opinión, los linfocitos escapan de los capilares sanguíneos de todo el cuerpo y vuelven a entrar en la circulación a través del torrente linfático”. Yoffey considera que la evidencia disponible, es muy a favor de que los linfocitos de la linfa del conducto torácico, son células que han entrado en la linfa, procedentes del tejido linfático: “… y una vez que los linfocitos se encuentran en el torrente sanguíneo, no vuelven a entrar en la linfa en cantidad apreciable, por lo que la linfa contiene pocos o ningún linfocito, antes de pasar por los ganglios linfáticos”. Algunas de las conclusiones a las que llega Yoffey (204) en su estudio son: 125 RESULTADOS 1-­‐ Los resultados encontrados, para un peso corporal de 10 Kg, en la salida media de linfocitos/hora fue de 211,6 millones, con un máximo de 875,4 millones y un mínimo de 30,2 millones 2-­‐ La entrada de linfocitos en sangre no se produce a una velocidad uniforme, sino que muestra una marcada fluctuación. 3-­‐ Los linfocitos del conducto torácico son nuevas células que han entrado en la linfa procedente de los tejidos linfoides. Entran en el torrente sanguíneo en cantidad suficiente para reemplazar a los que lo abandonan, a un promedio en el perro de 2,06 veces/día, y permanecen en él una media de 11,6 horas. Un nuevo estudio sobre “La constitución hemocitológica de conejos adultos machos en quince razas estándar” es llevado a cabo por varios autores, entre ellos, Casey y Pearce en 1936 (168). Se estudiaron 180 conejos, representativos de 15 razas estándar, la mayoría de 4-­‐10 meses de edad, todos siguieron la misma dieta alimenticia y se encontraban en condiciones físicas excelentes y libres de enfermedades. Se hicieron intentos de eliminar o mantener constantes variables tales como edad, sexo, estación del año, momento del examen, errores técnicos, alimentación, vivienda y enfermedades de los animales, y se realizaron tomas de sangre para conteo de células, durante un período promedio de 2 semanas. Las observaciones de catorce factores sanguíneos, entre ellos, eritrocitos, hemoglobina y células blancas de la sangre, se promediaron para cada animal, y los valores medios junto con la varianza se determinaron para cada raza y el grupo de conejos considerado como un todo. Los resultados mostraron una variación significativamente mayor entre las razas que dentro de la misma raza, lo que según los autores parecía indicar que en el conejo, los animales genéticamente relacionados tienen fórmulas de sangre similares, y los no relacionados genéticamente, fórmulas sanguíneas distintas. 126 RESULTADOS Las razas fueron divididas según el peso en grandes, intermedias y ligeras. Se tuvo en cuenta también el comportamiento frente a enfermedades y se observó que las distintas razas presentaron grandes diferencias respecto a la susceptibilidad a enfermedades experimentales y espontáneas. La influencia de los cambios estacionales en el recuento celular se tuvo en consideración cuando se hizo la observación, viéndose que la diferencia entre las razas no era debida a cambios estacionales. En base a todas estas consideraciones los autores (168) realizaron la evaluación final del estudio y formularon las siguientes conclusiones: • Que el análisis de la varianza mostró que la variación entre la raza fue significativamente mayor que la variación dentro de la raza. • Que no se detectaron variaciones en los monocitos, excepto cuando las razas se dividieron en pesadas y ligeras, ni tampoco hubo variación en los valores de neutrófilos/cmm; un gran número de razas tenían exactamente el mismo nivel medio de neutrófilos. Y concluyeron que: “las variaciones en la fórmula sanguínea, no podían ser atribuidas a ningún factor y eran debidas a una base hereditaria/genética”. En la misma línea de trabajo es el estudio llevado a cabo por F.R Sabin junto a Miller, Smithburn, Thomas y Hummel de 1936 sobre “los cambios en las células de la sangre y médula ósea de conejos en desarrollo” (206). Parten del supuesto de que los tejidos de la cavidad ósea no están asociados con la función de las células sanguíneas: “… más bien la primera función de este órgano está en relación con el desarrollo del hueso” y continúa con un repaso anatómico-­‐fisiológico del crecimiento del hueso en el conejo. Estudiaron la sangre de conejos jóvenes, empezando por las etapas fetales tardías, los cambios en el nacimiento y durante los 6 primeros meses de vida. De 132 conejos criados en el Instituto Rockefeller, se realizaron recuentos sanguíneos en 115, recuentos de médula ósea en 49 y estudio de sangre y 127 RESULTADOS médula ósea en 32 animales. En las etapas fetales y primeras dos semanas de vida la sangre se obtuvo por punción del corazón, después a partir de una vena de la oreja, y las preparaciones de médula ósea se obtuvieron a partir de fémur, tibia y húmero. La técnica supravital, que permite una mejor discriminación de los monocitos se utilizó para los recuentos de células sanguíneas, contabilizando un total de 100 células en cada animal. En todas las series y para todas las edades se contaron un total de 73.100 células. Respecto a los recuentos de células en médula ósea, los autores hacen una serie de advertencias: “… no pueden ser tan precisos como los recuentos sanguíneos, ya que debe hacerse una elección de campos; y sólo las zonas en las que las células están en una sola capa y ligeramente separadas unas de otras se pueden contar. Además, deberán cumplir ciertas normas como el tipo de fijador empleado, la ausencia de ácido en formol y el corte en secciones delgadas”. Los resultados (206) observados fueron los siguientes: 1. El número de células de la sangre del conejo es bajo en los últimos días fetales, con una tendencia al alza el día del nacimiento. 2. Los glóbulos blancos aumentan lentamente y no alcanzan los valores característicos del animal adulto hasta el 5º-­‐6º mes, y al igual que las células rojas hubo un fuerte aumento en el día del nacimiento, sobre todo debido al aumento de leucocitos neutrófilos. 3. Las curvas de linfocitos y monocitos son muy diferentes. El número de linfocitos mostró un aumento progresivo hasta los meses 5º-­‐6º de vida, siendo más rápido en el primer mes que a partir de entonces. En cambio, los monocitos aumentan rápidamente durante la primera semana de vida y se mantienen así en los siguientes meses, siendo por tanto el monocito la primera célula en el torrente sanguíneo que alcanza su cuota normal. 4. En el estudio del desarrollo de las células blanca, la característica leucopenia de etapas fetales se supera lentamente, y desaparece en el 6º mes de vida. Las diferentes cepas de células blancas, granulocitos, 128 RESULTADOS linfocitos y monocitos aparecen en la corriente sanguínea a ritmos diferentes. Figura 16. Células sanguíneas de 115 conejos jóvenes según los datos recogidos. Tomado de: Sabin y col., 1936 (206). Los datos recogidos sobre la médula ósea, muestran que el día del nacimiento, la médula ósea es relativamente inactiva. En el segundo día, se ven sinusoides dilatados llenos de células. Pasados tres días tiene lugar el inicio de la actividad medular, que se corresponde con una hiperplasia eritroide. Más importante, desde el punto de vista de la hematopoyesis, es el tipo de célula registrada con el nombre de célula primitiva, que es descrita en detalle: “Esta célula que se asemeja al linfocito pequeño, muestra menos signos de diferenciación que los linfocitos de los ganglios linfáticos, bazo y sangre. El estrecho borde de 129 RESULTADOS citoplasma casi no contiene gránulos visibles, el citoplasma tiene poca basofilia y el material mitocondrial debe estar finamente dividido. Los datos muestran que hasta el día 5º, estas células son numéricamente superiores al 5%”. Un hallazgo muy llamativo es la velocidad con la que el cuerpo joven es capaz de producir un número significativo de células rojas, en contraste con el retraso en la producción del correspondiente número de células blancas de la sangre. En la tercera semana de vida, la sangre del conejo tiene su cuota completa de células rojas, mientras que ésta no se alcanzará hasta el 6º mes en el caso de las células blancas. En estadios fetales tardíos, la médula ósea es relativamente quiescente, y por lo general predominantemente eritroide. Durante la segunda semana hay una marcada hiperplasia eritroide en médula, esto significa que durante un corto tiempo, el animal tiene que utilizar gran parte del espacio disponible en la cavidad medular para la producción de glóbulos rojos, y es durante esta segunda semana cuando tienen lugar el crecimiento óseo. Este comportamiento de la médula ósea les lleva a afirmar: “el crecimiento de los huesos tiene una influencia importante en la hematopoyesis”. Los autores (206) aceptan la existencia una célula madre común para todas las células blancas, y se plantea la cuestión de si esta célula madre es idéntica al linfocito o es un tipo menos diferenciado, asegurando: “Está claro que la célula madre es la Linfodocita de Pappenheimer, o el Hemoblasto linfoide de Jordam y Jonson”. Ruth Vortisch, otro de los autores dedicado al estudio de la tuberculosis, publica en 1937: “la relación monocito/linfocito (M/L) en la tuberculosis, con especial consideración a su comparación con el método de Schilling y el test de sedimentación de eritrocitos” (207). Parte del hecho de que el cuadro sanguíneo en el diagnóstico de la tuberculosis, es una excelente herramienta que va ganando importancia. Hace una revisión de la literatura, entre otros, menciona los trabajos de Gottlieb, 130 RESULTADOS para quien la monocitosis era una reacción defensiva frente a la tuberculosis, los trabajos de Zeyland (198) con cobayas y conejos, de Rezzesi que estudió el comportamiento del índice M/L en animales utilizando el método supravital de Sabin, y quien aseguraba que el aumento del índice M/L era señal de infección tuberculosa, y su reducción o negatividad aparecía en animales caquécticos. Dolgopol y Sitt, que estudiaron el cuadro sanguíneo tras la vacunación con BCG, y de Blackfan, Kenneth y Diamond, quienes investigan el comportamiento del monocito en la tuberculosis utilizando el colorante supravital. Vortisch realiza sus propios experimentos con la finalidad de aclarar el significado del índice M/L. Observó a 123 pacientes tuberculosos, a los que siguió por un período de tiempo no inferior a 6 meses, a fin de definir distintas etapas de la enfermedad. A intervalos regulares realizaba hemograma y sedimentación globular, además de los habituales exámenes clínicos, radiológicos y de laboratorio, prestando especial atención al índice M/L. Para lograr que los resultados fuesen comparables a los de los estudios americanos ya realizados, divide a los pacientes en cuatro grupos: I de tuberculosis severa, II media, III leve y IV inactiva, y hace un seguimiento del cuadro hemático en el curso del tratamiento. El principal objetivo de Vortisch es comparar los resultados de las pruebas sanguíneas que él realiza a los 123 pacientes tuberculosos, con los resultados de americanos como Sabin y Cunnigham, con el método habitual de Schilling y con la velocidad de sedimentación globular, destacando entre sus observaciones que el índice M/L de Sabin y Cunnigham coincidía con los valores de desviación a la izquierda y sedimentación globular de los recuentos sanguíneos pertenecientes a pacientes con hallazgos clínicos de tuberculosis activa. Como resultado final Vortisch (207) considera que: “… tanto en tuberculosis activa como inactiva los métodos de desviación a la izquierda y de sedimentación globular son más fiables que los métodos de respuesta no específicos utilizados por los autores americanos”. Donald Mainland y Basil K. Coady, del Departamento de Anatomía, Universidad de Dalhouse, Nueva Escocia, Canadá, publican en 1938: 131 RESULTADOS “Investigación sobre el tamaño celular, con especial atención a los linfocitos humanos” (208). A lo largo de la historia, muchos observadores han considerado que la medición de las células es un trabajo rentable, que las diferencias de tamaño en células o en partes celulares tienen valor en la clínica humana o la patología animal. Como ejemplo, tenemos la obra de Price-­‐Jones (año 1933) en glóbulos rojos humanos; o de Savage y colaboradores quienes mostraron que el tamaño de las cabezas de los espermatozoides de toros y caballos puede ser utilizados para indicar la esterilidad de éstos; o bien la de Quensel, cuya relación nucleolo/núcleo está siendo utilizada en la identificación de células malignas. Estudios previos realizados por Mainland (año 1935) en frotis de sangre humanos, mostraron que la variación en el tamaño de los linfocitos entre pacientes de distintas edades, de ambos sexos y en diversas enfermedades, no era apreciablemente mayor que la variación en los tamaños de linfocitos de personas sanas. En este documento (208) se presta más atención al significado e interpretación de las pruebas realizadas que a sus resultados numéricos. Se diseñaron una serie de experimentos para determinar hasta que punto factores controlables podrían explicar la gran variación entre los tamaños medios de linfocitos en diferentes extensiones. Las extensiones de sangre fueron secadas al aire y teñidas por el método de Wright, y los tamaños fueron expresados como diámetros medios. No hubo diferencias significativas debidas al secado de la película en una cámara de vidrio o sobre una llama de un quemador de laboratorio. Es concebible que el alcohol de la tinción de Wright podría alterar el tamaño celular, sin embargo, las diferencias encontradas examinadas por el análisis de la varianza, no revelaron ninguna relación con la técnica de tinción. El análisis de la varianza mostró que las diferencias en el tamaño medio de los linfocitos entre preparaciones hechas en diferentes ocasiones, incluso en diferentes días, no fue significativamente mayor que las diferencias en el tamaño medio de los linfocitos entre las gotas elaboradas a partir de la misma punción. 132 RESULTADOS Los experimentos realizados, indicaron que algo más que la velocidad de secado y diferencias controlables en la tinción deben ser consideradas para explicar las diferencias de tamaño de los linfocitos entre y dentro de las extensiones. Se encontró que los linfocitos eran más grandes en la periferia de la extensión que en el centro de la misma, y el examen de otras extensiones indicó que la distribución de los linfocitos en la extensión está relacionada con su tamaño y que en ella influyen fuerzas físicas y fenómenos asociados tales como la relación con la masa e impulso. Se estudió también la influencia de la temperatura en el tamaño, para ello cada linfocito fue tomado por separado haciendo tres medidas, a temperatura corporal (37º C), a temperatura ambiente (24-­‐27º C) y a baja temperatura (4-­‐13º C). Los cambios de tamaño observados a temperatura ambiente y a baja temperatura fueron inesperados, y su explicación no clara. Los autores (208) consideraron importante el estudio del tamaño de los linfocitos, y utilizando sangre humana teñida por el método de Wright y montada en solución salina isotónica teñida con violeta de genciana, determinaron los tamaños medios bien mediante micro-­‐proyección o con micrómetro ocular y obtuvieron los siguientes resultados: 1-­‐ Grandes diferencias de tamaño en los linfocitos de 29 extensiones de la misma persona (90-­‐100 linfocitos por extensión), que no podían ser atribuidas a las diferencias en la rapidez de secado o en el tiempo empleado en la tinción; ni a diferencias en las punciones realizadas en días diferentes o el orden de aparición de la gota en el pinchazo. 2-­‐ El diámetro del linfocito en la parte inferior del cristal era de 0-­‐56 veces mayor que en la parte superior, hecho que se atribuye a la caída más rápida de los linfocitos más grandes, concluyendo que factores físicos como la gravedad pueden producir una diferencia de tamaño importante. 3-­‐ Las diferencias de tamaños entre los linfocitos en solución salina y en extensiones puso de manifiesto que la variación entre las extensiones era mucho mayor que entre los soportes de solución salina, y 133 RESULTADOS consideraron que la solución salina proporciona un medio mucho mejor para detectar diferencias fisiológicas, patológicas o experimentales. En definitiva Mainland y Coady llegan a la siguiente conclusión una vez realizado el análisis de su trabajo: “Encontrar las causas de las diferencias reales o aparentes sería difícil. El punto principal a tener en cuenta es que las comparaciones de tamaños, por ejemplo, entre diferentes personas son legítimas sólo cuando el método de preparación y observación es constante, sino es así pueden introducirse serias discrepancias”. Los autores señalan un hecho de suma importancia, la necesidad de un tratamiento uniforme, estándar, cuando se realiza un experimento a fin de poder comparar resultados. Si damos la vuelta a la famosa frase de Einstein: “Si buscas resultados distintos, no hagas siempre lo mismo”, llegamos a la misma conclusión. En este caso, los autores consideran que el procedimiento debe estar bien estructurado y ser reproducible para poder ser comparado. Es necesario contrastar resultados en iguales condiciones de trabajo, solo así podremos comparar adecuadamente diferentes estudios y llegar a conclusiones fiables. Este principio que resulta tan obvio, ha llegado a tener tanta importancia, que en la actualidad, en la disciplina de anatomía patológica existe un texto, el Libro Blanco (209), donde se recoge detalladamente todos los procedimientos de interés. Su publicación se inició en 2009 y se actualiza cada dos años, siendo útil como guía en las tareas de gestión y apoyo documental. Se estructura en varios bloques: • Gestión y calidad: sistemas de información, catálogo de muestras y procedimientos, cargas laborales y costes por proceso, etc. • Aspectos metodológicos: actualización en citopatología y registros de tumores. • Aspectos legales: salud laboral, cesión de muestras y consentimiento informado. 134 RESULTADOS Así por ejemplo, el registro hospitalario de tumores, revisa el texto de 2009 y a partir de él presenta una nueva estrategia orientada a evolucionar desde la Red Nacional hacia un Club multidisciplinar abierto a otras especialidades, con el objetivo de ampliar la red, compartir, desarrollar propuestas en formación y promover la investigación. Perspectivas de futuro que necesariamente habrán de incluir y standarizar otros datos y métodos como son marcadores tumorales, tratamientos y evolución. “El Linfocito en la Inflamación Aguda”, es el título de un artículo de Fred Kolouch (210) del Laboratorio de Hematología, Departamento de Anatomía, Universidad de Minnesota, Minneapolis, de 1939. El autor introduce el tema haciendo un repaso de la literatura al respecto, destacando la aportación de Rich (211), que resume la postura de muchos histólogos y patólogos cuando afirma: “Estoy seguro de que todos los que participan en el estudio y enseñanza de la patología, están de acuerdo en que la completa ignorancia de la función de esta célula es una de las más humillantes y vergonzosas lagunas del conocimiento médico…Literalmente nada de importancia se sabe a cerca de las potencialidades de estas células, distintas de las que se mueven y se reproducen a sí mismas”. La explicación del rol del linfocito en reacciones titulares aceptada en las enseñanzas de los patólogos, puede ser ilustrada por una cita de la última edición del libro de texto de Patología de Boyd (212) en 1938: “En la inflamación crónica y en las últimas etapas de la inflamación aguda, los linfocitos pueden ser las principales células del exudado. Parece probable que la mayoría de las células redondas y pequeñas derivan de los tejidos y no de la sangre…Es notable, que somos tan ignorantes como para desconocer cuál es la exacta función de una célula que tiene un papel tan dominante en la infección crónica”. Muir (213) por su parte afirma: “No se puede decir que ellos (los linfocitos) no tengan poder fagocítico, pero éste es mínimo. Probablemente se agrandan cuando van a ejercer su función…Probablemente representan una respuesta al 135 RESULTADOS tipo más suave de irritación, pero su función exacta no se conoce. Migran, pero están más preocupados con la inflamación crónica”. Continúa con las opiniones expresadas entre otros por Cowdry sobre la migración de los linfocitos, Bailey, Metchnikoff, y Ehrlich que divide a los linfocitos grandes en hemo-­‐macrófagos y linfo-­‐macrófagos según se encuentren en sangre o linfa. Maximow (214), estudió el problema de la función del linfocito muy extensamente, expresando su punto de vista en la siguiente cita de un libro de texto de histología: “La cuestión de las posibles potencialidades de los pequeños linfocitos es de especial interés… Muchos investigadores creen que estas células son elementos específicamente diferenciados incapaces de un mayor desarrollo” Entre los observadores que han confirmado las observaciones de Metchnikoff y Maximow, se señalan los siguientes: Ziegel, que en sus estudios sobre el edema de la piel y tejidos, asegura que los linfocitos sufren cambios morfológicos y fagocitan; Schwarz; Helly; Fischer; Tschaschin; Downey; Bergel; Danchakoff y Seidlin; Stilwell; Lang; Bloon; y Watson entre otros. A partir del estudio de la literatura anterior, Kolouch (210) considera que es evidente que una importante parte en la formación del exudado inflamatorio celular agudo se adscribe al linfocito, y realiza una serie de experimentos con una técnica original, que permite la simultánea demostración de elementos histógenos y hematógenos, respecto al tiempo y la variación ambiental durante el proceso de la inflamación. Utilizó conejos adultos a los que se les inyecta vía subdérmica albúmina de huevo como sustancia irritante, señalando con violeta de metilo el punto de la inyección, y realiza mediante biopsias un muestreo del tejido inflamado, a determinados periodos de tiempo después de la inyección. Las biopsias obtenidas se fijan y tiñen por métodos habituales en hematología, que permiten una comparación de los elementos hematógenos reconocibles. A las 8 horas, el área inflamada muestra muchos pequeños linfocitos típicos, y a las 14 horas todas las etapas de transición entre linfocito y macrófago, 136 RESULTADOS indicadoras del proceso de transformación del linfocito están presentes. El citoplasma aumenta en cantidad y algunas células muestran pequeños pseudópodos citoplasmáticos, nos referimos a ellas como macrófagos hematógenos. A las 26 horas los macrófagos hematógenos presentan un patrón de cromatina similar a la de macrófagos histógenos. En 76 horas ambas células tienen el mismo aproximado tamaño y morfología. Figura 17. Clasmatocitos 1- Clasmatocitos normales del tejido conectivo subcutáneo. 2- Biopsia del tejido conectivo 2 horas después de un proceso inflamatorio agudo. Clasmatocitos activados mostrando fagocitosis. Clasmatocitos: fagocitos potenciales del tejido conectivo Tomado de: Kolouch, 1939, (210) Kolouch (210) llega dos importantes conclusiones: a) La mayoría de los macrófagos existentes en el exudado asociado con el proceso inflamatorio son de origen hematógeno. 137 RESULTADOS b) La trasformación de linfocito a macrófago ocurre pronto en el curso de la inflamación. A las 4 horas el linfocito origen de muchas células mononucleadas es en parte no visible, y después de las 18 horas de la aparición de la inflamación en un tejido, el linaje celular no puede ser trazado. Y describe someramente el proceso, reconociendo el desconocimiento existente: “Ambos tejidos, con inflamación aguda y crónica, poseen una atracción quimiotáctica positiva para linfocitos hematógenos. En los exudados inflamatorios agudos los linfocitos se trasforman en macrófagos, en contraste con la conservada morfología de éstos en los procesos crónicos. Los factores que subyacen a estos cambios no se conocen”. Los alemanes Branscheid y Ehrhardt escriben una de las publicaciones, a nuestro juicio, más originales e interesantes registrada en 1939 y que lleva por título “El linfocito sano en Jena y su relación con el contenido del suero en sangre” (215). Jena es una ciudad del centro-­‐este de Alemania, en el estado de Turingia, situada en el valle de río Saale, que es famosa por dos motivos; por poseer una de las universidades con más tradición cultural y científica de Alemania, fundada por Juan Federico “el Magnánimo”, y por su importante industria óptica. Branscheid y Ehrhardt consideran que uno de los campos más interesantes de las investigaciones en medicina es la patología geográfica, refiriéndose no a las enfermedades infecciosas o aquellas con ubicación unilateral relacionadas o inducidas por la dieta, como por ejemplo el déficit de vitaminas, sino aquellas en las que intervienen factores específicos del lugar todavía no conocidos. Por ejemplo, dicen: “la apendicitis, muy común en nuestro entorno, es prácticamente desconocida en Asia Oriental; la necrosis hemorrágica aguda pancreática abundante en nuestra región, varía según las zonas y el individuo y la esclerosis 138 RESULTADOS múltiple rara en Japón y China, presenta diferencias significativas entre los estados del Sur y el Norte de Alemania”. Los autores pretenden con este estudio investigar la influencia de factores localizados (geográficos) sobre la composición de la sangre, en particular de las células blancas, y el contenido en cationes del suero. Aceptan como valores normales para leucocitos, neutrófilos, eosinófilos, basófilos, linfocitos y monocitos, los de Naegeli y Schilling, que servirán de base para el estudio: Respecto a los linfocitos, su número varía según los autores de los distintos libros de texto consultados, así por ejemplo: Y hacen referencia a los diferentes valores que se presentan según los países: Las razones que se dan para explicar estas diferencias son muy variadas, pero en general, no fueron satisfactorias ni suficientemente convincentes para estos investigadores. Soler demostró que herbívoros alimentados con carne durante largo tiempo manifestaban una linfopenia, y asegura que los alimentos ricos en carbohidratos aumentan las cifras de linfocitos. No existe duda de la influencia 139 RESULTADOS que ejerce la dieta, y se tuvo en cuenta el componente climático que solo da una explicación parcial a estas diferencias. Sobre el índice leucocitario según la ubicación geográfica hay poca literatura. Wichline encuentra en los soldados estadounidenses a su llegada a Filipinas una media de 21,8 % linfocitos, que después de 8 meses sube a 26,6 % y después de 16 meses es del 33,3 %, y atribuye este cambio al clima tropical y el aumento de la intensidad de radiación solar como desencadenante. Considera que este es un factor a tener en cuenta, ya que se ha demostrado varias veces experimentalmente. Gänsslen realiza la observación en sí mismo, sus familiares y algunos empleados, que cambiaron su residencia de Tübinger a Frankfurt. Igualmente uno de los autores, Ehrhardt, observó que su linfocitemia, en Marburg fue del 36 %, y después de unos meses en Jena aumentó al 51 %, sin cambios en las demás condiciones de vida o dieta. Observaron (215) que el porcentaje de linfocitos en la población de Jena, era mayor que los valores aportados por Naegeli y Schilling, y calcularon un índice linfocitario medio en 200 personas completamente sanas, siendo excluidas aquellas con síntomas leves o inespecíficos, para no influir en la fórmula sanguínea. Además del preciso examen clínico, se realizó un control de la velocidad de sedimentación globular, retirándose del estudio, los hombres con valores de más de 7 mm y las mujeres con más de 10 mm en la primera hora. El valor medio de la cifra de linfocitos en estas 200 personas fue de 2.600 linfocitos, equivalente al 40 %. Aplicando la fórmula Gaussiniana se calculó una desviación constante, resultando que en condiciones normales, los linfocitos sanguíneos en la población de Jena suponen un 40 % con fluctuaciones entre 37-­‐47 %, consideradas dentro de los límites fisiológicos. A continuación los autores (215) inician la búsqueda de compuestos existentes en el área geográfica de Jena que pudieran explicar estos valores, interesándose por el hecho de que la procedencia de las fuentes de suministro de agua en Jena se originan en piedra caliza y dolomita, y fijando su atención en el filtro de arenisca utilizado, que permite mantener la cuota de minerales 140 RESULTADOS especialmente bicarbonato. Consideran que la dureza total del agua del servicio local, informada de 24º a 17,6º y con un 73,3 % de contenido de carbonato de calcio, podría ser una posible explicación. En vista a estos hallazgos Branscheid y Ehrhardt mantienen la hipótesis siguiente: “El aumento de iones de calcio da lugar a una pérdida de iones hidrógeno con un enriquecimiento en valencias alcalinas. Supongamos lo siguiente: debido al suministro continuo de bicarbonato en el agua potable y los alimentos producidos en la tierra, existe en todo el organismo de los habitantes de Jena una conversión a la alcalosis, y por tanto un mayor tono vagal inducido por ésta, que tiene su efecto sobre el recuento sanguíneo local”. Sin embargo, la determinación del contenido de cationes, potasio, calcio y el contenido de sodio en suero mostró valores dentro de la normalidad, tanto absolutos, como relativos, por lo que los autores reconocen no poder demostrar su hipótesis cuando dicen: “la relación entre los valores sanguíneos y el contenido de cationes en el suero todavía no ha podido ser demostrada”. En 1940 bajo el título: “El misterio de la desaparición de los linfocitos” se publica un artículo en La Revista Médica Británica en Diciembre de 1940 (216), que comienza planteando la incógnita existente sobre el destino de linfocitos de esta forma: “En una observación superficial, los linfocitos parecen ser una de las células más insignificante y poco interesantes del cuerpo. Sin embargo, el linfocito es el centro de un profundo misterio, que hasta ahora ha desconcertado a todos los anatomistas y fisiólogos en su intento de resolverlo. El misterio es el siguiente: en un animal como el perro, se estima que más de 200 millones de estas células se incorporan a la sangre, a través del conducto torácico cada hora, y sin embargo, el contenido linfocítico se mantiene constante. ¿Qué sucede con todas estas células?” Se sugieren tres alternativas explicativas: • Que se filtran por los capilares sanguíneos hacia los espacios de los tejidos, y desde aquí de nuevo entran en los capilares linfáticos, circulando de forma continua. Posibilidad que fue rechazada por Yoffey y 141 RESULTADOS Drinker, quienes aseguran que de 30 linfocitos, solo uno vuelve al sistema linfático; los restantes 29 deben ir a otro lugar. • Que los linfocitos se separan por filtración de la sangre en la membrana mucosa de tracto gastrointestinal y desde allí se incorporan a la luz intestinal. Pero al parecer esto ocurriría solo en un pequeño porcentaje de linfocitos. • La tercera alternativa es que son transportados por el torrente sanguíneo a la médula ósea, donde van a desempeñar un papel esencial en la diferenciación de los diversos tipos de corpúsculos sanguíneos. Según esta interpretación el linfocito es en realidad un tipo celular embrionario capaz de diferenciarse para formar células rojas y varios tipos de células blancas. Sin embargo, hay observaciones que lo ponen en duda, como la de Lewis quien asegura que los linfocitos permanecen siempre distintos de los otros tipos de células, sin tendencia a la transformación. En cambio Medawar apunta a la posibilidad de que los linfocitos requieran para ser activados algún factor presente en la médula ósea. El desconocido autor (216) concluye diciendo: “Tal vez la solución final al destino último de los linfocitos, se obtendrá cuando una técnica pueda marcarlos a fin de ser reconocidos donde quieran que vayan. Si el destino sigue siendo un misterio, su significado funcional no es menos oscuro. Hay alguna evidencia de que pueden elaborar anticuerpos, y las infiltraciones linfocíticas en infecciones víricas y bacterianas sugieren algún mecanismo interesante de defensa”. Esta publicación nos pone al día, sobre el estado del conocimiento respecto al linfocito a la fecha del escrito, 1940, las distintas corrientes de pensamiento y la existencia de una conciencia en cuanto al reconocimiento de las limitaciones técnicas del momento. Un estudio sobre “la diferenciación entre linfocitos sarcomatosos y leucémicos en ratones”, es llevado a cabo por varios autores: Kirschbaum, Gardner, Nahigian y Strong en 1940 (217). 142 RESULTADOS Se describe la naturaleza de los tumores (linfosarcoma o leucemia), como resultado de la inoculación de diferentes líneas de linfocitos neoplásicos en hospedadores. Se utilizaron ratas endogénicas de varias cepas (A, F, C3H, CBA y C57), mantenidas por apareamientos hermano-­‐hermana durante generaciones, y se establecieron 16 líneas de transferencia de enfermedad maligna linfoide y mieloide en 5 cepas puras de ratones inoculados. El concepto de la naturaleza neoplásica de la leucemia del ratón se reafirma con estos experimentos, y se obtienen los siguientes resultados: 1-­‐ En 12 líneas de transferencia de franca leucemia, aparecieron tumores subcutáneos cuando se inoculan células leucémicas bajo la piel, y en todos los casos de líneas leucémicas procedentes de casos con lesiones generalizadas, se produce leucemia 2-­‐ En la línea G, establecida de un caso espontáneo de linfosarcoma mediastínico sin enfermedad sistémica, las células procedentes de tumores subcutáneos e intraperitoneales provocados por inoculación en estos sitios, no se propagaron a través de la sangre y linfa, existiendo gradaciones entre líneas de linfosarcoma y leucemia en ratones. Células de la línea G producen linfosarcoma, otras líneas representan etapas intermedias entre linfosarcoma y leucemia 3-­‐ El periodo de latencia se prolongó cuando las células se inocularon vía intravenosa, al parecer el medio intravascular es menos favorable que el extravascular. 4-­‐ Observaciones adicionales mostraron que la incidencia de enfermedad linfoide neoplásica aumenta en ratones que reciben altas dosis de estrógenos. En vista a estos resultados, los autores (217) concluyen: “los hallazgos parecen indicar que son las células y no los hospedadores las responsables de los distintos tipos de crecimiento. En definitiva, nuestras observaciones indican una amplia gama en la capacidad de los distintos linfocitos malignos para invadir la sangre, 143 RESULTADOS dependiendo el carácter invasivo de la célula maligna original. Los factores responsables de este carácter invasivo están aún por determinar”. Christian Bomskov y Kart-­‐Heinz Kreft, de la Universidad de Freiburg, publican en 1940 un estudio sobre “la linfocitosis originada en el timo” (218). El timo es considerado un órgano con capacidad para producir una hormona estimulante de linfocitos. Para comprobar este punto, Bomskov y Kreft inician sus experimentos. Consideran que la edad infantil puede ser también definida en términos de un cuadro sanguíneo determinado, observando la característica linfocitosis de animales tratados con hormona del timo. Se conoce que el tejido del timo sufre un proceso involutivo a partir de la pubertad, que la curva de crecimiento del timo es coincidente con la del tejido linfoide, y que la etapa en la vida de los seres humanos y animales en la que el timo experimenta su mayor desarrollo es la anterior a la pubertad, período de crecimiento que se caracteriza por una linfocitosis fisiológica en la sangre del niño como expresión del efecto del timo. Así, el porcentaje de linfocitos varía a lo largo de la vida, siendo en la edad infantil del 50-­‐60 %, a los 7 años de aproximadamente el 40 %, a los 14 años del 32 % y en el adulto del 25 %. Bomskov y Kreft (218) cuestionan la participación del timo en la linfocitosis sanguínea. Examinaron la sangre de personas normales y ratas normales, y el examen se repitió después de la administración de la hormona del timo y la hormona timotropa de la hipófisis anterior. A partir de los hallazgos registrados en la rata normal durante el descenso fisiológico de los linfocitos con la edad, observaron un fenómeno paralelo en el ser humano, mostrando las ratas infantiles un aumento en el índice linfocitario respecto a las ratas adultas, y comprobaron que podían forzar este infantilismo, prestando linfocitos a la sangre en el animal adulto, con la administración de las hormonas tímica y/o timotropa que provoca una linfocitosis muy marcada en paralelo a una pronunciada leucocitosis. Los autores llegaron a demostrar que: 144 RESULTADOS ! En la rata infantil es característico una linfocitosis, es decir, un mayor número de linfocitos en sangre que en animal adulto. Esta linfocitosis infantil se corresponde con la linfocitosis de la edad del niño. ! Es posible en ratas adultas y en el hombre, provocar un infantilismo sanguíneo, mediante la administración de hormona tímica o bien hormona timotropa de la hipófisis anterior. ! Una única inyección de estas hormonas provoca una linfocitosis aguda, que en ratas fue del 92 % y en humanos del 36 %. 145 RESULTADOS 4.4. LA QUINTA DÉCADA (1941-­‐1950) Después de la Segunda Guerra Mundial (1939-­‐1945) y hasta la década de 1970, los países europeos experimentan un crecimiento económico acelerado, periodo que se conoce como “edad dorada del capitalismo” (219). Entre los factores que constituyen las bases para este crecimiento, hay que destacar el clima de cooperación existente dentro de las naciones y entre las naciones, sumado a una mayor intervención estatal en los asuntos económicos y sociales, lo que se denominó como “Nuevo Orden Económico Internacional”. En Bretoon Woods (Estados Unidos), tuvieron lugar varias reuniones internacionales, donde soviéticos, americanos y británicos diseñaron un “Nuevo Mapa Político Europeo de la Paz”. Se crearon las Naciones Unidas (ONU) con el propósito de erradicar la guerra como medio de solución a los conflictos entre naciones (220)(221). Como resultado de la Conferencia de Bretoon Woods en 1944, surgieron una serie de organismos institucionales, como el Banco Mundial (BM) para ayudar al crecimiento económico, el Fondo Monetario Internacional (FMI) en 1945 como ayuda al funcionamiento del sistema monetario internacional, y el Acuerdo General sobre Comercio y Aranceles (GATT) en 1947, que favorecía el libre comercio con la reducción de aranceles y de barreras arancelarias y para-­‐ arancelarias. En 1950, la población mundial alcanzaba los 2,3 billones, y los avances científicos iban a hacer posible la primera bomba cardiopulmonar, la televisión en color, el microchip, el descubrimiento de la estructura del ADN y el primer trasplante renal con éxito. Este es el marco económico y social donde se desarrolla y prolifera el interés por el conocimiento y donde un pequeño número de estudios experimentales van a ir destinados al conocimiento de las células sanguíneas, entre las que se encuentra el linfocito. En este campo, sin duda, el principal interés en esta década, se centra en el estudio de la relación existente entre anticuerpo y 146 RESULTADOS linfocito, y entre los estudios más novedosos figuran los que relacionan el linfocito con las alteraciones endocrinas y con las enfermedades psiquiátricas. La búsqueda en Scopus con lymphocyt* nos muestra un total de 321 documentos, un 0,0574% con respecto al total de documentos del siglo XX, lo que supone un notable incremento respecto a la década anterior (0,0125%). GRÁFICO 4 La enfermedad inflamatoria del apéndice cecal ha sido durante muchos años, y sigue siendo en esta quinta década, un acontecimiento sorpresivo, misterioso y temido. Quizás fue este afán en la búsqueda de la etiología y del papel desempeñado por la anatomía apendicular como causa de apendicitis, lo que movió a Meyer a realizar su estudio sobre “La hiperplasia linfocítica y lesiones “espontáneas” alimentarias”, en 1942, en el Departamento de Anatomía de la Universidad de Stanford (222). Los nódulos linfoides intestinales habían ya sido observados y descritos en años anteriores, generando toda serie de hipótesis discutibles en cuanto a su función o papel en la patología. En la descripción de nódulo linfocítico Gerlach en 1854, representó una glándula de Peyer solitaria, desprovistas de vellosidades y cubierta por epitelio muy bajo. 147 RESULTADOS Figura 18. Nódulo linfoide solitario de colon. A-glándulas de Lieberkühn. B- muscular de la mucosa. C- nódulo linfático solitario. D- submucosa. E- muscular. F- serosa. Tomado de: Meyer, 1942 (222) Ya Stöhr en 1882, estudiando la amígdala lingual, encontró un epitelio escamoso “enrarecido” con ganglios linfáticos numerosos, y la superficie epitelial cubierta por grandes masas agrupadas de células linfoides, que pensó eran patológicas, pero observando formaciones similares en animales sanos se convenció de lo contrario. Destacó además que los leucocitos que pasaban entre estas células epiteliales también las destruían. Figura 19. Sección vertical del epitelio de una glándula folicular de la lengua de un hombre adulto. 1- superficie epitelial escamosa. 2- espacios en la misma hechos por leucocitos. Tomado de: Meyer, 1942 (222) 148 RESULTADOS Rüdinger en 1891 al estudiar el apéndice humano, describió en primer lugar un epitelio invadido por linfocitos, y segundo el hecho de que cuando un nódulo linfático aumenta de tamaño se extiende más allá de los límites de la muscular de la mucosa, y entre las glándulas de Lieberkühn que quedan a los lados. Observaciones similares fueron realizadas por Davinoff en 1887 y Von Ebner en 1899, en la primera edición del libro de texto de histología de Böhm y Davinoff en 1901 (223), y en el atlas de Sobotta en 1902. Estudios posteriores de nódulos linfáticos de cavidad oral y tracto alimentario, encontraron una similar infiltración y destrucción del epitelio que ocurría siempre en nódulos hiperplásicos, y este fenómeno era frecuente y probablemente universal en apéndices sin enfermedad. Aschoff y Shelly en 1937, comunican idénticas observaciones después de estudiar apéndices extirpados accidentalmente. La ausencia de vellosidades, criptas y células caliciformes sobre nódulos linfoides y agregados, es conocida desde hace tiempo, y también que una marcada hiperplasia linfática puede ocurrir sin una alteración de la arquitectura de la mucosa y sin interferencia con la función de las estructuras afectadas. La atrofia y destrucción de células escamosas y epitelio del tracto digestivo, puede ofrecer una respuesta satisfactoria a la pregunta de Aschoff de: “… porqué los habitantes inofensivos del apéndice de repente pueden causar un ataque agudo”. Las observaciones de Meyer (222), no confirman la creencia de que la causa de la apendicitis se deba a condiciones mecánicas, como una ligera flexión de la porción distal del apéndice, subrayado por Aschoff, quien destacó la poca frecuencia de apendicitis reportada durante la hambruna en Rusia durante los años 1921-­‐1922, y mostró el efecto de la dieta sobre el tejido linfático. Como resultado de su estudio, Meyer asegura: “ No sólo el epitelio, sino la totalidad de la mucosa e incluso porciones de la submucosa, pueden ser afectadas de hiperplasia, incluyendo vellosidades, muscular de la mucosa y músculo liso, y en caso de una marcada hiperplasia difusa, todas estas estructuras pueden estar involucradas. La evidencia indica que el control de la hiperplasia linfática puede señalar un camino hacia el alivio de varias lesiones del tracto digestivo y debería 149 RESULTADOS tomarse en consideración”. Meyer en vista de sus observaciones, defiende que la hiperplasia linfoide es una causa de apendicitis, y considera la obstrucción local apendicular provocada por ésta como favorecedora de aquellos casos con alteración mecánica. Ehrich y Harris, publican sus resultados sobre “La formación de anticuerpos en los ganglios poplíteos del conejo” en 1942 (224). Entre los autores que han apoyado la idea de la formación de anticuerpos en los ganglios linfáticos, McMaster y Hudack (225), parecen ser los únicos que han presentados pruebas que pueden considerarse concluyentes. Detectaron aglutininas en la linfa una semana después de inyectar diversos antígenos, encontrando un título ligeramente más alto en los ganglios linfáticos que en el suero o tejidos del lugar de la inoculación. McMaster y Hudack manifiestan en su trabajo que las aglutininas se forman en los ganglios linfáticos de drenaje de los ratones. Las células que forman anticuerpos aún no han sido identificadas, aunque algunos autores sostienen que las células del sistema reticuloendotelial están involucradas. Rich, Lewis y Winttobe (226), sostienen que las grandes células mononucleadas del bazo, las cuales responden específicamente a la inyección parenteral de proteínas extrañas son sospechosas de estar relacionadas con el proceso de formación de anticuerpos, y aclaran: “no eran histiocitos sino que se asemejaban a linfoblastos, y aunque no eran linfoblastos en el sentido de células linfoides inmaduras, eran linfocitos agrandados en gran medida porque realizaban alguna función especial”. Ehrich y Harris creyeron conveniente, para un conocimiento más preciso, comparar la producción de anticuerpos y las células del ganglio linfático, y analizar los cambios morfológicos simultáneos en el tejido linfático. Los ganglios poplíteos eran los adecuados, ya que son los únicos que drenan las patas traseras y son lo suficientemente grandes para permitir un estudio 150 RESULTADOS separado de todas las partes del sistema: tejido local inyectado, linfa aferente, ganglio linfático poplíteo, linfa eferente y suero. Bacilos tifoideos y eritrocitos de oveja fueron los antígenos inyectados vía subcutánea en los pies de conejos Chinchilla machos. La subsiguiente formación de aglutininas y hemolisinas en el ganglio linfático poplíteo se comparó con el número de linfocitos en el linfático eferente y con los cambios histológicos ocurridos en dicho ganglio Los anticuerpos determinados fueron la aglutinina de fiebre tifoidea, hemolisina de eritrocitos y aglutinina de eritrocitos. En el estudio se valoró: a) El tiempo y sitio de aparición de anticuerpos. b) El título de anticuerpos relativos en linfa y suero. c) La relación del título de anticuerpos de los ganglios linfáticos con las células de los ganglios y el peso de los ganglios linfáticos. Los anticuerpos comienzan a aparecer en la linfa eferente de 2-­‐4 días después de la inyección del antígeno, alcanzando su título más alto pasados 6 días. Esto fue precedido por un fuerte aumento en la producción de linfocitos. La hiperplasia en el ganglio fue en principio de tipo difuso, pero más tarde fue sustituida por grandes centros germinales El número de células aumentó de un promedio de 17.000 por cmm antes de la inyección a un promedio de 50.000 a 60.000 al tercer día, manteniéndose este nivel durante al menos 3 semanas. El peso de los ganglios linfáticos se elevó de un promedio de 0,19 gramos a casi 1 gramo en el noveno día. Los resultados fueron similares con los dos antígenos empleados. 151 RESULTADOS Figura 20. Títulos de anticuerpo y número de linfocitos en el linfoide eferente comparado con el peso del ganglio linfático después de la inyección de bacilo tifoide en pie izquierdo y eritrocitos de oveja en pie derecho. Tomado de: Ehrich y Harris, 1942 (224). Cuando se estudió la naturaleza de las células en los ganglios mediante frotis linfáticos, se encontró que el porcentaje de pequeños linfocitos cae del 99% al 93% durante la primera semana del experimento, mientras que el porcentaje de linfocitos grandes aumenta de un 2-­‐3% a 6-­‐11%. En todos los casos los cambios morfológicos en los ganglios comenzaron con una infiltración granulocítica y aparición de algunos monocitos, seguidamente tiene lugar una hiperplasia linfoide más bien difusa, con o sin nódulos secundarios sólidos, siendo reemplazada en el cuarto día por una enorme hiperplasia con nódulos secundarios tipo Flemming (centros germinales), que duró hasta finalizar el experimento. 152 RESULTADOS En la interpretación de los resultados (224) hay que considerar dos cuestiones: si el lugar de producción de anticuerpos se corresponde con los centros germinales de los ganglios, y si son los linfocitos o bien células reticuloendoteliales las responsables de su producción. En general, las células reticuloendoteliales se mantuvieron en un discreto número, no aumentando de forma paralela a los títulos de anticuerpos. Por otro lado los linfocitos aumentaron enormemente su número, sobre todo en la linfa eferente, y esto tuvo lugar antes de que los anticuerpos hicieran su aparición. Estas observaciones conducen a Ehrich y Harris a manifestar que “…los linfocitos pueden desempeñar un importante papel en la formación de anticuerpos. Pueden tener la facultad de absorber antígenos disueltos o sus productos de degradación, y tal vez podrían entrar en acción sólo después de que la materia prima haya sido preparada adecuadamente por la acción de fagocitos u otros mecanismos”. Sea como fuere, los resultados se prestan poco a la idea de que los anticuerpos son productos de las células reticuloendoteliales por sí solas. Ehrich y Harris llaman la atención sobre el hecho de que la respuesta tisular que acompañó a la formación de anticuerpos fuera principalmente de tipo linfocítica, y apunta al linfocito como un factor importante en la formación de anticuerpos. En un continuo afán por conocer el destino de los linfocitos, Yoffey (227) en 1942 publica: “Enterectomia y linfocitos sanguíneos”. En un sujeto saludable, las células blancas de la sangre se mantienen a un nivel bastante constante. Para lograr este equilibrio, Naegeli en 1931 postulaba un control hormonal, pero el exhaustivo trabajo de Hoff en 1934 mostró la ignorancia que todavía se tenía sobre la naturaleza del mecanismo de control. Este desconocimiento se aplica a todas las células blancas sanguíneas, incluyendo linfocitos, a pesar de que para éstos existen algunos datos disponibles como el número de ellos que entran en la sangre (204). No obstante, seguimos sin saber el destino final de los linfocitos, aunque se han propuesto varias hipótesis. Bunting y Huston en 1921 (120), sugieren que los linfocitos desaparecen de la sangre por migración al lumen del tubo digestivo. Si esto fuera así, la eliminación de una gran parte del canal alimenticio debería 153 RESULTADOS interferir con el proceso de eliminación, y aumentar por tanto el número de linfocitos en sangre. Ésta es la hipótesis de trabajo de Yoffey. El experimento se realizó en perros, a los que se les extirpó la totalidad del intestino delgado distal al duodeno junto a la mayor parte del intestino grueso. Se extrae sangre en un primer momento, justo después de la intervención (A), y pasadas 2-­‐3 horas (B). Se realizaron recuentos totales y diferenciales de glóbulos blancos y rojos. En el intervalo de tiempo entre A y B, los linfocitos en sangre disminuyeron de 1.970 a 1.400 (B es el 71% de A), mientras que los neutrófilos elevaron su número de 2.170 a 13.150. Yoffey reconoce que el experimento es fácilmente criticable, y expresa sus dudas: “¿El tiempo entre A y B es suficiente para que ocurra un cambio apreciable en los linfocitos circulantes?”, “¿El intestino extirpado es suficiente para probar la hipótesis?”, “¿Hay un marcado cambio en el volumen de sangre durante el experimento?” Preguntas que reflejan la complejidad del problema, y que en vista de los resultados nada concluyentes del trabajo, recomiendan la realización de nuevos experimentos que tengan en consideración todos estos puntos. Los resultados (227) muestran que la enterectomía no es seguida por un cambio constante de los linfocitos en sangre, sino que aumentan o disminuyen aproximadamente con igual frecuencia. Se supone que tendría que ocurrir un aumento de linfocitos, ya que una disminución sería difícilmente explicable. Pero si los linfocitos abandonan la sangre de otra manera que no sea a través del intestino, la enterectomía no va a interferir con el proceso de eliminación, y el número de linfocitos circulantes no debería modificar su número o bien disminuir si no hay entrada para reemplazar a los que se pierden, pero no aumentar. Ya que en algunos experimentos hay un aumento después de la enterectomía, surge la posibilidad de otra entrada de linfocitos en sangre que no sea a través del conducto linfático, y se apunta al bazo como fuente adicional de linfocitos que pasarían directamente a la sangre, que al no utilizar la vía linfática, no estaría interferida por la enterectomía; sin embargo, los 154 RESULTADOS trabajos de Blalock en 1937, mostraron que la vía linfática es la principal fuente de linfocitos en sangre, ya que una obstrucción linfática completa da como resultado una disminución prácticamente a cero de linfocitos sanguíneos. El trabajo de Yoffey contiene demasiadas preguntas sin respuestas satisfactorias, que nos llevan a unos resultados no concluyentes. En definitiva seguimos sin conocer a dónde se dirigen, cómo abandonan la sangre, cuál es el destino final de los linfocitos. Sobre “La curva leucocitaria, el índice monocito/linfocito (M/L) y su importancia para el pronóstico y tratamiento de la tuberculosis infantil”, trata el trabajo de Eva Bergemann en 1943 (228). En 37 niños, con tuberculosis pulmonar y extrapulmonar, Bergemann estudia el comportamiento del hemograma en relación a las células blancas de la sangre, y la importancia y significado del índice monocito/linfocito (M/L), al que había que someter a evaluación antes de ser utilizado en clínica. El conocimiento obtenido en este trabajó coincidió esencialmente con otros hallazgos clínicos, sin que pueda interpretarse con ello una generalización de su uso a largo plazo. Considera que las investigaciones realizadas en niños pequeños no han tenido unos resultados prácticos aplicables. Este trabajo de Bergemann es otro de los muchos estudios de este tipo que durante varias décadas se realizan con la esperanza de encontrar un indicador fiable y de aplicación clínica en la lucha contra la temida tuberculosis, sin que hasta el momento los resultados obtenidos fueran de gran ayuda. En 1944 Harris, Grimm, Mertens y Ehrich, publican sus estudios sobre “El papel del linfocito en la formación de anticuerpos” (229). El trabajo anterior de Ehrich y Harris (224), apuntó al linfocito como un factor importante en la formación de anticuerpos. Ahora, el objetivo es determinar los anticuerpos dentro de los linfocitos, en la linfa eferente y en el ganglio linfático. 155 RESULTADOS Se utilizaron conejos Chinchilla, a los que se les inyectaron antígenos (bacilos tifoideos o eritrocitos de oveja) vía subcutánea en las patas traseras. Se valoró el titulo de anticuerpos en plasma linfático (linfoplasma) y en el extracto celular linfático, comparando las concentraciones en ambos medios. Los resultados mostraron que los valores en las concentraciones de anticuerpos fueron sistemáticamente mayores en los extractos celulares que en el linfoplasma, observándose la mayor diferencia a los cinco días. Surge la pregunta de si los anticuerpos se forman en el interior o en la superficie de los linfocitos, o bien si los anticuerpos de la linfa derivan de alguna otra fuente y secundariamente se absorben o adsorben por los linfocitos. Los experimentos realizados in vitro, de la absorción o adsorción de anticuerpos en linfocitos, ponen de manifiesto que los anticuerpos elaborados por linfocitos habían pasado al plasma linfático, sin embargo no se alcanzó el equilibrio entre ellos. Además no hubo absorción de anticuerpos por parte de los linfocitos, incluso cuando existía un título muy alto en el fluido linfático, y en ningún caso hubo disminución del título de anticuerpos en el linfoplasma durante la incubación. Con el fin de probar directamente la función de los linfocitos in vivo, se realizaron una serie de experimentos en conejos, a los cuales bajo anestesia se les aisló el ganglio linfático poplíteo y su vaso linfático. Después de ligar el vaso linfático eferente, se inyecta en el ganglio suero de conejo con un título alto de anticuerpos contra eritrocitos de oveja, y tras 3-­‐5 horas de incubación, se aspira la linfa desde el vaso linfático eferente. Las anotaciones (229) que realizan los autores una vez valorados los resultados fueron: • El título de anticuerpos en el extracto celular fue sustancialmente más alto y consistente que la del fluido. La incubación in vitro de los linfocitos que contienen una sola especie de anticuerpos con plasma 156 RESULTADOS linfático conteniendo otro, mostró que los anticuerpos pasan de las células al sobrenadante, para alcanzar un casi equilibrio. • Se obtuvieron resultados similares cuando los linfocitos normales se incubaron in vivo en su propio linfático, al que se había añadido fluido con anticuerpos. Se encontró de nuevo que los anticuerpos no fueron adsorbidos ni absorbidos por los linfocitos. Los resultados de este estudio robustecen las conclusiones recogidas en el documento anterior de Ehrich y Harris, y parecen mostrar por vez primera, que el linfocito tiene como función la formación de anticuerpos. Comparando la metodología de este estudio con el anterior de Ehrich y Harris de 1942, podemos advertir, en nuestra opinión, un gran avance, en la forma de abordar el estudio de la formación de anticuerpos. En esta segunda investigación, el análisis es más detallado, con pruebas directas más precisas de la medida de anticuerpos; por ejemplo, se compara el título de anticuerpos entre el extracto celular y el linfoplasma en una sola prueba, al mismo tiempo y utilizando los mismos reactivos, mientras que en primer trabajo realizado por Ehrich y Harris, las conclusiones se hicieron en base a la observación de la hiperplasia linfática en el ganglio regional donde tuvo lugar la inoculación del antígeno, el aumento de los linfocitos en ese ganglio y su coincidencia con la aparición de anticuerpos, hechos todos ellos mucho más generales y groseros, con una mayor dificultad en cuanto a la interpretación de los resultados y la emisión de conclusiones. No obstante, en este segundo trabajo, en algunos de los experimentos, como los realizados in vitro, no se ha considerado con toda su trascendencia el hecho de que los linfocitos son células vivas, y pequeñas modificaciones en sus condiciones fisiológicas pueden alterar su comportamiento. En 1944 sale a la luz otro de los trabajos de J.M.Yoffey (230), que junto a Josephine Parnell estudia “El contenido linfocítico de la médula ósea del conejo”. Se realiza una revisión de la literatura sobre la producción de linfocitos, en este 157 RESULTADOS sentido hay que recordar las aportaciones de Drinker y Yoffey (197), Sanders, Florey y Barnes, Kindred y de Erf en 1940 (231). Un resumen de los hechos esenciales en este conocimiento es el que sigue: los linfocitos entran diariamente en la sangre en gran número, y desaparecen de ella igualmente en gran número, por lo que el nivel de linfocitos circulantes permanece aproximadamente constante. Cómo salen y a dónde van estos linfocitos circulantes sigue siendo un misterio, un problema no resuelto. No parece que se destruyan en la sangre (Bunting y Huston, 1921), y tampoco se excretan en la luz del tracto alimentario. Los experimentos de Erf muestran que los linfocitos pueden desaparecer rápidamente de la sangre, incluso en ausencia de bazo y la mayor parte del tracto digestivo. El único destino que hasta el momento se ha sugerido es la médula ósea. Para su estudio se realiza una suspensión de células de la médula ósea en plasma, y esta suspensión es tratada exactamente igual que si fuese sangre, con el fin de obtener recuentos absolutos y diferenciales, teniendo en cuenta algunos factores que pueden alterar la observación, como que durante este proceso, algunas de las células más frágiles pueden ser destruidas por completo y el recuento diferencial pueda verse afectado. Se realiza una identificación celular, observando que la mayoría de linfocitos son pequeños, y no hay grandes linfocitos registrados en los recuentos diferenciales. La diferenciación entre linfocitos pequeños y medianos ha sido hecha en base a criterios morfológicos. Así por ejemplo, se observa que los núcleos de los pequeños linfocitos son regulares y característicamente policromáticos, mientras que los linfocitos medianos tienden a tener un núcleo más abierto respecto a la textura de la cromatina, es decir son leptocromáticos. (Fig. 21). La distinción entre linfocito pequeño y mediano es algo arbitraria: “… generalmente, las células denominadas linfocitos medianos son algo mayores que los pequeños linfocitos…” ya que no se han determinado unos límites en cuanto al tamaño para diferenciarlos, por ser difícil de realizar en la práctica. 158 RESULTADOS Figura 21. Diversos tipos de linfocitos. 1-15: pequeño linfocito; médula ósea. 16-20: pequeño linfocito; ganglio linfático 21-25: linfocitos medianos; médula ósea. 26: eritrocito 27-28: pequeño linfocito; sangre. 29-30: linfocito mediano; sangre Tomado de: Yoffey y Panell, 1944 (230). La aportación más importante de este trabajo es la descripción de una técnica para la obtención de recuento absoluto de células nucleadas de la médula ósea y la determinación porcentual de las células encontradas en la médula ósea costal de conejo donde se hallaron 469.000 células nucleadas por ml, de las cuales 61.000 eran linfocitos; el 75% de éstos eran linfocitos pequeños y el 25% medianos. Los autores realizan la siguiente observación: “el contenido total en linfocitos de la médula ósea del conejo es del mismo orden que el número de linfocitos que entran en la sangre todos los días a través del conducto torácico”, observación que nos lleva a formular las siguientes preguntas: ¿significa que la médula ósea es el único órgano productor de linfocitos? Si la 159 RESULTADOS respuesta fuese afirmativa ¿cual sería el papel de los restantes órganos linfoides? Preguntas para las que todavía no existían respuestas. La “relación existente entre linfocitos y cáncer”, se publicó en 1945 por Margaret A. Kelsall (232). Los linfocitos se han considerado en muchos estudios sobre el cáncer, pero prácticamente todos los experimentos se han desarrollado desde el punto de vista del papel de esa célula en la inmunidad a los trasplantes tumorales. Encontrada una relación directa entre la cantidad de tejido linfoide en el intestino y la incidencia de tumores mamarios en varias cepas de ratones, surgió la idea de que el linfocito podría contribuir a facilitar el crecimiento de tumores. Kelsall se basó en la asociación histológica existente entre la presencia de linfocitos y tumores, entre el crecimiento e incidencia de tumores y las condiciones que dan lugar a un aumento o disminución en el número de linfocitos en un tejido o en todo el cuerpo: “los linfocitos están presentes en las metástasis de carcinomas mamarios, el los tejidos que contienen el inicial tumor mamario y en focos tumorales causados por irritación” Analiza una serie de datos, recogidos sobre metástasis de carcinoma mamario en mujeres y en experimentos realizados en animales, que fueron interpretados como indicativos de una conexión entre tumores de mama y presencia de linfocitos. Entre estos datos figuran: “… las metástasis ocurren con mayor frecuencia en ganglios linfáticos, órgano cuyo componente principal son linfocitos”. Y añade como prueba: “Además, los linfocitos están presentes en las metástasis hepáticas, en focos neoplásicos y entre el tejido conectivo y el epitelio en la enfermedad de Paget. Ninguna célula es tan numerosa en los fluidos y órganos como el linfocito, y es un elemento histológico dominante en bazo y timo”. El aumento de linfocitos circulantes que se produce en algunos desequilibrios de las glándulas endocrinas son interpretados como una relación positiva entre linfocitos y desarrollo tumoral: “Las hormonas sexuales 160 RESULTADOS son factores conocidos en la producción de tumores, desequilibrios en ellas estimulan el crecimiento del tejido linfoide”. El término linfocitosis lo aplica, no sólo a un aumento de los linfocitos circulantes, sino también a un aumento en el número de linfocitos dentro de un órgano o tejido. Refiere la existencia datos que indican que un factor causante de tumores es la lesión irritante celular, ya sea química, física, por radiación o por ciertos parásitos y virus, y establece una posible relación etiológica entre linfocitos y cáncer: “una linfocitosis ocurre en algunas etapas de la sífilis, hecho que pudiera estar relacionado con el aumento de carcinoma de labio en sifilíticos”. Por otro lado, también alude a la relación entre linfopenia y disminución en la incidencia y crecimiento de tumores, como ocurre, por ejemplo, en la inanición crónica, donde disminuye el número de linfocitos en sangre y se observa una disminución en el crecimiento de tumores. La inanición, ha sido mencionada por muchos autores como causante de una reducción en el número de linfocitos y de una involución de órganos linfoides. Considera que la característica más evidente del pequeño linfocito es su enorme núcleo en proporción a la cantidad de citoplasma. La tinción basófila revela que los linfocitos contienen más ácido nucleico que la mayoría de células. Stowell (233), quien realizó una amplia revisión de la literatura sobre el papel del ácido timonucleico en los tumores, concluye que una perturbación en las nucleoproteínas constituye la base para una causa intracelular de neoplasia: “el linfocito puede jugar un papel en el cáncer, a través de la modificación de las nucleoproteínas, y ser la fuente de la mayor cantidad de ácido timonucleico encontrado en carcinomas” Este trabajo de Kelsall (232) pone de manifiesto una vez más, el desconocimiento que todavía existía sobre las funciones del linfocito y sobre la naturaleza de la célula cancerosa. Creemos que no explora todas las posibilidades respecto a las explicaciones de los hechos observados. En nuestra opinión la aportación más relevante, que aunque algo errónea es muy interesante, es la posibilidad de que el linfocito pueda tener un papel en la 161 RESULTADOS modificación de nucleoproteínas, la sugerencia de que la causa del cáncer podría encontrarse en el núcleo celular, concretamente en las nucleoproteínas. En el mismo año 1945, y en la misma revista científica Science, Kass publica un artículo sobre “la ocurrencia de suero normal de gammaglobulinas en linfocitos humanos” (234). Kass se lamenta de que a pesar del interés y la importancia del tema, hay muy poca información sobre el origen de las globulinas del suero, y expone el siguiente razonamiento: “Dado que la γ-­‐globulina sérica humana contiene casi toda la actividad de los anticuerpos del plasma, y aunque numerosos intentos de demostrar diferencias antigénicas y físicas entre globulinas inmunes y no-­‐ inmunes, no han tenido éxito, no parece haber duda de que los anticuerpos son globulinas séricas modificadas, y que en el hombre por lo menos, las principales globulinas modificadas son las γ-­‐globulinas. Si los anticuerpos se sintetizan en los linfocitos, el suero normal de γ-­‐globulinas debería también estar presente en ellos” Kass utiliza en su estudio una γ-­‐globulina purificada por él mismo, que en un 98-­‐100% era γ-­‐globulina pura al examen electroforético. Obtuvo anticuerpos de conejo preparados para esta altamente purificada γ-­‐globulina, observando que los extractos de linfocitos humanos reaccionan específicamente con estos antisueros. Realizó una suspensión con ganglios linfáticos mesentéricos tanto de hombres como mujeres, fallecidos por motivos diversos. El filtrado de dicha suspensión contenía un 95% de pequeños linfocitos y un 5% de grandes linfocitos. Con el filtrado se realizan varios experimentos, a fin de estudiar su comportamiento frente al suero y antisuero de γ-­‐globulinas. De las observaciones realizadas, Kass (234) concluye que: “La ocurrencia de γ-­‐ globulina normal en linfocitos, cuando se combina con la demostración de hiperplasia linfoide y se acompaña de la producción de anticuerpos, junto con el fallo de los linfocitos para adsorber anticuerpos in vitro, sugiere que la alteración 162 RESULTADOS específica de la γ-­‐globulina para hacer que la molécula se convierta en reactiva hacia un anticuerpo dado, se produce dentro de los linfocitos” Reinhardt y Chon Hao Li (235) publican, también en 1945, un breve artículo sobre “la disminución del contenido linfocítico del conducto torácico por hormonas adrenocorticotropas”. Reinhardt y Chon Hao Li llevan a cabo la administración de hormona adrenocortocotrópica (ACTH) a ratas hembras normales y adrenalectomizadas. Basan sus investigaciones en la evidencia de la existencia de una relación recíproca entre la función del córtex adrenal y el peso del tejido linfoide y tímico del cuerpo. La adrenalectomía da lugar a un aumento del peso del timo y ganglios linfáticos en la rata. En cambio, la estimulación del córtex adrenal por administración de ACTH o de extractos adrenales ha producido una rápida disminución en el peso de timo y ganglios linfáticos en el ratón y la rata, según demostraron por los estudios de Dougherty y White en 1943. Las mediciones realizadas por Reinhardt y Chon Hao Li (235) en el conducto torácico de las ratas para medir el flujo de linfocitos, antes y después de la inyección de hormonas, pusieron en evidencia que “la administración de ACTH produjo un rápido descenso del número de linfocitos en el conducto torácico. Esta disminución tuvo lugar en los primeros 15-­‐30 minutos, por lo general era mayor de un 50% y persistió durante la duración del experimento, de 4-­‐10 horas. En cambio, en el animal adrenalectomizado, la administración de ACTH no produjo un apreciable efecto”. Los autores sugieren la existencia de un control adrenal directo en el nivel de linfocitos existentes en sangre. En dicho control participarían la hipófisis anterior y corteza suprarrenal: … “debe desviar la atención a la función de la hipófisis anterior y corteza suprarrenal en diversos estados de enfermedad acompañados por linfopenias relativas y absolutas inexplicables hasta ahora”. Un nuevo estudio dirigido a la valoración de la “división mitótica y degeneración de linfocitos en las células del epitelio intestinal en el ratón”, es realizado por 163 RESULTADOS Warren Andrew y Nancy Valérie Andrew, del Departamento de Anatomía del Colegio de Médicos del Suroeste, Dallas, Texas en 1945 (236). Entre las numerosas funciones asignadas al linfocito, se encuentra la de protección del organismo frente a bacterias y toxinas, en palabras de Maximow: “es muy probable que la función de defensa de los nodos linfáticos, especialmente su capacidad para reducir la virulencia de microorganismos patógenos, se deba en parte a la actividad de los linfocitos”. Autores como Moor y Newport (237), aseguran que probablemente desempeñan un papel importante en la inmunidad, en los mecanismos de fijación de tóxicos celulares, especialmente toxinas bacterianas, produciendo a su vez sustancias antitóxicas. Entre otras funciones sugeridas para el linfocito está la de agente con actividad lipolítica, especialmente en tuberculosis, sífilis y actinomicosis, defiende Bergel (126). Según Murphy y Nakahara (106), el linfocito juega un importante papel en la resistencia al cáncer experimental en el ratón. Basados en estos estudios y en los de investigadores como Reding, Wiseman y Erf (174) entre otros, Andrew y Andrew, según las observaciones descritas en el presente trabajo, desarrollan un nuevo concepto de la posición y la actividad de los linfocitos en el epitelio intestinal. Utilizaron ratones negros de varias edades de la cepa C57, que se sacrifican después de una semana de permanencia en iguales condiciones de vida y alimentación. El estómago y duodeno se retiran y preparan para su estudio histológico, haciendo un recuento cuidadoso de las células para determinar las figuras mitóticas. 200 células epiteliales de vellosidades y 200 células de las criptas de Lieberkühn fueron estudiadas bajo aceite de inmersión, para cada uno de los 10 animales de experimentación. Observaron (236) lo siguiente: ! La abundancia de células linfoides en la capa epitelial. Un 6 a 16,7% de las células epiteliales en las criptas, muestran 1 linfocito dentro de su citoplasma, mientras que en las vellosidades un 4,5 a 36,5% de las células epiteliales contienen 1 linfocito. (Fig 22) 164 RESULTADOS ! Variedad en la apariencia de los linfocitos. Esta variedad es tan grande, que es necesario revisar casos individuales con aceite de inmersión, para identificar linfocitos alterados. ! Mayor frecuencia de mitosis de linfocitos dentro de las células de las criptas que dentro de las vellosidades. ! Los signos de degeneración del linfocito consisten en presencia de núcleo picnótico, vacuolización o reticulación del material nuclear, o bien, menos frecuentemente, agrupamiento del material basófilo del núcleo. De estas observaciones, los autores destacan hechos de interés como que el linfocito en la capa epitelial del duodeno del ratón es, al menos en gran parte, de posición intracelular, que existen a varios niveles en las células epiteliales, y por lo general, sufren un cambio degenerativo a medida que pasan de la parte basal hacia la luz intestinal. Describen los tipos de degeneración, que puede manifestarse como picnosis, como “agrupamiento” de la cromatina o como vacuolización del núcleo hasta la ruptura de la membrana nuclear. Aseguran que el paso de los linfocitos dentro de las células es pasivo; sin embargo, probablemente alcanzan la célula y penetran en ella con movimientos ameboides, no existiendo evidencia de que los linfocitos pasen más allá de la luz intestinal. Su destino normal es la degeneración dentro de la célula epitelial y su excreción a la luz. Respecto a la división mitótica manifiestan: “la división mitótica de los linfocitos, tanto normales como aberrantes, se produce abundantemente en las criptas de Lieberkühn, pero no en las vellosidades” y explican todos estos hechos:”… de la función de los linfocitos durante su paso a través del epitelio intestinal, es que son células defensivas, acorde con su localización y con la función del tejido linfoide en el organismo” 165 RESULTADOS Figura 22. Linfocito intraepitelial. Linfocito muy alterado justo antes de la extrusión en la luz intestinal. Picnosis e hichamiento del núcleo. Tomado de: Andrew, 1945 (236). Un hecho llamativo es la severidad con la que los autores abordan la existencia de una mayor frecuencia de mitosis de linfocitos dentro de las células de las criptas de Lieberkühn: “Hay cincuenta figuras mitóticas encontradas dentro de las 2000 células epiteliales de las criptas, y únicamente tres entre las 2000 células epiteliales de las vellosidades”. En su estudio, los autores dicen tomar como referencia la posición del núcleo para identificar a las células epiteliales; pero, ¿y si esta referencia se pierde como ocurre cuando una célula entra en mitosis? ¿Cómo diferenciar con seguridad las mitosis de linfocitos de las mitosis de las células epiteliales? Este punto es ampliamente debatido en la discusión. Los autores defienden su postura con argumentos como el elevado porcentaje de mitosis que se observan junto a otro núcleo celular y no en su lugar, o que los cromosomas observados en anafase son más pequeños que los esperados para células epiteliales. Sin embargo, Loreti (238), en un trabajo sobre la presencia de linfocitos en el epitelio de peces, describe la ocurrencia de mitosis abundantes en regiones de migración linfocitaria en amígdala y riñón, y asegura: “… son de células epiteliales”, y según Bremer y Weatherford (239) refiriéndose a la célula en mitosis, dicen: “el plano de división es un 166 RESULTADOS ángulo recto con el eje mayor de la célula, y después de la mitosis, los núcleos vuelven de nuevo a la posición basal. Los linfocitos que se abren camino entre las células epiteliales se ven con frecuencia excesivamente teñidos y pueden ser fácilmente confundidos con figuras de mitosis”. A pesar de que el tema no queda totalmente aclarado, Andrew y Andrew (236) plantean una hipótesis: “la degeneración y división mitótica de los linfocitos como fenómeno visible en las células epiteliales del duodeno del ratón puede representar una condición y estado de actividad de los linfocitos similar a la observada en los nódulos secundarios (centros germinales de Flemming) del tejido linfoide compacto”. Hoy sabemos un poco más sobre la biología y fisiología celular que nos puede ayudar en este dilema. Sabemos que la porción basal de las criptas contiene células pluripotenciales y que los enterocitos proceden de las mitosis activas de estas células indiferenciadas presentes en las criptas de Lieberkühn. También conocemos la importancia que tiene el número de linfocitos de las vellosidades intestinales. La linfocitosis intraepitelial es un marcador histológico de enfermedad celíaca, incluida en los criterios de la clasificación de Marsh para la celiaquía; y su importancia se entiende si se consideran algunas de las complicaciones de la enfermedad, como el linfoma de células T. Estos linfocitos tienen una función en efecto defensiva, como aseguran los autores, pero también intervienen en un complejo mecanismo inmunológico. Este trabajo tiene el mérito de ser uno de los trabajos pioneros en el estudio de los linfocitos intraepiteliales, y ser un punto de partida para futuros estudios. “El destino de las partículas antigénicas inyectadas en relación con la formación de anticuerpos”, es otra de las publicaciones que en 1946 realizaron Harris y Ehrich (240). Los resultados de varios tipos de experimentos realizados con anterioridad indicaban que los anticuerpos que se encontraban dentro de los linfocitos, no fueron absorbidos o concentrados desde el medio ambiente, sino que presumiblemente surgieron dentro del linfocito. Además, los anticuerpos no se encuentran en los granulocitos o macrófagos del exudado inflamatorio en el 167 RESULTADOS momento de mayor producción de anticuerpos; ello es la base de la creencia de que granulocitos o macrófagos no son el lugar de formación de anticuerpos. Harris y Ehrich establecen un razonamiento que les conducirá a una serie de experimentos: “Dado que los linfocitos no son células fagocíticas, y sin embargo la evidencia apunta a la síntesis de anticuerpos por esa célula, parece probable que las partículas antigénicas utilizadas en los experimentos deben someterse a un cambio en la estructura de tal manera que sean solubles”. Los autores (240) llevaron a cabo su estudio para analizar el tejido local y el tejido linfático, en el periodo entre la inyección del material antigénico y la aparición de anticuerpos en el ganglio. El método utilizado es el mismo que en experimentos anteriores. Los antígenos fueron eritrocitos de oveja y bacilos. Se obtienen extractos del lugar de la inoculación de antígenos y de los ganglios linfáticos de drenaje. Los test para determinar la concentración del antígeno específico, se realizan mediante inhibición de la hemólisis de eritrocitos de oveja, la inhibición de la aglutinación de los organismos bacilares y la fijación del complemento. Los resultados mostraron: 1-­‐ Que puede ser identificada una sustancia soluble de la misma especificidad inmunológica que el material antigénico inoculado, en extractos del tejido inyectado, en los ganglios linfáticos regionales y en la linfa eferente de ese nodo. 2-­‐ Que la concentración de este material soluble disminuye lentamente en el tejido inyectado en el curso de pocos días después de la inoculación, y desaparece cuando tiene lugar la aparición de anticuerpos. Se presenta evidencia de que la sustancia inmunológicamente activa deriva del material antigénico inyectado por un proceso fisiológico, mediante el cual los antígenos originariamente situados en las células son puestos a disposición de los linfocitos. 168 RESULTADOS Unos meses antes, en ese mismo año, Ehrich y Harris junto a Mertens publicaron otro de sus trabajos, esta vez sobre “la ausencia de anticuerpos en los macrófagos durante la máxima producción de los mismos” (241). En experimentos previos se descubrió que las células de los ganglios, en muchos casos contenían de 8-­‐16 veces más anticuerpos que el plasma, concluyéndose que el papel del linfocito era decisivo en la formación de anticuerpos. Cuando hablan de linfocitos se refieren al pequeño linfocito y sus habituales predecesores. Si otras células linfoides, tales como células plasmáticas, también producen anticuerpos u otras globulinas no ha sido determinado. Hasta ahora, el linfocito es la única célula en la que se han demostrado anticuerpos, sin embargo, no se ha descartado que otras células presentes en la inflamación también puedan sintetizarlos. En esta comunicación se trata de estudiar el papel del macrófago en la formación de anticuerpos. Se siguió el mismo método que en experimentos anteriores, empleando conejos y antígenos de bacilos de disentería. Las observaciones mostraron que: 1-­‐ Después de la inyección del antígeno de la disentería, en la pata trasera del conejo, se recuperaron cantidades considerables de anticuerpos en el nodo linfático poplíteo, mientras que el lugar de la inoculación rico en granulocitos y macrófagos, presentó cantidades insignificantes de anticuerpos. 2-­‐ La inyección de varios antígenos en la cavidad abdominal, no va seguida de la aparición de anticuerpos en los granulocitos y macrófagos aislados del exudado peritoneal, mientras que el fluido sobrenadante puso de manifiesto títulos paralelos a los encontrados en la sangre. Según Ehrich, Harris y Mertens (241), estos resultados parecen mostrar que el macrófago no sintetiza aglutininas contra la disentería (bacilos de la fiebre tifoidea): “De las observaciones realizadas se desprende que, al menos la síntesis 169 RESULTADOS de anticuerpos no es una función de los macrófagos”, puntualizan al final de su trabajo. Los autores, sin embargo, después de una discusión sobre el tema, llegan a un punto de encuentro con el descubrimiento que Metchnikoff realizó más de medio siglo antes. La teoría reticuloendotelial de la formación de anticuerpos, basada en las observaciones de Metchnikoff sobre el hecho de que las células reticuloendoteliales engullen y digieren el material antigénico, y que como resultado de esta fagocitosis y digestión tiene lugar la formación de anticuerpos, cobra ahora interés, a pesar de no existir evidencias, ya que nunca se han identificado los productos de la digestión de los macrófagos. Ehrich, Harris y Mertens (241) demuestran en estos experimentos que los macrófagos no desarrollan aglutininas contra los antígenos inoculados, pero ¿y si los anticuerpos sintetizados son de naturaleza distinta a las aglutininas? Pregunta que reconocen no estar en situación de contestar, pero dan algunas explicaciones que los acercan a Metchnikoff: “El hecho de que el citoplasma de estas células sea acidófilo también habla en contra de la síntesis de proteínas en ellos. Lo que no significa que granulocitos y macrófagos no tengan otra función en la formación de anticuerpos, de hecho, no nos cabe duda de que estas células juegan un importante papel en el complicado proceso que culmina con la salida de anticuerpos… Por ello tanto granulocitos como macrófagos tienen su lugar en la formación de anticuerpos”. En 1946, J Carse y E Slater (242) publican en la Revista de Neurología, Neurocirugía y Psiquiatría: “La linfocitosis después de la convulsión eléctrica”. Varios autores han comunicado cambios en el número de células blancas después de inducir artificialmente convulsiones, entre ellos Fattovich en 1937 y 1938, Blomquist en 1938, Lemmi en 1935, Horn en 1939 y Felici en 1940. La investigación descrita en este artículo, fue llevada acabo por J Carse sin el conocimiento de la obra de los autores anteriormente citados, y se inició en el año 1937. Se pretendía registrar los resultados a los pocos minutos de la convulsión, para lo cual en la serie de los 50 pacientes considerados en este 170 RESULTADOS trabajo, los recuentos se realizaron a intervalos de 3 minutos durante 21 minutos y a 30-­‐45 y 60 minutos, ya que los cambios más notables ocurren en el primer cuarto de hora. Se observó que inmediatamente después de la convulsión, durante los primeros 15 minutos, hay fluctuaciones violentas en el número de neutrófilos. Cuando se miden los valores medios de las 50 observaciones, en cada intervalo de tiempo no se encuentra un aumento o disminución consistente, y el análisis de la varianza muestra que las diferencias no son significativas. Ocurre de otro modo con los linfocitos, ya que, aunque también varían considerablemente después de la convulsión, hay una tendencia sistemática a aumentar su número hasta un máximo a los 6 minutos, y después a disminuir de manera gradual hasta llegar a valores normales. El análisis permite no dudar de que esta tendencia es significativa, y no se debe a error experimental o al azar. En vista del interés de este hallazgo, se propone el estudio del número de linfocitos cuando los pacientes se clasifican según sexo y diagnóstico. En los resultados se advierte que el aumento en el número de linfocitos es más marcado en hombres que en mujeres, y más marcado en pacientes esquizofrénicos que con otros trastornos. En su discusión, los autores (242) atribuyen la aparición de esta linfocitosis a un aumento en el flujo de linfa, que en el adulto puede observarse en la radiación actínica, exposición a rayos X, por efecto del calor seco y en grandes altitudes. Un punto que consideran de gran interés, es el hecho de que la linfocitosis es significativamente mayor en esquizofrénicos, considerándolo un hallazgo mas que se suma a la lista de alteraciones funcionales de estos enfermos, como las alteraciones en la regulación de la temperatura, presión arterial, metabolismo de glutatión, reacción a la insulina y al azúcar, y aseguran: “es un puzzle imposible en este momento de construir de forma coherente”. 171 RESULTADOS Son numerosas las comunicaciones realizadas durante esta década en relación a la llamada “Linfocitosis Infecciosa”. La lista de autores que escriben sobre esta enfermedad es interminable: Sita-­‐Lumsden, Barnes (243), Yanneth, Lieberman, Cohn, Novo Segura, Caldwell (244), Smith (245), Coffey, Honaker, Goldberg, Nevius, Klemola (246), Parrish, Larrain, y Del Solar (247), entre otros. Como ejemplo sirva la realizada por H.F MacInnis en 1946 (248). MacInnis resume la historia de la enfermedad desde las primeras referencias a la misma, hasta la fecha del presente artículo. El establecimiento de la linfocitosis infecciosa como entidad se atribuye a Smith (247), quien en Agosto de 1941 informó de dos casos de marcada leucocitosis con cifras de 44.900 y 98.000 leucocitos por ml y linfocitosis del 79% y 86%, correspondientes a dos niños que no presentaron signos ni síntomas. Referencias previas en la literatura incluyen las de Reyersbach y Lenert, quienes en febrero de 1941 informaron de 16 casos de una mononucleosis infecciosa típica que no tenía las manifestaciones de esta enfermedad, y con la única característica hematológica de una linfocitosis del 71-­‐93%. En 1929, Wilson y Cunningham describieron un caso de hiperleucocitosis en un adulto de 20 años sin signos ni síntomas clínicos que es al parecer, la primera referencia en la literatura. En 1943, Duncan comunicó el caso de un niño de 4 años con abdomen agudo, y en el que se hizo el diagnóstico provisional de leucemia linfática aguda. El recuento de glóbulos blancos subió de 45.000 a 110.000 a los tres días, con un 92% de linfocitos. El recuento diferencial y la duración del cuadro hematológico fue idéntico a los descritos previamente, excepto que en este caso había un inicio agudo, simulando una condición quirúrgica. En 1944, Smith (245) informó de cuatro nuevos casos en una familia, llegando a la conclusión de que la enfermedad es infecciosa y contagiosa, con un período de incubación de 12-­‐21 días. Finucare y Phillips en 1944 comunican la aparición de 21 casos en la sala infantil de un sanatorio tuberculoso. El recuento de leucocitos varió de 22.500 a 120.000 por ml, con una linfocitosis relativa del 62-­‐97%, y la duración media de la enfermedad fue de 4 a 7 semanas. Llegan la conclusión de que está 172 RESULTADOS causada por algún agente infeccioso, pero no pueden demostrar el supuesto agente responsable. MacInnis (248) a continuación, hace referencia al diagnóstico y diagnóstico diferencial de esta entidad. Los signos y síntomas son variables, pueden estar ausentes, ser muy leves, o se puede producir una severa reacción constitucional caracterizada por malestar general, fiebre, vómitos, síntomas referidos al sistema nervioso central, y a veces signos y síntomas que simulan un abdomen quirúrgico. El hallazgo más común es la infección de las vías aéreas superiores con linfadenopatías leves o ausentes. En la sangre se observa una leucocitosis usualmente de alto grado con una linfocitosis absoluta y relativa. Los linfocitos son pequeños y maduros y el test de Paul-­‐Bunnell es negativo. El principal diagnóstico diferencial debe hacerse con la reacción leucemoide encontrada en infecciones del tracto respiratorio superior, más frecuentemente por Pertussis, que presentan una linfocitosis y ocasionalmente una hiperleucocitosis. En casos dudosos, la demostración del bacilo de Bordet-­‐ Gengou establece el diagnóstico. El segundo diagnóstico diferencial es con la mononucleosis infecciosa, pero ésta presenta una aglutinación heterófila en el 75% de casos, y el cuadro hemático también ayuda a diferenciar ambas enfermedades. En la mononucleosis infecciosa el total de leucocitos rara vez es superior a 25.000 por ml, y el linfocito es de mayor tamaño que el visto en la linfocitosis infecciosa, de citoplasma abundante que se tiñe intensamente con la tinción de Wright, contiene gránulos y exhibe vacuolización. Una linfocitosis infecciosa aguda con severos síntomas constitucionales puede ser confundida con la etapa temprana de una leucemia linfática aguda, pero a diferencia de la linfocitosis infecciosa la leucemia progresa a anemia con una característica disminución de plaquetas y presencia de linfocitos inmaduros. La linfocitosis infecciosa es una enfermedad que despierta un gran interés en estos años, quizás motivado por su posible confusión con enfermedades como la mononucleosis infecciosa o la leucemia, siendo reiteradamente descrita y 173 RESULTADOS estudiada durante varias décadas. Conocida como enfermedad de Carl Smith, en la actualidad los pocos frecuentes casos registrados se ajustan a la descripción que Smith (247) hizo de ella en 1941. Es un trastorno raro, autolimitado, que se presenta preferentemente en niños menores de 10 años, y se caracteriza por una linfocitosis marcada asociada por lo general con una enfermedad febril inespecífica leve. Su etiología y patogenia continúan siendo desconocidas, su relación con enfermedades víricas ya fue indicada por algunos autores como Finucare y Phillips hace más de medio siglo. En 1947 se publica un trabajo sobre la “linfocitogénesis en los ganglios linfáticos humanos” firmado por Dorothy Sundberg (249). Los conocimientos sobre la importancia de los linfocitos en la producción de anticuerpos y en general en la respuesta inmune en animales, crean la necesidad de realizar nuevos estudios en el campo de la linfocitomorfosis. De los trabajos de Downey (250) y colaboradores se intuye que la linfocitogénesis en ganglios linfáticos humanos no leucémicos, probablemente es muy semejante a la observada en ganglios linfáticos normales de cobayas y conejos. En el trabajo de Sundberg, varios tipos celulares fueron motivo de estudio y son comparados con el mieloblasto de la médula ósea humana, con el linfocito maduro e inmaduro de ganglios linfáticos de conejos y cobayas y con los otros tipos de células linfoides encontradas en los ganglios linfáticos, sangre y médula ósea en casos de mononucleosis infecciosa, leucemia linfática crónica, y leucemia linfática aguda y subaguda. Se examinaron 55 ganglios linfáticos. De ellos 43 fueron de sujetos normales, 2 de pacientes con leucemia crónica, 9 de pacientes con mononucleosis infecciosa y 1 de un paciente con leucemia linfática subaguda. Para el estudio comparativo se utilizaron 11 ganglios de conejos y cobayas. Se realizaron improntas de los ganglios linfáticos y extensiones de sangre y médula ósea que fueron teñidas con May-­‐Grünwald-­‐Giemsa en combinación con la tinción de Wright. Los ganglios linfáticos se fijaron con el líquido de Helly, fueron 174 RESULTADOS cortados a 5 micras y teñidos con hematoxilina-­‐eosina o con tinción de Dominici. Se compararon las características morfológicas más frecuentemente encontradas en las células reticulares hematopoyéticas, en linfocitos reticulares, grandes linfocitos, mieloblastos y linfoblastos. La valoración se hizo según el tamaño celular, tamaño nuclear, forma nuclear, patrón de cromatina, características tintoriales, forma, número y tamaño de los nucleolos, características de la membrana nuclear y presencia de vacuolas. Figura 23. Tipos celulares de ganglios linfáticos. Tipos celulares de impronta de ganglios linfáticos humanos no leucémicos, los cuales muestran grados variables de hiperplasia. Todas las microfotografías están tomadas con la misma magnificación. 1- célula reticular indiferenciada. 2- célula reticular histiocitaria. 3- célula reticular hematopoyética. Linfocitos. Células plasmáticas 4 a 7- células reticulares hematopoyéticas. Tomado de: Sundberg, 1947, (249) 175 RESULTADOS Del resultado de estas observaciones Sundberg (249) señala como hechos de mayor interés los siguientes: 1. Los ganglios linfáticos humanos de sujetos normales, al igual que los de conejos y cobayas normales, no contienen células morfológicamente idénticas al mielobasto de médula ósea. 2. Las células linfoides inmaduras (células reticulares hematopoyéticas y linfocitos reticulares), progenitoras del linfocito, son morfológicamente distinguibles del mieloblasto. 3. Se pone de manifiesto la existencia de un dualismo morfológico y funcional en la hematopoyesis normal del adulto. Las células reticulares hematopoyéticas funcionan como una stem cell para los linfocitos, en cambio, el mielobasto funciona como la stem cell para los elementos mieloides. 4. Este dualismo morfológico y funcional prevalece, excepto en algunos casos de leucemia linfática aguda y subaguda, donde las células llamadas linfoblastos, muchas de ellas morfológicamente indistinguibles de mieloblastos, aparecen en sangre, ganglios linfáticos y médula ósea. Lo más interesante del trabajo de Sundberg (249) es la identificación de las dos principales vías de la hematopoyesis, mieloide y linfoide, usando como únicas herramientas el microscopio óptico y tinciones convencionales como las de May-­‐Grünwald-­‐Giemsa y Wright. Con algunas correcciones (pues no se considera válido el origen dual de las células sanguíneas, y se entiende actualmente que todas las células tienen un origen único y común en la médula ósea a partir de una única célula madre hematopoyética), tal clasificación sigue vigente. La mielopoyesis comprende a eritrocitos, plaquetas, leucocitos granulares y monocitos-­‐macrófagos, cuyo proceso de maduración se inicia y completa en la médula ósea, a diferencia de aquellos elementos cuyo proceso de diferenciación puede tener lugar fuera de médula ósea, la linfopoyesis (251). 176 RESULTADOS O.A Trowell, en un artículo publicado en la revista Nature en 1947 (252), realiza una síntesis de las aportaciones más notables en el estudio de la “función del linfocito”. La función del linfocito en los mamíferos ha sido durante mucho tiempo motivo de debate. Estas palabras del autor dan una idea del conocimiento que hasta entonces se tenía sobre el tema: “… al menos una definitiva función, y quizás la más importante, ha sido atribuida a ellos, la síntesis de γ-­‐globulina. El linfocito sintetiza y libera a la sangre la γ-­‐globulina, y también varios anticuerpos… los anticuerpos conocidos son simplemente modificaciones de γ-­‐globulinas. Esta actividad del tejido linfoide es controlada por una hormona del córtex adrenal”. A continuación hace referencia a la presentación que Ehrich, White y Dougherty hicieron en 1945 sobre el conocimiento del linfocito en la Academia de las Ciencias en Nueva York. Aproximadamente 200 millones de linfocitos/hora entran en el torrente sanguíneo, y desaparecen de la sangre a la misma velocidad, pero la manera cómo ocurre esto sigue siendo un misterio. En el proceso de destrucción de linfocitos, la γ-­‐globulina, supuestamente sintetizada por los linfocitos como un componente citoplasmático normal, es liberada y sale del sistema linfático a la sangre. La γ-­‐globulina, es una de las pocas proteínas plasmáticas, quizás la única, no manufacturada en el hígado. Ehrich (253) supone que “… los antígenos deben ser primero ingeridos y parcialmente digeridos por células reticuloendoteliales, y son estos productos de la digestión los que estimulan al linfocito para la formación del anticuerpo específico”. Por lo tanto, el linfocito elabora el anticuerpo específico en lugar de la normal γ-­‐globulina cuando se desintegra en los centros germinales. Los anticuerpos que aparecen durante el desarrollo de inmunidad a proteínas extrañas, bacterias y toxinas, son modificaciones específicas de la γ-­‐globulina plasmática normal, distinguiéndose únicamente en la configuración de la molécula, en el ordenamiento de los aminoácidos constituyentes Con sus estudios, White y Dougherty (254) demostraron que la hormona del córtex adrenal estimula la destrucción de linfocitos en el centro germinal, 177 RESULTADOS aumenta la liberación de γ-­‐globulinas y anticuerpos en la sangre, y disminuye el volumen de tejido linfoide total del cuerpo: “La cantidad de tejido linfoide es controlada por el córtex adrenal, el cual está bajo control de la pituitaria anterior. La normal involución del tejido linfoide debido a la edad (incluida la del timo), es debida a un aumento de actividad del cortex adrenal por estímulo de la pituitaria anterior”. Considerando otro punto de vista, señalan: “… los linfocitos representan un considerable sistema de almacenamiento de proteínas, las cuales pueden ser rápidamente puestas a disposición en estados de emergencia por el sistema pituitario-­‐adrenal” Observaciones que correlacionan el número de linfocitos circulantes y las situaciones de estrés, estimulan a H C Shands y J E Finesinger a interesarse por el estudio de los linfocitos en las enfermedades mentales. En 1947 (255) publican los resultados de su trabajo, bajo el título “los linfocitos en las psiconeurosis”. Los autores parten del hecho de la presencia de la queja de angustia, característica clínica relevante en las psiconeurosis, y se cuestionan si puede haber diferencias demostrables en el número absoluto de linfocitos entre los pacientes psiconeuróticos y los sujetos normales cuando se someten a diversas condiciones de estrés de la vida diaria. Basan su línea de pensamiento, en la teoría del “síndrome general de adaptación” indicada por Selye (256), la cual asegura que en respuesta al estrés, tienen lugar reacciones específicas e inespecíficas. Las reacciones no específicas constituyen lo que Selye llamó “reacción de alarma”, caracterizada por la respuesta de los tejidos linfoides y linfocitos circulantes, y que en caso de reacción grave, cursa con marcada linfocitopenia junto a cambios degenerativos en timo y órganos linfoides. Algunos autores, como Elmadjian y Pincus (257) han sugerido que se producen variaciones diarias del número de linfocitos en los seres humanos que se pueden correlacionar con el estrés de la vida diaria, que el linfocito es sensible a estas tensiones, y que una situación de estrés específica es cualitativamente diferentes en sujetos normales y psicóticos. 178 RESULTADOS Shands y Finesinger (255) con la esperanza de encontrar correlaciones fisiológicas en síntomas como la fatiga crónica, someten a un grupo de sus pacientes a situaciones de estrés, y para establecer un fondo de referencia, también realizan estudios sobre la variación diurna del recuento de linfocitos. Dicen establecer criterios arbitrarios sobre la base de los síntomas para diferenciar los sujetos normales de los psicóticos: “puede ser cierto que entre nuestros sujetos normales, pueda haber algunos psiconeuróticos leves, y quizás algún psicópata leve, pero respecto a los pacientes, estamos seguros que todos se clasifican como psiconeuróticos”. En el grupo de pacientes se inclinan a incluir principalmente pacientes con quejas de fatiga, y la gran mayoría corresponden a pacientes diagnosticados de histeria, neurosis de ansiedad, o bien síntomas mixtos de histeria y ansiedad, un paciente con depresión reactiva y otro con trastorno obsesivo compulsivo. El grupo considerado normal se compone de secretarias, estudiantes de medicina, médicos y marineros de los cuarteles locales de EE.UU, que mantienen un trabajo estable. Entre los métodos inductores de estrés que los autores emplean en el estudio, la más utilizada es la entrevista con y sin estrés. En las observaciones sobre la variación diurna, impresiona la gran diferencia existente de día a día y de hora a hora. Se pudo apreciar una pequeña, pero definida tendencia al aumento de los recuentos desde las 8,30 am hasta las 9,00 pm. No parece haber ninguna diferencia apreciable entre los pacientes con neurosis de ansiedad y los controles. Resultó interesante que el grupo de mujeres con histeria, tanto con ansiedad como sin ella, muestran niveles más bajos que los otros pacientes y controles, lo que podría indicar que la histeria es una condición cualitativamente diferente respecto a los mecanismos fisiológicos de la neurosis de ansiedad; sin embargo, el tratamiento estadístico de los datos relativos a las variaciones diurnas no resultan significativos. Cuando se analizan los resultados derivados de las situaciones de estrés, observamos una caída en los linfocitos en las situaciones de estrés comparada con las situaciones sin estrés, sin embargo la respuesta media a este tipo de 179 RESULTADOS estrés varía según la patología, ya que en los psicóticos aparece una linfocitosis relativa, mientras que en los psiconeuróticos se aprecia una linfopenia relativa. La histeria femenina fue el diagnóstico habitual de un amplio abanico de síntomas que incluían desfallecimientos, insomnio, pesadez abdominal, espasmos musculares, respiración entrecortada, dolor de cabeza, irritabilidad, pérdida de apetito y tendencia a causar problemas. Se llegaron a enumerar más de setenta y cinco síntomas propios de la histeria. Por tanto debemos preguntarnos ¿qué es la histeria? (258). Fue descrita tanto por Platón como por Hipócrates, pero no cabe duda de que con tantos síntomas posibles, la histeria era un estado donde cabía cualquier enfermedad no bien identificada. Parece ser que fueron los trabajos de Freud y Charcot, quienes al profundizar en el estudio de la mente, propician la desaparición de la histeria como enfermedad, a medida que las técnicas diagnósticas mejoran, el número de casos disminuye hasta desaparecer (258). Nos encontramos en 1947, ya se conocen los trabajos de Freud, y resulta curioso que los autores, Shands y Finesinger todavía consideren la histeria como una enfermedad con entidad propia, incluyendo en su estudio una gran mayoría de pacientes diagnosticados de histeria. Por otro lado, dentro de las teorías existentes sobre la etiología de las enfermedades mentales, la historia está llena de modelos que van siendo descartados conforme se logran avances que permiten un conocimiento más preciso del fenómeno. En este momento histórico, las teorías vigentes son la psicoanalista de Freud, el conductismo, la teoría biológica y la teoría que integra factores biológicos, psicológicos y sociales y que es la más aceptada en la actualidad. El trabajo de Shands y Finesinger (255) podemos considerarlo dentro de la línea de pensamiento biológico, y supone una muestra de la tendencia del pensamiento general ya existente, y que pocos años después será un hecho, ya que el término psiquiatría biológica se empezó a utilizar en 1953, y es a finales de los años 50 cuando se desarrollan las primeras drogas antipsicóticas y antidepresivas como la clorpromazina y la imipramina (259). 180 RESULTADOS También los estudios de Freeman y Elmajian (260), en 1947 abordan el tema de las enfermedades mentales: “La relación entre los niveles de azúcar en sangre y linfocitos en sujetos normales y psicóticos”. El trabajo se llevó a cabo en 20 sujetos normales y 35 psicóticos, principalmente esquizofrénicos, en los que se determinaron los niveles de glucosa en sangre y el número de linfocitos tras la administración de dos dosis de 50 gramos de glucosa a un intervalo de 30 minutos. Los autores observaron que: • En sujetos normales, los niveles en sangre de linfocitos y de glucosa varían inversamente, siendo la correlación de −0,85. • En los pacientes la correlación era mucho más pobre, con un coeficiente de −0,29, y un 43% de pacientes mostraron una relación anormal de carácter positivo. • El grupo de pacientes con la relación negativa (normal) linfocitos-­‐ glucosa, tendía a incluir sobre todo, aquellos con una sintomatología mal definida, mientras que el grupo con un resultado positivo (anormal) se correspondía más claramente con los subtipos clásicos de esquizofrenia. El grupo con la relación negativa (normal) linfocito-­‐ glucosa estaba integrado por pacientes que habían sufrido un periodo de hospitalización reciente, de 9 meses de media. En cambio los que mostraban una relación positiva (anormal), habían sido hospitalizados por un tiempo más prolongado, de 5 años de media. Según los autores, estas observaciones indicarían que la menor respuesta linfopénica y la relación positiva anormal en pacientes psicóticos, es debida a un fallo en el control del sistema adrenocortical. Un punto de interés en este estudio, es la comparación de los dos grupos de pacientes en función de la edad y duración de la hospitalización. En el grupo de respuesta normal, la edad media fue de 30 años, y en el grupo de respuesta anormal, la media fue de 38 años. ¿Puede estar asociada la diferente respuesta fisiológica a algún proceso relacionado con el envejecimiento? La diferencia de 181 RESULTADOS edad no es notoria, en cambio, los dos grupos muestran una gran diferencia respecto al grado de gravedad de la enfermedad, y sobre todo con la duración de la hospitalización. Un largo período de hospitalización indica una no recuperación y una tendencia hacia el deterioro, además una retirada prolongada del estrés y tensiones de la vida cotidiana tienen su efecto sobre los mecanismos endocrinos del organismo. Después de la exposición de este razonamiento, los autores (260) reconocen la incapacidad para resolver con garantías el dilema: “… es imposible actualmente determinar si la respuesta anormal es debida a factores inherentes a la psicosis, a la hospitalización o a ambos”, y consideran que sus resultados están de acuerdo con la hipótesis de que en las psicosis un alto porcentaje de sujetos sufren una insuficiencia suprarrenal “de un tipo particular”, existiendo en ellos una alteración fisiológica de origen no explicado, siendo la anormal relación linfocito-­‐glucosa una de sus manifestaciones. Una breve comunicación de 1948 sobre la “reacción linfocitopénica y la administración de glucosa en ratas timectomizadas”, tiene por autores a Sass-­‐ Kortsák y Vogt (261). A final de los años cuarenta ya se conoce que la administración de glucosa causa una disminución de linfocitos en sangre. Los trabajos de Wiseman y Akeroyd (262) en 1938, mostraron que las dietas ricas en carbohidratos producen una involución del tejido linfático y una reacción linfopénica en sangre. Unos años más tarde, Freeman y Elmadjian (263), mostraron que la administración de glucosa causa linfocitopenia y que ello no ocurre en animales adrenalectomizados. Se suponía que la hormona 11-­‐ oxicorticosterona producida en el córtex adrenal, era la causante de esta acción linfocitopénica, según los estudios llevados a cabo por Dougherty y White (254). También es conocido que después de la adrenalectomía se produce una hipertrofia del timo, mientras que después de la administración de ciertos corticosteroides suprarrenales el timo se atrofia. Parecía probable que el timo jugase algún papel en esta respuesta linfopénica, ya que este órgano parece tener una función secretora interna. Basados en estas premisas, Sass y Vogt, 182 RESULTADOS deciden estudiar la reacción linfopénica tras sobrecarga de glucosa en ratas timectomizadas. Para este estudio (261) siguieron el mismo método que utilizaron Elmajian y Freeman. Se realizan recuentos de control de linfocitos, sin administración de glucosa, durante varias generaciones, en ratas normales y timectomizadas de la misma edad y peso, observándose diferencias muy leves en el número de linfocitos. Posteriormente, los animales se mantienen en ayunas 24 horas, se administra glucosa, y se realizan nuevos recuentos. El número de linfocitos, tanto en animales timectomizados como no timectomizados, disminuye aproximadamente el 70% de su valor original a los 30 minutos, y es del 64-­‐ 56% a las 2-­‐3 horas, no existiendo diferencias entre animales normales y sin timo. Por tanto, Sass-­‐Kortsák y Vogt (261) demuestran que el timo no interviene en la disminución de los linfocitos que tiene lugar después de la administración de glucosa, y que la ausencia de reacción linfopénica después de la adrenalectomía, cuando el timo es hipertrófico, no puede ser relacionada con una función del timo, ya que la ausencia total del timo no influye en ello. Procedente del Laboratorio de Patología del Instituto Nacional de Salud, Servicio de Salud Pública de Maryland, es el siguiente trabajo que tiene por autores a Habel, Endikott, Bell y Spear (264), quienes en 1948 publican “la evidencia inmunológica del papel de los linfocitos en la formación de anticuerpos”. Los trabajos de Ehrich y Harris (229)(253) sobre formación de anticuerpos han provocado un gran interés en la búsqueda del lugar de formación de los anticuerpos, y parece que vale la pena tratar de confirmar los resultados experimentales en los que se han basado las interpretaciones inmunológicas. En apoyo a la teoría de la formación de anticuerpos por parte de los linfocitos, tenemos la evidencia de la aparición de anticuerpos en los ganglios linfáticos antes que en el suero, los mayores títulos de anticuerpos específicos en linfa y 183 RESULTADOS linfocitos del ganglio inoculado con antígeno y el alto título de anticuerpos en los linfocitos cuando se compara con el suero. Con este estudio, los autores tratan de confirmar los resultados de Ehrich y Harris, y validar algunas de las técnicas empleadas. Se recogen todos los datos obtenidos de 37 conejos inoculados con antígenos bacterianos (Samonella typhinurium y Pasteurella tularensis) desde el día 2 al 13 después de la administración del antígeno. La respuesta general es de un aumento de las células en la linfa de la zona inoculada, comparado con el ganglio no inoculado, que comienza en el primer día y continúa hasta el día 13. Los títulos de anticuerpos aparecen en el tercer día y aumentan hasta el sexto día, manteniéndose a niveles relativamente altos durante el periodo estudiado. Las observaciones histológicas de los tejidos inoculados mostraron, además de la reacción local, consistente en un absceso estéril, una linfadenitis regional seguida de una rápida y marcada hiperplasia folicular. En los ganglios, a los tres días aparece una proliferación folicular, que alcanza su máximo a los nueve días para disminuir a partir de entonces, alcanzando un aspecto de normalidad a los trece días. Una vez valorados los resultados, Habel, Endikott, Bell y Spear (264) aseguran que éstos no confirman el hallazgo de que los anticuerpos aglutinantes aparecen antes en la linfa que en suero, y que los altos títulos de anticuerpos en los extractos de ganglios linfáticos deben ser cuestionados, asimismo discuten la validez del método: “Cualquier método en que las células no se lavan, y los títulos se expresan en términos de volumen de células no parece válido, debido al arrastre en las paredes del tubo de anticuerpos a partir de pequeñas cantidades de linfa, pero altamente significativas”. Señalan que el alto contenido de anticuerpos en la linfa, puede explicarse, al menos en parte, por transferencia de los mismos desde la sangre, además, existen pocas evidencias significativas en los experimentos, para asegurar que los cuerpos de linfocitos o células de los ganglios linfáticos contengan grandes cantidades de anticuerpos, y que a pesar de haber utilizado técnicas inmunológicas más críticas y mejor controladas, las pruebas de la 184 RESULTADOS producción de anticuerpos por parte de los linfocitos deben considerarse incompletas Nos encontramos por tanto en un punto histórico algo confuso en cuanto a la fiabilidad y validez de los experimentos realizados. Habrá que esperar todavía algunos años más para que el lugar de producción de los anticuerpos pueda desvelarse, y en este avance va a ser fundamental e imprescindible contar con las técnicas resultantes de los avances tecnológicos que aseguren los experimentos. En 1949, se publicó un nuevo trabajo sobre producción de anticuerpos realizado por T.N Harris y Susana Harris. Se pretende investigar los “cambios histoquímicos en linfocitos durante la producción de anticuerpos en los ganglios linfáticos del conejo” (265). Las recientes aplicaciones de la histoquímica a la fisiología ofrecieron un nuevo enfoque para abordar el problema de la identificación de las células relacionadas con la producción de anticuerpos. Caspersson y Brachet (266)(267), demostraron por vez primera que las células que forman activamente proteínas, se caracterizan por grandes cantidades de ARN en su citoplasma. La aplicación de este descubrimiento, que los autores adaptan al presente estudio, se hace según el siguiente razonamiento: “Existe considerable evidencia de la producción de anticuerpos en el ganglio linfático poplíteo del conejo después de la inyección de material antigénico en el tejido drenado por ese nodo. Parece factible identificar las células que participan en esa síntesis proteica por medios histoquímicos, mediante la determinación de ARN y ADN, y su correlación con las concentraciones de anticuerpos en el ganglio linfático”. Harris y Harris inyectaron en las patas traseras de conejos varios tipos de sustancias, antigénicas y no antigénicas, como bacilos de disentería, eritrocitos de oveja, virus de la gripe, aceite de lanolina, minerales y gelatina (estas tres últimas sustancias son no antigénicas). Posteriormente se procedió a la recogida de muestras para su estudio histológico, químico y serológico, 185 RESULTADOS sacrificando los animales a diferentes intervalos (0-­‐2-­‐3-­‐4-­‐5-­‐7 y 9 días) después de la inyección de la sustancia elegida. Las observaciones (265) histológicas mostraron que, tras la inoculación de sustancias antigénicas, los linfáticos extirpados eran progresivamente más grandes, y aumentaron en tamaño casi exclusivamente debido al aumento del córtex. A los dos días, se observó un cuadro de intensa hiperplasia difusa, constituida por linfocitos pequeños intercalados con células reticulares. En el cuarto día comienza en la corteza de los ganglios linfáticos la formación de folículos. En el sexto día, la estructura folicular del nodo se estableció claramente. Este estudio histológico hizo posible identificar las células que contienen los gránulos de ARN, pero solo permitió una valoración subjetiva de las cantidades relativas de ARN en las diversas muestras de ganglios linfáticos. En el segundo día después de la inyección del antígeno, comienzan a aparecer nucleolos, gránulos citoplasmáticos y medias lunas teñidas con pironina, que se encuentran primero en células reticulares poco alteradas, más tarde en formas de transición, linfocitos jóvenes y posteriormente en los linfocitos más maduros. La identidad de este material teñido con pironina es ARN, como se demostró mediante digestión específica con Ribonucleasa libre de proteasas Se encontró asimismo que las concentraciones de ADN en el ganglio linfático no variaban significativamente entre las muestras analizadas. La concentración de ADN es una constante en todos los ganglios linfáticos dentro los límites de variación experimental. En cambio, la concentración de ARN mostró una considerable variación, con aumento de hasta más del doble de los valores normales, alcanzando su máximo en el día 4º-­‐5º, y seguido de su disminución en días posteriores. Se estudió la correlación de las concentraciones de ARN en relación con la concentración de anticuerpos para el antígeno inyectado. Los resultados muestran como el contenido de ARN del nodo se eleva un poco antes del título de anticuerpos, y alcanza su valor máximo aproximadamente al mismo tiempo 186 RESULTADOS que lo hacen éstos. A partir de entonces, la concentración de ARN disminuye más rápidamente que la de anticuerpos. Con materiales no antigénicos no se observa cambios en el ARN, a pesar de la severa inflamación local encontrada en el lugar de la administración de aceite de lanolina y gelatina, no se encontró un aumento del ARN en el nodo de drenaje. Los materiales no antigénicos, cuando se inyectan, no dan lugar a un aumento en el contenido de ARN en el nodo linfático. Figura 24. ARN y título de anticuerpos. Concentración de ARN y título de anticuerpos en ganglios linfáticos que drenan el sitio de inoculación de diversos antígenos. Cada punto representa la media de las muestras obtenidas de dos o tres conejos. Tomado de: Harris y Harris, 1949, (265). Con este trabajo de Harris y Harris se da un paso más en el conocimiento de los antígenos, anticuerpos y del fenómeno antígeno-­‐anticuerpo. Las sustancias antigénicas y no antigénicas quedan perfectamente definidas. Concede una gran importancia a la asociación entre ARN y la síntesis de proteína, imprescindible para futuros avances en este terreno. En la discusión realizada 187 RESULTADOS por los autores, hay que destacar algunos puntos de interés, como la comparación establecida entre sus propias experiencias y el trabajo de otros autores: “… que las estructuras manchadas con pironina encontradas en las formas de transición de células reticulares y linfocitos jóvenes son bastante similares a las descritas por Caspersson y Brachet” (267)(266) y la seguridad con la que afirman que el material teñido con pironina es ARN: “…queda bien establecido, ya que la reacción de tinción fue suprimida por el tratamiento con ribonucleasa libre de proteasas”. De especial importancia consideramos las observaciones histológicas que se advierten dos días después de la inoculación del antígeno, cuando aparece la tinción por pironina en nucleolos y estructuras citoplasmáticas en células dispersas a lo largo de la corteza ya aumentada de tamaño, que posteriormente se observa en células grandes, linfocitos jóvenes, y por último en linfocitos maduros. Se pone de manifiesto una continuidad desde la célula reticular al linfocito maduro, hallazgo consistente con las observaciones descritas por Sundberg (249), quien describió la linfogénesis en los ganglios linfáticos maduros. En base a estos hallazgos Harris y Harris proponen la siguiente sugerencia: “la estimulación antigénica puede ofrecer un método para el estudio del desarrollo celular en el ganglio, ya que constituye un medio para inducir linfocitopoyesis en los ganglios linfáticos, que puede ser controlado en tiempo e intensidad”. Ehrich, Drabkin y Forman, se dedican al estudio de la “relación entre ácidos nucleicos y la producción de anticuerpos en las células plasmáticas”, publicando resultados en 1949 (268). Es conocido ya por anteriores trabajos como los de Harris, Grimm, Mertens, Dougherty y White, que el linfocito está relacionado con la producción de anticuerpos (241)(229)(254). Otros autores como Bing y Plum (269), habían señalado que pacientes con hiperglobulinemia tenían un aumento de células plasmáticas en sus tejidos, y que los niveles más altos de globulinas eran encontrados en pacientes con mieloma de células plasmáticas, mientras que pacientes con leucemia linfática no mostraban aumento de globulinas. Fagraeus (270) apuntó la posibilidad de que fueran las células plasmáticas más 188 RESULTADOS que los linfocitos las responsables de la producción de anticuerpos. Estudios presentados en 1947, en las Jornadas de la Sociedad de Biología Experimental, en Cambridge, dieron lugar a la tesis de que la síntesis de proteínas está relacionada con los ácidos nucleicos. Gracias a métodos especiales de espectrografía y citología se han obtenido pruebas de la relación existente entre la multiplicación de cromosomas y la formación de ácido desoxirribonucleico (ADN), mientras que la producción de proteínas plasmáticas está relacionada con la de ácido ribonucleico (ARN), como quedó demostrado por los estudios de Caspersson (271)(267). Estos antecedentes llevan a Ehrich, Drabkin y Forman (268) a realizar un estudio comparativo de la formación de anticuerpos y los cambios en los ácidos nucleicos que ocurren en la linfa de los ganglios linfáticos. Se inyectó vacuna de la fiebre tifoidea a 41 conejos experimentales y a otros 9 conejos que sirvieron como controles, siendo sacrificados a distintos intervalos de tiempo, después de la administración del antígeno. Cuando se estudió el contenido de ADN y ARN de los ganglios linfáticos, se encontró que el incremento en el contenido de ADN va en paralelo al aumento de peso de estos ganglios, relacionado con la multiplicación activa de sus células, y que el pico de aumento de ARN, ocurrió entre el día 4º y 6º después de la administración del antígeno, cuando la formación de anticuerpos estaba en su máximo nivel. El estudio histológico realizado con secciones de ganglios teñidas con verde de metilo y pironina, colorantes utilizados para detectar ADN y ARN respectivamente, revelaron que durante los primeros seis días del experimento, la reacción celular fue principalmente a base de células plasmáticas, predominando el plasmoblasto en los tres primeros días, y que la mayor parte de ARN se encontraba contenido en las células plasmáticas. Los linfocitos comienzan a proliferar en suficiente número en los días 3º y 4º, y los centros germinales comienzan a aparecer en los días 4º-­‐5º, mostrando su máxima actividad en el 9º día, cuando la formación de anticuerpos y el ARN ya han tenido sus valores máximos. 189 RESULTADOS Estos resultados son interpretados por Ehrich, Drabkin y Forman como indicativos de que es la célula plasmática y no el linfocito, la responsable de la formación de anticuerpos (268). En su discusión los autores hacen referencia repetidamente a Fagraeus (270)(272), que ya en 1948 publicó un trabajo donde aseguraba que eran las células plasmáticas las productoras de anticuerpos. Bing, Fagraeus y Thorell (273), demostraron que la basofilia del citoplasma de las células plasmáticas se debía al RNA, que eran células muy diferentes de los histiocitos, y que en su maduración, la concentración de RNA en el citoplasma fue totalmente retenida en él, a diferencia de las otras células de la sangre. Los autores (268) dan algunas explicaciones a sus resultados diciendo: “nuestro hallazgo anterior de los anticuerpos en las células linfoides de la linfa eferente, ha sido interpretado como que los linfocitos pueden formar igualmente anticuerpos. En contra de esta interpretación está el hecho de que las leucemias linfáticas, a diferencia del mieloma, no se asocian con hiperglobulinemia. En el presente estudio, las secciones de los linfáticos eferentes teñidas con verde metilo y pironina, sugieren que algunas de las células linfoides que dejan los ganglios linfáticos durante el período de formación de anticuerpos son plasmoblastos y células plasmáticas”. Con lo que reconocen o intentan acercarse a los resultados anteriores de Fagraeus y de sus propias observaciones actuales. Fagraeus (272) defendía la idea de que las células plasmáticas eran derivadas de células reticuloendoteliales. Sus resultados con cultivos de tejidos le llevaron a creer sobre formación de anticuerpos que: “… se lleva a cabo al lado de y durante el desarrollo de células reticulares en células plasmáticas. Con la aparición de las células plasmáticas maduras el título vuelve a disminuir”. Ella concluye que los anticuerpos se formaron dentro de las células reticulares, y que en casos de intensa formación de anticuerpos, estas células se diferencian a células plasmáticas, considerando a la célula plasmática madura como el último eslabón de una cadena de acontecimientos: “… una célula que ya ha pasado la etapa de su mayor actividad funcional”. Es probable que la síntesis se 190 RESULTADOS lleve a cabo durante la etapa de maduración, y cese cuando se alcanza la madurez. Sin embargo los autores en sus experimentos, parecen demostrar que la concentración máxima de anticuerpo en las células plasmáticas se alcanza sólo cuando estas células están totalmente maduras. Es posible que en los experimentos de Fagraeus, debido a las manipulaciones a las que se somete el tejido, se dañen las células maduras más que los elementos inmaduros, por eso dice: “… las células plasmáticas maduras en el momento de la explantación producen menos anticuerpos que los cultivos con elementos inmaduros”. Durante el curso de una epidemia por una enfermedad infecciosa respiratoria conocida como infección por “virus X”, Small y Hadley (274) deciden estudiar las células linfoides: “Leucocitos en la infección por virus X”. Observaron células linfoides anormales en los frotis de sangre de pacientes con esta enfermedad, lo que les llevó al estudio sistemático del cuadro sanguíneo de estos enfermos, la mayoría estudiantes de medicina, enfermeras y empleados del hospital. Registraron una serie de 50 casos de este síndrome gripal en el Hospital y Sanatorio de Loma Linda, California, entre Noviembre de 1947 y enero de 1948. El estudio de la sangre de estos pacientes con “virus X”, reconocida como una gripe debida a virus influenza tipo A, mostró una leve leucopenia y la presencia de linfocitos anormales, con citoplasma basófilo. Los tipos celulares que en el estudio del frotis despertaron mayor interés fueron: ! Células tipo I, de hasta 18 μ de diámetro, con citoplasma azul oscuro granular. Núcleo grande que ocupa 2/3 del diámetro celular, de color púrpura oscuro y con cromatina de la textura habitual encontrada en el linfocito adulto. Células similares se han visto en pacientes con rubéola, mononucleosis infecciosa y sarampión. Algunas de ellas eran parecidas a las células plasmáticas, pero el núcleo no era por lo general excéntrico. ! Células tipo II, de igual tamaño que las anteriormente descritas, pero con citoplasma claro, transparente y azul, a menudo mucho más oscuro en la periferia celular, que parecían tener muy baja viscosidad citoplasmática, ya que corrían entre los eritrocitos y se adaptaban fácilmente al contorno irregular de objetos adyacentes. 191 RESULTADOS ! Células tipo III con alguna de las características nucleares descritas, pero con citoplasma vacuolado o fenestrado, de aspecto nebuloso o turbio con pequeños gránulos rojizos como los de los monocitos. Figura 25. Tipos celulares en la infección por virus X. Izquierda: figura 2. Derecha: figura 3 La figura 2 muestra una célula tipo II, con tendencia a plegarse alrededor de los eritrocitos adyacentes. En la figura 3 se muestra una célula tipo III, con vacuolización citoplasmática y apariencia espumosa. Tomado de: Small y Hadley, 1949, (274). Small y Hadley (274) aplicaron el término de “linfocito tóxico” a las células tipo I descritas: “El término indica una célula que aparece bajo una influencia tóxica o infecciosa, pero no es un linfoblasto, no es una célula necesariamente inmadura. El nombre célula de Turk se aplica a menudo a tales células. Aunque no se pueda decir que estas células aberrantes sean diagnósticas de gripe o cualquier otro síndrome clínico, sí indican algún proceso tóxico, normalmente vírico, o al menos una infección no bacteriana”. Considera que estos tipos celulares forman parte del espectro continuo de variaciones de las células linfoides, y no son tipos celulares diferentes. La existencia de variantes morfológicas del linfocito son vistas con relativa frecuencia en los estudios de sangre periférica. Se trata de células mononucleares de origen linfocítico, y que no son ni normales, ni leucémicas, 192 RESULTADOS tal y como lo refieren los autores Small y Hadley, y que a falta de un término mejor, se denominan linfocitos atípicos. La descripción que hizo Türk en 1907 de una célula atípica con núcleo relativamente inmaduro y citoplasma basófilo semejante al de la célula plasmática, adquirió relevancia en la bibliografía médica con el nombre de célula de Türk, pero hasta la fecha no existe un acuerdo general respecto a lo que es una célula de Türk. Así encontramos en la literatura médica, dos posibilidades para las células de Türk: una como sinónima del plasmocito, presente en sangre sólo en estados patológicos, y otra que se aplica al gran mononuclear ortobasófilo, leucoblasto o mieloblasto. Estamos sin embargo, bastante seguros de que las células que descubrió este autor, son linfocitos reactivos que pueden englobarse en el grupo de los virocitos (275). Los linfocitos atípicos, para los cuales Litwins y Leibowitz en 1951 (276) proponen la denominación de “virocitos”, son generalmente indistinguibles de los encontrados en la mononucleosis infecciosa, y también se registran en otras enfermedades infecciosas como pueden ser, parotiditis, sarampión, rubéola y herpes zoster. Los españoles Diaz-­‐Rubio y Garrachón Aguado, de la Facultad de Medicina de Cádiz, se centran en el estudio de la influencia que ejerce la sobrecarga de glucosa en los linfocitos, publicando dos de sus trabajos en 1949. En el primero de ellos (277), “Influencia de la sobrecarga de glucosa sobre los linfocitos en los enfermos de Addison, otras enfermedades endocrinas y en magullados”, consideran de especial interés el estudio de la linfocitopenia tras la sobrecarga de glucosa, sobre todo en los pacientes afectos de enfermedad de Addison, ya que aunque no pueden considerarse equiparables fisiológicamente a los animales adrenalectomizados, sí existen en ambos una base común que se manifiesta en múltiples aspectos como la linfocitosis y la hiperplasia de estructuras linfáticas, características de tales estados y relacionadas con la actividad cortical suprarrenal. Recogen los datos de cuatro enfermos de Addison, a los que realizan recuentos de linfocitos tras sobrecarga de glucosa, en distintos intervalos de tiempo. El comportamiento de la curva de linfocitos 193 RESULTADOS resultó dibujó un perfil plano con una discreta elevación a la 1 ½ hora del inicio de la prueba. Otras enfermedades endocrinas consideradas de interés por Diaz-­‐Rubio y Garrachón Aguado, son las tiroideas y sobre todo, las afecciones del sistema diencéfalo-­‐hipófisis-­‐corteza suprarrenal. Recogen los datos del estudio de dos enfermos hipotiroideos, un hipertiroideo por enfermedad de Basedow, dos enfermos de acromegalia, tres estados de hipofunción hipofisaria global y ocho casos de síndrome diencefálico. A diferencia de los enfermos addisonianos, en los estados tiroideos no se apreció ninguna alteración, tanto en los pacientes hipotiroideos como en el bassedowiano, la curva siguió el trayecto visto en los sujetos normales. En los casos tanto de hiperfunción como de hipofunción hipofisaria, el comportamiento de la curva linfocitaria fue francamente anormal. En los enfermos con afectación diencefálica, la conducta de la curva fue dispar, una respuesta normal y leve en unos, y plana o retardada en otros, independientemente cual fuese la conducta de la glicemia. En cuatro enfermos magullados por accidente traumático, con una intensa repercusión adrenal propia de este estado, se determinó también la curva linfocitaria y, a excepción de un caso, el resto mostró un aumento del número de linfocitos. Diaz-­‐Rubio y Garrachón Aguado (277), realizan en su exposición una serie de comentarios, bastante lúcidos y acertados. Hay que tener en cuenta que partiendo únicamente de la valoración de la curva linfocitaria, pretenden explorar el funcionamiento endocrino de algo tan sumamente complejo y desconocido hasta entonces como las hormonas, y no solamente las hormonas, sino los sistemas de estimulación y autorregulación que parten del hipotálamo, y todo ello utilizando métodos muy básicos y simples. Sirvan algunos de sus comentarios, donde se advierten parte de los razonamientos utilizados en la explicación de los resultados: “… aquí el estímulo que supone la glucosa, no va seguido de respuesta por parte de la corteza adrenal… La situación armoniza con los resultados obtenidos a título comparativo con el animal adrenalectomizado, e 194 RESULTADOS incluso permitiría proponer la prueba de la linfocitopenia provocada por la glucosa como método para conocer la capacidad de función de la corteza. Por lo demás no debe extrañar tales pequeñas diferencias en las curvas del enfermo de Addison y la rata adrenalectomizada, si se tiene presente la enorme diferencia entre ambos…otros elementos y factores son a considerar”. De gran interés nos parecen los comentarios explicativos cuando se refieren a las enfermedades hipofisarias y diencefálicas: “Sin embargo, y para explicar tal resultado en los enfermos hipopituitarios, baste con la consideración, no sólo de la considerable atrofia que la corteza sufre tras la hipofisectomía, sino de la situación de hipofunción adrenal en tales enfermos, si bien en ninguno de ambos casos la muerte se produzca por insuficiencia adrenal exclusivamente…”. En vista de los datos experimentales, Diaz-­‐Rubio y Garrachón Aguado (277) aseveran que, la acción linfopénica y regresiva linfática de la glucosa se ejerce en forma inmediata a través de la corteza adrenal, sin embargo, no se puede eliminar una vía indirecta a través de la hipófisis y del sistema nervioso, aunque la corteza adrenal ocuparía un papel central. Bajo el título “La linfocitopenia tras la administración de glucosa en diversos estados infecciosos”, Diaz-­‐Rubio y Garrachón Aguado publican un segundo trabajo (278). Las enfermedades infecciosas, no sólo aumentan las exigencias de la función suprarrenal, sino que en ellas tienen lugar situaciones de déficit adrenal, expresadas en ocasiones de manera patente y grave por estados de asuprarrenalismo agudo como el síndrome de Waterhouse-­‐Friederichsen. Por tanto, un estudio de la curva de linfocitos tras la administración de glucosa, en infecciones agudas y crónicas, podría poner de manifiesto una alteración en el mecanismo de regulación de los linfocitos, en el cual las suprarrenales podrían estar implicadas. Los enfermos objeto de estudio fueron explorados como cualquier otro paciente, realizándoseles recuentos de linfocitos, a determinados intervalos de tiempo tras la sobrecarga de glucosa. En los resultados, se observó una conducta dispar en las infecciones pulmonares agudas y sepsis por un lado, y 195 RESULTADOS las infecciones crónicas y focales por otro. Las infecciones agudas y sepsis presentan una curva patológica evidente, con un discreto descenso inicial, que es restituido una hora después e incluso paradójicamente sobrepasado en sus niveles normales. En el grupo de las infecciones crónicas y focales, se observa una conducta normal, con una disminución acusada al inicio seguida de recuperación. La explicación, a nuestro entender más acertada de todas las posibilidades expuestas por Diaz-­‐Rubio y Garrachón Aguado, es que la repuesta patológica observada en la sepsis y las situaciones agudas, expresan un trastorno en la regulación de la cifra de linfocitos condicionada por la insuficiencia cortical existente, responsable de la falta del efecto linfocitopénico de la glucosa. Los autores acertadamente reconocen la influencia de otros factores y la amplitud del problema: “… el sistema es más amplio, y estímulos vegetativos y a partir de otras glándulas endocrinas deben aquí ser considerados. No obstante, la experimentación y la clínica dan a la corteza adrenal, y fundamentalmente a la actividad secretora de 11-­‐corticosteroides, un papel central en este problema”. En 1950, otro estudio que pone en relación sistema endocrino y linfocitos es publicado por Colfer, Groot y Harris (279), del Laboratorio de Fisiología de la Universidad de Cambridge: “Glándula pituitaria y linfocitos sanguíneos” Dougherty y White (254) demostraron que la inyección de hormona adrenocorticotropa (ACTH) en ratones, ratas y conejos, provoca una caída temporal en sangre del número absoluto de linfocitos. Respuesta que parece ser mediada por la corteza suprarrenal. La administración de extractos de corteza suprarrenal produce una linfopenia similar, mientras que la inyección de ACTH en animales adrenalectomizados no tiene este efecto. Experimentos anteriores realizados por Harris (224)(229), en los que fue necesario inmovilizar a los conejos mediante abrazaderas por períodos de tiempo de hasta 2 horas, tuvieron que ser suspendidos al comprobarse que el estrés emocional originado en los animales inmovilizados, era suficiente para provocar linfopenia. 196 RESULTADOS Los autores de este trabajo pretenden explorar la función de la glándula pituitaria en la reacción al estrés emocional, los efectos de la estimulación eléctrica del hipotálamo y la hipófisis por una técnica que no requiera la inmovilización del animal. Se emplearon conejos a los que se les provoca estrés emocional mediante la administración repetida de corrientes farádicas. El temor a estas corrientes eléctricas provocaba en los animales movimientos de lucha, taquicardia y polipnea, claros signos de estrés. Parte de los animales fueron hipofisectomizados o bien sometidos a denervación de las suprarrenales. Se decidió también investigar el papel desempeñado por la médula suprarrenal que excitada por la emoción, es posible que juegue algún papel en la respuesta linfopénica. Colfer, Groot y Harris (279), observaron lo siguiente: • El estrés emocional en conejos normales, dio lugar a una marcada linfopenia, que fue máxima 3 horas después del estímulo. • En los animales hipofisectomizados, el estrés emocional no provocó linfopenia, en cambio, la administración intravenosa de extractos de pituitaria dio lugar a una linfopenia después de la hipofisectomía. • La denervación de las glándulas suprarrenales no alteró la respuesta linfopénica al estrés, y la administración intravenosa de adrenalina no provocó linfopenia en la mayoría de los animales, por lo que se deduce la poca intervención de la médula suprarrenal en la reacción linfopénica. Los autores asumen que la linfopenia al estrés es debida a la actuación de la glándula pituitaria anterior: “La respuesta linfopénica puede ser considerada como el indicador más rápido de la actividad de la hipófisis anterior, conocido en la actualidad”. La investigación realizada por Groot y Harris (280) y publicada en 1950 sobre “el control hipotalámico de la hipófisis anterior y los linfocitos sanguíneos”, es otro de los novedosos estudios realizados en esta década, sobre la influencia de las glándulas de secreción interna en el control de la linfocitosis. 197 RESULTADOS Ya es sabido que la secreción de la glándula pituitaria anterior es esencial para el desarrollo de una linfopenia aguda como resultado de estrés emocional, como lo indican los anteriores trabajos de Dougherty y White (254) y de Colfer, Groot y Harris (279). Se acepta que el estrés emocional estimula al sistema nervioso, éste a su vez a la adenohipófisis, ésta al córtex adrenal y como resultado aparece una linfopenia. Groot y Harris deciden estudiar: a) La estimulación eléctrica de varias regiones del hipotálamo y glándula pituitaria, para comprobar la respuesta linfopénica. Se estudian regiones como el tuber cinereum, cuerpos mamilares, neurohipófisis, pars distal, pars intermedia, zona tuberalis, tallo del infundíbulo y otras regiones como el quiasma óptico. b) Estudiar diferentes partes del hipotálamo, para determinar si la linfopenia que sigue al estrés emocional en conejos normales puede ser bloqueada. Esperan de esta forma, obtener las pruebas necesarias para controlar la secreción hipotalámica y determinar las vías anatómicas que intervienen en dicho control. Para evitar un estrés emocional indeseado en los animales, se utiliza un método de estimulación por control remoto, en conejos sueltos y conscientes. Las observaciones de Groot y Harris (280) indican que: • La estimulación de la región posterior del tuber cinereum o de los cuerpos mamilares en conejos no anestesiados y sin restricciones, dio lugar a linfopenia, similar en cuanto a tiempo de aparición y magnitud, a la observada después de un estímulo de estrés emocional, que la simpatectomía cervical no abolió dicha respuesta, y que la estimulación eléctrica de otras regiones del hipotálamo, de la pars y zona tuberalis, pars distal, intermedia y tallo infundibular de la glándula pituitaria, no provocan esta respuesta. 198 • RESULTADOS La respuesta linfopénica que sigue al estímulo de estrés emocional en conejos normales, fue abolida por lesiones en la zona tuberalis, y en la mayoría de casos fue abolida o disminuida por lesiones transversales en la parte posterior del tuber cinereum o cuerpos mamilares, mientras lesiones similares en la pars distal e intermedia, y lesiones con interrupción del tallo infundibular fueron compatibles con una respuesta normal. Los autores, según estos resultados admiten que: “la secreción pituitaria anterior, probablemente hormona adrenocorticotropa, está bajo control neural a través del hipotálamo y los vasos portales hipofisarios que discurren por el tallo hipofisario”. Un curioso estudio realizado por Tuttle y publicado en 1950, lleva por título: “15 años de estudio de linfocitosis oculta en 280 pacientes con hipercolesterolemia” (281). Del estudio y tratamiento durante un periodo de 15 años, de 280 pacientes con hipercolesterolemia, Tuttle descubre una triada de síntomas, que incluye: 1-­‐ Una linfocitosis relativa. 2-­‐ Hipercolesterolemia. 3-­‐ Hipotiroidismo Es sabido por investigaciones anteriores, entre ellas, las realizadas por Artom en perros tiroidectomizados y por Perlzweig en sujetos hipertiroideos (282), la existencia de una relación inversa entre el funcionamiento de la tiroides y los niveles de colesterol en sangre. Tuttle para determinar la existencia de esta linfocitosis que denominó oculta, hizo recuentos diferenciales de glóbulos blancos según el método de Arneth (283), clasificando a los linfocitos en grandes y pequeños, según la proporción relativa de citoplasma y núcleo, observando en todos los casos una linfocitosis relativa y en algunos casos una linfocitosis absoluta, que considera “oculta” en 199 RESULTADOS el recuento total de glóbulos blancos, normal en la sangre de estos pacientes. Para reconocer fácilmente esta linfocitosis oculta utiliza el “índice linfocítico”, que resulta al dividir por 30 el porcentaje de linfocitos encontrado en cada caso. Tuttle considera este desorden metabólico, la hipercolesterolemia, como una enfermedad degenerativa, ya que según ella: “Las enfermedades degenerativas son aquellas en las que están implicadas estructuras mesodérmicas, y los linfocitos son estructuras mesodérmicas”. Después de exponer de forma resumida 21 de los casos seleccionados al azar, revelar los síntomas clínicos más comunes, y establecer una serie de medidas terapéuticas encaminadas a corregir el desorden metabólico, como dieta, estimulantes tiroideos, control de peso, psicoterapia, tratamientos de bio-­‐oxidación y agentes lipotrópicos, manifiesta que: 1. En los casos de hipercolesterolemia e hipotiroidismo, se detecta una linfocitosis oculta. 2. La actividad tiroidea se refleja por una diferenciación celular de derivados mesodérmicos, expresada por el aumento selectivo de linfocitos. 3. Se obtienen resultados gratificantes mediante las medidas terapéuticas, con un re-­‐equilibrio de las células blancas, una disminución del colesterol sanguíneo y una corrección del metabolismo basal a niveles normales. 4. Un nuevo agente reductor de colesterol “Povon”, derivado de la lecitina del aceite de las semillas de girasol, es utilizado con éxito en los casos que no responden a hipocolesterolémicos habituales como colina, metionina e inositol. En este trabajo de Tuttle, los linfocitos son referidos como parte de la triada sintomática, como una consecuencia o un indicador de la alteración metabólica, la hipercolesterolemia. En realidad el motivo de estudio no es la linfocitosis, sino la alteración metabólica y se presta una gran importancia al tratamiento de la misma, realizando recomendaciones terapéuticas y hasta la “promoción” de un nuevo fármaco para disminuir los niveles de colesterol, el “Povon”. Por lo 200 RESULTADOS que, a nuestro entender, un título como “Presencia de linfocitosis en 280 pacientes hipercolesterolémicos en un estudio de 15 años”, reflejaría mejor el contenido del trabajo. Schweizer y Reber (284), en 1950, escriben en lengua germana sobre ”La formación de anticuerpos en los linfocitos”. En base a los estudios realizados en animales en los últimos años, la teoría que atribuye a los linfocitos capacidad para elaborar anticuerpo cobra importancia. En este sentido podemos nombrar los trabajos de McMaster y Kidd en 1937, y de McMaster y Hudack en 1935 (225). Schweizer y Reber utilizan gatos para sus investigaciones, y emplean como antígeno el bacilo tífico. Estudian la formación de anticuerpos en los primeros seis días después del estímulo antigénico, las fracciones de proteínas de los mismos y el número de linfocitos en la linfa del conducto torácico del gato. Observaron la aparición de aglutininas en la linfa tres días después de la inyección subcutánea de antígeno en la pata trasera del gato. Sin embargo, a los seis días, el título de aglutininas era significativamente mayor en el suero, y deducen que las aglutininas de la linfa son, en parte debidas al paso de anticuerpos a partir del suero. Cuando estudian las suspensiones de extractos de linfocitos del conducto torácico de gatos vacunados, no encuentran ni sustancias aglutinantes ni anticuerpos protectores, además el número de linfocitos en la linfa del conducto torácico se mantiene sin cambios durante los primeros días de la formación de anticuerpos, aumentando sólo ligeramente al sexto día. Asimismo, observan que el contenido de proteínas en suero y linfa se mantiene sin cambios en el primer día de formación de anticuerpos, y posteriormente discurren de forma paralela. En base a los resultados obtenidos, Schweizer y Reber (284) dicen que existe una falta de definición en relación con los linfocitos, no estando de acuerdo con la teoría de que los anticuerpos se forman en los linfocitos de los ganglios linfáticos regionales. 201 RESULTADOS Un tema ampliamente estudiado en años anteriores, es ahora, a finales de esta quinta década, motivo de nuevas investigaciones. Glenn, Bauer y Cresson, exponen sus observaciones sobre “la salida de linfocitos en la fístula del conducto torácico realizada experimentalmente” (285). Gracias a la utilización del cloruro de polivinilo y polietileno para la fabricación de cánulas, ha sido posible el estudio del conducto torácico, mediante la producción de fístulas, durante tiempos más prolongados que en años anteriores. Es sabido, el gran número de células, de las cuales el 98% son linfocitos, que pasan continuamente a sangre desde el conducto torácico. Yoffey (197) calculó, para un perro de unos 10 kilogramos, una circulación de linfocitos desde el conducto torácico a sangre, a razón de 200 millones de linfocitos por hora. Considerando las diversas teorías sobre el destino de los linfocitos, Ehrlich declaró: “Después de haber sido formado en el tejido linfático, el linfocito pasa a través de los vasos linfáticos, y posiblemente, también a través de las venas, al torrente sanguíneo donde circula durante varias horas. Después de este tiempo, algunos abandonan la sangre a través de las mucosas del tracto gastrointestinal, mientras que, probablemente la mayoría, regresan al tejido linfático a través de los vasos linfáticos periféricos. Es probable que muchos de estos linfocitos sigan y vuelvan a la sangre, de hecho este ciclo se puede repetir varias veces”. Glenn, Bauer y Cresson llevaron a cabo sus experimentos en 7 perros con fístula experimental del conducto torácico de 2 a 8 días de duración, a los que se realizó recuentos totales y diferenciales de células blancas, cada día, en sangre venosa periférica, determinando el flujo linfático simultáneamente. Observaron una caída en la concentración de linfocitos en sangre después de la realización de la fístula, lo que está de acuerdo con otros trabajos como los de Adams, Sanders y Lawrence (286) realizados en gatos. El hallazgo de una disminución de linfocitos después de varios días de drenaje linfático continuo, es considerado por los autores de gran interés, pero no aportan nada concluyente que explique el fenómeno. Tampoco queda claro la importancia del porcentaje de los distintos tamaños linfocitarios después de varios días de 202 RESULTADOS drenaje, reseñando la aparición de un aumento en el porcentaje de células con citoplasma profundamente basófilo, que puede interpretarse, según las observaciones de Wiseman, como indicación de que la caída de los linfocitos de salida fue a expensas del linfocito con citoplasma menos basófilo, el linfocito más viejo. Glenn, Bauer y Cresson (285), analizando los resultados creen necesario tomar en consideración los muchos factores que puedan influir en la evaluación de la salida de los linfocitos del conducto torácico. Ya en 1921 Bunting y Huston (120) abordan el problema del destino de los linfocitos, observando que:”… los linfocitos no muestran ninguna tendencia a la desintegración. Deben abandonar el torrente sanguíneo”. En nuestra opinión, la aportación más interesante de Glenn, Bauer y Cresson (285), en un tema repetidamente estudiado a lo largo de los años es la intuición de encontrase ante un proceso fisiológico muy complejo, en el que intervienen muchos factores todavía desconocidos (ellos reconocen que sólo destacan un único factor), y el reconocer no disponer de los medios técnicos para su completa resolución. En este sentido manifiestan: “… en experimentos de varios días, hay muchos factores que pueden influir en la producción de linfocitos. El hambre se sabe que tiene un efecto depresor… Factores distintos a la simple pérdida de linfocitos a través de la fístula del conducto torácico pueden haber influido en la salida de los linfocitos”. Con la introducción de las cánulas de plástico, el método utilizado mejora notablemente, y a pesar de los problemas que todavía se presentan como la deshidratación del animal o la contaminación de la cánula, suponen un avance respecto a las de vidrio, y posibilita un estudio durante un tiempo más prolongado; sin embargo, y a pesar de las mejoras, el estudio no es concluyente. 203 RESULTADOS 4.5. LA SEXTA DÉCADA DEL SIGLO XX (1951-­‐1960) En estos años va a producirse un acelerado crecimiento económico en América y Europa, incluso los países más perjudicados por la Segunda Guerra Mundial lograron mejorar su producción y economía. La gran expansión industrial, fundamentalmente en EE.UU se traducirá en un aumento del consumo por gran parte de la población, dando lugar al “American way of life”, una nueva forma de vida que se convertirá en modelo para el resto del mundo. El coche, los electrodomésticos y la televisión pasan a ser elementos cotidianos e indispensables en los hogares; surge la sociedad de consumo (287). La incorporación al mundo laboral de la mujer cambiará las relaciones familiares y los hábitos alimenticios de la familia, aparece la hamburguesa y la comida rápida. Como respuesta a esta nueva forma de vida nacen dos grandes movimientos contraculturales: el rock, representado por los Rolling Stone y los Beatles, y un movimiento juvenil y político que originado en la universidad francesa e italiana se extiende por toda Europa, y que en Francia provocó la mayor huelga general en la historia de este país, conocida como “Mayo Francés” (220). Esta década también queda marcada por el comienzo de una nueva lucha por los derechos civiles de los negros, la división de Alemania con un muro, las primeras acciones bélicas en Vietnam y el inicio de la carrera espacial con Yudi Gargarin que da la vuelta a la Tierra en 104 minutos. Coincidiendo con estos avances se aprecia un aumento significativo en el número de artículos publicados que hacen referencia al linfocito. La búsqueda en Scopus con la palabra lymphocyt* nos muestra un total de 1129 documentos, lo que supone un 0,202% del total de publicaciones registradas en el siglo XX. Podemos decir que el aumento es significativo, comparado con la década precedente (0,0574%). Se observa un predominio de la lengua inglesa como “lengua oficial científica”, sin embargo, se advierte también como fenómeno nuevo la aparición de otras 204 RESULTADOS lenguas distintas al alemán, que era hasta ahora la segunda lengua más utilizada, como por ejemplo la lengua rusa, checa, polaca o italiana en algunos de los trabajos, cuyo contenido no obstante sigue la línea general de motivos de estudio registrada en estos años. Los estudios aquí comentados dan una idea global de las tendencias y las publicaciones en este periodo de tiempo, y nos muestran como han ido evolucionando los trabajos sobre el linfocito, las principales líneas de investigación y los campos en los que el linfocito suscita interés. Si en la década anterior la novedad fueron los trabajos que relacionaban linfocito con enfermedades endocrinas y psiquiátricas, en esta década un gran número de estudios versan sobre la fisiopatología de las hormonas y su influencia en los linfocitos, y tímidamente aparecen las primeras publicaciones de genética aplicada a las células sanguíneas (288). Con el título “Localización de antígenos en células tisulares. VI. Destino de proteínas extrañas inyectadas en el ratón”, Coons, Leduc y Kaplan (289) publican en 1951 los resultados de sus investigaciones. Un breve resumen de trabajos realizados con anterioridad sirve como actualización sobre el tema. Mencionan, entre otros, a Sabin (290) quien encontró que la albúmina de huevo R-­‐salt-­‐azo-­‐bencidina, después de ser inyectada es ingerida por las células de Kupffer, macrófagos, neutrófilos, endotelio linfático, monocitos, células de médula ósea y fibroblastos, y a Kruse y McMaster (291) que llevaron a cabo un cuidadoso estudio con azoproteínas (suero de caballo y globulinas bovinas), demostrando que éstas son tomadas por el sistema reticuloendotelial y por los fibroblastos en todos los órganos, así como por células de los túbulos renales y músculo liso en cambio no se encontró en linfocitos ni en cerebro. Coons, Leduc y Kaplan (289) centran sus estudios en tres proteínas: albúmina de huevo cristalina, albúmina bovina cristalina y γ-­‐globulina humana. Emplearon ratones a los que inyectaron las proteínas a estudiar vía intravenosa, siendo sacrificados a intervalos variables de tiempo para el estudio histológico de sus órganos, mediante secciones congeladas de tejido en 205 RESULTADOS microscopio de fluorescencia. Gracias a una serie de modificaciones bioquímicas con componentes aromáticos azo y carbamido se hizo posible el uso del anticuerpo acoplado a isocianato de floresceína, quedando un precipitado inmune sobre el área de la sección de tejido que contiene el antígeno homólogo. El precipitado es fluorescente brillante bajo el microscopio e indica la presencia de antígeno, haciendo posible el estudio de la distribución en tejidos animales de sustancias extrañas, con la única condición de que sean lo suficientemente antigénicas como para estimular la producción de anticuerpos. Se presentan pruebas de que estas moléculas o fragmentos de ellas penetran en la membrana nuclear de varios tipos de células y se pueden encontrar en el núcleo. En los resultados (289) observaron que los lugares de depósito de las tres proteínas eran muy similares a pesar de las diferencias existentes en sus propiedades, y respecto a la distribución de las proteínas, se constató que el lugar con mayor depósito de antígeno era la adventicia arterial, especialmente de riñón. Un hallazgo constante era la presencia de las tres proteínas en el revestimiento endotelial de venas de riñón e hígado. El hallazgo más inesperado en el curso del experimento fue la presencia de proteínas extrañas en los núcleos de varios tipos celulares: “Si las observaciones no son el resultado de un artefacto, indican una relación íntima, hasta límites insospechados, entre sangre circulante y núcleo de algunas células”, puntualizan. Y añaden: “Las proteínas motivo de estudio se encontraron tanto en el citoplasma como en el núcleo de macrófagos, células reticulares y linfocitos, estos últimos sospechosos de la producción de anticuerpos”. En definitiva, el destino de las tres proteínas tras su administración intravenosa en el ratón, fueron las células del sistema reticuloendotelial, tejido conectivo, endotelio vascular, linfocitos de bazo y ganglios linfáticos, células de túbulos de riñón e hígado, y se encontró que cada una de las tres proteínas estaba presente en los núcleos de cada tipo de célula mencionada, a menudo en mayor concentración que en el citoplasma. 206 RESULTADOS El trabajo de Coons y sus colaboradores (289) es uno más de los numerosos estudios, que con más o menos acierto o fortuna abordan la complejidad de antígenos y anticuerpos, pero en realidad poco aporta al conocimiento de linfocito. Podemos clasificarlo como un estudio de tipo anatómico-­‐histológico-­‐ inmunológico, que siguen estando vigentes en esta sexta década. Quizás la contribución más importante de este trabajo es la mejora que introduce en el método para la realización de cortes de tejidos en congelación, necesarios para el microscopio de fluorescencia; sobre el tema, los autores manifiestan: “la manipulación de cortes de 4 μ de grosor es delicado, por lo que se modificó el método a fin de hacerlo más fácil”. Con este fin se construyó un criostato mucho más fácil de manejar que permite con una pequeña práctica cortar secciones congeladas no teñidas a 4 μ de grosor de forma rutinaria. Esta pequeña aportación tecnológica modificando un criostato para hacerlo más manejable, y que facilita el método de preparación de las muestras para fluorescencia va a favorecer e impulsar el desarrollo de posteriores estudios que utilizan esta técnica. De índole muy distinta es el trabajo de Weisberger y colaboradores (292): “El papel de los pulmones en la eliminación de los linfocitos transfundidos” de 1951. En muchas condiciones caracterizadas por la presencia de leucopenia o granulocitopenia sería deseable un aumento de los leucocitos circulantes y, sin embargo, ello no es posible debido al fallo en las transfusiones para aumentar el número de células blancas de la sangre de forma análoga a como se hace en el caso de células rojas en pacientes con anemia. Se ha sugerido, que es debido a la corta vida de los leucocitos o a posibles incompatibilidades de los grupos sanguíneos. En estudios previos, realizados por algunos autores de este grupo de trabajo (293)(294) se observó, que una transfusión de concentrado de granulocitos no producía un aumento de leucocitos, ya que en cuestión de minutos aparecía la leucopenia, y se mostró que era debido a una acción selectiva del pulmón que eliminaba las células transfundidas administradas de forma intravenosa o intraperitoneal. 207 RESULTADOS Weisberger y colaboradores (292) emplearon ratas, de las que obtuvieron linfocitos, los cuales fueron marcados con un trazador radiactivo (P32) para hacer posible su seguimiento. Los animales fueron sacrificados de 10 a 15 minutos después de la administración de suspensiones de estos linfocitos marcados para estudio de la radiactividad en pulmón, hígado, bazo y riñones, y comparando la distribución de la radiactividad con la existente en controles animales que la recibieron en forma de ión fosfato inorgánico, pudieron constatar lo siguiente: ! Una disminución en el número total de linfocitos circulantes después de la primera hora, siendo esta disminución inversamente proporcional al incremento de neutrófilos. ! Una afinidad selectividad del pulmón por los linfocitos radiactivos marcados con P32, que es fuerte, si se compara con la distribución de la radiactividad que sigue a la inyección de fósforo inorgánico administrada a los controles. ! Que la diferencia en la distribución de la radiactividad después de la administración de células desintegradas, indica que la mayoría de las células fueron eliminadas intactas. 208 RESULTADOS Figura 26. Valor del número de linfocitos después de la administración de una suspensión de linfocitos concentrados. Tomado de Weisberger y col, 1951 (292). Según estos resultados, los autores (292) consideraron que la administración de grandes cantidades de linfocitos no pudo aumentar el número de linfocitos circulantes, ni siquiera temporalmente. Después de la administración de linfocitos radiactivos intravenosos o intraarteriales en 17 animales, la mayoría de la radiactividad fue recuperada en los pulmones, hígado, bazo y riñones. De estos órganos, el pulmón y el bazo contenían cantidades de radioactividad muy superiores a las encontradas en los animales que recibieron dosis intravenosas de fósforo inorgánico. Los autores manifiestan: “Podemos concluir que el pulmón y el bazo son los órganos más activos en la eliminación de linfocitos transfundidos, y aunque una considerable cantidad de radioactividad fue detectada en bazo, es posible que debido a su pequeño tamaño no contribuya significativamente a la eliminación de linfocitos de la circulación. El papel del pulmón es de gran importancia si se considera que contiene más radiactividad que el hígado y su peso es inferior a una quinta parte del peso del hígado”, y elaboran algunas conclusiones, como que la disminución del número de 209 RESULTADOS linfocitos circulantes en ratas después de su administración intravascular es el resultado de la acción selectiva específica de los pulmones, y que tal vez hígado y riñones jueguen algún papel en la eliminación de los linfocitos transfundidos, pero indudablemente no son tan activos como los pulmones a este respecto. Este estudio (292) confirma la importancia de la barrera pulmonar en la transfusión de leucocitos, y señala al pulmón como el órgano principal involucrado en la eliminación de células transfundidas, existiendo la posibilidad de que los pulmones puedan actuar como órgano homeostático en el control del nivel de células blancas. Es probable que la eliminación de estas células de la circulación sea representativa del destino de los linfocitos administrados en las transfusiones clínicas usuales y que el pulmón pueda formar parte de un mecanismo fisiológico normal para controlar el número de leucocitos circulantes. El papel de las hormonas en la producción linfoide es un tema de plena actualidad a principio de los años cincuenta. “Efecto del ACTH y extracto suprarrenal en la médula ósea”, es el título de otro de los trabajos que encontramos publicados en 1951, figurando como principales autores Yoffey y Metcalf de la Universidad de Bristol (295). La existencia de una conexión entre tejido linfoide y corteza suprarrenal ha sido postulada desde que la clásica monografía de Addison apareció por primera vez en 1855 (296)(297). A partir de entonces, la información acumulada sobre la hipertrofia del tejido linfoide en la enfermedad es tal, que esta hipertrofia es considerada como una de las características de la enfermedad de Addison, siendo descrita como “estado linfático”. Dougherty y White (254) por su parte, llevaron a cabo un extenso estudio sobre la relación entre corteza suprarrenal y tejido linfoide desde 1943 a 1947, publicando una serie de artículos sobre el tema. Yoffey y Metcalf (295) admiten la ignorancia existente sobre el papel de los linfocitos y realizan un breve repaso sobre el nivel de linfocitos en sangre. Se 210 RESULTADOS sabe que se forman en el tejido linfoide y desde aquí, pueden dirigirse directa o indirectamente a la sangre. Una vez que los linfocitos han entrado en el torrente sanguíneo no permanecen allí por mucho tiempo y no existen pruebas satisfactorias de su destrucción en sangre, pero como un gran número de nuevos linfocitos están continuamente entrando en sangre, se debe asumir que un número equivalente de estos linfocitos deben abandonar el torrente sanguíneo, ya que de no ser así, el número de linfocitos no sería constante. Y se sabe que en el perro los linfocitos de la sangre se renuevan dos veces al día (197)(204), y de cuatro a cinco veces en el gato y conejo (298). La luz del tracto alimentario fue considerada el destino más probable de los linfocitos de la sangre, pero esta posibilidad fue desechada. En cuanto al origen de los elementos formes de la sangre en la médula ósea, con un contenido de linfocitos asombrosamente elevado, ha sido motivo de una de las controversias hematológicas de más larga duración, y la tendencia general ha sido ignorarla. Sopesando todos estos hechos, Yoffey y Metcalf consideran que merece la pena investigar el contenido de linfocitos en médula ósea después de la administración de ACTH y extractos corticales suprarrenales, y creen que el análisis cuantitativo sería lo más adecuado. La médula ósea es obtenida de cobayas sacrificados y desangrados a fin de eliminar el exceso de sangre en el animal. Se realizaron recuentos en la médula ósea de cuatro grupos de estudio: 1. Con médula ósea normal. 2. Seis horas después de la administración intraperitoneal de extracto de corteza suprarrenal (eschatin). 3. Seis horas después de la administración de ACTH de hipófisis. 4. En un grupo control de 10 cobayas a los que se les administró inyecciones intraperitoneales de solución salina. Los resultados (295) de estos experimentos se recogen en la Fig 27 que muestra la media de los valores del recuento de linfocitos tanto en porcentaje como en valor absoluto en médula ósea normal, en controles, y después de la 211 RESULTADOS administración de eschatin y ACTH. En estos dos últimos casos, el valor de los linfocitos es superior al de médula ósea normal. Figura 27. Contenido de linfocitos en médula ósea de animales normales y de experimentación. Tomado de Yoffey y Metcalf, 1951 (295). Se encontró que en el cobaya normal la médula ósea contenía aproximadamente 1236000 células nucleadas por ml, de las cuales 144800 fueron linfocitos, y que este alto contenido de linfocitos en médula ósea no es debido a contaminación sanguínea. No hubo un aumento demostrable de células dañadas en médula ósea después de la administración de ACTH o eschatin, pero sí un aumento estadísticamente significativo del contenido de linfocitos con eschatin, y un aumento casi alcanzando el nivel significativo en el caso de ACTH. Los autores (295) manifiestan como conclusiones: “Es posible que el aumento en la captación por médula ósea de los linfocitos sanguíneos pueda ser un factor contribuyente en la linfopenia que puede seguir a la administración de ACTH u hormonas esteroides. Cualquiera que sea el efecto de estas hormonas esteroides en el tejido linfoide de otras partes del cuerpo, no ha 212 RESULTADOS sido posible demostrar por métodos cuantitativos una acción destructiva de los linfocitos en médula ósea”. En estos años podemos apreciar un enorme interés por el conocimiento de la fisiopatología de las hormonas, especialmente hormonas hipofisarias y de las glándulas suprarrenales, sobre los elementos celulares de la sangre. Los trabajos recogidos en ésta (34 documentos = 3%) y la anterior década (9 documentos) son los primeros que abordan el tema, observándose ya en la siguiente década (años sesenta) un considerable aumento en el número de publicaciones al respecto (592 documentos). En 1952, Trowell (299) es el autor de: “La acción de la cortisona sobre los linfocitos in vitro”. Parece no haber duda sobre el efecto que tiene el estrés en el tejido linfoide, ya que la exposición de un animal a estrés emocional conduce en unas pocas horas a la muerte de los linfocitos. Este efecto parece estar mediado por la corteza suprarrenal, pues no se produce en el animal adrenalectomizado, y puede ser provocado mediante la inyección de extracto de corteza suprarrenal o de cualquiera de los esteroides adrenales C-­‐11 oxigenados, como por ejemplo cortisona. La pregunta que surge, es si los esteroides C-­‐11 oxigenados son tóxicos directos para los linfocitos o si el efecto es resultado de una acción indirecta, una cuestión que, según Trowell, puede ser resuelta mediante la observación de los linfocitos cultivados in vitro. Varias investigaciones realizadas con anterioridad habían dado resultados dispares; así, Heilman en 1945, trabajando con cultivos de ganglios linfáticos, encontró que la cortisona causa cierta disminución en la migración de los linfocitos y un aumento en la degeneración de los mismos después de 24 horas, en cambio, otros autores como Baldridge, Kligman, Lipnik y Pillsbury (300) no pudieron confirmar estas observaciones. Trowell da la siguiente explicación a esta discordancia: “los resultados tan dispares de estos y otros trabajos similares sugieren la existencia de dificultades técnicas desconocidas o no resueltas; de éstas, la principal consiste en mantener a los linfocitos in vitro sanos y vivos, aunque sea por unas horas. El método empleado para el cultivo no conseguía este objetivo, y los cultivos de control empleados por todos los autores mencionados contenían un 213 RESULTADOS considerable porcentaje de linfocitos muertos desde el inicio, que iba aumentando durante el tiempo de cultivo”. Señala también como posible causa el empleo de las preparaciones clínicas de esteroides que contienen aditivos como conservantes, disolventes o estabilizadores que pueden ser tóxicos para las células in vitro. El autor (299) dice haber superado estas dificultades y emplea una nueva técnica para el mantenimiento de linfocitos in vitro, basada en el cultivo de ganglios linfáticos intactos de ratas jóvenes sobre la superficie de un lecho húmedo de lana y algodón en una atmósfera oxigenada, reduciendo así el porcentaje de linfocitos muertos o degenerados aproximadamente a un 2%. Sobre estas bases examina el efecto de varios esteroides como cortisona, 11-­‐ deoxycorticosterona, acetato de 11-­‐doxycorticosterona, testosterona, estradiol y progesterona, y utiliza el valor del recuento de núcleos picnóticos como medida cuantitativa del efecto citotóxico. De sus observaciones obtiene los siguientes resultados: • Cuando la cortisona se inyecta a un animal, la destrucción linfocitaria es máxima a las seis horas. A baja concentración el efecto tóxico es ya evidente, pero el aumento 100 veces de la concentración de cortisona sólo incrementó ligeramente el efecto tóxico, a diferencia de lo que ocurre cuando se emplea cianuro, mercurio y rayos X bajo similares condiciones. Estos resultados sugieren que el tiempo, en lugar de la concentración, es el factor limitante, y que la acción tóxica intensa de la cortisona podría llegar a ser más tardía que la del cianuro, mercurio o rayos X. • Cuando la cortisona a una concentración media se deja actuar durante períodos más largos de tiempo, el efecto aumenta con el tiempo, siendo máximo a las 46 horas. • Estos resultados confirman el hecho de que los esteroides corticales tienen una acción citotóxica leve y retrasada en el tiempo sobre los linfocitos in vitro. No se encontraron cambios citológicos específicos en 214 RESULTADOS los linfocitos expuestos a cortisona, ya que las células simplemente sufrieron un proceso de degeneración picnótica, que constituye un proceso de autólisis no específica en una célula ya muerta, y los cambios citológicos son exactamente los mismos si las células mueren por cortisona, cianuro, rayos X, anoxia, disminución de glucosa o trauma mecánico. • Los resultados con otros esteroides muestran que el efecto linfotóxico es específico para algunos esteroides adrenales. Parece ser que el grupo HO.CH2.CO en C-­‐17 y el átomo de oxígeno en C-­‐11 son los implicados en la acción linfotóxica. Testosterona, estradiol y progesterona no tuvieron ningún efecto linfocitotóxico. Se sugiere que la sangre normal podría contener suficientes esteroides naturales que afecten a los linfocitos, y la cuestión es saber si el suero empleado en los experimentos contenía esteroides en suficiente cantidad como para influir en los resultados. Con esta finalidad se repiten algunos experimentos utilizando suero de ratas adrenalectomizadas. Los ganglios linfáticos cultivados bajo estas condiciones tenían un valor medio de linfocitos picnóticos de 3,3%, ligeramente superior al índice de linfocitos picnóticos en los ganglios linfáticos de ratas normales que se sitúa aproximadamente en el 1%. Estos resultados no apoyan la idea de que la picnosis encontrada en cultivos normales se deba a la presencia de hormona suprarrenal natural en los medios de cultivo, de hecho, el suero de los animales adrenalectomizados parece ser más tóxico que el suero normal. Las diferencias en la picnosis utilizando suero de animales adrenalectomizados y no adrenalectomizados no son estadísticamente significativas, y se concluye que con estos métodos no se detectaron esteroides linfotóxicos de origen adrenal en el suero normal. Con la esperanza de obtener información sobre el mecanismo de acción de la cortisona en el linfocito Trowell (299) en otra serie de experimentos añade al medio de cultivo una serie de sustancias como insulina, potasio, suero de una colonia de ratas diferentes, testosterona, purinas y pirimidinas. De todas ellas 215 RESULTADOS únicamente se encuentran diferencias significativas con el suero de ratas diferentes, que el autor considera necesario estudiar más detalladamente, y con la adición de purinas y pirimidinas que reducen ligeramente el efecto de la cortisona. El autor en la discusión da una posible explicación: “el efecto protector de estas dos sustancias parece demasiado pequeño para que tenga una gran importancia... La cortisona podría actuar por inhibición de la síntesis de purinas y pirimidinas necesarias para el recambio de los ácidos nucleicos, pero igualmente la cortisona podría simplemente hacer que los linfocitos fueran menos permeables a las purinas y pirimidinas del medio”. Y concluye diciendo: “Es difícil creer que una hormona natural sea específicamente tóxica para un tipo particular de célula, y que esto sea un efecto secundario fortuito, que en la reacción de alarma los linfocitos mueran en vano… quizás la clave de la función de los linfocitos se encuentre entre los productos de la lisis linfocitaria”. Nuestra opinión al respecto es que Trowell lleva acabo un laborioso trabajo, muy completo y detallado que aporta algunos conocimientos sobre la acción de varias hormonas en el linfocito. Da un gran paso cuando emplea una nueva técnica para el cultivo de linfocitos in vitro, que asegura: “reduce el porcentaje de linfocitos muertos”, concede importancia al factor tiempo de actuación de un fármaco “es el factor limitante” dice, frente al factor concentración considerado tradicionalmente el más importante, aclara que el modo de acción tóxica es inespecífico, “un proceso de autólisis no específica” común a muchas causas de muerte celular, identifica asimismo los grupos químicos implicados en la acción linfocitotóxica y asegura no encontrar esteroides linfotóxicos de origen adrenal en el suero normal. En esta línea de investigación sobre esteroides y tejido linfoide, se encuentra también el trabajo de Hungerford, Reinhardt y Hao Li (301) en 1952, de título: “Efectos de la cortisona e hidrocortisona en el número de linfocitos del conducto torácico”. Está bien establecido que la ACTH y ciertos corticoides adrenales pueden disminuir el número de linfocitos circulantes en sangre, y ello podría resultar 216 RESULTADOS de la disminución en el aporte de estas células por parte del conducto torácico, pero los intentos para demostrar esto último no han tenido éxito. Los autores deciden estudiar los efectos de los esteroides sintéticos, cortisona e hidrocortisona, sobre el contenido linfoide del conducto torácico en ratas normales y hipofisectomizadas. Se emplearon ratas de la cepa Lonsg-­‐Evans de aproximadamente 60 días de edad, normales e hipofisectomizadas, y se calculó el número total de linfocitos por muestra de linfa obtenida del saco linfático yugular, extrapolando el valor en 24 horas. En ratas normales, 4 horas después de la administración de 10 mg de hidrocortisona, se observó que el número total de linfocitos del conducto torácico se redujo significativamente (p<0,01) en comparación con los controles normales. Cuatro horas después de la administración de cortisona, había tenido lugar una disminución del número total de células, pero ésta era menos marcada que con hidrocortisona. En ratas hipofisectomizadas, se observó que la hidrocortisona produce una disminución altamente significativa de células en el flujo linfático, en comparación con el efecto de la cortisona. Este estudio (301) que compara el efecto de cortisona e hidrocortisona a dosis de 10 mg, muestra que a esta dosis, la hidrocortisona es mucho más eficaz que la cortisona en la disminución de los linfocitos del conducto torácico en ratas normales e hipofisectomizadas. Es de interés destacar que un estudio preliminar de la actividad relativa de la cortisona e hidrocortisona, usando como test la disminución de eosinófilos en sangre de ratas, mostró que la hidrocortisona es más eosinopénica que la cortisona. Teniendo en cuenta estas observaciones, los autores manifiestan su opinión al respecto: “estos resultados conducen a la hipótesis de que la linfocitopenia del conducto torácico producida por la ACTH es más probablemente dependiente de la liberación de hidrocortisona (o sustancia similar) que de la liberación de cortisona”, demostrando que la hidrocortisona produce una marcada linfopenia en el conducto torácico, efecto que es más marcado en ratas hipofisectomizadas que en normales, y que a niveles comparables, la hidrocortisona es significativamente más efectiva en producir linfocitopenia que la cortisona. 217 RESULTADOS Si Hungerford, Reinhardt y Hao Li trabajaron en el terreno de la fisiopatología, estudios de tipo morfológico a nivel celular son realizados por Hempelmann y Knowlton (302) que dirigen su interés a la investigación de la naturaleza de los gránulos del linfocito, publicando en 1953: “La naturaleza de las partículas de refracción teñidas con rojo neutro en los linfocitos de la sangre de humanos normales”. La existencia de partículas citoplasmáticas de refracción que se tiñen con rojo neutro en los linfocitos de sangre periférica humana es conocida desde que Gall (303) las describió en 1936. Se observó que el número de estas partículas aumentaba en ciertas condiciones, como en personas expuestas a radiaciones ionizantes o a ciertos tóxicos químicos, pero el significado de este aumento de cuerpos refractarios no está claro, ni se conoce su relación con el tejido dañado. Gall describe con detalle la apariencia de estas partículas: “su tamaño usual es considerablemente menor que 1μ, y su forma exacta está enmascarada por su fuerte refracción, pero normalmente son esféricas y a veces de forma irregular. En preparaciones de sangre fresca, estas partículas pueden verse con aparente libertad de movimiento en el citoplasma”. El estudio realizado por Hempelmann y Knowlton (302) recoge algunas de las propiedades ópticas de dichas partículas, como su color, índice de refracción, y absorción de luz ultravioleta, quedando también reflejado el efecto que tienen en los cuerpos de refracción los agentes químicos como solventes, ácidos y bases. Se tuvieron en cuenta asimismo, las reacciones histoquímicas durante el proceso de tinción de dichas partículas con lípidos, polisacáridos, ácidos nucleicos y tinción supravital, entre otros, y se llevó a cabo una valoración de la producción de cuerpos de refracción de linfocitos in vitro. El resultado del estudio indica claramente que el número promedio de partículas de refracción en linfocitos teñidos fue aproximadamente el doble que en células no teñidas, y que el rojo neutro fue más efectivo que el colorante verde janus. Muestran que las propiedades de las partículas de refracción son idénticas a aquellas previamente descritas por Gall, y el estudio de la naturaleza química de estos cuerpos indica que: 218 RESULTADOS a) Están compuestos por material lipídico, ya que se tiñen con colorantes de grasa, como sudán negro o ácido ósmico. b) Se encuentran en un estado físico relativamente estable, como lo indica su resistencia a la destrucción por cambios en el PH, c) No contienen grandes cantidades de polisacáridos o ácidos nucleicos. Los autores (302) resumen los resultados: “las partículas de refracción vistas en linfocitos y que se tiñen con rojo neutro, son birrefrigentes, su índice de refracción se sitúa entre 1,45 y 1,48, son solubles en disolventes de grasa, en alcohol, ácidos o bases fuertes, pero no en débiles, se tiñen con colorantes básicos y de grasa y su número aumenta por la acción del rojo neutro o tinción supravital”, y establecen una serie de conclusiones: “Estas propiedades se interpretan como indicativas de que las partículas vistas en linfocitos humanos se componen de material lipídico de tipo polar-­‐apolar, siendo probable que la mayor parte de la grasa sea de naturaleza fosfolipídica, y seguramente corresponden a las gotas de fosfolípidos que se encuentran en las células somáticas de otros tejidos en mamíferos”. También aparecen es esta década estudios sobre las propiedades o naturaleza de los linfocitos. En 1953, H. Harris (304) de la Escuela de Patología Sir William Dunn de Oxford, publica bajo el título “El movimiento de los linfocitos” sus observaciones al respecto. Ya en 1875, Ranvier describe el movimiento ameboide in vitro de linfocitos, que a partir de entonces es estudiado por diversos autores como Jolly (año 1898), Wolf (año 1901), Neumann (año 1903), Askanazy (305), McCutcheon (306) y Lewis y Webster (98), entre otros. Pero junto a estas observaciones positivas también existen otras que niegan la existencia de movimiento en los linfocitos, como las de Dixon y McCutcheon, las de Schade y Mayr o la de Coman, que encontraron que los linfocitos no exhiben quimiotaxis in vitro bajo las condiciones en las que fácilmente se demuestra quimiotaxis en los granulocitos. Harris (304), trata de resolver el dilema empleando un método de seguimiento fotomicrográfico que ya había empleado con anterioridad para 219 RESULTADOS el estudio de la quimiotaxis en granulocitos y monocitos y que ahora modifica para el estudio de los linfocitos. Obtiene linfocitos del conducto torácico del conejo y registra fotográficamente el movimiento de los mismos por exposición continua bajo iluminación de fondo oscuro. Los linfocitos eran de dos tipos, grandes y pequeños. Los grandes linfocitos variaban mucho de tamaño, y algunos presentaban una tinción característica atribuida a células inmaduras. Numerosas figuras de mitosis se observaron en los grandes linfocitos, pero ninguna entre los pequeños linfocitos. Figura 28. Grandes y pequeños linfocitos Muestra de grandes y pequeños linfocitos obtenidos por canulación del conducto torácico. (May-Griinwald-Giemsa. X 760.) Tomado de Harris, 1953 (304). Todas las células estaban vivas cuando fueron incorporadas a una película de plasma para su estudio, pero solamente el 20-­‐30 % eran móviles. La proporción de células móviles, por lo general comenzó a disminuir después de la primera hora de incubación, y a las 6 horas sólo fue vista una célula móvil ocasional. Durante este tiempo algunas células murieron. Se observó que tanto 220 RESULTADOS los grandes como los pequeños linfocitos eran capaces de moverse, si bien los linfocitos pequeños eran más activos, y existía una gran variación en la velocidad de movimiento de las diferentes células. Los caminos seguidos por las células eran bastantes irregulares, aunque no era raro que los pequeños linfocitos se desplazaran en línea recta. Los linfocitos no se adhieren a las superficies de vidrio, y esto permitió hacer una clara distinción entre linfocitos grandes y monocitos. Se probó el efecto que diversos microorganismos tenían sobre los linfocitos, y en ningún caso se pudo ver ninguna señal de quimiotaxis. Los linfocitos seguían moviéndose de manera completamente aleatoria y no había acumulación de células en o alrededor del objeto de prueba. Los monocitos ingerían rápidamente el grupo de bacterias, pero no fue nunca observada la fagocitosis por parte de linfocitos grandes o pequeños. Las células parasitadas por virus (virus de la encefalomielitis del ratón) tampoco fueron quimiotácticas para los linfocitos. En la discusión, Harris (304) realiza la siguiente reflexión: “… es posible que las condiciones adecuadas para la demostración de la quimiotaxis de linfocitos aún no se hayan encontrado, pero los resultados negativos de otros muchos autores, sugieren la posibilidad de que el linfocito no responda a estímulos quimiotácticos”. Dixon y McCutcheon (306) sugieren que las diferencias entre linfocitos y otros leucocitos pueden deberse a diferencias en la superficie de las células, pero investigaciones posteriores no las encontraron. Sin embargo, Harris cree que deben existir, ya que los linfocitos a diferencia de monocitos y granulocitos, no pueden adherirse al vidrio y tienen sólo un ligero contacto con la matriz en la que se mueven. Teniendo en cuenta los resultados del estudio, el autor expresa su opinión al respecto: “Si la capacidad de una célula para la quimiotaxis, depende de las propiedades en su superficie, esta diferencia entre la propiedades de la superficie de linfocitos y otros leucocitos, puede ayudar a explicar la aparente ausencia de quimiotaxis en el linfocito”. 221 RESULTADOS Las nuevas tecnologías son una opción tentadora para profundizar en el estudio de las células linfoides. En 1953, Nicholas Petrakis (307) publica sus observaciones sobre el contenido de ADN en linfocitos: “Estimación microespectrofotométrica del ácido desoxirribonucleico (ADN) contenido en linfocitos humanos normales y leucémicos”. El desarrollo de métodos espectrofotométricos para el estudio de los constituyentes celulares ofrece una herramienta importante en la investigación citológica, útil para reevaluar muchos conceptos basados hasta ahora únicamente en la morfología. A través de la microespectrofotometría se demostró la relación entre la síntesis de proteínas y los ácidos nucleicos, particularmente con respecto al ácido ribonucleico y las células plasmáticas en el mieloma. El ADN celular está confinado en los cromosomas y esta cantidad de ADN parece ser un valor constante en los núcleos de células somáticas diploides de los tejidos dentro de una especie, siendo el doble de la cantidad presente en los núcleos celulares haploides. Petrakis (307) emplea en su estudio el método de tinción de Feulgen (308) para estimar el contenido relativo de ADN en linfocitos individuales de muestras sanguíneas, ganglios linfáticos y médula ósea de sujetos normales y pacientes con formas agudas y crónicas de leucemia linfocítica. Los linfocitos se obtuvieron de la sangre de 9 sujetos normales y 15 leucémicos. Las células una vez teñidas con el reactivo de Feulgen se sometieron a hidrólisis y se fijaron en alcohol obteniéndose una cromatina de apariencia difusa y homogénea. Los frotis fueron examinados para la búsqueda de figuras mitóticas, pero el conteo de cromosomas tuvo ser abandonado por la pobre definición del material. Petrakis en sus resultados hace referencia a: 1-­‐ Linfocitos normales. Encuentra que en linfocitos normales y espermátides existe una relación directa entre contenido de ADN y tamaño celular en el ganglio linfático, pero no así en linfocitos circulantes. 222 RESULTADOS 2-­‐ Linfocitos leucémicos. Existe una gran variedad en el contenido de ADN de los linfocitos leucémicos, desde valores similares a los de linfocitos normales, hasta cuatro veces los valores de los linfocitos circulantes. La mayoría de los núcleos con valores elevados se encontraban en el rango intermedio entre los valores de los linfocitos periféricos (clase I) y con valores tetraploides (clase II). En definitiva, el trabajo de Petrakis (307) analiza el contenido de ADN, expresado en unidades arbitrarias, de los linfocitos individuales circulantes en sujetos normales y leucémicos, mediante una técnica microespectrofotométrica con tinción de Feulgen, encontrando que los pacientes leucémicos tienen un significativo aumento de linfocitos circulantes con valores de ADN elevados sobre los valores de células diploides normales. Los pacientes que mostraban estos valores elevados de ADN se encontraron en una fase clínica de exarcebación de la enfermedad. A continuación, discute el significado de estos hallazgos, y señala: “Los linfocitos normales tienen un número diploide de cromosomas. No se encontró ninguna alteración en el contenido de ADN entre los diferente tipos de linfocitos, lo que sugiere que la clasificación morfológica en pequeños y grandes linfocitos no es debida a diferencias en el contenido de ADN”. Y aclara que los estudios de médula ósea y ganglios linfáticos de pacientes leucémicos revelaron células con valores diploides, tetraploides e intermedios, dando posibles explicaciones desde el punto de vista citogenético a la presencia de núcleos con valores aumentados de ADN: ! Actividad mitótica. ! Poliploidía y politenia. La poliploidía es un aumento de cromosomas en una célula de forma progresiva, por ejemplo 2, 4, 8 veces el valor diploide de cromosomas (2N). La politenia se caracteriza por reduplicación de cromonemas sin reduplicación de cromosomas, dando como resultado cromosomas de mayor tamaño, sin aumento en el 223 RESULTADOS número total de cromosomas, pero con un contenido de ADN que puede estar marcadamente elevado, dependiendo del grado de politenia. ! Amitosis. Se caracteriza por una desigual separación de cromosomas. La extensa ocurrencia de este proceso debe reflejarse por la presencia de células con valores de ADN por debajo del promedio diploide, y al no observarse linfocitos con disminución de ADN, se sugiere que la amitosis no juega un significativo papel. La fascinación por los recientes descubrimientos sobre el material genético queda patente en trabajos de este tipo, donde el presente no es más que un ejemplo. Es en este año de 1953, cuando Watson y Crick (309)(310) realizan sus publicaciones en una serie de cinco artículos en la revista Nature, donde narran de forma cautelosa el descubrimiento que habían realizado: “… esta estructura tiene una novedosa característica, la cual la hace que tenga un considerable interés biológico”. Y detallaron: “es bastante probable que gran parte o toda la información genética de cualquier organismo se localice en los ácidos nucleicos, generalmente en el ADN, no obstante, también hay algunos pequeños virus que utilizan el ARN como material genético”. Fueron capaces de desvelar la estructura de la doble hélice de la molécula del ácido desoxirribonucleico (ADN) y formular los principios de almacenamiento y transmisión de la información hereditaria. El descubrimiento de Watson y Crick constituyó uno de los logros más importantes del siglo XX, iniciándose una nueva era de la biología molecular, en la que va a ser posible evaluar, cartografiar y modificar la actividad genética. Encontrar la estructura del ADN no fue un proceso sencillo ni fue solamente obra de los estudios de estos dos investigadores. Ya en 1869 el biólogo suizo Miesscher identifica un grupo de sustancias celulares que denominó nucleínas, a las que más tarde Altmann les daría el nombre de ácidos nucleicos. Por su parte Robert Feulgen (308)(311) en 1914 describió un método para revelar por tinción el ADN, basado en el colorante fucsina. La utilización de este método mostró la presencia de ADN en el núcleo de todas las células eucariotas, específicamente en los cromosomas. Durante los años veinte, 224 RESULTADOS Levene (312) analizó los componentes del ADN, encontrando que contenían cuatro bases, el azúcar desoxirribosa y un grupo fosfato. En 1952 Hershey y Chase (313) realizaron una serie de experimentos destinados a dilucidar cuál era material hereditario, marcando ADN y proteínas con isótopos radiactivos en un cultivo de virus indicaron que el ADN es el soporte físico de la herencia. El trabajo de Petrakis va a contribuir al conocimiento de la naturaleza más íntima del linfocito, su carga genética, y ayuda a entender las características básicas del linfocito normal y patológico, dando paso a nuevos campos por explorar como la genética de las neoplasias o las clasificaciones de tumores linfoides, y es una pequeña muestra de las aplicaciones en las que el descubrimiento del ADN va a intervenir, como por ejemplo: 1. Ingeniería genética (314). La investigación sobre el ADN tiene un impacto significativo, especialmente en el ámbito de la medicina, pero también en agricultura y ganadería. 2. Medicina forense. Mediante el estudio del perfil genético, método muy fiable en la identificación de personas víctimas de catástrofes o en pruebas de consanguinidad. 3. Nanotecnología (315). Utilizando el ADN como material estructural más que como portador de información biológica. 4. Historia, antropología y paleontología (316). Mediante el estudio de las secuencias de ADN se puede inferir la evolución histórica de los organismos, su filogenia, siendo útil también para el estudio de especies ya extinguidas (ADN fósil) Sobre el papel de la metionina y el ácido pantoténico en el linfocito, los investigadores Dinning, Neatrour y Day (317), de la Universidad de Arkansas, publican en 1954 sus resultados: “Interrelación del ácido pantoténico y metionina en la producción linfocitaria de la rata”. Existen en la literatura trabajos sobre la importancia del ácido pantoténico y metionina en la nutrición de la rata. Nelson (318) informó en 1947, que el 225 RESULTADOS crecimiento y supervivencia de las ratas con una dieta deficiente en ácido pantoténico mejoraba cuando el contenido de caseína de la dieta aumentaba del 24 al 64 %, y trabajos posteriores constataron que el mismo efecto se podía obtener adicionando metionina a la dieta de un animal deficiente en ácido pantoténico. En esta línea, Ludovici en 1951 (319) señala que la disminución de ácido pantoténico en ratas reduce la formación de anticuerpos en respuesta a un estímulo antigénico, y que ello mejora con la adición de metionina en la dieta. Surge así el interés por el vínculo entre ácido pantoténico y linfocito, ya que éste se considera relacionado con la producción de anticuerpos. Dinning en 1951 y 1952 (320)(321) publica los resultados de sus observaciones sobre la influencia de la dieta y producción de células blancas en la rata, mostrando que ratas con una dieta deficiente en metionina desarrollan leucopenia caracterizada por una disminución de los linfocitos periféricos. Para estudiar el tema más a fondo, los autores emplean ratas de ambos sexos sometidas a una dieta con un contenido de metionina del 0,27 % (dieta base). Las ratas se separan en varios grupos: a) dieta base sin suplemento b) dieta base con suplemento de DL-­‐metionina a dosis de 0,1%, 0,43% y 1,73 %. Estas condiciones se mantienen 55 días, durante los cuales se registran datos sobre el crecimiento, ingesta de alimentos y mortalidad, así como recuentos totales y diferenciales de leucocitos en sangre. Después de 48-­‐52 días se observó que: 1. cuando la dieta no contenía ácido pantoténico, el crecimiento de las ratas respondió al aumento máximo de metionina en la dieta. En contraste, cuando el ácido pantoténico estaba presente en la dieta, el suplemento de 0,43 % de metionina dió lugar a un máximo crecimiento. Parece pues, que en ausencia de ácido pantoténico el requerimiento de metionina para el crecimiento se incrementó. 226 RESULTADOS 2. las muertes ocurridas durante los 55 días de observación tuvieron lugar únicamente en los grupos que recibieron la dieta base y en aquellos con dieta base suplementada con 0,1 % de metionina, lo que indica una interrelación entre ácido pantoténico y metionina. 3. el número de linfocitos periféricos disminuyó notablemente en ratas deficientes en ácido pantoténico y metionina, y en ausencia de ácido pantoténico, era necesario un suplemento de 1,73 % de metionina para mantener los niveles normales de linfocitos en sangre. Los datos, analizados por el método de análisis de la varianza, mostraron un valor significativo para la interacción de ácido pantoténico y metionina sobre linfocitos de sangre periférica (p<0,01), lo que indica que la respuesta del linfocito a uno de estos dos nutrientes estaba condicionada por el otro. Los autores (317) aseguran: “… estos resultados demuestran la interrelación del ácido pantoténico y la metionina”, y sugieren dos posibles explicaciones de base bioquímica: 1. por una disminución de la coenzima A en los tejidos de ratas con déficit de ácido pantoténico, subsanada mediante una cantidad extra de metionina que resulta en una más eficiente conversión del ácido pantoténico en coenzima A. 2. que el ácido pantoténico intervenga en la transmetilación, ya que las necesidades de metionina para la formación de linfocitos están relacionados con la capacidad de metil donación. La vitamina B5 o ácido pantoténico es una vitamina hidrosoluble considerada un nutriente esencial. Fue descubierta por Roger J Williams (322) en 1931 como un cofactor de crecimiento de las levaduras. Es necesaria para la formación de la Coenzima A y se considera un nutriente crítico en el metabolismo y síntesis de carbohidratos, proteínas y grasas. Su nombre deriva del griego pantothen que significa “de todas partes”, pues hay pequeñas 227 RESULTADOS cantidades en casi todos los alimentos. Las primeras publicaciones sobre el ácido pantoténico son debidas a Williams, su descubridor: ! “El efecto del inositol, vitamina B1 y ácido pantoténico sobre el crecimiento de varias cepas de levaduras”, (323). ! “El ácido pantoténico como nutriente para plantas verdes”, (322). ! “La importancia del ácido pantoténico en procesos de fermentación, respiración y almacenamiento de glucógeno”, (324). A mediados de los años cuarenta se advierte un interés progresivamente creciente por el estudio del ácido pantoténico que tiene su máximo histórico en los años 50 y 60. No son muchos los estudios encontrados que relacionan ácido pantoténico y linfocito. Entre ellos: “el efecto de la deficiencia de ácido pantoténico sobre el cortex adrenal, timo, bazo y circulación de linfocitos en el ratón” de 1951 (325) y “el cortex adrenal de la rata deficitaria en ácido pantoténico; respuesta en eosinófilos y linfocitos” de 1952 (326). Sin embargo, el interés por su estudio se mantiene a través de los años como lo demuestran publicaciones más recientes, que ponen de relieve la importancia y trascendencia de los estudios iniciados por Dinning, Neatrour y Day: ! “el ácido pantoténico protege a las células Jurkat contra la apoptosis inducida por luz ultravioleta” de 2001 (327). Se induce la apoptosis de las células Jurkat (linfocitos T leucémicos humanos) mediate luz ultravioleta, pero la preincubación con ácido pantoténico de dichas células resulta en un aumento del contenido celular de glutation y en una protección parcial contra la apoptosis inducida por dicha luz. ! “la baja ingesta de Ca, ácido fólico, nicotínico, vitamina E, retinol, ß-­‐ caroteno y alto consumo de ácido pantoténico, biotina y riboflavina, se asoció significativamente con un aumento de la inestabilidad del genoma” de 2005(328). Estudio que asocia la ingesta en la dieta y el daño genómico en linfocitos usando como marcador la presencia de micronucleolos. 228 RESULTADOS ! “la deficiencia de ácido pantoténico puede aumentar la excreción urinaria de 2-­‐oxo-­‐ácidos y catabolitos de nicotinamida en ratas” de 2013 (329). Estudia la importancia del ácido pantoténico en el metabolismo de aminoácidos y ácidos grasos. Los estudios sobre la producción de anticuerpos mantienen su interés en esta y sucesivas décadas, creciendo lentamente en complejidad y dificultad. En 1955, Coons, Leduc y Connolly publican: “Método para la demostración histoquímica de anticuerpos específicos y su aplicación al estudio del conejo hiperinmune” (330). La introducción de marcadores visibles en el estudio de las reacciones inmunológicas por Heidelberger, Kendall y Soo Hoo (331)(332) permitieron a Marrack (333) la realización de un experimento en el que demostró que una molécula de anticuerpo podía acoplarse a una molécula de colorante sin pérdida de la capacidad para la reacción específica con el antígeno. Posteriormente se observó que compuestos fluorescentes podían conjugarse con moléculas de anticuerpo también sin pérdida de actividad, procedimiento que ha sido aplicado con éxito para el estudio de un gran número de antígenos. Los autores de este estudio (330) utilizan conejos albinos a los que se les inyecta por vía intravenosa γ-­‐globulina humana o bien ovoalbúmina cristalizada de gallina, tres días sucesivos cada semana durante meses, siendo sacrificados al quinto día después de la última inyección. Los órganos de estos animales fueron extraídos y conservados a −20ºC para posterior estudio histológico. Se prepararon soluciones de anticuerpos fluorescentes contra γ-­‐ globulina humana y ovoalbúmina, y los tejidos se observaron bajo el microscopio de fluorescencia. Los controles fueron preparados de igual forma pero con omisión del antígeno. El procedimiento seguido es expuesto con todo detalle, figurando tiempo y volúmenes de las distintas soluciones empleadas. Coons, Leduc y Connolly (330), describen un método en dos fases para la demostración histoquímica de anticuerpos específicos en las células utilizando 229 RESULTADOS secciones congeladas de tejidos. En una primera fase tiene lugar la reacción entre el anticuerpo existente en el tejido y el antígeno aplicado in vitro. En un segundo tiempo se lleva a cabo la detección de aquellas áreas en las que este antígeno ha sido absorbido específicamente mediante una reacción de precipitación, utilizando un anticuerpo marcado con fluoresceína y su examen bajo el microscopio que revela fluorescencia amarilla-­‐verde sobre las zonas donde se ha formado el precipitado. Los autores hacen referencia a lo que consideran tinción no específica y debido a la importancia de esta tinción no específica como factor de confusión, ésta es descrita en detalle. Para la identificación de las células que contienen anticuerpos por tinción convencional, se utilizaron secciones adyacentes (ya que la misma sección no funcionó para dos tinciones), una para el anticuerpo y otra para el estudio histológico mediante Giemsa. Las células que contienen anticuerpos están presentes en grandes grupos en la pulpa roja del bazo, y estos grupos son fáciles de ver también en las secciones adyacentes donde las células se pueden reconocer de forma inequívoca. En el conejo hiperinmune se determinan las células que contienen anticuerpos en varios órganos como bazo y sangre. El estudio revela que el anticuerpo contra la γ-­‐inmunoglobulina humana está presente en los grupos de células plasmáticas de la pulpa roja del bazo, áreas medulares de ganglios linfáticos, submucosa de íleon y tejido conectivo porta del hígado. Pequeñas cantidades de anticuerpos eran ocasionalmente vistos en las células de los folículos linfoides del bazo y ganglios linfáticos, de modo que no podía ser excluida una contribución menor de los linfocitos para la síntesis de anticuerpos. Fagraeus (272) en 1948 encontró que el tejido de pulpa roja esplénica que contienen infiltraciones de células plasmáticas produce anticuerpos in vitro, mientras explantes similares de pulpa blanca producen menos o ningún anticuerpos. Hallazgo que fue confirmado por otros autores como Keuning y van der Slikke (334). Coons, Leduc y Connolly, confirman con este trabajo los resultados obtenidos por Fagraeus, y declaran haciendo referencia a las células plasmáticas: “… el tejido que las contiene y está relativamente libre de otros tipos 230 RESULTADOS celulares, contiene más anticuerpo que tejidos similares sin células plasmáticas, y estos tejidos que las contienen continúan su síntesis de anticuerpos in vitro, mientras tejidos si ellas no lo hacen”. Por tanto, trabajos anteriores ya habían establecido la correlación entre células plasmáticas y síntesis in vitro de anticuerpos (272)(335). La demostración de que estas células individualmente contienen anticuerpos confirman estas observaciones. Esta es la gran aportación del trabajo de Coons, Leduc y Connolly (330), el reconocimiento del contenido de anticuerpos en las células plasmáticas individuales: “Los datos indican que el sitio principal de formación de anticuerpos es una familia de células que aparecen como respuesta al estímulo antigénico, y que esta respuesta consta de multiplicación celular, diferenciación y síntesis de una proteína específica, el anticuerpo. El miembro maduro de esta familia celular es la célula plasmática”. A pesar de los notables avances introducidos en el campo de la investigación linfoide, el destino de los linfocitos aún no se ha definido. Bajo el título “El efecto de la continua re-­‐infusión de linfa y linfocitos en la salida de los linfocitos del conducto torácico en ratas no anestesiadas”, Gowans en 1956 (336) da a conocer su trabajo sobre la cinética de los linfocitos. Muchos han sido los que se han dedicado al estudio del conducto torácico y los linfocitos que circulan por él. Entre ellos tenemos que citar el trabajo de Yoffey (204) en perros o los de Sanders, Florey y Barnes (298) en conejos y gatos, quienes aseguraban que el número de linfocitos que salían del conducto torácico era suficiente para reemplazar a todos los linfocitos de la sangre varias veces al día. Bollman, Cain y Grindlay (337) en 1948 hicieron un notable avance con la descripción de un método para la recogida continua de la linfa del conducto torácico de ratas anestesiadas por períodos de hasta 10 días. Por su parte, Mann e Higgins (338) en 1950 encuentran que cuando el drenaje de linfa se prolonga, la producción diaria de células se reduce progresivamente, de forma que al cuarto día después de la canulación había caído aproximadamente un 30% del valor inicial. Esta caída de la producción era independiente de la tasa de flujo y no era debido a trauma quirúrgico o pérdida 231 RESULTADOS de peso corporal. Mann e Higgins sugirieron que podía ser debido a la pérdida del animal de un pequeño número de células que son esenciales para mantener la producción de linfocitos. Gowans (336) describe en su trabajo un aparato para la re-­‐infusión intravenosa continua de la linfa y linfocitos en ratas no anestesiadas con fístula del conducto torácico. El aparato permite mediciones de la salida de los linfocitos del conducto torácico y reproduce la situación encontrada en el animal intacto con la devolución de toda la linfa a la sangre venosa. El primer objetivo de Gowans fue descubrir si la caída de la producción de linfocitos, descrita por Mann e Higgins, podría prevenirse mediante la devolución de toda la linfa y linfocitos. El segundo objetivo era explicar la caída en la producción de linfocitos que se produce cuando la linfa y linfocitos se pierden en el animal. Se hizo un estudio comparativo entre la salida de células del conducto torácico en las que la linfa y lifocitos vivos fueron reinfundidos y en ratas que recibieron una re-­‐infusión continua de linfa libre de células o con linfocitos muertos; con ello Gowans esperaba aportar una prueba de que la caída de células observada por Mann e Higgins (338) podría ser debida a la pérdida en el animal de células esenciales para el mantenimiento de la producción de linfocitos. En su experimento empleó ratas machos albinas, y el conducto torácico se canuló con un tubo de poliestileno (de 0,5 mm de diámetro interno) justo por debajo del diafragma. Este procedimiento drena la linfa desde hígado, intestino, músculos y piel de debajo del diafragma, pero no de los vasos linfáticos de extremidades, cabeza y cuello. El aparato mostrado en la Figura 29 fue diseñado para la re-­‐infusión vía intravenosa de toda la linfa aproximadamente a la misma velocidad a la que salía el fluido del conducto torácico. 232 RESULTADOS Figura 29 Aparato de reinfusión continua intravenosa de linfa y linfocitos. A y B: jeringas impulsadas eléctricamente. C: depósito de vidrio siliconado. D: bomba de rodillo. E: cánula del conducto torácico. F: cánula de retorno de linfa a vena femoral. G: conexión entre el circuito del relé de control y electrodos. H,I,J,K,L: diversos componentes electrónicos. Tomado de: Gowans 1956 (336). Se hicieron pruebas para verificar la viabilidad de los linfocitos, utilizando como único criterio satisfactorio de viabilidad la motilidad de las células in vitro. Se comparó el comportamiento in vitro de las células obtenidas a la salida de la bomba con las células que salían directamente del conducto torácico, para determinar si las células habían sido dañadas a su paso a través 233 RESULTADOS de la bomba. Las células vivas eran reconocidas por sus movimientos ameboides y los linfocitos muertos por su forma redondeada permanente y la formación de vesículas citoplasmáticas, siempre preludio de muerte celular. Aproximadamente el 80% de los linfocitos estaban vivos y móviles después de 24 horas de incubación y el 20% después de 48 horas. Ninguno sobrevivió al sexto día. De esta observación se dedujo que las células no fueron dañadas por el paso a través de la bomba y estaban vivas cuando entraron en la rata. Para comparar resultados también se realizó una re-­‐infusión de linfa con linfocitos muertos y una re-­‐infusión de linfa libre de células. En el primer caso los linfocitos eran irradiados con luz ultravioleta o bien calentados en un baño de agua entre 45ºC y 56ºC. En el segundo caso, la linfa es centrifugada y el sobrenadante libre de células se añade al depósito y es devuelto por la bomba a la rata. Gowans (336) obtiene los siguientes resultados: 1. Encuentra que la salida de linfocitos del conducto torácico era independiente de la tasa de flujo de la linfa y del peso corporal de la rata, y calculó que el número total de linfocitos en sangre debería ser sustituido unas once veces al día. El efecto del drenaje de linfa durante 7 días sobre la salida de linfocitos del conducto torácico produce una fuerte caída durante el tercero o cuarto día y se estabiliza entorno al quinto día en un 20-­‐30% del valor inicial, manteniéndose constante entre el día 7-­‐14 después de la canulación. Como la mayor caída se había producido al final del cuarto día fue en este momento cuando se estudió el efecto de la re-­‐infusión intravenosa. 2. Observa que la salida de linfocitos del conducto torácico cuando el animal es reinfundido permanece constante, y se compara con ratas que no recibieron la re-­‐infusión. La caída en la salida de linfocitos no fue debida a pérdida de líquido linfático, ya que la continua re-­‐infusión de linfa libre de células no lo impidió. Se pensó que esta pérdida celular a partir de una fístula del conducto torácico podría privar al animal de 234 RESULTADOS materiales esenciales para la síntesis de nuevos linfocitos. Si los linfocitos muertos suministraban sustancias que son reutilizadas en la formación de nuevas células, podría esperarse un mantenimiento en los linfocitos que salen del conducto torácico, sin embargo, no se evitó la caída en la producción de linfocitos. La salida media de 768 x 106 linfocitos/día desde el conducto torácico de ratas no anestesiadas coincide con las cifras publicadas por Mann e Higgins en 1950. El trabajo de Gowans confirma el hallazgo por estos autores de una profunda caída en la producción de linfocitos cuando el drenaje de la linfa se mantienen más de dos días, y Gowans encuentra que esta caída puede evitarse por la re-­‐ infusión intravenosa continua de linfa con linfocitos vivos: “El recuento de linfocitos en sangre y la salida del conducto torácico se aumenta o se mantienen sin cambios durante los cuatro días que duró el experimento. Parece que la entrada continua de linfocitos vivos en sangre es esencial para el mantenimiento de la salida de los linfocitos del conducto torácico” La interpretación más atractiva que realiza Gowans (336) respecto a la explicación de los resultados de sus experimentos es que la entrada de los linfocitos vivos en sangre es esencial para mantener la salida de linfocitos del conducto torácico: “Por ello hay que considerar que los linfocitos re-­‐infundidos ejercen su efecto dejando la sangre y retornando a los ganglios linfáticos”. Y apunta algunas posibilidades como que los linfocitos podrían morir en los ganglios linfáticos y liberar sustancias necesarias para las nuevas células, o bien que un número considerable de linfocitos podrían salir de la sangre y finalmente volver a entrar en el conducto torácico a través de ganglios linfáticos o de los espacios tisulares. Una tercera posibilidad sería que los ganglios linfáticos pueden volver a repoblarse a partir de la sangre por una pequeña proporción de linfocitos capaces de división celular, según Gowans: “el conducto torácico de ratas contiene aproximadamente un 5% de linfocitos grandes y medianos que puede observarse como se dividen in vitro, y posiblemente estas células son las que Mann e Higgins sugieren como responsables de mantener la producción de linfocitos”. Y reconoce las 235 RESULTADOS limitaciones existentes: “de momento se sabe muy poco sobre el ciclo de vida de los linfocitos para poder decantarnos por alguna de estas posibilidades”. Igual que el dilema sobre el destino de los linfocitos, el timo es otro de los temas pendientes presente en todas las décadas. “El origen tímico del factor plasmático estimulante de linfocitosis” es una de las publicaciones de Metcalf en 1956 (339). Metcalf había demostrado que el plasma de varias enfermedades era capaz de inducir una linfocitosis cuando se inoculaba en ratones recién nacidos (RN). Las enfermedades en las que se detectó esta actividad plasmática fueron: leucemia linfática crónica, linfosarcoma y mielofibrosis. La actividad plasmática estaba deprimida durante las remisiones naturales de la enfermedad y después de transfusiones de sangre entera, y aumentaba tras inyecciones intramusculares de adrenalina. El efecto de la linfocitosis en ratones inoculados se inhibía por el precalentamiento del plasma a 60ºC o por la administración concurrente de cortisona o estrógenos. También se había encontrado que las emulsiones de timo humano poseían una actividad estimulante de la linfocitosis cuando se inyectaban a ratones recién nacidos, y esta sustancia tímica estaba estrechamente relacionada con el factor estimulante de la linfocitosis que se encuentra en el plasma de las enfermedades mencionadas. Para el estudio Metcalf obtuvo muestras tisulares de las autopsias de personas muertas por causas accidentales, por leucemia linfática crónica y por linfosarcoma, preparando emulsiones a partir de las muestras de órganos utilizando sílice estéril, y a partir de material obtenido de ratones normales y leucémicos. Las emulsiones realizadas se probaron para ver su actividad estimulante de linfocitosis, y se calculó el contenido del órgano en la sustancia estimulante de linfocitosis (LSS), realizando diluciones seriadas de la emulsión y determinando la dilución más alta que es capaz de producir linfocitosis. Se determinó así la actividad estimulante de linfocitosis de diversos tejidos humanos y de ratón, mostrando que de todos los órganos probados sólo el 236 RESULTADOS timo y tiroides mostraron una actividad estimulante de linfocitosis, siendo la actividad de la glándula tiroides menor en grado. Las emulsiones de otros órganos no alteraban el cuadro sanguíneo periférico, con la excepción de las emulsiones de la glándula suprarrenal que produjeron linfopenia. El estudio de timos normales de personas de distintas edades, reveló que el tejido del timo en todas las edades posee una actividad estimulante de la linfocitosis para los ratones, y que el contenido en el timo de la sustancia estimulante se redujo con la edad. Metcalf (339) reconoce que: “fuera de toda duda, el timo humano normal contienen una sustancia (LSS) capaz de producir una linfocitosis en la cría de ratón, y que el contenido del timo en LSS disminuye con la edad”. Y cuando estudia el contenido en el timo de la leucemia linfática crónica, tanto en el hombre como en los ratones, Metcalf observa que: “existe un mayor contenido tímico de LSS que en el timo normal”. Figura 30 Contenido tímico total de sustancia estimulante de linfocitosis (LSS) expresadas logaritmicamente en unidades de ratón. Cada columna representa un ratón. Tomado de Metcalf.1956, (339). 237 RESULTADOS Metcalf también estudió la producción de LSS en cultivos de fragmentos de timo, utilizando timo de ratón normal; pretendía determinar si el tejido del timo en realidad produce éste LSS o simplemente actuaba como órgano de almacenamiento del mismo. Tras el cultivo del tejido tímico se aprecia un aumento definitivo en el contenido total de LSS, y supone pues que es resultado de una producción real de LSS por los fragmentos tisulares. Realiza un intento para identificar el tipo celular productor de LSS, ya que el timo contiene dos tipos principales de células: redondas y pequeñas con las características morfológicas de linfocitos y células epiteliales más grandes. Además contiene una red de células reticulares consideradas poco probables como fuente de LSS. Metcalf, para ello emulsiona fragmentos de tejido tímico cortical (rico en linfocitos) y de tejido medular (rico en células epiteliales), para determinar su LSS, y resalta la observación siguiente: “mientras que los fragmentos de médula fueron muy activos, los corticales no mostraron ninguna actividad, lo que indica que son las células epiteliales del timo la fuente de LSS, pero no se excluye que las células corticales puedan desempeñar algún rol asociado a la producción de LSS”. El trabajo de Metcalf (339) demuestra, según sus propias palabras: “que el timo produce una sustancia estimuladora de la linfocitosis, tanto en animales normales como leucémicos”, y pone de manifiesto la presencia de un LSS tímico y un factor plasmático (LSF), que presentan unas características comunes: “ambos, LSF plasmático y LSS tímico producen una linfocitosis en ratones inoculados, mientras que al mismo tiempo inhiben la actividad funcional del timo en la producción de LSS. Este LSS es idéntico que el factor estimulante encontrado en el plasma de ciertas enfermedades, particularmente leucemia linfática crónica y linfosarcoma” Los linfocitos son considerados células no fagocíticas por naturaleza, sin embargo se ha comunicado su capacidad para ingerir hemosiderina (341), lo que crea la necesidad de realizar nuevos experimentos para determinar este punto. Con el título “Estudios sobre la actividad fagocítica de linfocitos: la fagocitosis de tinta china administrada de forma intravenosa en humanos”, 238 RESULTADOS Koszewski, Emerick y Dicus (340) publican en 1957 los resultados de su trabajo. Las teorías existentes a estas alturas del siglo XX, atribuyen la actividad fagocítica de la sangre a monocitos y leucocitos polimorfonucleares. Los linfocitos son considerados células sin capacidad fagocítica. Cuestión que los autores tratan de comprobar mediante la administración de tinta china inyectada vía intravenosa. Se seleccionaron ocho pacientes con enfermedades crónicas intratables, los cuales recibieron diariamente una inyección intravenosa de tinta china a dosis de 10-­‐20 cc. Antes del inicio de la inyección, se examinó la sangre de cada paciente a intervalos regulares por un periodo de dos semanas para obtener los valores de base en el número de leucocitos, eritrocitos, hemoglobina, hematocrito y recuento diferencial de leucocitos. Una muestra de sangre fue teñida con 1% de safranina para mostrar la presencia de partículas de carbón en las células blancas de la sangre, tinción que permite distinguir fácilmente linfocitos, monocitos y células segmentadas. No se observaron cambios cuantitativos esenciales en los valores sanguíneos de pacientes que recibieron las inyecciones de tinta china. El único cambio notorio en la morfología de los leucocitos fue la presencia de partículas en el citoplasma de los leucocitos. En todos los ocho casos se mostró ingestión de partículas de carbón en los leucocitos circulantes. Los neutrófilos fueron las células más eficaces en el proceso de fagocitosis, y en todos los casos el porcentaje de polimorfonucleares que tomó tinta china fue superior al de linfocitos, y además contenían más inclusiones en el citoplasma que otras células. 239 RESULTADOS Figura 31 Porcentaje de neutrófilos, linfocitos y monocitos que fagocitan tinta china en el caso nº 3. Tomado de Koszewski, Emerick y Dicus, 1957, (340). La tasa de aparición de linfocitos fagocitantes varía con el individuo y también varía de día a día en el mismo individuo. Ante estos hechos, los autores aseguran: “Las inclusiones ocurren con tal regularidad que podemos hablar de actividad fagocítica de los linfocitos”. No obstante, el papel del linfocito en el cuerpo sigue siendo desconocido, aunque constituye el 20-­‐30 % de las células blancas circulantes. Su función es contemplada como un mecanismo principal en la inflamación crónica. Bergel (342) ha señalado repetidamente que la función del linfocito está relacionada con la digestión de las grasas, y que la inyección intraperitoneal o intrapleural de grasa da lugar a una marcada exudación mononuclear. Sin embargo, un estudio detallado con tinción supravital mostró que tal exudado está formado por macrófagos y monocitos. Ehrich (343) les asignó una función enzimática, ya que al ser ricos en adenosina podrían destruir productos tóxicos. El hecho de que los linfocitos participan en mecanismos de defensa ha sido demostrado por muchos autores, entre ellos Metalnikov y Toumanoff (344), Ehrich y Harris 240 RESULTADOS (345) ó Dougherty y White, que creían estaban involucrados en la producción o transporte de anticuerpos más que en la función de fagocitosis. La teoría de la fagocitosis fue mencionada por Metchnikoff (346)(31), quien divide a las células en micrófagos (leucocitos polimorfonucleares) y macrófagos (grandes mononucleares de distintos orígenes). La cuestión de si los linfocitos son o no fagocitos se complica por la sospecha de que estas células pueden transformarse en otras como monocitos, poliblastos, histiocitos libres o células reticulares. Maximow (347) asegura que el linfocito es capaz de incorporar colorantes después de haber sufrido una trasformación a poliblasto. Tampoco nos ayuda la experimentación en animales a decidir si el linfocito, como tal, o su trasformación, exhiben actividad fagocítica. Ingestión de partículas por linfocitos ha sido observada por Downey (348) en la sangre de conejos y por Stilwell (349) en la sangre de ranas después del uso de tinta china. Koszewski (341) observó inclusiones de hemosiderina en los linfocitos de pacientes con hemocromatosis, y el mismo fenómeno se observa en animales y humanos que reciben suspensiones de óxido de hierro parenteral. El trabajo realizado por Koszewski, Emerick y Dicus (340), utilizando tinta china en humanos, proporciona más datos de que los linfocitos son capaces de desarrollar una actividad fagocítica. El polimorfonuclear es la célula fagocítica más activa, pero muchos linfocitos también ingieren partículas de carbón en su citoplasma. El alto porcentaje de linfocitos fagocíticos habla en contra de una vía pasiva de incorporación de partículas de carbón y la cantidad de tinta china inyectada diariamente fue insuficiente para provocar un fenómeno de inclusiones por contacto, por que debe incorporarse por mecanismo activo de fagocitosis. La opinión de los autores es la siguiente: “La fagocitosis debe ser añadida a la limitada lista de posibles funciones del linfocito en el cuerpo humano”. La utilización del microscopio electrónico comienza a ser algo habitual en los centros de investigación, haciendo posible el estudio de la célula a nivel ultraestructural. Nowell y Berwick (350), en 1958 dan a conocer el resultado 241 RESULTADOS de su trabajo bajo el título: “La ultraestructura de la superficie de linfocitos normales y leucémicos en la rata”. Existe un creciente interés por conocer la superficie externa de células cancerosas, ejemplo de ello es el trabajo de Coman y colaboradores (351) que establece el hecho de que la adhesividad intercelular de las células malignas es menor que la de sus prototipos normales, y que esta diferencia probablemente juegue un papel importante en la capacidad invasiva de las células cancerosas. Ya que la adhesividad es un fenómeno de superficie, parecía razonable la existencia de alguna diferencia morfológica en la estructura de esta superficie que pueda ser demostrable a nivel ultramicroscópico. Es posible que estos cambios estén directamente relacionados con la disminución de la adhesividad de células malignas, aunque también pueden ser independientes de la adhesividad y reflejar algunas otras características de las células neoplásicas. En base a ello, Nowell y Berwick (350) deciden estudiar linfocitos normales y leucémicos, ya que la adhesividad de ambas células es baja, pueden separarse fácilmente de sus compañeras y estudiarse como células individuales. Comparan la superficie ultraestructural de linfocitos normales y leucémicos tanto de sangre como de ganglios linfáticos de ratas Wistar, que se convierten en leucémicas mediante trasplante de leucemia linfocítica. Los animales trasplantados con leucemia murieron en 14 días, presentando lesiones en hígado, bazo, timo y ganglios linfáticos del mediastino, y el estudio de las secciones de los ganglios mediastínicos revelaron sábanas de linfocitos pequeños y grandes, intercalados con un número variable de células reticulares con grandes núcleos vesiculares. Se hicieron secciones de todas las muestras para ser teñidas con Wright y con hematoxilina–eosina, para su estudio comparativo con el microscopio electrónico y el microscopio de luz. Al microscopio electrónico, los linfocitos, tanto grandes como pequeños, normales y neoplásicos, procedentes de sangre y ganglios linfáticos, mostraron una superficie fina de similar estructura, consistente en un par de desigualmente distribuidas partículas de 100-­‐300 Angstrom (Å) de diámetro. 242 RESULTADOS Además había células de gran tamaño que incluían células linfoides inmaduras, células reticulares, macrófagos y células plasmáticas. Muchas de ellas tenían una ultraestructura de superficie similar a la encontrada en los pequeños linfocitos, sin embargo un 10-­‐20% de estas grandes células mostraban características bien diferentes en su superficie, que los autores describen de la siguiente forma: “Eran células grandes, planas, normalmente de forma oval, y en su superficie aparecía un número variable de pequeños cráteres de un diámetro de 800-­‐1000 Å con un número variable de gránulos de 100-­‐200 Å, situados alrededor de una depresión central”. Estas grandes células, presumiblemente células reticulares, con cráteres en su superficie fueron observadas en todas las preparaciones excepto en aquellas procedentes de la sangre circulante de ratas normales, lo que podría indicar que estas células cráter presentan una adhesividad mayor que las otras células de los nodos. La diferencia observada en la ultraestructura de la superficie sugiere que un cambio en el estado neoplásico no es necesariamente acompañado por alteraciones en la superficie celular, al menos en linfocitos de rata. Figura 32. Célula cráter. Parte de una célula cráter de un ganglio linfático normal. La célula es ovoide y plana, y muestra una superficie salpicada de cráteres, consistentes en varias partículas de 100-200 A (flecha) alrededor de una depresión central. Tomado de Nowell y Berwick, 1958, (350). 243 RESULTADOS Cuando se examina sangre normal, pequeños y grandes linfocitos, así como eritrocitos tienen la misma configuración y ultraestructura de superficie que las células observadas en el ganglio linfático. Grandes células, presumiblemente granulocitos muestran una superficie gruesa nodular y ultraestructuralmente similar a las observad en el linfocito, pero únicamente se detectó una célula cráter en las numerosas muestras de sangre de rata normal. De todas las células examinadas, solo las células reticulares mostraron una superficie distinta, y estas células también mostraron mayor adhesividad, lo que sugiere que la ultraestructura de la superficie celular linfocitaria puede estar directamente relacionada con la adhesividad intercelular. En su estudio, Nowell y Berwick (350) no encontraron diferencias sustanciales entre la ultraestructura de la superficie linfocitaria de ratas normales y leucémicas, lo que podría indicar: “… que el estado de malignidad puede existir sin alteraciones concomitantes en la superficie ultraestructural celular”. De otro lado, añaden que los cambios estructurales de la superficie pueden reflejar diferencias específicas en la adhesividad intercelular: “En nuestras preparaciones únicamente células con diferencias ultraestructurales, las llamadas células cráter, fueron mucho más difíciles de aislar de ganglios linfáticos que los otros elementos celulares, sugiriendo que la adhesividad de las células cráter es mayor”. Y consideran sumamente interesante la identidad de estas células cráter: “No se encontraron en preparaciones de sangre circulante de animales normales y no son granulocitos ni monocitos. Aparecen en un número significativo en preparaciones de timo, y no se identifican con células plasmáticas ni con macrófagos”. Y añaden: “La evidencia indica que las células cráter, con su especial ultraestructura de superficie son células reticulares y su adhesividad intercelular es grande”. El microscopio electrónico utiliza electrones en lugar de fotones o luz visible, permitiendo alcanzar ampliaciones hasta 5000 veces mayores que los mejores 244 RESULTADOS microscopios ópticos, debido a que la longitud de onda de electrones es mucho menor que la de los fotones visibles. El primer microscopio electrónico fue diseñado por Ernst Ruska y Max Knoll entre 1925 y 1930, patentándose en Alemania en 1932. Los primeros modelos experimentales eran muy imperfectos, y sólo después de la Segunda Guerra Mundial se superaron sus problemas técnicos y se consiguieron cortes de tejido idóneos para ser utilizados con la nueva tecnología. Existen dos tipos principales, el de transmisión, que fue el primero diseñado, y el de barrido, de mejor calidad visual y prestaciones, desarrollado unos años más tarde, en 1942(171). La utilización del microscopio electrónico como medio de estudio e investigación es ya un hecho en la década anterior, la quinta década. La primera comunicación encontrada se remonta a 1939 y lleva por título “La localización de minerales en tejidos animales por el microscopio electrónico” (352). Tímidamente en los años posteriores va a ir aumentando el número de publicaciones y los temas objeto de estudio. Como ejemplos tenemos: ! “La contribución del microscopio electrónico a la medicina” en 1940 (353). ! Los estudios sobre morfología bacteriana; leptospiras, fusobacterias y bacteriófagos son descritos con gran detalle (354). ! “Estudios de microscopía electrónica de las células sanguíneas en órganos hematopoyéticos y exudados inflamatorios del hombre” de 1947 (355). ! “Estructura y desarrollo de virus observados al microscopio electrónico”(356). ! Estudios sobre la corteza cerebral o sobre el molusco contagioso (357)(358). Como podemos apreciar el microscopio electrónico se convierte en un elemento sumamente útil en medicina, biología, genética, microbiología y anatomía e imprescindible en el campo de la investigación. Pero será en la 245 RESULTADOS década siguiente cuando la utilización del microscopio electrónico tenga su mayor apogeo, ampliando su campo a otras disciplinas como la cristalografía o la ingeniería: “La textura de la superficie de los granos de arena. Una aplicación del microscopio electrónico: Glaciación” (359), “Autorradiografía de alta resolución. Métodos” (360) o “Investigación por el microscopio electrónico de la caseína de la leche en polvo” (361), son algunos ejemplos de su aplicación en campos diversos. En la actualidad su utilidad sigue patente, como lo demuestra el número de publicaciones y la extensión de su uso a los más diversos temas, como en cosmética: “Estudio sobre los efectos de acondicionamiento de champús naturales utilizando el microscopio de barrido” (362) o en aplicaciones ópticas: “Microestructura láser y microscopía de barrido de las capas compuestas por plasmapolymer-­‐plata” (363). Hecha esta breve exposición sobre el trayecto del microscopio electrónico en el curso de la historia, sólo nos queda encuadrar el trabajo de Nowell y Berwick, que publicado en 1958, se convierte en unos de los pocos estudios que tratan sobre la ultraestructura del linfocito, dando un gran paso en el campo científico al señalar la importancia de la adhesividad celular en la trasformación celular maligna. También Shelton y Dalton (364) en 1959, del Instituto Nacional de la Salud en Bethesda, optan por la utilización del microscopio electrónico y publican un breve artículo que lleva por título “Microscopía electrónica de la emperipolesis”. En 1958, Shelton (365)(366) describe la tendencia a la emperipolesis que exhiben las células del líquido ascítico del ratón con un tumor linfocítico. La pregunta que surge cuando se observan una o varias células dentro de otra célula, versa sobre las relaciones existentes entre la célula envolvente y la que ha sido engullida, si existe un contacto íntimo entre ellas o si se relacionan a través de la existencia de membranas que separan una célula de otra. Estas preguntas son respondidas en parte por las micrografías que acompañan a esta nota (Figura 33). 246 RESULTADOS El citoplasma claramente granular de los macrófagos puede ser fácilmente distinguido del citoplasma claro de las células tumorales contenidas en él, pero sin embargo, la naturaleza de la frontera existente entre los dos citoplasmas celulares no puede ser determinada por microscopía de fase. Cuando se mira con el microscopio electrónico, se ve que en su mayor parte los linfocitos malignos se separan del citoplasma de los macrófagos por dos membranas, una perteneciente al linfocito y otra al macrófago (Figura 33) y mientras la membrana de los linfocitos está intacta, hay zonas donde la membrana interna de los macrófagos parece entrar en contacto con la membrana del linfocito. Aunque la naturaleza de la membrana plasmática de las células no se conoce bien, no hay razón para pensar que la membrana interior de macrófagos sea distinta de la exterior. Existe datos que apoyan (367) la creencia de que es el linfocito el que provoca esta situación. Se ha observado a los linfocitos dentro de células hepáticas, huevos de anfibios, cultivos de fibroblastos, epitelio de riñón, intestino, tráquea y tiroides. Por tanto, el macrófago no es sino una de las muchas células en las que lo podemos encontrar. Shelton y Dalton (364) acaban su escrito con una pregunta sumamente interesante y de gran trascendencia para el futuro: “¿Es el linfocito una célula con una inmunidad tan peculiar, que le permite estar fuera y dentro de otras células, y dividirse dentro de ellas?” Y sentencian: “el linfocito sea quizás la célula peor comprendida de todas las células metazoarias”. 247 RESULTADOS Figura 33. Emperipolesis 2- Fina sección de un macrófago que contiene cinco células tumorales. Destaca la continuidad de la membrana externa del macrófago. La célula tumoral en el extremo derecho tiene la característica “cola” de linfocito en movimiento. 3- Detalle de la unión de cuatro de los linfocitos. Nótese lo que parece ser el citoplasma del macrófago en los espacios existentes entre las células tumorales. 4- Detalle de la membrana del macrófago y su relación con la membrana plasmática de la célula tumoral. Tomado de: Shelton y Dalton, 1959 (364). El estudio del hallazgo de “célula dentro de célula” comenzó con el deseo de comprender este complejo fenómeno y derivó en la observación de lo que Lewis denominó un proceso no fagocítico en la alimentación de células, en 248 RESULTADOS 1925 (368). El término emperipolesis fue acuñado por Humble, Jayne y Pulvertaft en 1956 (369) para describir el fenómeno de la presencia de linfocitos dentro de una célula, y ha sido observado en una gran variedad de condiciones, como en la enfermedad de Rosai-­‐Dorfman, hepatitis autoinmune y en megacariocitos que contienen neutrófilos segmentados. En sentido amplio, emperipolesis es la presencia de una célula intacta dentro del citoplasma de otra célula, y se considera que es una penetración activa de una célula en otra. Se ha pretendido vincular el fenómeno con la respuesta del sistema inmune a las células tumorales, pero en realidad no existe ninguna explicación convincente. La emperipolesis es una característica común en los linfomas malignos tipo Hodgkin y no Hodgkin. Su presencia en la enfermedad de Rosai-­‐ Dorfman (370) es característica de la enfermedad, pero se observa en muchas condiciones fisiológicas y patológicas, como lo atestiguan un importante número de publicaciones registradas recientemente: “la emperipolesis megacariocítica como hallazgo histológico en el síndrome mielodisplásico” (371), “ fagocitosis celular (canibalismo) en tumores humanos: actualización y revisión de la literatura” (372) o “la emperipolesis en la púrpura trombocitopénica inmune” (373), son algunas de ellas. En la búsqueda de los mecanismos subyacentes, Wang y Li (368) habían revelado que la emperipolesis es un medio para mediar la muerte de una célula tumoral por células asesinas naturales. Ellos demostraron la desintegración de células tumorales a través de una vía de degradación lisosomal después de la emperipolesis de las células asesinas naturales a nivel ultraestructural. Sin embargo también descubrieron que algunas células asesinas naturales fueron desintegradas por células tumorales a través de un mecanismo similar. El estudio ultraestructural ha permitido diferenciar la emperipolesis de otros fenómenos aparentemente semejantes como la entosis, considerada la invasión de una célula viva en el citoplasma de otra. El proceso fue descubierto por Overholtzer (374), y se ha observado afectando a más de dos células. Una célula atrapada por entosis es inicialmente viva y se puede dividir en el interior de la célula que la ha envuelto, pero son finalmente asesinadas. Este nuevo 249 RESULTADOS concepto de entosis abrirá la polémica sobre el significado real de emperipolesis y canibalismo celular, ya que la línea que los divide es muy tenue. Muchos científicos actualmente piensan que la entosis puede aportar beneficios al organismo, y que al ser un tipo de muerte celular no apoptótica, puede ser de ayuda en la lucha contra el cáncer y tumores mediante la oportuna modificación de estas células entóticas (372) (375). Procedente de la Universidad de Oxford, J.L Gowans (376) publica en 1959 uno de los estudios más interesantes de esta década: “La recirculación de los linfocitos de la sangre a los ganglios en la rata”. En todas las especies de mamíferos hasta ahora examinadas, un gran número de linfocitos entran en la sangre todos los días desde los principales vasos linfáticos. En la rata, por ejemplo, entran en la sangre suficientes linfocitos desde el conducto torácico para reemplazar a todos los que salen de ella y esto ocurre alrededor de once veces al día (336), estimación mínima de la sustitución real diaria, ya que no considera a todos los vasos linfáticos que drenan cabeza, cuello, extremidades y tórax, y que han sido tenido en cuenta por otros autores como Whaler y Widdicombe (377), quienes consideran que el tiempo medio de permanencia de los linfocitos en la sangre de la rata es menor de una hora. Muchas teorías han sido propuestas para explicar esta rápida rotación de los linfocitos en sangre, pero ninguna de ellas con un buen soporte experimental (378) (379). En un trabajo anterior, Gowans (336) demostró que con el fin de mantener la salida de linfocitos constante a partir de una fístula del conducto torácico en la rata sin anestesia, era necesario volver a infundir continuamente en la sangre todos los linfocitos que salían de la fístula. Cuando la linfa que se reinfundía no tenía linfocitos o contenía linfocitos muertos, tenía lugar una profunda caída en la salida de los linfocitos del conducto torácico, de forma similar a las observaciones descritas por Mann e Higgins (338) en ratas que no recibieron reemplazo intravenoso de células. 250 RESULTADOS Gowans (376) intenta ahora, en este trabajo, llegar a conocer los mecanismos por los que la infusión intravenosa de linfocitos vivos influye en la salida de los linfocitos del conducto torácico. Esencialmente lo que pretende es determinar el destino de los linfocitos del conducto torácico después de su transfusión a sangre. El método utilizado por Gowans en su trabajo anterior se extiende a éste, de tal forma, que los linfocitos del conducto torácico de un número de ratas donantes pueden ser transfundidos continuamente, durante largos períodos de tiempo, en la sangre de la rata receptora. Un requisito esencial es que los linfocitos transfundidos deben sobrevivir en el animal receptor, y para que dicha transfusión de linfocitos sea un homoinjerto, las ratas donantes y receptoras deben ser genéticamente idénticas. Aunque la idoneidad genética de las ratas no se comprobó, los resultados de los experimentos dejaron claro que los linfocitos transfundidos no habían sido afectados por la intervención de una reacción al injerto. La característica esencial del método es que permite que linfocitos y linfa sean inyectados en la sangre de la rata a un ritmo fisiológico durante muchos días. El aparato utilizado fue el mismo que Gowans utilizó en 1957, el conteo celular se realizó cada 2-­‐3 horas en la linfa de la rata donante, y cada muestra con un número conocido de linfocitos se añadió al depósito para la re-­‐infusión. La linfa y linfocitos del depósito se bombearon continuamente en la vena femoral de la rata receptora a una velocidad de 2-­‐4 ml/hora, siendo las ratas transfundidas con éxito por períodos de hasta siete días. Los linfocitos fueron etiquetados con P32 e inyectados vía subcutánea en las ratas donantes. También se realizó el etiquetado de linfocitos con timidina tritiada, método ideal para el estudio de la síntesis de ADN (ácido desoxirribonucleico) en las células individuales de mamíferos, ya que la timidina se incorpora exclusivamente al ADN, y en las autorradiografía, el tritio se localiza estrictamente sobre el núcleo de cada célula. Gowans efectúa una serie de transfusiones a diferentes intervalos, valora diferentes parámetros y realiza una serie de observaciones: 251 RESULTADOS ! 1-­‐Transfusión de linfocitos en el quinto día. Se llevó a cabo una transfusión intravenosa procedente de ratas donantes, que se inicia en el cuarto día después de la canulación en la rata destinataria, recogiéndose toda la linfa del conducto torácico de la rata receptora para recuento celular. Cuando la linfa era de compañeras de camada de la misma cepa endogénica, la salida de células de la rata receptora se elevó gradualmente y se estabilizó un punto por debajo de la salida inicial, pero dentro de valores normales. Por el contrario la transfusión de ratas encapuchadas a ratas albinas dio lugar a un aumento más lento en la salida que terminó bruscamente en el cuarto día, y a pesar de la continua transfusión de linfocitos, la producción disminuyó progresivamente a un nivel por debajo del inicio de la transfusión. Gowans demuestra así, que la caída en la salida de linfocitos descrito por Mann e Higgins en 1950 puede ser revertido por la transfusión de linfocitos de una rata a otra, siempre que la disparidad genética entre donante y receptor no sea demasiado grande. (Figura 34). ! Transfusión de linfocitos en el segundo día después de la canulación del conducto torácico. Se observó que la transfusión vigorosa de linfocitos junto con la re-­‐infusión continua de todos los linfocitos del propio conducto torácico del destinatario, aumenta la salida a un valor más del doble que el valor del primer día. ! Relación entre la entrada y salida de linfocitos. Se estudió en ratas cuya producción de linfocitos había caído a un nivel bajo después del drenaje de la linfa del conducto torácico (CT) durante cinco días. La salida de linfocitos de tales ratas presentó una línea de base adecuada para estudios cuantitativos: un número conocido de linfocitos se transfundía y se medía la salida de linfocitos del CT en muestras de linfa sucesivas durante cuatro horas. Se observó un aumento continuo en la salida de linfocitos durante las primeras cuatro horas después del inicio de la transfusión y una caída durante las cuatro horas siguientes al final de la transfusión. La producción total de linfocitos por encima de la estimada 252 RESULTADOS como línea de base asciende aproximadamente al 80% del número de células transfundidas. Figura 34. Valores medios de linfocitos/hora. La producción de linfocitos del conducto torácico de tres ratas albinas. Valores medios por hora para cada sucesiva colección de linfa en 24 horas. -Las ratas recibieron transfusiones de linfocitos de ratas albinas: ■--■; de ratas encapuchadas: ○--○; una rata no recibió ninguna transfusión: ▲--▲ -1,2, etc = períodos de transfusión a partir de sucesivas ratas donantes. Tomado de Gowans 1959, (376). ! Mecanismo por el que la entrada de linfocitos en sangre aumenta la salida en el conducto torácico. Según experimentos, la salida de linfocitos del CT varía directamente con en número de linfocitos transfundidos en la sangre. Se mencionan tres posibles mecanismos: que sustancias químicas derivadas de los linfocitos puedan ser 253 RESULTADOS esenciales para la producción de nuevos linfocitos en el animal receptor, o bien que las células transfundidas puedan repoblar los ganglios linfáticos de receptor y por sucesivas divisiones celulares dar lugar a nuevas poblaciones de linfocitos. La tercera posibilidad sería que los linfocitos transfundidos puedan recircular de nuevo de la sangre al conducto torácico, lo que implica que las células transfundidas y las células adicionales emergentes desde el CT son idénticas. ! Formación de nuevos linfocitos. Se puso a prueba mediante la infusión de timidina marcada con tritio en la vena femoral de una rata, mientras que los linfocitos se transfundieron simultáneamente en la otra vena. La realización de autorradiografía permitió evaluar la situación. Si el extra de linfocitos que sale del CT se había formado después del comienzo de la transfusión, su ADN estaría marcado con tritio. Se observó que solo el 2% de los pequeños linfocitos estaban marcados y en cambio lo estaban el más del 90% de los grandes linfocitos, lo que llevó a la conclusión de que casi todos los pequeños linfocitos de la rata eran células viejas en el sentido de que su ADN había sido formado antes del inicio de la transfusión. ! Recirculación de linfocitos. Los experimentos realizados sugieren que los linfocitos que fueron transfundidos en la sangre encontraron su camino de regreso a la linfa del CT. Esta hipótesis fue comprobada directamente por transfusión de linfocitos marcados con 32P en una rata, y siguiendo la aparición de radiactividad en los linfocitos que salen del CT, se observó un continuo aumento de la radiactividad en las células de comienzo en las primeras cuatro horas. ! Proporción de linfocitos recién formados normalmente presentes en la linfa del CT. La idea de que los linfocitos recirculan continuamente desde la sangre a la linfa proporcionaría una explicación satisfactoria a la aparente alta rotación de linfocitos en la sangre del animal normal, y esta explicación sería más convincente si se pudiera demostrar que el número de linfocitos de nueva formación cada día es solo una pequeña fracción del número total que normalmente sale del CT. Con este fin, se 254 RESULTADOS infundió timidina marcada con tritio a una rata en las primeras doce horas siguientes a la canulación de su CT, y se recogió la linfa en las sucesivas 24 horas durante cuatro días. La rata no recibió ninguna transfusión o re-­‐infusión de linfa. Las autorradiografía mostraron que durante los primeros 2 días, más del 90% de los grandes y medianos linfocitos, y solo aproximadamente 1% de los pequeños contenían ADN marcado. En su discusión Gowans (376) analiza todos estos resultados, y dice: “Se ha demostrado bajo una variedad de condiciones que la transfusión de linfocitos en sangre de una rata resulta en un aumento de salida de linfocitos en su CT. Este aumento se observa de 1 a 5 días después de la transfusión, y ha sido posible estimar que el aumento total en la salida asciende a aproximadamente el 80% del número de células transfundidas”. Prácticamente todos los pequeños linfocitos que aparecían en el CT contenían ADN sintetizado antes de la transfusión, por lo que Gowans asegura que: “El aumento de la producción no fue el resultado que un proceso que implique formación de nuevas células por mitosis, lo que sugiere fuertemente que las células transfundidas habían recirculado desde la sangre a la linfa”. Estos hallazgos experimentales sugieren que en el animal intacto, la gran mayoría de linfocitos pequeños del CT son células que han recirculado desde la sangre a la linfa y que este circuito se recorre en varias ocasiones por los linfocitos durante su tiempo de vida. La rápida rotación fisiológica sería más aparente que real, ya que las mismas células estarían continuamente desapareciendo de la sangre y apareciendo en ella. La existencia de una recirculación de linfocitos en el animal normal es fuertemente apoyada por la demostración de que la tasa de formación de nuevos linfocitos no sigue el ritmo de la velocidad a la que salen los linfocitos del CT. Los experimentos con timidina tritiada mostraron que una proporción muy pequeña de células en el CT fueron células recién formadas y que no existe un almacenamiento de pequeños linfocitos en el ganglio linfático. 255 RESULTADOS “La demostración de que los pequeños linfocitos recirculan deja el problema de su destino final sin resolver”, comenta el autor. Y, añade: “… si tienen una larga vida y un número relativamente pequeño muere cada día o son eliminados por cualquier otro proceso, entonces es razonable suponer que puedan tener un final convencional en las células del sistema reticuloendotelial, ya que tampoco hay evidencia convincente de que exista un especial mecanismo para eliminar los pequeños linfocitos decadentes”. Aunque Gowans, sumamente crítico con los resultados de su trabajo, considera no resuelto el problema del destino final de los linfocitos, la realidad es que da un paso gigantesco en este camino, y posiblemente no fuera plenamente consciente de la gran importancia de su descubrimiento. Solo tenemos que mirar hacia atrás y revisar los numerosos trabajos, los numerosos autores y las variadas teorías que intentan dar solución al problema del destino final de linfocitos. La cuestión del destino final de los linfocitos es abordada ya en 1921 por Bunting y Huston, en un intento de dar respuesta a preguntas tales como ¿A dónde van los linfocitos en su viaje por el torrente sanguíneo?, ¿se desintegran dentro de los vasos sanguíneos? Ellos consideraron que deberían abandonar el torrente sanguíneo y destruirse fuera de él. Yoffey es otro de los autores que dedican gran parte de su tiempo a resolver el enigma del destino final de los linfocitos (años 1933, 1935 y 1936), apuntando la posibilidad de que los linfocitos que salen del conducto torácico no sean células recién formadas, que pueda existir una continua recirculación de linfocitos desde la sangre a ganglios linfáticos y viceversa. En 1940, en: “El misterio de la desaparición de los linfocitos”, se sugieren tres alternativas explicativas, y se apunta: “Tal vez la solución final del destino último de los linfocitos, se obtendrá cuando una técnica pueda marcarlos a fin de ser reconocidos donde quiera que vayan”. A nuestro juicio, en realidad Gowans no solo contribuye a aclarar el destino final de los pequeños linfocitos, sino que identifica los linfocitos inmaduros como las células precursoras: “El destino de los grandes y medianos linfocitos, 256 RESULTADOS que normalmente comprenden menos del 5% de las células de la rata en la linfa del conducto torácico, no se puede deducir a partir del presente estudio… son conocidas por ser capaces de dividirse, y en los experimentos con timidina tritiada casi todas ellas fueron marcadas… el tamaño y la capacidad para dividirse son las únicas características que distinguen grandes y medianos de los pequeños linfocitos” Consideramos que con la exposición aquí realizada queda patente la gran importancia del trabajo de Gowans, su significado en la historia y su gran contribución a la ciencia. Con el título “La conversión de linfocitos con núcleos atípicos en células plasmáticas tipo Marschalkó”, el autor E.H Tompkins (380) nos desvela en 1960 sus observaciones al respecto. Tompkins (381) en trabajos de años anteriores, ya había comunicado la presencia de linfocitos pequeños con núcleos atípicos en secciones de tejido linfoide de varias especies animales. En el presente informe, el autor hace referencia a la transformación de estos linfocitos en células tipo Marschalkó: “Es una transformación que parece iniciarse dentro del núcleo “atípico” de los linfocitos antes de que los cambios citoplasmáticos sean detectables”. Los estudios se llevan a cabo con los tejidos de ratas machos cepa C3H de diferentes edades. Las tinciones empleadas son principalmente hematoxilina-­‐ eosina, pironina y verde metilo. Tompkins observa que: 1-­‐ Las características histoquímicas que caracterizan los núcleos de los linfocitos atípicos, también caracterizan a los núcleos de las formas tempranas del desarrollo de las células Marschalkó. El núcleo de la célula plasmática en su fase incipiente es similar al de linfocitos atípicos excepto por una mayor formación de grumos. Las características específicas de los núcleos de los linfocitos atípicos disminuyen a medida que se transforman en células plasmáticas. 257 RESULTADOS 2-­‐ Un material birrefrigente y una matriz nuclear de características peculiares se extiende desde el núcleo atípico al citoplasma durante el proceso de transformación. En la metamorfosis de los linfocitos con núcleo atípico en células plasmáticas tipo Marschalkó no se observa una división completa de estos linfocitos. Tompkins describe de esta forma el inicio de esta transformación (380): “El borde del citoplasma característico de los pequeños linfocitos se agranda, aumenta en densidad óptica y toma una tonalidad de color paja en las preparaciones no teñidas llegando a ser profundamente basófilo así como pirinofílico en preparaciones teñidas, haciéndose difícil separar núcleo de citoplasma ya que la profunda basofília es difusa. Cuando se llega a esta etapa es evidente que el desarrollo de las células plasmáticas tipo Marschalkó está en marcha”. 3-­‐ La intensidad de la birrefrigencia y la tinción nuclear disminuyen concomitantemente con estos cambios. La cantidad, densidad e intensidad de la basofilia y pirinofilia del citoplasma de las células plasmáticas en desarrollo, aumentan de forma paralela con estas alteraciones. Estas características aparecen primero en los límites núcleo-­‐citoplasma, se extienden periféricamente y disminuyen a medida que el núcleo se desintegra. Las zonas periféricas del citoplasma son las últimas en ser completamente abasofílicas Tiene lugar una ordenada continuidad de paso de material del núcleo al citoplasma, y pérdida de material desde el citoplasma al medio ambiente. El citoplasma se torna vacuolado, espumoso o irregular en los bordes en la última etapa, cuando el núcleo ha llegado a parecerse más al de un macrófago y el citoplasma ha perdido la mayor parte de su basofilia. Que las células plasmáticas aparecen por metamorfosis de otras células no es un hallazgo nuevo, existiendo argumentos a favor y en contra sobre la célula origen, postulándose el linfocito (382), las células reticulares (179)(272) y linfoblastos como posibles células precursoras. El propósito del trabajo de Tompkins no es sin embargo determinar la célula precursora, solamente se 258 RESULTADOS limita a describir los cambios morfológicos que ocurren en el proceso de transformación a partir de los linfocitos atípicos. Es sorprendente que Marschalkó concediera una gran importancia al núcleo de la células plasmáticas, considerándolo como un requisito para la diferenciación de las plasmáticas, y que Tompkins por estos estudios también considere la gran importancia que juega el núcleo, no solo importa la estructura del núcleo, sino que el núcleo parece ser el lugar de origen donde tienen lugar el estímulo que inicia el desarrollo de la célula plasmática. El trabajo de Tompkins (380) indica que el estímulo comienza dentro de los núcleos de los pequeños linfocitos y se extiende en una segunda etapa al citoplasma y del citoplasma al medio ambiente. El autor considera que la sugerencia de Dougherty (383) de que la reacción antígeno-­‐anticuerpo en los linfocitos inicia la producción de células plasmáticas debe ser considerada. Acabamos esta década constatando un gran interés por la célula plasmática. Esta célula ha sido uno de los principales objetivos de estudio en muchos de los artículos publicados en estos años, con un interés renovado desde que fue descrita a finales del siglo XIX, gracias a los trabajos de Fagraeus quien en 1948 describe en su tesis doctoral la producción de anticuerpos por dichas células plasmáticas. En 1877, Paul Ehrlich demuestra una metacromasia acentuada en unas células que llamó mastocitos tisulares. Posiblemente estas células correspondían a células plasmáticas, pero al no poderse asegurar existen muchas controversias al respecto (384). Dos años antes, Waldeyer, alumno de Virchow, había descrito células redondas con abundante citoplasma que llamó Plasmazellen para diferenciarlas de Flügzellen o células de citoplasma escaso. Es posible que estas Pasmazellen correspondiesen a lo que hoy conocemos como células plasmáticas, pero también existe polémica acerca de esto. Unna en 1891, utilizando el método del azul policromo, identificó unas células con basofilia citoplasmática, sin embargo, la basofilia es una propiedad no exclusiva de estas 259 RESULTADOS células, por lo que no existe seguridad de que Unna hubiera observado verdaderas células plasmáticas (385). Santiago Ramón y Cajal identificó las células plasmáticas en un estudio de condilomas sifilíticos, a las que llamó células embrionarias, células sifilíticas o corpúsculos cianófilos. Cajal las describe de esta forma: “… esférica, ovoide o ligeramente poliédrica con aristas redondeadas. Núcleo único, esférico… posee una red cromática floja, grosera… el núcleo es siempre excéntrico y hasta tangencial. El citoplasma es intensamente basófilo y la zona no teñida perinuclear contiene el aparato de Golgi” Cajal encontró dichas células en tejidos normales y patológicos y sugirió que podían ser linfocitos transformados (384)(386). A pesar de ser esta la primera descripción de célula plasmática, el nombre que quedó registrado no fue el de célula cianófila de Cajal, sino como lo había designado Waldeyer, célula plasmática. Cinco años después de la descripción de Cajal, Marschalkó detalló las características de las células plasmáticas, con su núcleo redondo-­‐oval, y su cromatina agrupada en 5-­‐ 8 grumos periféricos e hizo notar también que el citoplasma se acumulaba en la periferia de la célula dando lugar a la formación de un halo perinuclear típico ocupado por el aparato de Golgi (385). Por tanto, al parecer, la descripción de la célula plasmática madura podría ser original de Cajal, por lo que la célula plasmática diferenciada bien podría llamarse tipo Cajal-­‐Marschalkó. 260 RESULTADOS 4.6. LA SÉPTIMA DÉCADA (LOS AÑOS SESENTA) Los años sesenta se van a caracterizar por el inicio de cambios sociales y culturales que tenderán a expandirse hasta convertirse en nuevos modos de vida. Es el inicio de los movimientos ecologistas, los movimientos feministas contemporáneos, los movimientos en pro de la tolerancia y la legitimidad del otro, de la aceptación de las identidades y diversidades culturales, sexuales y étnicas (220) (219). En Occidente crece la sensibilidad ecológica. La “Primavera Silenciosa” de Rachel Carson (387) publicado en 1962, es el primer libro divulgativo sobre impacto ambiental, convertido en un clásico de la concienciación ecológica. Fue dignificada la diversidad cultural, gracias entre otros, a Martín Luther King y Malcolm X, y el nacimiento de movimientos antinucleares reactivan el pacifismo. En 1960 nace la anticoncepción hormonal oral, la conocida “píldora”, considerada con una perspectiva histórica, la creación cultural por excelencia que permitió una respuesta real a la amenaza de la sobrepoblación. En Estados Unidos nacería el más importante de los movimientos juveniles de estos años: los Hippies, herederos de la precedente generación Beat de los años cincuenta. Contra el consumo y el autoritarismo, proclamaban un regreso a la naturaleza, rechazaban el trabajo embrutecedor defendiendo un trabajo creativo y autónomo, siendo uno de los grupos más activos contra la guerra de Vietnam en los años sesenta (388). El 21 de julio de 1969 el hombre pisaba la Luna y, frente al televisor, la humanidad tomaba conciencia de un sentimiento de pertenencia a la Tierra que nunca antes había existido. En el mundo de la ciencia hay cambios importantes. Se consolida la cibernética y la teoría de sistemas, surge la física no lineal y las matemáticas de la complejidad. El estudio y la síntesis del ADN será clave en el conocimiento del origen de la vida, dando paso a la era de la genómica (389). Se realizan los primeros ensayos y experimentos de interconexión de computadoras en red 261 RESULTADOS orientadas a la defensa militar, pre-­‐configurando la actual red Internet, y el cielo terrestre empezaría a llenarse de “estrellas” artificiales: los satélites. Poco a poco aparecerán nuevos campos de interés y formarán parte de nosotros las nuevas tecnologías, lo que influirá sin duda en el número de estudios y en la forma de abordar los trabajos relacionados con el linfocito. La búsqueda en Scopus con la palabra lymphocyt* da como resultado un total de 12.334 documentos de 1961 a 1970, el 2,20% del total de publicaciones registradas sobre el linfocito en el siglo XX, un porcentaje muy superior al de la década precedente (0,0574%). El mayor número de trabajos proceden del Instituto Nacional del Cáncer seguido del Instituto Karolinska, siendo los países con mayor número de publicaciones Estados Unidos, Alemania y Reino Unido. GRÁFICO 5 La gráfica aquí presentada corresponde al número de documentos por años desde 1961 a 1970. Es una gráfica claramente ascendente, observándose como aparece una gran pendiente a partir de 1966. Así si en 1961 el número de documentos representa el 1,4% del total en esta década, en 1970 ascienden al 262 RESULTADOS 25,4%. La mayoría de publicaciones se encuadran en el área de medicina (76,2%), seguido por el bloque de bioquímica/genética/biología molecular (16,7%) e inmunología/microbiología (10%), y las revistas donde más se publica son Nature, Lancet, Blood, Transplantation y Journal of Immunology. Cuando realizamos un análisis más detallado de estudios por áreas, observamos que a mitad de década el interés por los estudios de bioquímica/genética/biología molecular se hace evidente. Así en 1964, el 19,3 % pertenecen a esta área de estudio. Esta tendencia se mantiene hasta finalizar la década. Los trabajos de Gowans sobre la recirculación de los linfocitos despiertan un gran interés y el deseo de continuar en esta línea de trabajo. En 1961, con el título: “Estudio sobre el destino de los linfocitos. Etiquetado de los pequeños linfocitos con timidina tritiada”, RG Murray y A Murray publican sus observaciones (390). Los autores consideran que pese a las extensas e intensas investigaciones, realizadas por Trowell (379)(367) y Gowans (376), la función de los linfocitos, particularmente de los pequeños linfocitos, sigue siendo un enigma. Un enfoque para solucionar este problema ha sido el etiquetaje de los linfocitos con diversas sustancias para poder seguir su distribución y comportamiento cuando se inyectan a un animal. Entre las diversas sustancias empleadas se encuentran fósforo radiactivo, cromo radiactivo, oro radiactivo y los colorantes fluorescentes, pero ninguno de estos métodos proporcionaba una localización celular precisa ni tenían la estabilidad necesaria que proporciona el marcaje con timidina tritiada considerado el método de elección. Este estudio (390) recoge una serie de ensayos a distintos tiempos y con diferentes dosis de timidina tritiada para el marcaje de pequeños linfocitos. Como animal de experimentación se utilizaron ratas Wistar que fueron inyectadas por vía intraperitoneal con timidina marcada. Después de varias inyecciones, fueron sacrificadas y sus órganos como el timo y el bazo fueron 263 RESULTADOS estudiados mediante improntas y suspensiones celulares. Según el marcado nuclear, los linfocitos se dividieron en pequeños linfocitos y todos los demás linfocitos. Al menos 400 células fueron contadas en cada determinación, y solo las células con la presencia de dos o más granulaciones fueron consideradas positivas. En las autorradiografía realizadas a los frotis de suspensiones de linfocitos, se observó que no había linfocitos pequeños marcados a los treinta minutos, lo que concuerda con los informes de otros autores. El etiquetado de medianos y grandes linfocitos varió entre el 60% y 80%, y dado que éstos constituían solo el 10-­‐20% de todos los linfocitos, el porcentaje de células marcadas no difiere sensiblemente de la de pequeños linfocitos marcados. Cuando las mismas ratas se estudian a mayor intervalo de tiempo, se observa un aumento del número de linfocitos marcados en sangre. El número de pequeños linfocitos marcados aumentó linealmente con el tiempo hasta aproximadamente 1 ½ día y solamente un poco a partir de entonces. En la rata se dice que el timo es una fuente importante de suministro de linfocitos a sangre (391)(392) lo que sugiere que muchos pequeños linfocitos recién formados permanecen durante algún tiempo en el timo. Las autorradiografía de improntas confirmaron en general los valores obtenidos en los frotis de las suspensiones, pero debido a las muchas variables adicionales a tener en cuenta poco valor se puede dar a estas estimaciones. En las autorradiografía de las secciones de los tejidos, la distribución de las células marcadas se estudió en timo, bazo, médula ósea, ganglio linfático y duodeno, después de la inyección única o repetida de H3 timidina. La corteza de timo presentó un aumento de células marcadas, pero ninguna con un carácter fuerte, no considerándose muy fiable. Los ganglios linfáticos tenían centros germinales llenos de células ligeramente marcadas al igual que la corteza del timo, mientras que la zona de alrededor del centro activo estaba repleta de linfocitos pequeños con un modesto número de ellos ligeramente marcados. Los centros germinales de la pulpa blanca del bazo tenían la misma apariencia 264 RESULTADOS que la de ganglios linfáticos. La médula ósea mostró un patrón semejante a la pulpa roja esplénica y el duodeno, con células fuertemente marcadas en las criptas y menos fuerte en las células que emigran a las vellosidades. Respecto a la viabilidad de las células inyectadas, era importante establecer si eran inmediatamente engullidas por células fagocíticas como células muertas o dañadas o si sobrevivían en condiciones normales. A los 15 minutos un gran número de pequeños linfocitos marcados se encontraron en pulmón, en buen estado, en la pared alveolar y no ingeridos por macrófagos, y a los 60 minutos habían desaparecido casi por completo. Unos pocos linfocitos marcados estaban en médula ósea, hígado, y ganglios mesentéricos en un intervalo de 15 minutos a 48 horas, la mayoría en buen estado y solo en raras ocasiones ingeridos por macrófagos. Figura 35. Autorradiografía de una suspensión de células del timo de rata. El tiempo total después de la primera inyección hasta el sacrificio fue de 37 horas. L= gran linfocito; N= medianos linfocitos; S= pequeños linfocitos; ?= célula presumiblemente rota parcialmente. Tomado de: Murray, 1961, (390) 265 RESULTADOS En la discusión, los autores cuestionan el origen de linfocitos y timocitos: “la elección del timo como fuente de linfocitos plantea la cuestión de la identidad de los linfocitos y timocitos… no hay evidencias indiscutibles de su diferencia”, afirmando que no se pueden separar por criterios morfológicos. Sin embargo, Diderholm y Fichtelius (393) señalan diferencias basadas en el diferente trato dado por el hospedador a la inyección de células del timo y células de los ganglios linfáticos. Pero consideran que estas diferencias no solucionarían el problema: “...si el timocito es una célula distinta al linfocito, quedará una cuestión igual de desconcertante, la desaparición y función del gran número de timocitos que se producen cada día”. Los autores (390) aclaran que la clasificación de las células linfoides en pequeñas, medianas y grandes, como se hace en este informe, es necesariamente arbitraria, ya que no existen límites claros entre ambas categorías, solamente las células menores de 7µ de diámetro y borde citoplasmático muy estrecho fueron incluidas como pequeños linfocitos. Consideran, que quizás el parámetro más valioso para medir la presencia o ausencia de síntesis de ADN marcado sea el tiempo, y añaden: “la ausencia de pequeños linfocitos marcados a los 30 minutos confirma la creencia de que estas células rara vez, tal vez nunca, se someten a mitosis, sino que son producto de la maduración de células algo mayores”. Siguiendo la línea de investigación referente a producción de anticuerpos, linfocitos y células plasmáticas es el trabajo de de E.H. Cooper (394): “Producción de linfocitos y células plasmáticas en la rata después de la inmunización con albúmina de suero humano” publicado en la revista Immunology del año 1961. Desde los trabajos realizados por los investigadores escandinavos Bjorneboe, Gormsen y Lundquist en 1947 (395) y por Fagraeus en 1948 (272), se han ido acumulando pruebas que evidencian una estrecha relación entre células plasmáticas y la producción de las proteínas de los anticuerpos. Por otro lado, 266 RESULTADOS las células plasmáticas no son el único tipo de célula presente en tejidos linfoides que participa en las reacciones a antígenos. En este trabajo, Cooper (394) analiza los cambios que ocurren en la proliferación celular y el metabolismo inducidos por la inmunización de ratones con albúmina de suero humano. Se tienen en cuenta tres perspectivas: • Estudio de la fístula que drena el conducto torácico en ratas normales e inmunizadas, que aporta información sobre el metabolismo y producción de linfocitos per se. • Estimación de la absorción de 32P en el ADN y ARN, para el estudio del patrón metabólico global de los ácidos nucleicos en diverso órganos in vivo. • Observaciones sobre el metabolismo del ADN de las células plasmáticas maduras en condiciones normales y en ratas inmunizadas, para la obtención de datos que ayuden a la interpretación de los cambios en la actividad específica de ácidos nucleicos en tejidos linfoides. Se emplearon ratas machos de la cepa Wistar para investigar los efectos de la inmunización con albúmina de suero humano y ratas hembras de la misma cepa para la producción de células plasmáticas normales. Como antígeno se empleó albúmina de suero humano (HSA) obtenida de los hemoderivados de varios laboratorios como el Instituto Lister. Cada rata fue inmunizada con 0,5 mgr de HSA por vía intramuscular en ambos muslos y posteriormente con precipitado de alumbre de HSA a intervalos de tres días cada uno con 0,5, 10 y 2 mgr respectivamente. La estimación de anticuerpos precipitables en suero se realizó por el método de Kabat y Mayer (396)(397). Tanto las ratas de control como las inmunizadas fueron inyectadas intramuscularmente 32P, siendo sacrificadas noventa minutos después, para medir la concentración de anticuerpos en suero. La medición de la radiactividad de 32P se realizó con un medidor tipo Geiger-­‐Müller, expresando la absorción de 32P en el ADNP y ARNP (fosfato de ADN y fosfato de ARN) como porcentaje relativo de las actividades específicas. El conducto torácico (CT) fue canulado y la linfa se recogió de ratas 267 RESULTADOS conscientes durante 24 horas, momento en que son sacrificadas. El número de linfocitos que sintetizan ADN activamente se detectó mediante la incubación de los linfocitos recién extraídos de los ganglios linfáticos que contenían timidina tritiada. También se realizan observaciones en células plasmáticas in vivo e in vitro. En los experimentos realizados con ratas vivas, se les inyectan tres dosis de timidina tritiada a intervalos de cuatro horas para ser posteriormente sacrificadas a distintos intervalos de tiempo en los siguientes catorce días, haciendo preparaciones a partir de los ganglios linfáticos para su estudio autorradiográfico. Para los estudios in vitro células de ganglios linfáticos y bazo se prepararon por incubación de los tejidos picados en medio de Hanks (398) con un contenido del 50% de suero de rata. En los resultados (394) se determina la salida media en 24 horas de los linfocitos de la fístula del CT en ratas control y ratas inmunes, observando que ambos valores se encuentran en el mismo rango, y que la proporción de grandes linfocitos es la misma en ambos grupos. También se observó por el estudio de las autorradiografias que el número de linfocitos que sintetizan ADN era similar en ambos grupos. No se observaron células plasmáticas en ninguna de las muestras de linfa de los dos grupos. Por otra parte, la inyección de HSA en las ratas canuladas no causa ningún cambio en la composición de las células de la linfa, tanto en ratas normales como hiperinmunes. En los recuentos de sangre periférica se pudo constatar un leve aumento de polimorfonucleares en las ratas inmunes, pero el número de linfocitos se mantuvo al mismo nivel en los dos grupos de ratas. En el estudio histológico realizado a ratas inmunes, tanto bazo y ganglios linfáticos mostraron una marcada proliferación de células plasmáticas, y el patrón folicular de ganglios linfáticos mostró una apariencia normal. Estos cambios apreciados en el tejido linfoide recuerdan a los descritos con anterioridad por Marshall y White (399) en conejos como reacción a antígenos particulados. 268 RESULTADOS Se investigó el metabolismo del ADN en células plasmáticas mediante la capacidad para incorporar timidina tritiada in vivo e in vitro. Las células plasmáticas maduras tienen una apariencia histológica bien definida que no fue distorsionada por la preparación de los frotis o improntas, pero si hubo una pérdida del detalle estructural de los diversos precursores de las células plasmáticas, que impidieron hacer un análisis de estas células blásticas. En los experimentos realizados in vitro de ganglios linfáticos y médula ósea no se observó marcaje en células plasmáticas o pequeños linfocitos. En vivo se analizaron los efectos de un corto período de exposición a timidina tritiada en las improntas de ganglios linfáticos, no observándose células plasmáticas maduras marcadas. El porcentaje medio de blastos marcados fue de 30-­‐36%. Dado que existe evidencia que la célula plasmática es una célula evolutiva final, se hizo un análisis autorradiográfico del porcentaje de células plasmáticas maduras marcadas en la población celular de ganglios linfáticos después de marcar células precursoras durante ocho horas. El resultado fue una evidente diferencia en el etiquetado de los pequeños linfocitos y las células plasmáticas. Un alto porcentaje de plasmáticas contenía timidina marcada a las 30 horas, y este porcentaje va disminuyendo en los doce días siguientes. Por el contrario, la población de pequeños linfocitos mostró un aumento más lento pero sostenido de células marcadas en un periodo de 14 días. El número de células en la linfa del conducto torácico que sintetizan ADN fue la misma en los dos grupos. La actividad específica de ADNP y ARNP en bazo y de ARNP en ganglio linfático fue significativamente mayor en ratas inmunizadas. Una considerable proliferación de células plasmáticas ocurrió en los ganglios linfáticos y bazo de ratas inmunizadas. El autor (394) realiza una extensa y detallada discusión sobre el tema, señalando algunos puntos de interés: “… no hubo ninguna alteración significativa en el total de linfocitos circulantes y la salida/hora del CT canulado, tanto en ratas inmunes como en ratas control”. La salida de células del CT es un indicador sensible de la producción y 269 RESULTADOS circulación de linfocitos, así pequeñas dosis de radiación causan una rápida y profunda caída de los linfocitos que salen de la fístula del CT, y las drogas radiomiméticas tienen el mismo efecto (400). Es posible por tanto, que el control homeostático del nivel de linfocitos circulantes ajuste la dinámica de la producción celular y compense cualquier disminución en los precursores de los pequeños linfocitos. En contra de esta hipótesis está el hecho de que no se produjo ningún cambio en el número de grandes linfocitos en los ganglios de ratas inmunes y que el ciclo de división celular de estos grandes linfocitos era similar a los controles. “Si las células plasmáticas son células maduras finales pueden también ser consideradas como una parte de la masa inerte de ADN del tejido”. Está bien establecido que la síntesis de ADN en células que se dividen no es un proceso continuo, ya que solo se producen durante un período determinado en el ciclo celular, según las observaciones de Howard y Pelc (401) y Lajtha, Oliver y Ellis (402). Los pequeños linfocitos son células finales y no se dividen in vitro (336), por lo que el ADN de estas células forma parte de la masa de ADN inerte. La actividad específica de ARNP es un parámetro más variable que el de ADNP. La relación exacta entre la producción de ARN y síntesis de proteínas en las células de mamíferos es aún desconocida, aunque existe una buena correlación entre basofília, contenido de ARN y síntesis de proteínas como lo asegura Brachet (403) uno de los estudiosos del tema. En el bazo y en menor medida en los ganglios de ratas inmunes, se produjo un aumento en la actividad específica ARNP y esto fue acompañado de un aumento en la población de células plasmáticas. Sin embargo, la presencia de células plasmáticas no está directamente relacionada con la renovación media del ARN del tejido. Esto sugiere que puede haber células plasmáticas de actividad metabólica variable y con las mismas características morfológicas. El alto porcentaje de células plasmáticas marcadas encontradas en las autorradiografias de los nodos a las 30 horas después de la inyección de timidina marcada, junto con la evidencia de que la célula plasmática es una 270 RESULTADOS célula final, indica que estas células marcadas deben haberse originado a partir de células precursoras durante el período de 48 horas después de la inyección del isótopo. Lo que indica que el mantenimiento de la población de células plasmáticas en un órgano requiere una entrada constante de nuevas células plasmáticas; por el contrario, los linfocitos con un larga vida, mostraron un lento aumento su porcentaje durante un período de 14 días. Por todo ello, es poco probable que las células plasmáticas deriven de los pequeños linfocitos, a menos que la diferenciación de pequeños linfocitos en células plasmáticas tenga lugar muy rápidamente después de la formación de los pequeños linfocitos a partir de sus precursores. Cooper consciente de las diversas dificultades existentes, considera la siguiente hipótesis como posible interpretación de los datos obtenidos: “la inmunización de ratas va acompañada sin duda de cambios en la población celular de los tejidos linfoides, que se caracteriza por un aumento de células plasmáticas. Puesto que estas células plasmáticas son células finales y su esperanza de vida intranodal parece ser corta, se deduce que para mantener la población ganglionar de estas células debe haber una producción continua de estas células por sus células precursoras”. Por lo tanto, el control de la división celular en los ganglios linfáticos y bazo de la rata inmune está influenciado por la producción de plasmáticas y existiendo evidencia de que la producción de linfocitos circulantes está dentro de los límites normales, nos indica que la proliferación de células plasmáticas es independiente de la producción de linfocitos. Ya en 1947 encontramos una publicación de la aplicación de la autorradiografía al estudio celular: “Autorradiografias de células aisladas” de Bayley (404) La radioactividad tiene la capacidad de impresionar una placa fotográfica, fenómeno descubierto por Becquerel en 1896 que utiliza la autorradiografía, y que nos va permitir seguir el destino de un compuesto radiactivo en el interior de un organismo y cuantificar el mismo, ya que la intensidad de la impresión de la placa fotográfica es proporcional a la cantidad de radioactividad presente en la muestra. Las aplicaciones de esta técnica son 271 RESULTADOS numerosas, encontrándose entre ellas el seguimiento de moléculas marcadas en el interior de una célula, como lo han hecho nuestros dos anteriores autores, Murray y Cooper, que utilizan moléculas biológicas a las que sustituyen un elemento por otro radiactivo, o bien incorporan un elemento radiactivo que las células o los tejidos emplean. No podemos seguir adelante sin mencionar a Chales Philippe Leblond (405), uno de los personajes más notables que contribuyó a configurar esta página de la historia. Leblond fue pionero en la técnica de la autorradiografía y descubridor de las células madres adultas. Nacido en Francia en 1910, en 1937 se incorporó al laboratorio de Síntesis Atómica de París, donde trabajó en la preparación de isótopos radiactivos para la identificación de moléculas en procesos biológicos. Descubridor de que el yodo se acumula en la glándula tiroides (406)(407), se interesó por una nueva técnica con la que se podían obtener imágenes de rayos X de moléculas marcadas con isótopos radiactivos. En 1941, en Montreal, usando yodo-­‐131 consiguió mejorar dicha técnica y localizar los folículos tiroideos. Después de la Segunda Guerra Mundial, en 1946, volvió a Montreal donde se dedicó a mejorar la resolución de las técnicas de autorradiografias. El uso de secciones delgadas junto a la incorporación de las capas de emulsión dio lugar a nuevos avances en la resolución, y la introducción de tritio fue un hito en la técnica. La autorradiografia de alta resolución desarrollada por Leblond se sigue usando hoy día para localizar genes y secuencias de ADN y ARN. Algunas de sus publicaciones son: ! “Localización histológica de elementos radiactivos”, de 1947 (408). ! “La timidina H3 como herramienta para la investigación de la renovación en las poblaciones celulares”, de 1959 (409). ! “Patrón de renovación de stem cells en tres epitelios: esófago, intestino y testículo”, de 1967 (410). Leblond pudo demostrar que la mayor parte de células y tejidos adultos están en continua renovación y calculó con gran precisión el ciclo de vida de muchos tipos celulares. Descubrió la presencia de células madre en el epitelio situado 272 RESULTADOS en la base de los tubos seminíferos del testículo, y también en tejidos que normalmente no se dividen como el esquelético, mediante la utilización de timidina tritiada, llegando a la conclusión de que en el cuerpo existen tres tipos de poblaciones celulares: " poblaciones estáticas, si poseen células que no se dividen " poblaciones expansivas, que contienen un pequeño porcentaje de células madres adultas y " poblaciones renovables, con predominio de de células madres. Resultados que han sentado las bases no solo de la investigación moderna de células madres, sino también de la biología celular moderna (410)(411). La médula ósea rica en células de diferente morfología es uno de los órganos a estudiar que más interés despierta y más dificultades plantea. Bajo el título “Células semejantes a linfocitos en la médula ósea de la rata” Charles Harris publica en 1961 sus observaciones al respecto (412). Existen dos períodos en la vida temprana de las ratas albinas cuando la población celular de la médula ósea (MO) sufre grandes cambios de manera espontánea. En una primera fase, que tiene lugar de 7-­‐28 días de edad, la población celular semejante a linfocitos varía del 10% a los 7 días a 45% a los 14 días, para pasar de nuevo al 10% en 28 días. Un observador podría interpretar esta linfocitosis medular en la rata de 14 días como una leucemia linfática. En una segunda fase, que tienen lugar entre la 6ª y 10ª semana de edad, el número de granulocitos se duplica. La observación de numerosas formas intermedias entre las formas semejantes al pequeño linfocito de la MO (en ratas de 28 días de edad) y normoblastos, pequeños granulocitos (en ratas de 6 a 8 semanas de edad), formas blásticas y células plasmáticas, lleva a Harris (en una publicación anterior) a considerar la existencia de una célula multipotencial y a representar la trasformación morfológica entre las células semejantes a los linfocitos y las otras formas (413). 273 RESULTADOS El estudio de Harris (412) hace referencia a la identificación de la célula linfocito-­‐like (semejante a linfocito) de la médula ósea de la rata y los factores que pueden influir en sus bruscas fluctuaciones numéricas. Se utilizaron ratas Wistar y se siguió el método de conteo utilizado y descrito en anteriores publicaciones (413)(414). Los fémures de los animales fueron abiertos, con un pincel se tomaron muestran de la MO y con él se pintaron tiras en el portaobjeto. Con este método, la hemodilución es insignificante y bajo el microscopio, la MO aparece como un pannus celular donde la distribución grasa y grado de celularidad están preservados. Todas las células, por lo general de 70-­‐100 son contadas en un campo, facilitando así las estimaciones del total y los recuentos diferenciales por unidad de área. El experimento se amplia con la utilización de cortisona que deprime los linfocitos-­‐like de la MO. Las ratas fueron sacrificadas a diferentes intervalos de tiempo, se realizó autopsia y estudio de MO. La cortisona provoca un retraso de aproximadamente una semana en el patrón esperado de linfocitosis de la MO, de modo que el aumento de las células linfocito-­‐like un 40% o más se produce a los 21 días de edad en lugar de a los 14 días, y a partir de este momento, la MO de ratas tratadas con cortisona y las no tratadas es indistinguible. En el estudio histológico de la MO se advierte una hipoplasia celular y un aumento de la cantidad de grasa en la MO de las ratas tratadas con cortisona. Se observa que el aumento o disminución en el número de células por unidad de área en la MO de controles y animales tratados, se debe principalmente a las fluctuaciones en el número de células semejantes a linfocitos y que la concentración de normoblastos se mantiene bastante constante. Los estudios microscópicos del timo revelaron un reemplazo de la arquitectura tímica normal por sábanas de linfocitos sueltos y ausencia de estructuras foliculares. La atrofia tímica persistió en las ratas de tres a siete días pos tratamiento, recuperándose después. El bazo no apareció sensiblemente afectado. 274 RESULTADOS Se valoró también el efecto de la timectomía en la MO de ratas lactantes y el efecto del Tio-­‐TEPA. Se llevó a cabo la timectomía en ratas de 7 días de edad. Todas las ratas sobrevivieron y no hubo diferencias con el grupo control. Datos que no descartan al timo como un posible sitio de origen de linfocitos-­‐like de la MO, pero tampoco lo confirman. El Tio-­‐TEPA es algo único entre los agentes quimioterápicos, ya que en los niveles más altos de la gama de dosis letal existe un efecto linfocitotóxico severo, mientras que a dosis inferiores a las letales no se observa ningún daño en los linfocitos. Se observó que con altas dosis apareció una necrosis severa del componente celular en el timo, bazo y ganglios linfáticos y una simultánea reducción en el número de linfocitos de sangre periférica y linfocitos-­‐like de MO. La respuesta paralela a cortisona y Tio-­‐TEPA por los linfocitos-­‐like de la MO y los linfocitos de los órganos linfáticos periféricos, sugieren al autor que: “… estos tipos celulares son biológicamente similares, de hecho pueden ser la misma célula en diferentes ubicaciones” En la discusión Harris (412) dice: “… de acuerdo con la teoría polifilética, una célula totipotencial no puede ser un pequeño linfocito, y viceversa, los pequeños linfocitos en la MO no pueden ser totipotenciales. Por esta razón originalmente hemos llamado a las pequeñas células multipotenciales de MO de la rata linfocitos-­‐like”. Células similares en otras especies incluyendo el hombre han sido llamadas pequeños linfocitos, hematogonias, linfocitos protoplásmicos y células libres primitivas: “… los muchos nombres que se han aplicado y que hacen a estas pequeñas y redondas células prácticamente anónimas, también sirve para ilustrar las diferentes líneas celulares con las que se ha asociado”. Según Harris, que esta célula es multipotencial se basa en un volumen considerable de pruebas. Ya Maximov en 1923 informó de la trasformación de linfocitos pequeños en granulocitos en los explantes de ganglios linfáticos incubados con una mezcla de sangre, plasma y células de MO, posteriormente Yoffey también describe la trasformación de pequeños linfocitos a grandes blastos (415), Rebuck demostró la trasformación de los pequeños linfocitos en grandes linfocitos, monocitos y macrófagos (416), y la conversión de linfocitos 275 RESULTADOS en células plasmáticas puede observarse en la MO de ratas que se recuperan de Tio-­‐TEPA, metrotexate o 6-­‐mercaptopurina. Así pues, parece haber abundantes pruebas de que una célula multipotencial semejante al pequeño linfocito existe en MO, y puesto que su reacción a la cortisona y Tio-­‐TEPA es idéntica a la del pequeño linfocito en otros órganos debería llamarse linfocito pequeño. Yoffey (415) después de descubrir una fase activa y otra inactiva del pequeño linfocito asegura: “si el estímulo activador no está presente, el pequeño linfocito permanece en fase inactiva durante un considerable período de tiempo”, que coincide con la opinión que da Harris según su propia experiencia: “mis propios estudios me han llevado a conclusiones similares, que el linfocito pequeño en la rata representa una fase de reposo de una célula totipotencial”. Y a continuación explica más detalladamente su punto de vista. Considera que el desarrollo de las células de la MO se debe en su mayor parte a precursores blásticos tradicionales. Sin embargo, cuando se altera la homeostasis, ya sea durante ciertas fases del crecimiento, por enfermedad o durante un proceso experimental, los pequeños linfocitos aumentan su tamaño, y durante esta fase se pueden encontrar integrados en éstas u otras formas celulares. El modus operandi que podría explicar la capacidad del linfocito para existir en fases activas e inactivas, se sugiere por los recientes estudios del metabolismo de los ácidos nucleicos. Resulta interesante la especulación que hace sobre algunas discrasias sanguíneas, como por ejemplo: ¿la súbita aparición de un gran número de blastos en las leucemias agudas podría representar la conversión de linfocitos pequeños a otra célula metabólicamente más activa y por tanto más tóxica, en lugar de corresponder a un clon derivado de un solo foco de células malignas? Y por el contrario ¿no podría el aumento gradual de linfocitos pequeños, indolentes, característicos de la leucemia linfática crónica, ser el resultado de una anormal presentación de esta célula en fase inactiva, y no resultado de una mayor proliferación? 276 RESULTADOS Preguntas que hoy día nos pueden sorprender, pero que tiene toda su lógica si nos situamos en ese momento de la historia. Cuando observamos un fenómeno que desconocemos, hemos de agotar todas las posibilidades explicativas, para posteriormente ir eliminando en medida de lo posible una a una. Y la medida de lo posible se nos queda todavía muy pobre, será años más tarde, la biología molecular y la genética las que nos indiquen el camino a seguir, y harán que surjan nuevas preguntas y se multipliquen los interrogantes en este campo. En este sentido el autor dice: “las especulaciones de este tipo carecen de sentido a menos que sean probadas mediante más experimentación. Se ofrecen para indicar que desde la teoría actual no podemos explicar interpretaciones contradictorias de los hechos existentes, nuevas bases de la interpretación deben ser buscadas”. Solo nos queda comentar algo sobre las teorías que explican el origen de la pluricelularidad: la monofilética y la polifilética a las que el autor hace referencia (417). La teoría monofilética fue propuesta por Darwin y considera que el origen de una unidad sistemática tiene lugar a partir de una misma fuente, se puede representar como las ramas de un árbol en el que todo se origina a partir de un mismo tronco. La teoría polifilética defiende en cambio que un grupo de seres vivos se originan a partir de varias líneas evolutivas y no de un solo ancestro. Traducido al terreno celular significaría la existencia de una célula madre por cada línea celular (418)(419). Actualmente la teoría aceptada universalmente es la monofilética que propone que las células derivan de una sola célula madre pluripotencial o stem cell. De esta célula madre surgirían dos líneas celulares, una célula madre mieloide y otra linfoide. Pese a que estas células madres poseen marcadores de identidad diferentes en su membrana plasmática, en estos años (cuando Harris realiza su estudio) no hemos llegado todavía a la era del esplendor de la biología molecular y la diferencia basada exclusivamente en criterios morfológicos es 277 RESULTADOS prácticamente imposible. En el trabajo, Harris está claro que confunde la morfología del linfocito con la de la célula madre, y quizás la linfocitosis a la que alude no es tal linfocitosis, sino un aumento de células madres que darán lugar a todas las demás células, ya que las células madres tienen la capacidad de dividirse y diferenciarse en diversos tipos celulares especializados y de auto-­‐renovarse para producir más células madres (411). El conocimiento y los avances en la morfología y patología del linfocito van a ser de gran ayuda en el estudio, diagnóstico, pronóstico y tratamiento de diversas enfermedades. Con el titulo: “El valor diagnóstico de los mucopolisacáridos aislados de la orina y de las inclusiones linfocitarias en el Gargolismo” Helen Muir, Ursula Mittwoch y Thomas Bitter (420) publicaron sus estudios en 1962, Londres. El síndrome de Hurler o gargolismo es una rara enfermedad determinada genéticamente (421) con herencia autonómica recesiva y ligada al sexo. Dos de los seis mucopolisacáridos sulfatados conocidos, el condroitin sulfato B y el heparitin sulfato, se acumulan en exceso en la mayoría de los órganos del cuerpo especialmente hígado y bazo, apareciendo como depósitos metacromáticos, sin embargo son muy solubles en fijadores de rutina, por lo que las biopsias hepáticas pueden mostrar vacuolas vacías a menos que sean fijados con técnicas especiales. Ambos mucopolisacáridos se excretan en grandes cantidades en la orina y son resistentes a la hialuronidasa. Se ha encontrado que un porcentaje de los linfocitos de estos pacientes contienen inclusiones citoplasmáticas anormales (422) que pueden ser vistas en preparaciones teñidas con May-­‐Grumwald-­‐Giemsa, y también con azul de toluidina. En el presente estudio (420) estas inclusiones se han correlacionado con la presencia de cantidades anormales en la orina de mucopolisacáridos resistentes a la hialuronidasa. Los mucopolisacáridos aislados de la orina se purificaron y aislaron mediante un método de precipitación. Para demostrar las inclusiones intracelulares, se 278 RESULTADOS obtuvieron muestras de sangre por punción de los pulpejos de los dedos utilizando el método del cubreobjetos (423). Tales preparaciones han demostrado ser superiores a las obtenidas en el portaobjetos para la preservación del detalle citológico, ya que los leucocitos están menos dañados cuando se extiende por fuerza capilar. Se fijan en metanol absoluto y se tiñen con May-­‐Grümwald-­‐Giemsa (MGG) y toluidina. Se elaboraron varias tablas en las que se recogieron los signos clínicos de los pacientes con inclusiones linfocíticas y mucopolisacáridos en orina o bien con inclusiones en linfocitos únicamente. El rango de edad de los pacientes es de 2 a 15 años. Todos presentaron un retraso mental más o menos severo y fue frecuente la presencia de opacidades en córnea, aumento de hígado y bazo, vértebras en cuñas y/o cifosis. Figura 36. Linfocitos con inclusiones. Preparación secada al aire y teñida con May-Grümwald-Giemsa. Tomado de: Muir y colab, 1962 (420) Se estudiaron 11 pacientes con hallazgos clínicos y radiológicos de gargolismo. En 7 de los pacientes que excretaban grandes cantidades de mucopolisacáridos resistentes a hialuronidasa, se encontraron inclusiones características en un 279 RESULTADOS porcentaje variable (5-­‐26%) de linfocitos. Estas inclusiones aparecen típicamente como vacuolas bien definidas con un centro granular profundamente teñido que puede tomar la forma de punto, coma o anillo (Figura 36). Las vacuolas se distribuyen típicamente en grupos, y con la tinción de MGG los gránulos aparecen rojos o violetas mientras que con azul de toluidina se tiñen metacromáticamente apareciendo de color púrpura en un citoplasma azul. La morfología de estas inclusiones se suele distinguir bien de los gránulos azurófilos de los linfocitos normales, pero el criterio final es su metacromasia con azul de toluidina, cosa que no ocurre con los gránulos azurófilos del citoplasma. Algunas de las vacuolas linfocitarias no tuvieron valor diagnóstico de gargolismo, ya que era imposible distinguirlas de aquellas que se ven en ciertas condiciones patológicas, como por ejemplo, la idiocia amaurótica, y tampoco podían diferenciarse de vacuolas normales. La excreción de mucopolisacáridos de los cuatro pacientes restantes fue tanto cualitativamente como cuantitativamente similares a la normal de los sujetos control, y no se encontraron inclusiones en los linfocitos de estos pacientes, descartándose el diagnóstico de gargolismo. En la discusión se hace referencia a una serie de entidades como la disóstosis encondral recesiva, el síndrome de Morquio-­‐Brailsford-­‐Ullrich, gargolismo, disóstosis epifisaria elástica y un número de observaciones de difícil clasificación que forman un grupo de enfermedades con una superposición clínica y radiológica hasta el punto de que han sido consideradas como una entidad nosológica. Se aborda la polémica sobre los gránulos linfocitarios que fueron descritos por Alder en 1939 en dos hermanos que en la pubertad desarrollaron osteocondronecrosis meta-­‐epifisaria como ocurre en la enfermedad de Ribbing o displasia ósea esclerosante (424). Reilly (425) en 1941 describe gránulos similares en cuatro de ocho pacientes con gargolismo, posteriormente se encontraron en el síndrome de Morquio-­‐Ulrich-­‐Brailsford, y hay autores como Undritz (426) que afirman que no son característicos de ninguna de estas enfermedades. Otros autores aseguran que no hay tales gránulos anormales en los enfermos estudiados de gargolismo, y que las 280 RESULTADOS células de un exudado inflamatorio inducido (427) muestran inclusiones morfológicamente similares a las aquí descritas. Los autores (420) añaden: “En los 22 casos de gargolismo examinados hasta ahora, los linfocitos de la sangre periférica contenían inclusiones características como las descritas por primera vez por Gasser y Zellweger (428) en tres pacientes con gargolismo”. Aunque ellos no usaron azul de toluidina, la descripción morfológica coincide con las presentes características descritas, por lo que Muir y Mittwoch mantienen que tales inclusiones metacromáticas han sido comunicadas hasta ahora solo en el gargolismo. El defecto metabólico origen de la enfermedad sigue siendo desconocido, y no se han encontrado diferencias bioquímicas entre las formas ligadas al sexo y las formas autonómicas recesivas a la fecha de esta publicación. “Las inclusiones metacromáticas de linfocitos parece ser una prueba diagnóstica, que hasta ahora ha sido completamente confirmada por los hallazgos bioquímicos de mucopolisacáridos sulfatados urinarios anormales”, es la conclusión a la que llegan sus autores. A día de hoy, siglo XXI, el diagnóstico de las mucopolisacaridosis se realiza mediante técnicas enzimáticas o genéticas. El análisis morfológico de los linfocitos, si se hace, es meramente una curiosidad. No sabemos los motivos de su olvido, quizás estas inclusiones sean poco específicas, aparezcan también en otras enfermedades no metabólicas, su apreciación morfológica quizás no sea tan sencilla como muestran los autores, o quizás sea un método tan sencillo y económico que no tenga cabida en esta sociedad de mercado. No lo sabemos, tendríamos que realizar un extenso y detallado estudio para poder contestar. Lo que si está fuera de duda es el espíritu investigador y pragmático de los autores en la búsqueda de un marcador específico, sencillo de realizar y de bajo coste, marcando así una línea de actuación muy futurista, y adelantándose a lo que unos años más tarde se conseguiría a un mayor coste monetario con técnicas de genética y biología molecular. Hoy día asistimos a la búsqueda de marcadores específicos en casi todas las enfermedades, marcadores no solo 281 RESULTADOS diagnósticos, sino también de pronóstico y tratamiento, donde la existencia de dianas terapéuticas más o menos específicas es un hecho. Las mucopolisacaridosis son enfermedades poco frecuentes, debidas a mutaciones genéticas y con una herencia normalmente autosómica recesiva. En 1900, en Edimburgo figura la primera descripción de un caso de mucopolisacaridosis. La enfermedad de Hurler toma su nombre de Gertrudis Hurler quien en 1919 publicó la historia clínica de la enfermedad (429). En 1952, Brante (430) aisló el mucopolisacárido dermatán sulfato del hígado de los pacientes con síndrome de Hurler, surgiendo el término de mucopolisacaridosis. En 1969, McKusick y colaboradores (431) proporcionan una clasificación numérica basada en el tipo de sustancia excretada en la orina y las características clínicas dominantes, y en 1970 el déficit enzimático es señalado por Neufeld y Fratantoni (432). Por otro lado el gargolismo, con una frecuencia inferior a 5/10.000 personas es considera actualmente una “enfermedad rara”, y como tal y durante siglos, este tipo de enfermedades muy poco frecuentes y que se acompañaban de alteraciones morfológicas importantes se podían ver en circos y espectáculos callejeros ambulantes, la única forma de supervivencia que tenían muchos de los afectados. El gargolismo fue una enfermedad de circo, rara, extraña, que despertaba curiosidad y morbo, como se recoge en “El hombre que compraba gigantes” del autor extremeño Luis Folgado (433), que ambientada en la España del siglo XIX, relata la vida de Agustín Luengo. Con el avance científico y el conocimiento etiológico de las enfermedades también se produce una comprensión de las mismas, una “normalización social” y su humanización. Actualmente este tipo de enfermedades poco frecuentes siguen siendo “enfermedades raras”, pero dentro de un orden conceptual muy diferente, son difíciles de abordar, ya que se requieren esfuerzos especiales, unificados y de coordinación internacional. El síndrome de Hurler figura en el listado de enfermedades raras de la Federación Española 282 RESULTADOS de Enfermedades Raras (FEDER), pero ha ganado su lugar en la medicina, respeto y dignidad (434). De naturaleza muy distinta, pero de gran interés es el trabajo de Milton H. Brown (435) que publica en 1962 un compendio sobre los resultados de estudios que relacionan inmunidad y radiaciones ionizantes: “Efecto de las radiaciones ionizantes sobre la inmunidad”. Hay abundante evidencia experimental, tanto en animales como en el hombre, de que la radiación ionizante tiene un efecto perjudicial sobre el sistema inmune. Grandes dosis de radiaciones ionizantes inducen graves “enfermedades por radiación” que se manifiestan con síntomas típicos. Dosis pequeñas pueden no presentar efectos visibles externos, y tener como única manifestación la afectación del sistema hematopoyético. Los efectos generales de la radiación van a depender no solo de la dosis total, sino también de la velocidad de absorción y la región corporal expuesta, así 1000 rem en una sola dosis sería por lo general fatal si todo el cuerpo estuviera expuesto, pero no habría efectos visibles en la mayoría de las personas si la misma dosis se aplica más o menos uniformemente durante un período de 30 años. Diferentes partes del cuerpo tiene también diferente sensibilidades, siendo las partes más radiosensibles el tejido linfoide, médula ósea, bazo, órganos reproductores y sistema gastrointestinal, y la sensibilidad varía de un individuo a otro. Los efectos de una irradiación corporal total aguda van a depender de la dosis. Con 25-­‐100 rem no se aprecia enfermedad visible, pero en la sangre se advierten cambios, con una caída de linfocitos y granulocitos. De 100-­‐200 rem se produce malestar y fatiga, puede que náuseas y vómitos y cambios más marcados en sangre. Después de la exposición a 200-­‐1000 rem, la supervivencia puede tener lugar en el rango inferior de esta dosis, pero será muy pobre en el superior. Aparecen náuseas, vómitos y hemorragias, con un aumento de la susceptibilidad a las infecciones y depresión marcada de los elementos sanguíneos con lo que la inmunidad se altera profundamente. Con más de 1000 rem la supervivencia es improbable. 283 RESULTADOS El sistema inmune es de una gran complejidad, Coons (436) mostró evidencia de que los anticuerpos están presentes en las células plasmáticas de la pulpa roja del bazo, área medular de ganglios linfáticos, submucosa de íleon y tejido conectivo portal de hígado, y también encontró pequeñas cantidades en células de folículos linfoides del bazo y ganglios linfáticos apuntando que los linfocitos pueden tener una pequeña contribución en la producción de anticuerpos. Parece pues que las células plasmáticas son las principales responsables de la síntesis de anticuerpos y que el gran linfocito es el principal antepasado de plasmablastos y células plasmáticas. Así pues, las células con el papel más importante en los mecanismos inmunitarios se encuentran en los tejidos más sensibles a la radiación. Se ha observado que la radiación provoca además cambios en el equilibrio normal de la microflora del tracto gastrointestinal, sistema respiratorio y la piel, y que este factor juega un importante papel en el curso y desenlace de la enfermedad por radiación. Con dosis subletales las bacterias del tracto gastrointestinal invaden tejidos y órganos dando lugar a una bacteriemia que puede conducir a la muerte, y el mismo fenómeno se ha observado en tracto respiratorio y piel. El daño en las defensas naturales del cuerpo conduce a un aumento de la permeabilidad para microorganismos y a una disminución de la fagocitosis por disminución del número de células y disminución de la eficacia bactericida de suero, sangre y piel (437)(438). Pero estas observaciones en el hombre son muy escasas. Después de la detonación de la bomba atómica en agosto de 1945, los habitantes de Hiroshima mostraron evidencia clínica y patológica de estos efectos, por manifestaciones tales como angina, neumonía, enteritis, colitis, sepsis y otras infecciones, pero no fue posible determinar la dosis, ni fueron posibles los exámenes bacteriológicos debidos a las condiciones del momento. Un seguimiento documentado completo fue posible en el pueblo de Rongelap, en las islas Marshall, que constituye unos de los pocos registros de los resultados a corto y largo plazo en el hombre después de la radiación corporal total (439)(440)(441)(442). En Rongelap, un grupo de 64 personas fueron 284 RESULTADOS expuestas a 175 rem y 18 personas a 69 rem. Aparte de náuseas y vómitos, el hallazgo más frecuente fue el efecto sobre las células blancas de la sangre, el número de linfocitos disminuyó rápidamente y se mantuvo bajo hasta la décima semana, no volviendo a la normalidad en algunos casos durante meses y años después. El recuento en el número de linfocitos fue el hallazgo de más valor, especialmente en las dosis más bajas, su disminución se produce en cuestión de horas después de la exposición. A pesar de la depresión en la hematopoyesis, en la gente de Rongelap no había ninguna otra señal de enfermedad relacionada con la exposición, solo los títulos de anticuerpos determinados a parainfluenza, virus sincitial respiratorio, psitacosis y fiebre Q, fueron algo menores en las personas expuestas. Así mientras los efectos en el sistema hematopoyético eran notables, no se apreció ningún tipo de perturbación en la resistencia normal de las personas expuestas. Investigaciones sobre el efecto de la radiación en el desarrollo de la inmunidad activa ha sido llevado a cabo en animales, y en todos los casos se han empleados antígenos no vivos como toxoides o células rojas de la sangre. Se realizaron un gran número de exposiciones en una amplia variedad de animales, con diferentes dosis de radiación y diferentes antígenos. Se observó que la respuesta inmune primaria era aproximadamente tres veces más radiosensible que la secundaria. La conclusión general que se puede extraer de los trabajos en animales es que la dosis de radiación ionizante subletal en el hospedador, incuestionablemente inhibe la producción de anticuerpos cuando se administra al comienzo de la inmunización, mientras su efecto no es tan marcado en el caso de la producción de inmunidad secundaria (443). Si estos resultados pueden ser extrapolados al humano, la inmunización activa debería llevarse a cabo mucho antes de la exposición a la radiación ionizante. En cuanto a la mejora de la inmunidad, debe tenerse en cuenta que en experimentos controlados en animales se han comunicados resultados favorables de la aplicación local de rayos X. En el hombre, dermatólogos y radiólogos pueden dar fe del efecto beneficioso de pequeñas dosis de radiación aplicadas localmente para controlar la infección, empleándose dosis que no 285 RESULTADOS superan los 75 rem, pero no existe todavía una explicación clara sobre este efecto potenciador de la inmunidad (438). Estudios pioneros en radiobiología demostraron el efecto perjudicial en animales de la radiación ionizante en la reducción a la resistencia de microorganismos patógenos. Hecho que posteriormente fue confirmado en varios estudios (437)(444). Los agentes estudiados incluyeron Cl. perfringens, Cl. tetani, Staphylococcus aureus, Streptococcus haemolyticus y C. diphtheriae. Una significativa recuperación de la capacidad de los sobrevivientes para luchar contra los agentes infecciosos tuvo lugar sobre la tercera semana después de la radiación, período conocido como de “resistencia mínima” que varía según los distintos animales. Es razonable suponer un efecto similar en el hombre, donde existe la necesidad de comenzar un tratamiento lo antes posible en aquellas víctimas de desastres nucleares. Además el tratamiento debería mantenerse durante todo el tiempo suficiente hasta que el sistema inmunológico se halla recuperado lo suficiente como para asegurar la supervivencia del individuo. Los efectos de la irradiación prolongada a bajos niveles en la respuesta inmune son generalmente menos perjudiciales que una dosis similar aguda (445). En conejos este tipo de radiación, no ocasionó una reducción significativa del título de anticuerpos ni una depresión de los niveles máximos. Por inferencia, los mecanismos inmunitarios de los individuos expuestos a dosis bajas de radiación en caso de un desastre atómico no se verían afectados de forma adversa. Finalmente Brown (435) da a conocer sus conclusiones: 1. Cuando las radiaciones ionizantes se aproximan a dosis letales y se administran en todo el cuerpo, disminuye la inmunidad del huésped a los habitantes bacterianos normales del tracto gastrointestinal, sistema respiratorio y piel y se reduce la formación de anticuerpos especialmente a una estimulación primaria. 286 RESULTADOS 2. El tratamiento de los individuos que manifiestan evidencia de infección después de la radiación debe mantenerse el tiempo suficiente para que el proceso de recuperación inmunológica tenga lugar, lo que puede durar semanas. 3. La inmunización activa debe llevarse a cabo antes de la exposición a la radiación ionizante. 4. Pequeñas cantidades de rayos X administrados localmente, mejoran la formación de anticuerpos y la resistencia de los animales de experimentación a ciertas infecciones. Rongelap es un atolón de Micronesia, formado por 61 islas y situado en el Océano Pacífico. Cuando la mayor parte del mundo vivía con el miedo de la guerra nuclear, para estas islas del pacífico, la guerra nuclear era una realidad (446)(447)(448). Desde 1946 a 1958, el ejército de los Estados Unidos llevó a cabo numerosas pruebas nucleares incluyendo la bomba de hidrógeno, sobre todo en el atolón de Bikini a unos 150 kilómetros de Rongelap (449). El 1 de marzo de 1954 explotó una bomba de hidrógeno de nombre “Bravo” que fue mil veces más potente que “Little Boy” lanzada en Hiroshima, lo que aumento el nivel de radiación media de fondo en todo el mundo. Los habitantes del atolón de Bikini fueron evacuados previamente, pero no así los de Rongelap a 150 kilómetros: “restos como de nieve cayeron durante el día y la noche…”. Los isleños de Rongelap no fueron evacuados hasta después de más de 48 horas de la explosión, lo que proporcionaría datos valiosos para evaluar el efecto de la radiación en los seres humanos (450). Treinta años después, el 95% de la población que había estado expuesta a la radiación había desarrollado cáncer de tiroides y una alta proporción de sus descendientes sufrían defectos genéticos (451)(452). Después de varios años de lucha, evacuaciones y reasentamientos de los isleños, en 1985 Greenpeace evacua a las personas de Rongelap y las asienta en una isla a 180 kilómetros, en la llamada “Operación Éxodo”. En septiembre de 1996, el Departamento de Interior de los E.U firma un acuerdo de reasentamiento asignando 45 millones de dólares a los isleños (453)(454). 287 RESULTADOS ¿Cómo se puede valorar la pérdida de salud y la destrucción genética de una población? ¿Quién pone el precio? El impacto del microscopio electrónico en la investigación se aprecia igualmente en el trabajo de Inman y Cooper (455) del Fondo Imperial para la Investigación del Cáncer, Londres, publicado en 1962: “Microscopía electrónica de linfocitos humanos estimulados por fitohemaglutinina”. Las células mononucleares de sangre periférica se someten a una reacción proliferativa intensa cuando se cultivan en presencia del mitógeno fitohemaglutinina (PHA). Esta proliferación se acompaña de un cambio marcado en la morfología de las células, observándose al microscopio de luz un aumento de tamaño. Las células trasformadas son móviles, sus núcleos aumentan, presentan uno o más nucléolos y aparecen vesículas de diferentes tamaños y densidad dispersas en sus citoplasmas. Los autores en este informe describen las principales características morfológicas vistas con el microscopio electrónico de los linfocitos estimulados por PHA. La fitohemaglutinina es una lecitina ampliamente distribuida entre legumbres y algunas oleaginosas. Se caracterizan por su habilidad para combinarse con receptores de membrana carbohidratos, además estimula inespecíficamente la proliferación de células T y esta característica mitogénica es la que le da su principal aplicación. Las células a estudiar se obtienen de sangre periférica o de la linfa del conducto torácico de pacientes sometidos a toracotomía, se cultivan según el método de Moorhead (456) y son preparadas de forma adecuada para su visualización bajo microscopio electrónico. La ultraestructura observada (455) en los linfocitos aislados de sangre periférica fue similar a la ya descrita por otros autores (457). Los pequeños linfocitos tienen un escaso y fino citoplasma y pocas organelas, éstas consisten en un puñado de mitocondrias grandes y ovaladas, alguna vesícula de origen micropinocítico, aisladas laminillas ergatoplásmicas y ocasionalmente una vacuola compuesta. Esta escasez de organelas citoplasmáticas era menos 288 RESULTADOS marcada en los grandes linfocitos. Las ultraestructura de los linfocitos de los cultivos de control no fue diferente de la de las células aisladas. En los cultivos celulares con PHA de tres días, se observó una mayor complejidad en el citoplasma de dichas células, mitocondrias más abundantes y a veces con formas alargadas o extrañas. El cambio citoplasmático más notable fue el aumento de contenido de ribosomas, apareciendo solos o agrupados en rosetas. Se observó también un aumento de la micropinocitosis y como resultado un aumento del número de vesículas citoplasmáticas. Hubo una clara disminución de la densidad de la cromatina en los núcleos de las células transformadas. (Figura 37) Figura 37. Linfocito transformado de un cultivo de tres días con PHA. La forma alargada de esta célula es indicativa de su estado móvil en el momento de la fijación. La disminución de la densidad de la cromatina en el núcleo (N) y los nucleolos bobinados son evidentes (nu). La doble membrana nuclear (NM) suele ser irregular. El citoplasma contiene numerosos ribosomas libres, mitocondrias (M), un aparato de Golgi ampliamente desarrollado (G) y numerosas vesículas. Hay evidencia de activa micropinocitosis (p). Tomado de: Inman y Cooper, 1962 (455). 289 RESULTADOS Los autores consideran la existencia de una considerable evidencia que sugiere que los pequeños linfocitos, células diploides en reposo, son las células principalmente estimuladas cuando se cultiva sangre con PHA. La linfa del conducto torácico contiene una población más uniforme que la sangre periférica de células mononucleares y la acción de la PHA en ellas produce una transformación similar a la observada en los cultivos de sangre. El gran aumento del contenido citoplasmático de ribosomas es indicativo de una síntesis activa de proteínas, que puede estar asociado con el crecimiento de las células y la síntesis de nuevas organelas. Cristales de proteínas y estructuras similares han sido comunicados en el citoplasma de las células linfoides leucémicas. Los autores (455) indican que: “Estas observaciones ponen de manifiesto el potencial latente de los linfocitos para cambiar su estructura y metabolismo, y este efecto es tan profundo que las células transformadas adquieren muchas de las características ultraestructurales de las células cancerosas”. En 1963, autores como Robert C. Mellors y Leonard Korngold (458) deciden continuar la línea de estudio sobre las inmunoglobluinas: “El origen celular de las inmunoglobulinas humanas”. Los autores dicen haber tenido la oportunidad durante años, de estudiar en secciones de tejido linfoide a familias de células que contienen y aparentemente forman inmunoglobulinas y anticuerpos (459). Existe además un número considerable de publicaciones (330)(460)(461)(462) que han presentando observaciones del origen celular de cada una de los tres tipos de inmunoglobulinas humanas (γ2, γ1M, γ1A). Las globulinas relacionadas antigénicamente se han encontrado en la región electroforética que se extiende desde la lenta γ2-­‐globulina a la β-­‐globulina (463). El origen común de estas inmunoglobulinas como muestran los autores en esta comunicación justifica su consideración como familia. 290 RESULTADOS Las inmunoglobulinas fueron aisladas de diversos modos, así las fracciones séricas ricas en γ2-­‐globulinas fueron obtenidas bien por electroforesis, cromatografía de intercambio iónico o fraccionamiento mediante alcohol frío. Las γ1A-­‐globulinas mediante electroforesis utilizando suero de pacientes con mieloma como material de partida, y las γ1M-­‐globulinas se aislaron mediante electroforesis a partir de sueros de pacientes con macroglobulinemia. Los antisueros se elaboraron de conejos inoculados con las correspondientes γ-­‐ globulinas, y se evaluaron por técnicas de inmunoelectroforesis y doble difusión en gel. Las fracciones de anticuerpos activos de los antisueros fueron separadas y conjugadas con una proteína ligada a isocianato de fluoresceína, obteniéndose así anticuerpos fluorescentes. La especificidad de la inmunofluorescencia conseguida se evaluó regularmente en secciones de tejido linfoide en los que existiese una población grande o exclusiva de células que contienen γ1M-­‐globulinas, como ocurre en la macroglobulinemia de Waldeström o la artritis reumatoide y también en aquellos con un predominio de células que contiene γ2-­‐globulinas. Los tejidos empleados procedían de piezas quirúrgicas y biopsias obtenidas de 279 pacientes, de los que 118 padecían artritis reumatoide y el resto diversas enfermedades no neoplásicas. Cada una de las inmunoglobulinas fue identificada en los centros germinales de los nodos linfoides de los ganglios linfáticos y bazo. Los centros conteniendo γ2-­‐globulinas fueron los más numerosos, seguidos de los que contenían γ1M-­‐ globulinas. La inmunoglobulina se localizó casi exclusivamente en el citoplasma de las células del centro germinal, pero también se pudo detectar en el entorno inmediato extracelular. Las células de los centros germinales que contienen inmunoglobulinas corresponden con tinciones convencionales a agregaciones esféricas o elípticas de células primitivas, que se distinguen por un núcleo pálido con nucleolo, contorno celular indistinto, citoplasma basófilo y pirinofílico y procesos celulares continuos y alargados. El manto de linfocitos pequeños maduros que característicamente rodea el centro germinal nunca mostró cantidades detectables de inmunoglobulinas. 291 RESULTADOS Cada una de las inmunoglobulinas (un tipo por célula) fue identificada en ocasiones en el citoplasma de las células primitivas de tamaño grande, intermedio y pequeño en el tejido linfoide de los cordones medulares de ganglios linfáticos o la pulpa roja del bazo. A las grandes células primitivas, muy semejantes a las vistas en los centros germinales les llamaron células reticulares primitivas, y consideran que las células de tamaño intermedio posiblemente corresponden a una etapa de transición a células plasmáticas con las que se asocian normalmente. Cada una de las inmunoglobulinas (un tipo por célula, con raras excepciones) se identificó en el citoplasma de las células plasmáticas inmaduras y maduras. Aquellas más maduras comprenden a las células tipo Marschalko y las de apariencia más distintiva contenían cuerpos de Russell en su citoplasma. La mayoría contenían γ2-­‐globulinas, y algunas parecían no contener cantidades detectables de cualquier inmunoglobulina. En las secciones convencionalmente teñidas, las células tipo Marschalko se ajustaba a la descripción clásica: forma ovalada o redonda, citoplasma profundamente basófilo o pirinofílico, distintivo contorno celular, núcleo excéntrico con grumos gruesos de cromatina y normalmente una zona pálida juxtanuclear. . Figura 38. Células plasmáticas 20- Dos células plasmáticas con cuerpos de Russell del bazo de conejo. 21- Dos células plasmáticas del bazo de ratón formando T2-globulina. Tomado de: Mellors y Korngold, 1963 (458) 292 RESULTADOS Los autores (458) realizan una amplia discusión sobre el tema señalando algunos puntos importantes. El resultado indica la presencia de dos familias relacionadas de células que forman inmunoglobulinas y anticuerpos en el tejido linfoide humano: células de centros germinales (células reticulares) y células plasmáticas. Reconocen el trabajo de Fagraeus: “… la función de las células plasmáticas ha sido ampliamente documentada por los trabajos de Fagraeus y sus seguidores”. Las investigaciones de Fagraeus (272) indicaron que la formación celular de anticuerpos en el conejo se llevó a cabo con y durante el desarrollo de células reticulares en plasmáticas, y los trabajos de Coons, Leduc y Connolly(436) con técnicas de inmunofluorescencia y los de Nossal (464) y Attardi (465) mostraron que la célula plasmática inmadura era la principal fuente de anticuerpos. Estudios con inmunofluorescencia que también indican que los centros germinales fueron el sitio de formación de γ-­‐ globulinas en el hombre y de anticuerpos en animales de experimentación. La opinión de que las células del centro germinal pueden convertirse en miembros de la familia de las células plasmáticas ha sido expresada por Nossal y apoyada por Congdon y Makinodan (466). Los autores (458) llaman la atención al hecho de la presencia de inmunoglobulinas en grandes células primitivas (células reticulares grandes), que se asemejan a las grandes células de los centros germinales, pero dicen, son células diferentes a ellas, y también comunican la existencia de células reticulares de tamaño intermedio y pequeño con inmunoglobulinas, a las que consideran posiblemente células en proceso de transición a células plasmáticas mucho más numerosas. El punto de vista expresado por Nossal, una vez revisadas las pruebas en pro y contra: “es difícil escapar a la conclusión de que la mayoría de pequeños linfocitos no parecen formar anticuerpos”, es apoyado por los autores en el presente estudio, ya que el manto de pequeños linfocitos que rodea al centro germinal, no contiene cantidades detectables de inmunoglobulinas. 293 RESULTADOS Otro de los puntos de discusión se refiere a la aparición de más de un tipo de inmunoglobulina en una célula. Globulinas γ1M y γ2 se identificaron en ocasiones en la misma célula plasmática grande o pequeña, inmadura o madura y en el mismo centro germinal del tejido linfoide, que contienen una abundante distribución “policlonal” de células que forman inmunoglobulinas de todos los tipos, pero con la excepción señalada de solo un tipo por célula. Los estudios con fluorescencia determinaron que la gran mayoría de las células mostraron una inmunoglobulina señalada por la presencia de una coloración, ya sea amarilla, verde o roja… pero de vez en cuando fueron mezclas de colores, y métodos más sensibles han demostrado la presencia de dos anticuerpos diferentes en la misma célula en animales de experimentación (467)(464). A lo que los autores (458) añaden: “Desde que este manuscrito fue completado, nuevos estudios realizados por Solomon, Fahey y Malmgren han sido publicados (468), y sus observaciones en el mieloma múltiple revelan una diversidad de formas morfológicas en células malignas que contienen cada tipo de proteína monoclonal, la coexistencia de proteína de Bence Jones y proteína de mieloma en la misma célula y la presencia del mismo tipo de proteína en todas o casi todas las células malignas de un paciente determinado. Este estudio y otro en curso con células normales, parecen ser coherentes entre sí y mutuamente complementarios”, pero que ellos solo pueden decir de momento que en general solo un tipo de inmunoglobulina estuvo presente en una célula individual o centro germinal, y que las inmunoglobulinas γ1M y γ2 fueron identificadas ocasionalmente en la misma célula plasmática y centro germinal. Debido a su alto contenido en linfocitos, el timo ha sido considerado durante mucho tiempo como parte del sistema linfático, pero las diferencias histológicas y celulares con respecto a otros órganos linfoides plantea dudas en este sentido. Con el título: “Papel del timo en la Inmunidad”, JF Miller en 1963 (65), desde el Instituto para la Investigación del Cáncer en Londres, publica los resultados de sus investigaciones. Miller comienza su exposición realizando un recuerdo sobre el desarrollo y la estructura del timo de los mamíferos. Hace referencia a Hammar y Maximow 294 RESULTADOS quienes a principios de siglo afirman que las pequeñas células del timo son de origen mesodérmico, derivadas de células mesenquimales, en cambio Maurer y Stöhr sostienen que las células redondas del timo surgen directamente desde el epitelio reticular. La controversia sobre el origen de los linfocitos del timo parece haber sido resuelta por Auerbach en 1961 (469), que utilizó técnicas de cultivo para analizar los eventos morfogenéticos en el desarrollo del timo, y nunca aparecieron células huéspedes migratorias, ni mesodermo tímico, ni un generalizado mesénquima pareció contribuir significativamente en la población celular inicial del timo. Un timo totalmente desarrollado se divide en lóbulos, y posee dos tipos principales de células: linfocitos y células reticulares. Los linfocitos se encuentran en la corteza donde eclipsan a las células reticulares, no existiendo diferencias significativas entre estos linfocitos y los presentes en otros lugares. Las células reticulares son más prominentes en la médula, son células de naturaleza epitelial, grandes, de contornos mal definidos y núcleo vesicular grande. En la médula destaca la presencia de los corpúsculos de Hassall, que pudieran proceder de células reticulares hipertróficas o grupos de células reticulares activas, tiene forma poligonal irregular, aplanados, hialinizados y dispuestos concéntricamente alrededor de un núcleo central de residuos nucleares. El timo difiere de otras estructuras linfoides en varios aspectos como son la presencia de células epiteliales-­‐reticulares, los corpúsculos de Hassall, la ausencia de centros germinales y la presencia de escasas células plasmáticas que no aumentan su número después de la estimulación antigénica. Además la linfopoyesis del timo no se ve afectada por la estimulación antigénica, la resección de otros órganos linfoides, la timectomía parcial o la presencia de uno o más injertos de timo. Miller (65) intenta buscar contestación a algunas preguntas. ¿Cual es el destino de los linfocitos producidos en la corteza del timo? Autores como Sainte-­‐Marie y Leblond (470,471) creen que los pequeños linfocitos formados en la corteza migrarían a través de la médula con el fin de entrar en la circulación. ¿Pueden los linfocitos del timo de hecho alcanzar la circulación? es un interesante 295 RESULTADOS motivo de disputa, y según Miller: “teóricamente, todos los linfocitos formados en el timo pueden morir sin entrar en la circulación. Así mientras hay pruebas inequívocas de que el timo produce linfocitos, hasta ahora no hay evidencia directa que muestre que tales linfocitos estén destinados a otros lugares”. ¿Es el timo un órgano con capacidad de reacción inmunológica? La reactividad inmunológica se asocia generalmente con órganos linfoides tales como el bazo y ganglios linfáticos, y es el pequeño linfocito el tipo más común de célula linfoide presente en tales órganos, en sangre y linfa, demostrándose recientemente ser una célula competente en inmunidad, que puede iniciar una reacción inmunológica cuando tiene el estímulo apropiado (472). Aunque el timo es rico en pequeños linfocitos, no muestra producción de anticuerpos y no responde a los estímulos antigénicos mediante la producción de células plasmáticas o centros germinales. Por lo tanto su comportamiento es totalmente diferente al de otras estructuras linfoides. ¿Significa esto que no hay células en el timo capaces de tomar parte en las reacciones inmunológicas? Algunos de los experimentos ideados para resolver esta cuestión se describen a continuación (65): • La inyección directa de antígenos en el timo se asocia con una reacción plasmocelular y producción de anticuerpos dentro del timo. Se sugirió que el fracaso del timo para reaccionar a un antígeno circulante, era debido a la presencia de una barrera existente entre la sangre y el parénquima del timo que impedía la entrada del antígeno al timo (473). • Células aisladas de timo de ratón pueden dar lugar a un ataque inmune (reacción injerto contra huésped) contra un huésped extraño que sería menos eficaz que las células de ratón de los ganglios linfáticos. Pero en el ratón es difícil separar el timo de las pequeñas adenopatías torácicas que están adheridas a la cápsula, por lo que dicha reacción podría haber sido debida a contaminación de células ganglionares. • En cobayas y conejos, las células del timo tomados de donantes presensibilizados o de donantes normales y sensibilizados in vitro con 296 RESULTADOS antígenos de Shigella, han producido poco o ningún anticuerpo en los receptores apropiados, en contraste con los linfocitos no tímicos (474). La cuestión, es si hay células en el timo con capacidad inmunológica. Si el timo es necesario para mantener la reactividad inmunológica en su conjunto, es posible que la timectomía reduzca tal reactividad. La timectomía se ha realizado en muchas especies y cepas de roedores adultos jóvenes y no ha afectado a la tasa de crecimiento, comportamiento de la cría, susceptibilidad a infecciones o longevidad. No obstante, se asocia con una disminución de los linfocitos circulantes, tanto en sangre como en el conducto torácico de aproximadamente un 30-­‐40 % por debajo de los niveles normales y con una disminución en el peso del bazo y ganglios linfáticos en un 25 %. Sin embargo, no se aprecia ninguna disminución significativa en las capacidades inmunológicas del animal timectomizado en la vida adulta, lo que sugiere que la glándula del timo no participa en el control de la respuesta inmune (475). Dado sin embargo que el timo es el principal órgano linfoide en el recién nacido (RN), es posible que la timectomía en el RN, en contraste con la timectomía del adulto, pueda estar asociada con alteraciones más graves. Miller (65) lleva a cabo timectomías en ratones RN, observando que dichos ratones se desarrollan normalmente durante las primeras semanas. Después del destete sin embargo, muchos mueren a causa de una enfermedad degenerativa caracterizada clínicamente por disminución de peso, postura encorvada, piel erizada, diarrea y muerte en unas semanas (síndrome de la pérdida). Cuanto más temprano se realiza la timectomía en la vida del ratón, más alta es la incidencia de este síndrome, variando su comienzo de cuatro semanas a cuatro meses. El ratón neonatal timectomizado característicamente muestra un nivel persistentemente bajo de linfocitos en sangre periférica, de 50-­‐90 % por debajo de lo normal, y una marcada depleción de los linfocitos de los tejidos linfoides, desaparece la estructura folicular de los ganglios linfáticos y los centros germinales están ausentes. Los espacios normalmente ocupados por folículos linfoides muestran células reticuloendoteliales, histiocitos, 297 RESULTADOS macrófagos con hemosiderina y solo unos pocos linfocitos dispersos, la mayoría inmaduros. Hay una marcada deficiencia de linfocitos pequeños y una moderada hiperplasia de células reticuloendoteliales. Figura 39. Ratones timectomizados El ratón de la derecha, timectomizado al nacimiento, muestra ocho semanas después signos del síndrome de la pérdida. El ratón de la izquierda es un compañero de camada timectomizado al nacimiento pero injertado a los siete días con tejido tímico en la región subcutánea de la axila. Tomado de: Miller, 1963 (65) Se probó la capacidad de los ratones timectomizados neonatalmente para producir anticuerpos (65), estudios que mostraron que el 50-­‐80 % de tales ratones dejaron de producir niveles detectables de anticuerpos contra el antígeno H de Salmonella Typhi, virus de la influenza muerto o a eritrocitos de oveja. Similares deficiencias fueron comunicadas en ratas y conejos timectomizados al nacer por Jankovic y Archer (476). Ratones sometidos a un simulacro de operación o a esplecnectomía en la etapa de RN, pudieron más tarde rechazar los injertos de piel extranjera al igual que los ratones normales. Sin embargo, más del 70 % de los ratones RN timectomizados fallan en el rechazo a injertos de piel, no solo de donantes estrechamente relacionados genéticamente con el receptor sino de donantes muy alejados como pueden ser las ratas (477), y por lo general mantienen el injerto hasta la muerte. Sin embargo la timectomía realizada después de una semana de edad ya no se asocia con una prolongación significativa en la supervivencia de los injertos de piel extranjeros. 298 RESULTADOS Figura 40. Ratón timectomizado e injertado. Ratón de ocho semanas timectomizado al nacimiento e injertado a las cuatro semanas con piel de un ratón de distinta cepa. Nótese el mechón exuberante de pelo del injerto. Tomado de: Miller, 1963 (65 ) Por tanto, según los resultados del estudio de Miller, la timectomía en el ratón RN se asoció con: a. Una mortal enfermedad que se acompaña de diarrea. b. Una marcada disminución de la población de los pequeños linfocitos en sangre y tejidos. c. Disminución de la capacidad para producir anticuerpos en el suero para algunos antígenos. d. Un fallo permanente en la capacidad para rechazar injertos de tejido extranjero. Miller se pregunta: “¿Cómo se pueden explicar estos resultados?”. Se podría postular que en los RN, las células con potencial inmunológico en órganos linfoides distintos del timo son escasas o inexistentes y a medida que el animal desarrolla estos órganos linfoides recibe desde el timo una ayuda, ya sea celular o no, que los dota de células inmunológicamente competentes. 299 RESULTADOS En el animal adulto no ocurren alteraciones graves, ya que los órganos linfoides pueden continuar funcionando de forma independiente del timo. Pero si su potencial inmunológico es agotado o destruido, ¿puede el timo ser necesario para restaurar este potencial? A continuación Miller (65) realiza experimentos con ratones timectomizados en la vida adulta y expuesto a radiación corporal total para intentar dar respuesta. Los resultados mostraron: 1-­‐ una marcada disminución de los linfocitos en la sangre y tejidos de los ratones timectomizados e irradiados, pero que tal disminución no ocurría en los controles irradiados pero no timectomizados a los dos meses después de la irradiación. 2-­‐ un fallo para producir una respuesta inmunológica ya sea primaria o secundaria a eritrocitos de oveja y para rechazar los injertos de piel de 4 a 6 semanas después de la irradiación en ratones timectomizados, pero que tales deficiencias no existían en los controles. Es evidente por tanto que la recuperación del tejido linfopoyético y de las funciones inmunes en el ratón adulto irradiado, depende de un mecanismo controlado por el timo, según Miller (65): “el timo adulto es esencial para reestablecer los mecanismos inmunológicos cuando el aparato inmunológico ha sido dañado”. A continuación fueron probados una serie de métodos por su capacidad para proteger contra los efectos de la timectomía neonatal. Así la suspensión de células de bazo o ganglios linfáticos de animales normales, se encontró eficaz en la prevención de la emanciación y en el reestablecimiento del recuento de linfocitos cuando se inyecta a ratones de la misma cepa dentro de una semana después de la timectomía. Del mismo modo, células linfoides pre-­‐sensibilizadas eran capaces de rechazar un injerto de piel establecido en un ratón timectomizado al nacimiento. Los ratones timectomizados al nacer e implantados una semana después con un timo intacto procedente de un donante RN, tenían un período de vida normal, poblaciones normales de 300 RESULTADOS linfocitos y podían rechazar injertos de piel extranjeros como los ratones normales (64). La protección proporcionada por los implantes de timo puede ser explicada por el supuesto de que el implante produce linfocitos que pueblan al hospedador y restauran las funciones inmunes. Si esto es así, se podría esperar encontrar en los tejidos linfoides células con las características citogenéticas del donante de timo. Se emplearon técnicas para identificar cromosomas y se encontró que la mayoría de las células en los tejidos linfoides y timo implantado eran del hospedador y por tanto no habían sido aportadas por el timo donante. El timo por tanto podría ejercer su influencia mediante la producción de un factor humoral que induce la maduración de dichas células, o bien permitiría la diferenciación de estas células dentro de propio medio ambiente, es decir, las células del hospedador invadirían el implante, se diferenciarían en él y a continuación colonizarían los tejidos linfoides. Sobre el posible papel del timo en la inmunidad Miller (65) dice: “Cualquier teoría actual que pretenda explicar el papel del timo en la inmunidad debe sufrir necesariamente de la falta de conocimientos fundamentales a cerca de muchos aspectos de la fisiología de los linfocitos en general y del timo en particular. Dando por hecho ciertos supuestos, nos quedamos con dos hipótesis alternativas que no son excluyentes entre sí: la hipótesis celular y la hipótesis humoral”. Un factor humoral elaborado específicamente por el timo, podría actuar de diversas maneras, por ejemplo, permitiendo la diferenciación de precursores linfoides, estimulando la linfopoyesis o la recirculación de linfocitos, o bien dotando a los linfocitos no tímicos de la propiedad de la competencia inmunológica, existiendo pruebas experimentales que apoyan esta última posibilidad (65)(478). Cualquiera que sea la hipótesis verdadera, lo que está claro es que el timo tiene algún papel en el reestablecimiento de la reactividad inmunológica. Dado que el timo es esencial para la restauración inmunológica después de la irradiación corporal total, parece probable que también sea responsable de la pérdida de la tolerancia en esas circunstancias. ¿Es posible 301 RESULTADOS que el timo sea responsable en el desarrollo de la auto-­‐tolerancia y por tanto en el desarrollo de las enfermedades autoinmunes? En el hombre existen evidencias de una posible relación del timo con anormalidades inmunológicas. La aplasia tímica o linfocitotisis esencial es un síndrome reconocible a la edad de tres meses, cuando aparecen la clínica de diarrea, erupciones morbiliformes, infecciones bacterianas y fúngicas, a-­‐ gamma-­‐globulinemia y linfopenia marcada. Es una enfermedad fatal a pesar del tratamiento con antibióticos y transfusiones sanguíneas. Se observa una marcada atrofia del tejido linfático y una moderada a marcada hiperplasia reticuloendotelial, el timo está atrófico, casi carente de los pequeños linfocitos y no existen cuerpos de Hassall. Otra alteración encontrada es la hiperplasia con formación de centros germinales en el timo que se asocia con trastornos de naturaleza autoinmune. Lesiones hiperplásicas han sido descritas en artritis reumatoide, tirotoxicosis y enfermedad de Addison, y en todas ellas se han encontrado anticuerpos circulantes. Algunas neoplasias del timo están asociadas con trastornos clínicos que muestran característicamente alteraciones inmunológicas, ya sean de naturaleza primaria o secundaria, como por ejemplo la anemia hemolítica aguda, leucopenia y trombocitopenia. Informes de asociación clínica entre miastenia gravis y lupus eritematoso sistémico sugieren la posibilidad de una etiología común. Miller (65) acaba su publicación con unas sencillas conclusiones: “La asociación entre trastornos inmunológicos y alteraciones del timo junto a los resultados obtenidos a partir de experimentos en ratones, apoyan sin duda el concepto de que el timo tienen un papel clave en el control de la reactividad inmunológica, pero realmente es necesario trabajar mucho más para evaluar su papel en las enfermedades autoinmunes”. Este (65) es uno de los primeros trabajos de Miller sobre timo e inmunidad, estudios que se extenderán hasta finales del siglo XX. Las primeras publicaciones, desde 1959 a 1961, están centradas en el estudio que relacionan la leucemia y el timo: 302 RESULTADOS ! “Papel del timo en la leucemia murina”, de 1959 (479). ! “Destino de los injertos de timos subcutáneos en ratones timectomizados inoculados con filtrados leucémicos” (480). Jacques Francis Albert Pierre Miller (481)(482) médico e investigador australiano nacido en Francia, es conocido por haber descubierto las funciones del timo y por la identificación en los mamíferos de dos tipos de linfocitos: linfocitos T y B. Miller comenzó sus estudios doctorales a principio de la década de los sesenta, en Londres, en el Instituto de Investigación Chester Beatty, donde estudió la patogénesis de la leucemia linfocítica en ratones y el papel del timo en la enfermedad (483). En aquellos años se creía que el timo era un órgano residual sin función, pero debemos a Miller el descubrimiento de la importancia del timo como órgano vital con funciones claves en el sistema inmune adaptativo. Observó como aquellos animales que nacían sin él eran incapaces de generar rechazo a tejidos ajenos y resistir infecciones. De vuelta a Australia en 1966 para liderar el Instituto de Investigación Médica Walter y Eliza Hall en Melbourne, Miller descubrió que los linfocitos de los mamíferos podían ser clasificados en T y B, y que interactúan para producir anticuerpos. Su trabajo también mostró que el timo produce linfocitos T y elimina los autorreactivos, aportaciones fundamentales que han contribuido a entender enfermedades como el cáncer, el SIDA, las enfermedades autoinmunes, el fenómeno del rechazo, la alergia o la inmunidad antiviral. Mäkelä (484) procedente del Departamento de Serología y Bacteriología de la universidad de Helsinki, publica en 1964 estudios sobre la calidad de los anticuerpos: “Estudios sobre la calidad de los anticuerpos neutralizantes de bacteriófagos producidos por células individuales. II. Evidencias que indican que células diferentes de un ganglio linfático producen diferentes clases de anticuerpos contra el antígeno polisacárido 0-­‐9,12 de Salmonella”. Muchos estudios in vitro han demostrado que cuando un animal ha sido inmunizado con dos o más preparaciones de antígenos diferente, A, B, etc…, algunas de sus células solo producen anticuerpos anti-­‐A y otras anti-­‐B 303 RESULTADOS (485)(461). Tratar de explicar el porqué de esta observación es el objetivo de Mäkelä. El estudio se hizo con el antígeno 0-­‐9,12 de Salmonella. Los animales fueron inmunizados con bacterias que portan el antígeno 0-­‐9,12 de Samonella, y las células linfoides aisladas fueron probadas para determinar su adherencia bacteriana con cepas que comparten el inmunógeno 9 o 12, método demostrado sensible para la demostración de anticuerpos 0 en células linfoides. El término 0-­‐9,12 indica que los antígenos 9 y 12 se encuentran originalmente presentes juntos y no mezclados por el investigador. Se utilizaron ratas albinas cepa Sprague-­‐Dawley como animal de experimentación y varias cepas de Salmonella (S. typhi, S. berta, S. strasbourg, S. typhimurium, S. adelaide y S. minnesota), utilizando para el test de adherencia bacterias móviles. Se utilizó una modificación del medio de Helmreich, Kern y Eisen (486), que no requiere CO2 en la atmósfera para mantener el PH. El medio de cultivo para las células linfoides contenía un 10% de suero de rata bebé y el 90% restante era una solución de diversos compuestos entre ellos fosfato sódico, magnesio, calcio, potasio, hierro, glucosa, insulina, colina, ácido fólico, nicotinamida, rifoflabina, tiamina, etc… y varios aminoácidos esenciales como cistina o lisina. Posteriormente se llevó a cabo la preparación e inmunización de las células de los nódulos linfáticos poplíteos. Todas las ratas se inmunizaron mediante inyección de bacterias en las almohadillas de las patas traseras, repitiendo la inoculación entre la cuarta y la sexta semana, siendo sacrificadas cinco días después de la segunda inyección. Se eliminó un ganglio poplíteo para realizar una suspensión celular y obtener una concentración celular final de 1-­‐5 x 106/ml, del que se obtenían pequeñas gotas para micromanipulación. El estudio del fenómeno de la adhesión se llevó a cabo en células individuales. Se aislaron de cuarenta a ochenta células para probar la adherencia bacteriana. Cerca de 102 bacterias móviles fueron añadidas en cada gota. En algunas de la 304 RESULTADOS gotitas se observó que entre una y veinte bacterias se unieron a la célula porque permanecieron inmóviles. La adherencia entre célula y bacteria se ha demostrado que es una reacción entre antígeno bacterianos y anticuerpos unidos a las células, y se anotó la adherencia bacteriana evidenciada por la motilidad celular, siendo positiva si tenían más de cinco células adheridas (Figura 41). . Figura 41. Fenómeno de la adherencia. Una célula linfoide grande con bacterias adheridas y un linfocito grande y dos pequeños libres de bacterias. Tomado de: Mäkelä, 1964, (484) El fenómeno de la adherencia fue estudiado en grandes poblaciones de células, registrándose el número de células positivas y el número de campos escaneados. La titulación de anticuerpos séricos se valoró mediante la aglutinación a simple vista. En los resultados, Mäkelä (484) valora los anticuerpos del suero y el estudio de las células individuales. Los títulos de anticuerpos en suero fueron bajos durante los cinco días después de la segunda dosis de antígeno. Había algunas 305 RESULTADOS moléculas de anticuerpos capaces de reaccionar con el antígeno 9, pero no con el 12 y viceversa. En los estudios con células individuales todos los animales estudiados fueron inmunizados con el antígeno 0-­‐9,12 cepa SH-­‐116 y las células probadas en la adherencia bacteriana con la cepa SH-­‐130 (antígenos 1, 4, 5, 12), SH-­‐118 (antígenos 9, 46) y SH-­‐125 (antígenos 9, 12), intentado así buscar si las células tenían una reactividad más fuerte con el antígeno 9 que con el 12 o viceversa. La cepa SH-­‐130 se utilizó para detectar las células que pudieran producir la fracción anti-­‐12 y la cepa SH-­‐118 la fracción anti-­‐9. Solo aquellas células que fueron negativas a al adhesión en la primera cepa se probaron con la segunda, y solo las células negativas con la primera y segunda se probaron con la tercera cepa. Así se detectaron seis categorías distintas de células: • Células negativas a todas las cepas. • Células negativas al antígeno 9 y 12, pero positivas a 9,12. • Células negativas al antígeno 9 pero positivas al 12, siendo desconocida su actividad al 9,12. • Células negativas al antígeno 12 y positivas al 9, siendo desconocida su actividad al 9,12. • Células positivas al antígeno 9, siendo desconocida la actividad a los otros anticuerpos. • Células positivas al antígeno 12, siendo desconocida la actividad a los otros anticuerpos. En el 23 % de las células se pudo demostrar actividad a anticuerpos. Los resultados muestran que las bacterias pueden adherirse a células formadoras de anticuerpos siempre que compartan una parte del antígeno 0 del inmunógeno. La técnica no permitió una estimación exacta del número de células positivas para anti-­‐9 y anti-­‐12, pero si se pudo hacer una estimación estadística. Parece claro que un número considerable de las células probadas producían anti-­‐9 o anti-­‐12, pero ninguna se demostró que produjera ambas, 306 RESULTADOS aunque no se excluye que puedan existir dobles productores que contengan tan poco anti-­‐9 o anti-­‐12 como para causar un fenómeno de adherencia. Estos experimentos confirmaron que, por regla general, las bacterias se adhieren a las células de los ganglios linfáticos solo si comparten antígenos con la cepa inmunogénica. No se observaron reacciones cruzadas entre los antígenos 1, 4, 5, 12 y 9,46, los cuales están relacionados con 0-­‐9,12. Los fenómenos de adherencia observados, se supone son causados por una reacción entre un antígeno 0 de la bacteria de ensayo y el anti-­‐0 producido por la célula. Cuando el inmunógeno y la cepa de prueba no muestran antígenos 0 no había fenómeno de adherencia, aunque fuera Salmonella. Los resultados muestran que la adherencia puede ocurrir cuando comparten solo una parte del antígeno 0. A pesar de la incertidumbre sobre el número de células que reacciona tanto con 0-­‐9 como con 0-­‐12, la principal pregunta de este estudio parece ser contestada: hay células que solo reacciona con 0-­‐9 y otras que solo lo hacen con 0-­‐12. El autor (484) explica de esta forma los resultados: “Solamente dos explicaciones razonables puedo argumentar para la observación de que diferentes células producen anticuerpos contra diferentes fracciones de la molécula de endotoxina. Una es que la molécula se digiere antes de llegar a la célula formadora de anticuerpos, con lo que algunas células recibirían la parte 0-­‐ 12 y otras la 0-­‐9, pero esta alternativa parece poco probable ya que la endotoxina no es muy fácil de digerir. La explicación alternativa es que la calidad de los anticuerpos producidos por una célula se determina, no por el antígeno, sino por la célula y el papel del antígeno es forzar a la célula para hacer más anticuerpo y tal vez multiplicarse”. Y reconoce que: “El presente experimento no prueba esta alternativa, pero los resultados constituyen una prueba más a su favor”. Ya en los años cincuenta la quimioterapia ha adquirido importancia en la práctica clínica, pero es esta década cuando inicia su imparable crecimiento. En 307 RESULTADOS 1964, con el título: “Inhibición de la respuesta linfocitaria a la inflamación con antimetabolitos” publica Arthur R. Page de la Universidad de Minnesota, Minneapolis (487). Poco se sabe a cerca de la respuesta inflamatoria de las células mononucleares, existiendo varios estudios que indican que son en su mayor parte linfocitos que emigran de los vasos y se trasforman en macrófagos (210). Que la emigración de linfocitos no comienza hasta 3-­‐4 horas después de iniciada la inflamación sugiere que esta respuesta pueda depender también de sustancias producidas en el sitio de la inflamación, o puede ser que las 3-­‐4 horas de retraso en la parición de linfocitos, sea el tiempo requerido para responder a un estímulo en lugar de un retraso en la producción de un estímulo. El hallazgo de que el tratamiento con 6-­‐mercaptopurina puede eliminar completamente la respuesta de los linfocitos a la inflamación sin alterar la respuesta de los neutrófilos es interesante, ya que indica que la emigración de los linfocitos depende del metabolismo normal de las purinas (488). El presente estudio estudia el uso de fármacos que interfieren específicamente con la producción de ADN, ARN o proteínas. Se utilizaron conejos jóvenes albinos de ambos sexos para todos los experimentos y ratones adultos para los experimentos que implican administración de puromicina. Se administraron diversas drogas: aminopterina, 5-­‐fluoracilo, 8-­‐azaguanina, ciclofosfamida (cytosan), cloromicetina, actinomicina D y puromicina. La inflamación se provocó administrando una solución de clara de huevo al 10 % en medio salino y una pequeña cantidad de tinta china, y la biopsia fue tomada a las 4, 8, 12, 24 y 48 horas después de la inyección de esta solución para su estudio al microscopio. Se observó que la respuesta inflamatoria en los animales fue uniforme, neutrófilos y linfocitos estaban presentes en la zona perivascular en las muestras de biopsia de 4 horas. A las 24 horas, las células mononucleares estaban dispersas y habían cambiado de los pequeños linfocitos, 308 RESULTADOS predominantes a las 4 horas a los grandes macrófagos. Una droga fue considerada efectiva cuando provocaba un bloqueo selectivo de los linfocitos en la respuesta inflamatoria. Se eligieron fármacos con la posibilidad de ser administrados por un período de 10 a 14 días y se determinó la dosis tóxica antes de probar su efecto sobre la respuesta inflamatoria. Cada animal tratado con el fármaco fue emparejado con un animal de control no tratado y las muestras de biopsias fueron evaluadas sin conocer su origen. Los conejos tratados con aminopterina, 5-­‐fluoracilo o ciclofosfamida administradas 14 días antes de la provocación de la inflamación presentan una alta tasa de mortalidad debido a la toxicidad de las drogas, y ninguno de los animales supervivientes al inicio de la inflamación mostraron anomalías en la migración celular. Los tratados con 8-­‐azaguanina y cloranfenicol tuvieron una respuesta normal a la inflamación. La puromicina igual que el cloranfenicol fue elegido debido a su capacidad para interferir con la incorporación de aminoácidos en la cadena polipeptídica. Puromicina se administró a ratones y la respuesta de los linfocitos a la inflamación fue bloqueada en todos los animales. Con actinomicina D, 2 conejos sobrevivieron durante 10 días a una dosis de 30 µg por kilo y día. Ambos mostraron una caída en el 50 % de los linfocitos circulantes y ambos un bloqueo total en la participación de los linfocitos en la inflamación. La inflamación fue normal cuando la droga se administra al mismo tiempo que el estímulo inflamatorio, pero la respuesta fue bloqueada, con ausencia de linfocitos, en los dos animales cuando se administra 72 horas antes del estímulo inflamatorio. A continuación se repitió el experimento pero utilizando una única dosis superior a 0,375 mg/kilo. La respuesta linfocitaria fue completamente bloqueada en todos los animales cuando la inflamación fue iniciada al mismo tiempo o en cualquier momento después de la administración de la droga. Todos estos animales a los que se les había administrado esta gran dosis murieron en 72 horas. 309 RESULTADOS El siguiente experimento fue diseñado para probar si una reacción inflamatoria preexistente se vería afectada por la actinomicina D en la emigración de linfocitos en otro sitio del mismo animal. Se mostró que algunos de los linfocitos circulantes en un animal con una reacción inflamatoria establecida son resistentes al bloqueo de la actinomicina D y capaz de participar en una respuesta inflamatoria inducida en otro lugar del animal, incluso en presencia de actinomicina D. Las muestras de biopsia de estas segundas reacciones fueron normales en dos casos, en otro caso los linfocitos estaban presentes pero en número disminuido. Estas observaciones sugieren que algunos de los linfocitos circulantes en un animal con una reacción inflamatoria establecida son resistentes a la acción de bloqueo de la actinomicina D y capaces de participar en una respuesta inflamatoria inducida. Las drogas aminopterina, 5-­‐fluoracilo y ciclosfamida fueron elegidas como fármacos que actúan principalmente por interferir con la síntesis de ADN. El hecho de que los niveles tóxicos de estas drogas fueran ineficaces en el bloqueo de los linfocitos, indica que la replicación del ADN no es un componente esencial en las primeras fases de la inflamación. La actinomicina D tiene una acción específica en el bloqueo de la síntesis de ARN. Cloromicetina y puromicina son fármacos que inhiben directamente la síntesis de proteínas por interferencia con la incorporación de aminoácidos en la cadena polipeptídica. La observación de que tanto actinomicina D como puromicina fueron eficaces en el bloqueo de la respuesta linfocitaria a la inflamación indica que la producción de proteínas es necesaria para la respuesta inflamatoria normal. Puromicina interfiere directamente con la producción de proteínas y actinomicina D lo hace indirectamente a través del ARN mensajero. En definitiva (487), en los ensayos realizados con Actinomicina D, 5-­‐fluoracilo, cytosan, 8-­‐azaguanina, cloromicetina, aminopterina, y puromicina para determinar la capacidad de alterar una respuesta inflamatoria, solamente actinomicina D y puromicina bloquearon específicamente la participación de linfocitos en la respuesta inflamatoria, los otros fármacos no tuvieron efecto. 310 RESULTADOS Una gran dosis de actinomicina D fue inmediatamente efectiva en el bloqueo de la respuesta de los linfocitos a la inflamación en conejos normales, pero fue ineficaz si una lesión inflamatoria previamente establecida estaba presente. El trabajo de Page (487) nos anticipa el camino que seguirá la quimioterapia en el futuro. El autor pretende en su trabajo obtener datos sobre la eficacia selectiva, en este caso sobre linfocitos, de diversos antimetabolitos y su relación con el mecanismo de acción de los mismos: “el hallazgo de que el tratamiento con 6-­‐mercaptopurina puede completamente eliminar la respuesta de los linfocitos a la inflamación sin alterar la respuesta de los neutrófilos, es interesante ya que indica que la emigración de los linfocitos depende del metabolismo normal de las purinas”. El conseguir compuestos quimioterapéuticos cada vez más específicos, o lo que es lo mismo hacer una terapia selectiva, más eficaz y con una menor repercusión en tipos celulares no deseados, son objetivos prioritarios en la nueva quimioterapia (489). Un primer artículo sobre quimioterapia aparece en 1910: “La quimioterapia de Ehrlich, una nueva ciencia” (490). La principal contribución de Ehrlich a la ciencia fue la teoría de las cadenas laterales, que establece las bases químicas para la especificidad de la respuesta inmunológica y explica como los receptores de las células se combinan con toxinas para producir cuerpos inmunes capaces de combatir la enfermedad. Pero Ehrlich también fue pionero en el campo de la quimioterapia, descubrió métodos para colorear leucocitos y sustancias con toxicidad selectiva como el descubrimiento en 1901 del compuesto 606 (fruto de 606 experimentos), que él mismo llamó la bala mágica o salvarsán. El 606 era una preparación de arsénico orgánico empleada en el tratamiento de la sífilis, mataba las espiroquetas pero en cambio resultaba inocuo para el cuerpo humano, es decir presentaba toxicidad selectiva (491). El método de trabajo del 606 sirvió como base y modelo para la obtención de nuevos preparados, siendo una de las técnicas que se emplean actualmente en la búsqueda de nuevos antibióticos y compuestos por parte de la industria farmacéutica. Para Ehrlich el 606 se trataba de: “un instante de buena suerte frente a siete años de desgracias”. 311 RESULTADOS La quimioterapia, se desarrolló a principios del siglo XX, aunque no fue pensada originalmente como tratamiento contra el cáncer. Las primeras publicaciones sobre quimioterapia eran en relación a la sífilis, como el trabajo de Nichols (492) en 1911: “Observaciones adicionales sobre ciertas características de la sífilis experimental en el conejo” o el de G M. Findlay (493) en 1932: “Quimioterapia: nuevos recursos en el tratamiento de la sífilis”. También la temida tuberculosis fue susceptible de tratamiento con quimioterapia, como lo atestiguan las numerosas publicaciones, entre ellas, las de G. Koga de 1916 (37), y con quimioterapia se trataban numerosas infecciones bacterianas (494). Del año 1922 es un artículo sobre “Las acciones organotróficas, bacteriotróficas y leucocitotróficas de determinados productos químicos orgánicos” de L D. Felton (495). Pero la era de la quimioterapia, tal como se conoce hoy día, se inició en los años cuarenta, los con los primeros usos del gas mostaza y drogas antagónicas del ácido fólico en el tratamiento del cáncer (496). Los primeros antimetabolitos usados fueron aminopterina y amethopterina (metotrexato), que indujeron la remisión en niños con leucemia linfoblástica aguda, estableciéndose así el siguiente principio: “los antimetabolitos pueden suprimir la proliferación de células malignas y reestablecer el funcionamiento normal de la médula ósea”, proclamado por defensores de la quimioterapia como Farber (497)(498). Farber se encontró con una gran resistencia para proseguir sus estudios, pues la comunidad médica tenía muy asumido que la leucemia era incurable. En 1948, un artículo de Farber (499) fue víctima del más absoluto ridículo. Una década más tarde, en el Instituto Nacional contra el cáncer de Estados Unidos (NCI) se descubrió que el tratamiento con metotrexato podía curar el coriocarcinoma, el primer tumor sólido curado con quimioterapia (500). En esta década, los años sesenta, ocurre otro importante avance en la terapia contra el cáncer. J. Holland, Freireich y Frei formulan la hipótesis de que la quimioterapia podría seguir una estrategia similar a la usada por la terapia antibiótica contra la tuberculosis: la combinación de drogas cada una con un 312 RESULTADOS mecanismo de acción diferente (501)(502). Método que se extendería al tratamiento de los linfomas Hodgkin y No-­‐Hodgkin (pauta MOPP). En 1964, desde el Instituto de Patología de las Fuerzas Armadas de Washington, Robert A. Prendergast (503) publica sus observaciones en relación a la especificidad celular: “La especificidad celular en la reacción al homoinjerto”. Los trabajos de Najarian y Feldman (504) habían demostrado la posibilidad de una transferencia específica en el rechazo de homoinjertos de piel en ratones y cobayas previamente no sensibilizados. Estos autores estudiaron que la transferencia pasiva de células linfoides (marcadas con timidina) sensibilizadas a tejidos homólogos en huéspedes isólogos no sensibilizados, provocaba un rechazo acelerado de los injertos de piel en los destinatarios. En sus varios experimentos, en ningún caso se encontró un número significativo de células marcadas en el sitio del rechazo del injerto, a pesar de que la transferencia se había hecho con un gran número de células. Este rechazo de tejidos homólogos sin la presencia física de las células sensibilizadas en el sitio del injerto sugirió que un agente humoral producido por la célula podría ser el responsable del rechazo. Estudios similares, sin embargo, no pudieron demostrar este punto, quedando en duda el papel del infiltrado mononuclear en las reacciones de hipersensibilidad retardada y en el rechazo de homoinjertos, que cuestiona la especificidad inmunológica de las células implicadas. Esta controversia sobre la especificidad inmunológica es el estímulo para que Prendergast inicie su trabajo, donde se investiga también la cinética celular implicada en la reacción. Prendergast (503) idea una técnica para el etiquetado permanente de las células dentro de los ganglios linfáticos regionales del drenaje de un homoinjerto. La instilación de pequeñas cantidades de timidina tritiada en la base del injerto aseguraría el marcado en esta población celular por vía aferente linfática, mientras el resto de las células del hospedador permanecerían sin marcar. 313 RESULTADOS Se utilizaron conejo de cepas chinchilla, albino y holandés. Injertos de 2x1 cm fueron intercambiados en las superficies dorsales de las orejas de los albinos y cinchilla, al mismo tiempo una segunda transferencia entre el mismo par de conejos y un injerto del conejo cepa holandés fueron colocados en el oído opuesto. Todos los injertos fueron de espesor completo de piel y manejados con una técnica aséptica. Injertos de control fueron intercambiados entre diferentes cepas, siendo examinados todos con frecuencia para identificar signos de rechazo o fracaso en la cura. Después de un periodo de curación de dos días, se inició una serie de dos inyecciones diarias de timidina tritiada en la base del injerto primario. Con base en evidencia macroscópica e histológica, los injertos fueron rechazados en el 7º a 8º día, siendo sacrificados los animales en el día del rechazo. Los tejidos extirpados se estudiaron histológicamente, así como los ganglios linfáticos cervicales, bazo, intestino delgado y los injertos en su espesor completo. Se realizaron autorradiografias en secciones de tejidos tratadas con tinciones adecuadas para su estudio. El porcentaje de células marcadas en los ganglios de drenaje y bazo se determinó contando 1000 células mononucleadas en cada tejido. 314 RESULTADOS Figura 42. La especificidad celular en la reacción al homoinjerto. Diseño general del experimento. Tomado de: Prendergast, 1964 (503) Prendergast (503) valora la apariencia del injerto primario, observando en todos los casos el patrón normal del rechazo. Las autorradiografias de este material mostraron que no había grandes cantidades de células mononucleares marcadas, que en el ganglio linfático de drenaje fue entorno al 18 %. La mayoría de las células marcadas se encontraron en la zona de la corteza reactiva subyacente al seno linfático, en el estroma profundo y en una alta proporción en las células endoteliales. El estudio histológico de rutina de los ganglios linfáticos no reveló ninguna diferencia entre los ganglios de drenaje del injerto primario y aquellos que drenaban los injertos de los controles. Esto indica que la manipulación del injerto primario mediante la inyección de múltiples dosis de isótopo no provocó una estimulación inespecífica del nodo de drenaje. 315 RESULTADOS Respecto a las células marcadas en injertos homólogos y de control, la mayor concentración de células marcadas tanto en homólogos como en los controles, se encontraban en los pequeños vasos sanguíneos dentro del injerto y en el tejido de fondo del injerto. Las características morfológicas y la posición de las células marcadas fueron similares en todos los aspectos a los observados en controles. Las células marcadas dentro de los pequeños vasos y en el infiltrado perivascular fueron pironina positivas, pero esta característica tinción se fue perdiendo a medida que nos alejamos de los vasos. No hubo evidencia de células marcadas en otros tejidos con actividad mitótica intensa, así el examen del epitelio intestinal, centros germinales de ganglios linfáticos y bazo no revelaron la presencia de células marcadas a pesar de la existencia de un gran número de figuras mitóticas en estos lugares. Ocasionalmente se observaron células marcadas en tejido linfoide distante de los sitios de rechazo. En secciones e improntas de linfáticos que drenan las zonas del injerto, el número de estas células fue algo mayor que el material obtenido de ganglios y bazo distantes al injerto. Ninguna de las células pironinofílicas activas mitóticamente de los nodos se marcaron y los frotis de sangre periférica mostraron de una a dos células mononucleares marcadas por mil. En la discusión, Prendergast (503) asegura que sus hallazgos están en contraste con los de Najarian y Feldman (504), que no pudieron demostrar la presencia de un número significativo de células marcadas en los sitios del injerto en ratones y cobayas. Una posible explicación de esta aparente disparidad podría estar relacionada con la vida útil de las células marcadas en el sitio del rechazo. Si este tiempo es demasiado corto, el porcentaje de células marcadas podría ser extremadamente pequeño. En el marco de las condiciones del estudio de Prendergast, un porcentaje relativamente constante de células marcadas desde el nodo estimulado sustituyen a las que ya han muerto o se han dividido. 316 RESULTADOS Estos resultados están de acuerdo con los datos publicados de estudios en relación con la transferencia de la hipersensibilidad retardada (505). Esta difusión del conocimiento inmunológico podría ser explicada por el paso libre de células linfoides desde los ganglios linfáticos a la circulación general y de nuevo al tejido linfoide como lo describió Gowans (336). El hallazgo de un pequeño número de células linfoides en ganglios y bazo distantes confirma esta recirculación de células linfoides. Otros estudios (506) han demostrado el rechazo acelerado específico en ratones, cobayas y conejos cuando se transfieren células sensibilizadas limitadas en cámaras de Millipore y por tanto sin acceso al injerto. Dado que el contacto directo entre las células sensibilizadas y el injerto no es necesario para lograr el rechazo acelerado, se deduce que algún factor humoral producido por las células de las cámaras tienen que haber sido el responsable del rechazo o ha trasferido sensibilidad a las células del huésped. El presente estudio (503) ofrece evidencia directa de que las células de los ganglios linfáticos regionales del drenaje del homoinjerto de piel establecido en un primer tiempo, participan en el infiltrado linfoide en sitios distantes del rechazo del injerto. Cantidades adecuadas de timidina se limitaron al nodo regional reactivo, y fue posible detectar la aparición de estas células marcadas en sitios remotos del mismo animal, obviando la necesidad de la transferencia de células. Bajo estas condiciones, los recuentos diferenciales de células linfoides no mostraron un predominio de células marcadas en el infiltrado acompañante del homoinjerto comparado con el control en el rechazo del injerto. Resultados que sugieren que la especificidad de la reacción de homoinjertos es probablemente dictado por algún factor humoral que actúa en consonancia con células linfoides para lograr el rechazo del injerto. A principios de esta década algunos estudios como los de Pearse o Glick inciden en las diferencias existentes entre timo y “la bursa”. Raymond D.A Peterson y Robert A. Good (507) presentan en 1965 uno de los trabajos relacionados con este tema: ”Diferencias morfológicas y de desarrollo entre las células del timo y de la bolsa de Fabricio del pollo”. 317 RESULTADOS El pollo tiene dos órganos linfoides distintos asociados al intestino, el timo y la bolsa de Fabricio. A pesar de que estos dos órganos se encuentran en extremos opuestos del tracto gastrointestinal, tienen ciertas similitudes que han llevado a los investigadores a considerarlos juntos. Los lóbulos del tejido linfoide que caracterizan a ambos se denominan “linfoepiteliales” porque parece surgir del epitelio que recubre el intestino. Microscópicamente los lóbulos de ambos consisten en una corteza de oscuros linfocitos y una médula con grandes células reticulares. Estas similitudes no tuvieron un significado biológico real hasta que Glick (508) descubrió el papel funcional de la bolsa de Fabricio, demostrando que la extirpación temprana de ella interfería con la posterior capacidad del pollo para formar anticuerpos circulantes. En cambio, se observó que la timectomía neonatal no afectaba a la capacidad del pollo para producir anticuerpos circulantes, aunque si tenía algún efecto sobre el desarrollo de la hipersensibilidad celular (509)(510). Auerbach y Ball (511)(512) demostraron que en el ratón, los linfocitos del timo cambian de grandes a pequeñas células durante su maduración y que este cambio de tamaño proporciona un índice útil de diferenciación. Los datos presentados por Peterson y Good (507) sugieren que el desarrollo del timo en el pollo se caracterizan por la maduración temprana de la población linfoide, que ocurre antes de la eclosión, mientras que la maduración de la células linfoides de la bolsa de Fabricio comenzaría más tarde, es más lenta y continúa más allá de la eclosión. Huevos de pollo fértiles fueron incubados a 37ºC, de ellos de 8 a 12 fueron retirados para estudio histológico a intervalos entre el décimo y vigésimo primer día de incubación. Pollos recién eclosionados fueron sacrificados a diferentes edades desde el primer día hasta la edad de cuatro meses. El timo y la bolsa de Fabricio eran retirados de cada pollo y preparados tanto para su estudio histológico como para el análisis de la distribución del tamaño celular de la población celular, siguiendo el método de Auerbach y Ball y utilizando un contador Coulter electrónico (513), que determina el volumen celular 318 RESULTADOS individual automáticamente y automáticamente recoge el número de células de diversos tamaños. Figura 43. Timo y bolsa de Fabricio de un pollo recién nacido. El timo (T) consiste en varios lóbulos unidos a cada arteria carótida. La bolsa de Fabricio (B) es una evaginación de la pared dorsal de la cloaca. Tomado de: Peterson y Good, 1965 (507) Peterson y Good (507) observaron que las células del timo desarrollaban sus características linfoides entre los días 12 a 14 de incubación y en ese momento se podían separar fácilmente del estroma para analizar su tamaño. Advierten que las células se van haciendo progresivamente más pequeñas, y entre el día 15 y 17 ocurre un importante cambio: las células adquieren su tamaño celular definitivo. En la bolsa de Fabricio el tejido linfoide aparece como grupos de células en racimos a lo largo de las criptas endodérmicas aproximadamente a los 14 días de incubación. Estos grupos más tarde serán los folículos linfoides que caracterizan la bolsa completamente desarrollada, y este tejido linfoide conserva su íntima relación con el estroma intestinal, a diferencia del tejido 319 RESULTADOS tímico. El examen microscópico reveló un predominio de células grandes, tanto a principio de la embrionación como durante toda la vida de la bolsa. El tamaño celular conseguido a los 17 días de incubación caracteriza el tamaño de la célula del timo por lo menos durante los primeros cuatro meses de la vida del pollo. La bolsa, por el contrario, se desarrolla plenamente después de la eclosión, y sus linfocitos permanecen grandes durante toda la vida de la bolsa. Por tanto existen dos diferencias entre las células linfoides de timo y bolsa de Fabricio: 1-­‐ El tiempo de desarrollo embriológico. 2-­‐ El tamaño. El tamaño de los linfocitos es una indicación de su etapa de diferenciación, las células más grandes están menos diferenciadas que las más pequeñas, y la dificultad de los linfocitos de la bolsa para abandonar el estroma cuando se agitan, también puede ser una manifestación de falta de madurez. Ackerman (514) demostró la presencia de desmosomas que conectaban las células epiteliales de la bolsa de Fabricio unas con otras y con las formas inmaduras de linfocitos. El tamaño de las células y la adhesividad son por tanto dos parámetros útiles que reflejan la diferenciación. Las diferencias morfológicas y de desarrollo entre ambos órganos merecen especial atención, pueden ser la clave para la comprensión de las diferencias funcionales aparentes entre timo y bolsa de Fabricio que se han dado a conocer gracias a los experimentos de ablación de los mismos. Si el timo ha pasado por sus principales etapas de desarrollo antes de la eclosión, cosa que sugieren los datos, también puede haber ejercido su principal función inmunológica antes de ese tiempo. Existen evidencias considerables de que las células son susceptibles a virus oncogénicos solo durante una etapa crítica de su diferenciación. En el momento de la eclosión la bolsa puede tener células linfoides susceptibles, mientras que los linfocitos tímicos, siendo más maduros en este momento ya no serían susceptibles a un virus inyectado en el período 320 RESULTADOS neonatal. Si el pollo se infecta en una etapa temprana de la embrionación, las células del timo podrían ser involucradas y la bursectomía neonatal por sí sola no sería capaz de prevenir la enfermedad. Los autores (507) consideran que: “Las diferencias funcionales entre timo y bolsa pueden ser medidas por las diferencias en el tiempo de desarrollo embriológico y el grado de maduración de los linfocitos en los dos órganos. Una explicación alternativa para las diferencias funcionales del timo y bolsa podría ser que las células de los dos órganos son básicamente diferentes. Los linfocitos tímicos pueden diferenciarse hacia un tipo celular y los de la bolsa hacia otro. Mantenemos esta posibilidad hasta que nuevos métodos permitan estudiar a estas células y sus derivadas”. Procedente de la Facultad de Medicina de la Universidad de Kyoto es esta publicación de 1966 de Masaichi Fukase (515): “Estudio sobre células responsables de hipersensibilidad retardada con especial referencia a los linfocitos”. En los años cuarenta, MW Chase (516) ya indicó que la hipersensibilidad retardada era transferida por las células vivas de los tejidos linfáticos, hecho confirmado posteriormente por numerosos investigadores (517)(518), lo que sugiere que los anticuerpos responsables de la hipersensibilidad retardada son de naturaleza celular. Sin embargo sigue siendo motivo de controversia qué tipo o tipos celulares de los tejidos linfáticos están involucrados y cómo participan. Los intentos realizados hasta ahora para resolver el problema se han abordados de distintas formas: 1-­‐ Pruebas de transferencia pasiva con linfocitos, neutrófilos y células de exudado peritoneal. 2-­‐ Neutralización de la tuberculina por linfocitos (vivos y modificados). 3-­‐ Intentos de romper o alterar la tolerancia inmunológica adquirida a la tuberculina mediante linfocitos 321 RESULTADOS 4-­‐ Desarrollo de hipersensibilidad a la tuberculina en conejos timectomizados. Los resultados de estos experimentos sugieren que son los linfocitos los responsables de esta hipersensibilidad. Fukase da un nuevo enfoque al problema mediante el trasplante de linfocitos puros, granulocitos y células peritoneales, en lugar de utilizar mezclas de diversos tipos celulares, en un intento de transferencia pasiva de hipersensibilidad. Su propósito es determinar el tipo celular implicado en la producción de anticuerpos en la hipersensibilidad retardada y aclarar el mecanismo por el cual estas células participan en el proceso. Conejos y cobayas son empleados en experimentos sobre la hipersensibilidad a la tuberculina y a dinitroclorobenceno (DNCB) respectivamente. Utiliza los mismos métodos de sensibilización a tuberculina y DNCB que ya utilizó en trabajos anteriores (519). Se realizaron las pruebas pertinentes para valorar la hipersensibilidad a tuberculina y a DNCB, y los procedimientos y criterios seguidos para valorar su neutralización (519). La inducción de la tolerancia inmunológica a la tuberculina fue realizada en el conejo mediante una única inyección de bacilos tuberculosos muertos por calor y suspendidos en aceite de parafina líquida. Conejos tolerantes fueron utilizados como receptores de linfocitos de 5-­‐6 semanas después de comprobar su no respuesta a al tuberculina tras la exposición a bacilos tuberculosos. Una tolerancia inmunológica nativa a la tuberculina en los conejos es bien conocida, mostrando éstos una gran variación individual, desde muy fuerte a muy débil o incluso negativa. De 150 conejos, seis no desarrollaron hipersensibilidad a la tuberculina, es una tolerancia inmunológica innata, la llamada “tolerancia inmunológica materna”. Conejos recién nacidos (RN) fueron timectomizados a las 24 horas del nacimiento realizándose un seguimiento semanal del número de linfocitos en ellos, comparándolo con el de conejos normales de la misma edad. Los conejos 322 RESULTADOS se dividieron en grupos y la sensibilización a la tuberculina se realizó a diferentes edades, entre 3 y 10 semanas después del nacimiento. Fukase (515) sigue cuatro vías para intentar detectar la célula o células responsables de la hipersensibilidad retardada: ! Transferencia pasiva con linfocitos sensibilizados, neutrófilos y células mononucleares de exudado peritoneal. ! Pruebas de neutralización a la tuberculina por linfocitos sensibilizados in vitro. ! Intentos de rotura de la tolerancia nativa y adquirida a la tuberculina con linfocitos no sensibilizados. ! Desarrollo de la hipersensibilidad a la tuberculina en conejos timectomizados. Se observó que la hipersensibilidad de tipo retardado a la tuberculina o a DNCB era 100 % transferible desde un donante sensibilizado a un destinatario no sensibilizado por linfocitos o células del exudado peritoneal, pero no transferible por neutrófilos. Es razonable concluir que en el mecanismo de transferencia de la hipersensibilidad estaban implicados no solo linfocitos, sino también otras células mononucleares (probablemente reticuloendoteliales). Estas células mostraron su capacidad de transferencia solo cuando estaban vivas, ya que la perdieron por vibración ultrasónica y por congelación– descongelación. La aplicación de suero obtenido a partir de donantes fuertemente sensibilizados, o de linfocitos o células del exudado peritoneal de donantes no sensibilizados no tuvo efecto sobre la sensibilidad de los receptores a tuberculina o a DNCB. También se observó una pérdida de la tolerancia inmunológica adquirida por transferencia de linfocitos vivos o alterados no sensibilizados. Los linfocitos obtenidos de conejos no sensibilizados se administran vía intravenosa a los destinatarios en dos ocasiones con intervalo de una semana, posteriormente bacilos tuberculosos muertos por calor se administran vía intramuscular a los 323 RESULTADOS destinatarios. Dos de tres conejos tolerantes desarrollaron hipersensibilidad a la tuberculina con un retraso de 4-­‐6 semanas, pero una semana después la hipersensibilidad se había desvanecido. Al igual que la tolerancia adquirida, la pérdida de la tolerancia inmunológica nativa fue provocada por la transferencia de linfocitos y células de ganglios linfáticos. Cuando los conejos tolerantes recibieron de 2-­‐4 transferencias de linfocitos o células de ganglios linfáticos, y fueron puestos a prueba nuevamente con bacilos tuberculosos, mostraron hipersensibilidad a la tuberculina con una duración de varias semanas, igual que ocurrió en los conejos con la tolerancia adquirida. Figura 44. Transferencia pasiva. La transferencia pasiva de la hipersensibilidad a la tuberculina a través de los linfocitos. Tomado de: Fukase, 1966 (515) 324 RESULTADOS Queda pues claro que el tipo de tolerancia nativa o falta de respuesta nativa a la tuberculina se modifica y trasforma en capacidad de respuesta con el trasplante de linfocitos normales o células de ganglios linfáticos, resultado que sugiere que las diferencias individuales en la respuesta inmunológica depende de la función de los linfocitos. Miller en 1961 (64) señaló que el timo juega un importante papel en el desarrollo de la respuesta inmunológica. Muchos investigadores ya lo han confirmado y presentado evidencia adicional de que el timo también participa en el desarrollo del tejido linfático, en particular en la producción de linfocitos. El desarrollo de la hipersensibilidad a la tuberculina se inhibió en conejos timectomizados antes de las seis semanas después del nacimiento, mientras que la mayoría de conejos normales y los timectomizados después de las siete semanas del nacimiento desarrollaron hipersensibilidad. La aparición de hipersensibilidad en un conejo timectomizado, sin embargo no siempre se correlaciona con un aumento en el número de linfocitos, en consecuencia es posible que la respuesta inmunológica de un animal dependa no solo del número de linfocitos sino también de la maduración inmunológica de éstos. En cualquier caso, asegura Fukase: “estos resultados proporcionan evidencia adicional de que los linfocitos son responsables de la hipersensibilidad a la tuberculina”. El autor (515) considera necesaria una nueva concepción del mecanismo de transferencia pasiva y pérdida de la tolerancia para comprender estos resultados. Cuando los linfocitos vivos se transfieren del donante al receptor, casi todos los linfocitos llegan a los órganos linfáticos, tal como Gowans (336) había indicado unos años antes. Los linfocitos fuertemente sensibilizados transferidos darán alguna información sobre la producción de anticuerpos a un número de células del propio receptor inmunológicamente competentes, células que se sensibilizarán de manera débil e incompleta, pero podrán reaccionar con la tuberculina y dar lugar a una reacción positiva en la piel. Sin embargo, si algún antígeno pasa con los linfocitos transferidos, las células del receptor se sensibilizan con mayor rapidez y con tanta fuerza como en el caso 325 RESULTADOS de la respuesta secundaria a un antígeno. Por otro lado cuando linfocitos sensibilizados rotos se transfieren al receptor, la información facilitada por ellos debe ser mucho menor que en caso de los vivos y el receptor sigue siendo negativo a la tuberculina, pero si el antígeno se introduce, las células débilmente sensibilizadas se sensibilizan con tanta fuerza y tan rápidamente como en una respuesta secundaria. Posible explicación del mecanismo de transferencia pasiva que ratifica las observaciones de Fukase: “Podemos decir que los linfocitos son responsables de la hipersensibilidad a la tuberculina” Tumores e inmunidad, un vínculo hasta ahora no totalmente comprendido es motivo de estudio para Fairley (520) en 1967, del Hospital de San Bartolomé en Londres: “La resistencia humana a los tumores”. En las enfermedades malignas existen dos problemas inmunológicos distintos. En primer lugar, está el efecto que la enfermedad maligna pueda tener sobre los mecanismos de la inmunidad normal, y en segundo lugar existe la posibilidad de una reacción inmune del anfitrión contra su propio tumor, que nos lleva a la cuestión de si el comportamiento maligno de la enfermedad puede ser modificado por procedimientos inmunológicos. Debido a las dificultades de este tipo de investigaciones en el hombre, hasta ahora la mayoría de trabajos realizados han sido hechos en animales. Fairley estudia el efecto de las enfermedades malignas en las reacciones inmunológicas a antígenos extraños. El deterioro del sistema inmune podría predisponer al desarrollo de la enfermedad maligna, y es por ello necesario considerar por separado los problemas en la formación de anticuerpos y los referentes a la inmunidad celular. Respecto a la formación de anticuerpos circulantes, dado que los anticuerpos se producen por células de la serie linfo-­‐ plasmocitaria no es de extrañar que sea en las enfermedades malignas primarias de estas células donde la formación de anticuerpos esté más gravemente afectada. La producción de anticuerpos en una respuesta primaria (el paciente recibe un antígeno que desconoce) está deteriorada en algunos casos tardíos de carcinoma y en estadios tempranos de enfermedades malignas 326 RESULTADOS primarias del tejido linforeticular, siendo el déficit inmunológico más marcado en las enfermedades malignas de linfocitos y células plasmáticas. Cuando probamos la respuesta secundaria (el paciente recibe un antígeno ya conocido) se observa una respuesta en la formación de anticuerpos muy deficiente y alterada en la leucemia linfocítica crónica y mielomatosis, mientras que la respuesta en otras reticulosis e incluso en casos avanzados de carcinoma es generalmente normal (521). La inmunidad celular ha sido estudiada midiendo las reacciones de hipersensibilidad retardada, rechazo de homoinjertos y mediante el test de transferencia de linfocitos de Brent y Medawar (522). Fairley expone los diversos trabajos de autores como Lamb (523), Hughes, Mackay, Levin (524), Robinson y Hochman (525) que ponen de manifiesto la existencia de una incompetencia inmunológica temprana en pacientes de reticulosis pero tardía en el caso de sarcoma o carcinomas. Si el deterioro de estas respuestas permite avanzar a la enfermedad o es simplemente el resultado de tal avance sigue siendo una incógnita. La reacción inmune del paciente contra su propio tumor se tiene que considerar en el contexto de la capacidad de los pacientes con enfermedades malignas para responder a un ataque con antígenos extraños, y existe la posibilidad de una reacción por parte del paciente contra su propio tumor, reacción que es más bien celular que humoral y que al igual que ocurre en los animales está mediada por linfocitos. Esto es un hecho evidenciado en muchos estudios sobre injertos tumorales en el hombre. Respecto a los homoinjertos tumorales, Southam (526) demostró que el trasplante subcutáneo de células cancerosas en personas normales, casi siempre tiene como resultado un crecimiento transitorio seguido de una regresión completa a la 3ª-­‐4ª semana, sin embargo, bajo ciertas condiciones, el homoinjerto de células malignas puede ser aceptado, y esto es particularmente probable cuando el receptor ha recibido fármacos inmunosupresores o cuando existe algún grado de histocompatibilidad entre donante y receptor, o si el receptor es de edad avanzada. Y se considera que es realmente peligroso en ciertas circunstancias 327 RESULTADOS trasplantar tejido maligno en receptores sanos además de éticamente injustificable. En los autoinjertos, la implantación subcutánea de células malignas o material de biopsia del propio tumor puede dar lugar a implantes dependiendo de las condiciones experimentales. Southam (527) encontró que el número de células necesarias para producir un autoinjerto con éxito estaba relacionado con la capacidad del paciente para rechazar un homoinjerto de células tumorales, lo que a su vez estaba relacionado con el estado inmunológico del paciente como se puede medir con la prueba de la tuberculina, la capacidad de desarrollar hipersensibilidad retardada a DNFB y la respuesta de los macrófagos en los exudados inflamatorios. Según Southam: “cuanto mayor sea el deterioro inmunológico, el número de células necesarias es menor para un autotrasplante tumoral”. También encontró que con la mezcla de células tumorales con los propios linfocitos del paciente había a menudo una inhibición del crecimiento en el sitio de la inoculación, lo que apoya firmemente la idea de que los linfocitos están relacionados con mecanismos de defensa tumorales. Fairley (520) analiza los estudios existentes sobre histología tumoral y el fenómeno de la regresión. La historia natural de la enfermedad maligna está relacionada con la morfología del tumor. Black y Speer (528) observaron que el pronóstico del carcinoma de mama y estómago se relaciona con el grado de diferenciación nuclear, la intensidad de la infiltración linfoide en el tumor primario y la presencia de histiocitosis sinusal en los ganglios linfáticos regionales. Cuanto mayor es el grado de diferenciación, infiltrado linfoide e histiocitosis sinusal mejor es el pronóstico. Lukes (529) demostró que una situación similar ocurría en la enfermedad de Hodgkin, donde la ausencia de linfocitos está asociada con mal pronóstico y enfermedad generalizada, en cambio, la proliferación linfoide e histiocítica se asocia a enfermedad localizada y un mejor pronóstico. Respecto a el fenómeno de la regresión espontánea están los datos aportados por Everson, Cole, Burchenal (530)(531) que recogieron los casos de 132 328 RESULTADOS leucemias agudas cuyos pacientes sobrevivieron más de cinco años, algunos de los cuales parecían haberse curado. En el caso del linfoma de Burkitt africano han tenido lugar remisiones de masas tumorales con ausencia de recurrencia con una única inyección de metotrexato o ciclofosfamida (532), siendo muy poco probable que el tratamiento haya acabado con todas las células malignas y ya que los resultados sugieren la existencia de células resistentes, se puede pensar que el mecanismo de eliminación tumoral podría ser inmunológico. Tras la experiencia seguida en los tumores primarios de rata descrita por el profesor Alexander (533), Nadler y Moore (534) trasladaron dicho estudio a pacientes con enfermedades malignas incurables. Una biopsia de tejido maligno fue tomada de cada paciente e implantada vía subcutánea en otro y viceversa. Después de 10 a 14 días se recogieron células blancas de los destinatarios de los injertos tumorales y se inyectaron de nuevo en los donantes. De 26 pacientes 7 se beneficiaron de este procedimiento y en 2 pacientes se obtuvo una completa remisión objetiva. “Por vez primera ahora parece que puede ser posible influir en el curso de la enfermedad maligna utilizando linfocitos inmunes, y no hay duda de que una mayor investigación de la actividad de los linfocitos en la enfermedad maligna aportará una información esencial sobre la resistencia a los tumores en el hombre”, con esta argumentación de Fairley, que parece más la de un visionario, se abre la puerta a todo un mundo sumamente complejo que poco a poco va a ir tomando cabida en varias ramas de especialidades tales como la oncología, inmunología, inmunoterapia, patología, las nuevas técnicas de biología molecular y genética como principales, que se van a conexionar y enlazar progresivamente hasta convertirse en todo un universo de difícil acceso al profano en la materia. Fairley ha comentado muy de pasada el problema de la ética en este tipo de estudios. Tema muy interesante y de una importancia clave para el futuro de la investigación tumoral. 329 RESULTADOS La ética estudia qué es lo moral, cómo se justifica racionalmente un sistema moral, y cómo se ha de aplicar posteriormente a nivel individual y social. Fernando Savater (535)en “Ética para Amador” la define como el arte de vivir, el saber vivir, por lo tanto el arte de discernir lo que nos conviene y lo que no nos conviene, y el Diccionario de la Lengua Española dice que es la parte de la filosofía que estudia el bien y el mal relacionado con el comportamiento humano y la moral. La bioética es una disciplina nueva (536). El origen del término se debe al pastor protestante y educador Fritz Jahr (537), que en 1927 usó el término Bio-­‐Ethik en un artículo sobre la relación ética del ser humano con las plantas y animales. El criterio ético fundamental que regula esta disciplina es el respeto al ser humano, a sus derechos, a su bien verdadero e integral: la dignidad de la persona, y en una visión más amplia la bioética no solo afectaría a aquellos organismos capaces de sentir miedo y dolor, sino también al ambiente en que se desarrolla la vida, relacionándose así con la ecología. Las primeras declaraciones de bioética surgen con posterioridad a la Segunda Guerra Mundial, tras el descubrimiento de los experimentos médicos llevados a cabo por el régimen hitleriano en los campos de concentración. Las atrocidades cometidas y reveladas en el Juicio de Nuremberg (538) dieron lugar al primer código que regulaba la investigación en humanos. La redacción del código de Nuremberg en 1948 será el primer protocolo de la historia sobre ética de la investigación en humanos donde se refleja el consentimiento voluntario de los sujetos en investigación. A partir de los años sesenta, hechos como la catástrofe de la talidomida (539), o el dilema planteado por el invento de la fístula de Scribner para la diálisis renal (540) que puso a los médicos en un dilema moral sobre quién podría recibir el tratamiento si éste fuese limitado, son estímulos que van acelerar las normativa sobre bioética. En 1964, la Sociedad Americana para los Órganos Internos Artificiales discutió el problema sobre selección de pacientes, finalidad del tratamiento, muerte con dignidad y selección de futuros trasplantados, lo que significó el comienzo de la bioética. 330 RESULTADOS En 1979, TL Beauchamp y JF Childress (541)(542) definen los cuatro principios de la bioética: autonomía, no maleficencia, beneficencia y justicia. En el ámbito médico el consentimiento informado es la máxima expresión del principio de autonomía, constituyendo un derecho de los pacientes y un deber del médico. La no maleficencia o la abstención de realizar actos que puedan causar daño o perjudicar a otros es un principio válido en todos los sectores de la vida humana. El Informe Belmont (543), creado por el departamento de Salud, Educación y Bienestar de los Estados Unidos con el título “Principios éticos y pautas para la protección de los seres humanos en la investigación”, es un importante documento histórico en el campo de la ética médica y continúa siendo una referencia esencial para los investigadores y grupos que trabajan en la investigación con humanos. La Asociación Médica Mundial (AMM) ha promulgado la Declaración de Helsinki (544) como una propuesta de principios éticos para la investigación médica en seres humanos. La Declaración se adoptó en 1964 en un intento de demostrar la capacidad autorreguladora de los médicos en el control de la investigación, pero diversos hechos desgraciados como el estudio de la sífilis de Tuskegee (545) confirmaron que dicha autorregulación no fue suficiente y se hacía necesaria una regulación externa, un control público. La quinta modificación de la Declaración de Helsinki (Edimburgo 2000) ha supuesto un cambio fundamental en la historia de este documento en un intento de aproximar los postulados éticos a la nueva realidad de la medicina y de la investigación médica. La última revisión ha tenido lugar en 2013, en Fortaleza, Brasil. La complejidad de muchos estudios se va ver favorecida por la actuación conjunta de diversos centros en varios países. Tal es el caso de: “Factores serológicos en trasplantes humanos”, título de un trabajo de investigación en el que participan varias instituciones como el Colegio Médico de Virginia, el Instituto de Investigación de las Enfermedades de la Sangre, la Clínica Nefrológica del Hospital Necker, el Centro de Investigaciones del Instituto Nacional de la Salud y de la Investigación Médica de la Universidad de París y el 331 RESULTADOS Departamento de Bacteriología e Inmunología de la Universidad del Estado de Nueva York en Buffalo, de 1967, entre cuyos autores figuran Félix T. Rapaport y Jean Dausset (546). La observación de que los leucocitos de la sangre se comportan como antígenos en el trasplante humano (547)(548) ha planteado la posibilidad de que determinantes antigénicos de estos leucocitos puedan ser un factor condicionante en la respuesta humana de histocompatibilidad. La primera parte de este informe (546) hace referencia a un estudio retrospectivo de la correlación entre antígenos de los grupos leucocitarios reconocidos por la batería de antisueros Hu-­‐1 (equivalente humano del sistema H-­‐2 del ratón) y el destino de 59 trasplantes renales. Los resultados serológicos de compatibilidad se correlacionan con el resultado clínico de dichos trasplantes con especial referencia a la supervivencia a largo plazo. La segunda parte del informe se ocupa de la caracterización de la respuesta de anticuerpos heterófilos asociada al rechazo de aloinjertos humanos. Anticuerpos dirigidos contra eritrocitos de oveja, cobaya y rata fueron detectados en 49 receptores de antígenos de trasplantes y aloinjertos de piel y en 22 receptores de trasplantes renales. Los resultados son comparados con determinaciones similares en 86 individuos normales, describiendo la especificidad y el carácter antigénico de tales anticuerpos, y se discute la aplicabilidad de esta respuesta de anticuerpos en la monitorización clínica de los trasplantes de órganos en el hombre. El grupo Hu-­‐1 se correlacionó con la clínica de 59 trasplantes renales realizados en el Colegio Médico de Virginia, Richmond y en el Hospital Necker de la Universidad de París. Había tres trasplantes dobles dicigóticos, 25 trasplantes de riñón de donantes maternos, 10 de donantes paternos y 20 de donantes hermanos en este grupo. Todos, donantes y receptores se sometieron a determinaciones completas de los grupos leucocitarios, los resultados se registraron a ciegas y sin conocer el estado clínico del trasplantado en el momento de la prueba. 332 RESULTADOS Las muestras de suero analizadas para anticuerpos heterófilos se obtuvieron de sujetos sensibilizados con antígenos de trasplantes y aloinjertos de piel, y de receptores de aloinjertos de riñón. Los receptores de antígenos y aloinjertos de piel fueron 49 voluntarios normales que formaban parte de un estudio sobre los efectos in vivo de eritrocitos y leucocitos de la sangre y sus fracciones. También se obtuvieron muestras de 22 receptores de trasplante renal de diversos hospitales, los cuales fueron observados y controlados serológicamente por un período de 1-­‐48 meses. Las muestras obtenidas de 86 voluntarios normales sirvieron como controles. Tres técnicas diferentes se utilizaron para la evaluación de la compatibilidad de los grupos leucocitarios entre donantes y receptores de aloinjertos renales: • Leucoaglutinación por el método de Dausset (549). • Linfocitotoxicidad por el método de Gorer y Gorman (550). • Fijación de plaquetas por complemento por el método de Shulman. Los receptores fueron clasificados en cuatro categorías principales en base a la compatibilidad de los grupos leucocitarios con los donantes: " Grupo A: incompatibilidades no detectables, incluso en sueros aislados de especificidad desconocida. " Grupo B: incompatibilidades detectables ocasionalmente con algunos sueros, sin evidencia de incompatibilidad de grupo. " Grupos C y D: una o más definidas incompatibilidades de grupos leucocitarios. El resultado clínico del trasplante renal en estos grupos de pacientes fue clasificado de acuerdo con dos diferentes conjuntos de criterios: 1. Criterios clínicos basados en la función renal del trasplante en individuos que sobreviven un mínimo de seis meses. El trasplante es considerado un éxito si el aclaramiento de creatinina >50cc/min, el nitrógeno ureico <30 mg/100 ml y la creatinina en suero < 2 mg/100 333 RESULTADOS ml. En base a estos criterios los pacientes fueron separados en cinco categorías desde A (excelente función renal) a D (insatisfactoria) o R (rechazo completo). 2. Tipo de crisis de rechazo. Una crisis de rechazo fue definida como cualquier aumento en sangre del nitrógeno ureico y/o creatinina sérica o una disminución del aclaramiento de creatinina que no pueda explicarse sobre la base de la necrosis tubular aguda, obstrucción urinaria, infección, oclusión vascular o aumento en la carga de nitrógeno. Se reconocieron cuatro grados de severidad: A (sin pérdida de función), B (un solo episodio de rechazo agudo de menos de 30 días que no requiere hemodiálisis), C (más de un episodio en los primeros cuatro meses) y D (un episodio de rechazo superior a 30 días o grave para requerir hemodiálisis). A los efectos de este estudio (546), las clases A y B se denominan compatibles, mientras C y D son incompatibles. Un año después de la intervención, 32 sujetos compatibles y 18 sujetos incompatibles estaban disponibles para el estudio. Resultados desfavorables (C, D y R) ocurrieron en el 55, 6% de sujetos incompatibles, pero solamente en el 15,7% de los receptores compatibles. La diferencia en la aceptación del trasplante se hizo aún mayor a medida que pasó el tiempo. Se observó 28 meses después del trasplante que el 69,3% de los sujetos con trasplante de donantes incompatibles rechazaron su injerto o estaban en proceso de rechazo, y únicamente el 8,3% de sujetos compatibles. También hubo una correlación positiva entre la compatibilidad con los grupos leucocitarios y el tipo de rechazo observado. Los resultados de los 59 trasplantes de riñón procedentes tanto de Richmond como del Hospital Necker de París, se resumen el la figura 45. Al final de los tres años, el 33% de los injertos incompatibles, pero únicamente el 8% de los injertos compatibles habían sido rechazados. Los resultados sugieren fuertemente que la oportunidad de supervivencia a largo plazo para el riñón es 334 RESULTADOS mayor en receptores compatibles. Hay que hacer notar que en tres casos de rechazo no se detectó evidencia de incompatibilidad con los grupos leucocitarios. Figura 45. Seguimiento del estado de 59 casos de trasplante renal. Correlación con la compatibilidad de los grupos leucocitarios. Tomado de: Rapaport, 1967 (546) Se amplió el estudio en los sujetos con al menos un aumento de 8 veces en el título de anticuerpos heterófilos. En algunos de ellos, ocasionalmente la hemaglutinina alcanzó títulos de 1:640 contra eritrocitos de oveja, de 1:1280 contra eritrocitos de cobaya y de 1:5120 contra eritrocitos de rata. Un pico en los títulos de hemaglutinina por lo general aparece una semana después del injerto de piel, generalmente en asociación con el rechazo del aloinjerto y la intensidad de la respuesta de anticuerpos parece ser proporcional a la intensidad de los estímulos antigénicos aplicados en cada individuo. No había 335 RESULTADOS ninguna relación entre la presencia o ausencia de incompatibilidad ABO entre donante y receptor el desarrollo de las respuestas de anticuerpos heterófilos. Hubo un aumento en el título de anticuerpos heterófilos en 20 de 22 receptores de riñón. Todos los individuos de este grupo estaban bajo terapia inmunosupresora con azatioprina y prednisona y la mayoría sufrieron al menos una crisis de rechazo durante el periodo de estudio. Los mayores títulos se observaron contra eritrocitos de rata. Es de particular interés que tales aumentos en los títulos de anticuerpos parecían tener una relación con el estado clínico de los pacientes, así los picos en los títulos eran asociados con crisis de rechazo inminentes o eran vistos después del rechazo del riñón y la nefrectomía. Con el fin de evaluar la especificidad de la respuesta de anticuerpos heterófilos observada después del trasplante, se utilizaron antisueros de alta titulación que fueron absorbidos con eritrocitos humanos A y B, con eritrocitos de oveja, cobaya, rata, buey y monos rhesus. El tratamiento con 2-­‐mercaptoetanol abolió la actividad heterófila hemaglutinante en 17 de 24 sueros, los 7 sueros restantes no fueron afectados. El resultado de este estudio (546) indica una particular correlación entre la compatibilidad de los grupos leucocitarios y la supervivencia y función del trasplante renal a largo plazo. Este hallazgo confirma la sugerencia de Porter (551) y colaboradores: “… el grado de compatibilidad del grupo leucocitario existente entre donante y receptor puede ser un factor determinante en la progresión de los cambios que ocurren en el rechazo crónico responsables del fallo del trasplante a largo plazo” Los resultados obtenidos en Richmond y París eran muy similares. A pesar de la evidencia de los resultados, los autores (546) muy cautos, dicen al respecto: “Aunque estos resultados son muy sugerentes, una evaluación definitiva del papel que el grado de compatibilidad de grupos leucocitarios tiene en el órgano trasplantado espera la acumulación de un mayor número de 336 RESULTADOS pacientes estudiados de forma prospectiva y la disponibilidad de sueros tipificados de naturaleza monoespecífica” Algunos sujetos rechazaron los trasplantes en ausencia de cualquier evidencia serológica de incompatibilidad leucocitaria, lo que indica que la batería de los antisueros que tenían a disposición fue incapaz de detectar todos las especificidades antigénicas que pueden condicionar el rechazo del trasplante en el hombre. Sin embargo, a pesar de ello, una correlación estadísticamente significativa se ha demostrado que existe entre la compatibilidad de grupo leucitario y la duración y calidad de la supervivencia del injerto renal en el hombre. Los autores (546) consideran que: “Los resultados justifican la inclusión de técnicas para determinar los grupos leucocitarios en la evaluación preoperatoria de donantes y receptores de trasplantes, lo que podrá permitir mejorar los resultados actuales y también evitar diferencias antigénicas que podrán reducir los requisitos para una terapia más efectiva con inmunosupresores en los trasplantes de órganos”. Con respecto al desarrollo de la respuesta de anticuerpos heterófilos se observó que no se relacionan con incompatibilidad ABO entre donante y receptor. Los experimentos de absorción cruzada indican que tales anticuerpos están dirigidos principalmente contra antígenos presentes en eritrocitos y son compartidos en grado variable con otras especies probadas. Las pruebas realizadas siguieren que tales anticuerpos no están dentro de la categoría Forssman y también que son distintos de los anticuerpos de Paul-­‐Bunnell. Dichos anticuerpos podrían constituir una respuesta a los antígenos que son liberados, alterados o expuestos como resultado de daño en los tejidos. La asociación del pico de estos anticuerpos con el rechazo de aloinjertos indica que la determinación de anticuerpos heterófilos en suero puede proporcionar una simple advertencia temprana para poder gestionar rápidamente la crisis del rechazo. Por tanto, este tipo de anticuerpos podría ser útil como método adicional en los protocolos de las pruebas de histocompatibilidad humana (552). 337 RESULTADOS Entre los autores de este estudio tenemos a Jean Dausset (553), descubridor del sistema HLA (Human Leucocyte Antigens). Su descubrimiento le valió el Premio Novel en 1980, compartido con Benacerraf y Snell. En 1958, Dausset (548) descubrió en la superficie de los glóbulos blancos pequeñas estructuras químicas capaces de provocar la aparición de un anticuerpo que se fija en ellas de forma específica; a este antígeno le denominó Mac, el primero aislado del sistema HLA. Dausset comprendió la importancia de estos antígenos en la defensa del organismo contra toda agresión externa e interna: “Las moléculas de HLA del donante, al ser distintas a las del receptor se perciben como extrañas e inducen una respuesta inmune que conduce al rechazo del trasplante”. Estos hallazgos le valieron el respeto de la comunidad científica y un elevado número de citas en las revistas médicas, figurando en más de 500 artículos. Unos años después, en 1965 da a conocer las leyes de la histocompatibilidad en el hombre por el sistema HLA (554)(555), afirmando que: “Si los leucocitos de dos personas son compatibles, los tejidos también lo son”. Demostró que el tiempo de supervivencia de los injertos era inversamente proporcional al número de incompatibilidades entre los genes HLA donante-­‐receptor. Clasificó a los HLA en dos grupos: los denominados de clase I (HLA-­‐A, HLA-­‐B, HLA-­‐C) presentes en la superficie de todas las células del organismo, y los de clase II (HLA-­‐DR, HLA-­‐DQ, HLA-­‐DP) que no se expresan más que en células implicadas en la respuesta inmune, y abordó el sistema HLA, no ya a nivel de los productos de los genes, sino de los mismos genes y sus funciones. En 1967 (556)(557) da a conocer la relación existente entre el sistema HLA y distintas enfermedades, como por ejemplo entre HLA-­‐B27 y la espondilitis anquilosante ó entre HLA-­‐ DQ2 y HLA-­‐DQ8 y la celiaquía, creando el concepto de “Medicina Predictiva”, entendida como el sistema que permite anticipar la aparición de una enfermedad y aplicar precozmente el tratamiento preventivo, si existe, o evitar los factores desencadenantes, si se conocen. 338 RESULTADOS Dausset (558) dio a conocer su descubrimiento a nivel mundial afirmando que: “Cada célula es portadora de la identidad de un individuo” y que “todos y cada uno de nosotros somos genéticamente diferentes y únicos”, por lo que la libertad en el respeto a la diferencia, es decir, a los individuos, es algo más que una reivindicación humanista: “… existe justificación hasta en nuestros genes”. En 1983 se creó el Centro de Estudios del Polimorfismo Humano, cuya central se encuentra en el Collége de France, que contribuye a descifrar el mapa del genoma humano y a identificar los genes relacionados con las enfermedades genéticas. Entre sus últimos estudios están los dirigidos a las moléculas HLA llamadas de tolerancia (HLA-­‐G) que inhiben la reacción de la madre contra su feto (559). Pero el conocimiento del linfocito como tal aportará nuevos datos que irán en beneficio de futuros estudios. Sobre la existencia de “Dos poblaciones de pequeños linfocitos” publican en 1968, G. Ruhenstroth-­‐Bauer y C. Lücke-­‐Huhle (560) del Instituto Max-­‐Planck de Bioquímica en Munich. Es conocida la existencia de dos poblaciones de linfocitos en el conducto torácico de la rata, de ellos aproximadamente un 5% son grandes linfocitos y un 95% pequeños linfocitos, que difieren no solo en aspectos morfológicos sino también biológicos (561). Además existen pruebas de que la población de pequeños linfocitos está constituida por dos subpoblaciones, siendo esto observado particularmente en roedores (562). El documento de Ruhenstroth-­‐ Bauer y Lücke-­‐Huhle proporciona datos que apoyan la existencia de dos poblaciones de pequeños linfocitos distintas morfológica y funcionalmente. Datos que se desprenden de las observaciones realizadas en la movilidad de los linfocitos en un campo electroforético y los cambios en el volumen celular asociados a cambios en la función de la población. Al estudiar la movilidad electroforética de los linfocitos de 308 ratas adultas, se observa la existencia de dos poblaciones de linfocitos. La gráfica obtenida es la suma de dos poblaciones de distinta movilidad que siguen una distribución Gaussiana. La población I, que comprende el 77% del total, tiene una movilidad 339 RESULTADOS electroforética con valores medios distintos a los de la población II, que constituye el 23% del total. La población de linfocitos electroforéticamente más lenta será llamada A y la más rápida B. En segundo lugar se midió el volumen de los linfocitos del conducto torácico de la rata, analizando la distribución de la misma manera que se hizo para la movilidad electroforética. Se observó que aquí también el total puede dividirse en dos poblaciones, cada una con una distribución Gaussiana distinta, así el 79% de las células tenía un volumen medio de 141±28 µ3 y el 21% de 196±24 µ3. Si tratamos los linfocitos como células esféricas, estos volúmenes se corresponden con un diámetro de 6,5µ y 7,2µ respectivamente. Los linfocitos A serían aquellos con mayor volumen y más lentos y los B se corresponderían con linfocitos más pequeños y más rápidos. Hipótesis que fue confirmada con la siguiente observación: de 1-­‐2 horas después de irradiar con 800 R todo el cuerpo de la rata, los linfocitos del conducto torácico son en la mayoría de los casos casi únicamente de tipo B cuando se examinan electroforéticamente, y las mediciones de volumen muestran que la población está compuesta casi en su totalidad por los linfocitos de menor volumen medio (Figura 46 y 47). Figura 46. Movilidad electroforética. Distribución según la movilidad electroforética de los linfocitos del conducto torácico de la rata. I, linfocitos tipo A; II, linfocitos tipo B. Tomado de: Ruhenstroth-Bauer y C. Lücke-Huhle, 1968 (560) 340 RESULTADOS Figura 47. Volumen. Distribución según el volumen de los linfocitos del conducto torácico de la rata. Tomado de: Ruhenstroth-Bauer y C. Lücke-Huhle, 1968 (560) Los autores (560) consideran que sus hallazgos están en armonía con los de Gowans y Knight (561), que indican que el 80% de los linfocitos recirculan tan rápidamente que estas células desaparecen en un tiempo relativamente corto en un drenaje prolongado, y que el restante 20% son más lentas, apareciendo después de cinco días de drenaje en número bajo pero detectable. Una distribución exacta del volumen de las células en sangre periférica no fue posible con los métodos utilizados, ya que la sangre era tratada con saponina para destruir los eritrocitos y esta saponina provocaba daños en la membrana del linfocito de forma que los métodos eléctricos de medición de volumen no daban resultados fiables. Sin embargo se pudo obtener información mediante otros dos tipos de estímulo: irradiación y etanol. Pocas horas después de la irradiación corporal total con 800 R, los linfocitos periféricos eran casi exclusivamente de tipo B. Con una inyección intravenosa o intraperitoneal de 0,2 ml de etanol al 96%, a los 15 minutos aparecían hasta un 30% de linfocitos tipo B en sangre, desapareciendo de la circulación una hora después, cuando la distribución electroforética vuelve a la normalidad. 341 RESULTADOS Con el fin de aclarar la distribución corporal de linfocitos, los linfocitos A y B del conducto torácico fueron marcados in vitro con adenosina tritiada y reinyectados vía intravenosa de nuevo en la rata (561). 24 horas después de inyectar linfocitos A marcados, los linfocitos periféricos contenían un 4% de células marcadas, que también fueron encontradas en bazo y ganglios, mientras que médula ósea y timo se mantuvieron libres de células marcadas. A las 24 horas después de la inyección de una mezcla de linfocitos A y B marcados en proporción 20:80, el 2% aproximadamente de linfocitos estaban marcados. A pesar del hecho de que los linfocitos B eran cuatro veces más numerosos que los A, la población total de linfocitos de la sangre contenía 20 veces más células A que B, lo que implica que los linfocitos B salían de la sangre mucho más rápidamente que los A. Una estimación aproximada fija en 19 horas para los A y 9 minutos para los B, lo que da lugar a que parezca mayor el número de linfocitos A que de B. Se plantea la cuestión de si los dos tipos morfológicos de pequeños linfocitos tienen funciones distintas. Para ello, las ratas fueron inmunizadas con eritrocitos de oveja y cinco días después los linfocitos del conducto torácico fueron probados para producción de anticuerpos contra eritrocitos de oveja relacionando el número de placas hemolíticas con el número de leucocitos probados. Aseguran (560) que a pesar de los errores experimentales, los resultados sugieren la existencia de una correlación entre el número de linfocitos A y el número de células productoras de anticuerpos, pero no se excluye la posibilidad de que la actividad inmunológica sea una propiedad de un tipo distinto de célula presente en cantidades muy pequeñas en la población examinada, y por lo tanto no detectable morfológicamente. En un pequeño resumen, los autores (560) aclaran que: “Nuestros experimentos están de acuerdo con la hipótesis de que el llamado pequeño linfocito de la rata constituye dos poblaciones de células, A y B, determinadas por la movilidad electroforética y el volumen celular medio de las mismas, distinto en A y B. En el conducto torácico de la rata estos dos tipos se encuentran en una proporción de 342 RESULTADOS 20:80, y se pudo demostrar que hay una correlación entre la proporción de linfocitos A y el número de células que responden a estímulos inmunológicos”. De índole muy distinta al anterior es este trabajo de R N, Allan y M K Alexander (563) del laboratorio de Patología de Warwick (Reino Unido) en 1968: “La diferencia de sexo en el recuento de leucocitos”. La llegada de técnicas más modernas de procesamiento de datos en el laboratorio clínico permite el análisis de una gran cantidad de muestras de forma rutinaria. En un estudio preliminar realizado con pacientes hospitalarios, se observó una posible diferencia entre sexos en el recuento diferencial de leucocitos, lo que motivó la necesidad de realizar un estudio más amplio con donantes sanos y pacientes hospitalizados. Se estudiaron 325 donantes sanos de edades comprendidas entre 18 y 65 años, que para el análisis estadístico fueron divididos en cuatro grupos según la edad (18-­‐29; 30-­‐39; 40-­‐49; 50-­‐65). También se analizó la sangre de pacientes hospitalarios de medicina general, cirugía, y otorrinolaringología. El estudio abarcó tres meses consecutivos, marzo, abril y mayo de 1967, y un total de 911 hombres y 864 mujeres cumplieron los criterios referentes al valor de hemoglobina y células blancas para ser incluidos en el estudio. Tabla II. Distribución de las muestras por edad y sexo. Donantes sanos y pacientes hospitalizados. Tomado de: Allan y Alexander, 1968 (563). 343 RESULTADOS Se tuvieron en cuenta una serie de factores para no influir en los resultados. Así, para evitar la tendencia a la subida en el recuento de glóbulos blancos al final de la tarde, solo se realizaron extracciones durante la mañana y a primera hora de la tarde, los frotis de sangre se realizaron de inmediato y el recuento de células blancas y la hemoglobina fueron estimadas en el mismo día. Un centenar de células fueron examinadas en cada frotis para el recuento diferencial de células blancas, y el sexo de los pacientes no fue una información disponible cuando las estimaciones se llevaron a cabo. Todos los resultados se plasmaron en tarjetas perforadas y se procesaron en un ordenador IBM 1440 programado para crear histogramas de los valores de hemoglobina y de los recuentos totales y diferenciales de células blancas para hombres y mujeres, con los correspondientes análisis estadísticos como los coeficientes de correlación y el análisis de la varianza. El análisis de la varianza se empleó para determinar si había una diferencia significativa en el recuento total y diferencial de células blancas respecto al sexo y en los distintos grupos de edad. El coeficiente de correlación entre el valor de la hemoglobina y el recuento total de glóbulos blancos en los dos sexos y por edades se realizó para determinar la existencia o no de relación en ambos parámetros. En los resultados obtenidos de los donantes sanos se observó que no había diferencias significativas en el total de células blancas en personas menores de 50 años. En el grupo de 50-­‐65 años, el recuento total en las mujeres fue significativamente menor que en hombres. Con el aumento de la edad en la mujer se observó una caída significativa en el recuento total de glóbulos blancos debido a una disminución del número de linfocitos y una menos marcada disminución de polimorfos. No hubo correlación significativa en este grupo de mujeres de 50-­‐65 años entre los valores de hemoglobina y el recuento de células blancas. No se observaron variaciones en el recuento total y diferencial con la edad en el sexo masculino. En el grupo de pacientes hospitalarios, la diferencia según el sexo en el recuento total de leucocitos fue significativa en el rango de edad de 30-­‐49 años y altamente significativa entre 50-­‐65 años. Había una correlación altamente 344 RESULTADOS significativa entre la hemoglobina y el recuento total de células blancas en las mujeres de edad comprendida entre 30 y 39 años y una correlación significativa en el grupo de edad de 40-­‐49 y 50-­‐65. Por lo que se puede decir que el análisis de un gran número de recuentos en pacientes hospitalarios ha confirmado la existencia de diferencias entre los sexos en el recuento total de leucocitos. En la discusión los autores (563) hacen referencia a otros estudios similares. El realizado por Previtera, Venezia y Vignale en el año 1965 (564) que a partir de un estudio de 735 hombres y 939 mujeres de edades comprendidas entre 1 y 25 años, encontraron en hombres entre 10 y 20 años un recuento de neutrófilos inferior y entre 5 y 25 años un recuento de linfocitos superior que en mujeres. Los resultados de estos tres autores (Previtera, Venezia y Vignale) están en contradicción con los comunicados por Osgood (565), Geller y Sukhanova en 1964 (566). En una investigación sobre los estándares hematológicos normales de 500 personas normales, Osgood llegó a la conclusión de la no existencia de diferencias respecto al sexo en el número total y recuento diferencial leucocitario a cualquier edad, sin embargo, la mayoría de sus sujetos eran menores de 30 años. Los autores dicen haber confirmado los resultados de Osgood, Ellis y Cohen, los cuales no encontraron diferencias significativas atribuibles al sexo en el recuento de leucocitos en el grupo de edad de 19-­‐30 años. En un estudio realizado a 439 rusos saludables, Geller y Sukhanova (566) no encontraron diferencias en el recuento de leucocitos en relación a edad y sexo, pero todos los estudiados fueron menores de 50 años. Otras causas fueron reconocidas como causa de variación en el recuento de leucocitos, como la variación diurna y una variación cíclica en los hombres. Las razones (563) de las diferencias en razón de sexo en la presente investigación no son satisfactoriamente explicadas. Así, entre los factores considerados está la posible influencia de las hormonas sexuales, aunque es conocido que la administración de ACTH o cortisona es seguida de un aumento 345 RESULTADOS de neutrófilos y una disminución de linfocitos, no hay evidencia de que los niveles plasmáticos de cortisol en los sexos, como se refleja por la excreción de 17-­‐cetosteroides, se altere con el incremento de edad, y además este patrón de cambio en el recuento de células no se ajusta a la encontrada en los donantes de sangre. La presencia de una correlación significativa entre nivel de hemoglobina y recuento de células blancas en varios grupos de edad de mujeres donantes y pacientes hospitalarios sugiere la posibilidad de un factor nutricional o metabólico que pueda influir en ambos parámetros, como puede ser la vitamina B12, el hierro o los folatos. En valor medio del hierro sérico disminuye con la edad en ambos sexos, y es más bajo en todas las edades en la mujer que en hombre, pero la posible relevancia de este hallazgo para los resultados de este estudio es incierta. El linfocito también interesa en el estudio etiológico de las enfermedades. La observación de una disminución del infiltrado linfocitario en la mucosa intestinal de la enfermedad de Whipple conduce a W.C. Watson, J.D. Maxwell y Anne Fergunson (567) a su estudio, que publican en 1969 un breve artículo titulado: “Los linfocitos en la enfermedad de Whipple”. El diagnóstico de la enfermedad de Whipple se basa en la presencia de macrófagos que contienen material PAS positivo en tejidos como el intestino y ganglios linfáticos regionales. Este material corresponde a los restos fagocitados de las paredes celulares de un organismo bacilar. Además los pacientes pueden ser curados con antibióticos, un prueba más de que las bacterias están de alguna forma implicadas en la enfermedad. La disminución de linfocitos en la enfermedad de Whipple contrasta con los hallazgos histológicos encontrados en otras patologías crónicas del intestino delgado (568). Hechos que sugieren que los factores del huésped pueden contribuir en la patogénesis de la enfermedad. Se estudiaron dos pacientes con enfermedad de Whipple diagnosticada, en remisión clínica y con un perfil inmunológico normal. Sus linfocitos mostraron 346 RESULTADOS una respuesta alterada a la estimulación con fitohemaglutinina según el método descrito por Tormey y Mueller en 1965 (569), por lo que los autores consideran la necesidad de un estudio más amplio: “Si esta anormalidad se encuentra en otros casos sería sugestiva de que la existencia de un fallo en la inmunidad mediada por linfocitos podría contribuir en la patogénesis de la enfermedad”. La enfermedad de Whipple es una infección multisistémica, afortunadamente poco frecuente, causada por una bacteria Gram positiva. Fue el patólogo George Hoyt Whipple quien la describió en 1907 (570) e hizo notar que estaba causada por una bacteria, lo que fue la base de estudios posteriores para identificarla. Whipple describió el curso clínico de la enfermedad en un hombre de 36 años. Debilidad, pérdida de peso, dolor abdominal, artritis múltiple y presencia de grasa en heces eran los principales síntomas, que refería padecer desde seis años antes cuando trabajaba como misionero en Turquía. En base a los hallazgos patológicos Whipple le dio el nombre de lipodistrofia intestinal y sugirió que los microorganismos encontrados en los ganglios linfáticos podían ser los causantes. Años más tarde Black Shaffer (571) descubrió que los macrófagos se coloreaban intensamente de magenta con la tinción de Schiff (PAS), prueba que hoy día sigue utilizándose para orientar el diagnóstico. En los años sesenta, varios autores, entre ellos Yardley y Hendrix (572) y Chears (573) de forma independiente corroboraron la presencia de estructuras bacilares en el intestino. Cuando Watson, Maxwell y Fergunson publican su artículo ya se conoce la causa de la enfermedad, y la aportación de éstos se limita a dar a conocer la observación de una disminución del infiltrado linfocitario en la mucosa intestinal y sugerir que los factores del huésped, es decir el sistema inmune, pueden contribuir a la patogenia de la enfermedad. Da la impresión de que los autores lo que pretenden es una llamada de atención en la búsqueda de nuevos casos, ya que se les presenta el problema de la dificultad del estudio debido al escaso número de pacientes (ellos presentan solo dos), y su intención queda clara cuando dicen: “si esta anormalidad se encuentra en otros casos, sería 347 RESULTADOS sugestiva de que la existencia de un fallo en la inmunidad mediada por linfocitos podría contribuir a su patogénesis”. La enfermedad de Whipple es un ejemplo de cómo la tecnología médica moderna ha contribuido al conocimiento preciso de las enfermedades y su tratamiento. Dos importantes hallazgos han contribuido a ampliar el conocimiento y mejorar el diagnóstico: 1-­‐ las técnicas de amplificación molecular del ARN ribosomal 16s (ARNr) de la bacteria causante trofheryma whippelli, mediante PCR (Reacción en Cadena de la Polimerasa) y 2-­‐ el cultivo de la bacteria, que a permitido probar diversos antibióticos in vitro y emplear el más efectivo en el tratamiento de los pacientes. A día de hoy tenemos constancia de la probabilidad de que la causa radique en deficiencias inmunológicas del paciente, como acertadamente señalaron los autores, que permiten que la bacteria se multiplique en los macrófagos y monocitos. Esta alteración se asocia con la replicación de la bacteria en los macrófagos y la apoptosis de los mismos que disemina la bacteria. Fenómenos relacionados con la expresión de la liberación de interleucina 16 (IL16) y una producción inadecuada de interleucina 12 (IL12) que incapacita para destruir antígenos bacterianos, añadido al hecho de que los anticuerpos neutralizan la producción de IL16, y por lo tanto permiten el desarrollo de la trofheryma whippelli. Y es posible que sea esta deficiencia inmunológica, propia del paciente o bien causada por la bacteria, la responsable, ya que la respuesta humoral no se muestra alterada. La obra de Whipple (574) puede enmarcarse en una época en la que se producen gran cantidad de hallazgos clínicos gracias al extraordinario auge que alcanza la “Medicina de Laboratorio”. Los principales temas de investigación de Whipple estuvieron relacionados con la anemia y con la fisiología y patología del hígado, por los que fue galardonado con el Premio Novel en 1934, y sin embargo su nombre ha quedado ligado (paradojas de la vida) a la enfermedad que casualmente describió. 348 RESULTADOS Es conocido que las enfermedades infecciosas en general provocan una alteración del equilibrio celular sanguíneo. Glade, Paltrowitz y Hirschhorn (575), procedentes del Departamento de Genética Médica de la Escuela de Medicina Monte Sinaí de Nueva York, publican en 1969sus observaciones sobre el tema: “Potencial linfoproliferativo de las enfermedades infecciosas”. Los años cincuenta han sido testigos de una rápida comprensión en la naturaleza de los linfocitos y su importancia en la sa