- Ninguna Categoria
Capítulo 3 3.1.- Lípidos 3.1.a.- Clasificación Los
Anuncio
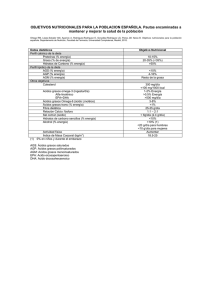
Capítulo 3 3.1.- Lípidos 3.1.a.- Clasificación Los lípidos son un conjunto de biomoléculas que coinciden en su hidrofobia como elemento común. Pueden ser lípidos complejos y simples, y dentro de estos últimos, están los ácidos grasos, que son ácidos orgánicos monocarboxílicos de cadena lineal, y con un número de átomos de carbono entre 4 y 24, siendo los más abundantes los comprendidos entre 14 y 22. La insaturación se refiere al doble enlace que pueden tener (CH=CH), lo que se puede indicar mediante la letra C y los subíndices m:n, siendo C el carbono, m el número total de átomos de carbono y n el de insaturaciones (por ejemplo, en el ácido oleico, con 18 carbonos y un solo enlace insaturado, la identificación sería C18:1). Otra denominación, más habitual, es la de nombrar la posición del doble enlace dentro de los últimos 7 carbonos de la cadena a partir del grupo metilo terminal, ocupando la posición 3 (serie n3 u Omega3) o la posición 6 (serie n6 u Omega6) (el linolénico sería un representante de los Omega3. El araquidónico sería un Omega6) (Tabla I. Tabla II). La digestión de las grasas se realiza en el intestino delgado, donde los triacilgliceroles de la grasa de la comida se transforman en diacilgliceroles, monoacilgliceroles y ácidos grasos por la lipasa pancreática. La secreción pancreática contiene esterasas que degradan los fosfolípidos (el enzima catalizador es la fosfolipasa A2, que se sintetiza en el páncreas como proenzima y se activa en el intestino por la tripsina). Nuestro organismo puede sintetizar la mayoría de los ácidos grasos a partir del acetil coenzima A procedente de los carbohidratos mediante un complejo multienzimático denominado sintetasa de ácidos grasos. Además, tenemos sistemas de elongación de cadena de ácidos grasos, que permiten aumentar los carbonos hasta 18 en el caso de los saturados y hasta 24 en los insaturados, y sistemas para introducir dobles enlaces mediante las desaturasas de ácidos grasos. Sin embargo, no se pueden introducir dobles enlaces en posiciones posteriores al C9. Pues bien, algunas biomoléculas de gran importancia para el organismo como las prostaglandinas y tromboxanos derivan de ácidos grasos que no pueden sistetizarse y hay que incorporarlos, por tanto, en la alimentación (Tabla III). El organismo humano puede sintetizar los ácidos grasos de la familia Omega-9, pero no puede sintetizar los de la familia Omega-6 u Omega-3. Entre ellos está el linoleico, el araquidónico y el linolénico. Los vegetales en cambio, pueden sintetizar los de la familia Omega-6 y algunos de ellos (especialmente las algas marinas microscópicas), pueden sintetizar la familia Omega-3. Los peces, por ejemplo, acumulan Omega-6 y Omega-3 y lo hacen a partir del plancton marino que consumen. Sabemos que el contenido y características del fosfolípido presente en las distintas membranas de las diversas células del organismo varía en función de las necesidades de cada órgano. De esta forma, cada fosfolípido, dependiendo de su estructura, tiene unas propiedades físico-químicas concretas que, al formar parte de la membrana, trasfiere a ésta. Según cuál sea el componente que predomine, la membrana será más o menos fluida, más o menos permeable a tal o cual sustancia, y sus movimientos serán más o menos fáciles. Por ello, hay que reseñar la importancia del porcentaje en que cada lípido se halla en la membrana, porcentaje que viene predeterminado genéticamente. Pues bien, en términos generales podemos decir que no hay una previsión genética sobre el ácido graso concreto que debe ocupar cada posición en cada uno de los fosfolípidos de la membrana lipídica, sino que sólo existe una generalidad sobre las características globales que deben tener. El determinante de cuál es el ácido graso individual que ocupe cada posición depende en gran medida de la dieta. De las distintas materias grasas, tienen predominio de ácidos grasos monoinsaturados los aceites de oliva, colza, aguacate, frutos secos como la almendra o la avellana, las margarinas y las mantecas hidrogenadas, mientras que son ricas en ácidos grasos poliinsaturados: aceites vegetales como el de soja, maíz, pepita de uva o de semillas de girasol, y los aceites marinos, extraídos de diferentes pescados n3, o consumidos en el conjunto de este tipo de alimentos. En el deportista son especialmente importantes los alimentos ricos en ácidos grasos n3 3.1.b.- Utilización de los lípidos durante el esfuerzo En el momento de la iniciación el ejercicio (paseo o carrera), los músculos en actividad aumentan la captación de los AGL. La cantidad de AGL conseguida por l as células musculares sobrepasa a la producida en la lipolisis, de manera que en los primeros 10- 15 minutos de comenzado el ejercicio disminuyen los AGL en el plasma. Entre los 15 y 20 minutos de iniciado un esfuerzo con el cicloergómetro a una intensidad del 40% del VO2max., el recambio de los AGL aumenta unas 4 veces, respecto al observado en reposo; y se incrementa hasta 6 veces, transcurridas 4 horas. Con intensidades inferiores al 40% del VO2max las necesidades energéticas se cubren solamente con las grasas. A medida que el ejercicio se hace más intenso, el porcentaje de AGL consumido desciende progresivamente a la vez que aumenta el de los carbohidratos. En las actividades moderadas cuya duración no supera las dos horas, los AGL son la principal fuente de energía. Cuando el esfuerzo es igual o superior al 85% del VO2max sólo el 25% del gasto se halla cubierto por los AGL, el otro 75% le corresponde a los carbohidratos (Sadur CN and Eckel RH 1982) La mejoría de la forma física disminuye las concentraciones plasmáticas de glicerol y AGL, a la vez que incrementa la oxidación total de las grasas. Por otra parte, sabemos que la captación y la oxidación de los AGL es directamente proporcional a su concentración plasmática. Por tanto, para que los AGL transportados en el plasma fueran los responsables de la donación de energía en los individuos entrenados, sería condición indispensable que el entrenamiento incrementase la captación de los AGL por parte de las fibras musculares en una cantidad igual o ligeramente inferior a su concentración plasmática. Por otro lado sabemos que la mejoría de la condición física producida por el entrenamiento, no aumenta la captación de los AGL por el músculo durante el ejercicio moderadamente intenso en el que intervienen las grandes masas musculares. También sabemos que en los deportistas bien entrenados, las tres cuartas partes de los AGL oxidados durante el ejercicio moderado no provienen de los AGL del plasma, sino de los triglicéridos musculares, justamente el 90 % (Martin et al 1993). 3.1.c.- Ácidos grasos “trans” Los ácidos grasos trans son los isómeros trans de las grasas insaturadas. La mayoría de los ácidos grasos insaturados que se presentan en los alimentos naturales están en forma "cis". Las siglas "cis" y "trans" se refieren a la posición que los átomos de hidrógeno tienen alrededor de los enlaces dobles de los ácidos grasos. Cuando los átomos de hidrógeno se encuentran en el mismo lado de la cadena, éstos se encuentran en la posición "cis"; cuando se encuentran en el lado opuesto de la cadena están en la posición "trans". El ácido oleico es un ejemplo de lo que es un "cis", mientras que el ácido elaídico es un ejemplo de un "trans". Los ácidos grasos "cis" presentan una estructura doblada mientras que los ácidos grasos "trans" son derechos. Si los triglicéridos que están siendo estudiados tienen un alto nivel de ácidos grasos "trans", ellos se juntarán fuertemente y se solidificarán formando cristales duros. Las grasas duras tienen mayores puntos de fusión. El punto de fusión de los ácidos grasos "trans" se encuentran más cerca de las formas saturadas que de las formas insaturadas denominadas "cis". Los ácidos grasos trans se forman en el proceso de hidrogenación que se realiza sobre las grasas para utilizarlas en diferentes alimentos. Una de las características principales de la hidrogenación es hacer más sólida la grasa, por ejemplo la de los aceites vegetales que se encuentra en estado líquido. Este proceso de hidrogenación promueve la frescura, le da mejor textura, mejor estabilidad y por lo tanto una mayor calidad al producto terminado. La ingesta de este tipo de ácidos no debe superar el 3% del total de calorías de la dieta Por otro lado, recientemente se presta una especial atención a un ácido graso “trans” que se encuentra en algunos alimentos, el llamado ácido linoleico conjugado (CLA). Los CLA son componentes naturales de los alimentos de origen animal derivados del ácido linoleico. Se encuentran en la grasa de la leche, los productos lácteos y la carne procedente de animales rumiantes. Recientemente, ha aumentado el interés por el contenido del CLA en la dieta como consecuencia de una serie de investigaciones (realizadas fundamentalmente con animales), cuyos resultados sugieren que estos componentes pueden ser beneficiosos para la salud. El CLA se descubrió en la University de Wisconsin en 1978. Su análisis nos proporciona los siguientes picos:a) isómeros delta 8,10 y delta 9,11; b) isómeros delta 11,13 y delta 10,12 y c) isómeros di-trans y di-cis. Algunos estudios relacionan la ingesta de ácido linoleico conjugado (CLA) con un efecto anabólico y lipolítico en deportistas (Thom E et al 2001). Se precisan unos 4 g diarios para este supuesto efecto. Sin embargo otros autores no han encontrado tal efecto (Zambell KL, 2001); (Kreider RB et al 2002). En otro estudio, se relaciona la ingesta de este derivado del ácido linoleico con la inmunidad, masa ósea y contenido mineral del hueso (Sebedio JL 1999). Parece que la ingesta del isómero 10 trans 12 cis del CLA, estimula la síntesis proteica muscular en ratas adultas (Arnal MA et al. 2001), mientras que el t10,c12-CLA aumenta la tasa de transporte de Calcio en células Caco-2 (Jewell C, and Cashman KD 2003), aunque en posteriores estudios no se confirma “in vivo” (Doyle L et al., 2005). La ingesta de 6 g/d de CLA (22,6% trans-10, cis-12; 23,6% cis-11, trans-13; 17,6% cis- 9, trans-11; 16,6% trans-8, cis-10; 7,7% trans-9, trans-11 ; y trans-10, trans-12 ; 11,9% otros isómeros), durante 28 días, aunque no produjo efectos estadísticamente significativos respecto a un aumento de masa muscular (deducida a partir de la masa magra obtenida por DEXA) o de fuerza (máxima carga admitida), o masa ósea (medida con DEXA), se comprobó una tendencia estadística (sin llegar a ser estadísticamente significativa) de aumento de masa ósea y muscular (Kreider RB et al. 2002). En algún trabajo aislado incluso se han llegado a encontrar efectos anabólicos con aumento de la masa muscular en culturistas (Lowery LM et al. 1998), aunque no está comprobado en otras investigaciones. No obstante, lo que sabemos es que los isómeros CLA reducen en cultivos de hueso la PGE2 (Li Y and Watkins BA 1998), la IGF-I en suero de ratas alimentadas con CLA (Li Y et al. 1999), la IL-6 en macrófagos peritoneales (Turek JJ et al. 1998) y LTB4 esplénico (Sugano M et al. 1998). Tabla IV En este sentido, sabemos que la biosíntesis exagerada de PGE2 puede disminuir la formación de hueso e incrementar su reabsorción (McCarthy et al., 1998). Efecto que ha sido estudiado por otros autores que, incluso, han encontrado un efecto sinérgico al añadir ácidos grasos n3 (Watkins BA and Seifert MF, 2000). Regulación del equilibrio n6/n3 Couet et al., 1997 realizaron un estudio dando a un grupo de 6 personas una dieta cuyos lípidos predominantes eran mantequilla, aceite de oliva y de semilla de lino y cambiando posteriormente a una dieta posterior cuya fuente de grasa era aceite de pescado. Comprobaron que tras la ingesta de aceite de pescado aumentó la oxidación de grasa como fuente energética, sin alterar la utilización de glucosa o proteínas, al tiempo que también observaron cambios en los fosfolípidos de membrana en eritrocitos (se hicieron más ricos en EPA y DHA). Finalmente, la disminución en la fluidez de la membrana eritrocitaria inducida por el ejercicio físico se minimiza cuando se ingiere una mezcla de vitaminas y ácidos grasos n-3, al menos en estudios realizados en caballos25. Este último estudio deja abierta la puerta a un doble mecanismo de acción de los ácidos grasos n-3 en relación con el estrés oxidativo potenciado por el ejercicio físico; la regulación al alza de enzimas como la gamma-glutamil-cisteinil ligasa y la glutation reductasa, lo que supone aumentar el glutation, y la acción sobre la fluidez de la membrana celular. Una investigación previa realizada por nuestro grupo con un lípido estructurado con ácido docosahexaenoico (DH) en posición 2 de un trigicérido (Brudy Plus), demostró que la ingesta continuada de 2 g de Brudy Plus por encima de tres semanas produce un incremento de la Capacidad Antioxidante Total del plasma de forma generalizada tanto en ciclistas de nivel competitivo como en el resto, siendo, asimismo, el daño oxidativo a lípidos menor. Por otro lado, el malonildialdehído (MDA) medido en el mismo estudio, mostraba que el aumento que provocaba el esfuerzo físico realizado antes de ingerir el Brudy Plus, era mayor que el experimentado después de tres semanas de ingesta y, además, era el mismo en ambos grupos de deportistas (aficionados y competitivos). Finalmente, también observamos que el daño al ADN medido con un marcador urinario (8-oxo-dG) disminuye tras la ingesta de Brudy Plus durante tres semanas. Todos estos datos son especialmente relevantes en el caso de deportistas aficionados cuyo nivel de daño oxidativo es mayor al no tener entrenadas las defensa antioxidantes del organismo y nos indica que el mayor efecto antioxidante del Brudy Plus se produce en los deportistas menos entrenados y más necesitados de este efecto. En una investigación posterior realizada en 59 varones, provocamos un estrés oxidativo inducido por el ejercicio físico aerobio (esfuerzo continuado al 75% de su consumo máximo de oxígeno durante 90 minutos), lo cual suponía una producción suficiente de especies reactivas como para conocer la capacidad antioxidante de cualquier manipulación dietética. Los resultados fueron: 1) Cambios en la membrana celular: a. El perfil de ácidos grasos en membrana eritrocitaria nos mostró un incremento de los ácidos grasos n-3 en su conjunto (a dosis de 500 mg, mientras que los n-6 descienden también de forma significativa. Como consecuencia el cociente n-6/n-3 experimenta descensos significativos dosis dependientes en los individuos que consumieron el producto a estudio. b. Al valorar cada ácido graso poliinsaturado por separado apreciamos un incremento significativo del porcentaje de DHA en la membrana del eritrocito tras el consumo de Brudy Plus®. Este incremento es tanto mayor cuanto mayor es la dosis de ingesta del Brudy Plus®, siendo significativas las comparaciones de las dos dosis mayores con respecto al placebo. El coeficiente de correlación lineal entre las variables incremento de DHA tras 1 mes de consumo de Brudy Plus® y la dosis ingerida es 0,414. c. Por otro lado, el EPA aumenta en la membrana del eritrocito tras el consumo de Brudy Plus®. Aunque este consumo es apreciable para cualquier dosis, solamente se hace significativo a partir de 2500 mg. Asimismo desciende el porcentaje de araquidónico en la membrana eritrocitaria tras el consumo de dosis de Brudy Plus® iguales o mayores a 1500 mg, siendo significativo para 1500 mg y para 3500 mg. En definitiva, el consumo de Brudy Plus® produce un incremento de PUFA n-3 (principalmente de DHA y, en menor medida, de EPA) en la membrana del eritrocito y este incremento es dosis dependiente. El ascenso de DHA y EPA produce un desplazamiento de la membrana del ARA disminuyendo su contenido eritrocitario al consumir dicho Brudy Plus®. Las diferencias entre las concentraciones de ambos ácidos grasos encontrada en nuestro perfil de la membrana de eritrocitos, son similares a las establecidas en la literatura médica en la que se conoce que al suministrar DHA dietético en dosis de 2 g se incrementa el DHA plasmático y el EPA, aunque éste último en menor medida. 2) Cambios en el daño oxidativo producido por un esfuerzo físico intenso a. Los cambios en la membrana celular tienen su expresión en los resultados obtenidos al medir la 8-OHdG excretada en orina como indicativo de daño oxidativo. Los resultados demuestran que el consumo diario de Brudy Plus® durante un mes no produce modificaciones en el daño oxidativo basal al ADN. En cambio este consumo sí genera una protección contra el daño oxidativo creado por una prueba de esfuerzo si se consume a dosis iguales o superiores a 1500 mg. Este efecto es idéntico para todas las dosis efectivas, no presentando un efecto dosis-respuesta sino una dosis umbral a partir de la cual el efecto es constante. Atendiendo al efecto acumulativo que presenta el DHA, cuando se suplementa en una dieta, pensamos que el uso de Brudy Plus en dosis de 500-1000 mg/d durante 6 a 12 meses, podría conseguir efectos iguales al experimentado con mayores dosis. Por otro lado, hemos comprobado que el consumo diario durante un mes de 3500 mg de Brudy Plus® neutraliza totalmente el daño oxidativo generado por la prueba de esfuerzo. Los resultados de nuestro grupo con el Brudy Plus se ven confirmados en la literatura científica más reciente con fuentes biodisponibles de DHA (Peoples GE et al., 2008; Walser B and Stebbins CL 2008). CONCLUSIONES Por todo ello, no existe fundamento para administrar grasa durante el esfuerzo al deportista, excepto en situaciones muy puntuales o en deportes extremos, como el montañismo (altitud y frío). El ejercicio en altitud parece que incrementa la utilización de glucosa como sustrato energético frente a los ácidos grasos libres (Butterfield GE et al., 1999) (Askew EW 2004) (Kayser B 1994), aunque un alto consumo de grasa (52%) en la dieta, favorece la resistencia a la hipoxia en ratas (Purshottam, T 1977). Este tipo de dietas ricas en grasa son bien toleradas por el deportista (364 g de grasa al día no produce aumento de cuerpos cetónicos ni alteraciones digestivas) (Nag PK et al., 1978). De hecho, en un estudio realizado en una expedición al Everest, los montañeros prefirieron una comida rica en grasas frente a otra rica en carbohidratos (Young PM et al.,1989). Lo que sí que es importante es equilibrar el tipo de grasas que se ingiere y, especialmente la relación n6/n3. Enlaces en inglés Muy interesante por su contenido en citas bibliográficas los ácidos grasos poliinsaturados http://lansbury.bwh.harvard.edu/polyunsaturated_fatty_acids.htm Generalidades de lípidos THE Medical Biochemistry Page Enlace muy importante para ver la alteración de las propiedades de la membrana celular en función del tipo de ácido graso que conforma el fosfolípido de membrana http://efaeducation.nih.gov/sig/klaus.pdf DHA Brudy Technology - Science of Omega 3 - El DHA EPA http://www.pdrhealth.com/drug_info/nmdrugprofiles/nutsupdrugs/eic_0104.shtml CLA http://www.jlr.org/cgi/content/full/40/8/1426 http://www.jlr.org/cgi/content/full/43/12/2112 Recomendaciones de la American Heart Org. en torno a la ingesta de n3. http://www.americanheart.org/presenter.jhtml?identifier=4632 Brudy Plus Brudy Technology - Science of Omega 3 - Bienvenido a Brudy Technology Bibliografía Askew EW Environmental and physical stress and nutrient req...[Am J Clin Nutr. 1995] - PubMed Result Butterfield GE et al., 1999 Nutrient requirements at high altitude. [Clin Sports Med. 1999] - PubMed Result Chardigny JM et al., 2003 Effect of conjugated FA on feed intake, body compo...[Lipids. 2003] - PubMed Result Doyle L et al., 2005 Effect of dietary supplementation with conjugated ...[Eur J Clin Nutr. 2005] - PubMed Result Jewell C, and Cashman KD 2003 The effect of conjugated linoleic acid and medium-...[Br J Nutr. 2003] - PubMed Result Kayser B 1994 Nutrition and energetics of exercise at altitude. ...[Sports Med. 1994] - PubMed Result Campbell B and Kreider RB 2008 Conjugated linoleic acids. [Curr Sports Med Rep. 2008 Jul-Aug] - PubMed Result Li Y and Watkins BA 1998 Conjugated linoleic acids alter bone fatty acid co...[Lipids. 1998] - PubMed Result Lowery LM, Appicelli PA and Lemon PWR. Conjugated linoleic acid enhances muscle size and strength gains in novice bodybuilders. Med. Sci. Sports Exercise 30:S182 (1998). Martin WH et al 1993 Effect of endurance training on plasma free fatty ...[Am J Physiol. 1993] - PubMed Result Nag PK et al., 1978 Optimal rate of work for mountaineers. [J Appl Physiol. 1978] - PubMed Result Peoples GH et al., 2008 Fish oil reduces heart rate and oxygen consumption...[J Cardiovasc Pharmacol. 2008] - PubMed Result Purshottam T et al., 1977 Altitude tolerance in rats in relation to carbohyd...[Aviat Space Environ Med. 1977] - PubMed Result Sadur CN and Eckel RH 1982 (acceso libre) Insulin stimulation of adipose tissue lipoprotein ...[J Clin Invest. 1982] - PubMed Result Sebedio JL et al., 1999 Recent advances in conjugated linoleic acid resear...[Curr Opin Clin Nutr Metab Care. 1999] PubMed Result Sugano M et al. 1998 Conjugated linoleic acid modulates tissue levels o...[Lipids. 1998] - PubMed Result Turek JJ, Li Y, Schoenlein IA, Allen KGD, Watkins BA: Modulation of macrophage cytokine production by conjugated linoleic acids is influenced by the dietary n-6:n-3 fatty acid ratio. J Nutr Biochem 9: 258–266, 1998 Walser B and Stebbins CL Omega-3 fatty acid supplementation enhances stroke...[Eur J Appl Physiol. 2008] - PubMed Result Watkins BA and Seifert MF. 2000 (acceso libre) http://www.jacn.org/cgi/content/full/19/4/478S Young PM et al., 1989 Operation Everest II: plasma lipid and hormonal re...[J Appl Physiol. 1989] - PubMed Result Zambell KL 2001 Conjugated linoleic acid supplementation in humans...[Lipids. 2001] - PubMed Result 3.2.- Hidratos de Carbono 3.2.a.- Generalidades La glucosa, también denominada dextrosa, se caracteriza por su sabor dulce y es el principal nutriente de las células, a las que llega por la sangre; no suele estar libre en los alimentos, sino formando oligo o polisacáridos, si bien se encuentra libre en frutas y otros vegetales, procedente de la hidrólisis del almidón. La fructosa o levulosa, está presente principalmente en las frutas (también en la miel, junto a la glucosa), y es el azúcar más dulce; la galactosa no existe generalmente libre, sino formando parte de otros azúcares, como la lactosa de la leche y algunos oligosacáridos. Entre los disacáridos, destacan: sacarosa, maltosa y lactosa. En los alimentos son mucho más abundantes que los monosacáridos. La sacarosa es el azúcar que denominamos como tal; se encuentra en la caña de azúcar y en la remolacha, que es de donde se extrae, así como en la fruta; esta formada por una molécula de glucosa y otra de fructosa, cuya unión se rompe fácilmente; es muy dulce y se digiere y absorbe con facilidad. La maltosa está formada por dos moléculas de glucosa y se genera principalmente a partir del almidón; es menos dulce que la sacarosa. La lactosa o azúcar de leche está formada por glucosa más galactosa; es poco dulce y se hidroliza por la lactasa, presente en el intestino, cuya ausencia da lugar a un trastorno denominado intolerancia a la lactosa En el momento de la iniciación del esfuerzo se origina una neta liberación de glucosa que cesa al cabo de unos minutos, para aumentar la captación. Durante el ejercicio, la utilización de la glucosa se correlaciona de forma curvilínea con la intensidad. En las primeras etapas del ejercicio de intensidad moderada el combustible utilizado por el músculo es la glucosa, de manera que cerca de la tercera o la cuarta parte del total de la energía consumida la proporciona este azúcar (Coggan 1991). A medida que el esfuerzo se hace más intenso, la captación de glucosa se incrementa, de manera que al llegar al 100% del VO2max el consumo puede alcanzar hasta 1,4 gr/minuto. En este momento, y como consecuencia de la glucogenolisis sobrevenida, la glucosa-6-fosfato produce la inhibición de la enzima hexocinasa, lo que favorece la acumulación de la hexosa en el músculo en actividad. Sin embargo, ni el aporte de insulina ni el de glucosa al músculo en actividad originado por el mayor flujo vascular, son suficientes para explicar el aumento de la captación de glucosa, por lo que, con toda seguridad, existen algunos factores locales que colaboran a ello, además del aumento de la disponibilidad de la glucosa. La activación de las enzimas oxidativas y de las glucolíticas y el cambio de lugar del GLUT 4 (transportador de la glucosa) desde su localización intracelular a la membrana plasmática influyen en el proceso, al facilitar la difusión de la glucosa y su posterior utilización. De cualquier manera, una buena disponibilidad del CH durante el ejercicio es indispensable para conseguir una captación eficaz por parte del músculo (Tsintzas K et al., 2000). 3.2.b.- Disponibilidad de la glucosa sanguínea y utilización del glucógeno muscular. Es un hecho perfectamente conocido, que la fatiga muscular originada por el ejercicio intenso de larga duración, se debe fundamentalmente a la disminución de los carbohidratos. Según eso, la administración de CH durante el ejercicio mejoraría el rendimiento deportivo (Tsintzas K et al., 2000). Esta mejoría se debería al aumento de la glucosa circulante y a la intensa oxidación de los CH cuando la concentración del glucógeno muscular disminuye a consecuencia del ejercicio. Pero también es posible que los efectos favorables tengan algo que ver con un aumento de la síntesis del glucógeno en las fibras musculares, facilitado por la gran oferta de glucosa exógena. La opinión mayoritaria de los científicos con experiencia en este campo es que, al menos en los ciclistas, cuando el ejercicio se realiza a intensidades entre el 70 y 75% del VO2max, la utilización del glucógeno muscular no se modifica (McConell G et al., 1994). Sin embargo, según (Tsintzas et al., 2000) el ejercicio en la cinta sin fin al 70% del VO2 pico, disminuye la utilización del glucógeno muscular, especialmente en las fibras del tipo I. 3.2.c.- Interacción entre el glucógeno muscular y la glucosa de la sangre durante el ejercicio. En los estadios iniciales del ejercicio el glucógeno muscular se degrada con bastante rapidez, siendo tanto mayor dicha degradación cuanto más intenso es el esfuerzo. Este hecho es debido a un aumento de la actividad de la enzima glucógeno fosforilasa. En ello desempeña un cometido muy importante la disponibilidad o abundancia del glucógeno para ser utilizado durante el esfuerzo, salvo cuando éste último es de gran intensidad, en cuyo caso, el aumento del recambio del ATP, los niveles elevados del fósforo (Pi), los del Ca intramuscular, y los de la adrenalina plasmática son los responsables directos de la estimulación de la glucogéno fosforilasa y por tanto, del incremento de la glucogenolisis. Aquí, la disponibilidad de glucógeno no posee efectos aditivos. Un factor muy interesante en la regulación de la glucogenolisis lo constituye el metabolismo de las grasa durante el ejercicio, de tal manera que el aumento de los ácidos grasos libres (AGL) en el plasma y por tanto, su disponibilidad; disminuye la utilización del glucógeno por el músculo (Dyck 1996) en tanto que la disminución de los AGL ejerce el efecto contrario. 3.2.d.- Carbohidratos durante la competición. La toma de CH durante la realización de un esfuerzo mayor de 90 minutos de duración y de intensidad superior al 70% de la VO2 max aumenta el rendimiento, ya que en los últimos momentos del ejercicio evita el descenso de la glucosa sanguínea, a la vez que facilita su oxidación, y disminuye la degradación del glucógeno (Yaspelkis BB et al., 2002). En esfuerzos intermitentes también es importante la utilización de CH (McGregor SJ et al., 1999). El consumo de carbohidratos durante el esfuerzo de resistencia atenúa el descenso de los niveles de glucógeno intramusculares, pero no afecta al rendimiento deportivo del atleta (Haff G 2000). Una comida con carbohidratos de alto índice glucémico (1 g/kg ) 65 minutos antes del esfuerzo, disminuye la glucemia y aumenta la lactacidemia con respecto a otra con bajo índice glucémico, pero no altera el rendimiento (Stannard SR et al., 2000). Es posible que la clase y la intensidad del ejercicio expliquen las diferencias observadas. En la última fase de las actividades de larga duración realizadas al 70-75% de la VO2 max, la glucosa debe acceder al músculo a razón de 1g/minuto. Parece ser que los CH en solución son los que producen los mejores resultados debido a su facilidad para abandonar la cavidad gástrica. La velocidad del vaciamiento gástrico depende de la concentración en CH y del volumen del líquido ingerido. Según Mitchell, con soluciones iguales o inferiores de CH al 12% se consigue que el estómago vacíe 1 g/min (Mitchell JB 1988). Nosotros hemos observado que el vaciamiento gástrico (medido por gammacámara tras marcar la bebida con isótopos), es muy dependiente de la densidad energética de la bebida (Martínez-Gonzálvez AB et al., 2005). Las bebidas deportivas se diseñaron con el objetivo de satisfacer con la ingesta de un único producto las necesidades de energía, líquidos y/o electrolitos de los deportistas profesionales, pero actualmente casi todos los aficionados a la práctica deportiva consumen este tipo de bebidas, porque se sienten atraídos por los mensajes que las acompañan, y recurren a ellas para mejorar su condición física o acelerar su recuperación. Existen tres tipos de bebidas deportivas (isotónicas, hipotónicas e hipertónicas) presentando generalmente unos componentes comunes como son: agua, carbohidratos (glucosa, fructosa, sacarosa, maltodextrina) y electrolitos (sodio, cloro, potasio, fósforo, magnesio, calcio,…); algunas marcas incluyen vitaminas, antioxidantes, edulcorantes y aromatizantes; y muy pocas, proteínas. La mayoría de bebidas comercializadas son isotónicas o hipotónicas. 1. Bebidas isotónicas: contienen similar concentración de solutos (azúcares y electrolitos, fundamentalmente) que la sangre (270 – 330 miliosmoles/litro), por ello el líquido ingerido sale del estómago al intestino delgado, y de éste al torrente sanguíneo sin dificultad, lo que favorece una rápida y óptima asimilación de sus componentes. Están indicadas en ejercicios intensos, de moderada y larga duración y/o en condiciones de alto calor ambiental, ayudando a evitar patología por calor, retrasar la fatiga, mejorar el rendimiento y acelerar la recuperación. Estudios recientes afirman que, incluso en ejercicios intensos de menos de 1 hora de duración (hasta este momento se afirmaba que sólo era necesario consumir agua durante los mismos), la ingesta de hidratos de carbono mejora el rendimiento. Añadir proteínas a las bebidas deportivas puede acelerar la síntesis de glucógeno muscular unas 4-6 horas y el efecto anabólico tras el ejercicio, lo cual sería muy adecuado para entrenamientos o competiciones sucesivas; lo aconsejable sería añadirlo a bebidas isotónicas, manteniendo su osmolaridad. 2. Bebidas hipotónicas: contienen una concentración de solutos menor que la sanguínea. El mejor ejemplo de bebida hipotónica es el agua, y la mayoría de bebidas existentes en el mercado. Proporcionan una pobre recuperación de iones y carbohidratos, no evitan la deshidratación en ejercicios intensos y de larga duración, y la absorción desde el intestino a sangre no es más rápida que en el caso de las bebidas isotónicas. 3. Bebidas hipertónicas: la concentración de solutos por unidad de volumen es mayor que la sanguínea. La principal indicación de este tipo de bebidas puede ser en la recuperación tras la actividad física, para la rápida recuperación del glucógeno muscular, ya que, normalmente, contienen una alta carga de carbohidratos (mayor del 10%). Las bebidas energetizantes se introdujeron en nuestro medio hace unos 4 años, y países como España, Inglaterra, Alemania y Austria son los mayores consumidores a nivel mundial. Su consumo suele darse con fines de diversión, y también asociado a la práctica de algún deporte como “rehidratante”; en este último caso, existe un considerable error de información, dado que estas bebidas, por su composición, son evidentemente estimulantes, y en modo alguno rehidratantes. Inconvenientes del consumo de estas bebidas: 1. En el etiquetado de algunas de ellas no constan algunos componentes. 2. El contenido de algunas de ellas puede ocasionar una deficiente absorción de líquidos y nutrientes desde el intestino, provocando molestias gastrointestinales. 3. No se deben consumir por personas hipertensas o con patología cardíaca por los estimulantes que contienen. 4. Las altas dosis de cafeína que contienen suelen producir cefalea. 5. Suelen ser muy costosas, y por su composición, no es conveniente su uso por parte de los atletas, pueden dar positivo en los controles de doping. 6. En deportistas, si se ingiere una vez iniciado el ejercicio, puede tener efecto laxante y diurético, pudiendo deteriorar el rendimiento y ser perjudicial para el sistema cardiovascular. RESUMEN Las bebidas deportivas se diseñaron con el objetivo de satisfacer con la ingesta de un único producto las necesidades de energía, líquidos y/o electrolitos de los deportistas profesionales, pero actualmente casi todos los aficionados a la práctica deportiva consumen este tipo de bebidas, porque se sienten atraídos por los mensajes que las acompañan, y recurren a ellas para mejorar su condición física o acelerar su recuperación. Existen tres tipos de bebidas deportivas (isotónicas, hipotónicas e hipertónicas) presentando generalmente unos componentes comunes como son: agua, carbohidratos (glucosa, fructosa, sacarosa, maltodextrina) y electrolitos (sodio, cloro, potasio, fósforo, magnesio, calcio,…); algunas marcas incluyen vitaminas, antioxidantes, edulcorantes y aromatizantes; y muy pocas, proteínas. La mayoría de bebidas comercializadas son isotónicas o hipotónicas. Son bebidas elaboradas con una base de agua, agregándole una alta concentración de hidratos de carbono y cafeína (el agua vehiculiza los nutrientes, los carbohidratos aportan energía y la cafeína estimula en sistema nervioso central); algunas también contienen hierbas (extractos de guaraná, ginseng, nueces de cola, hojas de hierba mate,..), proteínas y aminoácidos (ramificados como valina, leucina e isoleucina; arginina, taurina, glutamina…), creatina, carnitina, vitaminas y minerales, triglicéridos de cadena media, oxígeno, saliva de avispas (VAAMTM), pero en tan pequeñas cantidades que no producen efecto alguno sobre el rendimiento. Enlace del Consenso de FEMEDE sobre bebidas para deportistas (muy importante) http://femede.es/documentos/Consenso%20hidratacion%20ingles.pdf Enlace de consulta http://europa.eu.int/comm/food/fs/sc/scf/out64_en.pdf Bibliografía Coggan AR, 1991 Plasma glucose metabolism during exercise in human...[Sports Med. 1991] - PubMed Result Martínez-Gonzálvez AB et al., 2005 [Scintigraphic study of gastric emptying of rehydr...[Rev Esp Med Nucl. 2005 Jan-Feb] - PubMed Result McConell G et al., 1994 Effect of carbohydrate ingestion on glucose kineti...[J Appl Physiol. 1994] - PubMed Result McGregor SJ et al., 1999 The influence of intermittent high-intensity shutt...[J Sports Sci. 1999] - PubMed Result Mitchell JB et al., 1988 Effects of carbohydrate ingestion on gastric empty...[Med Sci Sports Exerc. 1988] - PubMed Result Stannard SR et al., 2000 The effect of glycemic index on plasma glucose and...[Int J Sport Nutr Exerc Metab. 2000] PubMed Result Tsintzas K et al., 2000 Effect of carbohydrate feeding during recovery fro...[Int J Sports Med. 2003] - PubMed Result Yaspelkis BB et al., 2002 Resistance training increases glucose uptake and t...[Acta Physiol Scand. 2002] - PubMed Result
 0
0
Anuncio
Documentos relacionados








Descargar
Anuncio
Añadir este documento a la recogida (s)
Puede agregar este documento a su colección de estudio (s)
Iniciar sesión Disponible sólo para usuarios autorizadosAñadir a este documento guardado
Puede agregar este documento a su lista guardada
Iniciar sesión Disponible sólo para usuarios autorizados