Virus y Cáncer Revisión Bibliográfica 2005-2009 Sandra Gómez Talavera Paloma Gallardo Culebradas Carmen Garcías de España Francisco Fernández-Capel Josep Mª Gaytán Sansa Miren Díez Zapirain Coordinador: Prof. Don Vicente Domínguez Rojas Catedrático del Departamento de Medicina Preventiva, Salud Pública e Historia de la Ciencia. Universidad Complutense de Madrid. Índice Virus de la Hepatitis C (VHC) Sandra Gómez Talavera • Mecanismos moleculares de la oncogénesis • Implicación directa de las proteínas del VHC • Otros mecanismos implicados en el efecto pro-cancerígeno de VHC • La carcinogénesis en la infección por VHC; un mecanismo diferente al de otros cánceres Virus de la Hepatitis B (VHB) • Patogénesis del Carcinoma Hepatocelulara ‐ Mecanismos moleculares de la oncogénesis ‐ Implicación directa del VHB en el hepatocarcinoma Paloma Gallardo Culebradas Virus de Epstein-Barr (VEB) • Epidemiología • Mecanismos moleculares de la oncogénesis ‐ Linfoma de Burkitt ‐ Carcinoma nasofaríngeo Carmen Garcías de España Virus del Herpes Humano 8 (VHH-8) • Virología ‐ Genoma de VHH-8 ‐ Patogenia de la infección • Epidemiología. Transmisión ‐ Epidemiología ‐ Transmisión • Enfermedades asociadas ‐ Primoinfección ‐ Sarcoma de Kaposi, SK ‐ Linfoma de Derrame Primario, LDP ‐ Enfermedad de Castleman Multicéntrica, ECM ‐ Otras asociaciones • Oncogénesis • Perspectivas de futuro Francisco Fernández-Capel Virus Linfotropo T Humano Tipo I (VLTH-I) • Mecanismos moleculares de la oncogénesis: ‐ Cofactores: Josep Mª Gaytán Sansa Miren Díez Zapirain Virus Del Papiloma Humano (VPH) • Cofactores infecciosos en el cáncer de cérvix • Mecanismos moleculares de la oncogénesis ‐ Capacidad transformadora del VPH ‐ Propiedades bioquímicas de las proteínas de VPH ‐ Propiedades biológicas de las proteínas de VPH • Evidencia experimental del rol de VPH mucosos de alto grado de en la conversión maligna y el cáncer cervical humano ‐ Requerimiento de la expresión génica de VPH para el crecimiento celular y la invasión ‐ Integración de secuencias de VPH ‐ Anomalías cromosómicas en los cánceres asociados a VPH ‐ Alteraciones de proto-oncogenes específicos VIRUS Y CÁNCER El cáncer es una de las cargas más importantes de la medicina moderna a nivel mundial y uno de los retos más interesantes en materia de investigación en la actualidad. Con el paso del tiempo se han ido conociendo distintos agentes que pueden influir en su origen. Tradicionalmente se ha asociado con radiaciones ionizantes y carcinógenos químicos pero el abanico se ha ido ampliando llegando a abarcar aspectos tan dispares como dieta o estilo de vida. Las infecciones crónicas suponen la causa de hasta un 20% de las enfermedades tumorales. De ellas las virales han adquirido una importancia creciente con el avance de las técnicas diagnósticas de biología molecular e inmunohistoquímica, que han permitido identificar cada vez más agentes en relación con enfermedades tumorales. Entre ellos los más destacados son: los Virus de la Hepatitis B y C, el Virus de Epstein Barr, el Virus del Herpes Humano 8, Virus Linfotropo T Humano Tipo 1 y el Virus de Papiloma Humano. El conocimiento de los diversos mecanismos que estos virus emplean para el desarrollo de distintos tumores es de gran importancia para encontrar nuevas dianas terapéuticas que permitan un manejo más específico de estas neoplasias. Este artículo hace una revisión de la literatura publicada en los últimos años sobre los principales virus oncogénicos identificados y los mecanismos de los que se valen para el desarrollo y promoción de los distintos tumores con los que se asocian. VIRUS DE LA HEPATITIS C (VHC) VHC es un virus RNA de la familia Flaviviridae. No se integra en el genoma del hospedador. El genoma viral codifica para las proteínas estructurales Core (C), proteína de la cápside y Envoltura (E), que son dos glicoproteínas E1 y E2; así como para las no estructurales (NS), que son NS 2,3,4 A, 5 A y 5 B. La región 5' no traducida y el gen que codifica el centro se mantienen muy constantes en los diversos genotipos. Dentro de la secuencia codificadora de E2 hay dos regiones hipervariables (que cambian en los distintos especimenes). Éstas son importantes para la capacidad del virus de evadir la neutralización del sistema inmune, conduciendo a la aparición de múltiples quasi especies genéticas variantes del VHC en el infectado. Existen seis genotipos de VHC. El genotipo más frecuente en España es el 1. La severidad del daño hepático no se relaciona con el genotipo ni cambian en el curso de la infección. El carcinoma hepatocelular (CHC) representa el 6% de los nuevos casos de cáncer diagnosticados en el mundo, éste constituye una de las neoplasias con menos posibilidades de curación y es la tercera causa más frecuente de muerte por cáncer a nivel mundial. Parece ser que la contribución del VHC al desarrollo de carcinoma hepatocelular aumenta en la edad media y en individuos mayores en países desarrollados y en algunos en desarrollo (1). El incremento de incidencia y mortalidad de CHC observado en la mayoría de países industrializados posiblemente refleje el incremento de la prevalencia de la infección por VHC (2). Mecanismos moleculares de la oncogénesis (2) Como la mayoría de los cánceres, la hepatocarcinogénesis es un proceso de múltiples pasos, incluyendo distintas alteraciones genéticas que finalmente conducen a la transformación maligna del hepatocito. El análisis detallado del desarrollo de CHC en animales de experimentación y la comparación de los resultados con CHC en humanos ha identificado una diversidad de alteraciones genómicas y moleculares en el desarrollo completo de hepatocarcinoma y en una menor extensión en las lesiones precursoras definidas como preneoplásicas. A pesar de los numerosos estudios realizados en este sentido, carecemos de un entendimiento coherente de los mecanismos de desarrollo de CHC. En particular, de la contribución de los diferentes factores etiológicos y de sus interacciones en hepatocarcinogénesis, que son aún pobremente entendidos. Además no tenemos identificadas las alteraciones genómicas y/o moleculares que podrían mejorar el diagnóstico o ser diana para intervenciones terapéuticas. La transformación maligna de los hepatocitos se cree que ocurre, a pesar de todos los agentes etiológicos, por un incremento en el recambio celular, inducido por la lesión hepática crónica y regeneración, en un contexto de inflamación y daño oxidativo al ADN; este microambiente favorece la aparición de alteraciones genéticas y epigenéticas, que incluyen activación de oncogenes, inactivación de genes supresores de tumores, así como sobreexpresión de factores de crecimiento y factores angiogénicos. Está demostrado que el principal factor de riesgo para el desarrollo de CHC es la cirrosis, apareciendo el 70-90% de CHC sobre un hígado cirrótico; siendo diferente el riesgo en función de la etiología, duración y grado de actividad. Existen al menos cuatro caminos que regulan la proliferación celular y /o la muerte celular que están alterados en el CHC; estos son: proteína del Retinoblastoma (pRb) y proteína p53 (genes supresor de tumores, freno a la proliferación), TGF-β (regulador del crecimiento celular) y β-catenina (componente de la vía de señalización de Wnt). • Expresión disregulada de β-catenina Mutación en el gen de β-catenina y/o alteraciones en la señal celular de Wnt parecen estar implicados en más del 50% de los casos de CHC. La señal Wnt/βcatenina está implicada en el control del desarrollo, adhesión y proliferación celular. Los niveles de β-catenina están regulados por la degradación dependiente del proteasoma, que está a su vez retrocontrolado por la actividad de APC (poliposis adenomatosa de colon), proteína Axin-1 y GSK- 3β. La acumulación de complejos de β-catenina nuclear conduce a la transcripción sin control de varios genes implicados en el control del ciclo celular. Un análisis multivariante ha demostrado peor pronóstico y mayor incidencia de recurrencias en estos casos (3). Se han descrito mutaciones de la β-catenina en un 18-41% de los CHC, mutaciones que la previenen de ser fosforilada y de este modo de ser degradada (3). Las mutaciones somáticas del gen APC son poco frecuentes y hay casos en los que existe mutación del gen Axin-1 con acumulación de β-catenina en ausencia de mutaciones en el gen de ésta. • El control de la expresión de los factores de crecimiento y la activación de sus caminos de señalización también juegan un papel importante en la hepatocarcinogénesis; ejemplo: IGF-II (factor de crecimiento de insulina tipo II), receptor de insulina, HGF (factor de crecimiento del hepatocito) y TGF-α/β. • También se ha descrito una sobre expresión de factores angiogénicos del endotelio vascular y angiopoyetina-2. • Es relativamente frecuente también encontrar pérdidas alélicas en los cromosomas: 1, 2q,4,5q,6,q,8,9 10q,11p,13q,14q,16, 17 y 22q; sugiriendo que éstos sitios pueden esconder genes supresores de tumores relevantes para la patogénesis del CHC. Hecho confirmado con estudios de genómica usando técnicas de hibridación.; aún así sólo unos pocos de los genes supresores de tumores localizados en esas regiones suprimidas están claramente involucrados en el subconjunto de CHCs; por ejemplo: el gen que codifica para p53 está localizado en el 17p, y el gen de la pRb está en el 13q (3). • Proteína del Retinoblastoma; está alterado en más del 50% de casos de CHC debido a cambios genéticos y epigenéticos. La pRb, su regulador y ciclina D están implicados en la progresión G1/S del ciclo celular. Se han descrito tanto mutaciones en el gen de la proteína reguladora, como su inactivación por metilación. Recientemente se ha descrito la sobreexpresión de Gankyrina, proteína que media la degradación de la pRb por el proteasoma 26. • El gen supresor de tumores p53, se encuentra frecuentemente alterado, habiéndose descrito mutaciones diversas en su posición y naturaleza; las cuales afectan entorno a 200 codones dispersos principalmente a lo largo de la región central del gen (3). La mutación G – T en la tercera base del codón 249 del gen que codifica para p53 se encuentra presente en 50- 70% de pacientes con CHC del sur de África y en áreas de China. Ésta mutación puntual (sustitución de una Arginina por Serina) y la expresión de una unión anómala al ADN de la p53 mutada, están asociadas con una elevada contaminación de la comida por la micotoxina aflatoxina B1. En otras regiones dónde los niveles de aflatoxina en la comida eran bajos o indetectable las mutaciones de p53 se muestran en un bajo porcentaje (< 4%) y pueden detectarse mutaciones genéticas no específicas. La relación entre p53 y p73 (que es también gen supresor de tumores) está también disregulada en CHC. No se han descrito mutaciones en el gen de p73, pero la isoforma con delección del amino terminal DNp73, que actúa cómo dominante trans-represor de ambos p53 y proteína pro-apoptótica TAp73, está acumulada progresivamente en hepatitis crónica, cirrosis y CHC, y confiere a las células del CHC un fenotipo quimioresistente. DNp73 es expresado por un promotor alternativo, que es controlado en los hepatocitos por NF-κB (factor de transcripción) y β-catenina. Esta observación conecta la activación de la β-catenina con la inactivación funcional de p53 y TAp73 que es mediada por la sobreexpresión de DNp73 y no requiere la mutación inactivadora del gen de p53. La pérdida de la pRb y p53 conducen a la inestabilidad genómica, que juega un importante papel permisivo en la transformación maligna. • Evidencias recientes sugieren que la disfunción de la telomerasa conduce a inestabilidad cromosómica de los telómeros, ocurriendo en los estadios tempranos de hepatocarcinogénesis; mientras la activación de la telomerasa, enzima que previene los acortamientos de los telómeros y prolonga la vida celular, ocurre tardíamente en la progresión a CHC. • Metilación de los promotores de varios genes individuales, ej: p73, APC, Ecadherina… podrían interferir con las uniones a factores de transcripción y otros mecanismos de regulación, lo que conllevaría un descenso en la exposición de los genes correspondientes. También se ha descrito metilación disbalanceada, áreas dentro de un mismo gen hipermetiladas, alternando con otras hipometiladas, en genes supresores de tumores. • Recientemente se ha descrito la implicación de los microRNAs en la hepatocarcinogénesis, los microRNAs son hebras de RNA monocatenario, formados por 21-25 nucleótidos capaces de regular la expresión postranscripcional de otros genes, entre los cuales se encuentran algunos oncogenes (ej.ras) y genes supresores de tumores (ej.PTEN) (3). En resumen, podemos observar como el Carcinoma Hepatocelular es un tumor genéticamente muy heterogéneo. El análisis amplio de las alteraciones genéticas en CHC muestra que éstas no están distribuidas al azar en los tumores; pero sí están claramente asociadas en grupo, y han permitido la definición de dos mecanismos principales de hepatocarcinogénesis: - Alteraciones genéticas acumuladas a través de inestabilidad cromosómica. - Activación de Wnt; en la cual la vía activada por la mutación de β-catenina es predominante. Éste es el más frecuentemente asociado con la ausencia de infección por VHB. Implicación directa de las proteínas del VHC Como el CHC asociado con la infección por el VHC ocurre tras muchos años de infección crónica (30 años de media, aunque se han observado periodos de latencia de 40-50 años) (4) y generalmente se ve precedido del desarrollo de cirrosis; el papel del daño hepático crónico (inflamación) seguido de regeneración, cirrosis y el desarrollo de CHC es la principal hipótesis. Pero el hecho de que sea extremadamente raro encontrar CHC en pacientes con hepatitis autoinmunes, en los cuales la inflamación severa en el hígado persiste indefinidamente, hace sospechar que es posible además que el VHC pueda contribuir por medio de mecanismos más directos en la transformación maligna de los hepatocitos (5); diferentes proteínas virales, sobretodo la proteína del core y NS5A han sido implicadas en la transformación y desarrollo de CHC. Las proteínas del VHC están implicadas en un amplio rango de actividades, incluyendo señalización celular, modulación de la traducción, transformación, apoptosis, ordenamientos de membrana, tráfico vesicular… Entre las múltiples interacciones que se producen entre el virus y los elementos celulares del hospedador, vamos a centrarnos en aquellos que son potencialmente relevantes para la transformación maligna de las células infectadas: • Proteína del core (2) Es la responsable de la traducción del RNA viral, para conseguir un virión completo (partícula viral completa, madura e infecciosa). La proteína del core interacciona con los genes supresores de tumores: p53, p73 y pRb, pero las consecuencias de estas interacciones no han sido bien aclaradas aún. La proteína del core también modula la expresión de la kinasa dependiente de ciclina (CDK), inhibidora de p21 (gen supresor tumores)/Waf1; p21/Waf1 es una diana transcripcional de p53 y regula las actividades del complejo ciclina/CDK involucrado en el control del ciclo celular y formación de tumor. La proteína del core del VHC es producida como una forma innata que es posteriormente procesada para producir la forma madura. La forma innata en el citoplasma incrementa la cantidad de p21/Waf1 por la activación de p53, y la forma madura en el núcleo disminuye la cantidad de p21/Waf1 por un camino independiente de la p53.También parece ser que las interacciones de la proteína del core del VHC con el receptor del factor de necrosis tumoral y el receptor de linfotoxina B están implicadas en la inhibición de la apoptosis. Asimismo se ha propuesto que la proteína del core presenta actividad inmunosupresora por medio de la interacción con los receptores del complemento en la célula T, lo que contribuye a la infección crónica. El mecanismo por medio del cual la proteína del core regula la transcripción parece ser indirecto, por medio de interacciones con moléculas citoplasmáticas (cascadas de activación, vía MAP kinasa). De hecho se ha observado que la expresión de la proteína del core induce la proliferación celular, síntesis de ADN y progresión del ciclo celular bien sola o en el contexto de la replicación del VHC. Variantes de la proteína del core que se han aislado del tumor hepático, pero no de los tejidos adyacentes no tumorales, son capaces de inhibir la señal de TGF-β; sugiriendo esto que durante la infección crónica las variantes virales que promueven la transformación celular, por suministrarle capacidad de expansión clonal a las células con resistencia a los efectos antiproliferativos del TGF-β, son seleccionadas activamente. Las cuasi especies del VHC parecen estar categorizadas en distintos tipos de células, como las células hepáticas o las células mononucleares de sangre periférica, y en el hígado se han aislado variantes virales diferentes en el tumor y regiones no tumorales, sugiriendo que esto podría contribuir a la carcinogénesis inducida por el VHC. Papel del TGF-β: Las señales inducidas por éste no tienen efecto únicamente en la proliferación, diferenciación y apoptosis, sino que además están relacionadas con los procesos de reparación hepática y de fibrogénesis por medio de su acción en la matriz extracelular. Los pacientes infectados con el VHC presentan niveles elevados de TGF-β, lo que se correlaciona con el grado de fibrosis. La proteína del core wild-type ha demostrado controlar la expresión del TGF-β a nivel trasnscripcional e inducir la síntesis del mismo en las células estrelladas hepáticas (células de Ito) promoviendo de este modo la fibrogénesis. El TGF-β puede al mismo tiempo favorecer la persistencia viral limitando la respuesta inmune antiviral. En resumen para conciliar éstos hallazgos aparentemente conflictivos sobre la acción de TGF-β, se cree que la proteína del core puede tener una acción dual sobre este sistema dependiendo de la fase del daño en la que se encuentre: - Infección reciente por VHC, contribuye a la fibrogénesis aumentando la síntesis de TGF-β. - Posteriormente, tras un largo periodo de fibrosis e inflamación y en el contexto de un hígado cirrótico, las variantes del core que contribuyen a la expansión clonal y la transformación celular por medio de la inhibición del camino antiproliferativo dependiente del TGF-β pueden surgir y ser seleccionadas. Las interacciones de la proteína del core con la membrana celular y las vesículas lipídicas, conducen a una alteración en el ensamblado y secreción de las lipoproteínas muy baja densidad (VLDL) y esteatosis hepática. En un estudio realizado con ratones transgénicos se observaron los efectos que producía la proteína del core sobre los hepatocitos: la proteína del core se localiza predominantemente en el citoplasma, asociada con las gotas de lípidos, pero también se encuentra presente en el núcleo y en la mitocondria. Esta produce un aumento de la actividad oxidativa a nivel hepático, incluso en ausencia de inflamación. Si la inflamación es inducida en un hígado con proteínas del core de VHC la producción de estrés oxidativo se intensifica a una extensión que no puede ser frenada por los mecanismos fisiológicos que lo antagonizan. Esto indica que la inflamación en infección crónica por VHC puede ser distinta a la que se observa en otras hepatitis, ej: la autoinmune. La base de la sobreexpresión de estrés oxidativo puede ser debida a una disfunción mitocondrial, por alteración a nivel del sistema de transferencia de electrones. La esteatosis hepática además podría actuar como un estímulo para la sobreproducción del estrés oxidativo (2;5). • Proteína E2 (2) El impacto de otras proteínas estructurales en la transformación maligna es más indirecto que el de la proteína del core. La glicoproteína E2 se ha visto que in vitro interfiere con la acción del interferón inhibiendo proteínas intermedias implicadas en el efecto del mismo. Además, una forma soluble de E2 interacciona con el marcador de superficie celular CD81 inhibiendo así la activación de los linfocitos T y células NK, in vitro. También interacciona con el receptor de las lipoproteínas de baja densidad (LDLR) en la superficie celular y activa una serie de señales intracelulares que promueven la proliferación y supervivencia celular. • Proteína NS3 (2) Es una proteína con un dominio serín proteasa, es capaz de inducir una transformación maligna de las células, pero no se conoce bien la conexión entre esta acción y el desarrollo de CHC. El efecto oncogénico de ésta podría incluir la interacción con p53. • Proteína NS5A (2) Está implicada en varias de las funciones celulares: apoptosis, transducción de señales, activación transcripcional y transformación celular. Estudios in vitro han demostrado que la interacción de ésta con proteína kinasa R (PKR), proteína protectora de la infección viral, dan resistencia al interferón, no conocido in vivo. Además estimula la IL-8, la cual inhibe el efecto antiviral del interferón. NS5A forma un complejo heteromérico con la proteína ligadora de la caja TATA (TBP) y la proteína p53 inhibiendo la unión de ambas a su ADN. Recientemente se ha observado que NS5A aumenta los niveles de β-catenina y estimula la transcripción dependiente de la misma, lo que sugiere que la señal Wnt/β-catenina es una diana común para las proteínas del VHB y VHC en el CHC humano. Todos éstos datos sobre funciones y propiedades de las diferentes proteínas codificadas por el VHC son potencialmente relevantes en el desarrollo de CHC y progresión tumoral; aún así es importante destacar que la mayoría de las alteraciones descritas se han identificado en sistemas artificiales, con un elevado nivel de exposición de proteínas virales; por tanto, estos hallazgos in vitro pueden no ser todos aplicables a la infección in vivo, dónde las proteínas se expresan en mucha menor concentración . De hecho aún no se sabe si y qué productos de los genes virales son necesarios in vivo para establecer CHC. Otra cuestión abierta es en qué momento, si es que este existe, el desarrollo del carcinoma se convierte en independiente del virus, así como el relativamente bajo y lento impacto que tiene en el desarrollo de CHC el tratamiento antiviral. Otros mecanismos implicados en el efecto pro-cancerígeno de VHC (6) Parece ser que el propio VHC es capaz de inducir por sí mismo la producción de radicales libres (que suponen un daño oxidativo al ADN) per se, no sólo por la acción de la inflamación. En la fase de iniciación del proceso los radicales de oxígeno podrían interactuar directamente con el ADN genómico, dañando genes que controlan el crecimiento de la célula y su diferenciación, o bien, ellos pueden aumentar la actividad cancerígena de xenobióticos (compuestos cuya estructura química en la naturaleza es poco frecuente o inexistente debido a que son compuestos sintetizados por el hombre en el laboratorio) facilitando su activación a compuestos reactivos. En fases progresivas de la carcinogénesis podrían estimular directamente el crecimiento de las células malignas. Entre los muchos radicales libres de oxígeno producidos en los procesos de inflamación, el más lesivo es el radical hidroxilo, que ha demostrados ser el responsable de modificar un determinado número de bases, incluyendo la formación de 8-hidroxi-desoxiguanosina (8 OHdG), que se produce por modificación de guanina, e induce una mutación puntual en la hebra de ADN y es por consiguiente un signo de daño en el ADN. Esta lesión en condiciones normales sufre un proceso de reparación específico por una proteína codificada por el gen OGG1 (tipo1a es el expresado principalmente en las células humanas). Se ha observado que en algunos cánceres humanos se encuentran alteraciones en los genes que codifican para estas proteínas implicadas en los procesos de reparación del daño al ADN. En cualquier caso reparado o no, el resultado del daño oxidativo in vivo permanece desconocido, mientras estudios indican que la modificación introducida por 8-OHdG persiste residualmente, acumulándose en el ADN de la célula, sugiriendo que esta lesión es mutagénica y por tanto potencialmente cancerígena. El daño hepático inducido por el virus hepatitis C se caracteriza por mayor formación de radicales libres, revelados por niveles elevados a nivel hepático y en suero de los productos de peroxidación lipídica y/o oxidación proteica. En el intento de establecer la relación entre esta actividad oxidativa y el proceso de carcinogénesis hepática se estudió en sujetos VHC positivo los niveles de 8-OHdG en el ADN de los leucocitos circulantes y se dedujo que existía un daño progresivo del genoma no sólo en pacientes con hepatitis crónica y cirrosis, sino también en los pacientes VHC positivo con niveles normales de alanino transaminasa (infección silente). Esto apoya los conocimientos que se tenían y muestra como los sujetos con transaminasas normales pueden presentar grados variables de daño hepático, además confirma la presencia de daño en el tejido en relación con la formación de radicales libres de oxígeno. Podemos deducir que el daño oxidativo en el ADN es un evento temprano en el daño hepático por VHC, acumulándose. Además se ha observado que el genotipo viral es un factor de riesgo para mayor severidad del daño hepático, en términos biológicos al menos; así se ha observado que el genotipo viral 1b, uno de los de peor pronóstico clínico, se asociaba a mayor daño oxidativo. No está claro cuáles son los factores del paciente que determinan la extensión del daño hepático mediado por VHC. Varía en función de la respuesta moduladora a la inflamación, polimorfismo del factor de necrosis tumoral (TNF) e IL-1, y el polimorfismo del gen OGG1. Es poco frecuente encontrar sujetos homocigotos para las mutaciones en el gen OGG1, siendo más frecuente encontrar heterocigotos, en cualquier caso estas mutaciones confieren un mayor riesgo de evolución a cirrosis o desarrollo de CHC. Tanto TNF-α como la IL-1B controlan el balance entre los mecanismos de proliferación y apoptosis, implicados en determinar daño celular hepático y progresión a cáncer. La carcinogénesis en la infección por VHC; un mecanismo diferente al de otros cánceres (5) En estudios de investigación de otros cánceres, la teoría de Kinzler y Vogelstein ha ganado popularidad; proponen que el desarrollo de cáncer colorectal es inducido por la acumulación de un juego completo de mutaciones genéticas celulares. Ellos dedujeron que la mutación en el gen APC por inactivación, este en K-ras por activación y este en p53 por inactivación acumuladas cooperarían hacia el desarrollo del tumor. Su teoría se hizo extensiva también al proceso de carcinogénesis de otros tumores y fue denominada: carcinogénesis tipo Vogelstein. Pues basándose en el efecto que hemos visto que la proteína del core tiene directamente en el desarrollo de CHC en infección crónica por VHC se podría introducir un mecanismo diferente de carcinogénesis en este caso. Debemos considerar un proceso gradual en la inducción de todos los cánceres, y es obligatorio para la hepatocarcinogénesis que varias mutaciones se acumulen en los hepatocitos, pero algunos de estos pasos, sin embargo, se pueden saltar en el desarrollo de CHC en VHC, a lo cual podría contribuir la proteína del core. Este conjunto de efectos alcanzados por la expresión de la proteína viral podrían ser responsables de la inducción del CHC, incluso en ausencia del juego completo de aberraciones genéticas, necesarias para la carcinogénesis. Considerándolo como un proceso tipo no Vogelstein para la inducción de CHC, podrían explicarse muchos eventos inusuales ocurridos en los portadores de VHC. De acuerdo a esta teoría no parece tan difícil como antes determinar porque se desarrolla con tanta incidencia CHC en la infección persistente por VHC. VIRUS DE LA HEPATITIS B (VHB) La diferencia en el curso evolutivo de la infección inicial por el VHB radica en la competencia de la respuesta inmune del huésped frente al virus (7). Se requiere una fuerte respuesta inmune para resolver la infección. Los linfocitos T citotóxicos específicos, primariamente dirigidos contra los antígenos del core, provocarán la lisis de los hepatocitos infectados dando lugar al daño hepático inmunomediado de la hepatitis aguda así como a la resolución del proceso infeccioso. Cuando esta respuesta inmune es insuficiente para resolver la infección, ya sea por inmadurez del sistema inmune como ocurre en neonatos, o por inmunodeficiencia de cualquier naturaleza, la infección persiste y se hace crónica (8). La mayoría de estos pacientes están asintomáticos, no obstante, algunos pacientes presentan a lo largo de la enfermedad reagudizaciones que pueden preceder a la desaparición del HBeAg y al desarrollo de anticuerpos contra el mismo, culminando en la remisión de la actividad de la hepatitis (seroconversión). Sin embargo en otras ocasiones, el resultado de estas reagudizaciones es únicamente la disminución de los niveles séricos del DNA viral acompañada de lisis hepatocitaria y mitosis. Paradójicamente podemos deducir que durante el curso de la infección crónica por el VHB, las mismas reagudizaciones que en unos pacientes pueden dar lugar a la seroconversión y a la remisión de la actividad de la hepatitis, en otros solo suponen ciclos continuos de inflamación y necrosis hepática que conducirán a la cirrosis y pueden dar lugar a la transformación maligna del hepatocito. En una pequeña proporción de estos pacientes, se desarrollará hepatocarcinoma sin cirrosis previa, y aunque este fenómeno está ampliamente reconocido entre los clínicos, las tasas en las que ocurre son bajas. Así las secuelas de la infección crónica por el virus B incluyen desde una leve a moderada fibrosis, cirrosis compensada, descompensación hepática, hasta el desarrollo de un hepatocarcinoma. Nos centraremos en la implicación del virus en el desarrollo del mismo. Patogénesis del Carcinoma Hepatocelular El VHB y el VHC son los dos principales agentes causantes del hepatocarcinoma, ya que ambos generan alteraciones en el genoma del huésped, siendo responsables del 80% de los casos, mientras que el otro 20% de los casos son atribuidos a otros factores de riesgo tales como abuso de alcohol, esteatosis, hemocromatosis y otros desórdenes metabólicos (3). De igual modo la mayoría de los tipos de cáncer, la hepatocarcinogénesis es un proceso en varios pasos de diferentes alteraciones genéticas que en última instancia darán lugar a transformación maligna del hepatocito. El mecanismo subyacente que lidera la transformación maligna de las células infectadas todavía no está claro. Mecanismos moleculares de la oncogénesis Se han documentado la presencia de distintas alteraciones cromosómicas en el hepatocarcinoma, algunas de las cuales están en relación directa con la infección por el VHB: • Se han detectado amplificaciones de los cromosomas 1q, 8q, 6p, y 17q • Entre los cromosomas más frecuentemente deleccionados en el hepatocarcinoma encontramos: 8p, 16q, 4q, 17p, y 13q. En los pobremente diferenciados 13q y 4q están significativamente representados. Estas regiones cromosómicas contienen elementos con un papel fundamental en el desarrollo de la hepatocarcinogénesis tales como el p53 (17q) o pRb (13q). De las distintas alteraciones moleculares implicadas en el desarrollo del hepatocarcinoma, la alteración del p53 tiene particular importancia por las múltiples funciones del mismo. El cual activándose en respuesta al estrés celular o al daño del ADN, intenta prevenir que el daño progrese, dando lugar a la reparación del ADN o a la entrada en apoptosis celular si la reparación no es posible. • Cambios epigenéticos, como las alteraciones de la metilación del ADN especialmente en las regiones promotoras de genes individuales, las cuales en contraste con las mutaciones somáticas, son capaces de regular la expresión génica sin cambios en la secuencia del ADN. La metilación de los promotores puede interferir con la unión de factores de transcripción y con otros mecanismos reguladores, lo que resulta en una disminución de la expresión del gen correspondiente. En el hepatocarcinoma observamos un desequilibrio en la metilación, dándose una hipometilación global de genoma acompañada de hipermetilaciones localizadas de los promotores de genes específicos, especialmente de genes oncosupresores. La importancia de estos cambios epigenéticos radica en que el patrón de metilación de algunos genes parece estar asociado con la infección del virus. Así la E-cadherina y GSTP1 están preferentemente metilados en el hepatocarcinoma asociado a VHB en comparación con el hepatocarcinoma asociado a VHC. Esto sugiere la participación de factores virales en el proceso de metilación. • Se han visto mecanismos de modulación epigenética específicos del VHB, así la proteína viral HBx, mediante la regulación de la actividad del promotor del gen de la DNA metiltransferasa (DNMTs), puede promover la hipermetilación de promotores de genes específicos. • En áreas tales como África Subsahariana y China, la exposición a la aflatoxina B1 es responsable de una incidencia muy alta de hepatocarcinoma. Y en estas áreas hay una alta proporción de p53 mutado en la tercera posición del codón 249, encontrándose una correlación positiva entre esta mutación y la exposición a aflatoxina. Esta mutación podría suponer una ventaja proliferativa para estas células, de tal manera que la presencia de la mutación en el p53 constituye un factor de pronóstico desfavorable respecto a las recaídas del hepatocarcinoma. • La vía de señalización Wnt/β-catenina también ha demostrado tener un papel en la regulación del desarrollo y diferenciación del tumor. El papel esencial en el hepatocarcinoma, es el aumento de la expresión y acumulación nuclear de βcateninas como característica de una activación de la señal de Wnt. Más del 62% de todos los hepatocarcinomas muestran esta disregulación de las β-cateninas, viéndose un peor pronóstico y una mayor tasa de recurrencias en pacientes con acumulación nuclear de β-cateninas. De manera que esta mutación oncogénica de las β-cateninas ha demostrado promover el desarrollo del hepatocarcinoma. Algunos artículos describen una alta asociación de la mutación con la exposición a Aflatoxina B1 (AFB1) e infección por VHC. • Otros importantes factores implicados son el TGF-β, proteínas Ras, pRb... Implicación directa del VHB en el hepatocarcinoma El hepatocarcinoma se establece en el seno de una hepatitis crónica, o de una cirrosis cuyos causantes principales son el VHB Y VHC. La inflamación es la forma básica de la patogenia de la hepatitis crónica que puede evolucionar a fibrosis nodular y progresar posteriormente a cirrosis y eventualmente a hepatocarcinoma. Globalmente, el VHB es la causa más frecuente de hepatocarcinoma. La incidencia anual de hepatocarcinoma en portadores de hepatitis B es del 0,5%, aumentando con la edad de manera que a los 70 años la incidencia es del 1%, siendo del 2,5% en pacientes con cirrosis conocida. Estudios de caso-control han demostrado que los portadores tienen un riesgo de 5-15 veces mayor de hepatocarcinoma que la población general. La gran mayoría de los hepatocarcinomas en portadores de VHB se han desarrollado en pacientes con cirrosis, pero el VHB también puede causar hepatocarcinoma en ausencia de cirrosis. El aumento de riesgo del hepatocarcinoma asociado a la infección por VHB, se aplica particularmente a áreas donde el VHB es endémico. En estas áreas la transmisión más usual es la vertical y el 90% de las infecciones siguen un curso crónico de la enfermedad. El VHB posee una estrategia de replicación que incluye la transcripción inversa de un RNA intermedio. En los pacientes con hepatitis crónica, secuencias de DNA del virus se ven integradas en el ADN celular del tejido tumoral y no en el no tumoral. Cuando el virus se integra, produce un amplio rango de cambios genéticos en el interior del genoma del huésped incluyendo delecciones, traslocaciones, la producción de transcriptos de fusión o recombinantes y una generalizada inestabilidad cromosómica. La integración del DNA viral se asocia con delecciones de segmentos cromosómicos muchos de los cuales contienen conocidos genes supresores tales como el p53, Rb, ciclina D1 y p16. Entre los diferentes genes de VHB, el HBx parece jugar el mayor papel causal en el hepatocarcinoma, ya que es el gen viral más comúnmente integrado. Entre los efectos patobiológicos del HBx está la coactivación transcripcional de los genes celulares y virales (modulación de la RNA polimerasa I y III), interaccionando con el sistema de reparación celular del DNA y generando una disregulación de los puntos de control del ciclo celular (interacciona con la función de p53. En conclusión, el hepatocarcinoma es la consecuencia del acúmulo de mutaciones somáticas en el genoma de las células hepáticas infectadas por el virus por una combinación de dos mecanismos: • Mediante la lisis celular y el estímulo de la mitosis que se dan continuamente en los ciclos de necrosis hepatocitaria y regeneración, liderando una acumulación de mutaciones necesarias para la transformación maligna. • Por el aumento de inestabilidad cromosómica mediante la inducción de proteínas recombinantes durante la hepatitis crónica. La carga viral de VHB es un importante determinante del riesgo de hepatocarcinoma, y las características genotípicas del virus aparecen para ejercer un efecto adicional en la carcinogénesis. La mayoría de los estudios del este y sudeste de Asia han demostrado que pacientes infectados con el genotipo C del VHB tienen un mayor riesgo de desarrollar hepatocarcinoma. De manera que el fenotipo C y la alta carga viral son predictores independientes de hepatocarcinoma. Los estudios sugieren que las variantes de los promotores del core y precore están asociadas con una enfermedad más severa del hígado y hepatocarcinoma. La prevalencia de la mutación del promotor del core es mayor entre pacientes crónicos de hepatitis B quienes desarrollan complicaciones de cirrosis y hepatocarcinoma. Controlando otros factores de confusión tales como la edad, fenotipo y la presencia de cirrosis, se ha visto que las variantes del promotor del core A1762T y G1764A están significativamente asociadas con el desarrollo del hepatocarcinoma. VIRUS DE EPSTEIN-BARR (VEB) (9) El virus Epstein-Barr (VEB) pertenece a la familia de los herpesvirus que infectan a gran parte de la población mundial. El virus establece una infección latente en los linfocitos tipo B y puede inducir la proliferación de los mismos. Aunque la mayoría de las infecciones por VEB son benignas, su potencial oncogénico está presente. El grupo IARC, que investiga la evaluación de riesgos cancerígenos para el ser humano, llegó a la conclusión de que existen pruebas suficientes que implican al VEB en la causalidad de múltiples enfermedades como linfoma de Burkitt, linfoma de Hodgkin, el carcinoma nasofaríngeo y la inmunosupresión relacionada con el linfoma. Epidemiología La epidemiología de la infección por VEB refleja la importancia de factores como la edad de exposición al virus. En general, la mayoría de las infecciones se producen en la infancia. Entre los factores que influyen, incluimos, contacto interinfantil (chupete), en el hogar, en el barrio… el nivel de higiene; la distribución de alimentos, y tal vez el contacto oral en adultos. Para las personas que escapan a la infección en la infancia, la adolescencia se asocia a riesgo de padecer infección (contacto oral) con una alta probabilidad de MI. Mecanismos moleculares de la oncogénesis El VEB ha desarrollado una serie de mecanismos oncogénicos intrínsecos que aseguran su supervivencia (Young y Murray, 2003; Dolcetti Masucci, 2003). Estos mecanismos incluyen su capacidad para transformar células B en líneas celulares linfoides en continuo crecimiento. La proteína LMP-1, que se expresa en el LH, y en el CNF, es en sí misma un oncogén. LMP-1 juega un papel central en la transformación y en la regulación de varios factores celulares. Lo que es realmente sorprendente es que la gran mayoría de la población es portadora de este virus, en gran parte de su vida, sin efectos perjudiciales, gracias a la protección de linfocitos T citotóxicos (LTC) específicos en constante vigilancia. Esta protección se ve más claramente cuando se somete a los portadores normales a inmunosupresión terapéutica para el trasplante de órganos. Una pequeña proporción, puede desarrollar una linfoproliferación espontánea, que puede evolucionar a un linfoma de células B VEB positivo. La infusión de LTC específicos para VEB puede invertir este proceso maligno (Rooney el al., 1995). Linfoma de Burkitt Linfoma de Burkitt (LB) es una neoplasia maligna que se produce endémicamente entre los niños pequeños, especialmente en varones, en el centro de África y Nueva Guinea y, esporádicamente, a cualquier edad en otras partes del mundo. Las zonas endémicas se correlacionan fuertemente con la epidemia de malaria. Se cree que el riesgo de LB es el aumento de la proliferación de los linfocitos B a comienzos de la infección por VEB, interactuando en el curso del efecto mitogénico de la malaria. En estas zonas, los niños se ven principalmente afectados, con una incidencia máxima en torno a los ocho años de edad. La mandíbula y los órganos abdominales son los más frecuentemente afectados. Estos tumores son poco frecuentes en el resto del mundo. Aproximadamente uno de cada tres de estos carcinomas LB no endémicos es VEB positivo, en comparación con casi el 100% de los endémicos. Como apunte podemos destacar que LB se produce como una neoplasia oportunista en la infección por el VIH, más comúnmente en niños. Independientemente de su origen geográfico, todos los LB muestran una de las siguientes translocaciones cromosómicas recíprocas. Todas ellas se relacionan con el brazo largo del cromosoma 8q: • cromosoma 14q en cualquiera de sus partes (75% de la casos) • brazo corto del cromosoma 2p (9%) • brazo largo del cromosoma 22q (16%) La parte del cromosoma 8 traslocada, incluye el oncogén c-myc, que, cuando se trasloca junto a genes de la cadena pesada del cromosoma 14q, o junto a los genes de la cadena ligera K del cromosoma 2p, o junto a genes de la cadena ligera de 22q, se activa. Este paso (es decir, la activación) se produce durante la generación de anticuerpos frente al VEB. La alteración de la actividad de c-myc, parece ser esencial en cuanto al desarrollo de células malignas cuya replicación conduce a LB. La asociación de VEB con LB se demuestra por la presencia del episoma del VEB en los tumores, que sólo expresan EBNA-1. En lo que a anticuerpos nos referimos, existe una importante elevación de IgG frente a VCA, EBNA, EA-R (en menor medida). El estudio epidemiológico más importante que relaciona la infección por VEB con el LB, es un estudio prospectivo basado en una cohorte de 42.000 los niños, de menos de 9 años, en el Nilo Occidental de Uganda, bajo los auspicios de la IARC (de Thé et al. en 1978). Carcinoma nasofaríngeo El carcinoma nasofaríngeo (CNF) se produce en todo el mundo. Estos tumores son clasificados histológicamente como carcinomas indiferenciados o pobremente diferenciados. La incidencia más alta está en el Extremo Oriente, principalmente en personas de origen chino y en poblaciones relacionadas que emigraron a diferentes lugares. Los datos difieren entre varias poblaciones chinas, dentro de la misma zona y son más altas entre los del Sur. El rango de edad de mayor incidencia entre los chinos de Hong Kong y Singapur se sitúa entre 20-50 años de edad, y posteriormente decae. Este tipo de curva de edad sugiere una etiología infecciosa, con exposición temprana. En Suecia en la década de 1980, por el contrario, el aumento se produjo cerca de dos décadas más tarde y continuó hasta el final de la vida. En los Estados Unidos, se ha señalado una distribución trimodal, que sugiere diferentes etiologías que operan en los diferentes grupos de edad en una población racialmente mixta. Poco se sabe acerca del proceso por el que el VEB se convierte en agente causal del CNF. Existe gran cantidad de estudios que tratan de dar una explicación a este fenómeno. Según Krishna SM et cols. (10), el fracaso del tratamiento en el carcinoma nasofaríngeo (CNF) se debe principalmente a la recurrencia local y regional. En la actualidad se carece de un marcador para identificar estos casos. VEGF es un potente mitógeno de células endoteliales y un importante agente inductor de la permeabilidad vascular. Desempeña un papel fundamental en el crecimiento tumoral y en la angiogénesis. Otros estudios orientales (11), comparan la expresión de etk/bmx en el carcinoma nasofaríngeo (CNF) y en lesiones nasofaríngeas benignas, con el fin de saber si la falta de expresión del receptor tirosina quinasa asociado con el desarrollo del CNF. El Departamento de Otorrinolaringología del Hospital de Shangay (12), asegura que VEB guarda estrecha relación con el carcinoma nasofaríngeo. La latencia de VEB durante la carcinogénesis del CNF, sigue siendo desconocida hoy en día. Este estudio trata de analizar la correlación existente. Varios informes (13) han demostrado que el gen barf-1 codificado por VEB tiene actividad oncogénica. Recientemente hemos informado de que barf-1 se expresa como un gen latente en la mayoría de los carcinomas nasofaríngeo, lo que sugiere que barf-1 puede tener un papel importante en la oncogénesis CNF. Del mismo modo en que en el caso de LB, los títulos elevados de anticuerpos IgG-VCA marcan una asociación con VEB, en el CNF, ocurre lo mismo. Dichos títulos se encuentran en más del 80% de los pacientes CNF. El hecho que certifica la presencia de VEB en los pacientes con CNF, es una vez más la biopsia tisular, donde se evidencia este tipo de virus en los tejidos extirpados. Para ello se utilizan varias técnicas moleculares. Ocurre mayormente en células epiteliales y no en linfocitos. El genoma viral es clonal y expresa sólo EBNA-1, LMP2 (su presencia predice recurrencia tras tratamiento), y LMP-1 de forma inconstante. Pathmanathan et al. (1995), representa uno de los principales estudios, que proporcionaron pruebas sólidas de que el VEB desempeña una función causal directa en el desarrollo del CNF. Un estudio serológico prospectivo realizado en Taiwán, valoró el papel de los anticuerpos VEB como predictivo sobre el desarrollo de CNF. Los objetivos fueron los hombres que residen en zonas con las tasas más altas de tumor. Otros autores confirman la importancia de la susceptibilidad genética y las exposiciones ambientales, en especial factores dietéticos (IARC, 1997). Linfoma de Hodgkin El linfoma de Hodgkin (LH) es una neoplasia relativamente común en adultosjóvenes occidentalizados, donde la curva de incidencia por edad es en general bimodal. Desde que VEB se identificó por primera vez, fue considerado un candidato para intervenir en la causalidad de este linfoma. La hipótesis de que la edad de contracción de la infección viral puede desempeñar un papel importante en el desarrollo de la LH, fue sugerida por las similitudes epidemiológicas entre LH en los adultos jóvenes y MI en poblaciones económicamente favorecidas. Para ambas enfermedades, el riesgo se ha asociado con la clase social más alta y con el tamaño pequeño de las familias. Estas similitudes sugieren que la exposición al agente causal se retrasa hasta la vida adulta-joven. Se encontró que las personas con antecedentes de MI tienen triplicado el riesgo de desarrollar LH. Además, una gran proporción de casos de LH tienen elevados los títulos de anticuerpos para varios antígenos del VEB: IgG-VCA y EA-D (en comparación con los controles). En general, del 30% al 40% de los casos de LH, elevan sus títulos. Un estudio piloto basado en dos casos de LH entre 25.802 personas utilizando muestras de suero, demostró que, en comparación con los casos controles, los anticuerpos frente a VEB se elevan de forma significativa antes del diagnóstico. Esto contrasta con otros tres herpesviruses cuyos títulos de anticuerpos no son elevados (Evans y Comstock, 1981). Este estudio se extendió para incluir el suero durante más de 240.000 personas, provenientes de cinco bancos de suero diferentes. En esta nuevo estudio ampliado, 43 personas que desarrollaron LH, 1 y 13 años después del inicio del estudio (Mueller et al., 1989). Los análisis de anticuerpos frente a VEB en dichos sueros confirmaron las conclusiones del proyecto piloto en que la proporción de pacientes con títulos de anticuerpos elevados, fue significativamente superior a que de los controles. Esto se asocia con un riesgo de 3 a 7 veces mayor de desarrollar LH. Más de la mitad (56%) de los sueros en los casos pre-LH había elevado títulos de uno o más anticuerpos frente a VEB, en comparación con el 35% de los casos controles. Un avance importante para vincular el VEB con LH se logró gracias a la hibridación molecular (Weiss et al., 1987). Los estudios demuestran que entre el 25% y 50% de los casos de LH, son VEB positivos (presencia del genoma viral o de productos virales). Por último, la relación con la historia de MI fue validado por Hjalgrim et al. (2003). En el seguimiento de 38.000 escandinavos con MI, los autores fueron capaces de reconocer la presencia del VEB en 29 biopsias en un total de 40 casos diagnosticados de LH, 2 o más años después del diagnóstico de MI. El RR de LH VEB positivos, en MI previamente diagnosticada fue de 2,8, y de 1,1 para VEB negativo. Estudios de la Sociedad Americana de Hematología, afirman que el linfoma de Hodgkin clásico se caracteriza por la presencia de células Reed-Sternberg (RS), que son transformadas y destinadas a la apoptosis, ya que no se hayan sometido con éxito a la reorganización de genes de inmunoglobulina. Varios mecanismos, incluida la infección latente por VEB, permiten que estas células puedan sobrevivir. Pese a los estudios realizados, en esta ocasión, del mismo modo que ocurre en el CNF, podemos afirmar que la relación biogénica tampoco es bien conocida. VIRUS DEL HERPES HUMANO 8 (VHH-8) En 1994 Chang y Moore publicaron el descubrimiento del último gammaherpesvirus patógeno humano, el VHH-8. Hoy, más de diez años y 2000 artículos después, se perfila como el último virus oncogénico descubierto y se estudia un espectro de enfermedades proliferativas en las que podría estar implicado. Se ha establecido claramente su asociación con el sarcoma de Kaposi (SK), la enfermedad de Castleman multicéntrica (ECM) y el linfoma de derrame primario (LDP). Sin embargo muchos aspectos sobre la infección y su oncogénesis se desconocen aún. Virología (14) El virus de herpes humano 8, también conocido como herpesvirus asociado a sarcoma de Kaposi (SKHV) es un gamma-2 virus o Rhadinovirus. Junto con HHV-4 o VEB, pertenece a la subfamilia Gammaherpesvirinae de la familia Herpesvirinae. Es un virus ADN y tiene una morfología típica de herpesvirus con una bicapa lipídica que envuelve a una cápside icosaédrica de unos 100-120nm compuesta por cuatro proteínas que nos permiten clasificar al virus en seis subtipos (A, B, C, D, E y N) con agrupación geográfica. Genoma de VHH-8 El genoma de VHH-8 es bicatenario de unas 170-270 kb. Consta de unos 26 genes altamente conservados entre los herpesvirus, que incluyen aquellos relacionados con: regulación genética, metabolismo nucleótido, replicación ADN, estructura del virión y transición entre ciclo lítico y latente. Según la terminología adoptada por Russo (15) se nombran con una K delante aquellos genes que no tienen homología con ningún otro herpesvirus. VHH-8 en particular tiene al menos doce genes homólogos a genes humanos que deben haber sido adquiridos de células huésped dada su escasez de intrones. Como veremos juegan un papel fundamental en su interacción con el hospedador. Patogenia de la infección En el huésped inmunocompetente su infección es benigna, por lo que VHH-8 sólo será patogénico cuando intervengan otros factores como inmunodepresión o coinfección por VIH. Infecta principalmente a linfocitos, monocitos y queratinocitos, sus reservorios naturales son las células B (CD19), aunque se ha informado de infectividad en células endoteliales, monocitos, epitelio glandular prostático, ganglios sensitivos dorsales y células fusiformes de SK. Éstas expresan CD34 (célula madre hematopoyética y marcador vascular endotelial) y marcadores linfáticos como D2-40, LYVE-1 y VEGFR-3, por lo que se discute sobre su origen. Algunas de las glicoproteínas virales de superficie se unen a receptores celulares para heparán sulfato o de integrina α3β1 y se produce la internalización del virus por endocitosis. El ciclo vital del virus se compone de dos fases: latente y lítica. En más del 90% de las células infectadas el virus entra en fase latente, su genoma se queda integrado en el núcleo celular en forma de episomas y se replica con los cromosomas expresando sólo algunos de sus genes como: LANA-1, v-ciclina, vFLIP o Kaposina que serán necesarios para mantener su forma latente y perpetuar así la infección. Se ha demostrado que los precursores endoteliales en sangre periférica actúan como reservorio para el virus. (16) Bajo los estímulos adecuados el virus puede pasar a fase lítica. Se produce la transcripción de genes y la síntesis activa de viriones además de otras proteínas necesarias para su interacción con el huésped como K1, vGCR, vIRF, vIL-6, vMIP. Como veremos, esto desempeña un papel fundamental en la oncogénesis (de forma más importante en ECM y SK y menor en LDP). Epidemiología. Transmisión Epidemiología Su distribución geográfica es limitada y paralela a la de SK. Distinguimos países con baja endemicidad como EEUU, Reino Unido, la mayoría de Europa del norte y Asia, donde la seroprevalencia oscila entre 2% a 9% en EEUU y 3% a 5% en Europa del Norte y Sudeste Asiático. Poblaciones de endemicidad intermedia en zonas mediterráneas de Italia y Grecia, con prevalencia de 14% a 30%. Por último hay alta endemicidad en algunos países de África central y del este (70%–90%) (17), donde SK es endémico, aunque es más baja en el sur y norte de África (10%–40%) (18). La seroprevalencia aumenta con la edad (en zonas endémicas se eleva desde la infancia, mientras que en áreas no endémicas no lo hace hasta después de la adolescencia), origen mediterráneo o de Europa del Este, infección por VIH, homosexualidad y otros comportamientos sexuales de alto riesgo (19). Transmisión (20) Los patrones de transmisión del virus se van perfilando al ir comprendiendo la patogenia y mejorando los test de detección. Hasta la fecha se sigue discutiendo acerca de la participación de ciertas vías. • Transmisión sexual Es mayor varones homosexuales activos (38% vs 0% en heterosexuales). Son factores de riesgo la presencia de enfermedades de transmisión sexual y el número de parejas masculinas. Ya que el virus se aísla con más frecuencia en saliva, las prácticas que incluyan sexo oral podrían suponer un factor de riesgo. La transmisión en secreciones sexuales masculinas es improbable ya que la prevalencia en semen es muy baja en seropositivos y seronegativos para VIH. En mujeres con SK de Zimbabwe se encontró ADN en secreciones cervicales de entre el 28% y 37% mientras que no se aisló en mujeres sin SK. • Transmisión sanguínea Sigue siendo objeto de debate la transmisión por vía sanguínea. Ésta se encontraría limitada por la naturaleza intracelular del virus, la intermitencia de su viremia y la baja tasa de virus circulante en asintomáticos seropositivos. Hladik et al confirmaron la transmisión por esta vía, con un riesgo de seroconversión máximo de 2.8% en 3-10 semanas. Afirman que la transmisión disminuye si se almacenan las muestras más de 4 días (riesgo acumulado 4.2%). Aseguran que los estudios anteriores que no apoyaban la asociación usaban muestras pequeñas o que incluían transfusiones acelulares (21). En España, la prevalencia en donantes de sangre se sitúa en 9.7% para VIH+ y en 13.1% en VIH-. Entre los varones homosexuales VIH+, 83.3% sin SK y 100% con SK fueron reactivos para antígenos latentes (22). • Transmisión familiar Se considera que la seropositividad conyugal o maternal aumenta el riesgo de transmisión. La transmisión vertical parece improbable bien en la gestación o en la lactancia y se limitaría a madres inmunocomprometidas. Sí se ha constatado la transmisión horizontal madre-hijo, por lo que se postula que la saliva es el medio de transmisión más trascendente entre los miembros de una familia. • Transplantes hematológicos y de órganos sólidos El estudio de la transmisión de VHH-8 en trasplantes está limitado ya que existen pocos estudios de seroprevalencia en donantes y receptores. Además, una vez constatada existen pocos procedimientos para monitorizar de forma diagnóstica o clínica a los pacientes. Cabe destacar que la seroconversión postransplante además de por transmisión del donante, puede deberse a reactivación de virus latente dada la inmunosupresión. La aparición de SK, a su vez, puede deberse a la transfusión de células infectadas en sangre o de reservorios tumorales desconocidos en órganos transplantados. La mayoría de estudios existente se han realizado en transplantes de riñón. La seroprevalencia en el primer año postransplante aumenta de 6.4% a 17.7% y es mayor que en los pocos estudios disponibles sobre transplante hepático. En un estudio en 2002 en el Hospital Gregorio Marañón de Madrid sobre la aparición de SK en pacientes seropositivos para herpesvirus transplantados con órganos sólidos la incidencia fue de 1:200 con mayor afectación de varones (6:1) (23). Se han encontrado afectaciones no neoplásicas en transplantados hematológicos como síndrome febril, aplasia de médula ósea o plasmocitosis que podrían asociarse a VHH-8. Enfermedades asociadas Primoinfección No se dispone de mucha información al respecto dada la falta de estudios de seroprevalencia y el desconocimiento de los síntomas que pudieran identificarla. Se ha visto que en pacientes inmunodeprimidos puede ser letal mientras que en pacientes sanos apenas muestra unos pocos síntomas inespecíficos como linfadenopatía, diarrea, rash o astenia. Sarcoma de Kaposi, SK El sarcoma de Kaposi es un tumor altamente vascularizado de estirpe policlonal, oligoclonal o monoclonal cuyo origen parece situarse en células endoteliales, linfoendoteliales o monocíticas. Afecta principalmente a la piel con posible extensión linfática y visceral. Las lesiones están constituidas por las características células fusiformes (Spindle Cells, SC). Se han descrito cuatro variedades de sarcoma de Kaposi: clásico o esporádico, asociado a SIDA, endémico y iatrogénico. Tienen características epidemiológicas y clínicas diferentes como la afectación anatómica o la agresividad pero en todos se ha descrito VHH-8 como agente etiológico necesario. Se ha demostrado que la coinfección con VIH es fundamental para el desarrollo de este tumor, por lo que su incidencia ha aumentado en las últimas décadas, aunque la variante asociada a SIDA ha sufrido una disminución debido al uso de Terapia Antiviral de Gran Actividad (TARGA), mientras que la variante iatrogénica ha aumentado en relación a los tratamientos inmunosupresores y los transplantes. Se considera una enfermedad definitoria de SIDA. Linfoma de Derrame Primario, LDP Es un raro subtipo de linfomas primarios de cavidades. Sus características son: carecen de morfología de linfoma de Burkitt, no presentan alteraciones en c-myc, carecen de reordenamientos en bcl-2 o p53, tienen una morfología distintiva de linfoma inmunoblástico y linfoma anaplásico, afectan principalmente a hombres, permanecen en la cavidad de origen, expresan CD45, frecuentemente faltan los antígenos asociados a las células B, tiene reordenamientos clonales en el gen de la inmunoglobulina y pueden albergar VEB. Suponen el 0.13% de las complicaciones del SIDA y son más frecuentes en pacientes que ya han padecido SK (24) (25). Enfermedad de Castleman Multicéntrica, ECM La enfermedad de Castleman es un raro desorden linfoproliferativo policlonal de células B del que se conoce poco. Su variante multicéntrica afecta a las células plasmáticas y tiene una presentación clínica y morfológica muy variable. El 90% de los pacientes con ECM en el contexto de SIDA son seropositivos para VHH-8, mientras que en pacientes no VIH representan el 40%. Es un dato indicativo que la mayoría de células patológicas no están infectadas por VHH-8, sino que sólo se limitan al manto folicular, lo cual nos hace pensar que existe un mecanismo de reclutamiento paracrino (vIL6) (26) (27). Otras asociaciones • Sarcoidosis: diversos estudios han sugerido una mayor prevalencia de SKHV en pacientes con sarcoidosis, aunque no se ha conseguido demostrar que sea mayor que la de la población de la zona geográfica (28). • Mieloma múltiple, MM: la etipatogenia del MM sigue siendo incierta. Se ha visto que influyen citocinas como IL-6, TNF e IL-1B y oncogenes como ras, c-myc y p53. Algunos autores han propuesto a VHH-8 como agente disregulador del sistema inmune en esta patología. Algunas series mostraron una mayor prevalencia de anticuerpos de fase lítica en estos pacientes, sin embargo otros estudios no fueron capaces de demostrar esta asociación al encontrar resultados contradictorios (29) (30) (31). • También se ha relacionado con aplasia medular, insuficiencia cardiaca o pénfigo vulgar sin encontrar resultados concluyentes. Oncogénesis VHH-8 es el último virus oncogénico humano descubierto, por lo que su patogenia no está aún clara (32). El proceso de malignización comprende varios mecanismos dependientes tanto del virus como de la célula. Podemos englobarlos en tres pilares fundamentales: control del ciclo celular-apoptosis, escape del sistema inmunológico y regulación del ciclo vital del virus. En muchos casos los procesos iniciados por una única proteína vírica alteran los tres de forma directa o indirecta, por lo que la descripción individualizada de cada proceso se hace compleja. Se ha otorgado un papel fundamental a algunos de sus genes (v-bcl-2, vIL-8R, vMIPs, vIL-6 y v-ciclina D) que son homólogos virales de los humanos y servirían de nexo entre el virus y la maquinaria celular, modificando la respuesta del huésped a su favor, como se describe a continuación. La forma de actuar se supone similar para SK, ECM y LDP, aunque se está trabajando para caracterizar cada una. En el caso de SK, y tomándolo como referencia, la malignización comienza como una respuesta angioproliferativa policlonal frente a VHH-8 producida por las citocinas proinflamatorias (IL-1, IL-6, TNF-α, IFN-γ b-FGF y VEGF) que producen activación de las células endoteliales y angiogénesis. Con el tiempo, alguna de las poblaciones se desmarcaría en un crecimiento oligoclonal que daría origen al tumor. Se ha visto que la infección por KSHV es necesaria pero no suficiente para el desarrollo de este tumor, por lo que se piensa que intervienen otros factores como el estado de inmunosupresión del huésped acompañado de una disregulación generalizada de la producción de citocinas, factores de crecimiento y moléculas de adhesión o la participación de la proteína Tat-VIH en caso de infección (33). La interacción del virus con la maquinaria celular es uno de los objetivos principales de la investigación en la actualidad y constantemente se descubren nuevos factores, mediadores y vías celulares etc. que podrían intervenir en la malignización de la infección. Su entendimiento es crucial para caracterizar por completo los procesos biológicos del virus, su relación con el huésped y el desarrollo de la enfermedad maligna. A continuación se describen los principales mecanismos que se han demostrado. • Antígeno Nuclear Asociado a Latencia tipo 1, LANA-1: es una proteína muy inmunogénica de gran importancia para mantener el virus en la forma quiescente y que juega un papel central en la malignización. Se expresa ampliamente en el ciclo latente pero también en el lítico, por lo que se usa como marcador de la enfermedad en lesiones así como para la serología. Son múltiples las funciones que se le atribuyen en el origen, mantenimiento y progresión de la enfermedad: ‐ Regula el ciclo celular al competir con E2F para unirse a la pRb, liberando a E2F para activar genes involucrados en progresión del ciclo. ‐ Disminuye la transcripción de p53 para escapar de la apoptosis. ‐ Bloquea diferenciación celular estabilizando la β-catenina por la vía Wnt y al secuestrar Glucógeno Sintasa Kinasa 3β (GSK-3β). A su vez, estimula la ciclina D1 (34) (35). ‐ Evita la salida del ciclo celular en G1 promovido por p16, BRD-4 y BRD-6 (34). ‐ En células de LDP, se produce la inactivación nuclear de la GSK-3, lo que reduce la fosforilación de c-myc y contribuye a su estabilización al disminuir su ubiquitinación. La β-catenina también activa c-myc, por lo que podría contribuir en este sentido (34). ‐ Incrementa el nivel de Kinasa Extracelular Regulada por señal (ERK-1) fosforilada y aumenta la fosforilación de c-myc por parte de ERK, sin intervención de la vía GSK-3, lo cual evita la apoptosis iniciada por c-myc (34). ‐ Hace de unión entre el genoma episomal del virus y el de la célula huésped durante la replicación. Además se une a los Complejos de Reconocimiento de Origen (CRO) que interaccionan en la región de las Repeticiones Terminales (RT) del genoma vírico, lo cual silenciaría la transcripción del gen de la fase lítica K1, situado en su vecindad. A su vez, impide la expresión de estos genes de la fase lítica al unirse a la proteína RBP-Jκ, que inactiva a RTA (36). ‐ Se une a la heterocromatina metafásica produciendo una redistribución de los centriolos a la periferia nuclear, lo que indica cambios extensos en el posicionamiento de los dominios cromosómicos (36). ‐ Activa el promotor de la transcriptasa inversa de la telomerasa (36). ‐ Puede producir la represión genética al inhibir la transcripción del factor ATF4/CREB-2 o al unirse a dos proteínas asociadas a los cromosomas como MeCP-2 y DEK (36). ‐ Al interactuar con la vía ubiquitina-proteasoma mediada por la ligasa E3 Sel-10, disregula el protooncogén Notch (vía ICN), relacionado con la activación de otros genes encargados de la regulación del ciclo, diferenciación y proliferación, lo que aumenta la replicación celular (36). • La interleucina 6 (IL-6) es un factor de crecimiento para células B del que VHH-8 es capaz de sintetizar un homólogo conocido como vIL-6 (K2). Es de gran importancia en la patogenia de ECM y LDP (1000 copias) y menor en SK (10-100 copias). A diferencia del humano, solo necesita la unión de gp130 y no de gp80. Favorece la angiogénesis por inducción de VEGF y a su vez estimula la producción de IL-6 celular. Zhang et al bloquearon la expresión de vIL-6 en células de LDP usando un antipéptido (PPMO) y se observó una disminución de los niveles de crecimiento celular, expresión de IL-6, ADN viral, RTA y LANA, así como un aumento de proteína p21, sin que se demostrara la relación causal (37). • VHH-8 induce una reprogramación en las células endoteliales para que expresen marcadores linfáticos como VEGFR-3 o podoplanina, mediada por la activación del promotor Prox1. La activación mediante gp130 de las vías de señalización JAK2/STAT-3 y PIK-3/Akt es necesaria para la activación de esta vía (38). • Disminuye el número de unidades del Complejo Mayor de Histocompatibilidad de clase I (CMH-I) en la superficie celular mediante dos proteínas transmembrana: MIR1 y MIR-2 (K3 y K5) que promueven su endocitosis y degradación lisosomal. Además MIR-2 disminuye la actividad de ICAM-1 y B7.2, proteínas accesorias para la estimulación T (20). • MIR-2 también interactúa con la cadherina del endotelio vascular promoviendo su ubiquitinación y eliminando así el mayor constituyente de las uniones intercelulares. Además reduce los niveles de α, β y γ catenina alterando así el citoesqueleto. Como resultado se modifica la permeabilidad celular y esto haría que las células respondieran con más intensidad a los factores de crecimiento y facilitaría la proliferación desorganizada de los vasos (35). • MIR-2, a su vez, mediante su actividad ligasa de ubiquitina E3 reduce la expresión en la superficie celular de los ligandos de MHC I relacionados con la cadena A (MICA), B (MICB) y el recién descrito ligando para NKp80, lecitina C inducida por activación (AICL o CLEC-2B), que son ligandos para los receptores de NK NKG2D y NKp80. Así se disminuye la sensibilidad frente a NK. Se sabe que en la primoinfección se activa el promotor de K5, lo que favorecería la infectividad en las primeras fases. MIR-2 además es capaz de unirse a CMH-I, CD86, CD1d, CD31PECAM-1, ALCAM, IFN-R, AICL… evitando así el reconocimiento por células de la inmunidad específica (CD8, CD4 y células T restringidas por CD1d) e innata (vías NKG-2D y NKp80 y receptor IFN) (39). • Entre los mecanismos que el virus utiliza para escapar a la respuesta inmune también se encuentran la reducción del número de epítopos presentados, la similitud de los mismos con péptidos endógenos y mutaciones en epítopos específicos que simulan una respuesta inmune adecuada en el huésped. Como muchos otros mecanismos manipulados por el virus, su forma de actuación es distinta en la fase latente y en la lítica. En este caso, el número de epítopos presentados así como la similitud de los mismos con péptidos endógenos es menor durante la fase quiescente (40). • El virus recurre también a la inhibición de la apoptosis inducida por NK (que actúan por la vía fas-CD95/Apo-1) expresando la proteína inhibitoria de vFLICe (vFLIP, K13) que impide la activación de procaspasa-8. Además vFLIP puede actuar en NK-kappaB y AP-1, que son esenciales para el crecimiento y supervivencia de la célula (41). • VHH-8 es capaz de producir una polarización de la respuesta inmune hacia la vertiente humoral reclutando linfocitos Th2 que producen IL-4 e IL-5, en detrimento de los linfocitos Th1 que producen IFN-γ. El gen K14 codifica una proteína de adhesión (OX-2) que también promueve la polarización Th2 y la producción de citocinas inflamatorias como IL-6 (20). • K1 es una glicoproteína transmembrana codificada por ORF1 de estructura similar al receptor de células B (RCB) que puede interactuar con múltiples ligandos que tengan dominios SH2, como lyn, syk, p85, PLCγ-2, RasGap, vav y grb-2. Su cola citoplasmática contiene una secuencia ITAM que dispara una vía dependiente de Syk kinasa y PI3K/Akt. Su activación en células B y células endoteliales las protege de la apoptosis. Si esta activación ocurre mediante ligandos superficiales a veces lleva a una internalización del receptor mediada por clatrina que se considera un mecanismo importante por el que la célula controla la intensidad y duración de la señal, pudiendo ser usado como amplificador (42). • La progresión tumoral se ve favorecida por citocinas y factores de crecimiento producidos por la infección VIH y la alteración del sistema inmune del huésped que conlleva. Además parece haber sinergismo entra ambas infecciones. Se ha visto que la replicación de VIH estimula la producción de VHH-8 en líneas celulares de LDP y en mononucleares de sangre periférica de pacientes con SK. Ha cobrado especial importancia la proteína VIH-Tat. Se ha demostrado que altera la producción de IL-6 celular y de su receptor, así como de la vías STAT-3, IL-4/STAT-6 o JAK/STAT lo que aumentaría la replicación y la carga viral (33). La activación del ciclo lítico de VHH-8 puede asimismo inducir un aumento en los niveles de replicación de VIH, lo cual aumenta la susceptibilidad celular para su infección, así como una permisividad temporal para la replicación del virus. • VHH-8 produce en la fase lítica tres citocinas, vCCl-1, vCCL-2 y vCCL-3 (vMIP-II, vMIP-II y vMIP-III) codificadas por los ORFs K6, K4 y K4.1, que están involucradas en la evasión inmune además de ser responsables de inducción angiogénica de forma autocrina y paracrina (43). ‐ Activan la respuesta Th2 mediante los receptores CCR-8, CCR-3 y CCR-4 y además son antagonistas de los receptores de la quimiotaxis de linfocitos Th1 y células NK (20). ‐ vCCL-2 se puede unir a muchos receptores de citocinas, incluyendo CXCR-4, CX3C y XCR-1, aunque sólo CCR-3 y CCR-8 pueden soportar la transducción de la señal de vCCL-2. Las señales de vCCl-1 vía CCR-8 y de vCCL-2 son agonistas CCR-4 y XCR-1. Las células endoteliales y las de SK expresan receptores CCR-8. Recientemente se ha demostrado el uso de esta vía para la supervivencia de células endoteliales y aumento de la replicación del virus. ‐ Las proteínas vCCL actúan de forma autocrina y paracrina suprimiendo la expresión de una proteína de la familia bcl-2, bim, que promueve la apoptosis inducida por estrés, lo que le permite escapar de la muerte celular que la activación del ciclo lítico induciría y así aumentar la producción viral. ‐ vCCL-2 y vCCL-3 son respectivamente los antagonistas y agonistas más potentes de XCR-1, linfotactina (44). • La producción autocrina por parte del virus de vGPCR (Receptor acoplado a Proteína G), un homólogo del receptor para IL-8, estimula la producción de factores celulares como VEGF, bFGF, IL-1β, IL-2, IL-4, IL-8 e IL-6, TNF-α, MIP-1, VCAM-1, ICAM-1 y selectina-E con actividad proangiogénica o proinflamatoria. A su vez, activa los factores de transcripción AP-1, NF-AT y NKκB y las vías Pyk-2/Lyn, ERK, p38, JNK y PI3K/Akt (43). Este último que parece ser el eje central de la sarcomatogénesis por este mecanismo: se activa el protooncogén TSC que fosforila la proteína Reb, relacionada con Ras y activa mTOR, que pone en marcha la maquinaria celular mediante la kinasa ribosómica p70 s6 (p70 S6K) y la proteína 1 de unión al factor eucariota de iniciación (4EBP-1) (45). La proteína K7 promueve la degradación de p53 y IκB al antagonizar a PLIC-1, un factor inhibidor de la degradación proteasómica. Además disregula la apoptosis al actuar en bcl-2 y en la ciclofilina moduladora de calcio del retículo endoplasmático. Se ha demostrado en ratones que interactúa con vGPCR induciendo su degradación proteasómica, lo cual reduciría su potencial oncogénico. Sin embargo, la expresión conjunta de ambos potencia la actividad antiapoptótica (46). • Las catepsinas parecen estar involucradas en la carcinogénesis de SK. A diferencia de otros tumores, su secreción están inhibida por la expresión del receptor de factor de crecimiento insulina-like/receptor manosa-6-fosfato (IGF-IIR/M6PR). Se ha visto que su rol no es evitar la apoptosis, por lo que puede estar relacionado con interferir en las cascadas de señalización intracelular, como VEGF, o regular la actividad de las metaloproteasas de la matriz (MMP) o de otros procesos proteolíticos (47). • El homólogo viral v-ciclina se une a CDK-6. Este complejo se diferencia de su homólogo celular en que: ‐ Tiene más actividad kinasa frene a la proteína Retinoblastoma (dejando libre E2F para que active la transcripción de los genes de la fase S) e histona H1. ‐ No sigue los mecanismos normales de regulación de los CDK (kinasas activadoras e inhibidores-CDK: p16, p21, p27), sino que mimetiza en parte los complejos activados CDK/ciclina de las fases G1 y S. ‐ Interacciona con un amplio abanico de CDKs, incluyendo CDK-2, CDK-4, CDK5 además de con otros sustratos: histonas H1, p27, p21, bcl-2, Id-2, Cdc25A, Cdc6, Orcl-1 y caldesmona. ‐ Está implicada en la disregulación de la expresión genética de la célula inhibiendo la activación genética mediada por STAT-3. ‐ Además puede unirse a CDK-9. Se ha descubierto recientemente su rol en la regulación de p53, fosforilándola en su dominio de transactivación (DTA). Otros virus como CMV, VHH-1, VEB la usan por su principal función de iniciación y elongación, aunque no se ha constatado su participación en la oncegénesis de VHH-8. Su potencial oncogénico reside fundamentalmente en sus interacciones con p53 y bcl-2, promoviendo el paso G1-S (48). • Kaposina: proviene de un gen transformador con alto potencial oncogénico. La isoforma B se expresa en todas las células infectadas y puede ‐ Activar la vía p38-MK2 y bloquear la degradación de mRNA que transcribe citocinas necesarias para la supervivencia de la célula. ‐ Codificar microRNAs (miRNA). Jugarían un papel importante en el mantenimiento de la infección latente y la oncogénesis. • vIRFs son cuatro proteínas homólogas al Factor de Regulación de Interferón (IRF). Al menos tres de ellas (vIRF 1-3) alteran la respuesta celular a interferones, así como otras proteínas reguladoras como TGF-β y p53. vIRF-3 (LANA-2) es una proteína multifuncional expresada en LDP y ECM, mientras que no se detecta en SK. Se une a MM-1 (Modulador de Myc 1) y deja libre a c-myc que estimula la actividad transcripcional del promotor de CDK-4 y posiblemente de los promotores de otros genes regulados por c-myc, lo cual conlleva la reentrada al ciclo celular. Además existen evidencias in vitro de que vIRF-3 puede interaccionar directamente con p53 inhibiendo su transcripción (49). • K7-vIAP (Proteína Inhibidora de la Apoptosis) es un homólogo a la proteína survivina que protege célula de la muerte celular programada. • En la zona asociada a latencia del genoma de VHH-8 se han descubierto doce genes que codifican microRNA. Se han identificado hasta 81 genes cuya expresión se ve afectada en presencia de estos miRNA. Cinco de los que más inhibición sufren son genes relacionados con la proliferación, inmunomodulación, angiogénesis y apoptosis: SPP-1, S100A2, ITM2A, PRG-1 y THBS-1 (trombospondina 1, un potente anti-angiogénico que actúa a través de TGFβ) (50). La entrada en el ciclo lítico, como ya hemos visto, juega un papel fundamental en la patogénesis del virus. Como muchos otros aspectos de la misma sigue sin esclarecerse por completo. Se sabe que ciertos estímulos desencadenan in vitro este paso en VHH-8 y VEB. Entre ellos Tetradecanoil Phorbol Acetato (TPA), butirato de sodio y otros ácidos grasos de cadena corta. Sin embargo, seguimos sin conocer cuáles son los mecanismos exactos por los que tiene lugar in vivo. Se cree que la reactivación en células quiescentes tiene lugar al alterarse el equilibrio entre señales que inducen o suprimen la replicación lítica, como en situaciones de hipoxia o por la producción de citocinas inflamatorias (IFN-γ y oncostatina-M) (20). Al entrar en el ciclo lítico, se activan vías pro-supervivencia (v-Bcl-2, inhibición de caspasas por la proteína survivina-like) para bloquear la lisis celular y permitir así una mayor producción viral, además de retrasar la muerte celular lo suficiente para que el virus pueda completar su ciclo lítico. Además las citocinas actúan a nivel paracrino, confiriendo así protección a las células de alrededor que no están en ciclo lítico. Así, una proteína de la fase lítica, coopera con las funciones de latencia para promover la patogenia o actuar de otras formas en células no infectadas (43). Se han descrito múltiples mecanismos que intervienen en la regulación del ciclo vital del virus y es otro de los principales objetivos de estudio en la actualidad. • La proteína Transactivadora de la Replicación y de la Transcripción (TRA) codificada por el ORF50, desempeña un papel central en el paso de la forma latente a la lítica. La sola expresión de RTA es necesaria y suficiente para disparar el proceso lítico al completo ya que actúa como factor de transcripción activando la expresión de múltiples genes diana entre los que están vIL-6, ORF57, el suyo propio y el RNA nuclear poliadenilado, PAN o nut-1. Además interacciona con proteínas reguladoras de la transcripción para maximizar el proceso, entre las que se encuentran la Proteína de Unión a CREB (CBP), el complejo remodelador de cromatina SWI/SNF y el coactivador mediado por TRAP y CSL, una diana de la vía Notch. Además se ha asociado a otras vías como Ras (que puede ser disparada por VEGF, PDGF o citocinas y quimiocinas), ERK-1/2 y MEK-1/2, JNK y p38 MAPK. Se ha encontrado que también responde a los niveles de adrenalina/noradrenalina o a los de dopamina y sus derivados (51). • En líneas celulares de LDP se ha demostrado que el estrés que induce el paso de célula B a célula plasmática en el Retículo Endoplasmático (RE) provoca la activación de la proteína Grp78/BiP del RE. La activación libera su ligando natural, la enzima requeridora de inositol (IRE-1), que forma un complejo capaz de catalizar el procesamiento de la proteína de unión X-box 1 (XBP-1). Esta proteína es uno de los desencadenantes del ciclo lítico, activa el gen ORF50 y produce RTA para inducir el cambio de ciclo (52). • RSK1 y RSK2, kinasas S6 de los ribosomas p90, juegan un papel fundamental en el ciclo lítico. Son intermediarias de la vía ERK-MAPK activadas por ORF45. La inducción de ERK termina con la activación de factores de transcripción que reprograman la expresión genética del huésped, por ejemplo AP-1. Las MAPSK más conocidas son: ERK1-/ERK-2, p38-MAPK y la proteínkinasa activada por estrés/c-jun N terminal kinasa (SAPK/JNK1/JNK2/JNK3). RSK-1/RSK-2 se activan en la entrada del virus a la célula y con la expresión de sus genes, mientras que en la reactivación se activa a través de ORF45 (53). • Recientemente se ha descubierto la participación de dos kinasas, Pim-1 y Pim-3, que fosforilan a LANA-1, deshabilitando su represión sobre el ciclo lítico. Su actividad está inducida en las reactivaciones mediadas por estímulos fisiológicos o químicos, como IFN-α, IFN-γ, IL-3 o IL-6, lo cual explicaría la reactivación del virus en una situación de inflamación (54). • Se ha comprobado en cultivos de células de LDP que cuando la célula entra en ciclo lítico cambia la edición RNA del gen K12 que produce kaposina por las adenosin deaminasas de RNA (ADAR-1) del huésped. Mientras en la fase latente se produce un transcrito sin editar y con actividad tumoral por incluir la Kaposina A. En el ciclo lítico se produce un aumento de la transcripción del gen, quizá porque las citocinas producidas en esta fase induzcan las ADAR-1, aunque los transcritos producidos están editados y no contienen Kaposina A, por lo que no tiene actividad tumoral. Por tanto, durante el ciclo latente aumenta la población de células infectadas, mientras que en el ciclo lítico, la función del transcrito editado está aún por determinar (55). Perspectivas de futuro El escaso conocimiento del que todavía se dispone acerca de la transmisión, biología, infectividad y patogenia de VHH-8 limita los avances en cuestiones de prevención, diagnóstico y tratamiento de los tumores con los que se relaciona. Muchos aspectos siguen sin estar esclarecidos como los factores de riesgo, predisposición genética o el potencial patógeno de diferentes genotipos. Conforme se vaya progresando en estas materias dispondremos de nuevos marcadores diagnósticos y nuevas dianas terapéuticas que ampliarán los campos de acción con los que enfrentarse a esta enfermedad. Dado el constante incremento en las tasas de infección por VIH y la relación causal existente con VHH-8, su estudio adquiere una importancia creciente en la medicina actual. VIRUS LINFOTROPO T HUMANO TIPO I (VLTH-I) Los principales virus de la familia Retroviridae con potencial oncogénico son: el Virus Linfotropo T Humano tipo I (VLTH-I), el virus homónimo tipo II (VLTH-II) e indirectamente el VIH. El primero de ellos es el más estudiado y el que se desarrollará con más detalle en este apartado. Distintos estudios lo asocian a la Leucemia/Linfoma de células T del Adulto (LTA) y a otro tipo de patologías como la Mielopatía asociada a VLTH-I, también llamada Paraparesia Espástica Tropical (MAH/PET). El VLTH-I infecta y transforma preferentemente Linfocitos T CD4. Esto último lo diferencia del VLTH-II que posee más afinidad por Linfocitos T CD8. Este segundo virus ha sido relacionado con la Leucemia Atípica de Células Peludas, pero los estudios más recientes no han corroborado dicha hipótesis (56). Por otro lado la coinfección VLTH-II + VIH sí parece aumentar la incidencia de la Leucemia/Linfoma de células T asociada/o a VLTH-II si se compara con los portadores de VLTH-II solamente (57). Mecanismos moleculares de la oncogénesis: El potencial oncogénico de este virus viene determinado y regulado por la activación y demás propiedades de la proteína Tax (pTax) (58). Ésta promueve la mutación genética en las células infectadas, inhibe el control del ciclo celular y dirige la expresión génica viral. Asimismo la pTax activa los factores de transcripción NFKB y SRF (59). También activa los genes celulares que codifican para: IL-2, receptor α de IL-2, GM-CSF y PTHRP. Además la activación de dicha proteína inhibe la expresión de los genes para la Polimerasa-β y la CDK inhibidora de p16 (60). La patogenia de la infección por VLTH-I viene determinada por los siguientes puntos (61;62): • La infección primaria a edad temprana. La inmadurez de la inmunidad de tipo 1 en el momento de la infección facilita el incremento de la carga viral. En proporción a este incremento se producirá una extraordinaria proliferación clonal de Linfocitos T infectados durante un largo periodo de tiempo. • La extensa expansión clonal de los Linfocitos T infectados que se autonomiza y junto a la inestabilidad genética orquestada por la pTax parece ser uno de los factores patogénicos más importantes. • La pérdida de la expresión de la pTax: Si se produce la pérdida de la producción de dicha proteína en uno de los clones, éste será invisible para la los Linfocitos Tc sanos del huésped. Este nuevo clon se considera ya premaligno pues escapará de la vigilancia de los Linfocitos Tc. Una vez desarrollada la LTA se evidencia la disminución de los títulos de Ac contra la pTax, no así los del resto de Ac contra el propio virus que sí que seguirán aumentando mientras se desarrolla la leucemia/linfoma; además en las propias células cancerosas de LTA se pone de manifiesto la ausencia de la expresión de dicha proteína. • Mutaciones genéticas adquiridas estocásticamente, que conllevan cambios genéticos y epigenéticos y conducirán a la malignidad del proceso. Cofactores: Existen una serie de factores que influyen en el desarrollo de LTA en portadores de VLTH-I, éstos serían los siguientes: • Género: Es bien conocido que los hombres tienen más probabilidad de desarrollar LTA que las mujeres aún siendo ellas las que poseen una mayor tasa de infección (63). La explicación a este hecho sería la existencia de diferencias según el género en la inmunidad celular (que es uno de los puntos clave en el control de la infección una vez establecida). En hombres portadores del virus se ha evidenciado una respuesta atenuada a la reacción de la tuberculina lo que evidencia una disminución en la respuesta inmune de tipo 1 (64). • Edad: En población joven no se han demostrado diferencias en la incidencia de LTA en relación con el género. En portadores asintomáticos, los hombres mayores de 30 años son más propensos a tener una alta carga viral que las mujeres. Pero no existen diferencias en población de menor edad (65). • Factores genéticos: Los distintos estudios realizados para poner de manifiesto la susceptibilidad genética para la malignidad inducida por VLTH-I, en relación con distintos fenotipos de HLA en pacientes con LTA, no han aportado resultados consistentes. Aún así estudios recientes (66) proponen la división de dos subgrupos en función de la división fenotípica identificada en el HLA en cuanto a la respuesta inmune celular a la infección por VLTH-I (in vitro). Así pues por un lado estarían los “Low responders” que se asocian al desarrollo de LTA. Y por otro los “High responders” que asociarían un mayor riesgo de desarrollar MAH/PET. De todos modos lo normal es que los portadores posean un HLA intermedio. • Estrongiloidiasis y desnutrición: Inicialmente se proponía que el riesgo de desarrollar LTA estaba asociado a desnutrición y exposición repetida a filariasis en la infancia. Ya que ambas condiciones eran comunes en las zonas endémicas de Japón. Por otro lado no se han observado diferencias en la prevalencia de Ac contra Strongyloides stercolaris entre los portadores de VLTH-I y los controles en Jamaica. De todos modos es evidente que una coinfección va en detrimento del huésped y se ha demostrado que en los portadores coinfectados la carga viral es más elevada. Este hecho parece ser debido a que la población coinfectada posee un sistema inmune alterado que disminuye la capacidad para controlar ambas infecciones. En individuos coinfectados aparece una disminución de la respuesta inmune tipo 2 (responsable del control contra parásitos), este hecho se pone de manifiesto con la medida en suero de IgE e IL-5 (67). Del mismo modo existe en población coinfectada, una alteración de la respuesta inmune de tipo 1, evidenciada por la disminución de anticuerpos anti-EBNA para VEB, con el consiguiente incremento de la carga viral de VLTH-I (68). Se considera que un paciente coinfectado posee alrededor de cinco veces más carga viral en comparación con los individuos no infectados por S. stercolaris (69). Como conclusión entenderemos que la estrongiloidiasis incrementa el riesgo de desarrollar LTA en individuos coinfectados con VLTH-I. VIRUS DEL PAPILOMA HUMANO (VPH) Se han identificado más de 100 tipos VPH de los cuales aproximadamente la mitad infectan el tracto genital. Los VPH genitales se dividen en bajo riesgo (tipos 6 y 11, por ejemplo) y alto riesgo (tipos 16, 18, 31, 33, 39, 45, 51, 52, 56, 58, 59, 66, 68, y posiblemente los tipos 26, 73 y 82) (9). Aproximadamente el 80% de los cánceres de canal anal y el 30% de los de vulva, vagina, pene y orofaríngeos pueden atribuirse al VPH (70). Cofactores infecciosos en el cáncer de cérvix (71) 1. Virus Herpes Simple (VHS): VHS-2 tiene mayor tropismo por los tejidos genitales y recurre con mayor frecuencia en el tracto genital que VHS-1. Un estudio de la IARC (Smith el al., 2002) que reunió datos de siete estudios de casos-controles diferentes llevados a cabo en Brasil, Colombia, Marruecos, Filipinas, España y Tailandia, encontró una mayor seroprevalenciade VHS-2 en 1158 casos de carcinoma epidermoide y 105 casos de adenocarcinoma que en 1117 controles; tras limitar el análisis a sujetos VPH-DNA positivas y ajustarlo para seropositividad para C. trachomatis, la asociación de anticuerpos séricos VHS-2 con carcinoma epidermoide (odds ratio, 2.2; 95% CI, 1.4-3.4) y adenoacarcinoma (odds ratio, 3.4; 95% CI, 1.5-7.7) era todavía significativa. 2. Otros Herpesvirus: los β-Herpervirus Citomegalovirus (CMV), Herpesvirus Humano tipo 6 (VHH-6) y VHH-7 han sido detectados en muestras cervicales. Se ha sugerido que sólo son hallazgos concomitantes más que co-factores en la carcinogénesis tumoral (Tran-Thanh y cols., 2002). Otros, sin embargo, han determinado que existe asociación entre CMV y neoplasia de cérvix (Koffa y cols., 1995), entre VHH-6 y cáncer cervical (Yadav y cols., 1996) y entre VHH-7 y neoplasia cervical de alto grado (Lanham y cols., 2001). También se han realizado estudios para conocer si existe relación entre los niveles de anticuerpos contra CMV y el desarrollo de estas neoplasias, pero los resultados obtenidos han sido conflictivos, ya que algunos han observado dicha asociación y otros no. El virus de Epstein-Barr (VEB), un γ-Herpesvirus, puede infectar células epiteliales y podría jugar un papel en el desarrollo del carcinoma nasofaríngeo. Los resultados obtenidos en diferentes estudios han sido confusos. 3. Chlamydia trachomatis: Está asociada a metaplasia escamosa y a ectopia hipertrófica. Ha sido bien estudiado como potencial cofactor en el desarrollo de cáncer cervical. Dos extensas investigaciones epidemiológicas han observado una asociación significativa entre anticuerpos de C. trachomatis y carcinoma epidermoide cervical (Koskela et al., 2000; Smith, J.S et al., 2004). Mecanismos moleculares de la oncogénesis (71) Capacidad transformadora del VPH - VPH mucosos: un grupo de VPH mucosotropos, incluyendo los tipos de alto riesgo 16 y 18, están asociados a más del 99% de los carcinomas cervicales. En estos cánceres, el DNA viral se encuentra, frecuentemente, integrado en cromosomas del huésped. Células epiteliales cervicales que contienen DNA VPH 16 tienen un crecimiento mayor en comparación a las células que mantienen el genoma viral en una localización extracromosómica. Esto se relaciona con la expresión incrementada de dos genes virales, E6 y E7. Las proteínas tempranas E6 y E7, se unen e inactivan el producto de un gen supresor de tumores, p53, y la proteína supresora de tumores de retinoblastoma, respectivamente. Es necesaria la expresión de los genes virales E6 y E7 para el continuo crecimiento de las líneas celulares derivadas del cáncer cervical. Este hecho apoya la hipótesis de que E6 y E7 están causalmente relacionados con la aparición y mantenimiento del cáncer de cérvix. La continua expresión de estas proteínas puede dar lugar al acumulo de mutaciones en el genoma celular, requerido para la conversión maligna consiguiente. Tanto E6 como E7 cooperan en la transformación de las células epiteliales, sin embargo, sólo vemos un fenotipo completamente maligno tras el cultivo prolongado de las células transformadas, hecho que apoya que la transformación inducida por VPH está formada por varios pasos. En un modelo de ratones transgénicos, se ha visto que, sólo la expresión de VPH 16 E6 es suficiente para inducir carcinomas (Song y cols., 1999). Por el contrario, E6 y E7 de tipos de VPH mucosos de bajo riesgo poseen muy poca actividad transformadora in vitro (Farr y cols., 1991; Sang & Barbosa, 1992). Reid y cols. en 2004 demostraron la presencia de genoma integrado de VPH 11 de bajo riesgo, pero no de VPH 6, en cánceres desarrollados en pacientes con papilomatosis respiratoria recurrente de inicio temprano (Reidy y cols., 2004). - VPH cutáneos: diversos estudios epidemiológicos han demostrado que un subgrupo de VPH cutáneos, también llamados tipo epidermoplasia verruciforme (EV)-VPH, se encuentran en cánceres de piel no-melanoma. El número de copias de DNA VPH-EV es mucho menor que una copia por célula (Iftner y cols., 2003). La mayoría de VPH tipo cutáneo expresan productos de los genes E6 y E7 que son estructuralmente similares a aquellos que aparecen en los tipos mucosos, pero su genoma no contiene un ORF E5 identificable (Pfister, 2003). Caldeira y cols. (2003) analizaron las propiedades in vitro de las proteínas E7 de los virus cutáneos EV(VPH 20 y28) y no EV-(VPH 10) que frecuentemente se encuentran en la piel. Se demostró que la E7 de VPH 38 se une e inactiva el supresor de tumores pRb e induce la pérdida del control de la transición G1/S. Por el contrario, la proteína E7 VPH 10 y VPH 20 no juegan ningún papel en las actividades transformadoras in vitro. Además, las E6 y E7 del VPH 38 demostraron su capacidad para inmortalizar queratinocitos humanos primarios, lo que sugiere que la infección por VPH 38 tiene un papel en la carcinogénesis de piel. Propiedades bioquímicas de las proteínas de VPH - E5: a la E5 de VPH de alto riesgo se le considera carcinogénica porque transforma fibroblastos murinos y queratinocitos en cultivos de tejidos, aumenta el potencial de inmortalización de E6 y E7 y, junto con E7, estimula la proliferación de células primarias humanas y de ratón. Se ha sugerido que EGFR, un receptor de tirosin-kinasa, media la actividad biológica de la proteína E5 de VPH 16, y plantean que la E5 activa EGFR e induce señales mitogénicas y transformación celular a través de este receptor (Tomakidi y cols., 2000). - E6: la asociación de E6 y p53 da lugar a la degradación de p53 a través de la reclutación de una ubiquitina ligasa, E6-AP, que da lugar a la inhibición de las actividades reguladoras trancripcionales de p53 en los cultivos de células de tejidos (Mietz y cols., 1992). - E7: es la más conocida por su habilidad para asociarse con la proteína supresora tumoral pRb. Promueve la degradación de la misma a través de un paso mediado por un proteasoma y elimina su capacidad para unirse e inactivar funcionalmente a los factores de transcripción celulares E2F. Además puede unirse a otras proteínas pocket relacionadas con pRb y también interacciona con diversos componentes de la familia E2F. La inactivación de las proteínas “pocket” por parte de E7 es necesaria pero no suficiente para provocar el potencial transformador de E7 (Kioyono y cols., 1998). Las proteínas E7 de alto riesgo forman un complejo con ciclinas e inactivan los inhibidores de asociados a kinasas p21 y p27 (Jones y cols., 1997). E7 se asocia con, o altera las actividades de múltiples factores celulares que normalmente contribuyen a la regulación del ciclo celular. - Las proteínas E6/E7 del VPH 16 y el erbB-2 cooperan en la transformación neoplásica de células orales epiteliales primarias: (5) en este estudio se ha estudiado el efecto de la cooperación entre VPH 16 y el receptor de tirosin kinasa erbB-2 en el complejo E-caderina/catenina y la transformación neoplásica del epitelio oral normal. Se ha determinado que la sobreexpresión aislada de ErbB-2 o E6/E7 no afecta al señalado complejo, por lo que no induce transformación en las células epiteliales orales. Sin embargo, la coexpresión de E6/E7 y erbB-2 disminuye la expresión de E-caderina y catenina. Los datos sugieren estos dos factores participan en la carcinogénsis, al menos en parte, a través de la conversión de la γ-catenina, que actúa como regulador transcripcional. Esto potencia a la ciclina D1, c-myc y otras oncoproteínas necesarias para la alteración de del complejo E- caderina/catenina y la transformación de las célula orales epiteliales normales. Propiedades biológicas de las proteínas de VPH • Inmortalización: las proteínas E6 y E7 de VPH mucosos de alto riesgo actúan de manera independiente o sinérgica para inmortalizar múltiples tipos celulares incluidos queratinocitos, epitelio cervical y células epiteliales mamarias. Sin embargo, las proteínas E6 y E7 de un solo tipo cutáneo EV-VPH, el 38 concretamente, ha demostrado inmortalizar queratinocitos humanos primarios (Caldeira y cols., 2003). Las proteínas E7 de tipos mucosos de alto riesgo (VPH 16, 18, 31) pero no de bajo riesgo (VPH 6, 11) cooperan con una proteína ras activada para transformar las células epiteliales cervicales humanas y el riñón de rata neonata (Storey y cols., 1988). La E7 de tipos de bajo riesgo mucosos VPH 6 y cutáneos EVVPH 8 han demostrado tener una actividad inmortalizadora más débil en comparación con los de alto riesgo VPH 16 (Halbert y cols., 1992; Schimitt y cols., 1994). Esta diferencia se relaciona con la capacidad de estas proteínas E7 para inducir la degradación de pRb más que con su afinidad por pRb (Giarrè y cols., 2001). La proteína E5 puede aumentar la inmortalización de los queratinocitos efectuada por E6 y E7. El mecanismo por el cual E6 contribuye a la inmortalización en controvertido. Algunos estudios han relacionado su potencial de inmortalización con su habilidad para inducir la expresión de telomerasas, ya que la actividad de las mismas está claramente inducida por E6. El nivel de actividad de la telomerasa en células E6-positivas aumenta más cuando se convierten en inmortales aunque los niveles de expresión de E6 no varíen (Fu y cols., 2003), lo que sugiere que existen otros eventos que contribuyen a la activación de la telomerasa. Otros estudios han relacionado el potencial inmortalizador de E6 en células epiteliales mamarias y queratinocitos con su inactivación de p53 (McMurray & McCance, 2004). • Inestabilidad genómica: ésta es otra característica distintiva de las células que expresan E6 y E7, lo cual ha sido observado en múltiples tipos celulares. Las anomalías incluyen monosomías y trisomías, roturas e interrupciones en la cromatina y cromosomas aberrantes. Algunas pérdidas alélicas se han asociado a con genes que podrían tener relación con transformaciones malignas y/o progresión. Por ejemplo, la pérdida del alelo 3p y 10p se asocian con la activación de telomerasas (Steenbergen y cols., 1998), lo cual constituye un paso crucial en la inmortalización mediada por VPH de alto riesgo. Las proteínas E6 y E7 de VPH 16, de alto riesgo (pero no VPH 6, de bajo riesgo) pueden inducir anomalías mitóticas. La habilidad de E6 para inducir inestabilidad genómica es probablemente reflejo de su habilidad para inhibir la función de p53, lo cual irrumpe en el normal proceso de reparación del DNA y da lugar al acumulo de cambios genéticos. La inestabilidad genómica producida por E7 puede ser reflejo de de su efecto sobre la biogénesis de los centrosomas y el consecuente defecto en la segregación de los cromosomas de las células hijas durante la división celular (Balsitas y cols., 2003). Otros estudios han demostrado que E7 puede inducir anomalías centrosómicas a través de mecanismos independientes a pRb (Duensing & Münger, 2003). Mediante la utilización de una línea celular VPH 16-positiva derivada de una lesión intraepitelial escamosa de bajo grado, se ha sugerido que la adquisición de la inestabilidad cromosómica está relacionada con la integración del genoma viral (Pett y cols., 2004). Se ha demostrado lo contrario en cultivos con queratinocitos humanos que contienen VPH 16, donde se vio inestabilidad genómica en ausencia de integración viral (Duensing y cols., 2001). • Respuestas al daño en el DNA: tanto E6 como E7 pueden obstaculizar las respuestas contra el daño en el DNA. Se cree que es debido a su actividad como inhibidoras de la detención del ciclo celular mediado por p53. Se relaciona con la capacidad de E6 para unirse e inhibir p53. Con E7, no sólo se relaciona con su habilidad para interrumpir la función de reguladora del ciclo celular de pRb, sino que también con su capacidad para inactivar p21, que es el inhibidor de kinasa ciclinadependiente que se induce cuando se activa p53 en respuesta a daño celular. Se piensa que la disrupción de las respuestas contra el daño celular contribuyen al acumulo de alteraciones genéticas en células VPH-positivas, incluyendo aquellas que podrían contribuir a la carcinogénesis. • Proliferación celular y diferenciación: mientras que en un epitelio estratificado normal las células suprabasales están retraídas del ciclo celular, VPH puede reprogramar las células suprabasales para mantener la síntesis de DNA que puede contribuir a la producción de virus a través de la amplificación del genoma viral. E7 es necesario y suficiente para este fenotipo. La inactivación aislada de pRb puede causar el mantenimiento de la síntesis de DNA (Balsitis y cols., 2003). E6 y E7 de alto riesgo modulan la apoptosis. La apoptosis inducida por E7 ocurre mediante pasos dependientes e independientes de p53. Evidencia experimental del rol de VPH mucosos de alto grado de en la conversión maligna y el cáncer cervical humano (71) Requerimiento de la expresión génica de VPH para el crecimiento celular y la invasión La expresión de las proteínas E6 y E7 es esencial para la inmortalización, pero son necesarios otros factores para adquirir un fenotipo totalmente transformado. Se ha visto que las células VPH inmortalizadas se convierten en carcinogénicas espontáneamente o después de ser tratadas con carcinógenos químicos o expuestas a irradiación-γ. La continua expresión de E7 de VPH 16 en presencia de ras es necesaria para el mantenimiento del fenotipo transformado en células primarias de roedores (Crook y cols., 1989). También se conoce el papel de la expresión constitutiva de c-fos en la transformación de las células del cáncer de cérvix (van Riggelen y cols., 2005). En una línea celular VPH 16 positiva llamada SiHa, se estudió la interferencia de la expresión de VPH 16 E6 y E7. Para esto se utilizó RNA short-interfering (siRNA) de E6. Se demostró que el E6siRNA disminuía los niveles de mRNA que codificaban E6 y E7 e inducía acumulo nuclear de p53 (Yosinouchi y cols., 2003). Integración de secuencias de VPH En la mayoría de los tumores invasivos, el genoma de los VPH de alto riesgo se integra en el genoma del huésped. Esta integración también puede verse en lesiones premalignas. Por el contrario, el DNA de VPH se localiza extracromosómicamente en lesiones benignas y de bajo riesgo. Es difícil encontrar VPH de bajo riesgo integrado en tumores. Sin embargo, Reidy y cols. (2004) estudiaron muestras de tejidos de pacientes con historia de papilomatosis laríngea recurrente benigna de inicio temprano que habían desarrollado cáncer. Encontraron VPH 11 integrado en estas muestras. In vitro, se ha sugerido la mayor habilidad de los VPH de alto riesgo para integrarse en el DNA del huésped en comparación con los tipos de bajo riesgo. Se considera que la integración es un evento molecular importante en la carcinogénesis inducida por VPH. La detección de la transcripción de genes tempranos por PCR es más sensible en cánceres y lesiones benignas o displasias cervicales, donde se ha demostrado que la presencia de genoma integrado se relaciona con la severidad de la enfermedad, en particular en VPH 18 (Hudelist y cols., 2004). En el cáncer de lengua, se ha demostrado que la presencia de genoma de VPH 16 fuera del cromosoma y un elevado número de copias están relacionadas con un mejor pronóstico (Mellin y cols., 2002). La integración de genoma de VPH afecta tanto a la expresión génica viral como del huésped. La expresión génica de VPH está regulada por activadores y represores transcripcionales virales y celulares. La regulación normal se altera debido a la integración viral, lo cual se traduce en la continua expresión de las proteínas E6 y E7 que da lugar al crecimiento selectivo. La integración se relaciona con la síntesis incrementada de la proteína E7. El hecho de que la integración de las secuencias de DNA de VPH tenga lugar en localizaciones predilectas de los cromosomas humanos ha sugerido un patrón de integración no aleatorio. En los carcinomas cervicales, Ferber y cols. (2003) observaron que la integración de VPH tenía lugar alrededor del gen hTERT, lo que daba lugar a un incremento en la expresión del mismo. Por otro lado, el DNA de VPH 18 se integra en las proximidades del gen c-myc en numerosos cánceres de cérvix, pero no se ha observado una regulación positiva en la expresión endógena del protooncogén. Algunos estudios citogenéticos y moleculares han demostrado que pueden encontrarse secuencias de DNA de VPH 16 y 18 integradas en loci cromosómicos conocidos como sitios frágiles en cánceres cervicales. Una de estas regiones, la FRA3B, abarca Tetrámeros Frágiles de Histidina (FHIT), gen supresor de tumores localizado en el cromosoma 3p. Se ha demostrado una clara asociación entre la pérdida de expresión de FHIT y la progresión de CIN VPH 16-positivo (Butler y cols., 2002). Se ha visto que los cánceres cervicales invasivos que expresan E6 y E7 de VPH de alto riesgo contenían una expresión normal de FHIT, mientras que sólo se detectaron unas pocas transcripciones de dichas proteínas cuando la expresión de FHIT era anormal. Esto sugiere que E6 y E7 podrían reprimirse en presencia de aberraciones de FHIT. Estos estudios favorecen la idea de que el genoma de VPH pueda interferir con funciones celulares críticas mediante la mutagénesis insercional. Algunas observaciones han demostrado que el fenotipo maligno no puede deberse únicamente a la expresión de genes de VPH. Se cree que la modificación de los genes del huésped que interfieren con la expresión o función de los genes virales podría contribuir a la evasión inmune, y a la progresión en invasión tumoral (zur Hausen, 1999). Anomalías cromosómicas en los cánceres asociados a VPH Se han descrito numerosos cambios cromosómicos en muchos cánceres asociados a VPH incluyendo el cervical, vulvar y de cabeza y cuello. Se han descrito aneuploidias en cánceres cervicales y sus precursores. La pérdida del brazo corto del cromosoma 3 y la ganancia en el brazo largo son los eventos más frecuentemente asociados con la progresión de lesiones de alto grado a cáncer cervical (Nishimura, M. y cols., 2000). Las alteraciones en 3q son las que más comúnmente se asocian a la severidad de la neoplasia intraepitelial anal, lo cual es similar a lo encontrado en cánceres de cérvix, lo que sugiere y paso molecular común para estas lesiones malignas asociadas a VPH. Se ha sugerido que la pérdida de un locus en el cromosoma X y la activación del encogen erb-B2 son importantes en la progresión de metástasis. Se encuentran más aberraciones cromosómicas en lesiones de alto grado con DNA de VPH integrado que en lesiones de bajo grado. Los genes p53 y pRb se encuentran mutados con menos frecuencia en tumores y líneas celulares VPH-positivas que en VPH-negativas (Tomasino y cols., 2003). De manera similar, la expresión de E6 de VPH 16 en cánceres de cabeza y cuello se relacionan con la ausencia de mutaciones en el gen p53 (Dai et al., 2004). La expresión de los genes E6 y E7 de VPH de alto riesgo en la capa basal induce inestabilidad y aneuploidía cromosómica. Se ha sugerido recientemente que este evento precede a la integración del genoma viral que, sucesivamente, provoca la continua expresión de los genes tempranos (Melsheimer y cols., 2004). Alteraciones de proto-oncogenes específicos - Ras: tanto en lesiones cervicales malignas como premalignas se han descrito mutaciones en la familia ras. Los niveles de mRNA de H- y n-ras eran significativamente más elevados en carcinomas cervicales que en tejido normal o CIN. Sin embargo, no se encontró relación entre la expresión de cada uno de los genes ras y la presencia de VPH (Mammas y cols., 2004). Por otro lado se han encontrado tasas más elevadas de mutación k-ras en el codón 12 en muestras de cérvix no canceroso que contenían tipos VPH de alto grado que en aquellas que estaban infectadas por VPH de bajo riesgo. - Myc: parece ser que los VPH se integran en las proximidades de c-myc, lo cual justifica la búsqueda de alteraciones en este protooncogén en lesiones asociadas con VPH. Se ha descrito una gran amplificación de c-myc en los cánceres cervicales en comparación a las lesiones cervicales benignas y premalignas (Abba y cols., 2004). Últimamente se ha añadido la participación de la proteína myc en la inmortalización inducida por VPH (Veldman y cols., 2003). Se ha visto que la E6 de VPH de alto riesgo está asociada a los complejos myc y activan el promotor hTERT. El antagonista específico de myc, mad, reprime la transactivación-E6 de hTERT. - Erb: la proteína y el mRNA HER-2/NEU fueron detectados en una gran proporción de adenocarcinomas cervicales. Esta expresión fue asociada en mayor medida con lesiones que contenían VPH 16 que con aquellas que contenían VPH 18. También se ha relacionado un incremento de la expresión ErbB-2 con la progresión de la enfermedad y se ha considerado que es un evento tardío en la carcinogénesis cervical (Kersemaekers y cols., 1999). - Otros: se ha descrito la metilación de genes virales o LCRs (long control region) de VPH de alto riesgo. El LCR de VPH 16 (Badal y cols., 2003) y 18 (Badal y cols., 2004) está hipermetilado en muestras cervicales normales y de bajo grado y progresivamente va disminuyendo su grado de metilación en muestras de alto grado. Bibliografía (1) SA Raza, GM Clifford and S Franceschi. Worlwide variation in the relative importance of hepatitis B and hepatitis C viruses in the hepatocellular carcinoma: a systematic review. British Journal of Cancer 96, 1127-1134. 29-1-2007. (2) M Levrero. Viral hepatitis and liver cancer: the case of case of hepatitis C. Oncogene 25, 3834-3847. 2006. (3) Alexander Tan, Shiou-Hwie Yeh, Chun-Jen Liu, Claudia Cheung and Pei-Jer Chen. Viral hepatocarcinogenesis: from infection to cancer. Liver International , 175-188. 2008. (4) Tong MG, EL-Farra MS, Rey Kes AR, et al. Clinical outcames after transfusion associated hepatitis C. NEJM 332, 1463-1465. 2009. (5) Kazuhiko Koike. Dysfunction of energy metabolism in hepatic cacinogenesis. Hepatits C virus contributes to hepatocarcinogenesis by modulating metabolic and intracelular signaling pathways. Journal of Gastroenterology and Hepatology 22[suplemento 1], 108-111. 2007. (6) F.Farinati, R.Cardin, M.Bortolami, P.Burra, F.P Russo, M.Rugge, et al. Hepatitis C virus: from oxygen free radicals to hepatocellular carcinoma. Journal of Viral hepatitis 14, 821-829. 2007. (7) Lee WM. Hepatitis B virus infection. NEJM 337, 1733-1745. 1997. (8) Surakit Pungpapong M, Ray KIM M, John J Poterucha M. Natural History of Hepatitis B Virus Infection: An Update for Clinicians. Mayo Clin Proc 82(8), 967975. 2007. (9) Nancy E.Mueller, Brenda M.Birmann, Julie Parsonnet, Mark H.Schiffman, Serri O.Stuver. Infectious Agents. Cancer epidemiology and Prevention. 2006. (10) Krishna SM, James S, Balaram P. Expression of VEGF as prognosticator in primary nasopharyngeal cancer and its relation to EBV status. Virus research . 1-9-2005. (11) Guo L, Guo Y, Xiao S. Expression of Etk/Bmx tyrosine kinase in the tumorigenicity of nasopharyngeal epithelium and its relation with EB virus infection and the apoptosis-related protein Bcl-2. Cancer lett. 8-2-2006. (12) Li X, Liu X, Li CY, Ding Y, Chau D, Li G, et al. Recombinant adeno-associated virus mediated RNA interference inhibits metastasis of nasopharyngeal cancer cells in vivo and in vitro by suppression of Epstein-Barr virus encoded LMP-1. International Journal of Oncology . 2006. (13) Martorelli D, Houali K, Caggiari L, Vaccher E, Barzan L, Franchin G, et al. Spontaneous T cell responses to Epstein-Barr virus-encoded BARF1 protein and derived peptides in patients with nasopharyngeal carcinoma: bases for improved immunotherapy. International journal of cancer . 1-9-2008. (14) Rimar D, Rimar Y, Keynan Y. Human Herpesvirus-8: Beyond Kaposi's. IMAJ 8. 2006. (15) Russo JJ, Bohenzky RA, Chien MC, Chen J, Yan M, Maddalena D, et al. Nucleotide sequence of the Kaposi sarcoma-associated herpesvirus (HHV8). Proc Natl Acad Sci U S A 93, 14862-14867. 1996. (16) Della Bella S, Taddeo A, Calabrò ML, Brambilla L, Bellinvia M, Bergamo E, et al. Peripheral Blood Endothelial Progenitors as Potential Reservoirs of Kaposi's Sarcoma-Associated Herpesvirus. PLoS ONE 3. 2009. (17) Johnston C, Orem J, Okuku F, Kalinaki M, Saracino M, Katongole-Mbidde E. Impact of HIV Infection and Kaposi Sarcoma on Human Herpesvirus-8 Mucosal Replication and Dissemination in Uganda. PLoS ONE . 20-1-2009. (18) Mbulaiteye SM, Pfeiffer RM, Dolan B, Tsang VCW, Noh J. Seroprevalence and Risk Factors for Human Herpesvirus 8 Infection, Rural Egypt. Emerg Infect Dis , 586591. 2008. (19) Adele CA, Elizabeth SF, Mariana M, Cavalheiro, Fabiana ST, Marcos S. Latent human herpesvirus - 8 (HHV-8) infection in female commercial sex workers from Imbituba, Santa Catarina, Brazil. Braz J Infect Dis 11, 9-11. 2007. (20) Edelman DC. Human herpesvirus 8--a novel human pathogen. Virology Journal . 29-0005. (21) Hladik W, Dollard SC, Mermin J, Fowlkes AL, Downing R, Amin MM, et al. Transmission of Human Herpesvirus 8 by Blood Transfusion. NEJM 355, 13311338. 28-9-2006. (22) Gambus G, Bourbolia D, Whitby D, Ruiz L, Weiss R, Casabona J. Seroprevalence of human herpes virus type 8 (HHV8) infection in Spain. Chicago (USA) 1998. (23) Munoz P, Alvarez P, de Ory F, Pozo F, Rivera M, Bouza E. Incidence and clinical characteristics of Kaposi sarcoma after solid organ transplantation in Spain: importance of seroconversion against HHV-8. Medicine 81, 293-304. 2002. (24) Adiguzel C, Bozkurt SU, Kaygusuz I, Uzay A, Tecimer T, Bayik M. Human herpes virus 8-unrelated primary effusion lymphoma-like lymphoma: report of a rare case and review of the literature. APMIS , 222-229. 2009. (25) Gessain A. [Human herpesvirus 8 (HHV-8): clinical and epidemiological aspects and clonality of associated tumors] [Article in French]. Bull Acad Natl Med 192, 1189-1204. 2008. (26) Powles T, Stebbing J, Bazeos A, Hatzimichael E, Mandalia S, Nelson M, et al. The role of immune suppression and HHV-8 in the increasing incidence of HIVassociated multicentric Castleman's disease. Ann Oncol 20, 775-779. 2009. (27) Chadburn A, Hyjek EM, Tam W, Liu Y, Rengifo T, Cesarman E, et al. Immunophenotypic analysis of the Kaposi sarcoma herpesvirus (KSHV; HHV-8)infected B cells in HIV+ multicentric Castleman disease (MCD). Histopathology 53, 513-524. 2008. (28) Knoell KA, Hendrix JD, Stoler MH, Patterson JW, Montes CM. Absence of human herpesvirus 8 in sarcoidosis and crohn disease granulomas. Arch Dermatol 141, 909-910. 2005. (29) Sadeghian MH, Katebi M, Ayatollahi H, Keramati MR. Immunohistochemical study association between human herpesvirus 8 and multiple myeloma. Int J Hematol 88, 283-286. 2008. (30) Csire M, Mikala G, Peto M, Jánosi J, Juhász A, Tordai A, et al. Detection of four lymphotropic herpesviruses in Hungarian patients with multiple myeloma and lymphoma. FEMS Immunol Med Microbiol 49, 62-67. 2007. (31) Tedeschi R, Luostarinen T, De Paoli P, Gislefoss RE, Tenkanen L, Virtamo J, et al. Joint Nordic prospective study on human herpesvirus 8 and multiple myeloma risk. Br J Cancer 93, 834-837. 3-10-2005. (32) Pyakurel P, Pak F, Mwakigonka AR, Kaaya E, Biberfeld P. KSHV/HHV-8 and HIV infection in Kaposi's sarcoma development. Infectious Agents And Cancer . 2-22007. (33) Zeng Y, Zhang X, Huang Z, Cheng L, Yao S, Qin D, et al. Intracellular Tat of Human Immunodeficiency Virus Type 1 Activates Lytic Cycle Replication of Kaposi's Sarcoma-Associated Herpesvirus: Role of JAK/STAT Signaling. J Virol 81, 2401-2417. 2007. (34) Liu J, Martin HJ, Liao G, Hayward SD. The Kaposi's Sarcoma-Associated Herpesvirus LANA Protein Stabilizes and Activates c-Myc. J Virol 81. 2007. (35) Mansouri M, Rose PP, Moses AV, Früh K. Remodeling of Endothelial Adherens Junctions by Kaposi's Sarcoma-Associated Herpesvirus. J Virol 81, 9615-9628. 2008. (36) Stuber S, Mattsson K, Flaberg E, Kati E, Markasz L, Sheldon JA. HHV-8 encoded LANA-1 alters the higher organization of the cell nucleus. Mol Cancer . 2007. (37) Zhang YJ, Bonaparte RS, Patel D, Stein DA, Iversen PL. Blockade of Viral Interleukin 6 Expression of Kaposi's Sarcoma-Associated Herpesvirus. Mol Cancer Ther.Author manuscript 7, 712-720. 1-3-2009. (38) Morris VA, Punjabi AS, Lagunoff M. Activation of Akt through gp130 Receptor Signaling Is Required for Kaposi's Sarcoma-Associated Herpesvirus-Induced Lymphatic Reprogramming of Endothelial Cells. J Virol 82, 8771-8779. 2008. (39) Thomas M, Boname JM, Field S, Nejentsev S, Salio M, Cerundolo V. Downregulation of NKG2D and NKp80 ligands by Kaposi's sarcoma-associated herpesvirus K5 protects against NK cell cytotoxicity. Proc Natl Acad Sci U S A 105, 1656-1661. 5-2-2008. (40) Vider-Shalit T, Fishbain V, Raffaeli S, Louzoun Y. Phase-Dependent Immune Evasion of Herpesviruses. J Virol 81, 9536-9545. 2007. (41) Wood NH, Feller L. The malignant potential of HIV-associated Kaposi sarcoma. Cancer Cell International , 1475-2867. 31-10-2008. (42) Lan K, Verma SC, Murakami M, Bajaj B, Kaul R, Robertson ES. Kaposi's sarcoma herpesvirus-encoded latency-associated nuclear antigen stabilizes intracellular activated Notch by targeting the Sel10 protein. Proc Natl Acad Sci U S A 104, 19287-16292. 9-10-2007. (43) Choi YB, Nicholas J. Autocrine and Paracrine Promotion of Cell Survival and Virus Replication by Human Herpesvirus 8 Chemokines. J Virol 83, 6501-6513. 2008. (44) Lüttichau HR. The herpesvirus 8 encoded chemokines vCCL2 (vMIP-II) and vCCL3 (vMIP-III) target the human but not the murine lymphotactin receptor. Virol J . 2008. (45) Montaner S. Akt/TSC/mTOR Activation by the KSHV G Protein-Coupled Receptor. Cell Cycle , 438-443. 15-2-2007. (46) Feng H, Dong X, Negaard A, Feng P. Kaposi's Sarcoma-Associated Herpesvirus K7 Induces Viral G Protein-Coupled Receptor Degradation and Reduces Its Tumorigenicity. PLoS Pathog . 2008. (47) Rose PR, Bogyo M, Moses AV, Früh K. Insulin-Like Growth Factor II ReceptorMediated Intracellular Retention of Cathepsin B Is Essential for Transformation of Endothelial Cells by Kaposi's Sarcoma-Associated Herpesvirus. J Virol 81, 80508062. 2007. (48) Chang PC, Li M. Kaposi's Sarcoma-Associated Herpesvirus K-Cyclin Interacts with Cdk9 and Stimulates Cdk9-Mediated Phosphorylation of p53 Tumor Suppressor. J Virol 82, 278-290. 2008. (49) Lubyova B, Kellum MJ, Frisancho JA, Pitha PM. Stimulation of c-Myc Transcriptional Activity by vIRF-3 of Kaposi Sarcoma-associated Herpesvirus. J.Biol.Chem. 282[44]. 2-11-2007. (50) Samols MA, Skalsky RL, Maldonado AM, Riva A, Lopez MC, Baker HV, et al. Identification of Cellular Genes Targeted by KSHV-Encoded MicroRNAs. PLoS Pathog 3. 2007. (51) Yu F, Harada JN, Brown HJ, Deng H, Song MJ, Wu T, et al. Systematic Identification of Cellular Signals Reactivating Kaposi Sarcoma–Associated Herpesvirus. PLoS Pathog 3. 2007. (52) Wilson SJ, Tsao EH, Web BLJ, Ye H, Dalton-Griffin L, Tsantoulas C, et al. X Box Binding Protein XBP-1s Transactivates the Kaposi´s Sarcoma-Associated herpesvirus (KSHV) ORF50 Promoter, Linking Plasma Cell Differentiation to KSHV Reactivation from Latency. J Virol 81, 13578-13586. 2007. (53) Kuang E, Tang Q, Maul GG, Zhu F. Activation of p90 Ribosomal S6 Kinase by ORF45 of Kaposi's Sarcoma-Associated Herpesvirus and Its Role in Viral Lytic Replication. J Virol 82, 1838-1850. 2008. (54) Cheng F, Weidner-Glunde M, Varjosalo M, Rainio EM, Lehtonen A, Schulz TF, et al. KSHV Reactivation from Latency Requires Pim-1 and Pim-3 Kinases to Inactivate the Latency-Associated Nuclear Antigen LANA. PLoS Pathog 5. 2009. (55) Gandy SZ, Linnstaedt SD, Muralidhar S, Cashman KA, Rosenthal LJ, Casey JL, et al. RNA Editing of the Human Herpesvirus 8 Kaposin Transcript Eliminates Its Transforming Activity and Is Induced during Lytic Replication. J Virol 81, 1354413551. 2007. (56) Orland JR WNCea. Mortality in a cohort of HTLV-I and HTLV-II infected blood donors. Transfusion , 43. 2003. (57) Poiesz B, Dube D, Dube S, Love J, Papsidero L, Uner A, et al. HTLV-II-associated cutaneous T-cell lymphoma in a patient with HIV-1 infection. N.Engl.J.Med. 342[13], 930-936. 30-3-2000. (58) Johnson JM, Harrod R, Franchini G. Molecular biology and pathogenesis of the human T-cell leukaemia/lymphotropic virus Type-1 (HTLV-1). Int.J.Exp.Pathol. 82[3], 135-147. 2001. (59) Jeang KT. Functional activities of the human T-cell leukemia virus type I Tax oncoprotein: cellular signaling through NF-kappa B. Cytokine Growth Factor Rev. 12[2-3], 207-217. 2001. (60) Gatza ML, Watt JC, Marriott SJ. Cellular transformation by the HTLV-I Tax protein, a jack-of-all-trades. Oncogene 22[33], 5141-5149. 11-8-2003. (61) Matsuoka M. Human T-cell leukemia virus type I and adult T-cell leukemia. Oncogene 22[33], 5131-5140. 11-8-2003. (62) Mortreux F, Gabet AS, Wattel E. Molecular and cellular aspects of HTLV-1 associated leukemogenesis in vivo. Leukemia 17[1], 26-38. 2003. (63) Arisawa K, Soda M, Endo S, Kurokawa K, Katamine S, Shimokawa I, et al. Evaluation of adult T-cell leukemia/lymphoma incidence and its impact on nonHodgkin lymphoma incidence in southwestern Japan. Int.J.Cancer 85[3], 319-324. 1-2-2000. (64) Hisada M, Okayama A, Spiegelman D, Mueller NE, Stuver SO. Sex-specific mortality from adult T-cell leukemia among carriers of human T-lymphotropic virus type I. Int.J.Cancer 91[4], 497-499. 15-2-2001. (65) Etoh K, Yamaguchi K, Tokudome S, Watanabe T, Okayama A, Stuver S, et al. Rapid quantification of HTLV-I provirus load: detection of monoclonal proliferation of HTLV-I-infected cells among blood donors. Int.J.Cancer 81[6], 859-864. 11-6-1999. (66) Yashiki S, Fujiyoshi T, Arima N, Osame M, Yoshinaga M, Nagata Y, et al. HLAA*26, HLA-B*4002, HLA-B*4006, and HLA-B*4801 alleles predispose to adult T cell leukemia: the limited recognition of HTLV type 1 tax peptide anchor motifs and epitopes to generate anti-HTLV type 1 tax CD8(+) cytotoxic T lymphocytes. AIDS Res.Hum.Retroviruses 17[11], 1047-1061. 20-7-2001. (67) Porto AF, Neva FA, Bittencourt H, Lisboa W, Thompson R, Alcantara L, et al. HTLV-1 decreases Th2 type of immune response in patients with strongyloidiasis. Parasite Immunol. 23[9], 503-507. 2001. (68) Satoh M, Kiyuna S, Shiroma Y, Toma H, Kokaze A, Sato Y. Predictive markers for development of strongyloidiasis in patients infected with both Strongyloides stercoralis and HTLV-1. Clin.Exp.Immunol. 133[3], 391-396. 2003. (69) Gabet AS, Mortreux F, Talarmin A, Plumelle Y, Leclercq I, Leroy A, et al. High circulating proviral load with oligoclonal expansion of HTLV-1 bearing T cells in HTLV-1 carriers with strongyloidiasis. Oncogene 19[43], 4954-4960. 12-10-2000. (70) Boyle P., Levin B. Etiology of Cancer. World Cancer Report 2008 . 2008. (71) IARC Working Group on theEvaluation of Carcinogenic Risks to Humans. Human Papillomaviruses. IARC Monographs on the Evaluationof Carcinogenic Risks to Humans 90. 2005.
 0
0
Anuncio
Documentos relacionados



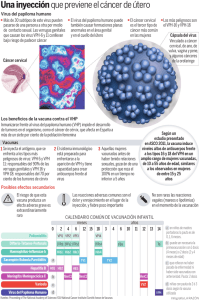




Descargar
Anuncio
Añadir este documento a la recogida (s)
Puede agregar este documento a su colección de estudio (s)
Iniciar sesión Disponible sólo para usuarios autorizadosAñadir a este documento guardado
Puede agregar este documento a su lista guardada
Iniciar sesión Disponible sólo para usuarios autorizados