Nombre: Fuentes Hernández José Gerardo Matricula
Anuncio

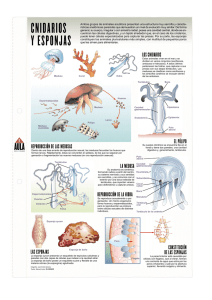
Nombre: Fuentes Hernández José Gerardo Matricula: 99221962 Teléfono: 044 (55) 18-39-11-09 Licenciatura: Hidrobiología División : Ciencias Biológicas y de la Salud Unidad: Universidad Autónoma Metropolitana Unidad Iztapalapa Trimestre lectivo 05-O Título del proyecto de investigación Proyecto de Servicio Social asociado a actividades disciplinarias realizadas por el profesor. Biología de los Organismos Planctónicos II (Zooplancton). Titulo del trabajo de Servicio Social Estudio de las medusas de Hydrozoa en la Bahía de Zihuatanejo, Guerrero, México. Asesor interno: M. en C. Carlos Álvarez Silva Lugar de realización: Laboratorio de Biología de organismos de Cuencas Hidrológicas. Anexo del edificio “S” AS – 206. Clave de registro: H.001.5 México, D.F., a 18 de noviembre de 2005. DR. A. OSCAR MONROY HERMOSILLO DIRECTOR DE LA DIVISIÓN DE CIENCIAS BIOLÓGICAS Y DE LA SALUD PRESENTE Estimado Dr. Monroy: Aprovecho la presente para saludarlo y al mismo tiempo informarle que el alumno JOSÉ GERARDO FUENTES HERNÁNDEZ, con matrícula 99221962, de la licenciatura en Hidrobiología, terminó satisfactoriamente su Servicio Social titulado “Estudio de las medusas de Hydrozoa en la Bahía de Zihuatanejo, Guerrero, México”. La fecha establecida para finalizar el Servicio Social era el 21 de julio de 2005, esto no fue posible debido a que el volumen de las muestras analizadas, rebasó las estimaciones en tiempo proyectadas. Por lo anterior, mucho le agradeceré a usted, tomar en cuenta este contratiempo en la conclusión del Servicio Social en forma extemporánea. Agradeciendo la fineza de sus acciones me pongo a sus órdenes para cualquier aclaración Atentamente “Casa abierta al tiempo” M. en C. Carlos Álvarez Silva Profesor Titular “C” de Tiempo Completo Departamento de Hidrobiología. No. Empleado: 12260 FORMATO PARA SER LLENADO POR EL ASESOR INTERNO Y EXTERNO PARA EL PROYECTO FINAL DE SERVICIO SOCIAL 1. Nombre y adscripción del asesor. M. en C. Carlos Álvarez Silva. Profesor Titular “C” de Tiempo Completo. Departamento de Hidrobiología. No. Empleado: 12260 2. La naturaleza del Proyecto del que procede el servicio social es: [ ] a) El proyecto de Servicio Social asociado a la investigación que se realiza en las áreas departamentales. [ ] Interno [ ] Externo [ ] Por convenio [ X ] b) El Proyecto de Servicio Social asociado a actividades disciplinarias realizadas por el asesor. 3. Nombre del Proyecto del que deriva el Servicio Social e institución que lo avala. El Proyecto de Servicio Social asociado a actividades disciplinarias realizadas por el asesor, relacionado con la UUEEAA Biología de los Organismos Planctónicos II (Zooplancton). 4. Justificar la vinculación del asesor con el proyecto del Servicio Social. El M. en C. Carlos Álvarez Silva, ha impartido en diversas ocasiones la materia de Biología de los Organismos Planctónicos II y de la misma manera ha dirigido diversos Servicios Sociales y Tesis de Licenciatura sobre distintos aspectos de medusas y otros organismos pláncticos. 5. Desglosar las actividades que desarrolló el asesor para favorecer el cumplimiento de los objetivos planteados en el proyecto inicial del Servicio Social • Se proporcionaron asesorías para la elaboración del proyecto de Servicio Social. • Se proporcionó información y orientación oportuna y eficiente al alumno sobre el objeto de su estudio. • Se supervisó y evaluó el cumplimiento de cada uno de los objetivos planteados. • Se dirigió y evaluó la elaboración del informe final. 6. ¿Cómo evalúa el desempeño del alumno prestador del Servicio Social? El alumno mostró un gran desempeño en el desarrollo del trabajo, no obstante que éste se prolongó más tiempo del señalado, el alumno siempre actuó con entusiasmo. 7. Anote las fortalezas y debilidades detectadas por usted con respecto a la formación del estudiante. Debido a que las últimas materias de su formación académica no estuvieron relacionadas directamente con medusas, considero que le hizo falta información en algunos temas de índole fisiológica, sin embargo esto quedó subsanado por la capacidad de trabajo y esfuerzo del estudiante. M. en C. Carlos Álvarez Silva Nombre: Fuentes Hernández José Gerardo, Matrícula: 99221962, Licenciatura: Hidrobiología. Título del Proyecto: Estudio de las medusas de Hydrozoa en la Bahía de Zihuatanejo, Guerrero, México. Registro: H.001.5, Fecha de entrega: Nombre de adscripción del asesor: M. en C. Carlos Álvarez Silva. Profesor Titular “C” de Tiempo Completo.Departamento de Hidrobiología, CBS, UAMI. No. Empleado: 12260. Resumen Durante los años de 1998 a 2000, se estudiaron las medusas de la Superclase Hydrozoa de la bahía de Zihuatanejo, Guerrero, México. Se analizaron 36 muestras de zooplancton provenientes de arrastres horizontales y que fueron recolectadas en cuatro diferentes muestreos: 6/Nov/98, 30/Mar/99, 15/Jun/99 y 25/Feb/00, utilizando una red de zooplancton que presentaba una boca de 30 cm., largo de 90 cm. y luz de malla de 250 µm. Se analizaron los parámetros fisicoquímicos (Temperatura y Salinidad) obtenidos en cada muestreo, para observar si existe o no una relación con la abundancia y distribución de las especies identificadas. Se identificaron 17 especies de medusas: Solmaris sp., Eutima spp., Rhopalonema velatum, Rhopalonema funerarium, Sarsia sp., Clytia uchidai, Liriope tetraphylla, Obelia spp., Ectopleura dumortieri, Dipurena brownei, Aglaura hemistoma, Solmundella bitentaculata, Eirene viridula, Eucheilota comata, Nausithoë spp., Hydractinia minima y Clytia simplex. El muestreo con el mayor número de especies de hidromedusas identificadas fue el realizado el 25/Feb/00 con 14 especies, seguido del realizado el 30/Mar/99 con 13 especies y el 15/Jun/99 con 9 especies. La máxima abundancia de medusas se registró en Febrero del 2000 con 557.03 org/m3, y no se presentó ninguna medusa en Noviembre de 1998. La especie más abundante fue Obelia spp., seguida de Aglaura hemistoma, Eucheilota comata y Clytia uchidai. La especie menos abundante fue Rhopalonema velatum. En este trabajo se incluye además, el listado taxonómico actualizado y la diagnosis de todas las especies identificadas, basado en Bouillon y Boero (2000). Visto Bueno: M. en C. Carlos Álvarez Silva Fuentes Hernández José Gerardo Estudio de las medusas de Hydrozoa en la Bahía de Zihuatanejo, Guerrero, México. Introducción Los organismos del Phylum Cnidaria y que pertenecen a la Superclase Hydrozoa, comúnmente conocidos como medusas, presentan una gran importancia en el medio marino, ya que son excelentes depredadores y se alimentan de cualquier organismo del zooplancton e incluso del necton (Gasca-Serrano, 1993). La importancia que tiene el estudio de la abundancia y la dinámica de las medusas, se enfoca en su gran mayoría, a que algunas especies son tóxicas, y estas toxinas provocan irritaciones en diversas partes de la piel de los bañistas, además, los compuestos y toxinas de las medusas se han estado empleando para la posible elaboración de medicamentos (Barnes y Horridge, 1965); (Tabrah et al., 1972). Por otro lado, sus mecanismos de desplazamiento son débiles, por lo que son importantes indicadores de masas de agua y corrientes; así mismo, presentan una distribución que está sujeta a restricciones definidas, relacionadas con su tolerancia a diferencias de salinidad, temperatura, densidad, productividad, etc. por lo que las especies son características de determinadas zonas (Russel, 1953). Se caracterizan por presentar simetría radial, tetrámera o polímera, y en ocasiones pude marcarse hasta simetría bilateral. Las formas y tamaños varían de especie a especie, las de tamaño grande pueden ser aplanadas o semiesféricas, las pequeñas esféricas, cuboidales o con forma de campana (Russel, 1953). Las medusas se caracterizan por presentar básicamente dos capas histológicas representadas por la epidermis y gastrodermis, ambas capas presentan diversos tipos de células. Su reproducción puede ser sexual o asexual presentando alternancia de generaciones o metagénesis; en la clase Hydrozoa la alternancia de generaciones está representada por la forma de pólipo, asexual y fija; y la forma medusa, sexual y libre nadadora. La fecundación se hace generalmente en el medio externo, aunque en algunos casos, se efectúa dentro de gónadas femeninas (Loman-Ramos, 2000). Objetivos • Analizar las muestras de zooplancton recolectadas en el periodo de 1998 al 2000 en la Bahía de Zihuatanejo, Guerrero, México. • Llevar acabo la identificación de las medusas presentes en la bahía. • Establecer la abundancia de cada una de las especies identificadas. • Identificar si su abundancia y distribución se encuentra en relación con algún parámetro ambiental como la salinidad y la temperatura. Metodología • Se analizaron 36 muestras de zooplancton provenientes de arrastres horizontales y que fueron recolectadas en cuatro diferentes muestreos: 6/Nov/98, 30/Mar/99, 15/Jun/99 y 25/Feb/00. Las muestras fueron recolectadas utilizando una red de zooplancton que presentaba una boca de 30 cm. largo de 90 cm. y luz de malla de 250 µm, los arrastres fueron aproximadamente de 5 minutos y se efectuaron de manera circular entre los primeros 60 y 90 cm. de la superficie, utilizando una lancha con motor fuera de borda. Las muestras fueron fijadas en formol 4% y neutralizadas con Borato de Sodio. • En el laboratorio se llevó acabo la separación de las medusas utilizando una caja de petri cuadriculada con la ayuda de unas pinzas de relojero; las medusas se colocaron en frascos de vidrio de tamaño pequeño con formol y glicerina al 4%, para posteriormente llevar acabo la identificación y cuantificación de las especies. Para resaltar estructuras con valor taxonómico se tiñeron con rosa de bengala. La identificación taxonómica se realizó en base a Bouillon y Boero (2000) y para la revisión de estructuras fisiológicas en medusas, se utilizaron los volúmenes de Mayer (1910). • Para obtener la abundancia (a) de cada una de las especies identificadas, fue necesario contabilizar los organismos por especie en cada una de las estaciones de cada muestra, y utilizando el volumen de agua filtrado de cada estación, se empleó la siguiente fórmula: a = # de org. / Vol. Filtrado m3. • Por otro lado, los datos obtenidos de temperatura y salinidad, se utilizaron para observar la posible relación con la presencia de algunas especies de medusas en las fechas de muestreo. Resultados • Análisis de muestras: Se identificaron 17 especies de medusas. Sólo se presentaron en 3 de los 4 muestreos analizados: 30/Mar/99, 15/Jun/99 y 25/Feb/00, debido a que en el del 6/Nov/98, no se encontró ningún organismo en las 9 estaciones analizadas. Cuatro organismos pertenecen a la Subclase Anthomedusae, seis a la Subclase Leptomedusae, cuatro a la Subclase Trachymedusae, dos a la Subclase Narcomedusae y una a la Subclase Coronatae. • Identificación de medusas: En la Fig 1. se presenta el listado taxonómico actualizado de las especies que se identificaron en este trabajo, basado en Bouillon y Boero (2000). Así mismo, las diagnosis de los ejemplares identificados. LISTADO TAXONÓMICO Phylum Cnidaria Superclase Hydrozoa Clase Hydroidomedusa Subclase Anthomedusae Familia Corynidae Dipurena brownei Sarsia sp. Familia Hydractiniidae Hydractinia minima (Antes: Podocoryne minima) Familia Tubulariidae Ectopleura dumortieri Subclase Leptomedusae Familia Campanulariidae Obelia spp. Clytia simplex (Antes: Phialidium simplex) Clytia uchidai (Antes: Phialidium uchidai) Familia Eirenidae Eirene viridula Eutima spp. Familia Loveneliidae Eucheilota comata Subclase Trachymedusae Familia Geryoniidae Liriope tetraphylla Familia Rhopalonematidae Aglaura hemistoma Rhopalonema velatum Rhopalonema funerarium Subclase Narcomedusae Familia Aeginidae Solmundella bitentaculata Familia Solmarisidae Solmaris sp. Subclase Coronatae Familia Nausithoidae Nausithoë spp HYDROMEDUSAS Subclase Anthomedusae: (Haeckel, 1879) Estas hidromedusas tienen considerables variaciones en su forma. La umbrela usualmente tiene forma de campana profunda. Las gónadas casi invariablemente están situadas en el estómago, muy raramente se extienden peradialmente en la subumbrela. Pueden tener o no ocelos y carecen de estatocistos. Los hidroides siempre son atecados. Familia Corynidae Esta familia de antomedusas posee una boca circular simple. Poseen cuatro canales radiales con las gónadas rodeando completamente el manubrio. Tienen de 2 a 4 tentáculos huecos marginales. Los ocelos se encuentran en el lado abaxial de los bulbos de los tentáculos. Dipurena brownei (Bigelow, 1909) El diámetro es de hasta 5 mm. No existe una proyección apical, sin embargo la mesoglea es gruesa. Posee gónadas en 1-5 anillos distintos. Tiene cuatro tentáculos largos suaves cerca de sus bases, el resto posee grupos de nematocistos ovales y colocados transversalmente arreglados de manera irregular. Tienen bulbos globulares con ocelos. Sarsia sp (Arai y Brinckmann-Voss, 1980) La umbrela posee una proyección apical pequeña y cónica. La umbrela tiene un diámetro que varía de 1.0 a 3.0 mm, y una altura de 2.0 a 6.0 mm; la mesoglea es muy delgada. El estómago es grande, tan largo como la cavidad umbelar, sin pedúnculo. La boca es sencilla, rodeada por un labio circular. El estómago posee una extensión apical ancha y triangular por encima del nivel de origen de los conductos radiales. Las gónadas ocupan casi toda la longitud del estomago, dejando descubierta únicamente una región basal corta. Los cuatro conductos radiales son sencillos y el conducto circular angosto. Los cuatro tentáculos marginales, en posición radial, llevan bulbos basales grandes, cada uno con un ocelo en su lado aboral. Los tentáculos cerca de su base se encuentran desnudos, pero a lo largo de toda su longitud, poseen anillos prominentes de nematocistos, y en el extremo de cada uno de los tentáculos se localiza un hinchamiento cubierto de nematocistos. Familia Hydractiniidae Boca con cuatro labios simples o ramificados y con racimos terminales de nematocistos. Canales radiales en número de cuatro. Gónadas en las paredes interradiales del estómago o en regiones próximas al canal radial. Con cuatro, ocho o más tentáculos marginales sólidos. Con o sin ocelli. Hydractinia minima (Trinci, 1903) (Antes: Podocoryne minima) La umbrela tiene forma de campana redondeada, ligeramente más alta que ancha, y por lo general con un engrosamiento apical pequeño. La mesoglea tiene un grosor regular. El velo es ancho. El estomago es cilíndrico y en forma de botella, y pende de un pedúnculo pequeño. Las gónadas se localizan en las zonas interradiales del estómago. Lleva cuatro tentáculos radiales sólidos. Los bulbos tentaculares basales son grandes y redondeados. Carece de ocelos. Familia Tubulariidae Son antomedusas con una umbrela circular simple. Poseen cuatro canales radiales y un manubrio que no se extiende mas allá del margen umbelar. Las gónadas rodean completamente el manubrio. Poseen cuatro o menos tentáculos marginales usualmente huecos. No poseen ocelos en los márgenes de los bulbos. Ectopleura dumortieri (Van Beneden, 1844) La umbrela es en forma de pera. El diámetro de la misma varía de .5 a 1.0 mm, y la altura de .5 a 1.5 mm; La mesoglea es bastante gruesa, especialmente en el poro aboral. El velo es angosto. La exumbrela posee 4 pares de líneas longitudinales de nematocistos, extendiéndose desde la base de los bulbos tentaculares hasta el ápice de la umbrela, donde todos convergen en un punto. El estómago es fusiforme y se extiende hasta el nivel de la apertura velar. No posee pedúnculo. La boca es sencilla en forma de tubo, y se encuentra armada de nematocistos. La boca se encuentra rodeando completamente al estómago dejando libre únicamente la boca. Los 4 conductos radiales y el conducto circular son angostos. Los cuatro tentáculos marginales radiales son cortos y con bulbos grandes. La superficie externa de los tentáculos es gruesa y cubierta de nematocistos, sus extremos distales se encuentran enrollados a manera de hélices. Subclase Leptomedusae (Haeckel, 1865) Son hidromedusas con la umbrela usualmente en forma hemisférica o aplanada. Las gónadas están situadas en los canales radiales. Poseen órganos sensoriales marginales, y cuando están presentes, se encuentran en forma de cordilios o vesículas marginales de origen ectodérmico. Ocasionalmente poseen ocelos. Familia Campanulariidae Son leptomedusas con un velo normal o reducido. Tienen un estómago pequeño carente de pedúnculo. Normalmente tienen cuatro canales radiales simples. Las gónadas rodean completamente a estos canales y están separadas del estómago. Los tentáculos son marginales y de naturaleza hueca, raramente sólidos. Carecen de poros excretores y cilios marginales o laterales. Tienen vesículas marginales cerradas y carecen de ocelos. Género Obelia: Obelia spp. (Péron y Lesueur, 1809) Son Campanulariidae con el velo reducido, con tentáculos marginales sólidos y con ocho 0litocistos. Obelia spp. tiene hasta 6 mm de ancho, aplanada y de mesoglea delgada. El velo es rudimentario, su estómago es corto y de base cuadrangular, la boca tiene cuatro labios cortos y simples. Posee gónadas redondas y de forma de saco situadas en medio de los canales radiales. Posee numerosos tentáculos cortos y sólidos de alguna manera rígidos y con un núcleo axial de una sola fila de células endodérmicas. Cada tentáculo tiene un pequeño bulbo basal y una corta prolongación de endodermo hacia la mesoglea del margen umbelar. Posee ocho litocistos radiales cada uno situado en el costado inferior del bulbo basal del tentáculo marginal, cada uno conteniendo una concreción. Clytia simplex (Browne, 1902) (Antes: Phialidium simplex) Alcanza hasta los 22 mm de ancho y tiene 10 mm de alto. El diámetro de la umbrela varía de 2.5 a 7.0 mm de largo. La mesoglea es de grosor regular. Es de forma de vidrio de reloj. Su estómago es corto con cuatro grandes labios fimbrados. Las gónadas están situados distalmente de un medio a tres cuartos a lo largo de los canales radiales los cuales son lineares y ligeramente doblados. Posee de 60 a 85 tentáculos con bulbos globulares basales y algunos bulbos jóvenes. Existe un estatocisto entre los tentáculos, estos últimos con una concreción cada uno. Se han visto especímenes con cuatro y hasta seis canales radiales. (Kramp, 1961). Clytia uchidai (Kramp, 1961) (Antes: Phialidium uchidai) Umbrela en forma de cúpula de escasa altura. El diámetro de la umbrela varía de 3 a 10 mm; la mesoglea es delgada. El estómago es pequeño, cuadrangular. Los cuatro conductos radiales, y el conducto circular son estrechos. Las gónadas se localizan sobre los cuatro conductos radiales, en las cercanías del conducto circular, y son de forma oval o alargada. Lleva de 16 a 28 tentáculos marginales pequeños. Posee de 24 a 42 estatocistos dispuestos irregularmente. Familia Eirenidae Son leptomedusas con un estómago pequeño y con un pedúnculo gástrico. Poseen cuatro ó seis canales radiales simples. Las gónadas se encuentran en los canales radiales y están restringidas a la subumbrela. Los tentáculos son huecos y marginales. Pueden tener o no poros excretores y de manera similar pueden tener o no tener cirrios laterales o marginales. Tienen un gran e indefinido número de vesículas marginales cerradas y carecen de ocelos. Los hidroides cuando son conocidos, son hidrantes con redes entre los tentáculos, la hidroteca posee un opérculo. Eirene viridula (Péron y Lesueur, 1809) Eirenidae con distintos pedúnculos gástricos; carece de cilios marginales o laterales, con o sin poros excretores; de 4 a 6 canales radiales; gónadas en la parte subumbelar de los canales radiales, no extendidas sobre el pedúnculo gástrico; numerosos estatocistos. Eutima spp. (Mc Crady, 1859) Eirenidae con distintos pedúnculos gástricos; con cirrios laterales (difícilmente observables, ya que frecuentemente se destruyen después de la fijación); con verrugas marginales hinchadas; sistema excretor externo; 4 canales radiales simples; gónadas en canales radiales. Familia Lovenellidae Leptomedusas con estómago pequeño. Carecen de pedúnculo y poseen cuatro canales radiales simples. Las gónadas se encuentran situadas en los canales radiales y están separadas del estómago. Los tentáculos son marginales y huecos. Carecen de poros excretores y poseen cirrios laterales o marginales. Poseen vesículas marginales cerradas y carecen de ocelos. Los hidroides cuando son conocidos, poseen una hidroteca con un opérculo. Eucheilota comata (Bigelow, 1909) La umbrela es más ancha que alta. El diámetro de la umbrela varía de 6.0 a 12.0 mm y la mesoglea es gruesa. Posee un estómago corto en forma de matraz. La boca posee 4 labios lobulados. En los ejemplares de mayor tamaño, se distingue un pedúnculo pequeño. Los cuatro conductos radiales y el conducto circular son estrechos. Las gónadas aparecen como pequeñas protuberancias globulares en la región media de los conductos radiales, y con el crecimiento de la medusa se alargan avanzando hacia el conducto circular, localizándose finalmente en la mitad distal de los conductos radiales. Lleva 17 tentáculos marginales y 23 bulbos rudimentarios. Los tentáculos marginales son pequeños, y se encuentran flaqueados por 1 a 3 pares de cirros laterales. Los cirros también se localizan a los lados de los bulbos rudimentarios más grandes, sin embargo, los bulbos pequeños carecen de ellos. Los bulbos tentaculares basales son prominentes. Los estatocistos varían de 7 a 14, son grandes y se encuentran distribuidos irregularmente. Subclase Trachymedusae (Haeckel, 1865) Hidromedusas con un margen umbelar completo y no dividido en lóbulos. Tienen un anillo de nematocistos marginal engrosado. Poseen canales radiales y gónadas usualmente confinadas a los canales radiales Los tentáculos son sólidos y marginales o pueden tener tentáculos tanto sólidos como huecos situados en el margen de la umbrela. Tienen clubs sensoriales con un eje ectodérmico el cual puede ser libre o confinado. FamiliaGeryoniidae Son traquimedusas con un estómago con pedúnculo. Tiene cuatro o seis canales radiales, además de canales centrípetos. Las gónadas son planas y de forma de hoja y están situadas en los canales radiales. Posee tentáculos marginales de dos tipos huecos y sólidos, además de clubs sensoriales marginales envueltos en la mesoglea. Liriope tetraphyla (Chamisso y Eysenhardt, 1821) La umbrela es hemisférica. El diámetro de la umbrela varía de .5 a 14.0 mm. En los ejemplares grandes la mesoglea gruesa. El velo es ancho. El estómago es pequeño, y se encuentra unido a un pedúnculo largo y de tamaño variable. En el pedúnculo se distinguen 4 bandas de músculos longitudinales. La boca posee 4 labios pequeños. Los cuatro conductos radiales y el conducto circular son anchos, además posee de 1 a 3 (o más) conductos centripetales ciegos, los cuales se localizan en cada uno de los cuadrantes. Las gónadas se localizan sobre los cuatro conductos radiales y observan una gran variedad de tamaños y formas, desde oval hasta circular y en forma de corazón. Lleva cuatro tentáculos radiales largos, huecos, con anillos de nematocistos en toda su longitud, y cuatro tentáculos interradiales pequeños, sólidos con protuberancias cubiertas con nematocistos en posición adaxial. Son ocho el número de estatocistos que poseen, y se localizan cerca de la base de los tentáculos. Familia Rhopalonematidae Son traquimedusas con un estómago estrecho que puede tener o no un pedúnculo. Usualmente tienen ocho canales radiales, raramente más y carecen de canales centripetales. Poseen gónadas situadas en los canales radiales. Los tentáculos marginales están distribuidos uniformemente, algunas veces se presentan en dos tipos y cada tentáculo es de forma estructural uniforme en ambos tipos. Los bulbos sensoriales marginales son libres y raramente encubiertos. Aglaura hemistoma (Péron y Lesueur, 1809) La umbrela tiene forma de campana, con el ápice plano. El diámetro de la umbrela varía de .5 a 4.0 mm, y la altura de .5 a 5.0 mm; La mesoglea es delgada, pero rígida. El velo es ancho. El estómago es pequeño y se encuentra unido a un pedúnculo cónico generalmente de longitud igual al radio de la umbrela. La boca posee cuatro labios sencillos y pequeños. El número de gónadas es de 8, en forma de salchicha y se encuentran localizadas sobre el pedúnculo, en el punto de unión de los 8 conductos radiales con el estómago. Los ocho conductos radiales son rectos y anchos, el conducto circular también es ancho. Aparecen de 48 a 85 tentáculos marginales sólidos y quebradizos, los cuales por lo regular completos se encuentran rotos, sin embargo cuando permanecen llegan a medir una longitud equivalente a 3/4 del diámetro de la umbrela y sus extremos distales poseen forma de bastón. Los estatocistos, en número de 8, se encuentran colocados entre los conductos radiales. Rhopalonema velatum (Gegenbaur, 1856) Umbrela en forma hemisférica; la mesoglea es delgada. El velo es muy ancho. El estómago es angosto y elongado, con base octagonal y contráctil, extendiéndose algunas veces hasta las cercanías del margen umbrelar. La boca posee 4 labios pequeños. Los 8 conductos radiales (4 radiales y 4 interradiales), y el conducto circular son estrechos. Posee 8 gónadas alargadas u ovales que se extienden a lo largo del tercio medio de los conductos radiales. Con 32 tentáculos marginales. Todos los tentáculos son quebradizos y regularmente se pierden en las maniobras de arrastre. Rhopalonema funerarium (Vanhöffen, 1902) Umbrela más o menos cónica, sin proyección apical; la mesoglea tiene un grosor regular. El velo es ancho. El estómago es pequeño. La boca posee labios pequeños. Posee 9 gónadas en forma de huso, elongadas, las cuales se extienden a lo largo de los extremos distales de los conductos radiales. Lleva 8 tentáculos marginales radiales, 8 tentáculos interradiales, y 16 tentáculos muy pequeños en posición adradial, éstos últimos semejan cirrios, y cada uno termina en una protuberancia globular. Los estatocistos aparecen en número de 32, y se localizan en la mitad de los espacios entre tentáculos y “cirrios”. Subclase Narcomedusae (Haeckel, 1879) Son hidromedusas con los lados de la umbrela dividida por surcos peroniales por el margen umbelar puede estar lobulado. Tienen un estómago amplio que abarca toda la zona periférica circular ó pueden tener bolsas periféricas. Carecen de canales radiales, pero pueden tener o no un sistema de canal periférico. Las gónadas están en las paredes del estómago. Los tentáculos son sólidos y marginales y permiten cierta distancia a la umbrela sobre el margen, algunas veces tienen pequeños tentáculos secundarios en el margen mismo. Los órganos sensoriales liberan a los clubes sensoriales del eje ectodérmico. Familia Aeginidae Narcomedusas con bolsas estomacales divididas y situadas interradialmente que contienen a las gónadas. Pueden tener o no un sistema de canal periférico. Los tentáculos primarios perradiales dejan a la umbrela entre las bolsas marginales, estas bolsas se extienden más allá de los puntos de origen de los tentáculos primarios. Pueden tener o no tentáculos secundarios en el margen de la umbrela. Solumndella bitentaculata (Quoy y Gaimard, 1833) La umbrela es hemisférica. El diámetro de la umbrela varía de .5 a 6.0 mm: la mesoglea es gruesa, especialmente en la región apical. El velo es ancho, con músculos circulares y potentes. El estómago es ancho y lenticular, con ocho bolsas gástricas, 2 entre cada par sucesivo de bandas perionales. Las bolsas gástricas son por lo general de forma rectangular, con los ángulos redondeados. Las gónadas se desarrollan en la pared ectodermal de la subumbrela, por debajo de las bolsas gástricas y en medusas de gran tamaño, adquieren la forma de un anillo que se localiza cerca de la periferia del estómago central. Lleva dos tentáculos opuestos muy largos, los cuales se proyectan desde la umbrela, cerca del ápice. Posee además dos surcos perionales profundos que se extienden a lo largo de la exumbrela, desde el margen velar hasta la base de cada uno de los tentáculos. Son 32 el número de estatocistos, 8 en cada cuadrante, aunque por lo general solo aparecen de 8 a 16. Familia Solmarisidae Sin bolsillos gástricos. Los productos sexuales se desarrollan como engrosamientos o divertículos en la pared central del estómago. Con o sin sistema canal periférico. Con o sin otoporpa. Género Solmaris: Solmaris sp. (Haeckel, 1879) Umbrela en forma de disco. La mesoglea es muy delgada. El velo es ancho. El estómago es plano. Las gónadas se encuentran organizadas en forma anular. Posee 30 lóbulos marginales de tamaño variable, cada uno de ellos con 1 a 2 estatocistos. Tiene 30 tentáculos marginales, la mayoría incompletos. Carece de sistema de conductos periféricos y otoporpa. Subclase Coronatae Familia Nausithoidae Género Nausithoë: Nausithöe spp. (Kölliker, 1853) La umbrela es de forma discoidal; la mesoglea es gruesa. Carece de velo. El disco central de la exumbrela es grueso, forma como un casquete lenticular con la superficie finamente punteada de gránulos, y se encuentra separado de la zona de pedalia periférica, mediante un surco anular. La zona de pedalia está compuesta por 16 engrosamientos radiales, 8 en los radios de los tentáculos, y 8 en los radios de los órganos sensoriales. Lleva 8 tentáculos marginales en posición adradial, los cuales se originan en fisuras profundas entre los lóbulos marginales. El estómago es central, y está conectado con un seno amplio. Las bolsas gástricas periféricas, se localizan en los radios de los órganos sensoriales y tentáculos, y se encuentran unidas por el conducto circular marginal en la línea axial de cada uno de los lóbulos marginales. La boca es sencilla, cruciforme y con 4 labios. Las gónadas son en número de 8, grandes y globulares, y se localizan en los 8 radios tentaculares, sobre el piso de la subumbrela en la zona del seno circular interno. • Abundancia: Para obtener la abundancia (a = # de org. / Vol. Filtrado m3) de los organismos de cada una de las fechas de muestreo, fue necesario en primer término, identificar y contabilizar cuántos organismos por especie estaban presentes en cada una de las estaciones de cada muestra, como se observa en las tablas 1 a 3. Fecha de muestreo: 30-Marzo-99 # de Estación 1 Especie Solmaris sp. 19 Eutima spp. 1 Rhopalonema velatum Rhopalonema funerarium Sarsia sp. Clytia uchidai Liriope tetraphylla Obelia spp. Ectopleura dumortieri Dipurena brownei Aglaura hemistoma Solmundella bitentaculata Eirene viridula Total por estación Tabla 1 # de organismos por estación 2 3 4 5 6 7 8 9 3 1 1 4 1 3 1 1 1 2 2 1 8 13 4 3 6 7 2 18 1 2 6 2 4 1 5 1 2 10 10 1 13 110 3 4 1 7 1 5 1 5 1 4 5 1 2 2 2 3 1 20 20 26 37 47 131 14 12 11 Total por especie 39 3 11 8 4 45 34 148 4 3 14 2 3 318 Total medusas Fecha de muestreo: 15-Junio-99 # de Estación Especie Solmaris sp. Eutima spp. Sarsia sp. Clytia uchidai Liriope tetraphylla Obelia spp. Aglaura hemistoma Eucheilota comata Nausithoë spp 1 # de organismos por estación 2 3 4 5 6 7 2 1 1 1 3 71 38 64 745 1 1 5 Total por estación 77 38 70 748 8 9 X 1 X 1 1 X 2 4 X 5 1 X 1 449 53 X 1889 4 4 X X X 8 Total por especie 3 4 1 14 2 3309 10 5 8 457 62 0 1895 9 3356 Total medusas Tabla 2 Fecha de muestreo: 25-Febrero-2000 # de Estación Especie Solmaris sp. Eutima spp. Rhopalonema velatum Sarsia sp. Clytia uchidai Clytia simplex Liriope tetraphylla Obelia spp. Dipurena brownei Aglaura hemistoma Solmundella bitentaculata Eirene viridula Eucheilota comata Hydractinia minima Total por estación Tabla 3 1 # de organismos por estación 2 3 4 5 6 7 1 5 1 1 16 8 9 1 7 2 7 3 4 35 1 1 2 14 28 4 5 15 2 1 30 5 3 3 2 2 4 16 95 181 99 185 481 179 15 14 35 1 3 6 51 6 7 1 5 1249 5 2 2 18 1859 4 1 73 34 2 25 105 628 664 220 157 17 43 9 8 2 2 4 Total por especie 8 39 125 711 848 452 278 211 545 198 2 74 12 3407 Total Medusas Posteriormente se utilizaron los parámetros fisicoquímicos obtenidos en cada muestreo, debido a que el volumen filtrado en cada estación es un dato indispensable para llevar acabo el cálculo de la abundancia. Por otro lado, los datos obtenidos de temperatura y salinidad, se utilizaron para observar la posible relación con la presencia de algunas especies de medusas en las fechas de muestreo. Estos datos se observan en las tablas 4 a 6 y gráficas 4 y 5. BAHÍA DE ZIHUATANEJO, GRO. 30 DE MARZO DE 1999 ESTACIÓN HORA T°C S VOL. FILTRADO UPS m3 1 10:08 25.0 30 7.75 2 10:35 25.0 32 8.49 3 10:48 25.0 32 6.37 4 11:06 25.0 32 3.99 5 11:23 26.0 32 6.36 6 11:40 26.0 32 6.45 7 11:55 26.0 32 6.56 8 12:15 26.0 33 5.33 9 12:35 27.0 32 9.47 Tabla 4 BAHÍA DE ZIHUATANEJO, GRO. ESTACIÓN HORA 1 2 3 4 5 6 7 8 9 Tabla 5 09:00 09:16 09:28 09:39 10:00 10:12 10:25 10:36 10:47 15 T°C 30.0 30.0 30.0 30.0 29.0 29.0 29.0 29.0 28.0 DE S UPS 34 32 28 32 30 29 30 30 30 JUNIO DE 1999 VOL. FILTRADO m3 14.14 16.27 13.57 8.84 12.88 17.82 14.93 15.80 11.39 BAHÍA DE ZIHUATANEJO, GRO. 25 DE FEBRERO DE 2000 ESTACIÓN HORA T°C S VOL. FILTRADO UPS m3 1 09:05 28.0 29 10.71 2 09:25 27.0 28 5.96 3 09:40 27.0 29 5.83 4 09:48 27.0 28 5.00 5 10:00 27.0 30 8.18 6 10:10 26.0 30 3.86 7 10:22 27.0 32 4.60 8 10:40 26.0 32 24.91 9 11:07 26.0 32 4.32 Tabla 6 Ya con los datos necesarios, y con la fórmula antes mencionada, se obtuvo la abundancia # de org / m3 de cada una de las especies identificadas en cada una de las estaciones, y con ello, la abundancia total de cada especie en las diferentes estaciones (tablas 7 a 9), observándose que Obelia spp. fue la especie que predominó con un número mayor de organismos por m3 en las fechas de muestreo, presentándose con mayor número en el muestreo realizado el 25/Feb/00 con una abundancia total por especie de 333.46 org/m3; seguido del muestreo realizado el 15/Jun/99 con 253.74 org/m3 y del realizado el 30/Mar/99 con 24.61 org/m3. Existieron diversas especies que presentaron una abundancia menor con respecto a Obelia spp., las cual varió de acuerdo a los diferentes muestreos; por mencionar algunas: 30/Mar/99 Clytia uchidai 7.11 org/m3, Liriope tetraphylla 5.97 org/m3, Solmaris sp. 5.73 org/m3, Solmundella bitentaculata 0.31 org/m3, destacando que esta última especie fue la que menor presencia tuvo en este muestreo. tabla 7, gráfica 1; 15/Jun/99 Clytia uchidai 0.92 org/m3, Aglaura hemistoma 0.72 org/m3, Sarsia sp. 0.06 org/m3, observándose que Sarsia sp. fue la especie que presentó menor abundancia total comparada con las diferentes especies identificadas en este muestreo. Paralelamente, se puede observar que los datos de abundancia total por especie obtenidos en esta fecha no varían demasiado (excepto Obelia spp.), es decir, las especies identificadas presentaron un número de organismos por m3 muy bajo en comparación con la mayoría de las especies identificadas en los otros dos muestreos, en los cuales se presentan rangos de datos más altos y diferenciados. tabla 8, gráfica 2; 25/Feb/00 Aglaura hemistoma 171.99 org/m3, Eucheilota comata 12.42 org/m3, Clytia uchidai 12.12 org/m3 y la especie con menor abundancia total en todos los muestreos Rhopalonema velatum con 0.04 org/m3. tabla 9, gráfica 3. La abundancia total de todas las especies en los 3 diferentes muestreos fue diferente, presentándose la mayor en Febrero del 2000 con 557.03 org/m3, seguido de 257.24 org/m3 en Junio de 1999 y 51.54 org/m3 en Marzo de 1999. La abundancia total de las diferentes especies por estación se realizó para resaltar en que estación de cada muestreo se presentó un mayor número de organismos. 30 de Marzo de 1999 Abun. Total por ABUNDANCIA #Org/m3 1 2 3 4 5 6 7 8 9 especie Solmaris sp. 2.45 0.35 0.63 0.75 0.94 0.31 0.00 0.19 0.11 5.73 Eutima spp. 0.13 0.00 0.16 0.25 0.00 0.00 0.00 0.00 0.00 0.54 Rhopalonema velatum 0.00 0.12 0.00 0.25 0.63 0.16 0.00 0.00 0.42 1.57 Rhopalonema funerarium 0.00 0.00 0.00 0.50 0.79 0.00 0.00 0.19 0.00 1.48 Sarsia sp. 0.00 0.24 0.00 0.25 0.16 0.00 0.00 0.00 0.00 0.64 Clytia uchidai 0.00 0.94 2.04 1.00 0.31 1.55 1.07 0.19 0.00 7.11 Liriope tetraphylla 0.00 0.35 0.94 1.75 1.57 0.16 0.15 0.94 0.11 5.97 Obelia spp. 0.00 0.24 0.00 4.51 2.04 17.05 0.76 0.00 0.00 24.61 Ectopleura dumortieri 0.00 0.12 0.00 0.00 0.00 0.47 0.00 0.00 0.00 0.58 Dipurena brownei 0.00 0.00 0.31 0.00 0.00 0.00 0.15 0.00 0.00 0.47 Aglaura hemistoma 0.00 0.00 0.00 0.00 0.31 0.47 0.00 0.75 0.53 2.06 Solmundella bitentaculata 0.00 0.00 0.00 0.00 0.31 0.00 0.00 0.00 0.00 0.31 Eirene viridula 0.00 0.00 0.00 0.00 0.31 0.16 0.00 0.00 0.00 0.47 2.58 2.36 4.08 9.27 7.39 20.31 2.13 2.25 1.16 51.54 Total por estación Tabla 7 Abun. total Abundancia total por especie 30/Mar/99 30.00 Eirene viridula Solmundella bitentaculata Aglaura hemistoma Dipurena brownei Liriope tetraphylla Clytia uchidai Sarsia sp. 0.00 Eutima spp. 5.00 Solmaris sp. 10.00 Rhopalonema funerarium 15.00 Rhopalonema velatum org/m3 20.00 Ectopleura dumortieri Obelia spp. 25.00 Gráfica 1 15 de Junio de 1999 Abun. Total por ABUNDANCIA #Org/m3 1 2 3 4 5 6 7 8 9 especie Solmaris sp. 0.00 0.00 0.15 0.11 0.00 0.00 0.00 0.00 0.00 0.26 Eutima spp. 0.00 0.00 0.07 0.11 0.08 0.00 0.00 0.06 0.00 0.33 Sarsia sp. 0.00 0.00 0.00 0.00 0.00 0.06 0.00 0.00 0.00 0.06 Clytia uchidai 0.00 0.00 0.22 0.00 0.16 0.22 0.00 0.32 0.00 0.92 Liriope tetraphylla 0.00 0.00 0.00 0.00 0.08 0.00 0.00 0.00 0.09 0.17 Obelia spp. 5.02 2.34 4.72 84.28 34.86 2.97 0.00 119.56 0.00 253.74 Aglaura hemistoma 0.07 0.00 0.00 0.11 0.31 0.22 0.00 0.00 0.00 0.72 Eucheilota comata 0.35 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.35 Nausithoë spp 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.70 0.70 5.45 2.34 5.16 84.62 35.48 3.48 0.79 257.24 Total por estación Tabla 8 0.00 119.94 Abun. total Abundancia total por especie 15/Jun/99 Obelia spp. 300.00 250.00 Nausithoë spp Eucheilota comata Clytia uchidai 0.00 Sarsia sp. 50.00 Eutima spp. Solmaris sp. 100.00 Aglaura hemistoma 150.00 Liriope tetraphylla org/m3 200.00 Gráfica 2 25 de Febrero de 2000 Abun. Total por ABUNDANCIA #Org/m3 3 4 5 6 7 8 9 especie Solmaris sp. 0.09 0.00 1 2 0.86 0.20 0.00 0.00 0.00 0.00 0.23 1.38 Eutima spp. 0.00 0.00 0.17 3.20 0.86 0.52 1.52 0.08 0.00 6.35 Rhopalonema velatum 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.04 0.00 0.04 Sarsia sp. 0.00 0.00 0.00 0.40 0.00 0.00 0.00 0.00 0.00 0.40 Clytia uchidai 0.00 0.67 0.51 2.80 3.42 1.04 0.00 0.20 3.47 12.12 Clytia simplex 0.37 0.00 0.00 6.00 0.00 0.00 0.00 0.00 0.00 6.37 Liriope tetraphylla 0.00 0.00 0.86 0.60 0.24 0.78 0.43 0.08 0.23 3.23 Obelia spp. 2.33 17.62 107.72 132.80 26.89 40.67 3.70 1.73 0.00 333.46 Dipurena brownei 0.00 0.67 0.34 3.00 1.71 0.00 0.00 5.73 Aglaura hemistoma 0.00 0.00 0.84 0.67 2.74 19.00 22.13 25.65 40.22 19.31 41.44 Solmundella bitentaculata 0.00 0.00 0.00 0.20 0.00 0.26 0.00 0.12 0.00 Eirene viridula 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.46 0.46 Eucheilota comata 0.00 0.34 8.75 1.20 0.00 1.81 0.00 0.32 0.00 12.42 Hydractinia minima 0.00 1.01 0.00 0.20 0.00 1.30 0.00 0.00 0.00 2.50 Total por estación Tabla 9 3.64 20.97 121.96 169.60 55.26 72.02 45.87 21.88 45.83 171.99 0.58 557.03 Abun. Total Dipurena brownei Liriope tetraphylla 0.00 Clytia simplex 50.00 Clytia uchidai 100.00 Eutima spp. 150.00 Sarsia sp. 200.00 Rhopalonema velatum 250.00 Solmaris sp. org/m3 300.00 Hydractinia minima 350.00 Eucheilota comata Obelia spp. 400.00 Aglaura hemistoma Solmundella bitentaculata Eirene viridula Abundancia total por especie 25/Feb/00 Gráfica 3 No todas las especies de medusas identificadas estuvieron presentes en todos los muestreos, como es el caso de Rhopalonema velatum, Dipurena brownei, Solmundella bitentaculata, Eirene viridula y Eucheilota comata, que solo se presentaron en dos fechas. Y del mismo modo, se identificaron otras especies que solo aparecieron en una sola fecha, como el caso de Ectopleura dumortieri el 30/Mar/99, Nausithoë spp. el 15/Jun/99, Hydractinia minima y Clytia simplex que se presentaron solo el 25/Feb/00, como se observa en la tabla 10. 30 de Marzo de 1999 Solmaris sp. Eutima spp. Rhopalonema velatum Rhopalonema funerarium Sarsia sp. Clytia uchidai Liriope tetraphylla Obelia spp. Ectopleura dumortieri Dipurena brownei Aglaura hemistoma Solmundella bitentaculata Eirene viridula 15 de Junio de 1999 25 de Febrero de 2000 Solmaris sp. Solmaris sp. Eutima spp. Eutima spp. Rhopalonema velatum Sarsia sp. Clytia uchidai Liriope tetraphylla Obelia spp. Aglaura hemistoma Eucheilota comata Nausithoë spp Sarsia sp. Clytia uchidai Liriope tetraphylla Obelia spp. Dipurena brownei Aglaura hemistoma Solmundella bitentaculata Eirene viridula Eucheilota comata Hydractinia minima Clytia simplex Tabla 10 • Relación de la abundancia y distribución de organismos con salinidad y temperatura: Se graficaron los valores de temperatura y salinidad de los diferentes muestreos (gráficas 4 y 5), observando que el 30/Mar/99 se registraron los valores más bajos de temperatura, pero la salinidad no vario demasiado, de 25 a 27 UPS, por lo que probablemente, la temperatura influye en la distribución de las especies que se identificaron en esta fecha, ya que fue este muestreo el que presento menor abundancia total de especies, aunque, sin descartar que Obelia spp. domino como en todos los demás muestreos, pero se presentó en menor número con respecto a Jun/99 y Feb/00. El 15/Jun/99 se registraron valores de salinidad más variados, de 28 a 34 UPS; sin embargo, los de temperatura fueron los más altos de los muestreos, esto debido seguramente a que el muestreo se realizó en verano. Probablemente estos valores tan variados de salinidad en las diferentes estaciones o las “altas” temperaturas registradas, impidieron que otras especies identificadas en este muestreo, presentaran gran abundancia como la que mostró Obelia spp., ya que ésta, fue la única representativa de esta fecha. Los valores registrados el 25/Feb/00 de temperatura: 26 a 28° C y de salinidad 28 a 32 UPS, aparentemente, comparados con los otros muestreos, son los rangos en los cuales se propagan mejor las medusas, ya que en esta fecha fue donde se registro una mayor abundancia de todas las especies identificadas, resaltando además la especie Aglaura hemistoma, que fue una de las que registro igualmente una gran abundancia. T° C Temperatura de muestreos 31.0 30.0 29.0 28.0 27.0 26.0 25.0 24.0 23.0 22.0 1 2 3 4 5 Estación 30-Mar-99 6 7 15-Jun-99 8 9 25-Feb-00 Gráfica 4 S a l i n i d a d U P S d e m u e s t r e o s 4 0 3 5 3 0 UPS 2 5 2 0 1 5 1 0 5 0 1 2 3 0 - M Gráfica 5 3 a r - 9 9 4 E s t a c i ó 5 n 1 5 - J u n - 9 9 6 7 8 2 5 - F e b - 0 0 9 Discusión y Conclusiones No todas las especies de medusas identificadas estuvieron presentes en todos los muestreos, 7 especies se presentaron en las tres fechas, 5 en dos de los muestreos y 5 en solo uno de los muestreos. En el muestreo realizado el 6/Nov/98, no se tomaron en cuenta los valores de los parámetros fisicoquímicos, debido a que cuando se llevo acabo la recopilación de estos datos, se presentó una abundante lluvia, por lo que los valores de salinidad y temperatura variaron constantemente al momento de tomar las lecturas. Con respecto a que no se encontró ningún organismo en las 9 estaciones analizadas, probablemente, el hecho de que estaba lloviendo demasiado, influyó en que los organismos se refugiaran en el fondo o en otros sitios, ya que por lo que se observó en el laboratorio, se percibieron pocos organismos que no fueran medusas (zooplancton en general) y que regularmente estuvieron presentes en los otros muestreos, capturados al momento del arrastre, siguiendo lo establecido por Alvariño (1972). Para obtener mejores resultados, es necesario tomar en cuenta las corrientes de agua presentes en la zona de estudio al momento del muestreo, así como la productividad de las masas de agua, esto para percibir las variantes de salinidad y temperatura, para mencionar con mayor precisión si los datos están o no en rangos normales. Diversos autores han utilizado estos parámetros para reforzar su análisis del área de estudio (Alvariño, 1967; Fernández-Álvarez, 1981; SeguraPuertas, 1984; Loman-Ramos, 2000). Uno de los principales problemas que se presentaron al momento de identificar las especies, tuvieron que ver con el poco conocimiento que se goza de los diferentes estadios de desarrollo de las diferentes especies de medusas, así como la poca bibliografía ilustrada que existe sobre este tema. Al mismo tiempo, el análisis de muestras con varios años de antigüedad, es un poco mas complejo, debido a que algunos organismos pierden parte de sus estructuras, por lo que son difíciles de identificar. El muestreo con el mayor número de especies de hidromedusas identificadas fue el realizado el 25/Feb/00 con 14 especies, seguido del realizado el 30/Mar/99 con 13 especies y el 15/Jun/99 con 9 especies; al mismo tiempo, las mayores abundancias por especie se registraron en el mismo orden respectivamente, sin embargo, a pesar de que el 30/Mar/99 registró mayor abundancia en las especies identificadas con respecto al 15/Jun/99, esta ultima fecha registró una abundancia total mayor, ya que la especie Obelia spp. dominó por mucho y en forma más marcada en esta fecha que en los otros muestreos, con respecto a otras especies. Se puede afirmar conforme a los datos obtenidos, que la especie dominante en la bahía es Obelia spp., ya que además de ser la medusa más cuantiosa en todos los muestreos, es la que presento mayor abundancia total: 25/Feb/00 con 333.46 org/m3, 15/Jun/99 con 253.74 org/m3 y 30/Mar/99 con 24.61 org/m3. Aglaura hemistoma, Eucheilota comata y Clytia uchidai además de ser la segunda, tercera y cuarta en abundancia total respectivamente, fueron unas de las mas frecuentes al presentarse en los tres muestreos, distribuyéndose de manera semejante, aparentemente, a lo largo del año; con excepción de la segunda, que solo se presentó en dos de estos. Por otro lado, la distribución y las variantes en los rangos de especies en el océano y en aguas costeras, pueden seguir un patrón previamente establecido por otros grupos del plancton que son depredadores de medusas. Los factores de temperatura y salinidad fueron importantes para la distribución de las medusas, observándose una mayor afinidad de estos grupos en aguas de temperaturas de medias a bajas al igual que de salinidad. Algunas especies son más tolerantes a las variaciones en temperatura y salinidad, sin embargo, esto varía significativamente de especie a especie. Se comparten estos resultados con los obtenidos por otros autores, en donde se concluye que la distribución de estos organismos, probablemente esta asociada con la entrada o salida de masas de agua o corrientes a la bahía, ya que con una masa de agua muy particular, puede variar significativamente la productividad, por lo que se recomienda para posteriores estudios tomarla en cuenta (Loman-Ramos, 2000; Russel, 1953). Así mismo, se recomienda analizar las muestras en el laboratorio en un tiempo corto, después de haber realizado el muestreo, esto para que al momento de identificar los organismos, no se pierdan estructuras esenciales y sea más fácil su identificación. Por último, se invita a consultar constantemente reportes científicos para actualización de la taxonomía, y de igual manera, para tener un mayor conocimiento de los diferentes estadios de desarrollo de las medusas. Criterios de evaluación 1. Presentación de avances en seminarios del área de investigación. 2. El trabajo se entregó de forma escrita y en los términos establecidos por el Consejo Divisional. 3. Se efectuó una exposición oral apoyada por diapositivas tomadas directamente de los ejemplares. Bibliografía Alvariño, A. 1967. Bathymetric distribution of Chaetognatha, Siphonophorae, Medusae and Ctenophorae off San Diego, California. Pacific Science., 21 (4): 474385. Alvariño, A. 1972. Zooplancton del Caribe, Golfo de México y regiones adyacentes del Pacífico. IV Congreso Nacional de Oceanografía México, 223-247. Arai, M. N. & Brinckmann-Voss A. 1980a – Hydromedusae of British Columbia and Puget Sound. Can. Bull. Fish. Aquat. Sci. 204, pp. 1-192. Barnes, W. P. & G. A. Horridge. 1965. A neuropharmacological active substance from jellyfish ganglia. J. Exp. Biol., 42: 257-267. Bigelow, H. B. 1909. Reports on the scientific results of the Expedition to the Eastern Tropical Pacific of “Albatros”, from October, 1904 to March 1905. Mem. Mus. Comp. Zool. Harv., 37: 1-245. Bouillon, J. & F. Boero. 2000. Sinopsis of the Families and Genera of the Hydromedusae of the World, with a list of the Worldwide Species. Thalassia Salentina., 24: 47-296. Browne, E. T. 1902. A preliminary report on Hidromedusae from the Falklam Islam. Ann. Mar. Nat. Hist., 9(7): 272-284. Chamizo, A. & Eysenhardt. C. G. 1821. De animalibus quibusdam e classe vermium Linneana, in circumnavigatione térrea. (1815-18). Fasc. 2. Nova Acta Phys. Med. Acad. Caesar. Leopold. Carol. Nat. Cur., 5 (2): 345-374. Fernández-Álvarez, O. 1981. Estudio preliminar sobre la distribución y abundancia de algunas especies de medusas (Cnidaria: Hydrozoa y Scyphozoa) del Golfo de Tehuantepec, México. Tesis Profesional. Facultad de Ciencias, UNAM., México. 81 pp. Gasca-Serrano, R. 1993. Especies y abundancia de sifonóforos (Cnidaria: Hydrozoa) en la Región Sur del Golfo de México. Carib. J. Sci., 29 (34): 220-225. Gegenbaur, C. 1856. Versuch eines Systemes der Medusen, mit Beschreibung neuer oder wening gekannter Formen; zugleich ein Beitrag… z. wiss. Zool., Bd. 8: 202-273. Haeckel, E. 1865. Über eine neue form des Generationswechsels bei den Medusen und üver die Verwandschaft der Geryoniden und Aeginiden. Monatsber. Königel. Akad. Wiss., Berlin, 85-94. Haeckel, E. 1879. Das System der Medusen. 2 vols. Gustav Fisher Verlag, Jena I., pp. 361-672. Kölliker, A. 1853. In: Gegenbaur, Kölliker y Müller. 1953; Berich über einige im Herbst 1852 in Messina angestellte vergleichende-anatomische Under suchungen. Z. Wiss. Zool. 4(3-4): 299-370. Kramp, P.L.. 1961. Synopsis of the Medusae of the World. Journal of the Marine Biological Association of the United Kingdom, 40: 1-469. Loman-Ramos, L. 2000. Medusas y Sifonóforos de la Bahía “La Ventosa”, Oaxaca, México (1987-1993). Informe de Servicio Social, Departamento de Hidrobiología, UAM-Iztapalapa. 1-6 pp. Mayer, A. G. 1910. Medusae of the world. Vols. I, II y III, Cargnegie Foundation, Washington, D.C. 735 pp. MacCrady, J. 1859. Gymnophtalma of Charleston Harbor. Proct. Elliott Soc. Nat. Hist.Charleston, South Carolina. 1. pp. 102-221. Péron, F. & C. A. Lesueur. 1809. Histoire générale et particuliere de tous les animaux que composent la famille des Méduses. Ann. Mus. Hist. Nat. 14:312-366. Quoy, J. R. C. & J. P. Gaimard. 1833. Voyage de 18 Astrolabe pendant 1826-29 sous le commandement de M. J. Dumont d’Urville. Zoologie, 4: 1-390. Russel, F. S. 1953. The Medusae of the British Isles: Anthomedusae, Leptomedusae, Limnomedusae, Trachymedusae and Narcomedusae. Cambrige University Press. Great Britain. 530 pp. Segura-Puertas, L. 1984. Morfología, Sistemática y Zoogeografía de las Medusas (Cnidaria: Hydrozoa y Scyphozoa) del Pacífico Tropical Oriental. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México. Publ. Esp. 8: 1-320. Tabrath, F., F. Kashiwagi & T. R. Norton. 1972. Antitumor activity in mice of four Coelenterate extracts. Int. J. Clin. Pharmacol Ther. Toxicol., 5(4): 420-422. Trinci, G. 1903. Di una nuova specie di Cytaeis gemmante del Golfo de Napoli. Mitt. Zool. Sta. Neapel, 16: 1-34. Van Beneden, P. J. 1844. Recherches sur l’Embriogénie des Tubularies, et l’Histoire Naturelle des différents Genres de cette Famille qui habitent la Côte d’Ostende. Mém. Acad. R. Belg., 17: 1-72. Vanhöffen, E. 1902. Die Acrasperen Medusen der deutschen Tiefsee Expedition 1898-1899 –I. Trachymedusen. Wiss. Engebn. “Valdivia”, 3: 1-52. ANEXOS Aglaura hemistoma Clytia simplex (antes Phialidium simplex) Dipurena brownei Clytia uchidai (antes Phialidium uchidai) Ectopleura dumortieri Eirene viridula Hydractinia minima (antes Podocoryne minima) Eucheilota comata Liriope tetraphylla Eutima spp. Nausithöe spp. Rhopalonema velatum Obelia spp. Sarsia sp. Rhopalonema funerarium Solmundella bitentaculata Solmaris sp.