Distribución espacial de Dinophysis spp. y detección de
Anuncio

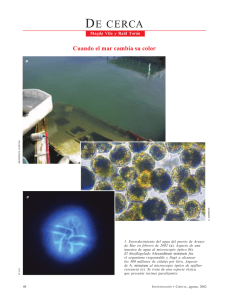
3.2 DISTRIBUCIÓN ESPACIAL DE Dinophysis spp Y DETECCIÓN DE TOXINAS VDM DISUELTAS EN EL AGUA, MEDIANTE SENSORES QUÍMICOS (X REGIÓN) (CONA-CF11 05-14) Gemita Pizarro1, César Alarcón1, José Franco2, Laura Escalera3, Beatriz Reguera3, Gastón Vidal1 & Leonardo Guzmán1 1 Instituto de Fomento Pesquero, Punta Arenas, Chile. 2 Unidad Asociada CSIC-IEO de Fitoplancton Tóxico, Vigo, España. 3 Instituto Español de Oceanografía (IEO), Vigo, España. INTRODUCCIÓN Las especies de dinoflagelados del género Dinophysis constituyen un serio problema para la salud pública y la actividad económica de recursos marinos de importancia comercial en el mundo y en Chile, dado que estas especies están asociadas a la producción de toxinas polietéreas diarreogénicas, como el ácido okadaico (AO) (Fig. 1) y hepatotóxicas, como las pectenotoxinas (PTXs) (Fig. 2) (Terao et al., 1986). En Chile este grupo de toxinas se conoce como veneno diarreico de los mariscos (VDM) por sus efectos sintomáticos en las personas que ingieren mariscos contaminados por ellas. Otro grupo de toxinas polietéreas, de efectos cardiotóxicos (Terao et al., 1990), son las yesotoxinas (YTXs) (Fig. 3), producidas por los dinoflagelados Protoceratium reticulatum, presente en la X (Proyecto FDI C703MR-01) y XI región (B. Reguera, com. pers.) y en menor cantidad Lingulodinium polyedrum (Satake et al., 1997; MacKenzie et al., 1998). Tradicionalmente se incluían las toxinas diarreogénicas (OA y derivados), las pectenotoxinas y las yesotoxinas en el mismo complejo denominado “Diarrhetic Shellfish Poisoning” (DSP, VDM en nuestro país). Hoy día se ha hecho una nueva clasificación y se han excluido las yesotoxinas y las pectenotoxinas del complejo DSP por no producir efectos diarreogénicos (Ogino et al., 1997). Las toxinas polietéreas son parcialmente liberadas al medio por los organismos en cultivos (Paz et al., 2004) y en el medio natural (MacKenzie et al., 2004). En Chile, la ocurrencia de toxinas diarreogénicas (AO y derivados) ha sido señalada en reiteradas oportunidades desde 1970 en el área del seno Reloncaví X región (Lembeye et al., 1993) y desde 1991 en la XI región (Anónimo, 1995). Durante 1999 y 2000 también ha sido detectada en el extremo norte de la XII región en niveles muy por debajo de la normativa por lo que no representan peligro para las personas (Guzmán et al., 2000, 2001). Durante el año 2005, los episodios de intoxicación de personas con síntomas de VDM fueron críticos durante los meses de verano en la X región. Además, ha habido casos de confusión en cuanto a la identificación del agente causante al coincidir la presencia de especies de Dinophysis en la columna de agua con un brote de Vibrio parahaemoliticus. Esta bacteria patógena produce los mismos síntomas del VDM al ingerir mariscos contaminados por ella. La principal diferencia estriba en que — 59 — Crucero CIMAR 11 las toxinas diarreicas de origen fitoplanctónico son termo-resistentes y las del Vibrio son termo-lábiles. Los recurrentes eventos de VDM en el seno Reloncaví (1970, 1979, 1986), fiordo Jacaf (1980, 1984), archipiélago Chonos (1991), estos dos últimos en la XI región, estuvieron asociados a la presencia de D. acuta (Lembeye at al., 1993) y aunque no fue confirmada como el organismo causante de la toxina éste ha sido el organismo más probable de producirla pues fue encontrado en 1991 en el tracto digestivo de mariscos contaminados con AO y dinofisistoxina-1 (DTX1) (predominante) (Zhao et al., 1993). En 1999 también fueron detectadas ambas toxinas en mariscos de la XI región pero en niveles trazas (Villarroel 2004) y en el 2000 sólo fue detectada DTX1 en la misma región (García et al., 2003). Dinophysis acuta, D. acuminata, D. rotundata y D. caudata, son dinoflagelados recurrentes en zonas templadas a nivel mundial, incluyendo algunas regiones de Chile. Dinophysis spp aparece principalmente durante los meses de primavera y verano en las regiones sur-austral de Chile (Uribe et al., 1998). Todas ellas son productoras de VDM en Europa (e.g. Lee et al., 1989; Andersen et al., 1996; Fernández et al., 2001; Marcaillou et al., 2001; Fernández et al., 2004; Fernández et al., 2005), Japón (e.g. Lee et al., 1989; Suzuki et al., 2001) y Nueva Zelanda (e.g. Suzuki et al., 2004; Mackenzie et al., 2005) de modo que constituyen potenciales agentes causantes de eventos tóxicos de VDM en Chile. Sin embargo, a la fecha los perfiles de toxinas de dichas especies son desconocidos en nuestro país, razón por la que es necesario dilucidar urgentemente, qué especies son los agentes causantes de los eventos VDM en Chile (Dinophysis spp, Protoceratium reticulatum, Prorocentrum spp) y la contribución de cada uno de ellos a estos episodios. Se ha encontrado que las mismas especies de Dinophysis presentan perfiles de toxinas diferentes de acuerdo a su procedencia. Tal es el caso de D. acuta, que en Irlanda presenta un perfil de toxinas con predominancia de dinofisistoxina-2 (DTX2), seguida de AO y PTX2 (Fernández-Puente et al., 2004), mientras que la misma especie en Nueva Zelanda presenta un perfil con predominio de PTX2, PTX11 y pequeñas cantidades de AO (Mackenzie et al., 2005). Así, D. acuta de Nueva Zelanda no presenta DTX2, a diferencia de D. acuta de Irlanda (James et al., 1999), España (Fernández et al., 2001) y Portugal (Vale, 2004). D. acuta en los países escandinavos presenta DTX1, toxina que ha sido detectada en los mariscos de la XI Región de Chile asociado a las proliferaciones de esta especie de Dinophysis. Además, una misma especie puede resultar no tóxica en ciertas partes del mundo, como es el caso de D. acuminata en la costa NE de USA (L. Maranda com. pers.) y constituir la principal fuente de toxinas VDM en Europa y Nueva Zelanda (Lee et al., 1989; Moroño et al., 2002; Fernández et al., 2001; Mackenzie et al., 2005). Más aún, existen especies productoras de toxinas que presentan cepas con diferentes grados de producción y perfil de toxinas procedentes de una misma localidad geográfica (e.g. Bravo et al., 2001). En el caso de Chile, el monitoreo para VPM está restringido a D. acuta y D. acuminata, debido a la falta de estudios del contenido y composición de las toxinas en otras especies. En este caso D. acuminata ha sido asociada a la producción de DTX1 en la XII región de Chile (Uribe et al., 2001). Las diferencias de perfiles de toxinas — 60 — entre las especies, e incluso entre cepas de la misma especie en distintas localidades tienen una gran repercusión en las labores de regulación y manejo de los recursos marisqueros susceptibles de ser contaminados por estas toxinas. Ante tantas incógnitas, la aplicación de novedosas técnicas, como los sensores químicos (SPATT) (Mackenzie et al., 2004), consistentes en pequeñas bolsitas rellenas con resina de adsorción pasiva de biotoxinas polietéreas, podría resultar de gran utilidad como herramienta de detección precoz de ficotoxinas en Chile. Estas bolsitas de resinas son utilizadas a modo de bivalvo artificial, lo que permite predecir estimativamente la acumulación de toxinas VDM y YTXs en los mariscos cuando sus concentraciones distan aún de los niveles de regulación no aptos para el consumo humano. La resina de los sensores es inocua para el medio ambiente y tras permanecer unos días en la columna de agua, son recuperados y conservados en el congelador hasta su análisis. Este modelo ha demostrado ser sensible y de muy bajo costo, ideal para ser probado con el fin de utilizarlo por largos períodos y en vastas extensiones geográficas como las regiones del sur austral de Chile. En los últimos años, gracias a las nuevas tecnologías de perfilamiento microestructural de la columna de agua con sistemas de última generación, se ha reportado frecuentemente que las especies de Dinophysis y otros dinoflagelados tóxicos como D. acuminata se pueden presentar en altas densidades en finas capas situadas en zonas de discontinuidad termohalina de la columna de agua. Estas capas finas se sitúan en profundidad variable y pueden alcanzar extensiones horizontales que pueden llegar a 25-30 km (Gentien et al., 2005). Así, la formación de las capas finas de alta densidad celular dependería de la estructura vertical de la columna de agua y de la intensidad de la estratificación. Esta línea de investigación está aún incipiente y su estudio representa grandes perspectivas para la comprensión de los ciclos de generación, persistencia y desaparición de los florecimientos de microalgas nocivas en Chile. HIPÓTESIS La composición de toxinas lipofílicas del fitoplancton presente en la X Región de Chile es diferente al encontrado en Europa y Nueva Zelanda y dependerá de la composición y abundancia de distintas especies toxigénicas. Se espera que los perfiles de toxinas sean similares en el fitoplancton de fiordos, canales y centros de cultivo dentro de la misma región. El perfil de toxinas estará asociado a la especie de Dinophysis predominante en el ensamble fitoplanctónico. En períodos de estratificación, las especies de Dinophysis presentan máximos celulares en capas óptimas, específicas de especie, determinadas por el gradiente de densidad (incremento sigma-T respecto a Z) de la columna de agua. Los sensores químicos podrían constituir una herramienta confiable, sensible y barata para los monitoreos de toxinas lipofílicas como alerta temprana de la presencia de VDM en los mariscos. — 61 — Crucero CIMAR 11 OBJETIVOS Describir la distribución espacial de Dinophysis spp en primavera-verano. Establecer la composición de toxinas lipofílicas presente en el fitoplancton (toxina particulada concentrada con red de arrastre) durante el período de muestreo en los concentrados de fitoplancton y asociarlo a la presencia de especies potencialmente tóxicas de dinoflagelados presentes durante diciembre-febrero de 2005-2006. Evaluar la utilidad de los sensores químicos en el monitoreo de toxinas VDM en la X región para su aplicabilidad potencial como alerta temprana de los niveles de toxicidad bioacumulados por recursos de importancia comercial. MATERIALES Y MÉTODOS Lugar de muestreo La recolección de fitoplancton cuali y cuantitativo así como la puesta de contenedores con resina en la columna de agua por un período de 5-7 días, fue realizada en 10 estaciones (Fig. 4). Las estaciones pertenecen a la Sección CP (estaciones 4, 5, 6 y 7); sección DH (estaciones 2, 3 y 5); Sección QL (estaciones 3, 5 y 6). El fitoplancton fue recolectado durante diciembre de 2005 y enero-febrero de 2006, en tanto las resinas fueron colocadas en diciembre de 2005 y enero-febrero de 2006. Los contenedores de resina colocados en diciembre no lograron ser recuperados debido a las condiciones climáticas, razón por la que no fueron analizados. Recolección del fitoplancton El fitoplancton cualitativo fue recolectado entre 0-30 m de profundidad con una red de arrastre con apertura de malla de 20 µm. Una alícuota de esta muestra fue fijada en lugol para el análisis microscópico cualitativo del fitoplancton. El fitoplancton cuantitativo fue recolectado a 0-5-10-20-30 m de profundidad con botellas oceanográficas Niskin. Una alícuota de 50 mL de estas muestras fueron fijadas con formalina para su posterior análisis en un microscopio invertido según método de Utermöhl (1931). Adsorción in situ de toxinas lipofílicas disueltas en el agua mediante resina DIAION: Preparación La determinación del perfil de toxinas disueltas en el agua de mar se realizó mediante la extracción de las toxinas del medio adsorbidas por resinas sintéticas porosas DIAION (HP-20, Supelco) de 250-850 mm de tamaño de partícula y adsorbente poliaromático. Previamente, la resina fue preparada de acuerdo a Mackenzie et al. (2004): 300 g de resina, se depositaron en un vaso de 4 L y se añadió 3 L de MeOH, lo cual se agitó a baja velocidad durante toda la noche. Se eliminó el MeOH en un embudo Buchner con papel filtro y presión de vacío suave, se lavó la resina con MQ y traspasó al vaso de 4 L. Se añadió aproximadamente unos 3 L de agua MQ, se agitó suavemente durante 30 min y se filtró a presión de vacío suave. Esta operación de lavado se repitió una vez más. Finalmente, se recogió la resina en 2 L de agua MQ y se almacenó en botellas plásticas a 4 ºC hasta su uso. — 62 — Se colocaron aproximadamente 0,5 g de resina húmeda activa, en el interior de bolsitas (Fig. 5), confeccionada con una red de 77 µm de apertura de malla. Las bolsitas, de dimensiones aproximadas de 60x60 mm, se confeccionaron con hilo de poliéster. Estos contenedores se expusieron durante 5-7 días a 3 m de profundidad en el mar (Fig. 4) próximos a líneas de cultivo de choritos. Una vez recuperados los contenedores fueron almacenados a 4 º C hasta el momento del análisis. Extracción de toxinas adsorbidas por la resina La extracción de las toxinas adsorbidas por las resinas durante su permanencia en la columna de agua se realizó siguiendo la metodología de Mackenzie et al. (2004) con algunas modificaciones. Una vez recuperados los contenedores de resina, la resina fue traspasada a un tubo provisto con una red de 20 en uno de sus extremos, donde fue lavada con abundante agua MQ (50-100 mL). Después de escurrir el exceso de agua, la resina fue traspasada a un vial de 20 mL con al menos 10-15 mL metanol 100%, donde permaneció al menos dos días a 4 ºC en oscuridad. Una vez extraída la toxina desde la resina, ésta fue traspasada a un tubo de elusión, cuyo extremo inferior se taponeó con un tubo de silicona provisto con un filtro en su interior (algodón hidrofóbico). Se reguló el flujo con una llave de paso. Transcurrido este tiempo, el metanol fue eluido a través del filtro de algodón para retener las partículas de resina. La resina se lavó dos a tres veces hasta completar 50 mL de metanol eluido. Los extractos metabólicos se conservaron refrigerados hasta el momento del análisis. Previo a inyectar la muestra en el LC-MS (20 µL), una alícuota de 4 mL se evaporó a baja presión a 40 ºC (Speed Vac Savant), durante 3 h, se resuspendió en 500 µL de metanol y se filtró por 0,45 µm. Análisis de toxinas por LC-MS Los análisis de toxinas se realizaron en un cromatógrafo líquido THERMO FINNIGAN acoplado a un espectrómetro de masas THERMO FINNIGAN LCQ-ADVANTAGE equipado con trampa iónica y una interfase de electro-espray iónico (ESI). El ESI fue operado en modo positivo y negativo (3,0 y 4,5 kV voltaje del espray, temperatura del capilar de 250 º C, flujo de gas de arrastre de 20 mL/min y flujo de gas auxiliar 10 mL/min). Las condiciones óptimas de ionización fueron establecidas en modo positivo y negativo con un estándar de ácido okadaico y fase móvil de acetato amónico 2 mM, pH 5,8 (A) y metanol (B) (30:70). La separación cromatográfica de las toxinas se realizó con una columna XTerra C18 (2,1 x 150 mm, 5 mm), y flujo de la fase móvil de 0,2 mL/min en el siguiente gradiente: 60% a 100% de B entre los 0 y 20 min, sostenido a 100% hasta el minuto 22, disminuyendo al 60% de B en el minuto 25 y sostenido hasta el minuto 30. A través de un microcapilar 50 mL del eluyente proveniente del LC se dirigieron a la interfase de pulverización iónica Los cromatogramas se registraron en un rango m/z de 200-2000 y el valor de detección iónico m/z para cada toxina fue: 827,5 [M+Na]+ y 803,5 [M-H]– para el AO y DTX2; 841,5 [M+Na]+ y 817,5 [M-H]– para el DTX1; 881 [M+Na]+ y 857,5 [M-H]–para la PTX2; 899 [M+Na]+ y 875,0 [M-H ]– para la PTX2SA y 1141 [M-2+H]– para la YTX. — 63 — Crucero CIMAR 11 La calibración del sistema de LC-MS y la cuantificación de las toxinas se realizaron con estándares de AO y PTX2 certificados por el Nacional Research Council de Canadá. La solución estándar de PTX1 fue donada por el Dr. T. Yasumoto (laboratorio de Investigación de los Alimentos de Japón), el patrón de DTX2 (1 mg/mL) por el Dr. P. Hess (Marine Institute, Galway, Irlanda), y los de PTX2SA y DTX1 (1 µg/mL) por el Dr. C. Miles (National Veterinary Institute, Oslo, Norway). La solución estándar de YTXs fue provista por el Dr. M. Satake, de la Universidad de Tohoku (Sendai, Japón). Las curvas de calibración se elaboraron con una solución de trabajo consistente en 0,60 ng/mL de AO, 0,59 ng/mL de DTX2 y 0,70 ng/mL de PTX2. Cada 15-20 muestras inyectadas, se generó una curva con 4 puntos obtenidos por una inyección de volumen variable de la solución de trabajo. La linealidad de las concentraciones de calibración usadas se confirmó estadísticamente siguiendo el método de Van-Trijp & Roos (1991). Se utilizó una solución cualitativa de trabajo con PTX2SA, PTX1 y DTX1 (0,25, 0,25 y 0,50 ng/mL, respectivamente) para la detección de estas toxinas en las muestras de fitoplancton. RESULTADOS Estándares Los cromatogramas de estándares de OA, DTX2, PTX2 y sus respectivos espectros m/z obtenidos por LC-MS se muestran en la figura 6. Así mismo se muestran los cromatogramas de estándares de PTX2SA, PTX1, dos toxinas que son producto de la transformación enzimática por parte de los moluscos que ingieren microalgas productoras de PTX2. Los cromatogramas y espectros de las toxinas DTX1 y YTX, ambas toxinas presentes en el sur de Chile se presentan en la Figura 7. Toxinas adsorbidas por las resinas La toxina presente con una mayor frecuencia en el área de estudio fue la PTX2 (Tabla I). Ejemplo de cromatograma y espectro m/z del extracto metanólico de la resina con PTX2, adsorbida después de permanecer una semana (del 21 de enero de 2006) en la columna de agua en la localidad de San Antonio (CP4) se presenta en la figura 8. PTX2SA (Fig. 9) fue encontrada en las resinas después de permanecer una semana (del 26 de enero de 2006 y 21 de enero de 2006) en la columna de agua de la localidad de Toralla (DH3) y en San Antonio (QL3) respectivamente. DTX1 fue otra de las toxinas del VDM, encontrada en LIN (CP7), Puluqui A (CP5) y en Calbuco (CP4). Un ejemplo del cromatograma para DTX1 del extracto metanólico de toxina adsorbida después de permanecer una semana (del 14 de febrero de 2006) en la columna de agua en LIN así como el espectro m/z respectivo obtenido por LC-MS, se presenta en la figura 10. Puluqui A (CP5) fue la localidad en la que se encontró trazas YTX (Fig. 11). La toxina fue adsorbida por la resina tras permanecer en la columna de agua durante la semana del 14 de febrero de 2006. — 64 — Fitoplancton cuali y cuantitativo Durante el período de muestreo, predominaron fuertemente las diatomeas (Tabla II) respecto a los dinoflagelados. Especies potencialmente tóxicas de Dinophysis sólo logró observarse en Yaldad (QL6), con 200 cél/L de D. acuminata. No obstante las muestras cualitativas de fitoplancton (Tabla III) revelaron la presencia de otras especies de Dinophysis: D. acuta y D. rotundata (Fig. 12). También se observó una especie similar a Protoceratium reticulatum, productora de YTXs en otras partes del mundo; sin embargo para establecer rigurosamente su identificación se requiere de un análisis de placas de su teca, actividad que será informada en la publicación final de esta expedición CIMAR 11. DISCUSIÓN La toxina predominante encontrada es PTX2, hallándose en trazas AO y YTX. La PTX2Sa, producto de transformación enzimática del PTX2, también fue encontrada en dos de las estaciones, sin embargo no es posible establecer su procedencia por cuanto su presencia en el agua puede ser debida al metabolismo de mariscos contaminados, zooplancton o incluso por mortalidad de las propias células fitoplanctónicas que la producen. De acuerdo a la hipótesis, existe DTX1 en las muestras de resina como segunda toxina importante y concuerda con resultados anteriormente reportados para Chile. Ninguna señal de la presencia de DTX2 fue encontrada. De acuerdo a las células fitoplanctónicas observadas en las muestras, existe una mejor representación de las Dinophysis en las muestras cualitativas que en las muestras cuantitativas. Sólo fue posible encontrar 200 cél/L de D. acuminata en la estación de Yaldad (QL6), en circunstancias que el listado de especies potencialmente tóxicas incluye también D. acuta y D. rotundata. La presencia de YTXs en trazas indica la presencia potencial del dinoflagelado Protoceratium reticulatum, sin embargo se requiere un estudio de placas de la teca de la especie sospechosa observada en muestras cualitativas para establecer rigurosamente su identidad. El método es suficientemente sensible para ser utilizado como alerta temprana de eventos del tipo VDM, no obstante lo anterior, para establecer una relación entre la cantidad de toxinas adsorbidas por la resinas con el contenido de toxina en los mariscos (i.e. bioensayo) de acuerdo a la normativa vigente, es necesario realizar una serie de tiempo controlando ambas variables. AGRADECIMIENTOS Investigación cofinanciada por el Centro de Estudios del Cuaternario, CEQUA (con sede en Punta Arenas, XII región), que ha hecho posible una estadía en el Centro Español de Oceanografía de Vigo, para realizar la investigación y análisis de toxinas lipofílicas mediante Cromatografía Líquida acoplada a un Espectrómetro de Masas. — 65 — Crucero CIMAR 11 REFERENCIAS ANDERSEN, P., B. HALD, & H. EMSHOLM (1996). Toxicity of Dinophysis in Danish coastal waters. In: Yasumoto, T. Oshima, Y. & Fukuyo, Y. (Eds.), Harmful and Toxic Algal Blooms. Intergovernmental Oceanographic Commission of UNESCO, Senadi, Japan, pp. 281-284. ANÓNIMO. (1995). COI-Segundo Taller Regional de Planificación Científica sobre Floraciones de Algas Nocivas en Sudamérica. Informes de reuniones de trabajo 123. Anexo IV: 19-20. BRAVO, I., M. L. FERNÁNDEZ, I. RAMILO & A. MARTÍNEZ (2001). Toxin composition of the toxic dinoagellate Prorocentrum lima isolated from different locations along the Galician coast (NW Spain). Toxicon 39 1.537-1.545. FERNÁNDEZ, M. L., B. REGUERA, I. RAMILO & A. MARTÍNEZ (2001). Toxin content of Dinophysis acuminata, D. acuta and D. caudata from Galician Rias Bajas. In: Hallegraeff, G., et al. (Eds), Harmful Algal Blooms 2000. Intergovernmental Oceanographic Commission of UNESCO 2001, pp. 360-363. FERNÁNDEZ, M. L., A. MIGUEZ, A. MOROÑO, F. ARÉVALO, Y. PAZOS, C. SALGADO, J. CORREA, J. BLANCO, S. GONZÁLEZ-GIL & B. REGUERA (2005). First report of pectenotoxin-2 in phytoplankton net-hauls and mussels from the Galician Rias Baixas (NW Spain) during proliferations of Dinophysis acuta and D. caudata. Toxicon. FERNÁNDEZ, P., M. J. FIDALGO, B. HAMILTON, A. FUREY & K. JAMES (2004). Studies of polyether toxins in the marine phytoplankton, Dinophysis acuta, in Ireland using multiple tandem mass spectrometry. Toxicon 44: 919-926. GARCÍA, C., P. PEREIRA, L. VALLE & N. LAGOS (2003). Quantitation of diarrhetic shellfish poisoning toxins in Chilean Mussell using pyrenyldiazomethane as fluorescent labelling reagent. Biol. Res. 36: 171-183. GENTIEN, P., P. DONAGHAY, H. YAMAZAKI, R. RAINE, B. REGUERA & T. OSBORN (2005). Harmful algal blooms in stratified environments. Oceanography. GUZMÁN, L., G. PIZARRO, R. IGOR, A. ATALAH, O. OYARZO, E. BARTICEVIC, C. ALARCÓN, J. FOPPIANO, N. BUTOROVIC, E. ALMONACID & M. RUIZ (2000). Difusión Programa Marea Roja en la XII Región de Magallanes y Antártica Chilena, Cuarta Etapa. Tomos I, II, III, IV. 274 pp. + figuras + fotografías + anexos. GUZMÁN, L., G. PIZARRO, M. I. BANCIELLA, R. IGOR, C. ALARCÓN, H. PACHECO, O. OYARZO, M. FARRÉ & N. BUTOROVIC (2001). Informe Final Subprograma de Monitoreo. Tomo I. 129 pág. + figuras y anexos. En: Difusión Programa Marea Roja en la Región de Magallanes y Antártica Chilena, IV Etapa. JAMES, K. J., A. G. BISHOP, R. DRAISCI, L. PALLESHI, C. MARCHIAFAVA, E. FERRETTI, M. SATAKE, & T. YASUMOTO (1999). Liquid chromatographic — 66 — methods for the isolation and identification of new pectenotoxin-2 analogues from marine phytoplankton and shellfish. J. Chromatogr. A 844: 53-65. LEE, J. S., T. T. IGARASHI, S. FRAGA, E. DAHL, P. HOVGAARD & T. YASUMOTO (1989). Determination of diarrhetic shellfish toxins in various dinoflagellate species. J. Appl. Phycol. 1: 147-152. LEMBEYE, G., T. YASUMOTO, J. ZHAO, R. FERNÁNDEZ, (1993). DSP OUTBREAK IN CHILEAN FIORDS. IN SMAYDA, T. J. & Y. SHIMIZU (eds.), Toxic Phytoplankton Blooms in the Sea. Elsevier, Amsterdam: 525-529. MACKENZIE, L., P. TRUMAN, M. SATAKE, T. YASUMOTO, J. ADAMSON, D. MOUNTFORT & D. WHITE (1998). Dinoflagellate blooms and associated DSPtoxicity in dhellfish in New Zealand. In: Reguera, B., J. Blanco, M. L. Fernández & T. Wyatt (Eds.). Harmful Algae. Xunta de Galicia & Intergovernmental Oceanographic Comisión of UNESCO. Santiago de Compostela, pp 74-77. MACKENZIE, L., V. BEUZENBERG, P. HOLLAND, P. MCNABB & A. SELWOOD (2004). Solid phase adsorption toxin tracking (SPATT): a new monitoring tool that simulates the biotoxin contamination of filter feeding bivalves. Toxicon 44: 901-918. MACKENZIE, L., V. BEUZENBERG, P. HOLLAND, P. MCNABB, T. SZUKI & A. SELWOOD (2005). Pectenotoxin and okadaic acid-based toxin profiles in Dinophysis acuta and Dinophysis acuminata from New Zealand. Harmful Algae 4: 75-85. MARCAILLOU, C., P. GENTIEN, M. LUNVEN, J. LE GRAND, F. MONDEGUER, M. M. DANILOU, M. P. CRASSOUS & A. YOUENOU (2001). Dinophysis acuminata distribution and specific toxin content in relation to mussel contamination. In: Hallegraeff, G. M., Blackburn, S. I., Bolch, C. J., Lewis, R. J. (Eds.), Harmful Algal Blooms 2000. Intergovernmental Oveanographic Commission of UNESCO, pp. 356-359. MOROÑO, ARÉVALO, F., M. L. FERNÁNDEZ, J. MANEIRO, Y. PAZOS, C. SALGADO & J. BLANCO (2002). Accumulation and transformation odf DSP toxins in mussels Mytilus galloprovincialis during a toxic episode caused by Dinophysis acuminata. Aquatic Toxicology 00: 1-12. OGINO, H., M. KUMAGAI & T. YASUMOTO (1997). Toxicologic evaluation of yessotoxin. Nat. Toxins 5: 255-259. PAZ, B., P. RIOBÓ, M. L. FERNÁNDEZ, S. FRAGA, J. M. FRANCO (2004). Production and release of yessotoxins by the dinoflagellates Protoceratium reticulatum and Lingulodinium polyedrum in culture.Toxicon.44: 251-258 . SATAKE, M., A. L. MACKENZIE & T. YASUMOTO (1997). Identification of Protoceratium reticulatum as the biogenetic origen of yessotoxin. Nat. Toxins. 5: 164-167. — 67 — Crucero CIMAR 11 SUZUKI, T., L. MACKENZIE, D. STIRLING & J. ADAMSON (2001). Pectenotoxin-2 seco acid: a toxin converted from pectenotoxin-2 by the New Zealand Greenshell mussel, Perna canaliculus. Toxicon 39: 507-514. SUZUKI, T., V. BEUZENBERG, L. MACKENZIE & M. A. QUILLIAM (2004). Discovery of okadaic acid esters in the toxic dinoflagellate Dinophysis acuta from New Zealand using liquid chromatography/tandem mass spectrometry. Rapid. Commun. Mass Spectrom. 18: 1.131-1.138. TERAO, K., E. ITO, M. OARADA, M. MURATA & T. YASUMOTO (1990). Histopathological studies on experimental marine toxin poisoning-5. The effects in mice of yessotoxin isolated from Patinopecten yessoensis and of a desulfated derivate. Toxicon 28: 1.095- 1.104. TERAO, K., E. ITO, T. YANAGI & T. YASUMOTO, (1986). Histopathological studies on experimental marine toxin poisoning. Part I: Ultraestructural changes in the small intestine and liver of sucking mice induced by dinophysis toxin-1 and pectenotoxine– 1. Toxicon 24: 1.141-1.151. URIBE, J. C., L. GUZMÁN, H. PACHECO, R. IGOR, G. PIZARRO, E. BARTICEVIC, M. FRANGOPULOS, C. ALARCÓN, T. HROMIC, C. LABBE, A. ATALAH & P. CHANG (1998). Informe Final Difusión Programa Marea Roja en la XII Región de Magallanes y Antártica Chilena, Primera Etapa Informe Presentado al Gobierno Regional. URIBE, J. C., C. GARCÍA, M. RIVAS, N. LAGOS (2001). First Report Diarrhetic Shellfish Toxins in Magellanic Fiords, Southern Chile. J. Shellfish Res. 20: 69-74 UTERMÖHL, H. (1931). Neue wege in der quantitativen Erfassung des Planktons (mit besonderer Berücksichtigung des Ultraplanktons). Verh.Int. Ver. Theor. Angew. Limnol. 5: 567-596. VALE, P. 2004. Differential dynamics of dinophysistoxins and pectenotoxins between blue mussel and common cockle : a phenomenon originating from the complex toxin profile of Dinophysis acuta. Toxicon 44:123:134. VAN-TRIJP, J. M. P. & A. H. ROOS (1991). Model for the calculation of calibration curves. Taken from the RIKILT Report 91.02, January 1991. RIKILT-DLO, Wageningen, NL. VILLARROEL, O. (2004). Detección de toxinas paralizante, diarreica y amnésica en mariscos de la XI región por cromatografía líquida de alta resolución (HPLC) y bioensayo en ratones. Cienc. Tecnol. Mar. 27: 33-42. ZHAO, J., G. LEMBEYE, G. CENCI, B. WALL & T. YASUMOTO (1993). Determination of okadaic acid and dinophysistoxin-1 in mussels from Chile, Italy and Ireland. In Smayda, T. J. & Y. Shimizu (eds.), Toxic Phytoplankton Blooms in the Sea. Elsevier, Amsterdam: 587-592. — 68 — Tabla I. Toxinas adsorbidas (ng/ bolsita) por las resinas que permanecieron durante una semana en la columna de agua en distintas localidades de la X región. Se indica la cantidad en peso seco de resina utilizada por bolsita. Lugar Calbuco Calbuco Puluqui A Puluqui B LIN Huenao Toralla Coñab San Antonio Q Viejo Yaldad Sección CP4 CP4 CP5 CP6 CP7 DH2 DH3 DH5 QL3 QL5 QL6 Fecha OA DTX2 14-feb-06 14-feb-06 trazas 14-feb-06 trazas 14-feb-06 14-feb-06 26-ene-06 26-ene-06 26-ene-06 21-ene-06 21-ene-06 trazas 21-ene-06 — 69 — PTX2 PTX2SA YTX DTX1 trazas 59 64 10 71 19 34 65 60 7 164 22 31 15 resina (g) peso seco 0,37 0,36 0,36 0,47 0,33 0,46 0,38 0,09 0,15 0,08 0,10 Crucero CIMAR 11 Tabla II. Resumen del fitoplancton cuantitativo presente en las estaciones muestreadas en el área de estudio en los meses de verano. CALBUCO dic-05 CALBUCO 14-feb-06 PULUQUI A 14-feb-06 PULUQUI B 14-Feb-06 LIN dic-05 LIN 14-feb-06 HUENAO dic-05 HUENAO 26-ene-06 TORALLA dic-05 TORALLA 26-ene-06 Profundidad Total Diatomeas Total Dinoflagelados D. acuminata Om 5m 10 m 20 m 30 m Om 5m 10 m 20 m 30 m Om 5m 10 m 20 m 30 m Om 5m 10 m 20 m 30 m Om 5m 10 m 20 m 30 m Om 5m 10 m 20 m 30 m Om 5m 10 m 20 m 30 m Om 5m 10 m 20 m 30 m Om 5m 10 m 20 m 30 m Om 5m 10 m 20 m 30 m 75.800 99.800 189.300 61.400 26.920 18.700 163.000 49.500 56.800 74.100 367.900 817.900 220.300 1.514.000 416.600 83.000 132.500 1.291.800 664.600 977.000 587.920 413.240 3.379.360 72.000 5.700 581.020 623.780 944.960 84.200 8.400 171.300 121.100 140.600 75.800 101.200 137.000 172.600 133.300 33.800 51.200 1.276.100 1.568.700 128.300 158.500 497.300 503.300 368.100 279.600 433.800 306.100 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 2600 800 400 0 0 0 0 0 0 0 0 0 0 0 0 200 600 0 0 0 0 700 100 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 — 70 — continuación Tabla II. Profundidad COÑA B dic-05 COÑA B 26-ene-06 SAN ANTONIO SAN ANTONIO dic-05 21-ene-06 Q. VIEJO dic-05 Q. VIEJO 21-ene-06 YALDAD dic-05 YALDAD 21-ene-06 Om 5m 10 m 20 m 30 m Om 5m 10 m 20 m 30 m Om 5m 10 m 20 m 30 m Om 5m 10 m 20 m 30 m Om 5m 10 m 20 m 30 m Om 5m 10 m 20 m 30 m Om 5m 10 m 20 m 30 m Om 5m 10 m 20 m 30 m — 71 — Total Diatomeas 1.498.000 725.800 800.600 694.500 296.000 1.789.960 2.625.760 1.373.400 812.000 300.300 66.900 225.900 75.100 66.600 89.400 985.900 137.700 171.600 253.600 465.800 227.600 823.900 179.700 547.000 54.100 1.116.500 236.500 811.500 214.900 106.400 128.200 118.100 134.100 20.100 10.300 148.800 38.000 85.400 24.400 20.000 Total Dinoflagelados 0 3400 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 200 400 400 500 0 D. acuminata 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 200 0 0 Crucero CIMAR 11 Tabla III. Ejemplo de especies de fitoplancton cualitativo presentes en las muestras recolectadas el 21-ene-2006 en la estación San Antonio (QL3). Diatomeas Asterionella japonica Coscinodiscus sp. Diploneis sp. Odontella sp. Paralia sulcata Pseudonitzschia sp. Thalassiosira rotula Dinoflagelados Ceratium furca Ceratium lineatum Dinophysis acuminata Dinophysis acuta Dinophysis rotundata Diplopsalis sp. Gonyaulax sp. Oblea sp. Prorocentrum micans Protoceratium sp. Protoperidinium claudicans Protoperidinium claudicans Protoperidinium conicum Pyrocystis lunula Scrippsiella sp. Silicoflagelado Dyctiocha speculum — 72 — Figura 1: Estructura del ácido okadaico (OA) y derivados: dinofisistoxinas (DTXs) y acyl derivados. Figura 2: Estructura de las Pectenotoxinas (PTXs) y pectenotoxinas seco ácido (PTXsSA). — 73 — Crucero CIMAR 11 R2 HO H O H O H O H H H H O R 1O NaO3SO n O H H H O H H H OH O H H O O H O H H O H Yessotoxin (YTX): n=1; R1= SO3Na; R2= 45-HydroxyYTX: n=1; R1= SO3Na; R2= 45, 46, 47-TrinorYTX n=1; R1= SO3Na; R2= HomoYTX: n=2; R1= SO3Na; R2= 45-HydroxyhomoYTX: n=2; R1= SO3Na; R2= OH H OH Figura 3: Estructura de las Yesotoxinas (YTXs) — 74 — 43° 4 5 6 7 Golfo de Ancud Se cci ón I C h s l a i l o é P a c í f i c o O c é a n o 42° S Sección CP DH 2 3 5 Golfo Corcovado Sección QL 6 5 3 Boca del Guafo Ba. Tictoc 44° 75° W 74° 73° Figura 4: Localidad de muestreo de fitoplancton y posición de bolsitas con resina en la columna de agua por un período de 5-7 días. Figura 5: Contenedor de resina utilizado en experimentos in situ para la adsorción de toxinas lipofílicas disueltas en el agua de mar: Bolsita confeccionada con red de 70 µm de tamaño de trama de 60x60 mm. — 75 — Crucero CIMAR 11 RT: 0.00 - 30.04 SM 7G AO DTX2 8.03 NL: 3.71 E6 m/z = 827.0-828.0 F: + c ESI Full ms [200.00-2000.00] MS 1P3 3000000 7.14 2000000 Intensity 1000000 0 12000000 0.88 2.0 2.99 3.73 4.29 9.40 10.05 6.08 11.99 0 14.73 15.45 13.19 16.82 17.71 18.99 27.60 26.47 20.68 21.89 22.13 23.34 24.14 24.79 29.22 11.91 NL: 1.21 E7 m/z = 881.0-882.0 F: + c ESI Full ms [200.00-2000.00] MS 1P3 10000000 8000000 6000000 4000000 2000000 0.71 1.53 2.34 3.24 4.21 4.94 0 0 9.72 7.62 8.19 6.08 8 6 4 2 12.95 13.84 14.4 11.34 10 12 66 18.19 20 26.07 26.80 27.84 28.97 24.38 19.56 20.28 21.33 22.37 18 16 14 22 28 26 24 30 Time (min) 1P3#171-201 RT: 6.98-8.11 AV:15 NL: 1.81E6 T: + c ESI Full ms [200.00-2000.00] 827.4 6 4 787.1 769.2 278.9 2 Relative Abundance 391.0 458.7 849.3 907.5 953.5 1046.8 606.6 696.2 0 100 1306.2 1265.1 1502.5 1698.7 1770.7 1954.6 1P3#292-298 RT: 11.83-12.07 AV:4 NL: 3.01E7 T: + c ESI Full ms [200.00-2000.00] 876.3 50 881.5 278.9 0 200 600 400 883.5 979.1 796.4 841.2 551.1 414.7 800 1200.2 1320.7 1437.6 1580.4 1656.3 1770.2 1400 1200 1000 1600 1969.9 2000 1800 m/z Figura 6: Cromatogramas de estándares de OA, DTX2, PTX2 y sus respectivos espectros m/z obtenidos por LC-MS. RT: 0.00 - 29.95 SM: 9G PTX2SA 7.27 Intensity 1000000 500000 2.66 1.28 1.69 0 3.79 4.92 5.53 8.16 22.43 11.16 9.30 11.72 10.11 10.11 Intensity 3 0.63 1.77 2.74 0 3.87 4.20 6.38 7.11 8.97 8.16 16.07 10.83 12.12 12.93 14.14 27.72 20.10 20.98 18.33 19.05 28.20 DTX1 1000000 500000 13.74 14.78 25.15 23.96 24.67 25.64 24.75 15.67 16.47 17.36 22.83 20.66 21.79 18.89 26.60 25.64 27.72 28.36 9.30 2000000 PTX1 1000000 2.58 0. 6 1.77 0 3.55 25.80 11.16 4.60 5.49 6.22 7.27 7.92 14.86 15.99 16.39 8.17 18.41 19.21 20.90 22.19 22.83 23.64 27.08 28.20 11.20 100 YTX 50 2.46 3.19 3.99 4.56 0.67 0 0 2 4 8.69 6.10 7.07 6 8 10 12 14 20.94 19.01 16.84 17.88 9.99 18 20 22.63 23.84 24.80 22 24 26.64 26 27.28 28.24 NL: 1.06 E6 m/z = 899.00-900.00 F: + c ESI Full ms [200.00-2000.00] MS 2P3 NL: 1.12 E6 m/z = 841.00-842.00 F: + c ESI Full ms [200.00-2000.00] MS 2P3 NL: 2.14 E6 m/z = 892.00-893.00 F: + c ESI Full ms [200.00-2000.00] MS 2P3 NL: 1.27 E6 m/z = 1141.00-1142.00 F: + c ESI Full ms [200.00-2000.00] MS 2P3 28 Time (min) Intensity 76 0 478.66 591.11 67 894.24 899.41 874.50 962.93 1095.64 534.64 676.30 7 863.47 983 0 1296.79 1402.53 1622.08 1836.83 1927.68 892.23 431.76 639.84 762.07 832.74 0 1000000 0 1949.18 841.44 1000000 2000000 1505.26 1633.13 1711.68 1 87.67 2P3#177-183 RT: 7.19-7.36 AV:3 NL: 2.14E6 T: + c ESI Full ms [200.00-2000.00] 570.46 468.74 400 600 893.28 898.36 1 779.25 884.40 800 3.04 141 1464.17 1314.07 1086.69 1000 1200 m/z 1400 1628.33 1511.40 1673.72 1600 2P3#227-232 RT: 9.22-9.38 AV:3 NL: 2.82E6 T: + c ESI Full ms [200.00-2000.00] 1878.74 2P3#271-280 RT: 10.96-11.28 AV:5 NL: 1.11E6 T: + c ESI Full ms [200.00-2000.00] 1846.29 1800 2P3#247-252 RT: 10.03-10.19 AV:3 NL: 1.51E6 T: + c ESI Full ms [200.00-2000.00] 2000 Figura 7: Cromatogramas de estándares de PTX2SA, DTX1, PTX1, YTX y sus respectivos espectros m/z obtenidos por LC-MS. — 76 — RT: 0.00 - 29.98 SM: 9G 11.73 NL: 4.14 E6 m/z = 881.00-882.00 F: + c ESI Full ms [200.00-2000.00] MS 23C_060711145934 4000000 3500000 PTX2 3000000 Intensity 1.93 2500000 2000000 24.49 3.30 4.27 1500000 8.65 6.14 1000000 9.06 13.27 10.03 14.73 7.19 22.56 15.86 500000 16.42 18.68 0.63 19.17 19.57 20.54 21.02 27.55 23.85 26.34 25.46 28.43 0 0 2 4 8 6 10 16 14 Time (min) 12 18 20 22 24 26 1530.54 1605.91 28 23C_060711145934 #283-295 11.49-11.89 AV: 6 NL: 1.21E7 T: + c ESI Full ms [200.00-2000.00] 876.21 12000000 Intensity 10000000 8000000 6000000 877.22 881.49 4000000 2000000 297.02 420.27 556.21 758.46 691.12 883.55 1121.35 1192.73 1349.00 1743.01 1871.53 1974.79 0 200 400 600 800 1000 1200 1400 1600 1800 2000 m/z Figura 8: Ejemplo de cromatograma (arriba) del extracto metanólico de resina con PTX2, adsorbida después de permanecer una semana (del 21 enero 2006) en la columna de agua en la localidad de San Antonio. Abajo se presenta el espectro m/z respectivo obtenido por LC-MS. RT: 0.00 - 30.05 SM: 9G Intensity NL: 3.26 E6 m/z = 899.00-900.00 F: + c ESI Full ms [200.00-2000.00] MS 27C Intensity 27C # 176-181 RT: 7.11-7.27 AV: 3 NL: 2.38E6 T: + c ESI Full ms [200.00-2000.00] Figura 9: Ejemplo de cromatograma (arriba) del extracto metanólico de resina con PTX2SA, adsorbida después de permanecer una semana (noviembre 2005) en la columna de agua en la localidad de Toralla. Abajo se presenta el espectro m/z respectivo obtenido por LC-MS. — 77 — Crucero CIMAR 11 RT: 0.00 - 30.01 SM: 9G 1.92 NL: 2.89 E6 m/z = 841.00-842.00 F: + c ESI Full ms [200.00-2000.00] MS 19C_060711082530 2800000 2600000 2400000 2.73 2200000 2000000 DTX1 Intensity 1800000 1600000 1400000 1200000 24.77 5.48 5.15 1000000 26.61 27.10 15.58 6.53 11.76 11.13 12.10 13.72 10.16 800000 600000 23.48 17.19 14.93 0.71 400000 25.25 9.43 4.10 3.21 18.48 21.79 20.42 21.39 22.19 28.22 200000 28.78 0 0 2 6 4 Intensity 19C_060711082530 #233-237 RT: 9.43-9.52 T: + c ESI Full ms [ 200.00-2000.00] 8 10 12 16 14 Time (min) 18 20 22 26 24 28 30 AV: 2 NL: 3.99E6 2500000 Figura 10: Ejemplo de cromatograma (arriba) del extracto metanólico de DTX1 adsorbida por la resina tras permanecer una semana (del 14 de febrero de 2006) en la columna de agua en LIN. Abajo se presenta el espectro m/z respectivo obtenido por LC-MS. RT: 0.00 - 30.05 SM: 9G 12.08 NL: 9.83 E5 m/z = 1141.00-1142.00 F: - c ESI Full ms [200.00-2000.00] MS 28C 900000 2.45 800000 YTX 13.05 Intensity 700000 600000 1.64 500000 13.94 11.11 400000 2.93 20.07 5.93 300000 200000 4.07 100000 15.71 16.92 17.25 18.05 4.63 7.63 21.85 25.88 18.94 24.91 20.72 10.30 27.00 23.54 27.89 9.33 0 0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 Time (min) Rlative Abundance 28C #274-278 RT: 11.11-11.11 AV: 2 NL: 1.18E6 T: - c ESI Full ms [ 200.00-2000.00] Figura 11: Ejemplo de cromatograma (arriba) del extracto metanólico de resina con YTX (trazas), adsorbida después de permanecer una semana (noviembre 2005) en la columna de agua en la localidad de Puluqui A. Abajo se presenta el espectro m/z respectivo obtenido por LC-MS. — 78 — Figura 12: Microfotografías de Dinophysis acuminata (izquierda), D. acuta (centro) y D. rotundata (derecha) en muestras cualitativas recolectadas el 21-en-2006 en la estación San Antonio (QL3). — 79 —