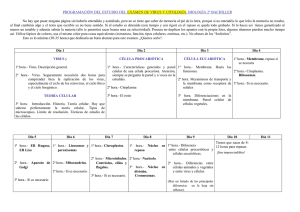
Temario Biología 2015-2016
Anuncio

BLOQUE I “LA BASE MOLECULAR Y FISICOQUÍMICA DE LA VIDA” 1.- BIOELEMENTOS Y BIOMOLÉCULAS La materia constituyente de los seres vivos está compuesta por moléculas, también denominadas biomoléculas, formadas a su vez por la unión de átomos de ciertos elementos químicos. Estos elementos presentes en las biomoléculas reciben el nombre de bioelementos o elementos biogénicos, de los que existen unos 70 diferentes. Los elementos biogénicos se unen por enlaces químicos para formar las moléculas constituyentes de los organismos vivos, que reciben el nombre de biomoléculas o principios inmediatos Atendiendo a su abundancia, que no a su importancia, los podemos agrupar en tres categorías: 1.1. BIOELEMENTOS PRIMARIOS O MAYORITARIOS. C, O, H, N. Constituyen más del 95% en masa del total de la materia viva. Las propiedades físico-químicas que los hacen tan adecuados para la vida son los siguientes: * Forman entre ellos con facilidad enlaces covalentes, compartiendo pares de electrones. * El C, N y O pueden compartir mas de un par de electrones, formando enlaces dobles o triples, lo cual les dota de una gran versatilidad para el enlace químico. * Son los elementos mas ligeros con capacidad de formar enlace covalente, por lo que dichos enlaces son muy estables. * Es particularmente significativa la capacidad del carbono para formar enlaces estables carbono-carbono, llegando a formarse largas cadenas carbonadas, que al unirse a diversos elementos químicos, confieren a los compuestos orgánicos una gran diversidad. * A causa de la configuración tetraédrica de los enlaces del carbono, los diferentes tipos de moléculas orgánicas tienen estructuras tridimensionales diferentes. * Los compuestos formados por éstos átomos en los organismos vivos se hallan en estado muy reducido. 1.2. BIOELEMENTOS SECUNDARIOS. S, P, Mg, Ca, Na, K, Cl. Los encontramos formando parte de todos los seres vivos, si bien en conjunto no superan, generalmente, el 4,5 % en masa total del organismo. Estudiaremos dos: 1.2.1. Azufre Se halla en dos aminoácidos (cisteína y metionina) presentes en casi todas las proteínas. También está presente en otras sustancias de interés biológico, como vitaminas del complejo vitamínico B y en la Coenzima A. Forma parte de los sulfatos, que son sales presentes en todos los seres vivos. 1.2.2. Fósforo. Es parte integrante de los nucleótidos, compuestos que forman parte integrante de los ácidos nucleicos y que constituyen sustancias de gran interés biológico, como muchos coenzimas ((NAD+, NADP+, etc). También forma parte de los fosfolípidos, sustancias fundamentales en la constitución de las membranas celulares. También forma parte de los fosfatos, sales minerales abundantes en los seres vivos. 1.3. OLIGOELEMENTOS. Se denomina de ésta forma al conjunto de elementos químicos que están presentes en los organismos de una manera vestigial (en conjunto, no representan mas allá del 0,5 % de la masa total del organismo. Pero no por eso dejan de ser importantes, pues su ausencia acarrea enfermedades conocidas como enfermedades carenciales; también un aumento de su contenido por encima de las cantidades requeridas por el organismo, suele provocar intoxicaciones. Se han podido aislar unos 60 oligoelementos en los seres vivos, pero solamente 14 de ellos pueden considerarse comunes para todos o gran parte de ellos, y reciben el nombre de oligoelementos esenciales: Fe, Mn, Cu, Co, Se, Zn, F, I, B, Si, V, Cr, Mo, Sn. Estudiaremos dos: 1.3.1. Hierro Es fundamental para la síntesis de la clorofila, actúa como catalizador en muchas reacciones químicas y forma parte de proteínas de funciones muy importantes, como los citocromos, que intervienen en la respiración celular, y la hemoglobina, que actúa como transportador de oxígeno. 1.3.2. Manganeso Interviene en la fotolisis del agua durante el proceso de fotosíntesis de las plantas. 2.- BIOMOLÉCULAS INORGÁNICAS 2.1. EL AGUA EN LOS SERES VIVOS. El agua es un componente muy importante de los seres vivos, ya que es la molécula que se encuentra en mayor proporción en todos los organismos (supone aproximadamente, entre el 50 y el 95% de su peso). La mayoría de las hipótesis actuales sobre el origen de la vida en la Tierra coinciden en aceptar que la vida, tal y como hoy la conocemos, se originó en el agua y las formas vivientes evolucionaron, en las primeras etapas, en el seno de los mares primitivos, Por otra parte, la conquista del medio terrestre sólo fue posible cuando algunos organismos consiguieron retener cierta cantidad de agua en sus estructuras corporales. En este sentido, el medio interno de los organismos terrestres no es otra cosa que un medio acuoso que permite el desarrollo de la vida celular en condiciones semejantes a las del medio marino donde surgió. 2.1.1. Estructura. La molécula de agua consta de un átomo de oxígeno y dos de hidrógeno. La unión entre el oxígeno y los hidrógenos se realiza mediante enlaces covalentes, en los que cada átomo de hidrógeno de una molécula comparte un par de electrones con el átomo de oxígeno. Sin embargo, como la electronegatividad (magnitud que mide la capacidad relativa de los átomos para atraer a los pares de electrones de los enlaces) del oxígeno es mayor que la del hidrógeno, los pares de electrones compartidos se ven atraídos con mas fuerza por el núcleo de oxígeno que por el de hidrógeno. Por otra parte, el oxígeno posee cuatro electrones mas sin compartir, lo que tiene dos consecuencias: * La geometría triangular que posee la molécula de agua, de manera que los átomos de hidrógeno forman respecto al oxígeno un ángulo de 104,5º. * La presencia de una carga negativa débil en la zona donde se sitúan los electrones no compartidos. Este último hecho, unido a la menor electronegatividad de los átomos de hidrógeno, crea una asimetría eléctrica en la molécula de agua que provoca la aparición de cargas eléctricas parciales opuestas (representadas por el símbolo ), de manera que la zona de electrones no compartidos del oxígeno es negativa y la zona donde se sitúan los hidrógenos es positiva. Por esta razón, y a pesar de ser eléctricamente neutra pues no presenta carga eléctrica neta, la molécula de agua tiene carácter dipolar. Esta polaridad favorece la interacción entre las moléculas de agua, de forma que la zona con carga eléctrica parcial negativa de una de ellas es atraída por la zona con carga parcial positiva de la otra, estableciéndose entre ambas un tipo de enlace denominado enlace o puente de hidrógeno, que convierte al agua en una sustancia altamente cohesiva, ya que cada molécula de agua puede establecer cuatro puentes de hidrógeno con otras tantas moléculas. También se pueden formar puentes de hidrógeno entre el agua y otras moléculas polares distintas (alcoholes, aminas, etc.) 2.1.2. Funciones del agua * Disolución. Debido a la polaridad de su molécula, el agua se puede interponer entre los iones de las redes cristalinas de los compuestos iónicos, lo que origina una disminución importante de la atracción entre ellos y provoca su separación y, en definitiva, su disolución. También puede formar puentes de hidrógeno con otras moléculas no iónicas, pero que tienen grupos polares (como muchas de las biomoléculas), y causar así mismo su disolución. Dado que las moléculas deben encontrarse disueltas en un medio líquido para reaccionar entre si, el agua desempeña un papel fundamental como medio donde tienen lugar las reacciones bioquímicas características de la actividad vital. La capacidad disolvente del agua es una de las propiedades mas importantes a la hora de justificar su protagonismo en los procesos biológicos, aunque también la hace muy susceptible a la contaminación por una gran variedad de sustancias, tanto naturales como procedentes de actividades humanas. * Termorregulación. El agua modera la temperatura interna de los seres vivos. El contenido en agua de los seres vivos amortigua las variaciones de su temperatura interna como consecuencia de cambios bruscos de la temperatura ambiental o por la producción de calor durante el metabolismo. Este efecto moderador favorece el desarrollo de las reacciones metabólicas. El agua necesita absorber mucho calor para aumentar 1ºC su temperatura. Del mismo modo, para que ésta descienda 1ºC, se ha de desprender de una gran cantidad de calor. El lento ascenso y descenso de la temperatura del agua se debe a su elevado calor específico. 2.2. LAS SALES MINERALES EN LOS SERES VIVOS. Los organismos vivos están también formados por sales minerales , compuestos inorgánicos que pueden ser solubles, se encuentran disociadas en iones, como es el caso del ClNa; o insolubles, se encuentran formando un precipitado que no se disocia, como por ejemplo el Ca3(PO4)2. En éste último caso, las sales se encuentran precipitadas y construyen estructuras sólidas en los seres vivientes, como esqueletos, caparazones o depósitos diversos. Lo mas normal, es que las sales minerales aparezcan disueltas y disociadas en sus iones componentes, aniones y cationes. 2.2.1. Funciones. * Estructural. Sales precipitadas. Los huesos, las conchas, los caparazones y las espículas de algunos organismos están formados por sales precipitadas, como fosfatos, carbonato cálcico e incluso sílice. Algunas inclusiones citoplasmáticas o los depósitos de algunas paredes celulares en órganos vegetales, como ciertas semillas y frutos, también están compuestos por sales minerales. * Reguladora. Sales disueltas. Contribuyen a la formación de las dispersiones del interior de los organismos. Entre los iones mas abundantes podemos citar: - Aniones: Sulfato, bicarbonato, fosfatos, nitratos y cloruros - Cationes: Na+, K+, Ca2+, Mg2+ * Osmosis. Todos los medios líquidos biológicos (sangre, plasma intersticial, líquido cefalorraquideo, etc) constituyen disoluciones de sales en agua de cuyo grado de concentración depende la estabilidad celular y la realización de algunas funciones fundamentales. Los procesos biológicos dependientes de la concentración de soluto (moléculas e iones) en agua se denominan osmóticos y tienen lugar cuando existen dos disoluciones de diferente concentración separadas por una membrana semipermeable que no deja pasar el soluto pero sí el disolvente. Se observa, entonces, el paso del disolvente (agua en los medios celulares) desde la disolución mas diluida (hipotónica o hipoosmótica) hacia la mas concentrada (hipertónica o hiperosmótica) a través de la membrana. Cuando el agua pasa a la disolución hipertónica, ésta se diluye, mientras que la disolución hipotónica se concentra al perderla. El proceso continua hasta que ambas disoluciones igualan su concentración, es decir, se hacen isotónicas o isoosmóticas. Para evitar el paso del agua sería necesario aplicar una presión, denominada presión osmótica, tanto mas intensa cuanto mayor fuera la diferencia de concentración entre ambas disoluciones. Como la membrana plasmática es semipermeable, es necesario mantener una concentración salina dentro de la célula igual a la del medio externo Para que ésta no tenga pérdida ni ganancia neta de agua. Si la concentración del medio intracelular es mayor que la del medio externo, la entrada excesiva de agua producirá un hinchamiento, conocido como turgencia celular, que puede provocar la rotura de la membrana y la muerte de la célula. Si, por el contrario, la concentración en el medio interno es menor que en el medio externo, la célula pierde agua y disminuye su volumen, proceso que recibe el nombre de plasmolisis y puede ocasionar también la muerte celular. La diálisis es un proceso relacionado con la ósmosis. En este caso, ademas del agua, la membrana semipermeable permite el paso de los solutos de baja masa molecular desde la disolución donde se encuentren en mayor concentración hasta la que los contiene en menor concentración. En realidad estas moléculas pasan por una simple difusión a favor de un gradiente de concentración. De ésta forma,si se tiene una disolución de dos tipos de sustancias, una de ellas de alta masa molecular y la otra de baja, se puede conseguir una separación parcial de ésta última. * Concepto de disolución tampón o “buffer”. Los organismos vivos no soportan variaciones del pH mayores de unas décimas de unidad, y por ello han desarrollado a lo largo de la evolución sistemas de tapón o “buffer” que mantienen el pH constante mediante mecanismos homeostáticos. Las variaciones del pH afectan en general a la estabilidad de las proteínas y, en concreto, influyen decisivamente en la actividad catalítica de los enzimas, pues presentan restos de aminoácidos que, en función del pH, pueden comportarse como compuestos ionizados o no, y generar cargas eléctricas que modifican profundamente la actividad biológica de la proteínas y demás compuestos biológicos. Los sistemas tampón, que tienden a impedir la variación del pH cuando se añaden cantidades moderadas de H+ o HO-, consisten en un par ácido-base conjugadas que actúan como dador y aceptor de protones, respectivamente. Las proteínas poseen gran capacidad tamponadora del pH, pero existen además otros tamponadores biológicos, como son el par carbónico-bicarbonato. Cada par conjugado presenta un pH determinado, en el cual la capacidad tamponadora es máxima. Este valor coincide en los tampones biológicos con el pH normal de los fluidos corporales, que suelen oscilar alrededor de 7. 3.- BIOMOLÉCULAS ORGÁNICAS 3.1. GLUCIDOS. Los glúcidos son moléculas orgánicas que contienen átomos de C, H, y O. A menudo se denominan azúcares, ya que muchos de ellos tiene sabor dulce, y también reciben el nombre de hidratos de carbono, porque su fórmula empírica es Cn(H2O)m parece corresponder a combinaciones de carbono con agua. Sin embargo, esta denominación resulta incorrecta, pues los glúcidos son polialcoholes con un grupo funcional carbonilo (grupo compuesto por un átomo de carbono y otro de oxígeno, unidos por un doble enlace, que constituye el grupo funcional de aldehídos y cetonas); es decir, constituyen polihidroxialdehídos o polihidroxicetonas. Esta composición química es común a todos los glúcidos. Se trata fundamentalmente, de moléculas energéticas, es decir, son utilizadas por los seres vivos como material para obtener energía, aunque también existen otros glúcidos no energéticos que cumplen diversas funciones. Se puede realizar una clasificación de los glúcidos atendiendo a los siguientes criterios: - Según el tipo de grupo funcional que poseen, se dividen en: º Aldosas: el grupo carbonilo es un aldehído º Cetosas: llevan un grupo cetona - Según su complejidad, se diferencian en : º Monosacáridos u osas. Son aldehídos o cetonas con dos o mas grupos hidroxilo (-OH). Comprenden los glúcidos mas simples y no pueden ser hidrolizados. º Osidos. Son moléculas mas grandes, formadas por la unión de varios monosacáridos. Pueden llevar a cabo reacciones de hidrólisis que liberan los monosacáridos componentes. Según el número de monosacáridos que presentan se clasifican en: Oligosacáridos. Compuestos por la unión de dos a nueve monosacáridos. Los mas importantes los disacáridos . Polisacáridos. Compuestos por un número elevado de monosacáridos. 3.1.1. Monosacáridos. Son los glúcidos más simples a partir de los cuales se forman los demás glúcidos. Generalmente se caracterizan por su sabor dulce y su solubilidad en el agua. La presencia del grupo carbonilo les confiere, además, propiedades reductoras. Para nombrarlos se antepone a la terminación –OSA, un prefijo que indique si poseen la función aldehído (ALDO-) o cetona (CETO-) y otro referente al número de átomos de carbono que posee la molécula: TRI (3 carbonos), TETRA (4 carbonos). PENTA (5 carbonos), HEXA (6 carbonos) etc. Los glúcidos, en general, y los monosacáridos, en particular, presentan una propiedad fundamental que se denomina estereoisomería. La estereoisomería consiste en la existencia de la molécula con la misma fórmula plana pero distinta estructura espacial. Esto sucede siempre que hay algún átomo de carbono asimétrico, es decir, un carbono que está unido a cuatro grupos distintos. Estos carbonos asimétricos son frecuentes en los glúcidos y se identifican en la molécula con un asterisco (*). Incluso en el glúcido mas sencillo, el gliceraldehído, se puede observar que el carbono 2 es asimétrico, pues se une a cuatro grupos diferentes Para representar en el plano del papel estas moléculas, se utilizan habitualmente la proyección de Fischer, en la que la cadena carbonada se dispone en vertical y los grupos unidos a los carbonos asimétricos se sitúan a la izquierda y a la derecha de éstos. Las proyecciones de Fischer resultan muy útiles sobre el papel, pero como ambos isómeros tienen la misma fórmula plana es preciso recurrir a modelos tridimensionales para diferenciarlos con claridad. Las disoluciones de estereoisómeros son capaces de desviar el plano de polarización a la derecha (isómero dextrógiro) o a la izquierda (isómero levógiro). En el primer caso se simboliza con el signo (+), y en el segundo, con el signo (-). Para nombrar los estereoisómeros de los monosacáridos se ha establecido el siguiente convenio: si al escribir la fórmula plana, según la proyección de Fischer, el grupo –OH del carbono asimétrico queda a la derecha y el grupo –H a la izquierda, el estereoisómero se denomina D. Si, por el contrario, el grupo –OH queda a la izquierda, el estereoisómero se llama L. No existe relación entre la actividad óptica, dextrógira o levógira, y el carácter D o L de un determinado estereoisómero. En otros glúcidos que contienen más carbonos asimétricos existe un número mayor de isómeros. En cada carbono asimétrico aparecen dos esteroisómeros, por lo que el número total de estereoisómeros en una molécula con n carbonos asimétricos será 2n. En éstos casos, la denominación D o L se establece por la posición del grupo –OH del último carbono asimétrico (D se sitúa a la derecha y L si está a la izquierda). Dentro de los estereoisómeros se pueden diferenciar aquellos que son imágenes especulares entre sí (denominados enantiomorfos o enantiomeros) y aquellos que no lo son (llamados epímeros). Triosas. Contienen tres átomos de carbono. Existen dos triosas: D-gliceraldehido y dihidroxiacetona. La dihidroxiacetona no presenta estereoisómeros, ya que no tiene carbonos asimétricos. Las dos triosas aparecen como productos intermedios en algunas reacciones metabólicas. Pentosas. Presentan cinco átomos de carbono. Algunas pentosas desempeñan funciones biológicas, como la D- ribosa (una aldopentosa) que es un componente fundamental de los ribonucleótidos que constituyen el RNA. Otra aldopentosa muy semejante, la desoxirribosa (sin grupo –OH en el carbono 2), se encuentra en los desoxirribonucleótidos que forman el DNA. Una cetopentosa, la D-ribulosa, participa en la fotosíntesis como la molécula sobre la que se fija el CO2 en la formación de moléculas orgánicas. Hexosas. Son monosacáridos que tienen seis átomos de carbono en su molécula. Presentan 24 esteroisómeros. Entre las aldohexosas destaca la glucosa, la molécula energética mas utilizada por los seres vivos. Se localiza en estado libre en el citoplasma celular, en el plasma sanguíneo (en concentraciones constantes) y en algunos frutos, como las uvas o los dátiles. Constituye, así mismo, la unidad componente de los polisacáridos mas comunes. Otra aldohexosa, la galactosa, se encuentra libre en la leche o formando parte de disacáridos como la lactosa (presente igualmente en la leche), de polisacáridos complejos y de ciertos glucolípidos. La fructosa, también denominada levulosa (por ser levógira), es una cetohexosa que se encuentra en estado libre en frutos y en algunos medios líquidos biológicos, como el semen, además de formar parte del disacárido sacarosa. * Formación de anillos en aldohexosas. La ciclación de las moléculas se produce al reaccionar el grupo aldehído del carbono uno con uno de los grupos hidroxilo (el carbono 5 en aldohexosas). Como consecuencia de ello se produce la pérdida de una molécula de agua y el establecimiento de un enlace hemiacetálico entre los dos carbonos que intervienen en la reacción. Además, la nueva estructura que aparece, llamada hemiacetal, transforma el carbono 1 en un nuevo carbono asimétrico que da lugar a dos estereoisómeros. Dicho carbono se denomina carbono anomérico y tiene un grupo hidroxilo, llamado –OH hemiacetálico. Los dos nuevos estereoisómeros reciben el nombre de anómeros y pueden ser y . Los anillos que se forman son heterocíclicos de 5 átomos de carbono denominados furanosas, o de seis átomos de carbono llamados piranosas. (Representaciones de Fischer). 3.1.2. Disacáridos Los disacáridos están formados por la unión de dos monosacáridos mediante un enlace O-glucosidico. Esta unión se denomina condensación o polimerización y se lleva a cabo al interaccionar dos grupos hidroxilo de dos moléculas distintas. Se produce entonces la liberación de una molécula de agua y la unión de los dos monosacáridos por el oxigeno de uno de los dos grupos hidroxilo implicados. Cuando los dos grupos –OH implicados en el enlace son anoméricos, el enlace se llama dicarbonilico; mientras que si uno de ellos no es anomérico, el enlace se llama monocarbonilico. En el primer caso, la molécula no posee poder reductor, pues éste deriva de la existencia de –OH anoméricos libres. En el segundo caso, el disacárido es reductor, ya que le queda un grupo anomérico libre, como los monosacáridos. Los disacáridos presentan las mismas propiedades que los monosacáridos: son solubles en agua, cristalizables, incoloros y tienen sabor dulce. Como disacáridos de interés biológico podemos citar los siguientes: * Sacarosa. Es el azúcar corriente de mesa. Se obtiene comercialmente de la remolacha o de la caña de azúcar. También está presente en numerosas frutas, como los dátiles y los higos, y en el néctar de muchas flores. Está formada por la unión de glucosa y fructosa. * Maltosa. Es el azucar de malta (cebada germinada); aparece cuando se descomponen los polisacáridos almidón y glucógeno por hidrólisis. Está formada por la unión de dos moléculas de -D-glucosa, por medio de un enlace O-glucosidico entre los carbonos 1 de la primera y 4 de la segunda; al quedar libre el carbono 1 de la segunda molécula de glucosa, en disolución acuosa puede sufrir mutorrotación y pasar a la forma -D-glucosa * Lactosa. Es el azúcar de la leche de los mamíferos. Está formada por -D-galactosa y D-glucosa que conserva libre el carbono anomérico, por lo que puede sufrir mutorrotación. * Celobiosa. Tampoco existe libre en la naturaleza, sino que aparece durante la descomposición del polisacárido celulosa. Está formada por dos moléculas de -D-glucosa unidas por un enlace o-glucosidico entre el carbono 1 de la primera y el 4 de la segunda. 3.1.3. Polisacáridos Están formados por largas cadenas de monosacáridos unidos mediante enlaces O-glucosidicos, por lo que son moléculas gigantescas o macromoléculas, estas cadenas pueden ser lineales o ramificadas. La formación de los polisacáridos a partir de los monosacáridos constituye un ejemplo de polimerización, en la que se libera una molécula de agua por cada enlace O-glucosidico formado. La rotura de éstos enlaces, pues, se realiza por hidrólisis. Al ser macromoléculas, los polisacáridos no se disuelven fácilmente en agua y pueden ser insolubles u originar dispersiones coloidales. Además, no son cristalinos ni tienen sabor dulce. Tampoco poseen carácter reductor, ya que no contienen carbonos anoméricos con grupos hidroxilo libres. Polisacáridos de reserva. - Almidón. Se encuentra en los amiloplastos de las células vegetales sobre todo en las semillas, las raíces y los tallos. También aparece en algunos protoctistas. El almidón se compone, en realidad, de dos moléculas: La -amilosa (cadena lineal) es un polímero de la -D-glucosa cuyas moléculas están unidas por enlaces O-glucosidicos entre el carbono 1 de una y el 4 de la siguiente, por lo que también se puede afirmar que se trata de un polímero de la maltosa. La cadena de amilosa adopta una conformación arrollada helicoidalmente que se empaqueta de forma irregular. La amilopectina (cadena ramificada) se diferencia de la anterior en que cada 25 ó 30 restos de glucosa aparece una ramificación de la cadena gracias a la formación de enlaces O-glucosidicos entre el carbono 1 de una glucosa y el 6 de la siguiente. La amilopectina tiene mayor masa molecular relativa que la amilosa, pudiendo llegar a contener más de 106 moléculas de glucosa. - Glucógeno Llamado también almidón animal, ya que constituye el polisacárido de reserva alimenticia de las células animales. Estructuralmente es muy similar a la amilopectina, pero con más ramificaciones, pues éstas aparecen cada 8 o 10 moléculas de glucosa de la cadena lineal. Polisacáridos estructurales - Celulosa. Es un polisacárido que constituye el principal componente de la pared de las células vegetales. Por eso se calcula que es el compuesto orgánico más abundante cuantitativamente del planeta: se calcula que cada año se sintetizan y degradan cerca de un billón de toneladas de celulosa en la biosfera. La celulosa tiene una estructura lineal constituida por unas 15.000 moléculas de D-glucosa unidas por enlace glucosidico (1-4), por lo que se puede decir también que es un polímero de la celobiosa. El enlace (1-4) confiere a la molécula una gran resistencia, y esa es una de las cualidades que han de tener las estructuras hechas a base de esta sustancia: las paredes de las células vegetales, cuyo componente principal es la celulosa, llegan a soportar 20 atmósferas de presión osmótica, y en las plantas leñosas, la celulosa es el principal componente de las estructuras de sostén. 3.2. LIPIDOS. 3.2.1. Lípidos saponificables. Todos los lípidos saponificables son ésteres y, por tanto, su hidrólisis produce el alcohol y un ácido caboxilico (A. Graso) * Acidos grasos. CH3-(CH2)n-COOH Los ácidos grasos son los componentes característicos de muchos lípidos y rara vez se encuentran libres en las células. Químicamente son ácidos orgánicos de cadena larga, que suelen tener un número par de átomos de carbono, generalmente entre 12 y 24. ENLACE ÉSTER: Radical ácido (COOH) + alcohol (-OH)……….Ester R-COOH + R´-OH……………. R´-COO-R´ + H2O En los animales y en las plantas superiores, los ácidos grasos mas abundantes son los que tienen 16 y 18 átomos de carbono, como son los ácidos palmítico, oleico, esteárico y linolénico. Las cadenas de los ácidos grasos pueden ser saturadas (no tienen dobles enlaces) o insaturadas (con uno o mas dobles enlaces). La longitud de la cadena carbonada y su grado de instauración, es decir, el número de dobles enlaces que contiene, influye de manera determinante en el punto de fusión de la molécula. Cuanto mas larga y saturada es la cadena, mayor es también el punto de fusión. Los compuestos totalmente saturados presentan cadenas extendidas que se pueden empaquetar estrechamente estableciendo interacciones de Van der Waals entre los átomos de las cadenas vecinas. Esto hace que a temperatura ambiente los ácidos grasos adquieran una consistencia cérea. Por el contrario, las cadenas insaturadas muestran angulaciones que no permiten un empaquetamiento tan fuerte como en las saturadas, por lo que las interacciones entre ellas son mas débiles y mas fáciles de desordenar por efecto de la energía térmica. Por esta razón, su punto de fusión es mas bajo y a temperatura ambiente son líquidos oleosos. Los ácidos grasos poseen una característica muy particular, su carácter anfipático. Al no tener grupos polares, la cadena carbonada resulta insoluble en el agua. Sin embargo, presenta en un extremo un grupo carboxilo (-COOH) que es claramente polar, lo que hace que en la molécula haya una pequeña parte soluble (hidrófila), llamada cabeza, y una porción larga, denominada cola, apolar e insoluble en agua (hidrófoba). Debido a ésta característica, al entrar en contacto con el agua, los ácidos grasos orientan las cabezas polares hacia ella mientras que las colas apolares se sitúan en la posición opuesta. Esto da lugar a la formación de estructuras esféricas (micelas) o en empalizada, en éste caso tanto en monocapas como en bicapas. En éstas últimas, las colas apolares se sitúan enfrentadas, quedando las cabezas polares hacia el exterior, en contacto con el agua. La disposición en monocapas se produce cuando los ácidos grasos se sitúan en la interfase entre el agua y un medio no polar (por ejemplo, el aire). Si los ácidos grasos se dispersan en el interior del agua se disponen en micelas o en bicapas. NOTA: Acidos grasos omega 3 (omega indica la posición del primer doble enlace contando desde el último carbono) Triacilglicéridos También se denominan triacilgliceroles, grasas o acilgliceridos. Están formadas por tres ácidos grasos unidos mediante enlace éster con el glicerol. Si los tres ácidos grasos son iguales se denominan triacilglicéridos simples. Cuando los ácidos grasos son diferentes reciben el nombre de triacilglicéridos mixtos. Las grasas naturales son mezclas de triacilglicéridos simples y mixtos. Las grasas sólidas (sebos, mantecas) poseen un punto de fusión superior a 40ºC, por lo que permanecen en ese estado a temperatura ambiente. En las grasas líquidas (aceites), por el contrario, el punto de fusión es inferior a 15ºC, y en las semisólidas (mantequillas, margarinas) se encuentran en un punto intermedio. Los triacilglicéridos pueden sufrir hidrólisis química o enzimática. - Hidrólisis química. Se lleva a cabo en la industria mediante tratamiento con álcalis (normalmente NaOH o KOH) sobre las grasas. Esta hidrólisis da glicerol y las sales sódicas o potásicas de los ácidos grasos, conocidas con el nombre de jabones. Debido a ello este proceso se conoce con el nombre de saponificación. El proceso inverso, es decir, la síntesis de estos lípidos mediante la unión del alcohol y el ácido, con liberación de una molécula de agua, se denomina esterificación. - Hidrólisis enzimática. Se realiza en los seres vivos mediante las enzimas lipasas dando lugar a glicerol y tres ácidos grasos. Este proceso se realiza en el tracto digestivo de los animales para digerir grasas ingeridas en la alimentación. Los triacilglicéridos desempeñan las siguientes funciones: - Son depósitos de energía química (metabólica) muy concentrada. - Proporcionan aislamiento térmico y físico (temperaturas bajas y protección de órganos). Céridos. Las ceras son moléculas estructurales que pueden encontrarse en algunos animales y, especialmente, en los vegetales. Son ésteres de ácidos grasos de cadena larga (de 16 a 36 átomos de carbono) con alcoholes también de cadena larga (de 16 a 30 átomos de carbono). En general son sólidas (poseen puntos de fusión de 60 a 100ºC) y totalmente insolubles en agua. Debido a su elevado grado de insolubilidad, las ceras desempeñan principalmente una función de protección, por lo que pueden aparecer recubriendo la superficie de órganos vegetales (frutos, tallos), como impermeabilizante de algunas estructuras tegumentarias de los animales (por ejemplo, las plumas de las aves) o en los panales construidos por las abejas. Fosfoglicéridos , Fosfolípidos o Glicerofosfolípidos. Son los principales componentes lipídicos de las membranas biológicas. El más sencillo es el ácido fosfatídico que está formado por el glicerol, dos ácidos grasos y ácido fosfórico. Los demás fosfolípidos tienen un compuesto polar, que contiene por lo menos un grupo amino o un grupo alcohol, unido al ácido fosfatídico. Los fosfolípidos se caracterizan, así mismo, por su comportamiento anfipático, es decir, tienen una parte polar (y, por tanto, soluble en agua) y otra apolar (insoluble). La primera corresponde a la porción dónde se sitúa el ácido fosfórico y el alcohol, mientras que la segunda corresponde al resto de la molécula (ácidos grasos). De ésta propiedad deriva su función biológica. Todas las membranas celulares están constituidas por una doble capa fosfolipídica en las que las colas apolares de ambas capas quedan enfrentadas entre sí, mientras que las cabezas polares se orientan hacia el medio externo e interno, ambos acuosos. Según el aminoalcohol presente, se diferencian varias clases de fosfolípidos: cefalina (o simplemente foosfatidiletanolamina) si el aminoalcohol es la etanolamina; lecitina (fosfatidilcolina) si contiene colina, y fosfatidilserina si tiene serina. Esfingolípidos Son componentes de las membranas celulares y, aunque, en general se encuentran en menor proporción que los fosfoglicéridos, abundan en el tejido nervioso. Al igual que ésos sus moléculas tienen carácter anfipático. Sos ésteres formados por la unión del alcohol esfingosina y un ácido graso mediante un enlace amida (Amina+grupo acilo=Amina ácida o Amida), que da lugar a una molécula llamada ceramida, a la que se une una molécula polar para constiruir un esfingolípido completo. Según la naturaleza de la molécula polar que se une a la ceramida se distinguen dos clases de esfingolípidos: - Esfingomielinas. Presentan una molécula de fosforilcolina o fosforiletanolamina (procendentes de la colina y etanolamina, respectivamente) unida a la ceramida. Son abundantes en las vainas de mielina que rodean los axones neuronales. - Glucoesfingolípidos o glucolípidos Su grupo polar es un glúcido. Estas moléculas desempeñan un papel fundamental como antígenos celulares, es decir, moléculas identificativas de células concretas que permiten el reconocimiento entre ellas. Los antígenos se relacionan con múltiples procesos, principalmente inmunitarios. Cuando el glúcido es un monosacárido (galactosa), los glucoesfingolípidos se denominan cerebrosidos mientras que si se trata de un polisacárido se llaman gangliósidos 3.2.2. Lípidos insaponificables Se incluyen en éste grupo los lípidos que no contienen ácidos grasos en su composición química (tampoco son ésteres) y, por tanto, no llevan a cabo la reacción de saponificación. Aunque se encuentran en los organismos vivos en menor cantidad que los lípidos saponificables, éste grupo comprende algunos compuestos biológicamente muy activos que desempeñan funciones importantes. Esteroides. Son derivados del hidrocarburo tetraciclico saturado denominado esterano o ciclopentanoperhidrofenantreno. Los esteroides se diferencian entre sí por el tipo y localización de sus grupos funcionales sustituyentes y por la presencia de dobles enlaces en los anillos. Un importante grupo de esteroides son los esteroles, que se caracterizan por tener un grupo alcohol (-OH) en el carbono 3 y una cadena hidrocarbonada en el carbono 17. Destaca entre ellos el colesterol que se encuentra en la membrana plasmática de las células animales, y unido a proteínas en el plasma sanguíneo. En otros eucariotas se encuentran esteroles de membrana similares, como por ejemplo, el estigmasterol en plantas y el ergosterol en hongos. En los animales el colesterol es el precursor de muchos esteroides, entre ellos: - Los ácidos biliares, cuya función es emulsionar las grasas en el intestino delgado, facilitando la acción de las lipasas y su posterior absorción. - La hormona sexual masculina, la testosterona, y las hormonas sexuales femeninas, el estradiol y la progesterona. - Las hormonas de la corteza suprarrenal, el cortisol y la aldosterona - La vitamina D, que regula el metabolismo del calcio y fosfato en vertebrados. 3.3. PROTEINAS 3.3.1. Aminoácidos: concepto y fórmula general Están formados por un átomo de carbono central, denominado carbono , al cual se unen un grupo carboxilo (COOH), un grupo amino (NH2) y un grupo R o cadena lateral variable según cada aminoácido. El carbono es un carbono asimétrico. Así pues, los aminoácidos presentan dos isómeros ópticos, uno de los cuales es imagen especular del otro y, por tanto, son enantiómeros. Si utilizamos como referencia el monosacárido gliceraldehido, los isómeros se clasifican en D y L. En los isómeros D, el grupo amino se sitúa a la derecha del carbono y en los isómeros L se localiza a la izquierda. Solamente los isómeros L constituyen las proteínas. En solución acuosa los aminoácidos suelen ionizarse y dan lugar a un ión dipolar, es decir, un ión con una parte cargada negativamente y otra con carga positiva. De éste modo, pueden actuar como ácidos y como bases. Las sustancias que presentan la capacidad de actuar como ácidos y como bases reciben el nombre de anfóteras. 3.3.2. Péptidos: formación del enlace peptídico Las proteínas son macromoléculas que resultan de la unión de aminoácidos. Esta unión se lleva a cabo mediante el enlace peptídico. Formalmente, se puede considerar como el resultado de la pérdida de agua entre el grupo carboxilo de un aminoácido y el -amino de otro. El conjunto de dos aminoácidos unidos por un enlace peptídico se llama dipéptido. Para péptidos formados por tres aminoácidos unidos por dos enlaces peptídicos se utiliza el nombre de tripéptido, y así sucesivamente. El nombre genérico de oligopéptido se emplea para designar un péptido de un número moderado de aminoácidos, sin especificar cuántos, y el de polipéptido designa, también sin especificar el número, al conjunto de muchos aminoácidos unidos por enlaces peptídicos. 3.3.3. Propiedades de las proteínas. Especificidad. A diferencia de otras moléculas como los glúcidos o los lípidos, las proteínas son moléculas específicas, es decir, cada especie biológica posee algunas proteínas que otros organismos no tienen. Incluso proteínas que presentan la misma función y una estructura tridimensional muy semejante suelen tener una secuencia peptídica algo diferente en distintos organismos. Este hecho posee una gran importancia, ya que el análisis de las semejanzas que existen entre algunas proteínas de diversos grupos de seres vivos permite llevar a cabo estudios filogenéticos y establecer el parentesco evolutivo entre especies. Por ejemplo, la hemoglobina es la proteína encargada del transporte de oxígeno en los eritrocitos de numerosas especies animales, pero en cada una de ellas tiene una secuencia de aminoácidos y una estructura tridimensional característica y, por tanto, sólo es funcional en los organismos en los que se ha sintetizado. Desnaturalización Se entiende por desnaturalización de una proteína un cambio estructural profundo, que normalmente conlleva la separación de las subunidades si las hay (pérdida de la estructura cuaternaria), pérdida de la estructura terciaria nativa y pérdida o grave alteración de la secundaria. Como la función de una proteína depende de su estructura, la desnaturalización conduce a la inactividad funcional de la proteína. Hay que tener en cuenta que la desnaturalización implica un cambio estructural extremo, ya que en muchos casos se dan cambios conformacionales mas suaves, sin que se altere radicalmente la estructura, que modulan la actividad de la proteína. La desnaturalización se puede producir por agentes físicos o químicos. El aumento de temperatura, por ejemplo, conduce a la desnaturalización de las proteínas. Entre las causas químicas de desnaturalización, la mas conocida es el cambio extremo del Ph. A valores muy bajos de Ph todos los aminoácidos ácidos están protonados y, por tanto, pierden su carga negativa. Análogamente, si el Ph es muy alto, los aminoácidos básicos pierden la carga positiva. 3.3.4. Niveles estructurales Estructura primaria Se denomina de esta forma la secuencia de los aminoácidos en la cadena polipéptídica. El número, tipo y orden o la secuencia de los aminoácidos que constituyen la estructura primaria son distintos en cada proteína. Siempre existe un extremo con un aminoácido cuyo grupo amino está libre y otro extremo con un aminoácido con su grupo carboxilo libre. Por convenio, los aminoácidos de la cadena se numeran comenzando por el que posee el extremo amino libre. En realidad la estructura primaria constituye una cadena de planos articulados, pues, como se ha visto, los enlaces peptídicos no pueden girar y los átomos de carbono, nitrógeno y oxígeno que participan en ellos se sitúan en un mismo plano. Estructura secundaria Aunque la disposición de la cadena polipeptídica es el espacio puede adoptar múltiples formas, existe una conformación mas estable que ninguna otra que, lógicamente, es la que se mantiene. Este plegamiento estable se denomina estructura secundaria, de la que existen dos tipos básicos. En las proteínas coexisten ambos, aunque uno de ellos puede predominar sobre el otro. Se ha comprobado que ciertas combinaciones de las dos estructuras son tan estables que se encuentran presentes en muchas proteínas, incluso con funciones distintas. Los dos tipos básicos de estructuras secundarias son la -hélice y la lámina plegada o lámina . - -hélice. Es te nombre alude a la -queratina, proteína muy abundante en las células de la epidermis, que presenta ésta característica estructura helicoidal. Consiste en un plegamiento en espiral de la cadena polipeptídica sobre sí misma. Este enrollamiento sigue el sentido de giro de las agujas del reloj y contiene 3,6 aminoácidos por cada vuelta. El plegamiento se mantiene estable por medio de puentes de hidrógeno entre el grupo –NH (que forma parte del enlace peptídico) de un aminoácido y el grupo –CO (que forma parte de otro enlace peptídico) del cuarto aminoácido que le sigue en la cadena lineal. Las cadenas laterales de los aminoácidos no intervienen en los enlaces y aparecen proyectadas hacia la parte externa de la -hélice. Debido a ésta falta de participación de las cadenas laterales, proteínas con estructuras primarias muy distintas pueden poseer la misma estructura secundaria. - Lámina plegada. También se conoce como estructura porque la queratina, presente en uñas, pelos y plumas, constituye un ejemplo característico. El plegamiento en éste caso, no origina una estructura helicoidal, sino una especie de fuelle o lámina plegada en zigzag, originada por el acoplamiento de segmentos de la misma cadena polipeptidica o de distintas cadenas, unidos entre sí por puentes de hidrógeno transversales análogos a los que estabilizan la -hélice. Las cadenas laterales (grupos R) de los aminoácidos se disponen alternativamente por encima y por debajo de esta estructura Estructura terciaria La estructura secundaria no es el último nivel de plegamiento de las proteínas, que adquieren una disposición espacial tridimensional estable característica. De la estructura terciaria depende la función de la proteína, por lo que cualquier cambio en la disposición de ésta estructura puede provocar la pérdida de su actividad biológica. La estructura terciaria es un conjunto de plegamientos característicos que se originan por la unión entre determinadas zonas de la cadena polipeptídica. Estas uniones se realizan entre las cadenas laterales R de los aminoácidos. Los enlaces pueden ser de cuatro tipos: - Puentes disulfuro. Constituyen fuertes enlaces covalentes entre dos grupos –SH que pertenecen a sendos aminoácidos cisteína. - Fuerzas electrostáticas. Se trata de enlaces de tipo iónico entre grupos con cargas eléctricas opuestas. Se producen, lógicamente, entre grupos R de aminoácidos ácidos (con carga negativa, -COO-) y aminoácidos básicos (con carga positiva, -NH+3) - Puentes de hidrógeno. Se establecen entre grupos polares no iónicos en los que existen cargas parciales en su cadena lateral. - Fuerzas de Van der Waals. Son las uniones mas débiles y se producen entre aminoácidos apolares. Estructura cuaternaria. En ocasiones existe otro nivel estructural por encima del anterior. Esto ocurre únicamente cuando la proteína está constituida por mas de una cadena polipeptídica, denominadas en éste caso subunidades proteícas. La estructura cuaternaria es, sencillamente, la disposición relativa que adoptan las subunidades proteícas entre sí. La unión entre ellas se realiza mediante los mismos tipos de enlaces que mantienen la estructura terciaria, establecidos esta vez entre las cadenas laterales de los aminoácidos de las distintas subunidades. 3.3.5. Funciones, indicando las proteínas más representativas. Estructural. Algunas proteínas confieren resistencia y fuerza a los tejidos. El colágeno forma parte de los huesos y los tendones; la -queratina constituye el pelo, las uñas y las plumas; y la elastina se encuentra en los ligamentos. De reserva Los aminoácidos obtenidos de la hidrólisis de las proteínas pueden ser utilizados en procesos de obtención de energía. La ovolabúmina es la principal proteína de la clara del huevo, y la lactoalbúmina de la leche. En la mayoría de las semillas de las plantas existen proteínas que se utilizan durante la germinación. De regulación. Algunas proteínas actúan en la regulación de procesos metabólicos. En éste grupo se encuentran algunas hormonas. La insulina es una hormona peptídica que favorece la absorción y utilización de la glucosa; el glucagón también es un péptido cuya función es estimular la degradación del glucógeno a glucosa en el hígado. Catalizadora. Los enzimas son las proteínas que controlan la velocidad de las reacciones que tienen lugar en los seres vivos. La lisozima cataliza la hidrólisis de los polisacáridos de la pared celular de algunas bacterias; la glucógeno-sintasa hace posible la síntesis de glucógeno a partir de glucosa. Defensiva. Existen proteínas que actúan defendiendo a los organismos de otros organismos patógenos. Las inmunoglobulinas o anticuerpos reconocen y neutralizan a los agentes patógenos que infectan el organismo. Transportadora. Algunas proteínas se unen a otras sustancias para transportarlas a los diferentes tejidos. La hemoglobina de la sangre transporta el oxígeno a los tejidos. Las lipopropteínas transportan lípidos. Contráctil. Existen proteínas que permiten a las células y los orgánulos contraerse y participar en distintos tipos de movimientos. La miosina y la actina permiten la contracción de los músculos; la tubulina forma los microtúbulos de los cilios y los flagelos. 3.4. BIOCTALIZADORES (ENZIMAS) Los enzimas son catalizadores biológicos que aceleran la velocidad de las reacciones químicas para que se aproximen al equilibrio (al menos, un millón de veces). Casi todas son proteínas globulares, aunque algunas moléculas de RNA (ribozimas) también muestran actividad catalítica. Los enzimas no alteran los equilibrios de la reacción química que catalizan, simplemente hacen que suceda mas deprisa. Las moléculas de enzima no se alteran al catalizar una reacción, sino que la misma molécula puede actuar varias veces. Una sola molécula de un enzima puede llegar a catalizar la reacción de decenas de miles de moléculas en un segundo. De ahí su eficacia a concentraciones muy pequeñas. Cada tipo de enzima es muy selectivo en relación con la reacción que cataliza. Su especificidad deriva de que cada enzima sólo puede reconocer, unirse y modificar a moléculas determinadas, que son los sustratos de la reacción. Todos los enzimas son proteínas, pero pueden estar formados únicamente por cadenas polipeptídicas o contener, además otro grupo no proteico. En éste último caso, la parte proteica recibe el nombre de apoenzima (da la especificidad), mientras que la parte no proteica se denomina grupo prostético cuando su unión al apoenzima es permanente, y cofactor cuando, como es habitual, no lo es. El apoenzima se encarga de proporcionar la estructura espacial específica que permite la unión a las moléculas sobre las que actúan los enzimas en las reacciones químicas y que se denominan sustratos. El cofactor o grupo prostético son los componentes enzimáticos que llevan a cabo la propia reacción, es decir, la catálisis en sentido estricto. El cofactor puede ser, sencillamente, un catión metálico, o una molécula orgánica, y en ese caso se llama coenzima. 3.4.1. Centro activo y actuación general de los enzimas El centro activo es una zona de la molécula formada por unos 10 aminoácidos especializados en la unión con los compuestos sobre los que actúan los enzimas. Estos compuestos se denominan sustratos. La unión del sustrato al enzima tiene lugar en ese sitio preciso: el centro activo. Esta formación del complejo enzima-sustrato no implica el establecimiento de enlaces covalentes, sino de interacciones iónicas, hidrofóbicas y puentes de hidrógeno. Hay por tanto una complementariedad entre centro activo y sustrato, de modo que si el sustrato posee carga negativa, en el centro activo suele haber aminoácidos con carga positiva en su cadena lateral, si en el sustrato hay átomos aceptores de puentes de hidrógeno, en el centro activo suele haber aminoácidos con grupos donadores en su cadena lateral. También debe existir una complementariedad estérica, es decir, que si el sustrato es voluminoso, el centro activo suele ser una cavidad con suficiente capacidad. Pero eso no significa que centro activo y sustrato tengan que ser exactamente complementarios. En un principio se pensó que el sustrato encajaría en el centro activo como una llave encaja en la cerradura. Esta suposición tenía algunas ventajas para explicar la especificidad de los enzimas, pero en la actualidad se prefiere admitir la idea de Koshland, según la cual el centro activo, que posee de antemano una cierta complementariedad con el sustrato, se adapta totalmente a él después de un primer contacto. La hipótesis de Koshland, que a menudo se conoce como ajuste inducido, tiene en cuenta la flexibilidad conformacional de las proteínas. Sería algo semejante a la introducción de la mano en un guante. La propia mano (que en ésta comparación equivaldría al sustrato) hace que el guante (equivalente al centro activo del enzima) se adapte mejor cuando se introduce en él. Los radicales de los aminoácidos del centro activo se unen al sustrato y consiguen debilitar sus enlaces provocando cambios energéticos que permiten alcanzar el estado de transición (estado en el que los enlaces de los reactivos están debilitados o rotos, pero aún no se han formado los nuevos). La unión es reversible, pues una parte del complejo enzima-sustrato (ES) se disocia: E + S ES Una vez formado el complejo enzima sustrato, el cofactor lleva a cabo la reacción y se obtiene le producto final (P): ES E + P El producto se libera del centro activo y la apoenzima queda libre para volver a unirse a nuevas moléculas de sustrato. 3.4.2. Inhibición enzimática: concepto y tipos. Los inhibidores enzimáticos son sustancias que disminuyen o anulan la actividad de un enzima. Se trata de un proceso inverso al de la activación. El enzima, previamente activo, disminuye su velocidad o incluso deja de actuar cuando aparece un inhibidor, el cual puede ser algún ión o también alguna molécula orgánica y, muy frecuentemente, el producto final de la reacción. La inhibición puede ser irreversible, cuando el inhibidor, que se denomina “veneno metabólico”, se une covalentemente al enzima alterando su estructura e inutilizándolo de forma permanente. La inhibición reversible, más común, tiene lugar cuando el enzima vuelve a tener actividad una vez eliminada la sustancia inhibidora. En éste caso, la unión del inhibidor con el enzima se realiza por enlaces no covalentes (iónicos o puentes de hidrógeno) mas fáciles de romper. Según el lugar de unión al enzima se diferencian dos tipos de inhibición reversible: competitiva y no competitiva. Inhibición competitiva. En ella, el inhibidor se une al centro activo impidiendo, por tanto la unión del sustrato. Existe una competencia entre ambos para ocupar el centro activo. Si éste es ocupado por el sustrato, la reacción se lleva a cabo, pero si es ocupado por el inhibidor no se produce, ya que el sustrato no puede entrar. El grado de inhibición dependerá de la proporción relativa entre sustrato e inhibidor. Para que exista este tipo de inhibición es necesario que el inhibidor se acople al centro activo, por lo que su forma espacial debe tener gran semejanza con la del sustrato. Por eso, a éstos inhibidores se les llama análogos metabólicos. Inhibición no competitiva. El inhibidor no compite con el sustrato, sino que se une en otra zona del enzima distinta del centro activo. Esta unión modifica la estructura del enzima al tiempo que dificulta el acoplamiento del sustrato. En ocasiones, el inhibidor se une al complejo enzima-sustrato, una vez creado éste, e impide la posterior formación del producto. Alosterismo. Existe una tercera forma de inhibición enzimática que la constituyen los enzimas alostéricos Estos enzimas están formados, en general, por varias subunidades llamadas protómeros, cada uno de los cuales posee un centro activo idéntico y otro centro llamado regulador, donde se acopla el inhibidor del enzima. Por otra parte, los enzimas alostéricos pueden adoptar dos estados según su conformación, llamados T y R. El estado T no muestra afinidad por el sustrato, por el contrario, el estado R la manifiesta muy alta. Estos estados son reversibles y en el mismo enzima no pueden existir los dos simultáneamente. Si en el centro regulador de algún protómero de la forma R se fija un inhibidor alostérico o modulador negativo, se pasa a la forma inactiva T. Inhibición feed-back. Puede ser considerada como una forma de efecto alostérico la llamada inhibición feed-back (retroalimentación). Consiste en que el metabolito final de una vía metabólica puede actuar como inhibidor del primer enzima de dicha vía, con lo cual dicho metabolito impide su propia formación. Este efecto se logra porque el metabolito final inhibidor es capaz de fijarse en el sitio regulador del primer enzima. 3.4.3. Clasificación. Según el tipo de reacción que catalizan, los enzimas se clasifican en las siguientes clases: Oxidorreductasas. Reacciones de oxidación-reducción con pérdida y ganancia de electrones. AH2 + B A + BH2 Las deshidrogenasas quitan H+ y electrones de una molécula; las oxidasas ceden electrones al oxígeno. Transferasas. Catalizan la transferencia de un sustrato a otro, de grupos funcionales o radicales distintos del hidrógeno. Transaminasas, transcarboxilasas, transmetilasas, etc. A-R + B A + B-R Hidrolasas. Catalizan reacciones de hidrólisis, introduciendo grupos OH y H. Carbohidrasas, esterasas, peptidasas, nucleasas. A + B A-OH + B-H Liasas. Catalizan reacciones de rotura y soldadura de sustratos, sin que intervenga el agua. Aminasas, carboxilasas, hidratasas, etc. C=A + B-H B.C-A-H Isomerasas. Transformación de un isómero en otro. A-B B-A Ligasas o sintasas. Unión de moléculas o de un grupo funcional a una molécula, utilizando la energía proporcionada por el ATP. 3.5. ACIDOS NUCLEICOS. Todos los organismos poseen unas moléculas que dirigen y controlan la síntesis de proteínas, proporcionando la información que determina su especificidad y características biológicas. Estas moléculas, que reciben el nombre de ácidos nucleicos, contienen las instrucciones necesarias para realizar los procesos vitales y son las responsables de todas las funciones básicas de los seres vivos. Podría decirse que los que un organismo es o puede llegar a ser, en términos biológicos, aparece “programado” en estas moléculas. 3.5.1. Nucleótidos. Los ácidos nucleicos son polímeros cuyas piezas elementales reciben el nombre de nucleótidos, los cuales, a su vez, están formados por la unión química de sustancias muy heterogeneas entre sí, que se enlazan siempre según una disposición determinada. Los nucleótidos, además de su función estructural, pueden desempeñar importantes funciones como coenzimas y como intermediarios en el metabolismo energético. Un nucleósido es una molécula constituida por la unión de una base nitrogenada y una pentosa. Las bases nitrogenadas son compuestos heterocíclicos con átomos de nitrógeno en el anillo y con carácter básico. Las bases presentes en los nucleosidos son de dos tipos: - Bases púricas. Derivadas del núcleo de la purina. En los ácidos nucleicos las bases púricas más abundantes son la adenina y la guanina. - Bases pirimidinicas. Derivadas del núcleo de la pirimidina, las mas abundantes son: citosina, timina y uracilo. Las pentosas, son monosacáridos de cinco átomos de carbono. A las bases nitrogenadas aparecen unidas dos posibles aldosas: la ribosa y la desoxirribosa. Para nombrar las estructuras de los nucleósidos, los átomos de las pentosas se numeran acompañando al número de una tilde. Las bases púricas se unen a las pentosas mediante un enlace N-glicosidico entre el carbono 1´ de la pentosa y el nitrógeno 9 de la base. Las bases pirimidinicas establecen el enlace N-glicosidico entre el carbono 1´ de la pentosa y el nitrógeno 1 de la base. Los nucleótidos son compuestos resultantes de la unión entre un nucleósido y moléculas de ácido fosfórico. Esta unión se realiza por esterificación de uno de los grupos alcohol de la pentosa, formándose un enlace éster entre la pentosa y el ácido fosfórico; los nucleótidos mas frecuentes son aquello en los que éste enlace se realiza sobre el carbono 5´ de la pentosa. Un nucleótido puede tener, una, dos o tres moléculas de ácido fosfórico enlazadas al carbono 5´ de la pentosa; por eso, al nombrar los nucleótidos se hace referencia al número de grupos fosfato que contiene. Se clasifican en desoxirribonucleotidos y ribonucleótidos según la pentosa y la base nitrogenada que los constituye. 3.5.2. ADN El ADN está formado por la unión de desoxirribonucleótidos, es decir, sus nucleótidos correspondientes contienen desoxirribosa. En cuanto a las bases nitrogenadas pueden ser, adenina, guanina, citosina y timina. Modelo de Watson y Crick. La secuencia polinucleotídica se dispone en el espacio en forma de una doble hélice, que presenta las siguientes características: - El ADN está constituido por dos cadenas polinucleotídicas unidas entre sí en toda su longitud. - Las dos cadenas son antiparalelas, lo que significa que el extremo 3´ de una de ellas se enfrenta con el extremo 5´ de la otra. - La unión entre las cadenas se realiza por medio de puentes de hidrógeno entre las bases nitrogenadas de ambas: concretamente, la adenina forma dos de éstos puentes con la timina y la guanina tres con la citosina. Resulta evidente que las dos cadenas no son idénticas, sino complementarias, ya que una de ellas tiene la secuencia de bases complementaria de la otra. - Las dos cadenas están enrolladas en espiral formando una doble hélice alrededor de un eje imaginario. Las bases nitrogenadas quedan en el interior de la doble hélice, mientras que los esqueletos pentosa-fosfato se sitúan en la parte externa. De ésta forma, las cargas negativas de los grupos fosfato se unen a las cargas positivas de cationes o de otras moléculas presentes en el medio, estabilizando la estructura. - Los planos de las bases nitrogenadas enfrentadas son paralelos entre sí y perpendiculares al eje de la hélice. - El enrollamiento de la doble hélice es plectonémico, es decir, las cadenas no se pueden separar sin desenrollarlas. - La doble hélice es dextrógira: el enrollamiento gira en el sentido de las agujas del reloj. - La anchura de la hélice es de 2nm, la longitud de cada vuelta es de 3,4 nm y cada 0,34 nm se encuentra un par de bases complementarias. Puede deducirse, por tanto, que existen 10 pares de nucleótidos por cada vuelta. 3.5.3. ARN El ARN, al igual que el ADN, es un polímero no ramificado, compuesto por una serie de nucleótidos unidos por enlaces éster fosfóricos. El ARN difiere del ADN en que la pentosa de los nucleótidos constituyentes es la ribosa, en lugar de la desoxirribosa, y en que las cuatro bases nitrogenadas mayoritarias presentes aparece el uracilo en lugar de la timina. El uracilo, como sucede con la timina, puede emparejarse con la adenina. Las cadenas de ARN son mas cortas que las del ADN y pueden aparecer tanto en el núcleo como en el citoplasma celular. Excepto en los reovirus, el ARN está constituido por una única cadena, aunque en el ARN transferente existe un apareamiento de bases intracatenario, lo que hace que aparezcan trazos bicatenarios, originados por el plegamiento de la cadena sobre sí misma. El ARN es la molécula encargada de sintetizar las proteínas específicas de un organismo realizando el mensaje genético codificado por el ADN. Así, mientras el ADN contiene la información, el ARN la utiliza para que se concretice en las proteínas específicas del individuo. ARN mensajero (ARNm). El ARNm es una copia de una parte del ADN (la que corresponde a cada gen o grupo de genes que vaya a expresarse) que será utilizada por los ribosomas como información para poder unir los aminoácidos en el orden adecuado y constituir una proteína concreta. Las cadenas de ARNm tienen una vida muy corta, ya que si no fueran destruidas (mediante enzimas ribonucleasas) la síntesis proteica se prolongaría indefinidamente y se originaría una superproducción de proteínas. Así, cuando se necesita sintetizar una proteína concreta, se fabrica de nuevo el ARNm correspondiente. El ARNm constituye, aproximadamente, entre el 3 y el 5% del total del ARN celular. ARN ribosómico (ARNr). Este ARN forma parte de los ribosomas y participa, por tanto, en el proceso de unión de los aminoácidos para sintetizar las proteínas. No constituye una molécula específica de cada organismo, pues, a diferencia del ARNm, no contiene información sobre la clase de proteínas que se va a sintetizar. Existen varias cadenas distintas de ARNr que se diferencian por su coeficiente de sedimentación (es una medida de la velocidad con que sedimenta una molécula u orgánulo cuando se somete a una fuerza centrífuga elevada). Las células procariotas y eucariotas tienen cadenas de ARNr ligeramente distintos. El ARNr es el tipo mas abundante, ya que corresponde al 80-85% del ARN celular total. ARN transferente (ARNt). Se encarga de transportar los aminoácidos presentes en el citoplasma celular hasta los ribosomas, donde se unirán para constituir las proteínas. Cada molécula de ARNt transporta un aminoácido especifico. Estas diferencias son debidas, fundamentalmente, a una secuencia de tres bases nitrogenadas, denominada anticodón, que varía entre los distintos ARNt. Los ARNt están formados por cadenas cortas (entre 70 y 90 nucleótidos) que contienen un 10% de bases nitrogenadas diferentes a las cuatro mayoritarias. Las moléculas de ARNt poseen una estructura secundaria muy característica, en la que existen tramos de doble cadena, por emparejamiento intracatenario. Estos tramos se denominan brazos y hay cuatro en cada molécula, aunque también puede aparecer un quinto brazo mas corto que los otros. En los extremos de tres de los brazos existen zonas sin emparejar que componen los denominados bucles. El extremo 3´ de la cadena tiene siempre la secuencia de bases CCA. A éste nucleótido terminal de adenina se une el aminoácido que va a ser transportado. En el extremo 5´ siempre existe un nucleótido de guanina con su grupo fosfato. El bucle 1 o brazo D, cuya secuencia es reconocida de manera específica por uno de los 20 enzimas, llamados aminoacil-ARNt sintetasas, encargados de unir cada aminoácido con su correspondiente molécula de ARNt. El bucle 3 o brazo TC, que actúa como lugar de reconocimiento del ribosoma. El bucle 2 situado en el extremo del brazo largo que contiene una secuencia de tres bases llamada anticodón. Cada ARNt “cargado” con su correspondiente aminoácido se une, mediante la región del anticodón, con tripletes de bases del ARNm (cada tres bases de ARNm definen un triplete o códón) en el proceso de la traducción del código genético que conduce a la síntesis de proteínas. BLOQUE II “MORFOLOGÍA, ESTRUCTURA Y FUNCIONES CELULARES” 1. LA TEORÍA CELULAR 1.1 LA CÉLULA: CARACTERÍSTICAS GENERALES Hace aproximadamente unos 350 años, concretamente en 1665, el inglés Robert Hooke, examinando al microscopio una laminilla de corcho, observó que estaba formada por pequeñas cavidades poliédricas a las que denominó células, aludiendo con ello al parecido con las celdillas de un panal. Ahora bien, el verdadero conocimiento de la célula y todo lo que ella representa, ha sido el resultado de una suma de descubrimientos sucesivos debidos a gran número de científicos. Ocupa un lugar destacado el botánico Brown (1831), que descubre el núcleo. En 1839 el fisiólogo Purkinje introduce el término de protoplasma para designar el liquido que llenaba la célula. En 1861 el alemán Max Schultze definía la célula como “un grumito de protoplasma en cuyo interior hay un núcleo”. Todos estos hallazgos suponen el pie donde descansan los trabajos posteriores de Schleiden y Schwan (botánico y zoólogo respectivamente) que les llevaron a determinar la teoría celular de los seres vivos que se resume en los siguientes puntos: Todos los organismos vivos están compuestos por células La célula es la unidad estructural y fisiológica de los seres vivos Las células constituyen las unidades básicas de la reproducción: cada célula procede de la división de otras preexistentes, siendo idéntica a estas genética, estructural y funcionalmente. La célula es la unidad de vida independiente mas elemental. 1.2.- MODELOS DE ORGANIZACIÓN CÉLULAR 1.2.1. PROCARIOTAS, EUCARIOTAS (ANIMAL Y VEGETAL) Se caracteriza porque carece de verdadero núcleo aislado por una membrana nuclear; su material cromosómico está formado por una doble hélice de DNA “desnudo”, que no se asocia con proteínas del tipo de las histonas, y se localiza en una región central, denominada nucleoide, sin separación del resto de protoplasma. Este se presenta poco diferenciado y, aunque existe un conjunto de sistemas membranosos formados por repliegues internos de la membrana plasmática, no se alcanza un alto grado de compartimentación. Contiene ribosomas, a veces posee flagelos, cromatoforos con actividad fotosintética, inclusiones lipídicas y todo el equipo enzimático que permite a éstos organismos comportarse como aerobios o anaerobios, fotosintéticos o quimiosintéticos, según los casos. La célula eucariota animal se caracteriza porque su protoplasma manifiesta un alto grado de complejidad y de organización; el núcleo está separado del citoplasma mediante una membrana nuclear y contiene el DNA asociado a las histonas con el fin de permitir su mejor empaquetamiento. El citoplasma contiene numerosos orgánulos y sistemas de membranas, como las mitocondrias, el R.E. etc que establecen una red de compartimentos celulares interrelacionados, que se reparten el trabajo metabólico, lo que permite mayor especialización y mas eficacia en el desempeño de sus funciones. La célula eucariota vegetal contiene los mismos orgánulos que la animal, con la diferencia de que posee una gruesa membrana de secreción de naturaleza celulósica y cloroplastos. No presenta centriolo y tiene gran número de vacuolas. * FORMA DE LA CÉLULA ANIMAL Y VEGETAL La forma celular es extraordinariamente variable, difícilmente se puede dar una forma general, podríamos, si cabe, hablar de una primitiva forma redonda que al irse especializando se ha adaptado a cada función especifica. Hay muchas clases distintas de células. Dentro de nuestro propio cuerpo hay mas de 100 tipos distintos de ellas. En una cucharilla de café con agua de un estanque se pueden encontrar cientos de ellas claramente distintos. Los vegetales están compuestos por células que superficialmente son bastante distintas a las de nuestro cuerpo; los insectos tienen muchas clases de células que no se encuentran en los vegetales ni en los vertebrados. De éste modo una de las características de las células es su diversidad. Generalmente las formas de vida libre son esféricas debido a la tensión superficial. En los tejidos celulares, al modificarse mutuamente sus membranas, adquieren forma poliédrica. Las hay estrelladas (C. nerviosas); fibrilares (musculares); planas (epitelios)..... * ENVUELTAS CELULARES + Pared celular. Las células vegetales poseen una envuelta externa a la membrana plasmática, altamente organizada y rígida, que constituye la pared celular y cumple las siguientes funciones: Confiere rigidez al vegetal y contribuye al mantenimiento de la forma celular Une las células adyacentes, conectando las células de los tejidos vegetales Posibilita el intercambio de fluidos y la comunicación intercelular Permite a las células vegetales vivir en el medio hipotónico de la planta, impidiendo que éstas se hinchen y lleguen a estallar Impermeabiliza la superficie vegetal en algunos tejidos gracias a la cutina y suberina para evitar las pérdidas de agua Sirve de barrera al paso de agentes patógenos La pared celular tiene dos componentes principales: Un componente cristalino, constituido por fibras de celulosa Un componente amorfo, formado por una matriz de pectinas, hemicelulosa, agua, sales minerales y, en algunos casos, ligninas En las células diferenciadas, la pared celular aparece como una estructura gruesa compuesta por varias capas que se van depositando a medida que madura la célula. Estas capas son: * Lámina media. Es la capa más externa y la primera que se forma después de la división celular, y puede ser compartida por las células adyacentes de un tejido. Está integrada por pectinas y proteínas, que se unen posteriormente a iones de Ca2+. * Pared primaria. Se trata de una gruesa capa de estructura fibrilar, situada por debajo de la lámina media hacia el interior de la célula. Está constituida, fundamentalmente, por largas fibras de celulosa cohesionadas por polisacáridos (hemicelulosa y pectinas) y glucoproteínas. * Pared secundaria. Es la capa mas interna y se encuentra por debajo de la pared primaria en algunos tipos especiales de células vegetales (t. De soprte o vasculares). Consta de una o varias capas fibrilares, semejantes en composición a la pared primaria, aunque contienen celulosa en mayor proporción y carecen de pectinas. En estas capas fibrilares, las miofibrillas de celulosa se ordenan paralelamente dando lugar a varias capas con diferente ordenación. En ocasiones entran a formar parte de su composición polímeros, como la lignina (xilema y esclerénquima), ceras y cutina (haz de las hojas) o suberina (corcho). En algunas paredes se observan también inclusiones minerales, principalmente carbonatos y sílice. La pared celular de las células vegetales no es continua, ya que presenta una serie de canales, los plasmodesmos, que permiten la comunicación y el intercambio de líquidos con sustancias disueltas y otras moléculas entre células adyacentes. Estos canales se forman en la división celular durante la síntesis de la pared, y aparecen en todas las células jóvenes. Los plasmodesmos pueden encontrarse en ocasiones en depresiones de la pared primaria (menos engrosada), en la que también se inhibe el depósito de pared secundaria. En éste caso se llaman punteaduras. + Membrana celular. La membrana plasmática es una estructura que rodea y limita completamente a la célula y constituye una “barrera” selectiva que controla el intercambio de sustancias desde el interior celular hacia el medio exterior circundante, y viceversa. La membrana plasmática posee la misma estructura en todas las células. En corte ultrafino aparece como dos bandas oscuras separadas por una banda clara. Esta organización es común, además, al resto de las membranas biológicas constituyentes o limitantes de los orgánulos celulares, por lo que se denomina unidad de membrana (o membrana unitaria). La estructura trilaminar observada en la unidad de membrana se corresponde con una bicapa lipídica con proteínas embebidas. Los lípidos se disponen en una bicapa con las zonas hidrófilas (grupos polares) hacia fuera, mientras que las zonas hidrófobas quedan enfrentadas hacia el interior: Las membranas presentan, por tanto, dos caras: una cara externa y una cara interna que, en el caso de la membrana plasmática, está en contacto con el citoplasma celular. Las proteínas pueden estar asociadas a la cara interna o externa, o ser transmembranales. Su composición es la siguiente: * Lípidos de membrana. Pertenecen fundamentalmente a tres categorías: fosfolípidos, glucolípidos y esteroles. Fosfolípidos: Son los lípidos mas abundantes en las membranas biológicas: presentan una zona hidrófila, que constituyen las denominadas cabezas polares (glicerina o glicerol en los fosfogliceridos), y una zona hidrófoba (ácidos grasos), que forman la cola apolar. Los fosfolípidos poseen, por tanto, un carácter anfipático. La estructura que forman los fosfolípidos no es rígida, sino que estas moléculas pueden presentar movimientos: - Difusión lateral: un fosfolípido intercambia su posición con otro situado en la misma monocapa, desplazándose lateralmente - Rotación de fosfolípidos las moléculas giran alrededor de su eje longitudinal - Flexión de las cadenas hidrocarbonadas: los fosfolípidos aumentan o disminuyen el grado de separación de las colas hidrofóbicas. - Flip-flop: un fosfolípido se desplaza verticalmente y ocupa un lugar en la monocapa opuesta. Este tipo de movimiento es muy poco frecuente. Glucolípidos: Son muy semejantes a los fosfolípidos, pero contiene oligosacáridos. En las células animales suelen ser derivados de esfingolípidos. En las células vegetales y procariotas, sin embargo, derivan de los fosfoglicéridos. Sólo aparecen en la cara externa de la membrana plasmática. Esteroles: Derivados del colesterol y presentes en la membrana plasmática de la célula eucariota, son mas abundantes, por lo general, en las células animales. Raramente aparecen en bacterias desprovistas de pared celular. El colesterol se encuentra en proporción elevada en las membranas de las células eucariotas. Las moléculas de colesterol se disponen intercaladas entre los fosfolípidos. Esta disposición limita la velocidad de los fosfolípidos, lo cual da estabilidad a la membrana y también reduce su permeabilidad * Proteínas de membrana. Las proteínas asociadas a la membrana pueden cumplir un papel meramente estructural, funciones de reconocimiento y adhesión, o bien estar implicadas en el transporte y el metabolismo celular. Según su grado de asociación a la membrana se clasifican en dos grupos: Integrales: Estas proteínas se asocian a la membrana mediante enlaces hidrófobos. Sólo pueden separase de la membrana si se destruye la bicapa. Dentro de éste grupo existen proteínas transmembranales y proteínas asociadas a la cara externa o a la cara interna de la membrana. Algunas proteínas presentan hidratos de carbono unidos a ellas covalentemente (glucoproteínas) y se disponen siempre en el lado externo de la membrana, como los glucolípidos. Periféricas: Son proteínas unidas a la membrana por enlaces de tipo iónico y se separan de ella con facilidad. Aparecen principalmente en la cara interna de la membrana. En éste grupo no existen proteínas transmembranales. Las proteínas de membrana presentan una cierta capacidad de movimiento: - Difusión lateral, a lo largo de la membrana - Rotación, alrededor de un eje perpendicular a la membrana. La cara citoplasmática y la cara externa de la bicapa lipídica no son iguales, sino que presentan algunas diferencias en su composición química. Estudios realizadoe en eritrocitos humanos han puesto de manifiesto las siguientes diferencias: * Por la cara externa predominan los fosfolípidos fosfatidilcolina y y esfingomielina, mientras que por la cara interna los mas abundantes son la fosfatidilserina y la fosfatidiletanolamina. * En la cara citoplasmática predomina una carga electroquímica negativa, por la abundancia de fosfatidilserina. * Los ácidos grasos de los fosfolípidos de la cara citoplasmática presentan más dobles enlaces que los de la cara externa y por ello están mas curvados. Por tanto, ésta cara de la membrana es más fluida que la externa. * Solo se encuentran glicolípidos en la cara externa de la bicapa. Debido a esto se dice que la membrana es asimétrica. Es frecuente que las células eucariotas presenten asociada a la parte exterior de la membrana plasmática una capa denominada glicocaliz, que está compuesta por cadenas de glúcidos unidas a las proteínas de membrana, a los fosfolípidos de la cara externa, o bien a ambos. La membrana mantiene la integridad estructural de la célula; pero además controla dos funciones básicas: El transporte de sustancias a su través Los contactos entre las células + Funciones de la membrana celular - Barrera selectiva. Regula el transporte y el intercambio de sustancias entre el citoplasma y el exterior de un modo selectivo, es decir, se trata de una membrana semipermeable. - - - Producción y control de gradientes electroquímicos. En la membrana se sitúan las proteínas y cadenas de transporte que regulan el intercambio de sustancias. Intercambio de señales. Detecta y transmite señales con el medio externo o con otras células. División celular. Está implicada en el control y desarrollo de la división celular o citocinesis. Adhesión.La membrana presenta proteínas que facilitan la unión y comunicación entre células adyacentes en organismos pluricelulares. Endocitosis y exocitosis 1.2.2. TRANSPORTE DE SUSTANCIA A TRAVÉS DE MEMBRANA. Para poder llevar a cabo todas las funciones metabólicas que tienen lugar en las células, éstas deben intercambiar sustancias con su medio exterior. Existen varios mecanismos de transporte y pueden ser útiles para moléculas pequeñas o bien para macromoléculas y partículas. + Transporte de moléculas pequeñas. Dentro de éste mecanismo de transporte distinguiremos entre transporte pasivo y transporte activo * Transporte pasivo. Es un tipo de transporte que no requiere gasto de energía y presenta dos modalidades: * Difusión simple. Ciertas moléculas pequeñas y sin carga electroquímica, como el oxígeno, el nitrógeno, el dióxido de carbono y también el agua, se difunden rápidamente a través de la bicapa lipídica, a favor de su gradiente de concentración. El paso tiene lugar aprovechando los huecos que aparecen por el desplazamiento de los fosfolípidos. * Difusión facilitada. Las moléculas que no pueden cruzar a través de la bicapa lipídica, como glúcidos, aminoácidos, iones....pasan a través de la membrana, a favor de gradiente electroquímico o de concentración, mediante proteínas transmembrana, que pueden ser: - Proteínas de canal, que forman poros acuosos por los cuales pasan determinados solutos. Algunos de éstos canales se abren mediante interacción con un ligando y se denominan canales regulados por ligando; otros se abren en respuesta a un cambio del potencial de la membrana y se denominan canales regulados por voltaje - Proteínas transportadoras, que se unen a la sustancia que se debe transportar y sufren un cambio de conformación para permitir el paso de ésta a través de la membrana. * Transporte activo. Tiene lugar en contra del gradiente de concentración o del gradiente electroquímico de las sustancias y por ello precisa energía. Este tipo de transporte se produce mediante proteínas transmembrana que hidrolizan ATP para obtener energía y, al mismo tiempo, alteran su conformación espacial para efectuar el transporte de las sustancias. Es el caso de las bombas de iones, como la de sodio-potasio: - Se incorpora un Na+ y se produce la hidrólisis de una molécula de ATP; la unión del fósforo provoca el cambio en la conformación de la proteína. - El cambio de conformación libera el Na+ en el exterior de la célula y la proteína queda en condiciones de aceptar K+. - La entrada del K+ produce la liberación del P, así se vuelve a la conformación inicial, se libera el K+ y la proteína puede aceptar Na+. + Transporte de macromoléculas y partículas. Los mecanismos anteriores no permiten el paso de moléculas grandes como polisacáridos o proteínas. En estos casos se utiliza otro tipo de sistemas de transporte: la exocitosis y la endocitosis. *Exocitosis. Tiene lugar cuando una macromolécula o una partícula debe pasar del interior al exterior de la célula. Las vesículas que se forman en este caso reciben el nombre de vesículas secretoras. Se desplazan hacia la membrana plasmática, se fusionan con ella y vierten su contenido al medio extracelular. Según el modo en que tiene lugar la secreción se distingue dos tipos de exocitosis: - Las vesículas se producen de manera regular en la célula y su contenido es vertido al exterior sin necesidad de que exista un estímulo. Es el caso de las proteínas que forman la matriz extracelular de los tejidos. - Las vesículas se producen y liberan su contenido al exterior sólo cuando la célula es estimulada por alguna señal extracelular. Un ejemplo de ello es la secreción de neurotransmisores en las neuronas. *Endocitosis. Es un proceso de incorporación de sustancias a la célula. En algunos casos, se ha observado que estas sustancias se concentran en pequeñas depresiones formadas en la superficie exterior de la membrana. Cuando se produce la invaginación y se forma la vesícula, ésta contiene una cantidad de sustancia superior a la que penetraría en la célula sino se produjera la concentración previa. - Cuando las vesículas alcanzan un tamaño superior a 150 nm y contienen partículas grandes, como restos celulares y microorganismos, se habla de fagocitosis. Se da en células con cierto grado de especialización, como ciertos protozoos. - Si las vesículas son de diámetro inferior a 150 nm y contienen fluidos, el proceso denomina pinocitosis. Es muy común en todo tipo de células. 1.2.3. CITOPLASMA Dentro del citoplasma celular está el citosol que es el medio acuoso en el que se encuentran inmersos los orgánulos celulares. Representa aproximadamente la mitad del volumen celular. Contiene una gran variedad de filamentos proteicos que le proporcionan una compleja estructura interna. El conjunto de estos filamentos constituye el citoesqueleto. + Citoesqueleto. Todas las células eucariotas tienen un esqueleto interno, el citoesqueleto, formado por una compleja red de filamentos proteicos que se extiende por todo el citoplasma. Las diversas funciones del citoesqueleto dependen sólo de tres tipos principales de fibras proteicas: Microfilamentos o filamentos de actina, microtúbulos y filamentos intermedios. * Microfilamentos o filamentos de actina. Están formados por proteínas como la actina y la miosina, que peden tener estructura globular o fibrosa. Los microfilamentos intervienen en las siguientes funciones: - Intervienen en la contracción muscular. En las células musculares los filamentos de actina se asocian con la miosina y otras proteínas para producir la contracción muscular - Intervienen en la locomoción celular y en la fagocitosis, mediante la formación de pseudopodos. Los pseudopodos son prolongaciones dinámicas de la superficie celular que contienen actina; en el interior de los pseudopodos los filamentos de actina se disponen de forma ordenada, con sus extremos en crecimiento hacia el exterior. - Producen corrientes citoplasmáticas o de ciclosis. En las células vegetales, las corrientes citoplasmáticas hacen girar el citoplasma alrededor de su vacuola central. - Forman el anillo contráctil. Durante la división celular, se forma alrededor del ecuador de la célula, justo por debajo de la membrana plasmática, un haz de filamentos de actina y miosina, conocido con el anillo contráctil. El estrechamiento del anillo conducirá a la separación de las células hijas. - Mantienen la estructura de las microvellosidades. Así, en el interior de las microvellosidades de las células del epitelio intestinal hay un haz muy organizado de filamentos paralelos de actina que mantienen su estructura rígida. - Contribuyen a reforzar la membrana plasmática. Muchas células animales poseen una densa red de filamentos de actina, justo por debajo de la membrana plasmática. Esta red constituye la corteza celular que da fuerza mecánica a la superficie de la célula y le permite cambiar de forma y moverse. * Filamentos intermedios. Son fibras proteicas resistentes que desempeñan una función estructural o mecánica en la célula. Son muy abundantes en los lugares donde las células están sometidas a tensiones mecánicas, como es el caso de los epitelios, a lo largo de los axones y en el citoplasma de las células musculares lisas. Hay muchos filamentos intermedios, ya que varían según el tipo celular. Entre ellos están: - Filamentos de queratina. En las células epiteliales. Estos filamentos forman una densa red en el interior de las células epiteliales y proporcionan a dichas células una gran parte de su fuerza mecánica. Muchos filamentos de queratina están anclados en regiones especializadas de la membrana plasmática (desmosomas). - Neurofilamentos. Se encuentran en los axones y dendritas de las neuronas. Estos filamentos intermedios están en estrecha asociación con los microtúbulos axonales y la resistencia y rigidez del axón podría deberse a estos complejos de neurofilamentos y microtúbulos. - Las laminas nucleares. Forman una lámina fibrosa en la superficie interna de la envoltura nuclear de todas las células. * Microtúbulos. Son cilindros huecos que están formados por moléculas de tubulina. Son los principales componentes del citoesqueleto de las células eucariotas. Pueden encontrarse dispersos por todo el citoplasma o bien formar estructuras estables tales como cilios, flagelos y centriolos. - Cilios y flagelos. Son prolongaciones móviles presentes en la superficie de muchos tipos celulares. Su función es permitir el desplazamiento de una célula aislada a través de un líquido o desplazar el líquido extracelular sobre la superficie de la célula. Los cilios son cortos y numerosos mientras que los flagelos son largos y escasos. Ambos presentan la misma estructura pero diferente tipo de movimiento. En un cilio o flagelo se pueden distinguir las siguientes partes: º Eje o axonema. Está rodeado por la membrana plasmática y en su interior tiene 2 microtúbulos centrales y 9 pares o dobletes de microtúbulos periféricos; todos ellos orientados paralelamente al eje principal del cilio o flagelo. Los dos microtúbulos centrales son completos y están rodeados por una delgada vaina. Cada doblete periférico está formado por un microtúbulo a completo y un microtúbulo b incompleto. El microtúbulo a presenta, a intervalos regulares, dos brazos formados por una proteína, la dineina, que se dirigen hacia el microtúbulo b del par adyacente. Cada doblete se une al adyacente mediante puentes formados por otra proteína, la nexina. Existen también fibras radiales que conectan cada doblete periférico con la vaina central. º Zona de transición. Corresponde a la base del cilio o flagelo. El par de microtúbulos centrales se interrumpen y en su lugar aparece la placa basal. º Corpúsculo basal. Está situado justo por debajo de la membrana plasmática y presenta la misma estructura que los centriolos. - Centriolos. Son estructuras formadas por nueve grupos de tres microtúbulos (tripletes). A excepción de los vegetales superiores todas las células eucariotas tienen un par de centriolos que forman parte del centrosoma. El centrosoma o centro celular se localiza al lado del núcleo y está formado por dos centriolos (diplosoma) dispuestos perpendicularmente entre sí y rodeados por un material amorfo, denominado material pericentriolar. Los microtubulos citoplasmáticos tienden airradiar en todas direcciones a partir del centrosoma. En general las funciones que realizan los microtúbulos son las siguientes: - Son los principales elementos estructurales y generadores del movimiento de los cilios y flagelos. - Dirigen el transporte de orgánulos en el citoplasma - Constituyen el huso mitótico - Determinan la forma de la célula - Determinan la localización celular del retículo endoplásmico y del complejo de Golgi - Intervienen en la organización de todos los filamentos del citoesqueleto. + Lisosomas. Son vesículas rodeadas de membrana que contiene enzimas hidrolíticas encargadas de las digestiones intracelulares. Contienen alrededor de unas 40 enzimas diferentes, todas ellas hidrolasas ácidas, cuyo pH óptimo es próximo a 5. Entre ellas están proteasas, nucleasas, glicosidasas, lipasas, fosfatasas, sulfatasas y fosfolipasas. La membrana lisosomal es resistente a la acción de los enzimas hidrolíticas. Contiene una bomba de protones que mantiene el pH ácido en el interior del lisosoma y proteínas de transporte que permiten que los productos resultantes de la digestión pasen al citoplasma. Los lisosomas se forman a partir de vesículas que se desprenden del complejo de Golgi. Hay dos tipos de lisosomas: º Lisosomas primarios: De reciente formación, proceden del complejo de Golgi y contienen diversas enzimas hidrolíticos º Lisosomas secundarios. En ellos tienen lugar procesos activos de digestión celular. Presentan un contenido heterogéneo que depende del tipo de nutrición de la célula. Los lisosomas están especializados en la digestión intracelular de nacromoléculas. Dependiendo de la procedencia del material implicado en la digestión se pueden distinguir dos procesos diferentes: *Heterofagia. Consiste en la digestión de material de origen exógeno que se incorpora a la célula por endocitosis. - Fagolisosomas. Se trata de orgánulos formados a partir de la unión de un lisosoma primario con una vacuola fagocítica, que contienen partículas alimenticias de naturaleza muy variada según el tipo de célula * Autofagia. Consiste en la digestión de material de origen endógeno, es decir, de partes de la propia célula. - Autofagolisosomas. En este caso, los lisososmas se fusionan con vacuolas autofagocíticas para eliminar restos celulares, como orgánulos dañados por sustancias tóxicas o que deben ser reciclados. * Cuerpos multivesiculares. (Su origen no es muy claro) Son lisosomas que, como su nombre indica, contienen en su interior numerosas vesículas. Puede tratarse de vesículas autofágicas o de endocitosis en el interior de lisosomas primarios, o de varios lisosomas primarios reunidos en una membrana común. Una vez finalizada la digestión celular, en los lisosomas secundarios quedan restos que no pueden ser aprovechados por la célula y son excretados al exterior, aunque en ciertos casos permanecen en la célula como cuerpos residuales o telolisomas. + Aparato de Golgi. Es un orgánulo común a todas lñas células eucariotas y está especialmente desarrollado en aquellas que tienen actividad secretora. Está formado por una serie de vesículas en forma de saco llamadas cisternas; en su interior se encuentra un espacio denominado lumen. Las cisternas se apilan en grupos de cuatro a seis y forman un dictiosoma. En un dictiosoma se distinguen dos caras: una cara cis o de formación que está relacionada con el RE; y una cara trans o de maduración que esta distendida formando un retículo tubular llamado red del trans Golgi. De este retículo surgen diferentes vesículas de transporte que se dirigen a sus destinos finales: la membrana plasmática, los lisosomas y las vesículas de secreción. Las funciones más importantes que realiza el aparato de Golgi son: - Interviene en los procesos de secreción. Las proteínas destinadas a ser secretadas al exterior son sintetizadas en el RE rugoso y posteriormente transferidas al complejo de Golgi, donde son empaquetadas en las vesículas de secreción. Dichas vesículas se fusionan con la membrana plasmática y vierten su contenido al exterior por exocitosis. Las proteínas pueden ser secretadas por dos rutas: a) Algunas proteínas son secretadas continuamente, ya que las vesículas secretoras una vez formadas se fusionan rápidamente con la membrana plasmática (ruta constitutiva). b) Otras proteínas son almacenadas en vesículas secretoras especiales y la secreción solo tiene lugar cuando la célula es estimulada por una señal extracelular, como por ejemplo por una hormona (ruta regulada) - Reciclaje de la membrana plasmática Durante la exocitosis, la membrana de las vesículas secretoras pasa a formar parte de la membrana plasmática. Esto permite reponer los componentes de la membrana que se incorporan a la célula por endocitosis, lo que constituye un reciclaje de la membrana plasmática. - Glicosilación. El complejo de Golgi contiene numerosas enzimas glicosil-transferasas que realizan reacciones de glicosilación, tanto en moléculas de lípidos como de proteínas, a medida que éstas van pasando a través del complejo de Golgi. En estas reacciones se eliminan algunos de los azúcares del oligosacárido añadido en el RE y se adicionan otros. - Formación de lisosomas. A partir de vesículas que surgen de la red del trans Golgi. - Formación de las vacuolas En las células vegetales. - Síntesis de componentes de la matriz extracelular en las células animales. - Síntesis de los componentes de la matriz de la pared celular en las células vegetales. - Formación del tabique telofásico en las células vegetales + Retículo endoplasmático liso y rugoso. Es un conjunto de cavidades, túbulos y vesículas conectados entre si y rodeados por una única membrana, que se prolonga formando la envoltura nuclear. El espacio limitado en el interior se llama lumen. Se distinguen dos zonas bien diferenciadas: Una zona que presenta ribosomas asociados a las membranas y que se denomina retículo endoplasmático rugoso (RER). Una zona que no presenta ribosomas y se denomina retículo endoplasmático liso (REL). Las funciones del RER son: - Síntesis y/o modificación de proteínas. Las proteínas sintetizadas en los ribosomas de la cara externa de la membrana del RER se almacenan en el retículo y son transportados por él hacia otros orgánulos. Durante su tránsito algunas proteínas son glicosiladas mediante la transferencia de glúcidos. - Almacenamiento de proteínas. En el lúmen del RER se almacenan las proteínas que han sido previamente sintetizadas. Las funciones del REL son: - Síntesis de fosfolípidos y colesterol necesarios para la formación de nuevas membranas celulares. Estos lípidos son luego distribuidos entre todas las membranas de la célula, mediante proteínas de intercambio o por medio de vesículas de transporte. - Síntesis de hormonas esteroideas. A partir del colesterol, en células especializadas en el metabolismo de lípidos. - Producción de partículas lipoproteicas. Los componentes lipídicos de dichas partículas se sintetizan en el REL, donde se unen a las proteínas que proceden del RER, para dar lugar a las lipoproteínas que serán excretadas. - Interviene en procesos de detoxificación. Metabolizando sustancias tóxicas liposolubles y compuestos perjudiciales producidos por el metabolismo, para evitar que se acumulen en las membranas celulares. Las reacciones de detoxificación transforman las sustancias tóxicas y los metabolitos insolubles en agua, en compuestos hidrosolubles que pueden abandonar la célula y ser eliminados por la orina. +. Mitocondrias. Son las centrales energéticas de todas las células eucarióticas, en ellas tiene lugar la respiración, proceso que implica la obtención de energía a partir de moléculas orgánicas y su conversión en moléculas de ATP. Su tamaño y forma es variable, dependiendo de su origen y su estado metabólico, aunque su forma se describe como alargada. Una célula eucariota típica contiene del orden de 2000 mitocondrias. Las mitocondrias poseen dos membranas: una membrana externa lisa y una membrana interna muy plegada cuyas invaginaciones reciben el nombre de crestas.Estas membranas definen dos compartimentos: el espacio intermembranoso entre ambas membranas y la matriz que está limitada por la membrana interna. La membrana externa es muy permeable a la mayor parte de las moléculas pequeñas e iones, porque contiene numerosas copias de una proteína transmembrana (porina) que forma grandes canales acuosos a través de la bicapa lipídica. La composición química del espacio intermembranoso, debido a la permeabilidad de la membrana externa, es equivalente a la del citosol. Las crestas de la membrana mitocondrial interna aumentan notablemente su superficie y por ello su actividad. Es prácticamente impermeable a las sustancias polares e iones; sólo es completamente permeable al O2, CO2 y H2O. Posse alrededor de un 75% de proteínas y un 25% de lípidos, lo que constituye un contenido en proteínas superior al de las otras membranas celulares: Se pueden distinguir tres tipos diferentes de proteínas: Proteínas transportadoras específicas, que regulan el paso de los metabolitos necesarios en la matriz (ADP, piruvato, ácidos grasos...) y en el exterior (ATP...). Las enzimas de la cadena respiratoria, que son esenciales para realizar la fosforilación oxidativa y generar la mayor parte del ATP de la célula animal. Un complejo enzimático denominado ATP-sintetasa que cataliza la producción de ATP a partir de ADP y Pi. La ATP-sintetasa es la estructura más compleja de la membrana mitocondrial interna. Contiene 8 cadenas polipeptídicas diferentes y en ella se distinguen dos componentes: F1 y F0. F1 es el componente que cataliza la síntesis de ATP y es una pequeña esfera proteica que está expuesta hacia la matriz mitocondrial. El componente F0 es una canal de protones que atraviesa la membrana. La matriz contiene una solución de apariencia gelatinosa, con menos de 50% de agua, q ue está formada por una mezcla muy concentrada de enzimas, diferentes sustratos, ADP, ATP e iones inorgánicos. Contiene también la maquinaria genética mitocondrial formada por varias copias idénticas de DNA circular, RNA y ribosomas. Entre las numerosas enzimas diferentes están: Las responsables del ciclo de Krebs y de la oxidación de los ácidos grasos. Las que intervienen en la replicación, transcripción y traducción del DNA mitocondrial. La mitocondria realiza las siguientes funciones: * La respiración celular. Consiste en la oxidación de las moléculas combustibles por el oxígeno molecular para obtener energía en forma de ATP. La mitocondria utiliza como combustibles mayoritarios los ácidos grasos y el piruvato producido en el citosol a partir de la glucosa. En la matriz de la mitocondria se realizan: - La descarboxilación oxidativa del piruvato a acetil-CoA. - La oxidación de los ácidos grasos a acetil-CoA mediante la -oxidación. - La oxidación del acetil-CoA a CO2 a través del ciclo de Krebs. Todas estas oxidaciones van asociadas a la reducción de los transportadores de electrones NAD+, y FAD a NADH y FADH2. En la membrana mitocondrial interna tiene lugar la síntesis de ATP por un proceso denominado fosforilación oxidativa, que consiste en lo siguiente: - Los electrones de alta energía del NADH y del FADH2 son transferidos a lo largo de la cadena respiratoria transportadora de electrones hasta el oxígeno molecular. Esta cadena respiratoria genera un gradiente de protones a través de la membrana mitocondrial interna. - La energía libre almacenada en el gradiente de protones se utiliza para sintetizar ATP. La ATP-sintetasa es el enzima que cataliza la fosforilación del ADP para formar ATP. * Síntesis de proteinas. En los ribosomas de la mitocondria se sintetizan las proteínas codificadas por el DNA mitocondrial. Estas proteínas constituyen solo el 5% de las proteínas mitocondriales; el resto de las proteínas de la mitocondria están codificadas por el DNA nuclear y se sintetizan en los ribosomas en el citosol. + Cloroplastos. Son orgánulos característicos de las células vegetales. En ellos tiene lugar la fotosíntesis, un proceso impulsado por la luz, en el que se fija CO2 para formar azúcares y otros compuestos orgánicos. Su forma y tamaño son variables. Su número también varía. Al igual que las mitocondrias, los cloroplastos están rodeados por dos membranas: la membrana externa y la membrana interna. El espacio entre ambas membranas se llama espacio intermembranoso. La región acuosa encerrada por la membrana interna se denomina estroma y es equivalente a la matriz mitocondria. En el interior del estroma se localiza una membrana continua denominada membrana tilacoidal, que encierra un espacio interno conocido como espacio tilacoidal. La membrana tilacoidal está muy plegada formando vesículas aplanadas denominadas tilacoides. Estas vesículas pueden encontrarse apiladas formando los grana o como vesículas individuales que atraviesan el estroma y conectan los grana entre si. La membrana externa es permeable a la mayor parte de las moléculas pequeñas; la membrana interna es prácticamente impermeable a la mayoría de las sustancias, pero contiene proteínas transportadoras. Ambas membranas son muy permeables al CO2, que es el sustrato para la síntesis de los hidratos de carbono. La membrana tilacoidal, es impermeable a la mayoría de las moléculas e iones. Contiene los pigmentos fotosintéticos que absorben la energía luminosa. Estos pigmentos son de dos tipos: Clorofilas y carotenoides. Las proteínas de la membrana tilacoidal se pueden clasificar en tres grupos: - Proteínas asociadas a los pigmentos que forman parte de los fotosistemas I y II. - Proteínas de la cadena fotosintética de transporte electrónico. - El complejo ATP-sintetasa, cuya estructura y función son semejantes al de la mitocondria. El estroma contiene una disolución concentrada de enzimas. Tambien contiene una o mas copias de DNA circular, RNA y ribosomas que intervienen en la síntesis de algunas proteínas del cloroplasto. Entre los enzimas están: - Las responsables del ciclo de reducción fotosintética del CO2 o ciclo de Calvin. - Las responsables de la reducción y asimilación de nitratos y sulfatos. - Las que intervienen en la replicación, transcripción y traducción del DNA del cloroplasto. Las funciones que realizan los cloroplastos son: * Fotosíntesis. Proceso en el que la energía de la luz puede utilizarse para convertir el CO2 en compuestos orgánicos, principalmente hidratos de carbono. La fotosíntesis es un proceso anabólico fundamental para todos los seres vivos. Mediante la fotosíntesis, las plantas verdes fabrican materia orgánica para ellas y para casi todos los organismos heterotrofos que dependen de ellas, a través de las cadenas alimenticias de la biosfera. Este proceso global se puede separar en dos fases: - Fase luminosa. La energía luminosa es captada por los pigmentos fotosintéticos y transformada en energía química en forma de ATP y NADPH. En este proceso se oxida agua para proporcionar los electrones que reducen el NADP+ a NADPH y se libera oxígeno molecular. Las reacciones luminosas de la fotosíntesis se localizan en la membrana tilacoidal. - Fase oscura. El ATP y el NADPH producidos en las reacciones de la fase luminosa, se utilizan como fuentes de energía y de poder reductor respectivamente, para convertir el CO2 atmosférico en carbohidratos. Lascreacciones oscuras de fijación y reducción del CO2 tienen lugar en el estroma del cloroplasto y se llevan a cabo mediante una secuencia de reacciones conocida como ciclo de Calvin. * Síntesis de proteínas. El DNA cloroplástico contiene información genética para la síntesis de algunas proteínas del cloroplasto; esta síntesis se lleva a cabo en los ribosomas del orgánulo. La mayor parte de las proteínas cloroplásticas están codificadas por el DNA nuclear y se sintetizan en los ribosomas libres en el citosol. + Vacuolas. Una vacuola es una vesícula muy grande lena de líquido y rodeada por una membrana denominada toniplasto. Junto con los cloroplastos y la pared celular, las vacuolas son componentes característicos de las células vegetales. El número y tamaño de las vacuolas varía según el tipo de célula y las distintas fases de desarrollo. La célula vegetal inmadura contiene gran cantidad de pequeñas vacuolas; en la célula madura, una sola vacuola puede llegar a ocupar hasta el 90% del volumen celular. El principal componente del líquido vacuolar es el agua; también pueden contener aceites y esencias. Las vacuolas contienen normalmente sales, azúcares y proteínas en disolución. Las vacuolas se forman por fusión de vesñiculas procedentes del A.de Golgi. Las funciones mas importantes que realizan son: Almacenan gran cantidad de sustancias: - Nutritivas, como las proteínas de reserva de muchas semillas (guisantes, judias...). - Productos de desecho tóxicos, que aislan del resto del citoplasma, como la nicotina y el opio. - Pigmentos antocianicos, como los que dan color a los pétalos de las flores. Poseen actividad digestiva. Pues pueden contener gran variedad de enzimas hidrolíticos. Están implicadas en la degradación de macromoléculas y en el reciclaje de los componentes celulares.Las vacuolas están estructural y funcionalmente relacionadas con los lisosomas. Regulan la presión de turgencia. La concentración de materiales disueltos dentro de la vacuola es muy alta, superior a la del citosol. La entrada de agua en la vacuola por ósmosis desde el citosol hace que esta se expanda y cree una presión hidrostñatica, o turgencia dentro de la célula. Esto está equilibrado por la resistencia mecánica que presenta la pared celular. El aumento de tamaño de las células se debe, en gran parte, a la acumulación de agua en sus vacuolas. Esto supone un sistema muy económico para el crecimiento de las células vegetales. + Ribosomas. Son pequeñas partículas donde tiene lugar la síntesis de proteínas en todos los seres vivos. Cada ribosoma está formado por una subunidad pequeña y una subunidad grande, que se disocian al final de cada proceso de síntesis de proteínas. Los ribosomas pueden encontrarse en: El citoplasma - Libres en el citosol - Unidos a la cara citosólica de las membranas de RE mediante la subunidad mayor. Las mitocondrias Los cloroplastos Su función más importante es: * Síntesis de proteínas. El ribosoma se une primero a un punto específico del mRNA. Luego el ribosoma se desplaza a lo largo de la molécula de mRNA y va traduciendo la secuencia de nucleótidos a la secuencia de aminoácidos de la proteína. Cuando un ribosoma llega al final del mensaje, la proteína recien sintetizada se libera y las dos subunidades del ribosoma se disocian del mRNA, quedando libres en el citosol. Generalmente varios ribosomas traducen simultáneamente la misma molécula de mRNA, dando lugar a un polisoma o polirribosoma. + Centrosoma. Se localiza al lado del núcleo y está formado por dos centriolos dispuestos perpendicularmente y rodeados por un material amorfo denominado material pericentriolar. Los microtubulos citoplasmáticos tienden a irradiar en todas direcciones a partir del centrosoma. 1.2.4 EL NÚCLEO CELULAR La estructura y composición del núcleo varían dependiendo del estadio del ciclo celular. Se distingue así entre núcleo interfásico, propio de la interfase (periodo que transcurre entre dos divisiones celulares), y núcleo mitótico o en división. + Envoltura nuclear. El núcleo se encuentra rodeado por una membrana doble: membrana nuclear externa y la membrana nuclear interna, separadas por un espacio perinuclear. La membrana nuclear externa está conectada y se contínua con el RER. Ambas membranas se comunican por medio de poros nucleares, a través de los cuales se produce el intercambio y transporte de moléculas grandes, como el RNA y proteínas, entre el nucleoplasma y el citoplasma. Cada poro está rodeado por un anillo integrado por 8 bloques proteicos dispuestos en forma octogonal. Entre el nucleoplasma y la membrana nuclear interna se observa, por último, una lámina nuclear fibrosa, constituida por filamentos intermedios. Su función parece ser la regulación de las interacciones entre la cromatina y la envoltura nuclear, la organización de ésta, así como su desaparición y nueva formación durante la mitosis. + Cromatina. Está compuesta por DNA plegado, asociado a proteínas básicas: las histonas. En la cromatina del núcleo interfásico, la doble hélice de DNA se asocia a histonas y forma complejos denominados nucleosomas. Se podría decir que el nucleosoma es la “unidad estructural” de la cromatina. De hecho, la fibra elemental de cromatina se observa al microscopio electrónico como una fibra de “cuentas” ensartadas semejante a un collar. Cada una de las “cuentas” equivale a un nucleosoma (DNA-histonas), y entre cada unidad hay un filamento de DNA libre que correspondería al “hilo del collar”. Las histonas son proteínas de bajo peso molecular que contienen una elevada proporción de aminoácidos básicos (lisina y arginina). Se dividen en dos grupos: * Histonas nucleosómicas. Componen un complejo proteico discoidal alrededor del cual se enrolla el DNA (dos vueltas de la doble hélice de DNA por nucleosoma). Cada uno de estos discos es un octámero formado por dos copias de cada histona (H2A, H2B, H3 y H4). Esta estructura constituye el nucleosoma. * Histona H1. Une los complejos nucleosómicos y es la responsable del plegamiento helicodal de la fibra elemental de cromatina. La cadena de nucleosomas es la fibra elemental o unidad de la cromatina. Las fibras complejas de cromatina se originarían por el superenrrollamiento de la cadena de nucleosoma según una disposición regular, en la que la histona H1 desempeña un papel esencial. Según la hipótesis mas aceptada hoy día, la cadena de nucleosomas se enrollaría helicoidalmente formando un solenoide o fibra de 30 nm, que contendría 6 nucleosomas por cada vuelta de hélice. Esta estructura estaría estabilizada por las histonas H1. En el núcleo interfásico hay dos tipos de cromatina: Eucromatina. De aspecto laxo o difuso, corresponde a zonas de cromatina activa, donde se produce la transcripción. Heterocromatina. Se visualiza al microscopio como áreas mas densas y homogéneas de cromatina altamente condensada, que corresponden a zonas inactivas que no se transcriben. + Nucleolo. Es una estructura esférica visible en el interior del núcleo interfásico, cuya función principal es la síntesis y ensamblaje de las subunidades ribosómicas. Algunas células presentan mas de un nucleolo. Está constituido por proteínas y ácidos nucleicos, DNA (1-3%) y RNA (1030%). Esta estructura interviene activamente en la transcripción de rRNA y su desarrollo en la célula parece guardar relación con la formación de ribosomas para la síntesis proteica. El DNA nucleolar que codifica para la síntesis de rRNA y puede formar parte de cromosomas distintos constituye el organizador nucleolar. Los rRNA recién sintetizados maduran y se ensamblan con las proteínas ribosómicas importadas del citoplasma, originando las subunidades ribosómicas que emigran al citoplasma, donde se ensamblan a su vez en presencia del mRNA. + Nucleoplasma. También llamado carioplasma, es el medio interno acuoso donde se encuentran inmersos los demás componentes nucleares. Está integrado por proteínas, principalmente enzimas relacionadas con el metabolismo de DNA y RNA. En numerosas ocasiones se han detectado en el núcleo interfásico algunos elementos citoesqueléticos (lámina nuclear) que podrían formar una trama que interacciona con el DNA de la cromatina y con la envoltura nuclear. + Cromosomas. Los cromosomas son estructuras cilíndricas que representan el grado mas elevado de empaquetamiento del DNA y, por tanto, de la cromatina en la célula. Durante la metafase, cada cromosoma aparece constituido por dos brazos o cromátidas iguales, unidos por el centrómero. Los centrómeros reciben también el nombre de constricciones primarias e incluyen los cinetocoros, placas de naturaleza proteica a las que están conectados los microtúbulos cromosómicos del huso mitótico. Según la posición del centrómero, los cromosomas se clasifican en metacéntricos, submetacéntricos, acrocéntricos y telocéntricos. Los extremos de las cromátidas se denominan telómeros y en ellos se encuentran secuencias repetitivas de DNA que tiene como función evitar la pérdida de información genética en la replicación y la fusión de cromosomas alterados (mantienen la individualidad de los cromosomas). Los satélites son zonas de forma aproximadamente esférica, separadas del cromosoma por constricciones secundarias. Los cromosomas presentan un nivel de complejidad estructural mayor que la cromatina del núcleo interfásico. El solenoide cromatinico constituye dominios en forma de bucles o lazos radiales que, a su vez, se enrollan helicoidalmente y experimentan sucesivos grados de compactación hasta dar lugar a las cromátidas del cromosoma metafásico. 2.- CICLO CELULAR: CONCEPTO Y CLASES A lo largo de la vida de las células se pueden observar distintos estadios que corresponden a la sucesión de diferentes actividades. Así, se define el ciclo celular como el conjunto de procesos que tienen lugar desde la división de una célula hasta la división de sus células descendientes. La duración y las características del ciclo celular son variables y dependen del tipo de célula y de las circunstancias en las que se desarrolla. A lo largo del ciclo celular, pueden distinguirse dos fases fundamentales: la división celular y la interfase celular, es decir, el periodo entre división y división. Cada una de estas fases se divide en etapas o periodos distintos. 2.1. DIVISIÓN CELULAR Es el proceso por el cual, a partir de una célula, que podemos denominar madre, se obtienen dos células hijas. Cada una de estas células hijas contiene una copia exacta de la información genética de la célula madre y, aproximadamente, la mitad del material citoplasmático de ésta. En los seres unicelulares, la división celular implica la generación de nuevos individuos y, por tanto, la perpetuación de la especie. En los seres pluricelulares, la división celular hace posible numerosos procesos, como: - El desarrollo de un individuo completo a partir de una célula inicial. - La aparición de distintos tejidos especializados. 2.2. INTERFASE CELULAR Es el periodo que suele abarcar el 90% del ciclo celular. Durante la interfase podemos distinguir varias etapas: G1, S y G2. 2.2.1. Etapa G1 Es la etapa comprendida entre la división celular y el inicio de la duplicación de los cromosomas. Durante G1 la actividad celular, que había disminuido durante la división, se recupera, y todos los procesos metabólicos se desarrollan a la velocidad marcada por los requerimientos celulares. En la célula pueden observarse importantes cambios morfológicos y funcionales. Aumenta el tamaño de las células. Cada célula hija crece hasta alcanzar el tamaño propio de su tipo celular, normalmente el que tenía la célula madre. En el caso de las células animales, el aumento de tamaño se produce por la síntesis de fragmentos de membrana y de otros componentes celulares. Sin embargo, en las células vegetales, la presencia de una pared rígida implica una serie de modificaciones, entre las cuales es fundamental un aumento de la turgencia celular. - El proceso se inicia con un reblandecimiento de la pared celular; al mismo tiempo, la célula incorpora agua al citoplasma. - Esto provoca la expansión del citoplasma y un incremento de la turgencia. - El citoplasma presiona sobre la pared reblandecida, que cede y permite el crecimiento de la célula. - Una vez se ha completado la expansión, la pared celular se refuerza con nuevos depósitos de celulosa. - Aumenta el número de estructuras celulares Se duplican las mitocondrias y los cloroplastos en los vegetales El RE aumenta de tamaño y origina el aparato de Golgi, nuevos lisosomas y vacuolas Se sintetizan proteínas que constituirán los ribosomas y el citoesqueleto. Se sintetizan todas las proteínas enzimáticas que se requieren para el control de éstos procesos. Se recupera lo forma habitual de las células Se “desmontan” las estructuras fibrosas que habían participado en el reparto de los cromosomas y el citoesqueleto se organiza del modo habitual La etapa G1 tiene una duración muy variable que oscila entre pocas horas y varios días y condiciona la duración del ciclo celular completo. 2.2.2. Etapa S Es una etapa clave en el ciclo celular, ya que en ellas se produce la síntesis de DNA para la duplicación de los cromosomas. Cada cadena de DNA actúa como molde en la síntesis de una nueva cadena, que será complementaria de la anterior. Al final de la etapa S, la duplicación de las dobles hélices da lugar a la formación de dos cromátidas que se mantienen unidas por el centrómero. 2.2.3. Etapa G2 Es la etapa que transcurre entre el final de la síntesis del DNA y el inicio de la mitosis. Como etapa precursora de la mitosis, durante G2 tiene lugar una serie de acontecimientos preparatorios de éste proceso: - Se empiezan a condensar los cromosomas - Se duplica el par de centriolos y se disponen cerca del núcleo - Se inician las modificaciones del citoesqueleto necesarias para facilitar su intervención en la mitosis. Al finalizar la etapa G2 los cromosomas se condensan y son visibles al microscopio óptico, de modo que pueden distinguirse fácilmente las dos cromátidas. En el centrómero se observan ya los cinetocoros que se unirán a los microtúbulos encargados de la distribución del material hereditario. 3.- MITOSIS Y CITOCINESIS. 3.1. MITOSIS La mitosis o división nuclear es el proceso mediante el cual se asegura que cada una de las células hijas reciba un núcleo con la información genética completa e idéntica a la de la célula progenitora. Aunque la mitosis constituye un proceso continuo suelen diferenciarse las siguientes fases: 3.1.1. Profase. Antes del comienzo de la profase, se producen en la célula varios fenómenos preparatorios para la mitosis, como la duplicación del par de centriolos (al inicio de la fase S), la duplicación de la información genética (de modo que al final de la fase S cada cromosoma estará formado por dos cromátidas unidas por el centrómero) y la reorganización del citoesqueleto y despolimerización de microtúbulos durante la fase G2. Durante la profase mitótica, el núcleo experimenta los siguientes cambios: - Condensación de la cromatina difusa, de manera que los cromosomas se van haciendo visibles progresivamente. - Migración de los cromosomas a la periferia nuclear - Desaparición paulatina del nucleolo - Comienzo de la formación del huso acromático: en las proximidades de cada par de centriolos, rodeados por el áster (conjunto de microtubulos muy cortos que irradian de los centriolos) y la centrosfera (zona clara que se localiza alrededor de cada pareja de centriolos, externamente al áster) se polimeriza una serie de microtúbulos polares que se conectan en la zona media y van alargándose a medida que los centriolos migran hacia polos opuestos. Al final de la profase desaparece la membrana nuclear. 3.1.2. Prometafase Esta etapa se caracteriza por la unión de los cromosomas al huso acromático. Cada cromosoma presenta dos cinetocoros, a ambos lados del centrómero, a los que se unen ciertos microtubulos denominados cromosómicos o cinetocóricos. De ésta forma el cromosoma está conectado a través de los microtúbulos con los dos polos del huso. 3.1.3. Metafase Los cromosomas, en su máximo grado de empaquetamiento, se disponen en el plano ecuatorial y el huso mitótico aparece claramente constituido por microtubulos polares y microtúbulos cinetocóricos. 3.1.4. Anafase Es una etapa muy corta que se caracteriza por: - Se produce la división de los centrómeros, que se separan llevando unidas sus cromátidas correspondientes - Tiene lugar el acortamiento progresivo de los microtubulos cromosómicos y la separación de las cromátidas de cada cromosoma que migran hacia los polos opuestos del huso mitótico. - Los microtubulos polares se alargan en el ecuador del huso, donde comienza a depositarse un material denso. 3.1.5. Telofase Constituye la fase final de la mitosis, que se caracteriza por: - La desaparición de los microtubulos cromosómicos, una vez que los cromosomas hijos han alcanzado los polos. - El comienzo de la desespiralización de los cromosomas. - Las masas de cromatina se rodean de una nueva envoltura nuclear (originada, fundamentalmente, a partir de fragmentos de la envoltura del núcleo parental) y se forman los núcleos hijos. - Aparecen de nuevo los nucleolos. 3.2. CITOCINESIS. La división del citoplasma se inicia habitualmente en la telofase. Se produce un reparto del citoplasma y los orgánulos celulares, y la célula comienza a sufrir una constricción en la zona ecuatorial (surco de división). En las células animales, la formación del surco de división implica una expansión de la membrana en esa zona y una contracción progresiva causada por un anillo periférico contráctil de microfilamentos de actina y miosina. Este anillo producirá, finalmente, la separación de las dos células hijas por estrangulación del citoplasma. En las células vegetales, el proceso es diferente, ya que la citocinesis no se produce por estrangulamiento, sino por la acumulación de vesículas procedentes del complejo de Golgi, que contienen elementos de la pared celular, en la zona media de la célula. Posteriormente, las vesículas se fusionan y entran en contacto con las paredes laterales de la célula parental. De ésta forma se origina un tabique o fragmoplasto que dará lugar a las membranas de las dos células hijas, separadas por la lámina media, en el ecuador de la célula. Por último, se depositará la pared primaria y, en algunos casos, la pared secundaria, dependiendo del tipo celular. La división celular origina dos células hijas genéticamente idénticas al individuo parental, por lo que en los organismos unicelulares este proceso puede considerarse una reproducción asexual. 4.- MEIOSIS Y GAMETOGÉNESIS. 4.1. MEIOSIS. La meiosis es un tipo especial de división del núcleo celular que origina cuatro núcleos haploides a partir de un núcleo diploide. En los individuos diploides es necesaria para la formación de los gametos durante la reproducción sexual y para el mantenimiento del número cromosómico de cada especie. La meiosis consta de dos divisiones sucesivas del núcleo, entre las cuales no se produce la duplicación del material genético (fase S). Estas divisiones se denominan primera división meiotica y segunda división meiotica. 4.1.1. Primera división meiotica o división reduccional En ella, los cromosomas homólogos se emparejan y posteriormente se separan para dar lugar a los núcleos hijos (que contendrán entonces un cromosoma de cada par de homólogos; cada cromosoma contiene todavía dos cromátidas, es decir, dos copias de la información genética). El reparto de los cromosomas de cada par de homólogos ocurre al azar, lo cual contribuye a la variabilidad genética de los gametos. Consta de cinco fases, denominadas del mismo modo que las fases mitóticas. La mas importante y la de mayor duración es la profase meiotica I. Profase meiotica I Es muy compleja y se ha dividido en seis etapas: - Preleptoteno. En ésta etapa, que en muchos casos se considera premeiotica, se ha producido ya la duplicación del ADN y, por tanto, la fase S. Los cromosomas apenas se distinguen. - Leptoteno. Los cromosomas , aún en proceso de espiralización, se unen a la membrana nuclear en zonas próximas a la localización de los centriolos a través de las placas de unión. Las dos cromátidas de cada cromosoma se unen estrechamente y comienza a formarse el huso mitótico. - Zigoteno. Los cromosomas homólogos se aparean entre sí, originándose en la zona de contacto una estructura denominada complejo sinaptonémico, constituido por una placa central densa y elementos laterales de estructura fibrilar. El complejo sinaptonémico es necesario para que se produzca, en la etapa siguiente, el entrecruzamiento o sobrecruzamiento de los cromosomas homólogos. En ésta fase cada pareja de cromosomas se llama bivalente o tétrada (contiene cuatro cromátidas). - Paquiteno. Una vez completados el apareamiento y la condensación de los cromosomas homólogos, tiene lugar el sobrecruzamiento (crossing-over) entre cromátidas no hermanas. Los puntos de sobrecruzamiento corresponden a los nódulos de recombinación que contienen los enzimas necesarios para el intercambio de genes entre las cromátidas de los dos cromosomas homólogos (endonucleasas, ADN polimerasas, ADN ligasa, y otras proteínas imprescindibles para que se produzca el sobrecruzamiento). Este proceso implica la rotura de la doble hélice en los fragmentos que va a ser intercambiados y una posterior fusión en su nueva localización. - Diploteno. Los cromosomas homólogos comienzan a separarse, aunque aún permanecen unidos en aquellos puntos donde ha tenido lugar el sobrecruzamiento y que se denominan quiasmas. Se inicia, así mismo, la desaparición de los complejos sinaptonémicos. Constituye la etapa mas larga de la meiosis, ya que puede durar días, e incluso años, como el caso de los ovocitos humanos. Al final de ésta etapa, los cromosomas empiezan a desespiralizarse y adquieren un aspecto mas laxo. - Diacinesis. Los cromosomas aparecen de nuevo condensados, alcanzando su máximo grado de empaquetamiento, y los quiasmas se van desplazando hacia los extremos del bivalente (fenómeno que se conoce como terminalización de los quiasmas). La membrana nuclear y el nucleolo comienzan a desaparecer. Prometafase meiotica I Se completa al final de ésta fase la desaparición de la membrana nuclear y el nucleolo, y empieza la unión de las parejas de cromosomas, totalmente condensados, a los microtubulos cinetocóricos. Metafase meiotica I Los bivalentes (parejas de cromosomas homólogos) se disponen en el plano ecuatorial. Solo se observan ya algunos quiasmas terminales. Anafase meiotica I. Se separan los bivalentes y cada uno de los cromosomas del par de homólogos, constituido a su vez por dos cromátidas, emigra hacia uno de los polos. Telofase meiotica I. La división nuclear concluye con la formación de dos núcleos hijos, cada uno de los cuales contiene un juego completo de cromosomas. En ésta fase reaparecen el nucleolo y la membrana nuclear. A continuación se produce la citocinesis o separación de las dos células hijas. En el periodo que transcurre entre la primera y la segunda división meiotica no hay fase de duplicación (S). 4.1.2. Segunda división meiotica. Como consecuencia de la primera división meiótica, cada núcleo hijo contiene un cromosoma de cada pareja de homólogos, constituido, a su vez, por dos cromátidas. La segunda división meiotica equivale a una mitosis normal, en la que las dos cromátidas de cada cromosoma se separan y emigran hacia los polos opuestos del huso acromático. Como tras la primera división meiótica se han originado dos células haploides, la segunda división meiotica dará como resultado cuatro células también haploides. Sin embargo, la dotación genética de cada una de las células es el fruto de la recombinación entre cromosomas homólogos (que tienen distinto origen, paterno y materno) y, por tanto, de la mezcla de caracteres genéticos diferentes. Esta segunda división consta, igual que la primera, de cinco fases, que se denominan: profase meiotica II, prometafase meiotica II, metafase meiotica II, anafase meiotica II y telofase meiotica II. 4.2. SIGNIFICADO BIOLÓGICO DE LA MEIOSIS. Cuando una célula se divide por mitosis, las dos células hijas producidas son genéticamente iguales a la original, lo cual significa que para aquellos organismos que se reproducen asexualmente las posibilidades de variabilidad genética y, en consecuencia, de adaptación y de evolución son muy limitadas. En los organismos con reproducción sexual, sin embargo, la fusión de dos núcleo haploides de distinta procedencia (es decir, genéticamente diferentes) originan un cigoto diploide con información genética diferente, producto de la recombinación de los dos núcleos parentales. La variabilidad genética generada en la reproducción sexual se debe a: - Las distintas posibilidades de reparto en la segregación de los cromosomas parentales en la primera división meiotica. - La recombinación y el intercambio de información genética producidos en la profase meiotica I. Los organismos unicelulares, en contacto directo con el entorno, se reproducen asexualmente cuando las condiciones ambientales son favorables, lo cual permite un incremento rápido de la población. Sin embargo, la posibilidad de producirse fenómenos sexuales, que ocurren a menudo ante cambios en el ambiente, aumenta considerablemente sus probabilidades de supervivencia e incluso de colonización de nuevos ambientes. En los seres mas complejos (animales y plantas superiores), las células de cada organismo se dividen asexualmente, pero la especie se reproduce sexualmente. 4.3. GAMETOGENESIS. La gametogénesis es la formación de gametos. Puede ser de dos tipos: espermatogenesis y ovogénesis. 4.3.1. Espermatogénesis. Es el proceso de formación de espermatozoides. Tiene lugar en el interior de los tubos seminíferos, existentes en los testículos, a partir de células germinales diploides llamadas espermatogonias, que se dividen (fase de multiplicación) activamente, por mitosis sucesivas, en cuanto el organismo alcanza la madurez sexual. Las espermatogonias de la última generación experimentan un aumento de tamaño (fase de crecimiento) y se convierten en espermatocitos de primer orden, que aún son diploides (2n). Cada espermatocito de primer orden sufre una meiosis en función de la cual se originan, en una primera división reduccional, dos espermatocitos de segundo orden, que son haploides (n). Cada uno de éstos experimentará una segunda división que producirá dos espermátidas haploides (n). Todo ello constituye la fase de maduración. Por último, las espermátidas, que son células inmóviles y redondeadas, sufren una serie de cambios profundos en su estructura (fase de diferenciación) y se transforman en espermatozoides. Estos cambios consisten en que el núcleo se desplaza hacia uno de los polos, el aparato de Golgi forma el acrosoma, las mitocondrias se disponen helicoidalmente, etc. 4.3.2. Ovogénesis. Es el proceso de formación y maduración de óvulos. Tiene lugar en el interior de los ovarios a partir de células germinales diploides denominadas ovogonias Las ovogonias, se dividen activamente por mitosis (fase de multiplicación) y crecen de tamaño, acumulando sustancias de reserva (fase de crecimiento) transformándose en oocitos de primer orden, también diploide (2n). Estos, inician la primera división de que consta una meiosis y originan dos células de tamaño diferente (fase de maduración). La mas grande constituye el oocito de segundo orden, haploide (n), y la otra, el primer corpúsculo polar. Tanto una como otra sufren la segunda división de que consta una meiosis. El oocito de segundo orden, vuelve a dar una célula grande que es la ovocélula y un segundo corpúsculo polar. El primer corpúsculo polar da lugar a otros dos. Todos los corpúsculos polares abortan y por tanto no son útiles como gametos femeninos. Sólo la ovocélula es la única célula válida para ello. 5.- CONCEPTO DE METABOLISMO: CATABOLISMO Y ANABOLISMO. El conjunto de reacciones químicas catalizadas por enzimas que se producen en el interior de las células de los seres vivos recibe el nombre de metabolismo. Las reacciones metabólicas se encuentran en su mayor parte totalmente interrelacionadas constituyendo las rutas o vías metabólicas, de modo que el producto de la primera reacción es el sustrato de la siguiente y así sucesivamente. En el metabolismo se distinguen dos fases: * Fase degradadora o catabolismo. Es el conjunto de reacciones que tienen por objeto la obtención de energía y moléculas precursoras a partir de otras mas complejas. Los procesos catabólicos son degradativos y producen energía química, por tanto, son reacciones exergónicas. Las rutas catabólicas más importantes son las siguientes: - Glucólisis. Es la ruta en la que la glucosa se degrada hasta producir un compuesto de tres átomos de carbono denominado piruvato. - - oxidación. Es el conjunto de reacciones en el que se produce la oxidación de los ácidos grasos hasta dar lugar a un compuesto de dos átomos de carbono denominado acetilcoenzima A (Acetil-CoA) - Transaminación y desaminación. Es el conjunto de procesos en los que tiene lugar la degradación de los aminoácidos mediante la separación del grupo amina del esqueleto carbonado. En general, las rutas catabólicas convergen hacia la formación de un compuesto de dos átomos de carbono denominado acetil-CoA. Este compuesto se incorpora a un ciclo que representa la ruta central de todo el metabolismo: el ciclo del ácido cítrico o ciclo de Krebs. * Fase de síntesis o anabolismo. Es el conjunto de reacciones que tienen por objeto la síntesis de moléculas complejas a partir de moléculas precursoras y energía. Los procesos anabólicos son sintéticos y consumen energía, por tanto, son reacciones endergónicas. Las principales rutas anabólicas son las siguientes: - Fotosíntesis. Consiste en la obtención de moléculas orgánicas (glúcidos) a partir de energía lumínica y de moléculas inorgánicas como CO2 y H2O. - Gluconeogénesis. Es la ruta de síntesis de glucosa a partir de sustancias precursoras, como por ejemplo el piruvato. Esta ruta se produce independientemente de la fotosíntesis. - Síntesis de ácidos grasos. Es el conjunto de reacciones en las que se obtienen ácidos grasos a partir de acetil-CoA. - Síntesis de aminoácidos. Es el conjunto de procesos en los que sintetizan aminoácidos a partir de piruvato, oxalacetato, etc. Ambas fases están íntimamente relacionadas, ya que la energía que se produce durante el catabolismo, así como las moléculas precursoras que se obtienen, son necesarias para el desarrollo de las reacciones del anabolismo. El ATP es la molécula rica en energía que se produce o se consume en éstos procesos. Por otro lado, también participan moléculas de NAD, NADP y FAD, que están relacionadas con la producción de ATP. En cada ruta metabólica se establece un balance energético, es decir, un recuento global del número de ATP, así como de moléculas de NAD, NADP y FAD. Las funciones del metabolismo son: º La obtención de energía química a partir de la degradación de las biomoléculas (glúcidos, lípidos y proteínas). º La obtención de moléculas precursoras, es decir, moléculas imprescindibles para la síntesis de las biomoléculas, como monosacáridos, ácidos grasos, aminoácidos. etc. º La síntesis de biomoléculas, como glúcidos, lípidos y proteínas, etc 6.- CATABOLISMO DE LOS GLÚCIDOS En la degradación completa de una molécula de glucosa se consume oxígeno y se obtienen como productos finales dióxido de carbono, agua y energía en forma de ATP. La glucosa se incorpora a las vías catabólicas directamente de la dieta, procedente de la hidrólisis de disacáridos de los alimentos, o de la degradación de polisacáridos de reserva. 6.1. GLUCOLISIS. Es la ruta metabólica que convierte a la glucosa en piruvato, con la producción de dos moléculas de ATP. Consiste en una secuencia de 10 reacciones catalizadas enzimaticamente, que se realizan en el citosol de la célula y comprende dos etapas diferentes: Una primera etapa preparatoria en la que la glucosa es fosforilada, dando lugar a dos moléculas de gliceraldehido-3-fosfato. En éste proceso se consumen dos moléculas de ATP en activar a la molécula de glucosa para su posterior catabolismo. Glucosa + 2 ATP 2 gliceraldehido-3-fosfato Una segunda etapa en la que las dos moléculas de gliceraldehido-3-fosfato son oxidadas por el NAD+ y a continuación convertidas en piruvato, con la producción de cuatro moléculas de ATP. Por tanto el rendimiento energético de la glucólisis es de dos ATP por molécula de glucosa. 2 gliceraldehido-3-fosfato + 2 NAD+ + 4 ADP + 2Pi 2 piruvato + 2 NADH + 4 ATP + 2 H2O Las reacciones de la glucólisis son: 1.- La glucosa es fosforilada por el ATP para formar glucosa-6-fosfato (G6P). Esta reacción catalizada por la hexoquinasa es irreversible. La fosforilación de la glucosa impide que ésta molécula pueda salir de la célula ya que los intermediarios fosforilados no pueden atravesar las membranas celulares. 2.- La glucosa-6-fosfato se isomeriza a fructosa-6-fosfato en presencia del enzima fosfoglucosaisomerasa . 3.- La fructosa-6-fosfato es fosforilada por el ATP para formar fructosa-1,6-bisfosfato con la intervención del enzima fosfofructoquinasa. 4.- La fructosa-1,6-bisfosfato, por medio del enzima aldolasa, se rompe en dos triosas: gliceraldehido-3-fosfato (G3P) y dihidroxiacetona fosfato (DHAP), que son dos intermediarios metabólicos importantes. 5.- Sólo el G3P continúa en la ruta glicolítica, pero el G3P y la DHAP son isómeros fácilmente interconvertibles: en el equilibrio de la reacción de interconversión de éstos dos azúcares está favorecida la formación de DHAP. Sin embargo, el continuo consumo de G3P en la etapa posterior de la glicólisis desplaza la reacción hacia la formación de G3P. Por tanto, el resultado de ésta primera etapa de la glicólisis es que una molécula de glucosa se ha transformado en dos moléculas de G3P y se han consumido 2 moléculas de ATP. 6.- La oxidación y fosforilación del gliceraldehido-3-fosfato es una reacción central de la glicólisis. En ella, el NAD+ actúa como agente oxidante y al mismo tiempo se incorpora una molécula de Pi al G3P para formar 1,3bisfosfoglicerato (1,3-BPG). En ésta reacción de óxido-reducción se obtiene el primer intermediario con un grupo “fosfato de alta energía de hidrólisis”. La reacción es catalizada por el enzima gliceraldehido-3-fosfato deshidrogenasa. 7.- El 1,3-BPG cede un grupo fosforilo al ADP formándose 3fosfoglicerato, en presencia del enzima fosfogliceratoquinasa y ATP. 8.- El 3-fosfoglicerato se convierte en 2-fosfoglicerato por medio del encima fosfogliceratomutasa. 9.- Por deshidratación del 2-fosfoglicerato se forma, por la enolasa, fosfoenolpiruvato. En ésta reacción se obtiene el segundo intermediario con un grupo “fosfato de alta energía de hidrólisis”. 10.- El fosfoenolpiruvato transfiere su grupo fosforilo al ADP formándose piruvato y ATP. Esta reacción es catalizada por la piruvatoquinasa. La reacción global del balance energético de la glicólisis es : H2O Glucosa + 2 ADP + 2 Pi + 2 NAD+ 2 piruvato + 2 ATP + 2 NADH + 2 H+ + 2 Así pues, respecto al rendimiento energético, se han gastado dos moléculas de ATP para iniciar el proceso y se han sintetizado cuatro, lo que da un rendimiento neto de dos moléculas de ATP. Por otro lado, se han obtenido también dos moléculas de NADH. La fase siguiente a la glucólisis es la degradación de las dos moléculas de piruvato. Esta etapa es distinta según se produzca en ausencia o presencia de oxígeno. - En ausencia de oxígeno, es decir, en condiciones anaeróbicas, el piruvato se transforma en lactato o en etanol mediante un proceso de fermentación. - En presencia de oxígeno, es decir, en condiciones aeróbicas, el piruvato sufre una oxidación y da lugar a acetil-CoA, NADH y CO2. 6.2. FERMENTACIONES. (Catabolismo del piruvato en ausencia de O2). La fermentación del piruvato puede ser de dos tipos, alcohólica o láctica. 6.2.1. Fermentación láctica. Determinados microorganismos como, por ejemplo, Lactobacillus bulgaricus, Lactobacillus casei, Streptococcus lactis, obtienen la energía de la lactosa de la leche mediante un proceso de fermentación anoxidativa, en la que se diferencian dos fases: * En la primera, la lactosa se hidroliza mediante el enzima lactasa, que suministra glucosa y galactosa; a su vez, la galactosa se isomeriza y se convierte en glucosa. * En la segunda fase, toda la glucosa se transforma en piruvato mediante la glucólisis; a partir de aquí, la etapa característica de la fermentación láctica consiste en la reducción del piruvato, que es el último aceptor de electrones, por el NADH y su transformación en ácido láctico, en presencia del enzima lactatodeshidrogenasa, que es el producto final de éste proceso. El balance energético es: Glucosa + 2 ADP + 2Pi 2 lactato + 2 ATP + 2 H2O En esta reacción se consume un NADH. Si se degradan dos moléculas de piruvato se consumen dos NADH. 6.2.2. Fermentación alcohólica Es una fermentación anoxidativa, levada a cabo, en éste caso, por determinadas levaduras del género Saccharomyces, que transforman la glucosa procedente de diversas fuentes hidrocarbonadas en alcohol etílico y CO2. El piruvato producido en la glucólisis sufre una descarboxilación, se transforma en acetaldehído (etanal) por la piruvatodescarboxilasa; el metabolito que actúa como último aceptor de electrones, y es reducido por el NADH a etanol en presencia del enzima alcoholdeshidrogenasa. El balance energético es: Glucosa + 2 ADP + 2 Pi + 2H+ 2 Etanol + 2 ATP + 2 CO2 + 2 H2O En ésta reacción se consume un NADH, por tanto, si se degradan dos moléculas de piruvato se consumen dos NADH. 6.3. REACCIONES PREVIAS AL CICLO DE KREBS. (Oxidación del piruvato) Es un proceso de degradación del piruvato hasta acetil-CoA, en presencia de oxígeno. Este proceso tiene lugar en la matriz mitocondrial. La molécula de piruvato pierde un grupo carboxilo en forma de CO2 y da lugar a acetil-CoA y a NADH. La reacción está catalizada por un grupo de tres enzimas, el complejo piruvato deshidrogenasa. Este complejo regula la velocidad de formación del acetil-CoA y, por tanto, la velocidad a la que se incorpora al ciclo del ácido cítrico. El balance energético es el siguiente: Glucosa + 2 ADP + 2Pi + 4 NAD+ 2 acetil-CoA + 2 ATP + 4 NADH + 4 H+ + “ CO2 + 2 H2O La degradación de una molécula de piruvato produce un NADH, por tanto, la degradación de dos moléculas de piruvato da lugar a dos NADH. 6.4. RESPIRACIÓN AEROBIA 6.4.1. Ciclo de Krebs El ciclo de Krebs es la ruta oxidativa final de la glucosa y de la mayoría de los combustibles metabólicos. Su función es oxidar el grupo acetilo del Acetil-CoA a CO2, al mismo tiempo que se reducen los transportadores de electrones NAD+ y FAD a NADH y FADH2. La posterior oxidación de NADH y FADH2 en la cadena respiratoria mitocondrial genera ATP. El ciclo de Krebs consta de una serie de ocho reacciones catalizadas enzimaticamente que se realizan en la matriz mitocondrial.. Estas reacciones son las siguientes: 1.- El grupo acetilo del acetil-CoA se condensa con el oxalacetato para dar citrato que es un ácido tricarboxilico. 2.- El citrato se isomeriza a isocitrato mediante una deshidratación seguida de una hidratación. En ésta transformación se forma un compuesto intermedio, el cis-aconitato. 3.- Por descarboxilación oxidativa del isocitrato, se forma alfacetoglutarato por medio del enzima isocitrato deshidrogenasa. Esta es la primera reacción en la que se produce NADH y también la primera reacción en la que se forma CO2. 4.- El alfa-cetoglutarato se descarboxila oxidativamente dando succinilCoA por medio del enzima alfa-cetoglutarato deshidrogenasa En ésta reacción se produce la segunda molécula de NADH y la segunda molécula de CO2. Hasta ese punto del ciclo se han formado dos moléculas de CO2, con lo que el grupo acetilo se ha oxidado completamente. 5.- El succinil-CoA se transforma en succinato en presencia del enzima succinil-CoA sintetasa. La energía que se libera en ésta reacción se utiliza para la síntesis de GTP. El GTP puede ceder su grupo fosfato al ADP y dar lugar a ATP. 6.- El succinato se oxida a fumarato, a la vez que se reduce el FAD a FADH2. 7.- El fumarato incorpora una molécula de agua, es decir, se hidrata y produce un isómero, el malato. El enzima que actúa es la fumarasa. 8.- En ésta última reacción del ciclo, el malato se oxida y, en presencia del enzima malato deshidrogenasa, origina oxalacetato y NADH. El balance energético es el siguiente: La oxidación de una molécula de acetil-CoA libera 1ATP, 3 NADH y 1 FADH2. Sin embargo, la degradación de dos moléculas de piruvato da lugar a dos moléculas de actil-CoA y, por tanto, se obtienen 2 ATP, 6 NADH y 2 FADH2. Como resultado de la oxidación de una molécula de glucosa a CO2 y H2O se obtienen: 4 ATP / 10 NADH / 2 FADH2 6.4.2. Cadena respiratoria: Fosforilación oxidativa El conjunto de moléculas transportadoras de electrones de la membrana mitocondrial interna constituye la cadena respiratoria. Estas moléculas pueden ser complejos enzimáticos que se encuentran fijos en la membrana o moléculas capaces de desplazarse por ella. La circulación de electrones por la cadena respiratoria se produce mediante reacciones de oxidación-reducción, ordenadas en serie. Cada componente de la cadena acepta los electrones del componente anterior y los transfiere al siguiente y así sucesivamente. El mecanismo de la cadena respiratoria es el siguiente: - Complejo I, complejo citocromo bc1 o NADH deshidrogenasa. Complejo enzimático que cataliza el paso de dos electrones del NADH a la ubiquinona. - Complejo II o succinato deshidrogenasa. Cataliza la transferencia de electrones procedentes del FADH2 a la ubiquinona. - Ubiquinona (UQ) o coenzima Q. Compuesto que se desplaza por la membrana y transfiere los electrones al complejo III. - Complejo III o ubiquinona-citocromo c oxidorreductasa. Cataliza el transporte de electrones desde la ubiquinona al citocromo c. - Complejo IV o citocromo oxidasa. Cataliza el transporte de electrones desde el citocromo c al oxígeno, se obtiene una molécula de agua. La cadena de transporte de electrones está íntimamente relacionada con la denominada Fosforilación oxidativa, que consiste en la incorporación de ácido fosfórico a una molécula de ADP para formar ATP, que almacena de esta forma la energía liberada en el proceso respiratorio. Esta energía liberada en forma de ATP puede ser fácilmente utilizada por el organismo para desarrollar el trabajo que exigen sus distintas funciones fisiológicas. Actualmente se considera que la fosforilación oxidativa tiene lugar en virtud de la hipótesis quimiosmótica. Según ésta hipótesis el mecanismo sería el siguiente: Los protones liberados al ionizarse los hidrógenos transportados por el NAD y/o el FAD, se van acumulando en el exterior de la membrana de las crestas mitocondriales, es decir, en la matriz. Esto crea una diferencia de potencial a ambos lados de la membrana por el predominio de cargas positivas en el lado externo (matriz). Ahora bien, el funcionamiento de la cadena de transporte electrónico provoca la expulsión activa de protones desde la matriz al espacio intermembranoso, los cuales retornan a la matriz a través de los factores de acoplamiento y al llegar a la porción F1 de dichos factores cambian la conformación molecular de la ATP-sintetasa. El mecanismo de síntesis de ATP ha sido interpretado de acuerdo con la hipótesis conformacional del siguiente modo. La ATP-sintetasa presenta dos centros activos con diferente afinidad para el ADP y el ácido fosfórico según su estado de conformación. Cuando no hay aporte energético, uno de los centros activos está ocupado por una molécula de ATP, mientras que el otro muestra gran afinidad por el ADP y el ácido fosfórico. Al producirse un aporte energético (funcionamiento de la cadena de transporte de electrones y paso de protones por las partículas F) se induce un cambio en la conformación del centro activo donde se halla alojado el ATP, que provoca su expulsión; mientras que el otro centro activo, que también cambia de conformación, se muestra entonces con capacidad catalítica para unir el ácido fosfórico al ADP y formar ATP, el cual quedará unido al enzima, hasta que un nuevo aporte energético provoque otro cambio de conformación y se repita el proceso. 6.4.3. Balance energético. A partir de una molécula de glucosa, se obtiene el siguiente balance energético: Glucólisis Oxidación del piruvato Ciclo ácido cítrico TOTAL Nº de ATP formados 2 Nº de NADH formados 2 N de FADH2 formados 2 2 4 6 10 2 2 Cada molécula de NADH da lugar a 3 ATP 10 x 3 = 30 moléculas de ATP Cada molécula de FADH2 da lugar a 2 ATP 2 x 2 = 4 moléculas de ATP 4 ATP + 30 ATP + 4 ATP = 38 moléculas de ATP La membrana interna mitocondrial de las células animales es impermeable al NADH citosólico y necesita un sistema de transporte específico. Este hecho hace oscilar el balance energético global del catabolismo de la glucosa entre 38 y 36 ATP. Para que pueda incorporarse a la cadena respiratoria los electrones procedentes del NADH del citosol existen unos sistemas de transporte especiales denominados lanzaderas. Las lanzaderas son unas rutas indirectas constituidas por diversos compuestos, que mediante su oxidación y reducción permiten el paso de los electrones al complejo I o al complejo III dependiendo de la lanzadera funcional. De éste modo, la incorporación de los electrones al complejo I da como resultado final 38 ATP, mientras que si se incorporan al complejo III se obtienen 36 ATP como resultado final 7.- ANABOLISMO AUTÓTROFO 7.1. FOTOSÍNTESIS. Consiste en la obtención de moléculas orgánicas (glúcidos) a partir de energía lumínica y de moléculas inorgánicas como CO2 y H2O. Los pigmentos fotosintéticos son moléculas especializadas en captar la energía lumínica. Entre los pigmentos fotosintéticos mas conocidos figuran las clorofilas presentes en los vegetales y en la cianobacterias.; las bacterioclorofilas, propias de la mayoría de los procariotas fotosintéticos, y los carotenoides. Las clorofilas y los demás pigmentos se asocian formando complejos proteicos llamados fotosistemas, que están integrados en las membranas de los tilacoides. Cada fostosistema está formado por dos partes: * El complejo antena. En él predominan los pigmentos; éstos capturan y ceden energía por la actuación de sus electrones excitándose por la energía solar y volviendo rápidamente a su estado original. La energía () se va canalizando de unos pigmentos a otros como si se tratara de un embudo hasta * El centro de reacción. Donde predominan las proteínas sobre los pigmentos; aquí se encuentra el pigmento diana, el primer aceptor de electrones y el primer donador de electrones. En los vegetales y en las cianobacterias hay dos tipos de fotosistemas llamados P680 (II) y P700 (I). En el desarrollo de la fotosíntesis se distinguen dos fases, una dependiente de la luz, en la que se forma ATP y NADPH y otra independiente de la luz, en la que se obtienen gúcidos. 7.1.1. Fase luminosa Esta fase tiene lugar en la membrana tilacoidal. Fotofosforilación no cíclica. Se produce en los organismos fotosintéticos oxigénicos, que contiene dos fotosistemas y se llama fotofosforilación no cíclica, porque los electrones siguen un recorrido abierto (esquema en “Z”) desde un donador primario, el agua, hasta un aceptor final, el NADP+ el transporte electrónico se produce de la siguiente manera: * Las moléculas antena del fotosistema II absorben los fotones de luz y transfieren la energía lumínica al P 680. * El P 680 cede electrones a una cadena transportadora de electrones constituida por la feofitina (Ph), que constituye el primer aceptor electrónico; plastoquinona (QA), molécula liposoluble muy similar a la que se encuentra en las cadenas mitocondriales; etc. * Estos electrones llegan al complejo del citocromo bf, el cual enlaza ambos fotosistemas. * Por efecto de la energía luminosa se produce la fotolisis de una molécula de agua y se ceden electrones que pasan al P 680 para que recupere los electrones transferidos a las moléculas transportadoras. De la rotura de la molécula de agua se libera O2 y H+. * Las moléculas antena del fotosistema I también absorben los fotones de luz, cuya energía se transfiere al P 700. * La pérdida de los electrones del P 700 es compensada por los electrones del fotosistema II. Los electrones son transportados mediante el complejo del citocromo bf que los transfiere a la plastocianina (PC) y ésta al P 700. * El P 700 cede electrones a una segunda cadena de moléculas transportadoras, entre las que se encuentran la filoquinona (A), la ferredoxina (Fd), etc. La última molécula de la cadena transfiere los electrones al NADP+, y da lugar al NADPH. * La transferencia de electrones va acompañada del bombeo de protones al espacio tilacoidal. Los protones vuelven al estroma a través del canal de la subunidad F0 del complejo ATP sintasa. ( La subunidad F0 se encuentra unida a la membrana tilacoidal y forma una estructura en canal por la que circulan los protones desde el espacio tilacoidal hasta el estroma). * En la subunidad F1 tiene lugar la reacción de síntesis de ATP a partir de ADP y del Pi. (La subunidad F1 se localiza en el lado del estroma y se mantiene unida a la membrana por F0. Contiene el sitio activo para la síntesis de ATP) El flujo de electrones descrito no es cíclico sino unidireccional, y como consecuencia de ello se producen ATP y NADPH que se utilizarán en la siguiente fase de la fotosíntesis (fase oscura) Fotofosforilación cíclica Cuando las células vegetales tienen suficiente NADPH pero siguen necesitando ATP, se produce una ruta alternativa en la que el flujo de electrones es cíclico. En ésta ruta solo participa el fotosistema I. * Las moléculas antena del fotosistema I absorben los fotones de luz, cuya energía se transfiere al P 700. * El P 700 cede electrones a la cadena formada por la filoquinona, la ferredoxina, etc. La ferredoxina los transfiere al complejo citocromo bf, por tanto, no se produce NADPH. * Los electrones transferidos al citocromo bf pasan a la plastocianina y de ésta al P 700, con lo que se compensa la pérdida de electrones de este centro de reacción. * Como consecuencia del flujo de electrones se origina un potencial electroquímico que proporciona la energía necesaria a la ATP sintasa para la síntesis de ATP o fotofosforilación cíclica. 7.1.2. Fase oscura (Ciclo de Calvin) En ésta fase se produce una secuencia cíclica de reacciones en las que el ATP y el NADPH procedentes de la fase luminosa proporcionan la energía necesaria y los protones para la reducción del CO2. Como consecuencia de ésta reducción, se sintetizan glúcidos como la glucosa. Esta fase también se denomina fase de fijación del CO2 debido a que se incorporan diversas moléculas de CO2 para la síntesis de glucidos. Esta fase se produce en el estroma. * El CO2 se condensa con la ribulosa- 1,5-bisfosfato, un glúcido de cinco átomos de carbono que se encuentra en el estroma del cloroplasto. De ésta unión se forma un compuesto inestable de seis átomos de carbono que se escinde en dos moléculas de 3-fosfoglicerato, de tres átomos de carbono. La reacción es catalizada por el enzima Ribulosa-1,5-bisfosfato carboxilasa/oxigenasa (Ribulosa1,5-difosfatocarboxilasa). El aumento de pH en el estroma como consecuencia del bombeo de protones al espacio tilacoidal favorece la actividad de este enzima. En la oscuridad desciende la velocidad con la que actúa y temperaturas elevadas disminuyen su afinidad por el CO2. * El 3-fosfoglicerato se reduce a gliceraldehido-3-fosfato mediante el consumo de ATP y NADPH. * A continuación, tienen lugar diversas reacciones a partir de las cuales la mayor parte del gliceraldehido-3-fosfato se utiliza para la regeneración de la ribulosa-1,5-bisfosfato, consumiendo ATP. El gliceraldehido-3-fosfato restante se emplea en la síntesis de hexosas como la glucosa. * El gliceraldehido-3-fosfato también puede incorporarse a la glucólisis. * Algunos de los compuestos intermedios de las reacciones son precursores de la sacarosa y el almidón: la sacarosa se sintetiza a partir de la fructosa-1,6bisfosfato en el citosol y el almidón a partir de la fructosa-6-fosfato en el estroma. Ecuación global del ciclo de Calvin: Para que se produzca la síntesis de una molécula de glucosa, se han de fijar seis moléculas de CO2. Por tanto, han de producirse seis vueltas del ciclo para que se fijen seis CO2: 6 CO2 + 18 ATP + 12 NADPH + 12 H2O Glucosa + 18 ADP + 18 Pi + 12 + NADP + 6H+ Explicación: - Para incorporar 6 moléculas de CO2, necesitamos 6 de Ribulosa 1,5 difosfato y se obtienen 12 de 3fosfoglicerato. - A partir de las 12 moléculas de gliceraldehido obtenido (35 carbonos) se produce la síntesis de 1 molécula de hexosa (6 carbonos) y 6 moléculas de Ribulosa 1,5 difosfato (30 carbonos) - Para poder fosforilar éstas 6 moléculas de ribulosa.fosfato son necesarias otras 6 de ATP Para obtener una molécula de hexosa a partir de CO2 los organismos fotosintéticos gastan 12 NADPH y 18 ATP. 7.2. QUIMIOSÍNTESIS En el proceso de quimiosíntesis, a diferencia de la fotosíntesis, la energía necesaria para llevar a cabo el anabolismo autótrofo procede de procesos de oxidación de moléculas inorgánicas sencillas. La quimiosínteis la realizan un reducido número de bacterias y en ella podemos distinguir dos fases: * En la primera fase se obtiene energía en forma de ATP y coenzimas reducidos, a partir de la oxidación de compuestos inorgánicos sencillo. * En la segunda, la energía obtenida en la fase anterior es utilizada para reducir los compuestos inorgánicos, transformándolos en compuestos orgánicos. Esta segunda fase es similar a la fase oscura de la fotosíntesis, tanto para el carbono como para el nitrógeno. BLOQUE III LA HERENCIA Y GENÉTICA MOLECULAR CONCEPTO DE GENÉTICA. BREVE RECORDATORIO DE LAS APORTAIONES DE MENDEL Es el campo de la Biología que busca comprender la herencia biológica que se transmite de generación en generación. 1ª Ley de Mendel: Si cruzamos dos seres homocigóticos para un determinado carácter, todos los descendientes serán heterocigóticose iguales entre sí. 2ª Ley de Mendel: Si cruzamos dos heterocigóticos de la F1 entre sí, veremos que en la descendencia (F2) obtenemos todos los genotipos y fenotipos posibles siguiendo unas proporciones concretas. 3ª Leu de Mendel:Si estudiamos cómo pasan a la descendencia dos caracteres diferentes veremos que éstos se heredan de forma independiente cumpliendo con la primera y segunda ley 1.- CONSERVACIÓN DE LA INFORMACIÓN GENÉTICA. 1.1. AUTODUPLICACIÓN O REPLICACIÓN DEL DNA EN PROCARIOTAS.. El DNA portador de la información genética debe transmitirse fielmente a cada una de las células hijas obtenidas tras la división celular. Por tanto, antes de producirse ésta, es imprescindible que el DNA pueda formar rèplicas exactas de sí mismo para disponer de dos copias iguales. Este proceso, conocido como replicación o autoduplicación, resulta fundamental para asegurar que todas las células de un organismo pluricelular mantienen la misma identidad. Cuando Watson y Crick elaboraron su modelo de doble hélice en 1953, indicaros también cuál podía ser el mecanismo para llevar a cabo la replicación del DNA: separación de las dos cadenas y síntesis de la cadena complementaria de cada una de ellas. Sin embargo, otros investigadores plantearon distintas hipótesis que dieron lugar a tres posibles formas de replicación: - Conservativa. La doble cadena original se mantiene y se sintetiza otra completamente nueva. - Semiconservativa. Es la propuesta por Watson y Crick. Una de las hebras de cada doble hélice procede de la original, mientras que la otra se sintetiza nuevamente. - Dispersiva. En cada doble hélice existen fragmentos de la original y fragmentos nuevos. Poco después. Matthew Meselson y Franklin Sthal demostraron experimentalmente que la hipótesis correcta es la semiconservativa: La precisión necesaria en la replicación del DNA implica un mecanismo complejo para asegurar la obtención de copias idénticas. Baste decir que para que un ser humano se desarrolle a partir del zigoto, se calcula que deben producirse unos mil billones de divisiones celulares con otras tantas replicaciones previas del DNA. Si consideramos que existen 3500 millones de pares de nucleótidos en el genoma humano, es posible hacerse una idea de la magnitud del proceso. La replicación se lleva a cabo durante la fase S del ciclo celular 1.1.1. Inicio de la replicación. La replicación comienza en ciertas zonas del DNA donde existen determinadas secuencias de nucleótidos. En primer lugar interviene un enzima helicasa que separa las dos hebras de DNA al romper los puentes de hidrógeno entre las bases nitrogenadas complementarias. La separación y desespiralización de las dos hebras genera tensiones en ellas, que son eliminadas por la intervención de otras enzimas, las topoisomerasas. Para ello cortan una de las dos hebras (topoisomerasa I) o las dos (topoisomerasa II o girasa). Una vez separadas, las dos hebras se mantienen así por la acción de las denominadas proteínas SSB (single strand-binding). 1.1.2. Formación de las nuevas hebras A continuación comienza la síntesis de las hebras complementarias sobre cada una de las originales. El proceso se lleva a cabo mediante el enzima DNA polimerasa III, que presenta las siguientes características: - Necesita una hebra molde de DNA, que recorre en sentido 3´ 5´ y sobre la que se sintetiza la hebra complementaria. - Une nucleótidos en sentido 5´ 3´; es decir, la nueva hebra formada crece en este sentido. Los nucleótidos que se van uniendo son los que se sitúan previamente frente a los correspondientes nucleótidos complementarios del DNA molde. - Utiliza nucleótidos trifosfato, los cuales proporcionan, al mismo tiempo, la energía necesaria para la unión. - No puede comenzar la síntesis por sí misma, pues sólo puede añadir nucleótidos sobre el extremo 3´ libre de una cadena polinucleótida previa. Por este motivo es necesario que exista una cadena corta de RNA (de 40 o 50 nucleótidos), denominada RNA cebador. El RNA cebador es sintetizado por el enzima primasa que, utilizando DNA como molde, sintetiza RNA complementario de éste. A medida que la doble hélice del DNA original se va separando (de manera semejante a una cremallera que se abre), se forma la llamada burbuja de replicación donde se produce la acción de la DNA polimerasa III. Existe una horquilla de replicación en cada extremo, pues el proceso es bidireccional, es decir, avanza en ambas direcciones. Dado que la DNA polimerasa III recorre el DNA molde en sentido 3´ 5´, la síntesis de una de las hebras es continua, ya que, a medida que se abre la doble hélice, el enzima va avanzando y añadiendo nuevos nucleótidos a la cadena en formación, denominada hebra conductora. Sin embargo, como la otra cadena es complementaria, la DNA polimerasa debería recorrerla en sentido 5´ 3´ , añadiendo nucleótidos a la hebra en formación en sentido 3´ 5´ lo cual no es posible. La síntesis, en este caso es discontinua y se produce en segmentos separados. Esta cadena se denomina hebra retardada, pues su síntesis es mas lenta que la de la hebra conductora. Los segmentos de DNA sintetizados de este modo se conocen como fragmentos de Okazaki y constan de 1000 a 2000 nucleótidos. Cada fragmento de Okazaki requiere un RNA cebador para iniciar la síntesis de una secuencia de nucleótidos. Posteriormente, tras la eliminación de los RNA cebadores, los fragmentos de Okazaki se unen gracias a la acción de los enzimas ligasas. La enzima DNA polimerasa I elimina después los RNA cebadores gracias a su acción exonucleasa (rotura de enlaces fosfodiester a partir de un extremo nucleótido libre). La misma enzima posee también actividad polimerasa, por lo que puede rellenar el hueco dejado por el RNA cebador eliminado. 1.1.3. Finalización. Por último, cada hebra recién sintetizada y la que ha servido de patrón se disponen enrolladas originando una doble hélice. A pesar de todas éstas etapas, el proceso de replicación es muy rápido. En Escherichia coli, por ejemplo, se unen 45000 nucleótidos por minuto 1.2. DIFERENCIAS ENTRE EL PROCESO REPLICATIVO EN PROCARIOTAS Y EUCARIOTAS. Las diferencias en la replicación del DNA entre las células procariotas y eucariotas no afectan al mecanismo fundamental. Entre estas diferencias se pueden destacar las siguientes: - Como el DNA de los eucariotas está asociado con las histonas, la replicación debe tener en cuenta la síntesis de estas proteínas. Se ha comprobado que las histonas originales se mantienen en la hebra conductora, mientras que se forman nuevas histonas que se unen a la hebra de DNA retardada. - El tamaño de los fragmentos de Okazaki es menor en los organismos eucariotas (100 a 200 nucleótidos) que en los procariotas (1000 a 2000 nucleótidos). - Existen tres DNA polimerasas en los procariotas y cinco en eucariotas. - La replicación tiene un único origen en los procariotas, mientras que en los eucariotas existen múltiples (cientos en cada cromosoma de mamíferos, lo que hace que haya varios miles en el conjunto de su genoma). Cada unidad de replicación se denomina replicón y produce la síntesis de fragmentos de 100 a 150 nucleótidos. La necesidad de numerosos puntos origen de la replicación resulta evidente, pues la cantidad de DNA en las células eucariotas es muchísimo mayor. Si sólo existiera un lugar de inicio, el proceso de replicación necesitaría varios meses para llevarse a cabo. - La velocidad de replicación en cada replicón es menor en los eucariotas (hasta 50 veces)) que en los procariotas. 2.- EXPRESIÓN DE LA INFORMACIÓN GENÉTICA 2.1. TRANSCRIPCIÓN DEL DNA EN EUCARIOTAS (SÍNTESIS DE ARNm). Durante el proceso se pueden distinguir las siguientes fases: 2.1.1. Inicio. En la cadena de DNA se encuentran unas secuencias especiales que reciben el nombre de secuencias promotoras o promotores. Estas secuencias se sitúan antes del primer nucleótido que debe ser transcrito y que identificaremos como nucleótido +1. Aproximadamente, las secuencias promotoras se suelen situar centradas en la posición –25, es decir, a unos 25 nucleótidos del lugar de inicio de síntesis de RNA. Esta secuencia ha sido identificada para numerosos genes y en numerosas especies y se observa una elevada coincidencia en la secuencia TATA; por ello se denomina caja TATA. Las proteínas denominadas factores de transcripción (TF) identifican las cajas y se unen a ellas, para facilitar, a continuación, la ubicación correcta de la RNA pol sobre la cadena de DNA. Seguidamente se inicia la síntesis de RNA a partir del nucleótido +1. En las células eucariotas existen tres tipos de rNA polimerasas especializadas en la síntesis de diferentes tipos de RNA: - La RNA pol I interviene en la síntesis de las subunidades grandes de los ribosomas. - La RNA pol II es la responsable de la síntesis de los precursores de los RNA mensajeros, que se traducirán a proteínas. - La RNA pol III controla la síntesis de los RNA de transferencia y de las subunidades pequeñas de los ribosomas. Vamos a seguir la descripción del proceso considerando el caso de la síntesis de un mRNA. 2.1.2. Elongación. La RNA pol II va recorriendo la doble hélice y utiliza como molde una de las dos cadenas. Esta cadena se va leyendo desde el extremo 3´ hacia el 5´. Al mismo tiempo, se van uniendo los ribonucleótidos uno tras otro y la cadena va creciendo en sentido 5´ 3´ . Los ribonucleótidos se van situando siguiendo la ley de complementariedad de bases, teniendo en cuenta que el ribonucleótido complementario de la adenina es el uracilo. Cuando se han transcrito una 30 bases del gen, al mRNA en formación se le añade una caperuza compuesta por un resto de guanosina metilada unida a un grupo trifosfato, (como se verá mas adelante, ésta caperuza es reconocida por los ribosomas como lugar de inicio de la traducción). A medida que se va desprendiendo la cadena de mRNA recién sintetizada, el DNA recupera su estructura espacial normal. 2.1.3. Terminación El proceso finaliza cuando el mRNA polimerasa II transcribe la secuencia TTATTT. Inmediatamente después actúa otro enzima que se llama poli-A polimerasa, que adiciona en el extremo 3´ del mRNA una cola “cola poli-A” formada por unos 150-200 nucleótidos de adenina. Esta molécula recién sintetizada, que presenta secuencias intrónicas y exónicas alternantes y posee caperuza y cola se llama mRNA transcrito primario. 2.1.4. Maduración. Debido a que los genes en eucariotas se encuentran fragmentados, los mRNA transcritos primarios contienen secuencias intercaladas (intrones) que no codifican para la síntesis de proteínas y, por tanto, deben ser eliminados. La supresión de éstas secuencias se realiza mediante un proceso de maduración que supone cortes entre los intrones y exones; los primeros se enrollan en lazos y se eliminan, mientras que los segundos se empalman yo forman una molécula de mRNA, que contiene todos los nucleótidos necesarios para la síntesis de proteínas. Los cortes en los transcritos primarios, tanto en los precursores de los tRNA y rRNA, se realizan con la ayuda de una proteína enzimática llamada ribonucleoproteína pequeña nuclear (RNPpn); el posterior empalme de los fragmentos exónicos se efectúa mediante el concurso de ligasas especificos. A continuación el mRNA se desplaza hacia el citoplasma para la síntesis de proteínas. Este transporte tiene lugar gracias al reconocimiento específico por parte de proteínas situadas en los poros de la envoltura nuclear que, mediante transporte activo, permiten el paso de mRNA. El tRNA y rRNA, transcritos mediante los enzimas RNA pol I y RNA pol III, sufren un proceso de maduración algo distinto, que incluye la adquisición de su configuración espacial correcta. Posteriormente, salen al citoplasma e intervienen, también, en la síntesis de proteínas. 2.2. DIFERENCIA CON PROCARIOTAS. En los procariotas los genes son continuos, es decir, las moléculas de mRNA son copias literales de los genes, que no necesitan ningún tipo de transformación previa a la traducción por los ribosomas. Sin embargo, como acabamos de decir, en eucariotas, existen secuencias intrónicas que no codifican para la síntesis de proteínas y exónicas que sí lo hacen; por lo que las secuencias intrónicas deben ser eliminadas en un proceso de maduración. 3.- EL CODIGO GENÉTICO Una vez obtenida una copia del mensaje genético en forma de cadena de mRNA, ésta dirige la síntesis de proteínas en los ribosomas. Para ello, éstos orgánulos interpretan la secuencia concreta de nucleótidos existente en la molécula de mRNA como la información necesaria para la unión de los aminoácidos precisos para construir la proteína específica. Se denomina código genético a la relación entre la secuencia de nucleótidos (o, mas concretamente, de bases nitrogenadas presentes en ellos) del mRNA y la secuencia de aminoácidos que constituye una proteína. Consiste, pues, en una equivalencia entre polímeros específicos. Uno de ellos, el RNA, tiene dispuestas sus bases nitrogenadas en una secuencia concreta que contiene la información que determina el orden en que han de engancharse los sucesivos aminoácidos que forman la cadena polipeptídica. Por tanto, los mRNA con secuencias de bases nitrogenadas distintas llevan información para la síntesis de proteínas diferentes. El código genético es, en definitiva, la clave que permite la traducción del mensaje genético a su forma funcional, las proteínas. Como sólo hay cuatro bases nitrogenadas distintas, las señales codificadoras para los 20 aminoácidos proteicos deben estar constituidas por más de una base. Si cada señal estuviera formada por dos bases nitrogenadas, solo codificarian 42 = 16 aminoácidos, por lo que aun quedarían aminoácidos sin codificar. Por tanto, cada señal que codifica para un aminoácidos está constituido por tres bases nitrogenadas consecutivas (un triplete), es decir, 43 = 64 tripletes de bases distintas. Los tripletes de bases del mRNA reciben el nombre de codones. Los tripletes del DNA correspondientes, que han sido transcritos, se denominan codógenos. Existen 61 codones codificadores de aminoácidos y 3 (UAA, UAG y UGA, llamados sin sentido) que señalan el final del mensaje y no especifican ningún aminoácido. Hay también un codón (AUG) que, además de codificar para el aminoácido metionina, es la señal de comienzo. 3.1. CARACTERÍSTICAS DEL CÓDIGO GENÉTICO. nitrogenadas. - Está formado por una secuencia lineal de bases - Entre los sucesivos codones no hay espacios ni separaciones de ningún tipo, ya que entre el tercer nucleótido de uno de ellos y el primero del siguiente existe la misma distancia que entre dos nucleótidos de un mismo codón. - Tiene carácter universal, ya que es el mismo código para todas las células de todas las especies. - El código está degenerado en el sentido matemático del término. Esto significa que no existe el mismo número de señales codificadoras en el RNA que aminoácidos van a ser codificados. Salvo el triptófano y la metionina, que están codificados por un único codón, los demás aminoácidos están codificados por mas de un triplete. Generalmente sólo se diferencian en la última base. La existencia de codones distintos que codifican el mismo aminoácido no constituye una falta de precisión o un fallo del código, ya que ante un mensaje de RNA concreto únicamente se forma una proteína específica, sin posibilidad de falsas interpretaciones. 3.2. TRADUCCIÓN Una vez transcrito el DNA, la molécula de mRNA formada contiene la información necesaria para la síntesis de la proteína correspondiente. La unión de los aminoácidos se realiza mediante enlaces peptídicos. La traducción se lleva a cabo en los ribosomas. Antes de que se inicie propiamente la síntesis de proteínas es preciso que los aminoácidos que van a ser unidos se activen. En ésta fase previa, que tiene lugar en el citoplasma y no en los ribosomas, cada aminoácido se une a una molécula de tRNA específica gracias a la acción de los enzimas aminoacil tRNA sintetasas. Para ello es necesario el aporte energético obtenido por la hidrólisis de ATP, que pasa a AMP y dos grupos fosfato. El aminoácido queda unido por su extremo carboxilo al extremo 3´ del tRNA (concretamente, al grupo hidroxilo del carbono 3´ del último nucleótido, que siempre lleva adenina). Las moléculas de tRNA actúan como sistemas intermediarios entre la secuencia de nucleótidos y la secuencia de aminoácidos, ya que, además de llevar unido un aminoácido en su extremo 3´, reconocen los nucleótidos del mRNA gracias a su anticodón complementario del codón correspondiente. Una vez activados los aminoácidos por la formación de los aminoacil tRNA tiene lugar la síntesis de proteínas, que comienza incluso cuando aún no ha acabado la formación del mRNA por completo. El proceso se lleva a cabo en tres etapas: 3.2.1. Iniciación. En la fase de iniciación, todo el sistema se prepara para llevar a cabo la síntesis proteica: el mRNA se une a los ribosomas citoplasmáticos, cuyas dos subunidades, que se encuentran separadas cuando no están realizando su función, deberán acoplarse. - En primer lugar, el mRNA se une por su extremo 5´ a la subunidad menor del ribosoma gracias a un factor proteico de iniciación (IF3). - A continuación se produce la fijación del primer aminoacil tRNA por la formación de puentes de hidrógeno entre las bases complementarias del anticodón del tRNA y las del codón del mRNA. El primer codón o codón de iniciación siempre es 5´AUG 3´ y, por tanto, el anticodón del primer tRNA es UAC. El aminoácido unido a este primer tRNA es la formil metionina, por lo que supuestamente todas las cadenas proteicas deberán empezar por éste. Dado que no es así, posteriormente éste primer aminoácido suele ser eliminado. En el proceso de fijación entre ambos RNA interviene otro factor de iniciación (IF2). - Por último, se produce el acoplamiento de la subunidad mayor del ribosoma, para lo cual se precisa otro factor de iniciación diferente (IF1). Queda formado así el denominado complejo de iniciación, que constituye la maquinaria sintetizadora activa. La porción de mRNA cubierta por el ribosoma corresponde a seis nucleótidos, es decir, a dos codones. Sobre el primero de ello, AUG, ya está situado el aminoacil tRNA correspondiente, en el lugar denominado sitio P. La zona donde se encuentra el segundo codón es el sitio A. El proceso de iniciación precisa energía, que se obtiene por la hidrólisis del GTP. 3.2.2. Elongación o alargamiento. En ésta etapa la cadena peptídica se sintetiza por la unión de los sucesivos aminoácidos que se van situando en el ribosoma transportados por los correspondientes tRNA. Para ello es necesario el desplazamiento del ribosoma a lo largo de la cadena del mRNA. En éste proceso se pueden diferenciar tres etapas: - Unión de un aminoacil tRNA al sitio A. Esto solo es posible si el anticodón del tRNA es complementario del codón del mRNA que se encuentra allí. En ésta subetapa se precisa la hidrólisis de GTP para proporcionar la energía necesaria y dos factores proteicos de elongación (EF-Ts y EF-Tu). - Formación del enlace peptídico. Una vez anclados los dos aminoacil tRNA, uno en el sitio P y otro en el sitio A, se produce la unión entre los dos aminoácidos gracias al enzima peptidil transferasa, localizado en la subunidad mayor del ribosoma. Al unirse el primer aminoácido (formil metionina) al segundo se desprende de se tRNA, el cual se libera del ribosoma, Se forma de ésta manera un dipéptido que permanece unido al segundo tRNA, el cual se localiza en el sitio A. - Translocación del dipéptido al sitio P. Se produce el desplazamiento del ribosoma sobre el mRNA en sentido 5´ 3´. Así, el segundo codón con el tRNA fijado sobre él (el cual lleva a su vez el dipéptido recién sintetizado) pasa al sitio P, quedando libre el sitio A, que es ocupado por el tercer codón del mRNA. Sobre éste se fija un nuevo aminoacil tRNA, con la participación de otro factor de elongación (EG-G). A continuación se forma un nuevo enlace peptídico entre éste aminoácido y el dipéptido situado en el sitio P, con lo que todo el proceso de translocación descrito comienza nuevamente. De este modo, mientras el ribosoma recorre el mRNA, los sucesivos aminoacil tRNA que se van fijando en el sitio A van incorporando su aminoácido correspondiente a la cadena peptídica en formación mediante la acción del enzima peptidil transferasa. En la fijación de cada tRNA se utiliza la energía suministrada por la hidrólisis de GTP. Debido a la complementariedad existente entre los anticodones del tRNA y los codones del mRNA, la secuencia de bases nitrogenadas de este último se refleja en la secuencia de aminoácidos de la proteína que se sintetiza. 3.2.3. Terminación. Existen tres codones de terminación (UAA, UAG y UGA) en el mRNA para los que no hay tRNA con los correspondientes anticodones. Por esta razón, cuando el ribosoma llega a uno de ellos, no se sitúa ningún aminoacil tRNA en el sitio A y la cadena peptídica se acaba. En esta fase intervienen unos factores de liberación: R1F para los codones de terminación UAA y UAG, y R2F para los codones UAA y UGA. Al situarse en el sitio A, estos factores hacen que el enzima peptidil transferasa libere el péptido del tRNA al que está unido, al hacer que reaccione el grupo carboxilo del último aminoácido con agua. También en éste proceso se utiliza la energía que proporciona el GTP. Como consecuencia del proceso de traducción se libera: - La cadena proteica que, conforme se ha ido sintetizando, ha adquirido su estructura secundaria y terciaria característica. - Las dos subunidades ribosómicas separadas. - El mRNA, que puede volver a ser utilizado, aunque por lo general es destruido inmediatamente e incluso, en ocasiones, se degrada según es “leído” por lo ribosomas. La velocidad de síntesis proteica es alta: se pueden unir hasta 1400 aminoácidos por minuto. 4.- GENES Y REGULACIÓN GÉNICA 4.1. CONCEPTO DE GEN. Es un segmento de DNA que, por regla general, contiene la información necesaria para la síntesis de una proteína. Desde el punto de vista de la Genética clásica, lo genes son las unidades estructurales y funcionales de la herencia, transmitidos de padres a hijos a través de los gametos. 4.2. REGULACIÓN DE LA EXPRESIÓN GÉNICA. La información genética codificada en la secuencia de nucleótidos del DNA origina la síntesis de las proteínas correspondientes, tras los procesos de transcripción y traducción. Sin embargo, la síntesis proteica no tiene lugar continuamente, ya que depende de las necesidades celulares. La producción excesiva de proteínas metabólicas resulta innecesaria y casi siempre ocasiona alteraciones importantes. Por tanto, para evitar el despilfarro de moléculas y energía, los genes sólo se expresan cuando es necesario sintetizar las proteínas adecuadas en cada momento de la vida celular. Las necesidades proteicas, y en particular, las enzimáticas dependen, de las variaciones del medio extra e intracelular. En los seres pluricelulares, por otra parte, los genes que se expresan son distintos según el tipo de célula de que se trate. De hecho, la diferenciación celular se basa en la activación e inhibición permanente de diversos genes que permite alcanzar un alto grado de especialización, origen de la existencia de diversos tejidos en un mismo organismo. De lo anterior se deduce que el control de la expresión génica es imprescindible en todos los seres vivos, aunque es mas complejo en los organismos pluricelulares, en los que se debe conseguir la especialización celular. La regulación de la expresión génica se realiza fundamentalmente sobre el proceso de transcripción, ya que si se controla la formación de mRNA se controla también la síntesis de las proteínas correspondientes. A principios de la década de 1960, Jacob y Monod propusieron un modelo para explicar la regulación de la transcripción en cepas mutantes de la bacteria Escherichia coli. De acuerdo con éste modelo, denominado del operón, existen unas proteínas reguladoras que controlan la transcripción de los genes que codifican para los enzimas implicados en una ruta metabólica determinada. Se distinguen cuatro tipos de genes: - Genes estructurales. Contienen la información para la síntesis de los enzimas que intervienen en la ruta metabólica. - Gen promotor. Está constituido por la secuencia de DNA donde se une la RNA polimerasa para comenzar la transcripción. - Gen operador. Es el lugar del DNA donde puede unirse una proteína reguladora e impedir la transcripción de los genes estructurales. Se sitúa inmediatamente delante de éstos. - Gen regulador. Sintetiza la proteína reguladora. Puede encontrarse en un lugar distinto a los otros genes del operón. Según se trate de una ruta catabólica o anabólica se diferencian dos tipos de regulación de la expresión génica: inducible y represible. 4.2.1. Sistema inducible. Es característico de los procesos catabólicos. La proteína sintetizada por el gen regulador es un represor activo que, al unirse al gen operador, impide la acción de la RNA polimerasa sobre los genes estructurales. Por tanto, no se formarán los enzimas de la ruta metabólica y ésta no se podrá realizar. Cuando aparece el sustrato inicial de ésta ruta, la proteína represora cambia su conformación, volviéndose inactiva, y deja de actuar sobre el gen operador. La inactivación del represor se consigue por la unión a una molécula inductora, que puede ser el mismo sustrato o un derivado de éste, de modo que se favorece la acción de la RNA polimerasa sobre los genes estructurales, los cuales se transcriben y sintetizan los enzimas correspondientes. Este mecanismo permite la síntesis de proteínas enzimáticas solamente cuando son necesarias. Hasta que no hay sustrato que catabolizar no existen los enzimas para hacerlo, pues el gen represor, unido al gen operador, impide la transcripción. Sin embargo, la presencia del sustrato inactiva al represor e induce la síntesis de los enzimas que llevan a cabo la ruta metabólica. Ej: El operón lac (de lactosa) constituye un sistema inducible que permite la regulación de la síntesis de los enzimas que intervienen en el catabolismo de la lactosa por Escherichia coli. Existen tres genes estructurales que codifican para los tres enzimas que participan en la ruta metabólica (una permeasa, un acetil transferasa y la beta-galactosidasa). La alolactosa, derivada de la lactosa, es el inductor que modifica la estructura del represor inactivándolo. La cantidad de los enzimas del operón lac aumenta cerca de 1000 veces, tan solo dos minutos después de la aparición de la lactosa, de los que se deduce que este sistema de activación génica es sumamente eficaz. 4.2.2. Sistema represible. Se da en procesos anabólicos. A diferencia del sistema inducible, el represor sintetizado por el gen regulador es inactivo y, por tanto, la RNA polimerasa actúa y los genes estructurales se transcriben. La molécula represora se activa únicamente al cambiar su conformación por la unión a un correpresor, el cual constituye el producto final de la ruta metabólica. La activación del represor permite su unión al gen operador impidiendo la transcripción de los genes estructurales. De esta forma se producen los enzimas que participan en la ruta anabólica hasta que existe suficiente cantidad del producto final. En este caso, el represor se hace activo y se inhibe la transcripción, con lo cual se evita una síntesis excesiva del producto. 5.- MUTACIONES GÉNICAS Y CÁNCER Las mutaciones génicas son las mutaciones en sentido estricto. Consisten en cambios químicos en el ADN, por lo que resulta imposible observarlas al microscopio. Las alteraciones afectan tanto a los genes estructurales como a los genes reguladores y pueden provoca cambios en un par de bases (microlesiones) o en un segmento génico (macrolesiones). Aparecen fundamentalmente por dos causas: Errores no corregidos durante la replicación del ADN La acción de determinados agentes físicos o químicos, como las radiaciones o algunas sustancias procedentes del exterior de la célula o del propio metabolismo, que alteran el ADN. Se distinguen varios tipos de mutaciones génicas: Mutaciones por sustitución de una base por otra distinta. Se dividen en dos tipos: las denominadas transiciones, cuando una base púrica es sustituida por otra base púrica o una pirimidínica es reemplazada por otra pirimidínica, y las transversiones, si se produce la sustitución de una base púrica por otra pirimidínica o viceversa. Mutaciones por pérdida o inserción de bases. Son más graves que las anteriores, ya que, a partir del punto de delección o de adicción, todos los tripletes de bases estarán cambiados. El mensaje codificado será totalmente diferente. Mutaciones por variaciones de lugar de algunos segmentos del ADN (transposiciones). Debido al desplazamiento de secuencias de la cadena nucleotídica aparecen nuevos tripletes, lo que modificará el mensaje genético. El cáncer es causado por un proceso de división celular sin control que provoca una multiplicación rápida y desorganizada de las células que conduce a la destrucción del tejido afectado e, incluso, a la invasión de otros órganos (metástasis). Aunque el desencadenamiento de un proceso cancerígeno intervienen diversos factores, hoy en día queda fuera de toda duda la relación que existe entre determinados cambios en el material genético y la aparición de células cancerosas, ya que con frecuencia se observan en ellas la presencia de alteraciones cromosómicas, como delecciones, translocaciones y roturas o uniones cromosómicas. Por otra parte, ciertos agentes mutagénicos también son cancerígenos, por ejemplo, radiaciones, ciertos virus y determinados productos químicos. No se conoce totalmente el proceso por el que una célula normal se transforma en cancerosa, pero se han logrado progresos importantes en su investigación. Básicamente se producen defectos en determinados genes que participan en la regulación de la división celular, por lo que ésta se descontrola y se vuelve caótica. En este proceso intervienen dos tipos de genes: * Oncogenes. Provocan un aumento de las señales que estimulan la división celular, sin que estén presentes los estímulos normales para ello. De ésta forma, se promueve la proliferación contínua de las células Actualmente se cree que los oncogenes proceden de otros genes, denominados protooncogenes, que codificn proteínas implicadas en determinadas etapas de la división celular. La alteración de los protooncogenes por agentes mutagénicos originaría los oncogenes activos. * Genes supresores de tumores. La mutación de éstos genes, que codifican proteínas inhibidoras de la división celular, estimula un aumento del ritmo reproductor de las células. Así, los agentes mutagénicos podrían actuar en ambos sentidos y es probable que para que se desarrolle un tumor sean necesarias varias mutaciones en diversos genes. Existen indicios, así mismo, de que en le proceso de transformación cancerosa de una célula intervienen otros agentes que la potenciarían favoreciendo la expresión de los oncogenes. Por otra parte, la mutación de los genes implicados en la corrección de errores del ADN evitaría la reparación de éstos tras la acción del agente mutagénico, en las primeras fases del proceso, y contribuiría notablemente al desarrollo definitivo del tumor. BLOQUE IV EL MUNDO DE LOS MICROORGANIMOS Y SUS APLICACIONES 1.- MICROBIOLOGÍA: CONCEPTO La microbiología es la ciencia que estudia los microorganismos, seres visibles únicamente con el microscopio, que son capaces, sin embargo, de llevar a cabo todas las funciones vitales que caracterizan a los seres vivos. Los microorganismos componen un grupo muy heterogéneo. Algunos, como las bacterias, poseen una estructura típicamente procariota, mientras que otros, los protistas y los hongos, tienen una estructura eucariota. Por lo general son unicelulares o aparecen formando asociaciones sencillas, y todos ellos, presentan la mas amplia gama de capacidades metabólicas entre los organismos vivientes, lo que les ha permitido colonizar una gran variedad de ambientes. Estos pequeños “animalucos” como los denominó Leeuwenhoek, pueden tener efectos perjudiciales para el ser humano, ya que son responsables de muchas enfermedades. Por otra parte, los microorganismos desempeñan un papel esencial en los ciclos biogeoquímicos de la naturaleza y su utilidad práctica en el ámbito de la Biotecnología es innegable (fermentaciones, eliminación de residuos, ingeniería genética...) 2. LOS MICROORGANISMOS EN LOS CINCO REINOS Reino Moneras. Son procariotas unicelulares. Bacterias Eubacterias (son las bacterias verdaderas, ya estudiadas) Arqueobacterias. Son organismos relacionados filogenéticamente con los eucariotas, que presentan, sin embargo, una organización procariota típica. Entre las características mas importantes que definen a éste grupo destacan las siguientes: - Organización general procariota - La pared celular no contiene peptidoglicano - La membrana presenta lípidos exclusivos en los que el glicerol se une por enlace de tipo éter a las cadenas laterales hidrofóbicas. - Realizan metabolismo fundamentalmente respiratorio en condiciones aeróbicas y anaeróbicas. - Se desarrollan en medios ambientales extremos. Esta característica permite distinguir tres grupos de arqueobacterias: especies halófilas, que proliferan en medios hipersalinos; termoacidófilas, propias de ambientes con temperaturas extremas o altamente ácidos; y metanógenas, bacterias que producen metano en ambientes anaerobios, como, por ejemplo, el estómago de los rumiantes, en los que contribuye a la digestión de los productos vegetales junto con otras bacterias celulolíticas e hidrolíticas. Cianofíceas. Unicelulares y procariotas. Se cree que fueron los primeros organismos fotosintéticos. Tienen pigmentos azules. Son autótrofas y hacen simbiosis con algunos helechos porque son capaces de fijar el nitrógeno atmosférico dándoselo a dicha planta. Mycoplasmas . Son unas células procariotas y habitualmente parásitos de animales y del hombre, en los que provocan diversas enfermedades como la neumonía atípica Algunos mycoplasmas contienen esteroles en su membrana, lo que le proporciona a esta estructura una rigidez mayor.. Su característica más importante es que carecen de pared celular, y por ello son difícil de atacar por los antibióticos. Reino Protoctista Protozoos. Son organismos móviles, heterótrofos y organotrófos, aunque comprenden algunas especies fotosintéticas con cloroplastos, como el grupo de los fitoflagelados. El sistema de locomoción presenta formas variadas: en algunas especies consiste en apéndices locomotores (flagelos o cilios), y en otras, en la formación de corrientes que originan unas prolongaciones hialinas del citoplasma denominadas pseudópodos. En algunos parásitos, el movimiento se produce por deslizamiento o flexión de la célula. En general, los protozoos se dividen asexualmente por escisión binaria, aunque también se puede dar la reproducción sexual cuando las condiciones ambientales son adversas (especialmente si hay escasez de nutrientes). Algunos protozoos presentan quistes de resistencia (células sin metabolismo apreciable de una gruesa cubierta) o esporas, formas de resistencia mas pequeñas que el individuo vegetativo y rodeadas de una envuelta característica, que se originan a menudo en unas células denominadas esporoblastos o esporoquistes. En función del tipo de locomoción, nutrición, ciclo vital y ecología, los protozoos se clasifican en distintos grupos que incluyen formas de vida libre y parásitas: ciliados, tienen cilios (paramecios); flagelados, tienen flagelos (Tripanosoma, que produce la enfermedad del sueño); rizopodos, con falsos pies (amebas); esporozoos, se reproducen por esporas (plasmodios) Algas microscópicas. Son organismos eucariotas con cloroplastos, orgánulos especializados en un tipo de metabolismo fotosintético y autótrofo, cuyo pigmento fundamental es la clorofila. La mayoría de las algas tienen pared celular, cuyo polímero fundamental es la celulosa o la quitina; otros grupos, sin embargo, carecen de pared celular. Los polímeros carbonados de reserva de la célula son muy variados y sólo en algunos grupos se sintetiza el almidón (como en las plantas superiores). Las algas son organismos acuáticos y forman los componentes fundamentales del fitoplancton, aunque también es posible encontrarlas en suelos, rocas, troncos de árboles o superficies húmedas. Entre los grupos mas característicos se encuentran: diatomeas, poseen un esqueleto formado por carbonato cálcico; euglenofitas, tienen dos flagelos; dinoflageladas, tienen flagelos y pigmentos rojos (producen a veces las mareas rojas). Reino Fungí Se suelen denominar hongos a un grupo de seres vivos muy heterogéneo. Todos ellos están constituidos por células eucariotas y son heterótrofos. Las diferencias entre los distintos tipos se establecen, básicamente, por su estructura y el modo en que captan el alimento. Los hongos ameboides tienen una estructura que recuerda a las amebas. Están formados por células sin pared y plurinucleadas, la mayoría de las especies captan el alimento por fagocitosis. El resto de los organismos conocidos como hongos corresponden a los hongos conocidos como lisotróficos (digestión externa). Estos tienen una estructura filamentosa, con pared celular exterior a la membrana plasmática. Son heterótrofos, y para su alimentación segregan las sustancias necesarias para la digestión del alimento y luego absorben los nutrientes resultado de la digestión. Levaduras. Algunos grupos de hongos tienen, de manera permanente, una estructura unicelular, también denominada levuriforme, y que se conocen con el nombre de levaduras. La célula de las levaduras tiene forma ovalada; hay especies con pared celular y especies que no la presentan. Las células poseen el resto de los orgánulos propios de una célula eucariota. Como inclusiones suele contener materiales de reserva como lípidos y polifosfatos. Las células pueden ser haploides o diploides, éstas últimas, algo mayores y mas activas que las primeras. 3.- PRIONES,VIROIDES Y PLÁSMIDOS.(FORMAS ACELULARES) Los viroides son moléculas de RNA monocatenario circular con forma de varilla, no asociado a proteínas. En los vegetales provocan enfermedades relacionadas principalmente con el crecimiento. Los viroides no codifican para proteínas y es probable que su efecto se produzca por la interacción con el genoma de la célula hospedadora o con su control. Los priones, están constituidos exclusivamente por proteínas. Se asocian a enfermedades degenerativas del sistema nervioso central, de desarrollo lento, en el ser humano y en los animales, como el prurito lumbar de las ovejas, la enfermedad de las vacas locas o la enfermedad de Creutzfeldt-Jacob, que produce demencia en el hombre. Los plásmidos son fragmentos de DNA bicatenario que contienen unos cuantos genes y se encuentran en el interior de algunas bacterias. Actúan y se replican de forma independiente al DNA bacteriano y pueden pasar de unas bacterias a otras. No producen enfermedades pero inducen pequeñas mutaciones en las bacterias. 3.1. ESTRUCTURA Y CLASIFICACIÓN DE LOS VIRUS 3.1.1 ATENDIENDO A SU ESTRUCTURA. La partícula vírica o virión está constituida por un fragmento de ácido nucleico encerrado en una cubierta proteica o capsida. Algunos virus presentan, además, una envoltura membranosa (envoltura lipoproteica), compuesta por una bicapa lipídica procedente de la célula hospedadora parasitada y asociada a proteínas víricas que proyectan salientes hacia el exterior denominadas espículas; estos virus se denominan virus con envoltura, en contraposición a los virus desnudos, que carecen de ella. El ácido nucleico de los virus puede ser DNA o RNA, ( nunca los dos), monocatenario (cadena sencilla) o bicatenario (cadena doble). La cápsida está formada por capsómeros, unidades estructurales constituidas por una o varias subunidades proteicas denominadas protómeros. Las proteínas de la cápsida se organizan regularmente alrededor del ácido nucleico, de manera que la nucleocápsida (cápsida + ácido nucleico) presenta una simetría determinada que caracteriza la morfología del virión. A) Virus con simetría helicoidal Son virus con forma de varilla, en los que los capsómeros, por lo general con un solo tipo de proteína, se disponen helicoidalmente alrededor del ácido nucleico. La mayor parte de ellos contienen RNA monocatenario (virus del mosaico del tabaco). B) Icosaedricos Los capsómeros que suelen ser de varios tipos, se ajustan formando un icosaedro regular (es decir, 20 caras triangulares y 12 vértices), y dejando un hueco central donde se sitúa el ácido nucleico fuertemente apelotonado. Algunos forman poliedros con más caras que el icosaedro, y algunos presentan fibras proteicas que sobresalen de la cápsida. La mayoría de ellos tienen envoltura lipopreteica: - Entre los virus con RNA monocatenario se pueden citar los de la rabia, el sarampión, la gripe y la rubéola. - Los retrovirus contienen RNA monocatenario y la enzima transcriptasa inversa. Al infectar la célula, transcriben el RNA en una molécula de DNA bicatenario que se une al DNA celular. Pertenecen a éste tipo de grupo el virus del SIDA y los productores de algunos tipos de cáncer. - Entre los virus con DNA bicatenario se pueden citar el grupo de los herpesvirus, como los del herpes y de la hepatitis. Hay también virus de células animales icosaedricos sin envoltura lipoproteica: - El virus de la polio humana tiene RNA monocatenario - La mayor parte de los reovirus (con RNA bicatenario) infectan células animales - Los virus que contienen DNA bicatenario suelen ser poco virulentos, como los adenovirus (de los resfriados) y los virus de las verrugas (papovirus) C) Complejos Con pequeñas variantes, responden a la siguiente estructura general: - Una cabeza de estructura icosaedrica que alberga el ácido nucleico. - Una cola de estructura helicoidal que constituye un cilindro hueco. - Un collar de capsómeros entre la cabeza y la cola. - Una placa basal, al final de la cola, con unos puntos de anclaje que sirven para fijar el virus a la membrana celular. De la placa salen también unas fibras proteicas que ayudan a la fijación del virus sobre la célula hospedadora. La mayor parte de los virus complejos contienen DNA bicatenario como los que infectan a las bacterias (bacteriófagos). 3.1.2. ATENDIENDO AL HOSPEDADOR Los virus animales y vegetales presentan, en su mayoría, simetría icosaédrica o helicoidal, y en muchos casos osn virus con envuelta. En los tipos animales se encuentran todo tipo de ácidos nucleicos, mientras que en los vegetales presentan mayoritariamente ARN. La infección vírica por ambos tipos de virus puede ocasionar la destrucción de la célula hospedadora (infección lítica) o alteraciones citológicas y de crecimiento en procesos en los que las partículas on liberadas lentamente por gemación (infección persistente). Se han descrito también virus animales que permanecen latentes (infección latente) en ciertas células del organismo y se reactivan en presencia de diversos estímulos. Los virus vegetales provocan infecciones cuyos síntomas visibles comprenden desde mosaicos (alternancia de zonas normales verdes con zonas amarillentas), moteados…hasta alteraciones en los brotes o en el desarrollo de la planta. Un mismo virus puede producir síntomas distintos en diversas especies. Algunos virus animales tienen la capacidad de transformar las células hospedadoras en células cancerosas (virus oncogénicos). Casi todos los virus animales relacionados con el cáncer son virus ADN, como los herpesvirus o el virus de la hepatitis B, con excepción de los retrovirus, que presentan ARN. 3.2.- CICLOS DE MULTIPLICACIÓN VÍRICA 3.2.1. CICLO DE LOS VIRUS VIRULENTOS O CICLO LÍTICO. + Ciclo de un bacteriofago Los pasos que se siguen en la infección de un bacteriofago son comunes a la mayoría de ellos: - Adsorción del virus e inyección de su ácido nucleico. Inicialmente el bacteriofago fija su cola a receptores específicos de la pared bacteriana, donde una enzima, localizada en la cola del virus, debilita los enlaces de las moléculas de la pared. Posteriormente se contrae la vaina helicoidal, lo que provoca la inyección del contenido de la cabeza a través del eje tubular de la cola del fago: el ácido nucleico penetra en la célula. - Entrada en actividad del ácido nucleico del virus. Una vez dentro, el virus interrumpe el normal funcionamiento de la célula y el DNA bacteriano se degrada. A partir de ese momento será el ácido nucleico del virus el que dicte las órdenes. Los genes víricos poseen la información necesaria para, utilizando los orgánulos y enzimas celulares, autoreplicarse y fabricar todos los componentes del virus. - Ensamblaje de nuevos virus. Todos los ácidos nucleicos replicados como el resto de los componentes víricos que se han sintetizado se ensamblan dando lugar a nuevos virus. - Liberación. En una bacteria pueden formarse unos 100 bacteriofagos, que producen lisozima, enzima que lisa la pared bacteriana. Debido a ello, se produce la ruptura de la pared y la muerte de la célula; los virus quedan libres para infectar nuevas bacterias. + Ciclo de un retrovirus El contacto entre las espículas de su envoltura membranosa y los receptores de la célula hospedadora (linfocitos T auxiliares), que median en la endocitosis, activa la captura endocítica; una vez en el interior del endosoma, la envoltura del virus se funde con la vesícula endocítica y arroja al virus infectivo al citoplasma del linfocito. Superado el mecanismo de penetración, se despoja de su cápsida proteica y queda libre la hebra de RNA y la transcriptasa inversa (retrotranscriptasa) que transporta. Inmediatamente después, se transcribe el RNA, y da lugar a una hebra de DNA, que se replica para formar una doble hélice y se inserta en el cromosoma celular, donde se comporta como un gen mas; se transcribe y se traduce por la maquinaria metabólica celular y origina nuevas copias de RNA vírico, proteínas de la cápsida y de le envoltura y enzimas retrotranscriptasas. Estos componentes se ensamblan, y los virus abandonan la célula mediante un proceso de gemación que les permite adquirir de nuevo su recubrimiento membranoso. 3.2.2. CICLO DE LOS VIRUS ATEMPERADOS O CICLO LISOGÉNICO. La mayoría de los virus bacterianos o bacteriofagos son virulentos, es decir, siguen un ciclo lítico. Sin embargo, los virus denominados atemperados, como el fago que infecta a Escherichia coli, pueden incorporar también su ácido nucleico al genoma del hospedador, replicándose con él (estado de profago), sin que se produzca la síntesis de los componentes virales y la liberación de la progenie viral. Solo ciertos agentes inductores, que por lo general son agentes físicos o químicos que dañan el DNA, provocan la liberación del ácido nucleico del virus, que seguirán entonces un ciclo lítico. 4.- BACTERIAS (FORMAS CELULARES) 4.1. FORMA Y TAMAÑO DE LAS BACTERIAS Son microorganismos unicelulares de dimensiones comprendidas entre 1 y 5 de diámetro. Su morfología externa puede ser variada, distinguiéndose: Cocos: De forma esférica Bacilos: En forma de bastoncillo Bacilococos: Intermedia entre las dos anteriores Vibrios: En forma de coma Espirilos: En forma de espiral o de escalera de caracol 4.2. ENVUELTAS CELULARES 4.2.1. Membrana plasmática. Tiene una estructura similar a la de la célula eucariota, con un espesor de 7,5 a 8 nm. Está formada por una doble capa de fosfolípidos, con proteínas incluidas. Los fosfolípidos presentan las regiones hidrofílicas polares hacia el exterior de la membrana y hacia el citoplasma. Las cadenas hidrofobas apolares de ácidos grasos se dirigen hacia el centro de la bicapa. La membrana contiene múltiples proteínas, entre ellas enzimas como permeasas y, en algunos grupos, pigmentos fotosintéticos, como bacterioclorofilas. Presenta unas invaginaciones a menudo muy largas y enrolladas, que reciben el nombre de mesosomas, su presencia supone para la célula procariota un aumento considerable de la superficie de la membrana; albergan en su seno, y a lo largo de la membrana mesosomica muchas enzimas; entre sus funciones podemos citar: Llevan a cabo la respiración celular Las bacterias fijadoras del nitrógeno, asimilan el nitrogeno en ellos. Las procariotas fotosintetizadoras tienen, también en los mesosomas, los pigmentos, enzimas y transportadores electrónicos encargados de llevar a cabo la fotosíntesis Asimismo, es el lugar donde residen los enzimas que llevan a cabo la duplicación del DNA. Las funciones de la membrana plasmática son: Regula el intercambio de sustancias entre el interior y el exterior celular Controla la síntesis y el crecimiento de la membrana y de la pared celular. Contribuye a la distribución de los componentes celulares hacia las células hijas durante la reproducción. 4.2.2. Pared celular. Es una cubierta rígida que recubre la membrana plasmática. Está constituida principalmente por mureina, un peptidoglicano formado por la unión de monómeros de N-acetilglucosamina, y ácido Nacetilmurámico (aminoazúcares que proceden de la sustitución del grupo –OH de algunas hexosas por un grupo NH2 ), moléculas que se unen por medio de enlaces Oglucosidicos. Existen dos tipos de pared que difieren en su grosor y forma. Fueron descubiertas por Hans Christian Gram (1884), el cual comprobó que podían diferenciarse según un método de tinción, que fue denominado tinción de Gram en su honor. Bacterias Gram-positivas. Se tiñen de color violeta y están constituidas por peptido-glicanos (cadenas de aminoazúcares unidos por péptidos de bajo nº de aa) y ácidos teicoicos (polímeros del glicerol), con un grosor de unos 500 A. Bacterias Gram-negativas. Se tiñen de color rojo. Disponen de una superficie celular más compleja, pues por encima de los peptido-glicanos tienen un mosaico de proteínas, lípidos y lipopolisacáridos. Posee un grosor de unos 100 A. Sus funciones mas importantes son: - Proporciona forma y rigidez a la célula - Protege a la célula de posibles ataques de patógenos o sustancias que podrían destruirla - Permite el paso de sustancias nutritivas y de desecho - Evita el estallido osmótico en medios hipotónicos. 4.2.3. Cápsula. Es una envoltura de grosor variable y de aspecto mucoso que puede formarse en algunas bacterias por la parte externa de la pared celular Está formada por cadenas de polisacáridos. Su función consiste en proporcionar resistencia a las bacterias frente a la sequedad ambiental, protegerlas del ataque de los virus y fijarlas al sustrato. También les confiere resistencia a sustancias tóxicas, como los antibióticos. 4.3. CITOPLASMA Tiene una estructura fibrosa similar al citoesqueleto de las células eucariotas. Está constituido por proteínas que se entrelazan para formar una matriz. El resto está formado por un 80% de agua. Contiene los siguientes elementos: a) Genóforo. Es el único cromosoma que poseen y está constituido por una doble cadena de DNA cerrada sobre si misma y muy replegada. Controla las funciones de la célula y transmite las características hereditarias a las células hijas. b) Ribosomas. Son responsables de la síntesis de proteínas. En las bacterias, la transcripción y la síntesis se llevan a cabo totalmente en el citoplasma. Su tamaño es mas pequeño que el de las células eucariotas. c) Vacuolas de gas. Son vesículas de aspecto cilíndrico. Favorecen la flotación de las bacterias que viven en un medio acuático. d) Carboxisomas. Son pequeñas estructuras membranosas que contienen ribulosa difosfato carboxilasa (rubisco), el principal enzima fijador del dióxido de carbono para el ciclo de Calvin. e) Clorosomas. Vesículas alargadas que tapizan que tapizan el interior de la membrana plasmática y contienen pigmentos verdes fotosintéticos. 4.4. PROCESOS PARASEXUALES Conjunto de mecanismos mediante los cuales las bacterias se intercambian fragmentos de DNA. a) Transducción. Intercambio de material genético entre bacterias que tiene lugar actuando como intermediario un virus bacteriofago. Estos virus se fijan a la membrana de las bacterias e inyectan dentro de ellas su filamento de DNA. Hay algunos bacteriofagos que pese a ello no producen trastornos en la bacteria hospedadora y tras su multiplicación en el interior de ellas, al abandonarlas capturan algo de su material genético que introducen en otra bacteria, con lo cual dicho fragmento cromosómico se puede incorporar al cromosoma de la nueva bacteria invadida que cambia asi su genotipo. b) Transformación. Intercambio de material genético en el cual fragmentos libres de un cromosoma bacteriano penetran por si mismos (sin intermediarios) en otra bacteria y se recombinan con su cromosoma anular; para ello es necesario que las bacterias donadoras lancen fuera de ellas el trozo de cromosoma que buscará a la receptora. c) Conjugación. La conjugación va unida a un fenómeno de sexualidad... en efecto, se pueden distinguir en las bacterias dos tipos sexuales: el dador (masculino) y el receptor (femenino). La diferencia entre ambas está determinada por el factor F, que lo poseen las bacterias dadoras (llamadas por ello F+) y del que carecen las receptoras (F-). Este factor no es otra cosa que un anillo de DNA distinto e independiente del cromosoma anular portador de los genes, pero que se autoduplica en cada división que sufren las bacterias, de forma análoga a como lo hace el cromosoma anular. Se inicia la conjugación con la aproximación de una bacteria dadora a una receptora estableciéndose entre ellas un puente citoplasmático de comunicación; seguidamente se rompe el cromosoma anular de la bacteria dadora transformándose en un filamento abierto que empieza a pasar a la bacteria receptora a través del puente citoplasmático. El cromosoma de la bacteria dadora rara vez se transmite completo. Generalmente no penetra en la receptora mas que una parte del mismo, pues las bacterias conjugadas se separan antes de que termine de pasar dicho cromosoma completamente. El fragmento de cromosoma que ha penetrado en la bacteria receptora se sitúa al lado de que ésta posee incorporándose a él o bien intercambiando entre ambos algún fragmento. En éste último caso la parte no incorporada acaba desapareciendo. En cualquier caso resulta que tras la conjugación, la bacteria receptora presenta un genoma mixto como consecuencia de la incorporación a su cromosoma de genes procedentes de la bacteria dadora. Esto le confiere unas características genéticas diferentes de las que poseía. Durante la conjugación pueden ocurrir dos casos respecto al factor F: * Que permanezca independiente del cromosoma anular, en cuyo caso este último no pasa a través del puente citoplasmático, haciéndolo solo el citado factor F, que de ésta forma convierte a la bacteria receptora de F- en F. * Que se una al cromosoma anular (previa rotura de éste), en cuyo caso es cuando se da la verdadera conjugación, pues entonces hay transferencia de un trozo de dicho cromosoma a la célula receptora, conservando ésta su condición de FLas bacterias Hfr, sin embargo, pueden transferir la totalidad o parte de su DNA cromosómico a través de las fimbrias a una bacteria F-. Para ello, previamente, deben duplicar su DNA cromosómico, junto con el episoma (cadenas de DNA doble y circulares) que lleva integrado, y una de las copias del cromosoma pueden trasladarse a una bacteria F- (generalmente solo pasan fragmentos cromosómicos debido a la fragilidad de las frimbias) 5. PRESENCIA DE LOS MICROORGANISMOS EN LOS PROCESOS INDUSTRIALES 5.1 QUESOS DE EXTREMADURA Las principales levaduras que intervienen en la elaboración del queso son las siguientes: Streptococcus thermophylus ; en cuanto a bacterias : Lactobacillus bulgaricus, Lactobacillus helveticus y Lactobacillus lactis. El queso es un producto obtenido a partir de la cuajada de la leche de vaca generalmente, aunque también se utiliza la de oveja y la de cabra. Las operaciones principales en la fabricación de éste producto son: el cuajado, el prensado, el salado y la maduración. La leche fresca se vierte en grandes recipientes y se pasteuriza, evitando que fermente; posteriormente se añaden una serie de productos tales como el ácido láctico y el cuajado líquido, extraido del cuajar (compartimento digestivo del estómago de los rumiantes) que permiten que la leche coagule; operación llamada cuajado. En éste proceso, la caseína (proteína láctea), precipita y forma flóculos, que constituyen la masa principal del queso. Para mejorar las condiciones de coagulación se adiciona sal y diversos colorantes. Tras ésta operación, queda un líquido residual, el suero, formado en gran parte por agua y un pigmento verdoso, el lactocromo, que mas tarde se separa de la cuajada. Esto se consigue cortando ésta última en diferentes trozos, que se presentan después, hasta que el suero escurre de los mismos (prensado). En la fase siguiente, se sumerge la cuajada en baños de salmuera (salado), para pasar a la última fase llamada de maduración en la que tiene lugar un proceso bioquímico realizado por diferentes bacterias y hongos. Éste se lleva a cabo en cámaras de fermentación especiales, en las que se controla la temperatura, la ventilación y el grado de humedad. 5.1.1. Queso de Los Ibores La zona de producción está localizada en las comarcas cacereñas de Los Ibores, Las Villuercas, La Jara y Trujillo. Se elabora a partir de leche entera y cruda, de cabras de las razas propias de la zona (serrana, verata y retinta); si bien, existen quesos de leche pasteurizada y mezcla. El ganado aprovecha los pastos naturales de la comarca que corresponde a la prolongación extremeña de los Montes de Toledo. Son suelos de escasa profundidad y fertilidad. Actualmente el ordeño está mecanizado (mejora la calidad e higiene del producto), la leche refrigerada, lo que permite elaborar ek queso una vez al día y, las instalaciones queseras modernizadas, por lo que el producto es de mejor calidad, mas uniforme y de mayor garantía sanitaria. Las características del queso so: forma cilíndrica, corteza semidura, pasta de color blanco marfil con ojos y una composición grasa de al menos un 45%. 5.1.2. Queso de La Serena La zona de producción corresponde al sureste de Extremadura en la comarca denominada La Serena donde las ovejas aprovechan los pastizales de secano. El queso se elabora, únicamente, con leche de oveja de raza merina. El colorante empleado en el proceso es un coagulante vegetal que se extrae por maceración en agua de los pistilos de la flor Cynara cardunculus. La leche se calienta ligeramente hasta los 28 o 30º y se mezcla con un extracto del coagulante macerado en agua desde el día anterior. Los microorganismos lipolíticos y proteolíticos dan lugar a la cuajada, que se corta en granos y se vierte en los moldes, donde es sometida a presión para conseguir el desuerado. Se sala con sal marina. Estos quesos presentan forma cilíndrica aplanada, de corteza blanda a semidura. La pasta puede presentar dos texturas: en las tortas, de semilíquida a líquida y, en los quesos, compacata y firme. 5.1.3. Torta del Casar El área donde se produce se encuentra en una planicie semiesteparia en la provincia de Cáceres, cuyo centro lo constituye Casar de Cáceres. Presenta un clima de inviernos cortos y suaves y veranos largos y calurosos, y lluvias irregulares concentradas en primavera y otoño. Son terrenos de baja productividad que ofrecen un buen desarrollo herbáceo. La leche es cruda de oveja, que se ordeña mecanizadamente dos veces al día y se mantiene a 4º hasta llegar a la quesería. El cuajado se obtiene mezclando la leche con el líquido resultante de la maceración en agua de los pistilos de la flor del cardo Cynara cardunculus. Una vez cuajada, se procede al corte en granos y se introduce en los moldes que, una vez llenos, se someten a presión para eliminar todo el suero. Se saca el queso del molde y se realiza el salado, bien directamente o sumergiéndolos en salmuera. La maduración del queso se realiza en cámaras a baja temperatura y alta humedad; al menos, durante dos meses en los que se voltean a diario. La pasta es de color entre blanco y amarillento y, la rextura –cualidad mas característica de éste queso- es altamente cremosa, llegándose a derramar por las grietas de la corteza semidura y el gusto intenso y desarrollado; fundente al paladar, muy poco salado y ligeramente amargo. 5.2 ELEMENTOS BIOLÓGICOS DE LA ELABORACIÓN DEL VINO CON DENOMINACIÓN DE ORIGEN RIBERA DEL GUADIANA. Las levaduras que intervienen en la elaboración del vino son: Saccharomyces cerevisiae, Sacchromyces bayanus y Saccharomyces acidifacienes.Kloeckera apiculata, Torulaspora rosei y Candida stellata La vid (Vitis vinífera) es un arbusto de cuyo tronco parten los brazos o sarmientos, que contienen a las hojas, flores y frutos. Las flores, agrupadas en inflorescencias tipo racimo son blancas y ligeramente perfumadas. Cuando se transforman en frutos dan lugar a los racimos de uvas. Entre las variedades de uvas más usadas con ésta denominación de origen destacan: Variedades blancas: Cayetana blanca, pardina y montúa. Variedades tintas: tempranillo, garnacha tinta y cabernet sauvignon. La materia prima del vino es la uva: Su piel y semilla contienen taninos, que en el caso de los vinos tintos le aportan aroma y color. La pulpa dará lugar al mosto, un líquido de densidad entre 1,06 y 1,15 Kg/l, que contendrá gran cantidad de agua con sustancias en disolución, como ácidos (tartárico, málico, cítrico y ascórbico), sales y en mayor concentración, dos glúcidos: glucosa y fructosa. 5.2.1 Fermentación alcohólca Es necesaria para que el mosto se convierta en vino. Se lleva a cabo por levaduras de las especies: Saccharomyces ellipsoideus y Kloeckera apiculata. Ambas anaerobias facultativas. Estos organismos utilizan los solutos del mosto como fuente de materia y energía que, en un ambiente propicio de temperatura (entre 13 y 35ºC para el vini blanco y 25 y 30 para el vino tinto), junto a una cierta cantidad de oxígeno+ y a un pH neutro o ligeramente ácido, se produce la fermentación entre 18 y 20 días. El componente fundamental del vino es el etanol, seguido de la glicerina; éste último es la responsable de la suavidad y aterciopelado del vino. Por otro lado, el ácido pirúvico no se transforma en acetaldehído, sino que da origen a productos secundarios como: ácido acético, succínico y cítrico. Estos ácidos contribuyen a las características organolépticas finales del vino y a su estabilidad biológica y fisicoquímica posterior. En los vinos tintos se realizan otras transformaciones de carácter secundario entre la que destaca la fermentación maloláctica, llevada a cabo por las bacterias de los géneros Streptococcus, Leuconostoc, Pediococcus y lactobacillus que se encuentran presentes en la uva cuando de recolecta. Esta fermentación es necesaria para la mejora en la calidad y en la estabilidad del vino, pues gracias a ella disminuye y se torna más suave, menos rojizo y de un aroma enriquecido en matices. Los vinos blancos son más agradables a la boca y no realizan ésta fermentación por lo que conservan una ligera acidez, cierto sabor afrutado y la frescura de lo nuevo. 5.2.2. Método para la elaboración del vino * Estrujado: Se aplastan los granos de uva y se libera el jugo, cuidando no romper el hollejo y las pepitas. El caldo obtenido se denomina mosto. * Desmangado: tras el prensado se lleva a cabo la decantación o filtración del mosto para eliminar las partes sólidas. * Encube: consiste en el paso del mosto al recipiente donde se va a producir la fermentación. A ésta fase se le llama también tumultuosa, por que parece que el mosto está en ebullición. * Trasiego o deslío: A continuación se pasa el vino de un recipiente a otro con el fin de separar las partes sólidas depositadas en el fondo del primer recipiente (fangos o lías) y airear el vino si fuera necesario. * Embotellado: se introduce el vino en las botellas definitivas, bien selladas y envejecidas a una temperatura de 13 a 18ºC, por un tiempo de 3 a 20 meses aproximadamente. 5.2.3. Denominación de origen Dentro de la extensa región que comprende la Denominación de origen” Ribera del Guadiana” nos encontramos con una interesante diversidad de microclimas y variedades de viñedos autóctonos que definen, desde el principio, la sorprendente calidad de éstos vinos. La Denominación de Origen fue constituida el 5 de Agosto de 1996 y, aprobado su Reglamento el 17 de Marzo de 1997, por la entonces Consejería de Agricultura, Pesca y Alimentación el 16 de Abril de 1999. Actualmente, la zona de producción abarca 1.666.400 Ha; de las cuales 14.500 Ha están inscritas en la Denominación. Los vinos de la Denominación de Origen “Ribera del Guadiana” cumplen los mínimos siguientes: “Crianza”: Destinadas a aquellos vinos que se someten acrianza y envejecimiento, debiendo prolongarse por plazo no inferior a 2 años naturales, de los cuales al menos 6 meses estuvieron en barrica de roble. “Reserva”: Destinadas a aquellos vinos muy seleccionados: - Tintos: crianza en envase de roble y botella durante un periodo total de 3 años como mínimo, con una duración en envase de roble de al menos 1 año. - Blancos y rosados: crianza en evase de roble y botella durante un periodo total de 2 años como mínimo, con una duración de crianza en envase de roble de al menos 6 meses. * “Gran reserva”: Destinados a vinos de grandes cosechas: - Tintos: crianza de 2 años como mínimo en envase de roble seguida y complementada de un envejecimiento en botella durante 3 años como mínimo. - Blancos y rosados: crianza en envase de roble y botella de 4 años como mínimo, con una duración de crianza en envase de roble de al menos 6 meses. 6.- BIOTECNOLOGÍA Es la disciplina basada en la utilización de los seres vivos o sus componentes, para realizar determinados procesos químicos con finalidad industrial o sanitaria. Los procesos propios de la biotecnología ya eran utilizados en la antigüedad; el uso de levaduras en la fabricación del vino y la cerveza eran conocidos desde hace unos 6000 años a.C En el siglo XX surge el término Biotecnología para definir el conjunto de técnicas que tienen como objetivo la explotación de recursos biológicos que pueden generar las células (animales y vegetales) y los microorganismos. Su nacimiento es consecuencia de los grandes avances logrados en las últimas décadas de el siglo XX por la ingeniería genética y la biología molecular. 6.1. INGENIERÍA GENÉTICA Se conoce como ingeniería genética al conjunto de técnicas basadas en la manipulación del DNA. Esta manipulación consiste básicamente en: Introducir nuevos genes en un genoma; eliminar los genes ya existentes en un genoma y modificar la información contenida en un gen determinado. En ocasiones, también se utiliza la expresión tecnología del DNA recombinante, porque muchas técnicas se basan en la recombinación de fragmentos de DNA. La recombinación génica es la unión de fragmentos de DNA que originalmente se encuentran separados porque corresponden a cromosomas distintos de la misma célula, a células distintas o, incluso, a organismos distintos. La incorporación de la ingeniería genética a la investigación aplicada ha permitido obtener, mediante microorganismos manipulados genéticamente, productos de gran interés para el hombre, como la insulina humana, la hormona del crecimiento humana, interferones, vacunas o enzimas de aplicación industrial. Así mismo, la manipulación del genoma de organismos superiores ha hecho posible la creación de especies de animales y vegetales transgénicos para mejorar la productividad agrícola y ganadera. 6.1.1. Técnicas básicas: enzimas de restricción y clonación Enzimas A partir de diversas especies de bacterias, levaduras, animales y, también, de virus se han aislado y catalogado numerosos enzimas que pueden utilizarse en el laboratorio a la hora de manipular el DNA. Estos enzimas permiten realizar muchas operaciones (cortar, copiar, pegar...) para obtener nuevas combinaciones de genes dentro de un organismo o bien transferir DNA de un organismo a otro. Algunos de estos enzimas son grandes polipéptidos, formados por varias subunidades con funciones distintas. Por ésta razón tienen mas de un tipo de actividad, como es el caso de la DNA pol I de E. Coli. Grupo de enzimas Actividad Ejemplo Añaden nucleótidos a pequeños fragmentos preexistentes o cebadores Exonucleasas Rompen los enlaces Y Endonucleasas fosfodiester entre los nucleótidos. Las exonucleasas rompen por los extremos de las cadenas y las endonucleasas, por el interior Endonucleasas de Reconocen una secuencia restricción de nucleótidos específica O restrictazas y rompen los enlaces entre ellos Hacen cortes en una de las Desoxirribonucleasas dos cadenas de la doble hélice Ligasas Forman enlaces fosfodiester entre un fosfato 5´ y un hidroxilo 3´ Transcriptasas inversas Sintetizan DNA a partir de RNA DNA polimerasas Transferasas terminales Añaden nucleótidos extremo 3´ DNA polimerasa I de E. coli DNA polimerasa I de E. coli Eco R I de E. coli. Desoxirribonucleasa I de páncreas de vaca Ligasa del Fago T4 Transcriptasa inversa del virus de la mieloblastosis aviar al Transferasa terminal del timo de ternera. CLONACIÓN Es un proceso de obtención de copias idénticas que pueden aplicarse en ámbitos muy diversos. Así, se pueden clonar fragmentos de DNA, células o individuos. La técnica de clonación de genes comprende varias etapas: Aislamiento y obtención del gen Selección del vector de clonación Formación de ADN recombinante Inclusión del ADN recombinante en una célula hospedadora Comprobación de la expresión del gen clonado y posterior selección de las células hospedadoras que los llevan. Las células hospedadoras pueden ser: Células bacterianas. El gen deseado, unido al vector se introduce en las bacterias, y al multiplicarse éstas se consigue un número muy elevado de copias de ese gen. Es la vía clásica de la clonación. Células eucariotas. El gen se une al vector de clonación apropiado y se introduce en células vegetales, animales o levaduras. 1. CLONACIÓN UTILIZANDO CÉLULAS BACTERIANAS Comprende las siguientes etapas: A.- Aislamiento y obtención del gen Se emplean varios procedimeintos según el tipo de gen que se quiera clona. Para clonar genes de organismos procariotas el procedimiento habitual es cortar el ADN cromosómico en sitios concretos, para obtener el gen que interesa, mediante las endonuclesas de restricción, enzimas que actúan como tijeras moleculares. B.-Selección del vector de clonación. Los vectores de clonación son pequeñas moléculas de ADN, generalmente circulares, que tiene capacidad de autorreplicarse dentro de las células hospedadoras, independientemente de los cromosomas de éstas. Se utilizan con frecuencia tres tipos de vectores de clonación: plásmidos, genomas de algunos virus y cromosomas artificiales. Plásmidos. Son moléculas de ADN circular, presentes en la mayoría de las bacterias, de menor tamaño que el cromosoma. Los plásmidos se replican con independencia del cromosoma bacteriano, ya que tiene su propio origen de replicación. Genomas de algunos virus. Se trata de pequeñas moléculas de ADN, que contienn su p`ropio origen de replicación. Cromosomas bacterianos artificiales. Son plásmidos que están diseñados para la clonación de fragmentos muy grandes de ADN, formados por centenares de miles de pares de bases. C.- Formación de una molécula de ADN recombinante. La unión covalente del gen a clonar con el vector de clonación mediante la ADN-ligasa produce ADN recombinante.. D.-Introducción EN la célula hospedadora. El ADN recombinante se introduce en una célula hospedadora, la cual proporciona la maquinaria necesaria para la replicación de las moléculas recombinantes. Las células hospedaoras deben cumplir una serie de requisitos para poder ser utilizadas en los procesos de clonación: Experimentar un crecimiento rápido No ser patógenas o peligrosas Er capaces de tomar o captar del medio externo moléculas de ADN e incorporarlas en el conjunto de su genoma Existen varios métodos para introducir DNA en células vivas, que difieren en función de los tipos de célula hospedadora, eucariota o procariota, y del vector de clonación: Transformación. Este procedimiento se utiliza principalmente en células procariotas. La célula capta moléculas de ADN que se encuentran en el medio externo, las introduce en su interior y las incorpora a su genoma mediante recombinación. La transformación es un fenómeno que ocurre espontáneamente en la naturaleza entre ciertos tipos de bacterias como mecanismo de recombinación genética. Sin embargo, se puede provocar también sometiendo a la célula procariota a una serie de tratamientos físicos y químicos, proceso que se denomina transformación artificial. Transducción. Este método consiste en introducir el ADN en la célula hospedadora utilizando como vector de clonación el genoma de un virus. Los virus son capaces de introducir su genoma en células hospedadoras, ya sean procariotas (virus bacterianos o fagos) o eucariotas (virus animales y vegetales). Cuando se emplea como vector de clonación el genoma de un virus lítico, es necesario suprimir antes los genes responsables de la lisis celular, ya que, de lo contrario, la célula hospedadora se lisaría como consecuencia de la infección vírica y no se podría realizar la clonación. Los virus lisogénicos pueden insertar su genoma en el cromosoma de las bacterias; por ésta razón, si se utilizan como vectores de clonación, se deberá modificar también artificialmente el genoma vírico para que se exprese el gen que se desea clonar sin inducir el ciclo lítico. E.- Comprobación de la expresión del gen clonado y selección de células hospedadoras que lo llevan. Cuando se introduce el vector con un gen en cultivo de bacterias, no todas ellas lo incorporan; por eso es necesario seleccionar las que sí lo han hecho y, además, lo expresan. Si los genes clonados son de origen bacteriano pueden expresarse en la bacteria hospedadora, pues la maquinaria de transcripciòn y traducción es semejante. En éste caso, se detecta directamente la proteína del gen de interés. Genoma humano: interés biológico El Proyecto Genoma Humano es, quizá, el programa de investigación biológica más ambicioso realizado hasta la fecha a escala mundial. Con un presupuesto de cerca de 375 000 millones de pesetas y una duración prevista de 15 años, el objetivo del proyecto consiste en conocer la localización precisa de cada gen en los 23 pares de cromosomas que componen el material hereditario humano y estudiar sus mecanismos de expresión. Desde su nacimiento en 1983, colaboran el Proyecto Genoma Humano laboratorios y centros de investigación de todo el mundo. Se calcula que el genoma humano contiene alrededor de 100 000 genes, es decir, unos 3000 millones de nucleótidos. Estos genes representan, aproximadamente, el 5-10% del contenido total de ADN; no se debe olvidar la existencia de muchos fragmentos repetidos y de otros aparentemente sin sentido. Si se pudiera extender y alinear todo el ADN de una persona, se podría rodear varias veces el ecuador terrestre. El Proyecto Genoma comprende siguientes objetivos: Determinar la secuencia completa del ADN humano Identificar los genes que constituyen el genoma humano Almacenar toda la información en base de datos. Crear y apliocar nuevas herramientas informáticas necesarias para analizar los datos obtenidos. Consideraciones éticas, legales y sociales de su conocimiento. Especies transgénicas: concepto Aquellas que son geneticamnte modificadas directa o indirectamente. Además de facilitar la obtención de productos importantes para el ser humano, la ingeniería genética permite crear organismos vivos cuyo genoma ha sido modificado artificialmente mediante la incorporación de DNA clonado en huevos fecundados de animales o en cultivos de células vegetales, si se trata de plantas. Estos organismos reciben el nombre de transgénicos. La utilización de organismos alterados genéticamente posee un interés enorme tanto desde una perspectiva comercial como médica. En el primer caso, con los animales transgénicos se persigue mejorar la producción y la calidad nutricional de las carnes, mientras que con las plantas transgénicas se espera potenciar la producción agrícola, al crear especies resistentes a plagas o con un valor nutritivo mayor. En cuanto a su interés médico, la utilización de animales transgénicos en la investigación biomédica como modelos vivos de enfermedades humanas permite, por ejemplo, comprobar la eficacia de nuevos medicamentos o vacunas. 6.2. REPERCUSIONES BIOTECNOLOGÍA SOCIALES Y VALORACIONES ÉTICAS DE LA Los avances experimentados por la biotecnología plantean graves problemas sociales, éticos y morales, sobre todos los derivados de la manipulación genética, no solo de especies animales y vegetales, sino también en la humana. Es evidente el enorme impacto que han tenido los resultados en el Proyecto Genoma Humano y sus aplicaciones futuras, tales como la disponibilidad de información sobre la composición génica de las poblaciones y la posibilidad de manipular el acervo génico. Esta nueva situación originará, sin duda, conflictos con normas éticas y legales vigentes en nuestra sociedad. Implicaciones sociales Uno de los aspectos sociales que mas preocupan del Proyecto Genoma Humano es la confidencialidad de la información. Cómo afectaría a la vida de una persona y a la de su familia el conocer que es portador de genes responsables de enfermedades de aparición tardía, como la Huntington, o la del Alzheimer, ¿sería éticamente correcto que ésta persona no informara a su pareja sobre la posibilidad de que alguno de sus hijos pudiera desarrollar la enfermedad ¿. Se están empezando a comercializar unos test para realizar, de manera sencilla, diagnósticos de algunas enfermedades, por ejemplo, la predisposición a desarrollar la enfermedad de Alzheimer; si el uso de éstos test se generaliza, podríamos, a corto plazo, encontrarnos con la exigencia de realizar tales pruebas para solicitar un trabajo. Dilemas éticos Alguno de los dilemas éticos que plantean éstas nuevas tecnologías puede resumirse en el planteamiento de algunas cuestiones. ¿ Qué harías si sabes que tu hijo puede tener alguna enfermedad, no solo de nacimiento, sino que la puede padecer a los 40 o 50 años de haber nacido?. ¿ Deben las compañías de seguros conocer la predisposición de una persona a padecer determinadas enfermedades y, por tanto, negarles la póliza ?. ¿Cómo afectaría a una industria el conocimiento de que una parte de los trabajadores están predispuestos a contraer un cáncer inducido por algún producto químico ?. Como punto de partida se acepta que las pruebas genéticas han de ser voluntarias y confidenciales y que sus resultados no deben emplearse con fines discriminatorios. Tras la clonación de la primera oveja clónica saltó la sorpresa y la preocupación en numerosos estamentos de la sociedad. La clonación de un ser humano había dejado de ser un tema de ciencia-ficción para pasar a ser algo científicamente posible, pero, evidentemente, éticamente rechazable. En España la clonación de embriones humanos está prohibida por la Ley de Reproducción Asistida. Nuestro nuevo Código Penal castiga con prisión de uno a cinco años “la creación de seres humanos idénticos por clonación “. BLOQUE V “ LA INMUNOLOGÍA Y SU APLICACIONES” 1.- CONCEPTOS DE INFECCIÓN, PATÓGENOS, VIRULENCIA, TOXINA Y BARRERAS El término infección implica acción competitiva y antagonista entre dos seres vivos, el huésped y el parásito. Si el parásito tiene éxito, aparece la enfermedad; cuando esto ocurre también recibe el nombre de agente patógeno. El término patogenicidad se refiere a la capacidad que tienen los microorganismos para penetrar en el huésped y producirle cambios fisiológicos y anatómicos, es decir, la enfermedad. El grado de patogenicidad de un microorganismo para producir una determinada enfermedad se denomina virulencia. La virulencia de un agente patógeno está influida no sólo por las propiedades inherentes al microorganismo, sino también por la capacidad del huésped para repeler la invasión y prevenir la enfermedad. Entre los factores de virulencia hay que destacar las toxinas, o sustancias venenosas para determinadas células del huésped. La acumulación de toxinas en la sangre de un individuo enfermo se conoce como toxemia. Por otra parte, el término infestación solo indica presencia de parásitos animales sobre o dentro del cuerpo del huésped. Así, un organismo infestante puede transmitir una infección como, por ejemplo, las pulgas que transmiten la peste, o los piojos el tifus. El sistema inmunitario se activa en respuesta a la invasión del organismo por agentes extraños a él. Sin embargo, antes de que se pongan en marcha estos mecanismos defensivos, los seres vivos cuentan con otros sistemas sumamente eficaces para protegerse de forma inmediata: son las denominadas defensas externas o barreras pasivas. 2.- MECANISMOS DE DEFENSA ORGÁNICA INTERNA 2.1. DEFENSAS INESPECÍFICAS Este sistema defensivo actúa con gran rapidez contra cualquier sustancia o agente extraño que invada el organismo. Comprende cuatro tipos de defensas inespecíficas: la inflamación, los fagocitos, el complemento y, si el agente invasor es un virus o células alteradas del propio organismo (por ejemplo, células tumorales, el interferón. 2.1.1. La inflamación Cuando una estructura extraña consigue atravesar las barreras pasivas, el organismo invadido reacciona rápidamente poniendo en marcha la respuesta inflamatoria, que, a su vez, activa y coordina las otras defensas inespecíficas. El proceso inflamatorio es un sistema muy eficaz para preparar el conjunto de mecanismos de respuesta que desencadenarán al penetrar en el organismo un antígeno invasor. La inflamación se manifiesta con unos síntomas característicos: calor, enrojecimiento, dolor y tumefacción de la zona afectada, y se lleva a cabo en cuatro etapas: A) Producción de un estímulo, que habitualmente consiste en la entrada de un microorganismo o de una sustancia tóxica, aunque también puede deberse a un traumatismo. B) Las células lesionadas liberan unas sustancias, que reciben el nombre de mediadores de la inflamación, entre las que destacan las siguientes: Factor de estimulación de la leucocitosis. Provoca un aumento del número de fagocitos circulantes al estimular su liberación a partir de la médula ósea. Leucotrienos. Producen una atracción (quimiotáxis) sobre los fagocitos (leucocitos con capacidad de fagocitar) y aumentan la permeabilidad de los capilares sanguíneos. Histamina y bradiquinina. Estos vasodilatadores aumentan también la permeabilidad capilar. Estimulan, así mismo, las terminaciones nerviosas, provocando la sensación de dolor. Prostaglandinas. Producen una vasodilatación prolongada y atraen y activan a los fagocitos. Por otra parte, aumentan la sensibilidad de los receptores del dolor a la histamina y la bradiquinina. También actúan como mediadores de la inflamación: Los componentes del complemento. Se trata de ciertas proteínas del plasma sanguíneo (concretamente las conocidas como fracciones C3a y C5a), que provocan vasodilatación y atraen y activan a los fagocitos. Productos bacterianos. Al igual que en el caso anterior, las sustancias procedentes del metabolismo de las bacterias y las moléculas liberadas cuando son destruidas atraen y activan a los fagocitos. C) Estos mediadores de la inflamación actúan sobre los capilares de la zona afectada, causando los siguientes efectos: Aumento de la cantidad de leucocitos circulantes en la sangre. Vasodilatación Aumento de la permeabilidad capilar Activación de los fagocitos Quimiotactismo sobre éstos D) Aparición de las consecuencias de los efectos anteriores: La presencia de un número mayor de leucocitos produce un aumento de las acciones defensivas que éstas células llevan a cabo. La vasodilatación incrementa el riego sanguíneo de la zona afectada y la afluencia de elementos defensivos, tanto leucocitos como moléculas de anticuerpos y del complemento. El aumento de la permeabilidad de los capilares favorece la salida de fagocitos, anticuerpos, complemento y fibrinógeno hacia los tejidos infectados. La salida de los fagocitos de los capilares sanguíneos se consigue mediante la separación de las células endoteliales que los forman. El fibrinógeno extravasado coagula y produce un aislamiento de la zona lesionada que dificulta la diseminación del agente invasor. La activación de los fagocitos hace que sean mas eficaces en la eliminación de los microorganismos invasores. El quimiotactismo resulta fundamental para localizar el lugar de la infección y atraer hacia él una gran cantidad de fagocitos que se desplazan hacia concentraciones crecientes de las sustancias quimiotácticas siguiendo su gradiente químico. 2.1.2. Los fagocitos Existen dos grupos de leucocitos: los que componen la serie mieloide y los que constituyen la serie linfloide. Los leucocitos de la serie mieloide se forman en la médula ósea. Pueden desplazarse por movimientos ameboides gracias a la emisión de pseudopodos y tienen capacidad de fagocitosis. Por esta razón también se denominan fagocitos. Se distinguen dos tipos: los granulocitos polimorfonucleares y los monocitos. Granulocitos polimorfonucleares. Poseen abundantes gránulos citoplasmáticos y un núcleo polilobulado. En dichos gránulos almacenan enzimas y compuestos químicos antimicrobianos, y según sus propiedades de tinción se subdividen en neutrófilos, eosinófilos y basófilos, cada uno de los cuales desempeña funciones diferentes. Monocitos. No poseen gránulos citoplasmáticos ni núcleo polilobulado. Cuando salen de los capilares sanguíneos hacia los tejidos circundantes aumentan su tamaño y su capacidad fagocítica y se transforman en macrófagos, que pueden desplazarse libremente o permanecer fijos. En éste último caso se denominan histiocitos y se distribuyen por todos los tejidos, aunque aparecen en mayor número en los alvéolos pulmonares, el bazo, el hígado (células de Kupffer) y la piel (células de Langerhans), constituyendo el sistema reticuloendotelial. Los leucocitos de la serie linfoide son los linfocitos. Aunque no tienen capacidad fagocítica, desempeñan un papel sumamente importante en la lucha contra los antígenos invasores al producir anticuerpos específicos. A) FUNCIONES DE LOS FAGOCITOS Constituyen una línea defensiva inespecífica de gran importancia, ya que se encargan de eliminar los microorganismos y cualquier estructura extraña de los tejidos invadidos en el proceso de la fagocitosis. Para que la fagocitosis sea eficaz es necesaria la activación previa de los fagocitos, lo cual se consigue gracias a los mediadores de la inflamación (prostaglandinas, componentes del complemento y productos bacterianos). También contribuyen a su activación ciertas sustancias producidas por los linfocitos. La activación consiste en la producción de moléculas glucoproteicas en la membrana celular del fagocito que aumentan su capacidad de adhesión a estructuras extrañas. Los primeros fagocitos en actuar son, generalmente, los histiocitos que se encuentran en el propio tejido invadido. Sin embargo, su capacidad fagocítica no es muy grande y enseguida comienza la acción de los neutrófilos sanguíneos, procedentes de los capilares. Estos tienen una vida corta (dos o tres días) y actúan en infecciones agudas. Posteriormente acuden macrófagos libres con mayor actividad fagocítica aún (mientras que un neutrófilo puede fagocitar de cinco a veinticinco microorganismos antes de morir, un macrófago puede eliminar hasta cien de ellos). La fagocitosis se realiza mucho mas eficientemente cuando se lleva a cabo contra una superficie, como, por ejemplo, la pared de un vaso sanguíneo o un coagulo de fibrina. El proceso fagocítico se divide en cuatro etapas: 1. Unión al elemento que va a ser fagocitado mediante receptores glucoproteicos de la membrana. Esta unión se ve favorecida por la presencia de unas moléculas, denominadas opsoninas, que sirven como puentes moleculares entre los receptores del fagocito y la pared bacteriana y facilitan el proceso de fagocitosis. Así, las bacterias que previamente estén recubiertas de opsoninas (bacterias opsonizadas) son fagocitadas con mayor facilidad. Las opsoninas mas importantes son los anticuerpos y algunos componentes del complemento que se han unido a los antígenos superficiales de las bacterias al detectarse la presencia de éstas. 2. Ingestión por medio de la emisión de psudopodos. Cuando dos psudopodos se unen dan lugar a un fagosoma en el que la estructura fagocitada queda encerrada. 3. Muerte y digestión intracelular del microorganismo por acción de las enzimas digestivas y oxidantes de los lisosomas, entre las que se encuentra la lisozima, que se liberan en el fagosoma, el cual pasa entonces a formar el fagolisosoma. 4. Expulsión de los restos no digeridos. El conjunto de restos bacterianos, fagocitos y otras células muertas constituye el pus, que supura en las heridas infectadas. Otra de las funciones de los fagocitos consiste en liberar productos tóxicos para los microorganismos que se encuentran en su cercanía. Los eosinófilos son los encargados de llevar a cabo éste proceso, denominado desgranulación, que resulta fundamental para combatir los agentes patógenos que, debido a su gran tamaño, no pueden ser fagocitados. Algunos leucocitos como los basófilos y mastocitos derivados de éstos, liberan histamina, un mediador de la inflamación. Los macrófagos cooperan con los linfocitos, produciendo sustancias que los activan, y favorecen el reconocimiento e identificación de los antígenos. 2.1.3. El complemento Es un sistema de proteínas (cerca de 30, a las que se llama componentes), presente en el plasma sanguíneo, cuya función defensiva se lleva a cabo con gran rapidez. La denominación de “complemento” deriva de la capacidad para complementar y potenciar la acción de los anticuerpos. Este sistema desempeña tres funciones muy importantes: Actúa como mediador de la inflamación. Esta acción se realiza mediante los componentes C3a y C5a. Interviene en la opsonización de células extrañas a un organismo para facilitar la acción de los fagocitos y de los anticuerpos. El componente C3b es el mas importante en este proceso. Provoca la lisis de las células invasoras por rotura de su membrana plasmática. En éste proceso participan varios componentes que forman un complejo proteico adherido a ella. Para llevar a cabo estas acciones debe producirse antes la activación del complemento, que consiste en la interacción de unos componentes con otros por medio de reacciones en cascada, de manera que el producto final de una de ellas actúa como enzima en la siguiente. Este mecanismo permite amplificar la respuesta considerablemente a partir de unas pocas moléculas iniciales. El proceso de activación del complemento puede iniciarse de dos formas: El proceso de activación del complemento puede iniciarse de dos formas: VÍA CLÁSICA Se produce por la aparición de anticuerpos unidos a los antígenos correspondientes. C1 se activa cuando se une a un complejo antígeno-anticuerpo. C1 activado divide a C2 en C2a y C2b y a C4 en C4a y C4b. C2b y C4b se combinan para formar una proteasa llamada C3 convertasa y divide a C3 en C3a y C3b. Debido a la necesidad de anticuerpos éste proceso está implicado en la inmunidad específica. VÍA ALTERNATIVA No es necesaria la presencia de anticuerpos, por lo que es importante en las primeras etapas de la infección cuando aún no se han producido éstos. Los antígenos, polisacáridos o los componentes de la pared celular reaccionan con C3b. Pequeñas cantidades de C3a y C3b se forman constantemente desde C3, pero sin activación, son pronto destruidas. C3b reacciona con unas proteínas (factor B,D y la properdina) formando C3 convertasa que divide a C3 en C3a y C3b. C3a es un mediador de la inflamación. C3b reacciona con otros componentes del complemento para formar C3 convertasa que forma de nuevo C3a y C3b. C3b se une a las superficies de los microorganismos. Los fagocitos tiene un sitio de unión para el C3b, por lo tanto, los microorganismos con C3b son más susceptibles a la fagocitosis (opsonización). El componente C3b desencadena el proceso denominado vía terminal del complemento (C5) La agregación de properdina a C3 convertasa da origen a C5 convertasa que divide a C5 en C5a y C5b. C5a es un mediador de la inflamación. C5b reacciona con otros componentes del complemento incluyendo C6, C7, C8 y C9 complejo de ataque de membrana formando un hoyo y provocando la lisis . La vía terminal del complemento participa también en la eritrolisis (rotura de la membrana de los glóbulos rojos) producida por incompatibilidad entre diferentes grupos sanguíneos tanto del sistema AB0 como del sistema Rh. 2.1.4. El interferón Las células infectadas por un virus sintetizan y liberan unas proteínas, conocidas con el nombre de interferón, que impiden que la infección se propague. El interferón lleva a cabo dos acciones básicas: Impide la replicación del virus en células infectadas que aún no han sido destruidas por la acción vírica. El interferón liberado se une a receptores de la membrana celular, lo que constituye una señal para la síntesis de unas enzimas llamadas proteínas antivíricas o AVP, que interfieren en la formación de las proteínas del virus. Activa unos linfocitos, denominados células asesinas naturales o células NK, capaces de reconocer células infectadas por virus o células cancerosas y eliminarlas. El interferón actúa, pues, en dos niveles: por un lado, evita la replicación vírica en células aún sanas y, por otro, favorece la destrucción de las células ya infectadas. Las moléculas del interferón realizan, además, otras acciones: activan a los macrófagos y linfocitos B, y modulan la síntesis de anticuerpos y otras sustancias reguladoras (linfoquinas). Es característica del interferón la acción protectora frente a todos los virus que puedan infectar a las células de los organismos de una determinada especie. Sin embargo, no tiene acción protectora en las células de los individuos de otra especie. Su especificidad, por tanto, se manifiesta a nivel de organismo invadido, no de virus. 2.2. DEFENSAS ESPECÍFICAS Se dirigen únicamente a un tipo concreto de antígenos. Para cada antígeno extraño que es detectado en un organismo se produce una respuesta que actúa específicamente contra él. Además de la especificidad, esta respuesta tiene otra característica importante: una vez detectado un antígeno, se crea una memoria inmunológica que permite rechazar al mismo antígeno, incluso con mayor efectividad, cuando vuelve a aparecer. Las células implicadas en esta respuesta específica son los linfocitos: 2.2.1. Linfocitos Son los leucocitos de la serie linfoide. A diferencia de los otros leucocitos no pueden formar pseudopodos y, por tanto, no fagocitan ni son móviles. Por este motivo producen una respuesta específica contra antígenos concretos. Existen tres tipos de linfocitos, cada uno de los cuales desempeña funciones distintas: Linfocitos B. Sintetizan proteínas específicas (anticuerpos) ante la presencia de antígenos. Dado que la mayoría de estos anticuerpos pueden difundirse por los líquidos orgánicos, la inmunidad que producen se denomina humoral. Linfocitos T. Son los responsables de la llamada inmunidad celular, pues no producen anticuerpos, sino que provocan la muerte de ciertas células alteradas y, por tanto, no deseadas (células diana). Así mismo, algunos linfocitos T regulan la actuación del sistema inmunitario. Linfocitos no-B no-T. También destruyen células diana, pero de una forma inespecífica. 2.2.2. Órganos linfoides Las células precursoras de los linfocitos, al igual que las de todas las células sanguíneas, se originan en la médula ósea y se convierten en linfocitos maduros en los órganos linfoides. Estos son los lugares donde se generan, acumulan e interaccionan los linfocitos. Se distinguen dos tipos de órganos linfoides: Órganos linfoides primarios. En ellos se produce la diferenciación de los linfocitos. En el caso de los linfocitos B, el órgano primario es la médula ósea, y en el de los linfocitos T, el timo, glándula situada detrás del esternón que alcanza su máximo desarrollo durante la pubertad para luego ir disminuyendo. Órganos linfoides secundarios. En ellos tiene lugar la acumulación e interacción de los diferentes tipos de linfocitos. Los principales son los ganglios linfáticos y el bazo, aunque también se consideran órganos linfoides secundarios el apéndice, las placas de Peyer intestinales, las amigdalas. 3.- ANTICUERPOS, INMUNOGLOBULINAS O GAMMAGLOBULINAS Son proteínas con una pequeña parte glucídica. Cada molécula de anticuerpo consta de cuatro cadenas polipeptídicas, dos de mayor tamaño (cadenas pesadas o H) y dos mas pequeñas (cadenas ligeras o L). La mayoría de los vertebrados posen sólo dos tipos de cadenas ligeras, denominadas kappa () y lambda (); pero existe un número mayor de cadenas pesadas: gamma ( ), mu (), alfa (), delta () y epsilon (), que definen las cinco clases de inmunoglobulinas presentes en los organismos: IgG, IgM, IgA, IgD e IgE. A su vez, cada una de las cadenas ligeras y pesadas incluye una región variable (VL y VH) cuya secuencia de aminoácidos es peculiar de cada anticuerpo; y una región constante (CL y CH) que mantienen la misma secuencia en todos los anticuerpos pertenecientes a la misma clase de inmunoglobulinas. Para describir cada una de éstas regiones tomamos como modelo la IgG, que tiene forma de “Y”, en cuya estructura se aprecia la doble condición de variación y constancia, responsable de su bifuncionalidad. Las regiones variables (VL, VH) comprenden la mitad de las cadenas ligeras (VL) y la cuarta parte de las cadenas pesadas (VH) y se localizan en la parte superior de la “Y”; pero además, existen ciertas zonas, denominadas hipervariables, cuya capacidad de modificación de las secuencias de aminoácidos determina la enorme diversidad de anticuerpos. Estos extremos hipervariables constituyen los sitios activos del anticuerpo, llamados paratopos, responsables de la combinación con el antígeno específico. Cada molécula de IgG posee dos paratopos y, por tanto, puede unirse con dos moléculas del mismo determinante antigénico o epitopo. La región constante (CL, CH) comprende la mitad de la cadena ligera (CL) y las tres cuartas partes de las cadenas pesadas (CH) y forma el pie de la “Y”; es la zona de la molécula que dirige su actividad biológica, es decir, regula las funciones que desempeñan las inmunoglobulinas dentro del sistema inmunitario, entre las que destacan las siguientes: Función metabólica, determina la vida media de la molécula. Función transplacentaria, participa en el transporte de determinados anticuerpos desde la madre al feto a través de la placenta. Función receptora, actúa como sistema de anclaje de los receptores de membrana de los linfocitos B. Función de anticuerpo circulante, es quizá la mas característica de todas, y representa la respuesta humoral frente al antígeno. Los anticuerpos neutralizan los efectos nocivos de las toxinas segregadas por algunas bacterias, en cuyo caso se comportan como antitoxinas que provocan la precipitación de las toxinas, y el precipitado se elimina por filtración renal o bien es fagocitado por un macrófago. En otros casos, los anticuerpos establecen uniones con los antígenos de superficie de diferentes células y produce aglutinación, como es el caso de las hemoglutininas liberadas contra los eritrocitos de otro grupo sanguíneo. 3.1. TIPOS DE ANTICUERPOS Según la composición de sus cadenas peptídicas se distinguen cinco clases de anticuerpos: 3.1.1. Inmunoglobulina G (IgG) Son monoméricas y se localizan en la sangre y en los líquidos extracelulares. Favorecen la fagocitosis, ya que actúan como opsoninas al unirse a los antígenos situados en la superficie de las estructuras microbianas. Posibilitan, además, la vía clásica de activación del complemento. Cuando los antígenos son exotoxinas, al unirse a ellas las neutraliza, por lo que también actúan como antitoxinas. Constituyen las inmunoglobulinas mas abundantes en la respuesta inmunitaria secundaria, que se produce al detectarse un antígeno con el cual ya ha existido un contacto previo. Son capaces de atravesar la placenta, por lo que inmunizan al feto contra los antígenos para los que la madre gestante es inmune. 3.1.2. Inmunoglobulias A (IgA) Pueden ser monoméricas o diméricas. Las primeras se localizan en la sangre y en otros líquidos extracelulares, mientras que las segundas se encuentran en secreciones como la saliva, las lágrimas, la leche y el mucus que recubre el interior del aparato respiratorio y el intestino. En éste caso, la IgA, denominada IgA secretora, se une a un componente segregado por las células epiteliales. Su función básica consiste en proteger las mucosas de la invasión microbiana. Esta acción es fundamental para evitar la fijación de los virus, pues las IgA se unen a los lugares de la cápsida del virus por lo que éstos se adhieren a las células del hospedador. 3.1.3. Inmunoglobulinas M (IgM) Son pentaméricas y se encuentran en la sangre y en los líquidos extracelulares así como sobre la superficie de los linfocitos B, donde reconocen a los antígenos. Son muy eficaces en la activación del complemento y provocan, además, la precipitación de antígenos solubles. Las IgM son los primeros anticuerpos que se producen tras la primera exposición a un antígeno nuevo (respuesta inmunitaria primaria). 3.1.4. Inmunoglobulinas D (IgD) Son monoméricas. Se localizan en la membrana de los linfocitos B, por lo que participan en el reconocimiento de los antígenos y en la estimulación de la producción de otros anticuerpos. 3.1.5. Inmunoglobulinas E (IgE) También aparecen como monómeros. Pueden encontrarse en la sangre y en los líquidos extracelulares. Al unirse a la membrana de los leucocitos eosinófilos provocan la desgranulación de éstos, es decir, la secreción de enzimas digestivas y de otras moléculas que actúan sobre estructuras que no pueden ser fagocitadas debido a su gran tamaño (parásitos, por ejemplo). Se unen, así mismo, a los basófilos y a los mastocitos, lo que causa también su desgranulación. Como consecuencia se producen alteraciones no deseadas (procesos alérgicos). 4.- RESPUESTA HUMORAL Y CELULAR 4.1. INMUNIDAD HUMORAL. Es el conjunto de mecanismos inmunitarios en los que intervienen proteínas específicas fabricadas contra los antígenos extraños que se difunden por los fluidos del organismo. Estas proteínas son los anticuerpos, y su difusión se realiza por la sangre, la linfa, los líquidos intersticiales y las secreciones donde llevan a cabo su acción. La síntesis de anticuerpos se realiza por los linfocitos B. 4.1.1. Linfocitos B Se forman y diferencian en la médula ósea. Allí adquieren capacidad para producir anticuerpos, moléculas específicas contra los antígenos invasores. Por ésta razón se dice que son inmunocompetentes. En la médula ósea se generan millones de linfocitos B, genéticamente diferentes, cada uno de los cuales fabricará distintos anticuerpos, capaces de unirse a la enorme cantidad de antígenos diferentes que entran en un organismo. Esto se consigue mediante la fragmentación de parte del ADN que lleva información para la síntesis de anticuerpos y el posterior reagrupamiento de los segmentos cortos formados (reordenamiento génico). Los genes implicados se localizan, en el ser humano, en los cromosomas 2, 14 y 22. De este modo, a partir de unos pocos genes iniciales se obtiene una gran cantidad de combinaciones génicas distintas, lo cual explica la enorme variedad de anticuerpos diferentes, aunque cada linfocito (y sus descendientes) produce un único tipo de anticuerpo específico. Algunos anticuerpos permanecen fijos en la membrana de los linfocitos B y otros se liberan. Por lo general, en el sistema inmunitario sólo hay unos pocos linfocitos B diferenciados que, además, se encuentran inactivos. Cuando aparece un antígeno, tiene lugar su unión a un anticuerpo de la membrana de un determinado linfocito B. Esta unión únicamente es posible si existe un acoplamiento espacial entre ambas moléculas; es decir, es necesaria una conformación complementaria entre la estructura del antígeno y la del anticuerpo.. El reconocimiento antigénico se basa, por tanto, en la especificidad de la unión con el anticuerpo correspondiente. Dada la enorme variedad de anticuerpos, siempre existe alguno capaz de reconocer a un antígeno concreto. La unión puede ser fuerte o débil. En el primer caso, la respuesta inmunitaria generada es mas intensa. Tras el reconocimiento del antígeno, los linfocitos B se activan. Su activación consiste en la rápida división de estas células para originar una serie o clon de células iguales productoras del mismo tipo de anticuerpo. Así, además de aumentar considerablemente el número de linfocitos después de la exposición al antígeno, todos ellos generan los anticuerpos específicos correspondientes. De esta forma, la propia aparición del antígeno sería la causa de una selección clonal por la que se obtiene un número elevado de linfocitos B productores de los anticuerpos necesarios contra ese antígeno. Esta teoría de la selección clonal, permite explicar por qué se producen grandes cantidades de anticuerpos específicos tras la introducción de un determinado antígeno. Es el mismo antígeno el que, por su unión específica a un linfocito concreto, de entre los millones existentes, induce a éste a originar, a partir de él, un clon encargado de rechazarlo. Los linfocitos que no contactan con el antígeno específico no se activan, pero siguen disponibles por si éste aparece en el futuro. La mayor parte de los linfocitos B activados se convierten en las llamadas células plasmáticas, de gran tamaño y con una enorme producción de anticuerpos (unas 10000 moléculas/célula/minuto). Sin embargo, algunos de ellos quedan como linfocitos B de memoria, que tienen una vida ilimitada u constituyen una reserva inmunitaria para nuevas exposiciones futuras al mismo antígeno. Tanto las células plasmáticas como las de memoria se acumulan en la zona cortical de los ganglios linfáticos, desde donde pasan a la linfa y de ésta a la sangre. La activación de los linfocitos B se intensifica mucho por la acción de las interleuquinas, unas sustancias segregadas por ciertos linfocitos T y por los macrófagos. 4.2. INMUNIDAD CELULAR Además de la acción de los anticuerpos sobre los antígenos, existe otro proceso defensivo, conocido como inmunidad celular o inmunidad mediada por células, sin producción de anticuerpos. Este proceso resulta muy eficaz en la destrucción de: Células extrañas a un organismo procedentes de otro individuo distinto, aunque sea de la misma especie. (Órganos trasplantados). Células propias tumorales Células infectadas por virus Células que contienen un microorganismo de crecimiento intracelular, como la bacteria Mycobacterium. Existen dos tipos de linfocitos implicados en ésta respuesta: los linfocitos T y los linfocitos no-B no-T. 4.2.1. Linfocitos T Los linfocitos T se diferencian en el timo y se dividen en dos grupos principales, T4 y T8. Linfocitos T4. Contienen en la membrana celular unas proteínas receptoras denominadas CD4. Se distinguen dos tipos de linfocitos T4: Linfocitos TH (cooperadores): su función consiste en estimular a otros linfocitos T y a los linfocitos B. Linfocitos TD : provocan un aumento del número y de la actividad de los macrófagos. Linfocitos T8. Poseen en su membrana unas proteínas conocidas como CD8. También se diferencian dos subgrupos: Linfocitos TC (citotóxicos): provocan la destrucción de las células diana. Linfocitos TS (supresores): evitan una respuesta inmunitaria excesiva o desproporcionada. Los linfocitos T actúan de forma específica contra células diana que portan antígenos extraños en su superficie. Poseen receptores de antígenos, moléculas que tienen una región constante unida a su membrana celular y una región variable encargada de reconocer y unirse al antígeno. A semejanza de los anticuerpos receptores de los linfocitos B, en las distintas estirpes celulares existen diferentes receptores antígenos. Sin embargo, a diferencia de aquellos, los receptores de los linfocitos T no pueden identificar antígenos libres y, además, deben reconocer simultáneamente el antígeno extraño y una molécula propia (autoantígeno) de la célula diana en la que se encuentran. El proceso se realiza de la siguiente forma: Una célula del hospedador, habitualmente un fagocito, captura un elemento extraño y tras su digestión intracelular sitúa alguno de sus antígenos en su propia membrana. Este determinante antigénico suele ser un pequeño péptido derivado de la molécula inmunógena inicial, procesada mediante una degradación parcial. En la membrana se combina con un autoantígeno, ya presente en ella, y da lugar a un complejo antigénico. Esto la convierte en una célula presentadora de antígenos. El complejo antígenico formado se une, entonces, al receptor antigénico del linfocito T produciendo la activación de éste. Los autoantígenos son glucoproteínas específicas de cada individuo, codificadas por una región del genoma que se denomina complejo principal (o mayor) de histocompatibilidad (MHC). En el ser humano esta región se encuentra en el cromosoma 6. Existen dos clases de autoantígenos codificados por el MHC: los de la clase I, que se encuentran en todas las células nucleadas del organismo; y los de la clase II, que sólo se localizan en las células presentadoras de antígenos y en los linfocitos B. Los receptores antigénicos de los linfocitos T8 (tanto los TC como los TS) únicamente son capaces de reconocer los complejos antigénicos formados por antígenos extraños y autoantígenos del MHC de la clase I. Por el contrario, los receptores antigénicos de los linfocitos T4 (TH y TD) sólo reconocen los de clase II. Una vez producido el reconocimiento por parte de los linfocitos T, éstos se activan. En el proceso interviene un complejo proteico de la membrana, llamado CD3, que transmite, de forma aún desconocida, una señal al interior del linfocito. En primer lugar se activan los linfocitos TH. Su activación se potencia por la presencia de la interleuquina 1, una sustancia que producen y liberan los macrófagos. Cuando los linfocitos TH se activan, se liberan a su vez otra sustancia (interleuquina 2), que incrementa considerablemente su estimulación. Es decir, se produce una autoactivación de los linfocitos TH por medio de la interlequina 2. La acción estimuladora de ésta sustancia es mas potente que la de la interleuquina 1. Además, durante la activación, se van formando en la membrana nuevos receptores para la interleuquina 2 (en estado inactivo existen 100 receptores/linfocito y al final de la activación puede haber hasta 12000 receptores/linfocito), que potencian aún mas su acción. Por otra parte, la producción de la interleuquina 2 activa también a los linfocitos TC y TS. 4.2.1. Funciones de los linfocitos T Tras su activación, los linfocitos T provocan una acción citotóxica y regulan la respuesta inmunitaria. Citotoxicidad. Consiste en la lisis de las células diana por los linfocitos TC. Las células diana pueden ser cualquier célula del propio individuo que tenga en su superficie complejos antigénicos formados por autoantígenos del MHC de la clase I y antígenos extraños (como ocurre en las células infectadas por virus), o bien antígenos anormales (caso de las células cancerosas). Tras la unión entre la célula diana y el linfocito TC, éste segrega unas proteínas, denominadas perforinas que producen unos orificios en la membrana celular, semejantes a los provocados por la acción del complemento. Como consecuencia, la célula diana se lisa y muere. También se puede producir la activación de las endonucleasas de la célula diana, lo que provoca igualmente su muerte. Regulación de la respuesta inmunitaria. Los linfocitos TH y TS desempeñan una función reguladora del sistema inmunitario para que éste actúe con la máxima eficacia impidiendo acciones inadecuadas (por falta o exceso de respuesta). Los linfocitos TH (cooperadores) intensifican la respuesta inmunitaria de tres formas distintas: linfocitos TC. Producen interleuquina 2, que estimula la activación de los Activan los linfocitos B mediante la secreción de interleuquinas 2, 4, 5 y 6, o uniéndose directamente a ellos. Aumentan la estimulación de los fagocitos gracias a la secreción de interleuquina 2. Por el contrario, los linfocitos TS (supresores) disminuyen la respuesta del sistema inmunitario de dos formas: Desactivan los linfocitos activados por medio de la formación de unos factores supresores. Esto sucede cuando se ha conseguido controlar a los agentes antigénicos o cuando la respuesta inmunitaria es excesiva y podría resultar peligrosa para el propio organismo. Producen una tolerancia inmunológica a los autoantígenos para evitar el ataque a las propias células del organismo. 4.2.2. Linfocitos no-B no-T. Las células conocidas con este nombre son minoritarias, pero desempeñan un papel importante en ciertos casos. A diferencia de los linfocitos B y T, éstas células, que poseen un mayor tamaño y gránulos citoplasmáticos, no reconocen al antígeno y tienen, por tanto, una actuación inespecífica. Por otra parte, tras la estimulación, su número no aumenta ni originan memoria inmunológica: Se diferencian dos tipos de éstas células: Células K o células asesinas. Atacan a las células recubiertas por anticuerpos y segregan perforinas, como los linfocitos TC, para destruirlas. A pesar de producir una muerte celular dependiente de la existencia de anticuerpos sobre la célula diana, las células K tienen una actuación inespecífica, ya que no diferencian los distintos tipos de anticuerpos. Celulas NK o células asesinas naturales. Actúan de manera semejante a las anteriores, destruyendo las células afectadas por virus y las cancerosas, así como las pertenecientes a los órganos transplantados. Como se ha dicho, el interferón producido por las células atacadas por virus favorece la actuación de las células NK. El estímulo para su actuación es la ausencia de las proteínas correctas del MHC de la clase I. Actualmente se cree que las células NK siempre están activas, pero se desactivan al unirse a las células normales del individuo (es decir, a las que tienen las proteínas normales del MHC de la clase I). 5.- LA MEMORIA INMUNE La inmunidad que un individuo desarrolla frente a determinados antígenos puede deberse a procesos naturales o bien a la aplicación de ciertas técnicas médicas. En el primer caso se habla de inmunidad natural, y en el segundo, de inmunidad artificial En cada uno de éstos tipos, la inmunidad puede ser pasiva, si se adquiere al recibir anticuerpos específicos fabricados por otro organismo, y activa, si el propio organismo sintetiza los anticuerpos tras la introducción del antígeno, gracias a sus defensas inmunitarias. Se distinguen, por tanto, los siguientes tipos de inmunidad: Inmunidad natural pasiva. Se adquiere por la transferencia de anticuerpos de la madre al feto y al niño lactante a través de la placenta y de la leche materna, respectivamente. De esta forma, el niño recibe anticuerpos de manera constante en un periodo durante el cual su sistema imnunológico aún no está desarrollado por completo. Inmunidad natural activa. Se produce después de superar una enfermedad infecciosa, ya que el organismo queda cargado con los anticuerpos sintetizados y, sobre todo, con linfocitos de memoria, por lo que durante un tiempo (en ocasiones toda la vida) se evita la reinfección. Inmunidad artificial pasiva y activa. También denominado proceso de inmunización, se obtiene estimulando el sistema inmunirario del individuo por medio de técnicas médicas. 5.1. INMUNIZACIÓN PASIVA. Consiste en la introducción en el organismo de anticuerpos sintetizados previamente por otra persona o por un animal, por lo que el sistema inmunitario del individuo receptor no necesita activarse. Este procedimiento tiene unas ventajas evidentes: Proporciona una protección inmediata, lo que puede ser importante en los casos en que se necesiten anticuerpos rápidamente (como en infecciones graves de desarrollo muy rápido). Al no ser necesaria la síntesis propia de anticuerpos, esta inmunidad resulta útil en individuos con deficiencias en su sistema inmunitario. Sin embargo, también presenta una serie de inconvenientes: Se obtiene una inmunidad de duración limitada hasta que los anticuerpos administrados desaparezcan del cuerpo del receptor. Existe el riesgo de transmitir las enfermedades padecidas por el individuo del que se han obtenido los anticuerpos. Si los anticuerpos proceden de la sangre de un animal pueden producirse reacciones de rechazo contra alguna proteína de éste. Para llevar a cabo la inmunización pasiva es preciso obtener previamente los anticuerpos. Para ello se inocula el antígeno correspondiente a un animal (frecuentemente se ha utilizado el caballo), que sintetiza los anticuerpos contra una enfermedad determinada. También se pueden obtener de la sangre de individuos hiperinmunes que posean gran cantidad de los anticuerpos que se precisan. En la inmunización pasiva se emplean dos tipos de sustancias: Sueros. También llamados antisueros, contienen solamente los anticuerpos específicos. Gammaglobulinas. Consisten en mezclas de anticuerpos obtenidos de la sangre de un organismo inmune, entre los que se encuentran los que interesan. 5.2. INMUNIZACIÓN ACTIVA. VACUNACIÓN Se basa en la estimulación artificial del propio sistema inmunológico de un individuo mediante el suministro de los antígenos necesarios para que el organismo sintetice por sí mismo los anticuerpos contra ellos. La vacunación es una forma de inmunización activa, ya que estimula las defensas inmunitarias frente a determinados agentes patógenos o sustancias dañinas, antes de desarrollarse la enfermedad, de manera que si el individuo queda expuesto de nuevo a éstos agentes patógenos, sus células de memoria sintetizarán rápidamente anticuerpos para combatirlos. Se trata , por tanto, de un método preventivo, que prepara el organismo ante un posible contacto con el agente patógeno correspondiente. La persona vacunada fabrica linfocitos de memoria que permiten producir una respuesta secundaria mas rápida y mucho mas intensa que la respuesta primaria cuando se expone a un microorganismo patógeno. Gracias a la vacunación, se han obtenido resultados espectaculares contra las enfermedades infecciosas, algunas de las cuales, como la viruela, han sido erradicadas. Sin embargo, no siempre es posible obtener las vacunas adecuadas; en la actualidad se están realizando numerosas investigaciones para desarrollar vacunas efectivas contra el Sida o la hepatitis C. 6.- CONCEPTOS DE HIPERSENSIBILIDAD, CÁNCER, SIDA Y TRANSPLANTES 6.1. HIPERSENSIBILIDAD Se produce cuando el sistema inmunológico desencadena una respuesta inmunitaria inadecuada que provoca lesiones en los tejidos del propio organismo. En la hipersensibilidad, los procesos inmunitarios no ocasionan un beneficio al organismo vivo, sino un efecto perjudicial. La acción lesiva puede ser causada tanto por anticuerpos como por linfocitos T. Según el efecto producido, el proceso inmunitario causante y el tiempo que tarda en aparecer tras el contacto con el antígeno, se distinguen cuatro tipos de hipersensibilidad. 6.1.1. Hipersensibilidad de tipo I (inmediata o anafiláctica) Conocida también como reacción alérgica o alergia, el desarrollo de esta hipersensibilidad es muy rápido, pues se produce entre los 10 y 20 minutos tras la exposición al antígeno, que en ese caso se denomina alérgeno. Entre los alergenos mas comunes se encuentran los siguientes: Veneno de abejas (concretamente, la fosfolipasa A). Proteínas de las células del polen. Esporas de mohos Heces de ácaros presentes en el polvo Pelo de animales Algunos medicamentos (como la penicilina) Ciertos alimentos (mariscos, cacahuetes) La reacción alérgica se desarrolla de este modo: La primera exposición al alérgeno provoca un estímulo en los linfocitos TH que, a su vez, inducen la activación de los linfocitos B. Estos fabrican inmunoglobulinas IgE, que se unen por su zona constante a leucocitos basófilos y a los mastocitos (células fijas localizadas cerca de los capilares sanguíneos), fijándose a la superficie externa de su membrana. Se dice entonces que se ha producido la sensibilización, la cual depende de la producción de IgE. La causa última de la sensibilización a alérgeno no se conoce. Parece ser, aunque aún no se ha demostrado, que la formación de las IgE depende, al menos en parte, de la información genética del individuo. También se ha atribuido a una deficiencia de los linfocitos TS encargados de evitar una actuación excesiva del sistema inmunitario. Una vez producida la sensibilización y tras una nueva exposición al alérgeno, éste se une a las IgE fijadas previamente a los mastocitos y basófilos, lo cual provoca su desgranulación, que consiste en la secreción de sustancias que se encuentran en el citoplasma celular. Para que se produzca la desgranulación es preciso que el alérgeno se una, al menos, a dos moléculas de IgE. Las sustancias expulsadas son mediadores de la inflamación (histamina, prostaglandinas y leucotrienos). Los síntomas y manifestaciones del proceso alérgico se derivan de la acción de éstas sustancias, que pueden desencadenar reacciones locales (inflamación cutánea con enrojecimiento, hinchazón y picor y aumento de la secreción nasal y lacrimal) o reacciones generalizadas (contracción de los músculos de bronquios o bronquíolos, que genera dificultades respiratorias, asma o bien una importante vasodilatación general). En éste último caso se puede producir la muerte por asfixia o por un descenso acusado de la presión sanguínea (shok anafiláctico). El tratamiento de los procesos alérgicos se basa en la administración de antihistamínicos, para neutralizar la acción de la histamina, y de otros medicamentos, para aumentar la presión sanguínea y dilatar los bronquios. La solución ideal, sin embargo, consiste en desensibilizar al individuo mediante técnicas de inmunoterapia. Para ello se realizan pruebas cutáneas con el fin de identificar al alérgeno responsable. Una vez detectado, se administra en dosis crecientes hasta conseguir la desensibilización. Al parecer, con éste método se forman anticuerpos bloqueantes IgG que al unirse al alérgeno impiden que éste se una a las IgE. También puede estar implicada la producción de linfocitos TS que provocan una disminución de la síntesis de IgE. 6.1.2. Hipersensibilidad de tipo II (Citotóxica). En éstas reacciones los anticuerpos IgM o IgG se unen a antígenos pertenecientes a células humanas, lo cual activa el complemento por la vía clásica y la lisis por la vía terminal. La activación también favorece la fagocitosis. Se encuandran en este tipo de hipersensibilidad algunas enfermedades autoinmunitarias en las que produce un fallo de la tolerancia inmunitaria, creándose anticuerpos contra antígenos propios. Suelen incluirse también en este grupo el rechazo a los órganos transplantados y la incompatibilidad entre diferentes grupos sanguíneos. 6.1.3. Hipersensibilidad de tipo III. (Mediada por complejos inmunitarios) Aunque presenta muchas semejanzas con la de tipo II, los antígenos a los que se unen los anticuerpos no forman parte de células, sino que circulan libres en la sangre. La reacción de hipersensibilidad se desencadena porque los complejos antígeno-anticuerpo creados no son eliminado por los macrófagos, como sucede normalmente. Se produce entonces una activación excesiva del complemento y una respuesta inflamatoria en la que se liberan enzimas que ejercen una acción destructiva sobre los tejidos. Los órganos afectados pueden sufrir daños importantes. Algunas enfermedades autoinmunitarias y ciertas infecciones crónicas están causadas también por ésta hipersensibilidad. Así mismo, se producen reacciones de éste tipo en la inmunización pasiva realizada con un suero procedente de un animal, debido a las proteínas que contiene (enfermedad del suero) 6.1.4. Hipersensibilidad de tipo IV (Retardada). Se denomina así porque su desarrollo se produce mas tarde que en los otros tipos (incluso varias semanas después de la exposición al antígeno). A diferencia del resto de las hipersensibilidades, en las de tipo IV no intervienen anticuerpos, sino un clon de células iguales. Tras una segunda exposición al mismo antígeno, los linfocitos TD de este clon celular liberan ciertas linfoquinas (regulan a los linfocitos T y B) que provocan la estimulación de los macrófagos y desencadenan un proceso inflamatorio. Este tipo de hipersensibilidad es el causante de la dermatitis de contacto originadas por algunos cosméticos, prendas de vestir, bisutería o plantas en contacto con la piel, cuyas proteínas se unen con haptenos presentes en dichos productos y originan los antígenos desencadenantes de la reacción. Dado que los síntomas (picor, erupción) son similares a los de la hipersensibilidad de tipo I, en ocasiones estas reacciones se denominan alergias de contacto, aunque el término alergia se debería aplicar a la primera. La reacción granulomatosa es un proceso grave de hipersensibilidad retardada, que produce daños importantes en los tejidos internos como consecuencia de la formación de nódulos en los que existe una inflamación continua con presencia de macrófagos y monocitos. Las lesiones tisulares que caracterizan a la tuberculosis y a la lepra se deben a reacciones granulomatosas. Alguno de los procesos de rechazo de órganos transplantados se consideran también reacciones de hipersensibilidad de tipo IV. 6.2. CÁNCER Un cáncer se inicia cuando ciertas células, por causas aún no suficientemente aclaradas, sufren una transformación que las induce a dividirse de manera rápida y descontrolada. Como consecuencia, se origina una masa de células sin las características propias (y, por tanto, las funciones) de las que proceden, con un metabolismo muy intenso y alterado. Esta masa de células anormales, que recibe el nombre de tumor, invade los tejidos circundantes causando su destrucción. Si su crecimiento es limitado se habla de tumor benigno. Por el contrario, cuando las células se dividen sin límite y se extienden a otros órganos, se denomina tumor maligno o cáncer. Se ha demostrado que las células transformadas, origen de los tumores, poseen en su superficie hay antígenos distintos a los de las células normales. Por ésta razón, el organismo no las considera como células propias y pone en marcha mecanismos inmunitarios, tanto inespecíficos como específicos, para destruirlas. Las defensas inespecíficas están constituidas por los macrófagos y por las células NK, que eliminan a las células tumorales mas eficazmente cuando tienen anticuerpos unidos a su membrana. También participan en la acción antitumoral sustancias citotóxicas, como el factor de necrosis tumoral (TNF) producido por varias células del sistema inmunitario. Las defensas específicas celulares consisten en la acción citotóxica causada por los linfocitos TC, que se unen a las células con antígenos alterados y provocan su destrucción. Además, los linfocitos TH liberan linfoquinas que favorecen la función de los linfocitos TC y la activación de los macrófagos y de los linfocitos B. Las defensas específicas humorales son los anticuerpos que, tras su unión a las células cancerígenas, favorecen la acción de los macrófagos y de las células NK, así como la activación del complemento por la vía clásica. De esta forma, según la teoría de la vigilancia inmunitaria, el sistema inmunitario se encargaría de eliminar estas peligrosas células antes de que se multiplicaran masivamente y pudieran desarrollar un cáncer. Este eficaz sistema, sin embargo, no siempre cumple su finalidad, pues en ocasiones se consigue eludir la acción inmunitaria y se produce un cáncer. Pese a que se ha especulado mucho en torno a la causa de éste fenómeno, aún no se ha podido elaborar una explicación satisfactoria. Las hipótesis mas aceptadas son las siguientes: Las células cancerosas producen y segregan moléculas que interfieren en las defensas inmunitarias, desactivando, por ejemplo, los linfocitos TC o provocando la estimulación de los linfocitos TS, lo cual disminuiría la eficacia de la respuesta. Los antígenos de las células alteradas no son detectados por el sistema inmunitario debido a la falta de algunas moléculas necesarias para la “presentación antigénica” o bien porque están ocultos por otras moléculas que dificultan su identificación. La existencia de una gran cantidad de antígenos en las células cancerosas provocaría el bloqueo, quizá irreversible, de todos los receptores de los linfocitos impidiendo de esta forma el reconocimiento de nuevas células alteradas. La respuesta inmunitaria se desencadena con lentitud y cuando puede ser efectiva, el tumor ya se ha desarrollado. Esto resulta evidente en casos de inmunodeficiencias debidas a la edad avanzada, a la infección por VIH o incluso a algunos tratamientos antitumorales que debilitan las defensas inmunitarias. Debido al desconocimiento de las causas exactas que determinan la aparición de células alteradas que escapan a la acción destructiva del sistema inmunitario, el tratamiento contra el cáncer se basa fundamentalmente en las siguientes técnicas: Extirpación quirúrgica Radioterapia Quimioterapia Administración de interferón Etc. 6.3. TRANSPLANTES Los transplantes de órganos constituyen un procedimiento quirúrgico cada vez mas frecuente. La mayor dificultad del transplante de órganos no reside en el proceso quirúrgico propiamente dicho, sino en un posible rechazo a los tejidos del donante por parte del organismo receptor. Este rechazo se produce como consecuencia de la respuesta inmunitaria contra los antígenos presentes en las células del órgano transplantado, que se detectan como extraños. Los antígenos responsables del rechazo son los autoantígenos del MHC que se encuentran en la superficie celular. Entre los individuos de una misma especie hay una gran variedad de autoantígenos del MHC. Los genes del MHC que codifican estos autoantígenos son numerosos. En el ser humano existen tres loci implicados en el MHC de clase I y, dado que un individuo concreto tiene dos alelos para cada uno de ellos, la determinación genética se lleva a cabo por seis alelos. También hay otros tres loci y, por tanto, seis alelos que codifican los autoantígenos del MHC de clase II. Como existen muchos alelos distintos posibles para cada uno de los loci , los genotipos del MHC, tanto de clase I como de clase II, son muy numerosos (se calcula que hay unas 10 000 combinaciones posibles). Si los autoantígenos del donante y del receptor no coinciden, se produce el rechazo, que comienza con el ataque de los linfocitos TC, los cuales causan la lisis de las células de los tejidos transplantados. Únicamente cuando los autoantígenos del MHC son idénticos (cosa que no sucede con frecuencia) no se produce el rechazo. Por este motivo, antes de llevar a cabo un transplante de órganos, es imprescindible realizar pruebas para asegurarse de que los autoantígenos de las células del donante y del receptor son iguales o muy semejantes. Además de la acción de los linfocitos TC, también intervienen en el rechazo de los órganos transplantados la respuesta específica humoral (anticuerpos) y la respuesta inespecífica (macrófagos, activación del complementos9. El proceso de rechazo puede iniciarse 48 horas después del transplante, debido a una hipersensibilidad de tipo II (rechazo hiperagudo), o bien al cabo de varias semanas o meses como resultado de una hipersensibilidad de tipo IV originada por el contacto prolongado con los linfocitos TD. Para evitar el rechazo se suministran al enfermo fármacos inmunosupresores que inhiben la respuesta inmunitaria, principalmente esteroides, que actúan sobre los macrófagos, y ciclosporina, que bloquea los receptores de las interleucinas. 6.4. INMUNODEFICIENCIAS ADQUIRIDAS En éste tipo de enfermedades la deficiencia inmunitaria no se transmite, sino que aparece en cualquier momento de la vida como consecuencia de diversos factores, entre los que figuran la leucemia, la exposición a radiaciones o el tratamiento durante largo tiempo con inmunosupresores como los esteroides. Así mismo, algunas enfermedades graves que debilitan el sistema inmunitario pueden provocar su aparición. Entre las inmunodeficiencias adquiridas mas graves, ya que no se dispone de un tratamiento efectivo para combatirlas, se encuentra el sida, producido por el VIH, que ataca y destruye los linfocitos TH. La inmunodeficiencia provocada impide una defensa eficaz contra los antígenos microbianos y contra las células tumorales, por lo que la persona afectada sufre infecciones importantes y desarrolla algunos tipos de cánceres. El VIH es un retrovirus, por lo que tiene ARN como material genético. En concreto, posee dos cadenas iguales de ARN monocatenario. Su cápsida es icosaédrica y tiene una compleja cubierta lipoproteica. El proceso infeccioso de la enfermedad se desarrolla de la siguiente manera: 1. El VIH penetra en el cuerpo de una persona sana procedente de otra infectada. El virus se localiza en la sangre y en otros fluidos orgánicos como el semen y las secreciones vaginales, desde donde accede al nuevo hospedador. Los principales mecanismos de transmisión son: Directamente a través de la sangre, por transfusiones o por medio de jeringuillas contaminadas. Mediante relaciones sexuales en los que el semen o las secreciones vaginales entran en contacto con microheridas y erosiones por las que el virus pueda introducirse. De madre a hijo, generalmente durante la gestación o el parto. 2. Una vez el virus alcanza el sistema circulatorio del nuevo hospedador se une a los linfocitos T4. La unión se efectúa entre una proteína determinada de la cubierta del virus (gp 120) y el receptor CD4 de los linfocitos T4, gracias a un reconocimiento específico entre ambas moléculas. También puede producirse la unión con los macrófagos. 3. Se produce luego la fusión de la envoltura del virus con la membrana celular del linfocito, y el ácido nucleico (ARN monocatenario) y la enzima transcriptasa inversa del virus penetran en el citoplasma del linfocito. 4. Gracias a la transcriptasa inversa (una ADN polimerasa) se forma ADN bicatenario a partir del ARN, que se incorpora al genoma del linfocito, y se construye un provirus, de modo que el ADN de éste, integrado en el ADN celular, se transmite a las células hijas cada vez que el linfocito se divide. En ésta etapa no se observan alteraciones en las células con el ADN vírico, pero todas tienen una copia de éste es su propio genoma. Se produce, además, una multiplicación lenta del virus, que se libera por gemación. Los linfocitos no sufren daños inmediatos, pero sí a largo plazo. 5. En cierto momento los linfocitos T4 infectados mueren. El descenso en su número es lo que provoca la inmunodeficiencia. Cuando existen menos de 150 linfocitos T4 por mm3 de sangre (en una persona sana hay 500/mm3), la deficiencia inmunitaria es grave y la muerte sobreviene por infecciones oportunistas, como tuberculosis pulmonar y neumonía, entre otras. También es frecuente la aparición de tumores raros, como el sarcoma de Kaposi, que afecta a los capilares sanguíneos de la piel.