Receptores y mecanismos neuromusculares periféricos.
Anuncio

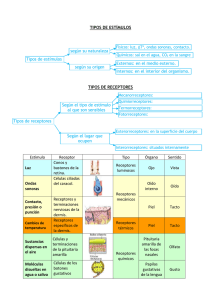
Universidad Andrés Bello Facultad de Odontología Fisiología Oral Texto de Apoyo; Receptores y mecanismos neuromusculares periféricos. Los sistemas sensoriales codifican cuatro atributos del estímulo que pueden ser correlacionados de forma cuantitativa con una sensación. Pese a la diversidad de sensaciones que experimentamos, todos los sistemas sensoriales transmiten cuatro tipos básicos de información cuando son estimulados: modalidad, localización, intensidad y duración. Juntos, estos cuatro atributos elementales de un estímulo originan una sensación. El hecho de que todos los sistemas sensoriales transmitan el mismo tipo de información puede ser la razón por la cual tienen una organización tan similar. Los cuatro atributos fundamentales de la experiencia sensorial son codificados dentro del sistema nervioso por subgrupos de neuronas especializadas. La modalidad define una clase general de estímulos, determinada por el tipo de energía transmitida por éste y por los receptores especializados para sentir esa energía (fig. 21-1). Los receptores, junto con sus vías centrales y sus áreas de destino en el cerebro, forman un sistema sensorial, y la actividad dentro de un sistema da origen a tipos específicos de sensaciones, como el tacto, el gusto, la visión o el oído. La localización del estímulo está representada por el conjunto de receptores sensoriales dentro del sistema que son activos. Los receptores están distribuidos de forma topográfica en un órgano sensorial, de tal manera que su actividad no sólo señala la modalidad del estímulo sino también su posición en el espacio y su tamaño. Como un estímulo activa muchos receptores de forma simultánea, la distribución de la población activa proporciona al cerebro una importante información sobre la sensación. La intensidad del estímulo está determinada por la amplitud de la respuesta de cada receptor, que refleja la cantidad total de energía del estímulo liberada al receptor. La duración del estímulo se define por el momento en el que comienza y termina la respuesta del receptor, y viene determinada por la rapidez a la cual la energía es recibida o perdida por el receptor. Por ello, tanto la intensidad como la duración del estímulo están representadas por los patrones de activación de las neuronas sensitivas activas. La modalidad sensorial viene determinada por la energía del estímulo. En cada sistema sensorial, el contacto inicial con el mundo se produce a través de estructuras nerviosas especializadas, llamadas receptores sensoriales. El receptor sensorial es la primera célula de cada vía sensitiva y transforma la energía del estímulo en energía eléctrica, estableciendo de esta manera un mecanismo de transmisión de señales común para todos los sistemas sensoriales. La señal eléctrica producida por el receptor recibe el nombre de potencial de receptor. La amplitud y la duración del potencial de receptor están relacionadas con la intensidad y la duración de la estimulación de ese receptor en particular. El proceso por el cual la energía específica del estímulo se convierte en señal recibe el nombre de transducción del estímulo. Los receptores están especializados morfológicamente para transformar formas específicas de energía. Cada receptor tiene una región anatómica especializada en la que se produce la transducción del estímulo. La mayoría de los receptores sensoriales son óptimamente selectivos para una única energía de estímulo, una propiedad denominada, especificidad del receptor. El estímulo particular que activa un receptor específico a un bajo nivel de energía fue denominado estímulo adecuado por Charles Sherrington. La especificidad de la respuesta en los receptores es la base del código de línea marcado, que es el mecanismo de codificación más importante para cada modalidad del estímulo. El hecho de que el receptor sea selectivo para un tipo particular de energía de estímulo significa que el axón del receptor funciona como una línea de comunicación específica para una modalidad; la actividad del axón transmite necesariamente información sobre un tipo particular de estímulo. La excitación de una neurona sensitiva en particular, ya sea de forma natural o artificialmente mediante estimulación eléctrica directa, desencadena la misma sensación. Por ejemplo, la estimulación eléctrica del nervio auditivo puede utilizarse en pacientes con sordera causada por una lesión de los receptores del oído interno. Cada clase de receptores sensoriales establece conexiones con estructuras particulares del sistema nervioso central, al menos en los primeros estadios del procesamiento de la información. Así la vista o el tacto se experimentan porque se activa una estructura particular del sistema nervioso central. La modalidad está representada, por lo tanto, por el conjunto de neuronas conectadas a una clase específica de receptores. Esos conjuntos de neuronas reciben el nombre de sistemas sensoriales, y forman los sistemas somatosensoriales, visual, auditivo, vestibular, olfatorio y gustativo. Los receptores transforman tipos específicos de energía en una señal eléctrica. Los seres humanos tienen cuatro clases de receptores, cada uno de los cuales es sensible primordialmente a una forma de energía física: mecánica, química, térmica o electromagnética (cuadro 21-1). Los mecanoreceptores del sistema somatosensorial intervienen en la sensibilidad táctil, la propiosensibilidad (extensión o contracción musculares) y la sensibilidad artrocinética, mientras que los mecanoreceptores del oído interno intervienen en la audición y el sentido del equilibrio. En la sensibilidad dolorosa, la sensibilidad al picor y los sentidos del gusto y del olfato intervienen quimioreceptores. Los termoreceptores de la piel notan la temperatura del cuerpo y también del aire ambiental, así como los objetos que nos tocan. Los seres humanos no poseen más que un tipo de receptor para la energía electromagnética: los fotoreceptores de la retina. Los mecanismos para la transformación de la energía del estímulo en el potencial de receptor varían según los tipos de estímulos físicos. Los mecanoreceptores notan la deformación física del tejido en el que se encuentran. La presión mecánica, como la ejercida sobre la piel o la extensión muscular, es transformada en energía eléctrica por el efecto físico del estímulo sobre los canales catíonicos de la membrana que están enlazados con el citoesqueleto (fig. 21-2 A). La estimulación mecánica deforma el receptor de la membrana y de esta forma abre los canales sensibles a la extensión y aumenta la conductancia iónica que despolariza el receptor (fig. 21- 2 B). El potencial de receptor despolarizante es por lo tanto similar, en cuanto a su mecanismo, al potencial postsináptico excitador. La amplitud del potencial de receptor es proporcional a la intensidad del estímulo; al abrir más canales iónicos y durante un mayor tiempo, una presión fuerte ejerce una mayor despolarización que una presión débil. La suspensión del estímulo alivia la sobrecarga mecánica sobre la membrana del receptor y hace que se cierren los canales sensibles al estiramiento. Los potenciales de receptor en los quimioreceptores y fotoreceptores son generados por segundos mensajeros intracelulares activados cuando la sustancia estimulante se enlaza a receptores de membrana acoplados a proteínas G (fig. 21- 3). Los segundos mensajeros producen cambios de conductancia localmente o en sitios remotos. Cada receptor responde a estrechos márgenes de energía del estímulo. Cada una de las principales modalidades tiene varias cualidades constitutivas o submodalidades. Por ejemplo, el gusto puede ser dulce, amargo, salado o agrio; los objetos que vemos difieren en color, forma y movimiento, y el tacto tiene las cualidades de temperatura, estructura y rigidez. Las submodalidades existen porque las distintas clases de receptores: quimioreceptores, mecanoreceptores, termoreceptores y fotoreceptores, no son homogéneas. En vez de ello, cada clase contiene varios receptores especializados que responden a limitados márgenes de energías del estímulo. El receptor se comporta como un filtro para unos estrechos márgenes, o anchura de banda, de energía. Por ejemplo, los fotoreceptores no son sensibles a todas las longitudes de onda de la luz, sino tan sólo a una pequeña parte del espectro. Decimos que los receptores están afinados para un estímulo adecuado, el único que activa un receptor de baja energía. A pesar de esto, la sensibilidad de una fibra nerviosa sensitiva para un tipo particular de estímulo no es absoluta. La distribución espacial de las neuronas sensitivas activadas por un estímulo transmite información sobre la localización del estímulo. La disposición espacial de los receptores activados dentro de un órgano sensorial transmite importante información relativa al estímulo. En las modalidades de sensación somática y de visión, la distribución espacial de los receptores transmite información sobre la localización del estímulo en el cuerpo o en el mundo exterior. En estas modalidades, el reconocimiento espacial implica tres cualidades perceptivas distintas: 1) la localización del sitio de estimulación sobre el cuerpo o de la fuente del estímulo en el espacio; 2) la discriminación del tamaño y de la forma de los objetos; y 3) la resolución del detalle fino del estímulo o el ambiente. Estas capacidades espaciales están ligadas a la estructura del campo receptivo de cada neurona sensitiva, que es el área dentro de la capa receptiva donde la estimulación excita la célula. La posición del campo receptivo es un factor importante en la percepción de la localización de un estímulo sobre el cuerpo. Los campos receptivos de las neuronas sensitivas de los sistemas somatosensorial y visual definen la resolución espacial de un estímulo. El campo receptivo de una neurona sensitiva en la sensación somática y en la visión asigna una localización topográfica específica a la información sensorial. Por ejemplo, el campo receptivo de un mecanoreceptor para el tacto es la región de la piel inervada directamente por los terminales de la neurona receptora, y así incluye toda el área de la piel a través de la cual un estímulo táctil puede ser conducido para alcanzar los terminales nerviosos (fig. 21-5). Cada receptor responde sólo a la estimulación dentro de su campo receptivo. Un estímulo que afecta a un área mayor que el campo receptivo de un receptor activará los receptores adyacentes. Por lo tanto, el tamaño de un estímulo influye en el número total de receptores que son estimulados. Un objeto grande, como una pelota de baloncesto que se mantiene sujeta con ambas manos, entrará en contacto y activará más receptores táctiles que un lapicero sujetado por los dedos pulgar e índice. La densidad de los receptores en una parte concreta del cuerpo determina hasta qué punto el sistema sensorial puede resolver el detalle de los estímulos en esa área. Una población densa de receptores permite una resolución más fina del detalle espacial, porque los receptores tienen campos receptivos más pequeños (fig. 21-6). Ahora bien, la resolución espacial de un sistema sensorial no es uniforme a través de toda la capa receptora. Por ejemplo, la discriminación espacial es muy aguda en las yemas de los dedos y en la parte central de la retina, donde hay muchos receptores sensitivos y donde los campos receptivos son pequeños. En otras regiones, como el tronco o los bordes externos de la retina, la información espacial señalada por los nervios individuales es menos precisa, porque los receptores son menos numerosos en esas áreas y por lo tanto tienen campos receptivos más grandes. Estas diferencias en la densidad de receptores se reflejan en el sistema nervioso central a través de los mapas del cuerpo creados por la disposición topográfica de las señales aferentes. En cada mapa las regiones del cuerpo más inervadas ocupan las áreas más grandes, mientras que las regiones escasamente inervadas ocupan áreas más pequeñas, debido al menor número de señales que reciben. La intensidad del estímulo en los nervios sensitivos es codificada por la frecuencia de los potenciales de acción. El cambio del potencial de membrana producido por el estímulo sensorial se transforma en un código de pulsación digital en el que la frecuencia de los potenciales de acción, refleja la amplitud del potencial de receptor. Los estímulos intensos provocan potenciales de receptor más grandes, que generan un mayor número y una mayor frecuencia de potenciales de acción. Además de aumentar la frecuencia de activación de las neuronas sensitivas individuales, los estímulos más intensos también activan un mayor número de receptores. Por ello, la intensidad de un estímulo está también codificada por el tamaño de la población receptora que responde. Estos códigos de población dependen del hecho de que los receptores individuales de un sistema sensorial difieren en cuanto a sus umbrales sensitivos. La mayoría de los sistemas sensoriales tienen por lo menos dos clases de receptores: de umbral bajo y de umbral alto. Según va aumentando la intensidad del estímulo, de débil a fuerte, responden primero los receptores de umbral bajo y después loas de umbral alto. La duración de una sensación viene determinada en parte por la velocidad de adaptación de los receptores. Las propiedades temporales de un estímulo están codificadas como cambios en la frecuencia de la actividad de la neurona sensitiva. Los estímulos aparecen, crecen en intensidad, fluctúan o permanecen estables y, finalmente, desaparecen. Muchos receptores señalan la velocidad a la que el estímulo aumenta o disminuye de intensidad mediante un rápido cambio en su ritmo de activación. Por ejemplo, cuando una varilla toca la piel, la activación inicial es proporcional tanto a la velocidad a la que se deprime la piel como a la cantidad total de presión. Durante una presión constante, el ritmo de activaciones disminuye a un nivel proporcional a la depresión de la piel. Las activaciones cesan cuando se retira la varilla. Así pues, las neuronas señalan importantes propiedades de los estímulos no sólo cuando lanzan impulsos sino también cuando dejan de hacerlo. Aunque la activación continua de una neurona sensitiva codifica la intensidad del estímulo, si éste persiste durante varios minutos sin cambiar su posición o su amplitud, su intensidad disminuye y se pierde la sensación. Esta disminución recibe el nombre de adaptación. Todos los receptores sensitivos se adaptan a una estimulación constante. Se piensa que esta adaptación constituye una importante base nerviosa de adaptación perceptiva en la que un estímulo constante deja de ser consciente. Los receptores pueden adaptarse en forma lenta o rápida. Los que responden a una estimulación prolongada y constante se conocen como receptores de adaptación lenta. Estos receptores son capaces de señalar magnitudes de estímulos durante varios minutos. La duración del estímulo es señalada por la despolarización y la generación persistente de potenciales de acción a lo largo del período de estimulación (fig. 21-9 A). Estos receptores se adaptan gradualmente a un estímulo como resultado de una activación lenta de los canales de Na+ o de Ca++ por el potencial de receptor despolarizante, o como resultado de la activación de canales de K+ dependientes del calcio. Algunos receptores dejan de activarse en respuesta a una estimulación de amplitud constante y sólo se activan cuando aumenta o disminuye la intensidad del estímulo. Estos receptores de adaptación rápida sólo responden al comienzo y al final de un estímulo, y señalan el ritmo o la velocidad de la estimulación (fig. 21-9 B). La adaptación de los receptores de adaptación rápida depende de dos factores. En primer lugar, en muchos de ellos la despolarización prolongada del potencial de receptor activa el mecanismo de generación de impulsos en el axón. En segundo lugar, la estructura del receptor filtra los componentes constantes del estímulo cambiando de forma, por lo que disminuyen la señal eléctrica generada por el receptor (fig. 21-10). La existencia de dos clases de receptores (sensores de adaptación lenta y rápida) demuestra otro importante principio de la codificación de las sensaciones. Los sistemas sensoriales detectan contrastes en estímulos individuales, es decir, cambios del patrón de estimulación en tiempo y espacio. Los receptores de adaptación rápida captan los derivados temporales de los estímulos (velocidad y aceleración) que señalan movimiento. Los ritmos de activación de estos receptores son proporcionales a la velocidad del movimiento y dejan de lanzar impulsos cuando el estímulo entra en fase de reposo. La activación de los receptores de adaptación rápida al comienzo y al final de la estimulación envía información sobre el medio sensorial cambiante al cerebro. Los sistemas sensoriales tienen un plan común. Los diferentes sistemas sensoriales utilizan códigos nerviosos similares para las propiedades de modalidad, localización, intensidad y duración de los estímulos físicos. Cuando una neurona sensitiva se activa comunica al cerebro que se ha recibido una determinada forma de energía en una localización específica del órgano sensorial. Los detalles del código del potencial de acción informarán al cerebro de cuánta energía se ha recibido en ese lugar, cuándo empezó, cuándo se detuvo y con qué rapidez cambió la intensidad. La información sensorial es transportada por grupos de neuronas sensitivas que actúan de forma conjunta La actividad de grupos completos de neuronas sensitivas está provocada por los miles y miles de estímulos que normalmente inciden a la vez en los receptores. Los mensajes de los sensores individuales son integrados, y no simplemente sumados, según convergen las señales en los centros de procesamiento del sistema nervioso central. La comprensión del modo en que la información sensitiva es transportada por receptores activados de manera simultánea es procesada en vías paralelas antes que se combinen en los centros más altos de la corteza cerebral constituye la base para la comprensión de la percepción sensorial. El procesamiento paralelo tiene una importancia particular en la visión, en la que casi todos los fotoreceptores de la retina reciben de forma simultánea estímulos luminosos de tono y de brillo diferentes. Para comprender una escena, el sistema visual necesita agrupar señales producidas por objetos individuales, separarlas y distinguir los objetos de interés del fondo común. Así en los seres humanos, todas las modalidades sensoriales, la más desarrollada es la visión, más de la mitad de la corteza procesa información visual. Los sistemas sensoriales procesan la información en una serie de núcleos de relevo Las vías que constituyen los sistemas sensoriales tienen una organización seriada. Los receptores proyectan sus señales a las neuronas de primer grado del sistema nervioso central, las cuales lo hacen a su vez a las de segundo grado y superiores. Esta secuencia de conexiones establece una jerarquía funcional muy clara. Por ejemplo, en el sistema somatosensorial, las fibras aferentes primarias convergen en neuronas de segundo orden, localizadas por lo general en el sistema nervioso central, y de allí en neuronas de tercer orden y superiores (fig. 21-11). Los núcleos de relevo sirven para procesar la información sensorial y determinan si se transmite o no a la corteza. Filtran ruidos o actividades esporádicas de las diferentes fibras y no transmiten más que las secuencias fuertes de actividad reiterada de las fibras sensitivas individuales o la actividad transmitida de forma simultánea por múltiples receptores. Las conexiones convergentes de los receptores sensitivos dentro del núcleo de relevo permiten a cada una de las neuronas de una orden superior la interpretación del mensaje sensorial en el contexto de la actividad de los canales de entrada vecinos. Como las neuronas receptoras, las que están en cada núcleo de relevo sensitivo tienen un campo receptivo. Este campo está definido por el grupo de células presinápticas que convergen en él. Los campos receptivos de las neuronas sensitivas de segundo orden o superior son mayores y más complejas que las neuronas receptoras. Son mayores porque reciben información convergente de muchos cientos de receptores, cada uno de ellos con un campo receptivo ligeramente diferente, pero superpuesto. Son más complejos porque son sensibles a características específicas del estímulo, como el movimiento en una dirección particular en el campo visual. Las interneuronas inhibidoras presentes dentro de cada núcleo de relevo ayudan a afinar el contraste entre los distintos estímulos. Al contrario que el campo receptivo uniformemente excitador del receptor sensorial, el campo receptivo de las neuronas sensitivas de órdenes superiores de los sistemas visual y somatosensorial tiene tanto regiones excitadoras como inhibidoras. La inhibición es producida por interneuronas inhibidoras en los núcleos de relevo. La región inhibidora de un campo receptivo es un medio importante para realzar el contraste entre distintos estímulos, y de esta forma otorga a los sistemas sensoriales una capacidad adicional para resolver el detalle espacial. Las interneuronas inhibidoras son activadas por tres vías distintas (fig. 21-12). La más importante es aquella en la que las fibras aferentes de los receptores o las neuronas de relevo de órdenes inferiores establecen conexiones con interneuronas inhibidoras que tienen conexiones con neuronas de proyección cercanas en el núcleo. Esta inhibición por alimentación anterógrada por fibras aferentes permite a las aferentes más activas reducir la salida de información de las neuronas de proyección adyacentes, menos activas. Esto permite lo que sherrington denominó singularidad de acción, una estrategia según la cual el ganador se lo lleva todo, que asegura la expresión de sólo una de dos o más respuestas competitivas. Las interneuronas inhibidoras también pueden ser activadas por las neuronas de proyección del núcleo de relevo a través de colaterales axónicas recurrentes procedentes de las neuronas de proyección. Esta inhibición por retroalimentación permite a las neuronas de salida más activas limitar la actividad de las menos activas. Estas redes inhibidoras crean zonas de actividad contrastante dentro del sistema nervioso central: una zona central de neuronas activas rodeadas de un anillo de neuronas menos activas (fig. 21-11 B). Además de los circuitos de alimentación anterógrada y retroalimentación locales para inhibición en un núcleo de relevo, las interneuronas inhibidoras pueden ser activadas por neuronas de sitios más distantes, como la corteza cerebral. De ésta manera, centros cerebrales más altos pueden controlar el flujo de información a través de los núcleos de relevo.