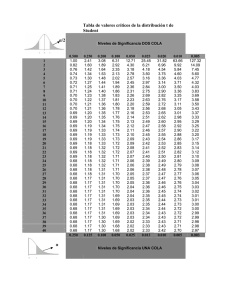
Dinámica poblacional y utilización ontogénica del hábitat por el
Anuncio

Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional Unidad Mérida Departamento de Recursos del Mar Dinámica poblacional y utilización ontogénica del hábitat por el caracol rosa (Strombus gigas), en el parque Xel-Há, Quintana Roo, México Tesis que presenta Biol. Joanne R. Peel Para obtener el grado de Maestra en Ciencias En la especialidad de Biología Marina Director de la Tesis: Dalila Aldana Aranda Mérida, Yucatán Noviembre del 2012 i Dedicatoria A mis padres Thomas y Petra Peel y mi hermana Jessika, por su apoyo incondicional durante toda la carrera. A mi pareja Jorge Saldaña, por regalar me la calma y seguridad en mi hogar durante este tiempo. A mi abuelo Edward Peel, por su apoyo económico y las palabras sabias. A mi tutor de la Universidad de Guadalajara, el Dr. Eduardo Juárez Carillo, por sus consejos como maestro y amigo. ii “There's a lady who is sure all that glitters is gold And she's buying a stairway to heaven. When she gets there she knows, if the stores are all closed With a word she can get what she came for. And she's buying a stairway to heaven. There's a sign on the wall but she wants to be sure 'Cause you know sometimes words have two meanings. In a tree by the brook, there's a songbird who sings, Sometimes all of our thoughts are misgiven. It makes me wonder. There's a feeling I get when I look to the west, And my spirit is crying for leaving. In my thoughts I have seen rings of smoke through the trees, And the voices of those who stand looking. It really makes me wonder. And it's whispered that soon if we all call the tune Then the piper will lead us to reason. And a new day will dawn for those who stand long And the forests will echo with laughter. If there's a bustle in your hedgerow, don't be alarmed now, It's just a spring clean for the May Queen. Yes, there are two paths you can go by, but in the long run There's still time to change the road you're on. And it makes me wonder. Your head is humming and it won't go, in case you don't know, The piper's calling you to join him, Dear lady, can you hear the wind blow, and did you know Your stairway lies on the whispering wind. And as we wind on down the road Our shadow is taller than our soul. There walks a lady we all know Who shines white light and wants to show How everything still turns to gold. And if you listen very hard The tune will come to you at last. When all is one and one is all To be a rock and not to roll. And she's buying a stairway to heaven.” Robert Plant iii Resumen En este trabajo se estudió la demografía, dinámica poblacional y movilidad del caracol rosa Strombus gigas mediante el método de captura-marcaje-recaptura en cuatro sitios de la caleta (Cueva, Centro, Bocana y Brazo Norte) y el análisis de frecuencia de tallas a lo largo del periodo 2005 a 2008 y 2009 a 2011, con la finalidad de determinar cómo esta especie hace uso de su hábitat y como se conectan las distintas etapas de su ciclo de vida. Se marcaron 1665 individuos entre 2005 y 2008 y 6627 entre 2009 y 2011. La abundancia de S. gigas incrementó, capturándose en promedio 219 ±49.0 caracoles por muestreo en el periodo 2005-2008 y 1096 ± 388.1 caracoles en el periodo 2009-2011. La población estuvo compuesta por juveniles en un 70% y la estructura de la población se mantuvo relativamente constante a través del tiempo. En los sitios Cueva, Centro y Brazo Norte se capturaron principalmente juveniles y en Bocana fueron en su mayoría adultos. La densidad reflejó una agregación natural sin impacto de pesca. El crecimiento de los caracoles de la caleta fue determinado directamente y se estimaron los parámetros de la ecuación de Von Bertalanffy. Éste fue alto en comparación con otros sitios del Caribe, pero comparable con otros estudios de marcaje-recaptura. El reclutamiento tuvo dos pulsos al año y fue máximo en los meses de junio a agosto, pero varía significativamente entre años y fue el factor con mayor impacto sobre el tamaño de población. La mortalidad reflejó niveles naturales, sin embargo fue muy alto en caracoles con una talla mayor a 200 mm. Los organismos realizaron transiciones en su hábitat lo cual fue asociado a su talla. La movilidad aumentó en organismos adultos y sub-adultos y fue mayor durante los meses de verano. Se encontraron evidencias que la alta mortalidad se puede atribuir a emigración. En base de la densidad, estructura de tallas y crecimiento se concluyó que la caleta de Xel-Há funciona como una zona de crianza para el caracol rosa. iv Abstract In the present work the demography, population dynamic and mobility of the queen conch Strombus gigas was studied, using a capture-mark-recapture sampling scheme and length frequency analysis, at four sites of the inlet of Xel-Há (Cueva, Centro, Bocana and Brazo Norte), through the years 2005 to 2008 and 2009 to 2011, in order to determine how S. gigas uses its habitat and how the different stages of its life cycle connect in it. One thousand six hundred and sixty five individuals were marked between 2005 and 2008 and 6627 between 2009 and 2011. S. gigas abundance increased from 219 ± 49.0 conchs per sample in 2005-2008 to 1096 ± 388.1 conchs in 2009-2011. The population was composed of 70% juveniles and population structure was relatively constant over time. At the sites Cueva, Centro and Brazo Norte mainly juveniles were captured and at Bocana mostly adults were encountered. The density of animals reflected a natural aggregation without fishing impact. Growth was determined directly and by estimating Von Bertalanffy growth parameters. Values were high compared with other locations of the Caribbean, but similar to studies conducted under mark-recapture sampling schemes. Recruitment peaked twice per year and was highest from June to August but showed significant differences between years and had a major impact over population size. Mortality reflected natural ranges, however was higher in conch bigger than 200 mm. Animals moved across their habitat, which was related to size, increasing in adults and sub-adults. Animals were more active during the summer months. There was evidence that high mortalities were linked with emigration. Based on the density, size structure and growth it was concluded that Xel-Há serves as a nursery for the queen conch. v Agradecimientos Al Consejo Nacional de Ciencia y Tecnología (Conacyt) por la beca N° 374674/2433757que me permitió realizar mis estudios de maestría. Al Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional por brindarme la oportunidad de realizar mis estudios de maestría y por el apoyo económico recibido. Al proyecto “Variación espacio-temporal del patrón reproductivo del caracol Strombus gigas en diferentes hábitats y su modelo biofísico de conectividad para el Caribe” del Conacyt con clave 50094 que permitió los monitoreo y la base de datos de 2005-2011 utilizada en esta tesis. A los fondos propios del laboratorio de Biología y cultivo de Moluscos del CINVESTAV IPN Unidad Mérida y a su personal que permitieron los monitoreos y la base de datos de 2001 a 2011 utilizada en esta tesis. A mi directora de tesis, la Dra. Dalila Aldana Aranda (alias La Doit), por haber me permitido desempeñar me en su laboratorio y por las múltiples oportunidades de difundir y presentar mi trabajo públicamente, brindando apoyo económico e intelectual. También le agradezco por las críticas duras, que me forzaron a reflejar profundamente y perseguir continuamente la perfección de este y futuros trabajos. Gracias. Al Dr. Pedro Ardisson Herrera, en su calidad de docente, le agradezco por despertar en mí nuevamente mi gran pasión por la ecología en muchas horas de clases, lectura y discusión. También le agradezco por haber me brindado su apoyo como un tutor intelectual y moral durante los últimos dos años y por la confianza que tuvo en mi persona. vi Al Dr. Ismael Mariño Tapia, como docente y sínodo, le agradezco por sus observaciones críticas y justas, su apoyo en todo momento, excelentes clases y el buen humor. A mi sínodo, el Dr. Alberto De Jesús Navarrete, por su esfuerzo de revisión de tesis y las correcciones y observaciones que recibí. A mi sínodo, el Dr. Richard Appeldoorn, por los consejos y observaciones sobre mi trabajo y la retroalimentación positiva. Al Dr. Erick Baqueiro Cárdenas por sus asesorías en el uso del programa FISAT II, interpretación de resultados y esfuerzo de revisión final de la sección de dinámica poblacional de esta tesis, así como por la oportunidad de discusión apasionada, porque me motivó a leer y practicar un poco más para eventualmente ganar la algún día con razón y argumentos fundamentados. A la coordinadora académica, la Dra. Silvia Salas Márquez, por el apoyo y orientación que me brindó en cuestiones académicas y a veces también sentimentales. A la secretaria, María Isabel Baqueiro, quien me acompañó desde el primer día hasta el último día a través de esta experiencia, siempre con una sonrisa, resolviendo todos los problemas con gran competencia. Al equipo de trabajo del laboratorio de Biología y Cultivo de Moluscos (Dra. Martha Enríquez, Manual Sánchez, Josefina Santos, Irma García, José Francisco Chávez, Wendy González, Ponchito, Panchito y Mariana) por los ánimos brindados, el apoyo y la rica comida. A mis docentes de maestría, Dr. Capurro, María Eugenia Vega, Jorge Herrera, Jorge Montero, Daniel Torruco, Gerado Gold y David Váldez, por el tiempo invertido en mi formación. vii A Ricardo Saénz Morales por permitirme realizar mi trabajo de campo en las instalaciones del parque Acuático Xel-Há y por su gran amistad. A Lic. Elizabeth Lugo, gerente executiva del parque Xel-Há, por el apoyo moral y económico recibido en el parque Xel-Há. A mi súper equipo de trabajo de Xel-Há, a Enrique May, Tere Rivas, Mario Hoil, Ángel Echeverría, Rafael Benítez, por sacrificar sangre y sudor junto conmigo en campo y haber hecho posible este gran trabajo. Agradezco mucho a mis padres, Thomas y Petra Peel, por apoyar me en cualquier manera imaginable, por sus buenos consejos y bondad, por su fe en mis habilidades, por animar me siempre perseguir mis sueños y por cultivar mi intuición y ética, entre mil cosas más que hicieron para mí. A mi hermana Jessika y su esposo Horst Beck, por patrocinar todas mis curiosidades electrónicas, los cuales me permitieron estar cerca con mi familia, aún estando lejos. A mi Abuelo Edward Peel, por su apoyo económico y por su confianza y fe en mis talentos académicos. A mi pareja, Jorge Saldaña, por su tolerancia, apoyo y amor durante esta travesía. A la compañera Wendy González por su amistad. A mi tutor y amigo de la UdeG, Eduardo Juárez Carillo, por ser buen amigo, por mandar papelitos ida y vuelta, por los buenos consejos y la gran empatía. A todos mis amistades, por las cenas, fiestas, platicas que me ayudaron a desconectar me en tiempos de gran presión. Al Dr. Canúl, médico del CINVESTAV, por mantener domado mi dolor de panza. viii Índice Dedicatoria………………………………………………………………………………...ii Resumen…..……………………………………………………………………………....iv Abstract………………………………………………………………………...……...…..v Índice……………………………………………………………………….……….……ix Lista de Tablas.……………………………………………………………...……...…....xii Lista de Figuras………………………………………………………………………….xiv 1. Introducción……………………………………………………………………………….1 2. Antecedentes………………………………………………………………………………4 2.1 Taxonomía………………………………………………………………………...…..4 2.2 Distribución geográfica……………………………………………………………..…4 2.3 Ciclo de vida y utilización del hábitat…………………………………………………5 2.4 Estudios de parámetros de población……………………………………………...…10 3. Problema central a Investigar………………………………………………………...…..19 4. Hipótesis………………………………………………………………………………....19 5. Objetivos…………………………………………………………………………………19 5.1 Objetivo General………………………………………………………………….….19 5.2 Objetivos particulares……………………………………………………………..…20 6. Justificación……………………………………………………………………………...20 7. Materiales y Métodos…………………………………………………………………….21 ix 7.1 Área de estudio…………………………………………………………………...….21 7.2 Métodos de Campo…………………………………………………………………..38 7.3 Métodos de análisis de datos……………………………………………………..…..41 7.3.1 Abundancia, Tamaño de Población y densidad……………………………41 7.3.2 Estructura de Tallas…………………………………………………...……42 7.3.3 Crecimiento por métodos directos……………………………………..…..46 7.3.4 Estimación de los parámetros de la ecuación de Von Bertalanffy……..….46 7.3.5 Reclutamiento………………………………………………………...……47 7.3.6 Sobrevivencia y Mortalidad………………………………………………..47 7.3.7 Movilidad…………………………………………………………………..49 8. Resultados………………………………………………………………………………..50 8.1 Demografía poblacional………………………………………….……………….….50 8.1.1 Abundancia, Tamaño de Población y densidad…………………………....50 8.1.2 Estructura de Tallas………………………………………………...………66 8.2 Crecimiento…………………………………………………………………………..86 8.2.1 Crecimiento de la heliconcha por métodos directos…………………….…86 8.2.2 Estimación de los parámetros de la ecuación de Von Bertalanffy…….….113 8.3 Dinámica Poblacional…………………………………………………………...….116 8.3.1 Reclutamiento…………………………………………………………….116 8.3.2 Sobrevivencia y Mortalidad…………………………………………..…..123 8.4 Movilidad…………………………………………………………………………...133 8.4.1 Dirección y frecuencia de transiciones…………………………………...133 8.4.2 Talla de transición…………………………………………………...……141 x 9. Discusión…………………………………………………………………………..……154 9.1 Demografía poblacional………………………………………………….….….…..154 9.1.1 Abundancia, Tamaño de Población y densidad………………….……….154 9.1.2 Estructura de Tallas……………………………………………….………158 9.2 Crecimiento…………………………………………………………………………160 9.2.1 Crecimiento de la heliconcha por métodos directos………………...……160 9.2.2 Estimación de los parámetros de crecimiento de la ecuación de Von Bertalanffy……………………………………………………………………...164 9.3 Dinámica Poblacional……………………………………………………………....169 9.3.1 Reclutamiento…………………………………………………………….169 9.3.2 Sobrevivencia y Mortalidad…………………………………..…………..170 9.4 Movilidad…………………………………………………………………………...172 9.4.1 Dirección y frecuencias de transiciones…………………………………..172 9.4.2 Tallas de transición…………………………………………….…………173 10. Conclusión………………………………………………………………….…………..175 11. Literatura Citada………………………………………………………………..………178 xi Lista de Tablas Tabla 1. Estudios de Población del caracol rosa S. gigas realizados en México……...………...13 Tabla 2. Estudios de Población del caracol rosa S. gigas realizados en la Caleta de Xel-Há, México………………………………………………………………...…………………………14 Tabla 3. Estudios de movimiento y migración del caracol rosa S. gigas realizados en el Caribe…………………………………………………………………………………………….16 Tabla 4. Estudios de crecimiento de S. gigas mediante métodos directos………….………..….17 Tabla 5. Estudios de crecimiento de S. gigas mediante análisis la estimación de los parámetros de la ecuación de von Bertalanffy.……………………………………………………………….18 Tabla 6. Granulometría de la caleta de Xel-Há (Xel-Há, 2008)………………….………….….37 Tabla 7. Abundancia media del caracol rosa S. gigas y desviación estándar (d.e.) en los periodos 2005-2008 y 2009-2011 en los cuatro sitios estudiados de la caleta de Xel-Há (Cueva, Centro, Bocana, Brazo Norte), obtenidas del número de individuos capturados y de las estimaciones del tamaño de población por los métodos de Jolly-Seber y Schnabel……..……………………...…57 Tabla 8. Abundancia media del caracol rosa S. gigas y desviación estándar (d.e.) en los periodos 2005-2008 y 2009-2011, en la caleta de Xel-Há, obtenidas del número de individuos capturados y de las estimaciones del tamaño de población por los métodos de Jolly-Seber y Schnabel……60 Tabla 9. Densidad (individuos ha-1) media poblacional del caracol rosa S. gigas y su desviación estándar (d.e.) en los periodos 2005-2008 y 2009-2011, en los cuatro sitios de la caleta de XelHá (Cueva, Centro, Bocana y Brazo Norte), obtenida del número de individuos capturados y por los métodos de Jolly-Seber y Schnabel……….……………………………………………….....62 Tabla 10. Densidad (individuos ha-1) media del caracol rosa S. gigas y su desviación estándar (d.e.) en los periodos 2005-2008 y 2009-2011, en la caleta de Xel-Há, obtenidas a partir del número de individuos capturados y por los métodos de Jolly-Seber y Schnabel…….………….65 Tabla 11. Análisis de varianza (Kruskal-Wallis) de medianas de longitud de heliconcha del caracol S. gigas entre cada uno de los cuatro sitios estudiados de la caleta de Xel-Há en el periodo de enero 2005 a septiembre 2011…..………………………………………………..….69 Tabla 12. Análisis de varianza (Kruskal-Wallis) de medianas de longitud de heliconcha del caracol S. gigas en el periodo 2005-2008 y 2009 2011 y en los sitios Cueva, Centro y Bocana de la caleta de Xel-Há………………………………………………...………………………….….69 Tabla 13. Análisis de varianza (Kruskal-Wallis) de medianas de longitud de heliconcha del caracol S. gigas en el periodo 2005-2008 y 2009 2011 en la caleta de Xel-Há…………......…72 xii Tabla 14. Análisis de varianza (Kruskal-Wallis) de medianas de del grosor de labio del caracol S. gigas entre los sitios Cueva, Centro, Bocana y Brazo Norte de la caleta de Xel-Há, en el periodo de enero 2005 a septiembre 2011……………………………………………………….75 Tabla 15. Análisis de varianza (Kruskal-Wallis) de medianas de del grosor de labio del caracol S. gigas en el periodo 2005-2008 y 2009 2011 y en los sitios Cueva, Centro y Bocana de la caleta de Xel-Há………………………………………………………………………………………..76 Tabla 16. Análisis de varianza (α=0.05) de las tasas de crecimiento medio (mm día⁻¹) del caracol S. gigas entre los sitios Bocana, Centro y Cueva, obtenidas en el periodo de enero 2005 a enero 2008………………………………………………………………………………………………90 Tabla 17. Análisis de varianza (α=0.05) de las tasas de crecimiento medio (mm día⁻¹) del caracol S. gigas entre los sitios Bocana, Centro, Brazo Norte y Cueva, obtenidas en el periodo de abril 2009 a septiembre 2011………………………………………………………………………….91 Tabla 18. Análisis de varianza (α=0.05) de las tasas de crecimiento medio (mm día⁻¹) del caracol S. gigas en la caleta de Xel-Há, entre los periodos de estudio 2005-2008 y 20092011………………………………………………………………………………………………92 Tabla 19. Densidad poblacional del caracol rosa S. gigas en el Caribe….................................156 Tabla 20. Tasas de crecimiento de S. gigas en el Caribe……………………………...……….162 Tabla 21. Parámetros de crecimiento de von Bertalanffy del caracol rosa, obtenidas en la caleta de Xel-Há……………………………………………………………………………………….165 Tabla 22. Parámetros de crecimiento de von Bertalanffy del caracol rosa en el Caribe…………………………………………………………………………………………...166 Tabla 23. Parámetros de crecimiento de von Bertalanffy para el grosor de labio del caracol rosa. …………………………………………………………………………………………………..167 Tabla 24. Mortalidad (z) del caracol rosa S. gigas en el Caribe estimado por curvas de captura de tallas convertidas………………………………………………………………………………..171 xiii Lista de Figuras Figura 1. Distribución geográfica del caracol rosa Strombus gigas………..................................5 Figura 2. Ciclo de vida del caracol rosa S. gigas………………………………………………..6 Figura 3. Ubicación geográfica del área de estudio, la Caleta de Xel-Há y de los sitios de Muestreo: Cueva, Brazo Norte, Centro y Bocana………………………………………………..22 Figura 4. Variación temporal de (A) la salinidad, (B) temperatura (°C), (C) oxígeno disuelto (mg/l) y (D) pH, en el fondo y la superficie de la columna de agua del sitio Cueva de la caleta de Xel-Há, durante el periodo de enero 2005 a septiembre 2011…………………………………...24 Figura 5. Variación temporal de (A) Nitritos (mg/l), (B) Nitratos (mg/l) (°C) y (C) Fósforo total (mg/l), en el fondo y la superficie de la columna de agua del sitio Cueva de la caleta de Xel-Há, durante el periodo de enero 2005 a septiembre 2011……………………………………………26 Figura 6. Variación temporal de (A) la salinidad, (B) temperatura (°C), (C) oxígeno disuelto (mg/l) y (D) pH, en el fondo y la superficie de la columna de agua del sitio Brazo Norte de la caleta de Xel-Há, durante el periodo de enero 2005 a septiembre 2011…………………………28 Figura 7. Variación temporal de (A) Nitritos (mg/l), (B) Nitratos (mg/l) (°C) y (C) Fósforo total (mg/l), en el fondo y la superficie de la columna de agua del sitio Brazo Norte de la caleta de Xel-Há, durante el periodo de enero 2005 a septiembre 2011…………………………………...30 Figura 8. Variación temporal de (A) la salinidad, (B) temperatura (°C), (C) oxígeno disuelto (mg/l) y (D) pH, en el fondo y la superficie de la columna de agua del sitio Centro de la caleta de Xel-Há, durante el periodo de mayo 2007 a septiembre 2011…………………………………...32 Figura 9. Variación temporal de (A) Nitritos (mg/l), (B) Nitratos (mg/l) (°C) y (C) Fósforo total (mg/l), en el fondo y la superficie de la columna de agua del sitio Centro de la caleta de Xel-Há, durante el periodo de mayo 2007 a septiembre 2011……………………………………………33 Figura 10. Variación temporal de (A) la salinidad, (B) temperatura (°C), (C) oxígeno disuelto (mg/l) y (D) pH, en el fondo y la superficie de la columna de agua del sitio Bocana de la caleta de Xel-Há, durante el periodo de enero 2005 a septiembre 2011………………………………..35 Figura 11. Variación temporal de (A) Nitritos (mg/l), (B) Nitratos (mg/l) (°C) y (C) Fósforo total (mg/l), en el fondo y la superficie de la columna de agua del sitio Bocana de la caleta de Xel-Há, durante el periodo de enero 2005 a septiembre 2011…………………………………...36 Figura 12. Granulometría de la caleta de Xel-Há…………………………………………...…..38 Figura 13. Método de marcaje del caracol rosa S. gigas………………………………….……39 Figura 14. Método para la medición de longitud de heliconcha del caracol rosa S. gigas ………………………….………………………………………………………………………...40 xiv Figura 15. Método de medición del grosor de labio del caracol rosa S. gigas…..……………..40 Figura 16. Número de caracoles S. gigas marcados en cada uno de los cuatro sitios estudiados de la Caleta de Xel-Há: (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, durante el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011………………………………....52 Figura 17. Número de caracoles S. gigas marcados en la Caleta de Xel-Há durante el periodo de (A) enero 2005 a enero 2008 y (B) de abril 2009 a septiembre 2011………………..…………..53 Figura 18. Número de caracoles S. gigas capturados (azul) y estimados mediante los métodos de Jolly-Seber (rojo) y Schnabel (verde), en cada uno de los cuatro sitios estudiados de la Caleta de Xel-Há: (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, durante el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011…………………………………………..56 Figura 19. Número de caracoles S. gigas capturados (azul) y estimados mediante los métodos de Jolly-Seber (rojo)y Schnabel (verde), en la Caleta de Xel-Há durante el periodo de (A) enero 2005 a enero 2008 y (B) de abril 2009 a septiembre 2011……………………………...……….59 Figura 20. Densidad poblacional (individuos ha⁻¹) del caracol rosa S. gigas en cada uno de los cuatro sitios estudiados de la Caleta de Xel-Há: (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, durante el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011………………………………………………………………………………………………61 Figura 21. Densidad poblacional (individuos ha⁻¹) del caracol rosa S. gigas en en la Caleta de Xel-Há durante el periodo de (A) enero 2005 a enero 2008 y (B) de abril 2009 a septiembre 2011……………………………………………………………………………………………....64 Figura 22. Diagrama de densidad de puntos, mostrando longitudes de heliconcha (mm) del caracol rosa S. gigas y abundancia por talla, en cada uno de los cuatro sitios de la Caleta de XelHá: (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, a través del periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011…………………………………………………..67 Figura 23. Diagrama de densidad de puntos, mostrando longitudes de heliconcha (mm) del caracol rosa S. gigas y abundancia por talla, en la Caleta de Xel-Há a través del periodo (A) enero 2005 a enero 2008 y (B) abril 2009 a septiembre 2011…….……………………...……..71 Figura 24. Diagrama de dispersión, mostrando la relación entre longitud de heliconcha (mm) y grosor de labio (mm) del caracol rosa S. gigas en cada uno de los cuatro sitios estudiados de la caleta de Xel-Há, (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, en el periodo de enero 2005 a septiembre………………………………………………………………………………..73 Figura 25. Diagrama de dispersión, mostrando la relación entre longitud de heliconcha (mm) y grosor de labio (mm) de la población del caracol rosa S. gigas en la caleta de Xel-Há, en los periodos (A) enero 2005 a enero 2008 y (B) abril 2009 a septiembre 2011……………..………74 Figura 26. Diagramas de caja mostrando las medidas de dispersión del grosor de labio (media, mediana, desviación estándar y valores extremos) del caracol rosa S. gigas a través del periodo enero 2005 a enero 2008 y de abril 2009 a septiembre 2011, en cada uno de los cuatro sitios xv estudiados de la Caleta de Xel-Há: (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte…………………………………………………………………………………………..…77 Figura 27. Diagramas de caja mostrando las medidas de dispersión del grosor de labio (media, mediana, desviación estándar y valores extremos) del caracol rosa S. gigas en la Caleta de XelHá a través del periodo (A) enero 2005 a enero 2008 y de (B) abril 2009 a septiembre 2011…………………………………………………………………………………………...….78 Figura 28. Abundancia relativa (%) del caracol rosa S. gigas por clases de talla, Adultos (grosor de labio ≥5 mm), Subadultos (grosor de labio < 5 mm) y Juveniles (sin labio), en cada uno de los cuatro sitios de la caleta de Xel-Há: (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, durante el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011……………..80 Figura 29. Abundancia relativa (%) del caracol rosa S. gigas por clases de talla, Adultos (grosor de labio ≥5 mm), Subadultos (grosor de labio < 5 mm) y Juveniles (sin labio), en la caleta de Xel-Há durante el periodo de (A) enero 2005 a enero 2008 y de (B) abril 2009 a septiembre 2011…………………………………………………………………………………………..…..81 Figura 30. Abundancia absoluta del caracol rosa S. gigas por clases de talla, Adultos (grosor de labio ≥5 mm), Subadultos (grosor de labio < 5 mm) y Juveniles (sin labio), en cada uno de los cuatro sitios de la Caleta de Xel-Há: (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, a través del periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011………………………………………………………………………………………………83 Figura 31. Abundancia absoluta del caracol rosa S. gigas por clases de talla, Adultos (grosor de labio ≥5 mm), Subadultos (grosor de labio < 5 mm) y Juveniles (sin labio), en la Caleta de XelHá, a través del periodo de (A) enero 2005 a enero 2008 y de (B) abril 2009 a septiembre 2011………………………………………………………………………………………………84 Figura 32. Análisis de componentes principales de la distribución espacial del caracol rosa S. gigas en cada uno de los cuatro sitios estudiados de la caleta de Xel-Há (Cueva, Centro, Bocana y Brazo Norte), durante el periodo de enero 2005 a septiembre 2011, en función de la longitud de heliconcha y del grosor de labio……………………………………………………………........85 Figura 33. Crecimiento medio (mm día-1) del caracol rosa S. gigas por clase de talla y desviación estándar, en los sitios (A) Cueva, (B) Centro y (C) Bocana, durante el periodo de enero 2005 a enero 2008…………………………………………………………………………87 Figura 34. Crecimiento medio (mm día -1) del caracol rosa S. gigas por clase de talla y desviación estándar, en los sitios (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, durante el periodo de abril 2009 a septiembre 2011……………………………………………………...89 Figura 35. Crecimiento medio (mm día -1) del caracol rosa S. gigas por clase de talla y desviación estándar, en la caleta de Xel-Há, durante el periodo de enero 2005 a enero 2008 (negro) y de abril 2009 a septiembre 2009 (gris)………………………………………………...92 xvi Figura 36. Incremento medio de longitud de heliconcha (LH) del caracol rosa S. gigas y edad estimada en años, en la caleta de Xel-Há durante el periodo de enero 2005 a enero 2008 (negro) y de abril 2009 a septiembre 2011 (gris)………………………………………………………...94 Figura 37. Media y desviación estándar del crecimiento (mm día -1) de individuos de S. gigas con tallas entre 90-99 mm, en los sitios Cueva, Centro y Brazo Norte (Caleta de Xel-Há), durante el periodo enero 2005 a septiembre 2011. ……………....................................................................95 Figura 38. Media y desviación estándar del crecimiento (mm día -1) de individuos de S. gigas con tallas entre 100-109 mm, en los sitios Cueva, Centro y Brazo Norte (Caleta de Xel-Há), durante el periodo enero 2005 a septiembre 2011......................................................................................97 Figura 39. Media y desviación estándar del crecimiento (mm día -1) de individuos de S. gigas con tallas entre 110-119 mm, en los sitios Cueva, Centro y Brazo Norte (Caleta de Xel-Há), durante el periodo enero 2005 a septiembre 2011………………………………………………………..99 Figura 40. Media y desviación estándar del crecimiento (mm día -1) de individuos de S. gigas con tallas entre 120-129 mm, en los sitios Cueva, Centro y Brazo Norte (Caleta de Xel-Há), durante el periodo enero 2005 a septiembre 2011………………..………………………………….….101 Figura 41. Media y desviación estándar del crecimiento (mm día -1) de individuos de S. gigas con tallas entre 130-139 mm, en cada uno de los cuatro sitios estudiados (Cueva, Centro, Bocana, Brazo Norte) de la caleta de Xel-Há, durante el periodo enero 2005 a septiembre 2011……………………………………………………………………………………………..103 Figura 42. Media y desviación estándar del crecimiento (mm día -1) de individuos de S. gigas con tallas entre 140-149 mm, en cada uno de los cuatro sitios estudiados (Cueva, Centro, Bocana, Brazo Norte) de la caleta de Xel-Há, durante el periodo enero 2005 a septiembre 2011………………………………………………………………………………………..……105 Figura 43. Media y desviación estándar del crecimiento (mm día -1) de individuos de S. gigas con tallas entre 150-159 mm, en cada uno de los cuatro sitios estudiados (Cueva, Centro, Bocana, Brazo Norte) de la caleta de Xel-Há, durante el periodo enero 2005 a septiembre 2011………………………………………………………………………………………….….107 Figura 44. Media y desviación estándar del crecimiento (mm día -1) de individuos de S. gigas con tallas entre 160-169 mm, en cada uno de los cuatro sitios estudiados (Cueva, Centro, Bocana, Brazo Norte) de la caleta de Xel-Há, durante el periodo enero 2005 a septiembre 2011……....109 Figura 45. Media y desviación estándar del crecimiento (mm día -1) de individuos de S. gigas con tallas entre 170-179 mm, en cada uno de los cuatro sitios estudiados (Cueva, Centro, Bocana, Brazo Norte) de la caleta de Xel-Há, durante el periodo enero 2005 a septiembre 2011……....111 Figura 46. Crecimiento del caracol rosa S. gigas en el periodo (A) 2005-2008, (B) 2009-2011, en la caleta Xel-Há obtenido mediante el método de Appeldoorn, utilizando incrementos obtenidos a partir de los datos de marcaje recaptura…………………………………………..112 xvii Figura 47. Crecimiento del caracol rosa S. gigas en el periodo (A) 2005-2008, (B) 2009-2011, en la caleta Xel-Há obtenido mediante el método de Appeldoorn, utilizando incrementos obtenidos de la progresión de modas…………………………………………………………...113 Figura 48. Crecimiento del grosor de labio del caracol rosa S. gigas en el periodo (A) 20052008 y (B) 2009-2011, en la caleta Xel-Há obtenidas mediante el método de Appeldoorn, utilizando incrementos obtenidos a partir de los datos de marcaje recaptura………………….114 Figura 49.A Frecuencia absoluta de longitud de heliconcha por clase de talla, de caracoles juveniles S. gigas en la caleta de Xel-Há, en el periodo de enero 2005 a diciembre 2005……………………………………………………………………………………………..116 Figura 49.B Frecuencia absoluta de longitud de heliconcha por clase de talla, de caracoles juveniles S. gigas en la caleta de Xel-Há, en el periodo de febrero 2006 a noviembre 2006…………………………………………………………………………………………..…117 Figura 49.C Frecuencia absoluta de longitud de heliconcha por clase de talla, de caracoles juveniles S. gigas en la caleta de Xel-Há, en el periodo de enero 2007 a enero 2008……..…118 Figura 49.D Frecuencia absoluta de longitud de heliconcha por clase de talla, de caracoles juveniles S. gigas en la caleta de Xel-Há, en el periodo de abril 2009 a octubre 2009………...119 Figura 49.E Frecuencia absoluta de longitud de heliconcha por clase de talla, de caracoles juveniles S. gigas en la caleta de Xel-Há, en el periodo de febrero 2010 a noviembre 2010……………………………………………………………………………………………..120 Figura 49.F Frecuencia absoluta de longitud de heliconcha por clase de talla, de caracoles juveniles S. gigas en la caleta de Xel-Há, en el periodo de enero 2011 a septiembre 2011……………………………………………………………………………………………..121 Figura 50. Porcentaje de reclutamiento de juveniles de S. gigas en la Caleta de Xel-Há, durante el periodo de (A) enero 2005 a enero 2008 y (B) abril 2009 a septiembre 2011, determinado por el método de análisis electrónico de frecuencia de longitud……………………………………122 Figura 51. Porcentaje medio y desviación estándar de la sobrevivencia finita (St=(Nt0/Nti)*100) del caracol rosa S. gigas en los cuatro sitios estudiados de la caleta de Xel-Há (Cueva, Centro, Bocana y Brazo Norte) en el periodo de (A) enero 2005 a enero 2008 y (B) abril 2009 a septiembre 2011………………………………………………………………………………...123 Figura 52. Porcentaje medio y desviación estándar de sobrevivencia finita (S t=(Nt0/Nti)*100) del caracol rosa S. gigas en los cuatro sitios estudiados de la caleta de Xel-Há (Cueva, Centro, Bocana y Brazo Norte) en el periodo de (A) enero 2005 a enero 2008 y (B) abril 2009 a septiembre 2011…………………………………………………………………………..….…124 Figura 53. Mortalidades finitas (zt=(-ln (Nt/N0)) medias y desviación estándar, del caracol rosa S. gigas en la Caleta de Xel-Há en el periodo de enero 2005 a enero 2008 (cuadrado) y abril 2009 a septiembre 2011 (círculo)…………………………………………………………………….125 xviii Figura 54. Número de individuos del caracol rosa S. gigas, que fueron encontrados muertos en los cuatro sitios estudiados de la caleta de Xel-Há (Cueva, Centro, Bocana y Brazo Norte) a través de los periodos de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011……………………………………………………………………………………………..126 Figura 55. Frecuencia relativa (%) de longitudes de heliconcha (mm) por clases de talla, del caracol rosa S. gigas sin labio (blanco) y con labio (negro), en la caleta de Xel-Há en el periodo de (A) enero 2005 a enero 2008 y de (B) abril 2009 a septiembre 2011……………………….127 Figura 56. Frecuencia relativa (%) del grosor de labio (mm) por clases de talla del caracol rosa S. gigas, en la caleta de Xel-Há en el periodo de (A) enero 2005 a enero 2008 y de (B) abril 2009 a septiembre 2011………………………………………………………………………………129 Figura 57. Curvas de captura de tallas convertidas (edad relativa), obtenidas de la frecuencia de distribución de longitud de heliconcha del caracol S. gigas en el periodo de (A) enero 2005 a enero 2008 de y (B) abril 2009 a septiembre 2011, en la caleta de Xel-Há…………………....130 Figura 58. Curvas de captura de tallas convertidas (edad relativa), obtenidas de la frecuencia de distribución del grosor de labio del caracol S. gigas en el periodo de (A) enero 2005 a enero 2008 de y (B) abril 2009 a septiembre 2011, en la caleta de Xel-Há………………………………...131 Figura 59. Abundancia relativa (%) de caracoles S. gigas capturados con marca originaria de Cueva, Centro, Bocana o Brazo Norte y sin marca, en los cuatro sitios estudiados de la caleta de Xel-Há (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011………………………………………………....133 Figura 60. Número de individuos de caracol S. gigas marcados en (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, que emigraron de su sitio de origen y destino de migración en la caleta de Xel-Há (Cueva, Centro, Bocana y Brazo Norte), a través del periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011……………………………………………...….135 Figura 61. Porcentaje de emigraciones de S. gigas marcados en la Cueva (Caleta de Xel-Há) por estación del año (Nortes, Secas y Lluvias), en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011………………………………………..…………………………...…..136 Figura 62. Porcentaje de emigraciones de S. gigas marcados en la Centro (Caleta de Xel-Há) por estación del año (Nortes, Secas y Lluvias), en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011…………………………………………………………………...137 Figura 63. Porcentaje de emigraciones de S. gigas marcados en la Bocana (Caleta de Xel-Há) por estación del año (Nortes, Secas y Lluvias), en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011…………………………………………………………………...138 Figura 64. Porcentaje de emigraciones de S. gigas marcados en la Brazo Norte (Caleta de XelHá) por estación del año (Nortes, Secas y Lluvias), en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011………………………………………………………………..139 xix Figura 65. Longitudes medias de heliconcha y desviación estándar de caracoles S. gigas marcados en (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, que emigraron de su sitio de origen, mostrando el destino de migración en la caleta de Xel-Há: Cueva (azul), Centro (verde), Bocana (morado) y Brazo Norte (rojo), a través del periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011……………………………………………………………...141 Figura 66. Medias y desviación estándar del grosor de labio de caracoles S. gigas marcados en (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, que emigraron de su sitio de origen, mostrando el destino de migración en la caleta de Xel-Há: Cueva (azul), Centro (verde), Bocana (morado) y Brazo Norte (rojo), a través del periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011………………………………………………………………………………143 Figura 67. Número de individuos marcados en Cueva, número de individuos que emigraron de Cueva, porcentaje de juveniles (caracoles sin labio), subadultos (caracoles con un grosor de labio < 5mm) y adultos (caracoles con un grosor de labio ≥ 5 mm) que participaron en las migraciones y destinos de migración: Centro (verde), Bocana (morado) o Brazo Norte (rojo), en el periodo de enero 2005 a septiembre 2011………………………………………………………………….145 Figura 68. Número de individuos marcados en Centro, número de individuos que emigraron de Centro, porcentaje de juveniles (caracoles sin labio), subadultos (caracoles con un grosor de labio < 5mm) y adultos (caracoles con un grosor de labio ≥ 5 mm) que participaron en las migraciones y destinos de migración: Cueva (azul), Bocana (morado) o Brazo Norte (rojo), en el periodo de enero 2005 a septiembre 2011……………………………………………………...146 Figura 69. Número de individuos marcados en Bocana, número de individuos que emigraron de Bocana, porcentaje de juveniles (caracoles sin labio), subadultos (caracoles con un grosor de labio < 5mm) y adultos (caracoles con un grosor de labio ≥ 5 mm) que participaron en las migraciones y destinos de migración: Cueva (azul), Centro (verde) o Brazo Norte (rojo), en el periodo de enero 2005 a septiembre 2011……………………………………………………...147 Figura 70. Número de individuos marcados en Brazo Norte, número de individuos que emigraron de Brazo Norte, porcentaje de juveniles (caracoles sin labio), subadultos (caracoles con un grosor de labio < 5mm) y adultos (caracoles con un grosor de labio ≥ 5 mm) que participaron en las migraciones y destinos de migración: Cueva (azul), Centro (verde) o Bocana (morado), en el periodo de enero 2005 a septiembre 2011……………………………………..148 Figura 71. Análisis de componentes principales del grosor de labio y longitud de heliconcha de caracoles S. gigas marcados en Cueva y recapturados en uno de los 4 sitios de muestreo de la caleta de Xel-Há (Cueva, Centro, Bocana o Brazo Norte), entre enero 2005 y septiembre 2011………………………………………………………………………………………….….149 Figura 72. Análisis de componentes principales del grosor de labio y longitud de heliconcha de caracoles S. gigas marcados en Centro y recapturados en uno de los 4 sitios de muestreo de la caleta de Xel-Há (Cueva, Centro, Bocana o Brazo Norte), entre enero 2005 y septiembre 2011.…………………………………………………………………………………………….150 Figura 73. Análisis de componentes principales del grosor de labio y longitud de heliconcha de caracoles S. gigas marcados en Bocana y recapturados en uno de los 4 sitios de muestreo de la xx caleta de Xel-Há (Cueva, Centro, Bocana o Brazo Norte), entre enero 2005 y septiembre 2011……………………………………………………………………………………..………151 Figura 74. Análisis de componentes principales del grosor de labio y longitud de heliconcha de caracoles S. gigas marcados en Brazo Norte y recapturados en uno de los 4 sitios de muestreo de la caleta de Xel-Há (Cueva, Centro, Bocana o Brazo Norte), entre enero 2005 y septiembre 2011………..……………………………………………………………………………………152 xxi 1. Introducción: El Caracol rosa Strombus gigas es un gasterópodo herbívoro comestible de gran tamaño (Abott, 1974), el cual representa uno de los recursos pesqueros de mayor importancia del Caribe (Brownell & Steveley, 1981; Chakalall & Cochrane, 1996) junto con la langosta espinosa (Theile, 2001). La creciente presión pesquera sobre las poblaciones rápidamente provocó la depleción del recurso en los años 1970s y 1980s (Stevely, 1979; Berg & Glazer, 1994; Stoner, 1996). En 1990 la especie fue incluida en el Apendicé II del Convenio Internacional de Comercialización de Especies en Peligro (CITES) y en 1994 fue clasificada como especie comercialmente amenazada por la Unión Internacional para la Conservación de la Naturaleza (IUCN) e incluida en la lista roja (Acosta, 2006). En México, había pesca de S. gigas en los estados de Quintana Roo y Yucatán. Tras la disminución de los volumenes de captura en 1988, se implementó una veda permanente para el estado de Yucatán, la cual sigue vigente hasta el día de hoy (Aldana-Aranda & Brulé-Desmarais, 1994; Aviso de Veda DOF 13 de febrero de 2009). En México la pesca de S. gigas se regula por la NOM-013-PESC (DOF, 21 de abril de 1995), cual entre otras dispociones establece una talla mínima de 200 mm. Como muchos de los demás organismos marinos, la especie está afectada por contaminación, disminución de la calidad del agua, desturbio y destrucción de su hábitat natural (Delgado et al., 2004) y muchas de las poblaciones no se han recuperado a pesar de las medidas de manejo, protección y rehabilitación (Glazer & Berg, 1994; Berg & Glazer, 1995; Stoner & Ray-Culp, 2000). El ciclo de vida del caracol rosa es complejo, presentando una etapa larvaria pelágica extensa, una etapa bentónica infaunal y una etapa epibentónica, lo que dificulta la evaluación de las poblaciones por métodos tradicionales empleados en la biología pesquera (Appeldoorn, 1987a). 1 Cada etapa del ciclo de vida está sujeta a distintos factores que influyen sobre su abundancia y distribución (CFMC QCSAMW, 1999). Se ha observado que el caracol rosa realiza diversas migraciones y se han descrito por lo menos dos: Migración de juveniles entre 50-100 mm (Stoner et al., 1988; De Jesús-Navarrete & Valencia-Beltrán, 2003; Danylchuck et al. 2003) y migraciones de organismos adultos (Robertson, 1959; Alcolado, 1976; Hesse, 1979; Appeldoorn, 1997). La ulterior fue asociada a la reproducción y se piensa que es de naturaleza estacional (Appeldoorn, 1997). La distribución de juveniles generalmente se encuentra restringida a ciertos sitios, por lo cual la mayoría de estudios presentan un sesgo de muestreo y abordan únicamente aspectos ecológicos de juveniles o aspectos ecológicos de adultos (Appeldoorn, 1987a), lo cual dificulta establecer relaciones en la conectividad entre juveniles y adultos. La talla de un organismo es un aspecto clave de su ecología, ya que determina su habilidad de explotar recursos y su posición en la cadena trófica y muchas especies realizan cambios en la utilización de su hábitat en función de su talla (Werner & Gilliam, 1984; Childress et al., 1994; Childress & Herrnkind, 2001; Bos et al., 2011). La caleta de Xel-Há ha sido utilizado desde 1995 como un parque ecoturístico bajo administración privada. Uno de los principales atractivos es la observación de la fauna marina en su hábitat natural, por lo que se prohíbe la extracción de cualquier organismo. La caleta se ha considerado como un santuario para el caracol rosa en la Riviera Maya, dado que representa un refugio importante para esta especie, encontrándose una población importante compuesta de organismos juveniles y adultos viejos (Baqueiro-Cárdenas & Aldana-Aranda, 2010). 2 El manejo sustentable, la rehabilitación y la protección de un recurso como S. gigas requieren de conocimientos biológicos y ecológicos. El estudio de la estructura y dinámica poblacional en el espacio y en el tiempo permiten entender como S. gigas hace uso de su hábitat y como se conectan las distintas etapas de su ciclo de vida. En el trabajo presente se estudiarán la estructura de tallas, la dinámica poblacional y la movilidad de la población epibentónica en el espacio y en el tiempo, mediante el método de capturamarcaje-recaptura y análisis de frecuencia de tallas, con la finalidad de determinar cómo S. gigas hace uso de su hábitat y cómo se conectan las distintas etapas de su ciclo de vida. 3 2. Antecedentes: 2.1 Taxonomía El caracol rosa es un invertebrado molusco, que pertenece a la clase de los gasterópodos, en el orden de los mesogasterópodos y es uno de 6 representantes de la familia Strombidae en el Atlántico Occidental (Abott, 1974). Los Strombidos representan un grupo monofilético, los cuales probablemente tienen su origen en la radiación del Indo Pacífico occidental (Latiolais et al., 2006; Kronenberg & Vermeij, 2002). La especie S. gigas fue descrita por Linnaeus en 1758 y una descripción más actual, la cual permite su identificación es dada por Abott (1974). Los Strombidos se caracterizan por poseer un pie estrecho con un opérculo terminal, el cual se asemeja a una uña. Tienen ojos pedunculados, una apertura larga y estrecha, un canal sifonal dentado hacía el final (muesca stromboide) y desarrollan un labio glaseado al alcanzar la madurez (Abott, 1974). La concha del caracol rosa S. gigas es de color pardo o beige en el exterior y de color rosa en el interior. Presentan 8 espinas en cada vuelta y los adultos pueden desarrollar entre 13 y 15 vueltas en su espira (Abott, 1974). 2.2 Distribución geográfica Su distribución geográfica se extiende desde el norte de las Bahamas a Florida y todo el Caribe hasta el norte de Sudamérica, incluyendo la costa de Brasil (Chakalall & Cochrane, 1996; Woods, 1995; Tewfik, 1996). Existen poblaciones aisladas alrededor de Bermudas (Theile, 2001) y en el Golfo de México (Aldana-Aranda & Brulé-Desmarais, 1994) (Fig. 1). 4 Figura 1. Distribución geográfica del caracol rosa Strombus gigas (azul turquesa). 2.3 Ciclo de vida y utilización del hábitat El caracol rosa tiene un ciclo de vida complejo con una fase larvaria extensa, una fase infaunal y una fase epibentónica de juvenil y de adulto (Fig. 2). 5 Figura 2. Ciclo de vida del caracol rosa S. gigas. Alcanza la madurez sexual a los 3.2 años y realiza la primera reproducción entre los 3.6 y 4 años (Appeldoorn, 1988). Crece rápidamente hasta alcanzar su longitud máxima de 24-29 cm. Después la concha inicia a formar un labio prominente y todo el crecimiento subsiguiente se manifiesta únicamente en el engrosamiento de la concha (Randall, 1964; Alcolado, 1976; Appeldoorn, 1994). El grosor del labio es un indicador de la edad y del estado de madurez. Appeldoorn (1988) y Aldana-Aranda & Frenkiel (2007) determinaron que el caracol está sexualmente maduro cuando presenta un grosor de labio mayor a 5 mm. Egan (1985) determinó el inicio de la reproducción a un grosor de labio de 4 mm, mientras Ávila-Poveda & BaqueiroCárdenas (2006) reportaron 13 mm como medida de madurez. La morfología, coloración y el tamaño de la concha pueden variar considerablemente, dependiendo de los factores ambientales 6 y genéticos (Randall, 1964; Alcolado, 1976; Appeldoorn, 1994), por lo cual la longitud de heliconcha no es un buen indicador del estado reproductor (Aldana-Aranda & Frenkiel, 2007). S. gigas es dioico y presenta fertilización interna. Las poblaciones en equilibrio mantienen una proporción de sexos de 1:1 (Randall, 1964; Davis et al., 1984). El número de huevos por puesta ha sido estimado entre 313,000 y 485,000 (Robertson, 1959; Randall, 1964; D’Asaro, 1965). Se ha observado que el caracol desova entre 4-8 veces en una temporada (Shawl & Davis, 2004). El caracol rosa deposita sus huevos de preferencia sobre un sustrato arenoso (Randall, 1964; D’Asaro, 1965; Pérez-Pérez & Aldana-Aranda, 2002; Pérez-Pérez & Aldana-Aranda, 2003; Glazer & Kidney, 2004), evitando pastos marinos (Glazer & Kidney, 2004). La estación reproductiva varía a través del Caribe, pero ocurre con mayor frecuencia durante los meses cálidos del año (Brownell, 1977; D'Asaro, 1965; Randall, 1964). Aldana-Aranda (2006) determinó mediante una revisión bibliográfica que hay reproducción del caracol en todas las localidades estudiadas del Caribe entre Junio y Agosto y en algunas localidades la reproducción parece ocurrir durante todo el año (Blakesley, 1977; Aldana-Aranda et al., 2003). La actividad reproductiva del caracol rosa se correlaciona positivamente con la temperatura (Stoner et al., 1992; Pérez-Pérez & Aldana-Aranda, 2002; Pérez-Pérez & Aldana-Aranda, 2003; Appeldoorn, 1990) y el fotoperiodo (Stoner et al., 1992; Pérez-Pérez & Aldana-Aranda, 2002; Pérez-Pérez & Aldana-Aranda, 2003). Las larvas de caracol eclosionan después de un mínimo de 100 horas de incubación (D’Asaro, 1964) y adoptan un estilo de vida pelágico planctotrófico. 7 La distribución y abundancia de las larvas depende de la estación del año (Chávez-Villegas et al., 2009) y las mayores abundancias se han observado en la estación de lluvias en Junio (Aldana-Aranda & Pérez-Pérez, 2007) y Agosto (De Jesús-Navarrete, 2001; Aldana-Aranda & Pérez-Pérez, 2007). Las corrientes han sido descritas como factor de dispersión y transporte de las larvas (Mitton et al., 1989; Stoner, et al. 1996; Stoner, 2003). Se piensa que las larvas se concentran sobre los principales ejes de los corrientes, creando los sitios de alta abundancia juvenil (“nursery grounds”) (Stoner, 2003). Dada la extensa fase planctónica (15-22 días), las larvas pueden ser transportadas por distancias considerables, hasta 900 km (Davis, et al., 1993). Después de 17-22 días en la columna de agua las larvas inician su asentamiento en el bentos y a los 28-33 días completan su metamorfosis (Brownell, 1977). El caracol es altamente específico en la selección de su hábitat durante esta etapa, requiere establecer el contacto con el fondo, por lo cual se asienta principalmente en sitios de baja profundidad (< 6 m) con alta abundancia de juveniles y responde a señales de calidad trófica del sitio, como la presencia de algas rojas, con el inicio de la metamorfosis (Stoner, 2003). El caracol recién reclutado tiene un tamaño de aproximadamente 2 mm y una concha blanca (Brownell, 1977). Después de su metamorfosis inicia una etapa infaunal, enterrándose en el sedimento donde permanece por aproximadamente un año (Ray & Stoner, 1994). Después de su reclutamiento al bentos los caracoles siguen haciendo decisiones relacionados con su hábitat los cuales están asociados a su sobrevivencia y crecimiento (Stoner, 2003). Los caracoles post-asentamiento muestran preferencias por sustratos de arenas medias y gruesas (Iversen et al., 1987; Iversen et al., 1994; De Jesús-Navarrete & Valencia-Beltrán, 2003), fragmentos de coral y sedimento no consolidado con alto contenido de materia orgánica (De 8 Jesús-Navarrete & Valencia-Beltrán, 2003). Más allá, se han encontrado mayores abundancias en las depresiones de arena (Iversen et al., 1987; Iversen et al., 1994). Los caracoles juveniles son altamente vulnerables a la predación (Iversen et al., 1987; Iversen et al., 1994; Ray & Stoner, 1994; Ray-Culp et al., 1999), la cual puede causar la mortalidad del 100% de los organismos con talla entre 20-40 mm, bajo condiciones experimentales (Ray et al, 1994). Cuando alcanzan una longitud de concha de 50-100 mm, los juveniles empiezan a surgir del sedimento y adoptan un estilo de vida epibentónico (Stoner & Ray, 1993). Se determinó de forma experimental que la mortalidad por predación disminuye en un 90% en los juveniles mayores a 50 mm longitud de heliconcha (Ray-Culp et al., 1999). Cuando el caracol alcanza tallas de 170 mm empieza a mostrar preferencias por praderas de pastos marinos (Randall, 1964; Stoner & Ray, 1993; Stoner et al., 1995; Stoner & Waite, 1990) de Thalassia testudinium (DíazAvalos, 1991) o Syringodium sp. (Hesse, 1979), así como por planicies de macroalgas (Danylchuck et al., 2003), pero también se ha observado sobre arenales con fragmentos de coral (De Jesús-Navarrete, 2000). Stoner (1989) determinó que la dieta del caracol juvenil se basa principalmente en detritos. Los caracoles juveniles generalmente se hallan asociados a zonas de baja profundidad (Iversen et al., 1987; Stoner & Schwarte, 1994) y su abundancia disminuye inversamente con ésta y se acerca a nula a profundidades mayores a 15 metros (Stoner & Schwarte, 1994). Al alcanzar mayor tamaño los caracoles empiezan a migrar hacía mayores profundidades (Appeldoorn, 1997). Wicklund et al. (1991) reporta que la mayor abundancia de adultos se halla a profundidades mayores de 6 metros, mientras Stoner & Schwarte (1994) detectan el 74% de los 9 organismos adultos entre los 10-18 metros. En zonas donde la profundidad es uniforme se ha observado que se alejan de la costa (Hesse, 1979). Los caracoles adultos muestran preferencia por los sustratos arena, arena gruesa, fragmentos de coral, escombro y sustratos rocosos en etapas de alimentación (Stoner & Sandt, 1992; Glazer & Kidney, 2004). No se han encontrado diferencias entre la dieta de juveniles y adultos (Stoner & Waite, 1991). En cuanto a su longevidad existen divergencias y se piensa que varía entre 5 y 20 años (Berg, 1976; Wefer & Killingley, 1980; Appeldoorn, 1988; Twefik et al., 1998). 2.4 Estudios de parámetros de Población Una población se define como un grupo de organismos que ocupan un espacio dado en un momento específico (Krebs, 1985). La población tiene diversas características de grupo y se distinguen tres tipos: Fundamentales, dinámicos y secundarios (Krebs, 1985). Entre las características fundamentales es el tamaño de población o densidad. Como características dinámicas se definen aquellas que influyen sobre el tamaño de población y son la natalidad o reclutamiento, la mortalidad, la inmigración y la emigración. Como características secundarias es posible delinear la distribución de edades, composición genética y patrón de distribución de los individuos en el espacio (Krebs, 1985). La evaluación del crecimiento es un elemento esencial en el estudio de las poblaciones de importancia pesquera, ya que provee información sobre la longevidad, distribución de edades y mortalidad (Eberhardt & Valle-Esquivel, 2008). En la actualidad existen más de 230 artículos científicos relacionados con S. gigas (Stoner, 1997). 10 Las publicaciones sobre esta especie comenzaron en los 1960s y se relacionan principalmente con la ecología y biología de esta especie. Dentro de los estudios poblacionales más transcendentes y completos realizados en el área del Caribe se halla el trabajo de Randall (1964). Éste trabajo cubre una revisión y el estudio de los siguientes aspectos de la ecología y biología de S. gigas: distribución de tallas, dimorfismo sexual, proporción de sexos, hábitat, crecimiento, movimientos, hábitos alimenticios, reproducción, desarrollo, predación y comensalismo. Robertson (1959) estudio los hábitos alimenticios del caracol. Robertson (1959) y D’Asaro (1965) contribuyeron en mayor grado al conocimiento la reproducción y el desarrollo de S. gigas. Berg (1975; 1976) estudió aspectos del comportamiento, crecimiento y longevidad. Hesse (1979) contribuyó significativamente al estudio de movilidad y migración, estudiando poblaciones en las islas de Turks & Caicos. En los años 1980s se intensifica el interés científico en el caracol rosa, junto con su demanda comercial. Uno de los trabajos más completos de ésta época es el trabajo de Iversen et al., (1987), realizado en las Bahamas, el cual cubre: Distribución de caracoles del primer año, migración y movimientos, densidad, crecimiento, sobrevivencia y mortalidad y otros aspectos de importancia pesquera. Appeldoorn (1987a) contribuye significativamente al desarrollo de los modelos de crecimiento y los métodos numéricos en el estudio del caracol rosa. Mitton et al. (1989) estudiaron conectividad genética de las poblaciones del Caribe, determinando que las poblaciones del caracol no son panmixticas y que existe aislamiento genético local. 11 A partir de los 1990s múltiples aspectos de la ecología de S. gigas han sido tratados por A.W. Stoner y sus colaboradores, entre estos migración (Stoner et al., 1988; Stoner, 1989; Stoner et al., 1992), distribución (Stoner & Schwarte, 1994; Stoner & Lally, 1994), alimentación (Stoner & Waite, 1991), predación (Ray-Culp et al., 1999), densidad y crecimiento (Ray & Stoner, 1994). Entre los trabajos más recientes destacan los estudios de conectividad de las poblaciones en el Caribe (Paris et al., 2008) y utilización del hábitat (Glazer et al., 2003; Doerr & Hill, 2007; Bissada-Gooding & Oxenford, 2010; Phillips et al., 2010). En México se realizaron varios estudios de población a partir de los años 1980s (Tabla 1). Las principales líneas de investigación han sido: Estructura poblacional, reclutamiento, mortalidad, crecimiento, reproducción, abundancia y distribución larval. 12 Tabla 1. Estudios de Población del caracol rosa S. gigas realizados en México. Autor Año Objetivo De la Torre Díaz Avalos Díaz Avalos De Jesús Navarrete et al. 1982 1986 1991 1992 Diagnóstico del recurso Crecimiento y mortalidad Tamaño y estructura de población Distribución juveniles y adultos QR QR QR QR QR P. Gavilán P. Gavilán P. Gavilán Chávez & Arreguín-Sánchez 1994 Dinámica poblacional QR Chinchorro De Jesús Navarrete et al. Domínguez-Viveros et al. 1994 1994 Crecimiento, reclutamiento y estructura poblacional Dinámica poblacional QR QR Chinchorro, P. Gavilán Chinchorro De Jesús Navarrete & Oliva Riviera 1997 QR P. Gavilán De Jesús Navarrete et al. 1999 Estructura, evaluación poblacional, juveniles Crecimiento, reclutamiento y estructura poblacional QR Domínguez-Viveros et al. 1999 Estado de poblaciones QR De Jesús Navarrete 2000 QR De Jesús Navarrete et al. De Jesús-Navarrete & Aldana-Aranda Oliva-Rivera & De Jesús-Navarrete Pérez-Pérez & Aldana-Aranda De Jesús Navarrete Aldana-Aranda et al. De Jesús Navarrete & Valencia-Beltrán 2000 Crecimiento en 4 ambientes Crecimiento, mortalidad y reclutamiento P. Gavilán Chinchorro, P. Gavilán Chinchorro, P. Gavilán QR P. Gavilán 2000 Distribución y abundancia larval QR Chinchorro 2000 Distribución y abundancia larval Distribución, abundancia, morfometría Distribución y abundancia larval Periodo de reproducción QR Chinchorro Yuc QR QR Alacranes Chinchorro Chinchorro QR Chinchorro De Jesús Navarrete et al. Pérez-Pérez & Aldana-Aranda 2003 Abundancia caracol primer año Estructura poblacional, cambios a través del tiempo QR Chinchorro 2003 Actividad reproductiva Yuc Alacranes Tello et al. Aldana-Aranda & PérezPérez De Jesús Navarrete & Pérez-Flores Delgado et al. Paris et al. Bravo-Castro et al. Chávez-Villegas et al. 2005 Genética poblacional Yuc+QR Península 2007 Abundancia y distribución larval Yuc Alacranes 2007 2008 2008 2010 2010 Distribución y abundancia larval Conectividad de poblaciones Conectividad de poblaciones Abundancia y distribución larval Abundancia larvas QR QR+ QR + Yuc QR QR + 2000 2001 2003 2003 Estado Zona B. Asención México+ Península QR SAM La población de caracol rosa en Xel-Há ha sido estudiada y monitoreada desde Noviembre 2001 hasta la actualidad. Previo al año 2001 se realizó un trabajo de tesis sobre el caracol rosa entre 13 los años 1990-1991 y se encontró una publicación correspondiente (Valle-Esquivel, 1998). Para la población de S. gigas de la caleta de Xel-Há existen estudios sobre crecimiento de la especie (Valle-Esquivel, 1998; Aldana-Aranda et al., 2003; Aldana-Aranda et al., 2005; BaqueiroCárdenas & Aldana-Aranda, 2010), estructura poblacional (Aldana-Aranda et al., 2003; AldanaAranda et al., 2005; Baqueiro-Cárdenas & Aldana-Aranda, 2010), tamaño de población (AldanaAranda et al., 2005; Baqueiro-Cárdenas & Aldana-Aranda, 2010), reclutamiento y mortalidad (Baqueiro-Cárdenas & Aldana-Aranda, 2010) (Tabla 2). Tabla 2. Estudios de Población del caracol rosa S. gigas realizados en la Caleta de Xel-Há, México. Autor Valle-Esquivel Año 1998 Periodo del Estudio Nov 1990-Jul 1991 Objetivos Crecimiento Aldana-Aranda et al. 2003 Nov 2001- Sep 2002 Estructura poblacional, crecimiento Aldana-Aranda et al. 2005 Nov 2001- Ago 2003 Estructura poblacional, Tamaño de población, crecimiento Baqueiro-Cárdenas & Aldana-Aranda 2010 Nov 2001-Ago 2005 Tamaño de población, estructura poblacional, crecimiento, reclutamiento, mortalidad Existe una variedad de métodos para estudiar el movimiento y la conectividad entre agregaciones juveniles y adultos (Bronwyn et al., 2003). En el estudio del caracol rosa éstos generalmente incluyen la documentación de cambios o diferencias en la estructura de tallas y abundancia en el tiempo o espacio (Randall, 1964; Alcolado 1976, Stoner et al., 1988; Stoner, 1989; Wicklund et al., 1991; Stoner & Lally, 1994), marcaje (Alcolado, 1976; Hesse, 1979; Stoner & Sandt, 1992; Stoner & Ray, 1993; Bissada-Gooding & Oxenford, 2010; Phillips et al., 2010), y rastreo por telemetría acústica (Glazer et al., 2003; Doerr & Hill, 2007; Bissada-Gooding & Oxenford, 2010; Phillips et al., 2010) (Tabla 3). 14 Randall (1964) fue uno de los primeros autores en describir un patrón de movilidad. Alcolado (1976) detectó segregación de juveniles y adultos en el espacio. Los patrones de migración por talla y el rango hogareño fueron descritos formalmente por primera vez por Hesse (1979). En los 1990s Stoner y sus colaboradores estudiaron la formación y movilidad de agregaciones juveniles. En el siglo XXI inició el estudio de los patrones de migración y utilización del hábitat por el método de telemetría acústica (Glazer et al., 2003; Bissada-Gooding & Oxenford, 2010; Phillips et al., 2010). Sin embargo, los autores únicamente abordan aspectos ecológicos de juveniles (Stoner et al., 1988; Stoner, 1989; Phillips et al., 2010) o de adultos (Glazer et al., 2003; Bissada-Gooding & Oxenford, 2010) y ningún trabajo aborda la transición entre las zonas de crianza y la distribución de adultos. 15 Tabla 3. Estudios de movimiento y migración del caracol rosa S. gigas realizados en el Caribe. Autor Año Sitio Método Campo Método estadístico Randall 1964 Islas Vírgenes Abundancia por talla sobre diferentes sustratos; Marcaje y seguimiento Talla media y sustrato Tasa de progresión (m/día) Porcentaje, talla y dirección de movimientos Hesse 1979 Turks & Caicos Marcaje con boya y posicionamiento de S. gigas sobre una cuadrícula Iversen et al. 1986 Bahamas Marcaje y recaptura, posicionamiento por tallas en diferentes ambientes Distribución de tallas por sitio Stoner et al. 1988 Bahamas Estacas en los límites de la agregaciones, medición de la tasa de progresión y dirección Mapa de la progresión, progresión (m/día), densidad Longitud y ancho de la agregación, tasa de progresión (m), dirección (°) Stoner 1989 Bahamas Densidad sobre cuadrantes y circulo, estacas en los límites de la agregaciones, medición de dirección y la tasa de progresión Wicklund et al. 1991 Bahamas Conteo de caracoles juveniles sobre cuadrícula Mapa de distribución Stoner & Sandt 1992 Bahamas Marcaje, recaptura diferentes ambientes (adultos) Abundancia sobre distintos sustratos por mes Stoner & Ray 1993 Bahamas Marcaje y posicionamiento de S. gigas en un círculo, dirección y tasa de progresión Densidad por zona y por mes Mapa de progresión, longitud y ancho de progresión, velocidad de progresión, densidad Stoner & Lally 1994 Bahamas Estacas en los límites de la agregaciones, medición de la tasa de progresión y dirección, densidad Glazer et al. 2003 Estados Unidos Telemetría acústica Velocidad y estacionalidad de movimientos, rangos hogareño (ha) Doerr & Hill 2007 Islas Vírgenes Telemetría acústica Movilidad juveniles y adultos BissadaGooding & Oxenford 2010 Barbados Telemetría acústica y marcaje tradicional Rango hogareño de adultos, tasa de movimiento/mes, fieldad de sitio, Phillips et al. 2010 Barbados Telemetría acústica y marcaje tradicional Rango hogareño de juveniles, tasa de movimiento/mes 16 El crecimiento del caracol rosa se ha evaluado tanto por métodos directos como indirectos. Tasas de crecimiento obtenidos de métodos directos son el resultado de estudios de Captura-MarcajeRecaptura (CMR) o se obtienen en encierros y consisten en la medición del aumento de talla por unidad de tiempo (Appeldoorn, 1987a). En la tabla 4 se muestran algunos autores que han estudiado el crecimiento del caracol mediante métodos directos. Tabla 4. Estudios de crecimiento de S. gigas mediante métodos directos. Año Autor Sitio 1964 Randall Virgin Islands, USA Alcolado Brownell Gibson et al. Weil & Laughlin Ray & Stoner De Jesús-Navarrete & Oliva-Rivera 1976 1977 1983 1984 1994 Encierro Cuba Encierro, diferentes ambientes Florida Keys, USA Encierro Belice CMR Venezuela CMR Exuma Cays, Bahamas Encierro Punta Gavilán, México CMR Banco Chinchorro, México Encierro, diferentes ambientes Punta Gavilán, México Encierro, diferentes ambientes México Laboratorio, dieta artificial 1997 2001 De Jesús-Navarrete 2002 De Jesús-Navarrete Moreno de la Torre & Aldana-Aranda Método 2005 Los métodos indirectos se refieren a aquellos métodos basados en el análisis de frecuencias de longitud y progresión de moda (Eberhardt & Valle-Esquivel, 2008). Requieren de muestras grandes para arrojar resultados fiables (Appeldoorn, 1987a). Existen diferentes modelos para estimar el crecimiento mediante los métodos de análisis de frecuencia de tallas, revisados por Pauly & Morgan (1987). 17 En el estudio de S. gigas el método más utilizado es la estimación de los parámetros de crecimiento a través de la ecuación de crecimiento de von Bertalanffy (VBGF), los cuales se pueden obtener tanto por análisis de frecuencia de tallas o a partir de incrementos de marcajerecaptura (Tabla 5). Tabla 5. Estudios de crecimiento de S. gigas mediante análisis la estimación de los parámetros de la ecuación de von Bertalanffy. Autor Alcolado Berg Berg Iversen et al. Strasdine Strasdine Strasdine Appeldoorn, Año 1976 1976 1976 1987 1988 1988 1988 1990 Localidad Cuba, varios sitios St. John, Islas Vírgenes St. Croix, Islas Vírgenes Berry Island, Bahamas Boca Chica, Belice Tres Cocos, Belice Water Caye, Belice La Parguera, Puerto Rico García 1991 San Andrés & Providencia, Colombia Marquéz 1993 Providencia & Santa Catalina, Columbia Rathier & Battaglya De Jesús-Navarrete et al. 1994 1995 Martinique Punta Gavilán, México Gallo et al. 1996 San Andrés & Providencia, Colombia Twefik De Jesús-Navarrete et al. 1996 2000 2001 Pedro Bank, Jamaica Punta Gavilán, México Punta Gavilán, México De Jesús-Navarrete Una revisión exhausta y discusión de los diferentes métodos y herramientas para el estudio de los parámetros poblacionales de S. gigas es dado por Eberhardt & Valle-Esquivel (2008). 18 3. Problema central a Investigar: En base de una revisión exhausta sobre la ecología del caracol rosa, se fijaron 2 áreas centrales de investigación: Dónde se estudiarán: La demografía (estructura, abundancia, tamaño poblacional, densidad de organismos) en el espacio y tiempo. La dinámica poblacional (reclutamiento, sobrevivencia, mortalidad, crecimiento) en el espacio y tiempo. La movilidad y utilización espacial del área de estudio en función a la talla de los organismos. 4. Hipótesis 4.1 La distribución de tallas muestra diferencias en el espacio. 4.2 Los organismos realizan transiciones en su hábitat los cuales dependen de su talla. 5. Objetivos 5.1 Objetivo General Estudiar la estructura, dinámica y movilidad de la población epibentónica en el espacio y en el tiempo, con la finalidad de determinar cómo S. gigas hace uso de su hábitat en las distintas etapas de su ciclo de vida. 19 5.2 Objetivos Particulares Determinar la abundancia, el tamaño y densidad de la población, así como la estructura de tallas en el espacio y tiempo, mediante el análisis de los datos de marcaje-recaptura. Determinar la tasa de crecimiento por clases de tallas del caracol rosa en Xel-Há, mediante el análisis de las frecuencias de talla y los datos de marcaje-recaptura. Determinar el patrón de reclutamiento, la mortalidad y sobrevivencia mediante el análisis de las frecuencias de talla y los datos de marcaje-recaptura. Identificar un patrón y una dirección de los movimientos entre las diferentes zonas de muestreo de la Caleta de Xel-Há, mediante el análisis de los datos de marcaje-recaptura. 6. Justificación El caracol rosa S. gigas es una especie amenazada por la pesca, pérdida y destrucción de su hábitat natural. Su manejo sustentable, su protección y rehabilitación requieren de conocimientos biológicos y ecológicos. La especie tiene un ciclo de vida complejo y aún no se entiende en su totalidad como las distintas etapas de éste se conectan en el espacio. El estudio de la estructura de tallas y la dinámica poblacional en el espacio y en el tiempo permiten entender como S. gigas hace uso de su hábitat y como se conectan las distintas etapas de su ciclo de vida en este. La caleta de Xel-Há es un santuario para esta especie donde cumple todo su ciclo de vida bajo condiciones naturales. 20 7. Materiales y Métodos 7.1 Área de Estudio Xel-Há se localiza en la costa Este de la península de Yucatán entre los paralelos 20°19'15'' y 20°18'50'' de latitud Norte y los meridianos 87°21'41'' y 87°21'15'' de longitud Oeste, a 16 Km al Noreste de la ciudad de Tulum en el Estado de Quintana Roo, México (Fig. 3). La caleta de Xel-Há es un cuerpo de agua constituido por la mezcla de agua subterránea con agua de mar con una superficie total de 14 ha, conectado directamente al mar Caribe a través de un canal natural que tiene un ancho promedio de 90 m y una longitud de 170 m, a través del cual se realiza el intercambio de agua del sistema, debido al efecto de las mareas. Presenta una zona centro y 2 apéndices: Brazo Norte y Brazo Sur. Su profundidad varía entre 0.5 m y 4.0 m. La caleta Xel-Há está circundada por manglar y asociada al sistema arrecifal mesoamericano. La región se encuentra bajo un régimen climático de tipo sub-húmedo con lluvias aisladas en verano e invierno. Conforme a los registros existentes de la estación meteorológica de Tulum, que es la más cercana al sitio, se tiene una temperatura media anual de 26°C, una precipitación media anual del orden de los 1,079 mm, con lluvias máximas en el verano y mínimas durante los meses de marzo y abril, siendo octubre el mes más lluvioso, con 178.9 mm y abril el menos lluvioso con 31.1 mm. El 71% de la lluvia anual ocurre en los meses de junio a noviembre, en tanto que el 29% restante se presenta en los meses de diciembre a mayo. La evaporación media anual es del orden de los 1,392 mm (Organismo de Cuenca Península de Yucatán Dirección Técnica, 2008). Los Muestreos se realizaron en 4 sitios de la Caleta: Cueva, Brazo Norte, Centro y Bocana (Fig. 3). 21 Figura 3. Ubicación geográfica del área de estudio, la Caleta de Xel-Há y de los sitios de Muestreo: Cueva, Brazo Norte, Centro y Bocana. El sitio de muestreo "Cueva" se encuentra en el brazo sur de la caleta e incluye una pequeña bahía rodeada de manglares (Rhizophora mangle). Tiene una superficie total de 6,000 m2. El aporte persistente de agua dulce y fría desde cuevas subterráneas mantiene una termo- y haloclina a una profundidad aproximada de 1.25 m de la superficie y condiciones marinas en el fondo (Organismo de Cuenca Península de Yucatán Dirección Técnica, 2008). El sitio tiene una profundidad de 1.5 a 3.5 m (Stieglitz et al., 2011). El fondo está compuesto por lodos y arenas finas, fragmentos de algas coralinas (Halimeda sp.) y rocas. Se hallan aisladamente densos parches de distintas macroalgas (Padina sp., Halimeda sp., Penicillus sp. Amphiroa sp., Acanthophora sp., Caulerpa sp., Dictyota sp.) y medusas invertidas (Cassiopea sp.). En las orillas el fondo está cubierto por hojas de mangle en descomposición. 22 En las figuras 4.A a 4.D se muestra la variación temporal de la salinidad, temperatura, oxígeno disuelto y pH, en el fondo y la superficie de la columna de agua del sitio Cueva, durante el periodo de enero 2005 a septiembre 2011. Los datos fueron proporcionados por el parque Xel-Há y obtenidos por el Laboratorio de Análisis de Calidad del Agua y Medio Ambiente S.A. de C.V. (Lab-ACAMA), utilizando las metodologías establecidas por las normas oficiales mexicanas (NOM) vigentes. La salinidad en el fondo fluctuó entre 10 y 32. La variación de la salinidad en superficie fue mayor con salinidades entre 2 y 30. La temperatura varió entre 25 y 30 °C y fue similar en el fondo y la superficie. La concentración de oxígeno fue similar en fondo y superficie y fluctuó entre 1.5-8 mg/l a lo largo del periodo de estudio. El pH mostró variaciones entre 2005 y 2009 con un rango de 6.5-7.8, pero fue constante a partir del año 2010 y neutro. 23 Figura 4. Variación temporal de (A) la salinidad, (B) temperatura (°C), (C) oxígeno disuelto (mg/l) y (D) pH, en el fondo y la superficie de la columna de agua del sitio Cueva de la caleta de Xel-Há, durante el periodo de enero 2005 a septiembre 2011. 24 En las figuras 5.A a 5.C se muestra la variación temporal de nitritos, nitratos y fósforo total, en el fondo y la superficie de la columna de agua del sitio Cueva, durante el periodo de enero 2005 a septiembre 2011. Los datos fueron proporcionados por el parque Xel-Há y obtenidos por LabACAMA, utilizando las metodologías establecidas por la NOM. Se observó un pico en la concentración de nitritos entre octubre 2006 y mayo 2007 en el fondo. El tiempo restante las concentraciones de nitritos fueron bajas y constantes. Las concentraciones de nitratos y fósforo total fueron similares en el fondo y la superficie. Se observó un pico en la concentración de nitratos en agosto 2007. La concentración de fósforo total tendió a incrementar a partir de marzo 2010. 25 Figura 5. Variación temporal de (A) Nitritos (mg/l), (B) Nitratos (mg/l) (°C) y (C) Fósforo total (mg/l), en el fondo y la superficie de la columna de agua del sitio Cueva de la caleta de Xel-Há, durante el periodo de enero 2005 a septiembre 2011. El sitio de muestreo “Brazo Norte” se encuentra en el apéndice norte de la Caleta. La superficie muestreada fue de de 6.000 m². La profundidad del sitio varía entre 0.5 m y 3.4 m (Stieglitz et al., 2011). El fondo se compone de lodo fino, grandes rocas, los cuales forman varios canales y 26 presenta cobertura densa de macroalgas. Durante el periodo de estudio se hallaron muchas medusas invertidas (Cassiopea sp.). La columna de agua es menos estratificada que en la Cueva pero también presenta un halo- y termoclina cerca de la superficie y condiciones marinas en el fondo (Organismo de Cuenca Península de Yucatán Dirección Técnica, 2008). En las figuras 6.A a 6.D se muestra la variación temporal de la salinidad, temperatura, oxígeno disuelto y pH, en el fondo y la superficie de la columna de agua del sitio Brazo Norte, durante el periodo de enero 2005 a septiembre 2011. Los datos fueron proporcionados por el parque Xel-Há y obtenidos por Lab-ACAMA, utilizando las metodologías establecidas por la NOM. La salinidad en el fondo generalmente fue mayor que en la superficie y fluctuó entre 24 y 35. En la superficie se observaron salinidades entre 11 y 40. La temperatura tuvo una variación entre 24 y 31 °C y fue similar en el fondo y la superficie. La concentración de oxígeno fue similar en fondo y superficie y fluctuó entre 2.5-8 mg/l a lo largo del periodo de estudio. El pH tuvo una variación entre 6.8 y 8.2 a lo largo del estudio. 27 Figura 6. Variación temporal de (A) la salinidad, (B) temperatura (°C), (C) oxígeno disuelto (mg/l) y (D) pH, en el fondo y la superficie de la columna de agua del sitio Brazo Norte de la caleta de Xel-Há, durante el periodo de enero 2005 a septiembre 2011. 28 En las figuras 7.A a 7.C se muestra la variación temporal de nitritos, nitratos y fósforo total, en el fondo y la superficie de la columna de agua del sitio Brazo Norte, durante el periodo de enero 2005 a septiembre 2011. Los datos fueron proporcionados por el parque Xel-Há y obtenidos por Lab-ACAMA, utilizando las metodologías establecidas por la NOM. Se observó un pico en la concentración de nitritos en abril 2006 en la superficie. El tiempo restante las concentraciones de nitritos fueron bajas y constantes. Las concentraciones de nitratos y fósforo total fueron similares en el fondo y la superficie. Se observó un pico en la concentración de nitratos en agosto 2007. La concentración de fósforo total tuvo picos en marzo 2010 y septiembre 2010. 29 Figura 7. Variación temporal de (A) Nitritos (mg/l), (B) Nitratos (mg/l) (°C) y (C) Fósforo total (mg/l), en el fondo y la superficie de la columna de agua del sitio Brazo Norte de la caleta de Xel-Há, durante el periodo de enero 2005 a septiembre 2011. El sitio de muestreo Centro tiene una superficie total de 23,000 m2 y profundidades entre 1.5 m a 2.5 m (Stieglitz et al., 2011). La columna de agua es fuertemente estratificada (Organismo de 30 Cuenca Península de Yucatán Dirección Técnica, 2008). Comprende una planicie extensa de arena y la parte central de ésta es cubierta por pastos marinos (Halodule sp.). Se hallan en forma aislada rocas grandes que sobresalen a la superficie. En las orillas se presentan zonas rocosas con parches aislados de diversas macroalgas. En las figuras 8.A a 8.D se muestra la variación temporal de la salinidad, temperatura, oxígeno disuelto y pH, la superficie de la columna de agua del sitio Centro, durante el periodo de mayo 2007 a septiembre 2011. Sólo se registraron datos de la condición del fondo en mayo 2007. Los datos fueron proporcionados por el parque Xel-Há y obtenidos por Lab-ACAMA, utilizando las metodologías establecidas por la NOM. La salinidad en superficie varió entre 14 y 26. La temperatura fluctuó entre 24 °C y 31 °C a través del periodo estudiado. La concentración más alta de oxígeno disuelto se observó en mayo 2007 con 7 mg/l y la más baja en abril 2008 con 3.8 mg/l. El pH varía entre 7 y 7.8. Entre marzo 2010 y septiembre 2011 el pH fue constante en 7.3. En las figuras 9.A a 9.C se muestra la variación temporal de nitritos, nitratos y fósforo total, en la superficie de la columna de agua del sitio Centro, durante el periodo de mayo 2007 a septiembre 2011. Sólo se registraron parámetros en el fondo en mayo 2007. Los datos fueron proporcionados por el parque Xel-Há. La concentración de nitritos fue baja y mostró variaciones de 0 - 0.02 mg/l a lo largo del periodo estudiado. La concentración de nitratos máxima se observó en diciembre 2009 y fue de 1.5 mg/l. Se observó un pico en la concentración de fósforo total en marzo 2010. 31 Figura 8. Variación temporal de (A) la salinidad, (B) temperatura (°C), (C) oxígeno disuelto (mg/l) y (D) pH, en el fondo y la superficie de la columna de agua del sitio Centro de la caleta de Xel-Há, durante el periodo de mayo 2007 a septiembre 2011. 32 Figura 9. Variación temporal de (A) Nitritos (mg/l), (B) Nitratos (mg/l) (°C) y (C) Fósforo total (mg/l), en el fondo y la superficie de la columna de agua del sitio Centro de la caleta de Xel-Há, durante el periodo de mayo 2007 a septiembre 2011. El área de muestreo Bocana comprende la parte de la caleta adyacente al canal que conecta esta al mar y los primeros 30 m del canal. La superficie muestreada fue de 10,000 m2. La profundidad 33 varía entre 0.5 m y 3.4 m (Stieglitz et al., 2011). El fondo está cubierto por arena, rocas pequeñas y fragmentos de coral. Se hallan pequeñas extensiones aisladas de pastos marinos (Thalassia sp.) y algunas cabezas de coral. La zona tiene mayor influencia marina y la termo- y haloclina se encuentra a menos de 1 m de profundidad (Organismo de Cuenca Península de Yucatán Dirección Técnica, 2008). En las figuras 10.A a 10.D se muestra la variación temporal de la salinidad, temperatura, oxígeno disuelto y pH, en el fondo y la superficie de la columna de agua del sitio Bocana, durante el periodo de enero 2005 a septiembre 2011. Los datos fueron proporcionados por el parque Xel-Há y obtenidos por Lab-ACAMA, utilizando las metodologías establecidas por la NOM. La salinidad registrada en el fondo exhibió fluctuaciones entre 25 y 40. En la superficie se obtuvieron salinidades entre 12 y 45. La temperatura mínima registrada fue de 25 °C y la máxima de 31 °C. En el fondo se observaron concentraciones de oxígeno disuelto con un rango entre 3.9 mg/l a 8mg/l. En la superficie el valor más bajo registrado fue de 2.9 mg/l y el más alto fue de 6.9 mg/l. El pH en el fondo fue mayor en el periodo estudiado en comparación con los valores registrados en superficie. 34 Figura 10. Variación temporal de (A) la salinidad, (B) temperatura (°C), (C) oxígeno disuelto (mg/l) y (D) pH, en el fondo y la superficie de la columna de agua del sitio Bocana de la caleta de Xel-Há, durante el periodo de enero 2005 a septiembre 2011. 35 Figura 11. Variación temporal de (A) Nitritos (mg/l), (B) Nitratos (mg/l) (°C) y (C) Fósforo total (mg/l), en el fondo y la superficie de la columna de agua del sitio Bocana de la caleta de Xel-Há, durante el periodo de enero 2005 a septiembre 2011. En la tabla 6 se muestran los resultados de un estudio granulométrico de la caleta de Xel-Há y en la figura 12 se muestra la distribución de los sedimentos en el espacio. Los datos fueron proporcionados por el parque Xel-Há. Se observó un gradiente en la composición del fondo, 36 donde se encontraron principalmente sedimentos arenosos en el área de Bocana con cero limos. En Centro se reportaron sedimentos del tipo areno-fangoso y fango-arenoso. Se observó que la cantidad de limos aumenta hacía el interior de la Caleta, mientras disminuye la proporción de arena en el sedimento. En el Brazo Norte se hallaron sedimentos del tipo fango-arenosos y arenofangosos con alto contenido de limo. Tabla 6. Granulometría de la caleta de Xel-Há (2008). Ubicación % Arcilla % Limo % Arena Clasificación Clave 1 2 3 4 5 6 7 8 9 10 11 12 13 14 7.08 7.08 9.08 7.08 7.08 7.08 1.36 5.08 3.08 7.08 11.08 7.08 7.08 12.36 0 0 0 0 4 14 40 37 39 17 31 6.7 13 36 92.92 92.92 90.92 92.92 88.92 78.92 58.2 58.2 58.2 76.2 58.2 86.2 80.2 51.48 Arena Arena Arena Arena Areno fangoso Areno fangoso Fango arenoso Fango arenoso Fango arenoso Fango arenoso Fango arenoso Areno fangoso Areno fangoso Fango A A A A AF AF FA FA FA FA FA AF AF FA 15 7.08 51 41.48 Fango con apariencia de sedimento F 37 Figura 12. Granulometría de la caleta de Xel-Há. 7.2 Métodos de Campo De enero 2005 a enero 2008 se realizaron 23 muestreos de captura-marcaje-recaptura en los sitios Bocana, Centro y Cueva. De abril 2009 a septiembre 2011 se realizaron 13 muestreos, aplicando el mismo método, en los sitios Bocana, Centro, Cueva y adicionalmente Brazo Norte. Durante el año 2005 los muestreos fueron mensuales y bimensuales de 2006 a 2011. La superficie muestreada fue de 10,000 m² en la Bocana, de 23,000 m² en el Centro y de 6,000 m² en la Cueva y Brazo Norte, respectivamente. Se colectaron todos los organismos posibles en 3 horas, por 3 buzos en buceo libre, en cada uno de los sitios. 38 Para su identificación, todos los individuos fueron marcados con una marca plástica, elaborada de cinta de impresión Dymo ®, con un número de identificación único. La marca se fijó a la espira del caracol con un cincho de plástico (Fig. 13). En cada muestreo se cuantificó el número de individuos marcados y no marcados y posteriormente se marcaron todos aquellos individuos sin marca. Se determinó para cada individuo la longitud de la heliconcha (Fig. 14) y el grosor de labio en los organismos que lo presentaban (Fig. 15), utilizando un vernier con una precisión de 1mm. Posteriormente todos los organismos fueron liberados en la zona donde se encontraron. Para minimizar el riesgo de perder marcas, se reemplazaron todas las marcas y cinchos con señales de deterioro por marcas nuevas idénticas. Figura 13. Método de marcaje del caracol rosa S. gigas. 39 Figura 14. Método para la medición de longitud de heliconcha del caracol rosa S. gigas. Figura 15. Método de medición del grosor de labio del caracol rosa S. gigas. 40 7.3 Métodos de análisis de datos 7.3.1 Abundancia, Tamaño de Población y densidad Se cuantificó el número de individuos marcados en cada uno de los cuatro sitios estudiados de la caleta de Xel-Há para cada mes de muestreo. Se utilizaron diagramas de barra para mostrar los resultados. Se cuantificó el número de caracoles marcados en la totalidad de la caleta de Xel-Há, para cada mes, sumando el número de individuos marcados en los sitios Cueva, Centro y Bocana en el periodo 2005-2008 y el número de individuos marcados en los sitios Cueva, Centro, Bocana y Brazo Norte en el periodo 2009-2011. Se utilizaron diagramas de barra para mostrar los resultados. Se determinó la abundancia del caracol S. gigas para cada uno de los cuatro sitios de muestreo (Bocana, Centro, Cueva y Brazo Norte) y para la totalidad de la caleta a través del tiempo (enero 2005 a enero 2008 y abril 2009 a septiembre 2011) por el método directo, cuantificando el número total de organismos capturados (marcados y no marcados). Con los datos obtenidos en el muestreo CMR y la proporción de organismos marcados y no marcados, se determinó el tamaño poblacional para cada sitio de muestreo y para la totalidad de la caleta, a través del tiempo, utilizando el método de Jolly-Seber (Jolly, 1965; Seber, 1965) y Schnabel (1938). Se utilizaron diagramas de barra para comparar la abundancia determinada mediante el método directo y las estimaciones del tamaño de población, obtenidas por los métodos de Schnabel y Jolly-Seber, para cada uno de los sitios estudiados y para la totalidad de la caleta de Xel-Há a través del tiempo. Posteriormente se calculó la abundancia media y 41 desviación estándar a partir de la abundancia determinada por el método directo, para cada sitio y para la totalidad de la caleta de Xel-Há en el periodo 2005-2008 y 2009-2011, respectivamente. La densidad relativa de la poblacional (individuos por hectárea) se derivó de la abundancia determinada mediante el método directo y del tamaño de población estimado por los métodos de Jolly-Seber y Schnabel. Se dividió el número de organismos observados y estimados entre el área de muestreo de cada uno de los cuatro sitios estudiados, para cada mes muestreado. Con la finalidad de comparar la densidad entre métodos y sitios, se utilizaron diagramas de barra. La densidad relativa de caracoles por hectárea en la totalidad de la caleta de Xel-Há fue derivada de la abundancia determinada mediante el método directo y derivado del tamaño de población estimado por los métodos de Jolly-Seber y Schnabel. Se dividió el número de organismos observados y estimados entre el área de muestreo, el cual correspondió a 3.9 ha en el periodo 2005-2008 y a 4.5 ha en el periodo 2009-2011. Se calculó la densidad media y desviación estándar a partir de la densidad estimada por el método directo y por los métodos de Schnabel y Jolly-Seber, para cada sitio y para la totalidad de la caleta de Xel-Há en los periodos 2005-2008 y 2009-2011, respectivamente. 7.3.2 Estructura de tallas Se determinó la estructura de tallas para longitud de heliconcha, en cada uno de los cuatro sitios estudiados de la caleta de Xel-Há, para cada mes estudiado, utilizando diagramas de densidad de puntos, los cuales muestran la abundancia por talla. 42 Con la finalidad de detectar diferencias significativas en la estructura de longitud de heliconcha y grosor de labio entre sitios de muestreo, se ejecutó un análisis de varianza no-paramétrico (Kruskal-Wallis), con un nivel de confianza de 95%, utilizando el programa Infostat/S. Posteriormente se compararon los sitios entre periodos mediante una análisis de varianza noparamétrico (Kruskal-Wallis) con un nivel de confianza de 95%. Se determinó la estructura de tallas para longitud de heliconcha, en la totalidad de la caleta de Xel-Há, para cada mes estudiado, utilizando diagramas de densidad de puntos, los cuales muestran la abundancia por talla. Con la finalidad de detectar diferencias significativas en la estructura de longitud de heliconcha y grosor de labio entre periodos de estudio, se ejecutó un análisis de varianza no-paramétrico (Kruskal-Wallis) con un nivel de confianza de 95%, utilizando el programa Infostat/S. Con el objetivo de determinar a cual talla se forma el labio del caracol rosa y si existen diferencias entre sitios, se utilizaron diagramas de dispersión, los cuales muestran la relación entre longitud de heliconcha (x) y grosor de labio (y). Se utilizaron todas las medidas obtenidas en cada uno de los cuatro sitios de estudio en ambos periodos. Con el objetivo de determinar a cual talla se forma el labio del caracol rosa en la caleta de XelHá y si existen diferencias entre periodos, se utilizaron diagramas de dispersión, los cuales muestran la relación entre longitud de heliconcha (x) y grosor de labio (y). Se utilizaron todas las medidas de obtenidas en la totalidad de la caleta de Xel-Há, en cada uno de los periodos de estudio (2005-2008 y 2009-2011), respectivamente. 43 Para detectar diferencias en la estructura de tallas a través del tiempo, con respecto al grosor de labio del caracol rosa, se calculó la media, mediana y desviación estándar de este, para cada mes muestreado, en cada uno de los cuatro sitios estudiados de la caleta de Xel-Há, así como para la totalidad de la caleta de Xel-Há. Se utilizaron diagramas de caja para mostrar la variabilidad del grosor de labio en el tiempo y espacio. Se calculó la abundancia relativa (%) y absoluta de organismos juveniles (sin labio), organismos subadultos (grosor de labio < 5mm) y organismos adultos (grosor de labio ≥ 5mm) para cada uno de los cuatro sitios estudiados de la caleta de Xel-Há y para la totalidad de la caleta de Xel-Há a través del tiempo. Se utilizaron diagramas de barra para mostrar los resultados. Se ejecutó un análisis de componentes principales, utilizando el programa Infostat/S, con la finalidad de determinar si existe una relación entre la distribución espacial del caracol rosa S. gigas en cada uno de los cuatro sitios estudiados de la caleta de Xel-Há (Cueva, Centro, Bocana y Brazo Norte) y la longitud de heliconcha y el grosor de labio. Se utilizaron todas las medidas morfométricas obtenidas entre enero 2005 y septiembre 2011. Se utilizó un biplot para mostrar los resultados. 7.3.3. Crecimiento por métodos directos El crecimiento fue calculado para todos los individuos recapturados en dos muestreos consecutivos. Se calculó para cada individuo el crecimiento por día en milímetros (mm), determinando el aumento en longitud de cada uno y dividiendo éste entre el número de días transcurridos entre la última observación y su recaptura. 44 Luego se calculó el crecimiento medio por día en mm y su desviación estándar por clase de talla y por sitio, utilizando 19 clases de talla con un intervalo de 10 mm, desde 60 mm a > 239 mm. Para detectar diferencias significativas en el crecimiento entre clases de talla y entre sitios, se ejecutó un ANOVA, con un nivel de confianza de 95%, para cada periodo de estudio, respectivamente. Se calculó el crecimiento medio por día y su desviación estándar para la totalidad de la caleta de Xel-Há por clase de talla, en el periodo 2005-2008 y 2009-2011, respectivamente. Para detectar diferencias significativas entre clases de talla y entre periodos se ejecutó un ANOVA con un nivel de confianza de 95%, para cada periodo de estudio. Con las tasas medias de crecimiento obtenidas para la totalidad de la caleta de Xel-Há, se calculó la curva de crecimiento y la edad estimada de los organismos para el periodo 2005-2008 y 20092011, respectivamente. En base de los resultados anteriormente obtenidos se eligieron todas las clases de tallas que mostraron un crecimiento medio mayor a 0.15 mm día -1, seleccionando las clases de talla de 6069 mm hasta 170-179 mm, con la finalidad de detectar diferencias en el crecimiento entre meses y entre los sitios. Se calculó la media y desviación de la tasa de crecimiento (mm día -1) por clase de talla a través del tiempo, mostrando el incremento entre dos muestreos consecutivos. Se utilizaron diagramas de puntos para mostrar los resultados. 45 7.3.4. Estimación de los parámetros de crecimiento de la ecuación de Von Bertalanffy Se calcularon los parámetros de crecimiento para la longitud de heliconcha y grosor de labio del caracol rosa, para el periodo de estudio 2005-2008 y 2009-2011, respectivamente, utilizando la ecuación de Von Bertalanffy (VGBF) con oscilación estacional (Pauly & Gauschütz, 1979), (1) L(t) = L∞ * (1- exp(-k*(t-t0)-(Ck/2π)*sin(2π*(t-ts))); donde L(t)= Longitud al tiempo t; L∞= Longitud asintótica (t=∞) ; k= constante de crecimiento; t= tiempo; t0= tiempo cero; C= oscilación. Se calcularon los parámetros de la ecuación directamente, creando mediante el programa FSAT II un archivo de incrementos de longitud, utilizando los datos de marcaje recaptura y las longitudes de heliconcha de caracoles recapturados en dos muestreos consecutivos. Sólo se incluyeron medidas de longitud de heliconcha de organismos con un aumento ≥ 1 mm. Para el 2005-2008 se analizaron 1405 pares de medidas de longitud y para el 2009-2011 se analizaron 4090 pares de medidas de longitud de heliconcha. Los parámetros de crecimiento fueron obtenidos utilizando el método de Appeldoorn (1987b) incluido en el paquete FISAT II. Posteriormente se calcularon los parámetros de la ecuación de Von Bertalanffy de forma indirecta. Primero se generó la distribución de frecuencia de longitud de heliconcha de la población de S. gigas, con la ayuda del programa Infostat/S, utilizando intervalos de 10 mm en un rango de 40 mm a 270 mm, para cada mes de muestreo y para cada periodo de estudio respectivamente. Se obtuvieron los incrementos de longitud mediante el análisis de progresión de modas, utilizando el método de Bhattacharya, seguido por el método de separación de los 46 componentes de distribución normal (NORMSEP) y finalmente el enlace de las medias obtenidas, utilizando las rutinas del programa FISAT II. Se utilizaron los aumentos de longitud para calcular el crecimiento mediante el método de Appeldoorn (1987b). Para calcular el crecimiento del grosor de labio se utilizaron los datos de marcaje-recaptura de 158 individuos con labio, recapturados en 2 muestreos consecutivos en el periodo 2005-2008 y de 687 individuos en el periodo 2009-2011. Los incrementos de grosor de labio fueron analizados directamente, utilizando el método de Appeldoorn (1987b) incluido en el paquete FISAT II. 7.3.5. Reclutamiento Con la finalidad de determinar los periodos de reclutamiento de juveniles, se generaron histogramas de la frecuencia de longitud de heliconcha de caracoles juveniles (organismos sin labio), utilizando un intervalo de 10 mm a partir de los 40 mm de longitud de heliconcha, para cada mes muestreado en el periodo de enero 2005 a septiembre 2011. Se determinó visualmente la aparición de organismos con una longitud menor a 100 mm. Se determinó el reclutamiento para el periodo 2005-2008 y 2009-2011, utilizando el programa FISAT II, introduciendo los parámetros de VGBF obtenidos por el método directo y la distribución de frecuencia de tallas de la población de caracoles S. gigas, para cada periodo respectivamente. 7.3.6. Sobrevivencia y Mortalidad Se calculó el índice de sobrevivencia finita (S) (Krebs, 1985), 47 (2) St = (Nt/N0)*100; donde St= Sobrevivencia en el tiempo t; Nt= Tamaño de población el tiempo t; N0= Número de individuos marcados en el tiempo 0; a partir de los historiales de recaptura (Jolly, 1965; Seber, 1965) de los caracoles, para cada lote (individuos marcados en el mismo muestreo) de organismos, para cada uno de los sitios estudiados de la caleta de Xel-Há (Cueva, Centro, Bocana y Brazo Norte) en el periodo 20052008 y 2009-2011, respectivamente. Se calculó el intervalo medio entre muestreos y se promedió la sobrevivencia de todos los lotes, utilizando el tiempo promedio transcurrido entre marcaje y recaptura. Se contabilizaron en cada mes de muestreo el número de caracoles muertos en cada uno de los cuatro sitios de muestreo (Cueva, Centro, Bocana y Brazo Norte). Se utilizó un diagrama de barras para mostrar los resultados. Se calculó el índice de sobrevivencia finito para la totalidad de la población de la caleta de XelHá, para el periodo 2005-2008 y 2009-2011 respectivamente, utilizando la misma metodología. A partir de los historiales de recaptura de los caracoles se calculó el índice de mortalidad instantáneo (Krebs, 1985), (3) Zt = -ln (Nt/N0); donde Zt= Índice de mortalidad en el tiempo t; Nt= Tamaño de población el tiempo t; N0= Número de individuos marcados en el tiempo 0; 48 para la totalidad de la población de la caleta de Xel-Há, en el periodo 2005-2008 y 2009-2011 respectivamente. Se calculó el intervalo medio entre muestreos y se promedió la mortalidad, utilizando el tiempo promedio transcurrido entre marcaje y recaptura. Se contabilizaron en cada mes de muestreo el número de caracoles muertos en cada uno de los cuatro sitios de muestreo (Cueva, Centro, Bocana y Brazo Norte). Se utilizó un diagrama de barras para mostrar los resultados. Se analizaron visualmente las frecuencias de longitudes de heliconcha por clases de talla del caracol rosa S. gigas sin labio y con labio en la caleta de Xel-Há, en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011 respectivamente y se determinó la clase modal. Posteriormente se estimó la mortalidad total instantánea (Z) mediante las curvas de captura de tallas convertidas (length converted catch curve), utilizando el programa FISAT II. Se utilizó la distribución de frecuencia de tallas de la población de S. gigas de la caleta de Xel-Há y los parámetros de VBGF correspondientes, obtenidos por el método directo, para el periodo 20052008 y 2009-2011, respectivamente. También se calculó la mortalidad total instantánea (Z), utilizando el mismo método, pero la distribución de frecuencia del grosor de labio y los parámetros de VBGF correspondientes al crecimiento de este. 7.3.7. Movilidad Para describir el patrón y dirección de movilidad se calculó el porcentaje de organismos noresidentes (marcados en otro sitio), residentes (marcados en el sitio) y no marcados en cada sitio de muestreo. Se utilizaron diagramas de área para mostrar los resultados. También se determinó el número de organismos que emigraron de su sitio de origen y su respectivo destino a través del 49 tiempo. Se evaluó el patrón de estacionalidad de las transiciones visualmente, utilizando diagramas de barra. Se calculó la media y desviación estándar de la longitud de heliconcha y del grosor de labio de los caracoles que emigraron de su sitio de marcaje a través del tiempo. Se ejecutó un análisis de varianza con un nivel de confianza de 95%, con el objetivo de determinar si existen diferencias significativas entre las medias de longitud de heliconcha, así como las medias del grosor de labio respectivamente, según el destino de sus desplazamientos. Se calculó el porcentaje (%) de organismos juveniles (sin labio), organismos subadultos (grosor de labio < 5mm) y organismos adultos (grosor de labio ≥ 5mm) que participaron en las transiciones entre sitio, para cada uno de los cuatro sitios estudiados. Con la finalidad de determinar si la talla influye sobre la distribución espacial de los caracoles emigrantes y residentes, se hizo un análisis de componentes principales, utilizando el programa Infostat/S, para los organismos marcados en cada uno de cuatros sitios de muestreo de la caleta de Xel-Há (Cueva, Centro, Bocana y Brazo Norte) respectivamente, introduciendo como variables el sitio de recaptura, la longitud de heliconcha y el grosor de labio. Se utilizó un biplot para mostrar los resultados. 50 8. Resultados 8.1 Demografía poblacional 8.1.1 Abundancia, Tamaño de Población y Densidad En las figuras 16.A a 16.D se muestra el número de individuos que fueron marcados en cada uno de los cuatro sitios estudiados de la Caleta de Xel-Há: Cueva (Fig.16.A), Centro (Fig. 16.B), Bocana (Fig. 16.C) y Brazo Norte (Fig. 16.D), durante el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011, con excepción de Brazo Norte, donde sólo se realizaron muestreos en el periodo 2009-2011. En la Cueva se marcaron 583 individuos entre enero 2005 y enero 2008 y 1976 individuos entre abril 2009 y septiembre 2011. El mayor número de individuos fue marcado en octubre 2009 y julio 2010, mientras el menor número de individuos fue marcado en abril 2005. Centro fue el sitio donde se capturaron y marcaron más individuos, con 680 caracoles entre enero 2005 y enero 2008 y 2126 individuos entre abril 2009 y septiembre 2011. El menor número de individuos fue capturado y marcado en Bocana, con un total de 402 caracoles entre enero 2005 y enero 2008 y 891 entre abril 2009 y septiembre 2011. En Brazo Norte se marcaron 1598 caracoles, variando entre 70 y 228 individuos entre muestreos. Para el periodo de abril 2009 a septiembre 2011 y para Cueva, Centro y Brazo Norte, el número de individuos marcados fluctuó, con un aumento entre octubre 2009 y septiembre 2010. 51 Figura 16. Número de caracoles S. gigas marcados en cada uno de los cuatro sitios estudiados de la Caleta de XelHá: (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, durante el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. 52 En la figura 17.A se muestra el número de individuos que fueron marcados en la Caleta de XelHá durante el periodo de enero 2005 a enero 2008, el cual corresponde a la suma de los individuos marcados en Cueva, Centro y Bocana. En la figura 17.B se presenta el número de individuos marcados en la Caleta de Xel-Há entre abril 2009 y septiembre 2011, equivalente a la suma de los individuos marcados en los sitios anteriormente mencionados más Brazo Norte. En el primer periodo se marcaron 1665 caracoles con un mínimo de 27 y un máximo de 118 entre muestreos. Para el periodo abril 2009 a septiembre 2011 se marcaron 6627 individuos con un mínimo de 298 y un máximo de 787, observándose un aumento entre octubre 2009 y septiembre 2010. Figura 17. Número de caracoles S. gigas marcados en la Caleta de Xel-Há durante el periodo de (A) enero 2005 a enero 2008 y (B) de abril 2009 a septiembre 2011. En las figuras 18.A a 18.D se muestra la abundancia de caracoles S. gigas, la cual se determinó el por el método directo, método de Schnabel y método de Jolly-Seber, para cada uno de los cuatro sitios estudiados de la caleta de Xel-Há: Cueva (Fig. 18.A), Centro (Fig. 18.B), Bocana (Fig. 18.C) y Brazo Norte (Fig. 18.D). Cueva, Centro y Bocana fueron estudiados durante el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. Brazo Norte sólo fue muestreado en el periodo de abril 2009 a 53 septiembre 2011. En general e independientemente del método utilizado, se observó para Cueva, Centro y en menor grado en Bocana, que la abundancia de caracoles fue menor en el periodo 2005-2008 en comparación con el periodo abril 2009 a septiembre 2011. También se observó, independientemente del periodo y sitio, que las abundancias determinadas por el método directo generalmente fueron menores en comparación con las dos estimaciones del tamaño de población. En Cueva la abundancia determinada por el método directo fue constante a través del periodo 2005-2008, pero incrementó de abril 2009 a noviembre 2010. El método de Schnabel mostró un incremento gradual en el tamaño de la población a través de los dos periodos de estudio con un máximo de 1692 individuos estimados en septiembre 2011. El método de Jolly-Seber mostró un tamaño de población fluctuante con tres picos (abril 2005, julio 2007 y enero 2011). En Centro la abundancia determinada por el método directo fue constante a través de ambos periodos, pero fue mayor de abril 2009 a septiembre 2011, con respeto al periodo anterior. El método de Schnabel mostró un incremento gradual en el tamaño de la población, alcanzando un máximo en septiembre 2011 con 2503 individuos. El método de Jolly-Seber mostró un tamaño de población constante entre enero 2005 y enero 2008, con excepción de un pico en mayo 2005. En el periodo 2009-2011 se observaron fluctuaciones marcadas en el tamaño de población por éste método. En Bocana la abundancia determinada por el método directo fue constante a través de los dos periodos. Por el método de Schnabel se observó un tamaño de población constante a través del periodo 2005-2008 y un incremento gradual para el segundo periodo, alcanzando un máximo en septiembre 2011 con 1808 individuos. El método de Jolly-Seber mostró fluctuaciones en el tamaño de población a través del periodo enero 2005 a enero 2008 con tres picos (mayo 2005, octubre 2005 y mayo 2006) y un aumento gradual a través del periodo abril 2009 a septiembre 2011. En Brazo Norte la abundancia determinada por el método directo fue constante. El método 54 de Schnabel mostró un incremento gradual en el tamaño de la población hasta septiembre 2011 con 1248 individuos y con el método de Jolly-Seber se obtuvo un tamaño de población variable con un pico en marzo 2011. 55 Figura 18. Número de caracoles S. gigas capturados (azul) y estimados mediante los métodos de Jolly-Seber (rojo) y Schnabel (verde), en cada uno de los cuatro sitios estudiados de la Caleta de Xel-Há: (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, durante el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. 56 En la tabla 7 se muestran las abundancias medias de S. gigas obtenidas del número de individuos capturados y de las estimaciones del tamaño de población por los métodos de Jolly-Seber y Schnabel en el periodo 2005-2008 y 2009-2011 en cada uno de los cuatro sitios estudiados de la caleta de Xel-Há: Cueva, Centro, Bocana y Brazo Norte. Independientemente del método utilizado se pudo observar que las abundancias aumentaron en el 2009-2011 con respecto al periodo anterior, donde Cueva y Centro presentaron las mayores abundancias medias. Las menores abundancias medias se obtuvieron en Bocana. Las abundancias medias de Brazo Norte fueron menores a las obtenidas en Centro y Cueva, pero mayores en comparación con Bocana. 57 58 En las figuras 19.A y 19.B se muestra el número de caracoles capturados y estimados por los métodos de Jolly-Seber y Schnabel en la Caleta de Xel-Há durante el periodo de enero 2005 a enero 2008 y de abril 2009 y septiembre 2011, respectivamente. En el primer periodo de estudio el número de individuos capturados y estimados fue obtenido a partir de los organismos observados en Cueva, Centro y Bocana. En el segundo periodo se obtuvo el número de individuos capturados y estimados de los organismos observados en Cueva, Centro, Bocana y Brazo Norte. En ambos periodos de estudio la abundancia determinada por el método directo generalmente fue menor en comparación con las dos estimaciones del tamaño de población. El método de Schnabel mostró un incremento gradual en el tamaño de la población, tanto a través del periodo 2005-2008, como en el periodo 2009-2011. El método de Jolly-Seber mostró un tamaño de población constante a través del periodo de enero 2005 a enero 2008, con excepción de un pico entre abril y mayo 2005. En el periodo 2009-2011 se observó un pico marcado, con un máximo entre noviembre 2010 y mayo 2011, utilizando éste método. 59 60 En la tabla 8 se muestran las abundancias medias de S. gigas obtenidas a partir del número de individuos capturados y de las estimaciones del tamaño de población por los métodos de JollySeber y Schnabel en el periodo 2005-2008 y 2009-2011, para toda la población de la caleta de Xel-Há. Se observó, independientemente del método utilizado, que las abundancias aumentaron entre 4 a 5 veces en el periodo 2009-2011 con respecto al periodo 2005-2008. En ambos periodos las medias obtenidas de las estimaciones del tamaño de población por los métodos de Schnabel y Jolly-Seber fueron similares entre sí y 4 a 5 veces mayores con respeto al número medio de organismos capturados. Tabla 8. Abundancia media del caracol rosa S. gigas y desviación estándar (d.e.) en los periodos 20052008 y 2009-2011, en la caleta de Xel-Há, obtenidas del número de individuos capturados y de las estimaciones del tamaño de población por los métodos de Jolly-Seber y Schnabel. Periodos Métodos Organismos capturados Jolly-Seber Schnabel 2005-2008 2009-2011 Abundancia media (d.e.) Abundancia media (d.e.) 219 (49.0) 897 (465.1) 1096 (388.1) 4303 (2699.0) 1002 (301.3) 4143 (1645.7) En las figuras 20.A a 20.D se muestra la densidad poblacional (individuos ha -1) del caracol rosa en cada uno de los cuatro sitios estudiados de la caleta de Xel-Há: Cueva (Fig. 20.A), Centro (Fig. 20.B). Bocana (Fig. 20.C) y Brazo Norte (Fig. 20.D), los cuales fueron estudiados durante el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011; con excepción de Brazo Norte, donde sólo se realizaron muestreos de abril 2009 a septiembre 2011. La densidad poblacional fue derivada del número de individuos capturados, así como del número de individuos estimados por los métodos de Schnabel y Jolly-Seber. En Cueva y Centro se observó un incremento en las densidades en el periodo abril 2009 a septiembre 2011 con respeto al 20052008, independientemente del método utilizado. Las mayores densidades fueron observadas en Cueva y Brazo Norte y las más bajas en Centro y Bocana. 61 Figura 20. Densidad poblacional (individuos ha⁻¹) del caracol rosa S. gigas en cada uno de los cuatro sitios estudiados de la Caleta de Xel-Há: (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, durante el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. 62 En la tabla 9 se muestran las densidades (individuos ha-1) medias de S. gigas en el periodo 20052008 y 2009-2011, para cada uno de los cuatro sitios estudiados de la caleta de Xel-Há: Cueva, Centro, Bocana y Brazo Norte. La densidad poblacional fue derivada del número de individuos capturados, así como del número de individuos estimados por los métodos de Schnabel y JollySeber. Independientemente del método utilizado, se pudo observar que las densidades aumentaron en el periodo 2009-2011 en los sitios Cueva, Centro y Bocana, con respecto al periodo 2005-2008. Las densidades mayores fueron observadas en Cueva para el periodo 20052008 y en Brazo Norte y Cueva para el periodo 2009-2011. Las densidades menores se obtuvieron en Centro en ambos periodos del estudio. Tabla 9. Densidad (individuos ha-1) media poblacional del caracol rosa S. gigas y su desviación estándar (d.e.) en los periodos 2005-2008 y 2009-2011, en los cuatro sitios de la caleta de Xel-Há (Cueva, Centro, Bocana y Brazo Norte), obtenida del número de individuos capturados y por los métodos de Jolly-Seber y Schnabel. Sitios Periodos Métodos Cueva 200520092008 2011 Densidad media (d.e.) Centro 200520092008 2011 Densidad media (d.e.) Bocana 200520092008 2011 Densidad media (d.e.) Brazo Norte 200520092008 2011 Densidad media (d.e.) Capturados 149 (36.0) 582 (227.1) 28 (6.7) 132 (55.3) 65.9 (34.89) 151.5 (62.61) - 484.5 (125.23) Jolly-Seber 808 (574.3) 1968 (1657.6) 102 (125.7) 366 (304.4) 280.0 (203.90) 525.5 (261.17) - 1270.5 (1069.77) Schnabel 388 (169.2) 1803 (953.1) 150 (65.5) 621 (353.6) 522.7 (103.82) 804.5 (471.92) - 1411.3 (546.29) En las figuras 21.A y 21.B se muestra la densidad de caracoles por hectárea en la caleta de XelHá en el periodo enero 2005 a enero 2008 y de abril 2009 a septiembre 2011, respectivamente. Ésta fue derivada del número de individuos capturados, así como del número de individuos estimados por los métodos de Schnabel y Jolly-Seber. La densidad de caracoles fue menor en el periodo de enero 2005 a enero 2008 en comparación con el periodo abril 2009 a septiembre 63 2011. En ambos periodos se observó que las densidades determinadas por el método directo fueron menores en comparación con las densidades obtenidas a partir de los métodos Schnabel y Jolly-Seber. La densidad obtenida por el método de Schnabel mostró un incremento gradual a través del tiempo en ambos periodos de estudio. Por el método de Jolly-Seber se obtuvieron densidades constantes a través del periodo 2005-2008, con excepción de un pico entre abril y mayo 2005. En el periodo 2009-2011 se observó un incremento del tamaño de población, con un máximo entre noviembre 2010 y marzo 2011. 64 65 En la tabla 10 se muestran las densidades medias (individuos por hectárea) de S. gigas en el periodo 2005-2008 y 2009-2011 para toda la caleta de Xel-Há. Éstas fueron derivadas del número de individuos capturados, así como del número de individuos estimados por los métodos de Schnabel y Jolly-Seber. Se observó, independientemente del método utilizado, que las densidades aumentaron entre 4 a 5 veces en el periodo 2009-2011 con respeto al periodo 20052008. En ambos periodos de estudio las densidades medias obtenidas por los métodos de Schnabel y Jolly-Seber fueron similares entre sí y 4-5 veces mayores con respeto a la densidad media obtenida por el método directo. Tabla 10. Densidad (individuos ha-1) media del caracol rosa S. gigas y su desviación estándar (d.e.) en los periodos 2005-2008 y 2009-2011, en la caleta de Xel-Há, obtenidas a partir del número de individuos capturados y por los métodos de Jolly-Seber y Schnabel. Periodo 2005-2008 2009-2011 Densidad media (d.e.) Densidad media (d.e.) Organismos capturados Jolly-Seber 56.0 (12.56) 209.7 (131.61) 243.5 (75.14) 809.2 (654.78) Schnabel 245.9 (92.57) 849.9 (433.36) Métodos 8.1.2 Estructura de Tallas En las figuras 22.A a 22.D se muestran diagramas de densidad de puntos de las longitudes de heliconcha (mm) del caracol rosa S. gigas y abundancia por talla en cada uno de los cuatro sitios estudiados de la caleta de Xel-Há: Cueva (Fig. 22.A), Centro (Fig. 22.B), Bocana (Fig. 22.C) y Brazo Norte (Fig. 22.D). Cueva, Centro y Bocana fueron estudiados durante el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. Brazo Norte sólo fue muestreado en el periodo de abril 2009 a septiembre 2011. El rango de tallas fue similar en los sitios Cueva, Centro y Bocana en el periodo 2005-2008. En Cueva se encontraron organismos entre 59-270 mm, en Centro entre 6266 259 mm y en Bocana entre 60-270 mm. La proporción de tallas fue similar en Cueva y Centro en el periodo de enero 2005 a enero 2008, observándose la mayoría de los organismos en un rango de tallas de 120 mm a 240 mm y una distribución homogénea a lo largo de este. En Cueva los organismos menores a 120 mm fueron más abundantes en comparación con Centro y Bocana. En Bocana los organismos con tallas entre 190 y 240 mm fueron un poco más abundantes en comparación con los otros sitios. En el periodo 2009-2011 se observaron diferencias en la estructura de tallas a través del tiempo y entre sitios. En Cueva se hallaron organismos más pequeños y en mayor abundancia en comparación con el periodo anterior. El rango de tallas fue de 45 mm a 242 mm. El rango de tallas en Centro fue de 60-254 mm y también se observó mayor abundancia en las tallas pequeñas en comparación con el periodo anterior. Por lo contrario, en Bocana no se encontraron organismos menores a 71 mm y los organismos menores a 120 mm permanecieron escasos. La talla máxima encontrada fue de 248 mm. El rango de tallas observado en Brazo Norte fue similar al rango observado en Centro, con caracoles entre 64-250 mm. En el periodo abril 2009 a septiembre 2011 se observó una estructura de tallas similar en los sitios Cueva, Centro y Brazo Norte, que fue muy variable a través del tiempo, observándose un aumento significativo en la abundancia de organismos con tallas entre 60 mm y 120 mm entre junio y octubre 2009. En febrero 2010 se observó alta abundancia de organismos con tallas entre 100-180 mm y en mayo 2010 entre 140-240 mm. En los siguientes meses muestreados se observó que los organismos con tallas entre 170-250 mm fueron los más abundantes en estos sitios. La proporción de tallas observada en Bocana en el mismo periodo fue constante a través del tiempo, con mayor abundancia de organismos con tallas entre 180 mm y 240 mm. Se observó un incremento gradual en la abundancia de éstas tallas, con máximo en septiembre 2011. 67 68 69 Se detectaron diferencias estadísticas significativas (p < 0.05) entre las medianas de la longitud de heliconcha del caracol rosa obtenidas en cada uno de los cuatro sitios estudiados (Tabla 11). Las menores medianas se obtuvieron para Cueva y Brazo Norte, con 172 mm y 183 mm, respectivamente. La mediana de los organismos capturados en Centro fue de 195 mm. Los caracoles de Bocana tuvieron la mediana mayor con 208 mm. Tabla 11. Análisis de varianza (Kruskal-Wallis) de medianas de longitud de heliconcha del caracol S. gigas entre cada uno de los cuatro sitios estudiados de la caleta de Xel-Há en el periodo de enero 2005 a septiembre 2011. Sitio N Medias D.E. Medianas Bocana 3499 202.12 25.64 208 Brazo Norte 3779 177.75 31.25 183 Centro 5410 187.22 32.31 195 Cueva 6627 168.71 34.47 172 *N-Tamaño de Muestra *D.E.- Desviación estándar *H- Estadístico de Kruskal-Wallis *p- Valor de p *Letras distintas indican diferencias significativas (p<0.05) H p Rango 2852.53 0.009 A B C D Se detectaron diferencias estadísticas significativas (p < 0.05) entre las medianas de la longitud de heliconcha, obtenidas en el periodo 2005-2008 y 2009-2011, en los tres sitios comparados (Cueva, Centro y Bocana) (Tabla 12). Se observó un incremento en las medianas para el periodo 2009-2011 en los tres sitios. Tabla 12. Análisis de varianza (Kruskal-Wallis) de medianas de longitud de heliconcha del caracol S. gigas en el periodo 2005-2008 y 2009 2011 y en los sitios Cueva, Centro y Bocana de la caleta de XelHá. Sitio Periodo N Medias D.E. Medianas H p Cueva Centro Bocana 2005-2008 2009-2011 2005-2008 2009-2011 2005-2008 2079 4548 1464 3946 1530 2009-2011 1969 *N-Tamaño de Muestra *H- Estadístico de Kruskal-Wallis 161.56 171.98 184.25 188.31 198.61 33.03 34.62 30.49 32.89 29.07 204.85 22.25 *D.E.- Desviación estándar *p- Valor de p 162 177 190 197 206 170.63 <0.0001 38.31 <0.0001 25.31 <0.0001 209 70 En las figuras 23.A y 23.B se muestran las longitudes de heliconcha (mm) del caracol rosa S. gigas y abundancia por talla, para toda la caleta de Xel-Há, durante el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011, respectivamente. En el primer periodo de estudio las medidas fueron obtenidas a partir de los organismos muestreados en Cueva, Centro y Bocana; y en el segundo periodo de los sitios anteriores más Brazo Norte. El rango de tallas tuvo una amplitud similar en ambos periodos. Sin embargo, la proporción de tallas varió considerablemente a través del tiempo. Entre enero 2005 y mayo 2005 la mayoría de los organismos se encontraban distribuidos en un rango de tallas entre 120 mm y 240 mm con mayor proporción de organismos entre 190 mm y 240 mm de longitud. A partir de junio 2005 disminuyó la abundancia de organismos entre 190 mm y 240 mm y se hallaron más organismos menores a 120 mm que en los meses anteriores. Esta proporción de tallas se mantuvo relativamente constante para el resto de este periodo de estudio. Los organismos con tallas menores a 100 mm eran escasos en todo el periodo 2005-2008. A partir de abril 2009 se observó un aumento en la abundancia de organismos entre 180 y 240 mm. Entre octubre 2009 y febrero 2010 se observó la aparición de muchos organismos con tallas entre 60 y 120 mm, los cuales aumentaron de talla en los meses siguientes. A partir de mayo 2011 los organismos con tallas entre 180-240 mm fueron los más abundantes en la caleta. 71 72 No se detectaron diferencias significativas entre las medianas de la longitud de heliconcha entre los dos periodos estudiados para la totalidad de la población de S. gigas en la caleta de Xel-Há (Tabla 13). Tabla 13. Análisis de varianza (Kruskal-Wallis) de medianas de longitud de heliconcha del caracol S. gigas en el periodo 2005-2008 y 2009 2011 en la caleta de Xel-Há. Periodo N 2005-2008 5074 2009-2011 14242 *N-Tamaño de Muestra *D.E.- Desviación estándar *H- Estadístico de Kruskal-Wallis Medias D.E. Medianas H p 179.28 182.58 34.9 33.61 185 190 38.28 0.6936 *p- Valor de p En las figuras 24.A a 24.D se muestra la relación entre longitud de heliconcha y grosor de labio del caracol rosa en cada uno de los cuatro sitios estudiados de la caleta de Xel-Há: Cueva (Fig. 24.A), Centro (Fig. 24.B), Bocana (Fig. 24.C) y Brazo Norte (Fig. 24.D), en el periodo de estudio 2005-2011; con excepción de Brazo Norte, donde sólo se realizaron muestreos entre 2009 y 2011. En los cuatro sitios estudiados se observaron caracoles con un labio formado entre 170 y 250 mm de longitud. En Bocana los individuos con un grosor de labio mayor a 20 mm fueron abundantes en comparación con los otros sitios. En Cueva y Brazo Norte los individuos con labio mayor a 20 mm fueron muy escasos y en Centro se observaron algunos. 73 Figura 24. Diagrama de dispersión, mostrando la relación entre longitud de heliconcha (mm) y grosor de labio (mm) del caracol rosa S. gigas en cada uno de los cuatro sitios estudiados de la caleta de Xel-Há, (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, en el periodo de enero 2005 a septiembre. En las figuras 25.A y 25.B se muestra la relación entre longitud de heliconcha y grosor de labio del caracol rosa en la caleta de Xel-Há durante el periodo de estudio enero 2005 a enero 2008 y abril 2009 a septiembre 2011, respectivamente. En el 2005-2008 las medidas fueron obtenidas a partir de los organismos muestreados en Cueva, Centro y Bocana y en el segundo periodo de los anteriores más Brazo Norte. En ambos periodos de estudio la mayoría de los organismos con un labio formado se hallaron en un rango de tallas entre 170 a 250 mm. En el periodo 2009-2011 se observaron más organismos con un labio formado en comparación con el periodo anterior. 74 Figura 25. Diagrama de dispersión, mostrando la relación entre longitud de heliconcha (mm) y grosor de labio (mm) de la población del caracol rosa S. gigas en la caleta de Xel-Há, en los periodos (A) enero 2005 a enero 2008 y (B) abril 2009 a septiembre 2011. En las figuras 26.A a 26.D se muestran diagramas de caja de las medidas de dispersión (medias, medianas, desviación estándar y valores extremos) del grosor de labio del caracol rosa en cada uno de los cuatro sitios de muestreo de la caleta de Xel-Há: Cueva (Fig. 26.A), Centro (Fig. 75 26.B), Bocana (Fig. 26.C) y Brazo Norte (Fig. 26.D), a través del periodo enero 2005 a enero 2008 y abril 2009 a septiembre 2011; con excepción del Brazo Norte, el cual sólo fue estudiado a partir de abril 2009. En el periodo 2005-2008 se observaron pocos individuos con labio en Cueva y Centro. En el 2009-2011 se observaron más organismos con labio en estos dos sitios, pero las medianas del grosor de labio generalmente fueron menores a 10 mm. En Brazo Norte el grosor de labio mediano fue comparable con Cueva y Centro. En Bocana se encontraron los organismos con mayor mediana del grosor de labio, observándose valores entre 11 mm y 33 mm en el periodo 2005-2008 y de 8 mm a 23 mm en el periodo de abril 2009 a septiembre 2011. No se detectaron diferencias estadísticas significativas (H0.05 = 1995.71; p = 0.9999) entre las medianas del grosor de labio en los cuatro sitios estudiados (Tabla 14).. Tabla 14. Análisis de varianza (Kruskal-Wallis) de medianas de del grosor de labio del caracol S. gigas entre los sitios Cueva, Centro, Bocana y Brazo Norte de la caleta de Xel-Há, en el periodo de enero 2005 a septiembre 2011. Sitio N Bocana 2379 Brazo Norte 1031 Centro 1619 Cueva 1051 *N-Tamaño de Muestra Medias D.E. Medianas 15.33 10.13 14 3.97 3.75 3 4.97 4.62 4 5.09 3.98 4 *D.E.- Desviación estándar H p 1995.71 >0.9999 Se detectaron diferencias estadísticas significativas (Tabla 15) entre las medianas del grosor de labio de S. gigas en los sitios Cueva, Centro y Bocana entre los dos periodos estudiados. Se observó que las medianas fueron menores en el periodo 2009-2011 en los tres sitios comparados. 76 Tabla 15. Análisis de varianza (Kruskal-Wallis) de medianas de del grosor de labio del caracol S. gigas en el periodo 2005-2008 y 2009 2011 y en los sitios Cueva, Centro y Bocana de la caleta de Xel-Há. Sitios Periodos N Medias D.E. Medianas H p Cueva 2005-2008 72 2009-2011 979 Centro 2005-2008 101 2009-2011 1518 Bocana 2005-2008 829 2009-2011 1550 *N-Tamaño de Muestra *H- Estadístico de Kruskal-Wallis 7.15 3.59 4.94 3.97 8.8 6.53 4.72 4.35 18.51 9.73 13.63 9.94 *D.E.- Desviación estándar *p- Valor de p 7 4 7 3 19 12 30.17 <0.0001 65.06 <0.0001 137.86 <0.0001 77 Figura 26. Diagramas de caja mostrando las medidas de dispersión del grosor de labio (media, mediana, desviación estándar y valores extremos) del caracol rosa S. gigas a través del periodo enero 2005 a enero 2008 y de abril 2009 a septiembre 2011, en cada uno de los cuatro sitios estudiados de la Caleta de Xel-Há: (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte. En las figuras 27.A y 27.B se muestran diagramas de caja de las medidas de dispersión (medias, medianas, desviación estándar y valores extremos) del grosor de labio del caracol rosa en la 78 caleta de Xel-Há, durante el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011, respectivamente. En el primer periodo de estudio las medidas fueron obtenidas a partir de los organismos muestreados en Cueva, Centro y Bocana. En el 2009-2011 se realizaron muestreos en los tres sitios anteriormente mencionados más el sitio Brazo Norte. Se observó que las medianas fueron menores en el segundo periodo de estudio (3-9 mm) con respecto al periodo 2005-2008 (11-23 mm), pero no se detectaron diferencias significativas en estas entre los dos periodos estudiados (H = 863.36; p = 0.999). Figura 27. Diagramas de caja mostrando las medidas de dispersión del grosor de labio (media, mediana, desviación estándar y valores extremos) del caracol rosa S. gigas en la Caleta de Xel-Há a través del periodo (A) enero 2005 a enero 2008 y de (B) abril 2009 a septiembre 2011. En las figuras 28.A a 28.D se muestra la abundancia relativa del caracol rosa S. gigas por clases de talla en cada uno de los cuatro sitios estudiados de la caleta de Xel-Há, Cueva (Fig. 28.A), Centro (Fig. 28.B), Bocana (Fig. 28.C) y Brazo Norte (Fig. 28.D), donde se clasificaron como adultos todos los individuos con un grosor de labio ≥ 5 mm, como subadultos los organismos con labio < 5 mm y como juveniles los caracoles sin labio. Los sitios Cueva, Centro y Bocana se estudiaron durante los periodos enero 2005 a enero 2008 y abril 2009 a septiembre 2011, mientras Brazo Norte sólo fue estudiado en el segundo periodo. Cueva y Centro mostraron una estructura de tallas similar, compuestos en más de un 85% de juveniles en el periodo 2005-2008. 79 Para el periodo 2009-2011 y para los sitios Cueva, Centro y Brazo Norte, se observó que la proporción de las tres clases de talla fue similar, con alta abundancia juvenil en estos sitios. Se registró un pico en ésta entre octubre 2009 y septiembre 2010. Bocana tuvo una composición distinta con mayor abundancia relativa de adultos. En Bocana, en el periodo 2005-2008, se observó que el 30 a 70 % de los organismos fueron adultos. La abundancia relativa de organismos adultos disminuyó en noviembre 2007 y enero 2008 alcanzando un mínimo de 15% en enero 2008. Durante el periodo de 2009-2011 la abundancia relativa de adultos aumentó con respecto al periodo anterior y fue de 50 a 90%. 80 Figura 28. Abundancia relativa (%) del caracol rosa S. gigas por clases de talla, Adultos (grosor de labio ≥5 mm), Subadultos (grosor de labio < 5 mm) y Juveniles (sin labio), en cada uno de los cuatro sitios de la caleta de Xel-Há: (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, durante el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. 81 En las figuras 29.A y 29.B se muestra la abundancia relativa del caracol rosa S. gigas por clases de tallas en la caleta de Xel-Há, durante los periodos enero 2005 a enero 2008 y abril 2009 a septiembre 2011, respectivamente. Se clasificaron como adultos todos los individuos con un grosor de labio ≥ 5 mm, como subadultos los organismos con labio < 5 mm y como juveniles los caracoles sin labio. Entre enero 2005 y noviembre 2010 más del 70% de los organismos muestreados fueron juveniles. A partir de enero 2011 ésta fracción presentó un disminución marcada hasta el 30%. Figura 29. Abundancia relativa (%) del caracol rosa S. gigas por clases de talla, Adultos (grosor de labio ≥5 mm), Subadultos (grosor de labio < 5 mm) y Juveniles (sin labio), en la caleta de Xel-Há durante el periodo de (A) enero 2005 a enero 2008 y de (B) abril 2009 a septiembre 2011. En las figuras 30.A a 30.D se muestra la abundancia absoluta del caracol rosa S. gigas por clases de tallas, donde se clasificaron como adultos todos los individuos con un grosor de labio ≥ 5 mm, como subadultos los organismos con labio < 5 mm y como juveniles los caracoles sin labio, en cada uno de los cuatro sitios estudiados de la caleta de Xel-Há, Cueva (Fig. 30.A), Centro (Fig. 30.B), Bocana (Fig. 30.C) y Brazo Norte (Fig. 30.D). Cueva, Centro y Bocana fueron estudiados en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. Brazo Norte sólo fue muestreado en el segundo periodo de estudio. Para el periodo 2005-2008, en Cueva y Centro, 82 la abundancia de adultos fue similar y muy baja. Se observó un aumento gradual en la abundancia de adultos entre abril 2009 y septiembre 2011 en los cuatro sitios estudiados. La mayor abundancia de adultos fue observada en Bocana, donde se observaron fluctuaciones en ésta, con picos entre julio y noviembre 2010 y julio y septiembre 2011. Los juveniles fueron más abundantes en Cueva y en Brazo Norte. Entre abril 2009 y septiembre 2011 se observó un incremento marcado en la abundancia de juveniles en Cueva, Centro y Brazo Norte, con pico en Julio 2010. 83 Figura 30. Abundancia absoluta del caracol rosa S. gigas por clases de talla, Adultos (grosor de labio ≥5 mm), Subadultos (grosor de labio < 5 mm) y Juveniles (sin labio), en cada uno de los cuatro sitios de la Caleta de Xel-Há: (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, a través del periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. 84 En las figuras 31.A y 31.B se muestra la abundancia absoluta del caracol rosa S. gigas por clases de tallas, donde se clasificaron como adultos todos los individuos con un grosor de labio ≥ 5 mm, como subadultos los organismos con labio < 5 mm y como juveniles los caracoles sin labio, en la caleta de Xel-Há, durante el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. En el primer periodo de estudio el número de individuos fue obtenido a partir de la suma de los sitios Cueva, Centro y Bocana, mientras en el segundo periodo fue obtenido a partir de la suma de los sitios anteriormente mencionados más Brazo Norte. En ambos periodos de estudio se observó que la abundancia de juveniles era mayor en comparación con la abundancia de adultos. La abundancia de juveniles fue casi constante de enero 2005 hasta junio 2009, pero aumentó significativamente entre octubre 2009 y marzo 2011. Por lo que respecta la abundancia de adultos, esta incrementó gradualmente de abril 2009, alcanzando un máximo en septiembre 2011. Figura 31. Abundancia absoluta del caracol rosa S. gigas por clases de talla, Adultos (grosor de labio ≥5 mm), Subadultos (grosor de labio < 5 mm) y Juveniles (sin labio), en la Caleta de Xel-Há, a través del periodo de (A) enero 2005 a enero 2008 y de (B) abril 2009 a septiembre 2011. 85 En la figura 32 se muestra el resultado del análisis de componentes principales de la distribución espacial del caracol rosa en cada uno de los cuatro sitios estudiados de la caleta de Xel-Há (Cueva, Centro, Bocana y Brazo Norte) en función de la longitud de heliconcha y el grosor de labio, en el periodo de enero 2005 a septiembre 2011. El eje 1 logró explicar el 72.5% de la variación en la distribución de los caracoles y fue definido en un 99% por la longitud de heliconcha. El eje 2 explicó el 27.5% de la distribución espacial de los caracoles y fue definido en un 99% por el grosor de labio. Se observó que la ocurrencia de caracoles en Bocana se asoció a un aumento en el grosor del labio. Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 68.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Grosor de labio Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 34.00 PC 2(27.5)%) Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 0.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Longitud de heliconcha Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version -34.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version -68.00 -68.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version -34.00 0.00 PC 1(72.5)%) 34.00 68.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Bocana Centro Student version Student version Student version Student version Grosor Student version Variables (Longitud de heliconcha, de Student labio)version Cueva Brazo Norte Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Figura 32. Análisis de componentes principales de la distribución espacial del caracol rosa S. gigas en cada uno de los cuatro sitios estudiados de la caleta de Xel-Há (Cueva, Centro, Bocana y Brazo Norte), durante el periodo de enero 2005 a septiembre 2011, en función de la longitud de heliconcha y del grosor de labio. 8.2 Crecimiento 8.2.1 Crecimiento de la heliconcha por métodos directos En las figuras 33.A a 33.C se muestran las tasas de crecimiento medio del caracol rosa por clase de talla y las desviaciones estándar, obtenidas durante el periodo de estudio enero 2005 a enero 2008 en los sitios: Cueva (Fig. 33.A), Centro (Fig. 33.B) y Bocana (33.C). En Cueva se observó 86 el mayor crecimiento en los organismos con una talla entre 70-79 mm. El crecimiento disminuyó en las clases de 80-89 mm a 100-109 mm y fue relativamente constante en las clases de tallas de 100 mm a 149 mm, con una tasa de crecimiento de 0.21 - 0.26 mm día-1. La tasa de crecimiento disminuyó gradualmente en los organismos mayores a 150 mm. Para Centro no se obtuvieron tasas de crecimiento de organismos menores a 90 mm. El crecimiento fue relativamente constante para las tallas de 110 mm a 159 mm, con tasas de 0.27 - 0.37 mm día-1. El crecimiento disminuyó gradualmente en organismos mayores a 160 mm. En Bocana se observaron las tasas de crecimiento más bajas en comparación con los otros dos sitios. La clase de talla más pequeña observada fueron los organismos entre 110-119 mm, los cuales crecieron en promedio 0.19 mm día-1. El crecimiento de organismos entre 110 mm y 179 mm fue variable. En los organismos mayores a 180 mm el crecimietno fue menor a 0.1 mm día -1. 87 Figura 33. Crecimiento medio (mm día-1) del caracol rosa S. gigas por clase de talla y desviación estándar, en los sitios (A) Cueva, (B) Centro y (C) Bocana, durante el periodo de enero 2005 a enero 2008. 88 En las figuras 34.A a 34.B se muestran las tasas de crecimiento medio del caracol rosa por clase de talla y las desviaciones estándar, obtenidas durante el periodo de estudio abril 2009 a septiembre 2011 en los sitios: Cueva (Fig. 34.A), Centro (Fig. 34.B), Bocana (34.C) y Brazo Norte (Fig. 34.D). El patrón de crecimiento observado en Cueva, Centro y Brazo Norte fue similar, con un crecimiento relativamente constante en los organismos entre 70 mm y 149 mm. El crecimiento mayor en éstas clases de talla se obtuvo en Centro, donde los organismos crecieron entre 0.26-0.38 mm día-1, seguido por Brazo Norte con 0.27-0.33 mm día-1 y Cueva con 0.25-0.30 mm día-1. Se observó que el crecimiento disminuyó gradualmente en organismos mayores a 150 mm en estas tres localidades. En Bocana el crecimiento de organismos menores a 160 mm mostró variabilidad y disminuyó gradualmente en organismos mayores a 160 mm. 89 Figura 34. Crecimiento medio (mm día-1) del caracol rosa S. gigas por clase de talla y desviación estándar, en los sitios (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, durante el periodo de abril 2009 a septiembre 2011. 90 En la tabla 16 se muestran los resultados del análisis de varianza de las tasas de crecimiento medio del caracol rosa, entre los sitios Bocana, Centro y Cueva en el periodo de enero 2005 a enero 2008. Se detectaron diferencias significativas (p<0.05) entre sitios en 9 de 15 clases de tallas. Las clases 70-79 mm, 80-89 mm y 100-109 mm no fueron comparadas, ya que se presentaron sólo en uno de los tres sitios estudiados. Tabla 16. Análisis de varianza (α=0.05) de las tasas de crecimiento medio (mm día⁻¹) del caracol S. gigas entre los sitios Bocana, Centro y Cueva, obtenidas en el periodo de enero 2005 a enero 2008. Clase (mm) F gl p 70-79 80-89 90-99 100-109 110-119 120-129 130-139 140-149 150-159 160-169 170-179 180-189 190-199 200-209 210-219 220-229 230-239 > 239 nd nd 0.18 nd 2.42 4.73 6.45 2.25 10.71 7.61 6.50 6.08 7.69 4.87 0.36 5.93 0.11 1.10 nd nd 1; 15 nd 2; 47 2; 71 2; 92 2; 137 2; 146 2; 164 2; 144 2; 138 2; 226 2; 225 2; 178 2; 106 2; 46 1; 7 nd nd 0.6774 nd 0.1004 0.0119* 0.0024* 0.1096 < 0.0001* 0.0007* 0.0020* 0.0030* 0.0006* 0.0085* 0.6957 0.0036* 0.8926 0.3353 *F- Estadístico de Fisher *p- Valor de p *gl- Grados de libertad *nd- no definido **- Diferencias estadísticas significativas En la tabla 17 se muestran los resultados del análisis de varianza de las tasas de crecimiento medio del caracol rosa, entre los sitios Bocana, Centro, Brazo Norte y Cueva, en el periodo de abril 2009 a septiembre 2011. Se detectaron diferencias significativas (p<0.05) entre sitios en 12 de 18 clases de tallas. 91 Tabla 17. Análisis de varianza (α=0.05) de las tasas de crecimiento medio (mm día⁻¹) del caracol S. gigas entre los sitios Bocana, Centro, Brazo Norte y Cueva, obtenidas en el periodo de abril 2009 a septiembre 2011. Clase (mm) F gl p 70-79 80-89 90-99 100-109 110-119 120-129 130-139 140-149 150-159 160-169 170-179 180-189 190-199 200-209 210-219 220-229 230-239 > 239 3.75 3.95 6.75 0.58 1.04 0.39 6.66 2.2 14.5 18.55 7.48 6.72 12.25 15.68 7.11 6.48 1.46 2.12 2; 31 3; 43 2; 70 3; 72 3; 135 3; 184 3; 248 3; 300 3; 409 3; 475 3; 474 3; 566 3; 511 3; 497 3; 356 3; 184 3; 40 3; 7 0.0356* 0.0147* 0.0021* 0.63 0.3789 0.7597 0.0002* 0.0883 <0.0001* <0.0001* 0.0001* 0.0002* <0.0001* <0.0001* 0.0001* 0.0003* 0.2418 0.2406 *F- Estadístico de Fisher *p- Valor de p *gl- Grados de libertad **- Diferencias estadísticas significativas En la figura 35 se muestra el crecimiento medio por día del caracol rosa por clases de talla obtenido en la caleta de Xel-Há durante el periodo 2005-2008 y 2009-2011. En el periodo 20052008 el crecimiento fue máximo en los organismos con una talla entre 70-79 mm con un incremento de 0.64 mm día -1 y disminuyó en los organismos con una longitud de 80 mm a 109 mm. El crecimiento fue relativamente constante en organismos con una talla entre 110 mm y 169 mm, observándose un incremento entre 0.20-0.24 mm día-1. El crecimiento disminuyó gradualmente en organismos mayores a 160 mm. En el periodo 2009-2011 el crecimiento incrementó gradualmente en los organismos menores a 110 mm y fue máximo en las clases de talla 110-119 mm y 120-129 mm, con una tasa de crecimiento de 0.32 mm día-1 respectivamente y disminuyó en las siguientes clases de talla. El crecimiento mostró diferencias estadísticas 92 significativas (p< 0.05) entre periodos en las clases de talla 70-79 mm, 80-89 mm y desde la clase de 110-119 mm hasta 190-199 mm (Tabla 18). Figura 35. Crecimiento medio (mm día-1) del caracol rosa S. gigas por clase de talla y desviación estándar, en la caleta de Xel-Há, durante el periodo de enero 2005 a enero 2008 (negro) y de abril 2009 a septiembre 2009 (gris). 93 Tabla 18. Análisis de varianza (α=0.05) de las tasas de crecimiento medio (mm día⁻¹) del caracol S. gigas en la caleta de Xel-Há, entre los periodos de estudio 2005-2008 y 2009-2011. Clase (mm) F gl p 70-79 80-89 90-99 100-109 110-119 120-129 130-139 140-149 150-159 160-169 170-179 180-189 190-199 200-209 210-219 220-229 230-239 > 239 10.46 8.12 2.33 2.81 19.57 29.98 19.53 10.75 13.34 4.04 5.35 4.54 4.17 0.96 0.01 1.64 0.02 0.71 1; 32 1; 47 1; 86 1;95 1; 182 1; 126 1;341 1; 438 1; 556 1; 640 1; 618 1; 705 1; 738 1; 723 1;535 1; 291 1; 87 1; 15 0.0029* 0.0065* 0.1303 0.0971 <0.0001* <0.0001* <0.0001* 0.0011* 0.0003* 0.045* 0.021* 0.0334* 0.0415* 0.3286 0.9345 0.2008 0.883 0.4121 *F- Estadístico de Fisher *p- Valor de p *gl- Grados de libertad **- Diferencias estadísticas significativas En la figura 36 se muestra el incremento medio en longitud de heliconcha (mm) y la estimación de la edad del caracol S. gigas en la caleta de Xel-Há calculado a partir de las tasas de crecimiento medio (Fig. 34). En ambos periodos de estudio el crecimiento fue ligeramente superior para los caracoles del periodo 2005-2008 con respecto al periodo 2009-2011. En ambos casos se observó un aumento rápido en longitud hasta alcanzar los 200 mm, que corresponde a una edad estimada de 2.2 años. A partir de esta talla el crecimiento disminuyó y fue similar en ambos periodos estudiados. 94 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 250 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student 225 version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 200 version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 175 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version LH (mm) Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 150 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 125 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 100 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 75 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student 50 version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 25 version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 0 5.00 4.75 4.50 4.25 4.00 3.75 3.50 3.25 3.00 2.75 2.50 2.25 2.00 1.75 1.50 1.25 1.00 0.75 0.50 0.25 0.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version EdadStudent version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Crecimiento 2005-2008 Crecimiento Student version Student version Student version Student version Student version Student version Student version Student2009-2011 version Student version Student version Student version version Student versiondeStudent version Student version (LH) Student Student Student versionestimada Student version Student FiguraStudent 36. Incremento medio longitud de heliconcha delversion caracol rosaversion S. gigas y edad en años, en version la caleta de Xel-Há durante el periodo de enero 2005 a enero 2008 (negro) y de abril 2009 a septiembre 2011 (gris). Con la finalidad de determinar si el crecimiento exhibe variación estacional y en base de los resultados anteriormente obtenidos (Fig. 35), se calculó el crecimiento por intervalo de muestreo para todas las clases de talla que tuvieron un incremento mayor a 0.15 mm día-1, seleccionando las clases de talla de 60-69 mm hasta 170-179 mm. En las clases de talla de 60-69 mm, 70-79 mm, 80-89 mm el número de observaciones fue insuficiente para determinar variación temporal, por lo cual fueron excluidos del análisis. En la figura 37 se muestra la media y desviación estándar del crecimiento (mm día -1) de individuos de caracol rosa con tallas entre 90-99 mm, en el periodo de enero 2005 a septiembre 2011, en los sitios Cueva, Centro y Brazo Norte. Bocana no se tomó en cuenta, dado que el número de observaciones en esta clase de talla fue insuficiente para su interpretación. Se calculó el aumento medio por día entre dos muestreos consecutivos. En Cueva las mayores tasas de crecimiento fueron observados entre octubre-diciembre 2005, septiembre-noviembre 2006, febrero-mayo 2010 y enero-marzo 2011. En Centro la tasa de crecimiento más alta fue observada 95 en el intervalo febrero-mayo 2010. En Brazo Norte la tasa más alta fue observada en el intervalo octubre 2009-febrero 2010. Figura 37. Media y desviación estándar del crecimiento (mm día-1) de individuos de S. gigas con tallas entre 90-99 mm, en los sitios Cueva, Centro y Brazo Norte (Caleta de Xel-Há), durante el periodo enero 2005 a septiembre 2011. 96 En la figura 38 se muestra la media y desviación estándar del crecimiento (mm día -1) de individuos de caracol rosa con tallas entre 100-109 mm, en los sitios Cueva, Centro y Brazo Norte (Caleta de Xel-Há), durante el periodo enero 2005 a septiembre 2011. Bocana no se tomó en cuenta, dado que el número de observaciones en esta clase de talla fue insuficiente para su interpretación. Se calculó el aumento medio por día entre dos muestreos consecutivos. En Cueva el crecimiento más alto fue observado en el intervalo agosto-septiembre 2005. En Centro sólo se obtuvieron 3 medias, la más alta fue observado entre febrero-mayo 2010. En Brazo Norte el crecimiento máximo fue observado en el intervalo julio-septiembre 2010. 97 Figura 38. Media y desviación estándar del crecimiento (mm día -1) de individuos de S. gigas con tallas entre 100109 mm, en los sitios Cueva, Centro y Brazo Norte (Caleta de Xel-Há), durante el periodo enero 2005 a septiembre 2011. En la figura 39 se muestra la media y desviación estándar del crecimiento (mm día -1) de individuos de caracol rosa con tallas entre 110-119 mm, en los sitios estudiados Cueva, Centro y Brazo Norte (Caleta de Xel-Há), durante el periodo enero 2005 a septiembre 2011. Bocana no se 98 tomó en cuenta, dado que el número de observaciones en esta clase de talla fue insuficiente para su interpretación. En Cueva se observaron tasas mayores o iguales a 0.4 mm día -1 en los intervalos de muestreo enero-febrero 2005, julio-agosto 2005, septiembre-noviembre 2006, febrero-mayo 2010 y marzo-mayo 2011. En Centro se observaron picos en el crecimiento entre los meses julio-agosto 2005 y febrero-mayo 2010. En Bocana sólo se obtuvieron 3 observaciones y el crecimiento más alto fue registrado entre febrero y mayo 2010. En Brazo Norte el crecimiento fue mayor entre marzo y mayo del 2011. 99 Figura 39. Media y desviación estándar del crecimiento (mm día-1) de individuos de S. gigas con tallas entre 110119 mm, en los sitios Cueva, Centro y Brazo Norte (Caleta de Xel-Há), durante el periodo enero 2005 a septiembre 2011. En la figura 40 se muestra la media y desviación estándar del crecimiento (mm día-1) de individuos de caracol rosa con tallas entre 120-129 mm, en los sitios estudiados Cueva, Centro y Brazo Norte (Caleta de Xel-Há), durante el periodo enero 2005 a septiembre 2011. Bocana no se 100 gráfico, dado que el número de observaciones en esta clase de talla fue insuficiente para su interpretación. En Cueva se observaron dos picos, con mayor crecimiento entre febrero-marzo 2005, septiembre-octubre 2005, septiembre-noviembre 2006, febrero-mayo 2010 y marzo-mayo 2011. En Centro y Brazo Norte el crecimiento fue relativamente constante a través del periodo estudiado. 101 Figura 40. Media y desviación estándar del crecimiento (mm día-1) de individuos de S. gigas con tallas entre 120129 mm, en los sitios Cueva, Centro y Brazo Norte (Caleta de Xel-Há), durante el periodo enero 2005 a septiembre 2011. En la figura 41 se muestra la media y desviación estándar del crecimiento (mm día -1) de individuos de caracol rosa con tallas entre 130-139 mm, en cada uno de los cuatro sitios estudiados (Cueva, Centro, Bocana, Brazo Norte) de la caleta de Xel-Há, durante el periodo 102 enero 2005 a septiembre 2011. En Cueva se observaron oscilaciones del crecimiento. Las tasas mayores fueron registradas entre febrero y marzo del 2005, septiembre y noviembre del 2006 y febrero y mayo del 2010. En Centro el crecimiento también mostró un pico entre octubre y diciembre del 2005. Para la Bocana se tienen pocas observaciones en este rango de tallas, sin embargo la tasa de crecimiento ha aumentado para estos organismos en esta localidad de febrero 2005 a marzo 2011, pasando de 0.02 a 0.2 mm díaˉ¹. En Brazo Norte el crecimiento fue relativamente constante a través del periodo estudiado. 103 Figura 41. Media y desviación estándar del crecimiento (mm día-1) de individuos de S. gigas con tallas entre 130139 mm, en cada uno de los cuatro sitios estudiados (Cueva, Centro, Bocana, Brazo Norte) de la caleta de Xel-Há, durante el periodo enero 2005 a septiembre 2011. 104 En la figura 42 se muestra la media y desviación estándar del crecimiento (mm día-1) de individuos de caracol rosa con tallas entre 140-149 mm, en cada uno de los cuatro sitios estudiados (Cueva, Centro, Bocana, Brazo Norte) de la caleta de Xel-Há, durante el periodo enero 2005 a septiembre 2011. Se observó un pico en el crecimiento entre febrero y marzo del 2005 en los sitios Cueva, Centro y Bocana. En Bocana y Brazo Norte el crecimiento fue relativamente constante a través del periodo de estudio, mientras que en Cueva y Centro presentó oscilaciones. 105 Figura 42. Media y desviación estándar del crecimiento (mm día-1) de individuos de S. gigas con tallas entre 140149 mm, en cada uno de los cuatro sitios estudiados (Cueva, Centro, Bocana, Brazo Norte) de la caleta de Xel-Há, durante el periodo enero 2005 a septiembre 2011. 106 En la figura 43 se muestra la media y desviación estándar del crecimiento (mm día -1) de individuos de caracol rosa con tallas entre 150-159 mm, en cada uno de los cuatro sitios estudiados (Cueva, Centro, Bocana, Brazo Norte) de la caleta de Xel-Há, durante el periodo enero 2005 a septiembre 2011. Se observó un pico en el crecimiento entre febrero y marzo del 2005 en los sitios Cueva, Centro y Bocana. El crecimiento fue relativamente constante durante el resto del periodo en Brazo Norte, con excepción de septiembre 2010. Para las otras tres localidades las tasas de crecimiento fluctuaron a través del tiempo sin aparente relación estacional. 107 Figura 43. Media y desviación estándar del crecimiento (mm día-1) de individuos de S. gigas con tallas entre 150159 mm, en cada uno de los cuatro sitios estudiados (Cueva, Centro, Bocana, Brazo Norte) de la caleta de Xel-Há, durante el periodo enero 2005 a septiembre 2011. 108 En la figura 44 se muestra la media y desviación estándar del crecimiento (mm día -1) de individuos de caracol rosa con tallas entre 160-169 mm, en cada uno de los cuatro sitios estudiados (Cueva, Centro, Bocana, Brazo Norte) de la caleta de Xel-Há, durante el periodo enero 2005 a septiembre 2011. En Cueva se observaron tasas altas en el intervalo de febrero a marzo 2005 y febrero a mayo 2010. En Centro se registraron tasas de crecimiento elevado entre abril y mayo 2005 y en el intervalo de noviembre 2010 a enero 2011. En Bocana el crecimiento fue bajo en comparación con los demás sitios y relativamente constante a lo largo del periodo estudiado. En Brazo Norte se observó crecimiento alto entre noviembre 2010 y enero 2011. 109 Figura 44. Media y desviación estándar del crecimiento (mm día-1) de individuos de S. gigas con tallas entre 160169 mm, en cada uno de los cuatro sitios estudiados (Cueva, Centro, Bocana, Brazo Norte) de la caleta de Xel-Há, durante el periodo enero 2005 a septiembre 2011. 110 En la figura 45 se muestra la media y desviación estándar del crecimiento (mm día -1) de individuos de caracol rosa con tallas entre 170-179 mm, en cada uno de los cuatro sitios estudiados (Cueva, Centro, Bocana, Brazo Norte) de la caleta de Xel-Há, durante el periodo enero 2005 a septiembre 2011. En Cueva y Centro se observaron tasas altas en el intervalo de febrero a marzo 2005. En Bocana el crecimiento fue bajo en comparación con los demás sitios estudiados y relativamente constante a lo largo del periodo de estudio. En Brazo Norte el crecimiento más alto fue registrado entre noviembre 2010 y septiembre 2010. La menor tasa de crecimiento se observó en septiembre 2010 en las localidades Centro, Brazo Norte y Bocana, siendo la excepción Cueva. 111 Figura 45. Media y desviación estándar del crecimiento (mm día-1) de individuos de S. gigas con tallas entre 170179 mm, en cada uno de los cuatro sitios estudiados (Cueva, Centro, Bocana, Brazo Norte) de la caleta de Xel-Há, durante el periodo enero 2005 a septiembre 2011. 112 8.2.2 Estimación de los parámetros de crecimiento de la ecuación de Von Bertalanffy Se determinaron los parámetros de crecimiento de la ecuación de Von Bertalanffy mediante el método de Appeldoorn (1987b). En las figuras 46.A y 46.B se muestra el crecimiento para el caracol S. gigas en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011, respectivamente. La tasa se calculó utilizando el método directo, utilizando los incrementos obtenidos mediante el muestreo de marcaje recaptura. En el 2005-2008, el crecimiento fue menor con un L∞= 280.51 mm y un k de 0.57 año-1 (r2 = 0.3150) en comparación con la tasa de crecimiento en el periodo 2009-2011 (L∞= 278.73 mm; k= 0.71 año -1; r2=0.6138). Fue posible determinar oscilación estacional (C) de la tasa de crecimiento para el periodo 2009-2011, la cual fue de una magnitud de 0.24. El punto invernal (WP) fue estimado en 0.25. Figura 46. Crecimiento del caracol rosa S. gigas en el periodo (A) 2005-2008, (B) 2009-2011, en la caleta Xel-Há obtenido mediante el método de Appeldoorn, utilizando incrementos obtenidos a partir de los datos de marcaje recaptura. 113 En las figuras 47.A y 47.B se muestra el crecimiento del caracol rosa S. gigas en la caleta de Xel-Há, para el periodo 2005-2008 y 2009-2011, respectivamente. Se utilizaron los incrementos de longitud obtenidos a partir del análisis de progresión de modas. Para el primer periodo se estimó una tasa de crecimiento de k= 0.32 año-1 y un L∞= 285.16 mm (r2 = 0.9661). Se obtuvo una oscilación de 0.61 y un WP= 0.11. El crecimiento fue mayor en el segundo periodo (L∞= 290.51 mm; k= 0.55 año -1; r2=0.8528), con respecto al 2005-2008. La oscilación estacional (C) de la tasa de crecimiento fue de 0.33 y el punto invernal fue de 0.09. Figura 47. Crecimiento del caracol rosa S. gigas en el periodo (A) 2005-2008, (B) 2009-2011, en la caleta Xel-Há obtenido mediante el método de Appeldoorn, utilizando incrementos obtenidos de la progresión de modas. En las figuras 48.A y 48.B se muestra el crecimiento del grosor de labio del caracol rosa en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011, respectivamente La tasa se calculó utilizando el método directo, utilizando los incrementos obtenidos mediante el muestreo de marcaje recaptura. Para el periodo 2005-2008 se obtuvo un L∞= 99.55 mm y un k= 0.27, con un r2= 0.5457. Para el periodo de 2009-2011 se obtuvo un crecimiento menor para el grosor de labio con un k= 0.05 (r2= 0.0995) y un L∞= 301.10 mm. Para ambas curvas se 114 determinó oscilación estacional. En el 2005-2008 se obtuvo una oscilación C = 0.19 y el punto invernal (WP) fue de 1. Para el 2009-2011 se obtuvo un C= 0.47 y un WP= 0.27. Figura 48. Crecimiento del grosor de labio del caracol rosa S. gigas en el periodo (A) 2005-2008 y (B) 2009-2011, en la caleta Xel-Há obtenidas mediante el método de Appeldoorn, utilizando incrementos obtenidos a partir de los datos de marcaje recaptura. . 115 8.3 Dinámica Poblacional 8.3.1 Reclutamiento En las figuras 49.A a 49.F se muestran histogramas de la frecuencia absoluta de longitud de heliconcha de caracoles juveniles para cada mes estudiado del año 2005 (Fig. 49.A), 2006 (Fig. 49.B), 2007-2008 (Fig. 49.C), 2009 (Fig. 49.D), 2010 (Fig. 49.E) y 2011 (Fig. 49.F), respectivamente. En el 2005 se observó la aparición de organismos <100 mm en todos los meses estudiados con excepción de febrero. El reclutamiento de juveniles fue máximo de agosto a octubre. En el 2006 el reclutamiento de organismos <100 mm fue alto en todo los meses estudiados, con máximo en mayo. En el 2007 se observó reclutamiento en enero, pero la incorporación de organismos <100 mm fue baja a lo largo del año y nulo en enero 2008. En el 2009 se observaron organismos < 100 mm en todos los meses estudiados y su abundancia fue alta en junio y octubre. En el 2010 sólo se observaron reclutas en febrero y pocos en julio. En el 2011 se observó baja abundancia de juveniles en todos los meses estudiados. 116 Figura 49.A Frecuencia absoluta de longitud de heliconcha por clase de talla, de caracoles juveniles S. gigas en la caleta de Xel-Há, en el periodo de enero 2005 a diciembre 2005. 117 Figura 49.B Frecuencia absoluta de longitud de heliconcha por clase de talla, de caracoles juveniles S. gigas en la caleta de Xel-Há, en el periodo de febrero 2006 a noviembre 2006. 118 Figura 49.C Frecuencia absoluta de longitud de heliconcha por clase de talla, de caracoles juveniles S. gigas en la caleta de Xel-Há, en el periodo de enero 2007 a enero 2008. 119 Figura 49.D Frecuencia absoluta de longitud de heliconcha por clase de talla, de caracoles juveniles S. gigas en la caleta de Xel-Há, en el periodo de abril 2009 a octubre 2009. 120 Figura 49.E Frecuencia absoluta de longitud de heliconcha por clase de talla, de caracoles juveniles S. gigas en la caleta de Xel-Há, en el periodo de febrero 2010 a noviembre 2010. 121 Figura 49.F Frecuencia absoluta de longitud de heliconcha por clase de talla, de caracoles juveniles S. gigas en la caleta de Xel-Há, en el periodo de enero 2011 a septiembre 2011. 122 En las figuras 50.A y 50.B se muestra el porcentaje de reclutamiento de juveniles de S. gigas en la Caleta de Xel-Há, determinado por el método de análisis electrónico de frecuencia de longitud, en el periodo 2005-2008 y 2009-2011, respectivamente. En ambos periodos de estudio se observaron dos pulsos de reclutamiento de juveniles. El reclutamiento fue continúo a lo largo del del año con excepción de diciembre. Para el periodo 2005-2008 el máximo reclutamiento se observó entre junio y agosto. El segundo pico de menor magnitud se registró entre Abril y Mayo. En el segundo periodo de estudio el máximo reclutamiento se observó entre junio y septiembre y el segundo pulso de menor magnitud fue registrado entre marzo y mayo. Figura 50. Porcentaje de reclutamiento de juveniles de S. gigas en la Caleta de Xel-Há, durante el periodo de (A) enero 2005 a enero 2008 y (B) abril 2009 a septiembre 2011, determinado por el método de análisis electrónico de frecuencia de longitud. 8.3.2 Sobrevivencia y Mortalidad En las figuras 51.A y 51.B se muestra el porcentaje medio de la sobrevivencia finita (St=(Nt0/Nti)*100) del caracol rosa S. gigas en los cuatro sitios estudiados de la caleta de Xel-Há (Cueva, Centro, Bocana y Brazo Norte), en el periodo enero 2005 a enero 2008 y abril 2009 a 123 septiembre 2011, respectivamente. Brazo Norte sólo fue estudiado en el periodo 2009-2011. No se observaron diferencias significativas en la sobrevivencia entre periodos, sin embargo hubo diferencias entre los sitios. Las menores tasas de sobrevivencia se observaron en ambos periodos en Bocana y Centro. Las mayores sobrevivencias se obtuvieron en Cueva y Brazo Norte. A partir de los 450 días después del marcado inicial de los caracoles, las sobrevivencias fueron iguales en los cuatro sitios con menos del 10% de sobrevivencia. Figura 51. Porcentaje medio y desviación estándar de la sobrevivencia finita (St=(Nt0/Nti)*100) del caracol rosa S. gigas en los cuatro sitios estudiados de la caleta de Xel-Há (Cueva, Centro, Bocana y Brazo Norte) en el periodo de (A) enero 2005 a enero 2008 y (B) abril 2009 a septiembre 2011. En la figura 52 se muestra el porcentaje medio de la sobrevivencia finita (St=(Nt0/Nti)*100) del caracol rosa S. gigas en la caleta de Xel-Há, en el periodo de 2005-2008 y 2009-2011. Las tasas 124 de sobrevivencia fueron ligeramente mayores en el periodo 2009-2011, con respecto al 20052008. En ambos periodos la sobrevivencia fue menor al 5% a partir de los 600 días después del marcado inicial. Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 50 45 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 40 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 35 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 30 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 25 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student 20 version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 15 version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 10 version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 5 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 0 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 1050 1000 950 900 850 800 750 700 650 600 550 500 450 400 350 300 250 200 150 100 50 0 % Sobrevivencia Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Días Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 2005-2008 2009-2011 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Figura 52. Porcentaje medio y desviación estándar de sobrevivencia finita (St=(Nt0/Nti)*100) del caracol rosa S. gigas en los cuatro sitios estudiados de la caleta de Xel-Há (Cueva, Centro, Bocana y Brazo Norte) en el periodo de (A) enero 2005 a enero 2008 y (B) abril 2009 a septiembre 2011. En la figura 53 se muestran las tasas de mortalidad finita (zt) del caracol rosa S. gigas en la caleta de Xel-Há en el periodo 2005-2008 y 2009-2011, respectivamente. La mortalidad media en el 2005-2008 varía entre 1.03 y 3.0, mientras en 2009-2011 varía entre 1.06 y 3.87. En el periodo 2009-2011 la mortalidad incrementó linealmente con el tiempo, mientras en el 2005-2008 incrementó de forma lineal hasta los 1.5 años después de la fecha del marcado inicial y posteriormente se mantuvo casi constante. Para el 2009-2011 se observaron mortalidades mayores entre los 1.5 años y 2.5 años con respeto al periodo anterior. 125 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 5 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 4 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 3 Zt Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 2 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 1 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 0 0.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 0.75 1.50 Años Student version 2.25 3.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 2005-2008 2009-2011 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Figura 53. Mortalidades finitas (zt=(-ln (Nt/N0)) medias y desviación estándar, del caracol rosa S. gigas en la Caleta de Xel-Há en el periodo de enero 2005 a enero 2008 (cuadrado) y abril 2009 a septiembre 2011 (círculo). En la figura 54 se muestra el número de individuos del caracol rosa S. gigas, que fueron encontrados muertos en cada uno de los cuatro sitios estudiados de la caleta de Xel-Há (Cueva, Centro, Bocana y Brazo Norte) en el periodo de enero 2005 a septiembre 2011. Se encontraron 54 individuos marcados muertos a lo largo del periodo de estudio. La mayoría de éstos se hallaron en el 2006 y en el 2010. El mes con mayor mortalidad fue julio 2007. El mayor número de caracoles muertos fue recuperado en Cueva, donde se hallaron 38. En Brazo Norte se encontraron 6 individuos, 5 en Centro y 4 en Bocana. 126 12 No. Individuos 10 8 Bocana 6 Centro 4 Brazo Norte Cueva 2 Abr-09 Jun-09 Oct-09 Feb-10 May-10 Jul-10 Sep-10 Nov-10 Ene-11 Mar-11 May-11 Jul-11 Sep-11 Ene-05 Feb-05 Mar-05 Abr-05 May-05 Jun-05 Jul-05 Ago-05 Sep-05 Oct-05 Dic-05 Feb-06 May-06 Jul-06 Sep-06 Nov-06 Ene-07 Mar-07 May-07 Jul-07 Sep-07 Nov-07 Ene-08 0 Figura 54. Número de individuos del caracol rosa S. gigas, que fueron encontrados muertos en los cuatro sitios estudiados de la caleta de Xel-Há (Cueva, Centro, Bocana y Brazo Norte) a través de los periodos de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. En las figuras 55.A y 55.B se muestra la frecuencia relativa (%) de longitudes de heliconcha por clases de talla del caracol rosa S. gigas sin labio y con labio en la caleta de Xel-Há, en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011, respectivamente. En el primer periodo se obtuvieron 5074 medidas de longitud, 4072 de organismos sin labio y 1002 de caracoles con labio. En el segundo periodo se obtuvieron 14242 medidas, 9164 de caracoles sin labio y 5078 de caracoles con labio. En ambos periodos se observó una distribución normal sesgada. La clase modal en ambos periodos fueron los caracoles con una longitud de 200-209 mm. La mayor frecuencia de organismos sin labio se observó en esta clase de talla en el periodo 2005-2008. En el 2009-2011 la clase de mayor frecuencia de organismos sin labio fueron los organismos con una talla de 180-189 mm. La clase modal para organismos con labio fue en ambos periodos la clase de 210-219 mm. La frecuencia de observaciones disminuyó marcadamente a partir de la clase de 200-209 mm en ambos periodos. 127 Figura 55. Frecuencia relativa (%) de longitudes de heliconcha (mm) por clases de talla, del caracol rosa S. gigas sin labio (blanco) y con labio (negro), en la caleta de Xel-Há en el periodo de (A) enero 2005 a enero 2008 y de (B) abril 2009 a septiembre 2011. En las figuras 56.A y 56.B se muestra la frecuencia relativa (%) del grosor de labio del caracol rosa S. gigas por clases de talla en la caleta de Xel-Há, en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011, respectivamente. En el primer periodo se obtuvieron 1002 128 medidas y en el segundo periodo 5078 mediciones del grosor de labio. En el primer periodo se obtuvieron medidas de 1mm hasta 44 mm. Se observó una distribución bimodal. La clase con mayor frecuencia fueron los caracoles con un grosor de labio de 5-6 mm. Las frecuencias de clases no disminuyeron hasta en los organismos con un grosor de labio de 31-32 mm. En el periodo 2009-2011 se observaron caracoles con un grosor de labio de 1 mm hasta 54 mm. La clase con mayor frecuencia fueron los caracoles con un grosor de labio de 1-2 mm y a partir de esta clase se observó una disminución gradual en las frecuencias de clase. 129 Figura 56. Frecuencia relativa (%) del grosor de labio (mm) por clases de talla del caracol rosa S.giga, en la caleta de Xel-Há en el periodo de (A) enero 2005 a enero 2008 y de (B) abril 2009 a septiembre 2011. En las figuras 57.A y 57.B se muestran curvas de tallas convertidas (edad relativa) de la distribución de frecuencia de longitud de heliconcha de los caracoles de la caleta de Xel-Há, en el periodo 2005-2008 y 2009-2011 respectivamente, las cuales fueron utilizadas para determinar 130 la mortalidad. Para el periodo 2005-2008 se estimó una mortalidad z= 1.46 entre los 2.08 y 6.89 años de edad relativa. Se observó que la mortalidad no fue constante. La mayor mortalidad se registró entre los 2.08 y 3.36 años de edad relativa, con un z= 2.40. Para los organismos con una edad relativa entre 4.21 y 6.89 años la mortalidad fue muy baja con una tasa de z= 0.55. Para el periodo 2009-2011 se obtuvo una mortalidad instantánea (z) de de 4.71, la cual se manifestó entre los 1.7 y 3.68 años de edad relativa. Figura 57. Curvas de captura de tallas convertidas (edad relativa), obtenidas de la frecuencia de distribución de longitud de heliconcha del caracol S. gigas en el periodo de (A) enero 2005 a enero 2008 de y (B) abril 2009 a septiembre 2011, en la caleta de Xel-Há. En las figuras 58.A y 58.B se muestran las curvas de tallas convertidas de la distribución de frecuencia del grosor de labio del caracol rosa en la caleta de Xel-Há para el periodo 2005-2008 y 2009-2011, respectivamente, las cuales fueron utilizadas para determinar la mortalidad. Para el periodo 2005-2008 se obtuvo una mortalidad de z= 2.39 entre los 1.14 y 2.57 años de edad relativa, con respecto a la formación del labio, y para el periodo 2009-2011 la mortalidad fue estimada en z= 1.97, la cual fue estimada para los 0.86-4.17 años de edad relativa. 131 Figura 58. Curvas de captura de tallas convertidas (edad relativa), obtenidas de la frecuencia de distribución del grosor de labio del caracol S. gigas en el periodo de (A) enero 2005 a enero 2008 de y (B) abril 2009 a septiembre 2011, en la caleta de Xel-Há. 132 8.4 Movilidad 8.4.1 Dirección y frecuencia de transiciones En las figuras 59.A a 59.D se muestra el porcentaje de organismos sin marcas y con marcas, así como el origen de las marcas, en cada uno de los cuatro sitios de muestreo de la caleta de XelHá: Cueva (Fig. 59.A), Centro (Fig. 59.B), Bocana (Fig. 59.C) y Brazo Norte (Fig. 59.D), en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. En Brazo Norte sólo se realizaron muestreos en el segundo periodo de estudio. Se observó en los cuatro sitios, que la mayoría de los organismos encontrados fueron animales sin marcas o animales marcados en el propio sitio. En Cueva y Brazo Norte el porcentaje de caracoles provenientes de otros sitios no superó el 5%. En Centro hasta el 20% de los caracoles capturados fueron originarios de otros sitios y la mayoría de éstos provinieron de Cueva y Brazo Norte y pocos fueron de Bocana. En Bocana hasta el 35% de los organismos capturados fueron organismos cuyo origen proviene de otros sitios (Centro, Cueva y Brazo Norte). 133 Figura 59. Abundancia relativa (%) de caracoles S. gigas marcados en Cueva (azul), Centro (verde), Bocana (morado) o Brazo Norte (rojo) y sin marca (SM), capturados en los cuatro sitios estudiados de la caleta de Xel-Há (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. 134 En las figuras 60.A a 60.D se muestra el número de individuos cuyo origen de su marca fueron los cuatro sitios de estudio de la caleta de Xel-Há, Cueva (Fig. 60.A), Centro (Fig. 60.B), Bocana (Fig. 60.C) y Brazo Norte (Fig. 60.D), que emigraron de su sitio de origen y los destinos de sus migraciones, a través del tiempo. En Brazo Norte sólo se realizaron muestreos en el segundo periodo de estudio. En Cueva se marcaron 2559 individuos durante el periodo estudiado, de los cuales 366 fueron recapturados en otro sitio; 214 se encontraron en Centro, 140 en Bocana y 12 en Brazo Norte. No se registró la emigración de individuos de Cueva a otros sitios hasta febrero 2006. El mayor número de desplazamientos fue observado en septiembre 2006, septiembre 2007 y enero 2008. Para el periodo 2009-2011 se observaron oscilaciones en el número de desplazamientos con picos en julio 2010 y septiembre 2011. Los organismos marcados en Centro mostraron el mayor número de desplazamientos en comparación con los caracoles marcados en los otros sitios, con 1064 emigraciones, de los cuales 977 tuvieron como destino la Bocana, 54 Brazo Norte y 33 Cueva. De los organismos marcados en Bocana solo 30 caracoles realizaron migraciones a los sitios del interior de la caleta; 25 organismos fueron recapturados en Centro, 3 en Brazo Norte y 2 en Cueva. De los organismos marcados en Brazo Norte un total de 245 emigró. La mayoría de los organismos tuvieron como destino el Centro (126 individuos) y la Bocana (108 caracoles). En los sitios Cueva, Centro y Brazo Norte se observaron picos en el número de transiciones a través del tiempo, generalmente con aumento en la actividad de los caracoles entre los meses de marzo y septiembre. 135 Figura 60. Número de individuos de caracol S. gigas marcados en (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, que emigraron de su sitio de origen, mostrando el destino de migración en la caleta de Xel-Há: Cueva (azul), Centro (verde), Bocana (morado) y Brazo Norte (rojo), a través del periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. 136 En la figura 61 se muestra el porcentaje de emigraciones de S. gigas marcados en Cueva durante la estación de Nortes (noviembre-febrero), Secas (marzo-junio) y Lluvias (julio-octubre), en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. El 57% de los caracoles realizó migraciones durante la estación de Lluvias. El 27% de las migraciones ocurrió en la estación de Secas y el 16% restante durante la estación de Nortes. Figura 61. Porcentaje de emigraciones de S. gigas marcados en la Cueva (Caleta de Xel-Há) por estación del año (Nortes, Secas y Lluvias), en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. 137 En la figura 62 se muestra el porcentaje de emigraciones de S. gigas marcados en Centro durante la estación de Nortes (noviembre-febrero), Secas (marzo-junio) y Lluvias (julio-octubre), en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. El 47% de los caracoles realizó migraciones durante la estación de Lluvias. El 25% de las migraciones ocurrió en la estación de Secas y el 28% restante durante la estación de Nortes. Figura 62. Porcentaje de emigraciones de S. gigas marcados en la Centro (Caleta de Xel-Há) por estación del año (Nortes, Secas y Lluvias), en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. 138 En la figura 63 se muestra el porcentaje de emigraciones de S. gigas marcados en Bocana durante la estación de Nortes (noviembre-febrero), Secas (marzo-junio) y Lluvias (julio-octubre), en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. El 42% de los caracoles realizó migraciones durante la estación de Nortes. El 39% de las migraciones ocurrió en la estación de Lluvias y el 19% durante la estación de Secas. Figura 63. Porcentaje de emigraciones de S. gigas marcados en la Bocana (Caleta de Xel-Há) por estación del año (Nortes, Secas y Lluvias), en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. 139 En la figura 64 se muestra el porcentaje de emigraciones de S. gigas marcados en Brazo Norte durante la estación de Nortes (noviembre-febrero), Secas (marzo-junio) y Lluvias (julio-octubre), en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. El 49% de las migraciones ocurrió en la estación de Lluvias, un 32% durante la estación de Secas y el 18% de los caracoles realizaron migraciones durante la estación de Nortes. Figura 64. Porcentaje de emigraciones de S. gigas marcados en la Brazo Norte (Caleta de Xel-Há) por estación del año (Nortes, Secas y Lluvias), en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. 140 8.4.2 Talla de transición En las figuras 65.A a 65.D se muestran las longitudes medias de heliconcha y desviación estándar de caracoles S. gigas cuyo origen de su marca fueron los cuatro sitios estudiados de la caleta de Xel-Há: Cueva (Fig. 65.A), Centro (Fig. 65.B), Bocana (Fig. 65.C) y Brazo Norte (Fig. 65.D), los cuales emigraron de su sitio de origen a losdestinos de migración, a través del tiempo. Brazo Norte sólo fue estudiado en el segundo periodo de estudio. Los caracoles que emigraron de Cueva a otros sitios presentaron tallas medias entre 165 mm y 225 mm. Se observó que las tallas fueron similares independientemente del destino y no se detectaron diferencias estadísticas significativas (F0.05[2; 364] = 2.34; p = 0.0982) entre éstas. Los organismos que emigraron de Centro a otros sitios tuvieron tallas medias entre 195 mm y 236 mm, con excepción de dos organismos que migraron a la Cueva, cuyas tallas fueron de 131mm y 150 mm respectivamente. Sin embargo no se detectaron diferencias estadísticas significativas (F0.05[2; 1036] = 2.76; p = 0.0637) en la talla media y los diferentes sitios de recaptura. Para los caracoles emigraron de Bocana a otros sitios, sus tallas medias fueron entre 162 mm y 233 mm. No se detectaron diferencias estadísticas significativas (F0.05[2; 29] = 1.68; p = 0.2046) entre las tallas medias y el sitio de recaptura. Los organismos originarios en Brazo Norte y recapturados en otro sitio de la caleta tuvieron tallas medias de 185 mm a 223 mm. No se detectaron diferencias estadísticas significativas (F0.05[2; 244] = 0.70; p = 0.4967) entre las tallas medias en los distintos sitios de recaptura. 141 Figura 65. Longitudes medias de heliconcha y desviación estándar de caracoles S. gigas marcados en (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, que emigraron de su sitio de origen, mostrando el destino de migración en la caleta de Xel-Há: Cueva (azul), Centro (verde), Bocana (morado) y Brazo Norte (rojo), a través del periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. 142 En las figuras 66.A a 66.D se muestran las medias y desviación estándar del grosor de labio de caracoles S. gigas cuyo origen de su marca fueron los cuatro sitios estudiados de la caleta de XelHá: Cueva (Fig. 66.A), Centro (Fig. 66.B), Bocana (Fig. 66.C) y Brazo Norte (Fig. 66.D), los cuales emigraron de su sitio de origen a otro sitio de la caleta, en el periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011; con excepción de Brazo Norte, ya que sólo fue estudiado en el segundo periodo. Los organismos recapturados en Bocana generalmente tuvieron un grosor de labio mayor en comparación con los organismos recapturados en los otros sitios, independientemente de su procedencia. Se detectaron diferencias estadísticas significativas entre el grosor de labio medio y destinos de migración para caracoles marcadosen estos Cueva (F0.05[2; 274] = 36.40; p < 0.0001), Centro (F0.05[2; 819] = 24.76; p < 0.0001) y Brazo Norte (F0.05[2; 208] = 36.56; p <0.0001). En el caso de los caracoles marcados en Bocana no se detectaron éstas diferencias (F0.05[2; 16] = 0.60; p = 0.5649). 143 Figura 66. Medias y desviación estándar del grosor de labio de caracoles S. gigas marcados en (A) Cueva, (B) Centro, (C) Bocana y (D) Brazo Norte, que emigraron de su sitio de origen, mostrando el destino de migración en la caleta de Xel-Há: Cueva (azul), Centro (verde), Bocana (morado) y Brazo Norte (rojo), a través del periodo de enero 2005 a enero 2008 y de abril 2009 a septiembre 2011. 144 En la figura 67 se muestra el número de individuos marcados en Cueva y número de individuos que emigraron de ésta, así como el porcentaje de juveniles (caracoles sin labio), subadultos (caracoles con un grosor de labio < 5mm) y adultos (caracoles con un grosor de labio ≥ 5 mm), que participaron en las migraciones y su respectivo destino en la caleta (Centro, Bocana o Brazo Norte), en el periodo de enero 2005 a septiembre 2011.Se registró, de una población total marcada de 2559 caracoles, la emigración de 366 individuos. El 25% de los caracoles que participaron en estas migraciones fueron juveniles, 17% subadultos y el 58% adultos. El 84% de los juveniles fue recapturado en Centro y el 16% en Bocana. De los subadultos el 76% fue recapturado en Centro, el 22% en Bocana y el 2% en Brazo Norte. El 53% de los adultos se recapturó en Bocana, 42% en Centro y 5% en Brazo Norte. 145 Figura 67. Número de individuos marcados en Cueva, número de individuos que emigraron de Cueva, porcentaje de juveniles (caracoles sin labio), subadultos (caracoles con un grosor de labio < 5mm) y adultos (caracoles con un grosor de labio ≥ 5 mm) que participaron en las migraciones y destinos de migración: Centro (verde), Bocana (morado) o Brazo Norte (rojo), en el periodo de enero 2005 a septiembre 2011. En la figura 68 se muestra el número de individuos marcados en Centro, el número de individuos que emigraron de Centro, así como el porcentaje de juveniles (caracoles sin labio), subadultos (caracoles con un grosor de labio < 5mm) y adultos (caracoles con un grosor de labio ≥ 5 mm), que participaron en las migraciones y su destino en la caleta (Cueva, Bocana o Brazo Norte), en el periodo de enero 2005 a septiembre 2011.De Centro se observó la emigración de 1064 individuos de un total de 2806 individuos marcados en este sitio. El 23% de los organismos que participaron en estas migraciones fueron juveniles, un 5% fueron subadultos, pero la mayoría (72%) fuero a adultos. El 98% de los juveniles emigrados de Centro fue recapturado en Centro y 146 2% en Cueva. El 81% de los subadultos se encontró en Bocana, 9% en Cueva y 10% en Brazo Norte. La mayoría de los adultos fue recapturado en Bocana (91%) y un 3 y 6% en Cueva y Brazo Norte, respectivamente. Figura 68. Número de individuos marcados en Centro, número de individuos que emigraron de Centro, porcentaje de juveniles (caracoles sin labio), subadultos (caracoles con un grosor de labio < 5mm) y adultos (caracoles con un grosor de labio ≥ 5 mm) que participaron en las migraciones y destinos de migración: Cueva (azul), Bocana (morado) o Brazo Norte (rojo), en el periodo de enero 2005 a septiembre 2011. En la figura 69 se muestra el número de individuos marcados en Bocana, el número de individuos que emigraron de Bocana, así como el porcentaje de juveniles (caracoles sin labio), subadultos (caracoles con un grosor de labio < 5mm) y adultos (caracoles con un grosor de labio ≥ 5 mm) que participaron en las migraciones y su destino en la caleta (Cueva, Centro o Brazo Norte), en el periodo de enero 2005 a septiembre 2011. De un total de 1293 individuos marcados 147 sólo 30 caracoles migraron al interior de la caleta. El 57% de los individuos fueron adultos y el 43% juveniles. El 8% de los juveniles fue recapturado en Cueva y el 92% en Centro. De los adultos, el 76% se encontrón en Centro, el 18% en Brazo Norte y un 6% en Cueva. Figura 69. Número de individuos marcados en Bocana, número de individuos que emigraron de Bocana, porcentaje de juveniles (caracoles sin labio), subadultos (caracoles con un grosor de labio < 5mm) y adultos (caracoles con un grosor de labio ≥ 5 mm) que participaron en las migraciones y destinos de migración: Cueva (azul), Centro (verde) o Brazo Norte (rojo), en el periodo de enero 2005 a septiembre 2011. En la figura 70 se muestra el número de individuos marcados en Brazo Norte, el número de individuos que emigraron de Brazo Norte, así como el porcentaje de juveniles (caracoles sin labio), subadultos (caracoles con un grosor de labio < 5mm) y adultos (caracoles con un grosor de labio ≥ 5 mm) que participaron en las migraciones y destinos de migración: Cueva (azul), Centro (verde) o Bocana (morado), en el periodo de enero 2005 a septiembre 2011. De un total de 1598 individuos marcados en este sitio se registró la emigración de 245 caracoles. El 15% 148 fueron juveniles, el 22% subadultos y un 63% adultos. El 8% de los juveniles se recapturó en Bocana y el 92% en Centro. El 76% de los subadultos se recapturó en Centro, 20% en Bocana y 4% en Cueva. De los adultos un 60% fue recapturado en Bocana, 33% en Centro y 7% en Cueva. Figura 70. Número de individuos marcados en Brazo Norte, número de individuos que emigraron de Brazo Norte, porcentaje de juveniles (caracoles sin labio), subadultos (caracoles con un grosor de labio < 5mm) y adultos (caracoles con un grosor de labio ≥ 5 mm) que participaron en las migraciones y destinos de migración: Cueva (azul), Centro (verde) o Bocana (morado), en el periodo de enero 2005 a septiembre 2011. En la figura 71 se muestra el resultado del análisis de componentes principales del grosor de labio y longitud de heliconcha de caracoles marcados en el sitio Cueva, comparando la talla de organismos recapturados en su sitio de origen y organismos que emigraron a otro sitio de la caleta de Xel-Há (Centro, Bocana, Brazo Norte) entre enero 2005 y septiembre 2011. El eje 1 logró explicar el 98.9% de la distribución de los organismos y el eje 2 el 1.1% de ésta. La longitud de heliconcha explicó en un 99% la variación en el eje 1. El grosor de labio explicó el 149 99% de la variación en el eje 2. Se observó que la distribución espacial de los organismos se asoció principalmente al eje 1. Los caracoles recapturados en su sitio de marcaje (Cueva) tuvieron generalmente menores tallas en comparación con los caracoles recapturados en Centro, Bocana y Brazo Norte. Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 45.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Grosor de labio Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 30.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 15.00 PC 2(1.1)%) Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Longitud de heliconcha Bocana Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Cueva Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 0.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Centro Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Brazo Norte Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version -15.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version -30.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version -45.00 -45.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version -22.50 0.00 PC 1(98.9)%) Student version Student version 22.50 45.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Figura 71. Análisis de componentes principales del grosor de labio y longitud de heliconcha de caracoles S. gigas marcados en Cueva y recapturados en uno de los 4 sitios de muestreo de la caleta de Xel-Há (Cueva, Centro, Bocana o Brazo Norte), entre enero 2005 y septiembre 2011. En la figura 72 se muestra el resultado del análisis de componentes principales del grosor de labio y longitud de heliconcha de caracoles marcados en el sitio Centro, comparando la talla de organismos recapturados en su sitio de origen y organismos que emigraron a otro sitio de la caleta de Xel-Há (Cueva, Bocana, Brazo Norte) entre enero 2005 y septiembre 2011. El componente principal 1 logró explicar el 88% de la variación de la distribución espacial de los caracoles y fue definido en 96 % por la longitud de heliconcha. El eje 2 explicó el 12% de la distribución espacial de los caracoles y fue en 96% determinado por el grosor de labio. Se observó que la distribución espacial se asoció principalmente al eje 1. Las tallas de los organismos recapturados en Centro fueron menores en comparación con los caracoles 150 recapturados en Cueva, Bocana y Brazo Norte. La distribución de organismos recapturados en Bocana también fue asociada a un incremento en el grosor de labio. Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Grosor de labio 27.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 13.50 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version PC 2(12.4)%) Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Bocana Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Centro Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 0.00 Brazo Norte Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Cueva Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Longitud de heliconcha Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version -13.50 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version -27.00 -27.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version -13.50 0.00 PC 1(87.6)%) Student version Student version 13.50 27.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Figura 72. Análisis de componentes principales del grosor de labio y longitud de heliconcha de caracoles S. gigas marcados en Centro y recapturados en uno de los 4 sitios de muestreo de la caleta de Xel-Há (Cueva, Centro, Bocana o Brazo Norte), entre enero 2005 y septiembre 2011. En la figura 73 se muestra el resultado del análisis de componentes principales del grosor de labio y longitud de heliconcha de caracoles marcados en el sitio Bocana, comparando la talla de organismos recapturados en su sitio de origen y organismos que emigraron a otro sitio de la caleta de Xel-Há (Cueva, Centro, Brazo Norte) entre enero 2005 y septiembre 2011. El componente 1 explicó el 93.7% de la distribución espacial de los organismos y fue determinado en un 93% por la variación en la longitud de la heliconcha de los organismos. El componente 2 explicó el 6.3% de la distribución y fue en un 93% definido por la variación en el grosor de labio de los caracoles. Se observó que la distribución de caracoles se asoció en primer lugar a al eje 1. Los organismos con mayor talla fueron los recapturados en Brazo Norte y los de menor talla en Cueva. 151 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Grosor de labio 31.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 15.50 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version PC 2(6.3)%) Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Bocana Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Centro Brazo Norte Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 0.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Cueva Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Longitud de heliconcha Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version -15.50 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version -31.00 -31.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version -15.50 0.00 PC 1(93.7)%) 15.50 31.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Figura 73. Análisis de componentes principales del grosor de labio y longitud de heliconcha de caracoles S. gigas marcados en Bocana y recapturados en uno de los 4 sitios de muestreo de la caleta de Xel-Há (Cueva, Centro, Bocana o Brazo Norte), entre enero 2005 y septiembre 2011. En la figura 74 se muestra el resultado del análisis de componentes principales del grosor de labio y longitud de heliconcha de caracoles marcados en el sitio Brazo Norte, comparando la talla de organismos recapturados en su sitio de origen y organismos que emigraron a otro sitio de la caleta de Xel-Há (Cueva, Centro, Bocana) entre enero 2005 y septiembre 2011. El componente principal 1 explicó el 96.8% de la variación y fue principalmente definido por la longitud de heliconcha. La variación en el eje 2 fue en 98% por la variación del grosor de labio y explicó el 3.2% de la distribución espacial de los organismos. Se observó que la distribución de caracoles se asoció principalmente al eje 1. Los caracoles recapturados en Brazo Norte se encontraron asociados a tallas menores, mientras los recapturados en Cueva, Centro y Bocana fueron asociados a tallas mayores. 152 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Grosor de labio 29.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 14.50 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version PC 2(3.2)%) Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Cueva Bocana Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Brazo Norte Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version 0.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Centro Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Longitud de heliconcha Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version -14.50 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version -29.00 -29.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version -14.50 0.00 PC 1(96.8)%) 14.50 29.00 Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Student version Figura 73. Análisis de componentes principales del grosor de labio y longitud de heliconcha de caracoles S. gigas marcados en Brazo Norte y recapturados en uno de los 4 sitios de muestreo de la caleta de Xel-Há (Cueva, Centro, Bocana o Brazo Norte), entre enero 2005 y septiembre 2011. 153 9. Discusión 9.1 Demografía poblacional 9.1.1 Abundancia, Tamaño de Población y Densidad Durante el periodo de estudio se observó un incremento significativo en la abundancia del caracol rosa en la caleta de Xel-Há, habiéndose marcado 1665 caracoles en el 2005-2008 y 6627 en el 2009-2011. En el segundo periodo se realizaron adicionalmente muestreos en Brazo Norte, donde se marcaron 1598 individuos. La abundancia aumentó en todos los sitios, por lo cual el incremento en la abundancia no sólo se puede atribuir a la ampliación del área de estudio, sino a un incremento poblacional. Aldana-Aranda et al. (2005) reportaron un total de 803 individuos marcados en la caleta en el periodo de 2001-2003, 480 marcados en Bocana y 323 en Cueva. En este estudio, particularmente para el periodo 2009-2011 la población de la Cueva aumentó 6 veces y la de la Bocana duplicó. La abundancia determinada por el método directo fue menor al número de organismos estimados por los métodos de Jolly-Seber y Schnabel. Se capturó una alta proporción de organismos sin marcas en cada muestreo (25-80%), lo cual indicó que la población fue mayor a la porción muestreada. Utilizando el método de Schnabel, se obtuvieron inicialmente estimaciones similares al método de Jolly-Seber, sin embargo resultó en un incremento constante en el tamaño de población. El método de Schnabel es un diseño para estudios de corto plazo, el cual supone una población cerrada (Pollock, 1980). El trabajo presente se puede considerar de tipo largo plazo con reproducción y emigración a lo largo del periodo de estudio, por lo cual se trata de una población abierta y no se cumplieron los supuestos de éste método. Por lo contrario, el método de Jolly-Seber (Jolly, 1965; Seber, 1965) es un diseño que permite estimar parámetros 154 poblacionales como sobrevivencia y nacimientos, resultando en una estimación dinámica a través del tiempo. Las fluctuaciones observadas en el tamaño de población en este estudio, podrían ser explicadas por la sensibilidad de este método al éxito de recaptura, donde si se recapturan muy pocos individuos, se sobreestima el tamaño de la población, mientras si se capturan en su mayoría organismos marcados, se subestima (Pollock, 1980). El éxito de recaptura en los dos periodos fue similar con 57.1% ± 9.64 % y 51.0 ± 20.23%, en el 2005-2008 y 2009-2011 respectivamente, por lo cual las diferencias en el tamaño de población no se pueden atribuir a un efecto de muestreo. Aldana-Aranda et al. (2005) estimaron el tamaño de población en Xel-Há por el método de Schnabel en 2086.6 ± 91.28 individuos para el 2001-2003. Esta cifra fue mayor al tamaño de población estimado durante 2005-2008, pero menor al estimado para 2009-2011. Baqueiro-Cárdenas & Aldana-Aranda (2010) estimaron mediante el método de Jolly-Seber un tamaño de población de 616 ± 1771 individuos, con grandes fluctuaciones, atribuyendo lo al hábito de S. gigas de enterrarse durante las tormentas, fenómeno también observado por Hesse (1979) y Stoner & Sandt (1992). En este estudio se obtuvo por el mismo método un tamaño 1.5 veces mayor durante 2005-2008 y siete veces mayor en el 2009-2011. Con la finalidad de evaluar el grado de conservación y densidad poblacional se obtuvieron estimaciones de ésta. Diferentes autores reportan la abundancia del caracol en densidad de individuos por hectárea (De Jesús-Navarrete et al., 1992; Stoner & Ray, 1993; Friedlander et al., 1994; Stoner & Schwarte, 1994; Stoner & Ray, 1996; De Jesús-Navarrete & Oliva-Riviera, 1997; Ríos-Lara et al., 1998; Pérez-Pérez & Aldana-Aranda, 1998; Posada et al., 1999; PérezPérez & Aldana-Aranda, 2000; Pérez-Pérez & Aldana-Aranda, 2003; INP-SAGARPA, 2008). En la tabla 19 se muestran valores de densidad poblacional para el Caribe. La densidad poblacional observada en la Caleta de Xel-Há fue similar en comparación a la mayoría de lo 155 reportado la literatura para el periodo 2005-2008 pero alta en el periodo 2009-2011, sin embargo resultó difícil comparar los valores obtenidos por las divergencias en las metodologías aplicadas en la región. La mayoría de los autores utilizan el método de Transectos para la estimación de la densidad (Berg & Glazer, 1991; Friedlander et al., 1994; Stoner & Schwarte, 1994; Stoner & Ray, 1996; Ríos-Lara et al., 1998; Pérez-Pérez & Aldana-Aranda, 1998; Posada et al., 1999; Pérez-Pérez & Aldana-Aranda, 2000; Pérez-Pérez & Aldana-Aranda, 2003; INP-SAGARPA, 2008) pero la extensión de estos puede varía considerablemente en los diferentes estudios. Stoner & Ray (1993), INP-SAGARPA (2008) y Phillips et al. (2011) aplican el método de cuadrante, contando el número de caracoles sobre un área circular. Sólo De Jesús-Navarrete et al. (1992) emplean el mismo método utilizado en el trabajo presente, contabilizando individuos marcados en un área delimitada. A pesar de que hacen uso del método de CMR y que estiman el tamaño de población, no relacionan este a la densidad de organismos en el área de estudio. A pesar de que fue difícil comparar los resultados obtenidos en este estudio con otros, son de gran valor, ya que en este trabajo se empleó una metodología repetitiva y de largo plazo, generando datos que permiten evaluar confiablemente la evolución de la población a través del tiempo. Las mayores densidades de este estudio se observaron en los sitios Cueva y Brazo Norte. Densidades poblacionales de 1000 a 2000 caracoles por hectárea se consideran normales para las agregaciones de juveniles (Hesse 1979; Stoner & Ray 1993; Stoner et al., 1996), densidades que se obtuvieron en estos dos sitios durante el periodo de 2009-2011. A pesar de que se registró la mayor abundancia de individuos en Centro, este sitio tuvo las menores densidades. El sitio Centro tuvo el área mayor muestreado con 23,000 m2, sin embargo se aplicó el mismo esfuerzo de muestreo como a los otros sitios. Es un sitio estructuralmente 156 heterogéneo que comprende una amplia zona arenosa en su centro. La densidad de organismos en Centro probablemente fue subestimado por el diseño de muestreo. Tabla 19. Densidad poblacional del Caracol rosa S. gigas en el Caribe. Densidad (Ind. ha⁻¹) 47 Autor, Año Ríos-Lara et al. 1998 Localidad Alacranes, Yucatán, México Pérez-Pérez & Aldana-Aranda 1998 Pérez-Pérez & Aldana-Aranda 2000 Pérez-Pérez & Aldana-Aranda 2003 De Jesús-Navarrete et al. 1992 Alacranes, Yucatán, México Método Transecto, extensión variables Transecto 100 x 10m Alacranes, Yucatán, México Transecto 100 x 10m 84 Alacranes, Yucatán, México Transecto 100 x 10m 180 No. Individuos / 3 Ha 30 Círculo d=200 m2 211 ± 35 Transecto tiempo/esfuerzo Transecto 1.2 Ha 79 ± 165 Berg & Glazer 1991 Punta Gavilán, Quintana Roo, México Banco Chinchorro, Quintana Roo, México Banco Cozumel, Quintana Roo, México Florida Keys, Florida, USA Friedlander et al. 1994 Virgin Islands, USA 17 Stoner & Ray 1993 Exuma Cays, Bahamas Lee Stocking Island, Bahamas Transecto, extensión variable Círculo de r= 4m 2000 Transecto 100m 18-88 Exuma Cays (sin pesca), Bahamas Exuma Cays (con pesca), Bahamas Jaragua Parque Nacional, República Domenicana Barbados Transecto de longitud variable, ancho 8m Transecto de longitud variable, ancho 8m Transecto 100m 34-270 Círculo de r= 4m 6000 Caleta No. Individuos/3.9 Ha Caleta No. Individuos/4.5 Ha Caleta No. Individuos estimados/3.9 Ha No. Individuos estimados/4.5 Ha No. Individuos estimados/3.9 Ha No. Individuos estimados/4.5 Ha 56.0 ± 12.56 243.5 ± 75.14 245.9 ± 92.57) 849.9 ± 433.36) 209.7 ± 131.61) 809.2 ± 654.78) INP-SAGARPA 2008 INP-SAGARPA 2008 Stoner & Schwarte 1994 Stoner & Ray 1996 Stoner & Ray 1996 Posada et al. 1999 Phillips et al. 2011 Trabajo Presente, Periodo 2005-2008 (Directo) Trabajo Presente, Periodo 2009-2011 (Directo) Trabajo Presente (Schnabel), Periodo 2005-2008 Trabajo Presente (Schnabel), Periodo 2009-2011 Trabajo Presente (Jolly-Seber), Periodo 2005-2008 Trabajo Presente (Jolly-Seber) Periodo 2009-2011 *r- radio *d- diámetro Caleta Caleta Caleta 72 1-3 2-88 4-114 157 9.1.2 Estructura de tallas En este estudio la proporción de tallas de la población se mantuvo constante desde enero 2005 hasta enero 2011 con más de 2/3 de la población compuesta por juveniles. Este resultado concuerda con lo observado por Aldana-Aranda et al. (2005), que reportaron 79.2% juveniles y subadultos en la caleta. A partir de enero 2011 aumentó la proporción de adultos y subadultos. Este cambio en la estructura de talla podría atribuirse a un reclutamiento masivo entre octubre 2009 y febrero 2010, resultando en la inversión de la estructura poblacional a partir del 2011. Stoner et al. (1996) atribuye variaciones en la estructura poblacional de S. gigas principalmente a variaciones en las condiciones ambientales (procesos hidrográficos costeros que afectan el reclutamiento) y cambios en las interacciones biológicas (predación) (Roughgarden et al., 1988). La población muestreada de la caleta de Xel-Há fue una población principalmente compuesta por juveniles. De Jesús-Navarrete et al. (2003) reportaron para Banco Chinchorro poblaciones compuestas en un 95% de juveniles, sin embargo atribuye la estructura poblacional al efecto de pesca. Tewfik & Bené (2000) reportan para áreas con pesca una relación de juveniles a adultos de 4:1 y para zonas protegidas una relación de 2:1. En Xel-Há la extracción de organismos está prohibida, sin embargo se ha observado de forma incidental, pero el aumento en la abundancia sugiere que no tiene impacto sobre la estructura de población. Hesse (1979) describe una población grande compuesta por más de 10,000 individuos distribuida en un área sin actividad pesquera con una relación de juveniles a adultos de 3:1, lo cual atribuyó a la gran movilidad de organismos adultos, los cuales abandonan frecuentemente el área de estudio. El valor reportado por Hesse (1979) fue similar a la razón observada en Xel-Há. La distribución de tallas mostró diferencias significativas en el espacio, observándose un gradiente de tallas a lo largo de la caleta, por lo cual fue posible afirmar la hipótesis (4.1) de 158 trabajo planteada de es este trabajo. La mayoría de los organismos encontrados en Cueva, Centro y Brazo Norte fueron juveniles, mientras en Bocana la mayoría fueron adultos y básicamente todos los organismos con un grosor de labio mayor a 20 mm se detectaron en este sitio. Los “nurseries” o criaderos se caracterizan principalmente por la alta abundancia de juveniles a densidades de 0.2-2 caracoles m-2 (Stoner & Ray, 1993), lo cual indica que Cueva, Centro y Brazo Norte están funcionado como zonas de crianza. Aldana-Aranda et al. (2005) observaron durante el periodo de estudio de 2001 a 2003 diferencias similares en Xel-Há, reportando para la Cueva el 76.5% juveniles y para la Bocana un 82.3% adultos y subadultos. Estos resultados sugieren que el gradiente de tallas en la caleta se mantuvo relativamente constante a través de los últimos 10 años. Stoner y Ray (1996) determinaron mediante el análisis de sedimentos que los criaderos de S. gigas tienen una distribución muy restringida y que típicamente son zonas de persistencia histórica, donde las agregaciones de juveniles se encuentran año tras año a través de décadas. La perseverancia de la población y de su estructura de tallas en el espacio y tiempo es otro indicio que Xel-Há está funcionando como una zona de crianza para el caracol rosa. Alcolado (1976) determinó que los criaderos se distinguen de otros sitios por ser áreas resguardadas de la corriente, con alta estabilidad de los sedimentos y fondos poblados por Thalassia y algas filamentos. La caleta de Xel-Há presenta un gradiente en la distribución de los sedimentos en la caleta, el cual concuerda con gradiente de tallas. Bocana presenta un fondo principalmente compuesto por arena y se ha observado que este está siendo afectado por la marejada por su cercanía al mar, por lo cual podría ser un factor que limita la distribución de los juveniles en este sitio. Por el otro lado, el caracol adulto, especialmente durante la época reproductiva, tiene preferencias por sustratos arenosos (Randall, 1964; D’Asaro, 1965; PérezPérez & Aldana-Aranda, 2002; Pérez-Pérez & Aldana-Aranda, 2003; Glazer & Kidney, 2004), lo 159 cual podría explicar la predominancia de adultos en Bocana. La alta abundancia de adultos en Bocana sugiere que esta zona no forma parte del criadero y que desempeña otro papel ecológico. Aldana-Aranda et al. (2005) documentaron previamente actividad reproductiva (cópula, desove y masas de huevo fertilizadas) en este sitio y Aldana-Aranda & Sánchez-Crespo (pers. Com.) la caracterizan como zona reproductora. Chávez-Villegas (2011) encontró larvas de S. gigas por debajo de la haloclina en Bocana. Las tallas de éstas correspondieron al estadio I según el criterio de clasificación de Davis et al. (1993), por lo cual el sitio Bocana se considera suministrador de larvas. A pesar de que el agua superficial de la caleta se mueve en dirección del mar, existen evidencias que el agua del fondo tiende a dirigirse a bajas velocidades hacia el interior de la caleta (Sáenz-Morales, com. pers.), por lo cual es probable que Bocana podría suministrar larvas a las zonas de crianza en el interior de la caleta bajo condiciones ambientales adecuadas. 9.2 Crecimiento 9.2.1 Crecimiento de la heliconcha por métodos directos Las tasas de crecimiento mostraron diferencias entre los dos periodos de estudio. Estudios experimentales indican que el crecimiento del caracol es inversamente denso-dependiente como resultado de la competencia por alimentos preferidos (Stoner, 1989; Stoner & Ray, 1993; Stoner & Davis, 1994). Sin embargo, en el trabajo presente el crecimiento fue mayor a mayores densidades, lo cual implica que el alimento no es un factor limitante. Alcolado (1976) identificó varios factores ambientales que afectan el crecimiento del caracol, entre estos la temperatura, la presencia de algas filamentosas y sedimentación. En Xel-Há se observó un incremento en la concentración de fósforo entre 2009 y 2011, con picos marcados en marzo y septiembre 2010. En los sistemas marinos el nitrógeno es el principal limitante de la productividad primaria (Ryther & Dunstan 1971, Oviatt et al. 1995), mientras en sistemas dulceacuícolas la 160 productividad generalmente es limitada por fósforo (Schindler, 1974). En las lagunas costeras el fósforo también presenta un factor limitante de la productividad primaria y esta es estrechamente vinculada con la productividad de los recursos pesqueros (Nixon, 1982). Las altas tasas de crecimiento indican que el hábitat no se encontró completamente saturado a través del periodo de estudio y sugiere que el tamaño de población no está siendo limitado por disponibilidad de alimentos (Stoner, 2003). Las diferencias en el crecimiento entre periodos podrían atribuirse a diferencias en la calidad del hábitat, probablemente ligados a la productividad primaria. También se encontraron diferencias entre las tasas de crecimiento en los cuatro sitios estudiados de la caleta de Xel-Há. Los caracoles en Bocana tuvieron en ambos periodos de estudio las tasas más bajas de crecimiento. En Centro se registraron las tasas más altas y en Brazo Norte y Cueva fueron similares entre sí. Los criaderos de S. gigas se caracterizan por altas tasas de crecimiento juvenil (Stoner, 2003), indicando que Cueva, Centro y Brazo Norte, probablemente funcionan como zonas de crianza. De Jesús-Navarrete (2001) estudió el crecimiento de S. gigas en diferentes ambientes y reconoció que a nivel poblacional el crecimiento de juveniles generalmente es mayor en zonas con vegetación. La cobertura de macroalgas fue alta en Brazo Norte y Cueva, y heterogénea en Centro. Centro fue el sitio con menor densidad de individuos y presentó las tasas de crecimiento más altas. Es posible que las diferencias en las tasas de crecimiento entre sitios se asocien a la cobertura vegetal y en menor grado a diferencias en la densidad de organismos. En la tabla 20 se presentan las tasas de crecimiento de S. gigas reportadas para otras localidades del Caribe. De Jesús-Navarrete (2001) estimó un crecimiento medio de 3.21 mm mes-1 (~0.1052 mm día⁻¹) en Punta Gavilán y 2.30 mm mes-1 (~0.075 mm día⁻¹) en Banco Chinchorro, manteniendo los caracoles en encierros a una densidad de 0.4 Ind m-². En otras áreas del Caribe 161 se observaron tasas de crecimiento similares (Randall, 1964; Alcolado, 1976; Brownell, 1977; Ray & Stoner, 1994). Las tasas de crecimiento obtenidas en este estudio fueron altas en comparación con otros sitios del Caribe, sin embargo fueron comparables con las tasas obtenidas bajo condiciones naturales, utilizando métodos de captura-marcaje-recaptura. Gibson et al. (1983) determinaron un crecimiento de 7.2 mm mes-1 (~ 0.236 mm día⁻¹) en Belice, De JesúsNavarrete & Oliva-Rivera (1997) 10 mm mes-1 (~ 0.327 mm día⁻¹) en Punta Gavilán, Phillips et al. (2011) 0.3 mm día-1 en Barbados y en Weil & Laughlin (1984) 15 mm mes-1 (~ 0.492 mm día⁻¹) en Venezuela. 162 Tabla 20. Tasas de crecimiento de S. gigas en el Caribe (*Valores calculados a partir de la información del artículo). Autor, Año Localidad Método Crecimiento (mm día-1) Randall, 1964 Virgin Islands, USA Encierre ~0.136* Alcolado, 1976 Cuba Encierre, diferentes ambientes ~0.108* Brownell, 1977 Florida Keys, USA Encierre ~0,147* Gibson et al., 1983 Belice CMR ~0.236* Ballantine & Appeldoorn, 1983 La Parguera, Puerto Rico Encierre Weil & Laughlin, 1984 Venezuela CMR ~0.492* Appeldoorn, 1985 La Parguera, Puerto Rico CMR ~0.016-0.2* Ray & Stoner, 1994 Exuma Cays, Bahamas Encierre 0.058 0.139 De Jesús-Navarrete & Oliva-Rivera, 1997 Punta Gavilán, México CMR ~0,327* De Jesús-Navarrete, 2001 Banco Chinchorro, México De Jesús-Navarrete, 2002 Punta Gavilán, México Moreno de la Torre & Aldana-Aranda, 2005 México Phillips et al., 2011 Barbados Encierre, diferentes ambientes Encierre, diferentes ambientes Condiciones de laboratorio, dieta artificial CMR 0.21 ~0.1052* ~0.075* 0.16-0.23 0.3 El crecimiento mostró oscilaciones a través del tiempo, las cuales fueron más marcadas en el periodo 2009-2011. Alcolado (1976) observó que el crecimiento de S. gigas presenta oscilaciones anuales, con crecimiento máximo entre mayo y septiembre, lo cual atribuyó a un incremento en la temperatura. Iversen (1893) y Appeldoorn (1985) registraron crecimiento máximo durante los meses de verano. En este estudio no fue posible atribuir las oscilaciones del crecimiento a un patrón estacional, utilizando el método directo. 163 9.2.2 Estimación de los parámetros de crecimiento la ecuación de Von Bertalanffy Con la finalidad de evaluar la dinámica poblacional se calcularon los parámetros de crecimiento de von Bertalanffy, utilizando el método de Appeldoorn (1987b), lo cual se hizo por un lado por métodos directos y por el otro lado por métodos indirectos. Por ambos métodos se obtuvo un crecimiento menor en el periodo 2005-2008. El método indirecto determinó tasas de crecimiento más bajas en comparación con las tasas obtenidas a partir del análisis de los datos de marcajerecaptura. El uso del método directo puede producir resultados altamente variables, ya que generalmente el número de recapturas es pequeño y el crecimiento a nivel de individuos puede ser altamente variable (Appeldoorn, 1987a). Más allá, dado que organismos más grandes son más conspicuos, existe el riesgo de producirse un sesgo de muestreo hacía organismos de crecimiento alto, lo cual lleva a la sobrestimación de los valores de k y subestimación de L∞ (Appeldoorn, 1987a). El método indirecto requiere de muestras muy grandes cuando la población está compuesta por varios cohortes, cuyas distribuciones de frecuencia se sobreponen (Appeldoorn, 1987a). En el trabajo presente, especialmente para el periodo 2005-2008, fue difícil distinguir las cohortes de la distribución de tallas por el tamaño reducido de la muestra. Se ha observado que el reclutamiento en la caleta fue continúo, sin embargo en el análisis de las frecuencias de tallas no se distinguieron más de dos ó tres cohortes por año. Para el análisis de progresión de moda se utilizaron las medidas de todos los organismos observados, incluyendo aquellos con labio, los cuales presentan poco o ningún crecimiento, por lo cual probablemente se subestimó el parámetro k de la ecuación de von Bertalanffy. El crecimiento de los caracoles de la caleta de Xel-Há ha sido evaluado previamente. Las tasas de crecimiento obtenidas en el trabajo presente fueron comparables con las obtenidas por ValleEsquivel (1998) y Aldana-Aranda et al. (2005), pero fueron muy altas en comparación con 164 estudios realizados por Aldana-Aranda et al. (2003) y Baqueiro-Cárdenas & Aldana-Aranda (2010) (Tabla 21). 165 Tabla 21. Parámetros de crecimiento de von Bertalanffy del caracol rosa, obtenidas en la caleta de XelHá. Localidad L∞ mm k año⁻¹ Método Autor, Año 617.97 0.23 Gulland-Holt Valle-Esquivel, 1998 295.64 0.77 Gulland-Holt Valle-Esquivel, 1998 341.74 376.79 0.58 0.48 Gulland-Holt Gulland-Holt Valle-Esquivel, 1998 Valle-Esquivel, 1998 329.99 0.58 Fabens Valle-Esquivel, 1998 249.69 1.5 Fabens Valle-Esquivel, 1998 278.16 263.87 0.86 0.99 Valle-Esquivel, 1998 Valle-Esquivel, 1998 Aldana-Aranda et al., 2003 Aldana-Aranda et al., 2003 Aldana-Aranda et al., 2003 Aldana-Aranda et al., 2005 Aldana-Aranda et al., 2005 Aldana-Aranda et al., 2005 Aldana-Aranda et al., 2005 Baqueiro-Cárdenas & Aldana-Aranda, 2010 Baqueiro-Cárdenas & Aldana-Aranda, 2010 Xel-há, Caleta+Acuario Xel-há, Caleta+Acuario Tallas Juveniles pequeños Juveniles grandes Juveniles pequeños+ grandes Todos Juveniles pequeños Juveniles grandes Juveniles pequeños+ grandes Todos Xel-há, Cueva Todos 203.22 0.00578 Xel-há, Bocana Todos 249.33 0.1724 Xel-há, Bocana+Cueva Todos 361.83 0.0394 Xel-há, Cueva Todos 340.50 0.038 Xel-há, Bocana Todos 335.98 0.015 Fabens Fabens FordWalford FordWalford FordWalford FordWalford FordWalford Xel-há, Cueva Todos 356.80 0.524 ELEFAN I Xel-há, Bocana Todos 340.00 0.42 ELEFAN I Xel-há, Caleta Juveniles 251.00 0.3 Appeldoorn Xel-há, Caleta Xel-há, Caleta (20052008) Xel-há, Caleta (20092011) Xel-há, Caleta (20052008) Xel-há, Caleta (20092011) Adultos 330.00 0.07 Todos 280.52 0.57 Todos 278.73 0.71 Todos 285.16 0.32 Todos 290.51 0.55 Appeldoorn Appeldoorn, CMR Appeldoorn CMR Appeldoorn, indirecto Appeldoorn indirecto Xel-há, Caleta+Acuario Xel-há, Caleta+Acuario Xel-há, Caleta+Acuario Xel-há, Caleta+Acuario Xel-há, Caleta+Acuario Xel-há, Caleta+Acuario Trabajo Presente Trabajo Presente Trabajo Presente Trabajo Presente 166 En comparación con otros sitios del Caribe (Tabla 22) el crecimiento de S. gigas fue alto en la caleta de Xel-Há. La tasa de crecimiento calculada para el periodo 2005-2008 fue comparable con el crecimiento de S. gigas observado en Cuba (Alcolado, 1976), Islas Vírgenes (Berg, 1976) y Punta Gavilán, México (De Jesús-Navarrete, 2000). Sin embargo, el crecimiento estimado para el periodo de 2009-2011 fue muy alto y sólo fue comparable con el crecimiento obtenido en Providencia y San Andrés, Colombia (García, 1991), así como las tasas de crecimiento obtenidas por Valle-Esquivel (1998) en la caleta de Xel-Há en los años 1990s. Tabla 22. Parámetros de crecimiento de von Bertalanffy del caracol rosa en el Caribe. L∞ mm k año⁻¹ 232.70-383.4 260.4 241.7 300 268 332 269 460 340 0.287-0.571 0.516 0.42 0.2 0.223 0.207 0.209 0.25 0.437 San Andrés & Providencia, Colombia 329.4 0.72 García, 1991 Providencia & Santa Catalina, Columbia Martinique Punta Gavilán, México 375 339 325.35 0.25 0.392 0.345 Marquéz, 1993 Rathier & Battaglya, 1994 De Jesús-Navarrete et al., 1995 350 221 307.08 311 278.4-287.5 0.27 0.58 0.44 0.28 0.21-0.4 Gallo et al. 1996 Twefik, 1996 De Jesús-Navarrete et al., 2000 De Jesús-Navarrete et al., 2000 De Jesús-Navarrete, 2001 Localidad Cuba, varios sitios St. John, Islas Vírgenes St. Croix, Islas Vírgenes Berry Island, Bahamas Boca Chica, Belice Tres Cocos, Belice Water Caye, Belice La Parguera, Puerto Rico La Parguera, Puerto Rico San Andrés & Providencia, Colombia Pedro Bank, Jamaica Punta Gavilán, México Punta Gavilán, México Punta Gavilán, México Autor, Año Alcolado, 1976 Berg, 1976 Berg, 1976 Iversen et al., 1987 Strasdine, 1988 Strasdine, 1988 Strasdine, 1988 Appeldoorn, 1990 Appeldoorn, 1990 El crecimiento determinado por estimación de los parámetros de Von Bertalanffy fue congruente con los resultados obtenidos por el método directo. 167 Para el grosor de labio no fue posible estimar el crecimiento mediante el análisis de frecuencias, dado que no fue posible discernir progresión de modas de la distribución de frecuencia de grosor, por lo cual el crecimiento del labio únicamente fue evaluado mediante el método directo utilizando el modelo de Appeldoorn (1987b). A pesar de que se utilizaron los datos de marcajerecaptura el resultado no fue concluyente. Se observó que el crecimiento era altamente variable entre individuos y al parecer también entre cohortes. En la tabla 23 se presentan los parámetros de crecimiento reportados por Appeldoorn (2003) en Puerto Rico y Baqueiro-Cárdenas & Aldana-Aranda (2010) en la caleta de Xel-Há en comparación con los resultados obtenidos en este estudio. El crecimiento obtenido por Appeldoorn (2003) corresponde a un aumento de 16.97 mm en un año (Lt(1)). Baqueiro-Cárdenas & Aldana-Aranda (2010) calcularon por el mismo método, un incremento de aproximadamente 7 mm en un año (Lt(1)). La tasa de crecimiento obtenido en este estudio para el periodo 2009-2011 corresponde a un incremento de aproximadamente 14 mm en un año (Lt(1)), lo cual se acerca al resultado obtenido por Appeldoorn a pesar de que L∞ se sobreestimó y k parece subestimado, que es una particularidad del método, como se discutió anteriormente. El desplazamiento de las medias del grosor de labio entre mayo y septiembre 2011 (Fig. 27), de 4 mm a 10 mm, apoya el resultado obtenido mediante la estimación de los parámetros de la ecuación de von Bertalanffy. Por lo contrario, el crecimiento obtenido durante el periodo 2005-2008 parece muy alto y es posible que fuera sobreestimado. Tabla 23. Parámetros de crecimiento de von Bertalanffy para el grosor de labio del caracol rosa. Localidad La Parguera, Puerto Rico L∞ mm 54.9 k año⁻¹ 0.37 Xel-Há Xel-Há (2005-2008) Xel-Há (2009-2011) 47.78 99.55 301.1 0.17 0.27 0.05 Autor, Año Appeldoorn, 2003 Baqueiro-Cárdenas & Aldana-Aranda, 2010 Trabajo presente Trabajo presente 168 De acuerdo a los resultados obtenidos en este trabajo la mayoría de los caracoles desarrolla un labio a una talla entre 200 y 220 mm, lo cual corresponde según los cálculos de crecimiento determinado por los distintos métodos a una edad aproximada entre 2.2 a 3.3 años. Tomando como medida de madurez un grosor de labio de 5 mm, como fue sugerido por Appeldoorn (1988) y Aldana-Aranda & Frenkiel (2007), y la tasa de crecimiento del grosor de labio obtenido en el periodo 2009-2011, los caracoles probablemente inician la reproducción a los 2.6 a 3.7 años de edad. Appeldoorn (2003) determinó que el labio se forma a una edad de 3.2 años y que la reproducción inicia a los 3.6 a 4.0 años de edad. 9.3 Dinámica Poblacional 9.3.1 Reclutamiento Se observaron grandes diferencias en el reclutamiento de juveniles al epibentos entre años. El reclutamiento de organismos menores a 100 mm fue moderado en 2005, alto en 2006, casi nulo en 2007, masivo en 2009, moderado en 2010 y bajo en 2011. Observaciones similares de la variabilidad del reclutamiento entre años fueron hechas por Alcolado (1976) en Cuba y Appeldoorn (1987a) en Puerto Rico. En las poblaciones de las Bahamas se ha observado que el tamaño de población y la abundancia de juveniles son principalmente regulados por el aporte de larvas competentes (Stoner, 2003), por lo cual se concluye que los cambios en el tamaño y la estructura de la población de la caleta de Xel-Há se deben en primer lugar a diferencias observadas en la magnitud del reclutamiento. El retro-cálculo del reclutamiento por el método de análisis de frecuencias de longitud determinó que hay reclutamiento a lo largo del año con excepción de diciembre, pero que el reclutamiento fue de mayor magnitud entre marzo y abril y junio a agosto en ambos periodos de estudio. Aldana-Aranda et al. (2005) documentaron actividad reproductiva (cópula, desove, masas de 169 huevos) en la caleta en marzo, abril, mayo y agosto, lo cual coincide con la observación de los picos de reclutamiento de juveniles en este estudio. En México (Pérez-Pérez & Aldana-Aranda, 2002; Pérez-Pérez & Aldana-Aranda, 2003) y en las Islas Vírgenes (Randall, 1964) se observó que la reproducción se lleva a cabo entre enero y octubre, lo cual se puede confirmar con la continuidad del reclutamiento observada en el trabajo presente. De Jesús-Navarrete et al. (1994) determinaron reclutamiento con dos picos, el mayor de marzo a agosto y el segundo de agosto a noviembre, utilizando el mismo método como en el trabajo presente. 9.3.2 Sobrevivencia y Mortalidad Se estudio la sobrevivencia y mortalidad del caracol rosa en la caleta de Xel-Há mediante el análisis de los datos de marcaje-recaptura. La recuperación de organismos fue menor al 40 % pasando 2 meses y menor del 20% después de un año en ambos periodos de estudio. Se detectaron diferencias en la sobrevivencia entre sitios, registrando sobrevivencias inferiores en Bocana y Centro y sobrevivencias altas en Cueva y Brazo Norte. La principal causa de mortalidad en los juveniles es la predación (Randall, 1964; Stoner & Ray, 1993) y puede variar entre un 10% (Ray & Stoner, 1994) hasta un 60% (Iversen et al., 1986). Ésta tiene gran influencia sobre la distribución y abundancia de los juveniles (Stoner & Ray, 1995). Los criaderos son generalmente hábitats de poca profundidad (<10 m) (Randall, 1964; Stoner & Waite, 1991) y se ha especulado que la preferencia por profundidades bajas brinda refugio ante predadores marinos de mayor tamaño (Iversen et al., 1986). En el caso de la caleta de Xel-Há la profundidad es relativamente homogénea, sin embargo la persistente haloclina podría desempeñar un papel similar al gradiente de profundidad, limitando el acceso a grandes predadores marinos al interior de la caleta. En Bocana y parcialmente Centro persiste mayor 170 influencia marina lo cual permite el acceso de peces grandes a estas zonas y podría ser un factor que influye sobre la sobrevivencia diferencial observada a lo largo de la caleta. La mortalidad en ambos periodos de estudio fue de z= 2.3 después de un año. Alcolado (1976) obtuvo a partir del análisis de marcaje-recaptura una sobrevivencia de 15-35% al año, con un z entre 1.06 y 1.90 año -1. Iversen et al. (1987) obtuvo por el mismo método una sobrevivencia de 57-80% a un mes y de 2-9% después de un año, así como una mortalidad z de estimado en 2.413.91 año-1. Appeldoorn (1987c) calculó mediante el método de Jolly-Seber una mortalidad total de z= 1.53 año-1 y una mortalidad natural de z= 1.05 año -1. La mortalidad obtenida en Xel-Há por este método se encuentra dentro de los rangos de lo previamente reportado. A lo largo del estudio se encontraron pocos caracoles muertos en la caleta. La mayoría se encontraron en Cueva y eran juveniles. El análisis de la frecuencia de talla sin embargo determinó que la mortalidad era mayor en organismos con tallas superiores a 200 mm. La mortalidad determinada por el método de tallas convertidas fue excepcionalmente alta en comparación con otros estudios (Tabla 24). En el caso de S. gigas la mortalidad generalmente disminuye con la edad y se estabiliza cuando los organismos alcanzan la madurez sexual y desarrollan el labio (Appeldoorn, 1988). Dado que no se encontraron muchos caracoles muertos en los límites de la caleta, no hay pesca regular y que la mortalidad se manifestó principalmente en organismos mayores a dos años, se atribuye esta mortalidad a la emigración de los organismos. 171 Tabla 24. Mortalidad (z) del caracol rosa S. gigas en el Caribe estimado por curvas de captura de tallas convertidas. Localidad La Praguera, Puerto Rico Punta Gavilán, México Labio (Grosor) Toda la población (Longitud) Xel-Há Sin Labio (Longitud) Xel-Há Con Labio (Longitud) Xel-Há Labio (Grosor) Toda la población (Longitud), 2005-2008 Toda la población (Longitud), 2009-2011 Labio (Grosor), 20052008 Labio (Grosor), 20092011 Xel-Há Xel-Há Xel-Há Xel-Há Organismos z año⁻¹ Autor, Año 1.66 Appeldoorn, 1988 2.01 De Jesús-Navarrete et al., 2000 Baqueiro-Cárdenas & Aldana0.55 Aranda, 2010 Baqueiro-Cárdenas & Aldana0.72 Aranda, 2010 Baqueiro-Cárdenas & Aldana0.72 Aranda, 2010 1.46 Trabajo presente 4.71 Trabajo presente 2.39 Trabajo presente 1.97 Trabajo presente 9.4 Movilidad 9.4.1 Dirección y frecuencia de transiciones En el trabajo presente se detectó movilidad de los organismos a lo largo de la caleta. Alrededor del 15% de los organismos marcados en Cueva, Centro y Brazo Norte presentaron movimientos de este tipo. Los organismos marcados en Brazo Norte y Cueva fueron principalmente recapturados en Centro y Bocana. Los organismos marcados en Centro fueron detectados con mayor frecuencia en Bocana. En el caso de los organismos con origen en Bocana fue menos del 3% de la población marcada que realizó movimientos hacía el interior de la caleta, por lo cual se concluye que los movimientos registrados no son al azar, sino que los caracoles se dirigen principalmente en dirección del mar Caribe. La actividad de los caracoles aumentó en los meses de verano, particularmente durante la estación de Lluvias. Hesse (1979) estudió la movilidad del 172 caracol rosa en las Islas Turks & Caicos y observó que los animales realizaron pocos movimientos entre noviembre y febrero, pero que fueron muy activos entre marzo y septiembre, cuando abandonaron las zonas de algas mixtas y se dirigieron hacia zonas coralinas o pastos marinos. Glazer et al. (2003) detectaron mediante un estudio telemétrico que la actividad de organismos adultos aumentó durante los meses de verano. Doerr & Hill (2007) también detectaron que los organismos adultos abandonaron el área de estudio con mayor frecuencia entre marzo y agosto, lo cual asociaron con incrementos en la temperatura con una magnitud de 2°C (Doerr & Hill, 2007). El patrón observado en este estudio coincide con lo reportado en la literatura. 9.4.2 Talla de Transición Se detectó una asociación entre las tallas de los organismos que emigraron de los sitios Cueva, Centro y Brazo Norte y su distribución posterior en la caleta. Los organismos emigrantes de estos sitios tuvieron tallas medias entre 208 mm y 212 mm y fueron principalmente adultos, pero también subadultos y en menor proporción juveniles. Los organismos con mayor grosor de labio generalmente se reubicaron en Bocana. Randall (1964) detectó que los caracoles juveniles presentan poca movilidad. Alcolado (1976) detectó que la distribución de caracoles juveniles normalmente es confinada a los criaderos, mientras los adultos tienen una distribución más amplia. Hesse (1979) señaló diferencias significativas en el rango hogareño (Homerange) del caracol rosa en función de la talla, determinando un rango de 1,000 m2 para organismos con tallas de 100-130 mm y de 2,500-5,000 m2 para organismos con tallas entre 130-160 mm. En su estudio no fue posible determinar el rango hogareño para organismos con tallas mayores a 170 mm, dado que estos utilizan áreas muy extensas, mayores a 5,000 m2. Por el método de rastreo con telemetría acústica Glazer et al. (2003) determinaron que el rango hogareño de caracoles 173 adultos es en promedio de 59,800 m2. Bissada-Gooding et al. (2010) determinaron por telemetría acústica y marcaje tradicional que los adultos en promedio utilizaban un área de 11,031 m2, pero algunos usan extensiones de 21,255 m2. La observación hecha en este trabajo coincide con Hesse (1979), que los organismos amplían su rango de distribución conforme crecen. Las tallas de los organismos con mayor movilidad concuerdan con los organismos que presentaron mayor mortalidad, la cual fue máxima en organismos con longitud mayor a 200 mm. En el caso de S. gigas la mortalidad disminuye y se estabiliza cuando los organismos desarrollan su labio (Appeldoorn, 1988). En Xel-Há no se encontraron muchos caracoles muertos en los límites del área de estudio y no hay pesca regular, sin embargo se observó que la mayoría de los movimientos observados fueron en dirección de Centro y Bocana, donde la caleta conecta con el mar Caribe, por lo cual se concluye que la alta mortalidad se atribuye a emigración y que es probable que muchos organismos abandonan el área de estudio al alcanzar la madurez. Previamente se han descrito migraciones de juveniles entre 50-100 mm (Stoner et al., 1988; De Jesús-Navarrete & Valencia-Beltrán, 2003; Danylchuck et al. 2003), los cuales fueron asociados con la conectividad de la etapa infaunal y la etapa epibentónica juvenil y de organismos adultos (Robertson, 1959; Alcolado, 1976; Hesse, 1979; Appeldoorn, 1997), la cual fue asociado a la reproducción y se piensa que es de naturaleza estacional (Appeldoorn, 1997). Se planteó la hipótesis de que los organismos realizan transiciones en su hábitat, las cuales dependen de su talla. Se pudo observar que la movilidad aumenta significativamente en organismos adultos, existiendo una relación entre la talla y movilidad ontogénica, estableciendo la conectividad espacial y estacional entre juveniles y adultos. 174 10. Conclusión A lo largo del periodo de estudio la abundancia y densidad del caracol S.gigas en la caleta de Xel-Há incrementaron, principalmente por un aumento en la abundancia de juveniles en los sitios Cueva, Centro y Brazo Norte. La densidad de caracoles en Xel-Há es alta con respecto a otros sitios del Caribe, pero similar a otras áreas naturales protegidas en las cuales no se tiene impacto pesquero; concluyéndose que la caleta desempeña un papel como área natural protegida y funciona como un santuario para el caracol rosa. La densidad en Cueva y Brazo Norte fue comparable con densidades de agregaciones reportadas en otras zonas de crianza de esta especie en el Caribe. La población de S.gigas de Xel-Há está compuesta principalmente por juveniles y su composición fue relativamente constante a través del tiempo. Se encontró un gradiente en la distribución por tallas en el espacio, observándose primariamente juveniles en Cueva, Centro y Brazo Norte y adultos en Bocana. Lo anterior posiblemente es resultado de un aumento en la movilidad de adultos y subadultos, confirmándose las hipótesis de trabajo planteadas. También se notaron diferencias en el tipo de sedimento, densidad de vegetación y espesor de la haloclina los cuales coinciden con el gradiente de tallas de esta especie, que han sido identificados como factores que afectan el crecimiento y sobrevivencia en juveniles y determinan las áreas aptas para la reproducción. Las tasas de crecimiento de S. gigas determinadas en Xel-Há por el método directo fueron altas en comparación con otros sitios del Caribe, pero comprables con estudios que emplearon el método de marcaje-recaptura. Las mayores tasas de crecimiento se obtuvieron en los sitios Centro, Brazo Norte y Cueva. Por lo que respecta a los parámetros de crecimiento de la ecuación 175 de Von Bertalanffy, estimados para los caracoles de la caleta, estos también fueron altos con respecto a otros sitios, pero comparables a valores obtenidos en estudios previos en Xel-Há. A pesar de un aumento significativo en la densidad poblacional de S. gigas en el 2009-2011, el crecimiento incrementó, indicando que el área, disponibilidad de alimentos y/o calidad del hábitat no son limitantes del crecimiento y tamaño de población. El reclutamiento fue continúo pero mostró fluctuaciones a en el tiempo, con picos de marzo a mayo y de junio a agosto, pero también varió en intensidad entre años, lo cual posiblemente tiene impacto sobre el tamaño de población. Por lo tanto se concluye que el reclutamiento podría ser uno de los factores de mayor importancia para la regulación del tamaño de población de S. gigas en la caleta de Xel-Há. Las tasas de mortalidad de S.gigas obtenidas en este trabajo presentaron valores dentro de los rangos reportados en la literatura. Sin embargo se detectó alta mortalidad en adultos, lo cual se pudo atribuir a un incremento en la movilidad de esta fracción de la población y su posible emigración fuera de la caleta y por ende fuera del área de estudio de este trabajo. Se observó que la movilidad de los organismos incrementó en los meses de verano, particularmente en la temporada de Lluvias. La migración observada en este estudio establece la conectividad entre la distribución de juveniles y adultos en el espacio. La alta abundancia de juveniles, la permanencia y continuidad de la población y de su estructura de tallas en el espacio y tiempo, así como las altas tasas de crecimiento encontradas en Cueva, Centro y Brazo Norte, son elementos para considerar que la caleta de Xel-Há está funcionando como zona de crianza para el caracol rosa. La alta abundancia de adultos en Bocana, sugiere que 176 esta zona no forma parte del criadero y que ésta desempeña otro papel ecológico y existen evidencias que es una zona de reproducción. Con base en el estudio de la estructura, dinámica y movilidad de la población epibentónica en el espacio y en el tiempo se pudo determinar que el caracol S.gigas hace uso la caleta de Xel-Há como zona de crianza, donde completa la etapa juvenil de su ciclo de vida y migra hacía los límites del área de estudio y posiblemente fuera de este al alcanzar la madurez sexual. Este estudio aporta elementos útiles para la conservación, manejo y rehabilitación de esta especie emblemática del Caribe e información útil a la gestión y el manejo sustentable del parque de XelHá, permitiendo la conservación de su biodiversidad y el uso ecoturístico del parque. 177 11. Literatura Citada A Abott, R. T. 1974. American Seashells; The Marine Molluska of the Atlantic and Pacific Coasts of North America. 2nd ed. Van Nostrand Reinhold Company, New York City, NY. 663 pp. Acosta, C.A. 2006. Impending trade suspensions of Caribbean Queen Conch under CITES: A case study on fishery impacts and potential for stock recovery. Fisheries 31: 601-606. Alcolado, P. M. 1976. Crecimiento, variaciones morfológicas de la concha y algunos datos biológicos del cobo Strombus gigas L. (Mollusca, Mesogastropoda). Acad. Cienc. Cub Ser. Oceanol. 34: 1-36. Aldana-Aranda, D. & T. Brulé. 1994. Estado actual de la pesquería, del cultivo y programa de investigación del caracol Strombus gigas en México. In: Appeldoorn, R.S. & B. Rodrigo (eds.) Queen Conch Biology, Fisheries and Mariculture. Fundación Científica Los Roques, Caracás, Venezuela. Aldana-Aranda, D, Baqueiro Cárdenas, E. & S. Manzanilla Naim. 2003. Mexican marine parks as a fishery management tool for the queen conch Strombus gigas. In: Aldana-Aranda, D. (ed.) El caracol rosa Strombus gigas: conocimiento integral para su manejo sustentable en el Caribe. México, CYTED-Press, pp. 101-108. Aldana-Aranda, D., M. Sánchez-Crespo, P. Reynaga Alvarez, V. Patiño-Suárez, A. GeorgeZamora & E.R. Baqueiro-Cárdenas. 2005. Crecimiento y temporada reproductiva del caracol rosa Strombus gigas en el parque Xel-Há, México. Proc. Gulf Carib. Fish. Inst.56: 741-754. Aldana Aranda, D. 2006. Overview reproductive pattern of Queen conch, Strombus gigas in different localities in the Caribbean. Proc. Gulf Carib. Fish. Inst. 57: 771-789. Aldana Aranda, D. & L. Frenkiel. 2007. Lip thickness of Strombus gigas, Mollusca Gastropoda versus maturity: a management measure. Proc. Gulf Carib. Fish. Inst. 58:431-442. Aldana-Aranda, D. & M. Perez-Perez. 2007. Abundance and distribution of Queen conch (Strombus gigas, Linne 1758) veligers of Alacranes reef, Yucatan, Mexico. J. Shellf. Res. 26: 5963. Appeldoorn, R.S. 1987a. Practical considerations in the assesment of Queen Conch Fisheries and Population dynamics. Proc. Gulf. Carib. Fish. Inst. 38: 307-324. Appeldoorn, R.S. 1987b. Modification of a seasonally oscillating growth function for use with mark-recapture data. J. Cons. CIEM 43: 194-198. Appeldoorn, R.S. 1987c.Assessment of mortality in an offshore population of queen conch, Strombus gigas L., in Southwest Puerto Rico. Fish. Bull. 85(4): 797-804. 178 Appeldoorn, R.S. 1988. Age determination, growth, mortality and age of first reproduction in adult queen conch, Strombus gigas L., off Puerto Rico. Fish. Res. 6: 363-378 Appeldoorn, R.S. 1990. Growth of Juvenile Queen Conch, Strombus gigas, L., off La Parguera, Puerto Rico. J. Shellfish Res. 9: 59-62. Appeldoorn, R.S. 1994. Spatial variability in the morphology of queen conch and its implication for management regulations. In: R.S. Appeldoorn & B. Rodriguez (eds.). Queen conch biology, fisheries and management. Fundación Científica Los Roques, Caracas, pp. 145-158. Appeldoorn, R.S. 1997. Deep Water Spatial Variability in the Morphology of the Queen conch and its Implication for Management Regulations. In: CFRAMP (eds.): Lobster and Conch Subproject Specification and Training Workshop Proceedings, 9-12 October, Kingston, Jamaica. CARICOM Fishery Research Document No. 19. Appeldoorn, R.S. 2003. Age determination, growth, mortality and age of first reproduction in adult Queen Conch, Strombus gigas L., off Puerto Rico. Fish. Res. 6 (4): 363-378. Ávila-Poveda O.H. & E.R. Baqueiro-Cárdenas. 2006. Size at sexual maturity in the queen conch Strombus gigas from Colombia. Bol. Invest. Mar. Cost. 35: 223-233. B Baqueiro-Cárdenas, E. & D. Aldana-Aranda. 2010. Histories of Success for the Conservation of Populations of Queen Conch (Strombus gigas). Proc. Gulf Carib. Inst. 62: 306-312. Berg, C. J. 1975. Behavior and ecology of conch (Superfamily Strombacea) on a deep subtidal algal plain. Bull. Mar. Sci. 25: 307-317. Berg, C.J. 1976. Growth of the queen conch Strombus gigas with a discussion of the practicality of its mariculture. Mar. Biol. 34 (3): 191-199. Berg, C.J. & R.A.Glazer. 1991. Current research on queen conch (Strombus gigas) in Florida Waters. Proc. Gulf Carib. Fish. Inst.40: 303-306. Berg, C. J., & R. A. Glazer. 1995. Stock assessment of a large marine gastropod (Strombus gigas) using randomized and stratified towed-diver censusing. ICES Marine Science Symposium 199: 247- 258. Berg, C. J. & D. A. Olsen. 1989. Conservation and management of queen conch (Strombus gigas) fisheries in the Caribbean. pp. 421-442. In: J. F. Caddy (ed.). Marine Invertebrate Fisheries: Their Assessment and Management. Wiley, New York. Bissada-Gooding, C. E. & H.A. Oxeford. 2010. Estimating Home Range and Density of a Queen Conch Aggregation using Acoustic Telemetry and Conventional Tagging. Proc. Gulf Carib. Inst. 62: 384-389. 179 Blakesley, H.L. 1977. A contribution to the fisheries and biology of the queen conch, Strombus gigas L. in Belize. Amer. Fish. Soc. Ann. Meeting 107. Bos, A.R., Gumanao, G.S., van Katwijk, M.M., Mueller, B., Saceda, M.M. & R.L.P. Tejada. 2011. Ontogenetic habitat shift, population growth, and burrowing behavior of the Indo-pacific beach star, Archaster typicus (Echinodermata; Asteroidea). Mar. Biol. 158 (3): 639-648. Bravo-Castro, H.M., Montero-Muñoz, J.L., & D. Aldana-Aranda. 2010. Abundance and distribution of queen conch (Strombus gigas) veligers of Quintana Roo, Peninsula of Yucatan, Mexico. Proc. Gulf Carib. Fish. Inst.62: 338-440. Bronwyn, M.G., Able, K.W, Brown, J.A., Eggleston, D.B. & P.F. Sheridan. 2003. Evidence of connectivity between juvenile and adult habitats for mobile marine fauna: an important component of nurseries. Mar. Ecol. Prog. Ser. 247: 281-295. Brownell, W.N. 1977. Reproduction, laboratory culture and growth of Strombus gigas, Strombus costatus and Strombus pugilis in Los Roques, Venezuela. Bull. Mar. Sci. 27:668-680. Brownell, W.N. & J.M. Stevely. 1981. The biology, fisheries and management of the Queen conch Strombus gigas. Mar Fish Rev, US Dept Comm. 43(7):1-12. C CFMC QCSAMW, 1999. Proceedings of the Queen Conch Stock Assessment and Management Workshop, Belize City, Belize March 15-22, 1999. Caribbean Fishery Management Council. Chakalall, B. & K.L. Cochrane. 1996. The Queen conch fisheries in the Caribbean – An approach to responsible fisheries management. In: Posada, J.M. & G. Garcia-Moliner (eds.): Proceedings of the First International Queen conch Conference, San Juan, Puerto Rico, 29-31 July 1996. Caribbean Fisheries ManagementCouncil. p. 60-76. Chávez, E. A. & F. Arreguín-Sánchez. 1994. Simulation modelling for conch fishery management, p. 125-136. En: Appeldoorn, R. S. & Q. Rodríguez (eds). Queen Conch Biology, Fisheries and Mariculture. Fundación Científica Los Roques, Caracas Venezuela, p.169-189. Chávez-Villegas, J.F, Cid-Becerra, J.A, Enríquez-Díaz, M.R., Montero-Muñoz, J.L. & D. Aldana-Aranda. 2009. Abundancia de Larvas Veliger de Strombus gigas (Linnaeus, 1758) en el Sistema Arrecifal Mesoamericano. Proc. Gulf Carib Fish. Inst. 62: 433-437. Childress, M.J. & W. F. Herrnkind. 1994. The Behavior of Juvenile Caribbean Spiny Lobster in Florida Bay: Seasonality, Ontogeny and Sociality. Childress, M.J. & W. F. Herrnkind. 2001. Influence of conspecifics on the ontogenetic habitat shift of juvenile Caribbean spiny lobsters. Mar. Fresh. Res. 52(8): 1077 – 1084. CITES, 2003. Review of significant trade in specimens of Appendix-II species. Report of the Convention on International Trade in Endangered Species of Wild Fauna and Flora. www.cites.org/eng/resols/12/12-8.shtml 180 D D'Asaro, C.N. 1965. Organogenesis, development and metamorphosis in the queen conch, Strombus gigas, with notes on breeding habits. Bull. Mar. Sci. 15:359-416. Danylchuk, A., R.A. Mudd, G. Ingelise, & K. Baldwin. 2003. Size-dependent habitat use of juvenile queen conch (Strombus gigas) in East Harbour Lobster and Conch Reserve, Turks and Caicos Islands, BWI. Proc. Gulf Carib. Fish. Inst. 54:241-249. Davis, M. & A.W. Stoner. 1993. Trophic cues induce metamorphosis of Queen conch larvae (Strombus gigas Linnaeus). J. Exp. Mar. Biol. Ecol. 180:83-102. Davis, M., B.A. Mitchell, J.L. Brown. 1984. Breeding behaviour of the queen conch S. gigas held in natural and enclosed habitat. J. Shell. Res. 4 (1): 17-21. Davis, M., Bolton, C.A. & A.W. Stoner. 1993. A comparison of larval development, growth, and shell morphology in three Caribbean Strombus species. Veliger 36:236-244. De Jesús-Navarrete, A., E. González, J. Oliva, A. Pelayo & G. Medina. 1992. Advances over some ecological aspects of queen conch Strombus gigas, L. in the southern Quintana Roo, Mexico. Proc. Gulf Carib. Fish. Inst. 45: 18-24. De Jesús-Navarrete, M., Oliva-Rivera, Medina-Quej, A. & J. A., Domínguez-Viveros. 1994. Crecimiento, reclutamiento y estructura poblacional del caracol rosado Strombus gigas, en Punta Gavilán Quintana Roo México. Proc. Gulf Carib. Fish. Inst. 46: 74-83. De Jesús-Navarrete, A. & J.J. Oliva-Riviera. 1997. Density, growth, and recruitment of the queen conch Strombus gigas L. (Gastropoda: Strombidae) in Quintana Roo, Mexico. Rev. Biol. Trop. 45: 797-801. De Jesús-Navarrete, A. & D. Aldana Aranda. 2000. Distribution and abundance of Strombus gigas veligers at six fishing sites on Banco Chinchorro, Quintana Roo, Mexico. J. Shellf. Res. 19(2):891-896. De Jesús-Navarrete, A., M. Domínguez-Viveros, A. Medina-Quej& y J. J. Oliva-Rivera. Crecimiento, mortalidad y reclutamiento del caracol Strombus gigas en Punta Gavilán, Q. Roó, México. Ciencia Pesquera 4: 1-4. De Jesús-Navarrete, A. 2001a. Crecimiento del caracol Strombus gigas (Gastropoda: Strombidae) en cuatro ambientes de Quintana Roo, México. Rev. Biol. Trop. 49: 85-91. De Jesús-Navarrete, A. 2001b. Distribución y abundancia de larvas veligeras de Strombus gigas en Banco Chinchorro, Quintana Roo, México. Proc.Gulf Carib.Fish. Inst. 52:174-183. De Jesús-Navarrete, A. & V. Valencia-Beltrán. 2003. Abundance of Strombus gigas zero-year class juveniles at Banco Chinchorro Biosphere Reserve, Quintana Roo, Mexico. Bull. Mar. Sci. 73: 232-240. 181 De Jesús-Navarrete, A. & M. Pérez-Flores. 2007. Distribución y Abundancia de Larvas de Strombidae en la Bahía de la Ascensión, Quintana Roo, México. Proc. Gulf Carib. Fish. Inst. 58: 388-393. De la Torre, A.R. 1982. Diagnóstico de la pesquería de caracoles en el estado de Quintana Roo. Centro de Investigaciones pesqueras de Isla Mujeres, Q. Roo, 25 pp. Delgado, G.A., Bartels, C. T., Glazer, R.A., Brown-Petersen, N.J. & K.J. McCarthy. 2004. Translocation as a strategy to rehabilitate the queen conch (Strombus gigas) population in the Florida Keys. Fish. Bull. 102(2): 278-288. Delgado, D.A., Glazer, R.A., Hawtof, D., Aldana-Aranda, D., Rodríguez-Gil, L.A & A. De Jesús-Navarrete. 2008. Do Queen Conch (Strombus gigas) Larvae Recruiting to the Florida Keys Originate from Upstream Sources? Evidence from Plankton and Drifter Studies. Proc. Gulf Carib. Fish. Inst. 59: 29-41. Díaz Ávalos, C.1986. Crecimiento y mortalidad de juveniles de caracol rosado Strombus gigas en Punta Gavilán, Q. Roo, INP. Serie documentos de trabajo, No. 3. 16 pp. Díaz Ávalos, C. 1991. Crecimiento y mortalidad de juveniles de caracol rosado Strombus gigas en Punta Gavilán, Q. Roo, México. Algunas consideraciones sobre el aprovechamento de su pesquería. Cienc. Pesqu. 8: 63-70. Doerr, J. C. & R. L. Hill. 2007. A Preliminary Analysis of Habitat Use, Movement, and Migration Patterns of Queen Conch, Strombus gigas, in St. John, USVI, Using Acoustic Tagging Techniques. Proc. Gulf. Carb. Fish. Inst. 60: 509-515. Domínguez-Viveros, M., De Jesús-Navarrete, A., Medina-Quej & J. Oliva-Rivera. 1994. Estado actual de la Población de Strombus gigas en la Zona Sur en Quintana Roo, México. Proc. Gulf. Carb. Fish. Inst. 46: 84-95. Domínguez-Viveros, M., Sosa-Cordero, E. & A. Medina-Quej. 1999. Abundancia y parámetros poblacionales del caracol Strombus gigas en Banco Chinchorro, Quintana Roo, Mexico. Proc. Gulf Carib. Fish. Inst. 45: 943-954. E Ehrhardt, N.M. & M. Valle-Esquivel. 2008. Conch (Strombus gigas) Stock assessment manual. Caribbean Fishery Management Council, San Juan, Puerto Rico. 128 pp. Egan, B.D. 1985. Aspects of the reproductive biology of Strombus gigas. MSc Thesis. Univ. British Colombia. F Friedlander, A., R.S. Appeldoorn & J. Beets. 1994. Spatial and temporal variations in stock abundance of queen conch, Strombus gigas, in the U.S. Virgin Islands, p. 51-60 In R.S. 182 Appeldoorn & B. Rodriguez (eds.). Queen Conch Biology, Fisheries and Mariculture, Fundacion Cientifica Los Roques, Caracas, Venezuela. G García, M.I. 1991. Biología y dínámica poblacional del caracol de pala Strombus gigas L. (Mollusca, Mesogastropoda) en las diferentes áreas del Archipiélago de San Andrés y Providencia. Tesis Biología. Facultad de Ciencias Universidad del Valle. Colombia. 166 pp Gibson, J., Strasdine, S. & R. González. 1983. The Status of the Queen Conch industry. Proc. Gulf Carib. Fish. Inst. 35: 43-45. Glazer, R.A. & C.J. Berg, Jr. 1994. Queen conch research in Florida: an overview. pp. 79-96. In: Appeldoorn, R.S. & B. Rodríguez (eds.). Queen conch biology, fisheries, and mariculture. Fundación Científica Los Roques. Glazer, R.A., G.A. Delgado & J.A. Kidney. 2003. Estimating queen conch (Strombus gigas) home ranges using acoustic telemetry: implications for the design of marine fishery reserves. Gulf Carib. Res.14(2):79-89. Glazer, R. & J. Kidney. 2004. Habitat associations of adult Queen Conch (Strombus gigas L.) in an unfished Florida Keys back reef: applications to essential fish habitat. Bull. Mar. Sci. 75(2):205-224. H Hesse, K.O. 1979. Movement and migration of the queen conch, Strombus gigas, in the Turks and Caicos Islands. Bull. Mar. Sci. 29:303-311. I INP-SAGARPA. 2010. Dictamen Técnico: Estimación de biomasa explotable de Strombus gigas en los bancos abiertos a la pesca en Quintana Roo, México: Banco Chinchorro y Banco de Cozumel. Temporada de Captura 2008-2009. Iversen, E.S., E.S. Rutherford, S.P. Bannerot & D.E. Jory. 1987. Biological data on Berry Islands (Bahamas) queen conch, Strombus gigas, with mariculture and fisheries management implications. Fish. Bull. 85: 299-310. Iversen, E.S., Jory, D.E. & D.J. Diresta. 1994. Research on First Year Queen Conch Relevant to Fisheries Management. Proc. Gulf Carib. Fish. Inst. 43: 498-505. J Jolly, G. M. 1965. Explicit estimates from capture recapture data with both death and immigration-stochastic model. Biometrika 52, 225-247. K 183 Krebs, C. J. 1985. Ecología: Estudio de la Distribución y Abundancia. Harla.México.753 p. Kronenberg, G.C. & G.J. Vermeij. 2002. Terestrombus and Tridentarius, new genera of IndoPacific Strombidae (Gastropoda), with comments on the included taxa and on shell characters in Strombidae. Vita Malacologica 1: 49–54. L Latiolais, J. M., Taylor, M.S., Roy, K. & M.E. Hellberg. 2006. A molecular phylogenetic analysis of strombid gastropod mprphological diversity. Mol. Phyl. Evol. 41: 436-444. M Márquez, E. 1993. Biología poblacional y pesuqra del caracol Strombus gigas Linneaus, 1785 en las Islas de Providencia y Santa Catalina. Univ. Jorge Tadeo Lozano. Tesis Profesional de Biología Marina. 102 pp. Mitton, J. B., C. J. Berg, Jr. & K. S. Orr. 1989. Population structure, larval dispersal, and gene flow in the queen conch, Strombus gigas, of the Caribbean. Biol. Bull. 177:356-362. Moreno de la Torre, R. & D. Aldana-Aranda. 2007. Crecimiento y eficiencia alimenticia del caracol rosado Strombus gigas (Linnaeus, 1758) juvenil alimentado con nueve alimentos balanceados conteniendo diferentes niveles de proteína y energía. Proc. Gulf Carib. Fish. Inst.58: 463-467 N Nixon, S.W. 1982. Nutrient dynamics, primary production and fisheries yields of lagoons. Oceanologica Acta: 357-371. O Oliva-Rivera, J. & A. De Jesús-Navarrete. 2000. Composición, distribución y abundancia de larvas de moluscos gasterópodos en el sur de Quintana Roo, México y norte de Belice. Rev. Biol. Trop. 48 (4): 777-783. Oviatt, C., Doering, P., Nowiclu, B., Reed, L., Cole, J. & J. Frithsen. 1995. An ecosystem level experiment on nutrient limitation in temperate coastal marine environments. Mar. Ecol. Prog. Ser. 116: 171-179. P Paris, C.B., Aldana-Aranda, D., Pérez-Pérez, M. & J. Kool. 2008. Connectivity of Queen conch, Strombus gigas, populations from Mexico. Proc. Int. Coral Reef Symp. 11: 439-443. Pauly, D. & G.R. Morgan (Eds.). 1987. Length-based Methods in Fisheries Research. ICLARM Conf. Proc. 13. International Center for Living Aquatic Resources Management, Manila, and Kuwait Institute for Scientific Research, Safat, 468 pp. 184 Pauly, D. & H. Gaschutz. 1979. A simple method for fitting oscillating length growth data, with a program for pocket calculators. International Council for the Exploration of the Sea, Copenhagen. Pérez-Pérez, M. & D. Aldana-Aranda. 1998. Análisis preliminar de la densidad del caracol rosado (Strombus gigas) en el arrecife Alacranes Yucatán, México. Proc. Gulf Carib. Fish. Inst. 50: 49- 65. Pérez-Pérez, M. & D. Aldana-Aranda. 2000. Distribución, abundancia, densidad y morfometría de Strombus gigas (Mesogastropoda:Strombidae) en el Arrecife Alacranes Yucatán, México. Rev. Biol. Trop. 48: 51-57. Pérez-Pérez, M. & D. Aldana-Aranda. 2003. Actividad de Strombus gigas (Mesogastropoda: Strombidae) en diferentes hábitats del arrecife Alacranes, Yucatán. Rev.Biol. Trop. 51: 119-126. Phillips, M. Bissada-Gooding, C. & H.A. Oxenford. 2010. Preliminary Investigation of the Movements, Density, and Growth of Juvenile Queen Conch in a Nursery Area in Barbados. Proc. Gulf Carib. Fish. Inst. 63: 427-434. Posada, J.M., R.I. Mateo & M. Nemth. 1999. Occurrence, abundance and length frequency distribution of queen conch, Strombus gigas (Gasteropoda) in shallow waters of the Jaragua National Park, Dominican Republic. Carib. J.of Sci. 35: 70-82. Pollock, K. H. 1980. Capture-Mark-Recaptures Models: A Review of current methods, assumptions and experimental design. North Carolina State University. 32 pp. R Randall, J.E. 1964. Contribution to the biology of the Queen conch Strombus gigas. Bull. Mar. Sci. Gulf Carib. 14:246-295 Rathier, I. & A. Battaglya. 1994. Assesment of the Martinique Queen Conch fishery and management prospective. En: Appeldoorn, R.S. & B. Rodriguez (eds.) Queen Conch biology, fisheries and mariculture. Fundación Los Roques, Caracás, Venezuela. Ray, M. & A. W. Stoner. 1994. Experimental analysis of growth and survivorship in a marine gastropod aggregation: balancing growth with safety in numbers. Mar. Ecol. Prog. Ser. 105:4759. Ray-Culp, M. Davis, M. & A.W. Stoner. 1999. Predation by xanthid crabs on early post settlement gastropods: the role of prey size, prey density, and habitat complexity. J. Exp. Mar. Biol. Ecol. 240: 303-321. Rios-Lara, G.V., K. Cervera-Cervera, J.C. Espinoza-Méndez, M. Pérez-Pérez, C. Zetina-Moguel, & F. Chable Ek. 1998. Estimación de las Densidades de langosta espinosa (Panulirus argus) y caracol rosa (Strombus gigas) en el área central del Arrecife Alacranes, Yucatán, México. Proc. Gulf Carib. Fish. Inst.50:104-127. 185 Robertson, R. 1959. Observations on the spawn and veligers of conchs (Strombus) in the Bahamas. Proc. Malacol. Soc. Lond. 33:164-171. Roughgarden, J., Gaines, S. & H. Possingham. 1988. Recruitment dynamics in complex life cycles. Science 228: 1460-1466. Ryther, J.H. & W.M. Dunstan. 1971. Nitrogen, phosphorus and eutrophication in the coastal marine environment. Science 171: 1008-1013. S Schindler, D.W. 1974. Eutrophication and recovery in experimental lakes. Science 184: 897-899. Schnabel, Z. E. 1938. The estimation of the total fish population of a lake. Amer. Math. Month. 45:348-352. Schoener, T.W. 1967. The ecological significance of sexual dimorphism in size in the lizard Anolis cospercus. Science 155: 474-477. Seber, G. A. F. 1965. A note on the multiple recapture census. Biometrika 52: 249-259. Shawl, A.L. & M. Davis. 2004. Captive Breeding behavior of four Strombidae conch. J. Shellf. Res. 23 (1): 157-164. Stevely, J.M. 1979. The biology and fishery of the queen conch (Strombus gigas); a review. Proc. Fourth Annu. Trop. Subtrop. Fish. Technol. Conf. Americas: 203-210. Stoner, A. W., Lipcius, R. N., Marshall, L. S. & A.T. Jr, Bardales. 1988. Synchronous emergence and mass migration in juvenile queen conch. Mar. Ecol. Prog. Ser. 49: 51-55. Stoner, A.W. 1989a. Density-dependent growth and grazing effects of the juvenile Queen conch Strombus gigas L. in a tropical seagrass meadow. J. Exp. Mar. Biol. Ecol. 130:119-133. Stoner, A.W. 1989b. Winter mass migration of juvenile queen conch Strombus gigas and their influence on the benthic environment. Mar. Ecol. Prog. Ser. 56:99–104 Stoner A.W. & J.M. Waite. 1990. Distribution and behavior of queen conch Strombus gigas relative to seagrass standing crop. Fish. Bull. 88:573-585. Stoner A.W. & J.H. Waite. 1991. Trophic biology of Strombus gigas in nursery habitats: diets and food sources in seagrass meadows. J. Mollusc. Stud. 57:451-460. Stoner, A.W., Sandt, V.J. & I.F. Boidron-Metairon. 1992. Seasonality of reprodutucive activity and abundance of veligers in queen conch, Strombus gigas. Fish. Bull. 90:161-170. Stoner, A. W. & V. J. Sandt. 1992. Population structure, seasonal movements and feeding of queen conch, Strombus gigas, in deep water habitats of the Bahamas. Bull. Mar. Sci. 51:287-300. 186 Stoner, A.W. & M. Ray. 1993. Aggregation dynamics in juvenile Queen conch (Strombus gigas): population structure, mortality, growth and migration. Mar. Biol. 116:571-582. Stoner, A. W., Davis, M. 1994. Experimental outplanting of juvenile queen conch, Strombus gigas: comparison of wild and hatchery-reared stocks. Fish. Bull. U.S. 92: 390-411. Stoner, A.W. & J. Lally. 1994. High-density aggregation in Queen conch Strombus gigas: formation, patterns, and ecological significance. Mar. Ecol. Prog. Ser. 106:73-84. Stoner, A.W. & K.C. Schwarte. 1994. Queen conch, Strombus gigas, reproductive stocks in the central Bahamas: distribution and probable sources. Fish. Bull. 92:171-179. Stoner, A. W.. J. Lin & M. D. Hanisak. 1995. Relationships between seagrass bed characteristics and juvenile queen conch (Strombus gigas) abundance in the Bahamas. J. Shellfish Res. 14:315323. Stoner, A. W., P. L. Pitts & R. A. Armstrong. 1996. The interaction of physical and biological factors in the large-scale distribution of juvenile queen conch in seagrass meadows. Bull Mar. Sci. 58:217-233. Stoner, A.W. 1996. Status of the Queen conch research fishery in the Caribbean Sea. In: Posada J.M. & A.Garcia- Moliner (eds.): Proceedings of the First International Queen conch Conference, San Juan, Puerto Rico, 29- 31 July 1996. Caribbean Fisheries Management Council. p. 23-39. Stoner, A.W. & M. Ray. 1996. Queen Conch, Strombus gigas, in fished and unfished locations of the Bahamas: effects of a marine fishery reserve on adults, juvenile and larval production. Fish. Bull. 94: 551-565. Stoner, A.W. 1997. The status of queen conch Strombus gigas research in the Caribbean. Mar. Fish. Res. 59(3): 14-22. Stoner, A.W. 2003. What constitutes essential nursery habitat for a marine species? A case study of habitat form and function for queen conch. Mar. Ecol. Prog. Ser. 257: 275-289. Stoner, A.W. & M. Ray-Culp. 2000. Evidence for Allee effects in an over-harvested marine gastropod: density-dependent mating and egg production. Mar. Ecol. Prog. Ser. 202: 297-302. Strasdine, S.1988. The queen conch fishery of Belize: an assessment of resource harvest sector and management. MsC. Thesis Univ. of British Columbia, Cánada, 216 pp. T Theile, S. 2001. Queen conch fisheries and their management in the Caribbean. TRAFFIC Tello, C.J.A., Rodríguez-Gil, L.A. & F. Rodríguez-Romero. 2005. Population genetics of the pink snail Strombus gigas in the Yucatan Peninsula: Implications for its management and fishery. Cienc. Mar. 31: 379-386. 187 Tewfik, A. 1996. An assessment of the biological characteristics, abundance and potential yield of the queen conch (Strombus gigas L.) fishery on the Pedro Bank off Jamaica. M. Sc. Thesis. Acadia University, Wolfville, Nova Scotia. 139p. Tewfik, A., Guzman, H.M. & G. Jacome. 1998. Assessment of the Queen conch Strombus gigas (Gastropoda: Strombidae) population in Cayos Cochinos, Honduras. Rev. Biol. Trop. 46.(4):137150. Tewfik. A. & C. Bené. 2000. Densities and age structure of fished versus protected populations of queen conch (Strombus gigas L.) in the Turks & Caicos Islands. Proc. Gulf Carib. Fish. Inst. 51: 60-79. V Valle-Esquivel, M. 1998. Growth of queen conch Strombus gigas in Xel-ha, Quintana Roo, Mexico. Proc. Gulf Carib. Fish. Inst. 50: 66-77. W Wicklund, R.I, Hepp, L.J. & G.A. Wenz. 1991. Preliminary studies on the early life history of the queen conch Strombus gigas in the Exuma Cays, Bahamas. Proc. Gulf Carib. Fish. Inst. 40: 283-298. Wilson, D.S. 1975. The adequacy of body size as a niche difference. Am. Nat. 109: 769-784. Weil, M.E. & G.R. Laughlin. 1984. Biology, Population Dynamics and reproduction of the queen conch Strombus gigas Linné in the Archipelago de los Roques National Park. J. Shellf. Res. 4: 45-62. Werner, E.E. & J.F. Gilliam. 1984. The ontogenetic niche and species interactions in sizestructured populations. An. Rev. Ecol. Syst. 15:393-425. Wefer, G. & J.S. Killingley. 1980. Growth histories of strombid snails from Bermuda recorded in their O-18 and C-13 profiles. Mar. Biol. 60:129–135 Woods, E.M. 1995. Study of the Fishery for Queen conch in Haiti. Marine Conservation Society. Ross-onWye, UK. 57pp. 188 189