Atenuación genética
Anuncio

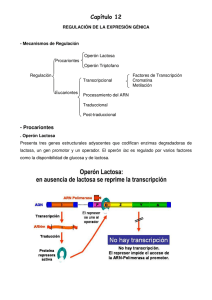
ATENUACIÓN Este proceso fue descubierto por Charles Yanofsky con el operón del triptófano de E. coli . La iniciación de la transcripción depende de un represor activado por triptófano que puede bloquear el acceso de la polimerasa del ARN al promotor. En la atenuación, la transcripción no se extiende siempre para producir ARN mensajero completo, esto es una atenuación. Se produce un ARN m corto. El que se dé atenuación depende de la estructura espacial del ARN y esta estructura depende de la frecuencia de traducción por ribosomas. Cuando hay suficiente triptófano el ARNm se traduce normalmente y adopta una estructura secundaria que da lugar a la terminación prematura de la transcripción. La escasez del triopófano retrasa la traducción desde el comienzo del mensaje y el ARN m adopta un estructura secudaria distinta que permita a la polimerasa continuar la transcripción. La escasez de triptófano estimula la expresión del operón del triptófano. Elimina al represor y evita la atenuación. La expresión del operón puede aumentar unas 600 veces. REGULACIÓN DE LA TRADUCCIÓN Una eficaz iniciación de la traducción exige que haya un grupo de seis a ocho nucleótidos, en su mayoría purinas, poco antes del triplete iniciador AUG. Este lugar de unión de robosomas en conocido como la secuencia de Shine y Dalgarno. Se traducen con más frecuencia los ARNm que tienen una secuencia de Shine y Dalgarno centrada ocho nucleótidos antes del triplete iniciador. Las mutaciones de esta secuencia pueden inhibir la traducción del ARNm. Sin embargo una secuencia de Shine y Delgardo bien situada no garantiza la iniciación correcta de la traducción. La eficacia de la secuencia de Shine y Delgarno puede ser modulada por proteínas ribosómicas que se unen a ellas y las bloquean. Cuando se producen más proteínas ribosómicas que ARNr, algunas de las que quedan libres se unen a las secuencias de S. y D. de esta manera las proteínas no se sintetizan más deprisa de lo que se usan para elaborar ribosomas. E. coli contiene varios operones y cada operón contiene una proteína clave que inhibe la traducción de todo el operón. Las proteínas clave se unen también al ARNr. Por lo tanto, la regulación de la traducción implica una competición de ambos ARN por las proteínas clave. DIFICULTADES INICIALES EN EL ESTUDIO DE LA REGULACIÓN GÉNICA EN PLANTAS Y ANIMALES SUPERIORES A finales de los sesentas los intentos de estudiar las señales que regulan los genes y como se coordina la expresión de tantas proteínas fracasó. Estas señales son hormonas esteroides que se unen a un receptor específico en el citoplasma, la señal se traslada al núcleo, donde se une a los cromosomas y se activa la síntesis de proteínas específicas. No se pueden hacer cruces genéticos entre células superiores cultivadas. 1