tema 9- sistema vacuolar- ribosomas
Anuncio

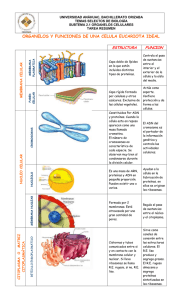
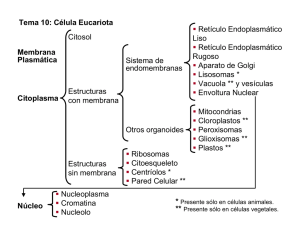
11. SISTEMA VACUOLAR- RIBOSOMAS 1. RETÍCULO ENDOPLASMÁTICO. ♦ MORFOLOGÍA ♦ FUNCIÓN. 2. APARATO DE GOLGI. ♦ ESTRUCTURA. ♦ FUNCIÓN. 1. RIBOSOMAS 2. LISOSOMAS. 3. PEROXISOMAS. 4. VACUOLAS. 1. RETÍCULO ENDOPLASMÁTICO. Es un conjunto de cavidades cerradas de 300 A de espesor que tienen partes dilatadas y de formas variables: láminas aplanadas, vesículas globulares, tubos contorneados. Forman el 10% del contenido celular, se comunican entre sí y forman una red continua, separada del hialoplasma por una membrana. ♦ ESTRUCTURA Hay dos tipos de RE: • RE rugoso, con ribosomas adosados en el lado de la membrana que da al hialoplasma, los ribosomas se unen por la subunidad grande a la membrana. • RE liso, sin ribosomas. Ambos coexisten en una misma célula e incluso en una misma cavidad. La membrana que forma el RE es semejante, aunque algo más delgada (50- 60 A) que la membrana plasmática. Constituye más de la mitad de todas las membranas presentes en la célula animal y tiene la particularidad de que se halla en continuidad estructural con la membrana nuclear. La composición química de la membrana, es también semejante, pero tiene menor cantidad de lípidos (30%) y mayor cantidad de proteínas (70%). Estas proteínas son en su mayoría enzimas, encargadas de transportar electrones, intervenir en el metabolismo de ácidos grasos, sintetizar fosfolípidos y esteroides, glucoproteínas y glucolípidos. Las cavidades contienen una solución acuosa rica en holo, gluco y lipoproteínas, aunque su composición depende de cada tipo celular, estado fisiológico y especie. ♦ FUNCIONES: LA MEMBRANA DEL RE RUGOSO: interviene en la síntesis de proteínas debido a los ribosomas que tiene adosados en su membrana. Las proteínas que se forman en el RER pueden tener dos destinos: • Formar parte de los productos de secreción celular en cuyo caso son transferidas al interior de las cavidades por donde circulan hasta ser llevadas o otros orgánulos o al exterior. 169 • Formar parte de las membranas celulares en cuyo caso quedan ancladas en la membrana del RER, para pasar posteriormente a formar parte de alguna otra membrana celular. Las proteínas que se sintetizan en el RER están glicosiladas, esto significa que se encuentran unidas a un oligosacárido, este oligosacárido puede estar formado por una o varias cadenas con osas diferentes según secuencias definidas. También interviene en el almacenamiento de proteínas, en su interior, se almacenan las proteínas que han sido previamente sintetizadas. Estas proteínas se hallan unidas a chaperonas (proteínas acompañantes, cuya función consiste en mantener la forma de otras proteínas, repararlas o eliminarlas), que acilitan el plegamiento y eitan su precipitación El RER se va a encontrar en todas las células eucariontes, excepto los glóbulos rojos, y su abundancia dependerá de la actividad celular, siendo mayor cuanto mayor sea también la actividad secretora de la célula (glándulas, neuronas...). LA MEMBRANA DEL RE LISO está involucrado en tres funciones diferentes: • Biosíntesis de lípidos: sobre todo fosfolípidos (a partir de glicerol-3-P y ácidos grasos) y colesterol, sólo los ácidos grasos se sintetizan en el hialoplasma, aunque aquí se elongan e insaturan, por lo tanto en el REL se sintetizan los componentes fundamentales de las membranas celulares; Estos son exportados en vesículas, lo mismo que el contenido de las cavidades; dichas vesículas se desprenden y se dirigen hacia la membrana de un orgánulo concreto o hacia la membrana plasmática en donde se integran. También aquí el colesterol se transforma en ácidos biliares y hormonas. Los lípidos sintetizados en el REL pueden unirse a proteínas que provienen del RER y formar lipoproteínas. Esta función implica que el REL será muy abundante en células especializadas en el metabolismo de lípidos, como los hepatocitos. • Detoxificación: en las membranas del REL existen enzimas capaces de eliminar la toxicidad d aquellas sustancias que resultan perjudiciales para la célula, ya sean producidas por ella misma, como hormonas esteroideas y el grupo hemo de la hemoglobina, consecuencia de su actividad vital o provenientes del medio externo, como insecticidas, pesticidas, conservantes, medicamentos... • Contracción muscular.: el REL es muy abundante en el músculo estriado (donde se denomina retículo sarcoplásmco). Su función en las células musculares consiste en acumular Ca 2+ en su interior y liberarlo en respuesta a estímulos nerviosos, para permitir así la contracción muscular. La pérdida de toxicidad se consigue transformando estas sustancias insolubles en otras solubles, que puedan abandonar la célula y ser excretadas por la orina. Esta función la realizan fundamentalmente las células del hígado, riñón, intestino, pulmones y piel. Las cavidades del RE forman un compartimento intracitoplasmico, en el cual se aíslan moléculas e iones, y se acumulan en ocasiones en gran cantidad. Estas cavidades al comunicarse entre sí, constituyen una vía por donde circula su contenido. Una vez que las moléculas han penetrado en las cavidades del retículo, estas ya no pueden salir, pero no permanecen en él, sino que las moléculas emigran en las cavidades y después las abandonan. Unas son almacenadas en otros compartimentos como los lisosomas (hidrolasas ácidas), granos de vitelo (proteínas) y otras son expulsadas al medio extracelular, después de pasar por el Aparato de Golgi, como es el caso de proteínas y glicoproteínas segregados por nº tipos celulares. De este modo la red de cavidades del retículo forma un compartimento en el cual circulan moléculas que actuarán en otra parte: en el interior o exterior de la célula. La segregación que se hace al interior de las cavidades puede ser de productos de origen endógeno, ya presentes en la célula o elaboradas por ella o de origen exógeno que provienen del medio extracelular; en las cavidades estos productos sufren a menudo transformaciones de diversos tipos. Estas sustancias de origen diverso se acumulan a aveces en grandes cantidades en las cavidades que quedan dilatadas, la mayoría de ellas se parecen a inclusiones amorfas o cristalinas más o menos voluminosas. Actividad 1: ¿Por qué crees que el RER está muy desarrollado en las células glandulares del páncreas? 170 Actividad 2: ¿Cuál crees que puede ser la causa de que los glóbulos rojos no presenten RER? Actividad 3: ¿Cuáles son las funciones celulares del colesterol? 2. APARATO DE GOLGI Fue descubierto por C. Golgi en 1898, está presente en todas las células excepto en los glóbulos rojos, se encuentra más desarrollado cuanto mayor es la actividad celular, disminuyendo hasta llegar a desaparecer en las fases finales de la vida de la célula. ♦ ESTRUCTURA La unidad básica del orgánulo es el sáculo, que consiste en una vesícula o cisterna aplanada. Cuando 8 o 4 sáculos se apilan, forman un dictiosoma, estos no están pegados, ya que hay una delgada capa de hialoplasma entre ellos. Además, pueden observarse toda una serie de vesículas más o menos esféricas a ambos lados y entre los sáculos. El conjunto de todos los dictiosomas y vesículas presente en una célula constituye el aparato de Golgi. La pila de sacos tiene una de sus caras dispuesta en las proximidades del RE llamada cara externa o formación o cis y la opuesta es la cara interna o secreción o trans. Entre el RE y la cara cis, se encuentra una población de vesículas pequeñas, denominadas vesículas de transición; y en la periferia de los dictiosomas, en la cara trans hay una serie d vesículas grandes, denominadas vesículas de secreción. Los sáculos están perforados de tanto en tanto, y partiendo de la periferia de estos, hay tubos de membrana que relacionan entre sí los diferentes dictiosomas de una misma célula. Las membranas que limitan los sáculos de la cara externa y las vesículas de transición son de espesor semejante al RE y los sáculos de la cara interna y vesículas de secreción son de espesor semejante a la membrana plasmática. En cuanto a su composición química, la membrana tiene una composición en lípidos intermedia entre el RE y la membrana plasmática. Existen enzimas relacionadas con la transferencia de grupos glucídicos y sulfato, específicas de estas membranas. Las cavidades tienen un contenido semejante a las cavidades del RE pero además hay muchos polisacáridos que no hay en el RE. 171 ♦ FUNCIONES • Ensamblaje de los productos de secreción: después de la secreción, las cadenas polipeptídicas destinadas a ser exportadas transitan por el AG y son embaladas en las vesículas y gránulos antes de ser descargadas al medio extracelular. Los productos de la secreción que llenan las cavidades d los sáculos son embalados dentro las vesículas de secreción que se forman a partir de los sáculos de la cara interna, ya sea por gemación del borde de los sáculos o por fragmentación de éstos. Al fusionarse, las vesículas dan los gránulos de secreción que emigran hacia la periferia celular. El desarrollo cronológico sería: la síntesis y la segregación a las cavidades del RE- varios minutos; su transporte hacía el AG- 10- 30 minutos; la migración de los gránulos de secreción hasta descargar su contenido- 1- 4 horas. • Glicosilación: gracias al equipamiento tan importante de sus membranas en glicosiltransferasas, el AG representa el sitio celular en donde se efectúa el mayor nº de glicosilaciones, interviniendo en la síntesis de glicoproteínas y glicolípidos. La síntesis de la parte heteropolisacárida de estas moléculas empieza n el RE, lo que hace el AG es elongar y terminar las cadenas de polisacáridos. Las glicoproteínas destinadas a la exportación celular no son liberadas a las cavidades hasta después de su síntesis. Las destinadas a la renovación de la membrana permanecen integradas en ella y las holoprotínas destinadas a la exportación fuera de la célula pasan inmediatamente después de su síntesis a las cavidades. • Producción de membrana: por fusión de la membrana de los gránulos d secreción con la membrana plasmática por exocitosis, contribuyendo a la producción de material para la membrana. Las porciones de membrana que se fusionan tienen la misma asimetría, ya que las cadenas de glicolípidos y glicoproteínas están en la cara interna del gránulo, que luego será la externa de la membrana plasmática. Durante la migración de los gránulos, su membrana cambia d composición asemejándose a la membrana plasmática. Participa en la formación de la pared celular vegetal y del glicocaliz en animales. Participa en la formación de los lisosomas. • • Con respecto a las cavidades, la segregación que se hace en las atañe principalmente a las moléculas de origen endógeno que llegan a las cisternas del retículo por vesículas de transición y enseguida son secretadas al medio extracelular, como son: holo, glico y lipoproteínas. En casos raros son holopolisacáridos como es el caso de la celulosa, elaborada en las cavidades del AG que formará las paredes celulares de algunas algas. Actividad 4: ¿Por qué se considera al complejo de Golgi el “director principal” del transporte de proteínas en la célula? 172 Actividad 5: Busca en el libro de texto u otros libros el origen del acrosoma en los espermatozoides. Actividad 6: Identifica en el dibujo los orgánulos celulares que aparecen señalados con un número e indica la función biológica de cada uno de ellos. Explica así mismo, que camino recorre una proteína sintetizada en el RER hasta que llega al exterior de la célula. 3. RIBOSOMAS Solo son observables al microscopio electrónico, los describió por primera vez Palade en 1953. Son partículas globulares de 150-200 A de diámetro. Aparecen en todas las células procariotas y eucariotas, situados en el hialoplasma, el núcleo carece de ellos, también aparecen en cloroplastos y mitocondrias. Están formados por dos subunidades de distinto tamaño, una grande y otra pequeña y pueden unirse en un polisoma por un filamento de unos 15 A d diámetro. Los ribosomas de procariotas son más pequeños que los de los eucariotas y sus subunidades son asimétricas y de distinta forma. Aislados los ribosomas forman una población heterogénea de partículas globulares que se caracterizan por su coeficiente de sedimentación: la ultracentrifugación de moléculas en solución en un medio adecuado, somete a los constituyentes a una elevada fuerza centrífuga (250.000 g) y que produce su sedimentación. Esta sedimentación se realiza a una velocidad que está en función no solo d la velocidad y duración de la ultracentrifugación, sino también del peso molecular y de la forma de la molécula o las partículas y que es el coeficiente d sedimentación. Los valores de este coeficiente en general son muy bajos y se expresan en unidades Svedberg (S) 1 S= 1. 10-13 cm/ sg Tipo celular PROCARIOTAS EUCARIO -TAS Coeficiente de S Subunidad grande Subunidad pequeña proteínas 70 S CS- 50 S ARNr 23 S- 3000 n ARNr 5 S- 120 n CS- 30 S ARNr 16 S- 1600 n 40% 80S CS- 60 SARNr 28 S ARNr 5 S CS- 40 S ARNr- 18 S 50% Colocados en un medio con poco Mg los ribosomas se disocian en sus subunidades, de distinto tamaño y diferente coeficiente de sedimentación. Esta disociación es reversible añadiendo pequeñas cantidades de Mg. Los ribosomas de eucariotas se unen a la membrana del RER por su subunidad de 60 S. Los ribosomas son estructuras porosas que contienen un 70% de agua, están formados por ARN r y proteínas que no son las mismas en las 2 subunidades, pueden tener estructura secundaria dentro de 173 cada cadena. Las proteínas ribosómicas representan el 50% del peso seco, son todas distintas y cada ribosoma contiene únicamente un ejemplar de cada una. En el caso de los procariotas, representan el 40% del peso seco, son poco conocidas, pero son distintas de los procariotas. La estructura molecular compleja de un ribosoma puede realizarse por autoacoplamiento de los numerosos materiales que los componen. Este Autoensamblaje se hace por interacción entre lugares estereoespecíficos que permiten a las moléculas reconocerse y encajar como las piezas de un puzle, pero en tres dimensiones. Las interacciones ARN-proteína son interacciones hidrófobas y de puentes de hidrógeno y no enlaces iónicos. El acoplamiento de las moléculas se realiza por etapas: 1º ciertas proteínas al ARN; 2º después de la formación de este complejo, se unen otras proteínas, estas ya en un orden establecido. Unidos los Acs. Nucléicos y proteínas forman un edificio con propiedades nuevas, ya es capaz de cooperación con otras moléculas, de polimerizar de manera programada los aminoácidos en una secuencia definida. En esta síntesis se ponen en juego los lugares estereoespecíficos propios del ribosoma, que l permiten fijar de manera reversible diversos ARNs y proteínas que van y vienen a lo largo de la síntesis. Actividad 7: En las mitocondrias y los cloroplastos de las células eucariotas se han detectado ribosomas 70 s. ¿qué hipótesis evolutiva apoya este hallazgo? Explícala. 4. LISOSOMAS Descritas por De Duve en 1949, son vesículas membranosas redondeadas que contienen en su interior una elevada concentración de enzimas hidrolíticas, denominadas hidrolasas ácidas, pues requieren un ambiente de pH bajo para su óptimo funcionamiento, resulta evidente la necesidad de mantener tales enzimas separadas por una membrana del resto del citosol, aunque su efectividad bajaría bastante, pues el citosol mantiene un pH ligerísimamente básico Teniendo en cuenta el contenido de los lisosomas es fácil deducir que están involucrados en la degradación de sustancias, lo que se conoce como digestión celular, bien para aprovechamiento de la célula, bien para ser vertidos al medio extracelular. Existen varias categorías de lisosomas dependiendo de su origen: a) Lisosomas primarios: son lisosomas reciente formación (250 A diámetro), por lo que todavía no han intervenido en proceso digestivo alguno y su contenido consiste en enzimas exclusivamente. Sus funciones son: • Generar los lisosomas secundarios. • Verter su contenido, por exocitosis, al exterior de la célula para llevar a cabo una digestión extracelular. Muy importante en los procesos de remodelación ósea y cartilaginosa en vertebrados, así como en el proceso de descomposición de sustancias por parte de las células de los hongos. Un caso especial es el acrosoma del espermatozoide, el cual contiene una enzima, la hialuronidasa, cuya función es abrir paso al espermatozoide en su camino hacia el óvulo. 174 b) Lisosomas secundarios: son lisosomas en actividad, es decir, contienen enzimas y sustancias en vías d degradación. Se pueden clasificar, a su vez, en tres tipos distintos, según el material que está siendo digerido, y cada tipo confiere a la célula diversas funciones: • Vacuolas digestivas: se forman cuando el fagosoma o endosoma se une a un lisosoma primario. Una vez digerido el material, los productos atraviesan la membrana del lisosoma y pasan al citosol para su aprovechamiento. Si la digestión no es completa se forman los denominados cuerpos residuales. Además de llevar a cabo la digestión intracelular, que pude ocupar tanto productos de nueva absorción por el organismo como proteínas reabsorbidas, por el riñón, por ejemplo, desempeñan funciones tales como la defensa contra patógenos, destruyéndolos una vez englobados (células macrófagas). Un caso especial de lisosomas secundarios son los denominados gránulos de aleurona en las semillas de ciertos vegetales; n ellos se acumulan sustancias de reserva y enzimas hidrolíticas que permanecen inactivas hasta que la semilla se hidrata y comienza la germinación, rindiendo entonces sustancias que serán utilizadas por el embrión. • Cuerpos multivesiculares: cuando se fusionan, no uno, sino numerosos endosomas. • Vacuolas autofágicas o autolisosomas: se originan al unirse un lisosoma primario con un autofagosoma. Este último consiste en un fagosoma que engloba estructuras o sustancias de la propia célula que ya no son necesarios. Este proceso de autodestrucción controlado, permite un equilibrio entre síntesis y destrucción, recambiándose todos los orgánulos de la célula excepto el ADN. El envejecimiento celular, se relaciona con la acumulación en los lisosomas de sustancias no expulsadas. Actividad 8: ¿Mediante qué proceso o gracias a que sistema mantiene en su interior el lisosoma el pH ácido necesario para que sus enzimas puedan actuar? Actividad 9: ¿Crees que hay diferencia en el contenido de las vesículas procedentes del retículo, las vesículas de transición y las de secreción? ¿En qué consisten esas diferencias? Actividad 10: ¿Qué clase de enzimas encierran los lisosomas? ¿Qué tipos morfológicos de lisosomas podemos observar en una célula? 5. PEROXISOMAS Fueron descritos por De Duve como vesículas de membrana similares en forma a los lisosomas, pero que, a diferencia de éstos, contienen enzimas oxidativas en lugar de hidrolasas ácidas. Las enzimas más 175 comunes son la uratooxidasa y la catalasa y, en ocasiones, están presentes en tan altas concentraciones que los peroxisomas aparecen en las microfotografías electrónicas conteniendo un corpúsculo central cristaloide. Los peroxisomas están involucrados en el metabolismo del oxígeno y parece ser que jugaron un papel importante en la evolución desde los procariontes a los eucariontes, permitiendo la supervivencia de los estados intermedios en un ambiente peligrosamente rico en oxígeno, mientras las mitocondrias alcanzaban cierta eficacia en el desempeño de sus funciones. De hecho, una vez que fue así, es decir, una vez que las mitocondrias desarrollaron plenamente su capacidad de utilizar el oxígeno en reacciones que rendían ATP, los peroxisomas se hicieron menos relevantes, lo que no impide que aún hoy se encuentren en todas las células eucariotas, eso sí, desempeñando funciones menores relacionadas con el oxígeno y que no llevan a cabo las mitocondrias. Su nombre se debe precisamente a que las oxidasas utilizan oxígeno molecular para extraer el hidrógeno de ciertos sustratos orgánicos (1), rindiendo peróxido de hidrógeno, que será utilizado, a su vez, por la catalasa para oxidar diversas sustancias (2), tales como fenoles, ácido fórmico, formaldehído y alcohol, algunas de ellas tóxicas. 1. formación del peróxido RH2 + O2 2. reacción “peroxidativa”: H2O + R´H2 R + H2O R´+ 2 H2O 3. Con altas concentraciones de peróxido: 2H2O2 2 H2O + O2 El desempeño de estas reacciones como vía de detoxificación hace que los peroxisomas sean muy importantes en las células del hígado y riñón. Si por alguna razón el peróxido se acumula en la célula, la catalasa procede a reducir su concentración mediante una reacción que rinde agua y oxígeno molecular (3). Igualmente importante, si no más, es la involucración de los peroxisomas en el denominado βmetabolismo o β- oxidación d los ácidos grasos. Las cadenas de los ácidos grasos son recortadas secuencialmente n moléculas de dos átomos de carbono que serán convertidas en acetil CoA para su exportación al citosol, donde se utilizarán en diversas reacciones de biosíntesis. Si bien en células animales esta cadena de reacciones tiene lugar tanto en mitocondrias como en peroxisomas, en levaduras y plantas sólo ocurre en los peroxisomas. De hecho, en las plantas se conocen otras dos funciones importantes desempeñadas por los peroxisomas. Una de ellas tiene que ver con su colaboración con cloroplastos, facilitando el intercambio de sustancias durante la denominada fotorrespiración, una reacción que fija carbono en forma de glúcidos y que usa O2 y libra CO2. La otra se refiere a su papel en la germinación de ciertas semillas, en las que colabora en la conversión de los ácidos grasos almacenados en glúcidos necesarios para el crecimiento de la nueva planta. Los peroxisomas involucrados en este último proceso son también llamados glioxisomas porque la serie de reacciones que tiene lugar es conocida como ciclo glioxílico. 176 6. VACUOLAS Se trata de estructuras de membrana con forma más o menos redondeada y que contienen sustancias inertes, es decir, que no están involucradas en proceso metabólico alguno. Las vacuolas están mucho más desarrolladas en vegetales que en animales. Concretamente, son pequeñas en las células jóvenes (o menos diferenciadas) y aumentan mucho de tamaño con la edad, de forma que incluso el citoplasma y el núcleo quedan desplazados y comprimidos contra la membrana plasmática. La delgada membrana que las forma se denomina tonoplasto. Son generalmente incoloras (las vacuolas de los pétalos de las flores están llenas de diferentes pigmentos, que las dotan de color). La función de las vacuolas en vegetales consiste, esencialmente, n almacenar sustancias: • Agua • • • Sustancias nutritivas: almidón, lípidos, proteínas. Los gránulos de aleurona son, según los autores, considerados vacuolas. Sustancias d desecho resultantes del metabolismo celular y que pueden resultar tóxicos para la planta si se encuentran libres en el citoplasma. Entre ellos, algunos muy conocidos como la nicotina Pigmentos ya mencionados. En animales, como hemos comentado, son menos importantes y se relacionan con los lisosomas: vacuolas digestivas y vacuolas autofágicas, vistas anteriormente. Quizás, eso sí, merezca destacar la presencia de un tipo especial de vacuolas, las vacuolas pulsátiles, en ciliados, organismos unicelulares en los que cumplen una función reguladora de la cantidad d agua que contiene la célula, ya que viven en ambientes hipotónicos y el agua entra en la célula de manera pasiva, de tal manera que estas vacuolas bombean el exceso de agua al exterior funcionando como una bomba d achique de agua que lo hace de manera regular como un palpito. Actividad 11: Explica por qué es necesario para la célula expulsar el agua en un medio hipoosmótico. 177