citoplasma organelas
Anuncio

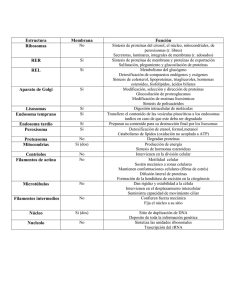
Citoplasma, organelas. Dr. Hernán G. Farina Universidad Nacional de Quilmes Laboratorio de Oncología Molecular El núcleo está rodeado por dos membranas, cada una de las cuales es una bicapa fosfolipídica que contiene muchos tipos diferentes de proteína. Las dos membranas nucleares se fusionan en los poros nucleares. Estos poros se encuentran formados por proteínas específicas y funcionan como canales que regulan el desplazamiento de sustancias entre el núcleo y el citosol. En el nucleolo presente en el núcleo, se sintetiza la mayor parte del ARN ribosómico y algunas proteínas ribosomales La subunidad ribosómica sintetizada pasa a través de un poro nuclear hasta el citosol. Existen proteínas denominadas láminas que forman una red bidimensional a lo largo de la superficie interna de la membrana interna que confiere forma y colabora en la distribución del ADN. El retículo endoplásmico (RE) es un compartimiento que forma una red de vesículas cerradas, limitadas por membrana e interconectadas. Su función principal es la síntesis de lípidos y proteínas de membrana. RE liso: Síntesis de Ac. Grasos y fosfolípidos. Las enzimas del RE liso de los hepatocitos modifican y detoxifican compuestos químicos hidrófobos (ej: pesticidas y carcinógenos), al convertirlos en productos conjugados más hidrosolubles que pueden excretarse del organismo. RE rugoso: En los ribosomas de esta organela se sintetizan determinadas proteínas de membrana y organelas, y la mayoría de las proteínas secretadas por la célula. El RE rugoso es especialmente abundante en las células especializadas para producir proteínas de secreción. La mayoría de las proteínas sintetizadas en el RE rugoso abandonan rápidamente la organela por vesículas de transporte, y son dirigidas hacia la cavidad luminal del Complejo de Golgi, formado por varios sacos aplanados de membrana. Transporte vesicular. Las vesículas de transporte emergen por gemación de un compartimiento y se fusionan con otro, al hacerlo, transportan material desde el lumen y la membrana del compartimiento dador, al lumen y la membrana del compartimiento de destino. Las vesículas de Golgi procesan y clasifican las proteínas de secreción y las proteínas de membrana Una gran parte de los glúcidos que fabrica el Golgi se unen como cadenas de oligosacáridos a las proteínas y lípidos que el ER le envía. Una vez adquiridas los oligosacáridos en el Golgi, son distribuídas mediante vesículas de transporte a otras partes de la célula. Aparato de Golgi: Procesamiento de proteínas Una vez modificadas, las proteínas de secreción son transportadas hacia el exterior por un segundo tipo de vesículas de transporte que brotan de la cara trans del golgi. Los lisosomas son organelas ácidas que contienen una batería de enzimas degradativas La planta energética Mitocondria de la célula: La Los intermediarios metabólicos parcialmente reducidos ingresan a la mitocondria como acetil-CoA y son completamente oxidados. Los estadios terminales de la degradación de la glucosa ocurren en la mitocondria. Son efectuados por enzimas localizadas en la matriz mitocondrial y las crestas. En la mitocondria se generan hasta 34 moléculas de ATP (de 36 totales) por molécula de glucosa. De forma similar, casi todo el ATP generado durante la oxidación de ácidos grasos se genera en la mitocondria. Mitocondria y apoptosis Las caspasas inician una cascada proteolítica de amplificación que finaliza en la fragmentación de los componentes celulares que desmantela la célula de forma rápida y limpia. Los restos celulares son rápidamente capturados y digeridos por otras células. Los peroxisomas degradan ácidos grasos y compuestos tóxicos Los ácidos grasos de cadena muy larga con más de unos 20 grupos CH2 son degradados exclusivamente en los peroxisomas; en las células de los mamíferos, los ácidos grasos de cadena mediana, de 10 a 20 grupos CH2, pueden degradarse tanto en los peroxisomas como en las mitocondrias. El peroxisoma es la organela más importante en la oxidación de ácidos grasos, con la consiguiente generación de importantes precursores para rutas biosintéticas. El citosol es la fracción rica en proteínas que queda luego de eliminar todas las organelas. Contiene numerosas enzimas solubles y tres tipos principales de filamentos proteicos: filamentos de actina, microtúbulos y filamentos intermedios. El citoesqueleto de las células eucariontes se compone de tres tipos de filamentos: -Filamentos intermedios: Proporcionan fuerza y resistencia al estrés mecánico. -Microtúbulos: determinan las posiciones de los orgánulos rodeados de membrana y dirigen el transporte intracelular. -Filamentos de actina: determinan la forma de la superficie celular y son necesarios para la locomoción. De la misma manera que necesitamos que nuestros ligamentos, huesos y músculos trabajen juntos, los tres sistemas de filamentos del citoesqueleto han de funcionar normalmente de forma colectiva, proporcionando a la célula su resistencia, su forma, y su capacidad de movimiento. Los microtúbulos actúan como autopistas moleculares facilitando el tráfico intracelular Estructura de una proteína motora (dineína) Drogas específicas de la actina Faloidina Se une y estabiliza los filamentos Citocalasina Encasqueta los extremos más de los filamentos Suinholida Fragmenta filamentos Latrunculina Se une a las subunidades e impide su polimerización Drogas específicas de los microtúbulos Taxol Se une y estabiliza los microtúbulos Colchicina, colcemida Se une a las subunidades e impide su polimerización Vimblastina, vincristina Se une a las subunidades e impide su polimerización Nocodazol Se une a las subunidades e impide su polimerización Células tumorales F3II Control Células F3II tratadas con taxol Células tumorales F3II tratadas con nocodazol