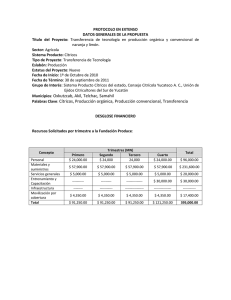
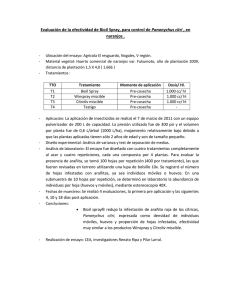
PD 6: Xanthomonas citri subsp. citri. NIMF 27, Anexo 6. Protocolos
Anuncio

E SP PD 6: Xanthomonas citri subsp. citri PROTOCOLOS DE DIAGNÓSTICO NORMAS INTERNACIONALES PARA MEDIDAS FITOSANITARIAS 27 NIMF 27 ANEXO 6 Producido por la Secretaría de la Convención Internacional de Protección Fitosanitaria (CIPF) Esta página se ha dejado en blanco intencionalmente Este protocolo de diagnóstico fue adoptado por el Comité de Normas, en nombre de la Comisión de Medidas Fitosanitarias en agosto de 2014. El anexo es una parte prescriptiva de la NIMF 27. NIMF 27 Protocolos de diagnóstico para las plagas reglamentadas PD 6: Xanthomonas citri subsp. citri Adoptado en 2014; publicado en 2016 ÍNDICE 1. Información sobre la plaga ...............................................................................................................2 2. Información taxonómica ...................................................................................................................2 3. Detección ..........................................................................................................................................3 3.1 Detección en plantas sintomáticas.....................................................................................3 3.1.1 Síntomas ............................................................................................................................3 3.1.2 Aislamiento .......................................................................................................................4 3.1.3 Detección serológica: inmunofluorescencia indirecta .......................................................4 3.1.4 Detección molecular..........................................................................................................5 3.1.4.1 Controles para las pruebas moleculares ............................................................................5 3.1.4.2 Extracción de ADN de tejido infectado de cítricos ...........................................................6 3.1.4.3 PCR convencional .............................................................................................................6 3.1.4.4 PCR en tiempo real ...........................................................................................................7 3.1.5 Interpretación de los resultados de la PCR convencional y en tiempo real .......................8 3.1.6 Detección mediante bioensayos ........................................................................................9 3.1.6.1 Prueba de inoculación en discos de hojas .........................................................................9 3.1.6.2 Enriquecimiento de hojas desprendidas ............................................................................9 3.2 Detección en plantas asintomáticas .................................................................................10 4. Identificación ..................................................................................................................................10 4.1 Métodos de PCR .............................................................................................................11 4.2 Detección serológica .......................................................................................................12 4.2.1 DAS-ELISA ....................................................................................................................12 4.2.2 ELISA indirecto ..............................................................................................................13 4.3 Pruebas de patogenicidad ................................................................................................13 4.4 Descripción y características bioquímicas ......................................................................13 4.5 Identificación molecular ..................................................................................................14 4.5.1 Análisis de secuencias multilocus ...................................................................................14 4.5.2 Identificación por rep-PCR .............................................................................................14 5. Registros .........................................................................................................................................15 6. Puntos de contacto para información adicional ..............................................................................15 7. Agradecimientos .............................................................................................................................16 8. Referencias .....................................................................................................................................16 9. Figuras ............................................................................................................................................20 Convención Internacional de Protección Fitosanitaria PD 6-1 PD 6 1. Protocolos de diagnóstico para plagas reglamentadas Información sobre la plaga Xanthomonas citri subsp. citri es el principal agente causal de la cancrosis bacteriana de los cítricos. Causa daños en muchas especies cultivadas de Rutaceae (EPPO, 1979) – principalmente Citrus spp., Fortunella spp. y Poncirus spp. – que se cultivan en las condiciones tropicales y subtropicales predominantes en muchos países de Asia, América del Sur, Oceanía y África, así como en Florida (Estados Unidos) (CABI, 2006; EPPO, 2006). Se han identificado cepas atípicas de X. citri subsp. citri, denominadas cepas A* y Aw (Sun et al., 2004; Vernière et al., 1998), con un rango de hospedantes restringido. La cepa A* afecta a Citrus aurantiifolia (lima mexicana) en condiciones naturales en Asia. La cepa Aw provoca cancrosis en Citrus aurantiifolia (lima mexicana) y Citrus macrophylla (Alemow) en Florida (Estados Unidos), bajo condiciones naturales (Cubero y Graham, 2002, 2004). Se ha constatado en forma experimental que ambas cepas causan lesiones atípicas en otras especies de cítricos (Escalon et al., 2013). La cancrosis bacteriana de los cítricos suele afectar a las plántulas y a árboles jóvenes y adultos de hospedantes susceptibles con crecimiento activo de vástagos y hojas desde finales del verano y durante el otoño en la mayoría de las áreas de cultivo de cítricos. Los cancros se forman en las hojas, los vástagos, las ramillas y los frutos de los hospedantes susceptibles. Las heridas, producidas por el viento, las espinas, los insectos y otros daños físicos y mecánicos, facilitan la infección de los tejidos maduros. Los ataques de Phyllocnistis citrella, el minador de las hojas de los cítricos, pueden aumentar la susceptibilidad de las hojas al cancro de los cítricos (Hall et al., 2010). X. citri subsp. citri puede sobrevivir en tejidos vegetales enfermos, como epifita en plantas hospedantes y no hospedantes, y como una saprofita en cubiertas vegetales de paja o en el suelo. No obstante, las lesiones invernantes, en particular las que presentan los vástagos angulosos, son la fuente más importante de inóculo para la estación siguiente. Los principales mecanismos de dispersión a corta distancia son la lluvia impulsada por el viento y las salpicaduras de agua en una misma planta o entre plantas: las bacterias se dispersan por el agua de lluvia que se desplaza sobre la superficie de las lesiones y que luego salpica los vástagos sanos (CABI, 2006). Se ha constatado que el movimiento de material de propagación infectado, incluyendo yemas, plántulas de portainjertos y árboles injertados, contribuye a la dispersión a larga distancia. No existe evidencia de que este patógeno sea transportado por la semilla (CABI, 2006). 2. Información taxonómica Nombre: Xanthomonas citri subsp. citri (Gabriel et al. 1989) Schaad et al. (2007) Sinónimos: Xanthomonas smithii subsp. citri Gabriel et al., 1989, Schaad et al., 2007 Xanthomonas axonopodis pv. citri (Hasse) Vauterin et al., 1995 Xanthomonas citri (ex Hasse, 1915) Gabriel et al., 1989 Xanthomonas campestris pv. aurantifolii Gabriel et al., 1989 Xanthomonas campestris pv. citri (Hasse) Dye, 1978 Xanthomonas citri f.sp. aurantifoliae Namekata y Oliveira, 1972 Pseudomonas citri Hasse, 1915 Posición taxonómica: Bacteria, Proteobacteria, Xanthomonadaceae Gammaproteobacteria, Xanthomonadales, Nombres comunes: cancro de los cítricos, cancrosis bacteriana de los cítricos, cáncer asiático Nota: Recientemente, X. axonopodis pv. citri (cepas del grupo A de X. campestris pv. citri) se ha reclasificado como X. citri subsp. citri. Se ha recuperado la nomenclatura de Gabriel et al. (1989) y el nombre aceptado del patógeno de la cancrosis bacteriana de los cítricos es actualmente X. citri subsp. citri (Bull et al., 2010; Schaad et al., 2006). Las cepas de los otros grupos de X. campestris PD 6-2 Convención Internacional de Protección Fitosanitaria Protocolos de diagnóstico para plagas reglamentadas PD 6 pv. citri se han reclasificado como Xanthomonas fuscans subsp. aurantifolii (grupos B, C y D) y Xanthomonas alfalfae subsp. citrumelonis (grupo E) (Schaad et al., 2006). 3. Detección 3.1 Detección en plantas sintomáticas El cancro de los cítricos se puede diagnosticar observando las características morfológicas de las colonias en medios nutritivos y mediante pruebas serológicas (por inmunofluorescencia (IF)), pruebas moleculares (mediante la reacción en cadena de la polimerasa (PCR)) y bioensayos en discos de hojas u hojas desprendidas. Todas las pruebas deben incluir controles negativos y positivos (los controles de referencia se describen en la sección 4). 3.1.1 Síntomas Los síntomas característicos de la enfermedad son costras o lesiones crateriformes en la cáscara de los frutos y en las hojas, los tallos y los vástagos. Los síntomas del cancro de los cítricos pueden darse en plántulas, en todas las estaciones y en árboles jóvenes, desde finales del verano y durante el otoño, cuando se produce un crecimiento abundante de vástagos angulosos (CABI, 2006) (figuras 1-4). La enfermedad se hace esporádica a medida que los árboles alcanzan el desarrollo completo de frutos, debido a que se producen menos vástagos angulosos y a que el tejido foliar más viejo y los frutos maduros son más resistentes a la infección por cancro de los cítricos en condiciones naturales. La severidad de la enfermedad también depende de la susceptibilidad de las especies vegetales hospedantes y cultivares (Goto, 1992). Síntomas en los frutos. Se desarrollan lesiones crateriformes en la superficie del fruto, ya sea dispersas en puntos aislados o formando patrones irregulares por la unión de varias lesiones. En los frutos jóvenes infectados se podrá observar la exudación de sustancias resinosas. Las lesiones nunca se extienden a través de la cáscara. Síntomas en las ramas. En condiciones secas, la mancha del cancro es corchosa o esponjosa, sobresale y presenta una superficie resquebrajada. En condiciones húmedas, la lesión aumenta rápidamente de tamaño, pero su superficie no se resquebraja y presenta un borde aceitoso. En los cultivares menos susceptibles, podrá formarse una capa de callo entre los tejidos enfermos y sanos. La cicatriz de un cancro podrá identificarse raspando la superficie rugosa con un cuchillo para retirar la capa corchosa exterior y revelar la presencia de lesiones de color marrón claro a marrón oscuro en los tejidos sanos de la corteza verde. El área afectada por el cambio de color puede tener formas diversas y su tamaño puede oscilar entre los 5 y los 10 mm, dependiendo de la susceptibilidad de la planta hospedante. Síntomas en las hojas. El primer síntoma es la aparición de manchas de color amarillo brillante en el envés de las hojas seguida de la aparición de lesiones protuberantes de color amarronado en ambas caras de las hojas, que se tornan rugosas, agrietadas y corchosas. El cancro podrá estar rodeado por un halo amarillo o clorótico humedecido. Los síntomas del cancro de los cítricos en las ramas, las hojas y los frutos se podrán confundir con síntomas similares en forma de costras o de manchas en las hojas provocados por otras bacterias u hongos que infectan a los cítricos o por desórdenes fisiológicos. Otras bacterias que pueden causar síntomas similares a los del cancro de los cítricos son X. alfalfae subsp. citrumelonis y X. fuscans subsp. aurantifolii. Ambas bacterias tienen un rango de hospedantes limitado, provocan síntomas menos agresivos y rara vez producen lesiones en los frutos (Schaad et al., 2005, 2006). Se ha informado que la sarna de los cítricos provocada por el hongo Elsinoë fawcettii produce síntomas similares a los del cancro de los cítricos, en particular en variedadeshospedantes que muestran resistencia a la sarna de los cítricos (Taylor et al., 2002), aunque en general sus lesiones son más secas y más irregulares que las del cancro de los cítricos y a veces carecen del halo amarillo característico. La sarna de los cítricos se puede diferenciar del cancro de los cítricos por la ausencia de exudado bacteriano. Convención Internacional de Protección Fitosanitaria PD 6-3 PD 6 Protocolos de diagnóstico para plagas reglamentadas 3.1.2 Aislamiento Para tener éxito en el aislamiento de X. citri subsp. citri a partir de material vegetal con síntomas es fundamental utilizar extractos de muestras recién preparados. El material vegetal debería analizarse lo antes posible después de su recolección; podrá almacenarse a una temperatura de entre 4 y 8 ºC hasta su procesamiento. Cuando los síntomas están muy avanzados o las condiciones ambientales no son favorables, el número de células cultivables de X. citri subsp. citri puede ser muy bajo y en su aislamiento pueden obtenerse placas saturadas con bacterias competidoras saprófitas o antagonistas. Debería ponerse especial atención en evitar confundir las colonias de X. citri subsp. citri con colonias de Pantoea agglomerans, que comúnmente también se suele aislar a partir de las lesiones de cancro y produce colonias morfológicamente similares en medios de cultivo bacteriológico estándar. Por lo general, P. agglomerans crece más rápido y las colonias son de un amarillo más intenso que el color amarillo o limón pálido de las colonias de X. citri subsp. citri. El aislamiento del organismo causal se puede efectuar mediante la siembra en estrías de extractos de lesiones sobre placas con un medio de cultivo adecuado, en el que las colonias de X. citri subsp. citri presentan un aspecto característico. Todavía no existen medios de cultivo selectivos exclusivamente para X. citri subsp. citri. Las lesiones se maceran en 0,5–1,0 ml de solución salina (agua destilada estéril con NaCl al 0,85 %, pH 7,0); cuando se requiera, se podrán desinfectar previamente con NaClO al 1 % durante 1 min, enjuagarse tres veces con agua destilada estéril y pulverizarse. Se siembra sobre un medio nutritivo una alíquota del extracto. Los medios de aislamiento general adecuados son el agar nutritivo complementado con un 0,1 % de glucosa (NGA), el agar levadura peptona glucosa o YPGA (extracto de levadura, 5 g; peptona Bacto™, 5 g; glucosa, 10 g; agar, 20 g; agua destilada, 1 litro; pH 7,0) y el medio Wakimoto (caldo de patata, 250 ml; sacarosa, 15 g; peptona, 5 g; Na2HPO4·12H2O, 0,8 g; Ca(NO3)2·7 H2O, 0,5 g; agar Bacto™, 20 g; agua destilada, 1 litro; pH 7,2). En caso necesario, se puede añadir cicloheximida esterilizada por filtración (100 mg/l) como fungicida después del autoclavado del medio de cultivo. Las colonias presentan, en los tres medios, morfología circular y convexa, con bordes lisos, y tienen aspecto mucoide y color amarillo crema. El crecimiento se evalúa tras incubar de tres a cinco días a 25–28°ºC. En las muestras de frutos comercializados, las bacterias pueden estar sometidas a factores adversos que podrán dificultar su cultivo, por lo que podrán requerirse períodos de incubación más prolongados, o bien las bacterias se pueden recuperar de las muestras mediante bioensayos, como se indica en la sección 3.1.6.2. La incorporación al medio de kasugamicina y cefalexina (medio semiselectivo KC o KCB) inhibe varias bacterias saprófitas y facilita el aislamiento del patógeno (Graham et al., 1989; Pruvost et al., 2005). En este protocolo de diagnóstico, los métodos (incluidas las menciones de nombres comerciales) se describen según se publicaron, ya que en los métodos publicados se define la sensibilidad, la especificidad y la reproducibilidad logrados inicialmente. La mención de nombres de sustancias químicas (p.ej., marcas comerciales) no implica su aprobación ni la exclusión de otras que también podrán ser adecuadas. Los procedimientos de laboratorio presentados en los protocolos podrán ajustarse a las prácticas establecidas en cada laboratorio, siempre que se validen adecuadamente. 3.1.3 Detección serológica: inmunofluorescencia indirecta Para la detección serológica (IF y ensayo de inmunoabsorción enzimática (ELISA)), es fundamental contar con controles adecuados para garantizar que los resultados de las pruebas sean confiables. Todas las pruebas deberían incluir controles negativos y positivos. Los controles positivos pueden consistir en una cepa de X. citri subsp. citri de referencia que se vuelve a suspender en un extracto de plantas hospedantes sanas (para la detección en material vegetal) o en una solución salina con tampón fosfato (PBS) (para la identificación de cultivos bacterianos). Los controles negativos deberían consistir en un extracto de plantas hospedantes sanas (para su detección en material vegetal) o en una suspensión de especies bacterianas distintas de la especie objetivo (para la identificación de cultivos bacterianos). PD 6-4 Convención Internacional de Protección Fitosanitaria Protocolos de diagnóstico para plagas reglamentadas PD 6 Para la detección serológica de células bacterianas, se extrae del cultivo fresco de la placa el contenido de un asa y se vuelve a suspender en 1 ml de PBS (NaCl, 8 g; KCl, 0,2 g; Na2HPO4·12H2O, 2,9 g; KH2PO4, 0,2 g; agua destilada, hasta 1 litro; pH 7,2) para obtener aproximadamente 108 unidades formadoras de colonias (ufc)/ml (EPPO, 2009). Para la detección serológica en tejido vegetal, deberían elegirse muestras con síntomas: vástagos, ramitas, hojas y frutos que presenten lesiones necróticas, o bien tejido de cancros en ramitas, ramas, el tronco o el pie. Las muestras se deberían procesar siguiendo el procedimiento general recomendado para la prueba serológica específica que vaya a utilizarse. Por lo general, el tejido vegetal se tritura en una solución tampón de maceración antioxidante recién elaborada (polivinilpirrolidona (PVP)-10, 20 g; manitol, 10 g; ácido ascórbico, 1,76 g; glutatión reducido, 3 g; PBS, 10 mM, 1 litro; pH 7,2) o en PBS (NaCl, 8 g; KCl, 0,2 g; Na2HPO4·12H2O, 2,9 g; KH2PO4, 0,2 g; agua destilada, hasta 1 litro; pH 7,2) antes de utilizarlo en las pruebas serológicas. Ambas soluciones se esterilizan por filtración con una membrana estéril de 0,22 µm. Se pipetean alícuotas de 25 µl de cada una de las preparaciones bacterianas o muestras vegetales que se someterán a la prueba y se depositan sobre un portaobjetos de microscopio con varias cavidades y recubierto de plástico. Tras dejarlas secar al aire, se fijan suavemente con calor sobre una llama. Se preparan portaobjetos distintos para cada bacteria o muestra de la prueba y también para los controles positivos y negativos, como los que se utilizan para el ELISA. Se diluye antisuero o anticuerpos monoclonales disponibles en el mercado en PBS (pH 7,2) y se añaden 25 µl de diluciones adecuadas a las cavidades de todos los portaobjetos. Los controles negativos pueden ser suero normal (no inmunizado) en una dilución y en PBS. Los portaobjetos se incuban durante 30 min en una cámara húmeda a temperatura ambiente. Se sacuden las gotitas fuera de los portaobjetos, se enjuagan con PBS y luego se lavan en PBS tres veces, durante cinco minutos cada vez. Los portaobjetos se secan cuidadosamente con material absorbente antes de depositar con pipeta en cada cavidad 25 µl del conjugado de isotiocianato de fluoresceína (FITC) y la gammaglobulina antiespecie apropiada en la dilución adecuada. Los portaobjetos se incuban durante 30 minutos a oscuras y a temperatura ambiente, se enjuagan, se lavan y se secan con material absorbente. Por último, se añaden 10 µl de tampón fosfato glicerol 0,1 mM (pH 7,6) con un reductor del apagamiento (antifading agent) a cada una de las cavidades, y a continuación se cubren con un cubreobjetos. Los portaobjetos se examinan con aceite de inmersión con un microscopio de fluorescencia a 600× o 1 000× aumentos. Bajo la luz ultravioleta del microscopio, el FITC presenta fluorescencia verde brillante. Si el control positivo, que contiene una bacteria conocida, muestra células bacterianas baciliformes fluorescentes y los controles negativos, que contienen suero normal y PBS, no muestran fluorescencia, se examinan las cavidades con las muestras en busca de células bacterianas fluorescentes con el tamaño y la forma de X. citri subsp. citri. Este método permite detectar aproximadamente 103 ufc/ml. 3.1.4 Detección molecular 3.1.4.1 Controles para las pruebas moleculares Para considerar confiable el resultado de la prueba, es fundamental emplear controles adecuados, que dependerán del tipo de prueba utilizada y del grado de certidumbre necesario. Para la PCR, deberían utilizarse, como mínimo, un control positivo de ácido nucleico, un control interno y un control negativo de amplificación (control sin molde). A continuación se describen estos controles y otros que deberían considerarse para todas las series de extracciones de ácidos nucleicos de las muestras de su prueba. Control positivo de ácido nucleico. Para controlar la eficiencia de la amplificación mediante PCR, se podrá utilizar como control ácido nucleico previamente preparado (almacenado), el ADN genómico completo o un control sintético (p. ej., un producto de PCR clonado). Controles internos. Para la PCR convencional y en tiempo real, debería incorporarse como control al protocolo de PCR un gen constitutivo vegetal (HKG), por ejemplo el COX (Weller et al., 2000), Convención Internacional de Protección Fitosanitaria PD 6-5 PD 6 Protocolos de diagnóstico para plagas reglamentadas ADN ribosomal (ADNr) 16S (Weisberg et al., 1991) o GADPH (Mafra et al., 2012), a fin de descartar la posibilidad de falsos negativos debido a deficiencias en la extracción del ácido nucleico o a su degradación, o a la presencia de inhibidores de la PCR. Control negativo de amplificación (control sin molde). Para la PCR convencional y en tiempo real, el agua de calidad apta para PCR que se utilizó para preparar la mezcla de reacción se añade en la fase de amplificación a fin de descartar falsos positivos por contaminación durante la preparación de la mezcla de reacción. Control positivo de extracción. Este control se utiliza para asegurar que el ácido nucleico del objetivo sea de calidad suficiente para la amplificación mediante PCR. El ácido nucleico se extrae de tejido infectado del hospedante o de tejido vegetal sano al que se ha añadido una concentración del objetivo igual a la considerada como límite de detección del protocolo. El control positivo debería ser aproximadamente una décima parte de la cantidad de tejido foliar utilizada por planta para la extracción de ADN. Para la PCR, deben adoptarse precauciones para evitar la contaminación cruzada por aerosoles procedentes del control positivo o de las muestras positivas. De ser necesario, el control positivo empleado en el laboratorio debería secuenciarse a fin de que esta secuencia se pueda comparar fácilmente con las secuencias obtenidas de los amplicones de la PCR del tamaño correcto. Otra opción es elaborar controles positivos sintéticos que contengan una secuencia conocida que, de nuevo, se puede comparar con los amplicones de la PCR del tamaño correcto. Control negativo de extracción. Este control se utiliza para controlar la contaminación durante la extracción del ácido nucleico y la reacción cruzada con el tejido hospedante. El control comprende ácido nucleico extraído de tejido no infectado del hospedante y posteriormente amplificado. Cuando se analicen grandes cantidades de muestras positivas se recomienda utilizar controles múltiples. 3.1.4.2 Extracción de ADN de tejido infectado de cítricos La extracción de ADN de tejido infectado de cítricos fue realizada por primera vez por Hartung et al. (1993) con un protocolo de extracción con bromuro de hexadeciltrimetilamonio (CTAB), pero hay métodos comerciales y un protocolo de extracción con isopropanol (para el que no se requiere fenol) que se han evaluado ampliamente (Llop et al., 1999). El ADN también se ha extraído satisfactoriamente de tejidos de cítricos mediante kits comerciales de extracción de ADN (p. ej., el equipo de purificación de ADN genómico Promega Wizard) (Coletta-Filho et al., 2006). En el protocolo de isopropanol, las lesiones o el material vegetal que se sospecha que puede estar infectado se cortan en pequeños trozos, se cubren con PBS y se introducen en un agitador giratorio durante 20 min a temperatura ambiente. El sobrenadante se filtra (para eliminar el material vegetal) y se centrifuga a 10 000 g durante 20 min. El sedimento se vuelve a suspender en 1 ml de PBS: 500 µl se guardan para un análisis ulterior o para su aislamiento directo en placas de agar, y 500 µl se centrifugan a 10 000 g durante 10 min. El sedimento se vuelve a suspender en 500 µl de tampón de extracción (200 mM de tris-HCl, pH 7,5; 250 mM de NaCl; 25 mM de etilendiaminotetraacético (EDTA); 0,5 % de dodecilsulfato sódico (SDS); 2 % de PVP), mediante un agitador de vórtice y se deja 1 h a temperatura ambiente con agitación continua. A continuación, la suspensión se centrifuga a 5 000 g durante 5 min y después se transfieren 450 µl del sobrenadante a un nuevo tubo y se mezclan con 450 µl de isopropanol. La suspensión se mezcla con suavidad y se deja reposar 1 h a temperatura ambiente. La precipitación se puede mejorar si se utiliza el coprecipitante Pellet Paint (Cubero et al., 2001). La suspensión se centrifuga a 13 000 g durante 10 min, se desecha el sobrenadante y se seca el sedimento. El sedimento se vuelve a suspender en 100 µl de agua. Se utiliza una muestra de 5 µl para un volumen de PCR de 50 µl. 3.1.4.3 PCR convencional Existen varios pares de cebadores para el diagnóstico de X. citri subsp. citri. Los cebadores 2 y 3 de Hartung et al. (1993) amplifican un fragmento de ADN polimórfico en la longitud de los fragmentos de restricción BamHI que es específico para X. citri subsp. citri; son los de uso más común en análisis PD 6-6 Convención Internacional de Protección Fitosanitaria Protocolos de diagnóstico para plagas reglamentadas PD 6 de material vegetal debido a su buena especificidad y sensibilidad (aproximadamente 102 ufc/ml). Los cebadores J-pth1 y J-pth2 amplifican un fragmento de 197 pares de bases (pb) de la señal de localización nuclear del gen de virulencia pthA en las cepas de Xanthomonas que producen los síntomas del cancro de los cítricos. Entre estas cepas se encuentran X. citri subsp. citri, X. fuscans subsp. aurantifolii y las cepas atípicas A* y Aw de X. citri subsp. citri, detectadas en Florida (Cubero y Graham, 2002). Se trata de cebadores universales, pero presentan menor sensibilidad (104 ufc/ml en material vegetal) que los de Hartung et al. (1993). No obstante, los cebadores de Hartung no detectan la cepa Aw ni ninguna de las cepas A* de X. citri subsp. citri, ni tampoco X. fuscans subsp. aurantifolii. En las situaciones en que se sospecha la presencia de cepas atípicas A * y Aw de X. citri subsp. citri -por ejemplo, cuando se observan síntomas del cancro de los cítricos en los hospedantes C. aurantiifolia (lima mexicana) y C. macrophylla (Alemow)- deben utilizarse ambos pares de cebadores. Protocolo de PCR de Hartung et al. (1993) Los cebadores son: 2 (inverso): 5′-CAC GGG TGC AAA AAA TCT-3′ 3 (directo): 5′-TGG TGT CGT CGC TTG TAT-3′. La mezcla de la PCR se prepara en un tubo estéril y está compuesta por tampón de PCR (50 mM de tris-HCl, pH 9; 20 mM de NaCl; 1 % de Triton X-100; 0,1 % de gelatina; 3 mM de MgCl2), 1 µM de cada cebador 2 y 3, 0,2 mM de cada desoxinucleótido trifosfato (dNTP) y 1,25 U de polimerasa Taq de ADN. Se añade un volumen de 5 µl de muestra del ADN extraído a 45 µl de la mezcla de la PCR para obtener un total de 50 µl por reacción. Las condiciones de la reacción consisten en una fase inicial de desnaturalización a 95 ºC durante 2 min, seguida de 35 ciclos de 95 ºC durante 60 s, 58 ºC durante 70 s y 72 ºC durante 75 s, y una fase de extensión final de 72 ºC durante 10 min. El tamaño de los amplicones es de 222 pb. Protocolo de PCR de Cubero y Graham (2002) Los cebadores son: J-pth1 (directo): 5′-CTT CAA CTC AAA CGCC GGA C-3′ J-pth2 (inverso): 5′-CAT CGC GCT GTT CGG GAG-3′. La mezcla de la PCR se prepara en un tubo estéril y está compuesta por tampón de Taq 1×, 3 mM de MgCl2), 1 µM de cada cebador J-pth1 y J-pth2, 0,2 mM de cada dNTP y 1 U de polimerasa Taq de ADN. Se añade un volumen de 2,5 µl de muestra del ADN extraído a 22,5 µl de la mezcla de la PCR para obtener un total de 25 µl por reacción. Las condiciones de la reacción consisten en una fase inicial de desnaturalización a 94 ºC durante 5 min seguida de 40 ciclos de 93 ºC durante 30 s, 58 ºC durante 30 s y 72 ºC durante 45 s, y una fase de extensión final de 72 ºC durante 10 min. El tamaño del amplicón es de 198 pb. También se han desarrollado protocolos basados en la PCR anidada, la inmunocaptura y la detección colorimétrica de productos de la PCR anidada para la detección directa y sensible de X. citri subsp. citri en plantas (Hartung et al., 1993). Se ha dado a conocer un examen comparativo de la sensibilidad de los distintos protocolos y cebadores en cultivos puros y extractos de frutos (Golmohammadi et al., 2007). 3.1.4.4 PCR en tiempo real Después de obtener el ADN del material vegetal mediante el protocolo anteriormente descrito por Llop et al. (1999), el sedimento se vuelve a suspender en 100 μl de agua ultrapura estéril y se almacena a -20 ºC hasta su utilización. Se diseñó un conjunto de cebadores, J-pth3 (5'-ACC GTC CCC TAC TTC AAC TCA A-3') y J-pth4 (5'-CGC ACC TCG AAC GAT TGC-3'), y la sonda TaqMan correspondiente (J-Taqpth2) (5'-ATG CGC CCA GCC CAA CGC-3') marcadas en el extremo 5′ con 6-carboxifluoresceína (FAM) y en el Convención Internacional de Protección Fitosanitaria PD 6-7 PD 6 Protocolos de diagnóstico para plagas reglamentadas extremo 3′ con tetrametilrodamina, a partir de secuencias del gen pth, un importante gen de virulencia utilizado en otros estudios para detectar específicamente cepas de X. citri subsp. citri (Cubero y Graham, 2005). Entre estas cepas se encuentran X. citri subsp. citri, X. fuscans subsp. aurantifolii y las cepas atípicas A* y Aw de X. citri subsp. citri detectadas en Florida. La PCR en tiempo real se efectúa añadiendo 2 µl de ADN molde a una mezcla de reacción que contiene 12,5 µl de QuantiMix Easy Kit, que contiene QuantiMix Easy Master Mix y MgCl 2 (50 mM), 1 µl de 10 µM de cebador directo (J-RTpth3), 1 µl de 10 µM de cebador inverso (J-RTpth4) y 0,5 µl de 10 µM de sonda TaqMan (J-Taqpth2), y completando con agua destilada estéril hasta un volumen final de reacción de 25 µl. El protocolo de PCR en tiempo real se ha desarrollado usando un sistema de detección de secuencia ABI PRISM 7000. Con otro equipo se han obtenido resultados similares (María López, com. pers., 2013). Las condiciones de amplificación para los cebadores y las sondas consisten en una fase inicial de activación de 15 min a 95 ºC seguida de 40 ciclos de 15 s a 95 ºC y 1 min a 60 ºC. Plant Print Diagnostics (http://www.plantprint.net) dispone de un equipo completo de PCR en tiempo real basado en este protocolo y que incluye la mezcla maestra y la enzima. Con la PCR en tiempo real se consigue una especificidad similar a la de los cebadores del gen pth utilizados en el método de PCR convencional (Cubero y Graham, 2002, 2005); asimismo, permite la detección fidedigna de aproximadamente 10 ufc de X. citri subsp. citri a partir de lesiones de hojas enfermas y de una dilución de células cultivadas (Mavrodieva et al., 2004). Este método se ha comparado recientemente con la PCR convencional y la PCR anidada (Golmohammadi et al., 2007) y se determinó una sensibilidad de detección de X. citri subsp. citri en lesiones de los frutos de 10 ufc/ml. 3.1.5 Interpretación de los resultados de la PCR convencional y en tiempo real PCR convencional La PCR específica del patógeno solo se considerará válida si se cumplen los siguientes criterios: el control positivo produce un amplicón del tamaño correcto para la bacteria, no se producen amplicones del tamaño correcto para la bacteria en el control negativo de extracción ni en el control negativo de amplificación. Si también se usan cebadores ADNr 16S de control interno, entonces el control negativo (tejido vegetal sano), si se utiliza, el control positivo y todas las muestras de la prueba producirán una banda de aproximadamente 1,6 kilobases (kb) (el tamaño de los amplicones dependerá de los cebadores ADNr 16S utilizados (Weisberg et al., 1991)). Téngase en cuenta que si se utilizan controles positivos sintéticos o plásmidos no se producirá una banda de 1,6 kb. Si no se logra la amplificación de las muestras con los cebadores de control interno, la causa puede ser, por ejemplo, una deficiente extracción de ADN, que no se incluyó el ácido nucleico en la mezcla de la reacción, que hay presencia de compuestos inhibidores de la PCR en el extracto de ADN o que el ADN se ha degradado. Una muestra se considerará positiva si produce un amplicón del tamaño correcto. PCR en tiempo real La PCR en tiempo real solo se considerará válida si se cumplen los siguientes criterios: el control positivo produce una curva de amplificación con los cebadores específicos del patógeno, no se observa ninguna curva de amplificación (esto es, el umbral de ciclo (Ct) es 40) con el control negativo de extracción ni con el control negativo de amplificación. Si también se usan cebadores COX de control interno, entonces el control negativo, si se utiliza, el control positivo y todas las muestras de la prueba deben producir una curva de amplificación. Si las muestras no producen una curva de amplificación con los cebadores de control interno, la causa puede ser, por ejemplo, una deficiente extracción de ADN, la no inclusión del ADN en la mezcla de la PD 6-8 Convención Internacional de Protección Fitosanitaria Protocolos de diagnóstico para plagas reglamentadas PD 6 reacción, la presencia de compuestos inhibidores de la PCR en el extracto de ADN o la degradación del ADN. Una muestra se considerará positiva si produce una curva de amplificación típica. Debe verificarse el umbral de ciclo en cada laboratorio cuando la prueba se realice por primera vez. 3.1.6 Detección mediante bioensayos 3.1.6.1 Prueba de inoculación en discos de hojas Esta prueba consiste en la inoculación de tejido foliar de cítricos susceptibles a X. citri subsp. citri con extractos de muestras de plantas enfermas y su incubación en condiciones adecuadas para la multiplicación bacteriana y el desarrollo de pústulas incipientes de la enfermedad. El procedimiento de este bioensayo comienza con la esterilización de placas de ELISA durante 15 minutos en un horno de microondas y la adición a los hoyos de la placa de 200 µl de agar al 1,5 % en agua estéril en una cámara de flujo laminar a temperatura ambiente. Se desinfecta la superficie de las hojas jóvenes de Citrus paradisi var. Duncan (pomelo) o de otros hospedantes susceptibles, por ejemplo Citrus aurantifolia (lima mexicana) o Poncirus trifoliata (naranjo trifoliado), con NAClO al 1 % durante 1 min. Las hojas deberían estar completamente desarrolladas pero no maduras ni duras. Las hojas se enjuagan tres veces con agua destilada estéril y a continuación se seca su superficie en una cámara de flujo laminar a temperatura ambiente. Los discos de hojas, extraídos mediante un perforador (desinfectado con etanol al 95 %), se colocan en el agar de agua con la superficie adaxial (el haz) hacia abajo en todos los hoyos. Se añaden 50 μl de macerado de lesiones de cancro de los cítricos (cuatro hoyos por cada muestra vegetal). Se utiliza una suspensión de 105 ufc/ml de X. citri subsp. citri como control positivo y una solución salina estéril como control negativo (cuatro repeticiones de cada una). Se tapan herméticamente las placas (p. ej., con Parafilm) para que alcancen una humedad relativa cercana al 100 %, y se incuban durante 12 días a 28 ºC con luz constante; se revisa su evolución con regularidad. A partir del tercer día se estudia la formación de pústulas incipientes de aspecto blanquecino en los discos de hojas por medio de un microscopio estereoscópico y aplicando las técnicas de aislamiento descritas en la sección 3.1.2 para X. citri subsp. citri. Los discos que no presenten síntomas se pueden analizar ulteriormente mediante su aislamiento en medios semiselectivos para detectar la presencia de bacterias vivas (Verdier et al., 2008). Después de 12 días, si hay presencia de X. citri subsp. citri, las células bacterianas se habrán multiplicado en el tejido vegetal y se podrán aislar en mayor número en otros medios. Este bioensayo es un método de diagnóstico sumamente específico y sensible (10 2 ufc/ml) (Verdier et al., 2008). 3.1.6.2 Enriquecimiento de hojas desprendidas X. citri subsp. citri también se puede enriquecer de forma selectiva en hojas desprendidas con heridas de C. paradisi var. Duncan (pomelo) o de otros hospedantes muy susceptibles, como C. aurantifolia (lima mexicana) o P. trifoliata (naranjo trifoliado). Las hojas terminales jóvenes de plantas cultivadas en invernaderos se lavan durante 10 minutos en agua corriente del grifo, se desinfecta su superficie con NAClO al 1 % durante 1 min y se enjuagan de forma abundante y asépticamente con agua destilada estéril. Se practican heridas asépticas en la superficie inferior de las hojas mediante punción con una aguja o haciendo pequeños cortes con un bisturí, y se colocan las hojas enteras, con su superficie inferior hacia arriba, en agua estéril con agar al 1 % en los hoyos de las placas de ELISA. Se añaden a las heridas gotitas de 10–20 µl de macerado de lesiones de cancro de los cítricos. Se utilizan controles positivos y negativos al igual que en los bioensayos de discos de hojas. Después de 4–12 días a 25 ºC en una incubadora iluminada, se evalúa el desarrollo de las pústulas. X. citri subsp. citri se puede aislar de las pústulas o del tejido foliar con heridas que no presenta síntomas, como se describió anteriormente (EPPO, 1998). Convención Internacional de Protección Fitosanitaria PD 6-9 PD 6 3.2 Protocolos de diagnóstico para plagas reglamentadas Detección en plantas asintomáticas La detección de X. citri subsp. citri en plantas asintomáticas puede realizarse mediante aislamiento y enriquecimiento en medios semiselectivos (véase a continuación), técnicas serológicas (inmunofluorescencia (sección 3.1.3)) y pruebas moleculares (sección 3.1.4). El aislamiento de X. citri subsp. citri a partir de plantas asintomáticas en medios semiselectivos puede realizarse lavando las muestras de hojas o frutos en tampón de peptona, concentrando el sobrenadante y, por último, sembrando en el medio (Verdier et al., 2008). Diez hojas o un fruto constituyen una muestra. Las muestras se agitan durante 20 min a temperatura ambiente en 50 ml de tampón de peptona (NaCl, 8,5 g; peptona, 1 g; Tween 20, 250 µl; agua destilada, 1 l; pH 7,2). Para muestras compuestas, se pueden utilizar 200 ml de tampón de peptona para 100 hojas. Los frutos (uno por muestra) se agitan durante 20 min a temperatura ambiente en bolsas estériles que contienen 50 ml de tampón de peptona. El sobrenadante se centrifuga a continuación a 6 000 g durante 20 min y se decanta y el sedimento se vuelve a suspender en 10 ml de solución salina al 0,85 %. Se siembran en estrías, por triplicado, cantidades iguales (100 µl) de diluciones a 1:100 y 1:1 000 de cada una de las suspensiones en medios semiselectivos XOS (sacarosa, 20 g; peptona, 2 g; glutamato monosódico, 5 g; Ca(NO3)2, 0,3 g; K2HPO4, 2 g; EDTA-Fe, 1 mg; cicloheximida, 100 mg; cefalexina, 20 mg; kasugamicina, 20 mg; violeta de metilo 2B, 0,3 mg; agar Bacto™, 17 g; agua destilada, 1 l; pH 7,0) (Monier, 1992). Tras incubar 5-6 días a 28 ºC, se evalúa el crecimiento de colonias, así como su tipo y morfología (sección 3.1.2). 4. Identificación La identificación de presuntas colonias de X. citri subsp. citri debería verificarse mediante varias técnicas, ya que es posible aislar de los cítricos otras especies de Xanthomonas, como X. fuscans subsp. aurantifolii y X. alfalfae subsp. citrumelonis. Además de la observación de las características morfológicas en medios nutritivos, hay otras técnicas: las pruebas serológicas, las pruebas moleculares, los bioensayos de discos de hojas u hojas desprendidas y las pruebas de patogenicidad. Para identificar un cultivo puro tiene que obtenerse como mínimo un resultado positivo en cada una de las tres técnicas siguientes: 1) PCR con dos pares de cebadores (sección 4.1); 2) una técnica serológica (inmunofluorescencia, ELISA en fase doble de anticuerpos (DAS-ELISA) o ELISA indirecto, secciones 4.2, y 4.2.1 y 4.2.2) que utiliza anticuerpos monoclonales específicos, y 3) pruebas de patogenicidad mediante la inoculación de cítricos hospedantes para cumplir los requisitos de los postulados de Koch (secciones 4.3 y 3.1.6). Se podrán efectuar pruebas adicionales (secciones 4.4 y 4.5) para caracterizar más detalladamente la cepa presente. Todas las pruebas deberán incluir controles negativos y positivos. En las secciones sucesivas se describen las técnicas recomendadas. Se pueden obtener cepas de referencia de X. citri subsp. citri de las siguientes colecciones, entre otras (se indican las cepas aisladas de X. citri subsp. citri recomendadas para su utilización como controles positivos): NCPPB 3234, de la Collection of Plant Pathogenic Bacteria, del Central Science Laboratory, York (Reino Unido). CFBP 2911, de la Collection Française de Bactéries Phytopathogènes, de la Station de Phytobactériologie de l'INRA, Angers (Francia) (se trata de una cepa A * de X. citri subsp. citri). ICMP 24, de la International Collection of Microorganisms from Plants, Landcare Research (Manaaki Whenua) New Zealand Ltd, Auckland (Nueva Zelandia). ATTC 49118, de la American Type Culture Collection, Manassas, Virginia (Estados Unidos). IBSBF 1594, de la Coleção de Culturas de Fitobactérias, del Instituto Biológico, del Laboratório de Bacteriologia Vegetal del Centro Experimental Central do Instituto Biológico (CEIB), Campinas (Brasil). PD 6-10 Convención Internacional de Protección Fitosanitaria Protocolos de diagnóstico para plagas reglamentadas PD 6 La autenticidad de las cepas solo se puede garantizar si se obtienen directamente de las colecciones de cultivos. 4.1 Métodos de PCR Se recomienda que, además del protocolo de PCR descrito en la sección 3.1.4.3, la identificación de cultivos puros de cepas sospechosas se confirme utilizando dos pares diferentes de cebadores. Uno de los pares debería ser el formado por los cebadores J-pth1/J-pth2 o J-Rxg/J-Rxc2 (Cubero y Graham, 2002) y el otro par el compuesto por Xac01/Xac02 (Coletto-Filho et al., 2005) o XACF/XACR (Park et al., 2006) (cuadro 1). Esto se debe a que se ha constatado que los pares de cebadores utilizados en la mayoría de los estudios publicados carecen de especificidad (Delcourt et al., 2013). Una confirmación adicional de la identificación puede obtenerse secuenciando los amplicones resultantes de la PCR y comparando sus secuencias con las de las cepas de X. citri subsp. citri depositadas en la base de datos GenBank del Centro Nacional de Información Biotecnológica de los Estados Unidos (NCBI). El protocolo de PCR de Cubero y Graham (2002) desarrolló cebadores de PCR para las regiones del espaciador transcrito interno (ITS) de los ADNr 16S y 23S específicos de X. citri subsp. citri. La variación de las secuencias del ITS permitió diseñar cebadores específicos para X. citri subsp. citri, los cuales detectan las cepas atípicas A* y Aw (Cubero y Graham, 2002). Estos cebadores son: J-Rxg: 5′-GCGTTGAGGCTGAGACATG-3′ J-RXc2: 5′-CAAGTTGCCTCGGAGCTATC-3′. La PCR se efectúa en mezclas de reacción de 25 μl que contienen tampón de Taq 1×, 1,5 mM de MgCl2, 0,04 μM de cebador J-RXg, 0,04 μM de cebador J-RXc2, 0,2 mM de cada dNTP y 1 U de polimerasa Taq de ADN. Las condiciones de la amplificación de la PCR son las mismas que las empleadas con los cebadores pthA descritas en la sección 3.1.4.3. El protocolo de PCR de Coletta-Filho et al. (2006) desarrolló cebadores basados en el complejo génico rpf. Estos cebadores son: Xac01: 5′-CGCCATCCCCACCACCACCACGAC-3′ Xac02: 5′-AACCGCTCAATGCCATCCACTTCA-3′. La PCR se efectúa en mezclas de reacción de 25 μl que contienen tampón de Taq 1×, 2,0 mM de MgCl2, 0,36 μM de cada cebador, 0,25 mM de cada dNTP y 1 U de polimerasa Taq de ADN. Las condiciones de la amplificación de la PCR consisten en una fase inicial de desnaturalización a 94 ºC durante 3 min, seguida de 36 ciclos de 94 ºC durante 45 s, 60 ºC durante 45 s y 72 ºC durante 45 s, y una fase de extensión final de 72 ºC durante 5 min. El tamaño de los amplicones es de 582 pb. El protocolo de PCR de Park et al. (2006) desarrolló cebadores basados en las secuencias génicas hrpW. Estos cebadores son: XACF: 5′- CGTCGCAATACGATTGGAAC-3′ XACR: 5′- CGGAGGCATTGTCGAAGGAA-3′. La PCR se efectúa en mezclas de reacción de 25 μl que contienen tampón de Taq 1×, 1,5 mM de MgCl2, 0,10 μM de cada cebador, 0,25 mM de cada dNTP, 0,01 % de gelatina y 2 U de polimerasa Taq de ADN. Las condiciones de la amplificación de la PCR consisten en una fase inicial de desnaturalización a 94 ºC durante 5 min, seguida de 30 ciclos de 94 ºC durante 15 s, 60 ºC durante 30 s y 72 ºC durante 30 s, y una fase de extensión final de 72 ºC durante 7 min. El tamaño de los amplicones es de 561 pb. Convención Internacional de Protección Fitosanitaria PD 6-11 PD 6 Protocolos de diagnóstico para plagas reglamentadas Cuadro 1. Resumen de los métodos de PCR descritos en el presente protocolo de diagnóstico. Los datos de especificidad se toman de Delcourt et al. (2013). * La detección no específica se refiere al porcentaje de Xanthomonas patogénicas y saprofitos cuyas pruebas dieron positivo. ** No dieron positivo con cepas saprófitas. 4.2 Par de cebadores Referencia Tamaño de los amplicones (pb) Detección de cepas de X. citri subsp. citri Detección no específica (%)* Límites de detección en material vegetal 2/3 Hartung et al. (1993) 224 No detecta w cepas A ni todas las cepas A* 17 10 ufc/ml J-pth1/J-pth2 Cubero y Graham (2002) 198 Todas las cepas 51 10 ufc/ml J-Rxg/J-Rxc2 Cubero y Graham (2002) 179 Todas las cepas 30 10 ufc/ml Xac01/Xac02 Coletto-Filho et al. (2005) 582 Todas las cepas 16 10 ufc/ml XACF/XACR Park et al. (2006) 561 Todas las cepas 6** No hay datos 2 4 4 4 Detección serológica Se recomienda utilizar diferentes anticuerpos para identificar cultivos puros, además del protocolo de inmunofluorescencia descrito en la sección 3.1.3. Asimismo, se puede recurrir al DAS-ELISA o ELISA indirecto como pruebas serológicas alternativas para identificar cultivos puros. 4.2.1 DAS-ELISA Para el DAS-ELISA, las placas de microtitulación se cubren con 100 µl/hoyo de tampón de recubrimiento de carbonato (Na2CO3, 1,59 g; NaHCO3, 2,93 g; NaN3, 0,2 g; agua destilada, 1 litro; pH 9,6) que contiene inmunoglobulinas (IgG) anti-X. citri subsp. citri debidamente diluidas y se incuban a 4 ºC durante la noche. Después de lavar las placas tres veces con PBS-Tween (NaCl, 8 g; KH2PO4, 0,2 g; Na2HPO4·12H2O, 2,9 g; KCl, 0,2 g; NaN3, 0,2 g; Tween 20, 0,25 ml; agua destilada, 1 litro; pH 7,4), se añaden (200 µl/hoyo) la muestra de la prueba, el control negativo (material vegetal sano) o el control positivo (cepa de referencia de X. citri subsp. citri). Las placas se incuban a 37 ºC durante 2 h. Después del lavado, se añade (200 µl/hoyo) una dilución apropiada de las IgG anti-X. citri subsp. citri conjugadas con fosfatasa alcalina en PBS-Tween y las placas se incuban a 37 ºC durante 2 h. Después del lavado, se añade (200 µl/hoyo) tampón sustrato de p-nitrofenil-fosfato (1 mg/ml) y las placas se incuban a temperatura ambiente durante 30–60 min. Se miden las absorbancias con un espectrofotómetro dotado de un filtro de 405 nm. El criterio para determinar que una muestra es positiva es que su densidad óptica (DO) sea el doble que la del control de material vegetal sano. El límite de detección del DAS-ELISA es de 104–105 ufc/ml (Civerolo y Fan, 1982). Este método no se recomienda para la detección directa en tejido vegetal. Existen anticuerpos monoclonales para ELISA, pero se aconseja utilizarlos únicamente para identificar cultivos puros debido a su escasa sensibilidad de detección en tejido vegetal. Se dispone de kits comerciales para la detección de X. citri subsp. citri mediante ELISA (p. ej., de Agdia, Inc.). Para los datos de especificidad, consúltese la información técnica facilitada por el fabricante. Se han constatado reacciones cruzadas de varios anticuerpos monoclonales con X. axonopodis pv. phaseoli, X. campestris pv. zinnea, X. alfalfae subsp. citrumelonis y Xanthomonas hortorum pv. pelargonii; no obstante, la presencia de estos patovares en cítricos es poco probable. PD 6-12 Convención Internacional de Protección Fitosanitaria Protocolos de diagnóstico para plagas reglamentadas PD 6 4.2.2 ELISA indirecto El ELISA indirecto con anticuerpos monoclonales descrito por Álvarez et al. (1991) se puede utilizar para identificar cultivos. Se comercializan equipos de ELISA que incluyen todos los componentes necesarios para la identificación de X. citri subsp. citri (p. ej., de Agdia, Inc.). En teoría, se pueden identificar todas las cepas de X. citri subsp. citri, pero se ha constatado que varias cepas fenotipicamente diferentes aisladas en Asia sudoccidental no reaccionan con los anticuerpos monoclonales disponibles (Vernière et al., 1998). Las suspensiones de cultivos puros se centrifugan durante 2 min a 10 000 g, aproximadamente, y el sobrenadante se desecha. Se añade 1 ml de PBS 1× y las células se vuelven a suspender mezclando en vórtex. Esta operación se repite dos veces más. Después del tercer lavado, las células se vuelven a suspender en tampón de recubrimiento. La concentración bacteriana se ajusta espectrofotométricamente a una DO600 de 0,01 (aproximadamente 2,5 × 107 ufc/ml). Se cargan porciones de las muestras en placas de microtitulación (dos hoyos por muestra, 100 µl/hoyo). Debería incluirse un control positivo (un cultivo o muestra de referencia facilitados por el fabricante) y un control de tampón negativo con otra bacteria. Las placas se incuban a 37 ºC durante la noche hasta su secado. Se añade 200 µl/hoyo de solución de bloqueo (5 % de leche desnatada en polvo en PBS). Las placas se incuban a temperatura ambiente durante 30 min y a continuación se lavan dos veces con PBS-Tween 1×. Se añade (100 µl/hoyo) el anticuerpo principal a la dilución apropiada en 2,5 % de leche en polvo en PBS-Tween. Las placas se incuban a temperatura ambiente durante 1 h y a continuación se lavan cinco veces con PBS-Tween 1×. Se añade (100 µl/hoyo) el conjugado enzimático a la dilución apropiada en 2,5 % de leche en polvo en PBS-Tween. Las placas se incuban a temperatura ambiente durante 1 h y a continuación se lavan cinco veces con PBS-Tween 1×. Se añade (100 µl/hoyo) solución de sustrato recién preparada que contiene 1 mg/ml de p-nitrofenil-fosfato en tampón de dietanolamina (pH 9,8). Las placas se incuban a temperatura ambiente durante 30–60 min. Se mide la DO con un espectrofotómetro dotado de un filtro de 405 nm. Las muestras positivas se determinan de la misma forma que para el DAS-ELISA. 4.3 Pruebas de patogenicidad Para confirmar el diagnóstico, debería determinarse la patogenicidad de X. citri subsp. citri para un conjunto de hospedantes indicadores como C. paradisi var. Duncan (pomelo), Citrus sinensis (naranjo dulce Valencia) o C. aurantiifolia (lima mexicana). La patogenicidad de colonias bacterianas puede demostrarse en ensayos en hojas de cultivares susceptibles de hospedantes del género Citrus mediante infiltración con una jeringuilla con o sin aguja. Es preferible utilizar hojas inmaduras, con una expansión del 50 al 70 %, debido a su mayor nivel de susceptibilidad. Las lesiones se desarrollan de 7 a 14 días después de la inoculación de hojas intactas o desprendidas (Francis et al., 2010; Koizumi, 1971) tras su incubación a 25 ºC con humedad elevada. Con estos ensayos, se puede distinguir perfectamente la reacción eruptiva de tipo calloso de X. citri subsp. citri. Se vuelven a suspender en agua destilada estéril bacterias cultivadas en medios líquidos o colonias de una placa de agar recién sembrada y se ajusta la concentración a 10 6–108 ufc/ml para su inoculación en los hospedantes. Deberían incluirse siempre controles negativos y positivos. Las plantas inoculadas con la cepa de control positivo deberían mantenerse apartadas de las plantas analizadas en la prueba. 4.4 Descripción y características bioquímicas X. citri subsp. citri es una bacteria gramnegativa, recta y baciliforme que mide 1,5-2,0 µm × 0,5-0,75 µm. Posee movilidad gracias a un único flagelo polar. Comparte numerosas propiedades fisiológicas y bioquímicas con otras especies del género Xanthomonas. Es quimioorganotrofa y aerobia obligada, con metabolismo oxidativo de la glucosa. Contiene el pigmento amarillo xantomonadina. En el cuadro 2 figuran algunas de las características bioquímicas distintivas de X. citri subsp. citri. Convención Internacional de Protección Fitosanitaria PD 6-13 PD 6 Protocolos de diagnóstico para plagas reglamentadas Cuadro 2. Principales características bioquímicas de Xanthomonas citri subsp. citri Prueba Resultado Catalasa + Oxidasa – o débil Reducción de nitratos – Hidrólisis de: almidón + caseína + Tween 80 + esculina + Licuación de la gelatina + Licuación del gel de pectato + Utilización de la asparagina – Necesidades para el crecimiento: 4.5 metionina + cisteína + 0,02 % (m/v) de cloruro de trifenil tetrazolio – Identificación molecular Se han caracterizado a nivel molecular las propiedades de diversas especies de xantomonas que infectan a los cítricos, incluidas X. citri subsp. citri y el género Xanthomonas en su conjunto, a fin de desarrollar métodos rápidos y precisos para su reclasificación e identificación. Los procedimientos incluyen la hibridación ADN-ADN (Vauterin et al., 1995), la identificación genómica (Hartung et al., 1987; Lazo et al., 1987), el análisis de secuencias multilocus (Young et al., 2008) y la rep-PCR (Cubero y Graham, 2002, 2004). 4.5.1 Análisis de secuencias multilocus Se ha utilizado un enfoque basado en el análisis de secuencias multilocus (MLSA) para la identificación específica de X. citri subsp. citri. (Almeida et al., 2010; Bui Thi Ngoc et al., 2010; Young et al., 2008). Los genes de mantenimiento se amplifican usando cebadores y condiciones de PCR como las descritas por Almeida et al. (2010), Bui Thi Ngoc et al. (2010) y Young et al., (2008). El MLSA consiste en la secuenciación de varios locus (por lo general, entre cuatro y ocho genes de mantenimiento) y la comparación de sus secuencias con secuencias de referencia de especies de Xanthomonas depositadas en bases de datos de nucleótidos; por ejemplo, la base de datos de fitomicroorganismos Plant Associated Microbes Database, PAMDB (http://genome.ppws.vt.edu/cgibin/MLST/home.pl) (Almeida et al., 2010) y el banco de datos de genotipos microbianos MLVAbank (https://bioinfo-prod.mpl.ird.fr/MLVA_bank/Genotyping/). 4.5.2 Identificación por rep-PCR La identificación mediante rep-PCR con cebadores diseñados a partir de elementos palindrómicos extragénicos repetitivos (REP) – secuencias intergénicas de consenso repetitivas de enterobacterias (ERIC) y el elemento BOX (Louws et al., 1994) – se puede utilizar para identificar y caracterizar cepas en condiciones de PCR determinadas (Cubero y Graham, 2002). El ADN se puede extraer de suspensiones bacterianas (absorbancia a 600 nm de 0,2 a 0,5) en un único paso con fenol-cloroformo-alcohol isoamílico. A continuación, se precipita en etanol y se vuelve a suspender en agua ultrapura. El ADN se almacena a -20 ºC hasta su uso. También se puede utilizar el procedimiento de extracción del ADN descrito en la sección 3.1.4.2. PD 6-14 Convención Internacional de Protección Fitosanitaria Protocolos de diagnóstico para plagas reglamentadas PD 6 La PCR BOX se efectúa en mezclas de reacción de 25 μl que contienen tampón de Taq 1×, 6 mM de MgCl2, 2,4 µM de cebador BOX1R (5′-CTACG-GCAAGGCGACGCTGCAG-3′) (Louws et al., 1994), 0,2 mM de cada dNTP, 2 U de polimerasa Taq de ADN y 5 µl de ADN extraído de cepas de Xanthomonas. Las condiciones de la reacción consisten en una fase inicial a 94 ºC durante 5 min, seguida de 40 ciclos de 94 ºC durante 30 s, 48 ºC durante 30 s y 72 ºC durante 1 min, y una fase de extensión final de 72 ºC durante 10 min. Los productos de PCR se analizan en geles de agarosa al 3 % en tampón de tris-acetato-EDTA (TAE) 1× (40 mM de Tris-acetato; 1 mM de EDTA; pH 8,0) teñidos con bromuro de etidio que se exponen durante 2 h a 110 V. La PCR ERIC se efectúa en mezclas de reacción de 25 μl que contienen tampón de Taq 1×, 3 mM de MgCl2, 1,2 µM de cebador ERIC1R (5′-ATGTAAGCTCCT-GGGGATTCAC-3′) y ERIC2 (5′AAGTAAGTGACT-GGGGTGAGCG-3′) (Louws et al., 1994), 0,2 mM de cada dNTP, 2 U de polimerasa Taq de ADN y 5 µl de ADN extraído de cepas de xantomonas. Las condiciones de la reacción son las mismas que se emplean para la PCR BOX. Para la visualización de los productos de la PCR se sigue el mismo procedimiento que para la PCR BOX. Las huellas genéticas (patrones de bandas) se pueden comparar y analizar a simple vista en busca de similitudes, pero también se pueden transformar los patrones en picos y compararse las cepas mediante un programa informático como BioNumerics (Applied Maths). La identificación debería basarse en la similitud con los patrones de las cepas de control (referencia) (sección 4). En las figuras 5 y 6 se representan esquemas para la detección y la identificación de Xanthomonas citri subsp. citri en material vegetal sintomático y asintomático, respectivamente. 5. Registros Los registros y las evidencias deberían conservarse según lo descrito en la NIMF 27 (Protocolos de diagnóstico para las plagas reglamentadas). En los casos en que los resultados de los diagnósticos puedan afectar negativamente a otras partes contratantes, se recomienda conservar, al menos durante un año, los cultivos de la muestra original de la plaga (etiquetados para facilitar la rastreabilidad), los especímenes conservados o preparados para observación microscópica, o los materiales de las pruebas (p. ej., fotografías de geles, copias impresas de los resultados de los ELISA o amplicones de la PCR), especialmente en casos de incumplimiento (NIMF 13 (Directrices para la notificación de incumplimiento y acción de emergencia)) o cuando las plagas se encuentren por primera vez en un país o área. 6. Puntos de contacto para información adicional Departamento de Laboratorios Biológicos, Dirección General de Servicios Agrícolas, Av. Millán 4703, CP 12900, Montevideo (Uruguay) (Enrique F. Verdier; correo electrónico: [email protected]; tel.: +598 23043992). Centro de Protección Vegetal y Biotecnología, Instituto Valenciano de Investigaciones Agrarias (IVIA), Carretera Moncada-Náquera km 4.5, 46113 Moncada (Valencia, España) (María M. López; correo electrónico: [email protected]; tel.: +34 963424000; fax: +34 963424001). Instituto Nacional de Investigación Agraria y Tecnología Alimentaria, INIA, Ctra. de La Coruña km 6, Madrid (España) (Jaime Cubero; correo electrónico: [email protected]; tel.: +34 913473900; fax: +34 913572293). Podrán presentar solicitudes de revisión de los protocolos de diagnóstico las organizaciones nacionales de protección fitosanitaria (ONPF), las organizaciones regionales de protección fitosanitaria (ORPF) o los órganos auxiliares de la Comisión de Medidas Fitosanitarias (CMF), por conducto de la Secretaría de la CIPF ([email protected]), que las remitirá al Grupo técnico sobre protocolos de diagnóstico (GTPD). Convención Internacional de Protección Fitosanitaria PD 6-15 PD 6 7. Protocolos de diagnóstico para plagas reglamentadas Agradecimientos El primer proyecto del presente protocolo fue redactado por el Sr. E. F. Verdier, del Departamento de Laboratorios Biológicos de la Dirección General de Servicios Agrícolas del Uruguay (véase la sección 6 para más información), y revisado por la Sra. R. Lanfranchi, del Laboratorio de Fitopatología del Servicio Nacional de Sanidad y Calidad Agroalimentaria (SENASA), Av. Ing. Huergo 1001 CP 1107, Buenos Aires (Argentina) (Rita Lanfranchi; correo electrónico: [email protected]; tel.: +5411 43621177 int. 118); el Sr. Ed Civerolo, del Departamento de Agricultura de los Estados Unidos (USDA), Estados Unidos (correo electrónico: [email protected]), y la Sra. M. M. López, del IVIA (España) (véase la sección 6 para más información). Asimismo, el Sr. J. Cubero, del INIA (España) (véase la sección 6 para más información), colaboró de forma destacada en la elaboración del presente protocolo. 8. Referencias La presente norma refiere a las Normas Internacionales para Medidas Fitosanitarias (NIMF). Las NIMF se encuentran disponibles en el Portal fitosanitario internacional (PFI) en https://www.ippc.int/core-activities/standards-setting/ispms. Almeida, N. F., Yan, S., Cai, R., Clarke, C. R., Morris, C. E., Schaad, N. W., Schuenzel, E. L., Lacy, G. H., Sun, X., Jones, J. B., Castillo, J. A., Bull, C. T., Leman, S., Guttman, D. S., Setubal, J. C. & Vinatzer, B. A. 2010. PAMDB, a multilocus sequence typing and analysis database and website for plant-associated microbes. Phytopathology, 100(3): 208–215. Álvarez, A. M., Benedict, A. A., Mizumoto, C. Y., Pollard, L. W. & Civerolo, E. L. 1991. Analysis of Xanthomonas campestris pv. citri and X.c. pv. citrumelo with monoclonal antibodies. Phytopathology, 81: 857–865. Bui Thi Ngoc, L., Vernière, C., Jouen, E., Ah-You, N., Lefeuvre, P., Chiroleu, F., Gagnevin, L. & Pruvost, O. 2010. Amplified fragment length polymorphism and multilocus sequence analysis-based genotypic relatedness among pathogenic variants of Xanthomonas citri pv. citri and Xanthomonas campestris pv. bilvae. International Journal of Systematic and Evolutionary Microbiology, 60(3): 515–525. Bull, C. T., De Boer, S. H., Denny, T. P., Firrao, G., Fischer-Le Saux, M., Saddler, G. S., Scortichini, M., Stead, D. E. & Takikawa, Y. 2010. Comprehensive list of names of plant pathogenic bacteria, 1980–2007. Journal of Plant Pathology, 92(3): 551–592. CABI. 2006. Crop protection compendium. Wallingford, UK, CABI. Civerolo, E. L. & Fan, F. 1982. Xanthomonas campestris pv. citri detection and identification by enzyme-linked immunosorbent assay. Plant Disease, 66: 231–236. Coletta‐Filho, H. D., Takita, M. A., Souza, A. A., Neto, J. R., Destefano, S. A. L., Hartung, J. S. & Machado, M. A. 2006. Primers based on the rpf gene region provide improved detection of Xanthomonas axonopodis pv. citri in naturally and artificially infected citrus plants. Journal of Applied Microbiology, 100(2): 279–285. Cubero, J. & Graham, J. H. 2002. Genetic relationship among worldwide strains of Xanthomonas causing canker in citrus species and design of new primers for their identification by PCR. Applied and Environmental Microbiology, 68: 1257–1264. Cubero, J. & Graham, J. H. 2004. The leucine-responsive regulatory protein (lrp) gene for characterization of the relationship among Xanthomonas species. International Journal of Systematic and Evolutionary Microbiology, 54: 429–437. Cubero, J. & Graham, J. H. 2005. Quantitative real time polymerase chain reaction for bacterial enumeration and allelic discrimination to differentiate Xanthomonas strains on citrus. Phytopathology, 95: 1333–1340. Cubero, J., Graham, J. H. & Gottwald, T. R. 2001. Quantitative PCR method for diagnosis of citrus bacterial canker. Applied and Environmental Microbiology, 67: 2849–2852. PD 6-16 Convención Internacional de Protección Fitosanitaria Protocolos de diagnóstico para plagas reglamentadas PD 6 Delcourt, S., Vernière, C., Boyer, C., Pruvost, O., Hostachy, B. & Robène-Soustrade, I. 2013. Revisiting the specificity of PCR primers for diagnostics of Xanthomonas citri pv. citri by experimental and in silico analyses. Plant Disease, 97(3): 373–378. Dye, D. W. 1978. Genus IX. Xanthomonas Dowson 1939. In: Young, J. M., Dye, D. W., Bradbury, J. F., Panagopoulos, C. G., & Robbs, C. F. A proposed nomenclature and classification for plant pathogenic bacteria. New Zealand Journal of Agricultural Research 21(1): 153-177. EPPO (European and Mediterranean Plant Protection Organization). 1979. Xanthomonas axonopodis pv. citri. Data Sheets on Quarantine Pests. EPPO A1 list No. 1. Paris, EPPO. EPPO (European and Mediterranean Plant Protection Organization). 1998. Phytosanitary procedure Xanthomonas axonopodis pv. citri. Inspection, test and survey methods. EPPO Standard PM 3/27(1). Paris, EPPO. EPPO (European and Mediterranean Plant Protection Organization). 2006. PQR database (versión 4.5). Paris, EPPO. EPPO (European and Mediterranean Plant Protection Organization). 2009. Indirect immunofluorescence test for plant pathogenic bacteria. EPPO Standard PM 7/97(1). Paris, EPPO. Escalon, A., Javegny, S., Vernière, C., Noël, L. D., Vital, K., Poussier, S., Hajri, A., Boureau, T., Pruvost, O., Arlat, M. & Gagnevin, L. 2013. Variations in type III effector repertoires, pathological phenotypes and host range of Xanthomonas citri pv. citri pathotypes. Molecular Plant Pathology, 14(5): 483–496. Francis, M. I., Pena, A. & Graham, J. H. 2010. Detached leaf inoculation of germplasm for rapid screening of resistance to citrus canker and citrus bacterial spot. European Journal of Plant Pathology, 127(4): 571–578. Gabriel, D. W., Kingsley, M. T., Hunter, J. E. & Gottwald, T. 1989. Reinstatement of Xanthomonas citri (ex Hasse) and X. phaseoli (ex Smith) to species and reclassification of all X. campestris pv. citri strains. International Journal of Systematic Bacteriology, 39(1): 14–22. Golmohammadi, M., Cubero, J., Peñalver, J., Quesada, J. M., López, M. M. & Llop P. 2007. Diagnosis of Xanthomonas axonopodis pv. citri, causal agent of citrus canker in commercial fruits by isolation and PCR based methods. Journal of Applied Microbiology, 103(6): 2309–2315. Goto, M. 1992. Citrus canker. In J. Kumer, H. S. Chaube, U. S. Singh and A. N. Mukhopadhyay, eds. Plant diseases of international importance, Vol. III, Diseases of fruit crops, pp. 170–208. Upper Saddle River, NJ, Prentice Hall. Graham, J., Gottwald, T. R., Civerolo, E. L. & McGuire, R. G. 1989. Population dynamics and survival of Xanthomonas campestris in soil in citrus nurseries in Maryland and Argentina. Plant Disease, 43(5): 423–427. Hall, D. G., Gottwald, T. R. & Bock, C. H. 2010. Exacerbation of citrus canker by citrus leafminer Phyllocnistis citrella in Florida. Florida Entomologist, 93(4): 558–566. Hartung, J. S. & Civerolo, E. L. 1987. Genomic fingerprinting of Xanthomonas campestris pv. citri strains from Asia, South America and Florida. Phytopathology, 77: 282–285. Hartung, J. S., Daniel, J. F., Pruvost, O. P. & Civerolo, E. L. 1993. Detection of Xanthomonas campestris pv. citri by the polymerase chain reaction method. Applied and Environmental Microbiology, 59(4): 1143–1148. Hasse, CH. 1915. Pseudomonas citri, the cause of citrus canker. A preliminary report. Journal of Agricultural Research 4, 97-100. Koizumi, M. 1971. A quantitative determination method for Xanthomonas citri by inoculation into detached citrus leaves. Bulletin of the Horticultural Research Station (Japan), Series B, 11: 167–182. Convención Internacional de Protección Fitosanitaria PD 6-17 PD 6 Protocolos de diagnóstico para plagas reglamentadas Lazo, G. R., Roffey, R. & Gabriel, D. W. 1987. Pathovars of Xanthomonas campestris are distinguishable by restriction fragment-length polymorphism. International Journal of Systematic Bacteriology, 37: 214–221. Llop, P., Caruso, P., Cubero, J., Morente, C. & López, M. M. 1999. A simple extraction procedure for efficient routine detection of pathogenic bacteria in plant material by polymerase chain reaction. Journal of Microbiology Methods, 37: 23–31. Louws, F. J., Fulbright, D. W., Taylor Stephens, C. & Bruijn, F.J. 1994. Specific genomic fingerprints of phytopathogenic Xanthomonas and Pseudomonas pathovars and strains generated with repetitive sequences and PCR. Applied and Environmental Microbiology, 60: 2286–2295. Mafra, V., Kubo, K. S., Alves-Ferreira, M., Ribeiro-Alves, M., Stuart, R. M., Boava, L. P., Rodrigues, C. M. & Machado, M. A. 2012. Reference genes for accurate transcript normalization in citrus genotypes under different experimental conditions. PloS One, 7(2), e31263. Mavrodieva, V., Levy, L. & Gabriel, D. W. 2004. Improved sampling methods for real-time polymerase chain reaction diagnosis of citrus canker from field samples. Phytopathology, 94: 61–68. Monier, L. 1992. Contribution à la mise au point d´un milieu de culture semi-sélectif pour la détection de Xanthomonas campestris pv. citri, agent du chancre bactérien des agrumes. Angers, France, École Nationale d´Ingénieurs des Travaux de l´Horticulture et du Paysage d´Angers, Institut de Recherches sur les Fruits et Agrumes (IRFA). 62 pp [in French]. Namekata, T. de Oliveira, AR. Comparative serological studies between Xanthomonas citri and a bacterium causing canker on Mexican lime. In Proceedings of International Conference on Plant Pathogenic Bacteria, Wageningen, The Netherlands, 1972, pp.151-152 Park, D., Hyun, J., Park, Y., Kim, J., Kang, H., Hahn, J. & Go, S. 2006. Sensitive and specific detection of Xanthomonas axonopodis pv. citri by PCR using pathovar specific primers based on hrpW gene sequences. Microbiological Research, 161(2): 145–149. Pruvost, O., Roumagnac, P., Gaube, C., Chiroleu, F. & Gagnevin, L. 2005. New media for the semiselective isolation and enumeration of Xanthomonas campestris pv. mangiferaeindicae, the causal agent of mango bacterial black spot. Journal of Applied Microbiology, 99(4): 803–815. Schaad, N. W., Postnikova, E., Lacy, G. H., Sechler, A., Agarkova, I., Stromberg, P. E., Stromberg, V. K. & Vidaver, A. K. 2005. Reclassification of Xanthomonas campestris pv. citri (ex Hasse 1915) Dye 1978 forms A, B/C/D, and E as X. smithii subsp. citri (ex Hasse) sp. nov. nom. rev. comb. nov., X. fuscans subsp. aurantifolii (ex Gabriel et al., 1989) sp. nov. nom. rev. comb. nov., and X. alfalfae subsp. citrumelo (ex Riker and Jones) Gabriel et al., 1989 sp. nov. nom. rev. comb. nov.; X. campestris pv. malvacearum (ex Smith 1901) Dye 1978 as X. smithii subsp. smithii nov. comb. nov. nom. nov.; X. campestris pv. alfalfae (ex Riker and Jones, 1935) Dye 1978 as X. alfalfae subsp. alfalfae (ex Riker et al., 1935) sp. nov. nom. rev.; and "var. fuscans" of X. campestris pv. phaseoli (ex Smith, 1987) Dye 1978 as X. fuscans subsp. fuscans sp. nov. Systematic and Applied Microbiology, 28: 494–518. Schaad, N. W., Postnikova, E., Lacy, G. H., Sechler, A., Agarkova, I., Stromberg, P. E., Stromberg, V. K. & Vidaver, A. K. 2006. Emended classification of xanthomonad pathogens on citrus. Systematic and Applied Microbiology, 29: 690–695. Schaad, N. W., Postnikova, E., Lacy, G., Sechler, A., Agarkova, I., Stromberg, P. E., Stromberg, V. K. & Vidaver, A. K. (2007). Xanthomonas alfalfae sp. nov., Xanthomonas citri sp. nov. and Xanthomonas fuscans sp. nov. In List of New Names and New Combinations Previously Effectively, but not Validly, Published, Validation List no. 115. International Journal of Systematic and Evolutionary Microbiology, 57: 893–897. Sun, X., Stall, R. E., Jones, J. B., Cubero, J., Gottwald, T. R., Graham, J. H., Dixon, W. D., Schubert, T. S., Chaloux, P. H., Stromberg, V. K., Lacy, G. H. & Sutton, B. D. 2004. PD 6-18 Convención Internacional de Protección Fitosanitaria Protocolos de diagnóstico para plagas reglamentadas PD 6 Detection and characterization of a new strain of citrus canker bacteria from Key/Mexican lime and alemow in South Florida. Plant Disease, 88(11): 1179–1188. Taylor, R. K., Tyson, J. L., Fullerton, R. A. & Hale, C. N. 2002. Molecular detection of exotic phytopathogenic bacteria: A case study involving canker-like symptoms on citrus. New Zealand Plant Protection, 55: 53–57. Vauterin, L., Hoste, B., Kersters, K. & Swings, J. 1995. Reclassification of Xanthomonas. International Journal of Systematic Bacteriology, 45: 472–489. Verdier, E., Zefferino, E. & Méndez, S. 2008. Survival of Xanthomonas axonopodis pv. citri on the surface of citrus fruit after postharvest treatment Fitopatologia, 43: 24–31. Vernière, C., Hartung, J. S., Pruvost, O. P., Civerolo, E. L., Álvarez, A. M., Maestri, P. & Luisetti, J. 1998. Characterization of phenotypically distinct strains of Xanthomonas axonopodis pv. citri from Southwest Asia. European Journal of Plant Pathology, 104: 477–487. Weisberg, W. G., Barns, S. M., Pelletier, B. A. & Lane, D. J. 1991. 16S ribosomal DNA amplification for phylogenetic study. Journal of Bacteriology, 173: 697–703. Weller, S. A., Elphinstone, J. G., Smith, N. C., Boonham, N., & Stead, D. E. 2000. Detection of Ralstonia solanacearum strains with a quantitative, multiplex, real-time, fluorogenic PCR (TaqMan) Assay. Applied and Environmental Microbiology, 66(7): 2853–2858. Young, J. M., Park, D. C., Shearman, H. M. & Fargier, E. 2008. A multilocus sequence analysis of the genus Xanthomonas. Systematic and Applied Microbiology, 31(5): 366–377. Convención Internacional de Protección Fitosanitaria PD 6-19 PD 6 9. Protocolos de diagnóstico para plagas reglamentadas Figuras Figura 1. Síntomas típicos del cancro de los cítricos en hojas, tallos y fruto de pomelo (Citrus paradisi). Figura 2. Síntomas del cancro de los cítricos en ramillas: lesiones incipientes en pomelo (Citrus paradisi). PD 6-20 Convención Internacional de Protección Fitosanitaria Protocolos de diagnóstico para plagas reglamentadas PD 6 Figura 3. Síntomas del cancro de los cítricos en frutos de naranjo dulce (Citrus sinensis) (izquierda) y de pomelo (Citrus paradisi) (centro y derecha). Figura 4. Síntomas del cancro de los cítricos en hojas de limonero (Citrus limon) agravadas por lesiones del minador de las hojas de los cítricos. Convención Internacional de Protección Fitosanitaria PD 6-21 PD 6 Protocolos de diagnóstico para plagas reglamentadas MATERIAL VEGETAL SINTOMÁTICO (sección 3.1.1) INOCULACIÓN EN DISCOS DE HOJAS Y HOJAS DESPRENDIDAS (sección 3.1.6) MEDIOS DE AISLAMIENTO (sección 3.1.2) PCR (sección 3.1.4) POSITIVO EN TODAS O ALGUNAS PRUEBAS DAS-ELISA E INMUNOFLUORES CENCIA (sección 3.1.3) NEGATIVO EN TODAS LAS PRUEBAS AISLAMIENTO DIRECTO (sección 3.1.2) COLONIAS CON MORFOLOGÍA TÍPICA (sección 3.1.2) SÍ PRUEBAS DE IDENTIFICACIÓN: (sección 4) Resultados positivos de PCR con dos pares diferentes de cebadores de PCR y una prueba serológica con anticuerpos monoclonales específicos (sección 4.2), seguido de una prueba de patogenicidad. Se pueden efectuar pruebas complementarias, como el MLSA (véase la sección 4.5). SÍ X. citri subsp. citri NO NO DETECTADO POSITIVO X. citri subsp. citri identificado Figura 5. Esquema para la detección e identificación de Xanthomonas citri subsp. citri en material vegetal sintomático. PD 6-22 Convención Internacional de Protección Fitosanitaria Protocolos de diagnóstico para plagas reglamentadas PD 6 MATERIAL VEGETAL ASINTOMÁTICO PRUEBAS DE DETECCIÓN: bioensayos en discos de hojas y hojas desprendidas, medio semiselectivos, inmunofluorescencia, pruebas de PCR (sección 3). POSITIVO EN AL MENOS UNA PRUEBA NEGATIVO EN TODAS LAS PRUEBAS AISLAMIENTO DIRECTO (sección 3.1.2) COLONIAS CON MORFOLOGÍA TÍPICA (sección 3.1.2) X. citri subsp. citri NO DETECTADO NO X. citri subsp. citri NO CONFIRMADO SÍ PRUEBAS DE IDENTIFICACIÓN: (sección 4) Resultados positivos de PCR con dos pares diferentes de cebadores de PCR y una prueba serológica con anticuerpos monoclonales específicos (sección 4.2), seguido de una prueba de patogenicidad. Se pueden efectuar pruebas complementarias, como el MLSA (véase la sección 4.5). SÍ NO POSITIVO X. citri subsp. citri identificado Figura 6. Esquema para la detección e identificación de Xanthomonas citri subsp. citri en material vegetal asintomático. Convención Internacional de Protección Fitosanitaria PD 6-23 PD 6 Protocolos de diagnóstico para plagas reglamentadas Historia de la publicación Esta no es una parte oficial de la norma Esta historia de la publicación se refiere sólo a la versión española. Para la historia completa de la publicación, consulte la versión en inglés de la norma. 2004-11 El CN añadió la cuestión Xanthomonas axonopodis pv. citri (2004-011) al programa de trabajo. La CMF-1 (2006) añadió la cuestión Xanthomonas axonopodis pv. citri (2004011) al tema: Bacterias (2006-005). 2012-11 El GTPD revisó el proyecto de protocolo. 2013-04 El CN aprobó, mediante decisión por vía electrónica, el proyecto de protocolo para consulta a los miembros (2013_eSC_May_12). 2013-07 Consulta a los miembros. 2014-02 Revisado y remitido por el GTPD al CN para que apruebe su adopción (2014_eTPDP_Feb_02). 2014-04 Remitido al CN para que apruebe su adopción mediante decisión por vía electrónica (2014_eSC_May_16). 2014-06 El CN aprobó el proyecto, mediante decisión por vía electrónica, para el período de notificación de 45 días (2014_eSC_Nov_03). 2014-07 El CN adoptó el PD en nombre de la CMF (no se recibieron objeciones formales). 2014-10 La Secretaría efectuó pequeñas correcciones en la redacción. 2014-11 La Secretaría efectuó pequeñas correcciones en la redacción. NIMF 27. Anexo 6 Xanthomonas citri subsp. citri (2014). Roma, CIPF, FAO. 2015-09 La Secretaría de la CIPF incorporó las enmiendas a tinta en conformidad con el procedimiento de revocación de las normas aprobado por la CMF-10 (2015). 2016-01 La secretaría de la CIPF hizo un cambio editorial menor. 2016-04 La CMF-11 ha tomado nota de los cambios editoriales efectuados en español por el grupo de examen de los idiomas. Última actualización de la historia de la publicación: 2016-08. PD 6-24 Convención Internacional de Protección Fitosanitaria Esta página se ha dejado en blanco intencionalmente CIPF La Convención Internacional de Protección Fitosanitaria (CIPF) es un acuerdo internacional de sanidad vegetal que tiene como objetivo proteger las plantas cultivadas y silvestres previniendo la introducción y propagación de plagas. Los viajes y el comercio internacional hoy son más abundantes que nunca antes. En el desplazamiento de personas y mercancías por todo el mundo, los acompañan organismos que representan riesgos para las plantas. La organización ++ Hay más de 180 partes contratantes de la CIPF ++ Cada parte contratante tiene una organización nacional de protección fitosanitaria (ONPF) y un contacto oficial de la CIPF ++ Nueve organizaciones regionales de protección fitosanitaria (ORPF) obran para facilitar la aplicación de la CIPF en los países ++ La CIPF se enlaza con las organizaciones internacionales pertinentes a fin de contribuir a la creación de capacidad regional y nacional ++ La Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO) proporciona la Secretaría de la CIPF Convención Internacional de Protección Fitosanitaria (CIPF) Viale delle Terme di Caracalla, 00153 Roma, Italia Tel. +39 06 5705 4812 - Fax: +39 06 5705 4819 Correo electrónico: [email protected] - Web: www.ippc.int