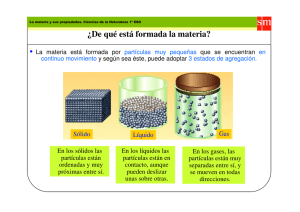
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU
Anuncio

CAPÍTULO 17
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU EMPLEO EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
289
DISEÑO DE PARTÍCULAS FÁGICAS
BIFUNCIONALES PARA SU EMPLEO
EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
HANLET C AMACHO1, ELAINE HONDARES1 Y MARTA DUEÑAS1
1
Centro de Ingienería Genética y Biotecnología. Ave. 31 e/ 158 y 190, Playa. AP 6162,
CP 10600, Ciudad de La Habana, Cuba.
INTRODUCCION
Cuando en 1985, George Smith [1] publicó el primer trabajo en el cual hacía uso
de los fagos filamentosos para la exposición de péptidos en su superficie, unió
en una misma partícula viral dos niveles de conocimientos, los genes y las
proteínas.Sin embargo, como ha pasado antes en la ciencia, no fue hasta 1990
[2] que este procedimiento comenzó a ser realmente explotado de manera extensiva e intensiva.
La metodología que se sigue para la exposición de partículas en la superficie de
los fagos tiene como característica principal que permite relacionar físicamente
el fenotipo y el genotipo de las moléculas expuestas en la superficie de la partícula
viral. De esta forma el ADN presente en el fago contiene la información que
codifica para la proteína o péptido expuesto en su cápsida. Esta propiedad la ha
convertido en una tecnología usada con múltiples fines.
En la última década, la construcción y selección de bibliotecas peptídicas mediante la expresión en la superficie de fagos filamentosos se ha convertido en
una tecnología de punta en muchos laboratorios del mundo. Varios investigadores han desarrollado sistemas de expresión [3], ensamblaje [4] y selección de
partículas [5,6] ajustados a sus intereses particulares por lo que existe en la
actualidad, como parte de la bibliografía científica, varios libros de procedimientos [7] y un arsenal de ejemplos de su empleo en tareas específicas [8-10].
También las partículas virales han sido usadas como inmunógenos, en este caso
segmentos cortos de varias proteínas han sido expuestos en la partícula del
fago M13 con el objetivo de inducir una respuesta humoral contra las proteínas
de la cápsida de numerosos parásitos y virus. La respuesta inmunológica a la
partícula viral M13 es T dependiente y no requiere ayudantes [11]. Varios
segmentos de diferentes proteínas de la cápsida del VIH han sido presentados
en fagos con el propósito de generar una vacuna [12-14].
290
CAPÍTULO 17
VIRUS DE INMUNODEFICIENCIA HUMANA
El virus de inmunodeficiencia humana tipo-1 (VIH-1), fue descrito por primera
vez en el año 1983 y posteriormente designado como el agente etiológico del
síndrome de inmunodeficiencia adquirida (SIDA).
Desde el descubrimiento de los lentivirus VIH-1 y VIH-2 como los agentes
causantes del SIDA, su organización genómica y sus proteínas han sido extensamente estudiadas para crear sistemas de diagnóstico que prevengan la transmisión por VIH, particularmente a través de los sumunistros de sangre. Desde
el primer ensayo de tipo ELISA en el año 1984 [15], estos han progresado
rápidamente.
El comportamiento serológico de un individuo infectado por VIH ha sido estudiado detalladamente. Esta información es utilizada para la detección de la infección y para monitorear la progresión de la enfermedad ya que el perfil
serológico persiste durante meses en los pacientes. El período de tiempo que
media entre la infección y la seroconversión se conoce como período de ventana.
Esencialmente todas las proteínas codificadas por el virus son antigénicas en
alguna fase del curso de la enfermedad. Sin embargo, las más inmunogénicas
son las proteínas estructurales, por lo que son las más utilizadas en el desarrollo
de sistemas de diagnóstico [16].
Los estudios de prevalencia y las concentraciones de anticuerpos demuestran
que los anticuerpos contra las proteínas de la envoltura constituyen marcadores primarios de la seroconversión y para la p41 persisten durante todas las
fases de la enfermedad. Los esfuerzos de desarrollar diagnósticos con el
producto del gen gag y el empleo de polipéptidos derivados de ADN
recombinante incluye toda la proteína p24 o un gran segmento de ella. Se han
realizado investigaciones con este antígeno que mostraron una reducción en
las concentraciones de anticuerpos medidos contra p24, lo cual se correlacionó
con la progresión de la enfermedad. Se ha postulado que este cambio es
debido a la aparición en la circulación del antígeno viral p24 [17]. Basados en
ellos, se han obtenido y evaluado un gran número de variantes de inmunoensayos
con la perspectiva de disponer de uno cada vez más sensible, rápido y asequible a la humanidad.
En los últimos años, se ha ido imponiendo una variante alternativa de desarrollo
que contempla inmunoensayos que no requieren equipamiento para su utilización, y se presentan en formatos rápidos, simples y de lectura visual, que utilizan suero o sangre total como muestra, y cuya ejecución puede ser hecha por
operadores sin un entrenamie nto especial.
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU EMPLEO EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
291
Los ensayos cromatográficos han sido uno de los sistemas analíticos más usados para la detección y/o determinación de sustancias, particularmente de interés biológico, entre ellos anticuerpos inducidos como respuesta a una infección
con patógenos, por ejemplo a la bacteria Helicobacter pylori y a la infección
por VIH. Estos sistemas son frecuentemente usados para el diagnóstico rápido.
Los inmunoensayos han sido desarrollados en estos contextos con muy buenos
resultados.
Además de los ensayos inmunocromatográficos se han desarrollado otros ensayos rápidos basados en la hemoaglutinación.
Un ensayo de aglutinación directa con partículas de latex que contiene una
proteína recombinante obtenida a partir de los genes env ha sido evaluado en
Zaire y autorizado por la FDA para su uso en Estados Unidos [18-19]. El suero
o la sangre completa diluida se mezcla con la partícula de latex a la cual se
encuentra unido el antígeno. En pocos minutos se puede detectar a simple vista
la aglutinación. En estudios de diferentes poblaciones han sido publicados datos
de sensibilidad y especificidad entre 99,3-99,7 % y entre 99,4-100 % respectivamente. Se informa en la literatura la aplicación de ensayos de aglutinación de
anticuerpos para diagnosticar HIV-1 y HIV-2 con una sensibilidad de 100 % y
una especificidad de 99 % [20].
LA SÍFILIS
La sífilis es una enfermedad infecciosa crónica sistémica con evolución en varias fases. Aunque la primera epidemia fue detectada en Europa hacia finales
del siglo XVI [21], el origen geográfico de la enfermedad es muy debatido.
Durante los primeros años del presente siglo la sífilis constituyó una de las
principales causas de enfermedades de origen neurológico y cardiovascular [22].
Desde el año 1946 se estableció un tratamiento antibiótico efectivo contra la
enfermedad, por lo que el número de casos sifilíticos registrados disminuyó
significativamente [23].
A partir del año 1985 se ha observado un incremento marcado en la incidencia
de la sífilis, por lo que actualmente se considera un importante problema de
salud mundial [24].
Adicionalmente, desde el aislamiento del virus de inmunodeficiencia humana
(VIH), numerosos trabajos han revelado la asociación epidemiológica de este
virus con la sífilis [22]. Se plantea que la lesión ulcerativa genital de la sífilis
constituye una puerta de entrada para la adquisición y transmisión del VIH
[25]. A su vez, tanto el tratamiento antibiótico habitual como el curso natural de
292
CAPÍTULO 17
la enfermedad se modifican en sifilíticos coinfectados con VIH [26]. En estos
casos se presentan frecuentemente complicaciones neurológicas inusuales de
la sífilis como la meningitis sifilítica y la vasculitis cerebral
La enfermedad es adquirida en más del 90 % de los casos por contacto sexual
y aproximadamente un 5 % mediante el contacto indirecto con objetos contaminados con secreciones de personas infectadas. La transmisión de la sífilis por
vía parenteral es poco frecuente por lo que se han publicado un número reducido
de casos [27]. Estudios de parejas sexuales enfermas revelaron que el riesgo
de infección es de un 10 a un 60 %. Esto indica que la transmisión ocurre en 1
de cada 3 personas expuestas a la enfermedad [28-29].
Las técnicas serológicas constituyen el soporte principal para el diagnóstico de
la Sífilis en todos sus estadíos, excepto en el período inicial de la lesión primaria,
en cuya etapa aún no hay presencia de reactividad serológica. Se ha comprobado que a partir de la segunda semana de la aparición del chancro comienzan a
detectarse anticuerpos antitreponémicos específicos del tipo IgM y a partir de
la cuarta semana son detectados altos títulos de IgG [30-31]. Por lo tanto durante la primera semana de la infección la microscopía de campo oscuro mediante el examen directo de las lesiones exudativas constituye el procedimiento
diagnóstico de elección. Este método proporciona la identificación de un organismo fino en espiral con morfología y motilidad características del T. pallidum.
La principal limitación de esta técnica es la necesidad de mantener la motilidad
de la bacteria para poder diferenciarla de las bacterias comensales no patógenas
localizadas en la mucosa oral y genital. Para establecer el diagnóstico diferencial en ocasiones es necesario emplear el microscopio de fluorescencia, que
utiliza anticuerpos policlonales y monoclonales contra el T. pallidum marcados
con fluoresceína.
Los procedimientos para el diagnóstico serológico de la sífilis están divididos en
dos grandes grupos. Usualmente para realizar el testaje masivo se utilizan las
técnicas serológicas cuantitativas no específicas de treponemas, entre las cuales las más utilizadas son la prueba reagínica rápida en plasma (RPR) y la del
laboratorio de referencia de enfermedades venéreas (VDRL). Un segundo grupo
de procedimientos serológicos incluye entre otros los de hemaglutinación del T.
pallidum (TPHA) y el de aabsorción de anticuerpos anti-T. pallidum con
inmunofluorescencia (FTA-ABS). Estos últimos ensayos son utilizados generalmente como técnicas diagnósticas confirmatorias, ya que permiten la detección de anticuerpos antitreponémicos específicos
La Hemaglutinación del T. Pallidum es una técnica muy utilizada, en ella, la
reactividad de los sueros comienza a detectarse a partir de la cuarta semana de
la infección, por lo que la sensibilidad para la sífilis primaria no tratada está en el
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU EMPLEO EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
293
rango del 64 al 80 %. Varios autores plantean que el TPHA es más sensible que
el FTA-ABS excepto durante las primeras 3 a 4 semanas de la infección,
estadio en el cual se observa el mayor número de falsos negativos.
En la actualidad está muy difundido para el testaje masivo de muestras el uso
combinado del VDRL y el TPHA lo que ha permitido aumentar la sensibilidad
hasta un 84 % para el diagnóstico de la enfermedad en el estadio primario.
El antígeno TmpA de 42 kD está reconocido como uno de los de mayor importancia en el diagnóstico serológico de la sífilis, debido a la gran sensibilidad y
especificidad que se obtiene cuando se emplea en ensayos serológicos. Una
particularidad interesante de los inmunoensayos desarrollados con este antígeno
es que permite diferenciar pacientes en infección activa de individuos curados
con anticuerpos circulantes. Se han desarrollado ELISAs con TmpA natural y
recombinante [32] con otros antígenos de T. Pallidum. Estos sistemas presentan sensibilidad y especificidad comparable con los valores respectivos reportados para TPHA y para FTA-ABS.
RECEPTORES Y LIGANDOS ESPECÍFICOS
DE CÉLULAS ROJAS HUMANAS
La hemaglutinación, una técnica diagnóstica muy vieja es aún usada debido a su
factibilidad y simplicidad. Existen muchos sistemas diagnósticos de aglutinación
en el mercado, para una gran variedad de enfermedades y estados [18-19]. La
gran mayoría emplea partículas de látex, recubiertas con antígenos ó anticuerpos,
en correspondencia con la naturaleza específica del diagnóstico a realizar.
La aglutinación de hematíes humanos ha sido utilizada durante mucho tiempo
en bancos de sangre y laboratorios clínicos, fundamentalmente en el reconocimiento de antisueros específicos contra algunos marcadores de este tipo de
células. Los receptores más comunes en la tipificación sanguínea son los
antígenos A,B,O y el factor Rh. La presencia de una u otra de estas moléculas
oligosacarídicas, confieren un fenotipo dado (A+/-, B+/-, AB+/- y O+/-). Estos receptores varían en su distribución poblacional y el fenotipo O está dado por la
ausencia de antígeno A y B, por lo que es imposible dirigir un reconocimiento
específico hacia los eritrocitos ocon el uso de ligandos para estos marcadores.
Existen antígenos eritrocitarios cuya variación fenotípica no se basa en la ausencia o presencia del mismo en la membrana de la célula roja, o si lo hace, la
frecuencia de aparición de variantes negativas es muy baja. Este es el caso de
los antígenos del sistema P y de la proteína mayoritaria de la superficie de los
hematíes, la glicoforina A (GFA).
294
CAPÍTULO 17
El antígeno P, está formado por una familia de glicoesfingolípidos, globósidos y
trihexosylceramidas que actúan como receptores de cepas de E. coli
pielonefríticas [33]. El sistema de antígenos P, formado por tres antígenos (P k ,
P 1 y P2 ), proporciona cinco fenotipos, P 1 , P 2 , P k 1 , P k 2 y P -. De estos P 1 y P 2 son los
más abundantes (75 % y 25 % respectivamente) y los P- los menos frecuentes
(< 0.0001%), de acuerdo con los informes de Race y Sanger en 1975 [34].
La otra proteína receptora, la GFA, pertenece a un grupo de sialoglicopéptidos
encontrados en eritrocitos humanos. Es el representante mayoritario de este
grupo (cerca del 75 % del total), y está compuesta por una única cadena
polipeptídica de 131 aminoácidos y por carbohidratos que conforman hasta el
60 % de su masa molecular. Los 70 aminoácidos del extremo amino-terminal
sobresalen de la doble capa lipídica de la membrana, y 16 de ellos están unidos
a carbohidratos. Se conoce, además, que el polimorfismo sanguíneo M ó N
radica en el primer y quinto aminoácidos del extremo amino terminal de la cadena polipeptídica [35]. La GFA aislada de individuos con subgrupo sanguíneo
MN contiene cantidades equimolares de cada especie. En todos los casos, el
segundo, tercer y cuarto aminoácido son O-glicosilados con un tetrasacárido
que contiene dos residuos de ácido N-acetil neuramínico (NANA). Fenotipos
negativos para esta glicoproteína ocurren a muy baja frecuencia en una población normal (< 1 %).
Los ligandos específicos para estos, y otros tipos de marcadores pueden ser
encontrados de forma natural o mediante la inmunización de animales de laboratorio para la obtención de antisueros policlonales y/o anticuerpos monoclonales.
Como parte de los ligandos naturales para los eritrocitos humanos se han
notificado proteínas bacterianas y virales, comúnmente denominadas
adhesinas. Estas se caracterizan por su capacidad de aglutinar células rojas
y han sido utilizadas durante varias décadas como sistema de clasificación
de fenotipos de estos microorganismos por sus patrones de aglutinación
especie-específico [36]. Existen algunas adhesinas propias de bacterias Gram
negativas, típicas de infecciones del tracto urinario y digestivo, con moléculas de adhesión bien caracterizadas [37]. De estas, las más propicias para
ser usadas como ligandos para sistemas diagnósticos son las F41 y la PapG.
La primera es una adhesina bacteriana de la cepa K88, específica contra
residuos proteicos de la GFA, su patrón de reconocimiento abarca las dos
variantes M y N. Específica contra esta proteína de los hematíes, es además
la proteína HN (hemaglutinina) del virus Sendai [38]. Esta es una
glicoproteína en la cual los residuos oligosacarídicos son muy importantes
en el reconocimiento a su receptor.
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU EMPLEO EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
295
La segunda adhesina antes mencionada, la PapG es la proteína responsable del
anclaje de las E. coli enterotoxigénicas (ETEC) [36]. La adhesina Pap G es
aparentemente una molécula con dos dominios. Los estudios de purificación de
variantes de Pap G con el extremo amino terminal truncado, con el empleo de
cromatografía de afinidad con Gal (1-4) Gal, han demostrado que dicho dominio
contiene el sitio de unión al receptor [36]. Además, la fusión del extremo 5’ del
gen pap G, que codifica para el sitio de unión al receptor, con el gen que codifica para la proteína de unión a la maltosa, resultó en un producto final soluble
que une tanto maltosa como Gal (1-4) Gal.
La PapG es, en las bacterias uropatogénicas, la proteína responsable del reconocimiento de su receptor en las células blanco. Este receptor es el carbohidrato α-D galactopiranosil-(1→4)-β-D galactopiranósido {Gal(1→4)Gal} presente
en las globoseries de los glicolípidos de las células que recubren el tracto urinario. Esta estructura está, además, presente en los eritrocitos humanos y su
reconocimiento es independiente de los antígenos A, B y O de grupo sanguíneo
y de los antígenos polisacarídicos Lewis. Basados en estas observaciones, el
grupo de Kallenius en 1980 descubrió que el antígeno reconocido por esta
adhesina debía corresponderse a algún antígeno frecuente en los eritrocitos,
debido a su capacidad de aglutinar a un gran número de estos, independientemente de clasificaciones sanguíneas y encontraron que la aglutinación con bacterias papG+ no ocurría para aquellos individuos con eritrocitos p-(40). De acuerdo
con la clasificación de antígenos eritrocitarios en humanos [34], individuos con
fenotipo p- en eritrocitos son muy raros en una población (< 0.0001 %). Además
esta adhesina es capaz de reconocer con la misma intensidad todas las otras
variantes de este antígeno (P 1 , P2 , Pk 1 y Pk 2 ).
Sobre la base de los elementos anteriormente expuestos fue interesante el empleo de partículas fágicas recombinantes que presenten en su cápsida péptidos
y/o proteínas específicas fusionadas a determinadas proteínas de su envoltura
que permiten su uso en sistemas diagnósticos. De acuerdo con esto, se estructuró
este trabajo sobre la base de los tres objetivos siguientes:
• Obtener un vector de expresión que permita la expresión de proteínas de
fusión con las proteínas p3 y p8 de la envoltura del fago filamentoso M13.
• Expresar y ensamblar partículas virales bifuncionales que tuvieran un motivo
antigénico y un sitio de unión a eritrocitos humanos (SUE).
Estudiar la capacidad aglutinante de estas partículas fágicas en muestras de
individuos infectados.
Se consideró oportuno fusionar al dominio N3 COOH-terminal de la proteína
p3 una proteína que fuera capaz de unirse a eritrocitos y a p8 un polipéptido
296
CAPÍTULO 17
antigénico, de importancia en diagnóstico serológico, representante de un agente infeccioso. Como modelo se determinó evaluar un péptido de 16 aminoácidos
de conocida importancia en el diagnóstico de infección por VIH 1 y una proteína utilizada en el diagnóstico serológico de la sífilis.
DESARROLLO
Hasta la fecha, se ha publicado la obtención de partículas fágicas en las que
todas sus proteínas p3 [5] y p8 [40] están modificadas por la fusión con péptidos
o proteínas heterólogas y otras en las cuales se produce una expresión concomitante de la molécula recombinante y la natural [41]. La primera de estas
variantes ha sido utilizada principalmente para p3, cuyas funciones biológicas,
no se ven afectadas significativamente por las fusiones a su extremo NH2 terminal. Sin embargo la inserción de péptidos de más de 10 aminoácidos en
todas las copias de p8 ha sido un fracaso debido a que no permite el ensamblaje
de la cápsida [42].
Para cumplir los objetivos planteados se decidió realizar la segunda variante, la
cual se puede lograr con un sistema de fagómidos, en los cuales el producto
final es la formación de partículas con una población mixta de proteínas fusionadas y sin modificar. En este trabajo fue necesario construir un vector fagómido
que tuviera información para los genes III y VIII del fago filamentoso M13 (o
parte de ellos), que permitan la expresión de proteínas de fusión en p3 y p8
respectivamente.
El nuevo vector (Figura 17.1) se desarrolló basado en el plásmido pFabIT, originalmente destinado a la expresión de fragmentos Fab fusionados a p3 [43].
Este vector contiene el promotor del operón lactosa (plac), el cual es regulado
por la presencia de su represor, codificado por el gen lacIq e inducido por lactosa
o un análogo como el isopropil-β-D-tiogalactopiranósido (IPTG). Además este
vector posee dos secuencias nucleotídicas que codifican cada una para la secuencia señal de la pectato-liasa B (pelB), lo que promueve la secreción de la
proteína al periplasma bacteriano. Contiene además, la información genética
para el dominio N3 del gen III del fago, un origen de replicación del fago f1 y el
gen que codifica para la b-lactamasa. Esta proteína es capaz de degragar la
ampicillina, por lo que las bacterias transformadas con este plásmido crecerán
en presencia de este antibiótico.
Se determinó realizar la clonación de dos proteínas con capacidad de unir
eritrocitos, para expresarlas fusionadas a la proteína p3 en la superficie del fago
M13. Una es la adhesina bacteriana PapG y la otra un fragmento de anticuerpo
de tipo scFv específico contra la glicoforina A humana (scFv aGFA).
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU EMPLEO EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
297
Figura 17.1. Representación esquemática de la construcción del vector pFACIB3/8 y la estrategia de clonación de los sitios de unión a eritrocitos y antígenos.
En el laboratorio de anticuerpos recombinantes de la división de Inmunotecnología
y Diagnósticos del Centro de Ingeniería Genética y biotecnología (CIGB) de La
Habana Cuba se amplificó la adhesina bacteriana y se clonó en un plásmido de
expresión en E. coli denominado pPACIB9+/PapG.9.12 (PCT/CU98/0005).
La otra molécula seleccionada fue un fragmento de anticuerpo de simple cadena (scFv) específico contra el marcador de los eritrocitos humanos, glicoforina
A (GFA). Este tipo de fragmento ha sido ampliamente explotado en los últimos
años, así como su exposición en la superficie de fagos filamentosos. En el laboratorio de anticuerpos recombinantes de la división de Inmunotecnología y Diagnósticos del CIGB, se obtuvo un fragmento de simple cadena a partir de un
anticuerpo monoclonal (AcM) específico contra la GFA, capaz de reconocer
tanto la GFA M como la N [44].
Las bandas de ADN fueron amplificadas y clonadas como se observa en la
Figura 17.1 y su clonación se evaluó por amplificación por la reacción en cadena
de polimerasa (RCP) con los oligonucleótidos específicos detallados en los materiales y métodos y por digestión enzimática con las enzimas de restricción
utilizadas para la clonación (ver Figura 17.2).
Dos de los clones recombinantes de cada construcción fueron seleccionados
para secuenciarlos según el método de Sanger, con el empleo de oligonucleótidos
diseñados con este fin que hibridan externamente a las regiones de clonación
298
CAPÍTULO 17
del pFACIB3/8. Las secuencias obtenidas para los dos genes coinciden con las
secuencias publicadas para el gen que codifica para la PapG [45] y para el
scFv aGFA[44].
Figura 17.2. Análisis de restricción de los plásmidos con los SUE.
Carrilera 1-3: Clones seleccionados de pFACIB3/8+PapG, carrilera 4: Patrón de
peso molecular 1kb (Gibco, BRL), Carrilera 5: pFACIB 3/8 lineal, carrilera 6-8:
Clones seleccionados de pFACIB3/8+scFv
Con el objetivo de estudiar la expresión y el ensamblaje de las proteínas de
fusión, se realizaron dos experimentos en paralelo. Se seleccionaron células de
E.coli que fueran capaces de expresar el pilus sexual o pili F’ de conjugación, lo
cual las hace susceptibles a ser infectadas por fagos filamentosos. En esta
investigación se trabajó con la cepa Top 10F’ y se transformaron con los clones
seleccionados de las construcciones PapG-p3 y scFv-p3.
Se realizó un Western blot con muestras de biomasa y partículas fágicas de
cada clon y se utilizó un AcM específico contra el dominio N3 de la p3 de
M13. En la Figura 17.3 se observa el resultado de este experimento. Tanto en
los clones que portaban la información para la proteína PapG-p3 (carrileras
1-4) como para los que contenían la de scFv-p3 (carrileras 6-9) se encontró el
reconocimiento específico de dos bandas, una correspondiente a la p3 nativa
y otra para la proteína de fusión. Se trabajó con el M13KO7 como control
positivo (carrilera 10). Los pesos moleculares observados no coinciden con
las tallas esperadas por secuencia, pero es conocido que debido al alto contenido de glicinas en los espaciadores entre los tres dominios de la p3, la migración es retardada hasta 20 kD (talla esperada 43 kD, talla observada 62 kD),
por lo que sólo se puede establecer una comparación entre las tallas de las
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU EMPLEO EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
299
proteínas, en la cual la PapG-p3>p3 nativa>scFv-p3. Como se puede apreciar, este es el patrón de migración encontrado en este ensayo lo que demostró la expresión y el ensamblaje de ambas proteínas de fusión en la bacteria
y en las partículas virales.
Figura 17.3. Análisis de reconocimiento por Western Blot de la proteína p3.
Carrileras 1 y 2 p3+PapG (Biomasa)
Carrileras 3 y 4 p3+PapG (Fagos)
Carrilera 5 Patrón de Peso Molecular
Carrileras 6 y 7 p3+scFv (Biomasa)
Carrileras 8 y 9 p3+scFv (Fagos)
Carrilera 10 M13KO7
Se escogió p8 para fusionarle la proteína o el péptido antigénico porque se
conoce que es una proteína estructural que se encuentra en la cápsida de los
bacteriófagos filamentosos en un elevado número de copias, a diferencia del
resto, que se encuentran en un número de 3 a 5 copias. No todas las p8 portarán la molécula inmunogénica, pero de esta forma se propicia una gran representación de la misma, lo cual es muy favorable para el reconocimiento antígeno/
anticuerpo. Esto permitiría que se forme un enrejado y aglutinen los eritrocitos
una vez que los mismos se unan al SUE-p3.
Se trabajó con dos variantes antigénicas para expresarlas fusionadas a la proteína p8 del fago M13:
1. La proteína TmpA, 42 kD.
2. Un péptido inmunodominante de la gp41 del VIH 1 (16 aminoácidos).
300
CAPÍTULO 17
Para ello se amplificó el gen TmpA a partir del vector pILTmpA donde había
sido clonado, según información de trabajos anteriores [34], con el empleo de la
RCP según el programa AcR55 y con oligonucleótidos (2979/2980), diseñados
con este fin, que aportan los sitios EcoRI/BamHI requeridos para la clonación.
Para la inserción de la secuencia codificante para los 16 aminoácidos de la
gp41 en el gen VIII, se diseñaron oligonucleótidos sintéticos, complementarios,
de 53 y 56 pb (péptido VIH-1 5’ y péptido VIH-1 3’, respectivamente).
Una vez preparados ambos segmentos de ADN y los vectores pFACIB3/8+PapG
y pFACIB3/8+scFv antiGFA digeridos con las enzimas EcoRI/BamHI se realizaron las reacciones de ligamiento para cuatro construcciones diferentes:
1⇒gp41+pFACIB3/8+PapG
2⇒gp41+pFACIB3/8+scFv antiGFA
3⇒TmpA+pFACIB3/8+PapG
4⇒TmpA+pFACIB3/8+scFv antiGFA
Se realizó una purificación preparativa del ADN del plásmido de los cuatro
clones y ese material se secuenció según el método de Sanger y con
oligonucleótidos previamente diseñados con este fin que hibridan externamente
los sitios de clonación. Las secuencias obtenidas coinciden con las esperadas
para el péptido de 16 aminoácidos de la gp41 y para el gen TmpA.
El chequeo de la expresión de las moléculas antigénicas fusionadas a p8 se
realizó por Western blot. Para ello se incubaron las muestras transferidas, correspondientes a la biomasa y los fagos de los clones 5, 7, 8 y 9 con un suero
policlonal anti-p41 obtenido en conejo y de los clones 1,2,3 y 4 con un AcM antiTmpA (CB.TMPA1) obtenido en ratón.
Tanto las muestras de biomasa como las partículas virales de todos los clones
mostraron la señal de reconocimiento lo que corroboró que se expresaron y
ensamblaron correctamente las moléculas antigénicas. Para el péptido de la
gp41 en los clones 5,7,8,9 en la Figura 17.4 se observa el reconocimiento de una
banda que migra por debajo de la banda correspondiente a 14 kD del patrón,
correspondiéndose con la talla esperada (aproximadamente 10 kD) de la proteína de fusión empleada por los invesatigadores. En este experimento se usaron
como controles negativos el M13KO7 y la biomasa obtenida a partir de un
cultivo de E. coli Top10 F’y como control positivo la gp41 recombinante.
En la Figura 17.5 se observan dos bandas de reconocimiento de la TmpA para
los clones 1, 2, 3 y 4. Las masas moleculares observadas son de 41 y 43 KD,
aproximadamente. Este resultado podría explicarse porque la secuencia
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU EMPLEO EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
301
nucleotídica usada en la clonación de la misma contenía información codificante
para el péptido señal de esta proteína de membrana externa y esta se clonó en
el pFACIB3/8 que también posee una secuencia señal. De modo que debido al
procesamiento post-traduccional se obtienen proteínas en las cuales los dos
péptidos señales han sido escindidos por la peptidasa señal una vez que la misma es translocada al periplasma y otras en las que sólo uno de ellos fue cortado
de modo que las mismas tienen diferencias en su masa molecular. Los controles
usados fueron biomasa de E.coli Top10 F’ y el fago auxiliador M13KO7 como
controles negativos y la proteína TmpA recombinante como control positivo.
Figura 17.4. Análisis de reconocimiento por Western Blot del antígeno del VIH1.
Carrilera 1: VIH1-papG clon 5, biomasa; Carrilera 2: VIH1-papG clon 7, biomasa,
Carrilera 3: VIH1-scFv clon 8, biomasa; Carrilera 4: VIH1-scFv clon 9, biomasa;
Carrilera 5: VIH1-papG clon 5, fagos; Carrilera 6: VIH1-papG clon 7, fagos;
Carrilera 7: VIH1-scFv clon 8, fagos; Carrilera 8: VIH1-scFv clon 9, fagos; Carrilera
9: M13KO7; Carrilera 10: Top 10 F’ biomasa; Carrilera 11: p41 de VIH1.
Para verificar los objetivos del trabajo, se estudió la capacidad de aglutinación
específica de las partículas recombinantes.
Los fagos recombinantes obtenidos como ya se ha analizado son partículas
bifuncionales. Por una parte tienen un SUE y por otra son reconocidos por
anticuerpos contra la gp41 del VIH-1 y contra la proteína TmpA de la bacteria
Treponema pallidum, causante de la sífilis. Estos deben ser capaces de aglutinar eritrocitos en presencia de los anticuerpos anteriormente mencionados.
Para estos estudios se necesitaba tener un mayor volumen de partículas fágicas
por lo que se procedió a realizar una purificación a gran escala de un clon de
302
CAPÍTULO 17
cada construcción a partir de 500ml de cultivo. A los fagos obtenidos se le realizó
un ensayo de tipo Western Blot, en el cual se verificó el reconocimiento de las
diferentes proteínas de fusion. También se titularon las partículas purificadas.
Figura 17.5. Análisis de reconocimiento por Western Blot de la expresión y ensamblaje de TmpA.
Carrilera 1: TmpA-papG clon 1, biomasa; carrilera 2: TmpA-papG clon 3, biomasa,
carrilera 3: TmpA-scFv clon 2, biomasa; carrilera 4: TmpA-scFv clon 4, biomasa;
carrilera 5: Top 10 F’ biomasa; carrilera 6: pTmpA, carrilera 7: M13KO7, carrilera
8: TmpA-papG clon 1, fagos, carrilera 9: TmpA-papG clon 3, fagos, carrilera 10:
TmpA-scFv clon 2, fagos, carrilera 11: TmpA-scFv clon 4, fagos,.
Inicialmente se evaluaron los dos clones que portaban la TmpA como antígeno
con eritrocitos de personas sanas O + mezclados con el sobrenadante de cultivo de
un hibridoma murino que produce un AcM antiTmpA y sueros de personas
infectadas con un titulo alto por VDRL. Los resultados obtenidos se muestran en
la Tabla 17.1 y como puede apreciarse los fagos del clón TmpA+pFACIB3/8+PapG
permitieron aglutinación de eritrocitos hasta una concentración de 109 ufc/ml con
el AcM antiTmpA mientras que los fagos del clón TmpA+pFACIB3/8+scFv
antiGFA sólo permitieron aglutinación a una concentración de 1010 ufc/ml. Esto
podría deberse a que el scFv aGFA ensamblado tiene menor afinidad por la GFA
que la PapG por el antígeno P. Además, como la TmpA es una proteína de 42 kD
puede encontrarse en un bajo número de copias en la partícula fágica.
Estos dos aspectos no facilitarían que se forme un enrejado lo suficientemente
grande que permita la aglutinación visual de los eritrocitos a una mayor dilución.
Basados en este resultado se determinó proseguir el estudio con el clon1.
Cuando se ensayó con sueros humanos positivos la magnitud de las aglutinaciones
se incrementó y se observaron reacciones bien definidas hasta una concentración de 109 ufc/ml en 3 minutos. Con 108 partículas también se observó aglutinación pero se necesitó más tiempo. En el suero humano positivo existe una
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU EMPLEO EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
303
mayor diversidad de anticuerpos que reconocen diferentes estructuras y secuencias en la TmpA, comparado con el único epítopo reconocido por el AcM.
Esto facilita el reconocimiento de la misma partícula viral por varias moléculas
de inmunoglobulinas que aumentarían las posibilidades de obtener una reacción
visual. Cuando se trabajó con los sueros negativos no se observó aglutinación
inespecífica a ninguna dilución de fagos.
Tabla 17.1. Ensayo de aglutinación con los fagos de los clones 1 y 2 con eritrocitos
humanos
a: Medidos en unidades formadoras de colonias por mililitro.
b: Suero humano negativo.
c: Suero humano positivo.
d: No ensayado.
Después de estudiadas las capacidades aglutinantes de los viriones con la TmpA,
se realizaron experimentos similares para los fagos que contienen la proteína de
fusión con el péptido de VIH. Primeramente, al igual que en el caso anterior, se
ensayó un sistema modelo de sangre reconstituida con eritrocitos de personas O +
y un suero policlonal anti-P41 obtenido en conejo. Luego se probaron sueros
humanos negativos y sueros humanos positivos. Los resultados obtenidos se
muestran en la Tabla 17.2 y tanto con el suero de conejo como con el suero
humano se obtuvieron muy buenas aglutinaciones (ver Figura17.6). En el ensayo
que se realizó con los sueros negativos no se observó ninguna reacción positiva a
ninguna dilución de fagos, lo que corroboró que no existe aglutinación inespecífica.
304
CAPÍTULO 17
Tabla 17.2. Ensayo de aglutinación con los fagos de los clones HIV con sangre
reconstituida
a: Suero humano negativo.
b: Suero humano positivo.
Figura 17.6. Aglutinación de eritrocitos con partículas recombinantes de los clones 7 y 8
1. Clon 7 (VIH1-papG, 1010 ufc/ml) + eritrocitos humanos O+ + suero VIH+.
2. Clon 7 (VIH1-papG, 1010 ufc/ml) + eritrocitos humanos O + + suero VIH+ (1:2.5).
3. Clon 8 (VIH1-scFv, 1010 ufc/ml) + eritrocitos humanos O+ + suero VIH+.
4. Clon 8 (VIH1-scFv, 1010 ufc/ml) + eritrocitos humanos O+ + suero VIH+(1:2.5).
5. Sin usar
6. Clon 7 (VIH1-papG, 1010 ufc/ml) + eritrocitos humanos O+.
7. Clon 8 (VIH1-scFv, 1010 ufc/ml) + eritrocitos humanos O+.
8. M13KO7 (1010 cfu/ml) + eritrocitos humanos O+ + suero VIH+
Posteriormente se realizó un ensayo preliminar con sangre directa extraída por
digito-puntura de pacientes seropositivos para VIH-1 con los fagos de los clones
VIH. En todos los casos se trabajó con una concentración de fagos de 10 10 ufc/ml.
Los resultados aparecen en laTabla 17.3. De siete pacientes evaluados, tres de
ellos dieron positivos con ambos fagos, dos de ellos dieron positivos con uno u
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU EMPLEO EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
305
otro reactivo pero no fue posible evaluarlos con ambos por insuficiente cantidad de sangre extraída; otros dos pacientes dieron negativos.
Tabla 17.3. Evaluación de la aglutinación con muestras de sangre de pacientes
de SIDA por VIH1.
Clon 7 ( PapG-p3)
a: No evaluado
En el caso de las partículas virales que contienen la papG como SUE, una
posible explicación puede estar dada por la presencia de anticuerpos específicos contra esta adhesina bacteriana que bloqueen el reconocimiento del antígeno
P. Debido a que esta proteína es característica de cepas uropatogénicas, personas con una infección urinaria crónica o reciente podrían poseer anticuerpos
neutralizantes como parte de su respuesta inmune. Sin embargo, esto no justifica los resultados con la segunda construcción, en la que la única posibilidad de
inhibición estaría dada por un fenotipo de GFA no reconocido por el anticuerpo
empleado. No obstante, es bastante improbable que ambas se den en el mismo
individuo en una frecuencia tan alta como la encontrada (2/7). Es probable que
la respuesta esté en el título serológico contra esta proteína en particular. Esta
hipótesis deberá ser comprobada experimentalmente en trabajos futuros.
Estos resultados iniciales son favorables, si se tiene en cuenta la novedad de la
tecnología y la originalidad del diseño empleado.
PROCEDIMIENTOS
Oligonucleótidos o cebadores
• genVIII 5’ NheI (4045)
5’GAGAATTCTGAGCTAGCTGCCCAACCAGCGATGGCCGCTGAGGGTGAATTC
• genVIII 3’ Asc I (4046)
5’ … TTGGCGCGCCTTATCAGCTTGCTTTCGAGGTGAATTTC …3’
306
CAPÍTULO 17
• gen PapG 5’ Sfi I (4041)
5’..CAGGCCCAGCCGGCCATGGCCGGATGGCACAATGTCATGTTTTATGC..3
• gen PapG 3’ Not I (4042)
5’…AAGGAAAAAAGCGGCCGCGGGGAAACTCAGAACCATAGTC …3’
• scFv 5’ Sfi I
5’…CAGGCCCAGCCGGCCGAGAATTCTGAGCTAGCTGCCCAACCA
GCGATGGCCGAGGTGCAGCTGTTGGAGTCT …3’
• scFv 3’ Not I (3920)
5' ...GATCC GAT CAG CAG CTG CTG GGC ATC TGG GGC TGC AGC
GGT AAA CTG TAT TGC GGGCC...
• Péptido VIH-1 5’ EcoR I
5' ...C GCA ATA CAG TTT ACC GCT GCA GCC CCA GAT GCC CAG
CAG CTG CTG ATC G...3’
• Péptido VIH-1 3’ BamH I
5’…AAGGAAAAAAGCGGCCGCTTT(TC)A(TG)(TC)TCCAGCTTGGT…3’
• gen TmpA 5’ EcoRI (2979)
5’... CGGAATTC ATG AAT GCT CAT ACG CTT GTG... 3’
• gen TmpA 3’ BamH I (2980)
5’... CGGGATC CCC TTC TTC GTT CAT AGA GG ...3’
• 5’ SalI (4683)
5’..CGACGCGTCGACATGAAATACCTATTGCCTACGGCAGCCGCTG
GATTGTTATTGCTAGCTGCC..3’
• 3’HindIII(4684)
5’..AGGGCCCAAGCTTTTAAGACTCCTTATTACGCAGTATGTTAGCA
AACGTCGAA..3’
• 5’pLac (1083)
5’.. AGCGGATAACAATTTCACACAGGA..3’
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU EMPLEO EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
307
1. CONSTRUCCIÓN DEL VECTOR FAGÓMIDO PFACIB3/8
1.1. PURIFICACIÓN PREPARATIVA DEL VECTOR (PLÁSMIDO), A EMPLEAR
La purificación de ADN se realizó usando columnas del kit Wizard Plus Midipreps
DNA Purification System (Promega, Madison, EE.UU.) de acuerdo a las orientaciones de los fabricantes.
1. Se partió de un cultivo de 3ml de LBAT con glucosa 1 %, el cual se inoculó
con colonias recombinantes(células transformadas con el pFabIT), de interés y se dejó crecer a 37 ºC durante 16 h. Este cultivo se usó como inóculo
para un cutivo de 50ml de LBAT con glucosa al 1 %, que se dejó crecer
también a 37 C durante 16 h.
2. Después de este tiempo el cultivo se centrifugó a 14000 x g por 10 min
(Hitachi SCT – 15 B).
3. El precipitado fue resuspendido en 3ml de solución de resuspensión(TrisHCl 50mM, pH 7,5; EDTA 10mM y RNAsa 10mg/ml), mediante vortex.
4. Luego se lisaron las células con 3ml de solución de lísis( NaOH 0,2M y
SDS al 1 %).
5. Posteriormente la reacción se neutralizó mediante la adición de solución de
neutralización(KAc 1,32M, pH 4.8).
6. La mezcla de reacción se centrifugó a 14000 x g por 15 min a 4 ºC y al
sobrenadante se le adicionaron 10 ml de la resina Wizard Midipreps DNA
Purification Resin.
7. Todo se aplicó a la columna mediante vacío.
8. La columna se lavó con 30ml de solución de lavado (KAc 80mM, Tris-HCl,
pH 7,5, EDTA 40 µM y etanol al 55 %).
9. El ADN fue eluido con 300ml de agua o TE 1X por centrifugación durante
20 seg a 10000xg.
10. Se comprobó la purificación con un gel de agarosa al 1,2 % (p/v). Se utilizó
un patrón de peso molecular adecuado.
1.2. CLONACIÓN DEL GEN VIII DEL FAGO EN EL PFABIT
1.2.1. AMPLIFICACIÓN DEL GEN DE ÍNTERES
1. Se amplificó el gen VIII del fago a partir del vector PC89 con los
oligonucleótidos 4045 y 4046, que hibrídan en los extremos 5’ y 3’ del gen.
Estos oligonucleótidos aportan los sitios de restricción NheI y AscI para su
posterior clonación.
308
CAPÍTULO 17
2. La amplificación se realizó con la siguiente mezcla:
ADN plasmídico
5,0 µL
Tampón de RCP 10x
10,0 µL
MgCl2 (25 mM)
10,0 µL
dNTPs (5 mM)
4,0 µL
oligonucleótido 5´ 4045 (20 mM)
1,0 µL
oligonucleótido 3´ 4046 (20 mM)
1,0 µL
Agua estéril
68,0 µL
3. La mezcla se colocó en el equipo de RCP, bajo las siguientes condiciones de
programa
94 ºC: 5 min, 30 ciclosy se añadió 1,0 µL de Taq ADN Polimerasa.
94 ºC: 1min, 30 ciclos
55 ºC: 1 min, 30 ciclos
72 ºC: 1 min, 30 ciclos
72 ºC: 10 min, 30 ciclos
4. La amplificación se comprobó mediante electroforesis en gel de agarosa al
1,2 % (p/v) con un patrón de peso molecular adecuado. El tamaño aproximado de la banda fue de 150 pb.
5. Se purificó la banda de ADN a partir de un gel de agarosa de bajo punto de
fusión al 1,2 % (p/v). Se cortó el bloque de agarosa correspondiente a la
banda y se colocó en un tubo de 1,5 mL. Se le adicionó 100 µL de solución
TE 1x e incubó a 75 ºC por 10 min para fundir la agarosa. Posteriormente se
procedió a la purificación de la banda según el juego de reactivos de promega
Wizart PCR preps, para la extracción de bandas de RCP, según las orientaciones del productor.
Nota 1. Todas las mezclas de RCP se recubrieron con parafina líquida.
Nota 2. Se evitó cortar agarosa en exceso porque disminuye el rendimiento del
ADN que se logra extraer.
Nota 3. Reactivos
Tampón 10x de RCP: KCl 500 mM, Tris-HCl100 mM pH=8,0
Taq DNA Polimerasa: 5 U/µL (Promega).
dNTPs (5 mM): 5 mM de cada nucleótido dATP, dCTP, dGTP y dTTP, a partir
de la solución 100 mM de Boehringer Mannheim, diluido con H2 O estéril.
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU EMPLEO EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
309
1.2.2. DIGESTIÓN Y REACCIÓN DE LIGAMIENTO
1. El segmento de ADN amplificado fue digerido conjuntamente con el
vector(pFabIT), con las enzimas NheI y AscI en presencia de soluciones
tampones de New England Biolabs y a las temperaturas recomendadas, de
la manera siguiente:
Se preparó el ADN a 50 ng/µL y la siguiente mezcla de reacción:
2,0 µg de ADN
40,0 µL
Tampón 10x NEB A
10,0 µL
BSA (1 mg/mL)
10,0 µL
Nhe I (50 U)
2,5 µL
Asc I (50 U)
5,0 µL
Agua estéril
32,5 µL
Se incubó a 37 ºC durante 2 h.
2. La banda y el vector digeridos se purificaron con el juego de reactivos de
Promega Wizart DNA clean up, según las orientaciones de los fabricantes,
para preparar a ambos para la reacción de ligamiento.
3. Para la reacción de ligamiento, se mezcló en un tubo de 1,5 mL el vector y la
banda digeridos.
Vector—NheI/AscI (50 ng/µL)
6,0 µL
Banda — NheI/AscI (50 ng/µL)
6,0 µL
Tampón 5x T4 ligasa
10,0 µL
Agua estéril
26,0 µL
T4 DNA ligasa (2 U Weiss)
2,0 µL
4. Se incubó a 16 ºC de 12-18 h y se purificó el producto de la reacción de igual
forma que en el paso 5.
1.2.3. T RANSFORMACIÓN Y CHEQUEO DE RECOMBINANTES
1. Posteriormente se procedió a la preparación y transformación de las células
de E.coli pertenecientes a la cepa Top 10 F’ con el plásmido resultante.
2. La preparación de las células consistió en preparar un cultivo de 3 ml en fase
estacionaria de E. Coli, el cual se empleó como inóculo para un volumen de
50 ml de medio LB.
310
CAPÍTULO 17
3. Este se colocó en zaranda a 37 ºC hasta alcanzar una DO530nm de alrededor
de 0,5 y luego se centrifugó a 3 500 rpm, durante 10 min.; el precipitado se
resuspendió en 5 ml de TFB1 frío (KAc 30 mM, MgCl2 50 mM, RbCl
100 mM, pH=5,8).
4. Posteriormente se añadieron 20 ml más de TFB1 y se incubó en hielo durante 10 min. Se centrifugó en iguales condiciones anteriores.
5. El sedimento fue resuspendido en 4 ml de TFB 2 (MOPS 10 mM, CaCl2
75 mM (2xH 2 O), RbCl 10 mM, glicerol al 15 %, pH=7,0) frío.
6. Las células se conservaron a -80 ºC.
7. Para la transformación de las células competentes se mezclaron 125 µl de
las mismas y 10 µl de ADN plasmídico (de una concentración aproximada
de 180 ng/µl).
8. Esta mezcla se incubó durante una hora en hielo y después se sometió a un
¨stress¨ térmico de 42 ºC por 2 min.
9. Se incubó nuevamente sobre hielo por 1 min. y se le añadió 1 ml de LB
líquido, y se puso durante una hora en movimiento, a 37 ºC.
10. Se sembraron 300 µl en medio sólido LB suplementado con los antibióticos
apropiados para la selección.
11. Para la selección de recombinantes se amplifica a partir de las colonias
mediante la RCP descrita en los pasos 2 y 3. El patrón de amplificación
esperado en los clones positivos es la amplificación de una banda de 150 pb.
Para el establecimiento de un vector final se debe corroborar la clonación
por secuencia de ADN.
12. El vector obtenido de esta clonación se denominó pFACIB3/8.
2. C LONACIÓN DE LOS GENES QUE CODIFICAN PARA LOS SUE
EN EL VECTOR PFACIB3/8
Se determinó realizar la clonación de dos proteínas con capacidad de unir
eritrocitos, para ser expresadas fusionadas a la proteína p3 en la superficie del
fago M13. Una es la adhesina bacteriana PapG y la otra un fragmento de
anticuerpo de tipo scFv específico contra la glicoforina A humana (scFv aGFA).
2.1. A MPLIFICACIÓN POR RCP DE LOS GENES QUE CODIFICAN
PARA LA PAPG Y PARA EL SCFV A GFA
El proceso de amplificación se realizo de igual forma que en el caso del gen
VIII, pero cambiando los oligonucleotidos, los cuales se diseñaron de acuerdo a
la secuencia registrada para cada gen en particular, y el ADN de partida.
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU EMPLEO EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
311
2.1.1. A MPLIFICACIÓN DEL GEN QUE CODIFICA PARA LA PAPG
La adhesina se amplificó por RCP a partir de una cepa de E. coli
pielonefríticas como se describe en 1.2 y 1.3 utilizando los oligonucleótidos
4041/4042 (Tabla 18.1). Estos oligonucleótidos introducen los sitios de restricción NotI y SfiI, necesarios para clonación del gen en el vector pFACIB3/8.
2.1.2. Amplificación del gen que codifica para el ScFv a GFA
Para la amplificación de este gen también se partió de un plásmido presente en
el laboratorio, (pPacib 9+/ scFv.9.11), y se realizó igualmente a través del procedimiento explicado en los pasos 1.2.1.2 y 3, con la pareja de oligonucleotidos
3920/GFA SfiI. Estos oligonucleotidos igualmente introducen los sitios de
restricción NotI y SfiI, los cuales se emplearan en la clonación del gen en el
vector pFACIB3/8, fusionados a P3.
2.2. DIGESTIÓN, REACCIÓN DE LIGAMIENTO, TRANSFORMACIÓN
Y CHEQUEO DE RECOMBINANTES
1. El proceso de digestión del vector pFACIB3/8 y de las bandas amplificadas
se realiza de manera secuencial debido a la diferencia de temperatura a las
cuales son activas.
2. El segmento de ADN amplificado fue digerido conjuntamente con el vector
a las temperaturas recomendadas, de la manera siguiente:
3. Se preparó el ADN a 50 ng/µL y se realizó la siguiente mezcla de reacción:
2.0 µg de ADN
40,0 µL
Tampón 10x NEB 3
10,0 µL
BSA (1 mg/mL)
10,0 µL
Not I (50 U)
2,5 µL
Agua estéril
32,5 µL
4. Se incubó a 37 ºC durante 2 h.
5. Este proceso se repite en las siguientes condiciones con una incubación a 50 ºC.
2,0 µg de ADN
40,0 µL
Tampón 10x NEB 2
10,0 µL
BSA (1 mg/mL)
10,0 µL
Sfi I (50 U)
2,5 µL
Agua estéril
32.5 µL
312
CAPÍTULO 17
6. La banda y el vector digeridos se purificaron con el juego de reactivos de
promega Wizart DNA clean up, según las orientaciones de los fabricantes,
para preparar a ambos para la reacción de ligamiento.
7. Para la reacción de ligamiento, se mezcló en un tubo de 1,5 mL el vector y la
banda digeridos.
Vector—Not I/Sfi I (50 ng/µL)
6,0 µL
Banda — Not I/Sfi I (50 ng/µL)
6,0 µL
Tampón 5x T4 ligasa
10,0 µL
Agua estéril
26,0 µL
T4 DNA ligasa (2 U Weiss)
2,0 µL
8. Se incubó a 16 ºC de 12-18 h y posteriormente se purificó el producto de la
reacción de igual forma que en el paso 6.
9. El chequeo de los recombinantes se realizó por digestión enzimática con las
enzimas empleadas en la clonación.
3. EVALUACIÓN DE LA EXPRESIÓN Y EL ENSAMBLAJE
DE LAS PROTEÍNAS DE ADHESIÓN EN LA PARTÍCULA VIRAL
Con el objetivo de estudiar la expresión y el ensamblaje de las proteínas de
fusión, se realizaron dos experimentos en paralelo. Se seleccionaron células de
E.coli que fueran capaces de expresar el pili F’ de conjugación, lo cual las hace
susceptibles a ser infectadas por fagos filamentosos. En este trabajo se empleó
la cepa Top 10F’ y se transformaron con los clones seleccionados de las construcciones PapG-p3 y scFv-p3.
Se transformaron las bacterias mediante la técnica de transformación de células competentes simillar al procedimiento descrito en el acápite 1.2.3 con los
clones seleccionados de cada construcción genética.
Nota 1: En este medio la tetraciclina es el antibiótico que permite la expresión del
pilis sexual, codificado por el operón tra A-Q y la adición de glucosa garantiza la
represión de la expresión basal de la proteína de fusión que, además de la posible
toxicidad que pueda manifestar su expresión para la célula [46], garantiza que no
haya expresión de p3, lo que podría provocar una retracción de los pili.
3.1. ESTUDIO DE EXPRESIÓN DE PROTEÍNAS
1. Se cultivaron cepas de E. coli transformadas con los plásmidos recombinantes
de interés hasta alcanzar una absorbancia a 530 nm de 0,5.
2. La inducción de los cultivos se realizó mediante la adición de 1 mM de IPTG,
permitiendo el crecimiento por, al menos, 6 horas.
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU EMPLEO EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
313
3. Se centrifugaron cantidades equivalentes de cada cultivo a 12 000 rpm
durante 3 min.
4. Al sobrenadante se le añadió TCA al 10 % y se incubó 30 min en hielo, el
precipitado se lavó un par de veces con etanol al 70 %.
5. El precipitado celular se resuspendió en 100 µl de solución de sacarosa al
20 % fría y de Tris/HCl 10 mM, pH=7,5 y se adicionaron inmediatamente
5 µl de EDTA 0,5 M, pH=8,0, incubándose en hielo 10 min..
6. Posteriormente se centrifugó a 10 000 rpm por 5 min, se resuspendió el
precipitado en agua destilada fría estéril, incubándose nuevamente 10 min
en hielo.
7. Se volvió a centrifugar en las mismas condiciones anteriores y se conservó
el sobrenadante como la fracción periplasmática y el precipitado como la
fracción intracelular.
8. Las muestras se mezclaron con 50 µl de solución tampón 2x (625 ml Tris 2
M, 2 ml SDS al 20 %, 1 ml de 2-mercaptoetanol, 2 ml de glicerol, 1 mg de
bromofenol azul) y se incubaron durante 20 min a 105 ºC.
9. A cada fracción se le realizó electroforesis en geles de poliacrilamida al 15 %
en presencia de SDS.
3.2. E XPRESIÓN DE LAS PROTEÍNAS RECOMBINANTES EN LA SUPERFICIE
DEL FAGO FILAMENTOSO
3.2.1. Preparación del ¨stock¨ de fagos auxiliadores
Para comprobar el ensamblaje de las proteínas recombinantes en la superficie
del fago se realizaron los siguientes procedimientos:
1. A un cultivo de células permisivas (capaces de ser infectadas por M13) (Top
10 F’ en este caso) en fase exponencial, se le adicionaron fagos a una concentración de 10 unidades formadoras de colonias (ufc) por bacteria.
2. A los 15 minutos se sembraron 15 ul de esta suspensión celular en placas LB
que contienen 20 ug/ml de kanamicina y se dejaron crecer por un tiempo de
16 horas.
3. Posteriormente se inoculó una colonia en 3 ml de cultivo suplementado con
la misma cantidad del antibiótico de selección y se dejó crecer con agitación
por seis horas a 37 ºC.
4. Este cultivo de 3 ml, a su vez, se utilizó como inóculo para un frasco de 300 ml
de LB con kanamicina que se dejó crecer con una agitación de 250 rpm
durante 10 horas a 37 ºC.
314
CAPÍTULO 17
5. Luego, el cultivo se centrifugó a 4300 rpm durante 20 minutos.
6. El sobrenadante de esta centrifugación se filtró por una membrana de poros
de 0,2 um y se conservó en alícuotas de 2 ml a -80ºC.
3.2.2. Titulación de fagos por unidades formadoras de colonias
1. Se preparó un cultivo de células permisivas (Top 10 F’) en fase exponencial
de crecimiento a 37 ºC.
2. Los sobrenadantes de cultivos que contienen fagos se diluyeron de forma
seriada en medio de cultivo LB y 10 ul de estas diluciones se mezclaron con
90 ul del cultivo de células indicadoras.
3. Se dejaron infectar durante 15 minutos a temperatura ambiente sin agitación
y luego se le adicionaron 100 ml de LB con tetraciclina y se incubó durante
15 minutos.
4. Posteriormente se sembraron en placas de medio sólido suplementado con
ampicillina y tetraciclina y se dejaron crecer por un tiempo de 16 h a 37 ºC.
5. Al cabo de este tiempo las colonias se contaron y el número de colonias
obtenidas en la máxima dilución fue el usado en el cálculo del título del fago.
6. Teniendo en cuenta los factores de dilución la expresión general sería :
C x 10n x FD x 103 [ufc/ml]
3
donde:
C: número de colonias contadas en la mayor dilución a la que se observa
crecimiento.
n: orden de la mayor dilución a la que se observa crecimiento.
FD: factor de dilución (siguiendo el procedimiento antes mencionado este
valor es 13).
3.2.3. Ensamblaje de las proteínas de interés en la superficie
del fago y purificación de los mismos
1. A partir de un cultivo de 3ml de LBAT y glucosa 1 % de E coli Top 10 F’
transformadas con el fagómido de interés (pFACIB3/8/PapG o pFACIB3/8/
ScFv), se inoculó un cultivo de 50 ml de 2xYT suplementado con tetraciclina
(20 µg/ml), ampicillina (50 µg/ml) y glucosa al 1 % , se dejó crecer a 37 ºC
con agitación hasta que alcanzó una DO 5 3 0 n m entre 0,5 y 0,7 (fase
exponencial).
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU EMPLEO EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
315
2. En estas condiciones se infectó con el fago auxiliador M13KO7, se adicionaron 10 partículas virales por célula (asumiendo una concentración
de 6,3 x 108 células por ml a esta densidad celular) y se dejó el cultivo en
agitación durante 30 min.
3. Luego se centrifugó el cultivo en tubos estériles a 3500 rpm durante 15 min.
4. El precipitado fue resuspendido en 500 ml de 2xYT con tetraciclina y
ampicillina a las concentraciones referidas anteriormente, además se le adicionó kanamicina (20 µg/ml) y el inductor IPTG (1mM), se dejó creciendo
durante 16 h a 37 ºC con agitación.
5. Pasado ese tiempo el cultivo se centrifugó a 2000 rpm por 20 min a 4 ºC.
6. Al sobrenadante se le adicionó el 25% del volumen total de solución de
precipitación (polietilenglicol 8 000 al 15% en 3,0 M de NaCl), incubándose
en hielo a 4°C durante 1 hcomo mínimo.
7. Luego se centrifugó a 6000 rpm durante 20 min.
8. El pellet fue resuspendido en 10 ml de TE y se procedió a una segunda
precipitación de los fagos en iguales condiciones que la primera precipitación.
9. Posteriormente se centrifugó a 10000 rpm por 10 min a 4 °C.
10. Los fagos que se encontraban en el precipitado fueron resuspendidos en
1ml de TE.
11. Opcionalmente se puede incubar la muestra a 70 °C por 10 min y luego
centrifugar a 12000 rpm durante 3 min. Se conserva el sobrenadante donde
se encuentran los fagos.
3.3 I NMUNODETECCIÓN DE LA EXPRESIÓN Y ENSAMBLAJE
DE LAS PROTEÍNAS DE INTERÉS
1. Se realizó la transferencia de muestras proteicas separadas en geles de
poliacrilamida hacia membranas de nitrocelulosa con un equipo de transferencia semi-seco (BioRad, EUA).
2. Las membranas fueron teñidas con rojo Ponceau para comprobar la transferencia proteíca y posteriormente desteñidas con agua.
3. Se bloqueó la membrana con una solución de leche al 2 % en PBS (Na2 HPO 4
10 mM, >NaCl 137 mM, KH2 PO4 1,7 mM, KCl 2,6 mM, pH=7,4) durante
una hora a temperatura ambiente.
316
CAPÍTULO 17
4. Se lavó tres veces con de Tween 20 al 0,05 % en PBS. Se incubó con el
antisuero apropiado (anti p3) previamente liberado de anticuerpos específicos contra proteínas de E. coli.
5. Se repitieron los lavados y se añadió un anti-conejo obtenido en carnero,
conjugado a peroxidasa (Sigma, EUA), diluido 1:5000 en PBS.
6. Se incubó por una hora a temperatura ambiente.
7. Se repitieron los lavados y se reveló con 0,5 mg/ml de diaminobenzidina
(DAB) y peróxido de hidrógeno al 0,015 %.
8. La reacción se detuvo al visualizar las bandas, por eliminación de la solución sustrato y lavado de la membrana con agua destilada. El resultado se
observa en la Figura 18.3.
4. C LONACIÓN DE LOS GENES QUE CODIFICAN
PARA LOS POLIPÉPTIDOS ANTIGÉNICOS EN EL VECT OR
PFACIB3/8, UNIDOS A
P8
4.1. P REPARACIÓN DEL GEN DE LA TMPA, 42 KDA,
PARA SU CLONACIÓN POSTERIOR
Para la clonación de este gen, el mismo se amplifico a partir del vector
pILTmpA donde había sido clonado en trabajos anteriores [34], con el empleo
de la RCP según los pasos 1.2.1.2 y 3 y usando oligonucleótidos (2979/2980)
diseñados con este fin, que aportan los sitios EcoRI/BamHI requeridos para
la clonación.
Las digestiones se realizaron igual a las publicadas. Se tuvo en cuenta la diferencia en las enzimas y los tampones de reacción.
4.2. PREPARACIÓN DE UN PÉPTIDO INMUNODOMINANTE DE LA GP41
DEL VIH 1 (16 AMINOÁCIDOS), PARA SU CLONACIÓN POSTERIOR
Para la inserción de la secuencia codificante para los 16 aminoácidos de la
gp41 en el gen VIII, se diseñaron oligonucleótidos sintéticos, complementarios, de 53 y 56 pb (péptido VIH-1 5’ y péptido VIH-1 3’, respectivamente).
Estos fueron desnaturalizados, hibridados a 70 °C y posteriormente fosforilados
sus extremos con el empleo de la PNK. De esta forma se construyó un adaptador sintético de doble cadena, listo para la clonación.
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU EMPLEO EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
317
4.3. E STRATEGIAS DE CLONACIÓN DE LOS POLIPÉPTIDOS
ANTIGÉNICOS OBTENIDOS
Una vez preparados ambos segmentos de ADN y los vectores pFACIB3/8+PapG
y pFACIB3/8+scFv aGFA digeridos con las enzimas EcoRI/BamHI se realizaron las reacciones de ligamiento de igual manera a la descrita anteriormente,
para cuatro construcciones diferentes:
1 ⇒ gp41+pFACIB3/8+PapG
2 ⇒ gp41+pFACIB3/8+scFv antiGFA
3 ⇒ TmpA+pFACIB3/8+PapG
4 ⇒ TmpA+pFACIB3/8+scFv antiGFA
4.4. CHEQUEO DE RECOMBINANTES
La búsqueda de recombinantes se realizó posterior a la transformación de la
cepa Top10F’ con las cuatro combinaciones por separado, a través de la digestión de los ADN plasmídicos purificados a partir de 12 colonias de cada construcción.
1. Para la clonación del péptido de VIH-1 el chequeo de recombinantes se
realizó por digestión con las enzimas NheI/AscI.
2. El resultado de la digestión se visualizó en una electroforesis de agarosa
normal 1,2 %.
3. Para la clonación de la TmpA en cada vector los resultados se evaluaron de
la misma forma, pero se utilizaron en este caso las enzimas de restricción
que se emplearon para la clonación del gen treponémico.
5. E VALUACIÓN DE LA EXPRESIÓN Y EL ENSAMBLAJE DE LOS PÉPTIDOS
ANTIGÉNICOS EN LA PARTÍCULA VIRAL
La expresión y ensamblaje de estos péptidos en la superficie del fago se realizaron de igual forma que en el caso de los péptidos de adhesión, teniendo en
cuenta las cuatro combinaciones de ensamblaje
(1⇒gp41+pFACIB3/8+PapG; 2⇒gp41+pFACIB3/8+scFvantiGFA ),
(3⇒TmpA+pFACIB3/8+PapG; 4⇒TmpA+pFACIB3/8+scFv antiGFA),
El chequeo del ensamblaje y la expresión se realizaron por western blot de igual
forma a la descrita en los pasos 3.3, con la diferencia en el empleo de los
anticuerpos, ya que en este caso se emplearon: un suero policlonal anti-p41
318
CAPÍTULO 17
obtenido en conejo para el caso de las combinaciones 1 y 2 (resultado Figura
18.5), y un AcM -antiTmpA (CB.TMPA1) obtenido en ratón para las combinaciones 3 y 4, respectivamente.
6.E STUDIO DE LAS PROPIEDADES AGLUTINANTES DE LOS BACTERIÓFAGOS
RECOMBINANTES OBTENIDOS
Previo a la determinación de la capacidad aglutinante de los fagos recombinantes
obtenidos, se procedió a la producción, purificación y titulación a gran escala
según lo descrito en los pasos 3.2.1 y 3.2.2, de un clon por cada una de las
cuatro combinaciones obtenidas. A dichos fagos se le realizó un Western Blot
para corroborar la presencia tanto de las adhesinas como de los péptidos
antigénicos en la superficie del fago.
6.1. E NSAYO DE AGLUTINACIÓN CON ERITROCITOS HUMANOS
1. Las partículas fágicas recombinantes purificadas, se ensayaron para su capacidad de aglutinar eritrocitos humanos.
2. Se prepararon diluciones seriadas de orden 10 en PBS (a partir de una concentración inicial de 1010 ufc/ml), en los pozos de placas de microtitulación, y
a un volumen final de 50 ul por pozo, de cada una de las combinaciones.
3. Se le adicionaron entonces a cada pozo 50 ul de una suspensión 1 % (vol/vol)
de eritrocitos humanos O+, mezclados con antisueros obtenidos en conejo
específicos para cada antígeno, con un antisuero humano conteniendo
anticuerpos específicos para el péptido de VIH-1, o para la TmpA, y sueros
no relacionados.
4. Las placas se incubaron entre 1 y 5 min a temperatura ambiente y luego se
evaluaron de forma visual.
6.2. E NSAYO DE AGLUTINACIÓN USANDO DIRECTAMENTE
SANGRE HUMANA
1. Se prepararon diluciones seriadas de orden 10 en PBS (a partir de una concentración inicial de 1010 ufc/ml), en los pozos de placas de microtitulación, y
a un volumen final de 50 ul por pozo de los fagos recombinantes 1 y 2.
2. Posteriormente, se adicionaron 50 ul de una dilución de sangre fresca proveniente de una extracción del dedo índice de la mano izquierda de individuos seropositivos para VIH-1, en cada pozo.
3. Las placas se incubaron entre 1 y 5 min a temperatura ambiente y se evaluaron de forma visual.
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU EMPLEO EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
319
REFERENCIAS
1. Smith GP. Filamentous fusion phage: Novel expression vectors that display cloned antigens
on the virion surface. Science 1985;228:1315-17.
2. McCafferty J, Griffiths AD, Winter G, Chiswell D J. Phage antibodies; filamentous phage
displaying antibody variable domains. Nature 1990;348:552-4.
3. Markland W, Roberts BL, Saxena MJ, Guterman S K, Ladner RC. Design, construction and
function of multicopy display vector using fusions to the major coat protein of bacteriophage
M13. Gene 1991;109:13-9.
4. Dueñas M, Borrebaeck CAK. Novel helper phage design: intergenis region affects the assembly
of bacteriophages and size of antibody libraries. FEMS, 1995;125:317-22.
5. Dueñas M, Borrebaeck CAK. Clonal selection and amplification of phage displayed antibodies
by linking antigenic recognition and phage replication. Bio/Technology 1994;12:999-1002.
6. Krebber C, Spada S, Desplancq D, Krebber A., Ge L, Pluckthun A. Selectively-infective
phage (SIP): A mechanistic dissection of a novel in vivo selection for protein-ligand interactions.
J Mol Biol 1997;268:607-18.
7. McCafferty J. Phage display of peptides and proteins. Academic Press1996;
8. Huang W, Petrosino J, Palzkill T. 1998. Display of functional beta-lactamase inhibitory
protein on the surface of M13 bacteriphage. Antimicrob agents Chemother1998;42:2893-7.
9. Prezzi C, Nuzzo M, Meola A., Delmastro P, Galfre G, Cortese R, et al. Selection of antigenic
and immunogenic mimics of hepatitis C virus using sera from patients. J of Immunology
1996;156:4504-13.
10. Folgori A, Tafi R, Felici F,Galfre G, Cortese R, Monaci P, et al. A general strategy to identify
mimotopoes of patological antigens using only random peptides libraries and human sera.
EMBO J 1994;13:2236-43.
11. Willis A, Perham R, Wraith D. Immunological properties of foreign peptides in multiple
display on a filamentous bacteriophages. Gene 1993;128:79-83.
12. Tsunetsugu-YokotaY, Tatsumi M, Robert V, Devaus C, Sprite B, Chermann J, et al. Expression
of an immunogenic region of HIV by a filamentous bacteriophage vector. Gene1991;99:323-6.
13. Minenkova O, Ilyichev A, Kishchenko G, Petrenko V. Design of specific immunogens using
filamentous phage as the carrier. Gene 1993;128:85-8.
14. Veronese FDM, Willis AE, Boyer-Thompson C, Appella E, Perham RN. Structural mimic ry
and enhanced immunogenecity of peptide epitopes displayed on filamentous bacteriophage.
J Mol Biol 1994;243:167-72.
15. Ward JW, Grindon AJ, Feorino PM. Laboratory and epidemiologic evaluation of an enzyme
inmunoassay for antibodies to HTLV-III. JAMA 1986;256:357-61.
16. Robey WG, Safai B, Oroszlan S, Arthur LO, Gonda MA. Characterization of envelope and
core structural gene products of HTLV-III with sera from AIDS patients. Science
1985;228:593-5.
17. Binley J, Klasse P, Cao Y, Jones I, Markowitz M, Ho D, et al.. Diferential regulation of the
antibody responses to gag and env proteins of HIV-1. J Virol 1997;71(4):2799-809.
320
CAPÍTULO 17
18. Francis HL, Kabeya M, Kaufuama N, Riggins C, Calebanders R. Comparison of sensitivities
and specificities of latex agglutination and an enzyme-linked inmunoabsorbent assay for
detection of antibodies to the human inmunodeficiency virus in African sera. J Clin Microbiol
1998;26:2462-4.
19. Quinn TC, Riggin CH, Kline RL, Francis H, Mulenga K. Rapid latex agglutination assay
using recombinant envelope polypeptide for the detection of antibody TO THE hiv. J Am
Med Assoc 1988;260:510-3.
20. Vercauteren G, Beelaert G, van der Groen G. Evaluation of an agglutination HIV-1+ 2 antibody
assay. J of Virol. Methods 1995;51:1-8.
21. Sell S, Norris SJ. The biology, pathology and immunology of syphilis. Int Rev Exp Pathol
1983;24:204-76.
22. Brandt AM. A social history of venereal disease in the United States since 1880. New York:
Oxford University Press; 1987.
23. Schroeter AL, Lucas JB, Price EV, Falcone VH. Treatment of early syphilis and reactivity of
serologic tests. JAMA 1972;221:471-6.
24. PerinePL. Syphilis and the endemic treponematoses. In: Strickland TG, editor. Hunters
tropical medicine, 6th ed. Philadelphia: WB Saunders Company;1984.p.247-56
25. Hook EW. Syphilis and HIV infection. J Infect Dis 1989;160:530-4.
26. Duncan WC. Failure of erythromycin to cure secondary syphilis in a patient with the human
immunodeficiency virus. Arch Dermatol 1989;125:82-4.
27. Farley A. The syphilis epidemic in Connecticut: relationship to drug use and prostitution.
Sex Transm Dis 1990;17:163-8.
28. Schroeter AL, Turner RH, Lucas JB, Brown W J. Therapy for incubating syphilis; effectiveness
of gonorrhea treatment. JAMA 1971; 218:711-3.
29. Schober PC, Gabriel G, White P, Felton WF, Thin RN. How infectious is syphilis?. J Vener
Dis 1983;59:217-9.
30. Wicher V, Zabek J, Wicher K. Pathogen-specific humoral response in Treponema palliduminfected humans, rabbits and guinea pigs. J Infect Dis 1991;163:830-6.
31. Baker- Zander SA, Roddy RA, Handsfield HH, Lukehart SA. IgG and IgM antibody reactivity to
antigens of treponema pallidum after of treatment of syphilis. Sex Transm Dis 1986;13:214-20.
32. Miranda A., Benítez J, Acevedo B, Domínguez MC, Villareal A, Palenzuela D, et al. A
comparison of VDRL and immunoassays developed with a recombinant TmpA antigen in
the screening of antibodies to Treponema pallidum. Ser and Immunoth. In infectious disease
1996;8:149-55.
33. Korhonen TK, Vaisanen V, Saxén H, Hultberg H, Svenson SB. P-antigen recognizing fimbriae
from human uropathogenic E. coli strains. Inf And Imm 1982;37:286-91.
34. Race, Sanger R. Blood groups in man, 6th ed. Blackwell: Oxford Press; 1975.
35. Brooks DE, Cavanagh J, Jayroe D, Janzen J, Snoek R, Trust T. Involvement of the MN
blood group antigen in shear-enhanced hemagglutination induced by the E. coli F41 adhesin.
Inf And Imm 1989;57:377-83.
36. Hultgren S, Lindberg F, Magnusson G, Kihlberg J, Tennent JM, Normark S. The PapG
adhesin of uropathogenic E. coli contains separate regions for receptor binding and for the
incorporation into the pilus. PNAS USA 1989;86. 4357- 61.
DISEÑO DE PARTÍCULAS FÁGICAS BIFUNCIONALES PARA SU EMPLEO EN SISTEMAS DE DIAGNÓSTICO RÁPIDO
321
37. Gaastra W, De Graf FK. Host-specifical fimbrial adhesins of noninvasive enterotoxigenics E.
coli strains. Microbiol Rev 1982; 46:129-61.
38. Kingsbury D W. Paramyxoviridae and their replication. Virology 1990;1:945-63.
39. Kallenius G, Mollby R, Svenson SB, Helin I, Hultberg H, Windberg J. Identification of
carbohydrate receptor recognized by uropathogenic E. coli. Infection (suppl.) 1980;3:288-93.
40. Felici F, Castagnoli L, Musacchio A, Jappelli R, Cesareni G. Selection of antibodies ligand
from a large library of oligopeptides expressed on a multivalentexposition vector. J Mol Biol
1991; 222:301-10.
41. Felici F, Luzzago A, Monaci P, Nicosia A, Sollazo M, Traboni C. Peptide and protein
display on the surface of filamentous bacteriophages. In: El-Gewely R, editor. Biotechnology
Anual Reviewvol 1. Amsterdam: Elsevier; 1995.p.149-83.
42. Iannolo G, Minenkova O, Petruzzelli R, Cesareni G. Modifying filamentous phage capsid:
Limits in the size of the major capsid protein. J Mol Biol 1995; 248:835-44.
43. Dueñas M, Fernández de Cossío ME, Hondares E, Díaz J, Ayala M, Bell H, et al. Displaying
erythrocyte binding sites and antigens as a diagnostic tool for
haemagglutination1999.(manuscrito en preparación).
44. Gavilondo J. Poster: “An antibody fragment-antigen recombinant fusion protein for a simple visual serologic diagnostic immunoassay of Syphilis”. Therapeutic Antibody Technology
1997;97, CA, USA .
45. Lund B, Lindberg F, Marklund B-I, Normark S. Proc Natl Acad ScUSA 1987; 84:5898-902.
46. Lama J, Carrasco L. Inducible expression of a toxic poliovirus membrane protein in E. coli:
Comparative studies using different expression systems based on T7 promoters. Bioch and
Biophy Research Communications 1992;188:972-81.