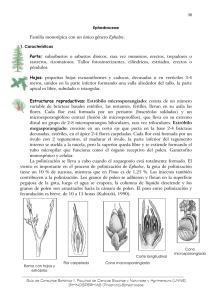
ÍNDICE 1. INTRODUCCIÓN
Anuncio

ÍNDICE 1. INTRODUCCIÓN………………………………………………..…………….…1 1.1. Hipótesis de trabajo……………………….………….…………………..……….2 1.2. Objetivo general…………………………………..…………………….………...2 1.3. Objetivos específicos…………………………….……….…………….….……..3 2. REVISIÓN BIBLIOGRÁFICA………………….…………………...………….4 2.1. Antecedentes de los cítricos………………….………………….…….…………4 2.1.1. Clasificación taxonómica de los cítricos……………………….….…………...4 2.1.2. Origen de las especies en evaluación...............................………….…………..4 2.2. Esterilidad en los cítricos……………………………..…………….……………5 2.3. Polinización en cítricos…………………………………………..………………6 2.3.1. Tipos de polinización……………………………………………………....…..8 2.3.1.1. Autopolinización…………………………………………….…………….....9 2.3.1.2. Polinización cruzada……………………………….……….………………...9 2.3.2. El polen y su transporte en los cítricos….…………………….………………10 2.4. Fecundación en cítricos………………………………………………………....11 2.5. El cuajado en cítricos…………………………………………………….……...12 2.6. Partenocárpia en cítricos…………………………………...…………………....14 2.7. La presencia de semillas en las mandarinas y su problemática………………....17 2.8. Utilización de productos con posible efecto polinicida……………….………...17 2.8.1. Aplicación de sales de cobre…………………………………………………..17 2.8.1.1. Características de las sales de cobre……………………………………..….19 2.8.2. Otros compuestos con capacidad polinicida………………………………..…20 3. MATERIALES Y MÉTODOS…………………………………………………24 3.1. Ubicación del ensayo…………………………………………………………....24 3.2. Caracterización climática y edáfica de la zona del ensayo……………………...24 3.3. Descripción del ensayo………………………………………………….……....26 3.4. Época de realización de los ensayos…………………………………………….27 3.5. Materiales utilizados…………………………………………………………….28 3.5.1. Materiales utilizados en campo……………………………………………….28 3.5.1.1. Material vegetal……………………………………………………………..28 3.5.1.2. Material instrumental………………………………………………………..29 3.5.2. Materiales utilizados en la observación de resultados………………………...29 3.6. Metodología de trabajo……………………………………………………….…30 3.7. Estados fenológicos en que se realizaron las labores…………………………...34 3.8. Determinación del término de la segunda caída fisiológica ……………...…….34 3.9. Diseño experimental……………………………………………………………36 2 4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS……………………..37 4.1. Análisis del efecto polinicida expresado en el número de semillas de cada fruto………………..…………………………………………...………...37 4.1.1. Influencia de la presencia de estambres y polen en los ensayos………….…...37 4.1.2. Influencia de los productos y fechas, y su posible efecto polinicida medido en el número de semillas….......................................…...……….…....43 4.2. Análisis del calibre de los frutos………………………………………..……….53 4.2.1. Influencia de la presencia de estambres y polen en los ensayos………….…...53 4.2.2. Influencia de los productos y las fechas de ensayo, en el calibre final del fruto………………………..………………….…………………………...56 5. CONCLUSIONES…...…………….………..…………………………………...61 6. RESUMEN………………………..…….………………………………………..62 7. ABSTRACT……………………….……….………………………..……………64 8. LITERATURA CITADA……………………...…………………….…………..65 ANEXOS 3 AGRADECIMIENTOS En primer lugar quiero agradecer a mi profesor guía don Ricardo Cautín por haberme dado la oportunidad de poder realizar este taller de licenciatura, por sus consejos, los que han sido esenciales para mi formación. A los productores por su generosa disposición para llevar a cabo los ensayos, tanto a la “Sociedad Comercial Agrícola Los Rosales de Santa Julia” como a la “Sociedad Agrícola Arancibia”. Por otra parte quiero dar una especial connotación a mis grandes amigos que forje en estos años en la Universidad, a Luís Felipe, Alfredo, Marcos, Rodrigo, Julio, Álvaro, Miguel, Claudio, Francisco, Cristián, Renán, Walter y Sebastián, con los que compartí grandes momentos, y con los que comprendí que la etapa de la Universidad no es solo aprender una disciplina para ganarse la vida, sino que además es aprender a vivir. También quiero agradecer en forma especial a personas muy especiales para mi como los son mi tío Grimaldo y mi tía Nena, por su preocupación por todos estos años en que este estuve lejos de casa, a mi primo Pablo, por su compañía y ayuda en mi última etapa de mi carrera. A mi abuelita Amanda y mi tía Carmen por todos estos años de apoyo, las tengo en el corazón, como también a mi tía Juana y mi tío Jorge. Un agradecimiento especial a Beña por su buena disposición en todos estos años de viajes, en que siempre me facilitaba las cosas, gracias. Y para el último he querido dejar a las personas más importantes de mi vida que son mis padres y mi hermana, a mis padres por su constante apoyo, comprensión, amor y 4 por haberme brindado la educación que hoy ya culmino. Gracias de todo corazón como también gracias a Dios por haberme bendecido con ustedes. A mi hermana, Ana María, por su gran apoyo y por todos los grandes momentos que hemos pasado, gracias. Y finalmente a Dios por todo, porque la fe y perseverancia son las mejores herramientas para alcanzar nuestras metas, como también pueden ser las mejores guías para los difíciles y largos senderos que nos depara la vida. “Mantén tus ojos en mi y la fe te estaré dando y veras que con paciencia podrás seguir caminando” Zaida C. de Ramón 5 1. INTRODUCCIÓN El cultivo de la mandarina es relativamente reciente en Chile (FIA, 2002). La superficie actual es de 1806 hectáreas de clementinas, localizadas principalmente en la IV región con 763 hectáreas, zona que aumentó sus plantaciones en un 6,422% (DIARIOPYME, 2005). Sigue en importancia la V región con 482 hectáreas y la región Metropolitana con 352 hectáreas, acumulando un incremento en las plantaciones a nivel nacional del 193%, según el últimos análisis del Catastro Frutícola Nacional del año 2004 (DIARIOPYME, 2005). La mayoría de las mandarinas que se cultivan corresponden al grupo de las clementinas y la principal variedad es Clemenules, exportándose el 35% de su producción a Inglaterra, Japón y Alemania. La producción debería seguir aumentando, ya que el 50% de las plantaciones están en etapa de formación, por lo tanto, no han alcanzado su máximo desarrollo (FIA, 2002). Tomando en cuenta los antecedentes anteriores y debido al auge que tiene la mandarina clementina en estos días y las perspectivas a futuro del cultivo, es necesario desarrollar nuevas técnicas para lograr mejores rendimientos y entregar al consumidor final un producto de calidad, para tener de esta forma ventajas competitivas respecto de otros productores del Hemisferio Sur. Resulta de vital importancia conocer todos los factores que afectan a la producción y calidad de la fruta del mandarino clementino. Dentro de los factores que afectan la calidad de la fruta y por ende la comercialización de las clementinas, esta la presencia de semillas en el fruto, lo cual es motivo de descarte para la comercialización del producto, ya que desvaloriza a la fruta. 6 Una alternativa que aparece para solucionar la presencia de semillas en los frutos de mandarinas clementinas, originadas por polinización cruzada con especies del grupo de híbridos, naranja común o blanca y/o limoneros, es la utilización de sales de cobre, las cuales tendrían la característica de impedir la germinación del grano de polen y/o el desarrollo del tubo polínico (AGUSTÍ, 2005)*. Esto llevaría a obtener fruta sin semillas, es decir, tendrían un efecto polinicida o la capacidad de formar frutos partenocárpicos. De ahí nace la necesidad de investigar más sobre el tema y llegar a precisar el verdadero efecto polinicida que puedan tener las sales de cobre. Este estudio se realizó en mandarina clementina cv. Clemenules polinizadas con polen del híbrido Fortune, el cual es un buen polinizante para el cv. Clemenules (SOLER, 1996), con la finalidad de entregar información sobre el verdadero efecto polinicida de las sales de cobre. 1.1. Hipótesis de trabajo: La aplicación de sales de cobre tiene efecto polinicida en la floración de mandarina clementina (Citrus clementina Hort. ex Tanaka). 1.2. Objetivo general: • Determinar la efectividad de productos de origen cúprico, en su efecto polinicida, en flores de mandarina clementina cv. Clemenules (Citrus clementina Hort. ex Tanaka). __________________________ AGUSTÍ, M. Doctor. Ing. Agr. 2005. Profesor de Citricultura y Fruticultura de la Universidad Politécnica de Valencia. Valencia, España. Comunicación personal. 7 1.3. Objetivos específicos: • Identificar entre cuatro distintos productos de origen cúprico, él o los de mayor eficiencia en cuanto a su efecto polinicida. • Determinar el momento fenológico de mejor resultado para la aplicación de los productos evaluados y lograr el efecto polinicida. • Determinar la incidencia de la polinización artificial en la formación de semillas en mandarina clementina y su efecto sobre el calibre final del fruto. 8 2. REVISIÓN BIBLIOGRÁFICA 2.1. Antecedentes de los cítricos: 2.1.1. Clasificación taxonómica de los cítricos Las especies con interés comercial de los cítricos pertenecen a la familia de las Rutaceas, subfamilia Aurantioideas. Esta se encuentra dentro de la división Embriophyta Siphonogama, subdivisión Angiospermae, clase Dicotyledonea, subclase Rosidae, superorden Rutanae, orden Rutales (AGUSTÍ, 2003). 2.1.2. Origen de las especies en evaluación El origen de las clementinas siempre ha sido controvertido. Para algunos autores (Trabut y Rebour), son descendientes de un cruzamiento casual entre el mandarino común y el naranjo amargo Granito, observado aproximadamente hace 90 años por el padre Clément en el orfelinato de Misserghine en Orange (Argelia). Para otros (Weber, Tanaka y Chapot), procede de una variedad de mandarina probablemente originada en el Extremo Oriente, ya que presenta un gran parecido con otras mandarinas, como la mandarina de Cantón que es, de hecho, también una Clementina. La cuenca mediterránea es la principal zona de producción de Clementinas; España, Marruecos y Argelia son los principales países productores (LOUSSERT, 1992). Con respecto a la clementina cv. Clemenules, según AGUSTÍ (2003), es una mutación espontánea de clementina Fina originada en Nules (Castellón) en 1953. Es tendiente a producir una floración más o menos uniforme, y posteriormente dos o tres más heterogéneas. Las hojas lanceoladas de distintos tamaños, son muy típicas de 9 clementina. El fruto es de buen calibre, de color naranja y propenso al bufado cuando alcanzan su madurez natural (SOLER, 1999). La mandarina Fortune, es un híbrido de mandarina clementina con mandarina Dancy, obtenido en la estación experimental de Indio, California, Estados Unidos (FURR, 1964). 2.2. Esterilidad en los cítricos: El término esterilidad indica la incapacidad para la reproducción mediante gametos, normalmente en la fecundación. En las plantas superiores, la esterilidad absoluta provoca normalmente una incapacidad completa para reproducirse de forma sexual, por medio de semillas. Sin embargo, en las plantas con embriones nucelares, la ausencia de semillas no requiere solamente una verdadera esterilidad generativa o sexual, sino también lo que puede llamarse como esterilidad nucelar (incapacidad para producir los embriones nucelares) (PUCHADES, 2002). Para AGUSTÍ (2004) la esterilidad puede verse inducida por causas climáticas, pero las que tienen verdadera importancia son las debidas a causas genéticas. Especial referencia merece la esterilidad citológica que afecta a los cultivares triploides. Por otra parte AGUSTÍ (2004), clasifica los tipos de esterilidad que se presentan en los árboles frutales en esterilidad gamética, la cual es una falta o deficiente desarrollo de estambres (androesterilidad), u ovarios (ginoesterilidad), la esterilidad citológica, lo que se refiere a alteraciones de la meiosis durante la gametogénesis y la esterilidad homogenética en la cual el polen no puede fecundar flores, ya sea del mismo cultivar (autoincompatibilidad), o de otro cultivar (incompatibilidad de cruce). 10 Según PUCHADES (2002) la esterilidad generativa se puede clasificar en dos: en esterilidad gamética y en aborto embrionario (embrión cigótico), y la esterilidad gamética puede ser dividida, asimismo, en esterilidad gamética relativa y esterilidad gamética absoluta. La esterilidad gamética relativa tiene relación con la incapacidad de formar semillas mediante la autopolinización, aunque si es por medio de polinización cruzada, la formación de semillas existe, ya que en estos casos, los gametos tanto femeninos como masculinos, son funcionales en polinización cruzada. Caso típico de este tipo de esterilidad son las mandarinas del grupo de las clementinas (PUCHADES, 2002). Los mandarinos clementinos, son auto incompatibles (BONO SOLER y BUJ, 2000), es decir, no producen semillas al ser polinizados con su propio polen, este sí es compatible con otras variedades como las de clementinas, produciendo numerosas semillas en sus frutos y viceversa. De los híbridos introducidos el más extendido fue el cv. Fortune que en polinización cruzada con cualquier variedad de clementina produce de 20 a 25 semillas por fruto en las dos variedades (ASÍNS et al., 2002). 2.3. Polinización en cítricos: La polinización se define como un acto mecánico de transferencia de polen a las partes femeninas de una flor (ROOT, 1976). También se describe como el acto de llevar el grano de polen desde la antera de una flor a los pistilos de otras flores, sean o no pertenecientes al mismo pie de planta, siendo la polinización el preludio de la fecundación (RALLO, 1986). Cuando se polinizan variedades que carecen de semillas o que las tienen en escaso número como mandarinas Satsumas, mandarinas clementinas o el cultivar Cadenera, 11 aparece un incremento de semillas en los frutos (GONZÁLEZ-SICILIA, 1968; MONSELISE, 1986). La mayoría de las especies cítricas comercialmente importantes no requieren polinización cruzada para producir una cosecha. Las excepciones a estos casos incluyen algunos híbridos de mandarino como el tangelo Orlando y el mandarino Robinson. La polinización es esencial, sin embargo, para la producción de semillas, o para estimular el crecimiento del ovario en los cultivares casi partenocárpicos como el naranjo dulce Hamlin (DAVIES y ALBRIGO, 1994). BEN-CHEIKH et al. (1997), observaron en el cultivar semillado Pineapple de naranjo dulce, que la polinización aumentaba la cantidad de giberelinas (GAs) en los ovarios en desarrollo. Luego, la emasculación de las flores redujo los niveles de GAs y causó una rápida abscisión del 100% de los ovarios, esto indicaba que un nivel alto de GAs, provocado por la polinización, era importante para evitar la caída de frutitos. Este efecto de caída fue parcialmente contrarrestado con polinización y aplicaciones de GA3. Además, en ovarios polinizados, aplicaciones con paclobutrazol (inhibidor de la síntesis de GAs) disminuyeron la cantidad de GAs y aumentaron la abscisión de ovarios, aunque el modelo de disminución de cuajado de frutos fue diferente a la repentina abscisión inducida por la emasculación (BEN-CHEIKH et al., 1997). En ensayos realizados por WALLACE y LEE (1999) también se demostró que la polinización es un factor principal para aumentar el cuajado, ya que en las mandarinas Imperial, Ellenor y Murcott, se observaron fuertes requerimientos de polinización manual para aumentar el cuajado. De estos cultivares específicos, Imperial respondió mejor a la polinización cruzada, Ellenor a la polinización con Murcott, y Murcott a cualquier polinización manual. 12 En estudios recientes, PUCHADES (2002), observó que la polinización tiene un gran efecto sobre el cuajado en mandarino híbrido Fortune. La polinización cruzada con mandarina del cv. Clemenules y autopolinización forzada mejoró drásticamente el cuajado y el diámetro, como consecuencia de la relación fructificación-semillas en frutos. La mandarina Fortune, en ausencia de este tipo de polinizaciones tuvo un cuajado muy bajo, por lo que cabía suponer que era incapaz de producir frutos partenocárpicos. En algunos casos, el diámetro fue excesivo, con lo cual se perdía calidad comercial. Por otra parte PUCHADES (2002), con datos de dos años en ensayos de polinización realizados con polen de mandarina clementina cv. Clemenules, sobre flores del cv. Fortune, obtuvo que los porcentajes de flores que quedaron en el árbol, es decir, que no abortaron, fueron mayores en los ensayos de polinización cruzada (53% y 80%); seguidos de la autopolinización manual (20% y 8%) y muy bajos en las flores sin aporte de polen por emasculación (0,5% y 1%) o embolsadas (4% y 0%). 2.3.1. Tipos de polinización La polinización de un pistilo puede efectuarse por el polen procedente de la misma planta o de otra distinta. En el primer caso hay autopolinización y en el segundo polinización cruzada. Para que, desde el punto de vista genético, haya diferencia entre ambos tipos de polinización, es preciso que la cruza tenga lugar entre plantas de constitución genética distinta, ya que la que se efectúa entre plantas de la misma variedad equivale a una autopolinización (PUCHADES, 2002). En general, cuando la autopolinización se efectúa en una variedad auto incompatible no tiene lugar la formación de semillas, pero sí cuando es auto compatible. También se forman semillas cuando se produce polinización cruzada entre variedades compatibles. Cruzamientos entre distintos parentales compatibles influyen en el número de semillas, e incluso en el tamaño de éstas (BUKIYA, 1990). 13 2.3.1.1. Autopolinización Es la transferencia de polen al estigma de la misma flor o a estigmas de diferentes flores de la misma planta, o a flores de diferentes plantas pero del mismo genotipo (ROOT, 1976). 2.3.1.2. Polinización cruzada ROOT (1976) la define como la transferencia del polen desde la flor de un cultivar, a otra planta de un cultivar diferente. Es decir, cuando la procedencia del polen a una flor es de otro pie de planta genéticamente diferente. Cuando se produce polinización cruzada entre variedades compatibles, sobretodo con los híbridos, provoca un aumento en la producción, debido a que hay un aumento importante de frutos cuajados, así como un aumento de su diámetro; todo ello acompañado por un aumento importantísimo del número de semillas por fruto (ZAURASHUILI, 1967). BONO et al. (2004) citan que las flores de las clementinas tienen altos porcentajes de polen y óvulos fértiles, pero no se produce fecundación, y por consiguiente formación de semillas, ni en una misma variedad (auto incompatibilidad) ni entre las distintas variedades del grupo (inter incompatibilidad). Ahora bien, cuando polinizan o son polinizadas por variedades de otros grupos (a excepción del grupo Navel y en menor medida el grupo Satsuma), aparecen semillas. Este grupo presenta partenocarpia, pero parece ser que la actividad de los polinizadores aumenta la producción. Las variedades son autoestériles por lo que sólo formarán semillas en presencia de fuentes de polen viable de otras variedades en las proximidades (MUÑOZ AYUSO y LABRADOR, 2005). 14 Además, todas las variedades mencionadas presentan semillas cuando se polinizan entre ellas y cuando son polinizadas por las variedades de otros grupos (a excepción del grupo Navel y en menor medida al grupo Satsuma); también provocan la aparición de semillas cuando polinizan a las variedades de los otros grupos con las excepciones mencionadas (MUÑOZ, AYUSO y LABRADOR, 2005). ZARAGOZA et al. (2005) citan que las variedades híbridas, Nova, Fortune y Ortanique inducen la formación de semillas en los frutos de otros híbridos, y en Clementinos y Satsumas, debido a la polinización cruzada. 2.3.2. El polen y su transporte en los cítricos PONS et al. (1995b), en un trabajo realizado con Clementina de Nules y Fortune, comprobaron la poca importancia que el viento tiene como vector de transporte de polen, indicando que son los insectos los que fundamentalmente intervienen en la polinización de los cítricos. Y en ese sentido, PONS et al. (1996), determinaron que las abejas melíferas (Aphis mellifera) representaron el 95,4% del total de insectos que visitaron las flores en un plantación del cv. Fortune. En términos similares se expresan FREE (1993) y BHATIA et al. (1994). Por su parte SOLER (2005)*, agrega que solamente es la polinización entomófila, y la efectuada por las abejas la de importancia en cítricos, ya que la producida tanto por el viento como por otros insectos carece de importancia. ____________________ *SOLER, J. Ing.Téc.Agr. 2005. Investigador IVIA y profesor Master En Citricultura de la Universidad Politécnica de Valencia. Valencia, España. 2005. Comunicación personal. 15 2.4. Fecundación en cítricos: Una vez que el polen se encuentra sobre el estigma, se produce la germinación del tubo polínico y este penetra vía micropilo en el saco embrionario del óvulo. Dos núcleos espermáticos se producen por el núcleo generativo del polen. Un núcleo espermático se fusiona con la oosfera formando un zigoto diploide, mientras que el otro núcleo espermático se fusiona con los dos núcleos polares del óvulo, y formarán el endosperma triploide. La fecundación de la oosfera ocurre dos o tres días después de la polinización bajo condiciones favorables, pero en algunos casos se ha informado un lapso de tres a cuatro semanas (SPIEGEL-ROY y GOLDSCHMIDT, 1996). La síntesis de citoquininas probablemente este relacionada con la mitosis tan intensa, sin embargo, no esta claro el origen de estas fitohormonas; puede que sean importadas de la raíz, o bien que las produzcan las mismas semillas (quizás el suspensor). El suspensor está muy relacionado con la biosíntesis de GAs detectadas en el endosperma en este período, hasta el punto de que en cultivo in vitro, se puede sustituir al suspensor por GAs exógenas (MATILLA, 2000). HERNANDEZ y PRIMO-MILLO (1990), determinaron el contenido endógeno de citoquininas, entre cultivares de cítricos con y sin semillas, y observaron que la polinización aumentó el contenido de esta fitohormona en los ovarios de todos los cultivares semillados, en comparación con los cultivares sin semillas. Esto indicaría que la fertilización de los óvulos es el principal sitio para la biosíntesis de esta hormona en estos frutos. ROHIDAS y CHAKRAWAR (1989) informan que el estigma es receptivo tres días antes de la antesis y se mantiene receptivo hasta seis u ocho días tras la apertura floral. En algunas variedades el estigma esta expuesto antes de la antesis por lo que puede ser polinizado. ETI y STOSSER (1992), observaron como cultivares de 16 mandarina clementina los tubos polínicos crecían normalmente tras la polinización cruzada, entrando en el núcleo por el micropilo tras un período variable de cinco a doce días. Sin embargo, en situaciones de autopolinización el crecimiento del tubo polínico se interrumpía en la parte superior del estilo, confirmando así la auto incompatibilidad en clementina. La regulación del crecimiento de los frutos depende en gran medida de las hormonas sintetizadas en las semillas, y de forma particular de las GAs. En muchas especies se ha demostrado que el número de frutos que cuajan, e incluso su tamaño individual correlaciona con el número de semillas que poseen. En los frutos sin semillas, la ausencia de polinización y de semillas detiene el crecimiento del fruto y provoca su abscisión. En algunos frutos como por ejemplo el guisante, pera, fresa y tomate, la aplicación de GA3 a frutos no polinizados sustituye el efecto de la polinización y las semillas y provoca la estimulación del desarrollo del fruto partenocárpico (GOODWIN, 1978). 2.5. El cuajado en cítricos: El cuajado, es el proceso que determina el tránsito del ovario de la flor a fruto. Este proceso está determinado en forma natural en variedades con semilla, por el crecimiento del tubo polínico y la fecundación del óvulo. Luego, las semillas son las que estimularán el crecimiento del fruto por medio de la síntesis de sustancias de crecimiento, principalmente giberelinas (GAs), dando como resultado frutos de mayor tamaño, por lo que el calibre final del fruto, medido como diámetro ecuatorial, es uno de los factores de mayor importancia para el consumidor, aunque el tamaño preferido puede variar de acuerdo a los requisitos de cada mercado (DELHOM y PRIMO-MILLO, 1989). 17 Comercialmente hay ciertos rangos preferidos para cada especie, siendo estos para mandarinas e híbridos entre 55-65 mm, como diámetro ecuatorial, para el caso de exportación, con un peso de 80 a 100 gramos (ORTÚZAR, 1999). El cuajado se define como la fase del desarrollo que marca la transición de una flor (ovario) a un frutito que se desarrollará hasta la maduración. Un fallo en el proceso de cuajado provoca la detención del crecimiento y, normalmente, a abscisión del ovario en el plazo de unos pocos días (GUARDIOLA, 2003). En los cítricos, el proceso de cuajado del fruto constituye la transición de ovario a fruto en desarrollo y se caracteriza por el paso de una fase de crecimiento lento a una fase de crecimiento rápido (TALÓN, 2000). La mayoría de las variedades con semillas cuajan con facilidad, pero en el caso de las variedades sin semillas el número de flores caídas es tan alto, que el número de frutos cosechados no supera el 10% de la floración inicial e incluso llega a ser tan bajo con valores de 0,1% (GOLDSCHMIDT y MONSELISE, 1977). Por está razón, se han desarrollado técnicas para aumentar el cuajado, con aplicaciones de ácido giberélico para algunas variedades (TALON, ZACARIAS y PRIMO-MILLO, 1992), y como el rayado, cuya eficacia parece general para todas las variedades ensayadas (AGUSTÍ et al., 1998). Clementina de Nules puede ser considerada una variedad con cuajado de fruto deficiente, pues en ausencia de polinización cruzada produce una cosecha escasa (MEHOUACHI, 2001). 18 2.6. Partenocarpia en cítricos: Según JUAN y PRIMO-MILLO (1989), la partenocarpia es la formación de frutos sin la formación de los óvulos y por tanto sin desarrollo de semillas, encontrándose en la actualidad muchas variedades cultivadas que tienen la capacidad de desarrollar frutos sin semillas (partenocárpicos). Por otro lado PRIMO-MILLO (1994) señala que la partenocarpia es un fenómeno que se produce en forma natural en muchas especies de cítricos y puede tener un origen genético o ambiental. Tras la fecundación, algunos óvulos se transforman en semillas que se desarrollan hasta su maduración. En las variedades partenocárpicas, los óvulos no son fecundados y, por tanto, las semillas no se desarrollan, quedando reducidas a pequeños rudimentos seminales (AGUSTÍ, 2000). NISTCH (1971) sugirió que el factor genético responsable de la partenocárpia, favorece el mantenimiento de un nivel hormonal relativamente alto en el ovario en el momento de la antesis e inmediatamente después de ella. El estímulo necesario para el cuajado de los frutos partenocárpicos, procede de las hormonas presentes en los óvulos o en el pericarpio (NISTCH, 1970). En la partenocarpia estimulativa, la polinización, germinación del polen y el crecimiento del tubo polínico en ausencia de fecundación, provee el estimulo suficiente para cuajar frutos sin semilla. Así, la autopolinización puede ejercer un estímulo suficiente en genotipos autos incompatibles para el cuajado de frutos sin semillas. Aplicación de GAs sustituye la necesidad de la polinización en Clementinas y otros híbridos de Citrus reticulata, conduciendo a la producción de frutos partenocárpicos (SPIEGEL-ROY y GOLDSCHMIDT, 1996). 19 En los frutos con semillas, su desarrollo depende de éstas. Su eliminación o la emasculación, que evita su formación, detienen el desarrollo del fruto y producen su abscisión, pero en estos casos la aplicación de hormonas restituye el crecimiento, dando lugar así a frutos partenocárpicos. En otros casos, la aplicación exógena de hormonas antes, durante o inmediatamente después de la antesis induce el crecimiento partenocárpico del fruto de flores intactas. Por lo tanto se puede deducir que son las hormonas producidas por las semillas las que regulan el crecimiento del fruto (AGUSTÍ, 2003). Para el caso de la mandarina Clementina lo que interesa es la obtención de fruta con un calibre apropiado y que sea partenocárpica, es decir, con ausencia de semillas. Si bien es cierto esta especie presenta una esterilidad gamética relativa, el cual se debe a fenómenos de autoincompatibilidad, en donde es incapaz de formar embriones por autopolinización, aunque los gametos son perfectamente funcionales con polinización cruzada (TALÓN, 1997). La importancia de las GAs en la estimulación del crecimiento del fruto está bien establecida. Por un lado, la actividad de la GA1 en Satsuma; por otro lado, el paclobutrazol, incrementa la abscisión de frutos; la abscisión inducida por al paclobutrazol puede ser revertida con aplicaciones de GA3 y aplicaciones de GA3 mejora el cuajado en la mandarina Clementina (Citrus reticulata) pero tienen una pequeña influencia sobre la mandarina Satsuma, la cual tiene una buena capacidad partenocárpica y de cuajado en forma natural (TALON, ZACARIAS y PRIMOMILLO, 1992). TALÓN, HEDDEN y PRIMO-MILLO (1990), sugieren que el potencial para cuajar frutos partenocárpicos es principalmente influenciado por el balance hormonal de los frutos, durante estados tardíos de la división celular y estados tempranos de la elongación celular. Es decir, la condición de baja partenocarpia parece esta asociado 20 con niveles bajos de actividad de GAs, baja capacidad para catabolizar ABA libre a ABA conjugado y una gran habilidad para conjugar AIA durante este período. Por otra parte GARCIA-PAPI y GARCIA-MARTINEZ (1979), aplicaron giberelinas a flores emasculadas de mandarina Monreal promoviendo el desarrollo de frutos partenocárpicos. Esto se puede interpretar en el sentido de que la presencia de giberelinas en el fruto, es esencial en al cuajado y que las diferentes respuestas obtenidas entre variedades puede deberse a diferencias en el contenido endógeno de estas sustancias en sus ovarios. Puede deducirse que son las semillas las responsables de la síntesis de giberelinas y en ausencia de ellas se ha postulado que son las paredes de los ovarios las que asumen este papel (MONSELISE, 1977). TALON, ZACARIAS y PRIMO-MILLO (1992) en un investigación realizada concluyeron que en mandarinas Satsumas niveles altos de giberelinas endógenas están asociadas a una elevada capacidad partenocárpica, y en clementinas los niveles bajos de giberelinas endógenas están asociados a una baja capacidad partenocárpica; por otra parte aplicaciones de giberelinas (GA3) aumenta el cuajado de Clementinas y tiene poca influencia en Satsumas; la inhibición de GA3 por aplicación de paclobutrazol resulta en la abscisión de frutos en ambas variedades. Se propone por estos autores que el GA3 endógeno controla el desarrollo partenocárpico en ambas variedades. La participación directa de fitohormonas como las giberelinas o citoquininas, serviría para el aporte de nutrientes y la regulación del desarrollo de los órganos (ADDICOTT, 1982). Otros autores han propuesto que las giberelinas podrían contrarrestar los efectos del ABA o del etileno en frutos de cítricos, lo que disminuiría la abscisión de éstos (ZACARÍAS et al., 1995). 21 2.7. La presencia de semillas en las mandarinas y su problemática: El problema de la presencia de semillas en el fruto comenzó con la desaparición de las plantaciones de mandarino Común, y con el comienzo del cultivo de mandarinos tales como Nova, Fortune, entre otros a nombrar. Hasta entonces la producción de mandarinas se había caracterizado por la ausencia prácticamente total de semillas, cualidad muy apreciada por los consumidores. Pero desde que se inició el cultivo de los mandarinos citados, con el objetivo de prolongar el período de producción de mandarinas y desestacionalizar la oferta, se empezó a detectar la presencia de semillas, y a medida que su producción iba en aumento, también se incremento la proporción de frutos con semillas; ello implica una disminución importante de la calidad y provoca que los frutos sean rechazados por la mayor parte de los consumidores o se vendan a precios más bajos (BONO, SOLER y BUJ, 2000). La presencia de semillas se debe principalmente a la polinización cruzada que tiene lugar entre los citados mandarinos y el resto de las variedades cultivadas (PONS et al., 1995a, 1995b y 1996). 2.8. Utilización de productos con posible efecto polinicida: 2.8.1. Aplicación de sales de cobre MESEJO et al. (2005) diseñaron un proyecto de investigación en tres etapas para solucionar el problema de la presencia de semillas en los frutos: determinar que sustancias podrían ser eficaces como gametocidas en condiciones in vitro, y comprobar si estás tenían la misma respuesta in vivo aplicándolas, primero a flores individuales y, finalmente, al árbol completo. De las sustancias analizadas in vitro, aplicadas a granos de polen del cv. Clemenules, se identificó que el CuSO4*5H2O a una concentración de 25 mg l-1 o superior, anuló la germinación de los granos de 22 polen cultivados, cuando cerca del 70% de los no tratados conseguían germinar (MESEJO et al., 2005). La acción gametocida del sulfato de cobre, radica en que impide la germinación de los granos de polen y si este germina interrumpe el desarrollo del tubo polínico. El tratamiento debe realizarse antes de la polinización, ya que los resultados obtenidos en aplicaciones de 24 h después de polinizada la flor, no mostraron ser eficaces en impedir la germinación del polen y/o el desarrollo del tubo polínico (AGUSTÍ, 2005)*. Tanto las aplicaciones a las flores individuales como al árbol completo, con la dosis de 25 mg l-1, redujo la cantidad de semillas en los frutos, pero se presenta el grave inconveniente que las flores no se encuentran todas el mismo estado de desarrollo en el momento del tratamiento y, por tanto cumplir con el requisito de que el tratamiento debe efectuarse antes de que ocurra la polinización resulta complicado. Para verificar cual es el mejor estado del árbol para realizar el tratamiento, se hicieron tratamientos en diferentes estados de floración del árbol, resultando el más eficaz cuando el árbol presentaba aproximadamente el 60% de las flores en antesis (MESEJO et al., 2005). En resumen la aplicación de 25 mg l-1 de CuSO4 * 5H2O cuando el 60% de las flores inician su apertura, se presentan como una técnica eficaz para reducir el número de semillas por fruto de las mandarinas cultivadas en condiciones de polinización cruzada. En las clementinas este tratamiento es compatible con la adición de ácido giberélico (5-10 mg l-1) para estimular el cuajado (MESEJO et al., 2005). __________________________ AGUSTÍ, M. Doctor. Ing. Agr. 2005. Profesor de Citricultura y Fruticultura de la Universidad Politécnica de Valencia. Valencia, España. Comunicación personal. 23 2.8.1.1. Características de las sales de cobre Con respecto a los productos cúpricos o sales de cobre se puede decir que son de amplio uso en la agricultura, y en especial en la fruticultura para el control de enfermedades causadas tanto por hongos como por bacterias. Son productos que actúan por contacto y son de uso preventivo, actuando principalmente a nivel del ciclo de Krebs o también desactivando proteínas, por lo que impiden la proliferación de estos patógenos (AFIPA, 2003). Entre los productos cúpricos mas usados tenemos el oxicloruro de cobre, cloruro de cobre, sulfato de cobre, óxido cuproso, hidróxido de cobre y óxido de cobre, los cuales vienen formulados como polvo mojable o suspensión concentrada (AFIPA, 2003). El oxicloruro de cobre es un fungicida inorgánico, de amplio espectro de acción, previniendo enfermedades sobre frutales, hortalizas, flores y ornamentales. Por su tamaño de partículas (90% entre 3-10 micrones) permite lograr una mejor cobertura, eficacia y adherencia (AFIPA, 2003). El óxido cuproso es un fungicida-bactericida inorgánico, que actúa por contacto y preventivamente sobre hongos y bacterias que atacan frutales y cultivos. El óxido cuproso es una de las formas mas activas del cobre como fungicida, lo cual le confiere una alta eficiencia la que se basa en el tamaño súper fino de las partículas de óxido cuproso, lo cual le permita alcanzar una mayor superficie de contacto, una mejor distribución del cobre y una mejor adherencia en la superficie (AFIPA, 1998). 24 2.8.2. Otros compuestos con capacidad polinicida Hay otros compuestos que se han utilizado en cultivos como los cereales o de tipo hortícola, para la producción de híbridos. Estos compuestos son sistemas bioquímicos que están basados en el uso de productos que cambian el comportamiento de la planta, como las hormonas femeneizantes, los inhibidores del desarrollo del polen o anteras, tanto en el tejido esporofítico como en el gametofítico (gametocida) y los inhibidores de la fertilidad del polen (CROSS y SCHULZ, 1997). Los agentes hibridizantes químicos (Chemical Hybridizing Agents CHAs), pertenecen a este último grupo y tomaron importancia desde 1950, cuando fue posible demostrar su selectividad para obtener plantas macho estériles, presentando ventajas por sobre el uso de la macho esterilidad citoplasmática, como acortar los tiempos de producción de un híbrido y no requerir de líneas restauradoras de fertilidad (Mc RAE, 1985). Estos agentes hibridizantes químicos tienen diferentes modos de acción. Estudios con LY195259 mostraron que su aplicación inhibe la formación de anteras (TSCHABOLD et al., 1988). A3C (Acetdine-3-carboxilic acid) afecta la funcionalidad del polen (MOGENSEN y LADYMAN, 1989). El uso de RH0007 (Fenridazon) actúa provocando el colapso de los granos de polen y su muerte debido a inhibición del ciclo bioquímico de la esporopolenina, componente fundamental de la exina, que bajo la acción del producto es un 80% más delgada que la normal (ELGHAZALY y JENSEN, 1990). En el caso de Clofencet se sabe que tiene actividad sistémica (NESVADBA y VYHNÁNEK, 2001), pero se tiene poca información. El modo de acción no ha sido informado, pero se cree que las microsporas abortan previo a la vacuolación (UAUY, 2001). 25 Dentro de estos citados compuestos esta el producto de nombre comercial Genesis®, es un agente hibridizante químico que tiene como ingrediente activo clofencet (sal potásica del ácido 2-(4-Clorofenil)-3-etilo-2,5-dihidro-5-oxo-4-Piridazincarboxílico ) que suprime el desarrollo normal de polen en la planta hembra, se utiliza en dosis de 3,4 a 6,7 kg x ha i.a, con un tope máximo de 11,2 kg x ha i.a por temporada con una ventana entre los siete y nueve grados de la escala de Feekes. Se realiza una aplicación única a menos que sea necesario reaplicar por lluvias que debe realizarse al menos 45 días antes de la cosecha de la semilla. Está en la categoría III de la clasificación toxicológica, es decir, posible cancerígeno humano (EPA, 1997). GAJU (2002) evaluó al producto Genesis® en trigo para provocar una esterilidad masculina y de esta forma favorecer la hibridación. Sus resultados fueron que con una dosis de 6,5 kg/ha de producto comercial aplicado en la segunda fecha de aplicación, correspondiente a plantas en 8,5 grados en la Escala de Feekes, las que presentaban tres nudos visibles sobre la superficie, la hoja bandera parcialmente desplegada y las espigas tenían 1,76 cm. promedio, obtuvo un 96% de macho esterilidad permitiendo su uso como gametocida (polinicida) para producir trigo hibrido. Por otra parte WONG, BLOUET y GUCKERT (1995) consideran que un 95% de MS es suficiente para la producción de semilla híbrida comercial. Por su parte BATCH et al. (1980), citan que en poroto de soja hay tres grupos químicos que han sido utilizados como productos gametocidas y específicamente polinicidas (provocan esterilidad a nivel del polen), siendo estos compuestos: derivados del etileno, derivados del arsénico y hormonas del crecimiento. De estos tres grupos se ha encontrado al compuesto etilo 4-fluoroxanilato como el que mejor resultado ha provocado en producir esterilidad masculina (ALI et al., 1999) en una dosis de 1000 ppm (MATHUR y LAL, 1999) en poroto de soja. 26 Como efectos adversos de los agentes gametocidas, se tiene que se necesitan altas dosis para obtener efectividad deseada, y por otra parte, se tiene que dosis altas requeridas del agente gametocida pueden afectar negativamente la fertilidad femenina o reducir el período de receptividad de la hembra, son muy específicos en la generación de macho esterilidad, obteniendo buenos resultados en una sola especie o variedad, no son lo suficientemente selectivos, ventanas pequeñas de aplicación limitan su uso práctico y deben evaluarse interacciones genotípicas, químicas, ambientales (Mc RAE, 1985; EDWARDS, 1986). VAN DER MEER y VAN BENNEKOM (1975) concluyen que ácido giberélico en dosis de 2% actúa como un efectivo polinicida, es decir, que provoca un esterilidad masculina reemplazando a la emasculación manual en cebolla. JAN y ROWELL (1981) en un estudio realizado con tres compuestos con capacidad gametocida (Ethrel, RH-532 y RH-2956), determinaron que el compuesto RH-2956, fue el mas efectivo en la dosis mas alta, y Ethrel fue solo efectivo en los primeros estados de desarrollo del trigo. BINGHAM y LUPTON (1987) indican que una 85% o más de cuaja del grano de trigo, depende en gran medida de la coordinación entre la emergencia de las espigas machos y el tiempo de receptividad de la hembra. Otros compuestos con características polinicidas son el ácido cítrico, nitrato cálcico y el ácido indolacético (AIA), en donde la aplicación de concentraciones crecientes de ácido cítrico hasta 20 mg l-1, nitrato cálcico hasta 15 g l-1 y de ácido indolacético hasta 50 mg l-1 reduce significativamente la germinación, hasta anularla, y la longitud del tubo polínico. A pesar del papel determinante del Ca+2 en la germinación del polen (GEITMANN, 1999) y en el crecimiento del tubo polínico, ambas situaciones estudiadas por BREWBAKER y KWACK (1963) en un medio con agarosa (EEO 27 media, Pronadisa) al 1%, ajustado a un ph de 5.4 ± 0.1. Estudios realizados por MESEJO et al. (2005), indican un papel inhibidor de este elemento en la germinación, efecto que ha sido registrado por otros autores (GONG, YANG y CAO, 1995). Los resultados existentes sobre el efecto del AIA indican, en todos los casos, un efecto positivo de la hormona sobre dicho crecimiento para concentraciones bajas (inferiores a 10 µM) (GEITMANN, 1997), pero la situación se invierte para concentraciones altas de la hormona, por encima de 100 µM, para las que se inhibe la germinación del polen y el desarrollo del tubo polínico (KWAN, HAMSON y CAMPBELL, 1969; GEITMANN, 1997). Por otra parte, la acción del ácido cítrico inhibiendo la germinación del polen e impidiendo el crecimiento del tubo polínico es compartida con otros ácidos tricarboxílicos (KWAN, HAMSON y CAMPBELL, 1969). 28 3. MATERIALES Y MÉTODOS 3.1. Ubicación del ensayo: La ubicación del ensayo fue en un huerto particular de mandarinas clementinas cv. Clemenules, propiedad de Sociedad Comercial Agrícola Los Rosales de Santa Julia, ubicado en el sector de San Isidro, provincia de Quillota, V región, Chile (32° 50’ latitud sur y 71° 13’ longitud oeste). Por otra parte, se trabajó con un huerto particular de mandarina cv. Fortune, perteneciente a la Sociedad Agrícola Arancibia, ubicado en el sector La Palma, provincia de Quillota, con el fin de obtener polen de este cultivar y ser utilizado en los árboles del cv. Clemenules para la polinización de sus flores. El huerto con los árboles del cultivar Clemenules, tenía una edad de plantación de cinco años aproximadamente, siendo Carrizo el patrón utilizado. La superficie de la parcela era de una hectárea aproximadamente, con riego por goteo, con un marco de plantación de seis metros entre hileras y tres metros sobre la hilera, en donde el destino de su fruta es el mercado de exportación. El huerto del cv. Fortune tenía una edad aproximada, en el momento de los ensayos, de tres años, con patrón Carrizo, riego por goteo, con un marco de plantación de seis metros entre hilera y tres metros sobre la hilera. Ambos huertos, al momento de los ensayos, se encontraban en buenas condiciones, es decir, sin ningún problema que les afectara, y que por lo tanto pudiera influir en los ensayos. 3.2. Caracterización climática y edáfica de la zona del ensayo: El agroclima Quillota se encuentra en el sector poniente del valle de Aconcagua, latitudes 32º50’S a 33º10’S constituyendo un clima muy local dentro de la zona (NOVOA et al., 1989). La localidad de Quillota se encuentra ubicada en la región de los valles transversales, correspondiendo su clima al tipo mediterráneo, el cual se 29 caracteriza por veranos secos y cálidos, moderados por vientos subtropicales: Los inviernos se presentan lluviosos debido a la acción del frente polar (MARTÍNEZ, 1981). ROMERO (1985) agrega que Quillota presenta una alternancia de las influencias marinas con las continentales. El régimen térmico se caracteriza por presentar una temperatura media anual de 15.3 °C, con una máxima media del mes más cálido (enero) de 27 °C, y una mínima media del mes más frío (julio) de 5.5 °C. La suma anual de temperaturas base 5°C es de 3700 grados día, y base 10 °C es de 1900 grados día. Las horas de frío, de marzo a noviembre llegan a 500. La temperatura media mensual se mantiene sobre 10ºC, y el periodo libre de heladas es de nueve meses, desde septiembre hasta mayo (NOVOA et al., 1989). El régimen hídrico se caracteriza por una precipitación anual promedio de 437 mm, siendo el mes de julio el más lluvioso con 125 mm. La evaporación media llega a 1361 mm anuales, con un máximo mensual en diciembre de 219.3 mm y un mínimo en junio de 36.1 mm. La estación seca es de ocho meses (NOVOA et al., 1989). Con respecto a las características edáficas de la zona de los ensayos, estas corresponden a un régimen de humedad xérico que limita hacia el norte con un régimen arídico y hacia el sur con régimen údico. En esta unidad, el desarrollo de los suelos es moderado, lo cual significa que hay dominancia de horizontes B de alteración (B cámbico) y con escasos horizontes B de iluviación (B argílico). Por otra parte, el contenido de materia orgánica del horizonte superficial, unido a una estructura bien desarrollada, determinan la presencia de un epipedón mólico, de tal manera que gran parte de los suelos de esta unidad, son Mollisols. Localmente, hay dominancia de materiales calcáreos que dan origen a suelos que presentan reacción al HCl a través de todo el perfil (LUZIO y ALCAYAGA, 1992). 30 Las características edáficas corresponden a la serie La Palma, es de tipo sedimentario, profundo de origen coluvial, formado a partir de sedimentos graníticos de los cerros, substrato constituido por gravas o piedras con material intersticial y suelo que le otorgan una permeabilidad moderada y buen drenaje (CASTRO, 2000). 3.3. Descripción del ensayo: En esta investigación se evaluaron cuatro sales de cobre en cuatro fechas diferentes. Para esto se contabilizó en la parte final de la investigación, el número de semillas por fruto, para llegar a concluir si los productos presentaron el efecto polinicida buscado, y por otra parte, se midió el calibre de cada fruto, para establecer la relación con la posible presencia de semillas en el fruto y el tamaño de éstos. Para llevar a cabo los diferentes ensayos, se utilizaron botones florales de mandarina clementina de Nules, y el polen de la mandarina Fortune, para polinizarlos. Para llevar a cabo lo anterior se trabajó con los siguientes grupos florales: GE/P: Flores emasculadas y polinizadas con polen del cv. Fortune. GSE/SP: Flores sin emasculación y sin polinización. GE/SP: Flores emasculadas y sin polinización. La forma de trabajo consistió en dejar 50 botones florales del Grupo GE/P y GSE/SP, para la aplicación de las cuatro sales de cobre en las cuatro distintas fechas, mientras que 50 botones florales del grupo GE/P y GE/SP se dejaron sin aplicación de ningún producto, como control de los tratamientos. Los tratamientos consistieron en aplicar los cuatro productos cúpricos o sales de cobre que fueron: sulfato de cobre, oxicloruro de cobre, óxido cuproso y cloruro de cobre, en una única dosis (20 ppm). Los productos se aplicaron en cuatro fechas distintas, siendo estas: 15 ddpf (días después de plena flor), 30 ddpf, 45 ddpf y 31 término de la segunda caída fisiológica de frutitos. El total de botones florales utilizados, incluyendo a todos los tratamientos, fue de 2000 botones florales, dividiéndose en 400 botones florales por producto, más 400 de testigo, sin producto. CUADRO 1. Tratamientos y número de flores por tratamiento. Sulfato de Oxicloruro Oxido Cloruro de Fecha Sin cobre de cobre cuproso cobre producto (P1) (P2) (P3) (P4) (control) G E/P G G E/P G SE/SP G E/P G SE/SP G E/P G SE/SP G E/P SE/SP G E/SP F1: 15 ddpf 50 50 50 50 50 50 50 50 50 50 F2: 30 ddpf 50 50 50 50 50 50 50 50 50 50 F3: 45 ddpf 50 50 50 50 50 50 50 50 50 50 F4:Término 50 50 50 50 50 50 50 50 50 50 2ª caída Total x 400 400 400 400 400 producto 3.4. Época de realización de los ensayos: Las labores en el huerto se iniciaron el 3 de noviembre del 2003, fecha en que el huerto estuvo disponible y que se marcó como el inicio de la plena flor, fecha clave para llevar a cabo la evaluación de los diferentes productos y de cada fecha de aplicación. Las diferentes labores de los ensayos se extendieron hasta el 2 de julio del 2004, fecha en que se culminó con la cosecha de la fruta perteneciente a cada ensayo. 32 3.5. Materiales utilizados: Los materiales utilizados se pueden dividir, en los usados en campo, correspondiente a la primera etapa de esta investigación, y los usados en la segunda etapa, que consistió en medir el diámetro ecuatorial de cada fruto con un pie de metro (calibre, en mm), y contabilizar el número de semillas de cada fruto. 3.5.1. Materiales utilizados en campo Los materiales utilizados en campo están relacionados con las labores de polinización, emasculación y embolsado de los botones florales, y a su vez con la preparación de la solución de agua más los diferentes productos cúpricos para su posterior aplicación. A continuación se presenta la lista de los diferentes materiales, tanto instrumentos de trabajo como material vegetal propio de la investigación, usados en esta primera etapa de la investigación, junto con una descripción de su uso. 3.5.1.1. Material vegetal 1.- Botón floral de mandarina clementina cv. Clemenules: Se trabajó en botón floral, debido a que en estado fenológico, que precede a la antesis, no hay riesgo de que la flor haya sido visitada por algún insecto, principalmente la abeja, y con eso se haya contaminado con polen de otra flor (Figura 1). 2.- Polen del híbrido cv. Fortune: Se utilizó como agente polinizante de clementina, lo cual en trabajos realizados por SOLER et al. (1996), demostraron la eficacia del cv. Fortune como polinizante para el cv. Clemenules, por lo que su uso fue ideal para poder llegar a determinar si existía algún efecto polinicida de alguno de los cuatro productos en evaluación, y además sirvió para ver la eficacia de este híbrido como polinizante de clementina, en las condiciones de la provincia de Quillota (Figura 2). 33 3.5.1.2. Material de trabajo 1.- Bolsas de malla de tul: Su objetivo principal fue el proteger a la flor de los insectos, principalmente abejas, para evitar que estos depositarán polen de otras especies en las flores tratadas, lo cual hubiera producido una distorsión de los resultados. 2.- Pincel Nº 5: Se utilizó para polinizar los botones florales de mandarina clementina con el polen del cv. Fortune, el cual se depositaba en un frasco cilíndrico para su transporte en campo. 3.- Pinza quirúrgica: Utilizada para la emasculación de los botones florales de mandarina clementina. 4.- Productos cúpricos: Se evaluaron cuatro productos cúpricos, en una dosis única, siendo los siguientes: oxicloruro de cobre, sulfato de cobre, óxido cuproso y cloruro de cobre. 3.5.2. Materiales utilizados en la medición de resultados En esta parte de la investigación se utilizaron un pie de metro digital, para medir el calibre (en mm), de los frutos de los diferentes ensayos, y un cuchillo para cortar y contabilizar el número de semillas de cada fruto. 3.6. Metodología de trabajo: 1.- Embolsado de flores: Correspondió a dejar al botón floral aislado para evitar la visitas de las abejas, principal agente polinizador en cítricos. Cada bolsa se hizo de 34 10x15 cm, como una forma de que la flor quedara con un espacio amplio para su desarrollo. 2.- Emasculación y polinización de flores: Consintió en la eliminación de los estambres de las flores del cv. Clemenules y su posterior polinización con polen del cv. Fortune, lo que se realizó de la siguiente forma: a) Recolección del polen de Fortune: El huerto de Fortune, al momento de que se realizó la recolección de polen se encontraba con diferentes estados fenológicos, entre los que se podían visualizar el de plena flor, de caída de pétalos, y frutitos cuajados (estadios 65, 67, 69 y 71 según escala BBCH) (los estados fenológicos en que se realizaron las labores serán especificados más adelante). El estado de interés para la recolección de polen es el de plena flor, teniendo la precaución que las anteras presentarán un color amarillo claro y no un café oscuro, las cuales presentaban una menor cantidad de polen viable. Una vez que se recolectaron las flores, se pusieron para su conservación en campo, en un pequeño conservador o aislante térmico, y de esta forma, tanto la flor como el polen se conservaron de mejor manera. Una vez recolectadas las flores, se procedió a la eliminación de todas las partes florales inservibles para el objetivo, dejando solamente los estambres, los cuales se colocaron en un recipiente abierto en su parte superior, con el fin de que las anteras abrieran y liberaran el polen. Una vez que las anteras presentaron un color amarillo más oscuro y al tocar éstas, las yemas de los dedos quedaban de color amarillo, se traspasaron a un tubo de color negro, para visualizar mejor la liberación del polen, en el cual se llevaron al huerto de mandarinas clementinas cv. Clemenules. b) Polinización de las flores: Para esto se tomaron flores en estado de botón floral (estadio 59), las cuales presentaban los pétalos cerrados y con una forma de bola alargada y hueca. Al momento de polinizarlas, se procedió a abrir el botón, para lo 35 cual se apretaba en la parte media a proximal de éste, emasculándolos con una pinza quirúrgica y polinizándolos con un pincel de pelo de camello Nº5 (Figura 3), y posteriormente se volvía a encerrar la flor con la malla de tul (Figura 4). c) Marcaje: Se hizo mediante el uso de una cinta de color amarillo sobresaliente, para facilitar la ubicación de los tratamientos, la cual fue amarrada con un alambre de cobre fino y dúctil, forrado en plástico, lo que facilitó su manejo en cuanto al realizar la sujeción de la bolsa a árbol sin provocar daño a este. c.1) Producto: Se abrevió como P, siendo asignadas de la siguiente forma: P1: oxicloruro de cobre. P2: sulfato de cobre. P3: óxido cuproso. P4: cloruro de cobre. La aplicación se realizó asperjando el producto directamente al frutito cuajado, procurando que el frutito en su totalidad quedara mojado. Las características de cada producto, correspondientes a sales de cobre se presentan en el siguiente cuadro: 36 CUADRO 2. Características químicas de las sales de cobre evaluadas. Sal de cobre Formula química Pureza Peso molecular Oxicloruro de CuCl2x3Cu(OH)2 87% p/p con un 13% 427,13 p/p cobre de ingredientes inertes. Sulfato de cobre Cu SO4*5H2O 98,0/99,0% como Cu 249,68 SO4*5H2O (Pentahidratado) y 24,5% como Cu metálico. Oxido cuproso Cu2O 60% como Cu2O y 50% 143,08 como Cu metálico. Cloruro de cobre CuCl2 98% de Cu metálico. 134,5 c.2) Fecha: Los productos fueron aplicados en las siguientes cuatro fechas: 15, 30, 45 ddpf y al término de la segunda caída fisiológica, siendo abreviadas como: F1: 15 días después de plena flor. F2: 30 días después de plena flor. F3: 45 días después de plena flor. F4: Término de la segunda caída fisiológica. c.3) Situación de la flor: Se refiere al estado del botón floral, lo que se relaciona con la presencia o ausencia de estambres y polen. Los diferentes grupos florales presentes en los ensayos fueron: GE/P: Flores emasculadas y polinizadas con polen del cv. Fortune. GSE/SP: Flores sin emasculación y sin polinización. GE/SP: Flores emasculadas y sin polinización. 37 Estambres con polen FIGURA 1. Botón floral de mandarina clementina cv. Clemenules. FIGURA 3. Polinización manual, con polen de mandarina cv. Fortune, en un botón floral de clementina cv. Clemenules. FIGURA 2. Polen del híbrido Fortune, al momento de la polinización. FIGURA 4. Botón floral de clementina cv. Clemenules, con malla para evitar la visita de abejas. 38 3.7. Estados fenológicos en que se realizaron las labores: Fue de vital importancia conocer el estado fenológico en que se encontraba el árbol al momento de realizar cada labor, propia de la investigación, ya que de esta manera es más fácil relacionar los resultados de cada tratamiento, y poder llegar determinar los efectos de estos mismos. Como apoyo se recurrió a la Escala BBCH para la descripción de los estadios fenológicos del desarrollo de los agrios (Género Citrus) (AGUSTÍ et al., 1995), (Anexo 1) a través de los códigos expresados en esta escala. CUADRO 3. Eventos fenológicos con sus respectivas labores. Labor Codificación escala BBCH. Embolsado de botón floral Estadios 56, 59 y menor grado 55 Polinización y emasculación Estadios 56, y en mayor grado 59 Aplicación de 15 ddpf Estadios 67, 69 y en menor grado 61 Aplicación de 30 ddpf Estadios 69, 71, 72, en menor grado 67, y 73 Aplicación de 45 ddpf Estadios 71,72 y en menor grado 73 Aplicación de término 2ª caída Estadios 74 y 79 Recolección frutos Estadios 81 y en menor grado 83 3.8. Determinación del término de la segunda caída fisiológica: Para determinar el término de la segunda caída fisiológica o de frutitos se basó en antecedentes bibliográficos, y también se hizo una observación a nivel de campo para determinar con una mayor exactitud el término de esta segunda caída fisiológica. Según AGUSTI (2000), el término de la segunda caída fisiológica sería 90 ddpf, por lo que para esta zona sería aproximadamente a mediados de enero. PUCHADES (2002) cita que la caída de capullos florales, de flores y de frutos en desarrollo se 39 hayan solapadas en el tiempo, su inicio y duración varían con los años y su intensidad global determina el porcentaje de frutos cuajados. Según lo anterior, la caída de diciembre (caída de junio para el hemisferio Norte), debería comenzar a fines de noviembre o en la primera quincena de diciembre, lo que se ratifica con la visualización de los primeros frutitos abortados el día 27 de noviembre en el huerto en donde se realizó la investigación, y el término de ésta es de difícil detección, según lo descrito anteriormente. Para tener un mejor criterio en determinar el término de la segunda caída fisiológica, se realizó una observación a nivel de campo, para poder determinar el momento cuando el árbol ya no abortara más frutitos. Lo anterior se logró por medio de la implementación de bolsas de plástico negras, debajo de la canopia de cinco árboles elegidos al azar, dentro de las doce hileras que contenían los árboles con los tratamientos. Se partió en el mes de diciembre realizando observaciones tres veces por semana, donde se iba viendo la magnitud de la caída de frutitos, hasta determinar el momento en que esta caída había llegado a su fin. Esto fue ocurriendo paulatinamente y cada vez se observaba una menor magnitud del aborto de frutitos, hasta que el día 19 de enero del 2004 fue cuando ya esta caída había disminuido en magnitud, ya que aun en algunos árboles, que no presentaban bolsas, se observaba la caída de frutitos pero en muy menor grado, por lo que se determinó el cese o término de la segunda caída fisiológica. Una vez determinado el fin de esta segunda caída se procedió a hacer la última aplicación de productos los días 20 y 21 de enero del 2004. 40 3.9. Diseño experimental: El diseño estadístico corresponde a un diseño completamente al azar (DCA) con arreglo factorial y distintas repeticiones, debido al aborto de frutitos, lo que hace que las 50 repeticiones iniciales se pierdan para cada tratamiento. El total de tratamientos es de 40, lo cual resulta de los diferentes factores en evaluación, siendo estos los siguientes: (4 Productos × 4 Fechas × 2 Condiciones de flor) + 8 controles = 40 Tratamientos. Con respecto al modelo matemático del DCA, este obedece a lo siguiente: Yi j = µi j + τ + ε Donde: Yi j: Valor observado en cada unidad experimental: número de semillas por fruto; calibre. µi j: Efecto de la media general sobre cada observación: gran media de número de semillas por fruto; gran media del calibre. τ: Efecto del tratamiento sobre cada observación. ε: Efecto del error experimental aleatorio sobre cada observación. Por intermedio de éste se podrá determinar si existe, al menos, una media de algún tratamiento que sea diferente a las demás, y de existir alguna diferencia significativa se realizará una separación de medias con el Test Tukey al 5%. 41 4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS 4.1. Análisis del efecto polinicida expresado en el número de semillas de cada fruto: 4.1.1. Influencia de la presencia de estambres y polen en los tratamientos Al presentar los datos del número de semillas para los estambres y el polen, se desprende que es el polen el que determina la presencia de semillas en los frutos de cada tratamiento, y no los estambres, por presentar los tratamientos con polen el mayor número de semillas (Cuadro 4 y Cuadro 5). CUADRO 4. Número de semillas en frutos de mandarina clementina cv. Clemenules para los tratamientos con o sin estambres sometidos a cuatro tratamientos gametocidas en base a cobre. Tratamientos Medias del Nº Semillas Diferencia Significativa Sin estambres 10,41 Con estambres 18,50 a b Letras iguales en la misma columna indica que no existe diferencia significativa entre tratamientos, según prueba de tukey (p= 0.05) CUADRO 5. Número de semillas en frutos de mandarina clementina cv. Clemenules para los tratamientos con o sin polen sometidos a cuatro tratamientos gametocidas en base a cobre. Tratamientos Medias del Nº Semillas Diferencia Significativa Sin polen 2,49 Con polen 31,40 a b Letras iguales en la misma columna indica que no existe diferencia significativa entre tratamientos, según prueba de tukey (p= 0.05) 42 En los cuadros 4 y 5 se puede observar que existe diferencia entre los ensayos que fueron emasculados y a los que no se emasculó, y entre los polinizados y los que no lo fueron. Esta diferencia observada para ambas condiciones florales, se debe exclusivamente a la presencia de polen en los ensayos. Hay que tener en cuenta al hacer el análisis de estos resultados, que las condiciones florales presentes en los diferentes ensayos eran: GE/P, GSE/SP y GE/SP. De éstas, las condiciones GE/P y GE/SP, ambas sin aplicación de productos, sirvieron como control para los demás tratamientos, en las cuatro fechas evaluadas. Para el caso de la condición GE/SP el número de semillas obtenido fue de cero, a diferencia de la condición GE/P, los que presentaron promedios elevados del número de semillas, tal como se observa en los siguientes cuadros (Cuadro 6 y Cuadro7). CUADRO 6. Número de semillas presentes en frutos de mandarina clementina cv. Clemenules de tratamientos con emasculación, sin polinización, y sin aplicación de productos gametocidas en base a cobre. Tratamientos Nº Semillas Diferencia Significativa GE/SP F1 0 a GE/SP F2 0 a GE/SP F3 0 a GE/SP F4 0 a Letras iguales en la misma columna indica que no existe diferencia significativa entre tratamientos, según prueba de tukey (p= 0.05) Para los tratamientos presentados en el cuadro 6, con flores emasculadas pero no polinizadas, se observa que el número de semillas es igual a cero, esto refleja que al no existir polen, proveniente de polinización cruzada o dirigida, no hay producción de semillas en el fruto. 43 Lo anterior esta dado por la auto incompatibilidad presente en las clementinas, lo cual significa que no se pueden autofecundar, debido a la esterilidad homogenética que poseen, en donde su polen no puede fecundar sus propias flores, resultando en una auto incompatibilidad (AGUSTÍ, 2004), pero si puede obtener semillas por medio de la polinización cruzada debido a que las clementinas tienen un alto porcentaje de polen y óvulos fértiles (PUCHADES, 2002). CUADRO 7. Número de semillas presentes en frutos de mandarina clementina cv. Clemenules de tratamientos con emasculación, polinización, y sin aplicación de productos gametocidas en base a cobre. Tratamientos Nº Semillas Diferencia Significativa GE/P F4 32,7 a GE/P F2 33,5 b GE/P F1 33,6 b GE/P F3 34,8 c Letras iguales en la misma columna indica que no existe diferencia significativa entre tratamientos, según prueba de tukey (p= 0.05) Los datos obtenidos en el cuadro 7 demuestran la capacidad como productor de polen del mandarino cv. Fortune, al mandarino clementino cv. Clemenules, y como controles de los tratamientos de las diferentes sales de cobre evaluadas, se puede determinar que bajo polinización cruzada las mandarinas clementinas producen semillas, lo que concuerda con ZARAGOZA et al. (2005), que citan que las variedades híbridas, Nova, Fortune y Ortanique inducen la formación de semillas en los frutos de otros híbridos y en Clementinos y Satsumas, lo que fundamenta el estudio del posible efecto polinicida de las sales de cobre en frutitos previamente polinizados con polen del cv. Fortune, lo que ratifica el elevado número de semillas obtenido para este caso, en que se realizó polinización y emasculación, y no se aplicó ningún producto. 44 BONO et al. (2004) agrega que no se produce fecundación, y por consiguiente formación de semillas, en una misma variedad (auto incompatibilidad) y tampoco entre las distintas variedades del grupo, produciéndose el fenómeno inter incompatibilidad. Ahora cuando polinizan o son polinizadas por variedades de otros grupos (a excepción del grupo Navel y en menor medida el grupo Satsuma), se producen semillas lo cual concuerda completamente con los resultados obtenidos, ya que los tratamientos polinizados con polen de Fortune presentaron mayores número de semillas. Por su parte SOLER et al. (1996), en estudios realizados sobre diferentes variedades de cítricos, en cuanto a su comportamiento frente a al polinización cruzada, llegaron a determinar la efectividad del cv. Fortune como donador de polen para el cv. Clemenules, obteniendo un número de 24 semillas/fruto, después de dos años de trabajo. Esto demuestra que los tratamientos que fueron polinizados, los que a su vez corresponde a las mismas flores sin estambres, son las que presentaron un mayor número de semillas por fruto, concordando con los trabajos anteriormente citados, y demostrando la eficacia del cv. Fortune como polinizante del cv. Clemenules. Esta media obtenida por SOLER et al. (1996) de 24 semillas/fruto es menor a la de esta investigación (31,4 semillas/fruto), lo cual puede ser atribuirle a las condiciones climáticas, ya que según AGUSTÍ (2004) la partenocarpia es afectada por las condiciones medioambientales, y hay cultivares que en ciertas zonas, como España, no han presentado semillas, pero si en otras zonas han presentado una débil partenocarpia, aunque esto sólo es un indicio que debe ser mas estudiado. GONZÁLEZ-SICILIA (1968) y MONSELISE (1986), citan que cuando se polinizan variedades que carecen de semillas o que las producen en un bajo número, como el caso de las clementinas, se genera un notable incremento de las semillas, lo que es observado en los resultados de ensayos polinizados. 45 También hay que destacar que los tratamientos sometidos a polinización tuvieron una menor abscisión de frutitos en comparación con los sin polinización, lo que se debe a que clementina presenta una gran caída de estructuras reproductivas (GOLDSCHMIDT y MONSELISE, 1977), y por otra parte está la mayor cantidad de fitohormonas sintetizadas, como citoquininas, las que son sintetizadas en el momento que ocurre la fertilización de los óvulos (HERNANDEZ y PRIMO-MILLO, 1990). Según MATILLA (2000) la síntesis de citoquininas esta relacionada con la mitosis intensa ocurrida después de la fecundación, siendo sintetizada en el suspensor, o también citan que pueden ser transportadas desde las raíces, no siendo claro su origen. Por otra parte PUCHADES (2002), con datos de dos años en ensayos de polinización realizados con polen de mandarina clementina cv. Clemenules, sobre flores del cv. Fortune, determinó que los porcentajes de flores que quedaron en el árbol y que no abortaron, fueron mayores en los ensayos de polinización cruzada (53% y 80%); seguidos de la autopolinización manual (20% y 8%) y muy bajos en las flores sin aporte de polen por emasculación (0,5% y 1%) o embolsadas (4% y 0%), lo que explica la mayor abscisión por parte de los frutos de los ensayos sin polinización, quedando un total de 1000, de las repeticiones iniciales, para los ensayos emasculados y polinizados, y un total de 800 para los ensayos sin emascular y sin polinización. Esta mayor abscisión por parte de los frutos de los tratamientos sin polinización, se debe a la ausencia de fitohormonas promotoras del desarrollo que se encuentran en las semillas, ya que el proceso de abscisión está mediatizado por los reguladores del crecimiento que pueden actuar de maneras distintas, o bien como inhibidores de abscisión (por ejemplo auxinas) o estimulantes (como el ABA y el etileno). La participación directa de otras fitohormonas como las giberelinas o las citoquininas no 46 es evidente, aunque parece que podrían actuar indirectamente aportando nutrientes o regulando el desarrollo de los órganos (ADDICOTT, 1982). Algunos autores han propuesto que las giberelinas podrían contrarrestar los efectos del ABA o del etileno en frutos de cítricos (ZACARÍAS et al., 1995) La diferencia obtenida en estos resultados, y la mayor cuaja por parte de los ensayos polinizados concuerda con los datos entregados por DELHOM y PRIMO-MILLO (1989), en que citan que algunas especies auto incompatibles de cítricos, muestran baja productividad, y la polinización cruzada con polen compatible da lugar a frutos con semillas que cuajan en gran proporción, que es el caso del cv. Clemenules polinizada con el polen del cv. Fortune. CHANDLER (1962) y PRALORAN (1977), manifiestan que en cítricos se presentan fenómenos de auto incompatibilidad, sobretodo en mandarino clementino, y ello ocurre aunque su polen y sus óvulos sean funcionales. Una razón a éste fenómeno, se atribuye a la excesiva longitud del pistilo, por lo que el tubo polínico no es capaz de alcanzar lo óvulos a tiempo para fecundarlos, esto explica porque en los ensayos que fueron polinizados con polen del cv. Fortune presentan las mayores medias en el número de semillas, y también deja claro que el cv. Clemenules es una variedad auto incompatible, que no se puede autopolinizar, por lo cual los ensayos con estambres y sin polen, presentaron el menor número de semillas, en que todos los tratamientos fue de cero, salvo unas tres excepciones, las que se deben a una falla en la malla que protegía a la flor, por lo que estas flores deben haber estado en contacto con abejas. Lo anterior se confirma con recientes trabajos realizado por MUÑOZ, AYUSO y LABRADOR (2005), citan que el grupo de las clementinas, al cual pertenece el cv. Clemenules, presentan partenocarpia, pero parece ser que la actividad de los polinizadores aumenta la producción. Por otra parte, señalan que las clementinas son autoestériles por lo que sólo formarán semillas en presencia de fuentes de polen 47 viable de otras variedades en las proximidades. PUCHADES (2002) es menos rígido con respecto al término autoesterilidad, indicando que este término también se puede denominar auto incompatibilidad o incapacidad para formar embriones zigóticos mediante la autopolinización. 4.1.2. Influencia de los productos y fechas, y su posible efecto polinicida medido en el número de semillas Referente a los productos y fechas, se presenta a continuación los resultados obtenidos en los diferentes tratamientos, correspondientes a la evaluación de la aplicación de cuatro sales de cobre en cuatro fechas (Cuadro 8). CUADRO 8. Número de semillas por fruto de mandarina clementina cv. Clemenules para cuatro tratamientos polinicidas en base a cobre. Producto Nº Semillas Diferencia Significativa Producto 1 (Oxicloruro de cobre) 13,99 b Producto 2 (Sulfato de cobre) 13,62 b Producto 3 (Oxido cuproso) 12,14 a Producto 4 (Cloruro de cobre) 11,38 a Sin Producto 21,12 c Letras iguales en la misma columna indica que no existe diferencia significativa entre tratamientos, según prueba de tukey (p= 0.05) Se observa que entre los cuatro productos evaluados, no hubo ninguno que tuviera el efecto polinicida deseado, es decir, que ninguno fue capaz de producir frutos totalmente partenocárpicos. De los cuatro productos, el cloruro de cobre y el óxido cuproso, no presentan diferencias significativas (Cuadro 8), presentando a su vez, el número más bajo de semillas por fruto. Si bien es cierto que no presentaron la 48 capacidad de eliminar las semillas originadas por la polinización manual, se ve una diferencia significativa con sulfato de cobre y oxicloruro de cobre. Esta diferencia del cloruro de cobre y el óxido cuproso con el sulfato de cobre y oxicloruro de cobre, se puede deber en un parte a que el cloruro de cobre es el que presenta una mayor riqueza de cobre, entre los cuatro productos, la que equivale a un 98% de cobre metálico, en comparación con los otros productos, los que presentan riquezas menores (AFIPA, 2003). Por otra parte, el óxido cuproso es una de las formas mas activas del cobre como fungicida, lo cual le confiere una alta eficiencia, la que se basa en el tamaño muy fino de las partículas del óxido cuproso, lo cual le permita alcanzar una mayor superficie de contacto, una mejor distribución del cobre y una mejor adherencia en la superficie (AFIPA, 1998). Otra razón, se puede deber a que el sulfato de cobre al momento de realizar las soluciones, fue el que presentó una mayor dificultad para disolverse en el agua, quedando un precipitado azul en la solución, lo que puede haber afectado su aplicación a los frutitos, no llegando a estos la cantidad necesaria de producto. Pero estos resultados se contraponen a los obtenidos por MESEJO et al. (2005), en donde según sus investigaciones, el sulfato de cobre fue el que presento un mayor efecto polinicida en pruebas in vitro, al igual que en las pruebas in vivo. A pesar de lo anterior, los fundamentos químicos antes dados, pueden tener alguna razón al momento de buscar las diferencias estadísticas presentadas entre los diferentes productos evaluados, aunque ninguno presentó la capacidad de formar frutos partenocárpicos, con la dosis de 20 ppm utilizada. A continuación se presentan los resultados obtenidos para cada fecha evaluada (Cuadro 9). 49 CUADRO 9. Número de semillas por fruto para cuatro fechas en mandarina clementina cv. Clemenules sometidos a cuatro tratamientos polinicidas en base a cobre. Fecha Nº Semillas Diferencia Significativa Fecha 1 (15 ddpf) 11,06 a Fecha 2 (30 ddpf) 12,77 Fecha 3 (45 ddpf) 16,88 c Fecha 4 (Término de 2da caída 17,09 c b fisiológica) Letras iguales en la misma columna indica que no existe diferencia significativa entre tratamientos, según prueba de tukey (p= 0.05) Referente a las fechas evaluadas, AGUSTÍ (2005)* menciona que estos productos actúan impidiendo la germinación del grano de polen y/o el desarrollo del tubo polínico, por lo que su efecto se logra si son aplicados antes que la flor sea polinizada, y según MESEJO et al. (2005) el mejor momento para realizar el tratamiento es con un 60% de la flores en antesis, con una dosis de 25 mg l-1. Teniendo en cuenta esto, se debe analizar que la aplicación de todos los productos evaluados se llevaron a cabo después de que las flores fueran polinizadas, por lo que los productos no pudieron lograr su efecto de impedir la germinación del polen o interrumpir el desarrollo del tubo polínico, debido a que el tiempo que paso entre la polinización de las flores y la primera aplicación de 15 ddpf, el polen ya había germinado y fecundado a los óvulos de la flor, produciendo semillas. _________________________ AGUSTÍ, M. Doctor. Ing. Agr. 2005. Profesor de Citricultura y Fruticultura de la Universidad Politécnica de Valencia. Valencia, España. Comunicación personal. 50 Por su parte, SPIEGEL-ROY y GOLDSCHMIDT (1996) dicen que la fecundación de la oosfera ocurre dos o tres días después de la polinización bajo condiciones favorables, pero en algunos casos se ha informado un lapso de tres a cuatro semanas, por lo que desde el momento en que se polinizaron las flores hasta que se aplicaron los productos, el polen ya había germinado, lo que queda demostrado con la gran cantidad de semillas presentes en los frutos que fueron polinizados. MESEJO et al. (2005) determinaron en sus ensayos de campo que dosis de 25 mg l-1 o superiores tenían efecto en reducir el número de semillas en frutos que habían sido polinizados. Esto, en comparación con la dosis de 20 mg l-1 utilizada en esta investigación demuestra que ésta pudo haber sido una dosis de una concentración baja, debido a que no tuvo el efecto polinicida buscado para producir frutos partenocárpicos. El que los productos no hayan tenido el efecto buscado se puede deber principalmente a la fecha de aplicación de estos, la cual fue muy tardía para lograr el efecto polinicida, ya que según resultados obtenidos por MESEJO et al. (2005), la aplicación de Cu SO4 * 5H2O solo tuvo el efecto polinicida en condiciones in vivo, cuando el producto fue aplicado antes de la polinización manual de las flores, y no después de 24 h de polinizadas. El momento de aplicación de estos productos con características gametocidas es sumamente importante, y pareciera que el éxito en lograr la muerte de los gametos masculinos radica principalmente en este factor por sobre otros, como la dosis a utilizar. Con respecto a esto BINGHAM y LUPTON (1987) se expresan en términos similares, indicando que la clave para realizar la aplicación se relaciona a su vez, con el tiempo de receptividad de la parte femenina, lo que coincide con lo expuesto por Mc RAE (1985) y EDWARDS (1986), que indican que una desventaja de estos 51 productos con capacidad gametocida, es su reducida ventana de aplicación, lo que disminuye su uso práctico. Este inconveniente se incrementa debido al modo de acción de las sales de cobre, las que actúan impidiendo la germinación del grano de polen y/o el desarrollo del tubo polínico, por lo que su efecto pasa a ser más bien de contacto. SOLER (2005)* agrega que el efecto de las sales de cobre pasan por tener un efecto más bien cáustico sobre el grano de polen y/o el tubo polínico, por lo que la aplicación debe ser en el momento adecuado y con un buen mojamiento. En estudios realizados por GAJU (2002) en trigo, se observó que el mejor resultado para provocar una macho esterilidad (MS) del 96%, (que según WONG, BLOUET y GUCKERT (1995) considera que un 95% de MS es suficiente para la producción de semilla híbrida comercial), fue en la segunda fecha de aplicación entre dos evaluadas, y que correspondió a plantas en 8,5 grados en la Escala de Feekes, las cuales presentaban tres nudos visibles sobre la superficie, la hoja bandera parcialmente desplegada y las espigas tenían 1,76 cm. de promedio, pero aún el proceso de floración no comenzaba, lo cual confirma que la aplicación de un producto polinicida o gametocida se debe realizar en un estado anterior a que el polen entre en contacto con el estigma de la flor, razón por la que los cuatro productos de esta investigación no tuvieron el efecto polinicida, ya que el polen ya había germinado y fecundado al ovulo de la flor. ____________________ *SOLER, J. Ing.Téc.Agr. 2005. Investigador IVIA y profesor Master En Citricultura de la Universidad Politécnica de Valencia. Valencia, España. 2005. Comunicación personal. 52 Por su parte LAL et al. (2005) determinaron en sus estudios que el compuesto etilo 4fluoroxanilato aumentó de un 20,03% a un 55,55% la esterilidad masculina en soja, en una dosis de 1000 ppm, y también concluyen que este porcentaje se puede aumentar si la aplicación se realiza en una fecha adecuada. VAN DER MEER y VAN BENNEKOM (1975) concluyen que dosis alta de ácido giberélico (2%) actúa como un efectivo polinicida, es decir, que provoca un esterilidad masculina, reemplazando a la emasculación manual en cebolla, esto sugiere que la dosis también debería ser mayor para lograr el efecto polinicida en las sales de cobre evaluadas, lo que se confirma con MESEJO et al. (2005) que determinaron que una dosis de 25 ppm provocaba un buena capacidad polinicida del sulfato de cobre, tanto aplicado a flores individuales como al árbol de mandarino clementino, dosis ésta más alta que la de 20 ppm utilizada en este estudio. Lo anterior concuerda con el trabajo realizado por JAN y ROWELL (1981) en un estudio de tres compuestos con capacidad gametocida (Ethrel, RH-532 y RH-2956) realizados en trigo, determinaron que el compuesto RH-2956 fue el más eficaz agente gametocida en causar esterilidad masculina (polinicida) a la dosis más alta ensayada, y que Ethrel sólo fue efectivo en las aplicaciones realizadas en los estados de menos desarrollo del trigo, y no en los mas avanzados cuando el polen ya tenía la capacidad de fecundar al los óvulos. Estudios en soja demostraron que el compuesto etilo 4-fluoroxanilato es el que mejor resultado ha provocado en producir esterilidad masculina (ALI et al., 1999), en dosis de 1000 ppm (MATHUR y LAL, 1999), dosis altas que demuestran que tienen una mayor efectividad en su acción polinicida o gametocida, por lo que la dosis usada en este estudio, también podría tener alguna incidencia, aunque no tan alta como la fecha de aplicación, en que los productos no tuvieron el efecto polinicida. Mc RAE (1985) y EDWARS (1986) citan que los productos gametocidas requieren de altas dosis para 53 lograr el efecto buscado, aunque estas altas dosis pueden afectar la fertilidad femenina o reducir el período de receptividad de la hembra, agregando que por lo general no son muy selectivos. Otros autores como BATCH et al. (1980), citan productos como derivados del etileno, derivados del arsénico y hormonas del crecimiento, como agentes polinicidas en soja, siendo dentro de las hormonas de crecimiento, el ácido indolacético (AIA) del que se tienen referencias de laboratorio (KWAN, HAMSON y CAMPBELL, 1969; GEITMANN, 1997). Según los datos entregados por estos autores, el AIA tiene un efecto de inhibir la germinación del polen y el crecimiento del tubo polínico con concentraciones altas del producto por sobre los 100 µM, ya que con concentraciones inferiores a 10 µM, el AIA tendría un efecto positivo para el crecimiento del polen, lo cual estaría indicando que la utilización de mayores concentraciones de sales de cobre, debería tener un mayor efecto polinicida, ya que aunque son compuestos de naturaleza distintas, ambos, al parecer, actúan de la misma forma sobre el polen. GEITMANN, (1999) en estudios realizados indica que concentraciones crecientes de ácido cítrico hasta 20 mg l-1, nitrato cálcico hasta 15 mg l-1 y de ácido indolacético hasta 50 mg l-1 reduce significativamente la germinación, hasta llegar anularla, como también impedir el desarrollo del tubo polínico. La utilización de ácidos tricarboxílicos, como el ya mencionado ácido cítrico, se basa en que estos productos también impiden la germinación del polen y el desarrollo del tubo polínico (KWAN, HAMSON y CAMPBELL, 1969), aunque MOGENSEN y LADYMAN, (1989) opinan que afecta en mayor parte la funcionalidad del grano de polen. El ensayar con mayores dosis de las sales de cobre sería una buena alternativa, ya que de haberse empleado una dosis mayor a las 20 ppm utilizadas en esta investigación, la 54 cantidad de semillas podría haber sido relativamente menor, coincidiendo con MESEJO et al. (2005), que probaron mayor dosis, equivalente a 25 ppm. También sería necesario saber si las sales de cobre afectan de igual manera a los granos de polen que aún no han sido liberados en las anteras, teniendo presente que tanto las clementinas como los híbridos actúan como dadores de polen, además de ser receptores y presentar semillas en sus frutos, ya que al momento de la aplicación del producto en condiciones de campo, la flor posee los granos de polen de sus estambres y los granos de polen que han sido depositado en su estigma, por lo que de actuar así se impediría que el polen que está en las anteras degenere y no polinice a otras flores de cítricos, para lo que tendría que actuar a nivel de la gametogénesis o inhibiendo la formación de anteras, como el producto LY195259 (TSCHABOLD et al., 1988). Para los productos y las fechas se pueden observar datos muy dispersos, lo que se debe al resultado erróneo obtenido para cada producto y fecha, o también al escaso efecto de éstos para producir frutos sin semillas, en cada repetición se obtuvo un numero considerable de semillas, indicando el nulo efecto de cada producto al igual de cada fecha evaluada con el fin de obtener frutos partenocárpicos, debido principalmente a que la fecha de aplicación de los productos no fue la adecuada. A lo anterior estaría el estudiar sobre productos con efecto sistémico como el denominado Clofencet, utilizado en hibridación de trigo, aunque se tiene poca información de su forma de acción (NESVADBA y VYHNÁNEK, 2001), pero se cree que su efecto sería que las microsporas abortan previo a la vacuolización (UAUY, 2001). Hay que tener en cuenta que dentro de los productos el mayor acercamiento a un posible efecto polinicida se obtiene en las dos primeras fechas de aplicación, siendo difícil de determinar bien el momento exacto, debido a los diferentes estados de 55 desarrollo en que se encuentran los flores en el árbol, lo cual se debe a que la floración en el cv. Clemenules se presenta larga y con un gran traslape de estados de desarrollo de sus estructuras reproductivas (ZARAGOZA, 2005)*. Debido a lo largo de la floración del cv. Clemenules y al traslape de estructuras que esto produce (SOLER, 1999), es difícil de detectar a nivel de campo, el momento óptimo para la aplicación de estas sales de cobre, o de cualquier compuesto que tenga alguna capacidad polinicida o gametocida. La mejor solución para este inconveniente es realizar varias aplicaciones en floración, como una manera de buscar un instante de la planta en que estos productos puedan surtir efecto y determinar el mejor momento de aplicación, teniendo en cuenta las condiciones de cada huerto y zona en particular (AGUSTÍ, 2005)*. Todos los ensayos que fueron polinizados y con aplicación de cualquiera de los cuatro productos, presentaron semillas, lo que indica el nulo efecto de éstos para formar frutos partenocárpicos (Figura 5). ___________________________ AGUSTÍ, M. Doctor. Ing. Agr. 2005. Profesor de Citricultura y Fruticultura de la Universidad Politécnica de Valencia. Valencia, España. Comunicación personal. ________________________ ZARAGOZA, S. Ing. Agr. 2005. Investigador del IVIA (Instituto Valenciano de Investigaciones Agrarias) y profesor del Master de Citricultura de la Universidad Politécnica de Valencia. Valencia, España. Comunicación personal. 56 OXICLORURO DE COBRE GE/P F2 P1 SULFATO DE COBRE GE/P F2 P2 FIGURA 5. Frutos de clementina cv. Clemenules, de diferentes tratamientos con cobre como producto polinicida, en donde se visualiza la presencia de semillas, situación que se da para todos los productos ensayados. OXIDO CUPROSO 54 mm GSE/SP F1P3 60 mm GE/P F1P3 FIGURA 6. Frutos de mandarina clementina cv. Clemenules, tratadas con el mismo producto polinicida y en igual fecha, diferenciándose en la presencia de polen. 57 4.2. Análisis del calibre de los frutos: 4.2.1. Influencia de la presencia de estambres y polen en los ensayos Se presentan los siguientes cuadros para realizar el análisis de estos dos factores, y su incidencia en el calibre final de los frutos (Cuadro 10 y Cuadro 11). CUADRO 10. Calibre (mm) de frutos para los tratamientos gametocidas en base a cobre con o sin estambres. Tratamientos Calibre Diferencia (mm) Significativa Sin estambres 57,75 a Con estambres 60,15 b Letras iguales en la misma columna indica que no existe diferencia significativa entre tratamientos, según prueba de tukey (p= 0.05) CUADRO 11. Calibre (mm) de frutos para los tratamientos gametocidas en base a cobre con o sin polen. Tratamientos Calibre Diferencia (mm) Significativa Sin polen 51,17 a Con polen 66,72 b Letras iguales en la misma columna indica que no existe diferencia significativa entre tratamientos, según prueba de tukey (p= 0.05) En este caso se puede observar que para los ensayos polinizados y emasculados (flores del grupo GE/P) en los ensayos con polen, se obtuvo la mayor media para el calibre en comparación con los ensayos sin emasculación y sin polinización (flores del grupo GSE/SP), que corresponden a los ensayos sin polen. Este mayor calibre por parte de los ensayos con polen se debe a la presencia de semillas debido a la 58 fecundación de los óvulos del ovario de la flor, producto de la polinización, la cual como define RALLO (1986), es el preludio de la fecundación, lo que conlleva a un gran incremento en el número de semillas por parte de los frutos de la mandarina clementina (GONZÁLEZ-SICILIA, 1968; MONSELISE, 1986). La presencia de las semillas de los ensayos polinizados es lo que le otorga a estos tratamientos un mayor calibre, debido a que cuando se produce polinización cruzada entre variedades compatibles, sobretodo con los híbridos, provoca un aumento en la producción, ya que hay un aumento importante de frutos cuajados, así como un aumento de su diámetro; todo ello acompañado por un aumento importantísimo del número de semillas por fruto (ZAURASHUILI, 1967). PUCHADES (2002) en un trabajo realizado para ver la influencia de la polinización en las características del híbrido Fortune, concluyo que la presencia de semillas, debido a la polinización cruzada, se relaciona con un mayor diámetro, el cual concuerda con BEN-CHEIKH et al. (1997) los que en ensayos realizados en naranjo semillado Pineapple, demostraron que la polinización aumentaba los niveles de giberelinas en los ovarios en desarrollo, como también disminuía la abscisión de éstos, lo que ocurrió con los ensayos sometidos a polinización, ya que ese mayor nivel de giberelinas en los ovarios en desarrollo fue lo que le dio el mayor potencial de crecimiento y por ende un mayor calibre final a estos frutos. PONS et al. (1995a), observaron un aumento en el tamaño del fruto relacionado linealmente con el contenido de semillas en polinización cruzada entre el cv. Fortune y el cv. Clemenules, lo que se relaciona con este trabajo. Los datos obtenidos en estos ensayos, determinan que bajo polinización cruzada una variedad auto incompatible como el cv. Clemenules, produce fruta con semillas, la que a su vez le otorga un mayor tamaño final a estos frutos, ya que la polinización es 59 esencial para la producción de semillas para el caso de variedades semilladas, o para estimular el crecimiento del ovario en variedades de débil partenocarpia, como el caso del cv. Clemenules (DAVIES y ALBRIGO, 1994). Por otra parte cuando se efectúa la autopolinización en una variedad auto incompatible, como el cv. Clemenules, no se producen semillas, ya que la auto incompatibilidad de esta variedad esta dada por la interrupción del crecimiento del tubo polínico en la parte superior del estilo (ETI y STOSSER, 1992), y aunque el estigma de la flor en las clementinas esté receptivo tres días antes de la antesis, aún así no se produce autofecundación, descartándose casos de cleistogamia (ROHIDAS y CHAKRAWAR, 1989). Esto resultados se relacionan y coinciden con los trabajos de ZARASHUILI (1967); WALLACE y LEE (1999); PUCHADES (2002) los cuales en sus trabajos demostraron que la polinización manual o cruzada, así como la aplicación de giberelinas exógenas aumenta el cuajado y el tamaño final en variedades de baja capacidad partenocárpica, como el caso de las clementinas, en que poseen una baja concentración de giberelinas endógenas (TALON, ZACARIAS y PRIMO-MILLO, 1992). También se puede observar que la media del calibre para los ensayos con polen es de 66, 72 mm, a diferencia de los ensayos sin polen en donde esta media es de 51,17 mm, por lo que las semillas le otorgan a los frutos un calibre ideal para su comercialización hacia el exterior, según los datos entregados por ORTÚZAR (1999), el que indica que para mandarinas e híbridos un diámetro ecuatorial entre 55-65 mm es apto para la exportación de la fruta, pero la presencia de semillas desvaloriza la calidad del fruto (PONS et al., 1995a), por lo que la solución a este problema es la aplicación de giberelinas exógenas, para mejorar el cuajado en mandarinas clementinas (TALON, ZACARIAS y PRIMO-MILLO, 1992), el rayado efectuado después del termino de la segunda caída fisiológica de frutitos (AGUSTÍ et al., 1998) 60 o la aplicación de auxinas de síntesis para aumentar el calibre final del fruto (AGUSTÍ, 2003). 4.2.2. Influencia de los productos y las fechas de ensayo, en el calibre final del fruto Se presentan los siguientes cuadros para realizar el análisis de estos dos factores, y su incidencia en el calibre final de los frutos (Cuadro 12 y Cuadro 13). CUADRO 12. Calibre (mm) de frutos de mandarina clementina cv. Clemenules en cuatro tratamientos de productos polinicidas. Producto Calibre Diferencia (mm) Significativa Producto 1 (Oxicloruro de cobre) 58,15 a Producto 2 (Sulfato de cobre) 57,93 a Producto 3 (Oxido cuproso) 58,34 a Producto 4 (Cloruro de cobre) 57,60 a Sin Producto 62,72 b Letras iguales en la misma columna indica que no existe diferencia significativa entre tratamientos, según prueba de tukey (p= 0.05) Los calibres obtenidos para los tratamientos no presentan diferencia, con la excepción de los ensayos sin productos, lo que se correlaciona con los datos observados para el número de semillas de estos tratamientos sin producto, que fueron los que presentaron el número de semillas más alto, por lo que este valor para el calibre concuerda con lo anterior, y se debe a que las semillas son las que estimulan el crecimiento del fruto por medio de la síntesis de sustancias de crecimiento, principalmente giberelinas (GAs) (DELHOM y PRIMO-MILLO, 1989), por lo que es de esperar que los ensayos sin producto presenten la media mas alta de calibre. 61 CUADRO 13. Calibre (mm) de frutos de mandarina clementina cv. Clemenules en cuatro fechas de aplicación de productos polinicidas en base a cobre. Fecha Calibre Diferencia (mm) Significativa Fecha 1 (15 ddpf) 57,86 a Fecha 2 (30 ddpf) 58,12 ab Fecha 3 (45 ddpf) 60,48 Fecha 4 (Término de 2da 59,33 c bc caída fisiológica) Letras iguales en la misma columna indica que no existe diferencia significativa entre tratamientos, según prueba de tukey (p= 0.05) En las fechas evaluadas, se observa que el menor número de semillas esta dada en la fecha 1, la que a su vez no presenta diferencia significativa con la fecha 2, lo cual también tiene cierta correlación con los datos analizados para el número de semillas, ya que la fecha 1 fue la que presento la menor media para el número de semillas, seguida de la fechas 2, 3 y 4, lo que tiene una estrecha relación con los datos obtenidos para el calibre. Esta correlación se da debido a que en las fechas que se obtuvieron el menor número de semillas (15 y 30 ddpf), lo que significa que dentro de las fechas evaluadas fueron las que presentaron las mejores condiciones para tratar de lograr el efecto polinicida, pero debido a este menor número de semillas por fruto se tiene un menor calibre también en estas fechas de aplicación (15 y 30 ddpf), teniendo una relación directa el mayor número de semillas con el calibre final del fruto, ya que según AGUSTÍ (2003) son las hormonas producidas por las semillas las que regulan el crecimiento del fruto. Esta relación directa entre el mayor número de semillas y el calibre final del fruto, se debe a que el cv. Clemenules se define como una variedad de partenocarpia débil 62 (GUARDIOLA, 1998) y de deficiente cuajado (MEHOUACHI, 2001), por lo que en ausencia de semillas son las paredes del ovario las que dan el estimulo de cuajar frutos partenocárpicos (NISTCH, 1970), que según HERNÁNDEZ y PRIMOMILLO (1990) corresponden principalmente a giberelinas y citoquininas, las que a su vez promueven la división celular. Por eso, la polinización cruzada aumenta el cuajado y el tamaño del fruto, observándose un aumento del tamaño del fruto relacionado linealmente con el contenido de semillas (PONS et al., 1995a), aunque resultan totalmente desechadas por la mayor parte de los consumidores o se venden a precios más bajos (BONO, SOLER y BUJ, 2000). Por medio de la polinización cruzada se fecundan los óvulos y se obtienen semillas, que son las responsables de la síntesis de giberelinas (MONSELISE, 1977), por eso que a mayor número de semillas mayor calibre, por lo que si los productos no tuvieron el efecto polinicida deseado, todos los ensayos polinizados presentarán medias de calibre mayores a los sin polinización, para todos los productos evaluados. Para cada producto de los ensayos polinizados, se presentan las dos primeras fechas como las con menor promedio de calibre, reafirmando los resultados obtenidos para las fechas evaluadas, en donde estas dos primeras fechas son las que presentaron la menor media para el número de semillas y como consecuencia esto trae un menor calibre, como ya se ha explicado, lo que se debe a la menor concentración de GAs, las que son importantes en la estimulación del crecimiento del fruto (TALON, ZACARIAS y PRIMO-MILLO, 1992). Las fechas ensayadas en esta investigación, al igual que los productos, no tuvieron el efecto polinicida, debido a lo tardío en que se realizaron las aplicaciones de los productos, lo que según MESEJO et al. (2005), se debe realizar en floración. Debido a esto se debe que los frutos de los ensayos con polen presentaran semillas, que en conjunto con los ensayos sin polen, dan medias de calibre que no presentan 63 diferencias significativas entre la fecha 1 y la fecha 2, la fecha 2 con la fecha 4 y la fecha 4 con la fecha 3. Esta igualdad de las medias de los calibres se relaciona a que para obtener la media se suman los ensayos polinizados y los no polinizados, y que la fecha 1 y 2 presenten diferencia significativa con las otras dos fechas, se debe a que en estas dos fechas se obtuvieron las medias menores para el número de semillas por fruto, como también para el calibre, debido a que las semillas regulan el crecimiento y desarrollo del fruto (AGUSTÍ, 2003), y las obtenidas por cruzamientos entre parentales diferentes las que le otorgan un mayor calibre al fruto, que es el caso de la polinización de las flores del mandarino clementino cv. Clemenules con el polen del híbrido Fortune (BUKIYA, 1990), lo que se aprecia en la Figura 6. El mayor calibre observado en el fruto polinizado (GE/P) se debe al nulo efecto presentado por las cuatro sales de cobre ensayadas, lo que se debió fundamentalmente, como ya se discutió anteriormente, a las fechas de aplicación evaluadas en esta investigación, las cuales fueron muy tardías, por lo que ya el polen había fecundado a los óvulos del ovario, produciendo semillas. Una fecha de aplicación anterior a que se produzca la fecundación de la flor debería presentar mejores resultados, lo cual es coincidente con el trabajo de GAJU (2002) y MESEJO et al. (2005). Según GOODWIN (1978) la regulación del crecimiento de los frutos depende en gran medida de las hormonas sintetizadas en las semillas, y de forma particular de las GAs. En muchas especies se ha demostrado que el número de frutos que cuajan, e incluso su tamaño individual correlaciona con el número de semillas que poseen. En los frutos sin semillas, la ausencia de polinización y de semillas detiene el crecimiento del fruto y provoca su abscisión, por lo que se entiende lo de la Figura 6, en donde se demuestra la diferencia de calibres entre dos frutos, diferencia dada por 64 la presencia de semillas en el fruto que fue polinizado, otorgándole una mayor concentración de hormonas que son las que finalmente estimulan su mayor desarrollo, a diferencia del fruto sin polinizar. En resumen, lo que se busca en las mandarinas clementinas es la obtención de fruta con un calibre apropiado y que sea partenocárpica, es decir, con ausencia de semillas. Si bien es cierto esta especie presenta una esterilidad gamética relativa, el cual se debe a fenómenos de auto incompatibilidad (BONO, SOLER y BUJ, 2000), al ser polinizada con el polen del cv. Fortune, se obtuvieron una gran cantidad de semillas por fruto, teniendo una media general de 31,4 semillas, lo que se presenta superior a las medias entregadas por ASÍNS et al. (2002), que son de 20 a 25 semillas por fruto. Por lo tanto, se puede decir que los mayores calibres se relacionan con la mayor presencia de semillas en los frutos, y por ende, por un efecto nulo de las sales de cobre en la obtención de frutos partenocárpicos en las fechas ensayadas, ya que la dosis está muy relacionada con otras concentraciones de productos entregados por autores como GEITMANN (1999), el cual da 20 ppm para ácido cítrico, 15 ppm para nitrato cálcico y de 50 ppm para el AIA, por lo que la dosis no es la responsable del nulo efecto como gentes polinicidas de las cuatro sales de cobre evaluadas, pero si las fechas como ya se ha discutido. Es necesario tener en cuenta el papel de otras sustancias que puedan tener algún efecto polinicida para próximas investigaciones, en donde su evaluación tanto en laboratorio como en condiciones de campo, se deben realizar aplicadamente, ya que algunos productos a concentraciones altas tienen un efecto distinto que a concentraciones más bajas, como el caso del AIA (GEITMANN, 1997), o como el ión Ca 2++ el que también presenta condiciones polinicidas (GONG, YANG y CAO, 1995), a pesar de resultar esencial para la germinación del polen. 65 5. CONCLUSIONES • No hubo efecto polinicida de ninguna sal cúprica ensayada. • De las cuatro sales de cobre evaluadas, el cloruro de cobre y el óxido cuproso, desarrollaron el menor número de semillas por fruto. • Ninguna de las fechas presentó el momento fenológico ideal para lograr frutos partenocárpicos. • El polen del mandarino cv. Fortune provoca la formación de semillas en los frutos del mandarino clementino cv. Clemenules, lo que conllevó a un mayor calibre final de estos frutos. 66 Taller de Licenciatura “EVALUACIÓN DE SALES DE COBRE PARA DETERMINAR SU CAPACIDAD POLINICIDA Y SU EFECTO SOBRE EL CALIBRE EN MANDARINA CLEMENTINA (Citrus clementina Hort. ex Tanaka) cv. CLEMENULES” 6. RESUMEN Uno de los grandes problemas de calidad que presenta, hoy en día, el cultivo del mandarino clementino es la presencia de semillas en sus frutos, situación que lo desvaloriza comercialmente. La presencia de semillas se debe a la polinización cruzada, aunque el mandarino clementino sea autoincompatible, y por lo tanto si se encuentra en una plantación monovarietal o alejado de fuentes de polen, no presenta semillas, sin embargo, si se presenta cerca de la plantación de alguna variedad cítrica capaz de ser fuente de polen, como puede ser el limonero (Citrus limón L.), las naranjas blancas o comunes (Citrus sinensis L.), o el grupo de los híbridos, como el caso del cv. Fortune, que es el que con mayor frecuencia ocasiona este problema, sus frutos presentan semillas. Como una opción para evitar la presencia de semillas en el fruto, aparece la capacidad gametocida o polinicida que poseen ciertos grupos químicos, la que esta referida a su capacidad de impedir la germinación del grano de polen y/o el desarrollo del tubo polínico, presentando el mejor resultado, según investigadores de la Universidad Politécnica de Valencia, el grupo de las sales de cobre, motivo por el cual, en esta investigación, se ensayaron cuatro productos cúpricos en una única dosis correspondiente a 20 ppm, siendo estos: oxicloruro de cobre, sulfato de cobre, óxido cuproso y cloruro de cobre. Estos productos se ensayaron en cuatro fechas distintas, que fueron 15, 30 y 45 ddpf y, al término de la segunda caída fisiológica, para ver cual era el mejor momento, para lograr el efecto polinicida. Para llevar a cabo los ensayos se utilizó al mandarino clementino cv. Clemenules, al que se les emascularon y polinizaron flores con polen del cv. Fortune, y a su vez se les dejaron flores sin polinización y sin emasculación, a las que se les aplicaron los cuatro productos evaluados para ver su efecto. Por otra parte, esta polinización sirvió para ver la incidencia del polen del cv. Fortune sobre producción de semillas y el calibre final, en los frutos de mandarina clementina cv. Clemenules. 67 Los resultados obtenidos fueron que ningún producto tuvo la capacidad polinicida buscada, en ninguna de las cuatro fechas, lo cual puede estar relacionado con las fechas, ya que los productos se aplicaron en un momento en que el polen ya había germinado y fecundado a los óvulos del ovario, por lo que los productos ya no tuvieron el efecto deseado. Finalmente, el polen del mandarino cv. Fortune produjo una gran cantidad de semillas en los frutos polinizados del cv. Clemenules, los que a su vez presentaron un mayor calibre y una menor abscisión que los frutos no polinizados. 68 7. ABSTRACT ASSESSMENT OF COPPER SALTS TO DETERMINE THEIR POLLENICIDAL CAPACITY AND EFFECT ON SIZE OF CLEMENTINE MANDARIN (Citrus clementina Hort. ex Tanaka) CV. “CLEMENULES” One of the largest quality concerns in clementine mandarin (Citrus clementina Hort. ex Tanaka) cv. “Clemenules” production is the presence of seeds in its fruits, which causes a commercial devaluation. The presence of seeds is due to cross-pollination since clementines are self-incompatible, and in an orchard whit just one variety or far from other pollen sources, there are no seeds. However, if the clementines are planted near other citrus varieties that can serves a sources of pollen, such as lemons (Citrus lemon L.), white or common oranges (Citrus sinensis L.), or hybrids like “Fortune”, which most often displays this problem, the fruit will contain seeds. To avoid the presence of seeds in the fruit, one option is the evaluation of the capacity of certain groups of chemicals as gametocides or pollenicides, referring to their ability to stop the germination of pollen grains or the development of pollen tubes. According to researchers from the Universidad Politécnica de Valencia, copper salts produce the best results. Four cupric products were tested in this study, at a dosage of 20 ppm: copper oxychloride, copper sulfate, cuprous oxide and copper chloride. These products were tested at four dates, 15, 30 and 45 days after full bloom and at the end of the second physiological fruit drop, to determine the optimal moment for achieving a pollenicidal effect. Clementine mandarins cv. “Clemenules” were used for these trials, in which the four products were applied to clementines whose flowers were emasculated and pollinated with pollen from “Fortune”, or were left without pollination and without emasculation. This pollination experiment was also useful in observing seed production and fruit size in clementines after pollination by “Fortune” mandarins. None of the copper salts demonstrated pollenicidal capacity on any of the four treatment dates in this study. This could be related to the application dates, if products were applied when the pollen had already germinated and fertilized the ovules within the ovaries, so that the treatments could not produce the desired effect. The pollen of the “Fortune” mandarin produced a great quantity of seeds in the pollinated “Clemenules” fruit, which were also larger and had lower rates of abscission than the unpollinated fruit. 69 8. LITERATURA CITADA ADDICOTT, F.T. 1982. Abscission. Berkeley. University of California, Press. ASOCIACIÓN NACIONAL DE FABRICANTES E IMPORTADORES DE PRODUCTOS FITOSANITAIORIOS AGRÍCOLAS. 2002-2003. Manual fitosanitario. Chile. AFIPA AG e IMPA. 1213p. ________. 1998-1999. Manual fitosanitario. Chile. AFIPA AG. 731p. AGUSTÍ, M. 2004. Fruticultura. Madrid. Mundi-Prensa. 493p. ________. 2003. Citricultura. Segunda edición. Madrid. Mundi prensa. 422p. ________. 2000. Citricultura. Madrid. Mundi-Prensa. 416p. ________.; ALMELA, V.; ZARAGOZA, S.; JUAN, M., TRENOR, I.; ALONSO, E. y PRIMO -MILLO, E. 1998. Técnicas para mejorar el tamaño del fruto de naranjas y mandarinas. Generalitat Valenciana. Conselleria de Agricultura, Pesca y Alimentación. pp 9-10 ________.; ZARAGOZA, S.; BLEIHOLDER, H.; BUHR, L.; HACK, H.; KLOSE, R y STAU, R. 1995. Codificación BBCH para Agrios, (on line). www.bba.de/veroeff/bbch/bbch.pdf ALI, A.J.; DEVAKUMAR, C.; ZAMAN, F.U. and SIDDIQ, E.A. 1999. Gametocida potency of ethyl 4' fluorooxanilate in rice. Ind. J. Genet.59: 3, 267-279. ASÍNS, M.J.; JUAREZ, J.; PINA, J.A.; PUCHADES, J.; CARBONELL, E.A. y NAVARRO, L. 2002. Nulessín, una nueva Clementina. Levante Agrícola. Nº 359: 36-40. BATCH, J.J.; PARRY, K.P.; ROWE, C.F.; LAWRENCE, D.K. and BROWN, M.J. 1980. Method of controlling pollen formation in monoecious and hemaphrodite plants using oxanilates and their derivatives. In: Plant growth regulators and herbicides, Antagonists-Recent advances. J C (ed.). Noyes Data corporation, New Jersey, pp 82-89. BATHIA, R.; GUPTA, D.; CHANDEL, J.S. and SHARMA, N.K. 1994. Relative abundance of insect visitors on flowers of major subtropical fruits in Himachal Pradesh and their effect on fruit set. Indian Journal of Agricultural Sciences. 65 (12): 907-912. 70 BEN-CHEIKH, W.; PEREZ-BOTELLA, J.; TADEO, F., TALÓN, M.; PRIMOMILLO, E. 1997. Pollination increases gibberellins levels in developing ovaries of ceded varieties of citrus. Plant. Physiology. 114: 557-564. BINGHAM, J. and LUPTON, F.G.H. 1987. Production of new varieties: an integrated research approach to plant breeding. In Lupton, F.G.H.(ed.), Wheat breeding: Its scientific basis. Londres. Chapman and Hall. 66p. BONO, R.; SOLER, J.; BUJ, A.; VILLALBA, D.; SALVIA, J., BELLVER, R. y MARES, M. 2004. El problema de la presencia de semillas en los frutos cítricos. Ficha Técnica, Serie Citricultura Nº5. Generalitat Valenciana, Conselleria D’ Agricultura, Pesca I Alimentació. pp. 1-4. ________., SOLER, J. y BUJ, A. 2000. Problemática de la presencia de semillas en los cítricos. IV congreso Cítricola de’l Horta Sud. Ediciones y promociones L.A.V. pp. 29-46. BREWBAKER, J.L. and KWACK, B.H. 1963. The essential role of calcium ion in pollen germination and pollen tube growth. Amer. J. Bot. 50: 859-865. BUKIYA, Z.M. 1990. Effect of the pollen of different pollinators on seed weight in early mandarins. Subtropicheskie-kul’tury. 1: 67-71. CASTRO, X. 2000. Evaluación del comportamiento que presenta la conducción de árboles de palto cv. Hass, en alta densidad. Taller Licenciatura Ing. Agr. Quillota, Universidad Católica de Valparaíso, Facultad Agronomía. 122 p. CHANDLER, W.H. 1962. Frutales de hoja perenne. México D.F. U.T.E.H.A. 666p. CROSS, J. and SCHULZ, P. 1997. Chemical induction of male sterility. In Shivana, K.R. y Sawhney, V.K. (ed.), Pollen biotechnology for crop production and improvement”. Cambridge University Press. Cambridge. (3): 218-237. DAVIES, F.S.; ALBRIGO, L.G. 1994. Cítricos. Zaragoza. Editorial Acribia S.A. 283p. DELHOM, M.J. y PRIMO-MILLO, E . 1989. Influencia de las hormonas en el cuajado del fruto de los agrios. Edita: Generalitat Valenciana. Conselleria d´ Agricultura y Pesca. 71 DIARIOPYME, 2005. Superficie plantada de Clementinas aumenta 193% según último análisis del Catastro Frutícola Nacional, (on line). www.diariopyme.cl DUMAS, C. 1984. Déterminisme du sexe floral. Parthenocarpie et sterilité mâle. Pollinisation et productions végétales. París. I.N.R.A. pp. 74-87. EDWARDS, I. 1986. Breeding of hybrid varieties of wheat. In Edwards, I.B., Feistritzer, W.P. y Kelly, A.F. Hybrid seed production of selected cereal, oil, and vegetable crops. Roma. FAO Plant Production and Protection Paper 82:3-24. EL-GHAZALY, G. and JENSEN, W. 1990. Development of wheat (Triticum aestivum) pollen wall before and after effect of a gametocide. Canadian Bot 68: 2509-2516 ENVIRONMENTAL PROTECTION AGENCY. 1997. Pesticide fact sheet. Washington,DC,EUA.(online).www.epa.gov/opprd001/factsheets/tralko xydim.pdf. ETI, S. and STOSSER, R. 1992. “Pollen tube growth and development of ovules en relation to fruit set in mandarin cv. Clementine (Citrus reticulata Blanco). Acta Horticulturae. Nº321: 621-625. FUNDACIÓN PARA LA INNOVACIÓN AGRARIA. 2002. Estrategia de innovación agraria para la producción de frutales de hoja persistente. Santiago. 72p. FREE, J.B. 1993. Rutaceae. In: Insect Pollination of crops. London. Academic Press. London. pp. 479-485. FURR, J.R. 1964. New Tangerines for desert. Calif. Citrog. 49: 266-276. GAJU, M. 2002. Estudio de la efectividad del agente Hibridizante GenesisR en cuatro cultivares de trigo. Tesis de Magíster en producción de cultivos. Santiago. Chile. Pontificia Universidad Católica de Chile. Facultad de Agronomía e Ingeniería Forestal. 42 p. GARCÍA-PAPÍ, M., GARCÍA-MARTINEZ, J.L 1979. Influence of gibberellic acid on early fruit development, diffusible growth substances and content of macronutrients in seedless Clementine mandarin. Scientia Hortic. 11, 337-347. 72 GEITMANN, A. 1999. The Rheological Properties of the Pollen Tube Cell Wall. In: Fertilization in Higher Plants. Cresti, M., Cai, G. y Moscatelli, A. (Eds.). Springer Verlag., 20: 283-302. ________. 1997. Growth and formation of the cell wall in pollen tubes of Nicotania tabacum and Petunia hybrida. Hänsel-Hohenhausen, egelsbach Frankfurt Washington. GOLDSCHMIDT, E.E and MONSELISE, S.P. 1977. “Physiological assumption toward the development of a citrus fruiting model”. Proc. Int. Soc. Citricultura. 2: 668-672. GONG, M., YANG, Z. and CAO, Z. 1995. Initiating effects of calcium on pollen germination and its regulative role in pollen tube growth. Biology Science. Life Science College, Peking University. Vol. 31, 2: 238-249 GONZALEZ-SICILIA, E. 1968. El cultivo de los agrios. Valencia, España. Editorial Bello. 814p. GOODWIN, P.B. 1978. Phytohormones and fruit growth. In: Phytohormones and Related Compounds. A comprehensive Treatise. Eds: Letham, D.S., Goodwin, P.B., y Higgins, T.J.V. Elsevier/North-Holland Biomedical Press. Amsterdam. Vol II: 175-214. GUARDIOLA, J.L. 2003. Fructificación, Producción y Tamaño del fruto. Curso de Citricultura. Sao Paulo, Brazil. pp. 159-196. ________. 1998. Nutrición mineral y producción Agrícola. In: Fructificación y utilización de nutrientes. VII Simposio Nacional-Ibérico sobre la nutrición mineral de las plantas. HERNANDEZ, F. ; PRIMO-MILLO, E. 1990. Studies on endogenous cytokines in citrus. J. Hortic. Sci. 65(5): 595-601. JACKSON, L.K. 1986. Seed development in Citrus. Citrus flowering fruit set and development. Florida, Cooperative Extension Service. University of Florida: 9-40. JAN, C.C. AND ROWELL. P.L. 1981. Response of wheat tillers at different growing stages to gametocide treatment. Department of Agronomy and Range Science, University of California, California, USA. Volume 30, number 2: 501-504. 73 JUAN, M., PRIMO-MILLO, E. 1989. Influencia de las hormonas en el cuajado del fruto de los agrios. Editorial Generalitat Valenciana, Conselleria d′Agricultura i pesca. KREZDORN, A.H. 1986. Flowering and Fruit Set of Citrus. Citrus Flowering, Fruit Set and Development. Ferguson. J.J. Florida Cooperative Extensión Service. University of Florida, 26-27 de Marzo de 1986. pp. 1-14. KWAN, S.C.; HAMSON, A.R. and CAMPBELL, W.F. 1969. The effects of different chemicals on pollen germination and tube growth in Allium cepa. L. Journal of the American Society for Horticultural Science, 94. 561-563. LAL, S.K., DEVKUMAR, C., SAPRA, R.L. and SINGH, K.P. 2005. Use of gametocide for emasculation in Soybean (Glycine max (L) Merr.) (on line). www.soygenetics.org/pdf/gametocide.pdf LOUSSERT, R. 1992. Los Agrios. Madrid. Mundi-Prensa. 319p. LUZIO, W. y ALCAYAGA, S. 1992. Mapa de asociaciones de grandes grupos de suelos de Chile. En: Agricultura Técnica 52 (4): 347-353. Mc RAE, DH. 1985. Advances in chemical hybridization. In Janick J (ed). Plant breeding review 3:169-191. MARTÍNEZ, A.R. 1981. Proyecto de implementación de un sistema de riego tecnificado en la Estación Experimental La Palma, Quillota. Tesis Ing. Agr. Quillota, Universidad Católica de Valparaíso, Facultad de Agronomía. 102p. MATILLA, J. 2000. “Germinación y dormición de las semillas”. Fundamentos de Fisiología Vegetal. Madrid. McGraw-Hill/Interamericana. pp. 435-450. MATHUR, D.S. and LAL, S.K. 1999. Chemical induction of male sterility in chickpea. Ind. J. Genet.59: 3, 379-380. MEHOUACHI, J. 2001. Control hormonal del creceimento y de la abscisión de los frutos cítricos. Efecto del estrés hídrico y de la disponibilidad de carbohidratos. Memoria para el grado de Doctor Ingeniero Agrónomo, Valencia, Universidad Politécnica de Valencia. 189p. MESEJO, C.; MARTÍNEZ-FUENTES, A.; REIG, C.; JUAN, M.; ALMELA, V. y AGUSTÍ, M. 2005. El control de la polinización cruzada en las mandarinas. Comunidad Valenciana Agraria (en prensa). 74 MOGENSEN, H.L. and LADYMAN, J.A.R. 1989. A structural study on the mode of action of CHA chemical hybridizing agent in wheat. Sexual Plant Reproduction 2 : 173-183. MONSELISE, S.P. 1986. Handbook of fruit set and development. Volumen 2. C.R.C. Inc. Boca Ratón. Florida. 2. pp. 87-107. Citrus. ________. 1977. Citrus fruit development: endogenous systems and external regulation. Proc. Int. Soc. Citricultura, 2: 664-668. MUÑOZ, A.F.; AYUSO, M.C. y LABRADOR, J. 2005. Polinización de cultivos. España. Mundi-Prensa. 232p. NESVADBA, Z. y VYHNÁNEK, T. 2001. Induction of hybrid wheat using gametocide Genesis® in the Czech Republic. Acta Univ. Agric. et Silvic. Mendel. Brun. 1, 27-35. NITSCH, J.P. 1971. Perennation through seeds and other structures: Fruit development. In: Plant Physiology, a treatise. Steward, F.C. Eds. Academic Press. London. 6A: 413-501. ________. 1970. Hormonal factors in growth and development. In: The Biochemistry of fruit and their products. Ed. Hulme A.C. Vol. 1. Academic Press, New York. NOVOA, R.; VILLASECA, R.; DEL CANTO, P.; ROVANET, J.; SIERRA, C., y DEL POZO, A. 1989. Mapa Agroclimatico de Chile. Santiago. INIA. 221 p. ORTÚZAR, J. 1999. La Calidad de los Frutos Cítricos y los Factores que la determinan. En: Avances en Citricultura: Manejo de Huertos y Postcosecha. Colección de Extensión, Facultad de Agronomía e Ingeniería Forestal, Pontificia Universidad Católica de Chile, Santiago. pp. 25-37. PONS, J.; PASTOR, J.; POLLS, M. y REVERTER, A.J. 1996. Polinización Cruzada en Cítricos. III. Polinización Entomofila. Efecto de Repelentes. Levante Agrícola. 4to Trimestre. Nº 337: 291-295. ________.; ________.; ________. y ________. 1995a. Polinización cruzada en cítricos. I. Incidencia Comercial. Levante Agrícola. 3er Trimestre. Nº 332: 201-207. 75 ________.; ________.; ________. y ________. 1995b. Polinización Cruzada en Cítricos. II. Efecto del Viento. Levante Agrícola. 4to Trimestre. Nº 333: 326-329. POUVREAU, J.C. 1984. . Cultures fruitiéres. Régions tropicales: Les Agrumes. In: Pollinisation el productions végétales . I.N.R.A. . París. pp. 411-422. PRALORAN, J.C. 1977. Los Agrios. Barcelona. Editorial Blume. 520p. PRIMO-MILLO, E. 1994. Regulación del cujado del fruto en los cítricos. Levante Agrícola. Nº 326: 8-16. PUCHADES, J. 2002. Influencia de la Polinización en las Características del Fruto de cv. Fortune. Trabajo Fin de Carrera, Valencia, Escuela Universitaria de Ingeniería Agrícola, Universidad Politécnica de Valencia. 129p. RALLO, J. 1986. Frutales y abejas. Madrid, Publicaciones de Extensión Agraria. 231p. ROHIDAS, S.B. and CHAKRAWAR, V.R. 1989. Studies on floral biology of some important Citrus species. II. Kagzi Lime (Citrus aurantifolia Swingle). Horticultural Journal. 2. 1, 20-25. ROMERO, H. 1985. Geografía de los Climas. Santiago, Instituto Geográfico Militar. 243p. ROOT, A. 1976. ABC y XYZ de la Apicultura. 15a. Ed. Hemisferio Sur. 670p. SOLER, J. 1999. Reconocimiento de cítricos en campo. Valencia, España. Edita Generalitat Valenciana. 187p. ________. VILLALBA, D.; CANALES. J.M.; BELLVER, R. y SALA, J. 1996. Formación de semillas. Polinización cruzada. Comunidad Valenciana Agraria. pp. 39-43. SPIEGEL-ROY, P., GOLDSCHMIDT, E.E. 1996. Biology of citrus. Cambridge, Ed. University press. TALON, M., 2000. Giberelinas. In: Fundamentos de Fisiología vegetal AzcónBieto, J y Talón, M, eds. Madrid. Mc Graw-Hill. pp. 325-343. ________. 1997. Regulación del cuajado del fruto en cítricos: Evidencias y conceptos. España. Levante Agrícola. Nº 338: 27-37. 76 ________.; ZACARÍAS, L., and PRIMO-MILLO, E. 1992. Gibberellins and parthenocarpic ability in developing ovaries of seedless mandarins. Plant. Physiology. 99: 1575-1581. ________.; HEDDEN, P., and PRIMO-MILLO, E. 1990. Gibberellins in Citrus sinensis: A comparison between ceded and seedless varieties. Plant growth Regul. 9: 201-206. TSCHABOLD, E.E.; HEIM, D.R.; BECK, J.R.; WRIGHT, J.L.; RAINEY, D.P.; TERANDO, N.H. and SCHWER, J.F. 1988. LY 195259, new chemical hybridizing agent for wheat. Crop Sci 28:4. 583-588 UAUY, C. 2001. Análisis de un Agente Hibridizante Químico para la Producción de Trigo Híbrido. Tesis Ingeniero Agrónomo. Santiago, Chile. Pontificia Universidad Católica de Chile. Facultad de Agronomía e Ingeniería Forestal. VAN DER MEER, Q.P. and VAN BENNEKOM. J.L. 1975. Gibberellic acid as a gametocide for the common onion (Allium cepa L.). II. The effect of GA 4/7 (on line). www.springerlink.com/app/home/contribution.asp. WALLACE, H., LEE, L.S. 1999. Pollen source, fruit set and xenia in mandarins. J. Hortic. Sci. Bio. 74(1): 82-86. WONG, M., BLOUET, A. and GUCKERT, A. 1995. Effectiveness of SC2053 as a chemical hybridizing agent for winter wheat, importance of developmental stages and doses of application. Plant Growth Reg 16: 243-248. ZACARÍAS, L.; TALÓN, M., BEN-CHEIKH, W.; LAFUENTE, M.T. and PRIMO-MILLO, E., 1995. Abscisic-acid increases in no growing and paclobutrazol-treated fruits of seedless mandarins. Physiology Plants 95: 613-619. ZARAGOZA, S.; PARDO, J.; IBAÑEZ, R. y MEDINA, A. 2005. Obtención de mandarinas tardías sin semillas mediante la irradiación de yemas. España. Levante Agrícola. Nº 374: 44-50. ZARASHUILI, R.M. 1967. Influence of bees on the yield of citrus trees on the commercial plantations of Georgia. XXI Int. Congr. Summ. Paper. 156, 159-163. 77 ANEXOS 78 ANEXO 1. Codificación BBCH de los estadios fenológicos del desarrollo d los agrios (Género Citrus) (AGUSTÍ et al., 1995). Estadio principal del desarrollo 0: Desarrollo de las yemas. 00 Reposo: Las yemas vegetativas y de inflorescencias están indiferenciadas, cerradas y cubiertas por escamas. 01 Comienzan hincharse las yemas. 02 Finaliza el hinchamiento de las yemas: las escamas verdes están ligeramente separadas. 03 Finaliza el hinchamiento de las yemas: las escamas verdes están ligeramente separadas. 07 Empieza la apertura de las yemas. 09 Los primordios foliares son visibles. Estadio principal del desarrollo 1: Desarrollo de las hojas. 10 Las primeras hojas empiezan a separarse: las escamas verdes están ligeramente abiertas y las hojas emergiendo. 11 Las primeras hojas son visibles. 15 Se hacen visibles mas hojas, pero sin alcanzar su tamaño final. 19 Las hojas alcanzan su tamaño final. Estadio principal del desarrollo 3: Desarrollo de los brotes. 31 Empieza a crecer el brote: se hace visible el tallo. 32 Los brotes alcanzan alrededor del 20% de su tamaño final. 39 Los brotes alcanzan alrededor del 90% de su tamaño final. 79 Estadio principal del desarrollo 5: Desarrollo de las flores. 51 Las yemas se hinchan: están cerradas y se hacen visibles las escamas, ligeramente verdes. 53 Las yemas revientan: las escamas se separan y se hacen visibles los primordios florales. 55 Las flores se hacen visibles: están todavía cerradas (botón verde) y se distribuyen aisladas o en racimos en inflorescencias con o sin hojas. 56 Los pétalos crecen; los sépalos envuelven la mitad de la corola (botón blanco). 57 Los sépalos se abren: se hacen visibles los extremos de los pétalos, todavía cerrados, de color blanco o amoratado. 59 La mayoría de las flores, con los pétalos cerrados, adquieren la forma de una bola hueca y alargada. Estadio principal del desarrollo 6: Floración. 60 Primeras flores abiertas. 61 Comienza la floración: alrededor del 10% de las flores están abiertas. 65 Plena floración: alrededor del 50% de las flores están abiertas. Empiezan a caer los primeros pétalos. 67 Las flores se marchitan: la mayoría de los pétalos están cayendo. 69 Fin de la floración: han caído todos los pétalos. Estadio principal del desarrollo 7: Desarrollo del fruto. 71 Cuajado: el ovario comienza a crecer; se inicia la caída de frutos jóvenes. 72 El fruto, verde, está rodeado por los sépalos a modo de corona. 73 Algunos frutos amarillean: se inicia la caída fisiológica de frutos. 74 El fruto alcanza alrededor del 40% del tamaño final. Adquieren un color verde oscuro. Finaliza la caída fisiológica de frutos. 80 79 El fruto alcanza alrededor del 90% de su tamaño final. Estadio principal del desarrollo 8: Maduración del fruto. 81 El fruto empieza a colorear (cambio de color). 83 El fruto está maduro para ser recolectado, aunque no ha adquirido su color característico. 85 Maduración avanzada; se va incrementando el color característico de cada cv. 89 Fruto maduro y apto para el consumo: tiene su sabor y firmeza naturales; comienza la senescencia y la abscisión. Estadio principal del desarrollo 9: Senescencia y comienzo del reposo. 91 Las brotaciones han completado su desarrollo; las hojas adquieren su plena tonalidad verde. 93 Las hojas viejas inician la senescencia y comienzan a caer. 97 Reposo invernal.