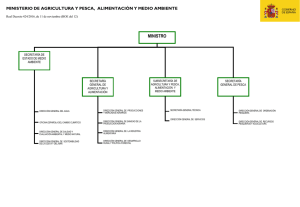
biodiversidad y manejo de la ictiofauna del archipiélago sabana
Anuncio

BIODIVERSIDAD Y MANEJO DE LA ICTIOFAUNA DEL ARCHIPIÉLAGO SABANA-CAMAGÜEY. Octubre del 2000 INFORME FINAL DEL PROYECTO BIODIVERSIDAD Y MANEJO DE LA ICTIOFAUNA DEL ARCHIPIELAGO SABANA-CAMAGÜEY. PROBLEMA NACIONAL DE CIENCIA Y TÉCNICA: Cambios Globales y evolución del medio ambiente cubano. Autores: Rodolfo Claro Madruga1, Karel Cantelar1, Fabián Pina Amargós2 y Juan P. García-Arteaga1 1 - Instituto de Oceanología, CITMA. 1ra. No. 18406, Rpto. Flores, Playa, La Habana 12100, Cuba. 2 - Centro de Investigaciones de Ecosistemas Costeros, CITMA, Cayo Coco, Ciego de Ávila, Cuba. Octubre del 2000 TABLA DE CONTENIDO RESUMEN..................................................................................................................................1 INTRODUCCIÓN ................................................................................................................. ...3 MATERIALES Y MÉTODOS............................................................................................. ....4 RESULTADOS...........................................................................................................................6 LA ICTIOFAUNA DE LOS ARRECIFES CORALINOS........................................................... 6 LA ICTIOFAUNA DE LOS FONDOS BLANDOS ...................................................................16 LA ICTIOFAUNA DE LOS MANGLARES...............................................................................18 DIVERSIDAD DE LA ICTIOFAUNA....................................................................................... 22 4.1 Diversidad de especies .................................................................................................22 4.2 Diversidad ecológica ....................................................................................................26 EVOLUCIÓN Y MANEJO DE LOS RECURSOS PESQUEROS............................................. 32 5.1 Organización, infraestructura y evolución histórica ................................................32 5.2 Distribución espacial de los recursos pesqueros y relación con los hábitat...............34 5.3 Esfuerzo de pesca ........................................................................................................35 5.31 Artes de arrastre ..........................................................................................................40 5.32 5.32 Nasas .................................................................................................................42 5.33 5.33 Nasas del alto.....................................................................................................43 5.34 5.34 Tranques ...........................................................................................................43 5.4 Volumen y composición de las capturas...................................................................... 43 5.5 Composición por especies de las capturas comerciales por artes.............................50 5.6 5.51 Capturas con chinchorros ................................................................................ 51 5.7 5.52 Capturas con nasas de placer...........................................................................58 5.8 5.53 Capturas con trasmallos.................................................................................. 61 5.6 Composición por tallas de las capturas .......................................................................62 MANEJO DE LA ICTIOFAUNA EN RELACIÓN CON EL TURISMO Y OTRAS OPCIONES....................................................................................................................70 6.1 Desarrollo del turismo ..................................................................................................70 6.2 La pesca recreativa.......................................................................................................71 LAS AREAS PROTEGIDAS MARINAS COMO MECANISMO DE PROTECCIÓN Y USO SOSTENIBLE DE LA CTIOFAUNA........................................73 DISCUSION ............................................................................................................................75 ELEMENTOS PARA UNA ESTRATEGIA DE MANEJO ................................................. 79 CONCLUSIONES ...................................................................................................................80 RECOMENDACIONES .........................................................................................................81 REFERENCIAS ......................................................................................................................83 RESUMEN En la década del 90 el Archipiélago Sabana-Camagüey sufrió grandes transformaciones ambientales como resultado de la acción de factores naturales y antrópicos. Al comparar la situación actual de las comunidades de peces con la existente a finales del 80, se comprobó una apreciable disminución de la diversidad de especies, y una reducción de tercer orden en la densidad y la biomasa de peces en los arrecifes coralinos y manglares. Tal degradación parece estar relacionada con la alarmante disminución de la cobertura de corales escleractínios, y una enorme proliferación de algas, las cuales cubren 70-80% del sustrato duro, impidiendo la recuperación de los corales y otros organismos sésiles.. Tal mortandad de corales parece ser consecuencia de las altas temperaturas provocadas por eventos ENSOs ocurridos en 1995, 1997 y 1998 lo que provocó el blanqueamiento masivo de los corales. Ello afecta la disponibilidad de refugios y alimentos para los peces, limitando sus condiciones de hábitat. En las macrolagunas de la porción oriental del Archipiélago la ictiofauna sufrió también graves afectaciones como resultado de la elevación de la salinidad por la interrupción del régimen de circulación y por el represamiento. Estos impactos además, han provocado la ruptura de los procesos ecológicos de varias especies comerciales, afectando su abundancia. La mayoría de las macrolagunas se encuentran afectadas también por la contaminación y los arrastres de redes de pesca, que destruyen parte de la biota superficial del fondo, remueven los sedimentos aumentando la turbidez y disminuyendo la fotosíntesis. La pesca comercial en la región ha disminuido como consecuencia de la degradación de los hábitat, en parte provocados por el uso de artes de pesca destructivos. Se sugiere una estrategia para el manejo con el fin de viabilizar la recuperación y uso sostenible y de los peces en la región en beneficio no solo de la pesca, sino también del turismo y otras opciones. ABSTRACT In the decade of the 90s the Archipelago Sabana-Camagüey have suffered big environmental transformations as a result of natural and antropogenic factors. When comparing the current situation of fish communities with the existed one at the end of the 80s, it is observed an appreciable decrease of species richness, and a reduction of third order in fish density and the biomass on coral reefs and mangroves. Such a degradation seems to be related with the alarming decrease of scleractinian coral covers, and an enormous proliferation of algae, which covers 70-80% of the hard substrate, impeding the recovery of the corals and other benthic organisms. Such coral mortalities seems to be the consequence of the high temperatures caused by the ENSOs events happened in 1995, 1997 and 1998, which caused the massive bleaching of corals. These changes have affected the availability of refuges and food for fishes, limiting their habitat conditions. In the inner waters of the eastern portion of the Archipelago, the ichthyofauna also suffered serious damages as a result of the increase of water salinity provoqued by the interruption of the circulation regime and damming of fresh waters. These impacts also caused the rupture of the ecological processes of several commercial species, affecting their abundance. Most of the lagoons are also affected by pollution and trawling of fishing nets that destroy part of the superficial biota of the bottom, remove the silts increasing water turbidity and diminishing the photosynthesis. The commercial fishing in the region has decreased, as a consequence of the habitat degradation, partially provoked by destructive fishing methods. Measures are suggested for the integrated and sustainable management of fish resources in the region, in order to benefit not only fishing activities, but tourism and other options. INTRODUCCIÓN Los ecosistemas costeros son altamente vulnerables a las alteraciones provocadas por el hombre y los cambios climáticos, por lo que requieren de la evaluación y monitoreo de sus condiciones ambientales como elemento básico para su conservación y manejo. En el Archipiélago SabanaCamagüey, como resultado de actividades para el desarrollo de la región, se han producido alteraciones del régimen hidrológico de las macrolagunas, que han provocado un incremento de la salinidad, cambios en la dinámica de los sedimentos, afectaciones notables de la fauna y flora, y otras alteraciones del medio ambiente marino (CUB/92/G31, 1997), con la consecuente disminución de la pesca en algunas áreas. Estos cuerpos de agua mantienen un estrecho intercambio con las zonas arrecifales exteriores, que influye sustancialmente sobre estos hábitat, los cuales sustentan la mayor parte de las pesquerías y el turismo. El aporte de nutrientes desde las aguas interiores, está provocando un incremento de la vegetación en los arrecifes coralinos poniendo en peligro su salud (CUB/92/G31, 1997) y, como consecuencia afectando la base alimentaria, el refugio y las condiciones de vida necesarias para los peces arrecifales, que constituyen la base económica de nuestras pesquerías. El valor estético de estos arrecifes depende de la abundancia y diversidad, tanto de los organismos sésiles (corales, gorgonias, esponjas, poliquetos, etc.) como de los peces. Estos últimos constituyen un importante elemento del paisaje submarino, ya que determinan en gran medida su colorido, movimiento, carisma y valor estético para uso turístico. Muchas de las especies arrecifales pasan parte de su ciclo de vida en las aguas interiores, y/o mantienen un intenso intercambio de energía entre los arrecifes y biotopos adyacentes, por lo que la degradación de estos últimos afecta seriamente a los arrecifes. En el Archipiélago se desarrolla una intensa actividad pesquera, basada principalmente en la captura de peces. Hasta la década del 80 la zona aportaba aproximadamente el 35% de la captura nacional de este renglón (Baisre, 1985, Giménez et al, 1992, Claro et al, 1994). Si bien existe bastante información estadística, un sistema de regulaciones y cierto control sobre la actividad pesquera, es pobre el conocimiento sobre los factores ecológicos que inciden sobre ésta en las condiciones actuales, así como de las influencias de los cambios ambientales sobre los recursos pesqueros. A fines de la década del 80, se realizó por parte de este colectivo un inventario de la ictiofauna en los arrecifes y manglares de la región (Claro y García-Arteaga, 1993, 1994), pero las grandes extensiones de las macrolagunas quedaron sin evaluar, y es en estas donde precisamente han ocurrido las mayores transformaciones ambientales. Por otra parte, estas últimas necesariamente se reflejan en la abundancia, composición y valor de los recursos pesqueros, y la propia actividad de pesca interviene en el complejo proceso de cambio que ocurre en el ecosistema. Al esclarecimiento de estos problemas estuvo orientado el presente proyecto, con el fin de contribuir al establecimiento de medidas para el uso sostenible de la biodiversidad y los recursos del Archipiélago Sabana-Camagüey. A tales efectos, se plantearon los siguientes objetivos específicos: 1- Concluir el inventario de la ictiofauna de los arrecifes coralinos, manglares, pastos marinos y fondos blandos del Archipiélago y sentar las bases para un programa de monitoreo. 2- Evaluar las alteraciones en la estructura de las comunidades de peces como resultado de los cambios ambientales provocados por las actividades antrópicas y eventos naturales. 3- Identificar especies y áreas que requieran protección o regulaciones pesqueras, y elaborar las medidas adecuadas para ello. 4- Aportar información para el planeamiento y manejo integrado de los arrecifes y otros hábitat neríticos. 5- Evaluar la evolución histórica y el estado actual de los recursos pesqueros, los efectos nocivos de los artes y métodos de pesca en los arrecifes y biotopos asociados, y elaborar recomendaciones para el manejo de dichos recursos. 6- Elaborar alternativas de manejo para compatibilizar el desarrollo sostenible del turismo marítimo, la pesca y otras actividades socioeconómicas. MATERIALES Y MÉTODOS Para el inventario y estudio de las comunidades de peces en los arrecifes y manglares se utilizó el método de censo visual con buceo autónomo (Brock, 1954; Rogers et al, 1994), mediante transeptos de 50 m de longitud y 2 m de ancho, en un grupo de estaciones seleccionadas de aquellas previamente evaluadas en 1988-1989 (Fig. 1). En los manglares se empleó el mismo método con las modificaciones de Claro y García-Arteaga (1993) para este hábitat. En cada estación se realizaron 8 transeptos. Los valores de densidad y biomasa se calcularon partir de la suma de todos los muestreos de cada estación, al igual que los indicadores de diversidad. Además se estimaron los valores promedios y sus desviaciones estándar para cada estación y hábitat. En los manglares se realizaron algunos muestreos con Rotenona para capturar la ictiofauna críptica. Los métodos utilizados y los resultados obtenidos en el inventario de los fondos blandos se presentaron en un informe parcial, el cual se adjunta (Anexo 1). Para el análisis de los recursos pesqueros se utilizaron los datos de capturas de las Empresas Pesqueras que operan en la región, obtenidos en el Dpto. de Estadísticas del Ministerio de Industria Pesquera, y otros fueron aportados por investigadores del Centro de Investigaciones Pesqueras. Se analizaron además los vales de recepción de la captura de la Asociación CAIMAR (Caibarién), y se realizaron encuestas a pescadores. El estudio de las capturas comerciales a bordo de embarcaciones de pesca se limitó a la Empresa de Caibarién, la más importante del ASC y una embarcación de Turiguanó, debido a limitaciones de recursos para la realización de muestreos. La riqueza de especies para cada subzona y hábitat se evaluó mediante curvas acumulativas del número de especies por número de individuos y por área muestreada, estimadas mediante 5 ó más combinaciones aleatorias de las unidades de muestreo y obteniendo la curva promedio para cada grupo muestral. El procesamiento de los datos para el análisis de la estructura de las comunidades se realizó mediante el programa de computación ESTRUCOM, (elaborado por J.P. García-Arteaga y R. Claro) que brinda la siguiente información: lista de especies en orden taxonómico y la densidad y biomasa de cada una y el total de la familia o para la categoría trófica, según sea necesario, Indice de diversidad de Shannon y Weaver (H'), Indice de Riqueza de Margalef (R1), Indice de equitatividad de Pielou (J) (Boesh, 1977), Indice de Jerarquía e Indice de frecuencia/abundancia (Mason y Brian,1974), estos dos últimos solo para conjuntos de muestras. Los datos originales de todos los muestreos, incluyendo los realizados en los manglares y arrecifes de todo el Archipiélago anteriores a este proyecto, se encuentran en una base de datos (Anexo 2), y los ejemplares de interés colectados se encuentran en las colecciones de referencia del Instituto de Oceanología. Fig. 1. Ubicación de las estaciones para los censos visuales en las pendientes (P) y crestas (C) arrecifales y en los manglares del Archipiélago Sabana-Camagüey. RESULTADOS Con el fin de evaluar los cambios ocurridos en los últimos años en los principales hábitat del ASC, se comparó la estructura de las comunidades de peces en los arrecifes coralinos y los manglares en el primer semestre del año 2000, con similares observaciones realizadas en 1988-1989 por parte de dos de los autores (Claro y García-Arteaga, 1993; 1994). Para ello se seleccionaron tres hábitat principales: las crestas y pendientes arrecifales y los manglares. Se repitieron en este segundo muestreo, un número de estaciones distribuidas a lo largo del archipiélago (Fig. 1). Considerando que la estructura de la comunidad guarda estrecha relación con el tipo de hábitat (Claro y GarcíaArteaga, 1994), los resultados se presentan por separado para cada uno. Los fondos blandos de las macrolagunas no habían sido estudiados anteriormente, por lo que se realizaron muestreos repetidos en algunas bahías durante el período 1995-1998 en que se sucedieron importantes cambios en el ecosistema como resultado de la construcción de viaductos sobre la plataforma. 1. LA ICTIOFAUNA DE LOS ARRECIFES CORALINOS. En el inventario realizado en 1988-1989 se registraron 132 especies de peces en 31 estaciones de las pendientes arrecifales, entre 15 y 25 m de profundidad. Tanto la diversidad como la densidad y biomasa de peces observada en ese período, se consideró relativamente alta, y muy superior a la encontrada por los mismos autores en plataformas impactadas por la sobrepesca y cambios ambientales desfavorables para los arrecifes coralinos, en las islas Martinica y Guadalupe (Claro y García-Arteaga, 1994; Claro, García-Arteaga, Bouchon-Navarro, Louis y Bouchon, 1998), aunque inferiores a los observados en los arrecifes del Archipiélago Jardines de la Reina y en los de la Florida (Claro, en prensa). En Abril del 2000 se repitieron 11 de aquellas estaciones, seleccionadas de forma que estuvieran representadas las principales formaciones arrecifales y distribuidas a lo largo de todo el Archipiélago (Fig. 1). La riqueza de especies en el arrecife frontal, expresada por las curvas acumulativas de número de especies por número de individuos, parecen ser iguales en los dos períodos estudiados (Fig. 2A). Sin embargo, cuando se comparan las curvas de número de especies por área muestreada acumulada (Fig. 2B), se observa claramente una mayor riqueza en el período 1988-89. Esta diferencia está relacionada con la gran disminución del número de individuos de las especies dominantes, razón por la cual el valor del índice de equitatividad es mayor en el 2000 (Tabla 1). Al comparar los valores promedios y acumulativos de aquellas estaciones que se repitieron durante el presente proyecto, observamos que la densidad y biomasa en este hábitat fueron tres veces menores que las observadas en 1988-89 (Tabla 1). Las figuras 3A y 3B muestran que, con excepción de las dos estaciones ubicadas en la región occidental del Archipiélago, y una estación en Cayo Fragoso (en la cual la densidad y biomasa fueron muy bajas en ambos períodos) en todas las demás la densidad fue superior en el primer inventario, con un incremento sustancial de dicha diferencia hacia la región oriental. La diferencia en biomasa fue aún mayor, manifestándose prácticamente en todas las estaciones. Tabla 1. Ver fichero Tablas apaisadas Figura 2. Curvas acumulativas de riqueza de especies en las pendientes arrecifales. A: Número de especies por número de individuos, B: Número de especies por área. La disminución en la densidad de peces se manifestó en todos las categorías tróficas (Fig. 4) con excepción de los omnívoros y fue más notoria en el caso de los planctófagos debido principalmente a la disminución de dos especies dominantes por su número en este hábitat, el cromis azul (Chromis cyanea) y la jenízara (Clepticus parrai), cuyas densidades bajaron en 6 veces y sus biomasas en 10. Sin embargo, estos pequeños peces no son determinantes en la biomasa total, la cual refleja mejor los cambios energéticos del ecosistema. La disminución de biomasa se debió principalmente al decrecimiento de los peces depredadores bentófagos e ictiófagos, entre los que se encuentran las principales especies comerciales. La densidad de tales carnívoros disminuyó sólo en 2,3 veces, pero su biomasa disminuyó en 5,3. Si consideramos sólo las especies comerciales de mayor calidad (Tabla 2) vemos que su densidad total disminuyó en 3,5 veces, mientras que su biomasa, en 8 veces, principalmente debido a la drástica caída de los pargos. Figura 3. Comparación de las densidades (A) y biomasas (B) de peces en 1988-89 y el año 2000, por estaciones en las pendientes arrecifales. La diferencia en las disminuciones de densidad y biomasa evidencian además un preocupante decrecimiento de las tallas, lo cual es indicio de aumento de la mortalidad total. Tanto la densidad como la biomasa de herbívoros disminuyó también, a pesar del desmesurado crecimiento de las poblaciones de algas. Tal decrecimiento se observó en todas las especies con excepción de Microspathodon chrysurus. El único grupo trófico que no disminuyó su abundancia fueron los omnívoros (Fig. 4), aunque en realidad en la mayoría de las especies hubo menor número de individuos en el 2000, con excepción de Stegastes partitus, la especie dominante en ambos períodos, y cuya abundancia aumentó en el último. Estas especies pueden actuar negativamente sobre la salud del arrecife, por su actividad como constructores de jardines de algas sobre heridas infligidas a los corales, lo que aumenta su vulnerabilidad a enfermedades infecciosas. En los arrecifes de crestas la riqueza de especies se comportó de forma similar al patrón observado en el arrecife frontal. La Fig. 5 muestra similar número de especies contra número de individuos en los dos períodos estudiados, pero mayor número de especies por igual área muestreada en el período 1988-89. También el número de especies por estación, y los valores de los índices de diversidad de Shannon y Weaver y el de Riqueza de Margalef fueron mayores en dicho momento (Tabla 3). Tabla 2. Densidad y biomasa de algunas familias de peces de relevante importancia comercial antes y después de la década del 90. Familias Densidad (ind./100m2) 1987-89 2000 Biomasa (g/100m2) 1987-89 2000 Jureles (Carangidae) 3,68 0,05 1146 14 Macarelas (Scomberomorus spp.) 0,27 0,06 635 103 Meros (Epinephelus spp. y Mycteroperca spp.) 2,71 1,7 2556 738 Pargos (Lutjanidae) 10,05 2,92 10347 854 Total 16,71 4,73 14684 1709 Fig. 4A. Densidad de peces en las pendientes arrecifales, por categorías tróficas. Oomnívoros; H - herbívoros; Pl - planctófagos; I - consumidores de invertebrados; Pe piscívoros; I+Pe - invertebrados + peces. Tabla 3 (Ver fichero: Tablas apaisadas) Fig. 4B. Biomasa de peces en las pendientes arrecifales, por categorías tróficas. Abreviaturas y leyenda igual que en 4A. Fig. 5A. Curvas acumulativas de riqueza de especies por número de individuos en las crestas arrecifales. En las crestas estudiadas del Archipiélago Sabana-Camagüey, Claro y García-Arteaga (1994) reportaron promedios de densidad y biomasa de 269 ind./100m2 y 12628 g/100m2, respectivamente, para 24 estaciones muestreadas durante los años 1988-89. La diversidad de especies fue casi tan alta como en el arrecife frontal, aunque con diferentes especies dominantes. Fig. 5B. Curvas acumulativas de riqueza de especies por área muestreada en las crestas arrecifales En los muestreos realizados en nueve de aquellas estaciones se observó similar decrecimiento de la abundancia que en las pendientes: en el año 2000 la densidad fue 2,8 veces menor y la biomasa 2,2. Igualmente, tales diferencias ocurrieron en casi todas las estaciones (Fig. 6) coincidiendo con mayores afectaciones de la biomasa en la región central y oriental de la zona. Fig. 6A Densidad de peces por estaciones en las crestas arrecifales, en 1988-89 y en el 2000. Fig. 6B. Biomasa de peces por estaciones en las crestas arrecifales, en 1988-89 y en el 2000. Tanto la densidad como la biomasa de peces en las crestas, disminuyeron en todas las categorías tróficas (Fig. 7), aunque fue notoria la caída de la biomasa de herbívoros en cinco veces, debido a la reducción en la abundancia de los barberos (Acanthurus spp.) y a la casi desaparición de especies de gran talla, como Scarus guacamaia, y el otrora abundante loro colirrojo, Sparisoma chrysopterum. Entre los bentófagos también se observó un decrecimiento grande, ocasionado en gran medida por una menor presencia de roncos (Haemulidae): 5,4 veces menos en el 2000. Evidentemente estos trascendentales cambios en la estructura de las comunidades guardan relación con la alta cobertura algas observada en todas las estaciones: 60-80% del substrato duro. Tanto en aguas someras como en la pendiente hasta 25 m de profundidad, el fondo estaba cubierto por una densa alfombra de algas, principalmente Dictyosphaeria cavernosa, Lobophora variegata, Cladophora spp., Halimeda spp., Penicillus spp. y Stypopodium zonale. En los años 1988-89, en ninguna estación se observó semejante cubrimiento algal. Si bien en esos muestreos no se trató de cuantificar su abundancia, ello se debió a que entonces su presencia no era significativa, mientras que el mismo personal sí observó una notable diferencia poco después, durante los muestreos realizados en 1991 en las islas Martinica y Guadalupe (Claro et al., 1998) donde ya en ese período se había producido una gran disminución de la biomasa de peces y un aumento inusitado de las algas, probablemente como consecuencia de la desaparición del erizo negro (Diadema antillarum) y de la sobrepesca de los peces herbívoros. Por el contrario, en los muestreos realizados en el año 2000, tanto en las pendientes arrecifales como en las crestas, se observó una gran cantidad de corales escleractinios y gorgonáceos muertos (no recientemente), cubiertos por el césped algal (Apéndice 1). Los corales vivos eran escasos (10-20% del fondo cubierto por estos), aunque en buen estado de salud. Se encontraron muy pocos corales con blanqueamiento u otras enfermedades. Fig. 7. Densidad (A) y biomasa (B) de peces por categorías tróficas, en las crestas arrecifales, en 1988-89 y en el 2000. Abreviaturas igual que en Fig. 4. 2. LA ICTIOFAUNA DE LOS FONDOS BLANDOS. La evolución y situación de los fondos blandos de las macrolagunas del Archipiélago SabanaCamagüey fueron descritas en un informe parcial presentado en 1998, el cual se adjunta a este documento (Anexo 1), por lo que a continuación se presenta solo un resumen de los principales resultados. Las macrolagunas del ASC se encuentran altamente degradadas por la acción conjunta de diversos factores antrópicos (CUB/92/G31, 1997) que han provocado serias afectaciones a la diversidad y abundancia de la ictiofauna. La interrupción del régimen hidrológico por los viaductos construidos para unir la isla principal con los cayos y estos entre sí, sumado al represamiento de las aguas fluviales que ya venía afectando a varias macrolagunas, provocó niveles letales de salinidad para la fauna marina en las bahías Los Perros, Jigüey y La Gloria, destruyendo además la vegetación del fondo, incrementando la turbidez del agua por la continua resuspensión de los sedimentos, y la acumulación de altas concentraciones de sulfuro de hidrógeno en el fondo (CUB/92/G31, 1997). Se demostró una drástica disminución de la riqueza de especies a partir de salinidades de 40‰, y la completa desaparición de la ictiofauna a más de 75‰. En los manglares afectados por salinidades superiores a 40‰ se encontró no sólo una disminución de la diversidad, densidad y biomasa de peces, sino además una notable reducción del reclutamiento de las especies comerciales. Las bahías Los Perros y La Gloria mostraron una extraordinariamente baja riqueza de especies (Fig. 8), lo cual se relaciona sin duda con las altas salinidades (hasta 83‰ en Bahía Los Perros en 1995). Las variaciones posteriores de la salinidad provocaron rápidos cambios en la riqueza de especies y su abundancia numérica. Predominaron en estas altas salinidades, especies de muy pequeña talla, no comerciales: las mojarritas (Eucinostomus spp.) y Cyprinodon variegatus. Esta última especie, típica de agua dulce, se encontró en salinidades de hasta 75‰, gracias a la virtual desaparición de sus depredadores, y a su notable tolerancia a los cambios de salinidad. La ictiofauna de carácter comercial prácticamente desapareció de esas bahías y se ha afectó seriamente en la Bahía Buenavista. Como resultado del incremento de la circulación a través de alcantarillas instaladas bajo el pedraplén y la apertura del dique Estero-Socorro, bajó la salinidad lo cual ha permitido cierta recuperación de algunas especies altamente tolerantes. La más alta diversidad especies se observó en El Puerto de Sagua, que presenta el mayor aporte de aguas fluviales y al mismo tiempo el mayor intercambio con el mar abierto. La mayor biomasa pesquera se observó sin embargo en la Bahía San Juan de los Remedios (Ver detalles en Informe Parcial del Proyecto de 1998, Anexo 1). 3. LA ICTIOFAUNA DE LOS MANGLARES. En los años 1988-89 se realizaron muestreos en 63 estaciones en los manglares de todo el archipiélago (Claro y García-Arteaga, 1993). En dichos muestreos se encontraron 87 especies de peces, con una alta densidad (1990 ind./100m2) gracias a la gran abundancia de manjúas (Jenkinsia lamprotaenia) y otros Clupeidae, los cuales constituyeron el 83% de los especímenes. No obstante, la biomasa, también relativamente alta, estuvo determinada por la presencia de pargos (46%) y roncos (40%). La mayoría de las especies que por el día se concentran en el manglar, se dispersan de noche por las macrolagunas y seibadales para alimentarse, por lo que la abundancia de tales peces está estrechamente vinculada a las condiciones ambientales de esos biotopos. Insertar Fig. 8 (Apaisada) Fig. 9. Curvas acumulativas de riqueza de especies en los manglares. En los muestreos realizados en abril del año 2000 en este hábitat, se encontró una riqueza de especies mayor que la encontrada en esas mismas estaciones en el período 1988-89 (Fig. 9), lo que se manifiesta al plotear el número de especies contra el número de individuos. Ello se debe a una gran disminución en la abundancia de las especies dominantes en ese biotopo, la manjúa, las sardinas y los roncos. La densidad de peces fue menor cuatro veces que la observada en 1988-89 en las mismas estaciones, mientras que la biomasa fue sólo 1,6 veces inferior (Tabla 4). Esta diferencia se debe a que fueron las especies de menor talla las más afectadas (principalmente Jenkinsia lamprotaenia y Anchoa liolepis.). Sólo en una estación (4M) la densidad y biomasa fueron superiores en el último muestreo (Fig. 10) gracias a la presencia de grandes cardúmenes de sardinas (Harengula spp.), las cuales raramente fueron observadas en las demás estaciones. Fig. 10. Densidad (A) y biomasa (B) de peces en los manglares, en 1988-89 y en el 2000. Fig. 11. Densidad (A) y biomasa (B) de peces por categorías tróficas en los manglares, en 198889 y en el 2000. Es necesario recalcar la gran disminución de las poblaciones de manjúa (6,2 veces menos) en comparación con el período anterior (Claro y García-Arteaga, 1993), ya que esta especie es la principal carnada para la pesca de bonito y albacora que se realiza en la zona oceánica adyacente al archipiélago. Los bentófagos, e ictiófagos, que son los gremios determinantes por su biomasa en este hábitat disminuyeron sustancialmente su abundancia. Los roncos (Haemulidae) redujeron su densidad en seis veces y su biomasa en cinco, mientras que los pargos redujeron su número y peso en cuatro veces. De la misma manera que en los arrecifes, los omnívoros, representados por las chopitas del género Stegastes, fueron el único grupo trófico que presentó mayor densidad y biomasa en el año 2000 (Fig. 11), en este caso gracias a la presencia de Stegastes leucostictus, que es generalmente la especie del género más abundante en el manglar. Tabla 4 (Ver Fichero: Tablas apaisadas) Fig. 11. Densidad (A) y biomasa (B) de peces por categorías tróficas en los manglares, en 198889 y en el 2000. En el caso de los herbívoros no se observaron diferencias entre los dos períodos. Este grupo y los omnívoros son los de menor significación en el manglar. Las causas de la disminución de peces en los manglares parece estar relacionada con la degradación de los fondos blandos y praderas donde se alimentan la gran mayoría de los peces que por el día se concentran en los manglares. El área productiva de ese ecosistema ha disminuido sustancialmente producto de los cambios en el régimen hidrológico (ver Anexo 1). Durante las inmersiones realizadas en los manglares en Mayo del 2000, fue perceptible, en mayoría de las estaciones una alta turbidez del agua, lo cual no fue un elemento llamativo en los muestreos realizados en 1988-89. Lamentablemente en ninguno de los dos casos fue posible la medición de este factor. El mismo guarda relación con los cambios ocurridos en los fondos blandos aledaños a los manglares. 4. DIVERSIDAD DE LA ICTIOFAUNA. 4.1 Diversidad de especies La Fig. 12 presenta una representación aproximada de la distribución de la diversidad de peces en Archipiélago Sabana-Camagüey, a partir de las informaciones obtenidas durante este proyecto y otras investigaciones anteriores, utilizando como criterio la existencia de cuatro niveles de concentración de la misma, tomando en cuenta la cantidad de especies por unidad de área y número de individuos: 1. Diversidad Alta. Generalmente se observa en los arrecifes frontales, arrecifes de parche, cabezos y en algunas crestas arrecifales, principalmente en su zona de embate. Pueden ser observadas entre 35 y 60 especies (promedio 40-50) en 1000 m2, mediante censo visual en horas del día. 2. Diversidad media. Usualmente se encuentra en arrecifes costeros, bancos arrecifales, algunas crestas pobres en refugios (principalmente en la zona trasera y superior de ésta) y en casi todos los manglares. Generalmente son visibles entre 20 y 40 especies (promedios 25-35) en 1000 m2 de observación. 3. Diversidad baja. Se encuentra principalmente en las praderas y fondos blandos. El número de especies por unidad de área es bajo, entre 2 y 20 especies (promedios entre 5 y 10) por 1500 m2 de área barrida con la rastra Otter (ver Materiales y métodos). No obstante, el número total de especies en una macrolaguna puede ser relativamente alto y su abundancia numérica más aún, debido a la gran extensión de éstos hábitat y a las migraciones diurnas y de forrajeo de los peces que se refugian en los arrecifes y manglares. 4. Diversidad pobre. Se manifiesta en los arenales extensos (playas y placeres exteriores cercanos al arrecife frontal) y en las macrolagunas afectadas por graves problemas antrópicos (contaminación, elevación de la salinidad, bajo contenido de oxígeno, etc.). El número de especies por 1500 m2 de rastreo fluctúa entre 0 y 10 (promedios de 3 a 6). La más alta diversidad de especies se encuentra en los arrecifes frontales, donde Claro y GarcíaArteaga (1994) censaron 132 especies mediante buceo diurno, de ellas 118 en las mismas estaciones muestreadas durante este trabajo, en el cual se encontró un número inferior (106 especies), a pesar de que el área muestreada fue mayor (Tabla 1). Los arrecifes de parche según los datos de Claro y García-Arteaga (1994) tienen similar diversidad de especies que las pendientes, pero son relativamente escasos en el ecosistema Sabana-Camagüey, mientras que casi toda la pendiente arrecifal que bordea la plataforma, entre 12-15 y 25 m se encuentra cubierta de arrecifes coralinos. Las especies dominantes en el arrecife frontal son: el chromis azul (Chromis cyanea), la chopita bicolor (Stegastes partitus), la doncella (Thalassoma bifasciatum) y el loro listado (Scarus croicensis) (Tabla 5). En los arrecifes de parche son más abundantes los roncos (Haemulidae) que constituyeron 43% de la biomasa y 59% de los individuos censados por Claro y García-Arteaga (1994). En las crestas arrecifales también puede ser alta la diversidad de especies, aunque usualmente menor que en los arrecifes frontales (Fig. 12). La densidad y biomasa son inferiores que en los arrecifes frontales y de parche (Tablas 1 y 3). Los herbívoros son aquí más abundantes que en las otras formaciones arrecifales. Predominan en este hábitat, las doncellas Thalassoma bifasciatum y Halichoeres bivittatus, así como la chopita negra, Stegastes dorsopunicans aunque por su peso, el ronco arará, Haemulon plumieri es la principal especie (Tabla 6). La diversidad de especies en los manglares es más variable y por lo general inferior a la de los arrecifes, pero pueden ocurrir densidades muy altas, gracias a la presencia de cardúmenes de sardinas (Jenkinsia lamprotaenia, Harengula spp.), cabezotes (Atherinomorus stipes) y anchoas (Anchoa spp.) que son las especies dominantes (Tabla 7), aunque por su peso los pargos son más importantes, principalmente el caballerote, Lutjanus griseus. Tabla 5. Relación de especies observadas en las pendientes arrecifales, densidad, biomasa e índices de Jerarquía (IJ) y de Frecuencia/abundancia-biomasa (F/a-b) para cada una. Biomasa FAMILIA Densidad F/a-b IJ 2 2 g/100m Especies #Ind/100m ORECTOLOBIDAE 0.01 174 Ginglymostoma cirratum 0.01 174 A/4-2 96 MURAENIDAE 0.02 134 Gymnothorax funebris 0.02 134 A/4-2 87 HOLOCENTRIDAE 2.29 181 Holocentrus ascensionis 0.13 25 A/3-3 53 Holocentrus marianus 0.16 4 A/3-4 51 Holocentrus rufus 1.30 116 A/2-2 17 Holocentrus vexillarius 0.28 13 A/3-3 42 Myripristis jacobus 0.43 23 A/3-3 33 AULOSTOMIDAE 0.07 7 Aulostomus maculatus 0.07 7 A/4-4 66 SERRANIDAE 3.43 707 Epinephelus cruentatus 0.36 47 A/3-3 36 Epinephelus fulvus 0.75 118 A/3-2 26 Epinephelus guttatus 0.31 89 A/3-2 40 Epinephelus striatus 0.07 71 A/4-3 67 Hypoplectrus guttavarius 0.03 menos de 1 A/4-4 83 Hypoplectrus indigo 0.08 2 A/4-4 64 Hypoplectrus nigricans 0.10 1 A/3-4 58 Hypoplectrus puella 0.88 10 A/3-3 21 Hypoplectrus unicolor 0.01 menos de 1 A/4-4 97 Liopropoma rubre 0.01 menos de 1 A/4-4 98 Mycteroperca bonaci 0.03 260 A/4-2 84 Mycteroperca interstitialis 0.01 2 A/4-4 99 Mycteroperca tigris 0.06 91 A/4-2 70 Mycteroperca venenosa 0.02 11 A/4-3 88 Serranus tabacarius 0.02 menos de 1 A/4-4 89 Serranus tigrinus 0.69 5 A/3-4 27 GRAMMISTIDAE 0.01 3 Rypticus saponaceus 0.01 3 A/4-4 90 GRAMMIDAE 2.72 menos de 1 Gramma loreto 2.68 menos de 1 A/2-4 10 Gramma melacara APOGONIDAE Apogon binotatus Apogon maculatus MALACANTHIDAE Malacanthus plumieri CARANGIDAE Caranx ruber LUTJANIDAE Lutjanus apodus Lutjanus cyanopterus Lutjanus jocu Ocyurus chrysurus HAEMULIDAE Anisotremus virginicus Haemulon aurolineatum Haemulon flavolineatum Haemulon plumieri Haemulon sciurus SPARIDAE Calamus bajonado Calamus penna Calamus pennatula Calamus sp. SCIAENIDAE Equetus acuminatus Equetus punctatus MULLIDAE Mulloidichthys martinicus Pseudopeneus maculatus KYPHOSIDAE Kyphosus incisor CHAETODONTIDAE Chaetodon aculeatus Chaetodon capistratus Chaetodon ocellatus Chaetodon sedentarius Chaetodon striatus POMACANTHIDAE Centropyge argi Holacanthus ciliaris Holacanthus isabelita Holacanthus tricolor Pomacanthus arcuatus Pomacanthus paru POMACENTRIDAE Abudefduf saxatilis 0.04 0.02 0.01 0.01 0.05 0.05 0.05 0.05 2.79 0.77 0.05 0.02 1.95 12.13 0.62 0.99 4.24 3.64 2.63 0.16 0.04 0.02 0.04 0.06 0.04 0.01 0.03 0.47 0.13 0.34 0.01 0.01 2.46 0.40 1.66 0.17 0.14 0.09 1.28 0.17 0.24 0.01 0.66 0.13 0.07 35.63 0.19 menos de 1 menos de 1 menos de 1 menos de 1 6 6 13 13 809 133 66 46 563 1857 166 28 961 431 271 28 9 6 4 9 1 menos de 1 menos de 1 39 18 21 10 10 388 3 25 6 350 3 295 menos de 1 42 4 57 108 84 134 14 A/4-4 79 A/4-4 A/4-4 100 101 A/4-4 73 A/4-3 74 A/3-2 A/4-3 A/4-3 A/2-2 25 75 91 12 A/3-2 A/3-3 A/2-1 A/2-2 A/2-2 30 19 7 8 11 A/4-3 A/4-4 A/4-4 A/4-3 80 92 81 71 A/4-4 A/4-4 102 85 A/3-3 A/3-3 54 39 A/4-3 103 A/3-4 A/2-3 A/3-4 A/3-2 A/4-4 34 14 49 52 59 A/3-4 A/3-3 A/4-4 A/3-3 A/3-2 A/4-2 50 45 104 28 55 68 A/3-3 47 Chromis cyaneus Chromis multilineatus Microspathodon chrysurus Stegastes dorsopunicans Stegastes leucostictus Stegastes partitus Stegastes planifrons Stegastes variabilis CIRRHITIDAE Amblycirhitus pinos LABRIDAE Bodianus rufus Clepticus parrai Halichoeres bivittatus Halichoeres garnotti Halichoeres maculipinna Halichoeres poeyi Lachnolaimus maximus Thalassoma bifasciatum SCARIDAE Scarus coelestinus Scarus croicensis Scarus taeniopterus Scarus vetula Sparisoma atomarium Sparisoma aurofrenatum Sparisoma chrysopterum Sparisoma rubripinne Sparisoma viride SPHYRAENIDAE Sphyraena barracuda OPISTHOGNATHIDAE Opisthognathus aurifrons LABRISOMIDAE Malacoctenus triangulatus BLENNIIDAE Ophioblennius atlanticus GOBIIDAE Coryphopterus eidolon Coryphopterus glaucofraenum Coryphopterus personatus Gobiosoma evelynae Gobiosoma genie ACANTHURIDAE Acanthurus bahianus Acanthurus coeruleus Acanthurus chirurgus 15.85 0.86 0.83 0.13 0.98 16.63 0.04 0.12 0.01 0.01 21.67 0.24 4.72 0.28 5.69 0.06 0.05 0.10 10.53 9.49 0.03 4.63 1.36 0.35 0.10 1.59 0.49 0.07 0.87 0.10 0.10 0.19 0.19 0.02 0.02 0.02 0.02 2.60 0.08 0.66 1.26 0.55 0.05 5.63 3.62 1.71 0.30 27 6 38 1 6 40 menos de 1 1 menos de 1 menos de 1 1525 15 1131 2 283 1 1 59 33 674 30 94 86 23 menos de 1 119 50 17 255 259 259 menos de 1 menos de 1 menos de 1 menos de 1 menos de 1 menos de 1 1 menos de 1 menos de 1 menos de 1 menos de 1 menos de 1 477 229 222 25 A/1-3 A/3-4 A/3-3 A/3-4 A/3-4 A/1-3 A/4-4 A/3-4 2 23 24 56 20 1 82 57 A/4-4 105 A/3-3 A/2-1 A/3-4 A/2-2 A/4-4 A/4-4 A/3-3 A/1-3 46 5 43 4 72 76 60 3 A/4-3 A/2-2 A/2-2 A/3-3 A/3-4 A/2-2 A/3-3 A/4-3 A/3-2 86 6 16 38 61 15 32 69 22 A/3-2 62 A/3-4 48 A/4-4 93 A/4-4 94 A/4-4 A/3-4 A/2-4 A/3-4 A/4-4 65 29 18 31 77 A/2-2 A/2-2 A/3-3 9 13 41 SCOMBRIDAE Scomberomorus maculatus Scomberomorus regalis SCORPAENIDAE Scorpaena plumieri BALISTIDAE Balistes vetula Cantherines pullus Melichthys niger Xanthichthys ringens TETRAODONTIDAE Canthigaster rostrata Sphoeroides spengleri 0.06 0.01 0.05 0.01 0.01 0.71 0.09 0.01 0.26 0.36 0.39 0.37 0.02 95 38 57 2 2 204 68 1 105 30 10 9 1 A/4-3 A/4-3 106 78 A/4-4 107 A/4-3 A/4-4 A/3-2 A/3-3 63 108 44 37 A/3-3 A/4-4 35 95 La ictiofauna de las praderas marinas es pobre en diversidad de especies, sobre todo durante las horas del día, y está asociada a la de los arrecifes y manglares, donde generalmente se refugian los peces que se alimentan en este biotopo de noche e incluso algunos por el día. No obstante, habitan en ellas de forma permanente un grupo de especies que utilizan como refugio la vegetación y turbidez del agua (Eucinostomus spp., Eugerres spp., Gerres cinereus, Archosargus rhomboidalis, Monacanthus spp. Hippocampus spp., etc. ) o que se entierran o camuflan en los sedimentos (Opsanus spp., Ogcocephalus nasutus, Achirus lineatus), así como especies pelágico-neríticas que forman cardúmenes como mecanismo defensivo (Albula vulpes, Elops saurus, Megalops atlanticus, Sphyraena guachancho, Selar crumenophthalmus, Caranx chrysos, Lutjanus synagris, etc.) (ver anexo 1). La mayoría de estas especies están también presentes e inclusive pueden ser más abundantes en los fondos blandos sin vegetación, que se encuentran en las más aguas interiores. Estas especies se caracterizan por su alta tolerancia a los cambios de las condiciones ambientales (principalmente de la salinidad) y generalmente sus poblaciones son numerosas. Aunque en condiciones normales la ictiofauna de estos biotopos es bastante homogénea, en las macrolagunas se observa una notable diferencia en la diversidad y abundancia de peces en relación con el grado de deterioro de las condiciones ambientales (ver Anexo 1), principalmente por la elevación de la salinidad, en las bahías Los Perros, Jigüey y La Gloria, así como en la parte oriental y sur de la Bahía de Buenavista. En este último caso, influye de forma notoria la contaminación, al igual que en algunas otras áreas del archipiélago. 4.2 Diversidad ecológica Como hemos visto anteriormente, las condiciones ambientales del Archipiélago Sabana-Camagüey han sufrido modificaciones que afectan las condiciones adecuadas para garantizar los procesos vitales para la alimentación, reproducción y crecimiento de las comunidades de peces y sus complejas interrelaciones. Así por ejemplo, la disminución de la complejidad topográfica del arrecife por la mortandad de corales, gorgonáceos y otros organismos sésiles asociados a estos, ha provocado una seria disminución de los organismos alimentarios y de la cantidad y calidad de los refugios necesarios para la vida de los peces. Las afectaciones sufridas en algunas macrolagunas por la elevación de la salinidad o por contaminación, han provocado una disminución sustancial del área para la producción biológica y los procesos vitales. El área de las bahías Los Perros, Jigüey y la Gloria, por mencionar solo las más impactadas, equivale a aproximadamente al 35 % del área total de aguas interiores del archipiélago. Por otra parte, la elevación de la salinidad provoca un mayor gasto de energía para el proceso de osmorregulación, en detrimento de la tasa de crecimiento y reproducción de los organismos. Tabla 6. Relación de especies observadas en las crestas arrecifales, densidad, biomasa e índices de Jerarquía (IJ) y de Frecuencia/abundancia-biomasa (F/a-b) para cada una. FAMILIAS Densidad Biomasa F/a-b IJ Especies g/100m2 #Ind/100m2 DASYATIDAE 0.03 398 Dasyatis americana 0.03 398 A/4-2 59 MURAENIDAE 0.01 1 Muraena miliaris 0.01 1 A/4-4 66 HOLOCENTRIDAE 2.27 196 Holocentrus ascensionis 0.41 98 A/3-2 27 Holocentrus rufus 0.58 67 A/3-2 21 Holocentrus vexillarius 1.28 31 A/2-3 12 AULOSTOMIDAE 0.01 menos de 1 Aulostomus maculatus 0.01 menos de 1 A/4-4 67 SERRANIDAE 0.08 8 Epinephelus cruentatus 0.03 4 A/4-4 60 Epinephelus fulvus 0.03 3 A/4-4 61 Epinephelus guttatus 0.01 1 A/4-4 68 Serranus tigrinus 0.01 menos de 1 A/4-4 69 CARANGIDAE 0.16 47 Caranx ruber 0.16 47 A/4-3 38 LUTJANIDAE 1.43 245 Lutjanus analis 0.01 10 A/3-3 70 Lutjanus apodus 1.07 156 A/4-4 14 Lutjanus griseus 0.07 13 A/3-3 48 Lutjanus jocu 0.01 6 A/4-3 71 Lutjanus mahogoni 0.04 5 A/4-3 54 Lutjanus synagris 0.01 2 A/4-4 72 Ocyurus chrysurus 0.22 53 A/3-2 33 HAEMULIDAE 6.97 1627 Anisotremus virginicus 0.03 9 A/4-3 62 Haemulon album 0.13 44 A/3-3 39 Haemulon aurolineatum 0.71 27 A/2-3 19 Haemulon carbonarium 0.05 5 A/4-3 50 Haemulon chrysergyreum 0.62 13 A/3-3 20 Haemulon flavolineatum 2.59 120 A/2-2 9 Haemulon parrai 0.09 20 A/3-3 46 Haemulon plumieri 0.93 1049 A/2-1 15 Haemulon sciurus 1.82 339 A/2-2 10 SPARIDAE 0.09 15 Calamus bajonado 0.03 8 A/4-3 63 Calamus pennatula Calamus sp. MULLIDAE Mulloidichthys martinicus Pseudopeneus maculatus KYPHOSIDAE Kyphosus incisor CHAETODONTIDAE Chaetodon capistratus Chaetodon ocellatus Chaetodon sedentarius Chaetodon striatus POMACANTHIDAE Holacanthus ciliaris Holacanthus tricolor Pomacanthus arcuatus Pomacanthus paru POMACENTRIDAE Abudefduf saxatilis Abudefduf taurus Chromis cyaneus Chromis multilineatus Microspathodon chrysurus Stegastes dorsopunicans Stegastes leucostictus Stegastes partitus Stegastes planifrons Stegastes variabilis Stegastes mellif CIRRHITIDAE Amblycirhitus pinos LABRIDAE Bodianus rufus Halichoeres bivittatus Halichoeres garnotti Halichoeres maculipinna Halichoeres poeyi Halichoeres radiatus Lachnolaimus maximus Thalassoma bifasciatum SCARIDAE Scarus croicensis Scarus guacamaia Scarus taeniopterus Scarus vetula Sparisoma aurofrenatum Sparisoma chrysopterum 0.04 0.03 0.25 0.13 0.12 0.07 0.07 0.33 0.12 0.08 0.01 0.12 0.17 0.04 0.04 0.05 0.04 15.04 2.92 0.01 0.05 0.29 3.20 6.04 0.57 1.59 0.12 0.22 0.03 0.01 0.01 27.81 0.12 7.68 0.83 0.74 0.58 0.21 0.01 17.64 6.76 3.34 0.01 0.20 0.37 0.50 0.24 4 3 13 9 4 34 34 10 3 2 1 4 37 2 2 21 13 197 77 1 menos de 1 2 82 26 3 4 1 1 menos de 1 menos de 1 menos de 1 169 15 58 8 9 8 1 7 64 360 27 13 3 8 23 17 A/4-4 A/4-4 55 64 A/3-3 A/3-4 40 41 A/3-3 49 A/3-4 A/3-4 A/4-4 A/3-4 42 47 73 43 A/4-4 A/4-4 A/4-3 A/4-3 56 57 51 58 A/2-2 A/4-4 A/4-4 A/3-4 A/2-2 A/2-3 A/3-4 A/2-4 A/3-4 A/3-4 A/4-4 8 74 52 30 6 3 23 11 44 34 65 A/4-4 75 A/3-3 A/1-2 A/2-3 A/2-3 A/3-3 A/3-4 A/4-3 A/1-2 45 2 16 18 22 35 76 1 A/2-3 A/4-3 A/3-4 A/3-3 A/3-3 A/3-3 5 77 36 28 24 31 Sparisoma radians Sparisoma rubripinne Sparisoma viride SPHYRAENIDAE Sphyraena barracuda LABRISOMIDAE Malacoctenus triangulatus BLENNIIDAE Ophioblennius atlanticus GOBIIDAE Coryphopterus glaucofraenum ACANTHURIDAE Acanthurus bahianus Acanthurus coeruleus Acanthurus chirurgus BALISTIDAE Balistes vetula Cantherines pullus TETRAODONTIDAE Canthigaster rostrata DIODONTIDAE Diodon holacanthus 0.05 0.83 1.22 0.33 0.33 0.02 0.01 0.47 0.47 0.02 0.01 7.37 4.13 3.00 0.24 0.67 0.20 0.47 0.01 0.01 0.01 0.01 menos de 1 93 176 720 720 menos de 1 menos de 1 1 1 menos de 1 menos de 1 589 308 266 15 72 59 13 menos de 1 menos de 1 2 2 A/4-4 A/2-2 A/2-2 53 17 13 A/3-1 29 A/4-4 78 A/3-1 25 A/4-4 79 A/2-2 A/2-2 A/3-3 4 7 32 A/3-2 A/3-3 37 26 A/4-4 80 A/4-4 81 Tabla 7. Relación de especies observadas en los manglares, densidad, biomasa e índices de Jerarquía (IJ) y de Frecuencia/abundancia-biomasa (F/a-b) para cada una. FAMILIAS Densidad Biomasa F/a-b IJ 2 2 Especies #Ind/100m g/100m DASYATIDAE 0.02 14 Urolophus jamaicensis 0.02 14 A/4-3 48 CLUPEIDAE 603.78 1639 Harengula clupeola 33.72 59 A/2-3 6 Harengula humeralis 112.91 1501 A/1-1 2 Jenkinsia lamprotaenia 456.92 78 A/1-3 1 Jenkinsia majua 0.23 1 A/4-4 30 ENGRAULIDAE 177.91 57 Anchoa hepsetus 94.19 49 A/1-3 3 Anchoa lyolepis 83.72 7 A/2-4 4 BELONIDAE 0.76 8 Strongylura notata 0.44 2 A/4-4 22 Tylosurus crocodilus 0.44 11 A/4-3 23 ATHERINIDAE 53.84 125 Atherinomorus stipes 53.84 125 A/2-2 5 SERRANIDAE 0.01 menos de 1 Hypoplectrus puella 0.01 menos de 1 A/4-4 59 CARANGIDAE 0.02 9 Caranx ruber 0.02 9 A/4-3 49 LUTJANIDAE Lutjanus analis Lutjanus apodus Lutjanus cyanopterus Lutjanus griseus Lutjanus jocu Lutjanus synagris Ocyurus chrysurus GERREIDAE Eucinostomus gula Eucinostomus jonesi Gerres cinereus Eucinostomus sp. HAEMULIDAE Anisotremus virginicus Haemulon aurolineatum Haemulon carbonarium Haemulon flavolineatum Haemulon parrai Haemulon sciurus Haemulon spp. SPARIDAE Archosargus rhomboidalis MULLIDAE Mulloidichthys martinicus CHAETODONTIDAE Chaetodon capistratus Chaetodon ocellatus Chaetodon striatus POMACANTHIDAE Holacanthus ciliaris Pomacanthus arcuatus Pomacanthus paru POMACENTRIDAE Abudefduf saxatilis Stegastes dorsopunicans Stegastes leucostictus Stegastes partitus Stegastes planifrons Stegastes variabilis LABRIDAE Halichoeres bivittatus Halichoeres maculipinna Thalassoma bifasciatum SCARIDAE Scarus croicensis Scarus guacamaia 27.42 0.34 7.56 0.37 17.76 1.35 0.03 0.01 10.43 7.66 0.06 2.63 0.08 12.66 0.33 0.67 0.66 2.28 0.42 7.55 0.76 0.02 0.02 0.01 0.01 0.34 0.30 0.01 0.02 0.15 0.01 0.01 0.13 8.41 6.43 0.13 1.60 0.03 0.02 0.19 0.34 0.14 0.03 0.16 1.10 0.93 0.01 4852 425 728 224 3257 212 5 1 206 30 menos de 1 176 menos de 1 1221 20 7 100 37 74 983 menos de 1 2 2 2 2 3 3 menos de 1 menos de 1 59 1 6 53 180 19 2 159 menos de 1 menos de 1 1 1 menos de 1 menos de 1 1 60 12 32 A/4-2 A/3-2 A/4-2 A/2-1 A/3-2 A/4-4 A/4-4 27 9 25 7 16 43 57 A/3-3 A/4-4 A/3-2 A/4-4 8 40 13 38 A/4-3 A/4-4 A/4-2 A/3-3 A/4-3 A/3-1 A/4-4 28 19 20 14 24 10 18 A/4-4 50 A/4-4 56 A/4-4 A/4-4 A/4-4 29 60 51 A/4-4 A/4-4 A/4-3 58 54 35 A/3-3 A/4-4 A/3-2 A/4-4 A/4-4 A/4-4 12 36 15 45 52 32 A/4-4 A/4-4 A/4-4 34 46 33 A/3-3 A/4-3 17 53 Scarus taeniopterus Sparisoma aurofrenatum Sparisoma chrysopterum Sparisoma viride SPHYRAENIDAE Sphyraena barracuda GOBIIDAE Coryphopterus eidolon ACANTHURIDAE Acanthurus bahianus Acanthurus coeruleus Acanthurus chirurgus DIODONTIDAE Diodon hystrix POECILLIDAE Gambusia spp. 0.05 0.01 0.07 0.03 0.49 0.49 0.03 0.03 0.70 0.13 0.22 0.35 0.03 0.03 7.40 7.40 menos de 1 2 13 1 294 294 menos de 1 menos de 1 088 11 37 40 140 140 1 1 A/4-4 A/4-4 A/4-3 A/4-4 41 55 39 44 A/4-2 21 A/4-4 47 A/4-3 A/4-3 A/4-63 37 31 26 A/4-2 42 A/3-4 11 Tales cuerpos de agua, aún cuando en condiciones no antropizadas pudieran ser poco productivos para la pesca por su poca profundidad, constituyen importantes fuentes de energía que se exporta a otros sitios del ecosistema, son importantes zonas de reclutamiento, cría y alimentación, donde se desarrollan las primeras etapas del ciclo de vida. Las zonas ubicadas al norte de dichas bahías eran zonas de cría de muchas especies comerciales que se pescan en las praderas marinas y arrecifes al norte de la cayería. La figura 13 resume las informaciones obtenidas sobre los sitios de concentración para el desove de varias especies comerciales (principalmente pargos), que usualmente también son utilizados por otros peces con semejante estrategia reproductiva (roncos, meros, bajonaos, jureles, pez perro, etc.). Fig. 13. Algunos sitios de congregación para el desove de pargos y meros. Esta estrategia ampliamente estudiada por varios autores (Johannes, 1978, Smith y Tyler, 1978; Barlow, 1981; Kaufman y Ebersole, 1984), consiste en producir el desove en el borde de la plataforma, generalmente en promontorios marinos o áreas de plataforma que se adentran en el océano, y donde el sistema de corrientes facilita la permanencia de los huevos y larvas en aguas oceánicas, evitando su inmediata entrada a las zonas arrecifales donde la depredación es elevada y de forma que las corrientes realicen el transporte y la dispersión de las larvas en un área extensa, mientras se produce el desarrollo larval. Al alcanzar estas el tamaño adecuado se incorporan a las zonas de plataforma aledaña donde se inicia su reclutamiento y posterior asentamiento y cría, ya en aguas someras tranquilas, preferentemente en el interior de las macrolagunas. La Fig. 14 representa, de forma esquemática, los procesos de reproducción, reclutamiento y cría de muchas de las especies neríticas de importancia pesquera (particularmente los pargos, roncos, jureles, etc.), tomando como base las informaciones obtenidas sobre la circulación, la distribución de las etapas juveniles y la pesca en la región central del Archipiélago. Muchas especies son reclutadas en las zonas bajas cercanas a los cayos y penetran a las lagunas por los canales que los separan, donde generalmente ocurre la etapa de cría. A medida que crecen, se dispersan más, buscando nuevas fuentes de alimentación. Debido a la topografía de estas bahías y el régimen de vientos y circulación, gran parte de la población se mueve en dirección oeste. Así, por ejemplo, aseguran algunos pescadores que gran parte de las biajaibas que se capturan en la Bahía San Juan de los Remedios, provienen de la Bahía de Buenavista. Por otra parte, el intrincado laberinto de canales que formaban los Cayos Cobos, Las Brujas, y otros de esa zona era, hasta la construcción del viaducto Caibarién-Santa María, una importante zona de reclutamiento directo de biajaiba, y también de pargo criollo, rabirrubia y otras especies. Ello explica porque, después de construirse el pedraplén disminuyó drásticamente la captura de dicha especie. Similar fenómeno puede ser la causa que impide la recuperación de las poblaciones de machuelo, tras su colapso, al parecer por sobrepesca, a partir de 1990, coincidiendo con el inicio de la construcción de dicho pedraplén. Igualmente, los organismos que logran sobrevivir a las deterioradas condiciones ambientales de las bahías La Gloria, Jigüey y Los Perros, encuentran una barrera en sus migraciones al encontrarse con los pedraplenes Jigüey-Cayo Romano y Turiguanó-Cayo Coco los cuales dificultan la migración de los preadultos y adultos hacia el océano al acercarse la etapa de reproducción. Es de esperar que el pedraplén que actualmente se construye para unir Cayo Santa María con Cayo Guillermo, provoque una situación aún peor para los recursos pesqueros en la zona y un deterioro mucho mayor en las Bahías Buenavista y San Juan de los Remedios, al cerrar más aún el reclutamiento a las zonas de cría. Fig. 14. Representación esquemática de los procesos de reproducción, reclutamiento y migraciones de los peces en la región central del Archipiélago Sabana-Camagüey. 5. EVOLUCIÓN Y MANEJO DE LOS RECURSOS PESQUEROS 5.1 Organización, infraestructura y evolución histórica En el Archipiélago Sabana-Camagüey operan las Asociaciones pesqueras de Matanzas (unidad pesquera de Cárdenas), Las Villas (Carahatas, La Panchita, La Isabela y Caibarién) Ciego de Ávila (Punta Alegre y Turiguanó), Camagüey (Nuevitas), Tunas (Manatí y Puerto Padre), Holguín (Gibara, Banes, Guatemala, Cayo Mambí y Moa) y Guantánamo (Baracoa) (Fig. 15). Fig. 15. Ubicación de las empresas de pesca que operan en el Archipiélago Sabana-Camagüey, y las asociaciones provinciales a las cuales se supeditan. El conjunto de todas estas unidades constituye la llamada Zona D de la plataforma. Las tres últimas empresas, aunque están ubicadas fuera del ASC, tradicionalmente han realizado parte de sus capturas en dicha región, debido a que sus áreas de pesca son de poca extensión y pobres recursos. También pescan en la plataforma suroriental de Cuba, mientras que la empresa de Camagüey, también realiza algunas capturas fuera del Archipiélago Sabana-Camagüey (hacia el este). Por otra parte, hasta el año 1984 varias empresas realizaban eventualmente capturas en la plataforma de Bahamas. A partir de 1985 el Ministerio de Industria Pesquera (MIP) adoptó medidas drásticas para evitar tal actividad (Cabrera y Obregón, 1986), aunque ocasionalmente, y de forma furtiva, algunos barcos realizan incursiones rápidas a dicha región. Tales capturas introducen cierto sesgo en la estadística de la pesca por zonas, pero su magnitud no altera sustancialmente el análisis sobre la evolución y situación de los recursos pesqueros en la Zona D, que incluye todas las áreas de pesca ubicadas entre Punta Hicacos y Punta Maisí. Ya desde mediados de la década del 70, las pesquerías en el ASC, y en general en todo el Archipiélago Cubano, alcanzaron y sobrepasaron los niveles de captura máxima sostenible, y algunos recursos iniciaron una etapa de sobrepesca (Baisre, 1993; Claro et al., 1994). Aunque se tomaron medidas para detener ese proceso (disminución del esfuerzo pesquero, regulaciones de tallas, vedas en los períodos de reproducción, cuotas, etc.), por la falta de vigilancia y control y en algunos casos por el afán de elevar la producción para satisfacer las demandas de la población, no se han podido evitar algunas afectaciones a los stocks. A ello contribuyen también factores ajenos a la pesca, como son la contaminación, los cambios provocados en el régimen hidrológico y la pesca furtiva. Por otra parte, el Archipiélago Sabana-Camagüey y la región norte de las provincias orientales son las áreas de plataforma más afectadas por la mal llamada “pesca deportiva”, que en la práctica no es más que pesca de subsistencia con una alta componente de especulación, especialmente en los últimos 10 años. Muchas de las embarcaciones particulares dedicadas a esta actividad utilizaban artes de pesca masivos (principalmente chinchorros y redes de sitio), en la mayoría de los casos obtenidos ilícitamente de las empresas estatales pesqueras. El establecimiento de la Organización Nacional de Inspección Pesquera (ONIP), ha contribuido sustancialmente a la reducción de esa actividad que afecta no solo a la pesca comercial, sino también a la conservación del ecosistema. No existe información concreta sobre el volumen de pesca que se obtiene por esa vía, pero resulta evidente que su magnitud es apreciable (ver epígrafe 6.2). Durante nuestros muestreos, pudimos constatar que los propios pescadores se apropian de cantidades apreciables de la captura, situación que ha mejorado con el establecimiento de una política de estimulación material, aunque aún se practica. Esta apropiación ilícita se produce fundamentalmente sobre las especies de mayor calidad y precio. En el caso de los barcos chinchorreros y rederos, se produce sobre los pargos (pargo criollo, cubera, biajaiba) y las vaquitas (Lactophrys trigonus), esta última llega a tierra en forma de filetes. Los barcos naseros, se apropian de parte de la captura de chernas y meros. A modo de ejemplo: de acuerdo con informaciones confidenciales obtenidas durante 1995-1996, calculamos que las embarcaciones de la Empresa Pesquera de Caibarién, desviaban en los años 1995-98, unas 150 TM de pescado anualmente. 5.2 Distribución espacial de los recursos pesqueros y su relación con los hábitats Las pesquerías en el Archipiélago Sabana-Camagüey se realizan en la zona marítima (tanto interior como exterior) que se extiende desde Península de Hicacos hasta Nuevitas, donde se obtienen 50005500 TM anuales. El tramo de Cayos Pajonal a cayo Sabinal, que aporta aproximadamente el 60% de toda la Zona D, es precisamente el área más sometida a la influencia de los cambios ecológicos que se están produciendo como consecuencia de las alteraciones en el régimen de circulación. La distribución de los recursos pesqueros como es lógico, guarda estrecha relación con la distribución de los biotopos a los cuales se han adaptado las especies objeto de explotación. A su vez, los artes de pesca que se emplean para ello, deben ajustarse a las condiciones físicas del medio. La Figura 16 ofrece una representación aproximada de la distribución de las principales áreas de pesca en el Archipiélago, que pudiera ser mejorada con el aporte de información más específica. Baisre (1984) definió tres complejos ecológicos en las aguas cubanas, basándose en criterios biológico-pesqueros: el complejo seibadal-arrecife, el complejo litoral estuarino y el de las aguas oceánicas. Al aplicar estos conceptos para el ecosistema marino del Archipiélago Sabana-Camagüey es necesario hacer algunas precisiones. En el litoral estuarino, al menos en las áreas más cercanas a la costa de la isla principal, donde predominan las influencias terrestres, el fondo es generalmente fangoso, cubierto por pastos marinos en la cercanía de las costas, donde la productividad biológica parece ser muy alta, la turbidez es generalmente elevada y las fluctuaciones de salinidad amplias, además en muchos sitios hay una fuerte componente de contaminación por residuales urbanos, de la industria azucarera y otras industrias. La ictiofauna de la región interior de gran parte de la cayería no es la característica de zonas estuarinas debido a la alta salinidad de estas macrolagunas. Es este complejo estuarino el que ha sufrido los trascendentales cambios ambientales provocados por las alteraciones antrópicas referidas anteriormente, y que han provocado la casi total desaparición de la pesca comercial en las Bahías Los Perros, Jigüey y La Gloria y en menor medida en Bahía de Buenavista. Las encuestas realizadas a los pescadores de mayor experiencia, atestiguan que hasta la década del 70 las mencionadas bahías, y especialmente la de Buenavista, eran ricas en langostas, cangrejo moro y muchas especies de peces que actualmente no existen en tales macrolagunas, como los pargos (Lutjanidae), jureles (Carangidae), roncos (Haemulidae), bajonaos (Sparidae), sierra, serrucho y pintada (Scomberomorus spp.), y en las áreas más cercanas a la costa, las lisas (Mugil spp.), corvinas (Bairdiella spp.), pataos y mojarras (Gerridae) y salema (Archosargus rhomboidalis). Actualmente solo los dos últimos grupos de especies se encuentran, con una abundancia mucho menor y de menores tallas. La alta salinidad en este caso no solo afecta a la base alimentaria de las pocas especies que la toleran, sino que además provoca altos gastos fisiológicos para el proceso de osmorregulación, en detrimento del crecimiento y la fecundidad. La mayor parte de las pesquerías que se realizaban en estas zonas afectadas, se han movido hacia los seibadales más alejados de la isla principal. Por otra parte, grandes extensiones de pastos marinos han desaparecido a causa de la eutroficación y la sedimentación, lo cual ha reducido también el área vital para algunas especies. En la década del 80, se obtenía en el complejo estuarino aproximadamente 30-35% de la captura total de la zona. En el quinquenio 1991-1995 disminuyó a aproximadamente 26 %, pero mucho mayor fue su disminución en cuanto a calidad. Las especies más abundantes y explotadas en estos biotopos son generalmente de baja calidad y precio (Tabla 8). Entre las principales se cuentan: jaibas (Callinectes spp.), pataos y mojarras (Gerridae), salema (Archosargus rhomboidalis), macabí (Albula vulpes), machuelo (Opisthonema oglinum), lisas y lisetas (Mugil spp.), raya (Dasyatis americana), guaguancho (Sphyraena guachancho), verrugato (Micropogonias furnieri), corvinas (Bairdiella spp.), sierra, serrucho y pintada (Scomberomorus spp.), robalo (Centropomus undecimalis) y sábalo (Megalops atlanticus). El machuelo era la única especie planctófaga de importancia en el área (aunque también consume organismos del meiobentos que obtienen del sedimento). Las poblaciones de dicha especie sufrieron un drástico colapso a partir de 1990, al parecer provocado por sobrepesca, aunque coincidió con la primera fase de construcción del viaducto Caibarién-Santa María, el cual provocó graves alteraciones de la calidad ambiental en la Bahía Buenavista (ver Anexo 1). Los principales artes de pesca empleados en las zonas estuarinas son las redes de arrastre (manteletas y chinchorros) y en mucho menor medida los trasmallos (red de agallas). En el complejo seibadal-arrecife que ocupa un área intermedia entre el estuarino y el océano, cuyos límites son poco precisos y variables, existe aún importante influencia de los aportes terrígenos, pero con marcada mezcla de aguas oceánicas, los fondos son generalmente fango-arenosos y arenofangosos, cubiertos de seibadal, con arrecifes de parche por su porción norte, e interrumpida por los manglares que forman o bordean los cayos y cuya ictiofauna juega un papel importante en las capturas que se obtienen en este complejo ecológico. La turbidez del agua es menor, la salinidad menos variable y en general las condiciones más estables que cerca de la costa (Fig. 16). Estas condiciones generalmente se extienden hacia el borde de la plataforma hasta los 10-12 m de profundidad, donde empiezan a predominar fondos arenosos con cabezos y arrecifes de parche. Tabla 8 . Composición de las capturas comerciales en el Archipiélago Sabana-Camagüey. 1Especies pelágico-oceánicas; 2 – del batial superior; 3- de la pendiente arrecifal; 4- del complejo seibadal-arrecife; 5- del complejo estuarino. Hábitat Nombre común Nombre científico 1986-90 Ton 1 1 1 3 3,4 3 2 1 3,4 2 4 1 5,4 5,4 4,5 4,5 4,5 4,5 5 4,3 4,3 3 4 3 4 5 4,3 4 4 Grupo 1 Aguja Tetrapturus spp., Istiophorus platypterus Castero Makaira nigricans Emperador Xiphias gladius Aguají Mycteroperca bonaci Cubera Lutjanus cyanopterus, L. Griseus Cherna criolla Epinephelus striatus Cherna del alto Epìnephelus mystacinus Dorado Coryphaena hippurus Guasa Epinephelus itajara Pargos del alto Lutjanus vivanus, L. Buccanella, Rhomboplites aurorubens Pargo criollo Lutjanus analis Peto Acanthocybium solanderi Pintada Scomberomorus regalis Sierra, serrucho Scomberomorus maculatus, S. cavalla Otros Total Grupo 1 Grupo 2 Biajaiba Lutjanus synagris Caballerote Lutjanus griseus Cají Lutjanus apodus Jocú Lutjanus jocu Lisa Mugil lisa, M. curema Perro Lachnolaimus maximus Rabirrubia Ocyurus chrysurus Otros Total Grupo 2 Grupo 3 Arigua Mycteroperca venenosa Bajonao Calamus spp. Cabrilla Epinephelus guttatus Cibíes Caranx ruber, C. bartholomaei Liseta Mugil curema, M. trichodon, M. hospes, Otros Total Grupo 3 Grupo 4 Cojínua Caranx fusus Chicharro Selar crumenophthalmus Jiguagua Caranx hippos 1991-94 % Ton % Diferencia Ton 0,907 <0.1 0,714 <0.1 0,194 0,000 0 0,270 <0.1 18,174 0,3 25,268 0,4 0,025 <0.1 0,050 <0.1 10,674 0,2 8,674 0,2 -0,025 0,221 7,501 16,595 100,979 1,8 3,405 0,1 2,550 <0.1 1,101 <0.1 39,943 0,7 15,722 0,4 1,001 <0.1 0,304 <0.1 0,051 <0.1 9,030 0,2 85,258 2,404 2,246 1,051 30,914 303,142 5,3 1,448 <0.1 24,256 0,4 32,586 0,6 166,991 3,8 0,000 0 1,640 <0.1 16,784 0,4 136,151 1,448 22,616 15,802 0,208 <0.1 552,379 9,8 26,203 256,662 0,6 5,8 -25,995 295.717 335,825 5,9 80,952 1,4 34,852 0,6 8,623 0,2 96,023 1,7 10,068 0,2 94,988 1,7 1,333 <0.1 662,665 11,7 287,678 6,5 56,757 1,3 1,554 <0.1 0,755 <0.1 12,649 0,3 0,589 <0.1 37,219 0,8 11,708 0,3 408,909 9,2 48,148 24,196 33,298 7,868 83,374 9,479 57,769 -10,375 253,756 0,166 <0.1 66,457 1,5 1,738 <0.1 8,723 0,2 21,679 0,5 3,800 4,011 43,759 35,007 54,860 3,966 70,468 45,496 43,730 76,539 0,1 1,2 0,8 0,8 1,3 3,577 243,776 0,1 4,3 27,180 125,942 0,6 2,8 -23,602 117,834 51,447 42,222 107,884 0,9 0,7 1,9 6,680 4,127 14,605 0,2 0,1 0,3 44,767 38,095 93,278 Hábitat Nombre común Nombre científico 4 4,5 4 3,4 2,4 4,5 4 4,3 2 5 5 5 5 5 4 4 4 4,3 3 3,4 4,5 4,5 5 4,3 5 4 5 3,4 5 Jurel-Gallego Medregal Palometa Salmonete Caranx latus Seriola fasciata Trachinotus spp. Mulloidichthys martinicus, Pseudopeneus maculatus Tiburones Condrichthyes varios Raya – chucho. Dasyatis americana, Aetobatus narinari y otros Ojanco Lutjanus mahogoni Arnillo Apsilus dentatus Otros Total Grupo 4 Grupo 5 Corvina Bairdiella spp. Guaguancho Sphyraena guachancho Salema Archosargus rhomboidalis Patao-Mojarra Eugerres spp., Gerres cinereus Robalo Centropomus undecimalis Sardina de ley Harengula humeralis Jallao Haemulon album Otros Total Grupo 5 Grupo 6 Agujón Tylosurus spp. Barbero Acanthurus spp. Carajuelo Holocentrus spp. Cochino Balistes vetula Chopa Kyphosus spp. Jorobado Selene vomer Macabí Albula vulpes Ronco Haemulon spp. Sábalo Megalops atlanticus Vaca Lactophrys spp. Verrugato Micropogonias furnieri Vieja lora Scarus spp. Spasisoma spp. Otros Total Grupo 6 Grupo 7 Machuelo Opisthonema oglinum Otros Total Grupo 7 Total Escama 1986-90 1991-94 Ton % 51,753 0,9 2,854 0,1 48,206 0,8 6,615 0,1 Ton % 37,209 0,8 5,934 0,1 4,342 0,1 3,073 0,1 Diferencia Ton 14,544 -3,080 43,864 3,543 381,973 6,7 882,816 15,6 322,474 7,2 643,397 14,5 59,500 239,420 16,127 0,3 35,179 0,8 3,884 0,1 20,752 0,5 35,392 0,6 108,099 2,4 1631,173 28,7 1199,199 26,9 -19,052 -16,868 -72,708 431,974 119,511 2,1 5,037 0,1 114,474 32,957 0,6 5,842 0,1 27,115 116,514 2,1 33,036 0,7 83,478 660,528 11,6 709,567 15,9 -49,038 21,532 0,4 2,519 0,1 19,013 69,816 1,2 228,819 5,1 -159,003 3,102 0,1 1,971 <0.1 1,131 0,000 0 120,767 2,7 -120,767 1023,960 18 1099,299 24,7 -75,339 13,566 0,2 13,248 0,2 32,058 0,6 106,621 1,9 82,248 1,4 4,760 0,1 287,537 5,1 439,728 7,7 53,798 0,9 29,971 0,5 15,205 0,3 25,968 0,5 38,995 0,7 1143,703 20,1 348,923 68,489 417,412 5675,069 6,1 1,2 7,4 16,486 2,092 5,324 5,054 284,844 0,523 50,411 348,315 16,165 1,018 32,935 7,276 202,969 972,888 0,4 -2,919 <0.1 11,156 0,1 26,735 0,1 101,567 6,4 -202,596 <0.1 4,237 1,1 237,126 7,8 91,413 0,4 37,633 <0.1 28,953 0,7 -17,730 0,2 18,692 4,6 -163,974 21,9 170,815 69,531 317,479 387,010 4449,909 1,6 279,392 7,1 -248,990 8,7 30,402 1225160 Aunque Baisre (1984) incluye en este complejo a la pendiente arrecifal que bordea la plataforma (desde los 10-12 m de profundidad hasta el veril, a 25-30 m), hemos encontrado grandes diferencias en la composición de la ictiofauna de ambos biotopos, y se utilizan para su captura diferentes artes y métodos de pesca (Fig. 17). En este complejo seibadal-arrecife, que incluye la ictiofauna de los manglares, arenales y playas, se realizan las principales pesquerías del Archipiélago, principalmente con chinchorros y redes de sitio sobre las praderas de seibadal y en las costas de los cayos. Las principales especies que aquí se obtienen son: langosta espinosa (Panulirus argus), pargo criollo (Lutjanus analis), biajaiba (L. synagris), cubera y caballerote (L. cyanopterus, y L. griseus), rabirrubia (O. chrysurus), roncos (Haemulon spp.), bajonaos (Sparidae), jureles (Caranx spp., Selar crumenophthalmus), raya (D. americana), obispo (Aetobatus narinari), sierra, serrucho y pintada (Scomberomorus spp.), tiburón gata (Ginglymostoma cirratum), peces cofre (Lactophrys spp). Algunas de las especies del complejo litoral estuarino se capturan también en este hábitat, aunque en menor proporción (pataos y mojarras, macabí, machuelo, sierra, serrucho y pintada, etc.). Los artes de pesca empleados para estos recursos son esencialmente los chinchorros y otros artes de arrastre (manteletas), algunos artes de sitio (tranques o corrales) y algunos barcos emplean como arte secundario los palangres para tiburones neríticos o redes de fondo para batoideos. Este complejo faunístico aportó aproximadamente el 56% de la captura en el período 86-90 y aumentó a 62% en el 91-95. En la pendiente arrecifal que bordea la plataforma se capturan algunas de las especies que se obtienen en el complejo anterior (roncos, bajonaos, rabirrubia y otros en pequeña proporción), pero predominan: el ronco arará, Haemulon plumieri (20% del peso total), los meros (Serranidae), principalmente la cherna criolla, Epinephelus striatus, (11%), y en menor porcentaje, la rabirrubia, Ocyurus chrysurus, y el cochino, Balistes vetula, los barberos (Acanthurus) y otras especies poco numerosas. En este hábitat se obtiene solo el 2-3% de la captura total del archipiélago, aunque en el quinquenio 1986-1990 era 4-5%. Aunque este hábitat posee la mayor diversidad de especies (ver epígrafe 1), predominan especies arrecifales de pequeña talla que no son atrapadas por las nasas. Muchas de ellas poseen gran valor ornamental. En el batial superior de la plataforma, se realizan dos tipos de pesca diferentes: la de pargos y meros del alto (entre 100 y 300 m) y la de tiburones (entre 100 y 600 m). Para la captura de pargos y meros en el batial superior se emplean las llamadas nasas del alto y el chambel, ambos artes con gran variedad de adaptaciones según la tradición local, los recursos disponibles, etc. En estas capturas se obtienen principalmente: pargo del alto (Lutjanus vivanus) y en menor proporción pargo sesí (L. buccanella), mero del alto (Epinephelus mystacinus), cotorro (Rhomboplites aurorubens) y otros de forma esporádica. En la pesca de tiburones predomina el marrajo, Hexanchus griseus. Esta pesquería se realiza con palangres de fondo de 50 a 100 anzuelos No. 0-11 a 0-13. También en la zona oceánica adyacente a la plataforma, pero en el estrato superficial (0-100 m) se realiza la captura con palangre de deriva, en la cual también predominan por su peso los tiburones. En la estadística de las capturas se mezclan los datos de ambas pesquerías, por lo que no es posible diferenciar cuales corresponden a cada hábitat o arte de pesca. También en dicha pesca con palangre de deriva se obtienen peces de pico (agujas, emperador, castero) cuyas capturas fluctuaron alrededor de las 100 TM anuales en el quinquenio 86-90 disminuyendo posteriormente a menos de la mitad. Además en esta pesca se captura el peto (Acanthocybium solanderi) y el dorado (Coryphaena hyppurus). Esta última especie llegó a aportar 150-180 TM en los años 1990-1991 pero descendió notablemente con posterioridad. La mayor parte de esta captura se obtiene al norte de las provincias orientales. El uso de estos artes ha estado limitado en los últimos 10 años por la disponibilidad de materiales para su construcción. 5.3 Esfuerzo de pesca. Según Giménez (1990), en la década del 80 la pesca comercial operaba en el ASC unas 460 embarcaciones de pesca (incluyendo enviadas y auxiliares), de las cuales 280 se dedicaban a la captura de "escama" (todos los peces menos los atunes) y 20 de túnidos. Considerando el efecto dañino de tal esfuerzo, desde mediados de ese período se inició un proceso de ordenamiento que redujo el número de barcos, sin afectación de la producción pesquera. La cifra total se redujo a 243 en 1996. La Asociación las Villas, disponía de 85 embarcaciones dedicadas a la captura de escama y redujo dicha cifra a 62 en 1995. En el último quinquenio el número de barcos se ha mantenido en niveles similares a 1995-1996. Para la captura de peces se emplean chinchorros y redes, nasas de placer y de veril, palangres de fondo y de deriva (principalmente para tiburones), tranques (artes de sitio), trasmallos para batoideos y para peces pequeños. 1988 Redes 17% Tranques 3% Nasas del alto 11% Eventuales 0% Chinchorros 54% Nasas de placer 15% 1994-95 Redes 22% Eventuales 2% Tranques 2% Nasas del Alto 9% Chinchorros 52% Nasas de placer 13% Fig.18. Capturas por artes de pesca en la empresa de pesca de Caibarién, en 1988 y promedio para el período 1994-98. En la empresa pesquera de Caibarién, en 1988 había 24 barcos (8 chinchorreros, 5 rederos, 7 con nasas de placer y palangres, 2 con nasas del alto y 2 con tranques), en 1995-1999 se redujo a 17 (5 chinchorreros, 8 con redes, 3 con nasas de placer, 1 con nasas del alto, y 1 con tranque). Durante el período 1995-1998 los barcos chinchorreros aportaron el 44% de la captura de escama (con solo 3-4 barcos pescando), y las redes (manteletas) aportaron el 25%. Los demás artes aportaron en conjunto el 26% (Fig. 18). Estas proporciones son parecidos a las existentes en 1988 con un esfuerzo mayor, aunque con una disminución notable en el uso de los tranques. Realmente la mayoría de las embarcaciones usan más de un arte: los “naseros” utilizan también palangres, que calan generalmente después de terminada la labor con las nasas. Algunos “rederos” utilizan tranques o palangres, u otro arte adicional. Los barcos que operan con Nasas del Alto, eventualmente pescan con redes para sardinas en las aguas interiores, o con palangres de fondo en aguas profundas. Por ello, es frecuente encontrar en los datos estadísticos, datos de captura de especies que no se corresponden con el arte principal de la embarcación. 5.31. Artes de arrastre. Aunque existe diferenciación en el control estadístico de las capturas entre “chinchorros” y “redes o manteletas”, la diferencia real radica en la longitud de tales artes y otros parámetros que determinan el esfuerzo, como el número de pescadores y el tamaño de las embarcaciones. Los primeros operan con dos barcos, que hasta 1998 eran del tipo Cayo Largo (18,3 m de eslora), los cuales están siendo sustituidos por embarcaciones de plástico más pequeñas (13 m de eslora). Generalmente los chinchorreros operan con 8 pescadores y los rederos con 5 pescadores o menos. Hasta hace algunos años los chinchorreros, empleaban paños de 300-500 brazas de longitud (740925 m) (Giménez, 1988). En los últimos años algunas tripulaciones emplean chinchorros de más de 1000 brazas (1850 m) de longitud. Debido a la drástica disminución de las poblaciones de peces en las Bahías Los Perros y Jigüey y la Gloria, el tamaño de las redes o manteletas (principal arte usado en esas Bahías) se ha incrementado de 250-400 brazas, hasta más de 2000. Los arrastres que antes se hacían con una embarcación pequeña por períodos de una a dos horas, en la segunda mitad de la década del 90 se extendieron a 10-12 horas, con dos embarcaciones. Aún así, las capturas son pobres y formadas por especies de baja calidad (principalmente mojarras, pataos y salema), por lo que ha disminuido sustancialmente la pesca en esas bahías. Este gran aumento en el tamaño de los artes de arrastre implica un incremento del esfuerzo que no se cuantifica en la estadística pesquera. Tuvimos la oportunidad de comparar la captura obtenida con chinchorros de diferente longitud. La Fig. 19A muestra que no existe correlación entre el largo del chinchorro y la producción por arrastre. La aparente tendencia de la línea de regresión (con muy bajo nivel de correlación) a aumentar la captura con el largo del chinchorro, es debida a una sola muestra con una alta captura, mientras que en todas las demás la biomasa se mantiene igual o inferior que con los chinchorros más pequeños. Por el contrario, la línea de regresión largo del chinchorro contra rendimiento por lance, muestra una tendencia a disminuir (Fig. 19B), aunque también aquí el coeficiente de correlación es muy bajo. Es decir, que el uso de chinchorros más largos no conlleva ventajas ni para la captura total ni para los rendimientos. Por el contrario, el uso de chinchorros de mayor longitud trae consecuencias negativas para los hábitats: al ser más lento el movimiento del arte, aumenta su peso y presión sobre el fondo y resulta mucho más difícil para los pescadores evitar los obstáculos, por lo que se destruyen los refugios de invertebrados y peces, se remueven los sedimentos, tornándolos cada vez más finos y aumentando el tiempo de resuspensión. El uso de chinchorros pequeños, aunque también dañino, permite una mayor velocidad de movimiento, menor escape de los peces y menor estrés para los que escapan, reduce el tiempo de arrastre, disminuye el costo del arte, el cual es alto y todos sus componentes deben adquirirse en MNC, y provoca menores las afectaciones al medio marino. Fig. 19. Capturas (A) y rendimientos (B) por lance de chinchorro en relación con su longitud. Otro elemento muy negativo que conlleva la pesca con chinchorros, es el uso de mandarrias para destruir o “aplanar” el fondo, en particular los corales (Fig. 20). Estas estructuras, al igual que las gorgonias, las esponjas, las rocas y otros elementos del relieve, no solo brindan refugio a los peces, sino que además constituyen un elemento fundamental en el proceso de desarrollo de la trama alimentaria. Esta destrucción de los hábitats conlleva necesariamente a una disminución de la productividad biológica y pesquera. Fig. 20. Destrucción premeditada de corales durante el arrastre de los chinchorros. Debido a la actual crisis económica del país, al menos durante la década del 90, usualmente no era posible adquirir los tamaños de mallas adecuados, para la selección de tallas, por lo que durante esos años y aún hoy, no se cumplen las regulaciones establecidas al efecto. Por otra parte, el uso de chinchorros impide la colocación en el área de artes más selectivos, como nasas o artes de enmalle. 5.32. Nasas Las llamadas “nasas de placer” en el caso del ASC, realmente se ubican en una estrecha franja de la pendiente arrecifal, muy cerca del borde de la plataforma (a 20-30 m de profundidad) y muy raramente en las zonas bajas de esta, como pudiera entenderse por su nombre. Por tanto, la captura está formada esencialmente por especies demersales de los arrecifes coralinos, generalmente separados de los pastos marinos, por extensos arenales. Las nasas de placer son rectangulares, construidas con armazón de cabillas de hierro corrugado y forradas con malla hexagonal de alambre galvanizado, aunque en los últimos años se ha iniciado el uso de mayas de plástico. Está regulado el uso de mallas de 35 x 60 mm, pero debido a la carencia de recursos, generalmente se construyen con los materiales disponibles, más frecuentemente con mallas de 25x35 o de 28x40 mm, que no permiten el escape de los peces pequeños. Según Giménez (1990), en las cuatro empresas con conforman la Asociación de Las Villas, la captura con nasas constituyó el 12% del total en el período 1985-1989. La empresa pesquera de Caibarién aportó entonces un 15%, proporción que se mantiene cercana a esta en los últimos años (Fig. 18). Tal actividad ejerce un efecto negativo sobre los arrecifes, pues las nasas rompen muchos corales y dañan otras colonias que forman parte del arrecife. Por otra parte, es usual que, cuando los pescadores concluyen su campaña y regresan a tierra, dejan sus nasas caladas, a veces por períodos de 6-10 días, tiempo durante el cual muchos peces mueren atrapados en estas. Tales artes no están acondicionados con partes de material degradable, por lo que al ser abandonadas o perdidas, continúan atrapando peces por largo tiempo. Afortunadamente, en los últimos años, por la carencia de materiales, los barcos naseros funcionan con un número de nasas inferior al normado, y su uso se prolonga por más tiempo. 5.33 Nasas del Alto Son pocas las embarcaciones que realizan la pesca del alto en la zona. Generalmente esta pesca se combina con el uso palangres de fondo, también en aguas profundas. Las nasas del alto por lo general se construyen de forma cilíndrica, con cintas de cañas de Castilla doble, aunque también en los últimos años se forran con malla plástica hexagonal. Son generalmente de mayor tamaño que las nasas de placer. Se utilizan para la captura de pargos y meros del alto, a profundidades de 100 a 300 m. Debido a limitaciones tecnológicas y económico-sociales, el uso de este arte después de un pequeño incremento en la década del 80, disminuyó sustancialmente. Giménez (1990) reportó que en el período 1985-1989 los naseros del alto aportaban aproximadamente el 3,3% de la captura de la Asociación Pesquera de Las Villas. Según los datos de desembarques de la empresa de Caibarién, en el período 1994-1998 los naseros del alto aportaron el 11% de la captura de dicha empresa, pero el análisis de la composición por especies demuestra que solo el 0.2-0,5% procede de las capturas con nasas del alto, mientras que del 2 al 4,5% corresponde a tiburones, obtenidos con palangres, y el resto, es de especies de plataforma capturadas con redes (rayas, sardinas, jureles, pargo, biajaiba, rabirrubia, etc.). La captura de sardinas se realiza, con el objetivo de cumplir los planes de producción y se usa principalmente como carnada, pues pocas veces se comercializa, por su pequeña talla. 5.34 Tranques Las redes de sitio, llamadas generalmente “tranques”, se utilizan de forma limitada en el Archipiélago Sabana-Camagüey. Según Giménez (1990), en el período 1985-1989, en la Empresa Las Villas, este arte aportaba solo 0.1% de la captura. En la Empresa de Caibarién su aporte fluctúa actualmente entre 2 y 3%, obtenido por una sola embarcación. Aunque está regulado el uso de los “tranques”, en ocasiones estos son colocados en las entradas de canales o zonas de paso de reproductores, lo cual puede afectar el reclutamiento de años posteriores. 5.4 Volumen y composición de las capturas La Zona D aporta aproximadamente el 20% de la captura nacional, no obstante, aquí se incluye la “morralla” que forma parte de la fauna acompañante del camarón en la Zona SE. Por tanto, en la realidad el aporte de la Zona D en productos pesqueros es aproximadamente 25%. Su principal renglón lo constituyen los peces, los cuales representan más del 70% de la pesca en la zona y aportan unas 7000-8000 TM anuales, o sea, aproximadamente el 35% del total de la plataforma. Hasta inicios de la década del 90 esta cifra era de 8000-9000 TM. Se incluyen aquí unas 100 especies, aunque solo unas 40 tienen cierta importancia pesquera. Más del 70% de dicha captura se obtiene propiamente en el Archipiélago Sabana-Camagüey, el resto al norte de las provincias orientales y más recientemente, parte también en el Archipiélago Jardines de la Reina. Las capturas totales de la Zona D promediaron 8407,792 TM/año en el período 1982 a 1990 y disminuyeron a 6611,743 (en 18%) en el período 1991-1994. Las empresas ubicadas en el Archipiélago Sabana-Camagüey, y que realizan todas sus pesquerías en esta zona (Empresas de Matanzas, Las Villas, Ciego de Avila y Camagüey) aportan el 70% de la captura de la Zona D, aunque además hay que considerar que las Empresas de las provincias orientales (Tunas y Holguín), también obtienen parte de sus capturas en dicha región (Fig. 21). TunasHolguín 26% Matanzas 8% Las Villas 30% Camagüey 12% C. de Ávila 24% Fig. 21. Proporciones de las capturas totales de peces por asociaciones, en el archipiélago Sabana-Camagüey. La Asociación Pesquera de Las Villas, que realiza todas sus operaciones en el ASC, obtiene el 3133% de esa captura, y la empresa de Caibarién, la mayor del área, aporta casi el 50% de la captura de la Asociación. Al igual que en el resto del país, las pesquerías en la Zona D, tuvieron su etapa de máximo crecimiento a partir de 1960 y hasta 1975, lo que condujo a algunas situaciones de sobrepesca, y un desarrollo desmesurado de la flota de pesca. Después de una notable disminución de las capturas totales de peces, en los años 1976-1981, éstas se incrementaron paulatinamente por la adopción de algunas medidas de manejo más racional, incorporación de recursos poco explotados hasta entonces (rayas, chuchos, machuelo) alcanzando ya en 1985 niveles que parecen indicar la captura máxima sostenible (Fig. 22). No obstante, estas capturas se realizaron con un excesivo número de embarcaciones, por lo que a partir de 1988 se inició por el MIP un proceso de disminución del número de barcos. No hemos podido obtener datos confiables de esfuerzo para cada empresa de los últimos años, no obstante los principales cambios ocurridos en la Empresa de Caibarién constituyen un ejemplo de lo que ocurrió en toda el área: hasta 1987 operaban como promedio 30 embarcaciones para la captura de escama, y al reducir el número en l988 y 1989, aumentaron los rendimientos y la captura total (Fig. 23). Ya en 1990 empezaron los efectos del Período Especial, con una notable disminución del esfuerzo que determinó una baja de las capturas. Sin embargo, a partir de 1996 se inicia un proceso de recuperación y se alcanza el nivel de esfuerzo de los años 1988 y 1989 (expresado en días/mar) con solo 17 embarcaciones, pero en este caso no ocurrió aumento de las capturas y los rendimientos, como era de esperar después de varios años de esfuerzo reducido. En nuestro criterio, ello se debe al deterioro del medio marino y los recursos pesqueros, debido a tres factores principales que actuaron casi al mismo tiempo: 1- el deterioro de las condiciones ambientales por factores naturales, sobre el que hemos hecho referencia, 2- el deterioro de los hábitats por factores antrópicos (pedraplenes, contaminación, represamiento, uso histórico de métodos y artes de pesca nocivos), y 3- el incremento de la pesca no comercial , que ha sustituido en gran medida a la pesca comercial. Fig. 22. Etapas de desarrollo de la pesca, capturas anuales de peces en la Zona D y curva de tendencia de dichas capturas. De cerca de 1500-1600 TM en los años 1985-1989, las capturas de la Empresa de Caibarién disminuyeron durante el Período Especial, a solo 331 TM en 1992 con una recuperación posterior, pero muy por debajo de los niveles anteriores (Fig. 24). Las principales reducciones se manifestaron en los grupos de calidad o precio II, III y VII. Las reducciones más drásticas se observaron en las poblaciones de biajaiba (Grupo II), y el machuelo (Grupo 7). Esta última pesquería, prácticamente desapareció. Fig. 23. Capturas de peces (A), esfuerzo (B) y rendimientos (C), en la empresa pesquera de Caibarién. La Tabla 8 presenta la composición de las capturas de "escama" (promedios de 1986-1990 y 19911994) de las empresas de pesca que operan de forma total y permanente en el Archipiélago SabanaCamagüey (Matanzas, Villa Clara, Ciego de Ávila y Camagüey). Las especies más importantes por el volumen de captura, son los batoideos (15%). En este renglón se incluyen varias especies, aunque Dasyatis americana es la especie predominante, seguida del obispo, Aetobatus narinari. Al nivel de especie, la primera es la que brinda los mayores volúmenes de pesca en el Archipiélago. Ya desde el período 1991-1994 se observa una disminución de los rendimientos y las tallas de los batoideos, con relación al período 1981-1990, a pesar de que el esfuerzo pesquero disminuyó en los últimos años (L. Espinosa, comunicación personal). Hasta fines de la década del 80 el machuelo (Opisthonema oglinum) era después de la raya, Dasyatis americana, la especie que mayor volumen aportaba, pero sus poblaciones colapsaron drásticamente desde 1990. Dicha especie estaba sometida a un alto nivel de explotación pesquera que pudiera haber sido una de las causas de su casi desaparición. Fig. 24 Capturas anuales por grupos de especies de la Empresa Pesquera de Caibarién. Esa pesquería cesó hace varios años y no se observan signos de recuperación de la especie, a pesar de su corto ciclo de vida y alta tolerancia a cambios ambientales, lo cual pudiera estar relacionado con el deterioro de su hábitat. Esta especie se utilizaba principalmente como carnada aunque se inició la producción en conservas enlatadas, para lo cual parece tener condiciones favorables. La biajaiba (Lutjanus synagris) que es una de las principales especies comerciales, sufrió la etapa de sobrepesca de la década del 70, pero se recuperó rápidamente. Posteriormente, a partir del año 1987 sus poblaciones aumentaron rápidamente. Tal incremento se produjo esencialmente por las capturas del establecimiento de Caibarién, que se obtienen en aguas interiores de la Bahía San Juan de los Remedios. Otras especies de diferentes grupos, que habitan en la misma región, incrementaron también sus capturas en el quinquenio 1986-90, aunque en menor medida: del grupo 1, la sierra y el serrucho (Scomberomrorus spp.), del grupo 2, la rabirrubia (Ocyurus chrysurus) y el caballerote (L. griseus), del grupo 5 los pataos y mojarras y otras. Tabla 8 . Composición de las capturas comerciales en el Archipiélago Sabana-Camagüey. 1Especies pelágico-oceánicas; 2 – del batial superior; 3- de la pendiente arrecifal; 4- del complejo seibadal-arrecife; 5- del complejo estuarino. Hábitat Nombre común Nombre científico 1986-90 Ton 1 1 1 3 3,4 3 2 1 3,4 2 4 1 5,4 5,4 4,5 4,5 4,5 4,5 5 4,3 4,3 3 4 3 4 5 4,3 4 Grupo 1 Aguja Tetrapturus spp., Istiophorus platypterus Castero Makaira nigricans Emperador Xiphias gladius Aguají Mycteroperca bonaci Cubera Lutjanus cyanopterus, L. Griseus Cherna criolla Epinephelus striatus Cherna del alto Epìnephelus mystacinus Dorado Coryphaena hippurus Guasa Epinephelus itajara Pargos del alto Lutjanus vivanus, L. Buccanella, Rhomboplites aurorubens Pargo criollo Lutjanus analis Peto Acanthocybium solanderi Pintada Scomberomorus regalis Sierra, serrucho Scomberomorus maculatus, S. cavalla Otros Total Grupo 1 Grupo 2 Biajaiba Lutjanus synagris Caballerote Lutjanus griseus Cají Lutjanus apodus Jocú Lutjanus jocu Lisa Mugil lisa, M. curema Perro Lachnolaimus maximus Rabirubia Ocyurus chrysurus Otros Total Grupo 2 Grupo 3 Arigua Mycteroperca venenosa Bajonao Calamus Spp. Cabrilla Epinephelus guttatus Cibíes Caranx ruber, C. Bartholomaei Liseta Mugil curema, M. trichodon, M. hospes, Otros Total Grupo 3 Grupo 4 Cojínua Caranx chrysos Chicharro Selar crumenophthalmus 1991-94 % Ton % Diferencia Ton 0,907 <0.1 0,714 <0.1 0,194 0,000 0 0,270 <0.1 18,174 0,3 25,268 0,4 0,025 <0.1 0,050 <0.1 10,674 0,2 8,674 0,2 -0,025 0,221 7,501 16,595 100,979 1,8 3,405 0,1 2,550 <0.1 1,101 <0.1 39,943 0,7 15,722 0,4 1,001 <0.1 0,304 <0.1 0,051 <0.1 9,030 0,2 85,258 2,404 2,246 1,051 30,914 303,142 5,3 1,448 <0.1 24,256 0,4 32,586 0,6 166,991 3,8 0,000 0 1,640 <0.1 16,784 0,4 136,151 1,448 22,616 15,802 0,208 <0.1 552,379 9,8 26,203 256,662 0,6 5,8 -25,995 295.717 335,825 5,9 80,952 1,4 34,852 0,6 8,623 0,2 96,023 1,7 10,068 0,2 94,988 1,7 1,333 <0.1 662,665 11,7 287,678 6,5 56,757 1,3 1,554 <0.1 0,755 <0.1 12,649 0,3 0,589 <0.1 37,219 0,8 11,708 0,3 408,909 9,2 48,148 24,196 33,298 7,868 83,374 9,479 57,769 -10,375 253,756 0,166 <0.1 66,457 1,5 1,738 <0.1 8,723 0,2 21,679 0,5 3,800 4,011 43,759 35,007 54,860 3,966 70,468 45,496 43,730 76,539 0,1 1,2 0,8 0,8 1,3 3,577 243,776 0,1 4,3 27,180 125,942 0,6 2,8 -23,602 117,834 51,447 42,222 0,9 0,7 6,680 4,127 0,2 0,1 44,767 38,095 4 4 4,5 4 3,4 2,4 4,5 4 4,3 2 5 5 5 5 5 4 4 4 4,3 3 3,4 4,5 4,5 5 4,3 5 4 5 3,4 5 Jiguagua Jurel-Gallego Medregal Palometa Salmonete Caranx hippos Caranx latus Seriola fasciata Trachinotus spp. Mulloidichthys martinicus, Pseudopeneus maculatus Tiburónes Condrichthyes varios Raya – chucho. Dasyatis americana, Aetobatus narinari y otros Ojanco Lutjanus mahogoni Arnillo Apsilus dentatus Otros Total Grupo 4 Grupo 5 Corvina Baidiella spp. Guaguancho Sphyraena guachancho Salema Orthopristis rhomboidalis Patao-Mojarra Eugerres spp., Gerres cinereus Robalo Centropomus undecimalis Sardina de ley Harengula humeralis Jallao Haemulon album Otros Total Grupo 5 Grupo 6 Agujón Tylosurus spp. Barbero Acanthurus spp. Carajuelo Holocentrus spp. Cochino Balistes vetula Chopa Kyphosus spp. Jorobado Selene vomer Macabí Albula vulpes Ronco Haemulon spp. Sábalo Megalops atlanticus Vaca Lactophrys spp. Verrugato Micropogonias furnieri Vieja lora Scarus spp. Spasisoma spp. Otros Total Grupo 6 Grupo 7 Machuelo Opisthonema oglinum Otros Total Grupo 7 Total Escama 107,884 51,753 2,854 48,206 6,615 1,9 0,9 0,1 0,8 0,1 0,3 0,8 0,1 0,1 0,1 93,278 14,544 -3,080 43,864 3,543 322,474 7,2 643,397 14,5 59,500 239,420 16,127 0,3 35,179 0,8 3,884 0,1 20,752 0,5 35,392 0,6 108,099 2,4 1631,173 28,7 1199,199 26,9 -19,052 -16,868 -72,708 431,974 381,973 6,7 882,816 15,6 14,605 37,209 5,934 4,342 3,073 119,511 2,1 5,037 0,1 114,474 32,957 0,6 5,842 0,1 27,115 116,514 2,1 33,036 0,7 83,478 660,528 11,6 709,567 15,9 -49,038 21,532 0,4 2,519 0,1 19,013 69,816 1,2 228,819 5,1 -159,003 3,102 0,1 1,971 <0.1 1,131 0,000 0 120,767 2,7 -120,767 1023,960 18 1099,299 24,7 -75,339 13,566 0,2 13,248 0,2 32,058 0,6 106,621 1,9 82,248 1,4 4,760 0,1 287,537 5,1 439,728 7,7 53,798 0,9 29,971 0,5 15,205 0,3 25,968 0,5 38,995 0,7 1143,703 20,1 348,923 68,489 417,412 5675,069 6,1 1,2 7,4 16,486 2,092 5,324 5,054 284,844 0,523 50,411 348,315 16,165 1,018 32,935 7,276 202,969 972,888 0,4 -2,919 <0.1 11,156 0,1 26,735 0,1 101,567 6,4 -202,596 <0.1 4,237 1,1 237,126 7,8 91,413 0,4 37,633 <0.1 28,953 0,7 -17,730 0,2 18,692 4,6 -163,974 21,9 170,815 69,531 317,479 387,010 4449,909 1,6 279,392 7,1 -248,990 8,7 30,402 1225160 Al parecer, el incremento de tales especies estuvo relacionado con cambios provocados en el ecosistema por el huracán Kate en 1986. Después del mismo, se hizo ostensible el aumento del área de praderas marinas (Thalassia testudinum) en las Bahías de Buenavista y San Juan de los Remedios, gracias a la evacuación parcial de los fangos que cubrían el fondo. El resurgimiento de los seibadales en las zonas menos profundas de la bahía, evidentemente favoreció el reclutamiento y aumento de las poblaciones de biajaiba. A juicio de varios pescadores de gran experiencia, paralelamente disminuyó la abundancia de roncos pequeños y otras especies no comerciales (morralla), aparentemente como resultado de la competencia con la biajaiba y otros depredadores. No obstante, gran parte del área permanece aún cubierta por los sedimentos, en particular en la parte central de la Bahía de Buenavista. La población de biajaiba disminuyó drásticamente en las Bahías Buenavista y San Juan de los Remedios a partir del cierre de la comunicación entre dichas bahías por el pedraplén que une a Caibarién con Cayo Santa María, el cual, durante su primera etapa de construcción (1989-1990) no tenía drenajes que permitieran el flujo de agua entre dichos cuerpos de agua. Esta especie tiene su fase de reclutamiento y cría en la bahía de Buenavista, desde donde se asume que se trasladan hacia la Bahía de San Juan de los Remedios con el crecimiento. Dicho pedraplén cerró el paso hacia el sur de Cayo Santa María, donde muchos pescadores de bonito obtenían la manjúa que utilizaban como carnada viva, la cual ha pasado a ser un factor limitante en dicha pesca. El pargo criollo (Lutjanus analis), también sufrió una importante disminución en sus desembarques a partir de 1990 (casi en 50%), aunque en este caso no solo por diezmo de su captura sino porque es una de las especies preferidas en el mercado negro, por lo que parte importante de la captura no se entregaba a las empresas. Los pargos, que se cuentan entre las especies de mayor calidad, en conjunto aportaban más de 900 TM (15-18% de la captura total en 1986-1990) en la zona D, y disminuyeron durante el período especial a unas 500 TM (promedio de 1991-1994), o sea en más de 40%. En la empresa de Caibarién disminuyeron en 30%. La pesca de pargos y meros del alto ha sufrido grandes fluctuaciones, pero nunca alcanzó su potencial. La mayor captura se obtuvo en 1987 cuando sobrepasó las 125 toneladas, pero actualmente se limita a 40-60 TM anuales. De acuerdo con las evaluaciones realizadas por GarcíaRodríguez (1984) la biomasa de pargos del alto al norte del Archipiélago Sabana-Camagüey es de unas 2700 ton. Resulta evidente que es este uno de los pocos recursos pesqueros del país con cierta potencialidad de incremento en sus capturas. 5.5 Composición por especies y tallas de las capturas comerciales por artes. Con el objetivo de conocer la composición de la ictiofauna en los principales complejos ecológicos, y los efectos de la pesca sobre la misma, se realizó un estudio detallado de la composición de las capturas que se obtienen con chinchorros y nasas por los barcos de la empresa de Caibarién, así como algunas capturas realizadas con trasmallos en la Bahía de Buenavista (Fig. 25). 5.51 Capturas con chinchorros Se analizaron 6 rastreos de chinchorros realizados en la Bahía de Nazábal, 15 en la Bahía San Juan de los Remedios y 8 en la plataforma exterior de los cayos al norte de Cayo Fragoso y de la Bahía de Buenavista (Fig. 25). Estos rastreos cubren prácticamente toda el área con profundidades mayores de 1,5 m es decir hasta donde pueden navegar aún con dificultad las embarcaciones. La mayoría de los rastreos se hicieron en la región este de la Bahía San Juan de los Remedios, al norte de Caibarién, que es la zona más intensamente pescada por los chinchorreros, y probablemente la que mayor esfuerzo pesquero soporta en toda la plataforma cubana. La Tabla 9 muestra los principales indicadores de las capturas obtenidas, separando la captura comercial de la de especies e individuos que no tienen talla o calidad comercial, y que por ello son considerados como "morralla". Fig. 25. Estaciones de muestreo de las capturas comerciales con chinchorros (Ch), nasas (N) y trasmallos (T). Tabla 9. Indicadores de los muestreos realizados en las capturas comerciales con chichorros. Estación Prof Area Km2 Ch7 Ch8 Ch9 Ch10 Ch11 Ch12 Ch13 Ch14 Ch15 Ch16 Ch17 Ch18 Ch19 Ch20 Ch21 2-3 2-2.3 1.8-3 2-2.5 2-2.5 1.8-2 2-4 2.5-5 1,042 1,645 0,823 1,042 1,042 1,372 0,823 1,371 0,823 1,371 3,888 3,888 3,672 2,160 0,900 Densidad Biomasa #Ind/Km2 Kg/Km2 Captura total 870 151,382 5358 438,321 42663 1361,021 3377 500,209 2798 580,897 1482 228,604 1652 487,102 4231 656,306 4352 801,108 2923 337,505 996 289,070 303 78,918 610 272,147 4751 1025,708 1642 473,108 No. Fam. No. Sp. H´ R1 J 15 19 20 21 11 18 13 16 10 18 20 12 16 12 14 34 30 30 38 15 32 20 25 22 36 50 26 30 26 24 3,64 3,13 1,85 3,13 2,49 3,53 1,64 2,18 2,45 3,65 4,04 3,63 2,70 2,96 1,69 3,36 2,21 1,92 3,14 1,45 2,82 1,83 1,92 1,78 2,92 3,79 2,33 2,61 1,88 2,18 0,71 0,64 0,38 0,60 0,64 0,71 0,38 0,47 0,55 0,71 0,72 0,77 0,55 0,63 0,37 Ch22 Ch30 Ch31 Ch32 Ch33 Ch34 Ch35 Ch36 Ch37 Ch38 Ch39 Ch40 Ch41 Ch42 Ch7 Ch8 Ch9 Ch10 Ch11 Ch12 Ch13 Ch14 Ch15 Ch16 Ch17 Ch18 Ch19 Ch20 Ch21 Ch22 Ch30 Ch31 Ch32 Ch33 Ch34 Ch35 Ch36 Ch37 Ch38 Ch39 Ch40 Ch41 Ch42 Totales Ch7 1-2.5 1.6-4 2-3.5 1.5-2 2-4 2-6 1.5-2 2-4 2-3 2-3 2 1.5-4 3-5 2-3 2-2.5 1.8-3 2-2.5 2-2.5 1.8-2 2-4 2.5-5 2-3 2-3 2-4 2-4 2-4 0.5-3 2-3 1-2.5 1.6-4 2-3.5 1.5-2 2-4 2-6 1.5-2 2-4 2-3 2-3 2 1.5-4 3-5 2-3 2,160 1,569 0,939 1,424 0,300 0,578 1,500 1,728 1,980 0,480 0,870 0,878 2,281 2,068 1,042 1,645 0,823 1,042 1,042 1,372 0,823 1,371 0,823 1,371 3,888 3,888 3,672 2,160 0,900 2,160 1,569 0,939 1,424 0,300 0,578 1,500 1,728 1,980 0,480 0,870 0,878 2,281 2,068 44,618 1,042 2672 586,880 799 272,650 4837 681,330 2148 500,341 4407 995,018 2571 711,334 8752 732,706 5951 855,249 1320 427,548 12935 2312,667 911 667,558 8939 880,963 3834 666,732 1686 486,094 Captura Comercial 378 135,480 1145 254,917 2129 335,881 629 396,166 816 500,984 835 200,527 1639 456,209 4150 583,056 2753 618,019 770 225,675 454 251,183 169 55,172 291 256,454 3737 958,241 1633 449,969 1647 473,190 381 232,584 1179 326,160 560 273,916 1163 616,685 512 382,031 3644 290,582 997 591,554 422 313,039 5779 1989,583 629 634,617 1536 416,406 3400 626,275 1028 444,014 1330 390,746 Morralla 492 16,070 16 17 18 16 10 17 13 12 12 10 14 13 12 8 31 27 35 35 18 29 26 22 20 23 26 27 22 15 3,60 2,39 2,88 3,48 2,79 3,66 1,80 2,60 3,42 3,19 3,22 2,65 2,15 2,84 2,10 1,77 2,83 2,94 1,64 2,66 1,83 1,52 1,67 1,75 2,60 2,01 1,60 1,19 0,73 0,57 0,56 0,68 0,67 0,75 0,38 0,58 0,79 0,71 0,69 0,56 0,48 0,73 11 15 13 12 12 8 11 12 6 9 11 7 10 11 12 13 9 7 9 7 10 6 7 8 8 4 7 11 7 26 20 19 19 18 17 14 18 20 11 15 19 18 15 23 20 21 13 17 21 11 16 17 12 13 12 10 17 20 13 65 3,46 2,34 3,08 2,94 2,67 2,80 1,57 2,08 2,37 3,20 3,32 3,12 1,60 2,54 1,65 3,24 1,74 2,71 3,13 2,19 3,03 1,26 1,86 2,49 2,85 2,68 3,07 1,90 2,24 3,92 2,20 1,65 1,67 1,82 1,64 1,28 1,63 1,52 0,90 1,39 1,67 1,82 1,39 1,70 1,81 1,70 1,30 1,58 2,07 1,18 1,83 1,27 1,02 1,24 0,96 0,99 1,54 1,47 1,09 4,04 0,80 0,55 0,73 0,70 0,65 0,74 0,38 0,48 0,68 0,82 0,78 0,75 0,41 0,56 0,38 0,74 0,47 0,66 0,71 0,63 0,76 0,31 0,52 0,67 0,80 0,81 0,75 0,44 0,61 0,65 10 19 2,10 2,00 0,49 Ch8 Ch9 Ch10 Ch11 Ch12 Ch13 Ch14 Ch15 Ch16 Ch17 Ch18 Ch19 Ch20 Ch21 Ch22 Ch30 Ch31 Ch32 Ch33 Ch34 Ch35 Ch36 Ch37 Ch38 Ch39 Ch40 Ch41 Ch42 Totales Promedio ±DE 2-2.5 1.8-3 2-2.5 2-2.5 1.8-2 2-4 2.5-5 2-3 2-3 2-4 2-4 2-4 2-0.5 2-3 1-2.5 1.6-4 2-3.5 1.5-2 2-4 2-6 1.5-2 2-4 2-3 2-3 2 1.5-4 3-5 1,645 0,823 1,042 1,042 1,372 0,823 1,371 0,823 1,371 3,888 3,888 3,672 2,160 0,900 2,160 1,569 0,939 1,424 0,300 0,578 1,500 1,728 1,980 0,483 0,870 0,878 2,280 2,068 44,619 4213 40535 2749 1983 647 13 82 1599 2152 542 135 319 1014 9 1025 417 3658 1588 3243 2059 5108 4954 898 7114 283 7403 435 658 2203 3287 7459 183,404 1025,140 104,043 79,914 28,077 30,893 73,249 183,089 111,831 37,887 23,746 15,693 67,468 23,139 113,690 40,066 355,169 226,425 378,333 329,303 442,124 263,695 114,508 321,172 32,941 464,558 40,482 42,080 127,099 178,213 214,355 14 15 18 19 15 2 9 10 17 16 13 11 9 4 13 14 17 17 10 15 13 8 10 10 14 13 7 6 20 24 30 25 24 2 10 18 28 26 17 21 18 4 20 22 32 30 17 25 25 12 18 20 22 22 9 10 12 4 20 7 3,05 1,63 2,45 1,52 2,65 0,99 2,71 1,21 3,15 2,72 2,92 2,19 1,94 1,75 2,97 3,04 2,66 3,03 2,44 3,38 2,06 2,18 3,03 3,06 2,76 2,19 1,73 2,08 1,49 1,53 2,53 2,18 2,35 0,29 1,32 1,64 2,34 2,26 1,77 1,96 1,53 1,00 1,71 2,24 2,64 2,60 1,61 2,35 1,86 0,84 1,57 1,62 2,64 1,66 0,80 0,86 0,71 0,36 0,50 0,33 0,58 0,99 0,81 0,29 0,65 0,58 0,71 0,50 0,47 0,87 0,69 0,68 0,53 0,62 0,60 0,73 0,44 0,61 0,73 0,71 0,62 0,49 0,55 0,63 Se obtuvieron en los muestreos realizados, 114 especies de peces, de los cuales 70 son aceptados para el consumo aunque solo unas 40 tienen cierta importancia económica (más de un kg/km2). La Fig. 26 muestra los valores de biomasa total y biomasa comercial, obtenidos en cada lance, así como otros indicadores de la estructura de la comunidad y la Fig. 27 los valores promedio para las tres regiones estudiadas. Los mejores rendimientos en biomasa por área se obtuvieron en la Bahía San Juan de los Remedios, seguida de las obtenidas en aguas exteriores al norte de Cayo Fragoso y de la Bahía Buenavista, mientras que en Bahía Nazabal se obtuvo la mayor abundancia de morralla, principalmente mojarritas, salema y machuelo, aunque con una notable diferencia entre estaciones. Las especies más importantes por su número en las capturas fueron los pataos y mojarras, pero el 95% de los ejemplares eran de tallas no comerciales, solo el 12 % del peso total correspondió a peces de tallas comerciales (Tabla 10). Aunque la mayoría de estas especies son de pequeña talla y no alcanzan nunca el tamaño comercial, si juegan un importante papel en el ecosistema al utilizar los pequeños organismos que habitan en los sedimentos y la vegetación acuática y transferir su energía a los niveles tróficos superiores, que son objeto de pesca. Entre las especies comerciales las más importantes por su abundancia y peso fueron: el pargo criollo (Lutjanus analis), la biajaiba (L. synagris) y la rabirrubia (Ocyurus chrysurus). No obstante, resulta preocupante el alto número de individuos juveniles de estas últimas dos especies, los cuales mueren durante la captura. Los pargos en su conjunto (Lutjanidae) conformaron el 20% de los individuos y el 41% del peso total de la captura comercial. Le siguen en importancia los jureles (Carangidae), entre los que se destaca el chicharro, Selar crumenophthalmus (10% de los individuos y 5,6% de la captura total en peso) y el cibí amarillo, Caranx bartholomaei (3,1% de la captura). A nivel de especie, la raya (Dasyatis americana) ocupó el tercer lugar, después del pargo criollo y la biajaiba. Estas dos últimas especies parecen ser más abundantes en la región de Caibarién que en otras del ASC. Fig. 26. Captura total (rectángulos claros) y captura comercial (rectángulos oscuros) de pces por área barrida por los chinchorros. Fig. 27. Rendimientos promedio (kg/km2) en las capturas con chinchorros realizadas en tres subzonas de pesca de la empresa pesquera de Caibarién. Tabla 10. Composición por especies, valores de densidad y biomasa, porcentajes correspondientes a morralla, e indicadores de Frecuencia/abundancia-biomasa (F/a-b) y de Jerarquía (IJ) por especie, de las capturas con chinchorros. ESPECIES DASYATIDAE Dasyatis americana Urolophus jamaicensis MYLIOBATIDAE Aetobatus narinari ELOPIDAE Elops saurus Megalops atlanticus ALBULIDAE Albula vulpes MURAENIDAE Gymnothorax funebris Gymnothorax moringa CLUPEIDAE Harengula clupeola Harengula humeralis Jenkinsia lamprotaenia Opisthonema oglinum ENGRAULIDAE Cetengraulis edentulus BATRACHOIDIDAE Opsanus beta OGCOCEPHALIDAE Ogcocephalus nasutus Ogcocephalus vespertilio EXOCOETIDAE Hemirhamphus brasiliensis Chiodorus atherinoides BELONIDAE Tylosurus cocodrilus HOLOCENTRIDAE Holocentrus ascensionis FISTULARIIDAE Fistularia tabacaria SYNGNATHIDAE Densidad % Biomasa % F/a-b IJ Ind/Km2 Morralla kg/Km2 Morralla 6,500 23,1 27,290 3,4 5,290 5,5 26,734 1,4 B/3-2 29 1,210 100,0 0,556 100,0 C/4-4 54 0,583 0,0 7,220 0,0 0,583 0,0 7,220 0,0 C/4-2 55 11,095 6,1 9,200 0,7 10,916 6,2 4,695 1,3 C/3-3 33 0,179 0,0 4,505 0,0 C/4-3 99 25,619 2,1 9,057 0,6 25,619 2,1 9,057 0,6 B/3-2 19 0,045 100,0 0,081 100,0 0,022 100,0 0,050 100,0 C/4-4 98 0,022 100,0 0,034 100,0 180,948 90,8 8,028 92,7 16,945 2,1 0,592 1,5 C/4-4 68 0,112 100,0 0,002 100,0 C/4-4 88 18,447 100,0 1,04 100,0 161,403 100,0 7,182 100,0 B/2-2 7 88,535 100,0 1,290 100,0 88,535 100,0 1,290 100,0 C/3-4 26 0,045 100,0 0,000 100,0 0,045 100,0 0,000 100,0 C/4-4 85 0,448 100,0 0,032 100,0 0,426 100,0 0,020 100,0 C/4-4 84 0,022 100,0 0,012 100,0 C/4-4 82 0,964 100,0 0,182 100,0 0,336 100,0 0,154 100,0 C/4-4 104 0,628 100,0 0,029 100,0 C/4-4 53 0,359 18,8 0,412 3,3 0,269 58,0 0,412 1,8 C/4-4 79 0,134 100,0 0,021 100,0 0,134 100,0 0,021 100,0 C/4-4 67 0,067 100,0 0,203 100,0 0,067 100,0 0,203 100,0 C/4-4 71 0,022 100,0 0,000 100,0 Hippocampus erectus CENTROPOMIDAE Centropomus undecimalis SERRANIDAE Diplectrum formosum Mycteroperca bonaci APOGONIDAE Astrapogon stellatus RACHYCENTRIDAE Rachycentron canadum ECHENEIDAE Echeneis naucrates CARANGIDAE Caranx bartholomaei Caranx chrysos Caranx hippos Caranx latus Caranx ruber Chloroscombrus chrysurus Decapterus punctatus Oligoplites saurus Selar crumenophthalmus Selene vomer Seriola dumerili Seriola rivoliana Trachinotus falcatus LUTJANIDAE Lutjanus analis Lutjanus apodus Lutjanus cyanopterus Lutjanus griseus Lutjanus jocu Lutjanus synagris Ocyurus chrysurus LOBOTIDAE Lobotes surinamensis GERREIDAE Eucinostomus gula Eucinostomus jonesi Gerres cinereus Eugerres plumieri Eucinostomus sp. Eucinostomus argenteus HAEMULIDAE Anisotremus virginicus Haemulon album Haemulon melanurum 0,022 0,403 0,403 0,045 0,022 0,022 0,022 0,022 0,291 0,291 0,157 0,157 499,697 26,717 18,088 2,040 17,483 32,657 5,178 8,383 5,312 376,488 4,864 0,919 0,022 3,564 748,717 197,893 1,457 1,098 38,574 0,090 291,516 248,325 0,067 0,067 835,728 22,011 56,640 59,509 183,615 508,797 5,155 414,659 0,852 1,412 0,381 100,0 0,0 0,0 0,0 0,0 0,0 100,0 100,0 38,5 38,5 100,0 100,0 24,4 8,1 8,1 17,6 8,3 23,8 98,7 100,0 100,0 22,8 53,0 36,6 0,0 94,3 29,1 4,0 50,8 0,0 2,5 75,0 19,3 73,4 0,0 0,0 95,2 100,0 100,0 81,1 84,2 100,0 100,0 61,3 28,9 6,3 0,0 0,000 2,439 2,439 0,040 0,002 0,038 0,000 0,000 1,732 1,732 0,014 0,014 69,833 16,646 5,012 0,403 2,178 7,080 0,380 0,193 0,495 29,862 1,041 6,036 0,381 0,390 216,465 93,441 0,307 7,032 11,686 0,026 72,735 33,066 0,028 0,028 23,113 0,431 1,890 3,741 7,898 9,006 0,147 32,146 0,203 0,940 0,063 100,0 0,0 0,0 0,0 0,0 0,0 100,0 100,0 0,4 0,4 100,0 100,0 6,5 1,3 2,1 5,4 3,6 9,8 94,1 100,0 100,0 6,9 19,1 0,0 0,0 86,7 9,4 1,0 40,9 0,0 1,0 60,6 4,6 53,2 0,0 0,0 88,5 100,0 100,0 66,9 81,9 100,0 100,0 37,3 7,9 1,9 0,0 C/4-4 73 C/4-4 62 C/4-4 75 C/4-4 80 C/4-4 60 C/4-4 63 B/2-2 16 B/3-3 21 B/4-4 47 B/3-3 28 B/2-2 14 C/3-4 37 C/3-4 81 C/3-4 34 B/1-2 5 C/4-4 61 C/4-2 91 C/4-4 102 C/4-4 48 A/2-1 C/4-4 C/4-2 B/2-2 C/4-4 A/1-1 A/2-2 4 45 59 13 95 1 3 C/4-4 53 C/4-4 B/3-3 B/3-3 C/4-4 C/3-4 C/3-4 83 23 30 44 46 C/4-4 48 C/4-3 105 B/3-3 18 Haemulon aurolineatum Haemulon flavolineatum Haemulon parrai Haemulon plumieri Haemulon sciurus Orthopristis chrysoptera Pomadasys crocro SPARIDAE Archosargus rhomboidalis Calamus bajonao Calamus calamus Calamus penna Calamus pennatula Calamus sp. SCIAENIDAE Bairdiella batabana Bairdiella ronchus MULLIDAE Mulloidichthys martinicus EPHIPPIDAE Chaetodipterus faber CHAETODONTIDAE Chaetodon ocellatus POMACANTHIDAE Pomacanthus arcuatus Pomacanthus paru LABRIDAE Halichoeres radiatus Lachnolaimus maximus SCARIDAE Sparisoma chrysopterum MUGILIDAE Mugil curema SPHYRAENIDAE Sphyraena barracuda Sphyraena guachancho ACANTHURIDAE Acanthurus bahianus Acanthurus coeruleus Acanthurus chirurgus SCOMBRIDAE Scomberomorus cavalla Scomberomorus maculatus Scomberomorus regalis DACTYLOPTERIDAE Dactylopterus volitans BOTHIDAE 22,638 1,166 418,716 262,804 118,122 94,856 0,022 196,391 102,320 12,260 0,269 58,837 24,812 5,335 3,631 3,631 32,747 10,086 10,086 12,776 12,776 0,560 0,560 2,981 2,241 0,740 0,672 0,045 0,628 142,531 142,531 0,224 0,224 17,012 16,452 0,560 40,099 0,448 0,448 43,685 7,778 0,359 1,009 6,522 0,022 0,022 0,022 100,0 100,0 98,3 62,4 55,9 100,0 100,0 70,7 81,6 5,9 16,7 87,9 36,0 25,2 34,6 34,6 100,0 11,1 11,1 80,7 80,7 100,0 100,0 51,9 40,0 87,9 53,3 0,0 57,1 75,9 75,9 100,0 100,0 12,3 9,3 100,0 67,7 100,0 85,0 70,5 21,0 0,0 2,2 26,5 100,0 100,0 100,0 0,709 0,034 23,698 18,240 10,816 5,223 0,001 19,861 6,120 3,801 0,047 6,319 2,946 1,073 0,431 0,431 1,193 1,523 1,523 2,192 2,192 0,039 0,039 0,548 0,474 0,074 0,292 0,023 0,269 13,991 13,991 0,025 0,025 7,477 7,367 0,109 4,431 0,036 0,055 4,699 4,192 0,490 0,547 3,186 0,008 0,008 0,001 100,0 100,0 95,6 37,4 39,8 100,0 100,0 44,2 61,8 1,5 3,8 73,1 22,7 8,0 15,8 15,8 100,0 5,2 5,2 15,4 15,4 100,0 100,0 29,3 20,6 84,6 12,3 0,0 13,3 70,2 70,2 100,0 100,0 57,1 56,5 100,0 51,1 100,0 87,8 54,0 8,8 0,0 0,3 12,4 100,0 100,0 100,0 C/4-4 C/4-4 B/2-2 A/1-2 B/2-2 C/2-2 C/4-4 52 69 8 2 9 22 78 B/2-3 B/3-3 C/4-4 C/2-2 A/3-3 C/3-3 17 25 65 77 15 74 C/4-4 C/2-3 42 72 B/3-3 24 C/4-4 66 C/3-3 38 C/4-3 C/4-4 39 49 C/4-4 C/4-4 76 51 B/2-2 6 B/4-3 B/3-3 36 20 C/4-4 C/4-4 B/2-3 58 57 12 C/4-3 B/4-3 B/3-3 94 43 27 C/4-4 86 Syacium micrurum SOLEIDAE Achirus lineatus BALISTIDAE Aluterus schoepfi Aluterus scriptus Balistes capriscus Balistes vetula Cantherines pullus Monacanthus ciliatus Monacanthus setifer OSTRACIIDAE Lactophrys bicaudalis Lactophrys polygonia Lactophrys quadricornis Lactophrys trigonus Lactophrys triqueter TETRAODONTIDAE Lagocephalus laevigatus Sphoeroides spengleri Sphoeroides testudineus DIODONTIDAE Chilomycterus schoepfi Diodon holacanthus Diodon hystrix XENOCONGRIDAE Chilorhinus suensoni 0,022 0,202 0,224 8,069 2,040 0,045 0,067 4,841 0,134 0,403 0,538 82,215 0,067 0,336 49,356 32,455 0,134 5,783 0,022 5,402 0,359 6,836 3,160 0,157 3,519 0,022 0,022 100,0 100,0 100,0 41,9 100,0 100,0 0,0 6,0 50,0 100,0 100,0 60,1 100,0 100,0 66,4 49,9 100,0 100,0 100,0 100,0 100,0 100,0 100,0 100,0 100,0 100,0 100,0 0,001 0,006 0,006 2,343 0,295 0,012 0,049 1,679 0,292 0,004 0,013 36,957 0,058 0,073 10,012 26,812 0,027 0,714 0,009 0,640 0,065 8,282 0,491 0,033 7,758 0,001 0,001 100,0 100,0 100,0 15,2 100,0 100,0 0,0 1,8 0,8 100,0 100,0 36,1 100,0 100,0 47,8 31,5 100,0 100,0 100,0 100,0 100,0 100,0 100,0 100,0 100,0 100,0 100,0 C/4-4 C/4-4 87 C/4-4 C/4-4 C/4-4 C/3-3 C/4-4 C/4-4 C/4-4 41 89 90 32 92 56 50 C/4-4 C/4-4 A/2-2 A/2-2 C/4-4 96 97 10 11 100 C/4-4 101 B/4-4 40 C/4-4 64 B/4-4 C/4-4 B/3-2 38 70 31 C/4-4 106 5.52 Capturas con nasas de placer. Se analizaron las capturas obtenidas por 360 nasas, correspondientes a 6 campañas de pesca de dos barcos, en la pendiente arrecifal de la zona al Norte de Cayo Fragoso y de la Bahía de Buenavista (Fig. 25). Se obtuvo una captura promedio de 5020±1816 g por nasa/dia (Tabla 11), cifra muy superior a la que se obtiene en las islas de las Antillas Menores (Munro, 1983; Gobert, 1990), donde por lo general no sobrepasan los 100 g por nasa. Tabla 11. Indicadores de los muestreos realizados en las capturas comerciales con nasas. Estación Prof No. Densidad Biomasa No. No. H´ R1 Nasas Ind/Nasa g/Nasa Fam. Spp. Captura Total N1 25-30 32 45 5406 19 42 3,19 3,91 N2 40-50 89 49 4881 17 43 1,58 3,48 N3 30 54 6 1995 15 39 3,36 4,52 N4 15 67 36 5655 14 31 2,83 2,67 N5 25 59 40 4596 16 45 3,29 3,92 N6 15 59 53 7586 14 33 2,36 2,76 360 39 5035 J 0,59 0,29 0,64 0,57 0,60 0,47 N1 N2 N3 N4 N5 N6 25-30 40-50 30 15 25 15 32 89 54 67 59 59 360 15 40-50 30 15 25 15 32 89 54 67 59 59 360 To Captura Comercial 10 2806 28 3557 3 1838 23 4877 17 3212 31 6091 20 3837 12 12 13 11 9 10 24 29 27 24 25 23 3,17 1,54 3,67 2,71 2,51 2,48 2,78 2,48 3,47 2,17 2,41 2,03 0,69 0,32 0,77 0,59 0,54 0,55 14 12 5 7 15 10 33 22 13 14 28 20 2,36 1,27 1,72 2,10 2,94 1,52 2,76 1,94 1,64 1,34 2,59 1,83 0,47 0,28 0,47 0,55 0,61 0,35 11±4 22±8 TOTAL N1 N2 N3 N4 N5 N6 TOTAL Promedio±DE 98 21 3 13 24 22 24 Morralla 13987 1324 156 778 1384 1495 2221 30±34 3187±5315 La Tabla 12 muestra los valores de la densidad y biomasa promedios por especies, los porcentajes de cada una que se consideran como morralla y los indicadores de frecuencia abundancia e índice de jerarquía. La especie dominante por su número y biomasa en las nasas fue el ronco arará (Haemulon plumieri), que constituyó el 57% de los individuos y el 36% por su peso. Le siguió en orden de importancia la rabirrubia (Ocyurus crhysurus) (6,8 y 7,7% por su número y peso respectivamente), el cají, Lutjanus apodus (2,1 y 2,9%) y la cherna criolla (0,7 y 6,8%), esta última, con muy baja densidad, pero relativamente alta biomasa gracias a su talla. Las poblaciones de esta especie se encuentran sobre-explotadas en todas las plataformas de Cuba (Claro et al., 1990) al igual que en todo el Gran Caribe (Sadovy, 1994). Tabla 12. Composición por especies, valores de densidad y biomasa, porcentajes correspondientes a morralla e índices de frecuencia-abundancia (F/a-b) y de jerarquía (IJ) de las capturas con nasas. ESPECIES Densidad Morralla Biomasa Morralla F/a-b IJ (Ind/nasa) (%) (g/nasa) (%) MURAENIDAE 0.1222 100.0 211.11 100.0 Gymnothorax funebris 0.0472 100.0 139.40 100.0 A/3-2 28 Gymnothorax moringa 0.0750 100.0 71.71 100.0 A/3-2 26 HOLOCENTRIDAE 1.1222 100.0 76.38 100.0 Holocentrus ascensionis 0.2778 100.0 27.26 100.0 B/3-3 21 Holocentrus rufus 0.7806 100.0 44.86 100.0 A/2-3 8 Myripristis jacobus 0.0639 100.0 4.26 100.0 B/3-4 35 AULOSTOMIDAE 0.0028 100.0 0.86 100.0 Aulostomus maculatus 0.0028 100.0 0.86 100.0 C/4-4 59 SERRANIDAE 1.2444 6.9 884.56 0.7 Ephinephelus adscensionis 0.0028 0.0 7.85 0.0 C/4-3 60 Epinephelus cruentatus 0.0639 26.1 14.36 9.9 B/3-3 31 Epinephelus fulvus 0.5167 7.5 103.30 2.7 A/2-2 11 Epinephelus guttatus Epinephelus morio Epinephelus striatus Hypoplectrus puella Mycteroperca bonaci Mycteroperca venenosa GRAMMISTIDAE Rypticus saponaceus MALACANTHIDAE Malacanthus plumieri CARANGIDAE Caranx bartholomaei Caranx fusus Caranx ruber LUTJANIDAE Lutjanus analis Lutjanus apodus Lutjanus buccanella Lutjanus griseus Lutjanus jocu Lutjanus vivanus Ocyurus chrysurus HAEMULIDAE Anisotremus virginicus Haemulon album Haemulon aurolineatum Haemulon chrysergyreum Haemulon flavolineatum Haemulon melanurum Haemulon plumieri Haemulon sciurus Haemulon striatum SPARIDAE Calamus bajonao Calamus calamus Calamus penna Calamus pennatula SCIAENIDAE Equetus lanceolatus MULLIDAE Mulloidichthys martinicus Pseudopeneus maculatus CHAETODONTIDAE Chaetodon aculeatus Chaetodon capistratus Chaetodon ocellatus Chaetodon sedentarius 0.3250 0.0056 0.2556 0.0111 0.0306 0.0333 0.0028 0.0028 0.1583 0.1583 0.2944 0.0194 0.0028 0.2722 3.5472 0.0083 0.8389 0.0167 0.0083 0.0111 0.0028 2.6611 26.7722 0.1361 0.0056 0.8167 0.0222 2.8583 0.0667 22.3361 0.5139 0.0167 0.6278 0.0139 0.0028 0.0139 0.5972 0.0028 0.0028 1.0972 0.1944 0.9028 1.2444 0.0111 0.8083 0.0944 0.0833 3.4 0.0 0.0 100.0 0.0 25.0 100.0 100.0 42.1 42.1 25.5 0.0 0.0 27.6 3.4 0.0 5.0 100.0 0.0 0.0 100.0 2.2 54.1 10.2 0.0 100.0 100.0 98.3 0.0 47.6 35.7 99.8 14.6 40.0 0.0 0.0 14.4 100.0 100.0 0.5 0.0 0.6 100.0 100.0 100.0 100.0 100.0 159.78 11.06 433.15 0.24 139.18 15.65 0.79 0.79 35.96 35.96 125.48 34.40 5.90 85.17 561.52 7.53 148.18 0.68 2.68 15.50 0.15 386.79 2072.80 29.46 3.17 33.50 1.42 146.43 12.78 1797.16 47.93 0.95 133.61 3.40 0.39 5.46 124.37 0.07 0.07 106.44 21.05 85.40 33.60 0.06 14.47 6.12 1.69 0.5 0.0 0.0 100.0 0.0 4.8 100.0 100.0 37.5 37.5 4.3 0.0 0.0 6.3 1.2 0.0 1.9 100.0 0.0 0.0 100.0 0.8 34.5 3.4 0.0 100.0 100.0 97.7 0.0 29.1 24.3 100.3 3.2 8.2 0.0 0.0 3.3 100.0 100.0 0.2 0.0 0.2 100.0 100.0 100.0 100.0 100.0 A/3-2 C/4-3 A/3-2 C/4-4 A/4-2 B/4-3 14 61 16 55 33 34 C/4-4 62 B/3-3 23 A/4-3 C/4-3 A/3-2 36 63 15 B/4-3 B/2-2 B/4-4 B/4-4 B/4-3 C/4-4 A/2-2 50 10 46 51 43 64 3 A/3-3 C/4-4 A/2-3 C/4-4 A/2-2 C/3-3 A/1-1 A/2-3 B/4-4 22 65 6 52 2 38 1 12 47 B/4-4 C/4-4 B/4-3 A/2-2 40 66 48 9 C/4-4 67 A/3-3 A/2-2 20 5 C/4-4 A/2-3 A/3-3 B/3-4 56 7 25 27 Chaetodon striatus POMACANTHIDAE Holacanthus ciliaris Holacanthus tricolor Pomacanthus arcuatus POMACENTRIDAE Abudefduf saxatilis Chromis multilineatus LABRIDAE Bodianus rufus Halichoeres garnotti Lachnolaimus maximus SCARIDAE Scarus taeniopterus Sparisoma aurofrenatum Sparisoma chrysopterum SPHYRAENIDAE Sphyraena barracuda ACANTHURIDAE Acanthurus bahianus Acanthurus coeruleus Acanthurus chirurgus BALISTIDAE Balistes vetula OSTRACIIDAE Lactophrys trigonus Lactophrys triqueter TETRAODONTIDAE Canthigaster rostrata 0.2472 0.0972 0.0306 0.0194 0.0472 0.0389 0.0250 0.0139 0.2556 0.0056 0.2361 0.0139 0.1917 0.0556 0.0222 0.1139 0.0111 0.0111 1.5389 0.3833 0.1111 1.0444 0.5389 0.5389 0.0194 0.0083 0.0111 0.0083 0.0083 100.0 14.3 0.0 71.6 0.0 100.0 100.0 100.0 72.8 0.0 78.8 0.0 34.8 35.0 50.1 31.7 100.0 100.0 46.2 84.8 7.5 36.2 3.1 3.1 0.0 0.0 0.0 100.0 100.0 11.26 65.19 13.24 1.62 50.33 1.49 1.17 0.32 32.79 1.71 14.52 16.56 19.70 4.46 0.94 14.29 42.50 42.50 290.04 29.03 37.14 223.88 327.51 327.51 12.46 6.99 5.47 0.44 0.44 100.0 1.4 0.0 57.4 0.0 100.0 100.0 100.0 31.6 0.0 71.4 0.0 19.4 24.7 44.9 16.1 100.0 100.0 22.0 82.2 3.2 17.4 0.4 0.4 0.0 0.0 0.0 100.0 100.0 A/3-3 18 B/4-3 B/4-4 B/3-2 37 42 32 B/4-4 B/4-4 41 49 B/4-4 A/3-3 B/4-3 57 19 39 B/3-4 C/4-4 B/3-3 30 53 29 B/4-3 44 B/3-3 A/3-3 A/2-2 17 24 4 A/2-2 13 B/4-3 B/4-3 54 45 C/4-4 58 5.53 Capturas con trasmallo Fueron analizadas 6 capturas con trasmallos, realizados en la parte sur de la Bahía de Buenavista (Fig. 24). Se obtuvo una captura promedio de 57.89 kg por 1000 m2 de área cubierta por el arte al formar un círculo encerrando los peces (Tabla 13). En la captura obtenida con este arte, la proporción de peces con tallas inferiores a la comercial es prácticamente nula, debido al alto poder de selectividad del mismo. Las especies dominantes fueron el patao (Eugerres plumieri), la mojarra (Gerres cinereus), y la picúa (Sphyraena barracuda), esta última recibida como S. guachancho. Tabla 13. Indicadores de los muestreos realizados en las capturas comerciales con trasmallos en Bahía de Buenavista. Estación Prof. Largo Densidad Biomasa No. No. H´ R1 J Trasmallo #Ind/100m2 kg/100m2 Fam. Spp. trasmallo trasmallo T23 2.5 1569 62,5 17,69 6 10 2,80 1,36 0,84 T24 2.5 939 240,6 75.53 4 6 0,81 0,64 0,31 T25 2-3 1428 342,5 77,81 13 16 2,04 1,68 0,51 18000 16000 14000 No. de peces 12000 10000 8000 6000 4000 2000 0 0 20 40 60 80 100 120 140 160 Longitud (cm) T26 1-1,5 T27 1-2 T28 1-2 T29 .5-1 Total Promedio ±DE 303 965 2501 1728 9433 171,8 347,1 16,8 365,7 198,7 221,0 142,2 61,92 91,44 9,61 71,25 49,18 57,89 31,56 4 8 7 9 17 7 3 5 9 11 13 30 10 4 0,74 1,88 3,10 2,74 3.43 0,70 0,95 1,85 1,29 2.63 0,32 0,59 0,90 0,74 0.70 5.6 Composición por tallas de las capturas En los 29 lances de chinchorros analizados, se capturaron 162970 peces de los cuales se midieron 157629. El 64.6% de ellos fueron identificados como morralla, por no considerarse comestibles o no alcanzar tallas comerciales (aproximadamente 18 cm en la mayoría de las especies) (Fig. 27). Este criterio se tomo a partir de la captura que se recibe realmente, ya que en la práctica, como se verá más adelante, muchos peces se reciben con tallas inferiores a la mínima legal por lo que la proporción de peces que no alcanzan talla comercial está cercana al 70%. Es decir, que por cada 5 individuos que se capturan, solo dos son aprovechados por la pesca. Sin embargo, los peces comerciales constituyeron el 75% del peso total de la captura, y el resto a la morralla. Se incluyen en la morralla los juveniles de especies comerciales tales como rabirrubia (Ocyurus chrysurus), biajaiba (Lutjanus synagris), pargo criollo (L. analis), roncos (Haemulon plumieri, H. sciurus), bajonaos (Calamus spp.), jureles (Caranx spp,), loros (Sparisoma spp, Scarus spp.), etc., aunque una alta proporción está formada por especies que no tienen aceptación comercial por su calidad o talla, tales como las mojarras (Eucinostomus spp.), algunos roncos pequeños (H. aurolineatum, H. flavolineatum), picúas, lijas, guanábanas, morenas, etc. Fig. 27. Composición por largos de las capturas con chinchorros en la empresa pesquera de Caibarién. La Tabla 14 relaciona las tallas mínimas legales establecidas por la Resolución correspondiente del MIP, y el porcentaje de ejemplares que en nuestros muestreos tuvieron tallas inferiores a esta, tanto en las capturas con chinchorros como en las nasas. Se incluyen en esta lista solo las especies para las cuales hemos obtenido suficiente información en nuestros muestreos. Se brinda la talla mínima en largo calculada para el peso mínimo tomando como base a ecuaciones largo-peso confiables, aunque lamentablemente no existen para todas las especies. Estas pudieran ser empleadas para un mejor manejo de estos recursos, aunque de todas formas se requiere de una revisión más detallada de las tallas mínimas considerando otros factores tales como la talla de maduración sexual, talla máxima, tasa de crecimiento, calidad para el comercio, etc. Tabla 14. Proporción de peces con tallas inferiores a la mínima legal según la Resolución de 1996 del MIP, en las capturas con chinchorros y nasas de la empresa pesquera de Caibarién (CAIMAR). Especies Talla mínima Proporción de Talla Legal según ind. con tallas Estimada Resolución del MIP inferiores a la Para el mínima peso (%) mínimo. Nombre común Nombre científico Peso Largo En nPor chorros (g) (onzas) (cm) Nasas Ecuación Largopeso Calamus sp. Bajonao 120 4 17 28.9 79.7 17 Elops saurus Banano 85 3 21 2.4 25 Acanthurus spp. Barberos 342 12 23 90.9 100 26 Lutjanus synagris Biajaiba 120 4 18 31 18 Lutjanus griseus Caballerote 170 6 22 12 22 Epinephelus guttatus Cabrilla 170 6 22 6.3 22 Lutjanus apodus Cají 170 6 19 19.7 7.5 21 Anisotremus virginicus Catalineta 170 6 34.7 18 Chapín (Torito) Lactophrys quadricornis 460 16 90.2 28 Chapín (Vaquita) Lactophrys trigonus y 460 16 29.5 29 otros Epinephelus striatus Cherna criolla 570 20 32 18.2 39 Selar crumenophthalmus 120 Chicharro 4 95.5 19 Pomacanthus spp. Chivirica 120 4 15 5 47.4 15 Chopa (Salema) Archosargus 120 4 95.8 17 rhomboidalis Caranx bartholomaei Cibí amarillo 230 8 22 7.5 22 Caranx ruber Cibí carbonero 230 8 22 54.4 57 22 Balistes vetula Cochino 230 8 31.6 19.5 21 Caranx fusus Cojinúa 170 6 36 22 Lutjanus cyanopterus Cubera 460 16 31 32 Bairdiella spp, Corvina 120 4 18 18 Epinephelus cruentatus Enjambre 170 6 37.5 22 Sphyraena guachancho Guaguancho 170 6 43 31-34 Guatívere Jallao Jiguagua Jurel o Gallego Macabí Machuelo Mojarra Pargo criollo Patao Pez perro Pintada Rabirrubia Ronco amarillo Epinephelus fulvus Haemulon album Caranx hippos Caranx latus Albula vulpes Opisthonema oglinum Gerres cinereus Lutjanus analis Eugerres spp. Lachnolaimus maximus Scomberomorus regalis Ocyurus chrysurus Haemulon sciurus 120 170 340 230 120 120 120 230 120 285 285 140 120 4 6 12 8 4 4 4 8 4 10 10 5 4 16 21 23 23 17 20 17 24 16 24 30 20 17 Ronco arará Ronco blanco Salmonetes Haemulon plumieri Haemulon parrai Mulloidichthys martinicus y Pseudopeneus maculatus Scomberomorus maculatus Sparisoma chrysopterum 120 120 4 4 17 17 70.8 80.7 85.8 18.5 18 120 4 18 53,5 85,4 19 420 16 36 38 85,4 34 170 6 22 7,5 100 19 Serrucho Vieja lora 8.8 10 40.5 57.4 4.2 70.8 100 15 100 60.7 16.2 99.4 56 58.9 85.8 61.7 19 21 25 20 20 17 24 21 25 36 21 18 La Fig. 28 muestra la distribución de frecuencias por tallas de algunas de las especies más importantes en las capturas. La alta proporción de juveniles de rabirrubia, biajaiba, bajonaos y otras especies, provoca una alta mortalidad que, sumada a las afectaciones ambientales, determinan la disminución paulatina de la productividad biológica de la zona. Los valores mostrados en la Fig. 28 se basan en los desembarques, pero en la práctica, gran número de ejemplares son recibidos con tallas inferiores a las mínimas legales o en algunos casos no están incluidos en la Resolución que regula las mismas, en algunos casos por falta de unidad o definición en el uso de los nombres comunes de las especies. Así por ejemplo, de las 9 especies de loros que pueden ser objeto de pesca, solo incluye la regulación el nombre "vieja lora" y no se especifica a que especie se refiere. Asumimos que se trata del Scarus guacamaia, el cual raramente cae en las capturas. Se establece para esa especie una talla mínima de 170 g (aproximadamente 22 cm). De la familia de los loros sin embargo, se capturan varias especies de muy diferente talla y crecimiento, lo cual no se considera por la regulación existente. La especie más abundante, tanto en la pesca con nasas como con chinchorros, es Sparisoma chrysopterum (loro colirrojo) de pequeña talla, el cual se recibe desde los 16-17 cm LT, y ningún ejemplar alcanza la talla mínima establecida para la “vieja lora” que es el único nombre común al cual se puede adjudicar tal captura. Para los barberos, la mencionada Resolución establece una talla mínima de 23 cm LH (6 onzas), que resulta más alta de lo necesario. En consecuencia, no se respeta dicha talla y se reciben peces desde 15-16 cm, a la cual muchos son juveniles. En el caso de los peces cofre (Lactophrys trigonus, L. quadricornis, L. triqueter), la regulación sólo menciona una especie, el "chapín", para la cual establece un peso de 460 g (28 cm). La principal especie en las capturas es L. trigonus, de mayor talla y crecimiento, para la cual pudiera ser adecuada la talla mínima establecida, sin embargo, L. quadricornis es de mucho menor talla por lo que el 90% de los individuos en las capturas no alcanzan dicha talla, aunque en la práctica se reciben casi todos los ejemplares mayores de 20 cm LT, violándose la resolución. La alta proporción de juveniles de rabirrubia, biajaiba, bajonaos y otras especies, provoca una alta mortalidad que, sumada a las afectaciones ambientales, determinan la disminución paulatina de la productividad biológica de la zona. Los valores mostrados en la Fig. 28 se basan en los desembarques, pero en la práctica, gran número de ejemplares son recibidos con tallas inferiores a las mínimas legales o en algunos casos no están incluidos en la Resolución que regula las mismas, en algunos casos por falta de unidad o definición en el uso de los nombres comunes de las especies. Así por ejemplo, de las 9 especies de loros que pueden ser objeto de pesca, solo incluye la regulación el nombre "vieja lora" y no se especifica a que especie se refiere. Asumimos que se trata del Scarus guacamaia, el cual raramente cae en las capturas. Se establece para esa especie una talla mínima de 170 g (aproximadamente 22 cm). De la familia de los loros sin embargo, se capturan varias especies de muy diferente talla y crecimiento, lo cual no se considera por la regulación existente. La especie más abundante, tanto en la pesca con nasas como con chinchorros, es Sparisoma chrysopterum (loro colirrojo) de pequeña talla, el cual se recibe desde los 16-17 cm LT, y ningún ejemplar alcanza la talla mínima establecida para la "vieja lora" que es el único nombre común al cual se puede adjudicar tal captura. En las capturas con nasas se obtuvieron 67 especies, de las cuales 43 son utilizadas para el consumo, aunque muchas de ellas se capturan en pequeña cantidad. Parece contradictorio que siendo el lugar de captura el de mayor diversidad de especies (la pendiente arrecifal) el número de especies es menor que en los fondos blandos. Ello se debe, por una parte, a que este hábitat cubre un área muy pequeña en comparación con el segundo, y por otra, a que muchos de sus habitantes son especies de muy pequeña talla, con hábitos crípticos, que entran y salen libremente de las nasas, o simplemente no penetran en ella. Fig. 28A. Composición por tallas de especies seleccionadas en las capturas con chinchorros. La línea vertical y el número sobre esta indican la talla mínima legal. Fig. 28B. Composición por tallas de especies seleccionadas en las capturas con chinchorros. Fig. 29A. Composición por tallas de algunas especies en las capturas con nasas de la empresa pesquera de Caibarién. Fig. 29B. Composición por tallas de algunas especies en las capturas con nasas de la empresa pesquera de Caibarién. En estas capturas también se encuentra un número apreciable de peces con tallas inferiores a las mínimas establecidas, aunque en mucho menor proporción (Fig. 29). El 47% de los ejemplares capturados fueron considerados como morralla, los cuales constituyeron el 24% del peso total de la captura. En este caso, la causa principal es el hecho de que se están utilizando mallas con tamaños inferiores a las establecidas por el MIP: en lugar de mallas de 35 x 60 mm, en la práctica la mayoría de las nasas tienen mallas de 25 x 35 o 28 x 40 mm. Una solución aceptada desde hace muchos años, pero cuya aplicación en la práctica se ha perdido, es el uso de las mallas adecuadas, al menos en las tapas de las nasas, lo cual pudiera solucionar en parte el problema con un menor costo, hasta que sea posible utilizar los materiales adecuados para el arte completo. No obstante, el efecto nocivo de este arte es mucho menor que el de los chinchorros, ya que durante la captura la mayoría de los peces son devueltos vivos al mar, con mucho menor daño físico que los capturados con chinchorros. Las mencionadas irregularidades en cuanto a la talla mínima legal, se manifiestan en otras especies que no relacionamos para no extender innecesariamente este documento. No obstante, otras deficiencias deben ser analizadas, como el hecho de que bajo un mismo nombre se incluyen especies con diferentes características biológicas. Por otra parte, las tallas mínimas se establecieron tomando como base al peso, el cual es difícil de comprobar en las condiciones materiales de los centros de acopio, aunque para muchas especies se da un largo estimado. Estos últimos en varios casos no se ajustan a la realidad, por lo que deben ser revisados, y las que no la tienen deben ser calculadas para facilitar su aplicación. Como resultado de las deficiencias de la regulación sobre las tallas mínimas de captura, se viola la resolución conscientemente, pues se reciben peces a tallas muy inferiores a la establecida y algunas de estas no son las adecuadas. En la realidad, para muchas de estas especies no existe información sobre las tallas de maduración sexual y crecimiento, por lo que las tallas mínimas legales no se ajustan a la necesidad real de proteger el recurso, sino más bien a conceptos de mercado. Aunque una parte de los peces pequeños es devuelta viva al agua, la mayor parte, al menos de los que se capturan con chinchorros, mueren o quedan tan afectados por la pesca que no pueden sobrevivir. Por tanto, de esta captura se derivan dos afectaciones serias a los recursos pesqueros: la eliminación de los juveniles, en detrimento de la producción pesquera futura, y la eliminación de una gran parte de la biomasa alimentaria para los depredadores, los cuales constituyen el recurso pesquero de mayor valor. Tal situación sólo puede ser resuelta con el establecimiento de Areas Marinas Protegidas y un adecuado sistema de vigilancia (más detalles en el epígrafe siguiente). La ictiofauna de la región ofrece otras opciones que no son explotadas. Son abundantes en algunas áreas, especies altamente cotizadas para la pesca de vara y carrete (spinning), la cual se practica con el método de “pescar y soltar” (catch and release), como el sábalo (Megalops atlanticus) y el macabí (Albula vulpes), las cuales son explotadas intensamente por la pesca comercial a pesar de su bajo valor de mercado, más bien por incrementar el volumen de las capturas que por su aporte económico. La captura comercial de macabí en el la Zona D superó las 500 TM anuales (350 en el ASC) en la década del 80, y disminuyó a menos de 100 como promedio en los últimos años (unas 50-70 TM en el ASC). Tal disminución evidentemente, es consecuencia del deterioro ocurrido en las zonas estuarinas del archipiélago. No obstante, aún se conservan suficientes recursos para el desarrollo de una pesca deportiva altamente cotizada y que puede brindar beneficios muy superiores a la pesca comercial. En el caso del sábalo, los datos estadísticos existentes no permiten tal comparación, pero sus capturas son inferiores y probablemente haya sufrido por igual las consecuencias ambientales. Durante nuestros muestreos encontramos poblaciones de ambas especies con perspectivas para esta pesca en la Bahía de Nazábal, donde en particular los sábalos alcanzan tallas superiores a 1 m. La pesca con chinchorro en esa bahía, si bien es productiva, está formada principalmente por especies de baja calidad y pequeña talla (la mayoría “morralla”), y destruye la posibilidad de explotación de un turismo especializado de mayores perspectivas. También son abundantes el macabí y el sábalo, en Punta Madruguilla al SW de Cayo Santa María, en la Bahía de Buenavista, en el Puerto de Sagua y las cercanías de La Panchita y Carahatas. Para la protección de este recurso, resulta necesario prohibir la captura en regiones seleccionadas, principalmente la que se realiza con cualquier tipo de redes (chinchorros, manteletas, trasmallos). La pesca deportiva de peces de pico (agujas, emperador, castero) pudiera ser una opción más para el turismo en la región. Este tipo de pesca al parecer no ha sido valorado, pero a juzgar por las rutas migratorias de estas especies, es probable su desarrollo. 6. MANEJO DE LA ICTIOFAUNA EN RELACIÓN CON EL TURISMO Y OTRAS OPCIONES 6.1 Desarrollo del turismo El Archipiélago Sabana-Camagüey es en la actualidad la región del país en la que se desarrolla con mayor intensidad el turismo internacional, fundamentalmente vinculado al medio marino. Algunas de las obras que se ejecutan para crear infraestructuras necesarias, provocan graves daños al medio marino, como se ha explicado anteriormente, en detrimento de la oferta que se brinda al turista. No se toman en cuenta algunos de los valores naturales que podrían incrementar y/o mantener la atracción turística. En las áreas dedicadas al turismo no existen regulaciones adecuadas ni una estrategia de educación dirigida a los guías de turismo y a los propios usuarios, para la protección de los arrecifes y sus comunidades asociadas por lo que, en áreas dedicadas al buceo contemplativo y de campismo se producen daños mecánicos a los mismos y una extracción incontrolada de corales pétreos, moluscos y otros organismos, se realiza pesca furtiva (con o sin licencias) que provoca estrés en las poblaciones de peces, se tiran anclas sobre los corales destruyendo muchos de ellos, se echa basura (latas, bolsas de polietileno, papeles, etc.) y se violan en general normas elementales de conservación, denotando un bajo nivel de cultura ambiental. Alcolado et al., (2000) describen en detalle el estado actual de deterioro de los arrecifes coralinos de la región, y en el primer capítulo de este informe se evalúa la situación actual de la ictiofauna que habita en esos arrecifes. El deterioro de su diversidad y abundancia conlleva a la eliminación de una de las opciones principales para el turismo: el buceo contemplativo, que cobra cada vez más adeptos en el Caribe. La ausencia de peces grandes en estos arrecifes y la conducta asustadiza de los que quedan, son consecuencia, por una parte, del deterioro de los arrecifes y por otra de la pesca comercial y fundamentalmente la pesca submarina, que se realiza con arpón, la cual se practica por un elevado número de personas, la mayoría hoy día, con licencias otorgadas por el MIP. Tal situación sólo puede ser resuelta con el establecimiento de Areas Marinas Protegidas y un adecuado sistema de vigilancia (más detalles en el epígrafe siguiente). La ictiofauna de la región ofrece otras opciones que no son explotadas. Son abundantes en algunas áreas, especies altamente cotizadas para la pesca de vara y carrite (spining), la cual se practica con el método de "pescar y soltar" (catch and release), como el sábalo (Megalops atlanticus) y el macabí (Albula vulpes), las cuales son explotadas intensamente por la pesca comercial a pesar de su bajo valor de mercado, más bien por incrementar el volumen de las capturas que por su aporte económico. La captura comercial de macabí en la Zona D susperó las 500 TM anuales (350 en el ASC) en la década del 80, y disminuyò a menos de 100 como promedio en los últimos años (unas 50-70 TM en el ASC). Tal disminución evidentemente, es consecuencia del deterioro ocurrido en las zonas estuarinas del archipiélago. No obstante, aún se conservan suficientes recursos para el desarrollo de una pesca deportiva altamente cotizada y que puede brindar beneficios muy superiores a la pesca comercial. En el caso del sábalo, los datos estadísticos existentes no permiten tal comparación, pero sus capturas son inferiores y probablemente haya sufrido por igual las consecuencias ambientales. Durante nuestros muestreos encontramos poblaciones de ambas especies con perspectivas para esta pesca en la Bahía de Nazabal, donde en particular los sábalos alcanzan tallas superiores a un m. La pesca con chinchorro en esa bahía, si bien es productiva, está formada principalmente por especies de baja calidad y pequeña talla (la mayoría "morralla"), y destruye la posibilidad de explotación de un turismo especializado de mayores perspectivas. También son abundantes el macabí y el sábalo en Punta Madruguilla, al SW de Cayo Santa María, en la Bahía de Buenavista, en el Puerto de Sagua y en las cercanías de La Panchita y Carahatas. Para la protección de este recurso, resulta necesario prohibir la captura en regiones seleccionadas, principalmente la que se realiza con cualquier tipo de redes (chinchorros, manteletas, trasmallos). La pesca deportiva de peces de pico (agujas, emperador, castero) pudiera ser una opción para el turismo en la región. Este tipo de pesca al parecer no ha sido valorado, pero a juzgar por las rutas migratorias de estas especies, es probable su desarrollo. 6.2 La pesca recreativa La llamada pesca "deportiva" o recreativa en Cuba, desde hace muchos años se ha convertido en una pesca de subsistencia, y muy en particular a partir del Período Especial. La mayor parte de la captura que se obtiene por esta vía se vende en el mercado negro, o en el mejor de los casos para el consumo familiar. La magnitud de la extracción pesquera que se obtiene por esta vía no se ha valorado, pero a juzgar por las informaciones obtenidas durante este proyecto, y de algunas estimaciones primarias, su volumen es apreciable y merece ser evaluado de forma más concreta si se pretende hacer un manejo de los recursos pesqueros. Desde la década del 60 la "pesca deportiva" fue organizada por el Instituto Nacional de Deportes, Educación Física y Recreación (INDER), aunque tal organización no incluía ningún tipo de control sobre la extracción pesquera. A partir del inicio del Período Especial aumentó extraordinariamente el número de pescadores, controlados o no por el INDER. De la pesca con cordel y anzuelo se pasó al uso de artes masivos (chinchorros, trasmallos, etc.), en la mayoría de los casos obtenidos ilegalmente de las empresas pesqueras o de los pescadores profesionales. En 1997 el MIP estableció el otorgamiento de licencias de pesca deportivo-recreativa, e inicia el control de dicha actividad a través de la ONIP, lo cual logró reducir tanto la extracción pesquera como el desvío de recursos. En Septiembre del 2000 según datos de la ONIP el número de licencias de pesca "deportivorecreativa" en todo el país ascendía a 4079 para la pesca submarina y 4070 para embarcaciones. Estas cifras fueron muy superiores en 1997 y disminuyen a un ritmo de 6-10% anualmente gracias a nuevas restricciones (reducción del tiempo de pesca a los fines de semana, aumento del precio de la licencia, mayor actividad de inspección). Estas licencias otorgan el derecho a extraer, por cada salida al mar, hasta 15 kg de pescado en zonas de gran interés económico-pesquero establecidas en el Reglamento de Pesca y hasta 30 kg en zonas abiertas de menor interés económico pesquero. Cuando se trate de especies de gran tamaño capturadas en las zonas antes referidas, cuyo peso unitario sea superior a los 15 kilogramos (33 libras) se autorizan hasta tres ejemplares por pescador, y no se establecen límites para las especies migratorias tales como castero, aguja, emperador, dorado, bonito o listado, comevíveres, albacora y atún aleta amarilla. En base al número licencias otorgadas por provincias, hemos estimado para el área del Archipiélago Sabana-Camagüey aproximadamente 670 licencias de pesca submarina y 840 para embarcaciones. Asumiendo que cada una realiza 50 salidas al año (una semanal), y en base a la cuota autorizada, se obtiene una captura de 1700-1800 TM al año. En nuestro criterio estas cifras son conservadoras, ya que una gran parte de los portadores de estas licencias utilizan el producto de la pesca para la venta a precios elevados o por apremiantes necesidades de consumo, por lo cual realizan tantas salidas como les sea posible. La captura obtenida por este tipo de pesca en la década del 90, antes del establecimiento de las licencias, puede haber sido varias veces superior, ya que: a) existía mayor necesidad de alimentos, b) no existía control ni restricciones sobre la actividad, c) se podían utilizar artes de mayor productividad. Aún hoy día son pocos los que cumplen con las cuotas de captura máxima establecidas y existe un número apreciable de personas que pescan sin licencia. A modo de ejemplo, durante una investigación de este problema en el Municipio Moa, el propio delegado del INDER y otros funcionarios, estimaron que el número de personas que pescan ilegalmente es igual o algo mayor que los que lo hacen con licencias y la captura obtenida por estas es muy superior a la de la pesca comercial. Aunque estas burdas estimaciones no pretenden caracterizar el problema, al menos dan una idea de la magnitud del mismo y su posible incidencia sobre los recursos pesqueros. Debe tomase en cuenta además, que esta actividad parece haber alcanzado su máximo auge en la segunda mitad de la década del 90, cuando ocurrió la degradación de los arrecifes coralinos a que hemos hecho referencia anteriormente, por lo que asumimos que su impacto sobre los recursos pesqueros debe haber sido considerable. Además de la extracción pesquera, esta actividad ejerce otros impactos indirectos sobre el medio marino: • La pesca submarina extrae los reproductores más grandes, genéticamente más aptos para el mantenimiento de las poblaciones, con mayor eficiencia que la pesca con artes convencionales. • La actividad de caza provoca reacciones defensivas en los peces (dispersión, conducta huidiza), que en las zonas utilizadas para el turismo disminuyen su valor estético-recreativo. • Muchos pescadores extraen organismos de valor ornamental (corales, gorgonias, conchas de moluscos), en muchos casos con fines lucrativos. • Se incrementa el número de embarcaciones que tiran anclas sobre los corales, el derrame de combustibles y lubricantes, la basura y otras formas de contaminación y degradación ambiental (sobre estas agresiones al medio no existen regulaciones adecuadas en el país, ni actividad de vigilancia). 7. LAS AREAS PROTEGIDAS MARINAS COMO MECANISMO DE PROTECCIÓN Y USO SOSTENIBLE DE LA ICTIOFAUNA. La experiencia internacional ha demostrado fehacientemente los beneficios que brindan las Áreas Protegidas Marinas a la pesca y para otras opciones de manejo de los recursos marinos. No se conoce en la literatura científica ningún caso en que estas reservas no hayan tenido resultados positivos para la pesca. Los beneficios son evidentes ya al cabo de 2-4 años después del establecimiento de la reserva (Alcalá, 1988; Bohnsack, 1994; Davis, 1989; FAO, 1994; García-Rubies y M. Zabala, 1990; Man et al., 1994; Plan Development Team 1990; Project Reefkeeper, 1992; Russ, 1985; 1996; Russ, G.R., y A.C. Alcalá. 1996; Russ, G.R., y A.C. Alcalá. En prensa; Shorthouse, 1990; While, 1986; y otros). Resulta pues contraproducente que en el ASC, la región marina del país con mayor desarrollo socioeconómico en la actualidad, no existan prácticamente reservas marinas que garanticen la protección y uso sostenible de sus recursos. Aunque existe una propuesta de todo un sistema integral de áreas protegidas, elaborado por el Proyecto PNUD/GEF CUB/92/G31 (CUB/92/G31, 1997), en la práctica solo se encuentra bajo régimen de protección: a) El área marina aledaña al Parque Nacional Caguanes, con muy bajo o ningún nivel de vigilancia y protección, b) Algunos puntos de pequeña área donde se prohibe la pesca submarina o la deportivo-recreativa, bajo la denominación de Zonas de uso y protección, establecidas por el MIP (categoría no contemplada en el Decreto-Ley de Areas Protegidas), c) Algunos sitios de cría de langosta, donde se prohibe solamente la pesca comercial de dicha especie. En la práctica, ninguna de las mencionadas áreas se ha definido con el objetivo de lograr la protección y/o uso sostenible de su biodiversidad, no existe suficiente información sobre sus valores naturales y no se ha elaborado un plan de manejo específico. El MIP, la principal institución encargada de la explotación y protección de dichas áreas, no prevé hasta ahora el establecimiento de Reservas de Pesca (Reservas de no-uso), como una vía de reconocida eficiencia para la protección de los recursos pesqueros. Se definen como Reservas de Pesca, áreas cerradas permanentemente para usos de consumo. Su objetivo es el de proteger segmentos de las poblaciones explotadas de la mortalidad por pesca, de forma que se pueda mantener, en áreas reducidas, comunidades de peces relativamente poco alteradas, así como la estructura por edades de las poblaciones. Estas áreas centrales protegen la diversidad genética intra e interespecífica y garantizan la biomasa del stock reproductor. La dispersión natural de los huevos y larvas garantizan el suministro de reclutas a las áreas de pesca adyacentes y el mantenimiento de capturas en estas, por el exceso y movimiento de los adultos. Los éxitos locales con las áreas protegidas marinas confirman que proteger de la pesca determinada área durante un tiempo suficientemente largo para rehabilitar su biomasa, conlleva a un incremento de las capturas en las áreas adyacentes, generalmente en exceso de aquellas necesarias para compensar la pérdida por la protección del área (Pauly y Christensen, 1996). Russ (1996), reconocido especialista en pesquerías tropicales platea que: “tenemos que aceptar reservas no óptimas, antes que dejar de establecerlas, pues no tenemos tiempo para esperar a que los científicos diseñen reservas perfectas”. En varios países se ha adoptado el criterio de que las Reservas de Pesca deben cubrir aproximadamente el 20% del área total de pesca (Plan Development Team, 1990; Bohnsack, 1990, Russ, 1996, y otros). Se recomienda que tales áreas cubran sectores transversales desde la costa hasta el océano abierto, de forma que estén representados todos los hábitat de la plataforma. Idealmente, estos hábitat deben estar representados de forma proporcional a como existen en toda la zona de pesca. El resto del área (80%), debe ser manejada de acuerdo con los sistemas tradicionales de regulaciones pesqueras. Si estas no existieren o fueren deficientes, se recomienda incrementar el área de las reservas a 30%. Otros criterios importantes deben ser analizados al diseñar y planificar las reservas: • Cada una debe ser lo suficientemente extensa como para incluir los hábitat necesarios para soportar el crecimiento de un stock de reproductores con una estructura de tamaños estable. Se considera que debe abarcar un mínimo de 20 millas náuticas de línea costera para garantizar los procesos biológicos de forma efectiva y facilitar su vigilancia y control. • Siempre que sea posible, las Reservas deben establecerse en zonas que incluyan áreas de desove en lugares donde las corrientes faciliten el transporte de las larvas y juveniles hacia las áreas de pesca • Deben estar ubicadas en lugares apartados de los grandes centros urbanos. • Preferentemente, deben ubicarse en lugares en los cuales se facilite el sistema de vigilancia, ya sea de forma directa (Unidades de Guarda Costas, Guardabosques) o en coordinación con otros sistemas de áreas protegidas (Parques Nacionales, Reservas Turísticas, Reservas Ecológicas, etc.). Una opción ideal resulta de la combinación de los objetivos pesqueros con otras opciones, como las actividades turísticas, tan importantes en el ASC. La existencia de un sistema de gobierno socialista establece condiciones excepcionales para el establecimiento de programas integrales de manejo de áreas marinas protegidas con múltiples objetivos. Son muchas las ventajas que ofrece esta opción de manejo. De forma resumida, se pueden mencionar las siguientes: • Protegen los stocks y hábitat de peces arrecifales de la sobrepesca y la explotación indiscriminada, al mantener protegidos parte de la población y sus hábitat • Garantizan la reposición de las poblaciones de peces y otros organismos objeto de explotación al proteger la capacidad reproductiva y mantener una reserva. • Constituyen un seguro contra las fallas del manejo, las catástrofes naturales y las alteraciones ambientales impredecibles. • Mantienen e incrementan los stocks de peces en las afueras de las reservas gracias a la dispersión desde estas de los nuevos reclutas y adultos a la población explotada. • Los peces alcanzan mayores tallas, mayor calidad y rendimientos más altos antes de salir de la reserva e incorporarse a la pesquería. • Preservan la abundancia, riqueza de especies y diversidad genética de las poblaciones de peces. • Mantienen el equilibrio natural de la comunidad de peces y la estructura por edades. • Permiten el mantenimiento de áreas naturales con alteraciones mínimas para la investigación pesquera, la educación y otros usos de no consumo. • Simplifican y reducen el costo de los sistemas de vigilancia. • Garantizan el reclutamiento en condiciones ambientales fluctuantes. • Permiten la existencia de especímenes de gran talla para la pesca deportiva (limitada en nuestro caso para el turismo internacional) y aumentan la atracción para el buceo contemplativo y otras opciones turísticas no extractivas. • Incrementan la flexibilidad de las normas de manejo y facilitan la cooperación entre diferentes agencias (Pesca, Turismo, Conservación, Educación ambiental, Investigación Científica, etc.). • Facilitan el uso no extractivo de los recursos, como el buceo contemplativo, la fotografía subacuática, y otras actividades que compensan económicamente con creces la pérdida de pesca si es que existiere en su etapa inicial. DISCUSION Los recursos marinos costeros de la zona tropical, por lo general muestran complejas, variadas y específicas relaciones con los diferentes biotopos que conforman el ecosistema nerítico. Cada especie posee adaptaciones de su ciclo biológico con el marco ambiental, por lo cual la ruptura de ese ciclo puede provocar grandes alteraciones en la estructura y funcionalidad de sus poblaciones y de la comunidad en general. Como se aprecia de los epígrafes anteriores, la diversidad biológica de la ictiofauna marina del ASC ha sufrido serias afectaciones en la última década. Ello es resultado de la acción conjunta de factores naturales y antrópicos que han provocado una notable disminución de la densidad y biomasa de las poblaciones de peces en todos los hábitat estudiados. La Tabla 15 resume los tipos de impactos a que está sometido el ecosistema y los recursos del Archipiélago Sabana-Camagüey y las consecuencias de estos. Tabla 15. Tipos de impactos sobre el medio marino en la zona costera del Archipiélago Sabana-Camagüey. Fuentes de impactos Impactos Directos Impactos Derivados Elevación de la temperatura del agua - Mortandad de corales y disminución de la complejidad estructural del substrato. - Sobrecrecimiento de algas - Disminución de los refugios para invertebrados y peces - Reducción de la productividad del fondo marino y, en consecuencia, de la abundancia de peces.- Reducción de la diversidad biológica. - Disminución del valor estético de los arrecifes, en detrimento del turismo. Interrupción del régimen de - Cambios en los procesos circulación en las macrolagunas y de sedimentación y del el represamiento. régimen hidrológico. - Elevación de la salinidad, mortandad de la fauna y flora, degradación general de la calidad del agua y el fondo marino. - Interrupción de los procesos ecológicos vitales - Reducción de las áreas aptas para la vida marina - Destrucción de las zonas de reclutamiento y cría, - Interrupción de las migraciones de peces - Reducción de la base alimentaria y el crecimiento de los organismos, - Interrupción del proceso de reproducción. - Reducción de la productividad del fondo y de la biomasa pesquera. Uso de artes de pesca nocivos. - Reducción de la disponibilidad de refugios para invertebrados y peces, - Disminución de la calidad y - Destrucción de los fondos marinos, - Aumento de la turbidez y disminución de la productividad primaria. - Captura excesiva de peces de pequeña talla, juveniles o no comerciales tallas de las capturas comerciales, - Reducción del reclutamiento Manejo no sostenible de los recursos pesqueros - Sobrepesca de especies y del ecosistema - Reducción de los recursos pesqueros - Reducción de las posibilidades del turismo ecológico - Reducción de los puestos de trabajo para personal vinculado a la pesca. Contaminación/toxicidad - Reducción del contenido de oxígeno en el agua. - Mortantades puntuales de peces y otros organismos, - Bioacumulación. - Degradación Biológica - Reducción de los recursos pesqueros - Reducción de la calidad de los productos pesqueros. Es evidente que la disminución de la cobertura de corales y como consecuencia el aumento del cubrimiento de algas en los arrecifes coralinos es resultado de la elevación de la temperatura durante los eventos ENSO ocurridos en 1995, cuando Carrodeguas et al, (1996) reportaron gran mortalidad de corales por blanqueamiento, y otros dos más intensos ocurridos en 1997 y 1998, cuando se registró una marcada anomalía térmica positiva al norte del Archipiélago según se observa en una imagen satelitaria que promedia el período Enero-Junio de 1998 (Wilkinson, 1998; Alcolado et al., 2000), y en los gráficos de temperatura superficial del mar al norte y sur de Cuba durante los años 1981 a Junio del 2000, confeccionados por S. Cerdeira (com. pers.) a partir de datos obtenidos de IGOSS (Integrated Global Ocean Service System). La disminución de la abundancia de corales escleractinios implica un grave decrecimiento de la disponibilidad de refugios tanto para los peces como para los organismos bentónicos que sirven de alimento a estos. Aún los planctívoros diurnos que se alimentan en la columna de agua, como el chromis azul (Chromis cyanea), dependen de la existencia de refugios en el arrecife para protegerse de sus depredadores u otros peligros y para descansar (Hobson, 1991). En el caso de los herbívoros, debe considerarse que la proliferación de algas no implica necesariamente una mejoría de su base alimentaria. Muchas de tales algas son disuasivas para los peces o no son consumidas por estos por otros motivos. Generalmente con el tamaño y densidad de la alfombra algal disminuye la posibilidad de ser consumidas por los herbívoros (Littler et al., 1983; Steneck y Watling, 1982; Hay, 1991). Los loros, por ejemplo, se alimentan de las pequeñas algas epífitas que crecen sobre los corales o las rocas, pero raramente consumen las algas crustosas y otras de mayor tamaño, a veces altamente calcificadas. Los barberos (Acanthuridae), pueden consumir las algas más grandes, pero también evaden aquellas que acumulan carbonatos. Varias de las especies de algas abundantes en las estaciones muestreadas (Apéndice 1) son consideradas como altamente disuasivas para los peces herbívoros (Hay, 1991). La existencia de refugios, que brindan los corales y otros organismos sésiles asociados a estos, evidentemente constituye un elemento crítico para los procesos ecológicos que ocurren en el arrecife, y su ausencia o disminución determina una reducción de la productividad biológica de todo el ecosistema, lo cual se refleja claramente en la densidad y biomasa de los peces que son los principales usuarios de tales recursos. En las macrolagunas del ASC también han ocurrido procesos naturales y/o producidos por la acción del hombre, que han provocado graves daños a los hábitat neríticos. Ya desde la década del 60 con el auge del represamiento de las aguas fluviales, se inició un proceso de salinización de algunas bahías. No obstante, dicho proceso estuvo amortiguado en parte por eventos naturales que impidieron que alcanzara niveles críticos, aunque con otras alteraciones del ecosistema. Así por ejemplo, el Huracán Flora, en 1962, trajo enormes precipitaciones y movimiento de los sedimentos. Grandes extensiones de praderas marinas quedaron sepultadas, eliminando un importante hábitat para las etapas tempranas de muchas especies de peces e invertebrados. Hasta esa época, según consenso de los pescadores más experimentados, en dichas bahías abundaban las langostas, pargos, biajaibas, jureles, tiburones, y muchas otras especies comerciales las cuales se fueron replegando hacia la porción norte, donde aún se mantenían las praderas marinas y mayor intercambio de aguas con el océano. Hasta la construcción de los pedraplenes se mantuvo la pesca comercial en la zona con aceptable productividad. En 1986 los fuertes vientos del huracán Kate provocaron la evacuación parcial de los sedimentos acumulados en la Bahía de Buenavista y poco después aparecieron de nuevo praderas marinas en las zonas menos profundas, cual facilitó el incremento las áreas de reclutamiento y cría de la biajaiba, cuya población aumentó y con ello la pesca de dicha especie tanto en la bahía de Buenavista como en San Juan de los Remedios. Pero las alteraciones más graves en dichas macrolagunas ocurrieron tras la construcción de los viaductos que atraviesan las bahías Jigüey y Los Perros y aunque en menor medida, el que une Caibarién con Cayo Santa María. La elevación de la salinidad hasta valores que sobrepasaron en 1995 los 83 partes por mil y grandes fluctuaciones posteriormente, han provocado: • Una catastrófica disminución de la diversidad de especies y del ecosistema; • condiciones desfavorables para la reproducción, reclutamiento y cría de la biajaiba, una de las principales especies comerciales en las bahías Buenavista y San Juan de los Remedios; • reducción o incluso eliminación del alimento fundamental del machuelo (que aportó hasta 1989 unas 350-400 TM anuales) con la casi total desaparición de ese recurso; • reducción o eliminación de la base alimentaria (plancton y bentos) para las etapas juveniles, tanto de las especies que habitan en las macrolagunas (pataos, mojarras, rayas, biajaiba, pargo criollo, caballerote, macabí, lisas, jureles, roncos, etc.), como de las especies que habitan fuera de la cayería (pargo criollo, biajaiba, jureles, roncos, rabirrubia, bajonaos, jureles, etc.); • que los animales que logren sobrevivir a la etapa juvenil no encuentren suficiente alimento al reducirse las poblaciones del macro y megabentos, del cual se alimentan la mayoría de las especies comerciales (rayas, pargos, langosta, etc.) con la consecuente disminución de su tasa de crecimiento y aumento de la mortalidad natural; • incremento sustancial en el gasto de energía de los peces y crustáceos que logran sobrevivir, para el proceso de osmorregulación, como resultado del aumento de la salinidad, lo que a su vez provoca reducción de la tasa de crecimiento, aumento de la edad de maduración sexual y disminución de la fecundidad, todo lo cual conduce a una reducción de la productividad biológica y pesquera del ecosistema; • limitación o cierre del acceso a las zonas de reproducción para las especies que habitan en la macrolaguna y para el regreso a las zonas de cría de las larvas y postlarvas de esos organismos (reclutamiento). De estos antecedentes se deduce la necesidad de establecer medidas para incrementar el volumen de circulación e intercambio de agua entre las macrolagunas y el océano, en toda la región de la cayería, y facilitar el flujo de aguas fluviátiles a las macrolagunas, como premisa fundamental para la conservación de sus recursos pesqueros y la diversidad de especies. Prácticamente todas las zonas pesqueras del mundo están hoy en estado de sobreexplotación o a un nivel cercano, y otras sufren afectaciones de sus ecosistemas como resultado de la fragmentación de los hábitat por la contaminación, la sedimentación, eutroficación, etc. Particularmente en las regiones tropicales esta situación se ve agudizada por la fragilidad de sus ecosistemas, fundamentalmente los arrecifes coralinos, que soportan la mayor parte de las pesquerías en esa zona, y que aportan las especies más valiosas. Al nivel mundial, más del 70% de las poblaciones de peces comerciales están en etapa de sobrepesca o cercana a ella (FAO, 1996) y las flotas comerciales han provocado “la extinción comercial” de algunas especies, y en algunos casos hasta la extinción biológica, e incluso han provocado la destrucción de importantes ecosistemas como los arrecifes coralinos y los pastos marinos. Aún en los sistemas de pesca mejor administrados (independientemente de que ninguno puede ser considerado perfecto), la extracción pesquera provoca alteraciones en la estructura y productividad de las poblaciones explotadas y daños al medio ambiente. La pesca, aún aquella que se realiza con un plan de manejo basado en regulaciones para proteger el recurso, tiende siempre a extraer en primera instancia a los individuos de mayor talla, que son generalmente los más aptos genéticamente, disminuyen paulatinamente la estructura de tallas de la población (y de la expectativa de vida de las especies), disminuyendo paralelamente su calidad como producto, altera la estructura de las comunidades y por tanto de su equilibrio ecológico, debido a la extracción no proporcional de sus componentes, provoca daños mecánicos al medio marino (las nasas y redes rompen los corales, arrancan esponjas y otros organismos que sirven de refugio o alimentación a los peces y crustáceos, etc.), provoca contaminación (por el achique de las sentinas de los barcos, las pinturas antioxidantes de los cascos de las embarcaciones, el vertimiento de residuales del procesamiento de productos pesqueros, etc.), facilita el acceso y actividad del hombre sobre otros recursos no pesqueros (extracción de organismos ornamentales, pesca ilegal de especies en peligro o protegidas, etc.) y muchas otras afectaciones al medio y los recursos marinos. Entre los problemas más comunes en las pesquerías tropicales, podemos mencionar (Bohnsack, 1990): • Potencial sobreexplotación del reclutamiento, debido a la insuficiencia de un adecuado stock de reproductores; • Incremento de la probabilidad de fallas en el reclutamiento debido a su carácter impredecible y a menor número de desoves y menor fecundidad, como resultado de ciclos de vida más cortos; • Pérdida de diversidad genética, lo que resulta en características biológicas no deseables de las poblaciones; • Sobreexplotación del crecimiento de las especies más valiosas; • Disminución de la abundancia de las poblaciones y de la talla media de sus individuos; • Pérdida de diversidad biológica; • Potencial ruptura de la estabilidad de las comunidades de peces y alteraciones permanentes de su estructura; • Selección natural más rápida en detrimento de la calidad genética, debido a la disminución de la vida de cada generación. En el caso del ASC, ha quedado demostrado que los métodos de administración pesquera adolecen de serias deficiencias (independientemente de la existencia de todo un sistema de regulaciones y vigilancia), que provocan degradación de los recursos pesqueros y del medio ambiente marino, todo lo cual se suma a las desfavorables alteraciones ambientales ocurridas en la última década como resultado de los cambios climáticos, y aquellas provocadas en el ecosistema por las construcciones para facilitar el desarrollo del turismo y por la contaminación. La eliminación del uso de métodos y artes de pesca nocivos y la limitación del esfuerzo pesquero a las posibilidades reales de producción del ecosistema resultan premisas indispensables para evitar la sobrepesca del mismo. Tales medidas deben incluir por supuesto la llamada pesca deportiva (realmente pesca de subsistencia) cuyo impacto sobre los recursos pesqueros parece ser mayor de lo que se presupone. . 8. ELEMENTOS PARA UNA ESTRATEGIA DE MANEJO. Se acepta hoy que los métodos actuales de explotación de los recursos, no son capaces de garantizar un manejo sostenible, dinámico y capaz de satisfacer las futuras demandas de alimentación, empleo y recreación, y al mismo tiempo mantener la diversidad biológica (Haughton y Jacobs, 1998). Por ello, se impone la necesidad de implementar nuevos enfoques de manejo integral, orientados a restaurar y mantener la productividad biológica natural, la biodiversidad y la integridad ecológica de todo el ecosistema costero y marino Como mínimo, un plan de manejo de los recursos marinos debe plantearse objetivos tales como: - Facilitar el uso múltiple de los hábitat y sus recursos; - Manejar la pesca de forma sostenible; - Crear empleos a largo plazo y satisfacer las necesidades materiales y espirituales de las poblaciones locales. Para lograr tales objetivos es necesario desarrollar estrategias orientadas a: • Crear una conciencia de propiedad del recurso, no del producto de la pesca, o del beneficio económico que brinda el turismo u otros usos. Si no se logra ese sentido de propiedad y responsabilidad no es posible garantizar límites en la explotación del recurso. Para ello es necesario vincular al pescador a la zona de pesca, mediante la asignación de derechos al uso del área, a través de licencias, creación de áreas de manejo locales, u otros mecanismos. Igualmente puede vincularse a las instalaciones turísticas costeras con los hábitat que utiliza. Paralelamente, las autoridades correspondientes deben garantizar la protección de esos derechos. • Manejar a la gente y los hábitat, no a los peces. Ello implica, controlar el uso de los métodos de pesca, más que captura y manejar las actividades que impactan a los hábitat. • Evitar cuellos de botella en el reclutamiento, para lo cual se requiere proteger los reproductores durante el proceso de desove, proteger el hábitat de los juveniles y de ser posible mejorar las condiciones para el reclutamiento en esos hábitat (por ej. mediante refugios artificiales, siembra de juveniles, etc.) • Maximizar las utilidades totales de los recursos que provee el hábitat, no solo el valor de la pesca. Una pesca sostenible, puede dar alimentos y beneficios económicos menores pero a largo plazo resulta más beneficiosa. Por otra parte el turismo puede dar grandes beneficios económicos si se combina adecuadamente con la extracción pesquera, siempre que no cause daños al ecosistema. Algunas formas de explotación no convencionales pueden elevar los beneficios que brindan los hábitat, como el uso de refugios artificiales para concentrar peces (ver detalles en Claro y García-Arteaga, 1990), tanto con fines pesqueros como turísticos, y la explotación controlada de peces ornamentales. • Crear mecanismos para lograr la participación pública en los programas de manejo y conservación, vinculando a las poblaciones locales. Para ello resulta indispensable crear una conciencia ambientalista tanto en los tomadores de decisiones al nivel local como de los usuarios de los hábitat y sus comunidades. Las Areas Protegidas Marinas, constituyen hoy la opción más aceptada y de mayor potencial para el manejo de una pesquería sostenible y disminuir los riesgos de sobreexplotación y otros daños a los recursos pesqueros, al tiempo que ofrece la posibilidad de otros usos altamente productivos, como el desarrollo del turismo ecológico, la educación ambiental, la investigación científica, etc. (Plan Development Team, 1990; Bohnsack, 1990; Pauly y Christensen, 1996; Russ, 1996). Pueden ser un mecanismo adecuado para implementar la estrategia descrita y en general para el manejo integrado de zonas costeras seleccionadas. CONCLUSIONES 1- La ictiofauna y en general las comunidades marinas de la zona costera del Archipiélago SabanaCamagüey han sufrido en la década del 90 graves afectaciones de su diversidad, disminución de sus poblaciones y reducción de los recursos pesqueros y estéticos, como resultado de la acción conjunta de factores ambientales naturales, provocados por cambios climáticos globales (principalmente elevación de la temperatura) y por factores antrópicos que provocaron drásticas alteraciones de las condiciones naturales. 2- La diversidad de especies, así como la densidad y biomasa de peces en los arrecifes coralinos del Archipiélago Sabana-Camagüey en la década del 90 sufrió graves reducciones, como resultado de la disminución de la cubierta coralina y del incremento de la cobertura algal provocada por mortandades masivas de corales, a causa de la elevación de la temperatura durante varios eventos ENSO, reduciendo la disponibilidad de refugio y alimento para los peces y otros organismos dependientes de tales condiciones. 3- La diversidad de especies y del ecosistema en las macrolagunas, así como la abundancia de peces y otros recursos pesqueros sufrió graves daños como resultado del incremento de la salinidad, provocada por el represamiento y la construcción de viaductos que interrumpieron el régimen de circulación de esos cuerpos de agua, provocando la degradación de los fondos y de las condiciones vitales para los procesos ecológicos (reclutamiento, cría, alimentación, crecimiento). 4- La alta turbidez del agua en las macrolagunas, y la reducción de su productividad biológica, parecen ser las causas que han provocado una seria disminución de la densidad y biomasa de peces en los manglares de los cayos. En las bahías más salinizadas, este factor afecta con mayor fuerza a la comunidad de peces del manglar, lo cual se refleja en especial sobre la abundancia de peces juveniles. 5- Las capturas en el Archipiélago Sabana-Camagüey ya desde la década del 70 alcanzaron el máximo nivel de explotación sostenible por dicho ecosistema. Los incrementos de captura obtenidos durante la década del 80 se sostenían principalmente por la explotación de especies de alta productividad pero de muy poco valor, como el machuelo, o de poblaciones biológicamente poco productivas, pero de gran talla y largo ciclo de vida, como los batoideos, las cuales se encuentran ya en fase de declinación. 6- Más de la mitad de las capturas de la zona NE procedían de las macrolagunas, o de las praderas marinas vinculadas a estas. Tales biotopos han sido seriamente degradadas por factores antrópicos, por lo que es preciso buscar alternativas para la sustitución de la pesca en dichas aguas interiores y emprender obras de mejoramiento y manejo ambiental para su recuperación. 7- La interrupción del intercambio de aguas entre el océano y las macrolagunas, además de causar serias alteraciones del medio marino, provoca la ruptura del ciclo vital de algunas especies comerciales, lo cual evidencia ser una de las causas de la actual decadencia de los recursos pesqueros. 8- El uso de métodos y artes de pesca nocivos (chinchorros, mallas de pequeños orificios, destrucción de corales, etc.) incide directamente en la productividad biológica y la economía del ecosistema, y es una de las causas directas de la degradación del medio y los recursos pesqueros. 9- La ausencia de Areas Protegidas Marinas adecuadamente diseñadas y manejadas en el Archipiélago constituye un factor limitante para la protección y uso sostenible del ecosistema y sus recursos pesqueros, el desarrollo del turismo y otras opciones, por lo que deben ser implementadas lo antes posible. RECOMENDACIONES 1. Elaborar una estrategia pesquera para enfrentar las consecuencias del deterioro del medio marino del ASC, como consecuencia de las actividades antrópicas y de las alteraciones desfavorables provocadas por cambios climáticos o naturales, a fin de desarrollar alternativas para recuperar o sustituir la pesca en aguas interiores por otras vías de producción (maricultivo, pesca del alto, selección de especies y tallas de valor, etc.). 2. Implementar lo antes posible el establecimiento de Areas Protegidas Marinas considerando todas las opciones de uso y protección para la conservación de la diversidad biológica. Integrar los intereses de uso turístico, pesca, investigación y educación ambiental en el diseño y planes de manejo de dichas áreas. Considerar los requisitos para las reservas de pesca, entre otros, la protección de los stocks de reproductores, las zonas de concentración para el desove, el reclutamiento y las áreas de cría. La propuesta elaborada al respecto por el Proyecto GEF/PNUD Sabana-Camagüey (CUB/92/G31, 1997) debe constituir un punto de partida para la implementación de las mismas. 3. Incrementar el número y superficie de los pasos de agua a través de todos los pedraplenes y entre los cayos que separan las macrolagunas del océano, para aumentar el intercambio de aguas y facilitar las migraciones de peces y otros procesos vitales. Evitar la construcción de nuevos viaductos que interrumpan el régimen de circulación natural. 4. Revisar y profundizar el sistema de las regulaciones para la protección ambiental marina en la región, de forma que cubra todo el espectro de actividades dañinas al medio ambiente.y exigir su cumplimiento. Sancionar el vertimiento al mar de hidrocarburos y todo tipo de sustancias tóxicas, basura, materia orgánica u otros contaminantes que puedan incrementar las concentraciones de nutrientes u otros elementos dañinos para la vida marina. 5. Disminuir paulatinamente el uso de los chinchorros hasta su sustitución total. Iniciar el proceso, de forma urgente, con la reducción de los mismos a un máximo de 500 brazas de largo, con los tamaños de malla que establecen las regulaciones pesqueras vigentes, y la adición de mecanismos de selección, preferentemente la corona selectora en el copo y como segundo paso, el vivero selector. Prohibir terminantemente la tenencia y uso de mandarrias y otros instrumentos utilizables para la destrucción de corales y otros refugios del fondo marino. Exigir el cumplimiento de las regulaciones sobre los tamaños de mallas en las nasas, o al menos en parte de las mismas. 6. Mantener un sistema de monitoreo permanente sobre el estado de las poblaciones de peces y del ecosistema en general, en las regiones afectadas por la construcción de viaductos en el mar (macrolagunas, arrecifes coralinos y manglares). Se recomienda el uso de las metodologías descritas en el Anexo 2 de este informe. 7. Establecer un control estadístico y un monitoreo permanente sobre las poblaciones de manjúa, especialmente en la región de Caibarién, que permitan acopiar información para el manejo y explotación racional de esta especie como base para el desarrollo de las pesquerías de bonito y albacora. 8. Disminuir el esfuerzo pesquero, sobre las especies en decadencia, particularmente los batoideos, y de aquellas con perspectivas para la pesca deportiva para el turismo internacional (macabí, sábalo, peces de pico). Incluir en el sistema de Areas Protegidas, zonas seleccionadas para esta actividad. 9. Establecer una regionalización de las zonas de pesca por Empresas y de ser posible por embarcación o colectivo de pescadores (según las condiciones locales), a fin de promover el interés por la conservación de los recursos pesqueros de cada región, mediante la concesión oficial de licencias u otro mecanismo legalmente establecido. 10. Establecer un sistema de control estadístico sobre la extracción pesquera que se realiza por la llamada "pesca deportiva", que contemple la magnitud de la captura, su composición por especies, áreas de captura (previamente definidas) y artes de pesca. Elaborar una estrategia para la disminución del esfuerzo pesquero de esa actividad. 11. Desarrollar programas de educación ambiental dirigidos especialmente a las personas y comunidades vinculadas al uso y conservación de los hábitat marinos y promover su participación en los programas de protección y manejo. Establecer un código ético de pesquería responsable. REFERENCIAS Alcalá, A. 1988. Effects of Marine Reserves on Coral Fish Abundance and Yields of Philippine coral reefs. AMBIO 17(3):194-199. Alcolado, P. M., R. Claro, B. Martínez-Daranas, G. Menéndez-Macías, P. García-Parrado, K. Cantelar-Ramos, J. Espinosa Sáez, R. del Valle, J. C. Martínez Iglesias y T. Neff. 2000. Estado general de los arrecifes coralinos de Cuba y propuestas de manejo ambiental. Informe Final de Proyecto, Instituto de Oceanología, CITMA. Alcolado, P.M., C. Jiménez, J. Espinosa, D. Ibarzábal, J.C. Martínez, R. del Valle, N. Martínez, A. Hernández, M. Abreu, L. Vega and E. Ramírez. 1990a. Aspectos ecológicos del acuatorio del noreste de la provincia de Villa Clara. En Estudios de los grupos insulares y zonas litorales del Archipiélago cubano con fines turísticos Cayos Francés, Cobo, Las Brujas, Ensenachos y Santa María. (J. Pérez, ed.). Ed. Cient. Téc. La Habana: 86-97. Alcolado, P.M., C. Jiménez, J. Espinosa, D. Ibarzábal, J.C. Martínez, R. del Valle, N. Martínez, A. Hernández, M. Abreu, L. Vega y E. Ramírez 1990b. Ecología Marina. En Estudios de los grupos insulares y zonas litorales del Archipiélago cubano con fines turísticos. Cayos Mégano grande, Cruz, Romano y Guajaba. (L. Fernández, R. Cañizares and H. Gómez, eds.). Ed. Cient. Téc. La Habana: 99-109. Alcolado, P.M. y L. Menéndez (en prensa): Mortalidad masiva de manglares: un caso en el norte de Cuba. Baisre, J. 1985. Los recursos pesqueros marinos de Cuba: fundamentos ecológicos y estrategias para su utilización. [resumen], Tesis de candidatura (Ph. D.) en Ciencias Biológicas, 17 pp., 9 ilus., 1 tabla. Boesch, D.F. (1977): Application of numerical classifications in ecological investigations of water pollution. Ecological Res. Ser., EPA-GOO/3-77-033, 115 pp. Baisre, J. 1993. Marine fishery resources of the Antilles: Lesser Antilles, Puerto Rico and Hispaniola, Jamaica, Cuba, FAO Fisheries Technical Paper, No. 326, Part 4, Cuba, Rome, pp. 181-235. Barlow, G.W. l981. Patterns of parental investment dispersal and size among coral-reef fishes. Environ. Biol. Fish., 6(1): 65-86. Bohnsack, J. A. 1994. How marine fishery reserves can improve reef fisheries. Proc. Gulf Caribb. Fish. Inst. 43: 217-241. Cabrera,V. y M. H. Obregón. 1986. Análisis preliminar de las pesquerías de escama en la costa norte de Camagüey. Archivo del Centro de Investigaciones Pesqueras, Ministerio Industria Pesquera, 12 pp. Carrodeguas, C. G. Arencibia, N. Capetillo y M. García. 1996, Decoloración de corales en el Archipiélago cubano. Rev. GeoCuba. Geomática y Ciencias Afines, 1-5 pp. Claro, R. En prensa. The Marine Ichthyofauna of Cuba, In R. Claro, K. Lindeman and L. Parenty eds., Ecology of The Marine Fishes of Cuba, Smithsonian Institution Press. Claro, R., J.A. Baisre y J.P. García-Arteaga 1994. VIII. Evolución y Manejo de los Recursos Pesqueros. 435-492 pp. En: Ecología de los Peces Marinos de Cuba (R. Claro Ed.), Instituto de Oceanología y Centro de Investigaciones de Quintana Roo, México. Claro, R., J.P. García-Arteaga, E. Valdés-Muñoz y L. M. Sierra. 1990. Alteraciones de las comunidades de peces en el Golfo de Batabanó, en relación con la explotación pesquera. En Asociaciones de peces en el Golfo de Batabanó (R. Claro, ed.), Editorial Academia, La Habana, pp. 50-66. Claro, R., y J.P. García-Arteaga (1993): Estructura de las comunidades de peces asociados a los manglares del grupo insular Sabana-Camagüey, Cuba. Revista de Oceanología y Ecología Tropical Avicennia, 0:60-82. Claro, R., y J.P. García-Arteaga (1991): Perspectivas para un programa de arrecifes artificiales para peces en la plataforma cubana. Editorial Academia, La Habana, 50 pp. Claro, R., y J.P. García-Arteaga (1994): Estructura de las comunidades de peces en los arrecifes del grupo insular Sabana-Camaguey, Cuba. Revista de Oceanología y Ecologia Tropical Avicennia, 2:83-107. Claro, R., J. P. García-Arteaga, y Bouchon-Navarro, M. Louis y C. Bouchon. 1998. La estructura de las comunidades de peces en los arrecifes de las Antillas Menores y Cuba. Revista de Oceanología y Ecologia Tropical Avicennia 8/9:69-86. Claro, R., A. García-Cagide, L.M. Sierra y J.P. García-Arteaga. 1990. Características biológico-pesqueras de la cherna criolla Epinephelus striatus (Bloch) (Pisces:Serranidae) en la plataforma cubana. Cien. Biol., 23:23-42. CUB/92/G31. 1997. Protección de la biodiversidad y establecimiento de un desarrollo sostenible en el Ecosistema Sabana-Camagüey. Síntesis de los estudios y estrategia. Informe final de Proyecto. Rept. GEF/UNDP/CUB/92/631. 257 pp. Davis, G. 1989. Designated Harvest Refugia: The Next Stage of Marine Fishery Management in California. CALCOFL Rep., 30:53-58. FAO, 1994. Review of the state of the world marine fishery resources. FAO Fish. Tech. Pap. 335, 136 p. García-Coll, I. 1984. Determinación de la edad y el crecimiento del bonito, Katsuwonus pelamis y la albacora Thunnus atlanticus en las regiones suroccidental y nororiental de Cuba y evaluación de las pesquerías de estas especies en La Coloma [inédito]. Tesis de candidatura (Ph. D.), Univ. Habana, 104 pp. García Rodríguez, J. 1978. Resultados de la pesca exploratoria del alto en la región central de la costa N de la plataforma cubana. (mecanografiado), 1er. Foro Científico Centro Investigaciones Pesqueras, La Habana, 8 pp, 4 tab. 14 fig. García-Rubies, A y M. Zabala. 1990. Effects of total fishing prohibition on the Rocky Fish Assemblages of Medes Islands Marine Reserve (NW Mediterranean). Fish Structure in Marine Reserves. Scientia Marina 54(4):317-328. Gimenez, E. 1988. La pesca con chinchorro de arrastre en la plataforma nororiental de Cuba (Zona D). Centro de Investigaciones Pesqueras, MIP, 15 pp. Gimenez, E. 1990. Análisis de las pesquerías de escama de la Empresa C.P.I. de Villa Clara. Centro de Investigaciones Pesqueras, MIP, 14 p, 10 tablas, 23 Figs. Gimenez, E., M. C. Sotolongo, A. Hondares y B. Pedroso. 1992. Ordenamiento Pesquero del recurso escama de la plataforma Nororiental de Cuba. Centro de Investigaciones Pesqueras, MIP, 29 p, 15 figs, 9 tablas. Gobert, B. 1990. Production relative des pêcheries côtières en Martinique. Centre ORSTOM de Fort -de-France Cedex (France), 3:181-191. Hay, M. E. 1991. Fish.Seaweed Interaction on Coral Reefs: Effects of Herbivorous Fishes and Adaptations of their Prey. In P.F. Sale ed. The Ecology of Fishes on Coral Reefs. Academic Press, Inc., pp. 86-119. Haughton, M. y N. Jacobs, 1998. The relationship between integrated fisheries management and marine biodiversity: Can the sustainable management of fisheries and marine biodiversity be mutually beneficial. Caribbean Workshop on Marine Biodiversity, Montego Bay, Jamaica, October 1998, 14 p. Hobson, E. S. 1991. Trophic relationships of fishes specialized to feed zooplankters above coral reefs. In P.F. Sale ed. The Ecology of Fishes on Coral Reefs. Academic Press, Inc., pp. 69-95. Johannes, R.E. (1978): Reproductive strategies of coastal marine fishes in the tropics. Env. Biol. Fish, 3(1)54-84. Kaufman, L.S. J.P. Ebersole (1984): Microtopography and the organization of two assemblages of coral reefs fishes in the West Indies. J. Exp. Mar. Biol. & Ecol., 78:253-268. Littler, M. M., P. R. Taylor and D.S. Littler. 1983. Algal resistance to herbivory on a Caribbean barrier reef. Coral Reefs 2:111-118. Lluis-Riera, M. (1984): Estudios hidrológicos de la plataforma nororiental de Cuba (zona D). Acad. Cien. Cuba, Inst. Oceanol.,44 pp. Man, A., R. Law y N.V.C. Polunin. 1994. Role of Marine Reserves in Recruitment to Reef Fisheries: A Metapopulation Model. Elsevier, :197-204. Martínez Canals, M., R. Pérez Díaz y A. Rodríguez Vargas. 1996. Caracterización metálica de los sedimentos superficiales y el nivel de contaminación por metales pesados e hidrocarburos del petróleo en el archipiélago Sabana-Camagüey. Archivo Instituto de Oceanología, 24 p. Mason, C. F. y R. J. Bryant 1974. The structure and diversity of the animal communities in a broadland reedswamp. J. Zool. , 172:289-302. Munro, J. L. 1983. Coral reef fish and fisheries of the Caribbean Sea. En Caribbean Coral Reef Fishery Resources (J.L. Munro, ed.), ICLARM Studies Review, 7:1-9. Obregón, M.H., E. Pozo y S. Valle. 1990. Las pesquerías de biajaiba (Lutjanus synagris) en la plataforma nororiental de Cuba. Resúmenes, II Congreso de Ciencias del Mar, 18-22 Junio, La Habana, Cuba, 151 p. Pauly, D., and V. Christensen. 1996. Rehabilitating Fished Ecosystems: Insights from the past. NAGA, The ICLARM Quarterly, July, 1996:13-14. Plan Development Team, 1990. The potential of marine fishery reserves for reef fish management in the U.S. Southern Atlantic. NOAA Technical Memorandum NMFS-SEFC-261, 40 p. Proyect Reefkeeper. 1992. The case for Marine Fishery Reserves in Fisheries Management. A conservation Action Guide based on a review of 21 scientific papers. Vol. 1, Executive Digest, 60 p. Rodas Fernández, L. y M. Rallo Garzón. 1998. Caracterización hidrodinámica de la bahía Jigüey. Archivo Instituto de Oceanología. Rogers, C. S., G. Garrison, R. Grober, Z. M. Hillis, M. A. Franke, eds. 1994. Coral reef monitoring manual for the Caribbean and Western Atlantic. Virgin Islands National Park, U.S: National Biological Survey, Russ, G. 1985. Effects of Protective Management on Coral Reef Fishes in the Central Philippines. Proc. Fifth Intern. Coral Reef Conference, Tahiti, Vol 4. Russ, R. G. 1996. Fisheries Management: What Chance on Coral Reefs?, Naga, July, 1996: 5-9. Russ, G.R., and A.C. Alcala. 1996. Do marine reserves export adult fish biomass? Evidence from Apo Island, central Philippines. Mar. Ecol. (Progr. Ser.) 132:1-9. Russ, G.R., and A.C. Alcalá. in press. Marine reserves rates and patterns of recovery and decline in abundance of large predatory fish. Ecol. Appl. Sadovy, Y. 1994. Grouper Stocks of the Western Central Atlantic: The Need for management needs. Proceeding of the 43rd Gulf and Caribbean Fisheries Institute:43-64. Shorthouse, B. 1990. The Great Barrier Reef Marine Park. How Does it Work for Fishermen?. Australian Fisheries 49(12):16-17. Smith, C.L. 1978. Coral reef fish community: a compromise view. Environ. Biol. Fish., 3(1):109-128. Sparre, P. y S.C. Venema. 1995. Introducción a la evaluación de los recursos pesqueros tropicales. Parte I - Manual. FAO, Documento Técnico de Pesca 306/1, Rev. 1, 419 p. Steneck, R. S. y L. Watling. 1982. Feeding capabilities and limitations of hervbivorous mollusks: a functional group approach. Mar. Biol. 68:299-319. White, A. T. 1986. Marine Reserves: How Effective as Management Strategies for Philippine, Indonesian and Malaysian Coral Reef Environments. Ocean Management, 10:137-159. Wilkinson, C. 1998. Status of coral reefs of the world. Global Coral Reef Monitoring Network. Australian Institute of Marine Science. 184 p.