REVISION DE GENETICA MENDELIANA Se presenta un repaso de
Anuncio

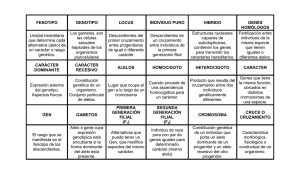
REVISION DE GENETICA MENDELIANA Se presenta un repaso de la base de la genética clásica, y se introducen los términos propios de la disciplina, para un comprensible mejoramiento animal a nivel del gendesde abajo hacia arriba. GENES, CROMOSOMAS Y GENOTIPOS La unidad básica de herencia es denominada gen. Los genes son segmentos de ácido desoxirribonucleico o ADN, la molécula compleja que forma el código genético para todos los seres vivos. Gen: La unidad física básica de herencia que consiste en una secuencia de ADN en una locación específica en un cromosoma. Los genes son secciones relativamente pequeñas de cromosomas y estos son largos hilos de ADN y proteínas asociadas presentes en el núcleo de cada célula del organismo. ADN: Acido desoxirribonucleico, molécula que conforma el código genético Los cromosomas se presentan de a pares, un cromosoma del par heredado del padre y otro heredado de la madre. El número de pares de cromosomas depende de la especie. Los humanos por ejemplo tienen 23 pares. Cromosoma: Uno de muchos hilos de ADN y proteínas asociadas presentes en el núcleo de cada célula. Homólogo: Un cromosoma del par correspondiente que tiene un loci. Los bovinos 30, los perros 39. Un par representativo de cromosomas homólogos es representado en la figura 3.1. En la figura 3.1 se muestran dos locus hipotéticos: “J” y “B”. Locus es una palabra en latín y denota el sitio que ocupa un gen en particular en el cromosoma. En cada locus hay un par de genes, un gen en el cromosoma paterno y un gen en el cromosoma materno. Los genes en un locus se expresa simbólicamente con una sola letra o combinación de ellas. Por ejemplo, los dos genes en el locus J en un individuo pueden ser designados J y j. Si un gen es representado con una letra mayúscula y el segundo con letra minúscula (u otra variante), implica que hay una diferencia química y funcional entre ellos. J y j son llamados alelos, formas alternativas de un gen encontradas en el locus J. Si los dos genes en el locus J fueran funcionalmente parecidos, ambos tendrían el mismo símbolo. Locus: La localización específica de un gen en un cromosoma. Alelo: Forma alternativa de un gen Aunque que hay solo dos genes en un locus particular en un individuo, los dos podrían ser un subconjunto, de una serie más grande de formas alternativas del gen. En otras palabras, puede haber alelos múltiples. Alelos múltiples: más de dos alelos posibles en un locus. En los perros por ejemplo, hay un locus que afecta el color del manto conocido como locus E (por extensión de la pigmentación). Puede haber tres alelos diferentes en el locus: E que causa la extensión completa del pigmento, (es decir no inhibe la pigmentación), Ebr que es atigrado), y e, el cual inhibe la pigmentación. Cualquier perro puede tener un máximo de dos de los tres alelos en la serie E. En el ejemplo hipotético mostrado en la figura 3.1, si hubiese cuatro alelos posibles en el locus B, podrían ser presentados como B, b, b’ y b’’. La combinación de genes en un locus en particular se denomina genotipo, especialmente un genotipo de un locus. Si J y j son los únicos alelos posibles en el locus J, entonces pueden haber tres genotipos: JJ, Jj y jj. Debido a que hay cuatro alelos posibles en el locus B, hay potencial para muchos más genotipos: BB, Bb, Bb’, Bb’’, bb, bb’,bb’’, b’b’, b’b’’ y b’’b’’. Si consideramos ambos loci (plural de locus) B y J juntos, entonces hay un número más grande de genotipos de dos locus: JJBB, JJBb, JJBb’,…jjb’’b’’ 30 genotipos de dos locus en total. Genotipo: La combinación de genes en un solo locus o en un número de loci. Hablamos de genotipos de un-locus, genotipos de dos-locus, y así sucesivamente. Un genotipo de un locus es considerado homocigoto si ambos genes en ese locus son funcionalmente iguales. Los genotipos JJ, jj, BB, bb, b’b’ y b’’b’’ son ejemplos de homocigotas. Genotipos de un locus que contengan genes funcionalmente diferentes son considerados heterocigotos. Los genotipos Jj,Bb,Bb’,Bb’’,bb’,bb’’ y b’b’’ son ejemplos de heterocigotos. Homocigoto: Genotipo de un locus que contiene genes funcionalmente idénticos. Heterocigoto: Genotipo de un locus que contiene genes funcionalmente diferentes. CÉLULAS GERMINALES Y SU FORMACIÓN La primera ley de Mendel es conocida como ley de segregación. Manifiesta que en la formación de las células germinales o gametos (en el macho, espermatozoide; y en la hembra, óvulo), los dos genes en un locus de la célula madre se separan y solo un gen es incorporado en cada célula germinal. Segregación: Separación de genes ligados durante la formación de células germinales Células germinales o gametos: Célula sexual – espermatozoide u óvulo. . Hoy llamamos al proceso que crea las células germinales meiosis. La meiosis es bastante complicada, involucra un número de pasos intrincados durante el cual no solo los genes sino cromosomas homólogos enteros se separan (mire la figura 3.2). Los gametos contienen sólo un gen del par. Meiosis: Proceso de formación de células germinales Los gametos que pueden ser obtenidos de varios genotipos de dos locus, se muestran en la figura 3.3. Note que cada gameto contiene solo un gen de cada locus. Mientras que el genotipo de dos locus originales contenía cuatro genes juntos, cada gameto contiene solo dos. Como regla, las células germinales contienen la mitad del número de cromosomas y por lo tanto poseen la mitad del número de genes de las células somáticas. El número de gametos que puede ser obtenido a partir del genotipo de los padres depende de cuan heterocigoto es el genotipo. Por ejemplo, el genotipo JJBB, es completamente homocigoto. Puede producir un solo tipo de gameto: JB. A partir del genotipo parcialmente heterocigoto JJBb pueden obtenerse dos tipos de gametos, y el genotipo completamente heterocigoto JjBb, puede producir cuatro tipos de gametos. La figura 3.3 ilustra la segunda ley de Mendel, la ley de distribución independiente. Distribución independiente: Segregación independiente de genes en loci diferentes Los genes se distribuyen independientemente durante la meiosis si todos los gametos posibles se forman en proporciones iguales. Para que esto ocurra, un gen dado de un determinado locus tiene que tener una probabilidad igual de estar presente en la misma célula germinal con cualquiera de los dos genes de otro locus. Como ejemplo, considere el genotipo JjBb en la figura 3.3. Los individuos JjBb, pueden producir cuatro gametos posibles: JB, Jb, jB, y jb. Si los cuatro gametos se producen en proporciones iguales (permitiendo alguna libertad de elección para la eventual variación), entonces estos genes se han distribuido independientemente. Sin embargo, si solo se producen los gametos JB y jb o si se producen a mayores frecuencias que los gametos Jb y jB, entonces la ley de la distribución independiente ha sido violada. En este caso, parecerá que el alelo J está “pegado” con el alelo B y el alelo j está análogamente pegado con el alelo b. Los loci que afectaban los caracteres que Mendel observaba en sus plantas de poroto todos se estaban en diferentes cromosomas. Los cromosomas se distribuyen independientemente (es decir, no hay tendencia en ciertos cromosomas de pegarse en la formación de células germinales), entonces los genes en esos cromosomas, también se distribuyen independientemente. Hoy en día sabemos que hay excepciones a la ley, pero son excepciones, no la regla. Las excepciones a la segunda ley de Mendel son causadas por el Ligamiento. Dos loci están ligados si están en el mismo cromosoma. Ligamiento: disposición de dos o más loci de interés en el mismo cromosoma. Debido a que los cromosomas homólogos enteros – no solamente los genes- se separan durante la meiosis, los genes en el mismo cromosoma tienden a terminar en el mismo gameto. Sin embargo, ésta es solamente una tendencia debido a un fenómeno llamado entrecruzamiento. El entrecruzamiento involucra un intercambio recíproco de segmentos de cromosomas entre homólogos y se produce durante la meiosis, previo al momento en que los cromosomas son separados para formar los gametos. Entrecruzamiento (crossing over): Intercambio recíproco de segmentos de cromosomas entre homólogos. El entrecruzamiento ocurre durante la meiosis previamente al momento en que los cromosomas homólogos son separados para formar gametos. La figura 3.4 representa cromosomas homólogos (a) antes del entrecruzamiento y (b) luego del entrecruzamiento. Los cromosomas en la figura 3.4 (a) tienen un patrón de fondo diferente (rayado y liso) para mostrar su origen en distintos progenitores. Note que antes del entrecruzamiento, los alelos J y B están ligados, como también los alelos j y b. En el proceso de entrecruzamiento se producen “roturas o rompimientos” mutuos en sitios idénticos en cada cromosoma, y los fragmentos son intercambiados. Debido a que el rompimiento o “rotura” en la figura 3.4 se produce entre los loci J y B, los genes en estos loci se recombinan y ahora están ligados con una nueva disposición. Un solo evento de entrecruzamiento es mostrado en la figura 3.4.Eventos de entrecruzamientos múltiple son comunes, y la probabilidad de recombinación de genes en cualquier a de dos loci ligados depende de la distancia entre los loci. Los loci que están alejados (como los loci J y B) es probable que se recombinen a menudo. Recombinación: Formación de una nueva combinación de genes en un cromosoma como resultado del entrecruzamiento. A fin práctico, los genes en estos loci se distribuirán independientemente, igual que lo harían si estuviesen juntos en diferentes cromosomas. La recombinación es menos probable para loci que están muy cercanos, porque la probabilidad de que ocurra una ruptura entre ellos es mucho menor. Estos loci estrechamente ligados, crean excepciones a la segunda ley de Mendel. Pero en las especies de mayor interés productivo, los genes están distribuidos en un gran número de cromosomas, y una vinculación estrecha entre dos loci de interés es relativamente rara. En general, podemos asumir que la distribución independiente y en los ejemplos utilizados en el recordatorio de este libro, así lo haremos. FORMACIÓN DEL EMBRIÓN Cuando un macho es satisfactoriamente apareado con una hembra, se unen el espermatozoide y el óvulo y se forma un embrión. En la jerga genética, decimos que los gametos del padre y la madre se combinan para formar un cigoto. Embrión: Organismo en los primeros estadios de desarrollo en una cáscara (aves) o útero (mamíferos) Cigoto: Célula formada de la unión de gametos masculinos y femeninos. Un cigoto tiene un juego completo de genes- la mitad del espermatozoide y la mitad del óvulo. Los cigotos son hijos. Tienen el número normal de genes y cromosomas, la mitad del padre y la mitad de la madre. El proceso que determina qué o cual óvulo madurará (se desarrolla fisiológicamente y hará dehiscencia ) y que espermatozoide tiene éxito en la fertilización del óvulo se denomina selección de gametos. Selección de gametos: Proceso que determina que óvulo madura y que espermatozoide es exitoso en la fertilización del óvulo. Algunos gametos contienen defectos genéticos que los hacen no viables. Estos gametos son naturalmente seleccionados en contra. Sin embargo, a pesar de esta forma de selección natural, la selección de gametos es esencialmente al azar. En otras palabras, casi todos los gametos tienen las mismas chances de producir un cigoto. Un recurso comúnmente utilizado para determinar los cigotos posibles de obtener del apareamiento de dos genotipos paternos cualquiera es el cuadrado de Punnett. El cuadrado de Punnett es una cuadrícula de dos dimensiones. A lo largo de la parte superior de la cuadrícula están listados los posibles gametos de un padre, a lo largo del lado izquierdo de la cuadrícula están listados los gametos posibles del otro padre. Dentro de las celdas de la cuadrícula se encuentran los posibles gametos del apareamiento. Son obtenidos por una simple combinación de los gametos de cada fila y columna del cuadrado. Un ejemplo de dos locus se muestra en la figura 3.5. En este ejemplo, un macho JjBb es apareado con una hembra JjBb. Cada padre puede producir cuatro gametos diferentes: JB, Jb, jB y jb, entonces hay cuatro filas y cuatro columnas en el cuadrado de Punnett, resultando en 16 celdas. Sin embargo, no todas las celdas contienen un cigoto único. Algunas celdas del cuadrado contienen el mismo genotipo. En este caso particular, hay nueve tipos distintos de cigotos. Si cada gameto listado a lo largo de la parte superior y al costado del cuadrado de Punnett se produce con la misma frecuencia, cada celda dentro del cuadrado se debería producir con la misma frecuencia. Entonces, es posible determinar la probabilidad de cualquier genotipo en particular en la progenie, observando la frecuencia de las celdas que contienen ese genotipo. Y si se sabe que fenotipo está asociado con cada genotipo – como es el caso de los caracteres de herencia simple no poligénicos – también se puede determinar las proporciones de los fenotipos esperadas en la progenie. Cuadrado de Punnett: cuadrícula de dos dimensiones usada para determinar los posibles cigotos a obtener de un apareamiento. El color del manto en ganado Shorthorn provee un buen ejemplo. El Shorthorn tiene tres colores básicos posibles: rojo, blanco y rosillo (combinación de pelo rojo y blanco). Estos colores son controlados por el locus R. Los individuos RR son rojos y los individuos rr son blancos y los Rr rosillos. El apareamiento de dos animales rosillos es ilustrado en el cuadrado de Punnett en la figura 3.6. Como está indicado en la frecuencia de las celdas que contienen cada genotipo, los tres genotipos y fenotipos de los hijos se deben producir en una proporción 1:2:1 – un rojo, dos rosillos, un blanco. Esta relación es una expectativa, no podemos decir que cada cuatro terneros de provenientes de apareamientos rosillos, uno será rojo, dos rosillos y uno blanco. Sin embargo, en promedio la proporción se producirá, y con un gran número de hijos de este apareamiento podremos anticipar los colores de los mantos para que se ajuste estrechamente a la proporción 1:2:1 La figura 3.6 es un ejemplo del cuadrado de Punnett de un locus. La figura 3.5 es un ejemplo de dos locus. El cuadrado de Punnett puede ser utilizado para ilustrar apareamientos que incluyan cualquier cantidad de loci. Pero con objetivos prácticos, los cuadrados que contienen más de unos pocos loci se vuelven “inmanejables”. Un cuadrado de Punnett que muestre el apareamiento de individuos heterocigotos en cuatro loci, tendría 256 celdas. LA ALEATORIEDAD DE LA HERENCIA Lo significativo de las leyes de Mendel recae en su explicación de la especial naturaleza de la herencia, las antiguas “partículas” son lo que ahora llamamos genes, y por ende la explicación de cómo se mantiene la variabilidad genética en la población. Previo a los descubrimientos de Mendel, la escuela del pensamiento genético más aceptada incluía una teoría de la herencia de la “mezcla”, en la cual la información hereditaria estaba contenida en fluidos, probablemente, incluso en la sangre, y era la mezcla de fluidos parentales la cual determinaba la estructura genética de un hijo. La teoría de la mezcla fue fatalmente incorrecta. No pudo explicar nunca porqué, luego de muchas generaciones de mezclar fluidos, los individuos de una población eran una mezcla similar. En otras palabras, no podía explicar porque hay tanta variación genética en la mayoría de las poblaciones y porque la variación no disminuye a través del tiempo. El trabajo de Mendel proveyó la respuesta y refutó la teoría de mezcla para siempre, aunque a raíz de eso hoy en día todavía usamos términos como porcentaje de sangre para describir el linaje de un animal. Para tener una mejor idea del efecto de la herencia mendeliana en la preservación de la variabilidad genética, considere un individuo que es heterocigoto en un loci de 100. Asumiendo la segregación y la distribución independiente, este individuo puede producir más de 1.2 x 1030 gametos únicos y diferentes. Y si estos individuos fueran apareados con otro individuo como él más de 5 x 10 47 cigotos únicos podrían ser posibles. Eso es 500 billones cigotos – no hay dos iguales. Estos números son tan grandes que son incomprensibles, y todavía subestiman los verdaderos números posibles. La mayoría de los animales domésticos son heterocigotos en mucho más de 100 loci. Un número mucho más realista de loci heterocigotos puede estar alrededor de los miles o decenas de miles. Los números resultantes para gametos posibles y cigotos son asombrosos. Calculando la cantidad de posibles gametos y cigotos Los siguientes Ejemplos muestran cómo usar un número estimativo de loci en los cuales un individuo es heterocigoto para determinar matemáticamente la cantidad de gametos únicos que el individuo puede producir. Un individuo con genotipo AABBCC no tiene loci heterocigoto y produce solo un tipo de gameto: ABC. Un individuo heterocigoto en un locus – digamos AaBBCC- puede producir dos gametos diferentes: ABC y aBC (note que en este ejemplo solamente el locus heterocigoto A contribuye a la variación de los gametos- los loci homocigotos B y C no lo hacen). Individuos heterocigotos en dos loci (AaBbCC) pueden producir cuatro tipos de gametos: ABC AbC aBC abC Y los individuos heterocigotos en tres loci ( AaBbCc) pueden producir ocho tipos de gametos: ABC ABc AbC Abc aBC aBc abC abc Este es un patrón que puede ser resumido en la siguiente fórmula: Numero de gametos únicos = 2 n Donde n es el número de loci en los cuales el individuo es heterocigoto. Por medio de un razonamiento similar, asumiendo solamente dos alelos posibles por locus, n Numero de cigotos únicos = 3 x 2 m Donde n es el número de loci en los cuales ambos padres son heterocigotos y m es el número de loci en los cuales solamente un progenitor es heterocigoto. En el ejemplo de un individuo heterocigoto en 100 loci, el número de gametos únicos posible es: n 2 100 =2 30 1.27 x 10 gametos únicos. Y si ese individuo es apareado con otro individuo igual que él, el número de cigotos únicos posibles es n n 100 0 100 x1 3 x2 =3 x2 =3 47 5.15 x 10 cigotos únicos Los procesos que aseguran la variabilidad genética son aleatorios (o casi aleatorios) por naturaleza. La distribución independiente de los genes durante la formación de células germinales es casi toda aleatoria; solamente un ligamiento estrecho previene la aleatoriedad completa. No hay forma de predecir que combinación de genes estará presente en un gameto en particular. Algunos gametos recibirán muestras favorables de genes; otros no lo harán. El proceso de selección de gametos en la formación del embrión es igualmente aleatorio. No hay forma de predecir la estructura genética del óvulo que será el próximo en madurar o predecir la estructura genética del espermatozoide que tiene éxito, entre millones, en la fertilización del óvulo. Usted puede pensar en el proceso aleatorio de distribución independiente y en la selección de gametos como dos procesos separados o como piezas de un solo proceso. De cualquier forma, el resultado es el mismo: la muestra de genes que recibe el hijo de sus padres es aleatoria. La aleatoriedad de la herencia es críticamente importante desde el punto de vista de la evolución, y como se verá más adelante, también es de vital importancia para el éxito de la selección artificial. Sin embargo, crea un problema para los criadores – reduce nuestra habilidad para controlar los resultados de los apareamientos. Podemos aumentar las probabilidades de tener una progenie superior apareando padres que sabemos que tienen valores de cría superiores, pero no tenemos el control sobre el muestreo mendeliano de los genes, los cual determina la estructura genética de la progenie. El hecho de que si el padre y la madre han producido un hijo sobresaliente en el pasado, no garantiza que producirán un hijo igualmente sobresaliente en el futuro. Igualmente, solo porque el primer apareamiento de dos individuos haya producido resultados menos deseables, eso no significa que no es posible obtener mejores resultados de este apareamiento. Muestreo mendeliano: Muestro aleatorio, de los genes parentales causados por la segregación y la distribución independiente de los genes durante la formación de células germinales y por la selección aleatoria de gametos en la formación del embrión. El muestreo mendeliano en peces es ilustrado en la figura 3.7 (Se eligieron peces como ejemplo porque tienen una alta fecundidad – un solo apareamiento produce muchos hijos). En (a) se aparean dos individuos con mérito genético inferior para la tasa de crecimiento. Los hijos de este apareamiento no son todos iguales porque el muestreo mendeliano ha causado que reciban diferentes conjuntos de genes de sus padres. Con respecto al mérito genético para la tasa de crecimiento (la escala horizontal en la figura 3.7), la progenie parece tener una distribución en forma de campana. La mayoría tiene mérito genético para la tasa de crecimiento próximo al mérito promedio de sus padres, el cual, en este caso es inferior. Algunos (aquellos en el extremo izquierdo de la distribución) son realmente pobres, pero unos pocos (aquellos en el extremo derecho de la distribución) son genéticamente capaces de un crecimiento bastante rápido. En (b) se aparean dos individuos con mérito genético para tasa de crecimiento superior. Nuevamente el muestreo mendeliano causa variación en los hijos. En este caso la mayoría de ellos son superiores – algunos extremadamente buenos – y unos pocos son inferiores. Note que a pesar de que el muestreo mendeliano causa una variación considerable en la progenie producida por un determinado apareamiento, la probabilidad de obtener un hijo superior es mayor cuando se aparean padres superiores entre sí que cuando se aparean padres inferiores. En algunos aspectos, a los hijos se le dan los genes de sus padres de una manera similar a la que se dan las cartas de un mazo: a veces se obtiene una buena mano, a veces se obtiene una mala. Estos es importante desde un punto de vista práctico para recordar y que muchos criadores no aprecian lo suficiente. La genética como un juego de cartas involucra chances – y en un cierto nivel – una cierta cantidad de suerte. Cuando pensamos que nosotros, como criadores, estamos en completo control, sobreestimamos seriamente nuestras habilidades. DOMINANCIA Y EPISTASIS Mendel descubrió que la expresión de un gen en un locus depende del otro gen presente en ese locus. Sus plantas de arvejas eran o altas o tan bajas que se consideran enanas. Las plantas enanas eran de genotipo tt, pero las plantas altas eran o TT o Tt (vea la figura 3.8). El gen para la pequeñez (t) producía un enano cuando estaba apareado con otro gen t. Pero cuando el alelo t era apareado con un alelo alto T, la planta no era de un tamaño intermedio como es de esperarse. En cambio, era tan alta como las plantas TT- el alelo t parecía no tener efecto alguno. Hoy decimos que el alelo T, es dominante sobre el alelo t. En los heterocigotos, el alelo T se expresa mientras que el alelo t no. Entonces, se dice que el alelo t es recesivo. Dominancia: Interacción entre genes en un solo locus tal que en los heterocigotos, un alelo tiene más efecto que el otro. El alelo con mayor efecto es dominante sobre su homólogo recesivo. El fenómeno de la dominancia es importante para el mejoramiento animal por dos razones. La primera se refiere a caracteres de herencia simple como los que estudiaba Mendel en sus arvejas. Para estos caracteres, la dominancia explica porque obtenemos varios fenotipos en proporciones particulares cuando hacemos apareamientos específicos. Entender la naturaleza de la dominancia en estas situaciones nos permite predecir los resultados de los apareamientos. Este capítulo contiene ejemplos que involucran caracteres de color del manto. La segunda razón involucra caracteres poligénicos. Para estos caracteres la dominancia es la fuente principal de vigor híbrido y de la depresión endogámica. (La epistasis, un concepto relacionado, es importante para el mejoramiento animal por las mismas razones). Los alelos dominantes son usualmente representados por una letra mayúscula y los alelos recesivos por una letra minúscula. En el locus J, el genotipo JJ es llamado genotipo homocigoto dominante, el genotipo Jj es el genotipo heterocigoto y el genotipo jj es el genotipo homocigoto recesivo. La letra o combinación de letras para representar un locus usualmente es una forma de abreviación relacionada con las características del gen dominante (por lo tanto el locus T para las plantas altas (tall en inglés) de Mendel versus las enanas). Desafortunadamente, la literatura genética está llena de excepciones a esta conversión: Mendel estudiaba loci que afectaban el color y forma de la semilla en las plantas de arvejas, y estos loci habían sido designados G y W, a pesar de que el color verde (de green) y la forma (wrinkle ) rugosa eran condiciones recesivas. Existen varias formas posibles de dominancia en un locus. Realmente, estas formas de dominancia no son fundamentalmente diferentes- varían solamente grado. Dominancia completa En las arvejas de Mendel, el modo de expresión de genes en el locus T era dominancia completa. Esta es la forma clásica de dominancia en la cual la expresión del genotipo heterocigoto no es diferente de la expresión del genotipo homocigota con dos genes dominantes. Los heterocigotos Tt y los homocigotas TT eran igualmente altos, fenotípicamente eran indistinguibles. La dominancia completa es común en una cantidad de caracteres de herencia simple en los animales. Ejemplos típicos en los bovinos son el carácter mocho (el alelo P para mocho es completamente dominante sobre el alelo p para astado) y el color del manto negro o rojo (el alelo B para negro es completamente dominante sobre el alelo b para rojo). Muchas condiciones letales, semiletales o deletéreas en animales, involucran dominancia completa, y el gen problemático es usualmente el alelo recesivo. Un ejemplo es el síndrome araña en las ovejas. El alelo S es el gen normal en el locus S. El alelo s es el gen recesivo responsable del síndrome araña- frecuente debilitamiento de las piernas en corderos. Los individuos SS y Ss son perfectamente normales. Solamente los corderos homocigotas recesivos ss muestran la condición de araña. Debido a que los genes deletéreos completamente recesivos pueden ser acarreados y difundidos por animales heterocigotos aparentemente normales, son de particular preocupación para los criadores. La dominancia completa es una de las formas de dominancia en las cuales los genotipos homocigotos y heterocigoto dominante tienen la misma expresión fenotípica. A diferencia del color del manto en los Shorthorn en los cuales cada genotipo (RR, Rr o rr) está asociado con un fenotipo distinto (rojo, rosillo o blanco), los caracteres afectados por la dominancia completa tienen más de un genotipo para un fenotipo. Por ejemplo, los bovinos mochos pueden ser PP o Pp. Como resultado la proporción clásica 1:2:1 esperada del apareamiento de dos heterocigotos no ocurrirá con dominancia completa. Para ver como resultan los apareamientos que involucran dominancia completa, considere cruzas de Angus y Hereford astado. El ganado Angus puro es mocho homocigota (PP). Por otro lado, los Hereford son astados. Los toros Angus apareados con vacas Hereford astadas (o viceversa) producen hijos todos heterocigotos pero fenotípicamente mochos, como puedes ver del cuadro de Punnett siguiente: tres Cuando se aparean cruzas Angus X Hereford entre ellos, se producen hijos mochos y astados en una proporción de tres mochos por un astado (vea el cuadro de Punnett siguiente). Dos de tres del tipo mocho serán heterocigotos, uno de serán mochos homocigotas. El carácter mocho no es el único carácter de herencia simple en el cual difieren Angus y Hereford. Los Hereford son colorados. La gran mayoría de los Angus son negros. La combinación de los loci P y B provee un ejemplo más complejo de dominancia completa. Asumiendo que los toros Angus son homocigotas para el gen de color negro, el apareamiento de toros Angus con vacas Hereford produce hijos todos negros y mochos. Cuando estos son apareados entre ellos, se producen nueve genotipos distintos, pero debido a la dominancia completa, se reconocen solo cuatro fenotipos: negro/mocho, negro/astado, rojo/mocho, rojo/astado. Esto sucederá en una relación de aproximadamente 9:3:3:1 Dominancia completa: Una forma de dominancia en la cual la expresión del genotipo heterocigoto es idéntica a la expresión del genotipo homocigota dominante. Dominancia Parcial La característica definitoria de la dominancia completa es que la expresión del genotipo heterocigota es idéntica a la expresión del genotipo homocigota dominante. Esto se muestra gráficamente en la figura 3.9. La línea horizontal representa una continuidad de la expresión génica. Si el locus J afectara, la altura (como el locus T de Mendel), entonces los puntos de la línea significarán mayores y mayores expresiones de la altura yendo de izquierda a derecha. Note que el punto en la línea para la expresión del heterocigota ( JJ’) es idéntico al punto representando la expresión del genotipo homocigota JJ. Por lo tanto, la dominancia es completa, y el alelo J es el dominante porque enmascara completamente la expresión del alelo J’ en el heterocigota. Dominancia parcial: Una forma de dominancia en la cual la expresión del heterocigota es intermedia entre las expresiones de los genotipos homocigotos y se acerca estrechamente a la expresión del genotipo homocigoto dominante La dominancia parcial, es representada en la figura 3.10. Con la dominancia parcial, la expresión del heterocigota es intermedia a la expresiones de los genotipos homocigotos y se parece más estrechamente a la expresión del genotipo homocigota dominante. En la figura 3.10, el genotipo JJ’ se encuentra en algún punto entre J’J’ y JJ. En (a) el heterocigoto se parece más al homocigota JJ. En este caso el alelo J es parcialmente dominante sobre el alelo J’ porque tiene una expresión mayor en el heterocigoto. J es el alelo dominante. La dominancia parcial también es mostrada en la figura (b), solo que en este caso el heterocigoto se encuentra más cerca de J’J’ que de JJ, haciendo a J’ el alelo dominante. Un ejemplo del mundo real de dominancia parcial es la condición conocida como HYPP (Parálisis hiperpotasémica periódica) en los caballos. HYPP causa episodios de temblores musculares que van desde sacudidas o temblores hasta colapso completo. En algunas instancias puede ser fatal. EL gen mutante que causa HYPP es heredado como parcialmente dominante. A pesar de que los signos clínicos de HYPP varían considerablemente entre los caballos, los síntomas son más severos para los animales HYPP homocigotos que para los heterocigotos. El HYPP es un caso particularmente interesante porque se esparce bastante rápido entre los caballos de espectáculo y placer en los Estados Unidos. Normalmente se esperaría que un gen dominante deletéreo, sea auto eliminatorio incluso si es solo parcialmente dominante, después de todo, se expresa en los heterocigotos. Sin embargo, el gen HYPP no fue rápidamente eliminado, porque (1) no es completamente letal y (2) los portadores a menudo exhiben musculatura pesada – característica deseada en las competencias de riendas. El gen ha persistido en caballos porque la tendencia de los criadores de seleccionar en su favor sin saberlo. Si compara la dominancia completa como se muestra en la figura 3.9, con la dominancia parcial como se representa en la figura 3.10, podrá ver que la forma de dominancia es definida por la posición del heterocigoto en relación a la posición de los dos homocigotos. Esto es cierto para cualquier forma de dominancia y lleva a una regla general: Para determinar el tipo de dominancia y el alelo dominante, hay que comparar las expresiones del heterocigoto con las expresiones de los genotipos homocigotos. Una forma fácil de hacer estos es usando un diagrama de línea como aquellos en las figuras 3.9 y 3.10. Ausencia de dominancia La ausencia de dominancia existe si la expresión del heterocigota está exactamente a mitad de camino entre las expresiones de los genotipos homocigotas. Ningún alelo es dominante en este caso porque ambos parecen tener una expresión igual en el heterocigoto. La ausencia de dominancia es representada en la figura 3.11. Para un ejemplo hipotético de la ausencia de dominancia, considere la resistencia a una enfermedad en particular – digamos tuberculosis. Si al exponerse al patógeno de la tuberculosis, los animales con dos copias del gen resistente a la tuberculosis (Tr) sobrevive el 100% de las veces, los animales con dos copias para el gen susceptible a la tuberculosis (Ts) y los r s heterocigotos (T T ) sobrevive el 70 % de las veces, entonces no existe dominancia en el locus. La expresión del heterocigoto está exactamente a mitad de camino entre las expresiones de los genotipos homocigotos. Ausencia de dominancia: Forma de dominancia en la cual la expresión de los heterocigotos está exactamente a mitad de camino entre las expresiones de los genotipos homocigotas. Sobredominancia La última forma de dominancia, sobredominancia, Sobre dominancia: Forma de es ilustrada en la figura 3.12. Con dominancia en la cual la sobredominancia, la expresión del heterocigoto expresión de los está fuera del rango definido por las expresiones heterocigotos esta fuera del de los genotipos homocigotas y de la expresión rango definido por las más cercana a la del genotipo homocigoto expresiones de los genotipos dominante. La sobredominancia a menudo se homocigotos y se acercan más caracteriza por tener un “heterocigoto superior”. a las expresiones de “Superior” probablemente no sea la mejor palabra genotipos homocigotos – “extremo” puede ser la correcta. En la figura dominante. 3.12 (a) el heterocigoto se encuentra a la derecha del genotipo JJ. J es entonces el alelo dominante. En (b) el heterocigoto se encuentra a la izquierda del genotipo J’J’, haciendo a J’ el alelo dominante. La supervivencia en ratas salvajes provee un ejemplo de sobredominancia. El gen para la resistencia al veneno anticoagulante warfarina es heredado como dominante con respecto a la resistencia al veneno. Ambos, homocigotos y heterocigotos no son afectados por warfarina. Desafortunadamente – al menos desde el punto de vista de las ratas- los homocigotas necesitan mayores niveles de vitamina K de lo que está disponible en dietas normales. Así en lugares donde la warfarina es utilizada, las ratas que no tienen el gen de la resistencia sucumben al envenenamiento, las ratas homocigotas para el gen sufren deficiencia de vitamina K y los heterocigotos permanecen sanos. Con respecto a la supervivencia el locus de la warfarina desarrolla sobredominancia. La sobredominancia es la forma más extrema de dominancia. Si sus cuatro formas son ordenadas por grado de dominancia, progresarían de ausencia de dominancia a dominancia parcial a dominancia completa a sobredominancia (figura 3.13) Errores comunes acerca de Dominancia El fenómeno de dominancia es a menudo incomprendido. Los estudiantes a menudo asumen, por ejemplo que los genes dominantes son “buenos” y que los recesivos son “malos”. En muchos casos tienen razón. Los genes recesivos letales y semiletales son claramente malos, y hay una razón para creer que la habilidad de los alelos más favorables de ser dominantes sobre alelos menos favorables es algo que ha evolucionado a través del tiempo (después de todo los dominantes desfavorables tenderían a ser eliminados por selección natural). Como veremos en los capítulos 7 y 17, la teoría del vigor híbrido y de la depresión endogámica está basada en la asunción de que los alelos dominantes son generalmente más favorables que los alelos recesivos. Pero hay excepciones a la regla. Claramente el gen HYPP en caballos, parcialmente dominante, no puede ser considerado favorable con respecto a la salud del animal. Tampoco pueden los genes para color de manto rojo o la presencia de cuernos en el ganado ser consideradas necesariamente desfavorables. Un error más fuerte es que los genes dominantes son más comunes que los genes recesivos. En realidad, los genes letales recesivos tienden a ser raros, porque son auto eliminatorios, y cualquier recesivo desfavorable tiende a convertirse en menos común porque son seleccionados en contra a través del tiempo. Sin embargo, nuevamente las excepciones abundan. Por ejemplo, en la población de ganado Hereford astado los genes dominantes para mocho y color del manto rojo son comunes al punto de ser los únicos alelos en sus respectivos loci. Puede ser cierto que generalmente los alelos dominantes son “mejores” y son más comunes que los alelos recesivos. Sin embargo, es importante recordar que estas características de alelos dominantes y recesivos no son parte de la definición de dominancia. La dominancia tiene que ver con la expresión relativa de los alelos en los heterocigotos - nada más. Epistasis La dominancia involucra interacción de genes en un solo locus porque afectan al fenotipo de un individuo. Los genes en diferentes loci también pueden interactuar, y este tipo de interacción es denominada epistasis. La epistasis puede ser definida como una interacción entre genes de diferentes loci, tal que la expresión de los genes en un locus depende de los alelos presentes en uno o más loci diferentes. Con respecto a los caracteres de herencia simple, la epistasis es como la dominancia, afecta los tipos y proporciones de fenotipos que podemos esperar de determinados apareamientos. Y como la dominancia, la epistasis es una fuente de vigor híbrido y depresión endogámica en caracteres poligénicos. Un ejemplo de epistasis en herencia simple que es relativamente fácil de entender es el color del manto en los Labradores retrievers. Los labradores tienen tres colores básicos: negro, chocolate y amarillo. Estos colores son determinados por genes en dos loci: El locus B (negro) y el locus E (extensión de la pigmentación), como sigue. B_E Epistasis: Interacción entre genes en loci diferentes tal que la expresión de los genes en un locus depende de los alelos presentes en uno o más de un loci. Negro bbE_ Chocolate __ ee Amarillo Los guiones en estos genotipos indican que los alelos pueden ser sustituidos sin cambiar el genotipo. Por ejemplo, los labradores negros pueden ser BBEE, BBEe, BbEE o BbEe. Los labradores amarillos pueden ser BBee, Bbee, o bbee. Note que la expresión de los genes en el locus negro depende de los alelos presentes en el locus de extensión. Entonces siempre y cuando haya al menos un alelo E en el locus de extensión, no aparece, la dominancia completa en el locus negro, con negro siendo dominante sobre chocolate. Sin embargo, si el genotipo en el locus de extensión es ee, entonces los genes en el locus negro son irrelevantes – todos los animales serán amarillos. Una muestra de los apareamientos de Labradores se puede ver en el cuadrado de Punnett en la figura 3.14. El apareamiento de dos animales totalmente heterocigotos negros es representado en (a). Este apareamiento produce una mezcla de cachorros negros, chocolate y amarillos. En (b) se aparean dos chocolates (bbEe). Debido a que chocolate es una condición recesiva, normalmente se esperaría que los chocolate de raza pura produzcan solo chocolates. Sin embargo, debido al efecto de la epistasis de los genes en el locus E, este apareamiento también produce cachorros amarillos. Solo los Labradores amarillos de pura raza (c) – un amarillo apareado con un amarillo producen solo amarillos. En este capítulo hemos rasgado la superficie de la genética. Se ha incluido aquí solamente la información mínima necesaria para la comprensión práctica del mejoramiento animal, dejando áreas enteras de la bioquímica del ADN y molécula relacionadas, la fisiología de la célula. HERENCIA RELACIONADA AL SEXO Para algunos caracteres, la expresión génica difiere en machos y hembras. Las causas son variadas, pero se pueden poner bajo 3 categorías: Ligadas al sexo; Limitadas al sexo; Herencia influenciada por el sexo. Herencia ligada al sexo En mamíferos un par de cromosomas comprenden los cromosomas sexuales, X e Y, con las hembras que tienen 2 cromosomas X y los machos que tienen un X y un Y. La hembra hereda un cromosoma X de cada uno de sus progenitores. Los machos heredan un cromosoma X de su madre y un cromosoma Y de su padre. (En las aves es a la inversa, los machos son XX y las hembras XY). Los cromosomas X e Y, aunque son miembros de un par, son bastante diferentes entre sí. No tienen correspondencia en las regiones del ADN, y los genes de estos cromosomas se dicen que son ligados al sexo. Debido a que los machos mamíferos son hemicigotas para los genes que aparecen en el cromosoma X (los machos tienen solo 1 copia), la herencia ligada al sexo difiere del patrón de la tradicional genética mendeliana. Un ejemplo de herencia ligada al sexo es la coloración “carey” o tricolor en gatos. El “carey” es una mezcla de colores que aparece en parches, siempre con algo de naranja, a menudo con blanco y negro o gris atigrado. (Con suficiente blanco es un gato Calico). Los carey ó “tricolor” son hembras (con muy raras excepciones), porque el locus para coloración naranja se encuentra en el cromosoma X. Las hembras con 2 cromosomas X pueden tener genotipos tanto OO, Oo, ó bien oo en el locus naranja (O del inglés orange). Las OO son naranjas, las oo son del color que dicten los otros loci (es decir que el alelo o no tiene efecto) y las Oo son carey o tricolor. La razón por la que ocurren los parches de color en los genotipos Oo es que un determinado cromosoma X es inactivado al azar en las células que se forman en el desarrollo temprano del embrión. En algunas células embrionarias, el cromosoma X derivado del padre es inactivado. Por lo tanto todas las células hijas tendrán inactivados los cromosomas X paternos. En otras células embrionarias y sus descendientes, el cromosoma X materno es el que se inactiva. Como resultado en ciertas áreas del cuerpo de individuos carey el alelo O es activo y el color naranja se expresa, pero en otras áreas el alelo o es activo y el color naranja no se expresa. Los machos tienen un solo cromosoma X y por lo tanto un solo alelo en el locus naranja. Un macho con genotipo O es naranja, mientras que con genotipo o no es naranja y no existen machos carey o tricolor. En resumen: Hembras Machos OO Naranja O Naranja Oo Carey o No naranja oo No naranja Herencia ligada al sexo: El patrón de herencia para genes localizados en los cromosomas sexuales. Hemicigotas: Tener solamente un gen de un par, por ejemplo un gen en el cromosoma X de los machos mamíferos. Herencia limitada al sexo En la herencia limitada al sexo la expresión fenotípica a un carácter se limita a uno de los sexos. Por ejemplo la producción de leche, se limita al sexo, los machos no producen leche, aunque portan genes para ello. Lo caracteres limitados al sexo, se piensa que son condicionados hormonalmente. Las hormonas femeninas permiten el desarrollo mamario y la producción de leche. Si los genes en los cromosomas sexuales están completamente involucrados, estos tienen probablemente solo un efecto indirecto a través de su influencia en la producción hormonal. Herencia limitada al sexo: Un patrón de herencia en el cual la expresión fenotípica se limita a un solo sexo. Herencia influenciada por el sexo En la herencia influenciada por el sexo, el modo de expresión del gen difiere entre machos y hembras. Por ejemplo un alelo se puede expresar como dominante en un sexo y como recesivo en el otro. La herencia de los tocos o cachos (vestigios de cuernos generalmente pequeños en el ganado bovino) es influenciada por el sexo. El alelo para los tocos, Sc del inglés scurs, es dominante en los machos y recesivo en las hembras. Así, si un macho porta solo una copia del alelo tendrá tocos, pero una hembra debe tener las dos copias del alelos para tener tocos. Hembras Machos Sn Sn Sin tocos Sn Sn Sin tocos Sc Sn Sin tocos Sc Sn Con tocos Sc Sc Con tocos Sc Sc Con tocos Un patrón idéntico afecta la expresión de los cuernos en ovinos. Note que al contrario que las características limitadas al sexo, las influenciadas por el sexo pueden aparecer en ambos sexos. Aunque generalmente, son más frecuentes en un sexo que en el otro. Herencia influenciada por el sexo: Un patrón de herencia en el cual el modo de expresión del gen difiere entre machos y hembras, por ejemplo un alelo puede ser dominante en machos y recesivo en hembras. GENÉTICA DE POBLACIONES Los principios mendelianos explican los mecanismos genéticos en individuos. Sin embargo, nuestra labor no es cambiar a los individuos, sino las poblaciones. Entonces debemos tomar los conocimientos sobre la herencia mendeliana y extenderlo del nivel del individuo al nivel de las poblaciones. FRECUENCIAS GÉNICAS Y GENOTÍPICAS Al describir un individuo para un carácter de herencia simple, nos referimos a genes específicos, o bien podemos describir su genotipo para uno o dos locus. Por ejemplo, podemos referirnos a la raza de gallinas Andaluza azul como si tuviera alelos negros (B), o blanco (b) en un locus que afecta el color de las plumas, o podemos decir que un pollo en particular tiene un genotipo heterocigoto Bb en ese locus. Sin embargo ¿Cómo describimos una población? ¿Cómo describimos un lote completo de gallinas andaluzas azules? La respuesta es utilizar la frecuencia de genes y genotipos. La frecuencia génica o frecuencia alélica es la frecuencia relativa de un alelo particular en una población. Es una medida de cuan común es el alelo en relación a otros alelos en ese locus. Las frecuencias relativas van de 0 a 1. Por ejemplo, si un alelo no existe en una Frecuencia génica o frecuencia población, su frecuencia es 0. Si es el único alelo alelica: Es la frecuencia en su locus en la población, su frecuencia génica relativa de un alelo particular es 1. Si alcanza al 35 % de los genes en el locus en en una población. la población, su frecuencia génica es 0.35. Cuando hay solo dos alelos posibles en un locus, la frecuencia del alelo “dominante” es comúnmente representada por la letra minúscula p y la frecuencia del alelo “recesivo” por la letra minúscula q. (Los términos dominante y recesivo están puestos entre comillas porque hay situaciones en que ningún alelo es dominante. En casos de ausencia de dominancia, la asignación de p o q para referirse a la frecuencia de un alelo en particular es arbitraria). Por ejemplo considere un lote de 100 andaluzas, 36 son negras (BB), 44 son azules (Bb, en realidad gris), 20 son blancas (bb). En el locus que afecta el color de las plumas de las andaluzas hay un total de 200 genes en esta población – dos genes por cada uno de los 100 individuos. Los 36 individuos negros, cada uno tiene 2 genes (o alelos) negros, los 44 azules cada uno tiene un gen negro, y los blancos no tienen genes negros. La cantidad total de genes negros en el lote es por lo tanto 2 x 36 + 44 =116, y la frecuencia génica del alelo negro es entonces 116 de 200 o en forma decimal, 0.58. Igualmente, no hay genes blancos en las gallinas negras, 44 genes blancos en las azules y 40 genes blancos en los individuos blancos, un total de 84 genes blancos en el lote. La frecuencia génica del alelo blanco es entonces 84 de 200 o 0.42. En cada caso hemos simplemente contado el número de genes de un tipo particular y dividido por el total de genes en un locus en la población. Las ecuaciones para las frecuencias génicas en nuestro ejemplo de las andaluzas pueden ser escritas así: Note que p + q = 1. Esto siempre será cierto si hay solo 2 alelos posibles en un locus. Si hay múltiples alelos en un locus, entonces la suma de las frecuencias génicas debe ser igual a 1. Por ejemplo, si los alelos posibles fueran B, b y b’ debemos llamar las frecuencias génicas de estos alelos p,q,r, respectivamente. Entonces p + q + r = 1. La frecuencia genotípica es la frecuencia relativa de un genotipo particular de un locus en una población. Las letras mayúsculas son utilizadas para anotar las frecuencias genotípicas. Con sólo dos alelos en un locus, P se refiere a la frecuencia genotípica del genotipo homocigota “dominante”, H se refiere a la Frecuencia genotípica: Es la frecuencia del genotipo heterocigoto y Q se refiere frecuencia relativa de un a la frecuencia del genotipo homocigota genotipo particular de un locus en una población. “recesivo”. En nuestro lote de andaluzas, hay 36 genotipos BB, 44 genotipos Bb y 20 genotipos bb de un total de 100 individuos. Por lo tanto: Para calcular frecuencias genotípicas, simplemente cuente el número de individuos en un genotipo particular y divide por el número total de individuos en una población. Note que P+H+Q = 1. Esto siempre será cierto si hay solo tres genotipos posibles posibles en un locus. Si hay más de tres genotipos posibles, entonces la suma de las frecuencias genotípicas de cada genotipo tiene que ser igual a 1. Por ejemplo si los alelos posibles fueran B,b y b’ con posibles genotipos de un locus BB,bb, b’b’, Bb, Bb’ y bb, podríamos llamar las frecuencias genotípicas P,Q,R,H (Bb) ,H( Bb’), y H (bb’),respectivamente. Entonces P+Q+R+H (Bb)+H( Bb’)+H (bb’) = 1. Hay un número de factores que afectan las frecuencias génicas y genotípicas en una población, entre ellos, las herramientas básicas para el mejoramiento animal: selección y sistemas de apareamiento. El estudio de estos factores compone la rama de la genética conocida como genética poblacional. En una población grande en apareamiento, en ausencia de selección, mutación o migración las frecuencias génicas y genotípicas se mantienen constantes de generación en generación, y las frecuencias genotípicas se relacionan con las frecuencias génicas por medio de fórmulas. LA LEY DE HARDY-WEINGBERG La ley de Hardy- Weinberg esencialmente dice que si no hay fuerzas para cambiar las frecuencias génicas y genotípicas en una población, esas frecuencias no van a cambiar. ¿Cuáles son las fuerzas que cambian las frecuencias génicas y genotípicas? La ley menciona específicamente la selección, mutación y migración. Sabemos que la selección modifica las frecuencias. De hecho, el propósito de la selección artificial es cambiar las frecuencias génicas. La mutación, el proceso que altera el ADN para crear nuevos alelos, tiene algún efecto sobre las frecuencias génicas y genotípicas, pero debido a que la mutación es un evento raro, el efecto es pequeño. La migración, es el movimiento de individuos hacia dentro o fuera de una población. La migración particularmente involucra la introducción de un número grande de individuos genéticamente diferentes en una población, puede tener grandes efectos en las frecuencias génicas y genotípicas. La ley de Hardy- Weinberg también expone que una población en equilibrio tiene que ser apareada de manera aleatoria. Las poblaciones pequeñas se convierten en consanguíneas rápidamente, y como hemos visto anteriormente, la consanguinidad cambia las frecuencias genotípicas incrementando la homocigocis. Y en las pequeñas poblaciones las frecuencias génicas pueden cambiar puramente por casualidad, fenómeno conocido como deriva genética. El apareamiento aleatorio implica la ausencia de cualquier esquema de apareamiento sistemático. En nuestro ejemplo del locus B, si no hubiésemos apareado de manera aleatoria, sino que hubiéramos aplicado una regla que diga que los genotipos BB solo pueden aparearse con otros genotipos BB, entonces podríamos haber esperado frecuencias genotípicas diferentes (y no estáticas). Entonces el apareamiento aleatorio es necesario para el equilibrio Hardy- Weinberg, y además como sugiere nuestro ejemplo, solo se necesita una población con apareamiento aleatorio para llegar al equilibrio. El apareamiento aleatorio entre los animales F1 creó un equilibrio en la generación F2. La derivación de las fórmulas Hardy- Weinberg relacionando frecuencias genotípicas con frecuencias génicas es simple. Si las frecuencias génicas en un locus particular en una población en equilibrio son p y q, entonces las fórmulas son las que aparecen en el siguiente cuadro de Punnett. p q p p2 pq q pq q2 No debemos confundir p2 2pq y q2 de Hardy- Weinberg con la razón 1:2:1 esperada cuando se aparean dos homocigotos. A pesar de que la razón 1:2:1 puede ser considerada una aplicación especial de la ley Hardy- Weinberg (cuando p=q=0.5), los dos conceptos son utilizados en contextos muy diferentes. ¿Existen verdaderamente poblaciones en equilibrio Hardy- Weinberg? Claramente no. Ninguna población es infinitamente grande, apareada perfectamente al azar o libre de selección natural y mutación. Algunas poblaciones se encuentran muy cercanas al equilibrio, sin embargo, que las fórmulas Hardy- Weinberg encajen bien puede ser de gran utilidad. Como un ejemplo de la utilidad Hardy- Weinberg, vayamos nuevamente a las poblaciones 1 y 2 y su descendencia F1. El objetivo de hacer este cruzamiento era mostrar como los cruzamientos en este caso incrementan la heterocigosis. Recuerde por la figura 4.4 que la frecuencia genotípica de los heterocigotas para el locus B en la población F1 era 0.74.Determinamos esto multiplicando las frecuencias génicas de las poblaciones de los padres. Lo que no sabemos son las frecuencias de los heterocigotas en esas poblaciones originales. Sin embargo podemos calcularlas asumiendo el equilibrio Hardy- Weinberg. Las frecuencias de las poblaciones heterocigotas en las poblaciones 1 y 2, como fue determinado por la fórmula Hardy- Weinberg son, H1 = 2p1q1 = 2(.8)(.2) = 32 H2 = 2p2q2 = 2(.1)(.9) = .18 La frecuencia heterocigota promedio en las poblaciones de los padres es (H1+H2) /2 ó (0.32 +0.18)/2 =0.25.Compare esa figura con la frecuencia de los heterocigotas en los cruzamientos F1 (0.74). Cruzando las poblaciones 1 y 2, la heterocigosis casi triplicó. Este es un ejemplo dramático de cómo el cruzamiento aumenta la heterocigosis. Si las poblaciones de los padres hubiesen sido más similares, (por ejemplo que sus frecuencias génicas en el locus B no hayan sido tan diferentes), el incremento de la heterocigosis a partir del cruzamiento hubiese sido menos sensacional pero todavía evidente. CARACTERES DE HERENCIA SIMPLE Y POLIGÉNICOS El aspecto principal de los caracteres de herencia simple es que los mismos son afectados sólo por unos pocos genes. El color del manto, la presencia de cuernos y defectos genéticos como el síndrome araña en las ovejas son ejemplos de caracteres de herencia simple. Un solo locus o como mucho unos cuantos loci están involucrados en su expresión. Hay dos características secundarias de los caracteres de herencia simple. Primero los fenotipos para estos caracteres tienden a ser, por naturaleza “unos u otros” o categóricos (descriptos estableciendo categorías). Un labrador es negro o chocolate o amarillo, una vaca es mocha o astada, un cordero tiene la condición de araña o no. El color del manto, la presencia de cuernos y el síndrome araña también son denominados caracteres cualitativos o categóricos por su expresión “unos u otros”. Es posible (aunque raro) que los caracteres de herencia simple sean cuantitativos, que tengan fenotipos que son medidos con números que oscilan más o menos continuamente de valores grandes a intermedios a pequeños. Un ejemplo es el peso corporal cuando es afectado por el gen para enanismo. El carácter es medido en kilos de peso, pero es de herencia simple. Segundo, los caracteres de herencia simple son muy poco afectados por el ambiente. Si un Labrador chocolate pasa mucho tiempo en el sol su color de manto se va a aclarar, pero todavía se lo va a poder distinguir de un labrador negro o amarillo. Su fenotipo es claramente chocolate. En contraste, los caracteres poligénicos son afectados por muchos genes. Ejemplos de caracteres poligénicos incluyen la tasa de crecimiento, la producción de leche, y velocidad. Se sabe poco sobre genes específicos que afecten estos caracteres y solo podemos concluir que hay muchos de ellos. Como los caracteres de herencia simple, los caracteres poligénicos comúnmente tienen características secundarias similares. Los fenotipos para caracteres poligénicos son usualmente descriptos por números. Hablamos de 200 kg de peso al destete, 15.000 kg de leche por lactancia, y 20 segundos para correr un cuarto de milla (300 m). En vez de ser o uno u otro por naturaleza o que caigan en categorías distintivas como lo hacen los fenotipos para caracteres de herencia simple, los fenotipos para caracteres poligénicos son típicamente cuantitativos o continuos en su expresión. La mayoría (pero no todos) los caracteres poligénicos son entonces caracteres cuantitativos. Los caracteres poligénicos son claramente afectados por el ambiente. Si a las vacas, cerdos u ovejas se les restringe el alimento, crecen más lentamente y producen menos leche. Si los caballos no son entrenados, no correrán rápido. Sin embargo, debemos ser cuidadosos, de no clasificar un carácter como de herencia simple o poligénico en base a características secundarias solamente. Hay algunos caracteres de herencia simple que tienen características secundarias de caracteres poligénicos y viceversa. El peso corporal, cuando es afectado por un gen de enanismo, es un ejemplo. Otro ejemplo es el carácter llamado distocia o dificultad al parto. Los fenotipos para la distocia a menudo caen en dos categorías: parto asistido o no asistido. Debido a la naturaleza de estos fenotipos, se podría asumir que la distocia es un carácter de herencia simple. Sin embargo, es poligénico porque es afectado por muchos genes. Muchos genes afectan el tamaño del feto (o los fetos), el tamaño del área pélvica de la madre, su perseverancia durante el parto, etc. Lo importante de recordar al decidir si un carácter es de herencia simple o poligénico es que las características secundarias no deberían ser un factor determinante – es la cantidad de genes involucrados lo que cuenta. A menudo se confunden los términos “de herencia simple” con “cualitativo” y “poligénico” con “cuantitativo”. No son sinónimos. La mayoría de los caracteres de herencia simple son cualitativos, y la mayoría de los caracteres poligénicos son cuantitativos. Pero el peso al destete, cuando es afectado por el gen del enanismo, es un ejemplo de carácter cuantitativo de herencia simple, y la distocia es un ejemplo de un carácter cualitativo poligénico. La herencia simple vs. poligénica se refiere a como se hereda el carácter. Cualitativo vs. cuantitativo se refiere a como se expresa el carácter. La distocia es un ejemplo de una clase especial de caracteres llamada caracteres umbral. Estos caracteres son poligénicos y exhiben fenotipos categóricos. Otros ejemplos de caracteres umbral son la fertilidad (como una medida del éxito o falla en la concepción), modo de andar (trotador o de paso). Los caracteres umbral presentan problemas particulares y serán discutidos luego. En general los caracteres poligénicos son más importantes en las especies productoras de alimento y fibra, – como tasa de crecimiento, fertilidad, producción de leche, etc. – los que determinan productividad y rentabilidad. Esto también sucede en especies recreacionales y de compañía también. Por ejemplo, la velocidad y resistencia, caracteres importantes para animales de carrera, son poligénicos – no hay un “gen de velocidad” o “gen de resistencia.” Sin embargo hay instancias donde los caracteres de herencia simple asumen importancia económica. Algunos mercados son sensibles al color de manto y plumas. En las poblaciones de ganado donde el alelo para mocho es raro, los animales mochos pueden ser particularmente valiosos. Y los animales con defectos genéticos heredados de manera simple son inevitablemente menos valiosos. Sin embargo, en general los caracteres de herencia simple son menos importantes que los caracteres poligénicos. Por lo tanto, aquí se hace énfasis en los caracteres poligénicos. CARACTERÍSTICAS COMUNES A CARACTERES DE HERNCIA SIMPLE Y CARACTERES POLIGÉNICOS. Los caracteres de herencia simple y poligénicos tienen mucho en común. Para comenzar, los genes que afectan a ambos tipos de caracteres están sujetos al mismo mecanismo mendeliano. Las leyes de Mendel de segregación y distribución independiente se aplica a los genes que influencian los caracteres poligénicos al igual que lo hacen a los genes que influencian los caracteres de herencia simple. La dominancia y epistasis también afectan a la expresión de los genes para ambos tipos de caracteres. Es cierto que muchos de los ejemplos prácticos utilizados para ilustrar el mecanismo de Mendel involucran caracteres de herencia simple. Esto es solo porque los genes que afectan estos caracteres son bien entendidos – después de todo hay pocos de ellos. Debido a que son muchos los genes que afectan los caracteres poligénicos, y a que el efecto de cada gen es tan pequeño, sabemos poco y nada sobre ellos. Entonces, es difícil utilizar caracteres poligénicos como ejemplos de la herencia mendeliana. Secundariamente, las herramientas básicas del mejoramiento animal – selección y apareamiento – son las mismas para ambos tipos caracteres (de herencia simple y poligénicos). Cuando los criadores seleccionan para cualquiera de los dos tipos de caracteres, están tratando de aumentar las frecuencias de los alelos favorables. Un criador que selecciona solamente animales mochos de un rebaño de animales donde hay mochos y astados va a aumentar la frecuencia para el alelo mocho en el rebaño. Igualmente, un criador que selecciona para área de ojo de bife (una medida de la musculatura) en una piara de cerdos va a aumentar la frecuencia de los genesdistribuidos entre muchos loci – que influencian favorablemente la musculatura. En el primer ejemplo, el criador seleccionó para un carácter de herencia simple, y en el segundo ejemplo seleccionó para un carácter poligénico, pero el efecto en las frecuencias génicas fue el mismo. Los sistemas de apareamiento, afectan las combinaciones de genes de la misma manera para caracteres de herencia simple y poligénicos. Cuando un criador de caballo cruza alazanes (CC) y cremellos o crema (ccr ccr) para producir palominos (C ccr), la frecuencia genotípica de los heterocigotas en el locus C aumenta. Igualmente cuando los criadores cruzan líneas no relacionadas o crían para producir vigor híbrido, la heterocigosis aumenta en muchos loci. Ya sea que los criadores hagan apareamientos específicos para afectar caracteres de herencia simple como el color del manto o cruzamientos para afectar al conjunto de caracteres poligénicos que responden al vigor híbrido, están usando los sistemas de apareamiento para crear combinaciones de genes deseables. Crema El gen Crema (Cr/cr) diluye los pigmentos colorados como los negros. Tiene un efecto al fenotipo ya con una parte de la pareja (Crcr), pero sí está doble (CrCr) lo afecta mucho más. El caballo alazán con un alelo crema es un palomino, es decir que tiene la capa amarilla y crines más claros. Con dos allelos resulta un caballo casí blanco con crines blancos y ojos azules claros y el color se llama Cremello.