RESPUESTAS AL TALLER SOBRE EFICACIA BIOLÓGICA
Y SELECCIÓN NATURAL
1. Eficacias biológicas relativas: WAA = 1 WAa = ½, y Waa = ¾. Este es un caso
de inferioridad del heterocigoto
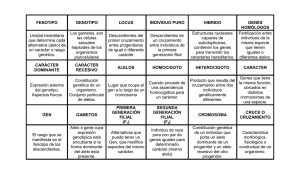
Genotipos
AA
Aa
Aa
Total
Frecuencias
cigóticas
iniciales
p2
2pq
q2
1
Eficacia
Biológica (W)
en términos
algebraicos
1+s
1
1+t
Proporciones
después de
selección
p2 (1 + s)
2pq (1)
q2 (1 + t)
1 + p2s + q2t
2pq (1)
1 + p2s + q2 t
q2 (1 + t)
1 + p2s + q2t
1
p2 (1 + s)
Frecuencias
1 + p2s + q2 t
genotípicas
después de la
selección
Frecuencia de
a después de
selección
q = pq + q2(1 + t)
1 + p2s + q2 t
q + q2t – q(1 + p2s + q2 t)
1 + p2s + q2 t
Cambio de q
∆q =
q en el
equilibrio
ˆq = ∆q = 0
=
=
Frecuencia
de a
q
q + q2t
1 + p2s + q2 t
pq(qt – ps)
1 + p2s + q2 t
⇒ qt = ps ⇒ qt – ps = 0 ⇒ qt – (1 – q) s = 0 ⇒ qt – s + qs = 0
⇒ qt + qs = s ⇒ q(t + s) = s ⇒ q = s / (t + s)
⇒ q en el equilibrio = s / (t + s)
reemplazando WAa = 0,5 equivalente a 1 en términos algebraicos, entonces
WAA = 1 + s => 0,5 + s = 1 y Waa = 1 + t => 0,5 + t = 0,75
s = 0,5 y t = 0,25
ˆq = 0,5 / (0,5 + 0,25) = 0,6667
2.
Distribución de
la población
antes de la
selección
diversificadora
Distribución de
la población
actuando la
selección
diversificadora
Distribución de
la población
después de la
selección
diversificadora
3.
Genotipo
AA
Aa
aa
# de
gametos
100
100
100
# de individuos que llegan a
adultos
3/4 = 75%
1/4 = 25%
0,2/4 = 5%
W
s
75/75 = 1
0
25/75 = 1/3 2/3
5/75 = 1/15 14/15
4.
No. Esta población no tiene reproducción diferencial. No hay diferencias
en la eficacia biológica de los diferentes fenotipos. Aunque el enunciado
no lo dice claramente, se deduce que no hay variación genética, por lo
tanto no hay un sustrato donde la selección pueda actuar.
5.
Si. En el caso de la selección direccional se favorece la proliferación de
un alelo a costa de otro. El resultado en ausencia de mutación será la
fijación del alelo favorecido y la perdida del menos eficaz. Cuando se
pierde el alelo con menor eficacia biológica, desaparece de la población
los genotipos homocigotos para tal alelo. En el caso de desventaja del
alelo dominante, la selección puede llevar a la extinción de los genotipos
homocigotos para tal alelo dominante y de los genotipos heterocigotos.
En el caso de desventaja del alelo recesivo, la selección no llevará a cero
la frecuencia del genotipo homocigoto recesivo porque el alelo recesivo
estará a salvo de la selección cuando se encuentre en condición
heterocigótica. El cruce entre heterocigotos siempre dará genotipos
homocigotos recesivos pero su frecuencia puede ser muy cercana a cero
a causa de su desventaja relativa respecto al alelo dominante. En el caso
de ausencia de dominancia la extinción de uno u otro alelo y la
consiguiente desaparición del genotipo homocigoto, dependerá de su
frecuencia inicial. Si la frecuencia es menor que 0,5 probablemente la
selección llevará a tal alelo hacia la extinción.
6.
ST/ST = 0,47; ST/AR = 1 y AR/AR = 0,62. Después de nueve
generaciones los genotipos heterocigotos ST/AR tenían la mayor eficacia
biológica; por lo tanto, este es un caso de heterosis (vigor híbrido o
sobredominancia).
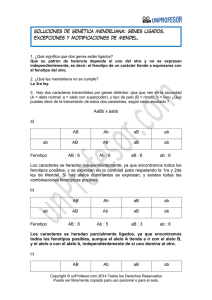
Genotipos
AR/AR
ST/AR
ST/ST
Total
Frecuencia
de AR
p
p2
Frecuencias
2pq
q2
1
cigóticas
iniciales
Eficacia
1–s
1
1–t
Biológica
(W)
p2 (1 – s)
2pq (1)
q2 (1 – t)
1 – p2s – q2t
Proporcione
s después
de la
selección
p2 (1 – s)
Frecuencias
2pq (1)
q2 (1 – t)
1
2
2
2
2
2
2
genotípicas 1 – p s – q t 1 – p s – q t 1 – p s – q t
después de
la selección
p=
p2 (1 – s) + pq
=
p – p2s
Frecuencia
2
2
1–ps–q t
– p2s – q2 t
de ST
después de
la selección
Cambio de p
∆p = p – p2s – p(1– p2s – q2 t)
= – pq(ps – qt)
2
2
1–ps–q t
1 – p2s – q2 t
p en el
ˆp = ∆p = 0
equilibrio
⇒ ps = q t ⇒ ps – q t = 0 ⇒ ps – (1 – p) t = 0 ⇒ ps – t + pt = 0
(ˆp)
⇒ ps + pt = t
⇒ p(s + t) = t ⇒ p = t / (s + t)
⇒ ˆp = t / (s + t)
reemplazan
do
WAR/AR = 0,62 ⇒ s = 1 – 0,62 = 0,38
WST/AR = 1 ⇒ s = 1 – 1 = 0
WST/ST = 0,47 ⇒ t = 1 – 0,47 = 0,53
⇒ ˆp = 0,53 / (0,53 + 0,38) = 0,58
7. (a) Ventaja del alelo dominante sobre el recesivo en condiciones de
dominancia completa:
AA
p2 2pq
Aa
q2
aa
⇒
Frecuencias
⇒
WAA =
WAa > Waa
Eficacias Biológicas Relativas
WAA =
WAa = 1 y Waa = (1 – saa)
pt+1 = [pt2(1) + ptqt(1)] / [pt2(1) + 2ptqt(1) + qt2(1 – saa)]
pt+1 = [pt(pt + qt)] / (pt2 + 2ptqt + qt2 – qt2saa)
como p + q = 1 y p2 + 2pq + q2 = 1
pt+1 = pt / (1 – qt2saa)
(b)
ventaja del alelo dominante sobre el recesivo en condiciones de
dominancia parcial:
AA Aa
p2 2pq
WAA >
aa
q2
⇒
WAa > Waa
WAA =
1; WAa = 1 – hsaa y Waa = (1 – saa)
Frecuencias
⇒
Eficacias Biológicas Relativas
h mide el grado de dominancia para la eficiacia biológica
pt+1 = [pt2(1) + ptqt(1 – hsaa)] / [pt2(1) + 2ptqt(1 – hsaa) + qt2(1 – saa)]
pt+1 = (pt2 + ptqt – ptqthsaa) / (pt2 + 2ptqt – 2ptqthsaa + qt2 – qt2saa)
como p + q = 1 y p2 + 2pq + q2 = 1
pt+1 = (pt – ptqthsaa) / (1 – 2ptqthsaa – qt2saa)
8. Este es un caso de heterosis (vigor híbrido, sobredominancia)
WAA = 1 – s ; WAa = 1,0; Waa = 1 – t
WAA = 0,9; WAa = 1,0; Waa = 0,7
s = 1 – 0,9 = 0,1; t = 1 – 0,7 = 0,3
Frecuencia inicial de A = pt = 0,5 y de a = qt = 0,5
Genotipos
AA
Aa
aa
Total
Frecuencias
cigóticas
iniciales
pt2
2ptqt
qt2
1
Eficacia
Biológica
(W)
1–s
1
1–t
Proporcione
s después
de la
selección
pt2 (1 – s)
2ptqt (1)
qt2 (1 – t)
1 – pt2s – qt2 t
Frecuencias
genotípicas
después de
la selección
pt2 (1 – s)
1 – pt2s – qt2t
2ptqt (1)
1 – pt2s – qt2t
qt2 (1 – t)
1 – pt2s – qt2t
1
Frecuencia
de A
después de
la selección
pt+1 =
reemplazan
do
pt+1 =
pt2 (1 – s) + ptqt
1 – pt2s – qt2t
0,5 – (0,5)2(0,1)
1 – (0,5)2(0,1) – (0,5)2(0,3)
=
=
Frecuencia
de A
pt = 0,5
pt – pt2s
1 – pt2s – qt2t
0,53
9. (a) Wi → 0,7/0,7; 0,2/0,7; 0,5/0,7; 0,5/0,7; 0,6/0,7; 0,5/0,7
(b) W media = (0,35*1,0) + (0,05*0,28571) + (0,10*0,71429) + (0,20*0,71429)
+ (0,15*0,85714) + (0,15*0,71429) = 0,814287
10. La hipótesis supone un vigor híbrido (heterosis o dominancia) en las
poblaciones de pinos; los individuos que porten mayor cantidad de loci
aloenzimáticos heterocigóticos tendrían mayor tasa de crecimiento y
supervivencia que las plantas más homocigóticas. Para probar tal hipótesis
se recomienda un análisis comparativo de las frecuencias genotípicas de
varios loci aloenzimáticos entre poblaciones con distintas tasas de
crecimiento y supervivencia. Como ejemplo, se pueden tomar muestras de
hojas de 30 individuos escogidos al azar de cada una de las poblaciones
previamente catalogadas como de alta y baja tasa de crecimiento y alta y
baja supervivencia. Las muestras se llevan al laboratorio donde se obtienen
extractos crudos celulares utilizando soluciones de lisis que contienen
preservantes de la actividad enzimática. Tales extractos se someten a
electroforesis en geles de almidón o acetato de celulosa de tal manera que
las proteínas citoplasmáticas se separan de acuerdo con sus cargas netas.
Las corridas electroforéticas deben ser muy cuidadosas para no dañar la
actividad enzimática. Posteriormente, se visualizan las posiciones de
migración de cada proteína al efectuar “in situ” la reacción enzimática
acoplada a un sistema de coloración. Para cada locus se cuentan
directamente los individuos heterocigotos y los homocigotos. Se calculan
las frecuencias genotípicas y alélicas y se emplean pruebas estadísticas
apropiadas para comparar las frecuencias de cada población bajo la
hipótesis enunciada en el problema.
11. La curva correspondiente es la B. El enunciado del problema indica que los
homocigotos recesivos van escaseando paulatinamente, por lo tanto se
puede asumir que el alelo favorecido por la selección es el dominante. El
alelo dominante aumenta rápidamente su frecuencia (exponencialmente) ya
que está en los homocigotos dominantes y en los heterocigotos. En
consecuencia el alelo recesivo presente en los homocigotos disminuye
rápidamente. El alelo recesivo también se encuentra en los individuos
heterocigotos, pero en estos casos aumenta su frecuencia, “arrastrado” por
la ventaja selectiva que tiene el alelo dominante. A medida que los
homocigotos recesivos disminuyen en la población la selección va
perdiendo fuerza porque se incrementa de manera relativa la presencia del
alelo recesivo en los heterocigotos, quienes manifiestan el fenotipo
dominante. En los heterocigotos el alelo recesivo no es detectado por la
selección; este siempre contribuirá a formar individuos homocigotos
recesivos en las generaciones siguientes en cantidades muy pequeñas,
siendo la selección sobre éstos cada vez más ineficaz.
 0
0