Mastitis y Células Somáticas: factores no nutricionales que alteran
Anuncio

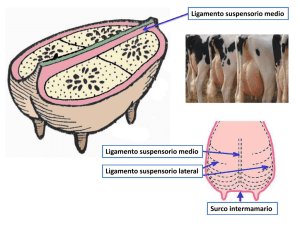
Mastitis y Células Somáticas: factores no nutricionales que alteran la composición láctea Patricia Cervantes Acosta, Antonio Hernández B., Diana P. Bonilla S., José Manuel Martínez H., Carlos Lamothe Z. Facultad de Medicina Veterinaria y Zootecnia, Universidad Veracruzana [email protected] Introducción Los alimentos de origen animal son importantes por tres razones principales. La primera es simplemente por su peso relativo en el sector de la economía global. Otra es porque el valor total de la producción mundial de carne, leche y huevos, la cría y captura de pescado supera 730 dólares millones de dólares anuales lo que se traduce en alrededor de $ 109 dólares por cada hombre, mujer y niño en la Tierra (FAOSTAT; www.census.gov). Además de que la demanda de leche, carne, huevos y pescado ha aumentado rápidamente en los países en desarrollo durante las últimas décadas, especialmente en economías de crecimiento rápido. Este crecimiento se prevé que siga durante las próximas décadas y por ello en el caso de la ganadería se permitió acuñar el término de la Revolución Ganadera (Delgado et al 1999). En particular y mediante la perspectiva científica, las propiedades físico-químicas de la leche tienen gran importancia desde el punto de vista de su interpretación: Como atributos de la capacidad nutricional del alimento, parte sustancial de los indicadores de la calidad de la materia prima y elemento decisivo en el proceso de elaboración de productos lácteos de calidad (Hernández, 2004). Pero, aún con los grandes avances en la selección genética, sistemas de alimentación, instalaciones y prácticas de ordeño, la mayor parte de rebaños de alta producción tienen un aumento significativo de un grupo de enfermedades multifactorial enfermedades conocidas como "Enfermedades de la producción". Entre estas enfermedades, la mastitis es la de mayor importancia económica debido a que lleva a pérdidas drásticas de leche, prematura eliminación selectiva de vacas genéticamente superiores, precio de fármacos, servicios veterinarios, el aumento personal para mano de obra, retención de la leche post-tratamiento, mejora genética reducida, cambios en la higiene y calidad de la composición de la leche, deterioro de las propiedades tecnológicas de la leche y disminución del desempeño reproductivo (Batavani et al., 2007). Se consideran varios los factores que contribuyen a variaciones en la composición de la leche y por lo tanto la calidad del producto, estas incluyen la etapa de lactancia de la vaca, raza, manejo de la nutrición, factores estacionales y cambios patológicos asociados con mastitis. Este documento se relacionará directamente con el problema de la mastitis. Composición de la leche y desarrollo humano Desde el punto de vista nutricional, la leche es uno de los alimentos más completos que existe en la naturaleza, tanto por el equilibrio y diversidad entre sus diferentes componentes, como por la alta digestibilidad y absorción de los mismos. Está compuesta de agua, carbohidratos (lactosa), grasa, proteína, minerales y vitaminas y otros componentes menores. Aunque cada componente se puede analizar por separado, es importante señalar que su secreción se lleva a cabo como una mezcla compleja de dichos componentes, ya que a su vez es una emulsión de glóbulos grasos, una suspensión de micelas de caseína (caseína, calcio, fósforo) y una solución o fase acuosa que contiene las sustancias solubles lactosa, proteínas del suero y minerales. Se agrega la presencia de gases y de células somáticas como los leucocitos y restos de células del epitelio mamario. Es por ello que se considera que sus propiedades e importancia son más considerables y complejas que cada uno de sus componentes individuales, (Chung y Norman, 1999). Estableciendo una visión solo comercial del problema, la lechería es un caso típico para aplicar los conceptos sobre las ventajas preferenciales de una región o país con relación a otro, es decir que produzcan los que tienen condiciones para ello y que compren los excedentes los que no alcanzan dicho estatus, un enfoque recurrente para el bloque de los países y regiones exportadoras. Sin embargo, el sector lechero constituye una importante fuente de trabajo y de subsistencia para millones de personas en todo el planeta con implicaciones sociales, económicas y ambientales y prueba de ello es que ha sido el área más sensible en el conjunto de acuerdos y regulaciones internacionales de los últimos 40 años, desde la OMC al ALCA, (IFPRI, 2003). Obtención de leche en los sistemas tropicales. La industria láctea está soportada principalmente en el uso del pastoreo estacional. Práctica que fomenta un suministro irregular de leche a los procesadores en términos de volumen y calidad. Los fabricantes de productos lácteos que acopian a partir de sistemas de lechería basados en pasturas, tienen el desafío de una demanda creciente de derivados lácteos de alta calidad y la leche que adquieren frecuentemente no es consistente en volumen ni en las características de calidad para su procesamiento de acuerdo a las temporadas climáticas (Petrovski y Kiro, 2006). Es importante tener en cuenta que la vaca lechera y los rumiantes en general, tienen un metabolismo integral que se inicia con la rumia, pasa por el decisivo papel del rumen en la transformación de los forrajes en sustancias asimilables por el organismo, continúa con la degradación de la proteína microbiana y otras sustancias en el intestino, se adicionan las características peculiares del metabolismo hepático en relación a la síntesis de glucosa y su papel en la síntesis de la leche, y otros eventos bioquímicos y finaliza con la alta complejidad de la síntesis y formación de los componentes lácteos a nivel de la glándula mamaria. La distribución preferencial de los nutrientes hacia la ubre durante el proceso de la lactación, conocido como homeoresis, no parece ser igual en un animal rústico o adaptado a las condiciones del trópico, que en una vaca especializada, genéticamente preparada para una alta producción (Kenelly, 1999). La elevada especialización productiva obtenida en diferentes razas de bovinos y caprinos es un ejemplo del trabajo genético y otros factores, dirigidos por el hombre hacia fines productivos, pero que induce a su vez problemas de comportamientos biológicos anormales como el estrés y enfermedades asociadas. La estrategia del desarrollo lechero en las condiciones del trópico debe tener en cuenta estos elementos en la búsqueda de genotipos y sistemas de manejo, que logren integrar los mejores indicadores bioproductivos y no ir solo a la búsqueda de altos rendimientos lecheros (Brian y Spencer, 2000). ¿Qué es la mastitis? La mastitis es una inflamación de la glándula mamaria causada comúnmente por microorganismos causantes de mastitis y rara vez por trauma físico o químico, se caracteriza por cambios patológicos en el tejido mamario; aumento en el número de células somáticas, cambios físicos, químicos y microbiológicos de la leche. Se sabe que en el ganado lechero, la mastitis clínica y subclínica afecta la composición y las propiedades de la fabricación de derivados lácteos. Es así que las infecciones bacterianas son por mucho la causa más común de mastitis en ganado lechero y se sabe mucho acerca del efecto de estas infecciones bacterianas intramamarias sobre la composición de la leche, pero ¿Cómo se producen estos cambios? Es un aspecto que se entiende un poco menos (Petrovski y Kiro, 2006). La mastitis de tipo subclínico particularmente es una de las condiciones de enfermedad mas persistente y ampliamente considerada en todo el mundo para el control de higiene y calidad de la leche de vaca, ya que influye en la producción total de leche y modifica la composición y su utilidad tecnológica (Ogola et al., 2007). En la vaca el conteo de células somáticas (CCS) se utiliza como un buen predictor de mastitis subclínica, ya que esta variable es un componente importante en términos de calidad, higiene y control de mastitis, un conteo elevado en leche cruda siempre se asocia a alteraciones en la calidad de las proteínas, cambios en la composición delos ácidos grasos, lactosa concentraciones de iones y minerales, incremento en la actividad enzimática y elevación del pH (Rajèeviè et al., 2003) Glándula mamaria y los cambios en la composición de la leche Linzell y Peaker (1971) señalan que en la granja la composición de la leche en el tanque es notablemente constante, especialmente en lactosa, componente que generalmente se supone no varía su concentración dentro de condiciones fisiológicas y desempeña un papel decisivo en la regulación del volumen de secreción. Así que el efecto en la composición constante de leche en tanque, se debe a un efecto de dilución de la leche a granel sobre la leche afectada. Para un mejor entendimiento de como ocurren los cambios en la composición de la leche se requieren conocimientos básicos de la morfología y fisiología de la glándula mamaria. Generalidades de la morfología de la glándula mamaria La glándula mamaria de la vaca consiste de cuatro glándulas separadas, localizadas en la región ventral o inferior de la vaca. Cada glándula tiene una teta y cada teta un orificio. La piel de las glándulas está cubierta de pelo a excepción de las tetas. Las mitades derecha e izquierda están separadas completamente, externamente esto esta indicado por un surco que se observa en la región inferior de la glándula. Los cuartos traseros, aportan del 55 al 60% de la leche producida, y son el 55 al 60% del peso de la glándula y son usualmente más cortas que los cuartos delanteros. Las tetas o papilas mamarias. La función de las tetas, es únicamente para la secreción y único medio del becerro para recibir la leche. Usualmente sólo una teta drena a una glándula. Las tetas de la vaca, usualmente no tienen pelo, glándulas sebáceas o sudoríparas. El tamaño y forma es independiente del tamaño y forma o producción de leche de la glándula. El promedio de tamaño para las tetas anteriores es de cerca de 6.6. cm. de largo y 2.9 cm. de diámetro, y para las tetas posteriores es de 5.2 cm. de largo y 2.6 cm. de diámetro. Tetas supernumerarias. Alrededor del 50% de las vacas tienen tetas extras, y se llaman supernumerarias. Algunas de estas tetas extras, abren hacia una glándula “normal”, pero muchas de ellas no. Generalmente son removidas en el primer año de edad. Una pseudoteta no tiene luz hacia el canal, y además no se conectan a la estructura interna de la glándula. Conducto del canal (ducto papilar). Funciona como el único orificio de la glándula entre el sistema secretor interno de la leche y el medio ambiente. Este canal es la principal barrera contra las infecciones. Esta cubierto con una epidermis parecida a la piel. Este canal se cierra por músculos del esfínter situados alrededor del canal. Con el incremento en número de lactaciones, el canal disminuye la potencia y se alarga la longitud del conducto. Roseta de Furstenburgs. Es una capa mucosa del conducto que cubre la porción final del conducto. Puede cubrir hasta la apertura del canal debido a que la presión dentro de la glándula es máxima. Este puede ser el punto de mayor entrada de leucocitos hacia la cubierta de la teta y entrar hacia la cisterna de la teta. Anillo cricoideo (Pliegue anular). Es la región en el final próximo de la cisterna de la teta, esta marca él límite entre la cisterna de la teta y la cisterna de la glándula. Cisterna de la teta (Seno papilar). Es la cavidad dentro de la teta, es continua a la cisterna de la glándula. Esta cisterna está cubierta con numerosas capas circulares y longitudinales en la mucosa, que forman paquetes en la pared interna de la teta. Cisterna de la glándula - (Seno lactóforo), también llamado cisterna de la ubre. Se abre directamente hacia la cisterna de la teta. Ocasionalmente se forma un septum entre la teta, la cisterna de la glándula y el cuarto puede ser ciego y puede ser corregido quirúrgicamente. La función de la cisterna es de almacenamiento de leche (sostiene ~ 100 – 400 ml). La cisterna de la glándula varía en tamaño y forma. Tiene forma de bolsa en la cisterna y al final del conducto de la glándula. Tejidos de la glándula Internamente la glándula mamaria esta compuesta por una entramado de conductos formados de células epiteliales que finalizan en un gran cantidad de racimos lóbulo-alveolares sitios donde ocurre la secreción láctea. Los tejidos que se encuentran en el interior de la glándula son: Tejido conjuntivo – tejido fibroso y graso Tejido secretor – células epiteliales secretoras o células productoras de leche. La cantidad de estos tejidos varía de animal a animal. El tejido secretor esta organizado en lóbulos, formados a su vez de muchos lobulillos. Cada lóbulo contiene alrededor de 150,220 alvéolos microscópicos. Alvéolos – (acinis) Son estructuras en forma de saco, y allí tiene lugar la síntesis y secreción láctea. Cada alveolo es una discreta unidad productora de leche. Se estima que tiene espacio para ~ 1/5 de una gota de leche, el lumen del alveolo está revestido por una sola capa de células epiteliales secretoras. Cada alvéolo esta rodeado de células mioepiteliales que se contraen en respuesta a la hormona oxitocina, como resultado se expulsa la leche fuera del lumen alveolar hacia los pequeños ductos. Por fuera, las células mioepiteliales están rodeadas de una membrana basal de tejido conectivo. El lecho capilar por fuera del alveolo es parte de tejido estromal entre el alvéolo. Un pequeño grupo de alvéolos, puede visualizarse como un racimo de uvas, con el tallo actuando como un pequeño conducto que llega desde el alveolo. Una capa simple de células epiteliales secretoras polarizadas rodea cada alvéolo dentro de estos racimos. Cada alvéolo se encuentra revestido por una capa de células secretoras que producen los componentes de la leche y los liberan dentro del lumen de los alvéolos. La cantidad de leche producida es proporcional al número de células secretoras presentes en la glándula y por lo tanto, al área desde la cual la leche puede liberarse dentro del sistema de conductos. Estas células se encuentran altamente especializadas para la producción de los componentes de la leche. Ciertos genes, que son específicos para las características de producción de leche, se expresan solamente (son funcionales) en estas células. A medida que las células secretoras liberan sólidos adicionales dentro del lumen alveolar, el agua pasa dentro de los alvéolos para mantener la correcta concentración o presión osmótica. Luego del secado, las células secretoras sufren un proceso llamado "involución" en el que disminuyen de tamaño y retornan al estado de descanso esperando el estímulo del próximo parto. Sistema de conductos. Los ductos son los túbulos por donde se drena la leche desde los alvéolos hasta la cisterna de la leche. Se organizan en: Ductos primarios o interlobulares, drenan hacia el lóbulo. Están revestidos con dos capas de células no secretoras y tienen muchas células mioepiteliales. Ductos intralobulares, están dentro de cada lóbulo y drenan a varias regiones del lóbulo. Ductos Interlobulares o secundarios, drenan a múltiples lóbulos. Están revestidos con una capa de células secretoras y rodeados de células mioepiteliales, participan eyección de la leche inducida por oxitocina. Ductos intralobulillares, son pequeños ductos dentro de un lóbulo intercalado o terciario, tiene salida hacia el alveolo. Esta clasificación sólo sirve para proveer información de cómo están organizados los ductos, sin que se considere uniforme el sistema de ramificación de los ductos. La glándula mamaria posee por supuesto una compleja red vascular, arterial, nerviosa, linfática y los receptores para hormonas de la lactancia, que permiten el funcionamiento correcto de la glándula tanto para la lactogenésis como la lactopoyésis. Síntesis de los componentes lácteos. Metabolismo de la glándula mamaria. Cambios relacionados con la lactación. Una característica sorprendente de la adaptación metabólica relacionada con el inicio de la lactación, es el aumento considerable de la ingestión de agua y alimentos, acompañado de hipertrofia del conducto intestinal para permitir una absorción más rápida de nutrientes. También existe hipertrofia de la glándula mamaria, hígado y corazón. Son numerosos tejidos que para cubrir las necesidades metabólicas de la lactación intervienen en la absorción y movilización de nutrientes. Es así que los requisitos de los tejidos periféricos se reducen con el fin de asegurar una disponibilidad adecuada de nutrientes para la síntesis de leche. Este equilibrio metabólico entre la glándula mamaria y los nutrientes del organismo está regulado principalmente por el sistema nervioso central, mediante hormonas, neuropétidos y neurotransmisores (Hurley, 2002). Estructura y función de las células mioepiteliales A nivel ultraestructural las células epiteliales mamarias en lactación están muy diferenciadas y son fáciles de distinguir de las células inactivas. Los precursores en la sangre entran a las células a través de las membranas basales y laterales y la leche se descarga por la membrana apical hacia el lumen. Las células epiteliales mamarias son típicas células secretoras que tienen un retículo endoplásmico (RE) y un aparato de Golgi bien desarrollados. Las proteínas de exportación sintetizadas en el retículo endoplásmico rugoso (RER), se modifican en el aparato de Golgi, donde se añade también las mayoría de los componentes no grasos de la leche. Se originan vesículas secretoras en el aparato de Golgi para llevar la composición de la leche a la superficie de la célula. La glucólisis, las síntesis de ácidos grasos y la activación de aminoácidos se llevan a cabo en el citosol. La transferencia de energía de los sustratos oxidables al adenosintrifosfato ocurre en las mitocondrias. El citrato y los compuestos utilizados en la síntesis de aminoácidos no esenciales también se sintetizan en las mitocondrias. Los lisosomas contienen enzimas hidrolíticas y tienen una función importante en la destrucción celular durante la involución, al final de la lactación (Hurley, 2002). Síntesis y secreción de los componentes lácteos. 1. Agua. El contenido de agua depende de la síntesis de lactosa. Sin ella, la leche sería una secreción viscosa compuesta mayormente por lípidos y proteínas y extremadamente difícil extraerla de la glándula. Después del nacimiento, los mamíferos neonatos no son capaces de valerse por sí mismos, y sin el aporte del agua de la leche se deshidratarían rápidamente. El contenido varía en las distintas especies, siendo baja en mamíferos marinos y muy alta en el humano y otras, en la vaca es cerca del 87%. 2. Grasa. La grasa láctea está formada de una compleja mezcla de lípidos: Son fundamentales los triglicéridos que están compuestos de tres ácidos grasos en unión covalente con una molécula de glicerol por uniones ester y constituyen el 98% de la misma. En el caso del bovino, los triglicéridos contienen ácidos grasos de cadena corta (C4-C10), intermedia (C12-C16) y larga (Cn>16). La síntesis se origina por dos vías: A partir de los ácidos grasos de las lipoproteínas de la sangre, que son transportadas hasta el tejido mamario donde son liberados e integrados esencialmente como triglicéridos, (Hurley, 2002), mientras que los de cadena corta se sintetizan en la glándula mamaria por síntesis de novo, a partir del acetato y beta-hidroxibutirato, obtenidos de la fermentación ruminal y los ácidos de longitud de cadena intermedia llegan por ambos caminos, (AFRC, 1998). La secreción se inicia en el retículo endoplásmico rugoso con la agregación de los pequeños glóbulos grasos, los cuales están primariamente compuestos de un glóbulo de triglicéridos rodeados por una membrana bicapa de lípido, semejante a la membrana apical de las células epiteliales, (Moore y Christie, 1979), que ayuda a estabilizar los glóbulos grasos en emulsión dentro del medioambiente acuoso de la leche. El incremento de tamaño y las características eléctricas hacen que dicho glóbulo emigre a la pared apical de la célula epitelial donde se funde con esta y es expulsado al exterior en un proceso de tipo apócrino. Muchos de estos glóbulos grasos acarrean algunas de las proteínas lácteas, por lo que la crema también contiene una pequeña cantidad de proteína en adición a los componentes de la grasa de la leche; estas proteínas contribuyen a las características de textura de la crema. 3. Proteínas. Las proteínas totales de la leche están compuestas de numerosas proteínas específicas. El grupo primario de estas son las caseínas, que son moléculas distintas pero con una estructura similar en forma de micelas. En la vaca la calidad de estas proteínas es la principal razón por la que se considera un alimento importante para la alimentación humana, (Hurley, 2002). Las moléculas de caseína contienen agua y sales en forma de fosfocaseinato de calcio. Por la habilidad para separarse fácilmente otros componentes de la leche, la estructura micelar de la caseína, es una parte importante para la digestión en el estómago e intestino del lactante y la base para muchos productos lácteos en la industria (tal como la industria del queso), (Jensen 1995). Con respecto a las proteínas séricas, estas varían en cada especie, periodo de lactancia, infecciones intramamarias y otros factores. Las mayores proteínas séricas son la alfalactoglobulina y la -lactoalbúmina, esta última muy importante en la síntesis de lactosa y su presencia es central en el proceso de síntesis de la leche. Otras proteínas séricas son las inmunoglobulinas (especialmente altas en el calostro) y la albúmina sérica o seroproteína. En las proteínas séricas se incluye un gran número de enzimas, hormonas, factores de crecimiento, transportadores de nutrientes, factores de resistencia a enfermedades y otros (Eigel et al., 1984). Tabla 1. Clasificación y características de las principales proteínas lácteas. Proteína Caseínas alfa-Caseína kapa- Caseína beta- Caseína gama- Caseína Proteínas Séricas alfa-Lactalbúmina beta-Lactoglobulina Albúmina sérica Lactoferrina Inmunoglobulinas: IgG1 IgG2 IgM IgA % Aproximado de proteína en leche descremada (mg/dl) ~80 45-55 8-15 25-35 3-7 ~20 2-5 7-12 0.7-1.3 0.2-0.8 % Aproximado de proteína en leche entera (mg/dl) 30 15 10 3.5 1.2 6-7 1.2 3.2 0.4 0.2 1-2 0.2-0.5 0.1-0.2 0.05-0.10 0.6 0.12 0.04 0.13 Punto Isoeléctrico Peso Molecular aproximado (kilo Daltons) 4.1 4.1 4.5 5.8-6.0 23 19 24 -- 5.1 5.3 4.7 7.8 14 18 68 80 ----- 160 160 ~1,000 ~400 Fuente: Jensen, 1995 La mayor parte de las proteínas incluyendo las caseínas, -lactoglobulina y la lactoalbúmina, se sintetizan en las células epiteliales de la glándula mamaria. Las inmunoglobulinas y la albúmina sérica son absorbidas desde la sangre, con excepción de algunas inmunoglobulinas que son sintetizadas por linfocitos que residen en el tejido mamario, y que son las que proveen a la glándula mamaria de inmunidad local (Rook, 1961). Para la síntesis proteica, los aminoácidos son absorbidos a través de la membrana basal de la célula por varios sistemas específicos de transporte de estos. Una vez dentro de la célula, los aminoácidos son unidos por enlaces covalentes a los polisomas del Retículo Endoplasmático Rugoso (RER) para formar caseína, -lactoglobulina, y -lactoalbúmina y proteínas unidas a la membrana, tales como las que se encargan del contacto célula-célula y de enzimas unidas a la membrana (Hurley 2002). Las diferentes caseínas se integran en el Aparato de Golgi desde moléculas de caseína simples, iones de calcio y de fósforo formando las micelas. Las proteínas que permanecen en la estructura de célula, se sintetizan en los propios ribosomas e incluyen enzimas, proteínas estructurales tales como queratina y otras proteínas celulares, (Mustafa, 2001). La tabla 1 muestra las diferencias fisicoquímicas de las principales proteínas lácteas. 4. Lactosa. Es el carbohidrato mayor de la leche en casi todas de las especies y prácticamente el único en la vaca, cabra y búfalo, es un disacárido compuesto de los monosacáridos D-glucosa y Dgalactosa unidos en una ligadura beta-1,4-glicosídico. Otros carbohidratos que se encuentran en la leche, pero a muy bajas concentraciones son la glucosa (cerca de 0.1 mM) y galactosa libre (cerca de 0.2 mM), también se incluyen amino azúcar, azúcar fosfato, oligosacáridos ácidos y neutros y azúcares nucleótidos. Algunos oligosacáridos complejos son importantes para mantener la microflora del intestino del neonato (tal como el factor bífido identificado en el humano). La lactosa juega el papel más importante en la síntesis y secreción de la leche, ya que por una parte es el principal componente osmótico y por tanto responsable de la formación de la fase acuosa de la misma, influyendo de forma decisiva en el volumen total. Por otra parte, más del 50% de la glucosa circulante en el organismo de los rumiantes en lactación se utiliza para la síntesis de lactosa, lo que hace a dicha especie metabolicamente dependiente de la síntesis de glucosa hepática y en última instancia de la síntesis de lactosa a nivel de la glándula mamaria, (Vilotte, 2002). Su síntesis se realiza en la célula epitelial mamaria a nivel del aparato de Golgi, por la enzima lactosa sintetasa en un complejo proceso formado por la interacción de la proteína galactosil transferasa que se encuentra unida a la membrana de Golgi y la proteína lactoalbúmina que fluye como parte del proceso de secreción láctea por dicho organelo, (Hurley, 2002). Una vez que la lactosa se sintetiza a nivel de Golgi, se produce un flujo de agua por ósmosis para diluir la misma y mantener un equilibrio isotónico entre la leche y el plasma sanguíneo, dada la gran proporción de agua en la leche, su secreción viene determinada por la formación de lactosa, junto con otros iones, como el potasio, cloruro y sodio (Wattiaux y Armentano, 2002). Las vesículas de Golgi se funden con la membrana apical y vierten su contenido al lumen alveolar, en un proceso conocido como exocitosis o pinocitosis reversa, que contribuye a la restitución continua de dicha membrana apical. 5. Minerales y otras sustancias. La mayor concentración de minerales encontrados en la leche son calcio y fósforo, requeridos en gran cantidad para el rápido crecimiento de huesos y desarrollo de tejidos blandos del neonato. Ambos minerales se asocian con la estructura de las micelas de caseína, y consecuentemente en el suero su contenido es relativamente menor que en leche entera, (Hurley, 2002). Tanto los macro como microelementos, no se sintetizan en el organismo, sino que pasan a la leche por difusión desde la sangre o como parte de otras moléculas más complejas. La leche también contiene diversas vitaminas liposolubles e hidrosolubles. Las más importantes del primer grupo son, la vitaminas A, D, E, y K, que se encuentran principalmente en la grasa, aunque en menor medida la vitamina K. Las principales vitaminas hidrosolubles son las que integran el complejo B, y la vitamina C, que se localizan en la fase acuosa. Existe un alto número de componentes y factores bioactivos tales como hormonas y factores de crecimiento, enzimas, proteínas celulares y otros, (Jensen, 1995). La presencia de células somáticas, principalmente diversos tipos de leucocitos o células de la defensa, tienen una gran importancia como expresión de la integridad del tejido epitelial y por su asociación con los procesos infecciosos como la mastitis y los cambios fisiológicos en el inicio y final de la lactación. Aunque en menor cuantía, también se localizan restos de células epiteliales. Factores asociados a la producción y composición láctea. Los resultados de la selección genética, apropiada nutrición y manejo adecuado de la lactancia son parte clave en la obtención de la eficiencia económica a las explotaciones lecheras, pero no siempre alta producción y alimentos concentrados significan la mejor opción económica ni fisiológica, (Stelwagen, 2001). 1. Factores genéticos: Raza y potencial productivo Las diferencias en la cantidad de leche y el contenido de grasa, lactosa y proteínas relacionadas a la raza de la vaca, estado de lactancia y número de lactancia ha sido bastante documentado en países de ganadería desarrollada. También está bien establecido el efecto del ambiente y manejo, la frecuencia y tipo de ordeño, y la presencia de enfermedades de la ubre (Dodd, 1984). 2. Salud de la ubre y trastornos asociados a la lactación en la vaca lechera. 2.1. Mastitis La causa más común para un sacrificio temprano de las vacas lecheras son los problemas de salud de la glándula mamaria, además de problemas de fertilidad. El 26.5% de las vacas lecheras sacrificadas en el continente americano se deben a trastornos ocasionados por la mastitis. El paso rápido de los leucocitos sanguíneos a la luz alveolar es de los mecanismos naturales más importantes de defensa contra la mastitis. En al caso de una glándula mamaria sana se puede observar un contenido menor de 100,000 leucocitos/mL en la leche. El contenido de leucocitos aumenta como una respuesta a los microorganismos invasores y otros efectos de la irritación local, por lo cual este criterio es el más común para evaluar la afectación de la ubre. La mastitis reduce la producción láctea entre un 5-25%, así como el contenido de grasa, caseína, lactosa y sólidos en general e incrementa la concentración de proteínas del suero, y los electrolitos sodio y cloro, por lo cual reduce la calidad y el valor industrial de la leche. 2.2. Síndrome de Leche Anormal En condiciones normales de lactación, la leche es isosmótica con el plasma sanguíneo, ello significa que la presión osmótica generada principalmente por la lactosa, y los electrolitos sodio, potasio, y cloruros se iguala con la sangre. Para mantener dicho osmolaridad, la disminución de uno o más componente implica cambios de los demás en sentido inverso. Generalmente, las concentraciones de lactosa y potasio disminuyen en la etapa calostral, etapa final de la lactación y también en presencia de un cuadro de mastitis, mientras el sodio y el cloruro lo hacen en sentido inverso. En condiciones fisiológicamente normales es común encontrar correlaciones entre dichos componentes Sin embargo este comportamiento no siempre se ajusta a lo descrito por la literatura científica, (Ponce y Bell, 1986; Hernández, 2002) Ponce (1996), reportó en Cuba la existencia de alteraciones variadas en la composición láctea, y en su aptitud para el tratamiento industrial, observándose con mayor frecuencia durante la época de seca y en zonas de ese país donde se localizan las razas lecheras más especializadas, fenómeno más recurrente cuando se incrementaron las limitaciones de alimentos en el periodo de seca. La denominación de Síndrome de Leche Anormal (SILA), establecida por Ponce y colaboradores en 1996, se asienta en el conjunto de resultados acumulados en Cuba durante más de 20 años, y establece que muchas de estas alteraciones se asocian a múltiples causas relacionadas con la alimentación y las mayores demandas nutricionales de las razas lecheras mas especializadas, y que se expresan en diversas alteraciones de la leche, (Ponce y Hernández, 2001; Hernández 2004). En la tabla 2 se clasifican los criterios para identificar un cuadro SILA. Tabla 2. Criterios establecidos para identificar un cuadro de SILA a partir de la leche Tipo de Alteración Indicador de Alarma Inestabilidad térmica Acidez titulable menor a 0.12 g%. Prueba del alcohol positiva. Depresión en el contenido de sólidos. Relación Proteína bruta/caseína menor del 75%. Generalmente alto contenido de urea y nnp. Grasa variable. pH mayor de 6.75 Alteración en sólidos Proteína bruta menor de 2.90g%. Caseína menor de 2.20g%. Lactosa menor de 4.6g%. Punto crioscópico menor de 420 mgc. Desequilibrio mineral Calcio normal, Fósforo menor de 80 mg%, Magnesio menor de 9 mg%, Potasio mayor de 150 mg% Fuente: Ponce, et al., (1999), Hernández, (2004). Patogénesis del cambio en la composición de la leche durante la mastitis Hay una serie de funciones que pueden ser interrumpidos durante la infección intramamaria y una serie de mecanismos que puede ser perturbado, el resultado es difícil de predecir y según Petrovski y Kiro (2006) depende de lo siguiente: 1) la gravedad de la infección, el efecto varía desde muy poco hasta la inhibición completa de la secreción de leche y depende del organismo que cause la mastitis, su virulencia y resistencia del huésped. 2) Severidad de la infección, que puede ser localizada en algunos alvéolos o abarcar a todos los alvéolos. La cuantificación de este efecto es difícil porque en la mayoría de los casos, sólo una glándula está infectada; por lo tanto, el efecto se diluye cuando las mediciones se realizan sobre una base de toda la glándula. Hay una considerable variabilidad que depende de las características individuales de cada animal, raza (en el caso de razas mixtas en el mismo rebaño), edad y etapa de lactancia. 3) Alteración de la actividad metabólica de las células productoras de leche, aspecto que lleva a la reducción de la síntesis de leche y la interferencia con el balance iónico, hasta la reducción de la concentración de las hormonas galactopoyéticas o por el aumento en la concentración de alguna hormona inhibitoria y/ó un mediador inflamatorio. 4) Interferencia con la disponibilidad de precursores para la síntesis de leche debido a: anorexia, disminución del flujo sanguíneo en la glándula mamaria o desequilibrio hormonal. 5) Interrupción de la integridad epitelial, abriendo la vía paracelular. 6) Descomposición de los componentes de la leche debido a los leucocitos y a las enzimas de los organismos que causan mastitis. Como parte del mecanismo de defensa de la vaca, a la nueva infección intramamaria sigue rápidamente una afluencia de leucocitos y el aumento de los recuentos de células somáticas en la leche. El aumento en la permeabilidad de las uniones intercelulares a través de las capas epiteliales y endoteliales se debe a productos de la reacción inflamatoria tales como histamina, TNF, IFN-g y las proteínas de fase aguda. Este aumento en la permeabilidad es una parte importante del proceso inflamatorio, ya que permite que componentes inmunes lleguen al sitio de infección. Benites et al. (2002) Establecieron que el aumento de la diapédesis paracelular de los leucocitos a través de las células epiteliales reduce la integridad de las uniones intercelulares y por lo tanto el intercambio de los componentes precursores entre la sangre y la leche a través de la vía paracelular. Los leucocitos predominantes presentes en la leche bajo tales circunstancias son neutrófilos polimorfonucleares (PMN). Ellos son responsables de los conteos altos de células somáticas (CCS) que es característico de leche colectada que se asocia a muchos de los cambios en la composición de la leche producida con mastitis. Además, algunas toxinas microbianas y enzimas de las células dañadas causan lesiones las células secretoras. Las concentraciones de N-acetil-b - D-glucosaminidase (NAG-ase) en la leche se incrementan con la aparición de mastitis, y un examen histológico de las glándulas colectadas reveló necrosis de células epiteliales. Tal daño celular puede producir "conductos" en el epitelio mamario que puede conducir a cambios en la composición de la leche y un cortocircuito en el potencial eléctrico de sangre-leche de la misma manera como apertura de uniones intercelulares estrechas. Por ejemplo, la lactosa, que es sintetizada exclusivamente por las células epiteliales mamarias, escapa parcialmente a la circulación sanguínea debido al daño de la barrera de sangreleche. Simultáneamente, se incrementan las concentraciones de los componentes proporcionados por la vía sanguínea en la leche de los cuartos afectados, tales como la albúmina sérica y iones de sodio (Na +) y cloro (Cl-). La filtración de a través de las uniones intracelualres estrechas causadas por mastitis se acompañan por una disminución en la tasa de síntesis de leche y la secreción de los componentes principales de la leche, mientras que la secreción de otras proteínas como la lactoferrina se regula simultáneamente. La concentración de las caseínas en cuartos infectados se reduce debido a una reducción de la secreción e incremento en la destrucción por proteasas proporcionadas por vía sanguínea en las proteínas de la leche y grasa, por ejemplo la plasmina. Algunos de los organismos causantes de mastitis se adhieren a la superficie de las células epiteliales y forman colonias. Estas áreas localizadas probablemente están más expuestas a concentraciones mayores de productos tóxicos que las no afectadas. Esto se debe las endotoxinas microbianas fallan al afectar directamente al tejido mamario (Shuster et al. 1991) concluyen que la disminución en la síntesis de leche durante la mastitis inducida por endotoxinas se debe probablemente a una respuesta patofisiológica de la vaca y que esta es inducida por mediadores inflamatorios, como los metabolitos de citoquinas y araquidonatos producidos por los leucocitos mamarias en interacción con las endotoxinas. En casos extremos, las células pueden resultar tan dañadas que se eliminan y entonces aparecen espacios en el epitelio. El proceso de reparación del tejido dañado de la glándula mamaria se logra por la proliferación de tejido fibroso, lo que se denomina fibrosis. La fibrosis es el comienzo de la formación de las cicatrices y puede iniciarse durante la respuesta inflamatoria o desarrollarse desde una dilatación quística, que es un tipo de reacción de reparación que forma quistes en los acinis dilatados de la glándula. El proceso de reparación reemplaza permanentemente al tejido glandular por tejido conectivo y en consecuencia esto conduce a la reducción en la producción de leche. A lo anterior se suma que restos de tejido, organismos causantes de mastitis, el fibrinógeno que llega hacia los espacios intersticiales y que se convierte en fibrina y los leucocitos forman coágulos que ocluyen los conductos de drenaje de las áreas afectadas. En este caso la secreción se acumula en el lumen y provoca involución local de la zona afectada y apertura de vías paracelulares. Es así que durante el proceso inflamatorio se implican tres mecanismos en el cambio de composición de leche: disminución en la síntesis, aumento de la permeabilidad de la barrera láctea y un aumento de la actividad proteolítico/enzimática en leche. De esta forma, la leche de una glándula infectada estará compuesta de secreción de áreas poco afectadas hacia otras que ya no pueden producir leche. En otras áreas, incluso el epitelio puede estar destruido y la complejidad de estos efectos significa que será muy difícil interpretar los cambios en la composición de la leche. Efectos de la mastitis sobre la producción de leche y composición. Efectos sobre la producción de leche. La infección intramamaria, aunque restringida a nivel subclínico, se ha reportado que influye negativamente en la producción de leche. Esta reducción es probable se deba en gran parte a daños físicos de las células epiteliales de la glándula afectada y una consecuente reducción en la capacidad de síntesis y secreción de la glándula en su conjunto. Por lo que cualquier falla de la capacidad de la glándula mamaria para sintetizar y secretar lactosa es de particular importancia dado el papel clave de lactosa como el regulador osmótico de volumen de leche. Sin embargo, no toda la represión de la producción de leche asociada con mastitis se debe atribuir al daño del epitelio mamario. Shuster et al., (1991) llegaron a la conclusión de que la hipogalactia en cuartos afectados se debe al efecto sistémico de la mastitis de cuartos afectados y puede ocurrir ya sea por la absorción sistémica de un inhibidor de la producción de leche de las áreas afectados que actua sobre los cuartos no afectados . Un número de mediadores inflamatorios, incluyendo citoquinas y metabolitos del ácido araquidónico; cambios en las concentraciones de hormonas estimulantes o inhibidores y disponibilidad reducida de precursores de la leche pueden desempeñar un papel importante. La producción de leche de los cuartos afectados es más evidente que en los no afectados, resultante de una inflamación localizada. Otras posibles explicaciones para la represión local de producción de leche incluyen el efecto directo de los mediadores inflamatorios producidos localmente, leucocitosis y el edema mamario localizado. La situación es más compleja, como generalmente se acepta, que cuartos no infectados pueden incrementar la producción y compensar en parte la disminución de la producción por los cuartos infectados. Sin embargo, esta compensación puede ocurrir sólo después de que la infección se eliminada. Por otro lado, el aumento de la permeabilidad de la barrera de sangre-leche en los cuartos afectados conduce a una disminución en el volumen y la concentración de los componentes de la leche, pero no toda su disminución en la producción de leche se debe a reducción en la síntesis, algo de ello es el resultado de la fuga desde la glándula hacia la circulación. También se plantea la hipótesis de que parte de la disminución en la producción se debe a la creciente demanda de energía del sistema inmune para combatir la infección, disminución del apetito asociados a la inflamación y reducción de la ingesta debido al dolor y disminución del movimiento. Efectos sobre la composición de la leche. Sobre el contenido de proteína. Es generalmente aceptado que durante la mastitis proteínas lácteas aumentan debido a la llegada de proteínas transmitidas por la sangre (como la albúmina, inmunoglobulinas, las proteínas menores del suero menores, transferirrinas, a-macroglobulina, asociada a una disminución de caseínas. Según Auldist et al. (1995) y Auldist y Hubble (1998) este aumento de proteínas séricas se puede atribuir a una interrupción en la integridad de los epitelios mamarias causada por las toxinas microbianas y apertura de las uniones intercelulares y que la disminución en la concentración de la caseína durante la mastitis es en gran parte debido a la degradación postsecretoria de la caseína por las proteasas procedentes de organismos que causan mastitis, leucocitos o la sangre y en parte a una reducción en la síntesis y secreción de caseína como consecuencia del daño físico en las células epiteliales mamarias por las toxinas microbianas. Esto tiene implicaciones importantes para el potencial de transformación industrial de la leche, particularmente, pero no exclusivamente para la fabricación de queso. Por otro lado, las proteínas de suero de leche sintetizados de novo son relativamente resistentes al ataque proteolítico. Sin embargo, en leche con alto CCS es una evidente la disminución de -lactoalbúmina y -lactoglobulina. Esto sería, en parte debido al deterioro de la función de síntesis y secreción celular y en parte a la fuga de estas proteínas fuera de la leche y hacia el fluido extra-celular a través de las vías de paracelular que proliferan durante la mastitis, ya que se han reportado concentraciones elevadas de -lactoalbúmina en la sangre de vacas con elevados CCS. El aumento en las concentraciones de lactoferrina durante la infección intramamaria, posiblemente relacionado con la función inmune de esta proteína. Efectos sobre el contenido de grasa. El efecto de la mastitis sobre las características de la grasa de la leche no ha sido tan ampliamente estudiado como las proteínas de la leche. Según Bruckmaier et al. (2004) el aumento en la concentración de grasa indica que hay una reducción en la síntesis de lactosa y en el volumen de leche con una síntesis de grasa deprimida ligeramente, ó que la reducción sólo se debe a un menor volumen de leche. La fuga de la lactosa de la leche se llevará consigo agua y el volumen de la secreción se reduce en la glándula disminuirá. Las gotas de grasas sin embargo son relativamente grandes en relación con los espacios intercelulares, se mantienen dentro del alvéolo y en consecuencia aumenta su concentración. La membrana del glóbulo de grasa de la leche es susceptible a la acción de la enzima lipasa, producida por los leucocitos que invaden la glándula mamaria en respuesta a la infección, resultando en la ruptura de los triglicéridos, oxidación de ácidos grasos y perdida del sabor. Se asume que la leche con alto CCS es más susceptible a la lipólisis espontánea. Una probable explicación de los factores que aumentan la hidrólisis de los triacilgliceroles en la gota de grasa durante la mastitis, es que este proceso se acentúa por la adición de componentes de suero de la sangre (iones Na + y Cl-) a la leche durante la mastitis. Efectos sobre el contenido de lactosa. Es aceptado que la mastitis provoca una disminución en la concentración de lactosa de leche y qu esto se debe al daño a las células epiteliales alveolares. Por su papel clave de lactosa como regulador osmótico del volumen de leche, su disminución es poco probable que sea a causa de la reducción de la síntesis y secreción a nivel celular y sólo podría estar deprimida, debido a que se incrementa la afluencia de electrolitos durante la mastitis, esto es porque el agua podría entrar en las células sólo en cantidades suficientes para mantener el equilibrio osmótico. La razón más probable para la depresión de lactosa es la fuga de la lactosa de la leche a través de las vías de paracelular que proliferan durante la mastitis. Por lo tanto, las concentraciones de lactosa baja dependen de la severidad del daño a las uniones intercelulares estrechas. Ya que la lactosa es sintetizado en la glándula mamaria y no secretada en cantidades significativas a través de la superficie de baso-lateral del epitelio mamario, ni metabolizada en otras partes del cuerpo, el nivel de lactosa plasmática prove una medida de la tasa de fuga de material desde el lumen de la glándula mamaria en la corriente sanguínea. La lactosa también puede deprimirse si se impide el mecanismo de regulación de la concentración de los iones principales, potasio, sodio, cloruro y bicarbonato, lo que aumentará la concentración de iones por la necesidad de mantener el equilibrio osmótico disminuyendo la concentración de lactosa. Además, según Auldist et al. (1995), muchos de los organismos que causan mastitis comunes son capaces de fermentar la lactosa y una baja concentración de lactosa en leche pueden deberse en parte a las actividades de estos organismos. Efectos sobre el contenido de minerales. El contenido iónico de leche varía marcadamente del fluido extracelular con el de dentro de los grupos de acinis de la glándula mamaria. La concentración de potasio es relativamente más alta que la de sodio leche contiene una alta concentración de potasio en relación con el sodio. La concentración de muchos minerales se altera durante la mastitis, y estos cambios pueden desempeñar un papel importante en la determinación de la calidad de la fabricación de derivados lácteos y el diagnóstico de mastitis subclínica. El potasio, que es el mineral más abundante en la leche, se fuga a través de la vía paracelular y disminuye su concentración, mientras que el sodio que en la sangre está en cantidades elevadas, se libera hacia la leche hasta concentraciones por encima de lo normal. Las concentraciones de cloruros en la leche de vacas con mastitis subclínica se elevan probablemente por el flujo de componentes sanguíneas de la sangre a la leche durante la infección. Contaminación bacteriana coincidente. La contaminación bacteriana puede provocar alteraciones en la composición de la leche por la acción de enzimas hidrolíticas liberadas por las bacterias. La contaminación bacteriana de la leche a granel con mastitis subclínica contribuye a un recuento de bacterias totales, pero normalmente no excede las 10.000 UFC/mL. Una contaminación bacteriana más significativa de la leche puede ocurrir cuando la leche de vacas con mastitis clínica se admite en el acopio a granel. Bibliografía consultada: 1. AFRC 1998. Technical Committee on Responses to Nutrients. Response in the yield of Milk Constituents to the Intake of Nutrient by Dairy Cows. Edit. Acribia. España. 2. Batavani, R. A., Asri, S. y Naebzadeh, H. 2007. The effect of subclinical mastitis on milk composition in dairy cows. Iranian Journal of Veterinary Research, Vol. 8, No. 3, Ser. No. 20. 3. Bruckmaier R.M., O., C.E., Blum, J.W., Fractionized milk composition in dairy cows with subclinical mastitis. Veterinary medicine-Czech, 2004. 8(283-290). 4. Chung P.S. Norman L. J. 1999. Glándula mamaria y lactación. Fisiología de los animales domésticos de Dukes, de Swenson M.J., Reece W.O. Cáp. VI. Endocrinología, reproducción y lactación. Pág. 711. LIMUSA. México. 5. Delgado, C., Rosegrant, M., Steinfeld, H., Ehui, S., and Courbois, C. (1999) “Livestock to 2020 – The 6. Next Food Revolution”. Food, Agriculture and the Environment Discussion Paper 28. IFPRI/FAO/ILRI. 7. Dodd F.H. 1984. Herd management effects on compositional quality. In: Castle, M.E. y Gunn, R.G. (Eds) Milk Compositional Quality and its Importance in Future Markets. British Society of Animal Production Occasional Publication No 9, pp. 77–83. 8. Eigel W.N, Butler J. E, Ernstrom C.A, Farrel H.M. Jr, Harkalkar V.R, Jennes R, Whitney R.M. 1984. Nomenclature of proteins of cow’s milk: fifth edition. J Dairy Sci, 67: 1599-1631 9. FAOSTAT, 2009, 2010. FAO (Food and Agricultural Organization of the United Nations). Rome, Italy. http://faostat.fao.org. 10. Hernández R. 2004. Caracterización, diagnóstico y corrección de alteraciones en las características fisicoquímicas de la leche en rebaños Holstein-Friesian. Tesis Doctoral. CENSA. Cuba. 11. Hurley W.L. 2002. Topics Areas in Lactation Biology. Lessons of Department of Animal Sciences. University of Illinois. Urbana-Champaign. 12. Jensen RG. 1995. Handbook of Milk Composition, Academic Press, San Diego. 13. Kennelly J.J., Glimm, D.R y Ozimek, L. 1999. Milk composition in the cow. Proceedings of the 1999 Cornell Nutrition Conference for Feed Manufacturers. Rochester, N.Y. 14. Linzell, J., energéticas, M, mecanismos de secreción de leche. Opiniones fisiológicas, 1971. 3 51: p. 564-597. 15. Mustafa A.F. 2001. Lactation Biology. Lectures of Courses on Line. Department of Animal Science. Mc Donald of McGill University, Ca. 16. Ogola H, Shitandi A y Nanua J. 2007. Effect of mastitis on raw milk compositional quality. J Vet Sci. 8(3):237-242. 17. Petrovski R. y Kiro, Stefanov Emanuel. 2006. Milk composition changes during mastitis. http://www.milkproduction.com/Library/?q=hidden:meta:category:Animal_health;Udder_health 18. Ponce P. y Bell L. 1984. Estudio de la lactación en vacas del cruce 3/4 H x ¼ C en Cuba. Rev. Cub. Cienc. Vet. 15:305-312 19. Ponce P. y Hernández R. 2001: Propiedades físico–químicas do leite e sua associacao com transtornos metabólicos e alteracoes na glandula mamária. In Uso do leite para monitorar a nutricao e o metabolismo de vacas leiteiras. Gráfica da Universidade Federal do Rio Grande do Sul, Porto Alegre. 20. Rajèeviè Marija, Potoènik Klemen y Levstek Joze. 2003. Correlations Between Somatic Cells Count and Milk Composition with Regard to the Season. Agriculturae Conspectus Scientificus, Vol. 68 No. 3 (221-226) 21. Rook A.J.F. 1961 Variations in the chemical composition of the milk of the cow. Dairy Science Abstracts 23, 251–258, 303–308 22. Shuster, D.E., et al., Suppression of Milk Production During Endotoxin-Induced Mastitis. J. Dairy Sci., 1991. 74(11): p. 3763-3774. 23. Stelwagen K. 2001. Effect of milking frequency on mammary functioning and shape of the lactation curve. JDS E. Suppl.: E204-E211. 24. Vilotte J.L. 2002. Lowering the milk lactose contents in vivo: potential interests, strategies and physiological consequences. Reprod. Nutr. Dev. 42, 127 132 25. Wattiaux M. y Armentano L. 2002. Esenciales lecheras. Instituto Babcock para la investigación y desarrollo internacional de la industria lechera. Universidad de Wisconsin – Madison http://babcock.cals.wisc.edu/spanish/default_spn.html