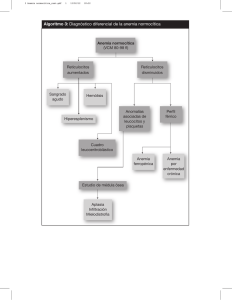
Farmacología del Hierro
Anuncio

Farmacología del Hierro Hoover Canaval, MD Hernán Pérez, DDS Diego Rincón, MD Jorge Vargas, MD Aprobado y Recomendado por el Anemia Working Group Latin America (AWGLA) y la Asociación Latinoamericana de Farmacología (ALF) Hierro Hoover O. Canaval Erazo, MD Profesor Auxiliar - Departamento de Ginecología y Obstetricia Facultad de Salud – Universidad del Valle. Gerente de la Unidad Estratégica de Servicios de Ginecología y Obstetricia - Hospital Universitario del Valle “Evaristo García” E.S.E. Cali – Colombia. Presidente del Anemia Working Group Latin America. Hernán Pérez Torres, DDS, Farmacólogo Profesor Facultad de Medicina Universidad Nacional de Colombia Profesor Facultad de Medicina Universidad «El Bosque» Diego Rincón Castillo, MD, MSc Farmacología Profesor Auxiliar - Seccion de Farmacologia Departamento de Ciencias Basicas Facultad de Salud Universidad Industrial de Santander Bucaramanga, Colombia Jorge Vargas, MD, Farmacólogo Ex-profesor de Farmacología Universidades Militar «Nueva Granada», Javeriana y Nacional de Colombia Asesor en farmacología para industria farmacéutica 1 Hierro 2 Hierro 1. Liberación del hierro Jorge Vargas 7 2. Absorción del hierro Jorge Vargas 22 3. Distribución del hierro Jorge Vargas 35 4. Metabolismo del hierro Hernán Pérez 43 5. Hierro y función cerebral Diego Rincón Castillo 69 6. Síndrome anemia cardio-renal Diego Rincón Castillo 87 7. Anemia y embarazo Hoover O. Canaval 99 8. Ferropenia, desarrollo y crecimiento en la infancia Jorge Vargas 114 9. Profilaxis de la anemia ferropénica Jorge Vargas 120 10. Hierro y estrés oxidativo Diego Rincón Castillo 127 3 Hierro 4 Hierro La deficiencia subclínica de hierro y la expresión clínica franca de la anemia ferropénica son de amplia prevalencia en Latinoamérica. Al revisar la literatura, es evidente la gravedad de la deficiencia de hierro, por ejemplo, su nefasto impacto sobre el desarrollo cerebral. Por estas razones, la anemia por deficiencia de hierro dejó de ser una situación en la cual solamente se busca alcanzar rangos normales en los parámetros clínicos y de laboratorio. Se han dado valiosos pasos en la conformación de equipos de trabajo para combatir la anemia por deficiencia de hierro. Entre ellos tenemos la Organización Panamericana de la Salud (OPS) y el Anemia Working Group Latin America (AWGLA). La realización de la presente obra contó con la notable participación de Hoover Canaval, actual presidente para Colombia y Latinoamérica del AWGLA. Igualmente, se han sumado en su realización, Hernán Pérez, Diego Rincón y Jorge Vargas, miembros de la Asociación Latinoamericana de Farmacología (ALF), quienes plasman su experiencia docente e investigativa. Este libro está destinado a fortalecer el entendimiento de las bases farmacológicas de la terapéutica con hierro y los avances en el desarrollo galénico, para lograr una óptima administración oral o parenteral. La difusión de estos conocimientos en nuestros países permitirá obtener el máximo beneficio en la lucha contra la anemia por deficiencia de hierro. Jorge Olarte, QF, PhD Profesor Emérito de la Universidad Nacional de Colombia Secretario de la ALF 5 Jorge Vargas E l hierro es un elemento químico de símbolo Fe cuyo número atómico es 26 y su peso atómico es 55.847. El hierro es el cuarto elemento más abundante en la corteza terrestre (5%), es un metal maleable, tenaz, de color plateado y magnético. 1 El hierro tiene una fuerte reactividad, esencial en muchos procesos metabólicos, pero también puede ser potencialmente nocivo. Por ejemplo, puede participar en varias reacciones capaces de producir radicales libres que pueden lesionar componentes celulares, especialmente los hierros bivalentes (Fe2+). El hierro es considerado como uno de los nutrientes más importantes, y muchas enfermedades están relacionadas ya sea con su deficiencia o sobrecarga. Aproximadamente 70% del hierro en el cuerpo está localizado en el hem o heme, participando en la función de la hemoglobina, mioglobina y el citocromo P450. El hierro propiamente dicho tiene muchas fun- ciones catalíticas a través de la acción conjunta hierro-azufre. Estudios recientes muestran que el hierro y/o heme juegan papeles significativos inclusive en la regulación genética. No solamente se ha identificado al hierro sino también al hem como reguladores importantes de la activación genética a través de la sensibilidad al oxígeno. Por ejemplo, el hierro controla la respuesta al oxígeno de la actividad del Factor 1 Inducible por Hipoxia (HIF-1), el cual es uno de los reguladores clave de la homeostasis del oxígeno. El HIF1 regula las respuestas fisiológicas a los niveles bajos de oxígeno (hipoxia) y la fisiopatología del ataque cardíaco, el cáncer, el accidente cerebrovascular y la neumopatía crónica. Se ha encontrado que las hemoproteínas en los procariotes tales como FixL, Dos, y HemAT son sensores de oxígeno. Investigadores japoneses informan el hallazgo de la Bach1, la cual parece ser la primera evidencia de regulación genética mediada por heme y oxígeno en nosotros, los vertebrados 2. Hierro Notables hallazgos como el mencionado, amplían nuestra concepción sobre la importancia biológica que adquiere un adecuado suministro de hierro. El hierro se encuentra en el medio ambiente principalmente como óxido o hidróxido férrico o como polímeros. En este estado, su disponibilidad biológica está limitada a menos que sea solubilizado por ácidos o quelatos. Por ejemplo, para satisfacer sus necesidades, las bacterias y algunas plantas elaboran agentes quelantes con alta afinidad que extraen el hierro del medio circundante. La mayoría de los mamíferos tiene poca dificultad para adquirir hierro; esta capacidad se explica por una ingesta más abundante de hierro y tal vez por una mayor eficiencia en su absorción. Sin embargo, el hombre es una excepción ya que, si bien la ingesta total de hierro con la dieta supera los requerimientos, su biodisponibilidad es limitada.3 1. Requerimientos de hierro Los requerimientos de hierro están determinados por las pérdidas fisiológicas obligadas y por las necesidades impuestas por el crecimiento. Así, el hombre adulto requiere sólo 13 µg/kg/día (alrede- 8 dor de 1 mg), mientras que una mujer en edad fértil, unos 21 µg/ kg/día (1,4 mg). En los dos últimos trimestres del embarazo, los requerimientos aumentan hasta 80 µg/kg/día (5 a 6 mg) y son similares para el lactante por su rápido crecimiento. Estos requerimientos deben ser considerados en el contexto de la cantidad de hierro disponible para su absorción. En los países desarrollados, el contenido de hierro en la dieta es de unos 6 mg/1.000 kcal; esta cantidad coloca la ingesta diaria promedio del hombre adulto entre 12 y 20 mg y de la mujer adulta entre 8 y 15 mg.3 La anemia por deficiencia de hierro es la carencia nutricional de mayor prevalencia en el mundo, especialmente en los países en desarrollo. Los niños son particularmente vulnerables, ya que se asocia con un alto riesgo de deterioro a largo plazo en el desarrollo mental y motor. Ellos también sufren de menores puntajes en las pruebas de coeficiente intelectual, falta de concentración, transcursos de atención corta y fácil distractibilidad1. Particularmente preocupante es que los déficit de desarrollo asociados con la anemia por deficiencia de hierro han mos- Hierro trado ser irreversibles5. Los niños entre 6 y 24 meses son un grupo particularmente de alto riesgo para el desarrollo de deficiencia de hierro debido al bajo contenido de hierro biodisponible en los alimentos de la lactancia de los países en desarrollo. Además, los requerimientos son especialmente altos en mujeres embarazadas, lactantes, niños menores y adolescentes, quienes están en mayor riesgo de padecer deficiencia de hierro. En los países en desarrollo, la principal causa de deficiencia de hierro es la baja biodisponibilidad del hierro de la dieta. Las consecuencias de la deficiencia de hierro son muchas y serias, afectando no solamente la salud de los individuos sino también el desarrollo de las sociedades y los países. La prevención y el control de la deficiencia de hierro y la anemia en todos los grupos de una población con diferentes requerimientos de hierro implican coordinar diferentes intervenciones. En ciertos períodos de la vida, especialmente durante el embarazo y en los niños desde los 6 meses de edad, los requerimientos de hierro son altos. Para las mujeres embarazadas, el abordaje actual favorece la suplementación de hierro-folato durante el emba- razo pero los resultados en términos de salud pública no son alentadores. La suplementación de hierro-folato preventiva semanal de mujeres durante su vida reproductiva, cuya eficacia está reconocida, ofrece una alternativa promisoria; su impacto en términos de salud pública actualmente se encuentra en evaluación.3 El hierro corporal presente al nacer es suficiente para los requerimientos fisiológicos de los bebés de peso normal al nacimiento durante los primeros seis meses de vida. Después de esto, el lactante rápidamente se vuelve dependiente de un suministro adecuado de hierro dietario fácilmente absorbible. El contenido de hierro corporal se debe incrementar en ~70% dentro de los 4 y 12 meses de edad 1. Con base en un modelo factorial, se ha estimado que el requerimiento diario promedio para el hierro absorbido desde los 712 meses de edad es 0.69 mg 2. Se necesita un poco menos de hierro después de los 12 meses de edad, ~0.63 mg/día para niños de 18 meses de edad. Los lactantes alimentados con leche materna con peso normal al nacimiento, rara vez desarrollan deficiencia de hierro antes de los 6 meses de edad. Sin embargo, el riesgo 9 Hierro se incrementa rápidamente durante los siguientes 3 meses en aquellos que continúan alimentándose con leche materna si otros ítems dietarios no incluyen una fuente rica en hierro altamente biodisponible.3 2. Fuentes de hierro en la dieta La carne animal es la fuente más fácilmente disponible de hierro para los humanos, porque contiene heme, el cual siempre es bien absorbido y promueve la absorción de otro hierro dietario (no heme). Los alimentos vegetales a menudo son ricos en factores que hacen del hierro no heme menos disponible para la absorción. Todo el hierro no heme en un metal que es soluble en los jugos gástricos ingresa a un depósito común.3 La promoción de la alimentación con leche materna es un elemento esencial de las intervenciones nutricionales durante la lactancia y la niñez temprana. Cualquier intento de suplir las necesidades de hierro adicionales con la introducción de alimentos complementarios debe asegurar que estos no reemplazan la leche materna disponible. Esto es especialmente importante en el segundo año de vida, el tiempo en que los requerimientos de 10 hierro son los más altos. Las cantidades de alimentos complementarios consumidas pueden ser relativamente pequeñas. Los alimentos ricos en hierro (más de 5 mg/100 g) son las vísceras, como hígado y corazón, levadura de cerveza, germen de trigo, yema de huevo, ostras y ciertas legumbres y frutas secas; los alimentos deficientes en hierro (menos de 1 mg/100 g) incluyen leche y productos lácteos y la mayoría de los vegetales no verdes. El contenido de hierro en los alimentos es afectado por la forma de preparación, ya que puede ser agregado por contaminación con suciedad o por su cocción en recipientes de hierro (Tabla 1).6 Tabla 1. Algunas fuentes alimenticias de hierro.6 Alimentos Porción Hierro (mg) • Hígado de res • Hígado de pollo • Carne de res • Morcilla • Yema de huevo • Tortilla de maíz • Granos (fríjol, lenteja, poroto, garbanzo) • Hojas verdes (espinaca) • Melón • Agua de coco • Leche de coco 3 oz. 2 oz. 3 oz. 1 oz. 1 u. 1 u. ½ taza 7.5 6.0 2.7 18.0 1.2 3.9 2.1 ½ taza 3.0 ¼ fruta 1 vaso 1 taza 1.4 1.4 3.6 Hierro 3. Hierro oral 3.1. Sales ferrosas Evidentemente, la primera opción, además de mejorar la dieta, es suplir el hierro por vía oral. El promedio de la dosis de hierro para adultos, requerido por día para el tratamiento de la anemia es de 200 mg (2-3 mg/kg), niños entre 15-30 kg requieren la mitad de la dosis de los adultos, niños más pequeños requieren 5 mg/kg/día, la profilaxis y las deficiencias nutricionales leves de hierro, como por ejemplo en el embarazo, pueden ser manejadas con dosis de 30-60 mg/día. Existen diversas preparaciones de sales y compuestos de hierro. El sulfato ferroso es una sal hidratada la cual contiene 20% de hierro elemental, el fumarato ferroso contiene 33% de hierro elemental y es moderadamente soluble en agua, el gluconato ferroso contiene 12% de hierro. La cantidad de hierro elemental, es más importante, que la masa total de la sal de hierro en la tableta. Variaciones específicas en las diferentes sales ferrosas, tienen relativamente poco efecto en la biodisponibilidad.7 Si se administra en su forma ferrosa (Fe2+)-como las sales ferrosas-, tiene que ser oxidado para poder incorporarse a las proteínas de transporte, transferrina y ferritina, respectivamente. Este proceso de oxidación causa la liberación de radicales libres, capaces de producir los efectos nocivos, tales como peroxidación lipídica y por consiguiente daño celular. El proceso es diferente si se administra en forma de hierro férrico (Fe3+), debido a que este no necesita oxidarse para ligar a las proteínas de transporte, y por lo tanto no genera liberación de radicales libres, disminuyendo la presentación de efectos secundarios.7 La dosis usada está regulada entre los beneficios terapéuticos deseados y los efectos tóxicos presentados, este es uno de los inconvenientes más frecuentes encontrados con el uso de las sales ferrosas. La intolerancia a las preparaciones orales de hierro se debe principalmente a la cantidad de hierro soluble en el tracto gastrointestinal superior, los efectos secundarios incluyen pirosis, náuseas, plenitud gástrica, estreñimiento y diarrea. Estos efectos secundarios pueden presentarse en aproximadamente el 26% de los pacientes y se incrementa aproximadamente al 40% cuando se dobla la dosis de hierro.7 11 Hierro Cualquier sobredosis de la sal puede superar fácilmente la “barrera de la mucosa” para causar toxicidad aguda [en los Estados Unidos, el Centro de Control de Intoxicaciones informó en 1997 que durante la década de 19861996 hubo 100.000 informes de intoxicación aguda por hierro en niños menores de 6 años, lo que subraya los posibles peligros del uso indiscriminado.8 3.2. Sales férricas Las sales férricas tradicionalmente no han sido preferidas sobre las sales ferrosas ya que el hierro férrico requiere primero la reducción a la forma ferrosa en la luz intestinal y usualmente esta capacidad de reducción no es suficiente para reducir las dosis de hierro terapéuticamente administrado. La biodisponibilidad del hierro de las sales férricas es 3 a 4 veces menor que la del sulfato ferroso. Mientras que 100 mg de hierro de sulfato ferroso/día son suficientes para una compensación oral óptima de terapia de hierro en adultos y para producir tasas de regeneración de hemoglobina iniciales de aproximadamente 0.26 g/100 mL/día, 400 a 1000 mg de hierro férrico/día son necesarios para el mismo efecto terapéutico 12 debido a la pobre biodisponibilidad del hierro férrico. Las sales férricas, sin embargo, llevan la ventaja inherente de un pobre potencial de intoxicación dada la limitada disponibilidad del contenido gástrico. Otras propiedades son esencialmente similares a las sales ferrosas.9 3.3. Hierro aminoquelado El hierro aminoquelado resulta de la unión covalente del hierro en forma ferrosa (Fe2+) a un ligante orgánico, lo cual reduce la carga del catión y provee alguna protección espacial en el sitio de la unión, lo que podría disminuir la toxicidad gastrointestinal debida a la irritación local. El grado de protección inherente en la molécula variará de acuerdo a la fortaleza de la unión del ligante, logrando bloquear: al menos un lado en la estructura espacial del ión metálico, o la potencial carga remanente del catión de hierro, o las reacciones de radicales libres comprometidos, los cuales contribuyen a los efectos tóxicos en el organismo.7 El hierro amino-ácido quelado corresponde a conjugados de hierro ferroso o férrico con aminoácidos. Aunque se han formulado Hierro numerosos conjugados, los más estudiados de estos son el bis-glicinato ferroso (hierro elemental 20%), trisglicinato férrico y glicina sulfato ferroso. Ellos no tienen efecto sobre el color o el sabor de los productos alimenticios. Además, tienen bajas propiedades pro-oxidantes.10 La molécula resultante es un metal quelado, donde las uniones de quelación deben ser lo suficientemente fuertes, para resistir a la división por efecto de la digestión o por los reactivos naturales presentes en los alimentos (como ácido fítico, fosfatos, ácido tánico), y además proteger los átomos del mineral para ser absorbidos y utilizados.7 Los ligantes orgánicos más frecuentemente utilizados son los aminoácidos, debido a que estos son predominantemente captados en el yeyuno, donde las proteínas son típicamente hidrolizadas y absorbidas.7 Estos compuestos quelados teóricamente disminuyen los efectos secundarios descritos con relación a otras presentaciones ferrosas (Fe2+), pero no en relación con los complejos polimaltosados, evidenciándose en la práctica clínica que persisten fre- cuentes molestias gastrointestinales.7 La biodisponibilidad de los hierros aminoquelados presenta variaciones marcadas con relación a otros suplementos, disminuyendo la absorción y el hierro disponible para la síntesis de la hemoglobina, por esta razón la FDA solo lo recomienda para la fortificación de alimentos, pero no como un suplemento terapéutico, prefiriéndose otras opciones de hierro disponibles.7 Para que un quelado sea nutricionalmente funcional, este debe tener un bajo peso molecular. El hierro bis-glicinato tiene un peso molecular menor a los 800 Da.11 3.4. Complejo de hierro polimaltosado (III) (IPC) El complejo de hidróxido de hierro férrico (Fe3+) polimaltosado no iónico (IPC) es un complejo hidrosoluble de hidróxido de hierro férrico (Fe3+) polinuclear y dextrina parcialmente hidrolizada (polimaltosa). La superficie de los núcleos de hidróxido de hierro (Fe3+) polinuclear está rodeada de varias moléculas de polimaltosa de unión no covalente, lo que resulta en un complejo con un peso molecular total de aproximadamente 52.300 Da. Esta molécula es tan 13 Hierro grande que la difusión a través de la membrana de la mucosa es unas 40 veces menor que aquella de la unidad del hierro(II) hexacuo. El complejo es estable y no libera hierro iónico bajo condiciones fisiológicas.7 Las propiedades farmacológicas y el potencial toxicológico del IPC son diferentes a las que se han observado con el sulfato ferroso. El potencial prácticamente no tóxico del IPC se asume que se debe al hecho de que existe un transporte activo de hierro y una tasa determinante del intercambio con ligantes, en vez de una difusión pasiva, y que este proceso ocurre con total ausencia de iones libres de hierro en todo momento. Se sugiere que el IPC transfiere el hierro cuando entra en contacto con los sitios de unión del hierro en la superficie de las células mucosas. Un complejo mixto entre el hierro, el ligando polimatosa y los sitios de unión de las células mucosas se forma para que se lleve a cabo un proceso de absorción (intercambio competitivo de ligandos). No es posible demostrar la bioequivalencia del IPC aplicando los métodos usuales de determinación del «área bajo la 14 curva del plasma» del hierro, esto debido a que el IPC tiene un comportamiento de absorción completamente diferente a las sales de hierro (Fe2+), resultando en una absorción con niveles de hierro sérico diferentes, así como distintas constantes de consumo y eliminación de hierro sérico y diferentes volúmenes de distribución. En conjunto, se puede decir que las pruebas de las preparaciones de hierro complejo de hidróxido de hierro(III) polimaltosado y las preparaciones de sales ferrosas, esencialmente se comportan en forma diferente. Las diferencias en los hallazgos toxicológicos así como en el comportamiento farmacocinético se pueden explicar y pronosticar con base en las propiedades físico-químicas de las preparaciones.7 La toxicidad aguda del IPC es baja; aproximadamente 10 veces menor que la del sulfato ferroso. La LD50 de IPC en ratones o ratas cuando se les administra por vía oral es > 2000mg/kg de peso corporal. Debe mencionarse que la principal parte de los depósitos de hierro del IPC se encontraron en el sistema retículo-endotelial (SRE) y no en el parénquima. Esto representa una ventaja esencial de Hierro este compuesto, la peroxidación lipídica radical inducida por el hierro, que sólo ocurre en el parénquima, no se evidencia con esta preparación. Por consiguiente, con el IPC no se esperan lesiones en el hígado, lo que ha sido confirmado por medio de resultados experimentales e histológicos. Estudios de interacción in-vitro, demostraron que el IPC es apropiado para administración oral simultánea con otras drogas sin afectar la absorción del hierro o de los otros componentes. En un reporte de Kaltwasser y Werner (1993) se resumen los resultados de varios estudios, realizados en la investigación de la absorción intestinal de diversas preparaciones de hierro tomando en consideración la forma de consumo (con o sin comidas), los autores concluyeron que comidas tomadas simultáneamente con preparados de hierro tienen una influencia negativa en la absorción de hierro de sales ferrosas, pero muestran un efecto favorable en el caso de la absorción de hierro del IPC. El IPC debido a su mecanismo de absorción fisiológicamente controlado, tiene efectos secundarios y toxicidad mucho menores que otras formas de suplementos de hierro, manteniendo unos óptimos niveles de absorción y biodisponibilidad que aseguran la respuesta terapéutica esperada. De hecho, sus particulares ventajas y el grado de aceptación alcanzado lo convierten en líder mundial dentro de las preparaciones de hierro. Además, es importante destacar que el alto grado de tecnología requerido obliga a dudar de preparaciones que fabricantes diferentes al original ofrecen sin tener la equivalencia sustentada.7 3.5. Hierro carbonilo El hierro carbonilo es una preparación de partículas pequeñas de hierro metálico altamente purificado. “Carbonilo” describe el proceso de manufactura de las partículas de hierro (de gas pentacarbonilo de hierro). Dado el pequeño tamaño de las partículas (<5 mcm) el ácido del estómago solubiliza este hierro. En el proceso de esta solubilización, los iones H+ son consumidos por lo tanto, incrementando el pH. Igualmente, como resultado la absorción del hierro es lenta (permitiendo la liberación continua de 1 a 2 días) y autolimitada por la tasa de secreción de ácido por la mucosa gástrica.9 15 Hierro El hierro carbonilo es mucho menos tóxico que las formas ionizadas de hierro. En humanos, la dosis letal de SF es ~200 mg/kg. Estudios de toxicidad de hierro carbonilo en animales, demostraron una dosis letal de 50000 a 60000 mg/kg (comparados con 200 mg/ kg de SF). En una reciente serie de casos no se informó toxicidad seria en todos los 33 pacientes con ingestiones de hierro carbonilo promedio de 11.2 mg/kg.13 3.6. Otras formulaciones de hierro Otras preparaciones de hierro están en diversas etapas de desarrollo, se destacan las preparaciones basadas en heme. La hemoglobina como una fuente de hierro fue promovida sobre la base de la alta biodisponibilidad del hierro heme. Sin embargo, el contenido de hierro de la hemoglobina es 0.34%. Como resultado, 300 mg de hemoglobina se requieren para liberar 1 mg de hierro elemental el cual conduce a grandes volúmenes y costos inhibitorios. Preparaciones más recientes en estudio incluyen oxalato ferroso, sulfato ferroso microencapsulado y fumarato ferroso microencapsulado. El oxalato ferroso ha sido encontrado recientemente que tiene 16 buena eficacia y baja toxicidad en estudios conducidos en cerdos. Recientemente, un suplemento que contiene fumarato ferroso microencapsulado (más ácido ascórbico) ha sido desarrollado y puede ser dispersado sobre cualquier alimento complementario. El hierro siendo encapsulado no cambia el color y sabor del alimento y se ha encontrado que es igualmente biodisponible al SF (31). De forma similar, una preparación de sulfato ferroso microencapsulada con fosfolípidos se encontró que tiene biodisponibilidad equivalente al SF.14-16 En términos de eficacia, todas las preparaciones de hierro disponibles son efectivas aunque el tiempo de la respuesta puede variar. Los aminoácidos quelados de hierro ofrecen teóricamente la mayor ventaja. De ellos, se destaca que El IPC ha probado tener menos efectos colaterales. El hierro carbonilo no ha sido evaluado extensamente en niños pero si se extrapolan los datos de los adultos es seguro y efectivo. 4. Hierro parenteral Cuando la terapia oral es insuficiente y/o se requiere mayor velo- Hierro cidad en la respuesta, la administración de hierro parenteral es una alternativa efectiva. Se recurre a esta vía en las siguientes condiciones: malabsorción del hierro (esprue tropical, síndrome intestino corto, etc.), intolerancia severa al hierro oral, suplementación en nutrición parenteral total, pacientes con insuficiencia renal crónica que presentan deficiencia funcional de hierro (pacientes en hemodiálisis, pacientes con depósitos bajos de hierro), anemia moderada a severa en el embarazo, terapia conjunta con eritropoyetina, programas de autotransfusión predepósito, necesidad de recuperación rápida de los niveles de hemoglobina y/o depósitos de hierro (pre y post quirúrgico, postparto, embarazo), enfermedad inflamatoria intestinal. En las áreas donde más experiencia y difusión del hierro parenteral existe son la nefrología, la gineco-obstetricia y los programas de autotransfusión. Existen tres formas de hierro parenteral: hierro dextrano, gluconato férrico sódico y hierro sacarato.17 4.1. Hierro dextrano Es una solución coloide de oxihidróxido férrico con dextrano polimerizado, que puede ser administrado por vía intravenosa o intra- muscular, con un peso molecular de ± 96.000 Da. Administrado intravenosamente en dosis menores de 500 mg, el hierro dextrano es depurado exponencialmente, con una vida media plasmática de 6 horas; cuando se administran dosis iguales o mayores de 1g la depuración por las células reticuloendoteliales es constante en 10-20 mg/hora. Esta tasa de depuración lenta resulta en una coloración marrón del plasma por varios días y en una elevación del hierro plasmático por una a dos semanas. Una vez el hierro es liberado del dextrano en el SRE, es incorporado en los depósitos o transportado por la transferrina a la médula ósea. La tasa de liberación es variable, mientras que una porción del hierro procesado está rápidamente disponible para la médula ósea, una fracción significativa es únicamente incorporada al hierro de los depósitos. Todo el hierro es eventualmente liberado, aunque se requieren muchos meses antes de que este proceso se complete. Durante este tiempo, la apariencia visible de los depósitos de hierro dextrano en el SRE puede confundir a los clínicos que evalúan el estado de hierro del paciente. 17 Hierro La aplicación intramuscular de hierro dextrano, únicamente puede ser iniciada después de realizada una dosis de prueba de 0.5 ml (25 mg de hierro). Si no se presentan reacciones adversas, se puede llevar a cabo la aplicación de la dosis total. Se informan con frecuencia reacciones locales, incluyendo dolor crónico y coloración local de la piel. Una dosis de prueba también debe preceder la administración intravenosa de la dosis terapéutica del hierro dextrano. El paciente debe ser observado durante la administración para signos de anafilaxis inmediata, y por una hora después de la aplicación para cualquier signo de inestabilidad vascular o hipersensibilidad, incluyendo dificultad respiratoria, hipotensión, taquicardia o dolor torácico. Cuando el paciente recibe crónicamente dextrano, una dosis de prueba debe ser aplicada previa a cada infusión, debido a que la hipersensibilidad puede aparecer en cualquier momento. Igualmente, se observan reacciones de hipersensibilidad retardada, especialmente en pacientes con artritis reumatoidea o con historia de alergias. Fiebre, malestar general, 18 linfadenopatías, artralgias, y urticaria pueden presentarse días o semanas posteriores a la aplicación. Una vez la hipersensibilidad es documentada, la terapia con hierro dextrano debe ser abandonada. Sus reacciones anafilácticas críticas constituyen el riesgo más serio, ocurren en cerca del 0.1-1% de los pacientes; esta condición ha hecho que se prefieran otras formas de hierro parenteral para ser utilizadas. La reacción puede tener un desenlace fatal aún con tratamiento adecuado, es por este motivo que su utilización en diálisis en los EE.UU. en los últimos años ha sido gradualmente suspendida y reemplazado por formas más seguras como el hierro sacarato.17 En agosto de 2004, en Dinamarca, las autoridades tuvieron que intervenir al observarse que no se estaba presentando información objetiva y completa sobre el hierro dextrano. Puntualmente, se exigió a los fabricantes que: a) no es aceptable que se afirme que el hierro dextrano es una formulación especialmente fraccionada de hierro de bajo peso molecular, b) no se puede promocionar que el peso molecular del hierro dextrano es similar al hierro sacarato o sacarosa. Hierro 4.2. Hierro gluconato El complejo de gluconato férrico sódico, tiene un peso molecular de aproximadamente 350.000 ± 23.000 Da, contiene el mismo centro de hidróxido de hierro que el hierro dextrano, pero utiliza el gluconato para estabilizar y solubilizar el compuesto; este carbohidrato puede tener también un potencial anafiláctico. Las reacciones anafilácticas con hierro gluconato, ocurren con menor frecuencia e intensidad que con hierro dextrano, se han reportado hasta en el 0.8% de los pacientes en algunas series.17 4.3. Hierro sacarato Consiste en un complejo de hierro polinuclear similar a la ferritina (complejo proteico de hierro (Fe3+) con hidróxido fosfato), en que el ligando de proteína apoferritina es sustituido por un compuesto carbohidratado. Esta sustitución es necesaria debido a que la ferritina posee propiedades antigénicas al ser aplicada por vía parenteral. Contiene el hierro en forma no iónica, como un complejo soluble en agua de hierro (Fe3+) con hidróxido de sacarato. Los filamentos polinucleares de hierro (Fe3+)– hidróxido se hallan rodeados periféricamente por un gran número de moléculas de sacarato unidas por enlaces no covalentes. De este modo, se forma un complejo férrico de alto peso molecular (PM), aproximadamente 43 kDa, que no es excretado por vía renal. La estabilidad del complejo es óptima lo que asegura que, en condiciones fisiológicas, no se libere hierro no iónico. Los aspectos sobre absorción y distribución del hierro sacarato se presentan en el capítulo 2: absorción del hierro.17 Referencias 1. www.lenntech.com 2. Fujita H, et al. Molecular biology of iron in nutritional science. Nippon Eiseigaku Zasshi. 2003 May;58(2):24853. 3. Hillman RS. Hematopoietic agents: growth factors, minerals, and vitamins. In, Goodman and Gilman’s The Pharmacological basis of therapeutics, 10th ed. (Hardman JG, Limbird LE, eds.) McGraw-Hill, New York, 2001, p14871517 4. National Family Health Survey (NFHS-2), India, 1998-99. Mumbai, International Institute for Population Studies and ORC Macro, 2000. 5. Technical consultation on strategies for prevention and control of iron deficiency anaemia amongst under three children in India. Indian Pediatr 2002;39:640-647. 6. www.paho.org/Spanish/DD/PUB/cre- 19 Hierro cer-nutricion.pdf 7. Delgado A. Terapia oral. Revista Latinoamericana de Farmacología. 2004;p. 22-26. 8. Preventing iron poisoning in Children. FDA Backgrounder. 1997 9. Nagpal J, Choudhury P. Iron Formulations in Pediatric Practice. Indian Pediatrics 2004; 41:807-815 10. Lindsay HA. Advantages and Limitations of Iron Amino Acid Chelates as Iron fortificants.Nutr Rev 2002.;60: S18-S21 11. Hertrampf E, Olivares M. Iron amino acid chelates. Int J Vitam Nutr Res. 2004 Nov;74(6):435-43. 12. Muller A Geisser P. Iron Pharmacokinetics after administration of ferrichydroxide-polymaltose complexes in rats. Arneim Forsch Drug Res 1984; 34:1560-1569. 13. Spiller HA, Wahlen HS, Stephens TL, Krenzelok EP, Benson B, Peterson J, Dellinger JA. Multi-center retrospective evaluation of carbonyl iron ingestions. Vet Hum Toxicol 2002; 44: 2829. 14. Nenortiene P, Sapragoniene M, Stankevicius A, Matusevicius AP, Daunoras G. Preparation, analysis and anti-anemic action of peroral powders with ferrous oxalate. Ferosol-1. Medicina (Kaunas) 2002; 38: 63-68. 15. Zlotkin S, Arthur P, Antwi KY, Yeung G. Treatment of anemia with microencapsulated ferrous fumarate plus ascorbic acid supplied as sprinkles to complementary (weaning) foods. Am J Clin Nutr 2001; 74: 791-795. 16. Olivares M. Bioavailability of microencapsulated ferrous sulfate in milk. Nutrition 2002; 18: 285-286. 17. Delgado A. Terapia parenteral. Revista Latinoamericana de Farmacología. 2004;p. 27-30. 20 Lecturas complementarias hierro oral Braunwald, E; et.al. Harrison’s principles of internal medicine. 15th ed. Vol. 1. New York:McGraw-Hill. 2001 pp. 660 DeMaeyer, EM; et.al. Preventing and controlling iron deficiency anaemia through primary health care:A guide for health administrators and programme managers. WHO. 1989 Dugdale, M; et.al. Anemia. Obstet Gynecol Clin North Am, 2001; 28 (2): 363 – 81 Hardman, JG; Goodman and Gilman’s the pharmacological basis of therapeutics. 10th ed. New York:McGraw-Hill. 2001 pp. 149 Jacobs, P; et.al. Absorption of iron polymaltose and ferrous sulphate in rats and humans.S Afr Med J, 1979; 55 (10): 1065 – 72 Jacobs, P; et.al. Better tolerance of Iron polymaltose complex compared with ferrous sulphate in the treatment of Anaemia. Hematology, 2000; 5 (1): 77 – 83 Jacobs, P; et.al. Oral iron therapy in human subjects, comparative absorption between ferrous salts and iron polymaltose. J Med, 1984; 15 (5): 367 - 77 Jacobs, P; et.al. The bioavailability of an iron polymaltose complex for treatment of iron deficiency. J Med, 1979; 10 (4): 279 – 85 Jeppsen, RB; et.al. Toxicology and safety of Ferrochel and other iron amino acid chelotes. Archivos Latinoamericanos de Nutrición, 2001; 51 (1): 26 – 34 Kaltwasser, JP; et.al. Bioavailability and therapeutic efficacy of bivalent and trivalent iron preparations. Drug Res, 1987; 37 (1) (I): 122 – 9 Langstaff, RJ;et.al. Treatment of iron-defi- Hierro ciency anaemia: a lower incidence of adverse effects with ferrum hausmann than ferrous sulphate. Br J Clin Res, 1993; 4: 191 – 198 Naude, S; et.al. Iron supplementation in preterm infants: a study comparing the effect and tolerance of a Fe2+ and a nonionic FeIII compound. J Clin Pharmacol, 2000; 40 (12) (2): 1447 – 51 Pestaner, JP; et.al. Ferrous sulfate toxicity: a review of autopsy findings. Biol Trace Elem Res, 1999; 69 (3): 191 – 8 Pineda, O; et.al. Effectiveness of iron amino acid chelate on the treatment of iron deficiency anemia in adolescents. J Appl Nutr, 1994; 46 (1): 1 – 13 Ramírez F, et.al. Anemia ferropénica en niños, rapidez de la respuesta al tratamiento con polimaltosado férrico (IPC) y ferritina (F o Proteína férrica). Data on file Skikne, B; et.al. Bovine ferritin iron bioavailability in man. Eur J Clin Invest 1997;27:228-33 Toumainen, TP; et.al. Oral supplementation with ferrous sulfate but not with non-ionic: Iron polymaltose complex increases the susceptibility of plasma lipoproteins to oxidation. Nutr Res, 1999; 19 (8): 1121 – 32 Lecturas complementarias hierro parenteral Al-Mómen, AK; et.al. Intravenous iron sucrose complex in the treatment of iron deficiency anemia during pregnancy. Eur J Obstetrics Gynecol 1996;69:121-4 Braunwald, E; et.al. Harrison’s principles of internal medicine. 15th ed. Vol. 1. New York:McGraw-Hill. 2001 pp. 660 Danielson, BG; et.al. Pharmacokinetics of iron(III)-hydroxide sucrose complex after a single intravenous dose in healthy volunteers. Drug res 1996;46(6):615-21 Fishbane,S; et.al. The comparative safety of intravenous iron Dextran, iron saccharate, and sodium ferric gluconate. Seminars in Dialysis 2000; 13(6):381-4 Geisser, P; et.al. Structure / histotoxicity relationship of parenteral iron preparations. Drug Res, 1992; 42: (12): 1439 - 52 Hamstra, RD ; et.al. Intravenous iron Dextran in clinical medicine. Jama 1980;243:1726-31 Harju, E. Clinical pharmacokinetics of iron preparations. Clin Pharmacokinet 1989; 17(2):69-8 Hardman, JG; Goodman and Gilman’s the pharmacological basis of therapeutics. 10th ed. New York:McGraw-Hill. 2001 pp. 149 Johnson, C; et.al. Intravenous iron products. ANNA Journal 1999 ;26(5) :522-4 Van Wyck, DB; et.al. Safety and efficacy of iron sucrose in patients sensitive to iron Dextran: North American clinical trial. Am J Kidney Dis 2000; 36(1):88-97 Wood, JK; et.al. The metabolism of irondextran given as a total-dose infusion to iron deficient Jamaican subjects. Br J Haematol, 1968; 14 (2): 119 - 29 Yee, J; et.al. Iron sucrose: The oldest iron therapy becomes new. Am J Kidney Dis 2002;40:1111-1121 Zanen, AL; et.al. Oversaturation of trnasferrin after intravenous ferric gluconate (Ferrlecit) in haemodialysis patients. Nephrol Dial Transplant 1996; 11:820-4 21 Hierro Jorge Vargas 1. Control molecular A unque la cantidad de hie rro extraída de la dieta es pequeña, la regulación de la absorción intestinal del hierro es crítica porque los humanos no tenemos una vía fisiológica para la excreción. Las células de la cripta duodenal detectan los requerimientos de hierro del organismo y son programadas por esta información a medida que maduran a enterocitos absortivos. Los enterocitos que cubren las vellosidades absortivas cercanos a la unión gastroduodenal son responsables de la absorción de todo el hierro. El hierro debe pasar desde la luz intestinal a través de las membranas apical y basolateral del enterocito para alcanzar el plasma. El hierro obtenido de los alimentos no está unido a la transferrina, y no existe función para la transferrina dentro de la luz del intestino. En vez de ello, el bajo pH del efluente gástrico ayuda a disolver el hierro ingerido y proporciona un medio rico en protones. Esto faci22 lita la reducción enzimática del hierro férrico a su forma ferrosa por una ferrireductasa en el borde en cepillo intestinal. El transportador metálico divalente 1 (DMT1; conocido anteriormente como Nramp2 o DCT1), es una proteína que transfiere el hierro a través de la membrana apical hacia la célula por medio de un proceso acoplado a protones. El DMT1 no es específico del hierro, puede transportar una amplia variedad de iones metálicos divalentes, incluyendo manganeso, cobalto, cobre, zinc, cadmio y plomo. El hierro heme es captado por un proceso separado que no está bien caracterizado. En el interior del enterocito absortivo, el hierro tiene dos posibles destinos: puede ser almacenado como ferritina, o puede ser transferido a través de la membrana basolateral para alcanzar el plasma. Estos no son mutuamente excluyentes, y el factor determinante es probablemente un «punto definido» de absorción del hierro que Hierro fue establecido cuando el enterocito se desarrolló de una célula de la cripta. El hierro que permanece en forma de ferritina a medida que el enterocito completa su limitado ciclo de vida exfoliará y abandonará el organismo a través del tracto gastrointestinal. Este proceso representa un mecanismo importante de pérdida de hierro. El transportador basolateral en el enterocito de hierro no ha sido identificado definitivamente, pero una proteína recientemente descrita, Ireg1, es un probable candidato. Además, la ceruloplasmina también tiene un importante papel en el metabolismo del hierro. La absorción del hierro intestinal está regulada en varias formas. Primero, puede ser modulada por la cantidad de hierro recientemente consumido en la dieta, un mecanismo referido como el regulador dietario. Durante varios días después de un bolo de hierro dietario, los enterocitos absortivos son resistentes a adquirir hierro adicional. Este fenómeno ha sido llamado «bloqueo de la mucosa». Esta acción de bloqueo probablemente resulta de la acumulación de hierro intracelular, la cual conduciría a que al enterocito detecte que se han satisfecho sus requerimientos de punto definido. Esto puede ocurrir incluso en la presencia de deficiencia sistémica de hierro. Un segundo mecanismo regulador también detecta los niveles de hierro, pero responde al hierro corporal total, más que al hierro de la dieta. Este mecanismo ha sido denominado regulador de los depósitos. Es capaz de cambiar la cantidad de hierro absorbida en una extensión limitada. El tercer mecanismo regulatorio, conocido como el regulador eritropoyético, no responde a los niveles de hierro. Más bien, modula la absorción de hierro en respuesta a los requerimientos para la eritropoyesis. El regulador eritropoyético tiene una mayor capacidad de incrementar la absorción de hierro que el regulador de los depósitos. Es lógico que el eritrón debe tener alguna influencia sobre la tasa de absorción de hierro intestinal, ya que la mayoría del hierro corporal es usado para la eritropoyesis. Se desconoce cómo se cumple esto. El regulador eritropoyético probablemente involucra una señal soluble que es llevada por el plasma desde la médula ósea al intestino. 23 Hierro En años recientes, se ha avanzado de forma notable en el conocimiento del control de la homeostasis del hierro a partir de estudios moleculares del metabolismo del hierro del mamífero. La captación de hierro plasmático por la transferrina involucra la unión de la transferrina rica en hierro (FeTf) por la proteína receptora transferrina (TfR) sobre la superficie celular, seguida por la endocitosis del complejo FeTf-TfR con la subsecuente liberación de hierro de la vesícula endocitótica a un depósito de hierro intracelular lábil (LIP). El hierro del LIP es utilizado para las necesidades metabólicas inmediatas de la célula (proteínas heme, citocromos, enzimas mitocondriales, etc.) o almacenado en la proteína de depósito del hierro ferritina (Ft). Las necesidades metabólicas de una célula de mamífero deficiente de hierro requiere la producción incrementada de TfR para facilitar la captación de hierro y la supresión de la síntesis de ferritina. Por el contrario, en caso de exceso de hierro, se necesita aumentar la síntesis de ferritina para prevenir una expansión potencialmente tóxica de LIP, mientras que al mismo tiempo la producción de TfR debe ser suspendida para prevenir la captación de hierro innecesario. 24 Ambos procesos son regulados simultáneamente por un elegante control de traducción recíproco y coordinado que involucra el ARNm citoplásmico por el elemento que responde al hierro (IRE) y el mecanismo de la proteína que responde al hierro (IRP). Se han descrito proteínas responsables de exportar el hierro a través de la membrana basolateral de la mucosa intestinal. Ireg1 codifica una proteína con múltiples dominios transmembrana y su ARNm contiene una estructura IRE que se une específicamente a IRP1 y IRP2. Los niveles de Ireg1 ARNm se correlacionan positivamente con tres condiciones independientes asociadas con incremento de la absorción de hierro: deficiencia de hierro, hipoxia y atransferrinemia. Otra proteína involucrada en la exportación de hierro desde la célula intestinal es la hefaestina, una ferroxidasa unida transmembrana la cual como aspecto interesante tiene una identidad del 50% con la ceruloplasmina. La hefaestina no se localiza en la membrana basolateral, y su relación con la Ireg1 en la actualidad no está clara. Sin embargo, su actividad ferroxidasa putativa debe facilitar la Hierro salida de hierro desde la célula creando un gradiente de concentración transmembrana de hierro ferroso. Igualmente, se ha descrito una reductasa férrica en la mucosa del duodeno, la cual es una citocromo reductasa que contiene un hem tipo b y se asume que es responsable de la actividad reductora férrica de la mucosa duodenal, requerida para el transporte de hierro luminal asociada con el DMT1. Otro descubrimiento reciente que se suma al fascinante fenómeno de la fina regulación del hierro es la hepcidina. Este regulador parece comunicar el estado de hierro corporal y la demanda para la eritropoyesis al intestino, y a su vez, modula la absorción del hierro intestinal. La hepcidina fue purificada de la sangre y orina humana como un péptido antimicrobiano y se encontró que se expresa predominantemente en el hígado. Se ha asociado la falta de expresión de la hepcidina con la sobrecarga de hierro y la sobreexpresión de hepcidina produce anemia por deficiencia de hierro en ratones. Además, las concentraciones de hepcidina disminuyen en los ratones alimentados con una die- ta alta en hierro. Estas observaciones apoyan el papel de la hepcidina como una señal que limita la absorción intestinal del hierro. La expresión de la hepcidina es afectada también por la hipoxia y la inflamación y es disminuida en los pacientes con hemocromatosis. Así, la relación entre estado de hierro corporal y hepcidina está alterada en los pacientes con hemocromatosis. Por lo que vemos, todavía queda mucho por investigar en la regulación de la hepcidina por el hierro, la hipoxia y la inflamación. 2. Hierro oral 2.1. Compuestos solubles en agua Los compuestos de hierro solubles en agua incluyen el sulfato ferroso. Su solubilidad es instantánea en el estómago. La absorción puede variar de aproximadamente un 1% a quizás un 50%, según el estado nutricional de hierro del individuo, la presencia de promotores e inhibidores de absorción del hierro en la comida y el contenido de hierro de la comida. La desventaja del sulfato ferroso es que reacciona fácilmente 25 Hierro con otras sustancias que existen naturalmente en la matriz alimentaria. Esto puede causar cambios sensoriales (sabor, color y olor) debido a la oxidación de grasas (rancidez). El sulfato ferroso también puede modificar las propiedades físicas del producto final hecho con los alimentos fortificados y precipitarse como complejos de hierro insolubles cuando se usa en preparaciones líquidas. Estas sales tienen una biodisponibilidad uniformemente buena. Sin embargo, la biodisponibilidad disminuye marcadamente en la presencia de inhibidores en la dieta como fitatos, ácido tánico, etc. 2.2. Sales férricas Se diferencian de las sales ferrosas por no contener el hierro en forma ferrosa (Fe2+), y del complejo de hidróxido férrico y polimaltosa (IPC) por no venir en forma de un complejo. La biodisponibilidad de hierro de las sales férricas es 3 a 4 veces menor que la del sulfato ferroso. Mientras que 100 mg de hierro de sulfato ferroso/día son suficientes para una compensación oral óptima de terapia de hierro en 26 adultos y para producir tasas de regeneración de hemoglobina iniciales de aproximadamente 0.26 g/100 mL/día, 400 a 1000 mg de hierro férrico/día son necesarios para el mismo efecto terapéutico debido a la pobre biodisponibilidad del hierro férrico. Las sales férricas, sin embargo, llevan la ventaja inherente de un pobre potencial de intoxicación dada la limitada disponibilidad del contenido gástrico. Otras propiedades son esencialmente similares a las sales ferrosas. 2.3. Poco solubles en agua/solubles en soluciones ácidas Estos compuestos se disuelven lentamente en la concentración ácida normal del estómago. El fumarato ferroso es el compuesto principal en esta categoría. Se absorbe tan bien como el sulfato ferroso en los adultos y adolescentes, pero los datos recientes indican que se absorbe menos en las personas con una concentración de ácido gástrico inferior, en particular los niños pequeños. La ventaja de este compuesto es que interactúa menos con la matriz alimentaria, y causa menos cambios sensoriales. Hierro 2.4. Insolubles en agua/poco solubles en soluciones ácidas Este grupo reúne los siguientes compuestos: 1) el hierro elemental, del cual existen tres tipos: a) reducido, b) electrolítico, y c) hierro carbonilo; 2) el pirofosfato férrico; y 3) el ortofosfato férrico. Estos compuestos son usados ampliamente por la industria de los alimentos en los países industrializados porque son bastante inertes y tienen efectos muy pequeños sobre las propiedades sensoriales de los alimentos. Sin embargo, su aporte a la absorción de hierro es dudosa debido a sus muy bajos niveles de solubilidad y absorción. 2.5. Compuestos quelados El compuesto quelado de hierro al cual se hace referencia más comúnmente es el NaFeEDTA (etilendiaminotetraacetato ferrosódico). La ventaja principal del uso del NaFeEDTA en la fortificación de alimentos es que, en esta forma, el hierro está protegido de los inhibidores de absorción del hierro de los alimentos en el estómago. La absorción de hierro a partir del NaFeEDTA agregado a los alimentos hechos con harinas de cereales de alta extracción o a una comida que contenga fitato es dos a tres veces mayor que en el caso del sulfato ferroso. Aunque no promueve la oxidación de grasas (rancidez) en la harina de trigo almacenada, el NaFeEDTA puede causar cambios de color inadmisibles en algunos vehículos alimentarios. Se ha determinado que la absorción de hierro a partir de bisglicinato ferroso es 1.1 a 5.0 veces mayor que la absorción de sulfato ferroso, pero inferior a la absorción de NaFeEDTA en estudios comparativos. El bisglicinato ferroso tiende a causar reacciones no deseadas sobre el color y la oxidación de grasas (rancidez) en las harinas de cereal almacenadas, lo cual limita su uso en estos alimentos. Sin embargo, parece ser útil para fortificar la leche. El trisglicinato férrico causa menos reacciones en los alimentos, pero su biodisponibilidad es mucho menor que la del bisglicinato ferroso. 2.6. Complejo de hierro polimaltosado (III) (IPC) En el capítulo 1 se presenta en forma amplia las características farmacológicas de esta formulación de hierro. Respecto a otras características interesantes, el 27 Hierro complejo de hierro polimaltosado (IPC) y el sulfato ferroso (SF) han demostrado tener biodisponibilidad equivalente en lactantes. La absorción del IPC no es afectada por los alimentos o la leche, lo que permite la administración independientemente del tiempo de la alimentación. Igualmente, hasta la fecha, no hay informe de ninguna interacción con alimentos o medicamentos. 2.7. Hierro carbonilo Estudios realizados en niños han mostrado que el IPC no tiene efectos adversos sobre los niveles de cobre y zinc; lo que sí se ha visto con SF después de un mes de suplementación. Del mismo modo, el IPC no tiñe los dientes como el SF y se presenta una tasa más baja de efectos colaterales respecto al SF. 3. Hierro parenteral El hierro del IPC se absorbe en el intestino a través de un intercambio competitivo autolimitado de ligandos, por lo que el sistema de transporte intestinal se satura en caso de una sobredosis. La intoxicación accidental con IPC por lo tanto se observa rara vez. Muller, et al. observaron que la LD50 del sulfato ferroso (SF) es 350 mg/kg, mientras que la LD50 de IPC podría estar registrada incluso a dosis mayores de 2000 mg/kg. 12 28 Como una fortificación de alimentos, el hierro carbonilo ha mostrado ser bien absorbido y utilizado en la síntesis de la hemoglobina, tanto en animales experimentales como en humanos (25). Sus ventajas incluyen ausencia de cambio en el color o sabor de los alimentos y ambientalmente estable. De las tres formas de hierro parenteral, revisaremos los aspectos sobre la absorción del hierro dextrano, el gluconato férrico sódico y el hierro sacarato. 3.1. Hierro dextrano Cuando se utiliza por vía intramuscular profunda, es movilizado gradualmente por los linfáticos y transportado al sistema reticuloendotelial (SRE), el hierro luego es liberado de la molécula de dextrano. Una proporción variable (1050%) puede ser fijada localmente en el músculo por varias semanas o meses, especialmente si hubo reacción inflamatoria. La administración intravenosa (en la cual no hay absorción) proporciona una respuesta más adecuada y es la preferida. Una limitación importan- Hierro te es la posibilidad de desencadenar reacciones anafilácticas. 3.2. Hierro gluconato Los niveles plasmáticos (concentración máxima), varían significativamente dependiendo de la dosis y la velocidad de administración; la concentración máxima se encuentra cuando de administran 125 mg en 7 minutos y tiene un valor de 19 mg/L. El volumen inicial de distribución es de 6 L. La vida media de eliminación terminal para el hierro unido es de aproximadamente 1 hora. La depuración total es de 3.02 - 5.35 L/ h. El área bajo la curva del hierro unido varía de 17.5 - 35.6 mg-h/L. Aproximadamente el 80% del hierro es entregado a la transferrina como un hierro iónico dentro de las 24 horas posteriores a la administración. Con relación al hierro dextrano el gluconato tiene una mayor y completa utilización. El tamaño y la rápida disociación pueden aumentar el riesgo de toxicidad aguda debida al hierro libre ionizado. En 1996, Zanen y Colaboradores, publicaron un estudio sobre el aumento excesivamente rápido de la liberación de hierro por el gluconato férrico sódico, encon- trando que los pacientes que presentan disminución de la presión arterial y flushing posterior a la administración del gluconato, tienen una sobre-saturación de la transferrina mayor al 100%, lo que lleva a un aumento en la toxicidad del hierro debido al hierro libre. 3.3. Hierro sacarato El hierro sacarato es captado casi exclusivamente por el sistema reticulo endotelial (SRE) hepático, el bazo y la médula ósea, lo mismo que por la transferrina y la apoferritina. Es rápidamente metabolizado subsecuentemente y disponible para la eritropoyesis. Como la mayor parte de los depósitos de hierro se encuentran en el SRE, y no en el parénquima, el hierro sacarato posee la ventaja de no provocar peroxidación lipídica inducida por los radicales de hierro. La farmacocinética del hierro sacarato muestra que se obtienen niveles máximos de hierro, de alrededor de 538 µmol/L, 10 minutos después de la inyección de 100 mg de hierro. El volumen de distribución del compartimiento central es de aproximadamente 3L. El volumen de distribución en el estadío previo es de alrededor de 8L. Langström y col (1995) investigaron las propiedades fármaco- 29 Hierro cinéticas del hierro sacarato, alrededor de 5 minutos después de la inyección se encontró un alto nivel de actividad no sólo en el hígado sino también a nivel de la médula ósea, indicando una alta concentración del hierro en estas áreas. En conclusión, es posible afirmar que el hierro del hierro sacarato es muy rápidamente utilizado para la eritropoyesis. Estas propiedades del hierro sacarato resultan especialmente útiles en pacientes con requerimientos clínicos de rápida liberación de hierro a los depósitos de hierro, en pacientes con anemia por deficiencia de hierro debida a administración insuficiente de hierro o pérdida exagerada del mismo, que no toleran o que responden insuficientemente a la terapia con hierro oral o en pacientes con mal absorción o mala tolerancia del hierro por vía oral. En los pacientes en tratamiento con eritropoyetina (rHuEpo), la optimización de la eritropoyesis es capaz de reducir significativamente, gracias al hierro, los requerimientos de rHuEpo. Comparado con los complejos de hierro del tipo lábil y débil, como son por ejemplo el gluconato férrico o el citrato férrico, el hierro sacarato ofrece la ventaja de ser rápida- 30 mente utilizable para la eritropoyesis. En relación con los efectos colaterales potenciales, la menor masa molecular de hierro sacarato puede considerarse una ventaja sobre los complejos de hierro del tipo robusto y fuerte. Debido a la ausencia de biopolímeros lentamente degradables, la frecuencia de reacciones alérgicas inducidas por la aplicación i.v. de hierro sacarato es muy baja. En un estudio que involucro 8100 pacientes/año, con mas de 160.000 dosis de 100 mg de hierro sacarato, no se presento ninguna reacción adversa que pusiera en peligro la vida, documentando la seguridad de esta forma de presentación. La FDA lo aprobó para su uso en USA, donde es ampliamente utilizado en los pacientes en diálisis y prediálisis. Recientemente su uso en ginecología y obstetricia ha mostrado beneficios clínicos importantes, sustentado en más de 100 publicaciones que evidencian resultados satisfactorios. 4. Aumentadores de la absorción del hierro no heme El ácido ascórbico es el promotor más eficiente de la absorción del hierro no heme. Tanto los jugos de Hierro frutas como el ácido ascórbico cristalino son efectivos. Las propiedades aumentadoras están relacionadas con la dosis y dependen de la presencia de ácido ascórbico en la luz del tracto gastrointestinal superior. El ácido ascórbico inyectado intravenosamente no tiene efecto. Se ha sugerido que otros ácidos orgánicos también pueden aumentar la absorción de hierro. Sin embargo, los efectos de otros ácidos orgánicos sobre la biodisponibilidad del hierro no han sido estudiados rigurosamente. Los experimentos que se han hecho han mostrado resultados inconsistentes. Algunos investigadores informaron modestas propiedades aumentadoras para ácido láctico, cítrico, málico y tartárico. Se han usado dos quelados de hierro para promover la absorción adecuada del hierro de comidas vegetales. El hierro en el EDTA de hierro sódico se absorbe dos a tres veces también como el hierro de los alimentos, cuando se agrega a comidas inhibitorias. No es adecuado para la fortificación de alimentos complementarios porque no es posible agregar suficiente hierro sin exceder la ingesta diaria aceptable para éste. Alguna evidencia sugiere que los quelados de aminoácidos, tales como bisglicinato de hierro, pueden ser útiles para la fortificación de productos lácteos. Información disponible no es suficiente para hacer recomendaciones del posible uso de estos compuestos. En resumen, la incorporación del heme o la adición de ácido ascórbico y una sal de hierro soluble son los únicos métodos prácticos establecidos para proporcionar cantidades adecuadas de hierro biodisponible en alimentos complementarios. Otro compuesto que mejora la absorción del hierro es el Na2EDTA (etilendiaminotetraacetato disódico), el cual quela, o une, fácilmente el hierro solubilizado en el estómago y el intestino. El Na2EDTA aumenta la absorción de hierro de dos a tres veces en los regímenes alimentarios que contienen cantidades altas de inhibidores de la absorción del hierro, siempre que el hierro provenga de fuentes fácilmente solubles en agua (por ejemplo, sulfato ferroso). Si el régimen alimentario es bajo en inhibidores de la absorción, el efecto estimulante del Na2EDTA tiene poca importancia, pero si el régimen alimentario es alto en inhibidores de la absorción 31 Hierro del hierro (por ejemplo, en las harinas de cereal de alta extracción), el Na2EDTA es una buena opción. 5. Eliminación o degradación de inhibidores Los principales inhibidores de absorción de hierro, presentes en los alimentos, son los fitatos, cuya presencia es abundante en los cereales y algunas leguminosas, y los polifenoles, que están presentes en el té, el café, el chocolate y los granos de sorgo. Estos inhibidores de la absorción forman complejos insolubles con el hierro, lo que hace que este no esté disponible para la absorción. La desfitinización podría ser una estrategia particularmente útil para mejorar la absorción del hierro de los cereales, los alimentos complementarios y la leche de soya. Sin embargo, debe extraerse prácticamente todo el fitato para obtener un aumento importante de la absorción de hierro. Lecturas complementarias control molecular Anderson GJ, et al. The ceruloplasmin homolog hephaestin and the control of intestinal iron absorption. Blood Cells Mol Dis. 2002 Nov-Dec;29(3):367-75. Barisani D, Conte D. Transferrin receptor 1 32 (TfR1) and putative stimulator of Fe transport (SFT) expression in iron deficiency and overload: an overview. Blood Cells Mol Dis. 2002 Nov-Dec;29(3):498505. Boccio J, et al. Iron metabolism: current concepts of an essential micronutrient. Arch Latinoam Nutr. 2003 Jun;53(2):119-32. Boccio J, et al. Current knowledge of iron metabolism. Biol Trace Elem Res. 2003 Jun;92(3):189-212. Chung J, Wessling-Resnick M. Molecular mechanisms and regulation of iron transport. Crit Rev Clin Lab Sci. 2003 Apr;40(2):151-82. Conrad ME, et al. Pathways of iron absorption. Blood Cells Mol Dis. 2002 NovDec;29(3):336-55. Danielson BG, et al. (1996) Iron therapy with special emphasis on intravenous administration. ISBN-85819-223-6 Eisenstein RS, et al. Novel roles for iron regulatory proteins in the adaptive response to iron deficiency. J Nutr. 2003 May;133(5 Suppl 1):1510S-6S. Frazer DM, et al. The orchestration of body iron intake: how and where do enterocytes receive their cues? Blood Cells Mol Dis. 2003 May-Jun;30(3):288-97. Ganz T. Hepcidin, a key regulator of iron metabolism and mediator of anemia of inflammation. Blood. 2003 Aug 1;102(3):783-8. Epub 2003 Mar 27. Guix P, et al. Molecular aspects of duodenal iron absorption. Gastroenterol Hepatol. 2003 Feb;26(2):86-93. Hallberg L, et al. Perspectives on iron absorption. Blood Cells Mol Dis. 2002 NovDec;29(3):562-73. Leong WI, et al. Hepcidin, the recently identified peptide that appears to regulate iron absorption. J Nutr. 2004 Jan;134(1):1-4. Miret S, Simpson RJ, McKie AT. Physiology and molecular biology of dietary iron absorption. Annu Rev Nutr. 2003;23:283301. Epub 2003 Feb 26. Hierro Morgan EH, et al. Mechanisms and regulation of intestinal iron absorption. Blood Cells Mol Dis. 2002 Nov-Dec;29(3):38499. Wessling-Resnick M. A possible link between hepcidin and regulation of dietary iron absorption. Nutr Rev. 2002 Nov;60(11):371-4. Lecturas complementarias hierro oral Braunwald, E; et.al. Harrison’s principles of internal medicine. 15th ed. Vol. 1. New York:McGraw-Hill. 2001 pp. 660 DeMaeyer, EM; et.al. Preventing and controlling iron deficiency anaemia through primary health care:A guide for health administrators and programme managers. WHO. 1989 Dugdale, M; et.al. Anemia. Obstet Gynecol Clin North Am, 2001; 28 (2): 363 – 81 Hardman, JG; Goodman and Gilman’s the pharmacological basis of therapeutics. 10th ed. New York:McGraw-Hill. 2001 pp. 149 Jacobs, P; et.al. Absorption of iron polymaltose and ferrous sulphate in rats and humans.S Afr Med J, 1979; 55 (10): 1065 – 72 Jacobs, P; et.al. Better tolerance of Iron polymaltose complex compared with ferrous sulphate in the treatment of Anaemia. Hematology, 2000; 5 (1): 77 – 83 Jacobs, P; et.al. Oral iron therapy in human subjects, comparative absorption between ferrous salts and iron polymaltose. J Med, 1984; 15 (5): 367 - 77 Jacobs, P; et.al. The bioavailability of an iron polymaltose complex for treatment of iron deficiency. J Med, 1979; 10 (4): 279 – 85 Jeppsen, RB; et.al. Toxicology and safety of Ferrochel and other iron amino acid chelotes. Archivos Latinoamericanos de Nutrición, 2001; 51 (1): 26 – 34 Kaltwasser, JP; et.al. Bioavailability and therapeutic efficacy of bivalent and trivalent iron preparations. Drug Res, 1987; 37 (1) (I): 122 – 9 Langstaff, RJ;et.al. Treatment of iron-deficiency anaemia: a lower incidence of adverse effects with ferrum hausmann than ferrous sulphate. Br J Clin Res, 1993; 4: 191 – 198 Naude, S; et.al. Iron supplementation in preterm infants: a study comparing the effect and tolerance of a Fe2+ and a nonionic FeIII compound. J Clin Pharmacol, 2000; 40 (12) (2): 1447 – 51 Pestaner, JP; et.al. Ferrous sulfate toxicity: a review of autopsy findings. Biol Trace Elem Res, 1999; 69 (3): 191 – 8 Pineda, O; et.al. Effectiveness of iron amino acid chelate on the treatment of iron deficiency anemia in adolescents. J Appl Nutr, 1994; 46 (1): 1 – 13 Ramírez F, et.al. Anemia ferropénica en niños, rapidez de la respuesta al tratamiento con polimaltosado férrico (IPC) y ferritina (F o Proteína férrica). Data on file Skikne, B; et.al. Bovine ferritin iron bioavailability in man. Eur J Clin Invest 1997;27:228-33 Toumainen, TP; et.al. Oral supplementation with ferrous sulfate but not with non-ionic: Iron polymaltose complex increases the susceptibility of plasma lipoproteins to oxidation. Nutr Res, 1999; 19 (8): 1121 – 32 Lecturas complementarias hierro parenteral Al-Mómen, AK; et.al. Intravenous iron sucrose complex in the treatment of iron deficiency anemia during pregnancy. Eur J Obstetrics Gynecol 1996;69:121-4 Braunwald, E; et.al. Harrison’s principles of 33 Hierro internal medicine. 15th ed. Vol. 1. New York:McGraw-Hill. 2001 pp. 660 Danielson, BG; et.al. Pharmacokinetics of iron(III)-hydroxide sucrose complex after a single intravenous dose in healthy volunteers. Drug res 1996;46(6): 615-21 Fishbane,S; et.al. The comparative safety of intravenous iron Dextran, iron saccharate, and sodium ferric gluconate. Seminars in Dialysis 2000; 13(6):381-4 Geisser, P; et.al. Structure / histotoxicity relationship of parenteral iron preparations. Drug Res, 1992; 42: (12): 1439 - 52 Hamstra, RD ; et.al. Intravenous iron Dextran in clinical medicine. Jama 1980;243:1726-31 Harju, E. Clinical pharmacokinetics of iron preparations. Clin Pharmacokinet 1989; 17(2):69-8 Hardman, JG; Goodman and Gilman’s the pharmacological basis of therapeutics. 10th ed. New York:McGraw-Hill. 2001 pp. 149 Johnson, C; et.al. Intravenous iron products. ANNA Journal 1999 ;26(5) :522-4 Van Wyck, DB; et.al. Safety and efficacy of 34 iron sucrose in patients sensitive to iron Dextran: North American clinical trial. Am J Kidney Dis 2000; 36(1):88-97 Wood, JK; et.al. The metabolism of irondextran given as a total-dose infusion to iron deficient Jamaican subjects. Br J Haematol, 1968; 14 (2): 119 - 29 Yee, J; et.al. Iron sucrose: The oldest iron therapy becomes new. Am J Kidney Dis 2002;40:1111-1121 Zanen, AL; et.al. Oversaturation of trnasferrin after intravenous ferric gluconate (Ferrlecit) in haemodialysis patients. Nephrol Dial Transplant 1996; 11:820-4 Lecturas complementarias aumentadores de la absorción del hierro no heme y eliminación o degradación de inhibidores Compuestos de hierro para la fortificación de alimentos: Guías para América Latina y el Caribe. 2002. Publicado originalmente en inglés: Ntr Rev NUREA8 60(7) [II]S50-S61 (Jul 2002). Hierro Jorge Vargas L El intercambio interno del hierro se realiza mediante la transferrina plasmática. El hierro es entregado por la transferrina a los sitios intracelulares mediante receptores específicos para la transferrina que se encuentran en la membrana plasmática. a distribución del hierro en los tejidos se muestra en la Tabla 1. Los hombres adultos normalmente tienen 35 a 45 mg de hierro por kg de peso corporal. Las mujeres premenopáusicas tienen depósitos de hierro más bajos como resultado de su pérdida recurrente de sangre a través de la menstruación. Más de dos tercios del contenido de hierro corporal es incorporado en la hemoglobina en el desarrollo de los precursores eritroides y los eritrocitos maduros. El flujo de hierro a través del plasma llega a un total de 30 a 40 mg diarios en el adulto (alrededor de 0,46 mg/kg de peso corporal). La circulación interna principal del hierro incluye al eritrón y las célu- Tabla 1. Distribución del hierro en adultos. Tipo de Hierro Concentración (mg Fe/kg) Mujeres Hombres 28 4 1 1 31 5 1 1 Transporte de hierro Transferrina <1 (0,2) <1 (0,2) Depósito de hierro Ferritina Hemosiderina 8 4 4 2 Total 50 40 Hierro funcional Hemoglobina (Hb) Mioglobina Enzimas Hem Enzimas no-Hem 35 Hierro las reticuloendoteliales. Alrededor del 80% del hierro presente en el plasma pasa a la médula eritroide para ser incorporado a los nuevos eritrocitos; normalmente, éstos circulan durante unos 120 días antes de ser catabolizados por el reticuloendotelio. En este momento, una fracción del hierro retorna de inmediato al plasma unido a la transferrina, mientras que otra fracción pasa a los depósitos de ferritina de las células reticuloendoteliales y regresa a la circulación en forma más gradual.1 1. Transporte Prácticamente todas las células y organismos requieren hierro para llevar a cabo los procesos celulares básicos. En la respiración, las proteínas del hierro capturan energía liberada de la oxidación de los alimentos sintetizando compuestos de alta energía, tales como el NADH, que son usados en el metabolismo celular. El hierro también permite que la hemoglobina de los eritrocitos una y transporte oxígeno a los tejidos por todo el cuerpo. Ya que el hierro es indispensable para la respiración y el transporte del oxígeno, no sorprende que la adquisición celular del hierro no haya sido dejada al azar. Ciertamente, pue- 36 de ser un verdadero desafío para los organismos obtener suficiente hierro debido a que mucho del hierro ambiental es muy insoluble. Por esto, se encuentran sistemas elaborados para la captación y distribución del hierro dentro de los organismos en todos los reinos vivos.2 Transportadores específicos del hierro están presentes en las células epiteliales que cubren el duodeno, y estos transportadores son mucho más abundantes en los animales deficientes de hierro. Los mamíferos – incluyendo a los humanos – tienen la capacidad de comunicar sus necesidades de hierro al duodeno.3,4 Los mecanismos moleculares por los cuales se detecta la necesidad de hierro y se activan respuestas que restauren el equilibrio o balance del hierro son materia de intensa investigación. La regulación anormal de la captación del hierro es la causa de la hemocromatosis, en la cual se capta hierro en exceso lo que conduce eventualmente a sobrecarga de hierro y hepatopatía, cardiopatía y daño en otros órganos.3,5 Una vez que el hierro ha cruzado las barreras celulares del duodeno, es liberado en la sangre y Hierro circula hacia los tejidos del cuerpo. Sin embargo, no es práctico que el hierro libre elemental circule porque el hierro puede unirse indiscriminadamente a muchas proteínas. Por lo tanto, el hierro es transportado en una forma no reactiva por una proteína transportadora específica, la transferrina.6 La mayoría del hierro que logra acceso a la sangre circulante se une estrechamente a la transferrina sérica, una proteína abundante que une uno (monoférrica) o dos átomos de hierro férrico (diférrica u holotransferrina) con alta afinidad. Cuando el hierro férrico se une a la transferrina, éste no es reactivo, lo que significa que no participa en la transferencia de electrones y no pone en riesgo, por decirlo así, a otras proteínas y las paredes de los vasos sanguíneos con su reactividad. Igualmente, como la transferrina en una proteína grande, permanece en la circulación en vez de excretarse en la orina al pasar por el riñón. De este modo, las células y tejidos que necesitan rellenar sus depósitos de hierro lo pueden hacer captando la transferrina que contiene hierro unido. Las células cumplen esta tarea sintetizando receptores de transferrina, proteí- nas que están presentes como pares (dímeros) sobre la superficie celular. Las células deficientes de hierro contienen muchos más receptores de transferrina que las que tienen sus reservas completas, lo que indica que las células tienen la capacidad de detectar sus propias necesidades de hierro e incrementar la síntesis del receptor de transferrina cuando consumen su hierro.7 2. Transferrina El nombre transferrina fue dado originalmente a la proteína sérica que une y transporta el hierro para distribuirlo a las células. Actualmente también se aplica a una familia más amplia de proteínas homólogas que incluyen transferina sérica (sTf), lactoferrina (Lf), ovotransferrina (oTf), y melanotransferrina (mTf).8,9 Únicamente la sTf tiene una función transportadora comprobada, pero todas las transferrinas controlan los niveles de hierro libre en donde se encuentran. Lf, por ejemplo, está presente en la leche, muchas otras secreciones exocrinas, y en los leucocitos, y une el hierro con incluso mayor intensidad que la sTf10 y lo retiene a un pH más bajo;11 su función parece 37 Hierro ser principalmente protectora.12 El papel de mTf, la cual se encuentra en formas unida a membrana y soluble,13 es incierto pero claramente se relaciona de alguna manera con el control del hierro. Todas estas proteínas son glucoproteínas de cadena simple, con 670– 690 residuos de aminoácidos y un peso molecular aproximado de 80 kDa.14 El sitio de unión está exquisitamente configurado para unir el hierro como Fe3+, esto se destaca por el hecho de que aunque la constante de unión para el Fe3+ es de aproximadamente 1020, la del Fe2+ es de sólo unos 103. El hierro es liberado en el contexto de la endocitosis mediada por receptor, en la cual la sTf cargada con hierro se une a los receptores de la superficie celular, es internalizada, libera su hierro dentro del endosoma, y es posteriormente retornada a la superficie para continuar sus rondas de unión y transporte.16,17 Al parecer, dos factores operan en la facilitación de la liberación del hierro. Primero, existe evidencia de que el receptor propiamente dicho puede actuar selectivamente sobre el sitio de unión. Segundo, el pH en el endosoma es cercano a 5.5, substancialmente 38 inferior al del pH extracelular de 7.4, y a este pH menor la sTf espontáneamente comienza a liberar el hierro.18 3. Hemopexina El heme (también conocido como hem o hemo), es usado en una amplia variedad de procesos biológicos, incluyendo la respiración y la transferencia de energía. El recambio de las proteínas heme, especialmente hemoglobina, conduce a la liberación del heme en los líquidos extracelulares, con potencialmente consecuencias severas para la salud. Así como el hierro libre, el heme es una fuente de hierro esencial para los patógenos bacterianos invasores19 y es altamente tóxico debido a su capacidad de catalizar la formación de radicales libres. Los niveles de heme usualmente son bajos pero pueden llegar a ser peligrosamente altos en condiciones de enfermedad hemolítica. La protección está dada por la hemopexina, una glucoproteína sérica de 60 kDa que captura al heme con una afinidad muy alta del torrente sanguíneo,20 lo transporta a receptores específicos sobre los hepatocitos, donde sufre endocitosis mediada por re- Hierro ceptor, y libera el heme unido en las células.21 Sirve por esto tanto para proteger contra la toxicidad del heme como para la conservación y reciclamiento del hierro. La hemopexina une una sola molécula de heme. El heme se une a dos residuos histidina. Como en el caso de la liberación del hierro de la transferrina, el heme es liberado de la hemopexina en el ambiente de pH bajo del endosoma, después de la endocitosis mediada por receptor.21 4. Almacenamiento El exceso de hierro se deposita o almacena intracelularmente como ferritina y hemosiderina, fundamentalmente en el sistema retículo-endotelial (SRE) del bazo, el hígado y la médula ósea. La molécula de apoferritina es un heteropolímero de 24 subunidades de 2 tipos diferentes: L y H, con un peso molecular de 20 kDa cada una, formadas por 4 cadenas helicoidales. Las variaciones en el contenido de subunidades que componen la molécula determinan la existencia de diferentes isoferritinas, las que se dividen en 2 grandes grupos: isoferritinas ácidas (ricas en cadenas H) localiza- das en el corazón, los glóbulos rojos, los linfocitos y los monocitos, y las isoferritinas básicas (ricas en cadenas L) predominantes en el hígado, el bazo, la placenta y los granulocitos. Las subunidades se organizan entre sí de manera tal que forman una estructura esférica que rodea a los cristales de hierro. Esta cubierta proteica posee en su entramado 6 poros de carácter hidrofílico y tamaño suficiente para permitir el paso de monosacáridos, flavinmononucleótidos, ácido ascórbico o desferroxamina. Se plantea que estos poros tienen una función catalizadora para la síntesis de los cristales de hierro y su incorporación al interior de la molécula de ferritina. La función fundamental de la ferritina es garantizar el depósito intracelular de hierro para su posterior utilización en la síntesis de las proteínas y enzimas. Este proceso implica la unión del hierro dentro de los canales de la cubierta proteica, seguido por la entrada y formación de un núcleo de hierro en el centro de la molécula. Una vez formado un pequeño núcleo de hierro sobre su superficie, puede ocurrir la oxidación de los restantes átomos del metal a medida que se incorporan. 39 Hierro Se han observado diferencias entre la velocidad de captación de hierro por las diferentes isoferritinas; así las isoferritinas ricas en cadenas H tienen una mayor velocidad de captación y se ha demostrado que ésta es precisamente la función de este tipo de subunidad. No obstante, las cadenas H y L cooperan en la captación del hierro, las subunidades H promueven la oxidación del hierro y las L, la formación del núcleo. Tanto el depósito de hierro como su liberación a la circulación son muy rápidos, e interviene en este último proceso el flavinmononucleótido. El hierro es liberado en forma ferrosa y convertido en férrico por la ceruloplasmina plasmática, para que sea captado por la transferrina que lo transporta y distribuye al resto del organismo.1 La hemosiderina está químicamente emparentada con la ferritina, de la que se diferencia por su insolubilidad en agua. Aunque ambas proteínas son inmunológicamente idénticas, la hemosiderina contiene un por ciento mayor de hierro (30 %) y en la microscopia se observa como agregados de moléculas de ferritina con una conformación diferente de los cristales de hierro.1 40 El volumen de las reservas de hierro es muy variable, pero generalmente se considera que un hombre adulto normal tiene entre 500 y 1 500 mg y una mujer entre 300 y 1 000 mg, aunque estos valores dependen notablemente del estado nutricional del individuo. 5. Regulación La vía fundamental de captación celular de hierro es la unión y subsecuente internalización de la transferrina cargada con hierro por su receptor. La cantidad de hierro que penetra a la célula por esta vía está relacionada con el número de receptores de transferrina presentes en la superficie celular. Una vez dentro, el hierro es utilizado para sus múltiples funciones o almacenado en forma de ferritina o hemosiderina. Por lo tanto, cuando las necesidades de hierro de la célula aumentan, se produce un incremento en la síntesis de receptores de transferrina y, en el caso contrario, cuando hay un exceso de hierro, ocurre un aumento de la síntesis de ferritina. Esto se logra mediante un estricto sistema de control al nivel postranscripcional. Tanto la expresión del receptor de transferrina como de la ferriti- Hierro na son reguladas en función de la disponibilidad y demanda de hierro para asegurar la homeostasia celular. En esta regulación está implicada una proteína citosólica de aproximadamente 98 kDa de peso molecular, altamente conservada a lo largo de la evolución, conocida como factor regulador de hierro (IRF) o proteína de unión al elemento de respuesta al hierro (IRE-BP). Esta proteína posee un centro 4Fe-4S que le permite cambiar entre 2 actividades diferentes en dependencia del nivel de hierro celular; así cuando los niveles de hierro son bajos, el centro se disocia y la apoproteína se une a una estructura tallo-lazo específica en el RNA mensajero (mRNA) del receptor de transferrina y de la ferritina, conocida como elemento de respuesta al hierro (IRE). Esta misma proteína se convierte en una aconitasa citosólica con un centro 4Fe-4S en células cargadas de hierro.1 Por su parte, la región 3´no traducida del mRNA del receptor de transferrina contiene 5 IREs; en este caso, la unión del IRF protege los mRNA de la degradación, con lo cual estimula la expresión del receptor. Cuando los niveles intracelulares de hierro están elevados, el IRF se disocia de los IREs, con lo que aumenta la traducción del mRNA de la ferritina y se acelera la degradación del mRNA de los receptores de transferrina. Así la interacción del IRF/IRE regula la expresión de estas proteínas en direcciones opuestas por 2 mecanismos diferentes, con lo cual se logra mantener el equilibrio entre la captación y almacenamiento intracelular del hierro. Mecanismos similares están implicados en la regulación de otras proteínas que participan en la distribución del hierro.1 Referencias Existe un IRE localizado cerca del extremo 5´terminal, de la región 5´no traducida de los mRNA de las cadenas L y H de la ferritina. La unión del IRF a este IRE inhibe la traducción del mRNA de la ferritina por interferencia en el orden de unión de los factores de iniciación de la traducción.1 1. Hillman RS. Hematopoietic agents: growth factors, minerals, and vitamins. In, Goodman and Gilman’s The Pharmacological basis of therapeutics, 10th ed. (Hardman JG, Limbird LE, eds.) McGraw-Hill, New York, 2001, p14871517 2. Rouault TA. How Mammals Acquire and Distribute Iron Needed for OxygenBased Metabolism.2003;1(3):326-328. 41 Hierro 3. Anderson GJ, Powell LW (2002) HFE and non-HFE hemochromatosis. Int J Hematol 76: 203–207. 4. Andrews NC (2002) Metal transporters and disease. Curr Opin Chem Biol 6: 181–186. 5. Ajioka RS, Kushner JP (2003) Clinical consequences of iron overload in hemochromatosis homozygotes. Blood 101: 3351–3353. 6. Baker HM, Anderson BF, Baker EN (2003) Dealing with iron: Common structural principles in proteins that transport iron and heme. Proc Natl Acad Sci U S A 100: 3579–3583. 7. Aisen P (1994) Iron metabolism: An evolutionary perspective. In: Brock JH, Halliday JW, Powell LW, editors. Iron metabolism in health and disease. London: W. B. Saunders. pp. 1–30. 8. Aisen, P. & Harris, D. C. (1989) in Iron Carriers and Iron Proteins, ed. Loehr, T. M. (VCH, New York), pp. 241–351. 9. Baker, E. N. (1994) Adv. Inorg. Chem. 41, 389–463. 10. Aisen, P. & Liebman, A. (1972) Biochim. Biophys. Acta 257, 314–323. 11. Baker, E. N., Baker, H. M. & Kidd, R. D. 42 (2002) Biochem. Cell Biol. 80, 27–34. 12. Sanchez, L., Calvo, M. & Brock, J. H. (1992) Arch. Dis. Child. 67, 657–661. 13. Sekyere, E., Food, M. R. & Richardson, D. R. (2002) FEBS Lett. 512, 350–352. 14. Baker HM, Anderson BF, Baker EN. Dealing with iron: Common structural principles in proteins that transport iron and heme. 2003; 100( 7): 3579–3583. 15. Harris, W. R. (1986) J. Inorg. Biochem. 27, 41–52. 16. Klausner, R. D., Ashwell, J. V., Van Renswoude, J. B., Harford, J. & Bridges, K. (1983) Proc. Natl. Acad. Sci. USA 80, 2263–2267. 17. Octave, J.-N., Schneider, Y.-J., Trouet, A. & Crichton, R. R. (1983) Trends Biochem. Sci. 8, 217–220. 18. He, Q.-Y. & Mason, A. B. (2002) in Molecular and Cellular Iron Transport, ed. Templeton, D. M. (Dekker, New York), pp. 95–123. 19. Lee, B. C. (1995) Mol. Microbiol. 18, 383–390. 20. Hrkal, Z., Vodrazka, Z.&Kalousek, I. (1974) Eur. J. Biochem. 43, 73–78. 21. Smith, A.&Hunt, R. T. (1990) Eur. J. Cell Biol. 53, 234–245. Hierro Hernán Pérez C omo se sabe el hierro es un elemento esencial para la vida puesto que participa en casi todos los procesos de oxido-reducción. Forma parte importante del ciclo de Krebs, de la respiración celular y participa en el trasporte de electrones en los citocromos celulares. Está presente en diferentes enzimas relacionadas con el mantenimiento de la integridad celular como son las catalasas, perioxidasas y oxigenasas(1). El hierro es controlado por un importante sistema regulador debido a su alto potencial redox y su facilidad para estimular la formación de sustancias muy tóxicas y reactivas. El hombre es un organismo aeróbico, es decir que requiere oxígeno para vivir. Nuestro organismo cuenta con hemoglobina para el transporte de O2, desde el medio externo hasta el medio interno, de allí pasa a cada célula para concluir el proceso de respiración celular. El hierro es el constituyente principal, el núcleo de la molécula de hemo- globina y también de la míoglobina, la cual transporta al oxígeno dentro de la célula muscular, hasta el interior de las mitocondrias. Es un oligoelemento, un individuo de 70 Kg de peso tiene solo unos cuatro gramos de hierro. El hierro también actúa como activador de numerosas enzimas. Generalmente la hemoglobina tiene una vida de 120 días. Pero el hierro no se pierde, es enviado de nuevo a la médula ósea para fabricar nueva hemoglobina, mientras que el resto de esta molécula es convertido en bilis y enviado al hígado para completar el proceso y eliminarla por la vía digestiva, convirtiéndose en parte de la materia fecal. Eventualmente el hígado puede actuar como reservorio de las moléculas de hierro. El hierro forma parte de dos compartimientos en el organismo: A. funcional que incluye la hemoglobina, mioglobina, transferrina y las enzimas que requieren hierro como cofactor o como grupo prostético en forma iónica o como 43 Hierro Enzimas Mioglobina Hierro de depósito (20%) Ferritina Hemosiderina Hierro de transporte (0,1 - 0,2 %) transferina Hemoglobina Hierro activo 80 % Enzimas Hemoglobina 65 % Catalasas Mioglobina 10 % Peroxidasas 5% Citocromos FIgura 1. Contenido total de hierro en un individuo normal (Forrellat M y Cols Rev Cubana Hematol Inmunol Hemoter 2000;16(3)149 grupo hemo. B: depósito conformado por la ferritina y la hemosiderina que forman las reservas orgánicas del metal. El paso de hierro entre estos dos compartimientos se realiza mediante un proceso específico y eficiente. El hierro se puede considerar como un metal que se requiere en cantidades relativamente pequeñas como un cofactor para muchas enzimas, pero en concentraciones más altas en la hemoglo44 bina hace que los requerimientos del organismo se alcancen con mayor dificultad y el desbalance sea más frecuente. El contenido normal de hierro en la mujer es de 3,5 gramos en la mujer y de 4 a 5 gramos en el hombre(2) En personas con nutrición normal aproximadamente el 65% del metal forma parte de la hemoglobina, 10% forma parte de la mioglobina, de las enzimas el 5% y como hierro de depósito el 20%; menos del 1% se une a la transferrina como hie- Hierro Enzimas 5 mg/día Mioglobina 2.700 mg 24 mg/día Hemoglobina 1 - 2 mg/día Intestino 1.000 mg Depósito (ferritina) Pérdida 1 - 2 mg/día Transporte de hierro (transferrina) CINÉTICA DEL HIERRO Figura 2. El hierro es permanentemente conservado y reciclado para utilizarlo en enzimas hem y no-hem. rro de circulante(3). De la cantidad diaria de hierro que se moviliza solo se pierde una pequeña cantidad a través de las heces, orina y sudor. La recuperación de la perdida se realiza a través de la ingestión de alimentos a pesar que esta es muy baja. El adulto normal contiene 2 gramos de hierro en su hemoglobina que de acuerdo con la vida media de los eritrocitos se ceden a los fagocitos del sistema retículo endotelial (SRE) los cuales en cantidad un poco diferente son eliminados diariamente en hombres y mujeres. El sistema retículo endotelial también recibe un sobrante de hierro de origen eritropoyético. Del hierro contenido en el SER una pequeña parte está en equilibrio con el compartimiento de depósito y la transferrina lo transporta 45 Hierro hasta la médula ósea para la síntesis de hemoglobina. La médula requiere una cantidad diaria de 25 mg de los cuales 92% provienen del SER y solo 8% se originan en absorción intestinal. Se considera que aproximadamente 7 mg mantienen en equilibrio entre la circulación y los depósitos señalados(4). Solamente un porcentaje pequeño de hierro total diario ingresa o sale de los depósitos de acuerdo con su concentración. Por tanto el transporte intercelular de hierro como parte de un proceso de reutilización es cuantitativamente más importante que la absorción intestinal. La concentración más importante de hierro se encuentra en los eritrocitos ya que estas células contienen cerca del 80% de la cantidad total del organismo. El SER recicla una cantidad importante de hierro de las células rojas envejecidas cercana a la cantidad que utiliza el eritrocito para la nueva producción de hemoglobina. El hierro es un metal de transición y su utilidad biológica depende de su capacidad para existir en diversos estados de oxidación y para formar diferentes complejos. Las sales ferrosas son relativamente solubles a pH 7 pero en 46 presencia de oxígeno son rápidamente oxidados a la casi insoluble forma férrica. La química biológica del hierro es casi tan amplia como la de sus quelatos (la mayoría como ligandos de proteínas) A pesar de la pobre disponibilidad de hierro libre o sea el que no está unido a proteínas u otros quelantes fuertes este metal es tóxico debido a su capacidad como catalizador de reacciones de radicales libres los cuales pueden producir lesiones celulares y titulares(5). El primer aspecto a tener en cuenta en la farmacocinética del hierro es su difusión a través de las membranas en su forma ferrosa y su almacenamiento y transporte en forma férrica. El organismo tiene dos clases de reservas de hierro, el esencial y el exceso que se mantiene en depósitos. La fracción esencial esta dominada por la hemoglobina, proteína que contiene cuatro átomos de hierro por molécula. Otra forma de hierro esencial es la mioglobina y otras enzimas dependientes de hierro hem (derivado porfirínico que contiene hierro) y no hem. La ferritina es un depósito soluble de hierro intracelular Hierro y la apoferritina es la proteína de almacenamiento de hierro depositado en las células de la mucosa intestinal y como macrófago en hígado, bazo y huesos. La apoferritina, está constituida por 24 subunidades de polipéptidos con un peso molecular de 450.000 y se sintetiza de acuerdo con las concentraciones de hierro libre. La ferritina y la apoferritina forman un depósito para el óxido férrico fosfatado e hidratado polinuclear. Cuando las concentraciones de apoferritina son bajas se inhibe y se equilibra en el momento en el que el hierro fijado se transforma en transferrina. Por el contrario si las concentraciones son altas se produce apoferritina para asegurar la eliminación del exceso de hierro y proteger los órganos de efectos tóxicos. Más del 30% de la ferritina puede ser hierro, la ferritina agregada denominada hemosiderina constituye cerca del 33% de las reservas normales y aumenta de acuerdo como lo hacen las reservas. Los sitios de almacenamiento de hierro son el sistema retículo endotelial y los hepatocitos. Como la ferritina que se localiza en el plasma está en equilibrio con la almacenada en esos sitios la con- centración sérica de ferritina plasmática se puede utilizar para valorar las reservas totales de hierro en el organismo. 1. Proteínas reguladoras del metabolismo del hierro La transferrina es la proteína transportadora de hierro en plasma y líquidos extravasculares (6)Es una glicoproteína con PM de 80 kDa sintetizada en el hígado que posee dos dominios homólogos de fijación para el hierro férrico (Fe3+) es un polipéptido simple constituido molecularmente por un dominio N-terminal y un dominio C-terminal que tienen sitios de unión al hierro, pero dos cadenas de olisacáridos de unión se encuentran unidos al dominio C-terminal. La molécula se relaciona con otras proteínas la ovotransferrina y la melanotransferrina producida por las células de melanoma y la lactoferrina. La lactoferrina tiene una estructura similar a la transferrina, se secreta en la leche y se sintetiza en granulocitos. Estudios recientes indican que la lactoferrina puede actuar como sitio especifico de unión del DNA y podría intervenir en la activación del proceso de transcripción (7)La afinidad de la transferrina por el hierro es muy alta, pero esta unión 47 Hierro RECEPTOR DE TRANSFERRINA Fe Fe Fe Fe Tr Tr Tr= Transferrina Tripsina Membrana Plasmática PO4 - Ser Ser - PO4 Figura 3. Representación esquemática del receptor de transferrina (Worwood M Annals Nestle 1995;53:30) requiere de la participación de un anión. Algunos estudios sugieren que los residuos de tirosina e histidina se relacionan con la unión del hierro o histidina y arginina en la unión aniónica (5) Los genes de transferrina, lactoferrina, melanotransferrina y el receptor de transferrina se encuentran en el cromosoma. (8). Los mecanismos exactos de la incorporación del hierro a la transferrina y su paso a las células del epitelio intestinal o del sistema retículoendotelial son desconocidos. La ferroxidasa dependiente 48 del cobre puede jugar papel importante. Ciertas evidencias indican que la proteína podría estar relacionada con la movilización de hierro de los depósitos titulares para producir transferrina diférrica. (9) 1.1. Receptor de Transferrina A pesar que Jandl JH y Katz JH plantearon la existencia de un receptor específico para la transferrina (10) solo hasta 1984 se logró la clonación de su receptor. Recientemente se clonó el receptor 2 (TfR2-alfa), (11). Hierro CICLO DE LA TRANSFERRINA Espacio Extracelular (pH 7.4) Fe2 TF Transportador de hierro ferroso (DMT1) Apo= TF TFR Membrana celular Endosoma Bomba de protones H- H+ Citoplasma Fe Liberado Mitocondria DMT Ferritina Células no eritroides Hemosiderina Endosoma acidifado pH 5.6 Figura 4. en la grafica se muestra la tranferrina cargada de hierro(Fe2Tf) unida a receptores de transferrina(TfR) sobre la superficie de células eritroides (Andrews N Disorders of Iron Metabolism N. England J Med.2000;342:364) La estructura primaria del receptor TfR2-alfa es similar al receptor Tf1 sus dominios extracelulares son considerablemente homólogos y su expresión es alta en hígado y células eritroides. El receptor de Tf es una glicoproteína constituida por dos subunidades unidas por un puente disulfuro simple. La molécula se extiende a través de la membrana celular y tiene una cola citoplasmática. El receptor plasmático soluble de transferrina detectable en circulación carece de los dominio citosólico y transmembrana del monómero. (12) La molécula receptora puede unir dos moléculas de transferrina. Si la molécula de transferrina es diférrica (ejem: si está unida a dos moléculas de hierro) la molécula receptora de transferrina puede transportar a través de la membrana cuatro átomos de hierro unidos a transferrina. La afinidad del receptor de transferrina por la transferrina depende del conteni49 Hierro do de hierro de la transferrina y del pH. El receptor tiene mayor afinidad con la transferrina de dos átomos de hierro que con la que tiene uno solo; la afinidad de este receptor por la apotransferrina es muy baja ya que esta no tiene nada de hierro. La concentración del receptor circulante depende del grado de eritropoyesis e incrementa cuando hay deficiencia de hierro. El receptor de transferrina tiene papel muy importante en el suministro de hierro a la célula ya que la afinidad del receptor por el complejo hierro-transferrina al pH alcalino de la sangre depende de la carga de hierro a la proteína. El complejo hierro-transferrinareceptor difunde a la célula mediante proceso de endocitosis. El paso de un pH alcalino a un pH ácido del endosoma induce cambios en la estabilidad del complejo lo que causa disociación espontánea de los átomos de hierro. La transferrina se mantiene unida al receptor hasta un nuevo cambio de pH, por el contrario en la membrana produce ruptura del complejo y liberación de transferrina que queda disponible de nuevo para la captación y transporte del hierro circulante. La liberación intracelular del hierro unido a la transferrina es secuencial. La primera molécula es liberada por el pH ácido del citosol, pero la segunda requiere ATP para su liberación. Las células humanas modulan la expresión de los receptores de transferrina y ferritina intracelular DISTRIBUCIÓN Y CINETICA DEL HIERRO COMPARTIMIENTO Hemoglobina Mioglobina Enzima Heme Enzima no Heme Depósito intracelular(ferritina) Hierro lábil intracelular (H.Q Transporte intercelular (transferrina) 50 HIERRO (gramos) TOTAL (%) 2.7 0.2 0.008 < 0.0001 1.0 0.07 0.003 66 3 0.1 —— 30 1 0.1 Hierro en respuesta a la administración de hierro. Cuando las concentraciones de hierro son altas disminuye la síntesis de receptores de transferrina e incrementa la producción de ferritina. Por el contrario en casos de deficiencia de hierro, las células expresan un mayor número de receptores de transferrina y disminuyen las concentraciones de ferritina para aumentar la captación y prevenir la movilización de hierro a los depósitos. El aislamiento de genes para el receptor de transferrina y ferritina humana ha permitido un mejor conocimiento de las bases moleculares de su regulación. 2. Fisiología transferrina/hierro La suma de todos los sitios de fijación de hierro a la transferrina constituye la capacidad total de unión de hierro al plasma. Normalmente cerca de una tercera parte de la transferrina unida al hierro se saturan. Por tanto con excepción de lo que ocurre en casos de sobrecarga de hierro en los que todos los sitios de fijación de la transferrina están ocupados, en la circulación prácticamente no hay hierro no fijado a transferrina. La distribución plasmática y tisular del hierro se puede investigar utilizando marcadores radioactivos (13). El sujeto recibe transferrina autóloga marcada con hierro radioactivo que puede ser monitoreado y las muestras de sangre se pueden analizar en ciertos intervalos de tiempo para determinar la velocidad de pérdida de marcador radioactivo. Los diversos estudios ferro cinéticos indican que la vida media normal del hierro en la circulación es cercana a los 75 minutos (14) La cantidad absoluta de hierro liberado de la transferrina por unidad de tiempo corresponde al volumen de recambio plasmático de hierro. Los estudios con trazadores radioactivos indican que por lo menos 8% del hierro unido a la transferrina circulante es liberado a la médula ósea e incorporado en los eritrocitos recién formados. Otros sitios importantes de liberación de hierro son: hígado el cual es un depósito primario para el hierro y el bazo. El hierro hepático se encuentra tanto en las células reticuloendoteliales como en los hepatocitos. Las células del SRE toman hierro principalmente por fagocitosis y por ruptura de células rojas viejas y estas células extraen el hierro de hem y vuelven a la circulación unidas a la transferrina. Los hepatocitos captan el hierro por dos vías diferen- 51 Hierro tes: la primera por endocitosis de transferrina mediada por receptor y adicionalmente pueden captar hierro iónico por un proceso independiente de la transferrina(15). 2.1. Ferritina Esta proteína es un importante depósito soluble de hierro dentro de la célula (16) La estructura puede considerarse como una concha de apoproteína constituida por 24 unidades de dos clases: H (heavy) y L (Light). La subunidad L contiene 174 aminoácidos y la subunidad H tiene 182 aminoácidos.La ferritina en tejidos humanos tiene las dos subunidades en diferentes porcentajes. Las isoferritinas ricas en subunidades H son más ácidas y se localizan en corazón, células rojas, linfocitos y monocitos. Las isoferritinas ricas en subunidades L son más básicas y predominan en hígado, bazo, placenta y granulocitos. La subunidad de ferritina tiene cinco hélices y una larga estructura interhelicoidal. Dicha estructura y el residuo N-terminal se encuentran en el exterior de la proteína y con un C-terminal dentro de la concha. Las dos cadenas H y L tienen igual conformación (17). 52 La función más importante de la ferritina es como depósito intracelular de hierro que puede ser utilizado para la síntesis de proteínas y enzimas que contienen hierro. El proceso involucra la unión inicial a sitios dentro de los canales de estas proteínas seguida de entrada a la proteína y formación de un depósito de hierro. Cuando pequeños núcleos de hierro se han reducido se produce una oxidación directamente en la superficie del depósito de hierro (17) Se han observado diferencias en la velocidad de recaptación de hierro con proporciones variables de las subunidades H y L con isoferritinas ricas en H con alta velocidad de recaptación de hierro (18). Con la disponibilidad de ferritinas recombinantes, se ha demostrado que la recaptación de hierro es función de la subunidad H; sin embargo las cadenas H y L tienen propiedades que participan en la captación de hierro, las subunidades H promueven la oxidación del Fe y las subunidades L estimulan la nucleación (19). Parece que las ferritinas ricas en subunidades L como las que están en hígado o bazo juegan papel importante en los depósitos de Hierro proteínas. Además hay evidencias que la sobrecarga de hierro causa incremento en el porcentaje de subunidades L y la deficiencia de hierro incremento en la proporción de subunidades H. Se han encontrado isoferritinas ricas en H en células con grandes requerimientos de hierro para síntesis hem como las células rojas nucleadas o células del músculo cardíaco, pero también tienen funciones que no tienen relación con fijación del hierro. La capacidad de isoferritinas ricas en H para inhibir hematopoyesis se ha investigado extensamente (20). Posiblemente estas funciones están relacionadas con interferencia en la captación de hierro por las células. Pequeñas cantidades de ferritina circulante y su concentración plasmática se relacionan con los niveles de hierro depositados en el organismo(21) Esta proteína circulante curiosamente es rica en subunidades L, tiene poco hierro y contiene moléculas glicosiladas. Lo anterior puede indicar que algunas de estas ferritinas son secretadas más no liberadas por células muertas o tejido lesionado, aunque no hay evidencia de un gen específico que regule dicha secreción. 2.2. Ferrocinética y médula ósea Debido al papel tan importante de la médula ósea en el aclaramiento del hierro marcado de la circulación, la farmacocinética permite observar su actividad eritropoyética. Aquellos factores que aumentan la producción de eritrocitos incrementan el recambio plasmático de hierro (PIT) Enfermedades del tipo anemias hemolíticas como la esferocitosis hereditaria y la de células peludas inducen rápida liberación de hierro a la médula ósea. Por el contrario trastornos relacionados con reducción en la producción de células rojas prolongan el PIT. Cuando los eritrocitos se producen y liberan en la circulación en forma normal la eritropoyesis se conoce como “efectiva” Sin embargo pacientes con ciertas anemias hemolíticas las células rojas nacientes consideradas anormales son destruidas antes que salgan de la cavidad medular. En estas condiciones la eritropoyesis es ineficaz lo cual significa que los precursores eritropoyéticos han fallado en su objetivo primario como es la liberación de eritrocitos intactos a la circulación. El perfil ferrocinético en tales casos muestra una rápida remoción de 53 Hierro hierro de la transferrina con una entrada demorada del marcador en el pool de hemoglobina de células rojas circulantes. La beta-talasemia es ejemplo importante de esta clase de anemia hemolítica con eritropoyesis ineficaz. En esta enfermedad la alteración de la eritropoyesis se relaciona con aumento marcado del PIT. unirse a proteínas de bajo peso molecular o aminoácidos, nucleótidos, ciertos azúcares y grupos sulfhidrilos. La reincorporación del hierro a la circulación requiere de una reoxidación, proceso en el que participan enzimas llamadas ferroxidasas 1 y 2 que contienen cobre. Por lo anterior la deficiencia de cobre efectos como disminución de hierro plasmático y anemia por deficiencia de hierro. 2.3. Captación celular del hierro Todos los tejidos y células tienen receptor específico para la transferrina que se expresa en la superficie celular regulando la captación de hierro de acuerdo a las necesidades. La mayor concentración de receptores se localiza en los eritroblastos en los cuales el hierro es captado por las mitocondrias y es incluido en las moléculas de protoporfirina durante la síntesis del grupo hemo. A medida que madura el glóbulo rojo el número de receptores disminuye debido a que las necesidades de hierro para la síntesis de hemoglobina son cada vez menores. 54 El Hem grupo prostético de numerosas proteínas que contienen hierro, se sintetiza en las mitocondrias a partir del hierro ferroso y de la protoporfirina IX. El hierro de la ferritina también puede ser utilizado para la síntesis de Hem. La disponibilidad de hierro regula la síntesis de Hem y de hemoproteinas en la mitocondria, ya que estas son necesarias para el transporte y depósito de O2 en la hemoglobina y mioglobina, transporte de electrones (citocromos) y diferentes reacciones de oxidación (citocromo-oxidasas, citocromo P450). 2.4. Papel del hierro intracelular 2.5. Captación de hierro no unido a transferrina A nivel intracelular el hierro al estado ferroso es liberado de la transferían formando complejos al A pesar que existen evidencias que el ciclo de la transferrina es importante para la captación de Hierro hierro por el eritron (22) hay tejidos que pueden tomar hierro por mecanismos alternativos. Algunos pacientes y especies mutantes tienen poca o carecen de transferrina circulante (23) A pesar del desarrollo de anemia hipocrómica, microcítica, los tejidos no eritroides son aparentemente normales. Cuando las células rojas presentan deficiencia de hierro, los niveles de hierro sérico (hierro no unido a transferrina) están elevados y el exceso de hierro es depositado en el hígado. Según Ponka y Schulman (22)las células no eri- troides dependen menos de la transferrina debido a que sus modestas necesidades de hierro pueden ser contrarestadas por el recambio de ferritina endógena y de hierro hem. Las células rojas son más vulnerables debido a la mayor utilización de hierro para formar hemoglobina (Ponka) El ciclo de la transferrina puede ser utilizado para incrementar la captación de hierro por tejidos de gran demanda de este elemento. La sobrecarga de hierro produce saturación total de transferri- CÉLULAS GERMINATIVAS HEMATOPOYÉTICAS Célula embrionaria pluripotente CFU - GEMM BFU - E CFU - GM Célula embrionaria linfoide CFU - Eo CFU - Baso CFU - Meg CFU - E Neutrófilo Monocito Eosinófilo Basófilo Célula T Célula B Plaqueta Eritrocito Figura 5. Esquema de la maduración de las células germinativas hematopoyeticas (Christensen RD Ped in Review 1991;244:12) 55 Hierro na y de hierro circulante no unido a transferrina de bajo peso molecular(24) Este hierro está ligeramente unido a albúmina, citrato, aminoácidos y azucares y se comporta en forma diferente del hierro unido a transferrina. Tejidos no hematopoyéticos especialmente el hígado, órganos endocrinos, riñones y corazón captan preferencialmente este hierro. Hierro marcado con o sin capacidad de fijación a transferrina administrado en animales presenta patrones de distribución diferentes (25) En animales normales los tejidos hematopoyéticos son los primeros sitios de captación. Cuando los sitios de transferrina libre no existen, la mayoría del hierro se deposita en hígado y páncreas lo cual indica que estos órganos son utilizados como reservorios en casos de sobrecarga de hierro. Esta forma de distribución es similar a la que se observa en la hemocromatosis idiopática. Estas observaciones respaldan la idea de que las vías de la transferrina son importantes para llenar las necesidades del eritrocito, pero no son esenciales para la captación de hierro por todos los tejidos. 56 Kaplan y cols (26) investigaron la incorporación del hierro del citrato FeNH 4 observando que la captación independiente de transferrina aumenta en proporción directa con la concentración de este compuesto en forma similar a la captación hepática de hierro no unido a transferrina en pacientes con saturación de transferrina. Los autores piensan que se trata de una vía de protección alternativa que permite remover el metal tóxico de la circulación. Randell y cols 27) han descrito captación similar en células HepG2 y demuestran que esto es reversible por adición de compuestos quelantes. Inman y Wessling y Resnick (15)describen un mecanismo de captación de hierro no unido a transferrina en células eritroleucémicas con características diferentes. En ausencia de transferrina férrica la captación de hierro por células K562 es muy sensible al tratamiento con tripsina lo que sugiere que se requiere de una proteína acarreadora. Medios con altas concentraciones de hierro no aumentan la captación celular del metal. Según Pollack (28) este transporte puede ser acompañado por el mismo mecanismo responsable del Hierro paso de hierro al exterior de los endosomas en el citoplasma. Estos dos procesos cumplen la misma función. El transporte de hierro endosomal se puede orientar a transportar hierro de un compartimiento extracelular endositosado dentro del citoplasma. Este tipo de transporte puede observarse en la superficie celular previamente a la endocitosis mediada por receptor con capacidad para transportar hierro en cantidad moderada. Esta actividad no está restringida a células eritroides. De acuerdo con los estudios de Hamazaki y Glass (29) los linfocitos periféricos humanos PHA-estimulados poseen mecanismos similares para la captación de hierro no dependiente de transferrina. 3. Eliminacion del hierro No existe un mecanismo específico para la eliminación del hierro. Se pierden pequeñas cantidades por la exfoliación de las células de la mucosa intestinal, en las heces, y se pierden cantidades muy limitadas en bilis, orina y sudor. Estas pérdidas pueden no ser superiores a 1 miligramo de hierro por día. Debido a que la capacidad del organismo para eliminar el hierro es muy limitada, el mayor porcentaje de eliminación se produce por descamación de células que contienen hierro en la mucosa gastrointestinal y en la epidermis. Durante el embarazo se presenta una situación especial ya que en este periodo la madre cede al feto y a los tejidos placentarios el hierro necesario para su desarrollo normal (30). También hay pérdida importante de hierro por sangrado durante el parto y durante la lactancia. Se considera que 300 mg son los requerimientos del feto durante los nueve meses, la placenta y el cordón umbilical 100 mg. Durante el parto se pierden 100 mg y durante seis meses de lactancia 100 mg se pierden también 100 mg de hierro. Lo anterior suma 600 mg como pérdida total de hierro durante el embarazo que requiere de tratamiento especifico para no generar deficiencia de dicho elemento (30) Bothwell y Cols han revisado estudios muy extensos utilizando marcadores de hierro con 55Fe (31) En hombres blancos americanos, surafricanos de origen indio y mestizos venezolanos el promedio diario de eliminación de hierro varia solo entre 0.95 y 1.02 57 Hierro mg. La pérdida estimada de piel, gastrointestinal y pérdidas urinarias son de 0.2, 0.3, 0.6 y 0.1 mg respectivamente y la suma de estos componentes individuales es similar a la pérdida total estimada por marcadores del hierro corporal total. En mujeres premenopáusicas la pérdida es cercana a 1.6 mg/día y esta coincide con los resultados de dos amplios estudios en los que se demuestra que la pérdida promedio mensual de sangre es aproximadamente de 0.5-0.6 mg/día (31). El hierro de depósito en el organismo sufre solo muy pequeños cambios en su eliminación. En sujetos con deficiencia de hierro se eliminan entre 0.5 mg/día y en sujetos con sobrecarga se pierden 1.5 mg/día. En infantes y jóvenes se han realizado pocos estudios, pero Bothwell y cols que la pérdida de hierro gastrointestinal puede ser mayor que en el adulto. Otros factores importantes de la pérdida de hierro son: la donación de sangre, infecciones parasitarias y las microhemorragias producidas por medicamentos como los AINE (31). 4. Hierro y sistema hematopoyetico Las células sanguíneas del adulto se forman con excepción de los 58 linfocitos, en la médula ósea. Durante el ciclo fetal la producción celular se inicia en el saco vitelino y posteriormente en el hígado y el bazo. Al quinto mes de gestación se inicia la hematopoyesis en la médula ósea que van a reemplazar a las células mencionadas. Al nacimiento la hematopoyesis esplénica y hepática desaparecen, pero puede reaparecer durante los primeros años de vida cuando hay aumento en la demanda de células sanguíneas. Al nacer el tejido hematopoyético ocupa casi todo el tejido óseo y este disminuye gradualmente hasta que en el adulto normal se distribuye solo en huesos planos como son las vértebras, esternón, costillas y pelvis. La función más importante de la médula ósea es la producción de células sanguíneas diferenciadas que son derivadas de células troncales multipotenciales con capacidad de renovación y con capacidad de diferenciación. Dicha capacidad facilita la producción de células unipotenciales y originan líneas celulares: eritropoyéticas, granulopoyéticas y trombopoyéticas. Las células unipotenciales están en una etapa activa del ciclo celular con capacidad de autorenovación y proliferación por Hierro tiempo prolongado. Estas células necesitan estímulo humoral de ciertas sustancias con capacidad hematopoyética. La sustancia más conocida es la eritropoyetina que es sintetizada por el riñón en respuesta a la hipoxemia y que estimula la producción de eritrocitos. Se sabe también de la existencia de otras sustancias como la leucopoyetinas y trombopoyetinas. Las enfermedades que comprometen a la médula ósea alterando la producción normal de células sanguíneas, se pueden relacionar con falla primaria de las células troncales, sustitución de tejido medular por tejido anormal o con alteración de factores estimulantes. Como se sabe la producción de eritrocitos es estimulada por la eritropoyetina. El proceso de maduración de los precursores de la línea roja en la médula ósea se caracteriza por la producción progresiva de hemoglobina en el interior de las células y por condensación del núcleo hasta su estrusión que se produce en el tiempo inmediatamente anterior a que el eritrocito se libere a la circulación. La hemoglobina se sintetiza durante el proceso de maduración PAPEL REGULADOR DEL RIÑON EN LA ERITROPOYESIS Célula mader CFU GERM.M BFU - E CFU - B Proeritroblasto E. Basófilo E. Policromatófilo E. Ortocrómico Reticulocito Eritrocito EPO O2 MO: médula ósea EPO: eritropoyetina Figura 6. Regulación renal de la eritropoyesis 59 Hierro de los eritrocitos y para su formación es fundamental la disponibilidad de hierro que es componente importante de la molécula. El hierro es entregado a las células precursoras por la proteína específica de transporte como es la transferrina e insertado en el anillo de la porfirina para formar el grupo heme. Como ya se señaló el hierro no utilizado para la síntesis de hemoglobina se almacena como depósito acoplado a la proteína de almacenamiento que es la ferritina. Para cumplir con la función de transporte de oxígeno a los tejidos, la hemoglobina debe combinarse en forma reversible con este captándolo a nivel pulmonar y liberándolo a los tejidos. La afinidad de la hemoglobina por el oxígeno puede cambiar lo cual se relaciona con mayor o menos facilidad para entregarlo a los tejidos. La membrana del eritrocito es igual a la de otras células pero se diferencia en su gran capacidad para el intercambio de gases y capacidad de deformación que le facilita el paso a través de los capilares. En la membrana del eritrocito se expresan sus antígenos de los cuales se han identificado más de 100, pero los más impor- 60 tantes en la práctica son los del sistema ABO y Rh. La ausencia del sistema ABO en un individuo determina la presencia del anticuerpo correspondiente en plasma (anti-A o antiB) lo que se considera de interés crítico en una transfusión sanguínea. El desarrollo de anticuerpos para los otros sistemas se produce solamente cuando alguien que carece del antígeno se expone a eritrocitos que lo contienen. Lo anterior permite explicar la enfermedad hemolítica del recién nacido cuya madre se sensibiliza a antígenos presentes en el feto de los cuales ella carece. Estos anticuerpos pueden cruzar la placenta y destruir los eritrocitos fetales. La deficiencia de diferentes elementos de la dieta como el hierro, vitamina B12, ácido fólico y proteínas pueden causar disminución de la producción de eritrocitos, lo cual implica reducción de su número en sangre periférica. Debido a que la deficiencia de hierro o los trastornos en su utilización son las causas más frecuentes de anemia es importante conocer muy bien su metabolismo. El hierro es un elemento esencial de la molécula de hemoglobi- Hierro na por tanto cualquier disminución del aporte por debajo de los requerimientos mínimos puede causar reducción en la síntesis de hemoglobina. La mayor concentración de hierro en el organismo se localiza en la hemoglobina o almacenado como ferritina o hemosiderina en el sistema retículo-endotelial (SER). El balance de hierro está determinado por la cantidad absorbida, por los requerimientos y las pérdidas. El hierro tiene importantes funciones en diferentes procesos celulares y sistémicos relacionados con: 1. Transporte y liberación de O2 a los tejidos como elemento esencial de la hemoglobina del eritrocito. 2. Síntesis de hemoproteinas como mioglobina molecular relacionadas con funciones específicas de depósitos. 3. Regulación de la síntesis mitocondrial del hem. 4. Utilización celular de O2 como componente de citocromos y oxidasas mitocondriales como el citocromo P450, citocromooxidasa, catalasas, peroxidasas. 5. Regulación de diferentes funciones metabólicas formando parte de flavoproteinas con hierro no hem, como oxidasas, deshidrogenasas, glicerofosfato deshidrogenasa, y NADH deshidrogenasa. También como componente de la ribonucleótido reductasa lo cual explica la importancia del hierro en la síntesis de ácidos nucleicos. Si la deficiencia de hierro es severa afecta en primer lugar la producción de glóbulos rojos con el desarrollo de una anemia con características propias, la anemia ferropénica. Esta patología carencial determina la iniciación de terapia férrica y su corrección. Si la anemia inicial no se corrige y se mantiene la pérdida de hierro en el tiempo, se alteran las otras funciones celulares enzimáticos y metabólicas del metal y por tanto el impacto en la salud es más grave que la misma anemia. La carencia crónica de hierro afecta el metabolismo muscular, independientemente de la hipoxia tisular por la anemia y produce importantes alteraciones en SNC. Si la deficiencia de hierro se presenta en los niños con el tiempo pueden ocurrir trastornos cognoscitivos, del aprendizaje, alteraciones de la memoria, problemas con- 61 Hierro ductuales y de comportamiento. Adicionalmente se pueden presentar síntomas neurológicos, como cefaleas, irritabilidad y alteraciones en el metabolismo de las catecolaminas y funciones dopaminérgicas en SNC. Otros problemas relacionados con la deficiencia crónica de hierro comprometen al tejido epitelial con la producción de koiloniquia, estomatitis angular y atrofia papilar de la lengua. En casos de deficiencia de hierro se altera también la regulación de la temperatura corporal y la producción de calor, problemas relacionados posiblemente con trastorno del metabolismo hormonal tiroideo en especial con la triiodotironina. Por tanto disminuye la capacidad de trabajo y ello se acompaña de debilidad y fatiga. Estos fenómenos se han relacionado con enzimas mitocondriales dependientes de hierro como son ciertas deshidrogenadas y otras que afectan funciones mitocondriales que son Fe-dependientes. 4.1. Eritropoyesis Es el proceso de formación de los eritrocitos (Figuras 5 y 6) a partir de las células madre, llamadas 62 también pluripotentes debido a su capacidad para producir todas las células de la sangre. Algunas de estas se diferencian hacia la primera célula de la línea eritroide que es la BFU-E y posteriormente la CFU-E células que se identifican en cultivos celulares debido a sus características de crecimiento. La BFU-E forma grandes colonias de eritrocitos maduros después de 2-4 semanas de cultivo; la CFU-E produce pequeñas colonias a los 7 días de cultivo in vitro. Una vez madura la CFU se forma el proeritroblasto que es primer precursor que se reconoce morfológicamente en la médula ósea. En las siguientes divisiones aparecen precursores más pequeños con núcleos progresivamente más pequeños, de cromatina más condensada y con mayor cantidad de hemoglobina en el citoplasma, razón por la que su apariencia es cada vez más rosada. Estas células se denominan eritroblasto básofilo, eritroblasto policromatófilo y eritroblasto ortocromático. Cuando se pierde el núcleo recibe el nombre de reticulocito. Esta célula no se divide pero se caracteriza por la presencia de poliribosomas y mitocondrias para continuar con la síntesis de hemo- Hierro globina. El reticulocito permanece 1-2 días en la médula y durante 1 a 2 días circula en la sangre hasta convertirse en eritrocito maduro; la vida media del eritrocito es de 120 días. La sustancia más importante en la regulación de la eritropoyesis es la eritropoyetina (EPO) una glucoproteína acídica formada por 166 residuos de aminoácidos y cuyo peso molecular es de 34.000 Da. El ácido sialico es un componente importante en sus efectos biológicos y su pérdida trae como resultado pérdida importante de su actividad. En cuanto al origen la EPO aparece inicialmente en el saco vitelino alrededor de los 14 días de gestación con la producción de eritrocitos macrocíticos nucleados. La eritropoyesis fetal típica se inicia fundamentalmente en el hígado y luego en el bazo entre las 6 y 8 semanas de gestación. Con excepción de lo que ocurre en el saco vitelino la eritropoyesis fetal, está principalmente bajo control de la EPO durante la segunda mitad de la gestación. Las concentraciones de la EPO aumentan progresivamente durante el tercer trimestre en forma regulada; este incremento de la EPO coincide con el aumento del hematocrito necesario para atender las demandas crecientes de oxígeno del feto en desarrollo, dado el medio intrauterino relativamente hipóxico. El mecanismo de producción y regulador de la EPO probablemente se localiza a nivel hepático y renal; se ha sugerido que la circulación fetal puede influir en el sitio de producción de la EPO. El estímulo para la producción de EPO se relaciona con disminución de la tensión de oxígeno en el tejido renal. En el feto la sangre oxigenada de la placenta llega a través de la vena umbilical y pasa en alto porcentaje por el sistema venoso al corazón y a su vez este lo envía al hígado, proceso que deja a este último órgano en un medio relativamente hipóxico, lo que permite que el sensor de oxígeno hepático participe la producción de EPO. En la fase final del tercer trimestre el conducto venoso se estrecha y envía cantidades cada vez más mayores de sangre oxigenada al hígado lo que permitió concluir que la constricción del conducto venoso se correlaciona con el cambio del sitio de producción de la EPO de hepática a renal. 63 Hierro Las células del sistema retículo endotelial también tienen capacidad de producir EPO y posiblemente sean responsables de la mayor producción de EPO fetal. La pobre participación del riñón en la producción de EPO en el feto se demostró en la sangre de fetos con agenesia renal (síndrome de Potter) en los que el hematocrito y la concentración sérica de EPO es normal. Después del nacimiento, se suspende el flujo sanguíneo placentario por lo que se cierra el conducto venoso y disminuye la irrigación hepática quedando este órgano solo con la arteria hepática y la vena porta, mientras aumenta el volumen y contenido de oxígeno de la sangre a los riñones. Los llamados sensores de oxígeno del riñón son más sensibles a la hipoxia que los del hígado, lo cual convierte al riñón en sitio extrauterino más importante de la producción de EPO. Se sugiere que el sensor de oxígeno sería una proteína con una fracción Hem sensible a los cambios en la tensión de oxígeno a los cuales responderían las células productoras de EPO. En esta forma la EPO se considera el enlace entre el sensor de oxí- 64 geno del riñón y los tejidos hematopoyéticos. Posteriormente a su secreción, la EPO se desplaza por la sangre a su blanco el tejido hematopoyético. La EPO actúa especialmente sobre la CFU-E, célula que posee gran número de receptores para ella. Sin EPO la CFU-E sufre muerte celular o apoptosis, por el contrario si hay EPO esta se fija a las células progenitoras o germinativas eritroides primitivas (CFU-E) y a las germinativas eritroides maduras estimula su proliferación y su diferenciación a proeritroblastos. Otros factores que influyen en la eritropoyesis son: • Hierro, vitaminas (B12, fólico, vitamina C, piridoxina o B6) • IL-3 (interleuquina-3) • GM-CSF (factor estimulante de colonias de granulocitos) • Tiroxina y andrógenos De acuerdo con el orden de madurez en la eritropoyesis hay participación de tres progenitores eritroides: BFU-E primitiva: con gran capacidad proliferativa que dependería de la cantidad de EPO disponible (P Gallangher y col) Sin em- Hierro bargo aún no se conoce la cantidad de EPO que requiere la BFUE primitiva en vivo, ya que ni la hipoxia ni la hipertransfusión alteran su crecimiento. BFU-E madura: su maduración y diferenciación disminuyen su capacidad proliferativa y aumentan su capacidad proliferativa? BFU-E: no posee gran capacidad proliferativa y tiene obligatoriamente necesidad de EPO. Después de una o dos divisiones se convierte en pronormoblasto que se consideran eritrocitos primitivos reconocibles. La diferenciación de las células germinativas eritroides a partir del estadio primitivo hasta alcanzar el estado de células progenitoras maduras se caracteriza por disminución en la necesidad de factores promotores de maduración eritroide y por la dependencia de la EPO para la maduración. La acción de la EPO en la célula germinativa eritroide madura es mediada por la unión con receptores específicos de la superficie celular. Se afirma que no existen reservas preformadas de EPO; estudios respecto a la regulación del gen de EPO han demostrado aumento importante en el RNA mensajero de EPO (RNAm) posterior a la exposición a hipoxia. La lentitud en el cambio de la producción de EPO hepática a renal puede desempeñar papel importante en la patogenia de la anemia del prematuro, debido a que disminuye la capacidad de respuesta de los sensores de oxígeno hepático a la hipoxia. Los sensores de oxígeno renales inmaduros, también pueden contribuir a este tipo de anemia. Otro factor que puede contribuir a la anemia del prematuro son las transfusiones durante las primeras semanas de vida; en este procedimiento se reemplaza la hemoglobina fetal por la del adulto. La hemoglobina del adulto cede el oxígeno a los tejidos más fácilmente que la hemoglobina fetal , por lo que disminuye el estímulo hipóxico para estimular la producción de EPO. Algunos autores han demostrado que lactantes con concentraciones más altas de hemoglobina adulta, presentabas cifras de EPO más bajas que lactantes con el mismo hematocrito pero concentraciones más altas de hemoglo- 65 Hierro bina fetal. Existen importantes diferencias entre el periodo de vida de los eritrocitos del neonato que es de 70 días y el de los adultos que es de 120 días, lo cual exige estímulos mayores en la eritropoyesis. Durante la maduración de los eritroblastos aumenta la concentración de hemoglobina en el citoplasma. La hemoglobina se forma por la unión de cuatro cadenas globínicas a cuatro grupos hemo. La síntesis del hemo se produce principalmente en la mitocondria y el grupo hemo es el resultado de la combinación de la protoporfirina IX con el hierro. Como se sabe el hierro necesario para la síntesis de hemoglobina es transportado a los eritroblastos por la transferrina que se une a receptores de membrana específicos. El metabolismo del eritrocito se dirige a mantener el equilibrio osmótico, por la elevada concentración de proteína intracelular representada por la hemoglobina y a mantener el hierro del grupo hemo en estado reducido (Fe++) ya que al oxidarse (Fe+++) la hemoglobina pierde su capacidad para unir el oxígeno. Por tanto el hierro es un elemento esencial para la función de numerosas proteínas ce- 66 lulares involucradas en el transporte de oxígeno y en la transferencia de electrones. Es un nutriente fundamental y su deficiencia tiene un espectro muy amplio de riesgos que incluyen trastornos en el desarrollo físico, alteraciones en el desarrollo cognoscitivo, alteraciones en el proceso de aprendizaje, disminución en la actividad física y en la productividad laboral, alteraciones inmunológicas, incremento del riesgo de enfermedades infecciosas y otras numerosas alteraciones(31) ; además es considerada la causa más frecuente de anemia nutricional en humanos. Por lo anterior el conocer las importantes funciones del hierro ha llevado a estimular el interés por el diagnóstico temprano de su deficiencia y como prevenirla. Referencias 1. Andrews NC, Bridge KR. Disorders of iron metabolism and sideroblastic anemia. Nathan and Oski”s Hematology of infancy and childhood 5a Ed Philadelphia: WB Saunders, 1998:423 2. Resfun AB, Schreiner BBI. Regulation of iron balance by absorption and excretion . Scand J Gastroenterology 1984;19:867 3. Wick M, Pinggera W, Lehmann P. Iron metabolism, diagnosis and therapy of anemias 3a Ed New York: Springer, 1996. Hierro 4. Fairbanks V, Klee G. Biochemical aspects Of hematology. Textbook of clinical chemistry. Tietz. Philadelphia WB Saunders, 1986. 5. Crichton RR, Charloteaux-Wauters M. Iron transport and storage. Eur J Biochem 1987;164:485 6. de Jong G, van Dijk JP, van Eijk HG. The biology of transferrin Clin Chim Acta 1990;190:1 7. He J, Furmansky P. Sequence specificity and transcriptional activation in the binding of lactoferrin to DNA. Nature 1995;373:721 8. Human Gene Mapping II Cytogenet Cell Genet 1991, Supplements 9. Osaki S , Johnson DA, Frieden E. The mobilization of iron from the perfused mammalian liver by a serum copper enzyme ferroxidase. I J Biol Chem 1971;246:3018 10. Jandl JH, Katz JH. The plasma to cell cycle of transferrin J Clin Invest 1963;42:314 11. Kawabata H, Germain R, Koffler H Phillip. Transferrin receptor 2 support cell growth both in iron chelated cultured cells and in vivo. J Biol Chem 2000; 275:16618 12. Cook JD, Skikne BS, Bines RD. Serum transferring receptor. Annu Rev Med 1993;44:63 13. Paterson S, Armstrong NJ, Iacopetta BJ, McArdle HJ, Morgan EH. Intravesicular pH and iron uptake by immature erithroid cells. Journal of Cellular Physiology 1984; 120:225 14. Huff RL, Hennessey TG, Austin RE, Garcia JF, Roberts BM, Lawrence JH. Plasma and red cell iron turnover in normal subjects and in patients having various hematopoietic disorders. Journal of Clinical Investigation 1950;29:1041 15. Inman RS, Wessling-Resnick M. Cha- racterization of transferring-independent iron transport in K562 cells. Unique properties provide evidence for multiple pathways of iron uptake. J Biol Chem 1993;268:8521 16. Worwood M. Ferritin. Blood Rev;4:259 17. Andrews SC, Arosio P, Bottke W. Structure, function and evolution of ferritins. J Inorg Biochem 1992;47:161 18. Wagstaff M, Worwood M, Jacobs A. Properties of human tissue isoferritins. Biochem J 1978;173:969 19. Levi S, Yewdall SJ, Harrison PM. Evidence of H and L chains have cooperative roles in the iron uptake mechanisms of human ferritin Biochem J 1992;288:591 20. Broxmeyer HE. H-Ferritin: a regularity cytokine that down-modulates cell proliferation. J Lab Clin Med 1992;120:367 21. Worwood M. Serum ferritin. Clin Sci 1986;70:215 22. Ponka P, Schulman HM. Regulation of heme biosynthesis:distinct features in erythroid cells. Stem Cells 11 (supplement 1) 24-35 23. Heilmeyer L. Atransferrinemias. Acta Haematologica 1966;36-40 24. Hershko C, Graham G, Bates G, Rachmilewitz E. Non specific serum iron in thalassemia: an abnormal serum iron fraction of potential toxicity Br J Haematol 1978;40:255 25. Craven CM, Alexander J, Eldridge M, Kushner JP, Bernstein S, Kaplan J. Tissue distribution and clearance kinetics of non-transferrin-bound iron in the hypotransferrinemic mouse: a rodent model for hemochromatosis. Proceedings of the National Academy of Sciences 1987;84:3457 26. Kaplan J, Jordan I, Sturrock A. Regulation of the transferring-independent iron transport system in cultured cells 67 Hierro J Biol Chemistry 1991;266:2997 27. Randell EW, Parkes JG, Olivieri NF, Templeton DM. Uptake of non-tranferrin bound iron by both reductive and non-reductive processes is modulated by intracellular iron. J Biol Chemistry 1994;269:16046 28. Pollack S Receptor mediated iron uptake and intracellular iron transport. American Journal of Hematology 1992;39:113 29. Hamazaki S, Glass J. Non-tranferrin dependent 59Fe uptake in phytohemagglutinin stimulated human peripheral lymphocytes. Experimental Hematology 1992;20:436 30. Bothwell TH, Charlton RW, Cook JD, Finch CA Iron metabolism in man : Blackwell Scientific Publications 1979. 31. Viteri E F Prevention of Iron Deficiency The National Academies Press 1998; 45-102 68 Monografías y Revisiones 1. Forrellat Barrios M, Gautier du Defaix H, Fernández N Metabolismo del Hierro. Rev Cubana Hematol Inmunol Hemoter 2000; 16(3): 149-60 2. Worwood Mark. Regulation of iron metabolism. Annales Nestlé 1995; 53:3039 3. Malgor LA, Valsecia ME. Farmacología de las anemias carenciales. Cápitulo 2 pags 26-31 4. El médico interactivo. Diario electrónico de la sanidad No 892 Enero 2003 5. Rojas Escalante E, Luchtenberg G, Sola H. Eritropoyetina Recombinante: Actualización. Rev Hospital Mat. Inf Ramón Sarda 1996 No2 Hierro Diego Rincón Resumen E l hierro es considerado un metal esencial no solo para el crecimiento normal, sino también para el desarrollo mental y motor normal del individuo. Siendo tan crucial, su deficiencia es padecida por una gran proporción de la población mundial y además gran parte de ella se acompaña de anemia. Ante este cuadro, el hierro juega un papel de capital importancia en un órgano esencial como es el cerebro, ya que en este órgano es donde el hierro alcanza su mayor concentración, alcanzándola en más del 80% de los casos en la primera década de la vida. La concentración a nivel cerebral del hierro no es homogénea, lo que indica que existen áreas con una mayor concentración como es la Sustancia Nigra y parte del Núcleo Estriado, y es allí donde podemos correlacionar el porque su deficiencia se asocia con alteraciones de diversos neurotransmisores como son la Dopamina y la Noradrenalina; y todo esto repercutirá en el deterioro de la función neurológica del individuo. Hierro y función cerebral El hierro en algunos aspectos ha sido considerado como un metal traza – requerido en muy pequeñas cantidades – pero indiscutiblemente es un elemento esencial, crítico en la función de todas las células; presente en un sinnúmero de sistemas moleculares, que van desde servir como portador de oxígeno y electrones; hasta catalizador en la oxigenación, hidroxilación y otros procesos vitales como en el desarrollo y la proliferación celular.(1) El principal papel del hierro en mamíferos es la de transportar Oxígeno como parte del grupo hemo que, a la vez es parte de una proteína, la hemoglobina; además el hierro es un elemento vital en la acción enzimática, involucrando allí al sistema de citocromos en la mitocondria. Todo este tipo de reacciones químicas hacen que se genere una serie de radicales li69 Hierro bres tales como el O2 y el OH, radicales que pueden potencialmente ser nocivos en el daño de los diferentes componentes celulares(2). Proceso que será explicado en el siguiente capítulo. El Hierro es esencial no solo para el crecimiento normal, sino también para su desarrollo tanto en la esfera motora como en la mental, así como en la función cognoscitiva(3). El Hierro puede jugar un papel importante en la función cerebral, ya que el cerebro es un sitio muy significativo de concentración del metal en el cuerpo humano. En ciertas regiones del cerebro, la concentración puede ser incluso más alta que en el hígado; v.gr. 21.3 mg/100 mg de peso fresco comparada con 13.4 mg/100 mg para el hígado (3). Es tan importante este metal que en los primeros años de vida, el 80% del total del metal que existe en el adulto fue almacenado en su cerebro durante la primera década de la vida (3). El Hierro puede afectar potencialmente la función cerebral de dos formas; la primera como evidencia del daño secundario a un 70 déficit del metal, y la segunda forma como evidencia de un daño por sobrecarga del metal; de las dos formas, la primera es la más importante y la más común. Se considera que la Deficiencia de Hierro (DHi) es el problema nutricional más prevalente, ya que cerca de 1.500 a 2.000 millones de personas están afectadas por el. Según la Organización Mundial de la Salud (OMS), está deficiencia afecta cuando menos al 20 – 25% de todos los lactantes menores; al 43% de los niños hasta los 4 años y al 37% de los niños entre los 5 – 12 años de edad,(WHO, 1992). La DHi se puede producir de varias maneras; la primera, cuando la absorción es insuficiente para satisfacer las necesidades; la segunda, puede ser debida a una ingesta inadecuada; la tercera, una reducción de la biodisponibilidad del hierro en la dieta o un incremento en la necesidad o una pérdida crónica de sangre; si la DHi se prolonga, llevará a una anemia por deficiencia de hierro (ADHi). La anemia por deficiencia de Hierro (ADHi) es un importante problema de Salud Pública, por el Hierro impacto tanto en el desarrollo psicológico y físico, como en el comportamiento y rendimiento laboral. Las personas más susceptibles a desarrollar (ADHi) son los niños lactantes, preescolares, adolescentes y las mujeres en edad fértil, sobre todo la mujer embarazada. Para ampliar el concepto de la anemia por deficiencia de hierro, podemos definir como un déficit de hierro corporal total al producido cuando los requerimientos de hierro exceden el suministro fisiológico de este o por la pérdida del mismo, lo que resultará en un balance negativo. Se distinguen dos etapas en esta deficiencia: • Deficiencia Latente de Hierro: Allí hay disminución de los depósitos de hierro; disminuye la ferritina sérica; aumenta la concentración de protoporfirina eritrocítica; disminuye la saturación de transferrina y los valores de hemoglobina permanecen normales. • Deficiencia Manifiesta de Hierro = Anemia: Una vez agotados los depósitos de hierro, la producción de hemoglobina y otros compues- tos es limitada, se reduce la ferritina, aumenta la concentración de protorfirina eritrocítica; se reduce la saturación de transferrina, reduciéndose los valores de hemoglobina. La deficiencia latente de hierro (DLH) es un problema que no solo se presenta en los niños sino también en los adolescentes. Un estudio realizado en Japón, indica que hasta el 71.8% de las jóvenes en edad escolar presentan DLH tres años después de su primera menstruación, (Kagamimori y cols., 1988). A nivel mundial, debido a la menstruación y al embarazo, más del 51% de las mujeres en edad fértil, pueden no tener, o tener muy bajas las reservas de hierro. En las comunidades occidentales, el 71.8% de las mujeres premenopáusicas no embarazadas tienen depleción de los depósitos de hierro y carecen de reservas, aunque solo un 2 a 3% de ellas presenta anemia ferropénica (1). Varios síntomas observados como cansancio, agotamiento, nerviosismo, falta de energía, poca capacidad de concentración, dificultad para encontrar las palabras adecuadas, dolor de cabeza en las mañanas, disforia de- 71 Hierro presiva, labilidad psicológica, disminución de la eficiencia, pérdida del apetito, aumento de la susceptibilidad a la infección y falta de memoria, suelen relacionarse con la anemia. Se supone, que estos síntomas son producidos por una menor capacidad de los eritrocitos para transportar oxígeno, pero allí el hierro por sí solo tiene un impacto importante en el cerebro, y por consiguiente, en los problemas mentales, eso quiere decir, que perfectamente todos estos síntomas pueden presentarse aun en pacientes que aunque tengan deficiencia de hierro no se encuentran anémicos. En 1984, Tucker y cols., realizaron un estudio en 69 estudiantes diestros, midiendo niveles de hierro y ferritina séricos e investigaron la actividad del cerebro tanto en estado de reposo como en actividad cognoscitiva. Los resultados fueron sorprendentes: tanto la activación del hemisferio izquierdo como el rendimiento cognoscitivo dependieron del nivel de hierro; determinaron que entre menor fuera el nivel de ferritina más débil era la activación no sólo del hemisferio izquierdo sino de la región occipital de ambos hemisferios. Esto significa que si el nivel de ferritina sérica es bajo, el he- 72 misferio dominante, en su totalidad, y los centros de memoria del área óptica de ambos hemisferios se activan en menor grado. Estos centros, como el área de lenguaje visual y la de lenguaje sensorial del hemisferio izquierdo son esenciales para la memoria, allí es evidente que la deficiencia de hierro puede llevar a un deterioro de la misma (1). Se observo también una correlación entre el nivel de hierro y la actividad cognoscitiva, sobre todo en la fluidez de lenguaje, medida por la capacidad del individuo de crear palabras que comenzaran y terminaran por letras específicas, cuando los niveles de hierro sean bajos se observara un menor grado de activación de las áreas del lenguaje de los hemisferios dominantes. Las bases biológicas del compromiso comportamental y del desarrollo cognoscitivo en infantes con DHi puede incluir las siguientes posibilidades: • El hierro desempeña una función importante en los sistemas dopaminérgicos, y su deficiencia puede acarrear anormalidades en el metabolismo del neurotransmisor. Hierro • En segundo lugar, el nivel de hierro influye en la mielinización de las terminaciones nerviosas. • Su deficiencia puede llevar a alteraciones en el metabolismo energético cerebral. Youdim en 1989, demostró que el intercambio de hierro a nivel cerebral es muy lento y la capacidad del cerebro para almacenar hierro es significativamente menor que la del hígado; sin embargo, el cerebro tiene una mayor tendencia que el hígado a retener el hierro y así evitar su depleción. El aumento de hierro después de una suplementación de este mineral es mucho más rápido a nivel hepático que a nivel cerebral y el hígado además presenta los niveles de hierro más altos. La lentitud de los cambios de los niveles de hierro a nivel cerebral son debidos al proceso por el cual el hierro atraviesa la barrera hematoencefálica (BHE); ya que esta barrera sólo permite la captación del suplemento de hierro en caso de deficiencia. En el cerebro existe un sistema utilizado para la adquisición de hierro, desde el “complejo” del plasma (receptor de transferrina (Tf), mecanismo que sirve para la dispersión o movilización del hierro, así como de un mecanismo para el almacenamiento de hierro específico de la célula (Isoformas H y L de ferritina), las cuales están heterogéneamente distribuidas y no todas las regiones del cerebro son igualmente sensibles a una alteración en el estado de hierro corporal. Estudios en la deficiencia de hierro postnatal involucrando las isoformas de ferritina (H:L) en el cerebro de cerdos y de roedores revelaron un dramático efecto en la DHi sobre la expresión de estas dos subunidades de la molécula de ferritina (4,26). Los cambios en la expresión de la proteína parecen estar reguladas por el Sistema del Elemento de Respuesta al Hierro – Proteína Reguladora del Hierro (IRE –IRP), el cual parece ser el “complejo” funcional de hierro en cada célula (5,6). La BHE provee de un efectivo punto de regulación para la movilización del hierro desde el “complejo” plasmático al líquido cefalorraquídeo,; el papel del líquido cefalorraquídeo no está bien entendido; los niveles de hierro se han descrito que son muy altos en este líquido en cerebros perinatales, y en individuos mayores se sugiere que el rango de concen- 73 Hierro tración está entre 15 y 25 mg/L en humanos; dichas concentraciones son 5 a 10 veces más bajas que las concentraciones correspondientes al plasma; mientras que los plexos coroideos son también una fuente de movimiento de hierro dentro y fuera del cerebro. No todas las regiones cerebrales contienen la misma concentración de hierro, con relación a la que se presenta en los ganglios basales, sustancia nigra, y núcleo cerebelar profundo, los cuales son particularmente ricos en hierro. Las más altas concentraciones se encuentran en el globus pallidus, núcleo caudado, putamen y sustancia nigra. Estas áreas del cerebro que son ricas en hierro en la rata adulta no lo fueron en los primeros 60 días de vida; lo mismo sucede en el cerebro del humano, donde por ejemplo, la sustancia nigra no es rica en hierro sino hasta que alcanza una edad de 12 a 15 años (4). La concentración de hierro es muy alta en el cerebro al nacer, disminuye gradualmente y comienza a incrementar sus niveles de manera coincidente con el inicio de la mielinización además de 74 observarse una expresión incrementada del RNAm del receptor de transferrina. El hierro que obtiene el cerebro primariamente por la vía del receptor de transferrina (Tf) y de receptores expresados de Tf en las células endoteliales de la microvasculatura cerebral, parecen tener un papel regulador para los astrocitos adyacentes en la regulación del ingreso a través de la BHE. (Figura 1). La tasa de ingreso dl hierro se afecta por el estado iónico del hierro; existe una tasa incrementada cuando el estado del hierro es bajo y una tasa disminuida cuando este es alto, además, este mecanismo de ingreso es altamente selectivo y no refleja la permeabilidad total de la BHE. Se ha observado una distribución heterogénea del hierro en el cerebro, lo que puede muy bien ser el resultado de una regulación regional del ingreso y proceso de redistribución que es dependiente de la distribución del receptor de Tf y de dos nuevas proteínas transportadoras de hierro; el transportador de metal divalente 1 (DMT1- cotransporte protón/hierro, el cual remueve el hierro de los endosomas acidificados) y un Hierro Neurona Astrocito MTP Microglia Oligodendrocito FrtR Células ependimales Frt MTP DMT-1 TfR DMT-1 Tf BHE LCR Plexo coroideo Ventrículo Figura 1. Papel de la barrera hematoencefálica (BHE) en el transporte de hierro dentro del cerebro. Diagrama adaptado de Beard, John. Iron deficiency alters brain development and functioning. J Nutr. 133:1468S-1472S, 2003. DMT-1: transportador de metal divalente 1; Tf: transferrina; TfR: receptor de transferrina; MTP: proteína transportadora de metal o ferroportina; FrtR: Receptor de Ferritina; Frt: ferritina. transportador de metaloproteína 1(MPT1/ferroportina/Ireg1, o exportador de hierro celular) (7). Dichas dos proteínas se piensa que están involucradas o que son responsables del ingreso y salida del hierro de la célula. La distribución regional de la proteína del Tf y los niveles de RNAm prestan soporte al concepto que el volumen de proteína del Tf cerebral es translocado a aquellas regiones donde las localizaciones del Tf y del RNAm del Tf sean distintas. Estudios en infantes humanos de conducción nerviosa proveen de alguna evidencia que el efecto de la deficiencia de hierro sobre el funcionamiento neuronal es irreversible. Dallman (8,9) demostró dos décadas atrás que las ratas jóvenes privadas del hierro en la vida postnatal temprana tenían significativamente menor (27%) contenido de hierro cerebral total que los controles de 28 días de nacidos y que eran completamente resistentes a la restauración de su complemento normal de hierro cerebral, luego de una agresiva administración en la dieta por 45 días, este concepto se mantuvo por todo este tiempo, hasta que 75 Hierro otros investigadores lograron demostrar que el cerebro es totalmente sensible a la depleción y administración de hierro de la dieta y que además usa un sinnúmero de mecanismos para regular de la manera más homeostática el flujo de hierro(10,11). Muchos investigadores se han esforzado en considerar la cuestión de los “períodos críticos” del desarrollo que absolutamente requieren de un adecuado hierro nutritivo para un “normal” desarrollo; aunque el curso del desarrollo en la rata es más corto que en el humano, en ambas, hay una secuencia en lo concerniente a la migración celular, mielinización significativa, diferenciación celular e incremento en la expresión de neuropéptidos; todo esto parece ocurrir en humanos de los 3 a los 16 meses postparto y en las ratas de los 7 a los 25 días postparto (4). Las diferentes necesidades regionales para el hierro en el cerebro durante diferentes estados del neurodesarrollo podrían dar a conocer una sensibilidad diferencial de las regiones cerebrales a la depleción nutricional del hierro. Por ejemplo, en estudios realizados en roedores durante los períodos medio y posterior de nacidos (equivalentes a 6 – 12 meses en 76 el humano), fue muy significante el descenso en el 25% en la corteza, estriado, y en el cerebro posterior al contenido de hierro con un corto período de alimentación con una dieta baja en hierro. En contraste, fue solamente un descenso del 5% en el contenido de hierro en el tálamo. Durante la deficiencia de hierro postdestete fue comparable la declinación de un 20 – 30% en corteza, estriado, y cerebelo, pero allí el tálamo también fue sensible a la deficiencia de hierro en la dieta con un descenso del 20% en la concentración de hierro (10,11). De manera importante, se debe considerar que la restauración del hierro cerebral con un agresivo ingreso de hierro en la dieta, da como resultado una serie de anormalidades en el metabolismo de la dopamina (DA) y en los comportamientos relacionados con ella, v.gr: la sensibilidad de una región cerebral a la pérdida de hierro durante el desarrollo, el cual, es similar al que está relacionado a los requerimientos de hierro para el desarrollo regional durante este período; todo esto contrasta con la percepción que el hierro cerebral es “resistente” a la depleción de hierro, ya que experimentos hechos por Pinero y cols. (12) de- Hierro mostraron claramente que en los roedores, los tratamientos dietarios pueden disminuir el hierro cerebral en 10 días y volver a llenarse en 14 días, por supuesto los datos en el humano no se han realizado por lo que las conclusiones al respecto son inciertas. ¿Qué hay de cierto en la irreversibilidad de los efectos a la deficiencia de hierro en la infancia? Todo esto se basa en las siguientes características: • La vasta mayoría de estudios en humanos se han centrado en infantes entre 12 – 24 meses de edad con DHi sin examinar cuidadosamente a niños que fueran mayores. • Los modelos animales muestran muy claramente anormalidades irreversibles como resultado de deficiencia al hierro tanto en la gestación como en la lactancia temprana. • Informes de deficiencia de hierro en adolescentes y adultos, generalmente muestran una normalización en el funcionamiento cerebral y en los comportamientos, con la corrección de la deficiencia de hierro. Recientemente fue descrita una condición clínica que fue llamada como Síndrome de las Piernas Intranquilas (Restless Legs Síndrome) el cual está relacionado con el déficit del contenido de hierro cerebral y su metabolismo; las imágenes por MRI (Imagen por Resonancia Magnética) demostraron una disminución en el contenido de hierro tanto en la sustancia nigra como en el núcleo rojo; observando que la severidad en la disminución del contenido de hierro estaba correlacionado con la severidad de los síntomas. Un número de pacientes son verdaderamente resistentes al ingreso del hierro por la dieta pero los síntomas se resuelven en la mayoría de los casos con altas dosis de hierro dextrano intravenoso; además es interesante observar como algunos pacientes responden a agonistas de DA o a L-DOPA si el síndrome es diagnosticado primariamente (13). Para ampliar un poco más, la distribución del hierro en el cerebro es enteramente heterogénea, ya se habían nombrado algunas áreas, como la sustancia nigra, el núcleo rojo, el núcleo cerebelar profundo, pero además contamos con áreas también muy ricas en este metal como son el núcleo accumbens, y una porción del hipocampo que pueden ser muy cer- 77 Hierro 78 canas sus concentraciones a la del hígado. Ahora, un examen de la proteína transferrina y de los niveles de RNAm, sugieren un sistema altamente regulado para el ingreso, distribución, y almacenamiento del hierro en el tejido neural. Esta distribución es sensible a los estados de neurodesarrollo, actividad metabólica, y, por supuesto, a las enfermedades neurodegenerativas asociadas con el envejecimiento. La función del hierro en el cerebro se ha focalizado en a-) metabolismo del oligodendrocito y la mielinización, b-) en el metabolismo de la monoamina, y c-) en el metabolismo del GABA. Hill noto una gran similaridad en la distribución del hierro cerebral con regiones del cerebro que recibían un influjo de fibras del ácido g-amino butírico (GABA) y además eran predominantemente regiones cerebrales dopaminérgicas. Esta colocalización del hierro con “sistemas” de neurotransmisores particulares sugiere que es una asociación de tipo funcional (14). síntesis de neurotransmisores, incluyendo la triptófano hidroxilasa y la tirosina hidroxilasa, también como en el catabolismo de los mismos. El hierro también es cofactor para la ribonucleótido reductasa, la cual es la etapa límite en la síntesis de DNA, y su función en determinadas vías de expresión génica para determinadas proteínas relacionadas con el metabolismo del hierro celular (15). El Hierro es requerido para la propia mielinización de la médula espinal y la sustancia blanca de los pliegues cerebelares, además de ser un cofactor para un número de enzimas involucradas en la El metaboismo de hierro intraneuronal involucra la incorporación de hierro en las enzimas del transporte de electrones; síntesis y empaquetamiento de neurotransmisores; además del ingreso El tipo de células predominante que contienen hierro en el cerebro del ratón, la rata, el mono, el cerdo, y el humano es el oligodendrocito (16). Los oligodendrocitos son responsables de la síntesis de ácidos grasos y de colesterol para la mielina; una pérdida del hierro disponible para los oligodendrocitos está asociado con una disminución en las cantidades y en la composición de la mielina. La alteración en la maduración de los oligodendrocitos está asociado con una falla para acumular el hierro. Hierro y degradación de los neurotransmisores. En adición a todo esto, los efectos secundarios sobre la reducción de peróxidos, metabolismo de aminoácidos, y elongación y desaturación de los ácidos grasos tiene implicaciones para los mecanismos potenciales de acción del hierro sobre la función neuronal. Mackler y cols. (17) demostraron concentraciones normales de los citocromos mitocondriales y de las tasas de fosforilación oxidativa en cerebros de ratas con DHi, aunado a eso, la actividad enzimática de la tirosina hidroxilasa, triptófano hidroxilasa, monoamino oxidasa, succinato hidroxilasa, y el citocromo C eran normales en cerebros de animales con DHi. Datos más recientes, en una evaluación por inmunohistoquímica de la citocromo C oxidasa en neonatos con DHi mostraron una dramática disminución en el hipocampo (18), y los niveles de monoaminas eran sensibles a las variaciones del estado del hierro en el cerebro. Los tractos dopaminérgicos parecen ser consistentemente sensibles a la deficiencia de hierro en regiones del cerebro (19,20); Youdim y cols. demostraron muy bajas densidades de los receptores D2 de Dopamina en el estria- do de ratas, y como se había hecho evidente anteriormente, estudios más recientes demostraron que el núcleo accumbens era también sensible a los efectos de la DHi. Además de todo esto, la densidad del transportador de DA está significativamente disminuido en el estriado y en el núcleo accumbens, los cuales son campos terminales de neuronas originadas en la sustancia nigra y en el tegmento ventral (1,5). In vivo, los estudios han demostrado, que la DA extracelular está elevada en el estriado de ratas con DHi y retornan a los niveles normales cuando el contenido de hierro cerebral y el estado del hierro retornan a lo normal. Experimentos farmacológicos hechos con cocaína – un inhibidor del transporte de dopamina - demostró una sensibilidad reducida, lo que implica cambios en ambos, tanto en la densidad del transportador como en su funcionamiento; otros datos además demuestran que ciertos receptores de dopamina en el estriado y en la sustancia nigra no son restaurados a los niveles normales si la DHi ocurre durante el período neonatal, lo cual contrasta con anteriores observaciones hechas en animales viejos donde la ad- 79 Hierro ministración de hierro normaliza el metabolismo de la dopamina. Es por esto que las alteraciones en el metabolismo de la dopamina tanto en el tracto mesolímbico como en el tracto nigroestriatal podría fácilmente estar relacionado con una percepción y una motivación alterada. Los sistemas serotoninérgicos y noradrenérgicos son sistemas bioquímicos similares a la dopamina y forman una familia de transportadores de monoamina responsables del ingreso de estos neurotransmisores en las neuronas presinápticas. El sistema dopaminérgico se desarrolla rápidamente durante la vida postnatal temprana con un rápido incremento en el número y densidad de los transportadores de DA y de receptores en las áreas terminales en la pubertad temprana. Dichas proyecciones de monoaminas juegan un papel importante en la organización del crecimiento axonal y en la formación de sinapsis durante los estados tempranos del crecimiento cerebral, para luego convertirse en su papel más tradicional de neurotransmisión con la edad. El papel del hierro, o de otros micronutrientes, en esta gran mezcla de conexiones neuronales con el desa- 80 rrollo no es bien conocido; ante esta evidencia, se presenta un gran conflicto, ya que no se sabe si estos sistemas monoaminérgicos son o no sensibles a los cambios en el estado del hierro a nivel cerebral, ya que por ejemplo, estudios hechos en el ratón demostraron significativamente densidades muy bajas de los transportadores de serotonina en el estriado si había DHi, mientras que estudios más recientes usando menos ligando específico no observaron tales efectos (5). Sinaptosomas de ratas deficientes de hierro fueron menos capaces de captar la serotonina marcada con Tritio (H3) que aquellos sinaptosomas de animales con suficiente concentración de hierro (20). En experimentos in vivo por microdiálisis en ratas jóvenes deficientes de hierro revelaron una elevada concentración a nivel intersticial de norepinefrina, mientras que medidas directas de homogenatos tanto en el ratón como en la rata no mostraron diferencias relativas a los controles en la dieta (21). En otros estudios realizados, donde se hizo tolerancia al frío y termorregulación que las mujeres anémicas deficientes de hierro y ratas tenían un elevado nivel plasmático de NA (22). Esto Hierro es más consistente con una más rápida pérdida de NA desde las cisternas del sistema nervioso simpático periférico y que son sugestivas de un efecto de la deficiencia de hierro sobre el mecanismo de entrada de la monoamina. Esto es importante de recalcar que los transportadores de serotonina, NA y DA son todos miembros de la misma familia de cotransportadores del Na + y muestran similares características con respecto a la regulación y translocación (4). La deficiencia de hierro in útero y en el destete está asociada con una disminución significativa en la actividad de la glutamato descarboxilasa, glutamato deshidrogenasa, y la GABA transaminasa (5). Las últimas dos enzimas son las responsables de la síntesis y degradación del GABA. Las concentraciones del GABA están elevadas en la deficiencia de hierro por la dieta tanto en el hipocampo, estriado, y globus pallidus (5) y puede estar relacionado con alteraciones en el metabolismo del manganeso cerebral. Ahora, con base en todas las investigaciones hechas en la búsqueda del papel realizado por el hierro en el metabolismo de los neurotransmisores, se conoce que el hierro es esencial para un número de enzimas involucradas en la síntesis del neurotransmisor incluyendo triptófano hidroxilasa (serotonina) y tirosina hidroxilasa (Noradrenalina (NA) y DA), además el hierro es un cofactor para la ribonucleótido reductasa, y es esencial para el funcionamiento de un número de reacciones de transferencia de electrones relacionadas tanto con el metabolismo de los lípidos como con el metabolismo energético cerebral (4). El hierro está relacionado con la actividad de la monoamino oxidasa, enzima crucial para las propias tasas de degradación de estos neurotransmisores. Para poder correlacionar al hierro con estos sistemas se dan las siguientes observaciones fundamentales: i) El hierro está colocalizado con las neuronas dopaminérgicas por todas las partes del cerebro (4). ii) La DA extracelular y la NA están elevadas en cerebros de ratas con DHi; pero no otros neurotransmisores. iii) Como la concentración del hierro cerebral cae debido a la restricción del hierro por la dieta, hay una disminución en la densidad de los receptores D2 y D1 y en los transportadores de DA en el estriado (23). iv) La pérdida del hierro cerebral con la deficiencia 81 Hierro de hierro en la dieta es específico de región y lleva a un efecto heterogéneo sobre la neurobiología de la DA, v.gr: en las regiones donde el hierro no falta, no hay cambios en la biología de la DA. v) Los efectos de la deficiencia de hierro sobre la DA cerebral no son debidos a la anemia, per se, ya que la anemia hemolítica en ausencia de deficiencia de hierro no produce estas anormalidades en la neurobiología de la DA (4). Correlacionando el hierro cerebral con la biología del neurotransmisor podemos determinar algunos cambios en el comportamiento y esto puede ser visto, cuando tenemos animales e infantes humanos con DHi que tienen cambios en el comportamiento los cuales son resistentes a la terapia con hierro. Los cambios comportamentales fueron robustamente asociados con cambios en la concentración de DA central y de hierro (12, 23). Recientes análisis del comportamiento se han venido realizando, con determinaciones de DA y de hierro cerebral, revelando algunas relevantes relaciones (4): • Análisis de regresión multivariado de la actividad espontánea 82 demostró que tanto como el 65% de la variabilidad en la exploración de los ambientes nuevos está asociado con los niveles de hierro en el tallo cerebral ventral, y la densidad de receptores de DA D1 en el cerebro medio, núcleo caudado y putamen. • Análisis multivariado de comportamientos similares a la ansiedad demostraron que cerca del 45% de la varianza en la latencia para moverse a un ambiente “seguro” puede ser atribuido a la variación en la densidad de transportadores de DA y de receptores D 2 en el núcleo accumbens. • En la deficiencia de hierro predestete y en la deficiencia de hierro postdestete en ratas, resulto en una exploración disminuida y movimiento disminuido. El suministro nuevamente de hierro, resulto en una normalización de varios de estos comportamientos, hasta poder alcanzar una normalización de la mayoría de las alteraciones en la biología de la DA. • Hay una falla para liberar hierro en el cerebro durante períodos particularmente sensibles, evidenciado en anormalidades bioquímicas en cerebros de infantes con deficiencia de hie- Hierro rro. Los investigadores lograron demostrar una velocidad de conducción nerviosa enlentecida usando para ello estudios de potenciales evocados auditivos en infantes de 6 meses de edad. Luego de 12 meses de terapia con hierro con los índices hematológicos corregidos del estado de hierro, la velocidad de conducción nerviosa enlentecida permaneció anormal y no mostró una indicación de normalización. Las dos posibles explicaciones ante estos hechos son; la primera, es que la DHi ocurrida en estos infantes sucedió durante un “período crítico” del desarrollo con cambios que resultaron irreversibles asociados con hipo-mielinización y cambios en el metabolismo de las monoaminas; y la segunda explicación, es que esto no es debido a mediciones sensibles al estado del hierro, sino que otros factores ambientales en los primeros 6 meses de la vida de los infantes conducen hacia otras vías del neurodesarrollo (24). De esta manera, cualquier “puente” directo entre la literatura de estudios en animales y las mediciones hechas en infantes, niños y adolescentes será muy difícil de establecer. La otra forma como el cerebro puede verse afectado por el hierro es si se presenta acumulación de este, situación que es referenciada como Neurodegeneración con acumulación de hierro cerebral (NBIA, Neurodegeneration with brain iron accumulation, formalmente llamado Síndrome de Hallervorden – Spatz) (25). El nuevo término NBIA está favorecido a la luz de las actividades poco éticas tanto de Hallervorden y Spatz antes y durante la II Guerra Mundial. La NBIA enmarca un grupo de algunos desórdenes extrapiramidales caracterizados por una evidencia radiográfica de acumulación de hierro en el cerebro, usualmente en los ganglios basales, aquí también se incluyen la neuroferritinopatías y la aceruloplasminemia. La enfermedad más relevante es la neurodegeneración asociada a la pantotenato – kinasa (PKAN) causada por mutaciones en el gen PKAN2. La prevalencia estimada es de 1 – 3/ 1.000.000 la cual ha sido sugerida basándose en los casos observados en la población. El sello característico de la PKAN es la disfunción extrapiramidal que 83 Hierro incluye la distonía, la rigidez, y la coreoatetosis. El cuadro de inicio puede tener una forma clásica, la cual está presente en la primera década de la vida; y una segunda forma atípica, la cual está más comúnmente en la segunda o tercera década de la vida. La pérdida de la ambulación en la forma clásica frecuentemente ocurre en los 10 a 15 años del inicio, mientras que en la forma atípica ocurre en los 15 a 40 años del inicio. Las características que lo corroboran son: • Tracto corticoespinal involucrado 1. Espasticidad 2. Signos de extensión de los dedos • Degeneración retinal o atrofia óptica • Acantocitosis de los eritrocitos • Fracción de Lipoproteína prebeta plasmática baja o ausente • Historia familiar consistente con herencia autosómica recesiva (puede incluir cosanguinidad). Antes de disponer de la MRI, el NBIA era un diagnóstico postmortem. El Síndrome de HallervordenSpatz se caracterizaba por la apa- 84 rición de una pigmentación rojiza –café en el globus pallidus y en la zona reticular de la sustancia nigra. El hierro es el mayor componente de este pigmento. En el PKAN, la acumulación de hierro es específica del globus pallidus y la sustancia nigra. Dichas áreas contienen aproximadamente tres veces la cantidad normal de hierro. El metabolismo del hierro a nivel sistémico es normal sin observarse un incremento en el nivel global del hierro cerebral. El PKAN se atribuye a la deficiencia o ausencia de la pantotenato kinasa 2, la cual se codifica por el PKAN 2 en el cromosoma 20p13 – p12.3, el cual es uno de los cuatro genes para la pantotenato kinasa humana. La pantotenato kinasa es una enzima reguladora esencial en la biosíntesis de la CoA, catalizando la fosforilación citosólica del ácido pantoténico (vitamina B5), la N – pantotenoil – cisteína, y la panteteína. La deficiencia de esta enzima, se piensa que causa la acumulación de la N – pantotenoli – cisteína y de panteteína, los cuales pueden producir directamente una toxicidad celular o un daño por la vía de los radicales libres como queladores del hierro. La deficiencia de la PAK 2 resulta en una depleción de la Hierro CoA y por lo tanto presentara un defecto en la biosíntesis de la membrana donde sea mayor los requerimientos de la CoA. Referencias 1. Hierro (Complejo de Hidróxido de Hierro (III) y Polimaltosa IPC). Monografía del producto. 2. ADAMSON, John W.; Iron deficiency and other hypoproliferative anemias; Principles of Internal Medicine of Kasper, Braunwald, Fauci et al.; 16th Edition; 2005; page: 586 – 591. 3. VALDIRI, Ernesto; VARGAS, Jorge; Alteraciones por Deficiencia de Hierro; Revista Latinoamericana de Farmacología y Terapéutica; No. 2; Junio 2004; pag. 17 –21. 4. BEARD, John; Iron Deficiency Alters Brain Development and Functioning; Journal of Nutrition; 133: 1468S – 1472S, 2003. 5. BEARD, John L.; CONNOR, James R.; Iron Status and Neural Functioning; Annu. Rev. Nutr. ; 23: 41 – 58 ; 2003. 6. THEIL, Elizabeth C.; Ferritin: At the Crossroads of Iron and Oxygen Metabolism; Journal of Nutrition 133: 1549S – 1553S, 2003. 7. SIDDAPPA, A.J.; RAO, R.B. et al; Developmental changes in the expression of IRP and iron proteins in perinatal rat brain; J. Neurosci. Res.; 68: 761 – 775; 2002. 8. DALLMAN, P.R.; SIIMES, A.M.; MANIES, E.C.; Brain Iron: persistent deficiency following short term iron deprivation in the young rat; Br. J. Nutr. 31: 209 – 215, 1975. 9. DALLMAN, P.R.; SPIRITO, R.A.; Brain Iron in the rat: Extremely slow turno- ver in normal rat may explain the long – lasting effects of early iron – deficiency; J. Nutr. 107: 1075 – 81; 1997. 10. ERIKSON, K.M.; PINERO, D.; CONNOR, J.; BEARD, J.L.; Iron Status and distribution of iron in the brain of developing rats. J. Nutr. 127: 2030 – 38; 1997. 11. PINERO, D.J.; LI, N.Q.; CONNOR, J.R.; BEARD, J.L.; Alterations in brain iron metabolism in response to dietary iron changes; J. Nutr. 130: 254 – 63; 2000. 12. PINERO, D.J.; JONES, B.; BEARD, J.L.; Alterations in brain iron metabolism in response to dietary iron changes; J.Nutr. 130: 254 – 263; 2000. 13. EARLEY, C.; ALLEN, R.; BEARD, J.L.; CONNOR, J.R.; Insight into the pathophysiology of restless legs syndrome; J. Neurosci. Res. 62: 623 – 628; 2000. 14. BEARD, J.L.; CONNOR, J.R.; JONES, B.C.; Iron in the brain; Nutr. Rev.; 51(6): 157-70; 1993. 15. EISENSTEIN, R.S.; Iron regulatory proteins and the molecular control of mammalian iron metabolism; Annu. Rev. Nutr. 20: 627 – 62; 2000. 16. CONNOR, J.R.; FINE, R.E.; Development of transferrin – positive oligodendrocytes in the rat central nervous system; J. Neurosci. Res. 17: 51 – 59; 1987. 17. MACKLER, B.; PERSON, R.; MILLER, L.R.; FINCH, C.A.; Iron deficiency in the rat: effects on phenylalanine metabolism; Pediatr. Res. 13: 1010 – 11; 1979. 18. SIDDAPPA, A.J. et al; Developmental changes in yhe expression of iron regulatory proteins and iron transport proteins in the perinatal rat brain; J. Neurosci. Res. 68(6): 761 – 75; 2002. 19. BEN-SCHACHAR, D.; FINBERG, J.P.M.; YOUDIM, M.B.H.; The effect of iron chelators on dopamine D2 recep- 85 Hierro tors; J. Neurochem. 45: 999 – 1005; 1985. 20. ERIKSON, K.M.; JONES, B.C.; BEARD, B.L.; Iron deficiency alters dopamine transporter functioning in rat striatum; J. Nutr. 130: 2831 – 37; 2000. 21. KWIK-URIBE, C.L.; et al; Chronic marginal iron intakes during early development in mice result in persistent changes in dopamine metabolism and myelin composition; J. Nutr. 130(11): 2821 – 30; 2000. 22. BEARD, J.L.; BOREL, M.J.; DERR,J.; Impaired thermoregulation and thyroid function in iron deficiency anemia; Am. J. Clin. Nutr. 52: 813 – 819; 1990. 23. FELT, B.; LOZOFF, B.; Brain iron and behavior of rats are not normalized by 86 treatment of iron deficiency anemia during early development; J. Nutr. 126: 693 – 701; 1996. 24. GRANTHAM-MCGREGOR, S; ANI, C.; A review of studies on the effect of iron deficiency on cognitive development in children. J. Nutr. 131: 649S – 66S; 2001. 25. GREGORY, Allison M.; HAYFLICK, Susan J.; Neurodegeneration with Brain Iron Accumulation; Orphanet Encyclopedia; September 2004. 26. HAN, Jian; DAY, Jonathan; Connor, James; BEARD, John; H an L ferritin subunit mRNA expression differs in brains of control and iron – deficient rats; J. Nutr. 132: 2769 – 2774, 2002. Hierro Diego Rincón Resumen L a anemia es considerada como un problema mayor en pacientes con insuficiencia renal crónica, ha sido corregida en parte por el desarrollo y posterior administración de la eritropoyetina recombinante humana (Epo), y al posterior conocimiento de que la Epo debe siempre administrarse asociada al hierro IV, debido al fenómeno de déficit funcional de hierro que se presenta en los pacientes con insuficiencia renal crónica. Aunque la anemia no ha sido considerada como un contribuyente común de la falla cardíaca, de alguna manera la anemia puede causar daño cardiaco que eventualmente puede progresar hacia la falla cardiaca y cuando se hace una relación entre falla cardiaca e insuficiencia renal, se ha encontrado que la anemia es muy común en la asociación entre falla cardiaca severa y su resistencia a la medicación; la corrección con eritropoyetina y hierro endovenoso permitieron observar un marcado mejoramien- to en el estado funcional de los pacientes y por supuesto en su función cardiaca; disminuyendo las hospitalizaciones, la administración de diuréticos, y la consiguiente mejoría de su función renal. A esta relación, se le ha denominado como Síndrome Anemia Cardio- Renal (1), donde se establece que el tratamiento de la anemia puede ser vital en el mejoramiento y progresión tanto de la falla cardiaca como de la insuficiencia renal. Sindrome anemia cardio- renal Al observar la literatura, podemos apreciar como dentro de las características clínicas de la falla cardiaca no aparece registrada la anemia como factor de riesgo y tampoco como rasgo clínico, v.gr: dentro de los factores que precipitan la falla cardiaca están la hipertensión no controlada, las arritmias cardiacas, los factores ambientales, la terapia inadecuada, la infección pulmonar, el estrés emocional, la sobrecarga de líquidos, el infarto de miocardio, la rup87 Hierro tura de una cuerda tendinosa, los desórdenes endocrinos (tirotoxicosis). Si vemos los diferentes signos y síntomas de la falla cardiaca precipitada por una disfunción ventricular derecha están el dolor abdominal, anorexia, náusea, constipación, ascitis, edema periférico, distensión venosa yugular, reflujo hepatoyugular, hepatomegalia. Si se encuentra ante una falla precipitada por una disfunción ventricular izquierda, se daría una disnea precipitada por el ejercicio, disnea paroxística nocturna, ortopnea, taquipnea, tos, hemoptisis, edema pulmonar, ruido S3 en galope, efusión pleural, respiración de Cheyne Stokes; y si desea puede encontrar hasta hallazgos no específicos como son: Intolerancia al ejercicio, fatiga, debilidad, nocturia, síntomas del SNC, taquicardia, palidez, cianosis de los dedos, cardiomegalia (2). Ahora bien, si eso es lo observable en la falla cardiaca, también podemos apreciar que en la insuficiencia renal, la anemia aparece como parámetro clínico en la clasificación de la insuficiencia, pero no se le da a ésta la importancia de su papel de agravamiento de la sintomatología de la insuficiencia; v.gr: en una insuficiencia re- 88 nal mínima el paciente generalmente cursa asintomático; cuando ya se hace leve la insuficiencia puede ser asintomática o llega a presentar nocturia, hipertensión, y una leve anemia. En una insuficiencia moderada ya el cuadro clínico se hace más florido, con nocturia, fatiga, intolerancia al frío, alteraciones del gusto, anorexia, hiperfosfatemia, hipocalcemia, hipercalemia, acidosis metabólica, empeoramiento de la anemia, inhabilidad en ajustar los cambios en el ingreso del sodio; y si ya se pasa a una insuficiencia renal severa, se presentará malestar general, pérdida de energía, prurito, náusea intratable y vómito, calambres en miembros inferiores, mioclonus, asterixis, alteración del sensorio, convulsiones, empeoramiento de los parámetros de laboratorio (3). Es por esto que hacia 1999, un grupo de médicos nefrólogos y cardiólogos liderados por el Dr. Donald Silverberg de la Universidad de Tel Aviv, Israel, “descubrieron” una relación no reconocida o por lo menos no apreciada con anterioridad, que involucraba la falla cardiaca congestiva, la insuficiencia renal crónica, y la anemia. La falla cardiaca congestiva puede ser la causa o puede empeo- Hierro rar la anemia y la insuficiencia renal crónica; a la vez, la insuficiencia renal puede causar o empeorar la anemia y la falla cardiaca; y la anemia puede causar o empeorar las dos anteriores; a este circulo vicioso se le denomino como Síndrome Anemia Cardio-Renal. Este modelo sugiere que si se da el tratamiento adecuado a la falla cardiaca y a la anemia prevendría o demoraría la progresión de la falla cardiaca y la insuficiencia renal; de otro lado, esto también sugiere, que a menos que la anemia y la falla cardiaca sean tratadas, ambas la falla cardiaca y la insuficiencia renal seguirían progresando. La pregunta de si la anemia puede causar la falla cardiaca congestiva?; ya es conocida, que si la anemia es lo suficientemente severa, puede causar falla cardiaca hasta en individuos normales (4). La hipoxia tisular y la vasodilatación periférica en la anemia causa un descenso de la presión sanguínea con un incremento de la respuesta simpática, por lo cual, producirá taquicardia, un incremento del volumen de gasto, hay vasoconstricción renal, y disminuye el flujo sanguíneo renal, con retención de sal y agua. La reduc- ción del flujo sanguíneo renal también causa un incremento en la secreción de renina, de angiotensina, aldosterona, y de la hormona antidiurética (ADH), además de aumentar la vasoconstricción renal y la retención de sal y agua, incrementa el volumen plasmático. La Noradrenalina, renina, angiotensina y la aldosterona son todas tóxicas para las células renales y cardiacas. Todas estas conducen a una dilatación ventricular e hipertrofia, y eventualmente a la muerte de la célula miocárdica y a falla cardiaca (5). Algunos estudios de pacientes que han estado bajo hemodiálisis y diálisis peritoneal han mostrado una asociación entre la anemia y un incremento en la morbimortalidad (6). En pacientes con falla renal en prediálisis, la anemia causa hipertrofia ventricular izquierda e incremento del índice de masa ventricular izquierda, lo que causa cardiomegalia debido a ambos, a un volumen incrementado de fin de sístole ventricular izquierdo y a un volumen incrementado de fin de diástole ventricular izquierdo, causando todo esto, una función sistólica cardiaca reducida detectada por una fracción de eyección ventricular izquierda reducida (7,8). Desde 1986 cuando se ini- 89 Hierro cio el uso de la eritropoyetina para tratar la anemia de los pacientes con diálisis, se encontró que los síntomas cardiacos tales como respiración corta y fatiga en esfuerzo o aun en reposo, mejoraban con la terapia. (8,9). Aunque la anemia es conocida por ser el factor de mayor riesgo para la falla cardiaca en pacientes con diálisis. (10), no se sabe nada acerca de la respuesta de la eritropoyetina en los pacientes con falla cardiaca anémica que no han sido dializados. Desde 1994, varios grupos médicos iniciaron el tratamiento de la anemia en pacientes con hemodiálisis, diálisis peritoneal e insuficiencia renal con eritropoyetina y hierro IV, principalmente en forma de hierro sacarato. La importancia del hierro IV en la falla renal comprende dos aspectos: la anemia por deficiencia de hierro, la cual es común en los pacientes con falla renal y en pacientes con diálisis, responden al hierro IV mejor que con el hierro oral, el cual es pobremente absorbido y poco tolerado; y cuando estos pacientes son tratados con eritropoyetina, la producción de células rojas, y la necesidad de hierro para la síntesis de hemoglobina, se incrementa en forma dramática; pero 90 esta necesidad frecuentemente no puede ser suplida de manera rápida y suficiente por los depósitos de hierro normal, permitiendo tan solo la depleción de estos. Sin el hierro IV, la eritropoyetina frecuentemente falla en la tarea de incrementar los niveles de hemoglobina. Cuando se usa a la vez con el hierro IV, la eritropoyetina incrementa los niveles de hemoglobina a valores superiores, y actualmente por esto, la eritropoyetina puede ser administrada en menores dosis. Este último concepto tiene gran importancia, ya que la eritropoyetina es excesivamente costosa. No sólo por esta limitación, sino por sus propiedades, el hierro más usado en esta indicación es el hierro sacarato. Danielson et al. investigaron las propiedades farmacéuticas del hierro sacarosa en 12 voluntarios sanos después de suministrarles hierro sacarosa en una dosis única IV que contenía 100 mg Fe. Las concentraciones de hierro en plasma y orina se comprobaron mediante muestreos repetidos, antes y después de la administración durante 24 horas. Los resultados obtenidos en este estudio en relación con el rápido aumento de la concentración de hierro sérico total y la eliminación renal mínima Hierro de hierro sacarosa confirman los resultados obtenidos en un estudio anterior en pacientes con déficit de hierro (Anatkov y Gekova). Se encontró que la eliminación renal de hierro sacarosa era menor al 5% de la dosis administrada y la fracción mayor fue excretada durante el primer día como lo demuestra Danielson et al. (1996b). Después de la inyección de hierro sacarosa marcado radioactivamente en cerdos, Langström et al. encontraron que cinco minutos después de la inyección ya se observó un nivel de actividad alto no sólo en el hígado sino también en la médula ósea, indicando una concentración elevada de hierro marcado en estas áreas. Por lo tanto, el hierro proveniente del hierro sacarosa es de muy rápida disponibilidad para la eritropoyesis. Las propiedades del hierro sacarosa son cruciales en pacientes con una necesidad clínica de liberación rápida de hierro a los depósitos de hierro; con anemia por déficit férrico debido a un aporte insuficiente de hierro o a una pérdida de hierro, que no toleran o responden insuficientemente a la terapia con hierro oral, o con una malabsorción al hierro oral o falta de cumplimiento en el tratamien- to con hierro oral. En los pacientes en tratamiento con eritropoyetina recombinante, la optimización de la eritropoyesis puede reducir significativamente los requerimientos de eritropoyetina así como los costos del tratamiento. ¿Qué tan común es la anemia en pacientes con falla cardiaca, y qué tan importante es ésta? Un estudio de Silverberg (11) en el 2000, en 142 pacientes, encontró que más de la mitad de los pacientes tenía una hemoglobina < 12 g/dL. Se observó como el promedio de los niveles de hemoglobina cayeron en pacientes con anemia, en los cuales la severidad de la falla cardiaca se había incrementado; tan solo el 9.1% de estos pacientes con bajo compromiso (Clase I NYHA (Clasificación de la New York Heart Association) eran anémicos, comparados con el 79.1% de aquellos con un alto compromiso (Clase IV NYHA). El promedio de los niveles de hemoglobina cayo de un promedio de 13.7 gr/dL en los de bajo compromiso a un promedio de 10.9 gr/dL en el grupo de mayor severidad. Asociado a todo esto, casi la mitad de los pacientes tenía evidencia de falla renal, determinada por una creatinina sérica por encima de 1.5 mg/dL, lo que hace que la pre- 91 Hierro valencia y la severidad de la anemia se incrementen con el incremento de las cifras de creatinina sérica. A pesar de observar una condición común en los pacientes con falla cardiaca, y que algunos estudios como el estudio Framingham en 1987 mencionaban a la anemia como un factor de riesgo para desarrollar la falla cardiaca (12); la Asociación Americana del Corazón (AHA) en 1999 dentro de las recomendaciones hechas para la falla cardiaca congestiva no incluyo una simple referencia a la anemia, ni tampoco lo hicieron en las recomendaciones hechas en el 2001 (13,14). Aun en el 2003, revisiones hechas para diagnóstico, investigación, y tratamiento de la falla cardiaca han fallado en mencionar a la anemia como causal (15). Un estudio reciente, encontró que cerca del 29% de los pacientes con falla cardiaca en un servicio de medicina interna y cardiología eran anémicos, y casi ninguno había sido diagnosticado y menos había sido tratado (16). Luego de evidenciar la asociación entre la anemia y la falla cardiaca, algunos grupos decidieron 92 determinar si la anemia era la causante o empeoraba la falla cardiaca, o era tan solo un indicador de una enfermedad crónica; optando por tratar al paciente anémico con falla cardiaca, conscientes de los posibles daños al intentar corregir la anemia; ya que algunos estudios controlados al azar, no mostraban ningún beneficio y en algunos mostraban hasta empeoramiento del pronóstico del cuadro cardiaco (17). Ante este panorama, un grupo de 26 pacientes con falla cardiaca y anémicos fueron tratados, encontrando que en 3 meses, su clasificación funcional de NYHA había mejorado, las hospitalizaciones habían caído dramáticamente, su depuración de creatinina, la cual venía cayendo a razón de 1 ml/min/mes, se estabilizó o en su mayoría mejoró, y por lo tanto, las dosis de furosemida oral o IV fueron marcadamente reducidas. Como dato adicional, la fracción de eyección ventricular izquierda también mejoró (11). Ante el escepticismo generado a partir de esta asociación, un sinnúmero de publicaciones empezaron a evidenciar lo común que era encontrar la anemia en los pacientes con falla cardiaca, y esta anemia, estaba estrechamente re- Hierro lacionada con la severidad de la enfermedad cardiaca, la mortalidad, la duración y el número de hospitalizaciones, además del compromiso de la función renal. (18,19). En estudios posteriores, se decidió realizar un estudio controlado al azar de 32 pacientes anémicos con falla cardiaca, 16 de los cuales se les corrigió la anemia y a los otros 16 no (20). En el grupo tratado, como en el primer estudio mencionado, la clase NYHA, la fracción de eyección ventricular izquierda, hospitalizaciones, y dosis de furosemida, todas mejoraron, y la función renal se mantuvo sin cambios. En contraste, en el grupo control, todas las cinco mediciones se deterioraron. Cuatro de los pacientes con la anemia sin tratar fallecieron, mientras la falla cardiaca siguió su progresión, mientras ningún paciente murió del grupo tratado. En otros estudios, pacientes que evidenciaban esta triada y que eran tratados con la combinación de eritropoyetina y hierro IV, se observo que la respuesta en los pacientes diabéticos y en los no diabéticos eran similar en todos los aspectos, aun en el deterioro de la tasa funcional renal. Esto sor- prende, ya que se espera que los pacientes diabéticos presenten una falla más rápida, asociado con la nefropatía diabética, pero este no fue el caso, por lo cual, lo que se cuestiona es si esta “nefropatía diabética progresiva” puede por lo menos ser el resultado de una anemia pobremente tratada y de una falla cardiaca inadecuadamente tratada. Vale la pena mencionar que el manejo será mucho mejor si tanto los nefrólogos como los cardiólogos trabajan al tiempo sobre el mismo paciente, esto se basa en el hecho en que los cardiólogos al tratar una insuficiencia lo hacen con su gran arsenal terapéutico, es decir, allí incluyen los diuréticos; los inhibidores de la enzima convertidora de angiotensina; los bloqueadores beta, tales como el carvedilol, metoprolol, o bisoprolol, los cuales han demostrado ser útiles en las insuficiencias cardiacas moderadas o severas; los bloqueadores de receptores de angiotensina; y el diurético aldospirona. El uso de todos estos agentes, entre los nefrólogos dista mucho de ser el ideal en pacientes con insuficiencia cardiaca a quienes se les ha asociado una enfermedad cardiovascular. 93 Hierro La prevalencia de la anemia en diferentes estudios de falla cardiaca puede variar considerablemente, con un rango tan amplio que va desde el 3 al 61%; esta variación puede estar sujeta a varios factores como son la población estudiada, la definición que se hace de anemia, y lo que tal vez se observa con una alta frecuencia, es que el médico no incluye dentro de los diagnósticos, el de anemia, a menos que la hemoglobina se encuentre extremadamente baja. Tomando más sin embargo los datos, se sugiere que cerca del 40% de los pacientes con falla cardiaca presentan una hemoglobina < 12gr/dL (16). En varios estudios se ha evaluado la mortalidad encontrada en los pacientes con falla cardiaca anémica v.s. aquellos pacientes con falla cardiaca pero sin anemia, encontrándose que estos últimos registran una menor tasa de mortalidad que los pacientes inicialmente nombrados; asociado con esto, se ha podido establecer que las hospitalizaciones eran más frecuentes y más prolongadas para aquellos pacientes que presentaban falla cardiaca con anemia que en los no anémicos; ahora bien, en algunos de los pacientes en que se relacionaba anemia con 94 falla cardiaca, la función renal era independiente de los dos problemas anteriores, aunque, la falla renal es considerada un factor de riesgo para la hospitalización y la mortalidad asociada con la severidad de la falla cardiaca (18,19). Así la anemia y la insuficiencia renal son aditivas en sus efectos negativos sobre todos aquellos aspectos relacionados con la falla cardiaca. En un estudio controlado con placebo de falla cardiaca severa, los tratados con eritropoyetina por tres meses tenían un marcado incremento en la utilización de oxígeno durante el ejercicio máximo (prueba de función cardiaca y buen predictor de sobrevivida), también como un incremento en la capacidad al ejercicio y calidad de vida (21). El incremento en la utilización de oxígeno estaba directamente relacionada con el incremento en la hemoglobina; en aquellos con un incrementado volumen plasmático, la corrección de la anemia estaba asociado con un incremento en la masa de células rojas a lo normal y una marcada disminución en el volumen plasmático. También se ha observado como en estudios de falla cardiaca se- Hierro vera, se ha corregido la anemia con eritropoyetina combinada con hierro endovenoso, observándose una marcada caída en las hospitalizaciones y en el uso de dosis de diuréticos, estabilizándose la función renal (22). Existe otra anormalidad que se adiciona a la anemia y que puede estar asociada a la ecuación establecida para la falla cardiaca, y es que gran parte de los pacientes con falla cardiaca anémica se encuentran mal nutridos, con un bajo índice de masa corporal, un bajo colesterol sanguíneo y una albúmina sérica disminuida (23). Estudios en diálisis y en cáncer han mostrado que la anemia puede causar mal nutrición y que el tratamiento de la anemia con eritropoyetina puede mejorar esto (24). La caquexia frecuentemente se ve en la falla cardiaca, lo que podría estar particularmente relacionada con la anemia. Los pacientes con falla cardiaca anémica son los que están más estrechamente relacionados con la mayor severidad de la falla cardiaca. Ellos están más relacionados con la presencia de hiponatremia, altos niveles del péptido natriurético beta (parámetro que refleja el compromiso de la falla cardiaca), y signos de inflamación tales como la Proteína C Reactiva (PCR), además del Factor de Necrosis Tumoral Alfa (a-TNF). Todo esto sugiere que una de las razones de que el tratamiento de la falla cardiaca no haya sido exitoso puede ser que la anemia haya sido ignorada. El hecho que la función renal sea estabilizada luego de la corrección de la anemia, es consistente con el reciente hallazgo de que la anemia es un factor de riesgo independiente para la progresión de la falla renal en la falla cardiaca (25). La causa más común de la anemia en la falla cardiaca es probablemente dada por la asociación con la insuficiencia renal, causada por el reducido flujo sanguíneo renal que se origina de la falla cardiaca. Esto da como resultado una inapropiada baja secreción de eritropoyetina. En adición a esto, las citoquinas tales como el a-TNF, secretado en grandes cantidades desde el daño cardiaco, puede contribuir a la anemia mientras esto deprime la producción de eritopoyetina en el riñón, interfiere con la habilidad de la eritropoyetina para hacer células rojas en la médula ósea, e interfiere con la liberación del hierro desde los sitios de almacenamiento en las cé- 95 Hierro lulas del sistema retículo endotelial y su liberación hacia la médula ósea (26). La falla cardiaca generalmente no es considerada una causa común de la insuficiencia renal progresiva; sin embargo, la mitad de todos los pacientes con falla cardiaca tienen una enfermedad renal moderada o severa, evaluado por una depuración de creatinina < 60 ml/min/1.73 m2 o una creatinina sérica mayor o igual a 1.5 mg/ dL (27). Los pacientes con falla cardiaca no controlada pierden la función renal a una tasa promedio de 1 ml/min/mes (27). Cerca del 60% de los pacientes que inician diálisis tienen falla cardiaca (28), esto podría significar que un muy amplio porcentaje de tales pacientes podrían tener su función renal estable, si su falla cardiaca fuera bien controlada; ahora bien, se observa que la anemia no es tratada de manera temprana en la mayoría de los pacientes con falla cardiaca o con insuficiencia renal. La anemia puede contribuir a otras enfermedades cardiovasculares, por ejemplo, en la falla arterial coronaria, donde la anemia ha sido considerada como un factor de riesgo común e independiente 96 para mortalidad de este tipo de evento cardiovascular (29); en un estudio de 689 pacientes con enfermedad coronaria, el 20 % de ellos cursaba con anemia, pero de estos, cerca del 50% eventualmente fallecieron. La mortalidad a dos años fue de 55% para aquellos con Hb <11gr/dL, y de solamente el 3% para aquellos con Hb >14gr/dL, además la anemia fue un factor de riesgo independiente para la mortalidad en análisis multivariado. Ante todo esto, ahora la anemia es considerada en la población sana como un factor de riesgo para la enfermedad cardiovascular, eventos coronarios, falla cardiaca y para mortalidad (30,31). Referencias 1. SILVERBERG, D.S.; WEXLER, D.; IAINA, A.; The role of Anemia in Congestive Heart Failure and Chronic Kidney Insufficiency, The Cardio-Renal Anemia Syndrome; Persp. in Biol. and Med, 47(4): 575-589; 2004. 2. JOHNSON, Julie A.; PARKER, Robert; Heart Failure Chapter 11; Dipiro et al. Pharmacotherapy, A Pathophysiologic Approach; Fourth Edition; Ed. Appleton & Lange,1999. 3. St. PETER, Wendy L.; LEWIS, Matthew; Chronic Renal Insufficiency and End-Stage Renal Disease, Chapter 42; Dipiro et al. Pharmacotherapy, A Pathophysiologic Approach; Fourth Edition; Ed. Appleton & Lange, 1999. Hierro 4. ANAND, I.S., et al.; Patogénesis of edema in chronic anemia: Studies of body water and sodium, renal function, haemodynamics and plasma hormones. Br. Heart J. 70:357-62, 1993. 5. KATZ, A.M.; The cardiomyopathy of overload: An unnatural growth response in the hypertropied heart; Ann. Intern. Med. ; 121: 363-71; 1994. 6. COLLINS, A. J. ; Influence of target hemoglobin in dyalisis patients on morbidity and mortality. Kidney Int. 61 (Suppl 80):S44-48; 2002. 7. LEVIN, A.; The relationship of hemoglobin level and survival: Direct or indirect effects. Neph Dial Transplant 17 (Suppl 5): 8-13; 2002. 8. LOW, I., et al.; Echocardiographic findings in patients on maintenance hemodialysis substituted with recombinant human erythropoietin. Clin. Nephrol. 31: 26-30; 1989. 9. LOW-FRIEDERICH, I., et al.; Therapy with recombinant human erythropoietin reduce cardiac size and improves heart function in chronic hemodialysis patients; Am.J. Nephrol. 11:54-60; 1991. 10. COLLINS, A.J.; Anaemia management prior to dialysis: cardiovascular benefits and cost-benefit observations. Nephrol Dial Transplant 18(Suppl 2):ii26; 2003. 11. SILVERBERG, D.S.;et al.; The use of subcutaneous erythropoietin and intravenous iron for the treatment of the anemia of severa, resistant congestive herat failure improves cardiac and renal function, functional cardiac class, and markedly reduces hospitalization. J. Am. Coll. Cardiol. 35: 1737-44; 2000. 12. KANNEL, W.B. Epidemiology and prevention of cardiac failure: Framingham Study insights. Eur. Heart J. 8(Suppl F): 23-29; 1987. 13. HUNT, S.A.; ACC/AHA guidelines for the evaluation and management of chronic heart failure: Executive summary. Circulation 104: 2996-3007; 2001. 14. PACKER, M., et al.; Consensus recommendations for the management of chronic heart failure. Am. J. Cardiol. 83: 1A-38A; 1999. 15. JESSUP, M.; BROZENA, S.; Heart Failure; N. Engl. J. Med. 348:2007-18; 2003. 16. TANG, W.H.W; et al; Anemia in ambulatory patients with chronic heart failure: A single-center clinical experience derived from electronic medical records. J. Am. Coll. Cardiol. 41 (Suppl): 157A, 2003. 17. BESARAB, A., et al; The effects of normal as compared with low hematocrit values in patients with cardiac disease who are receiving hemodialysis and epoetin. N. Engl. J. Med. 339: 584-90; 1998. 18. AL-AHMAD, A.; et al; Reduced kidney function and anemia as risk factors for mortality in patients with left ventricular dysfunction. J.Am. Coll. Cardiol 8:955-62; 2001. 19. McCLELLAN, W.M.; et al; Anemia and renal insufficiency are independent risk factors for death among patients with congestive heart failure admitted to community hospitals: A populationbased study. J. Am. Soc. Nephrol. 13: 1928-36; 2002. 20. SILVERBERG, D.S.; The effect of correction of mild anemia in severe, resistant congestive heart failure using subcutaneous erythropoietin and intravenous iron: A randomized controlled study. J. Am. Coll. Cardiol 37: 1775-1780; 2001. 21. MANCINI, D.M.; Effect of erythropoietin on exercise capacity in patients with moderate to severe chronic heart failure. Circulation 107:294-99; 2003. 97 Hierro 22. NOVAK, B.; Anemia management in an outpatient heart failure center. Abstract. J. Card. Fail. 9(Suppl 1):386; 2003. 23. HORWICH, T.B.; Anemia is associated with worse symptoms, greater impairment in functional capacity and a significant increase in mortality in patients with advanced heart failure; J. Am. Coll. Cardiol. 39: 1780-86, 2002. 24. RIEDEL, E. ; Amino acid and alphaketo acid metabolism depends on oxygen availability in chronic hemodialysis patients; Clin Nephrol 53(Suppl 1): S56-60; 2000. 25. BUTLER, J. et al.; Relationship between heart failure and development of worsening renal function among hospitalized patients. Am. Heart J. 147: 331-38; 2004. 26. IVERSON, P.O.; Decreased hematopoiesis in bone marrow of mice with congestive heart failura. Am. J. Physiol. Regul. Integr. Comp. Physiol 282:R166- 72; 2002. 27. HILLEGE, H.L.; et al; Accelerate de- 98 cline and prognostic impact of renal function after myocardial infarction and the benefits of ACE inhibition: The CATS randomized trial. Eur. J. Heart Failure 24: 412-20; 2003. 28. AVORN, J.; et al.; Nephrologist care and mortality in patients with chronic renal insufficiency. Arch. Int. Med. 162:2002-6; 2002. 29. REINECKE, H.; et al; Haemoglobinrelated mortality in patients undergoing percutaneous coronary interventions. Eur. Heart J. 24: 2147-50; 2003. 30. JURKOVITZ, C.T.; et al; Association of high serum creatinine and anemia increases the risk of coronary events: Results from the prospective community-based atherosclerotic risk in communities (ARIC) study; J. Am. Soc. Nephrol. 14: 2919 – 25; 2003. 31. SANDGREN, P.E.; et al.; Chronic anemia as a risk factor for the incidence of new heart failure in the general Medicare population. Abstract. J. Am. Geriatric Soc. 51 (Suppl): P263; 2003. Hierro Hoover O. Canaval L a anemia durante la ges tación es un gran proble ma de salud pública, mucho mayor en los países en vías de desarrollo, donde contribuye notoriamente a incrementar la morbilidad y mortalidad materna y feto-neonatal. Es muy importante, conocer la realidad nutricional de nuestros pueblos, para contribuir a disminuir los riesgos pluricarenciales, pretendiendo obtener el mejor resultado perinatal. La anemia ferropriva es la más prevalente de las anemias en el embarazo, se cree que causa alrededor de un 95% de los casos clínicos durante este importante período, motivo por el cual este capítulo se centrará fundamentalmente en esta patología. Muchas situaciones condicionan la aparición de una anemia durante la gestación. El parasitismo intestinal y la malaria son consideradas dos de las causas más importantes en la etiología. Algunos grupos se hacen más vulnerables, tal es el caso de las fumadoras, las vegetarianas, las deportistas y aquellas que consumen altas cantidades de café. Por supuesto el pertenecer a un bajo nivel socio económico será un condicionante fuerte para desarrollar anemia durante el embarazo. La malaria y la anemia de células falciformes son alteraciones importantes que aumentan las demandas tanto de hierro como de ácido fólico. El hierro presente en todas las células del ser humano, cumple varias funciones vitales: transportador de oxígeno desde los pulmones a los tejidos, en forma de hemoglobina (Hb), usándose como un facilitador de oxígeno. Almacenado en el músculo en forma de mioglobina, sirve como un medio de transporte de electrones de las células en la forma de citocromo y actúa como parte integral de las reacciones enzimáticas en varios tejidos. La OMS desde 1968 clasificó la anemia en la gestación determinándola a nivel del mar en aquellas mujeres cuyos valores de Hb eran inferiores a 11,0 g/dL en el 99 Hierro 1er y 3er trimestre e inferiores a 10,5 g/dL para el 2º trimestre del embarazo. Considerando desde esa época que era importante para el 1er trimestre iniciar la suplementación con hierro y folatos solo a aquellas que inicien su gestación con valores menores o iguales a 11,0 g/dL (anemia), y ya para los dos trimestres finales se deberá suplir preferiblemente a todas las embarazadas. Similar recomendación efectuó el “Center for Disease Control and Prevention” (CDC) de Atlanta y el Colegio Americano de Obstetras y Ginecólogos. 1. La masa del eritrocito El aumento en el volumen de masa eritrocitaria (RBC) no empieza hasta la semana 20 de gestación, desde entonces aumenta más rápidamente que el volumen plasmático (VP), hasta la semana 28, posteriormente el incremento es menor, hasta el término del embarazo. La masa eritrocitaria al final de la gestación llega a ser superior en un 30% que en la mujer no embarazada. En el postparto temprano, el RBC sigue siendo 10% aproximadamente superior a los niveles de la no embarazada durante 2 semanas, y regresan a lo normal alrededor de la 6ª se- 100 mana. Esta disminución se correlaciona principalmente con la pérdida de sangre durante el parto y a un descenso en la producción de eritrocitos. La eritropoyesis regresa a su nivel normal hacia finales del postparto, alrededor de la 8ª semana en el puerperio. El aumento de RBC durante el embarazo es producto de una interacción compleja de factores hormonales y fisiológicos, ajustado con la producción de la eritropoyetina. En una gestación normal, el nivel del eritropoyetina empieza a incrementarse lentamente desde la semana 15, pero solo se observan los efectos del estímulo en la RBC entre las semanas 18 a 20, estableciéndose la actividad máxima entre las 20 y 29 semanas, y ello se relaciona con el aumento máximo en el flujo sanguíneo y el mayor consumo de oxígeno basal. El nivel de eritropoyetina empieza a disminuir lentamente después del nacimiento a pesar de la pérdida de sangre en el parto. La deficiencia de hierro es aún más frecuente que la anemia como tal establecida, situación que se hace más grave en el embarazo, inclusive en mujeres con adecuados depósitos de hierro. Hierro La masa eritrocitaria (RBC) disminuye en el primer trimestre del embarazo, para luego aumentar, y alrededor de la semana 30 llega a valores similares a los del estado no grávido, por último aumenta mucho más hasta el final de la gestación, siendo considerablemente mayor en las gestaciones múltiples. Entonces al término de la gestación se puede apreciar un incremento de un 150% del volumen plasmático y en un 120 a 125% de la RBC, en relación a el estado no gravídico. Dado que el tiempo de vida de los eritrocitos no se modifica en la gestación, los cambios señalados se asocian a un incremento en la tasa eritropoyética. Uno de los elementos involucrados en la reducción de la hemoglobina (Hb) en el primer trimestre es una disminución en la eritropoyetina (Epo) sérica, lo que se traduce, junto al aumento en el volumen plasmático en el 1er y 2º trimestre, en un grado de hemodilución funcional, entre tanto que la Epo aumenta desde la semana 20 en adelante en forma regular, en todo embarazo normal. Cuando los depósitos de hierro materno decrecen, el número de receptores de transferrina placentarios aumenta para favorecer una mayor captación de hierro. Por otra parte un aumento en la transferencia de hierro al feto se produce por una mayor síntesis de ferritina placentaria. Sin embargo estos mecanismos homeostáticos para el hierro en la interfase fetoplacentaria siempre se verán afectados y serán vulnerables por estados deficitarios de hierro en la madre. 2. Manifestaciones de la deficiencia de hierro Las embarazadas con déficit de hierro tienen glóbulos rojos microcíticos e hipocrómicos. En la gestación las anemias ferroprivas presentes durante los dos primeros trimestres, están asociadas a un riesgo dos veces mayor para presentar parto pretérmino y hasta tres veces mayor para tener recién nacidos con bajo peso. Existe evidencia por estudios randomizados controlados, que la suplementación de hierro disminuye la incidencia de anemias ferroprivas durante el embarazo, principalmente los desarrollados en países no industrializados, Adicionalmente se encuentran en la literatura, algunos estudios en los que la suplementación rutinaria de manera universal, de hierro durante la gestación, pudiese llegar a producir efectos adversos en la salud ma- 101 Hierro terna o fetal, pero estos no son concluyentes. 3. Prevalencia y riesgos de anemia por déficit de hierro En la mujer embarazada la expansión del volumen sanguíneo es aproximadamente de un 35% y el crecimiento fetal, placentario y de otros tejidos maternos, elevan hasta tres veces las demandas de hierro en el segundo y tercer trimestre en aproximadamente 5.0 mg hierro /día. Aunque la menstruación ya no se sucede y la absorción de hierro se incrementa notoriamente durante la gestación, muchas mujeres que no consumen suplementación farmacológica de hierro pueden no sostener sus reservas de hierro, particularmente durante los dos últimos trimestres. En el puerperio las concentraciones de hierro en el feto y la placenta se contabilizan como una pérdida de las reservas maternas, pero por fortuna parte del hierro de ese volumen sanguíneo incrementado puede volver a conformar las reservas de la madre. La suplementación farmacológica de hierro debe entonces ser selectiva y fruto del estudio juicioso de nuestras pacientes, lo cual nos va a permitir determinar a cuales pacientes suplimos y 102 cuando es el momento oportuno para ello. Una de las razones principales para no ordenar suplementación a base de compuestos con hierro es la percepción por parte de la paciente o del profesional de la salud que el aporte extra de hierro pudiese llegar a ocasionar alteraciones en el bienestar materno o perinatal, igualmente limita fuertemente la presencia de efectos secundarios indeseables (Estreñimiento, náuseas, emesis, epigastralgia, etc.). 4. Concentración de hemoglobina y hematocrito La definición de anemia se da cuando los valores corresponden a una disminución del 37% del valor esperado para mujeres sanas en edad reproductiva, y siempre se debe considerar que los valores tanto de Hb como del Hcto difieren entre hombres, mujeres, edad, embarazo, semanas de gestación (Tabla 1, Figura 1), esta distribución además difiere por altitud, tabaquismo y raza. En las gestantes la Hb disminuye en el segundo trimestre debido principalmente a la expansión del volumen sanguíneo (en algunos textos descrita como anemia fisiológica); en embarazadas que no consumen hierro suplementario o Hierro Tabla 1. Concentraciones máximas de Hemoglobina y valores de Hematocrito para anemia. Mujeres no embarazadas y Mujeres lactando Hb (g/dL) Hcto (%) 11.8 12.0 12.0 35.7 35.9 35.7 Semanas de gestación 12 16 20 24 28 32 36 40 11.0 10.6 10.5 10.5 10.7 11.0 11.4 11.9 33.0 32.0 32.0 32.0 32.0 33.0 34.0 36.0 Trimestre Primero Segundo Tercero 11.0 10.5 11.0 33.0 32.0 33.0 Edad (años) 12 a <15 15 a <18 > 18 Mujeres embarazadas 14.0 Hemoglobina, g/dL 13.5 Percentil 50 13.0 12.5 Percentil 5 12.0 11.5 11.0 10.5 Figura 1. Variación de la concentración de hemoglobina según la semana de gestación. 0 4 8 12 16 20 24 28 32 36 40 Semanas de gestación 103 Hierro aquellas cuyos aportes provenientes de la dieta son inadecuados, se puede observar el sostenimiento de valores bajos de Hb en el tercer trimestre, contrario a lo que ocurre con las embarazadas que consumen hierro adicional, cuyos valores se pueden incrementar gradualmente entre el segundo y tercer trimestre logrando en la mayoría, los valores previos a la etapa pregestacional. Para mujeres embarazadas, una prueba que resulte con 3 desviaciones estándar más elevada que lo esperado para la población referencia (Hb > 15.0 g/dL o un Hcto > 45%) particularmente en el segundo trimestre, probablemente indique pobre ex- pansión del volumen sanguíneo. Concentraciones altas de Hb y Hcto han sido asociadas con hipertensión y morbilidad perinatal incrementada: restricción del crecimiento intrauterino (RCIU), muerte fetal, partos pretérminos y niños con bajo peso al nacer (BPN). Algunos estudios han demostrado que embarazadas con Hcto superior o igual a 43% entre la semanas 26 – 30 de gestación tenían un riesgo dos veces mayor para desarrollar parto pretérmino y cuatro veces mayor para RCIU, que aquellas mujeres que presentaban valores de Hcto entre 33 – 36%. Las mujeres que viven en zonas de gran altitud (mayores o Tabla 2. Ajuste de la concentración máxima de hemoglobina y del valor de hematocrito para anemia. Altitud (Pies) Hb (g/dL) Hcto (%) 3.000 – 3.999 4.000 – 4.999 5.000 – 5.999 6.000 – 6.999 7.000 – 7.999 8.000 – 8.999 9.000 – 9.999 10.000 – 11.000 +0.2 +0.3 +0.5 +0.7 +1.0 +1.3 +1.6 +2.0 +0.5 +1.0 +1.5 +2.0 +3.0 +4.0 +5.0 +6.0 +0.3 +0.5 +0.7 +0.3 +1.0 +1.5 +2.0 +1.0 Consumo de cigarrillos 0.5 -<1.0 paquetes /día 1.0 -<2.0 paquetes /día > 2.0 paquetes /día Todas las fumadoras 104 Hierro iguales a 3.000 pies), así como aquellas que consumen cigarrillo generan un incremento en los valores de Hb y de hematocrito. Tabla 2. den aumentar su concentración de manera independiente al estado del hierro, llegando a producir la elevación de ferritina un enmascaramiento del depletamiento del hierro de reserva. 5. Ferritina Mujeres en edad reproductiva que poseen una ferritina baja (menor o igual a 12 ug/L) por deficiencia de hierro, la sensibilidad de la prueba es del 75%, y la especificidad del 98%; cuando la concentración de ferritina es menor a 12 ug/L, la sensibilidad para deficiencia de hierro es del 61% y la especificidad es del 100%. Aunque las bajas concentraciones de ferritina son un indicador temprano de bajas reservas de hierro, ha sido cuestionado que los valores normales de ferritina durante el primer y segundo trimestre puedan llegar a predecir un adecuado estado del hierro en la parte final de la gestación. Otros factores diferentes al nivel de reserva de hierro pueden interferir en los resultados individuales de la ferritina, por ser esta un reactante de fase aguda. La infección crónica, la inflamación o las enfermedades que causan daño tisular o de órganos (Ej. Hepatitis, cirrosis, neoplasias, o artritis) pue- 6. Efecto de la anemia sobre la morbi-mortalidad materna Aproximadamente 600.000 mujeres mueren cada año como resultado de complicaciones de la gestación, la mayoría son prevenibles. En el mundo, la tasa de mortalidad materna es de 390 por 100.000 embarazos, la mayoría de estas muertes en países en vías de desarrollo. Estudios retrospectivos muestran una relación entre esta mortalidad y la presencia de anemia en las gestantes. Se sugiere fuertemente que la anemia materna se encuentra asociada a incremento del riesgo a padecer infección, fatiga y pérdidas sanguíneas incrementadas durante el parto y puerperio. En América Latina se estima que el 3% de las muertes maternas son atribuibles directamente a la anemia y el número de días de vida perdidos por la anemia materna (por 100.000 nacidos vivos y por año) es del orden de 114.495 días, situación que debe considerarse de gran preocupación. En un estudio de 105 Hierro la OMS que incluye 66 reportes de 33 países, se sitúa a la anemia como responsable directa de las muertes maternas en el 26% de los informes y como causa indirecta en las restantes. Vale la pena anotar que los criterios usados para definir la anemia difieren entre los diferentes estudios y la percepción del obstetra es en la mayoría de los casos la que determina el diagnóstico. La causa directa de la muerte por anemia es la descompensación cardiaca. Cuando la Hb es menor de 8 g/dL, se incrementa la acidosis láctica y se presenta disnea en reposo. La falla cardiaca suele ocurrir cuando la Hb es menor de 4 g/dL. La anemia aguda puede ser una causa primaria de muerte, como las crisis hemolíticas de la anemia de células falciformes, mientras que la anemia crónica es un factor contribuyente, especialmente como consecuencia de hemorragia o infección. La mortalidad en gestantes con hematocrito menor de 14%, es de 27% sin transfusión y de 1,7% al transfundirse. Hay un incremento marcado de la mortalidad materna cuando la Hb desciende por debajo 5 g/dL y alcanza a superar el 50% en situaciones donde la Hb es menor de 3 g/ 106 dL. En mujeres con alto riesgo de infección, el hecho de tener anemia crónica aumenta sustancialmente el riesgo de morir (RR 2,1 CI 95% 1,3-3,4). En la adolescencia (menores de 19 años), la OMS ha estimado que la incidencia de anemia puede ser hasta de un 45%. Igualmente importante es el aumento en las dietas vegetarianas y la ingesta de comidas “Light” entre las adolescentes, que conlleva a un incremento en el riesgo de padecer anemia crónica. La gestación en mujeres adolescentes potencia el riesgo de complicaciones asociadas, siendo un factor causal de mucha importancia la presencia de anemia en la embarazada menor de 20 años. 7. Causas de anemia asociadas con mortalidad materna La anemia durante la gestación es multifactorial. La anemia más común en nuestro medio es la anemia por deficiencia de hierro. La anemia hemolítica es frecuente en zonas tropicales donde la malaria está igualmente presente. Pero aún en áreas endémicas para malaria, la anemia por deficiencia nutricional del hierro es la que aporta el mayor compo- Hierro nente de la mortalidad materna por anemia severa. 8. Anemia ferropénica y duración de la gestación En varias publicaciones se encuentran afirmaciones referente a que la anemia ferrropénica en el embarazo temprano se asocia con la presencia de niños con bajo peso al nacer y a mayor incidencia de partos de pretérmino. La anemia (Hb menor de 10,4 g/dL) diagnosticada entre las semanas 13 y 24 de gestación presenta un riesgo relativo (RR) de 1,18 a 1,75 para desarrollar parto pretérmino, bajo peso al nacer y mortalidad perinatal. Al tomar en consideración múltiples variables de confusión, y definir la anemia en la gestación como variable principal, se puede encontrar que el riesgo de presentar parto de pretérmino y tener hijos con bajo peso al nacer es tres veces mayor en pacientes con anemia por deficiencia de hierro, comparada con aquellas que no presentan esta patología. 9. Anemia materna: salud perinatal y neonatal El Proyecto Colaborativo Perinatal (PCP), que incluyó 50.000 embarazos mostró que tanto los ba- jos (menores de 29%) como los altos (mayores de 39%) niveles de hematocrito se asociaron con muerte fetal, parto pretérmino, niños con bajo peso al nacer y mayor número de complicaciones médicas del embarazo (no especificadas). En anemias leves (Hcto 33%) el RR de las complicaciones es muy bajo (0,33) pero en las anemias severas (Hcto < 25%) el RR es mucho mayor (de 2 a 3). Una asociación entre anemia materna y bajos puntajes de Apgar ha sido también documentada. Altas concentraciones de Hb materna durante el parto se relacionan con mejores puntajes de Apgar y con menor riesgo de asfixia neonatal. Las mujeres con anemia ferropénica tienen un riesgo incrementado de presentar neonatos con bajo peso al nacer y prematurez, producto de una restricción en el crecimiento intrauterino. Se sugiere que la anemia ferropénica puede ser un factor de pobre resultado perinatal cuando se presenta temprano en la gestación. 10. Impacto de la suplementación de hierro en la gestación Hay pocas dudas acerca de los beneficios de suplir con hierro a 107 Hierro las embarazadas. Aún en los países industrializados, se sabe que esta conducta incrementa los niveles de Hb, de ferritina, del volumen eritrocitario, del hierro sérico y de la saturación de transferrina. Suplir con hierro a las gestantes a partir del segundo trimestre, resulta siempre en mejores niveles de Hb y de ferritina en el postparto. No se ha encontrado relación entre niveles de Hb materna y de sangre de cordón, sin embargo, sí se relaciona con los niveles de ferritina en la sangre de los niños a los dos meses de edad. Mucho más condicionante es la suplementación de hierro durante el embarazo, en regiones donde la prevalencia de la anemia ferropénica es alrededor del 40%, tal y como está registrado para América Latina, situación que es frecuente encontrar en muchas regiones de Colombia. 11. Principios terapéuticos generales La eficacia del tratamiento con hierro se evalúa a través de la medición de reticulocitos, Hcto, Hb y los índices eritrocitarios (Información obtenible de un hemograma computarizado). Confirmada la respuesta adecuada al tra- 108 tamiento instaurado, éste debe continuarse hasta restablecer los valores normales de Hb y Hcto y se extenderá en caso de pretender repletar los depósitos tisulares hasta tres meses posparto. Para muchas embarazadas con pobres reservas de hierro la suplementación farmacológica regular desde el principio del 2º trimestre no alcanza a revelar incrementos de los valores de hemoglobina, pues la ingesta del hierro es apenas la necesaria para sustituir las necesidades fetales, por ello no logramos observar variaciones significantes en la respuesta por laboratorio. Sin embargo el resultado fisiológico se está sucediendo. 11.1. Hierro oral Los altos requerimientos fisiológicos de hierro en el embarazo son por lo general difíciles de alcanzar con la mayoría de las dietas de la población, por tanto la mujer embarazada debe recibir suplementos de hierro desde el inicio del 2º trimestre, para prevenir la anemia; una dosis promedio entre 30 a 60 mg/día de hierro elemental o realizar tratamientos formales en el caso que se diagnostique anemia, situación en la que se recomienda suministrar una Hierro dosis de 60 a 120 mg/día de hierro elemental desde el primer trimestre. El hierro oral está indicado como primera línea en casos de anemias leves a moderadas con Hb superior a 9.0 g/dL y Hcto mayor de 27%. Las indicaciones para la suplementación de hierro en el embarazo siempre deben contemplar la prevalencia de anemia en la región donde vive la paciente, en las zonas donde la prevalencia de anemia en gestantes es menor al 40% la dosis recomendada es de 60 mg/día de hierro elemental durante los últimos seis meses de la gestación, pero si reside en un área con prevalencia mayor al 40% de anemia en el embarazo la recomendación es de mínimo 90 - 120 mg/día de hierro elemental durante los últimos seis meses de la gestación y continuar durante los tres primeros meses pos-parto (Esta última recomendación aplica para todos los casos). La duración total y la dosis calculada dependerán del grado de anemia de la paciente, y tendrán que ser individualizadas de acuerdo a la clasificación. La absorción de hierro se incrementa hasta 10 veces en la mujer embarazada, pero los valores de este incremento varían desde un 1.5 % hacia la semana 12 hasta un 14.6 % al final de la gestación. Los suplementos de hierro oral que se encuentran disponibles en formas muy solubles en agua y/o en soluciones ácidas diluidas, como la que encontramos en el estómago, presentan una mayor biodisponibilidad, sin embargo hay que considerar la tolerancia gástrica y la presencia de otros efectos colaterales que pueden contribuir positiva o negativamente en la absorción. Estos aspectos dependen fundamentalmente de dos condiciones, la forma iónica o no iónica y si el mecanismo de absorción es activo o pasivo; los compuestos que vienen en forma no iónica (férrica), los cuales se absorben en forma activa tienen menos efectos secundarios y por lo tanto una mayor adherencia al tratamiento, condición que es importante considerar pues el resultado final es fruto de la continuidad en la toma. Los efectos adversos son principalmente de tipo gastrointestinal, entre los cuales se cuentan la pirosis, náuseas, epigastralgia, diarrea o estreñimiento y la tinción de los dientes. Para reducir estos efectos colaterales pueden admi- 109 Hierro nistrarse inicialmente dosis bajas para comprobar la ausencia de los efectos secundarios e ir incrementándolas cada 1 a 2 semanas hasta alcanzar la dosis deseada. En dosificaciones de 200 mg de una sal ferrosa, los efectos adversos aparecen hasta en el 25 % de las pacientes, este porcentaje aumenta hasta el 40% si se duplica la dosis. La adición de fructosa u otros azúcares como la polimaltosa, contribuyen a incrementar la tolerabilidad de los preparados de hierro oral y la adición de ácido ascórbico favorece una mejor absorción intestinal y elevar los niveles en la médula ósea. La dosis terapéutica de hierro elemental es de 60 a 120 mg/día, una vez al día o fraccionada en 2 o 3 tomas, hecho que se podría lograr más fácilmente con preparados líquidos, y que para muchas embarazadas favorece la tolerancia a largo plazo. Entre los preparados de hierro, los estudios clínicos realizados con hierro polimaltosado (IPC) demuestran que el efecto es equivalente al de las sales ferrosas. Sin embargo, el IPC produce un número significativamente menor de efectos adversos gastrointestinales, en comparación con las sales 110 ferrosas. De hecho, se puede prevenir adecuadamente la anemia durante el embarazo con el tratamiento con IPC más ácido fólico, aún durante el segundo trimestre del embarazo, cuando los requerimientos de hierro son más altos. Su buen grado de tolerabilidad y la ausencia de efectos secundarios adversos observados mejora considerablemente el cumplimiento del tratamiento. 11.2. Hierro parenteral Como alternativa importante para el manejo de la anemia ferropénica se encuentra el hierro en preparaciones parenterales, con la ventaja de lograr una recuperación más rápida de los depósitos tisulares. Las indicaciones más frecuentes en obstetricia son las gestaciones con anemia ferropénica moderada o severa, alteraciones del tracto gastrointestinal que afecten fuertemente la absorción, presencia de efectos colaterales intolerables por parte de las pacientes con el uso de hierro oral, contraindicaciones para transfusión de hemoderivados, terapia conjunta con eritropoyetina, falta de adherencia a la terapia oral, rechazo al tratamiento oral, programas de autotransfusión predepósito como gestaciones que cur- Hierro sen con miomatosis severa, placenta previa y/o ácreta, etc., las cuales presentan altísimo riesgo de hemorragia masiva intraparto e intra cesárea, así como en el posparto inmediato. De los hierros parenterales disponibles, el hierro sacarato o sucrosa, presenta un excelente perfil de seguridad, su utilización durante la gestación tiene gran respaldo en publicaciones y estudios randomizados. Es un medicamento clasificado en la categoría B para uso en obstetricia, siempre debe tenerse una indicación clara para iniciar su aplicación. La seguridad del medicamento ha sido documentada ampliamente en pacientes con insuficiencia renal crónica en manejo concomitante con eritropoyetina y en múltiples estudios para el manejo de anemia ferropénica en mujeres gestantes y durante el posparto. Los efectos adversos del hierro sacarato son muy escasos, en la mayoría de las oportunidades es bien tolerado. Los efectos indeseables suceden en menos del 1% de las pacientes, según lo demostrado en diferentes ensayos clínicos, los más observados son: el sabor metálico durante la infusión, prurito, cefalea, febrículas, astenia y malestar general de corta duración, con mucha menor frecuencia se ha reportado hipotensión de fácil manejo, dolor torácico, náuseas, vómito, dolor abdominal, elevación de enzimas hepáticas, vértigo, calambres en extremidades, disnea pasajera, tos y reacción en el sitio de aplicación. El hierro sacarosa logra incrementar los niveles de hemoglobina rápidamente, permitiendo preparar a la gestante mucho más rápido, favoreciendo las condiciones durante el desarrollo fetal y preparto. Una dosis diaria de 200 a 300 mg de hierro sacarosa logra incrementar en 1 a 1,5 g/dL el valor de Hb. Se recomienda aplicar dosis de 100 – 200 – 300 mg/ día, unas dos o tres veces por semana, y evaluar los niveles de Hb una semana después de la última dosis suministrada. Por sus características fisicoquímicas el hierro sacarosa se debe disolver en solución salina normal al 0,9% (SSN). El autor con base en su experiencia, recomienda diluir cada ampolla de 100 mg en 100 ml de SSN e infundir en 30 a 60 minutos. Así como ejemplo práctico se tendría que una dosis de 300 mg de hierro sacarosa se diluirían en 300 ml de SSN y se infundirían en 3 horas, cada 3 días 111 Hierro hasta obtener el valor deseado tanto en Hb, Hcto, como en ferritina. La recomendación general según los niveles de hemoglobina, para iniciar el manejo de hierro parenteral en la anemia del embarazo es encontrar una hemoglobina menor de 10,5 g/dL, para pacientes embarazadas con hemoglobinas entre 10,5 y 12 g/dL, utilizar hierro oral a una dosis de 100 a 120 mg/día, y con hemoglobinas mayores de 12 g/dL, emplear una dosis de 60 -100 mg/día de hierro oral, las cuales podrían suministrarse día de por medio o 3 dosis por semana. Referencias 1. Allen LH. Anemia and iron deficiency: effects on pregnancy outcome. Am J Clin Nutr 2000; 71 suppl: 1280S-4S. 2. Allen LH., Pregnancy and iron deficiency: Unresoved Issues. Nutr Rev 1997; 55(4): 91-101. 3. Bolton FG, Street MJ, Pace AJ. Changes in erythrocyte volume and shape in pregnancy. Obstet Gynecol Surv 1983; 38:461. 4. Beguin Y, et al. Blunted erythropoietin production and decreased erythropiesis in early pregnancy. Blood 1991; 78: 89-93. 5. Bruce D, Bigelow N. Automated reticulocyte analysis. Clinical Practice and Associated New Parameters. Hematol Oncol Clin North Am 1994; 8:617-629. 6. Canaval H., Cifuentes R., y col. Obs- 112 tetricia. Tomo I. Centro Editorial Catorse. Cali. 5ª Edición, cap: Nutrición en el Embarazo. 2000: 79-100. 7. Canaval H., Cifuentes R., Ginecología y Obstetricia Basadas en las Evidencias. Canaval H., Editorial Distribuna, cap: Anemia en el Embarazo. Cali. 2002; 231-239. 8. Canaval H. Cifuentes R., Lomanto A. Texto de Ginecología y Obstetricia. Sociedad Colombiana de Ginecología y Obstetricia. Primera Edición, cap: Anemia en el Embarazo. 2004. 9. Centers for Disease Control and Prevention. Criteria for anemia in children and childbearing- aged women. MMWR Morb Mortal Wkly Rep 1989;38:400–404. 10. Cohen JH, Hass JD. Hemoglobin correction factors for estimating the prevalence of iron deficiency anemia in pregnant women residing at high altitudes in Bolivia. Pan Am J Pub Health 1999; 6:392–396. 11. De Leeuw NKW, Brunton L. Maternal hematologic changes, iron metabolism, and anemias in pregnancy. In Goodwin JW, Godden JO, Chance GW et al (eds): Perinatal Medicine, 1976 Baltimore, Williams & Wilkins, pp 425–446. 12. Dirren H, Logman MHGM, Barclay DV, Freire WB. Altitude correction for hemoglobin. Eur J Clin Nutr 1994; 48: 625–632. 13. Galen RS, Gambino SR. Beyond normality: the predictive value and efficiency of medical diagnosis. New York: John Wiley & Sons, 1975. 14. Goodlin RC: Why treat physiologic anemias? J Reprod Med 1982; 27: 639. 15. Horowitz JJ, Laros RK: Anemia and pregnancy: A review of the pathophysiology, diagnosis, and treatment. J Contin Educ Obstet Gyne- Hierro col 1979; 29(2): 9. 16. Kitay DZ: Assessing anemia in the pregnant patient. Contemp Ob/Gyn 1973; 2: 17. 17. Krause J. The automated white blood differential, a current perspective. Hematol Oncol ClinNorth Am 1994; 8:605-617. 18. Medicina Interna. Farreras y Rozman. 1995 Medicina Interna. Rodes Teixidor. 19. Miale JB. Application of statistics and quality assurance in hematology. En Miale JB. Ed. Laboratory Medicine Hematology. The C.V Mosby Company 1982; 367-387. 20. Morrison JC. Anemias and hemoglobinopathies. In Ryan GM Jr (ed): Ambulatory Care in Obstetrics and Gynecology, New York, Grune & Stratton, 1980; pp 205–238. 21. Morrison JC, Roberts WE: Evaluation of anemia in pregnancy. In Kitay DZ (ed): Hematologic problems in pregnancy. Philadelphia, McGraw Hill, 1985 22. Morrison JC: Red blood cell disorders. In Gleicher N (ed): Principles of Medical Therapy in Pregnancy. New York, Plenum Press, 1985. 23. PATH, OMNI. Anemia detection in health services Guidelines for program managers. 1996. 24. Restrepo M. Cifras del Hemograma normal en Antioquia. Antioquia Médica. 1970; 20: 95. 25. Ryan DH. Automated Analysis of Blood Cells. En Hoffman R., Benz E. Eds. Hematology. Basic principles and practice, Boston : Churchill Livingstone Inc. 26. Scholl, T. O., Hediger, M. L., Fischer, R. L. & Shearer, J. W. Anemia vs iron defi- ciency: increased risk of preterm delivery in a prospective study. Am J Clin Nutr 1992; 55: 985–988. 27. Steer P., Maternal haemoglobin concentration and birth weight. Am J Clin Nutr 2000; 71 suppl: 1285S-7S. 28. Stoltzfus, Rj; Dreyfuss, ML. Guidelines for the use of iron supplements to prevent and treat iron deficiency anemia. INACG, 2003. 29. The INACG Steering Committee. Document by Dr. Penelope Nestel. Adjusting Hemoglobin values in program surveys. 2003. 30. United Nations Children´s Fund, United Nations University, World Health Organization. Iron deficiency Anaemia assessment, prevention, and control: A guide for programme managers. 2001. 31. Wagner Patrick. La Anemia: Consideraciones Fisiopatológicas, Clínicas y Terapéuticas. Anemia Working Group Latinoamérica. Lima – Perú. 2004. 32. Wallerstein R: Iron metabolism and iron deficiency in pregnancy. Clin Haematol 1973; 2: 453. 33. WHO/UNICEF/UNU, Iron deficiency anaemia, assessment, prevention and control: a guide for programme managers. WHO/NHD/01.3. Geneva: WHO, 2001. 34. World Health Organization. The prevalence of anaemia in women: a tabulation of available information. 2nd ed. Geneva: WHO, 1992. 35. Zhou, L. M., Yang, W. W., Hua, J. Z., Deng, C. Q., Tao, X. & Stolzfus, R. J. Relation of hemoglobin measured at different times in pregnancy to preterm birth and low birth weight in Shanghai, China. 1998. 113 Hierro Jorge Vargas S e calcula que alrededor de 100 millones de personas en el continente americano sufren de anemia ferropénica, siendo las mujeres embarazadas y los niños pequeños quienes presentan las prevalencias más altas. Además, es posible que por cada anémico exista por lo menos una persona más que presenta deficiencia de hierro, por lo que la magnitud del problema se torna indescriptible. En los países del Caribe se notifican prevalencias del orden de 60% entre las mujeres embarazadas. Infortunadamente, pocos países cuentan con información detallada acerca de la prevalencia de anemia. Así, Ecuador, por ejemplo, notificó una prevalencia nacional de 70% en los niños de 6-12 meses de edad, y de 45% en aquellos de 12-24 meses. Cuba informó que 64% de los niños de 1-3 años sufren de anemia; en Misiones, Argentina, la prevalencia es de 55% en los niños de 9-24 meses, y en México, de 50.7% en una muestra de 152 niños cuya edad oscilaba 114 entre los 6 y los 36 meses. En general, la población más afectada corresponde a los recién nacidos de bajo peso, los menores de dos años y las mujeres embarazadas.1 La deficiencia de hierro se asocia con alteraciones en muchos procesos metabólicos que pueden tener impacto en la función cerebral; entre ellos están el transporte de electrones en la mitocondria, la síntesis y degradación de neurotransmisores, la síntesis proteica, la organogénesis y otras. Es necesario separar los aspectos del desarrollo de la deficiencia de hierro y la función nerviosa de los aspectos de la deficiencia de hierro que puede ocurrir en cualquier momento de la vida. 1. Lactantes Existen diferencias sustanciales entre los sexos en la Hb y otros indicadores del estado de hierro durante la lactancia. Algunas de estas pueden ser determinadas genéticamente, mientras que otras parecen reflejar una inciden- Hierro cia incrementada de deficiencia de hierro verdadera en los niños.5 examinar los efectos de la de la DH y la ADH sobre los desenlaces mentales. 2. Escolares El hematocrito (Ht), como un indicador de la anemia en individuos y poblaciones, muestra variaciones en relación a los factores mesológicos y genéticos. La anemia por deficiencia de hierro (ADH) es un problema importante de salud pública que se calcula afecta a más de 2.000 millones de personas en todo el mundo. El hierro es un nutriente esencial no solamente para el crecimiento normal, la salud y la sobrevida de los niños, sino también para su desarrollo mental y motor normal así como para una buena función cognoscitiva. La anemia por deficiencia de hierro (ADH) se asocia con desempeño significativamente más pobre en las escalas de desarrollo psicomotor y mental y en las calificaciones de comportamiento de los lactantes, puntajes más bajos sobre las pruebas de función cognoscitiva en niños preescolares y puntajes más bajos sobre las pruebas de función cognoscitiva y pruebas de avance educativo en niños escolares. Se han llevado a cabo numerosos estudios para 3. Alteraciones ocasionadas por la anemia El cerebro es el sitio más significativo de concentración de hierro en el cuerpo humano. En ciertas regiones del cerebro, la concentración de hierro es igual o incluso más alta que en el hígado, «considerado el depósito de hierro del organismo». La concentración de hierro puede llegar a ser tan alta como 21.3 mg/100 mg de peso fresco comparada con 13.4 mg/100 mg para el hígado. La importancia del hierro durante los primeros años de vida se vuelve más evidente cuando se considera que 80% del total de hierro en el cerebro que se encuentra en los adultos ha sido almacenado en sus cerebros durante la primera década de la vida. El cerebro de un niño se desarrolla durante los 9 meses del embarazo y en el primer mes de vida. Cuando se mide el log de la concentración de ADN, el incremento más dramático de neuronas ocurre durante el embarazo pero continúa en los primeros años de vida. 115 Hierro 3.1. Lactantes y niños jóvenes (6-24 meses) Los lactantes y niños jóvenes con ADH a menudo muestran dificultad con el lenguaje, pobre coordinación motora y del equilibrio, y quizás más evidente, calificaciones más pobres en atención, capacidad de respuesta y evaluaciones del humor. Se ha postulado, que la atención más pobre, el desempeño más pobre sobre la tareas motoras, o ambos, pueden mediar los puntajes más bajos sobre las pruebas de desarrollo mental. La mayoría de los estudios de intervención en los cuales se ha administrado tratamiento con hierro durante menos de 2 semanas utilizaron un diseño doble ciego, aleatorio, controlado con placebo. 3.2. Pre-escolares (2-5 años) Los beneficios cognoscitivos del tratamiento con hierro en niños preescolares son más aparentes que en los lactantes. Aunque todavía no se comprenden las razones para esto, puede ser que las pruebas disponibles para uso en niños mayores son inherentemente más sensibles o que los niños preescolares han pasado la edad crítica en la cual la ADH puede tener efectos de larga duración. 116 Se piensa que las principales áreas que se afectan en los preescolares son la atención, el entusiasmo y la motivación más que las capacidades cognoscitivas básicas. La distinción entre atención y problemas de adquisición de conceptos es de particular interés porque las dificultades en la atención pueden ocultar capacidades cognoscitivas en los niños. Tales dificultades pueden, a largo plazo, resultar en desempeño más pobre sobre las pruebas de función cognoscitiva. También es posible que la atención más pobre y la irritabilidad sean reacciones de los niños a tareas más allá de sus capacidades. 3.3. Escolares y adolescentes (5-16 años) Existe fuerte evidencia que entre los niños escolares, los puntajes inicialmente más bajos sobre las pruebas del área cognoscitiva o el logro escolar debidos a la ADH pueden ser mejorados y en algunos casos incluso revertidos después del tratamiento con hierro. Una razón para esta evidencia puede ser el gran número de ensayos controlados con placebo, los cuales pudieron capturar los efectos del tratamiento. Otra razón puede ser la sensibilidad incre- Hierro mentada de las pruebas usadas. Alternativamente, podría ser que los efectos de la ADH en los niños escolares son más transitorios que en los lactantes y por lo tanto responden más a los efectos del tratamiento con hierro. Así como con los estudios en lactantes y pre-escolares, parece haber una indicación de alteraciones en la atención y el comportamiento en niños con ADH. Es posible que los puntajes más bajos en la basal en la función cognoscitiva en niños con DH y ADH fueran el resultado de insuficiencia hematológica más temprana en la vida. En verdad, si los factores que predisponen a los niños escolares a la DH o la ADH no son recientes, existe una probabilidad incrementada de que estos niños hayan sido anémicos antes, como lactantes y como preescolares (niños jóvenes). 4. Suplementación Existen diversos preparados de suplementos de hierro. Se puede administrar al niño una dosis de 3 a 6 mg/kg/día en la forma de sulfato ferroso antes del desayuno. Sin embargo, se presentan efectos colaterales indeseables que sobrevienen habitualmente cuan- do se toman preparados iónicos de sales de hierro tales como irritación gastrointestinal, manchas en los dientes, sabor metálico y reducción de la capacidad de absorción del hierro debido a interacciones con alimentos o medicamentos que contienen grupos fenólicos (p. ej. las tetraciclinas). El hierro polimaltosado es un complejo esencialmente no iónico que no produce los efectos de las sales ferrosas. Además de una tolerancia excelente, asegura un margen de seguridad elevado. La toxicidad oral aguda del complejo de hierro polimaltosado es tan baja que la posibilidad de intoxicación con hierro debida a una sobredosis accidental puede ser excluida, a diferencia de las sales ferrosas. La biodisponibilidad del complejo de hierro polimaltosado no iónico, es superior a la de las sales de hierro. Esto resulta en una utilización rápida del hierro administrado en la síntesis de hemoglobina y de mioglobina, así como en el restablecimiento de los depósitos de hierro. Simultáneamente, mejoran los síntomas observados en los estados de deficiencia de hierro. La dosis de hierro polimaltosado es de 20 gotas al día (1 117 Hierro gota = 2,5 mg) por kg de peso, según prescripción médica o 1 cucharadita (5 ml) de jarabe al día, según prescripción médica. En los casos de anemia aguda se puede incrementar la dosis a 2 gotas/kg, la cual no incrementa los efectos secundarios. Un incremento en los niveles de hemoglobina mayor de 1.0 g/dL (10.0 g/L) por cuatro semanas es diagnóstico de anemia por deficiencia de hierro y amerita la continuación de la terapia por dos a tres meses adicionales para aprovisionar apropiadamente los depósitos de hierro.11 Durante este tiempo, se puede proporcionar intervención dietaria adicional y educación al paciente. Si la anemia recurre, se justifica un revisión para identificar la fuente de pérdida de sangre oculta. Estudios en modelos de rata demostraron que la anemia por deficiencia de hierro en la vida temprana causa una deficiencia en los receptores de dopamina que no podría ser corregida revirtiendo la anemia.15,16 Por lo tanto, es imperativo intentar prevenir la deficiencia de hierro en los niños antes del segundo año de vida. Las estrategias para la prevención de la anemia por deficiencia de hierro 118 pueden reducir la probabilidades de desarrollar la enfermedad. Ellas son: 1) Alimentar con leche materna durante los primeros seis a 12 meses de edad. 2) Si se usa fórmula, únicamente tomar fórmula fortificada con hierro. 3) No tomar leche de vaca durante el primer año de vida debido al incremento en el sangrado gastrointestinal oculto. 4) Cuando se introducen alimentos sólidos a los cuatro a seis meses de edad, se debe hacer con cereales enriquecidos con hierro. Referencias 1. Oski FA, Honig AS. The effects of therapy on the developmental scores of iron-deficient infants. J Pediatr. 1978 Jan;92(1):21-5. 2. Oski FA, et al. Effect of iron therapy on behavior performance in nonanemic, iron-deficient infants. Pediatrics. 1983 Jun;71(6):877-80. 3. Idjradinata P, Pollitt E (1993) Reversal of developmental delays in iron-deficient anemic infants treated with iron. Lancet 341:1-4. 4. Walter T, et al. Iron deficiency anemia: adverse effects on infant psychomotor development. Pediatrics. 1989 Jul;84(1):7-17. 5. Earl RO, Woteki CE. Iron deficiency anemia: recommended guidelines for the prevention, detection, and management among U.S. children and women of childbearing age. Washington, D.C.: National Academy Press, 1993. 6. Eden AN, Mir MA. Iron deficiency in 1- Hierro to 3-year old children. A pediatric failure? Arch Pediatr Adolesc Med 1997;151:986-8. 7. Siberry GK, Iannone R, eds. The Harriet Lane handbook. 15th ed. St. Louis: Mosby, 2000. 8. Oski FA. Iron deficiency in infancy and childhood. N Engl J Med 1993;329:190-3. 9. Patton WN, Cave RJ, Harris RI. A study of changes in red cell volume and haemoglobin concentration during phlebotomy induced iron deficiency and iron repletion using the Technion H1. Clin Lab Haematol 1991;13:15361. 10. Bessman JD, Feinstein DI. Quantitative anisocytosis as a discriminant between iron deficiency and thalassemia minor. Blood 1979;53:288-93. 11. Reeves JD. Prediction of therapeutic response to iron. In: Oski FA, Pearson HA, eds. Iron nutrition revisited: infancy, childhood, adolescence. Columbus, Ohio: Ross Laboratories, 1981:114-25. 12. Walter T, DeAndraca I, Chadud P, Pe- rales CG. Iron deficiency anemia. Pediatrics 1989;84:7-17. 13. Lozoff B, Brittenham GM, Wolf AW, McClish DK, Kuhnert PM, Jimenez E, et al. Iron deficiency anemia and iron therapy effects on infant developmental test performance. Pediatrics 1987;79:981-95 [Published erratum appears in Pediatrics 1988;81:683]. 14. Lozoff B, Jimenez E, Wolf AW. Longterm developmental outcome of infants with iron deficiency. N Engl J Med 1991;325:687-94. 15. Ben-Shachar D, Ashkenazi R, Youdim MB. Longterm consequences of early iron-deficiency on dopaminergic neurotransmission in rats. Int J Dev Neurosci 1986;4:81-8. 16. Yehuda S, Youdim ME, Mostofsky DI. Brain irondeficiency causes reduced learning capacity in rats. Pharmacol Biochem Behav 1986;25:141-4. 17. American Academy of Pediatrics Committee on Nutrition. The use of whole cow’s milk in infancy. Pediatrics 1992;89:1105-9. 119 Hierro Jorge Vargas E ntre las medidas que se han propuesto para prevenir la anemia por deficiencia de hierro se cuentan: 1) suplementación con hierro medicinal, 2) educación y otra medidas que contribuyan a elevar la ingesta de hierro dietario, 3) el control de las infecciones e infestaciones parasitarias y la fortificación de la alimentación básica con con hierro. 1. Suplementación con hierro medicinal La principal ventaja que ofrece es la mejoría rápida del estado de hierro. Además, puede ser dirigida a grupos de población con mayor necesidad de hierro o mayor riesgo de presentar deficiencia de hierro. La experiencia muestra que la suplementación de hierro tiene una mayor posibilidad de éxito cuando se dirige a grupos específicos, debido a que el cubrimiento de la población completa es virtualmente imposible, excepto cuando hay un sistema de distribución de salud excepcionalmente efectivo, y de to120 dos modos es innecesario. Los programas de suplementación logran el mayor éxito cuando se concentran en grupos de alto riesgo como mujeres embarazadas, lactantes y niños pre-escolares, y sobre “audiencias cautivas”, como escolares o trabajadores que pueden recibir la suplementación en la escuela y el trabajo. Sin embargo, la efectividad está restringida por dos factores importantes: los efectos colaterales gastrointestinales del hierro oral y la dificultad de sostener la motivación por 2 a 3 meses en los “pacientes”, ya que ellos no se perciben a sí mismos como enfermos. 1.1. Lactantes Entre las poblaciones rurales en la mayoría de países en desarrollo, la alimentación con leche materna exitosa y prolongada es la regla; lo contrario es más típico de las áreas urbanas, donde se deben concentrar los esfuerzos para promover la alimentación con leche materna. La leche materna es Hierro adecuada para cubrir los requerimientos de hierro dietario de los lactantes de peso normal al nacer hasta los 6 meses de edad. Los infantes de bajo peso al nacer pueden, sin embargo, requerir suplementación de hierro ya desde los 2 meses de edad. Los lactantes que requieren profilaxis de hierro pueden tomar 2-4 gotas/día de hierro polimaltosado. 1.2. Pre-escolares La suplementación de hierro en niños preescolares también es importante y requiere planeación especial. Los suplementos pueden ser administrados por los responsables de los centros infantiles, liberando así al trabajador de la salud primaria para otras tareas. Además, por supuesto, se debe aprovechar cualquier oportunidad para supervisar la ingesta de los suplementos en las clínicas pediátricas y durante otros contactos con el servicio de salud. Algunos programas promueven cursos de 2 – 3 meses de una o más tabletas de hierro al día. Aunque tal régimen es necesario para corregir la anemia severa, es dudoso si se requiere esta duración para la anemia leve a moderada que usualmente se encuentra en este grupo de edad. Para asegurar un cubrimiento más uniforme de hierro y tasas de cumplimiento más altas, puede ser más práctico darle a los niños un curso de 2 – 3 semanas con base en una dosis más baja (30 mg de hierro elemental al día en tableta o forma líquida) varias veces al año. Nuevamente, es importante tener en cuenta que los efectos colaterales pueden tener un impacto negativo en la adherencia al tratamiento. Por lo tanto, una preparación como hierro polimaltosado puede administrarse profilácticamente a dosis de 4-6 gotas/día. 1.3. Escolares Los niños en edad escolar usualmente no tienen la misma alta prevalencia de anemia como los niños preescolares. Los mejores cursos son cortos, como para los niños preescolares; la dosis diaria debe ser entre 30 – 60 mg de hierro elemental, dependiendo de la edad y peso del niño. El hierro polimaltosado que tiene una mejor tolerabilidad se puede administrar a dosis de 4-6 gotas/día. 1.4. Mujeres embarazadas Las mujeres embarazadas son un grupo prioritario. La dosis dia- 121 Hierro ria recomendada es de dos tabletas, cada una conteniendo 60 mg de hierro elemental más 250 mg de folato, tomados durante toda la segunda mitad del embarazo. El principal obstáculo para la suplementación de hierro es el pobre cumplimiento con el tratamiento. A menudo esto se debe a efectos colaterales, pero también puede provenir de la falta de conocimiento por las mujeres de que ellas tienen una necesidad real de hierro durante el embarazo. También se puede administrar profilácticamente hierro polimaltosado a dosis de 20-40 gotas/día o 5-10 mL/día de jarabe. Las mujeres deben ser convencidas de la importancia del hierro para su salud y la del ser en gestación. Es importante ser persuasivos mediante la adquisición de habilidades en las técnicas de comunicación y motivación por parte del equipo de salud. 2. Modificación dietaria La ingesta de hierro dietario puede incrementarse de dos formas. La primera es asegurar que las personas consuman mayores cantidades de sus alimentos habituales, de tal manera que satisfagan sus necesidades energéticas. Ya que no se necesitan cam- 122 bios cualitativos en la dieta, este abordaje puede parecer sencillo, pero involucra incrementar el poder de compra de los habitantes, lo cual está más allá de las capacidades del sector de la salud solo. No obstante, la importancia práctica de esta estrategia no debe ser subestimada, particularmente en situaciones donde puede ser difícil mejorar la biodisponibilidad de hierro ingerido. El aumento de la biodisponibilidad del hierro ingerido, más que su cantidad total, es el segundo abordaje básico de la manipulación dietaria. Existen cierto número de estrategias disponibles, cada una con sus ventajas y desventajas, pero todas se basan en promover la ingesta de aumentadores de la absorción de hierro, incluyendo hierro hem, o en la reducción de la ingestión de inhibidores de la absorción tales como taninos y ácido fítico. Una recomendación de incluir carne en la dieta con el fin de combatir la anemia por deficiencia de hierro se debe hacer teniendo en cuenta las limitaciones no sólo económicas sino también culturales. Por lo anterior, los esfuerzos para incrementar el contenido de ácido ascórbico en la dieta tienen una Hierro mayor posibilidad de éxito. La vitamina C tiene un notable efecto sobre la absorción del hierro no hem, siendo dependiente de la dosis. En muchas zonas rurales, los vegetales y frutas se consumen sólo infrecuentemente y en cantidades pequeñas. Por esto, persuadir a las familias a que agreguen estos alimentos a su dieta básicamente de almidón puede tener un impacto considerable. Aproximadamente 50-80% de la vitamina C originalmente presente en los alimentos se puede perder durante la cocción. Más aún, el contenido de vitamina C de los alimentos que se cocinan y se dejan en reposo disminuye considerablemente; el recalentamiento lo reduce todavía más. 3. Control de infecciones virales, bacterianas y parasitarias El cuidado efectivo, curativo oportuno, podría disminuir las consecuencias nutricionales adversas de la enfermedad viral y bacteriana. Aunque el número de episodios infecciosos probablemente se reduzca, los servicios curativos apropiados pueden al menos contribuir con una reducción en la duración y la severidad de las infecciones. Esto solo ayudaría a mejorar el estado de hierro incluso si no existe incremento en el consumo de hierro dietario. En especial, los niños preescolares, se beneficiarían de tales mejorías en el cuidado de la salud. Es vital educar a la familia acerca de prácticas de alimentación apropiadas durante y después del período de una enfermedad infecciosa. Esto es especialmente importante donde la preocupación son los niños pequeños, ya que ellos a menudo son dejados con dietas pobres cuando están enfermos. Los trabajadores de la salud primaria necesitan convencer a la familia de darle a los niños enfermos tanto líquido como sea posible y continuar alimentándolos tanto como lo toleren con una persuasión gentil. La alimentación con leche materna no debe ser interrumpida. La inmunización continúa ganando aceptación y el cuidado de salud primaria puede fortalecer esta actividad considerablemente. Sin embargo, todavía no existen vacunas disponibles contra las infecciones gastrointestinales y respiratorias más frecuentes. El control de estas in- 123 Hierro fecciones requiere medidas de salud pública preventivas. Los hematelmintos (Ancylostoma y Necator) y Schistosoma juegan un papel en la etiología de la anemia al causar pérdida crónica de sangre. El papel de otros parásitos intestinales comunes está menos esclarecido. Existe alguna evidencia de que cierto número de parásitos pueden interferir con la absorción de algunos nutrientes, particularmente si la carga de gusanos es alta; se ha mostrado que Giardia reduce la absorción de hierro. Desde un punto de vista de salud y nutricional es indeseable albergar parásitos, y frecuentemente se dan recomendaciones sobre hacer una desparasitación rutinaria como parte del cuidado de salud primario. Esto no significa que es la mejor forma de tratar el problema. La desparasitación en la ausencia de esfuerzos simultáneos para erradicar el reservorio de la infección es seguida rápidamente por reinfestación y una necesidad renovada de desparasitación. La desparasitación per se puede ser efectiva temporalmente en la disminución de la carga parasitaria pero puede ser sólo de beneficio menor en términos del nivel de 124 hemoglobina. La provisión de hierro adicional, ya sea a través de la suplementación con hierro medicinal o por fortificación de los alimentos, resulta en un incremento mucho mayor en la concentración de hemoglobina, aun cuando no se haga desparasitación. En casos individuales de anemia severa resultante de una carga de parásitos intensa, el tratamiento obviamente debe incluir la desparasitación. 4. Fortificación de alimentos La fortificación de alimentos ampliamente consumidos y procesados con hierro es la base del control de la anemia en muchos países. Es una de las formas más efectivas de prevenir la deficiencia de hierro. Puede ser dirigida para alcanzar algunos o todos los grupos poblacionales, y no necesariamente requiere la cooperación del individuo. El costo inicial es modesto, y los gastos recurrentes son menores que aquellos de la suplementación. La principal dificultad es identificar un alimento adecuado para ser fortificado y, de igual importancia, una forma de hierro que sea absorbida adecuadamente sin alterar el sabor o la apariencia del alimento. La población objetivo necesita, por su- Hierro puesto, acostumbrarse a la alimentación fortificada y debe estar en capacidad de comprarla. La fortificación con hierro es más difícil técnicamente que la fortificación con otros nutrientes porque las formas biodisponibles de hierro son químicamente reactivas y tienden a producir cambios indeseables en los alimentos. Por ejemplo, las sales ferrosas solubles a menudo producen cambios en el color, formando complejos con compuestos de azufre, taninos, polifenoles y otras sustancias. La decoloración es particularmente indeseable cuando el alimento a ser fortificado es de color claro. Además, los compuestos de hierro reactivo catalizan reacciones oxidativas, dando como resultado sabores y aromas indeseables. Ya que es improbable que las personas acepten un alimento fortificado en el cual se puede detectar el hierro agregado, los programas de fortificación han tendido a confiar sobre compuestos de hierro inertes que son pobremente absorbidos y por lo tanto más o menos inefectivos. Hoy en día, sin embargo, se están haciendo grandes esfuerzos para encontrar formas más adecuadas de fortificación de hierro. El sulfato ferroso ha sido usado extensamente para la fortificación del pan y otros productos de panadería que son almacenados solo por períodos cortos. Cuando tales elementos fortificados se almacenan por unos pocos meses, ellos desarrollan un aroma rancio. El uso de lactato ferroso y gluconato ferroso está limitado, por razones económicas, a la fortificación de productos como fórmulas lácteas para lactantes basados en soya. El hierro elemental también es ligeramente más costoso que el sulfato ferroso. De buena biodisponibilidad y estabilidad, es usado frecuentemente para fortificar la harina de trigo y el pan en Norteamérica y Europa occidental. En países industrializados, los alimentos más frecuentemente fortificados son la harina de trigo y el pan, las comidas de maíz, los productos lácteos incluyendo fórmulas para lactantes y los alimentos de destete (cereales para lactantes). En Chile, se ha observado que el uso de leche de fórmula fortificada con hierro y ácido ascórbico reduce la prevalencia de anemia a los 15 meses de edad a menos del 2%, comparada con 125 Hierro 28% entre lactantes que recibieron fórmula no fortificada. Se ha encontrado que el EDTA (tetraacetato diamino etileno), ampliamente usado en la industria de procesamiento de alimentos como un agente quelante, es un efectivo aumentador de la biodisponibilidad de hierro. La sal de hierro de EDTA ha sido utilizada exitosamente para fortificar el azúcar en Guatemala (13 mg de hierro / 100 g de azúcar). Los ensayos en campo han mostrado que su consumo (40 g por día por persona) mejora el estado de hierro de la población. Los costos del azúcar fortificado son aproximadamente 2% más que el azúcar no fortificado. Se debe reconocer que aunque la fortificación de alimentos con hierro posee pocos o ningún problema en el mundo desarrollado, se requiere una infraestructura industrial que no existe en algunos países en desarrollo, particularmente en África. Referencias 1. World Health Organization. Malnutrition: the global picture. Geneva: The Organization; 2000. 126 2. Grantham-McGregor S, Ani C. A review of studies on the effect of iron deficiency on cognitive development in children. J Nutr 2001;131:649S-668S. 3. Pollitt E. Iron deficiency and cognitive function. Ann Rev Nutr 1993;13:52137. 4. Lozoff B, Jimenez MD, Hagen J, Mollen E, Wolf AW. Poorer behavioral and developmental outcome more than 10 years after treatment for iron deficiency in infancy. Pediatrics 2000;105:E51. 5. Zlotkin SH, Ste-Marie M, Kopelman H, Jones A, Adam J. The prevalence of iron depletion and iron-deficiency anaemia in a randomly selected group of infants from four Canadian cities. Nutr Res 1996:729-33. 6. Willows N, Dewailly E, Grey-Donald K. Anemia and iron status in inuit infants from Northern Quebec. Can J Public Health 2000:91;407-10. 7. Willows N, Morel J, Grey-Donald K. Prevalence of anemia among James Bay Cree infants of Northern Quebec. CMAJ 2000:162(3);323-6. 8. Yip R. The challenge of improving iron nutrition: limitations and potentials of major intervention approaches. Eur J Clin Nutr 1997;51:516-24. 9. Dallman PR. Changing iron needs from birth through adolescence. In: Fomon SJ, Zlotkin SH, editors. Nutritional anemias. Nestle Nutrition Workshop Series. New York: Vevey/Raven Press; 1992. p. 29-38. 10. Saarinen UM. Need for iron supplementation in infants on prolonged breastfeeding. J Pediatr 1978; 93:177-80. Hierro Diego Rincón Resumen C onocido el hierro como un elemento esencial para la vida, sus concentraciones elevadas son consideradas como un factor de riesgo para algunas enfermedades y el estrés oxidativo generado, se da como una causa de algunas de estas enfermedades. De allí que el estrés oxidativo generado por el hierro vaya ganando atención, en enfermedades tan importantes como son el cáncer, enfermedades neurodegenerativas como son el Parkinson y el Alzheimer, además de enfermedades cardiovasculares. Aunque el hierro puede participar en las reacciones oxidativas para generar radicales libres bajo condiciones in vitro, su papel in vivo en las causas o en la progresión de dichas enfermedades es por el momento cuestionable. Hierro y estrés oxidativo El hierro es considerado un elemento esencial para todos los organismos vivientes; y es clave en la función del transporte y almacenamiento de moléculas de oxígeno, v.g. hemoglobina y mioglobina ; de algunas enzimas que catalizan las reacciones redox requeridas para la generación de energía, v.g. citocromos; la producción de varios metabolitos intermediarios, y para la defensa del huésped; v.g.: nicotinamida adenina dinucleótido fosfato [NADPH] (1). El ser humano estrictamente conserva el hierro, reciclando este de los eritrocitos que envejecen y de otras fuentes. La pérdida de hierro en un adulto varón típico es tan pequeña que este se puede conseguir con la absorción solamente de 1 – 2 mg de hierro al día (2). Para llegar a una comparación significativa, podemos establecer que el hierro corporal total en un varón adulto es de 3000 – 4000 mg y que diariamente sus requerimientos para la eritropoieisis es cerca de 20 mg. Tal conservación de hierro es esencial ya que algunas dietas humanas contienen tan solo la cantidad de hierro para reempla127 Hierro zar pequeñas pérdidas, sin embargo, cuando el hierro dietario es más abundante, la absorción es apropiadamente atenuada. Un mecanismo homeostático importante es prevenir la absorción excesiva de hierro en el intestino delgado proximal y regular la tasa de hierro liberado de los macrófagos involucrados en el “reciclamiento”. El hierro celular que no es usado por otras ferroproteínas se acumula en forma de ferritina cuya capacidad para el hierro es limitada, y puede ser sobrepasada luego de un período prolongado de exceso de hierro; esto puede ilustrarse muy bien, en aquellos pacientes con severas formas de hemacromatosis o con sobrecarga de hierro, cuyo hierro corporal total es mayor de 5-10 veces lo normal, allí el hierro tisular en exceso causa un amplio daño orgánico. Otra posibilidad, es que una rápida liberación de hierro desde los macrófagos crearía una sobrecarga de hierro local y podría causar un daño tisular localizado. Los efectos tóxicos del hierro libre se adscriben a su habilidad para catalizar la generación de radicales libres reactivos (1). En células sanas, los iones de hierro nunca se encuentran en un 128 estado “neutro”, sino que se encuentran generalmente quelado, usualmente por proteínas. Si el hierro está siendo transportado o almacenado, este debe estar quelado por vías específicas que “desacoplan” el ciclo redox (v,g,: por transferrina o por ferritina). Cuando el hierro se dirige hacia el ciclo redox (v.g.; en los citocromos, o peroxidasas), este es fuertemente retenido en el área circundante del sitio activo de la proteína (3). Una de las acciones más devastadoras del hierro activo libre del ciclo redox en la célula es la iniciación de la peroxidación lipídica. Esta peroxidación es una reacción en cadena de radicales libres entre los grupos acilo de los ácidos grasos poliinsaturados en las membranas celulares y el oxígeno molecular. Esto dejará una membrana disfuncional y una posible muerte celular. Quizás lo más interesante de las proteínas que contienen hierro, son aquellas que pueden liberar hierro activo libre de redox donde ocurre una reducción de 1 – electrón mediado por radicales libres (ferritina, aconitasa), o que puede tomar una nueva actividad biológica, tal como la transformación de la aconitasa citosólica en la proteína de respuesta al hierro – 1, una proteína que puede modular la eficiencia de la Hierro translación del código de RNAm de la proteína de almacenamiento de hierro (ferritina) (4). De manera más amplia se puede explicar cómo se asocia la propiedad pro-oxidante del hierro en su generación de radicales libres; el hierro unido es transportado en el cuerpo por la transferrina, mientras que el exceso de hierro es almacenado como ferritina y hemosiderina. Una pequeña cantidad del hierro corporal también se encuentra como el hierro no unido a transferrina (NTBI) y está asociado con ligandos de bajo peso molecular tal como ATP, ADP, GTP y citrato. Todas las células vivientes transportan NTBI desde el exterior de la membrana plasmática a varios organelos intracelulares (5,6). El transporte de hierro por células del epitelio bronquial humano (HBE) se involucran como una ferri-reductasa, v.gr.; reducción del ión férrico al estado ferroso. La reducción es la forma limitante de la rata de ingreso de hierro por determinadas células, y una rata lenta de la recaptación de Fe+3 a Fe+2 por las células del HBE sugiere que la reducción del Fe+3 limita el transporte del hierro más que la translocación del metal a través de la membrana plasmática. Las células del HBE toman el hierro de una manera dependiente del tiempo y de la concentración. La recaptación del hierro está inhibida por la presencia de deferoxamina, un quelante extracelular del hierro, comparable con lo que se encuentra en otros tipos celulares (7). La inhibición de la recaptación resulta directamente en la quelación del Fe+3, lo cual previene ambos la reducción a Fe+2 y el transporte celular. Ya que los queladores ferrosos también inhiben efectivamente el transporte de hierro férrico, el Fe+2 más que el Fe+3 es favorecido en la captación del metal (8). Así, el NTBI es transportado solamente después de la reducción del Fe+3, lo cual ocurre antes de la unión del metal y la translocación de la membrana por las proteínas transportadoras de metales catiónicos (9). Algunas células pueden usar pequeños compuestos orgánicos tal como el malato y el lactato para reducir el Fe+3 y facilitar la actividad de la ferri-reductasa(10). En las células del HBE, sin embargo, los ácidos orgánicos no promueven sino que además disminuyen la recaptación de hierro, probablemente por la formación de la coordinación estable de complejos con el Fe+3. En contraste, la supe- 129 Hierro róxido dismutasa de Manganeso (MnSOD) extracelular está disminuida por ambos, tanto por la reducción química de Fe+3 a Fe+2 y por el transporte de hierro en el interior de las células. De manera similar, la expresión de la superóxido dismutasa-1 (SOD-1) en el citoplasma está bloqueada la recaptación de hierro por las células. Estos experimentos indican un papel crítico de ·O2¯ en la membrana celular para la reducción del hierro Fe+3 . Un número de fuentes de generación del ·O2¯ cercanas a la membrana plasmática están presentes en células de mamíferos incluyendo la mitocondria, la NAD(P)H oxidorreductosa, xantina oxidasa, y un sistema de oxidasas de función mixta. El sistema de generación de ·O2¯ está bien caracterizado en la membrana, así como la NAD(P)H oxidorreductasa, ambos pueden reducir el hierro (11). Otra fuente de ·O2¯ en la membrana celular son las proteínas de intercambio aniónico – 2 (AE2); estas AE2 funcionan como un intercambiador de transmembrana electroneutro para cloro y el HCO3¯ (12). Cuando se presenta la eliminación extracelular HCO3¯ 130 previene ambos la reducción de hierro y el transporte de hierro por medio de las células HBE. Luego de una exposición a una alta concentración de hierro, la mayoría de células rápidamente regulan a la baja al receptor de transferían (13). Así, el transporte de hierro dependiente de transferrina usualmente no contribuye al papel antioxidante de la ferritina en el secuestro de metal por la ferritina intracelular para el almacenamiento y la detoxificación, tal como la NTBI, la cual puede ser incorporada dentro de la ferritina (14) . Dichos estudios, soportan el papel del AE2 en la recaptación del NTBI por las células epiteliales de la vía aérea por el secuestro subsecuente. Se han observado diferencias entre pulmones sanos y con enfermedad lo que sugiere un desequilibrio del hierro en la enfermedad que causa estrés oxidativo en el tracto respiratorio inferior. El metabolismo del hierro normal en el pulmón puede ser interrumpido en una variedad de vías. Entre estos se incluye la acumulación del metal por introducción de un quelante inapropiado (v.gr: bacteria, bleomicina o fibras de asbestos) y la inhibición de proteínas Hierro responsable para la recaptación del metal por las células respiratorias (v.gr: neumonitis luego de una aspiración de aceite de avión). En estas circunstancias, la concentración de hierro en el tracto respiratorio inferior está elevada, frecuentemente en asociación con elevaciones en la ferritina pulmonar. El mecanismo del transporte de hierro dentro de las células pulmonares para el secuestro por la ferritina es similar al involucrado en el AE2 (Figura 1). Todo esto soporta un nuevo concepto, y es que el AE2 es un componente obligatorio de la actividad de ferri-reductasa requerida para el transporte del NTBI por la célula epitelial respiratoria. El transporte del NTBI pudiera servir para una o varias funciones: la adquisición de un hierro metabólico o la remoción del metal catalíticamente activo desde el epitelio. En la superficie epitelial del pulmón, el último mecanismo puede servir primariamente para disminuir el estrés oxidativo además de soportar los requerimientos nutricionales ya que el metal a nivel apical es redistribuido a la superficie basal de la célula. Un efecto antioxidante, pudiera ser realizado a través de la disipación del ·O2¯, depurando el metal para el almacenamiento en una forma menos reactiva catalíticamente, o para la transferencia a un comportamiento vascular para la distribución a otros sitios de la economía corporal. Fe +3 + O- 2 Fe +2 Vía aérea AE2 HCO3 NRAMP2 (DMT1) Célula epitelial Espacio intersticial .O2 Endotelio Ferritina Figura 1. Cambios del AE2 generados por la superóxido de la célula para el bicarbonato extracelular. La superóxido reduce el hierro férrico a ferroso, y el producto ferroso es transportado al interior de la célula por el Nramp 2( DMT1) donde es retenido, p.ej. en la ferritina. DMT1: Transportador de Metal Divalente-1. (Adaptada de Ghio AJ, Nozik-Grayck E, Turi J, Jaspers I, Mercatante DR, Kole R, Piantadosi CA. Superoxide-dependent iron uptake: a new role for anion exchange protein 2. Am J Respir Cell Mol Biol. 2003 Dec;29(6):653-60). 131 Hierro El total del NTBI es pequeño en las personas sanas pero puede ser levemente elevada en personas con riesgo para sobrecarga de hierro (v.g: hemocromatosis, â – talasemia, y otras condiciones de enfermedad). Con base en la complicación del hierro en la reacción Fenton ( v.gr: produciendo radicales hidroxilo desde el H2O2) bajo condiciones in vitro, los complejos de hierro de bajo peso molecular pueden presumiblemente estar involucrados en esta reacción in vivo. Las radicales hidroxilo son las especies más reactivas de radicales de oxígeno (ROS) y se conocen por tener la habilidad de reaccionar con constituyentes celulares que incluyen residuos de aminoácidos y bases de purina / pirimidina en el DNA; y además, los radicales hidroxilo son también capaces de atacar los lípidos de la membrana celular causando peroxidación lipídica (15). Ya que la capacidad corporal para almacenar hierro es más que adecuada en individuos normales, y el hierro en circulación está estrechamente unido a la transferrina, solamente una insignificante cantidad de hierro catalítico está presente en el cuerpo. Basados en 132 los resultados de un reciente estudio que mostró que no había relación entre el NTBI y la oxidación de la Lipoproteína de baja densidad (LDL) en 77 hombres y mujeres con un estado de hierro normal (niveles séricos de ferritina de 42 – 96 µg/L). (16) Durante el estres oxidativo, sin embargo, el hierro puede ser liberado de sus proteínas, además induciendo daño oxidativo. Las ratas tratadas con el ácido 5-aminolevulítico (ALA), un precursor del grupo hemo que se acumula en la porfiria intermitente y en la intoxicación por plomo, tiene incrementada la liberación de hierro desde la ferritina de una manera dependiente del tiempo y de la dosis, lo cual puede estar relacionado con su comprometido generación de radicales libres. Con base en la inhibición de la liberación de hierro promovida por el ALA por la superóxido dismutasa (SOD), el O2 involucrado fue sugestible en la liberación de hierro desde la ferritina (17). Aunque los datos no indican que el estrés oxidativo induzca la liberación de hierro de las proteínas in vivo, esto puede parcialmente explicar porque los altos niveles de colesterol (que es una condición de estrés oxidativo) y el hierro tengan un Hierro efecto sinérgico causando aterosclerosis (15). El estres oxidativo y el daño por radicales libres a los tejidos puede estar involucrado en el desarrollo de enfermedades tales como el cáncer, enfermedades neurodegenerativas, y enfermedades cardiovasculares (18). La acumulación incrementada de hierro en los tejidos está asociada con la patogénesis de algunas enfermedades, aunque la extensión de su asociación es dependiente del tipo de células comprometidas. Por ejemplo, células que tienen una alta capacidad antioxidante, tal como los hepatocitos, son menos susceptibles, que las células del cerebro, las cuales tienen una baja capacidad antioxidante, son más susceptibles para el estres oxidativo catalizado por el hierro. Por ejemplo, las concentraciones de SOD y de catalasa son 7 y 140 veces mayor en el hígado que en el cerebro, respectivamente (4). A pesar de la alta capacidad antioxidante de los hepatocitos, la acumulación de hierro en el hígado luego de la administración de hierro dextrano tiene un efecto significante sobre sustancias reactivas al ácido tiobarbitúrico (TBARS), que son indicado- res de la peroxidación lipídica; este efecto sugiere que el hierro está involucrado en el daño oxidativo en el hígado (19). Ya que la mutagénesis causada por el daño oxidativo del DNA ha sido ampliamente reconocido como la etapa primaria para el crecimiento del cáncer, la corrección entre el hierro, ROS, y el cáncer ha recibido últimamente gran atención. Hace más o menos dos décadas atrás, por diversos trabajos, mostraron que el almacenamiento del hierro corporal y el ingreso de hierro en la dieta estaban positivamente asociados con posteriores riesgos de cáncer de colon en el hombre (20). Bajo condiciones in vitro, ambos, tanto el hierro no-hemo (NTAférrico) y el hierro hemo (hemoglobina) inducen daño del DNA en las células de colon humano células HT29), indicando como el hierro de la dieta está involucrado en el cáncer de colon (21). El riesgo para adenoma colónico fue de 4.3 veces mayor en individuos con niveles de ferritina sérico de 157 a 399 ng/mL en individiuos en el “cuartil” más bajo (8 - 43 ng/mL), aún después del ajuste para otros factores de riesgo (22); y como contraste, una dieta baja en hierro in- 133 Hierro crementa la incidencia del adenoma de pulmón en un 86% (23). El hígado es considerado el sitio primario para la acumulación de hierro, el cáncer de hígado está comúnmente asociado con hemocromatosis. Fischer y cols. (24), examinaron recientemente el efecto del estado del hierro sobre el estres oxidativo y el factor nuclear Kappa B (NF – êB) en hígados de ratas tratados con un proliferador de peroxisomas hepatogénico. Como se esperaba, se observó un moderado incremento en el estres oxidativo, indicado por el incremento de los marcadores de peroxidación lipídica, en ratas que consumieron una dieta rica en hierro comparada con las ratas que consumieron una dieta baja en hierro, pero no se observó ningún efecto sobre la doble hélice de DNA. Una dieta rica en hierro incrementa la actividad del NF – êB, el cual está influenciado por el estres oxidativo, especialmente por el H2O2. La evidencia común sugiere que el estres oxidativo puede activar el NF – êB por disociación de su proteína inhibitoria en el citosol. El NF – êB ingresa en el núcleo y altera la transcripción de genes implicado en la carcinogénesis. 134 Los resultados allí expuestos no soportan el papel del hierro en la inducción del cáncer a través de la activación del NF – êB, y pudiera llevar a la inducción por otros mecanismos; y esto es posible, ya que el radical hidroxilo producido por exceso de hierro puede no tener un efecto similar al H2O2 sobre la actividad del NF – êB, o que este requiera de concentraciones extremas de hierro como las que se observan en la hemocromatosis. El factor de necrosis tumoral – á (TNF – á) protege del daño oxidativo en los hepatocitos expuestos al tert-butil hidroperóxido por inducción de la síntesis de proteína ferritina de cadena pesada (H-ferritina) y hace que este menos disponible el hierro no unido a proteína en ratas que consumieron una dieta normal (25). El exceso de hierro puede inducir daño oxidativo por depleción de la defensa antioxidante corporal. La sobrecarga de hierro inducida por la alimentación de hierro carbonilo para reducir las tasas de concentración de antioxidante (vitamina E y ácido ascórbico en plasma) en el cuerpo, pero no afectaron la peroxidación lipídica, sugiere que el estres oxidativo inducido por el hierro puede estar Hierro relacionado con la depleción de antioxidantes (26). Existe alguna evidencia, que presenta el efecto benéfico de la vitamina E en la reducción del riesgo de cáncer inducido por el hierro, lo que soporta esta hipótesis. Ratas alimentadas con una dieta rica en hierro (280 ppm) incrementó los peróxidos en las heces y disminuyó las concentraciones de tocoferol en el colonocito comparadas con las ratas que se alimentaron con 35 ppm en la dieta (27). Cuando se adiciona tocoferol gamma no solamente se reduce los peróxidos lipídicos fecales, sino que también se reduce la expresión del rasp21, una proteína oncogénica expresada en el cáncer de colon inducida químicamente y se sobreexpresa en pacientes con cáncer de colon avanzado. Estos datos sugiere que la vitamina E contrarresta el efecto de un alto estres oxidativo inducido por el hierro. El colon está expuesto a grandes cantidades de hierro no absorbido y de esta manera reacciona de forma oxidativa. Cuando los humanos consumen 19 mg/día de hierro por (2) semanas, se incrementó cinco veces la concentración de hierro libre y un 40% se incrementó en radicales libres en las heces comparado con el pe- ríodo de pre-suplementación (28). El exceso de hierro no absorbido puede así causar daño en la mucosa y consecuentemente cáncer de colon. Por todo esto, es posible que el hiero esté involucrado en el daño por radicales libres pero no necesariamente induce cáncer la defensa antioxidante corporal y el proceso de reparación del DNA. Sin ser bien entendido, en la enfermedad de Parkinson y la de Alzheimer se han asociado con elevadas concentraciones de hierro en el cerebro relativas a la proteína de almacenamiento, la ferritina. Un incremento en los niveles de hierro sin un cambio concomitante en la ferritina provee de un hierro libre para la generación de radicales libres. Los cerebros de pacientes con enfermedad de Alzheimer muestran una interrupción en el metabolismo de hierro; así, hay una acumulación de hierro en las placas seniles y hay una distribución alterada del transporte del hierro y en las proteínas de almacenamiento. Además, en estudios porst-mortem los cerebros de pacientes con enfermedad de Parkinson revelaron acumulación de hierro en la sustancia negra. El hierro juega un papel casual en la 135 Hierro muerte celular, pero esta es controversial; la ferritina – H juega un papel en la protección del cerebro al daño, ayudando a conseguir bajar la concentración de hierro intracelular. Los cerebros de ratones mutantes tienen menos de la mitad de los niveles de ferritina-H pero poseen similar contenido de hierro comparado con el ratón “tipo – salvaje” (wild-type). El alto estrés oxidativo informado en términos de una actividad reducida de SOD acoplado con unas proteínas oxidadas incrementadas y mediciones de apoptosis incrementadas (Bax y caspasa – 3) en las neuronas de los modelos mutantes. La protección contra el 1-metil-4-fenil-1,2,3,6-tetrapiridina (MPTP) – induce al Parkinson por la administración oral de un quelante de metal o por expresión transgénico de la ferritina, soportan el papel del hierro en las enfermedades neurodegenerativas (29). Los cambios en la homeostasis del hierro son el mayor factor contribuyente en las enfermedades neurodegenerativas, pero no está claro el porqué estos cambios ocurren con la vejez. No hay datos que sugieran que el exceso de hierro en el cuerpo pueda ser un 136 factor de riesgo para el desarrollo de desórdenes neurológicos, a pesar de la falta de investigaciones en el área, los estudios hasta ahora realizados muestran que la terapia de quelación de metal es una aproximación promisoria para mejorar la calidad de vida de la población anciana. Los radicales libres pueden también inducir oxidación lipídica y aterosclerosis, son causas directamente relacionadas con enfermedad cardiovascular. El hierro de la transferrina puede ser liberado a valores de pH que pueden ser vistos en lesiones ateroscleróticas en la pared arterial (levemente inferior al pH fisiológico) y puede causar oxidación de las lipoproteínas de baja densidad (LDL). En estudios in vitro se demostró que la lipólisis inducida por la lipoproteín lipasa reducía el pH fisiológico de 7.4 a 7.0 y estos eventos lipolíticos eran el blanco de la liberación de hierro, lo cual a su vez incrementaba la oxidación de LDL (30). Conocedores de la química de unión del hierro a la transferrina dependiente del pH, es razonable sospechar que el hierro involucra- Hierro do es causante del daño oxidativo aún en individuos con almacenamiento normal de hierro, sin embargo, no hay información disponible sobre cuál es el pH que se encuentra en una lesión aterosclerótica in vivo. Estudios basados en ratas de enfermedad cardiovascular entre sexos a 10 años, revelaron en el estudio de Sullivan (31) como el primero que propuso que la mujer estaba más protegida a estas enfermedades que los hombres por su menor almacenamiento de hierro corporal antes de la menopausia, y es interesante de notar, que los factores conocidos que tienen un efecto sobre la enfermedad cardiovascular también son conocidos que afectan el hierro. Por ejemplo, la relación negativa de enfermedad cardiovascular al uso del ácido acetil-salicílico y la relación positiva al uso de anticonceptivos orales pueden estar indirectamente relacionados a la pérdida de hierro en estas situaciones. Podemos adicionar a esto, que los donadores de sangre experimentaron menos infartos al miocardio (0,7%) que los que no donadores (9,8%) (18), indirectamente sugieren que la reducción del hierro corporal puede ser responsable para la protección. Hace unos años, un estudio mostró que el almacenamiento de hierro elevado con ferrina sérica > 200 µg/ L tenía 2,2 veces más riesgo de un infarto agudo de miocardio que en aquellos hombres que tenían niveles de ferritina sérica < 200 µg/L (32). Contrario a lo arriba expuesto, otros estudios no encuentran tales relaciones, v.gr.: El Estudio de Riesgo Aterosclerótico en Comunidades (ARIC) mostró que después del ajuste para edad, estado de fumador, hipertensión, LDL, colesterol, diabetes, no había asociación entre ferritina sérica y la arterosclerosis asintomática de la carótida (33) . En tres estudios controlados en sujetos entre los 24 a los 65 años, no informaron de ninguna asociación entre el hierro (ferritina sérica y NTBI) y oxidación de LDL, como causa de aterosclerosis (34). Aunque el hierro está involucrado en la generación in vitro de radicales libres, su participación en las reacciones oxidativas in vivo es por ahora cuestionable. Es comúnmente aceptado ques el daño por radicales libres está involucrado en algunas enfermedades, pero no hay una evidencia directa que esté disponible para indicar 137 Hierro que el exceso de hierro es el causante de dichas enfermedades. Hay una fuerte evidencia para explicar el hierro cerebral en enfermedades neurológicas y para el exceso del ingreso dietario de este metal en cáncer colorectal además del involucrado en la enfermedad cardiovascular. Referencias 1. GRANZ, Tomas; Hepcidin, a key regulador of iron metabolism and mediator of anemia of inflammation; Blood; 1 August 2003; Vol 102; No. 3, 783 – 788. 2. ANDREWS, N.C.; Disorders of Iron Metabolism; N. Engl. J. of Med.; 1999, 341: 1986 – 1995. 3. McCORD, Joe M.; Iron, Free Radicals, and Oxidative Injury; The Journal of Nutrition; Nov 2004; 134; 11: 3171S – 3172S. 4. CRICHTON, R.R.; WILMET,S.; LEGSSYER, R.; WARD, R.J.; Molecular and cellular mechanisms of iron homeostasis and toxicity in mammalian cells; J. Inorg. Biochem.; 2002, 91: 9 -18. 5. MUIR, W.A.; HOPFER,U.; KING, M.; Iron transport across brush – border membranes from normal and iron – deficient mouse upper small intestine; J. Biol. Chem.; 1984; 259: 4896 – 4903. 6. WRIGHT, T.L.; BRISSOT, P.; MA, W-L.; WEISIGER,R.A.; Characterization of non – transferrin- bound iron clearance by rat liver; J. Biol. Chem.; 1986; 261: 10909 – 10914. 7. RICHARDSON, D.R.; BAKER, E.; Intermediate steps in cellular iron uptake from transferring. Detection of a cytoplasmic pool of iron, free of transfe- 138 rring; J. Biol. Chem.; 1992; 267: 21384 – 21389. 8. RIEDEL, H-D; REMUS, A.J.; FITSCHER, B.A.; STREMMEL, W.; Characterization and partial purification of a ferrireductase from human duodenal microvillus membranes. Biochem. J.; 1995; 309: 745 – 748. 9. RAJA, K.B.; SIMPSON, R.J.; PETERS, T.J.; Investigation of a role for reduction in ferric iron uptake by mouse duodenum. Biochim. Biophys. Acta; 1992; 1135: 141 – 146. 10. VAN DUIJN, M.; VAN DER ZEE, J.; VANSTEVENINCK, J.; VAN DEN BROEK, P.J.A.; Ascorbate stimulates ferricyanide reduction in HL-60 cells through a mechanism distinct from the NADH-dependent plasma membrane reductase. J. Biol. Chem.; 1998; 273: 13415 – 13420. 11. LESUISSE, E.; CASTERAS-SIMON, M.; LABBE, P.; Evidence for the Saccharomyces cerevisae ferrireductase system being a multicomponent elctron transport chain. J. Biol. Chem.; 1996; 261: 13578 – 13583. 12. WANG, Z.; SCHULTHEIS, P:J.; SHULL, G.E.; Three N-terminal variants of the AE2 Cl / HCO3 exchanger are encoded by mRNAs transcribed from alternative promoters. J. Biol. Chem.; 1996; 271: 7835 – 7843. 13. LEIBOLD, E.A.; GUO, B.; Iron-dependent regulation of ferritin and transferring receptor expression by the ironresponsive element binding protein. Annu. Rev. Nutr. 1992; 12: 345 – 368. 14. BARISANI, D.; BERG, C.L. ; WESSLING-RESNICK, M.; GOLLAN, J.L.; Evidence for a low Km transporter for non-transferrin-bound iron in isolated rat hepatocytes. Am. J. Physiol.; 1995; 269: G570 – G576. 15. REDDY, M.B.; CLARK, L.; Iron, Oxidative Stress, and Disease Risk; Nutri- Hierro tion Reviews, 2004; Vol 62, No. 3; 120 – 124. 16. DERSTINE, J.L.; MURRAY-KOLB, L.E.; YU-POTH, S.; HARGROVE, R.L.; et al; Iron status in association with cardiovascular disease risk in 3 controlled feeding studies. Am. J. Clin. Nutr. 2003; 77: 56 – 62. 17. OTEIZA, P.I.; KLEINMAN, C.G.; DEMASI, M.; BECHARA, E.J.H.; 5-Aminolevulininc acid induces iron release from ferritin. Arch. Biochem. Biophys. 1995; 36: 607 – 611. 18. TUOMAINEN, T.P.; SALONEN, R.; et al; Cohort study of relation between donating blood and risk of myocardial infarction in 2682 men in eastern Finland. BMJ, 1997; 314: 793 -794. 19. MUNTANE, J.; MITJAVILA, M.T.; et al; Dietary lipid and iron status modulate lipid peroxidation in rats with induced adjuvant arthritis. J. Nutr. 1995; 125: 1930 – 1937. 20. STEVENS, R.G.; JONES, D.Y.; et al; Body iron stores and the risk of cancer. N. Engl. J. Med. 1988; 319: 1047 – 1052. 21. GLEI, M.; LATUNDE-DADA, G.O.; KLINDER, A.; et al.; Iron-overload induces oxidative DNA damage in the human colon carcinoma cell line HT29 clone 19A. Mutat. Res. 2002; 519: 151 – 161. 22. NELSON, R.L.; DAVIS, F.G. ; et al ; Body iron stores and risk of colonic neoplasia. J. Nat. Cancer Inst. 1994; 86: 455 – 460. 23. OMARA, F.O.; BLAKLEY, B.R.; Influence of low dietary iron and iron overload on urethane – induced lung tumors in mice. Can. J. Vet. Res. 1993; 57: 209 – 211. 24. FISCHER,J.G.; GLAUERT, H.P.; et al; Moderate iron overload enhances lipid peroxidation in livers of rates, but does not affect NF-êB activation indu- ced by the peroxisome proliferators, Wy-14,643. J. Nutr. 2002; 132: 2525 – 2531. 25. HAGEN, K.; ECKES, K.; MELEFORS, Ö.; HULTCRANTZ, R.; Iron overload decreases the protective effect of tumour necrosis factor-á on rat hepatocytes exposed to oxidative stress. Scand. J. Gastroenterol. 2002; 6: 725 – 731. 26. DABBAGH, A.J.; MANNION, T.; LYNCH, S.M.; FREI, B.; The effect of iron overload on rat plasma and liver oxidant status in vivo. Biochem J. 1994; 300: 799 – 803. 27. STONE, W.L.; PAPAS, A.M.; et al; The influence of dietary iron and tocopherols on oxidative stress and ras-p21 levels in the colon. Cancer Detect. Prev. 2002; 26: 78 – 84. 28. LUND, E.K.; WHARF, S.G.; et al; Oral ferrous sulfate supplements increase the free radical-generating capacity in feces from healthy volunteers. Am. J. Clin. Nutr. 1999; 69: 250 -255. 29. KAUR, D.; YANTIRI, F.; RAJAGOPALAN, S. ; et al ; Genetic or pharmacological iron chelation prevents MPTP-induced neurotoxicity in vivo : a novel therapy for Parkinson´s disease. Neuron. 2003; 37:899 – 909. 30. BALAGOPALAKRISHNA, C.; PAKA, L.; PILLARISETTI, S.; GOLDBERG, I.J.; Lipolysis – induced iron release fron differic transferring: possible role of lipoprotein lipase in LDL oxidation. J. Lipid Res. 1999; 40: 1347 – 1355. 31. SULLIVAN, J.L.; Iron and the sex difference in heart disease risk. Lancet. 1981; 1: 1293 – 1294. 32. SALONEN, J.T.; NYYSSÖNEN, K.; KORPELA, H.; et al; High stored iron levels are associated with excess risk of myocardial infarction in Eastern Finnish men. Circulation. 1992; 86: 803 – 811. 139 Hierro 33. RAURAMMAA, R.; VÄISÄNEN, S.; MERCURI, M. ; et al; Association of risk factors and body iron status to carotid atherosclerosis in middleaged Eastern Finnish men. Eur. Heart J. 1994; 15: 1020 – 1027. 140 34. MOORE, M.; FOLSOM, A.R.; BARNES, R.W.; ECKFELDT, J.H.; No association between serum ferritin and asymptomatic carotid atherosclerosis. Am. J. Epidemiol. 1995; 141: 719 – 723. Este libro hace parte del Programa de Educación Médica Continuada del AWGLA www.awgla.com