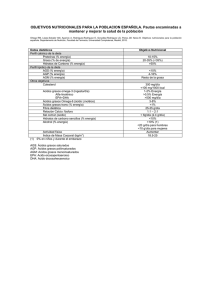
El origen de los lípidos en la leche. - UAM-I
Anuncio

El origen de los l¶³pidos en la leche
Cecilia Aranda Batalla y Martha Del Prado Mariquez
Unidad de investigaci¶on en nutrici¶on. Hospital de pediatr¶³a
Centro medico nacional siglo XXI, IMSS
Cambios en el metabolismo materno durante la lactancia.
Para el establecimiento de la lactancia el cuerpo materno se prepara a partir del primer d¶³a de embarazo, per¶³odo en el cual la madre acumula reservas
energ¶eticas, principalmente grasa en el tejido adiposo, que son movilizadas durante la lactancia para contribuir a la producci¶
on de la leche. Una vez
que se establece la lactancia, el cuerpo materno sufre una serie de cambios en el metabolismo de varios tejidos con el ¯n de proporcionar los sustratos
necesarios a la gl¶
andula mamaria para que se lleve a cabo la s¶³ntesis de leche, entre estos cambios se
encuentran:
Recibido: 26 de mayo de 2003.
Aceptado: 3 de junio de 2003.
Resumen.
Durante la lactancia la leche cubre las demandas de
los reci¶en nacidos favoreciendo un crecimiento y desarrollo adecuado. En la leche, la principal fuente de energ¶³a son los triacilgliceroles contenidos en
los gl¶
obulos de grasa, constituidos por ¶acidos grasos de cadena corta (< 8 C), de cadena media (8-14
C) y de cadena larga (¸ 16 C).
El patr¶
on de los ¶acidos grasos presentes en la leche
var¶³a por la in°uencia de diferentes factores: especie,
el tipo de dieta, epoca del a~
no, la duraci¶on de la gestaci¶
on, la etapa de lactancia y la hora del d¶³a. La dieta juega un papel importante en la s¶³ntesis y la captaci¶
on de l¶³pidos en la gl¶andula mamaria, en roedores el consumo de la dieta habitual (alta en carbohidratos) aumenta la lipog¶enesis en gl¶andula mamaria mientras que una dieta con alto contenido de l¶³pidos disminuye la lipog¶enesis al tiempo que aumenta la captaci¶on de l¶³pidos circulantes.
1. La hipertro¯a del h¶³gado, que est¶
a acompa~
nada
por cambios en la actividad de las enzimas que
intervienen en el metabolismo de carbohidratos
y l¶³pidos. El h¶³gado convierte una gran proporci¶
on de los ¶
acidos grasos de cadena larga provenientes de la dieta en l¶³pidos esteri¯cados y disminuye la proporci¶
on de estos que es convertida en cuerpos cet¶
onicos, con el ¯n de proporcionar l¶³pidos a la gl¶
andula mamaria en el inmediato estado post{absortivo.
Palabras clave: gl¶andula mamaria, lactancia, lipog¶enesis, lipoprote¶³na lipasa, l¶³pidos de la
dieta.
2. La hipertro¯a del intestino delgado que acompa~
nada por la hiperplasia de la mucosa intestinal aumenta la capacidad absortiva del
intestino.
Introducci¶
on.
La importancia de la lactancia radica en que los reci¶en nacidos se nutren con la leche de su madre, hasta que son capaces de ingerir alimentos s¶olidos, reduciendo as¶³ el riesgo a enfermedades infecciosas y alergias. La leche posee una elevada densidad energ¶etica
y un equilibrio de nutrimentos que satisfacen las necesidades de los reci¶en nacidos y les permite un crecimiento adecuado; adem¶as es un importante veh¶³culo para transferir inmunoglobulinas, enzimas, hormonas, agentes anti{in°amatorios y algunos factores no espec¶³¯cos (lisozima, folato, lactoperoxidasa)
que inter¯eren con el desarrollo bacteriano o potencian la actividad inmune de anticuerpos (Villalpando, 1993, Rodr¶³guez- Palmero, 1999).
3. El aumento en la movilizaci¶
on de los l¶³pidos acumulados durante la gestaci¶
on y la disminuci¶on
en la s¶³ntesis y captaci¶
on de l¶³pidos en el tejido
adiposo, tienen como resultado que menos glucosa y triacilgliceroles sean captados por el tejido adiposo y m¶
as ¶
acidos grasos no esteri¯cados y glicerol sean liberados a la circulaci¶on para ser captados por la gl¶
andula mamaria.
Estos cambios metab¶
olicos se acompa~
nan por el aumento en el consumo de alimento, el cual se encuen65
66
tra en funci¶
on del aumento en el requerimiento de
energ¶³a asociado con la s¶³ntesis de leche. En estudios con animales, se ha observado que estos pueden llegan a triplicar su consumo cal¶orico durante la lactancia (Allen, 1994; Neville, 2001; Willianson, 1980, 1986).
Lactancia
La lactancia es la fase ¯nal del ciclo reproductivo de
los mam¶³feros y se caracteriza por un aumento en
la demanda de sustratos por la gl¶andula mamaria
para llevar a cabo la s¶³ntesis de los constituyentes de
la leche (lactosa, l¶³pidos y prote¶³nas) en las c¶elulas
epiteliales de los alv¶eolos.
La lactog¶enesis es una serie de procesos dentro de las
c¶elulas alveolares de la gl¶andula mamaria mediante
los cuales se sintetizan los constituyentes de la leche.
En las primeras horas posteriores al parto, las c¶elulas alveolares llegan a desarrollar una citolog¶³a semejante a las c¶elulas ex¶ocrinas, en la base de la c¶elula se puede observar una gran cantidad de ribosomas libres, que son remplazados por membranas del
ret¶³culo endopl¶
asmico rugoso, las cuales son evidentes 48 horas despu¶es. En este tiempo el epitelio alveolar adquiere una forma cuboidal e incrementa su actividad secretora, la cual puede iniciarse con la repentina disminuci¶on o cese de la liberaci¶on de progesterona y con la evacuaci¶on de los productos que
forman el calostro (Mather, 1998).
El alv¶eolo se distiende en presencia de la leche y la
contracci¶
on de las c¶elulas mioepiteliales ayudan a la
propulsi¶
on de la leche desde el lumen alveolar hasta
los ductos y senos, donde la leche se encuentra bajo
presi¶
on y disponible para la succi¶on (Neville, 1999;
Jensen, 1995). El ritmo de secreci¶on de la leche es
alto cuando la presi¶on en los conductos y alv¶eolos
alcanza un nivel muy bajo, a medida que la presi¶
on
aumenta el ritmo de producci¶on disminuye.
ContactoS 48, 65{70 (2003)
tancia, en la primera parte de la lactancia presenta su m¶
axima velocidad y disminuye gradualmente a lo largo de este per¶³odo.
Durante la lactancia la gl¶
andula mamaria capta de
la circulaci¶
on: glucosa, amino¶
acidos y ¶
acidos grasos
no esteri¯cados, para la s¶³ntesis de los componentes
de la leche (Shenna, 2000; Williamson, 1986).
Se han de¯nido cinco v¶³as espec¶³¯cas (Fig. 1), que
act¶
uan paralelamente, para la s¶³ntesis y secreci¶
on de
la leche en la gl¶
andula mamaria. Estas v¶³as transforman a los precursores derivados de la sangre en
los constituyentes de la leche (Neville, 1983, 1997,
Shenna, 2000).
Mecanismo I (Exocitosis): Las micelas de
case¶³na, las prote¶³nas, el calcio, el fosfato y el
citrato son empaquetados en ves¶³culas secretoras. Estas ves¶³culas se mueven hacia el ¶
apice de
la c¶elula, se fusionan con la membrana celular
y liberan su contenido hacia la luz alveolar por
exocitosis.
Mecanismo II (S¶³ntesis y secreci¶
on de l¶³pidos): Los triacilgliceroles (TAG) sintetizados en
el citosol y en el ret¶³culo endopl¶
asmico liso, coalescen en vacuolas y se dirigen al ¶
apice de la
c¶elula, ah¶³ son envueltos por una porci¶
on de la
membrana apical y son separados de la c¶elula como gl¶
obulos de grasa.
Mecanismo III (Secreci¶
on de iones y agua): El sodio, el potasio y el
agua atraviesan la membrana libremente. El agua lo hace de acuerdo a un gradiente osm¶
otico impuesto por la lactosa. Los electrolitos se mueven bajo su gradiente
electroqu¶³mico.
El mantenimiento de la secreci¶on l¶actea depende del
est¶³mulo mec¶
anico de la succi¶on recibido en las terminaciones nerviosas del pez¶on, el cual produce impulsos nerviosos que llegan a la m¶edula espinal, siguen la v¶³a espinotal¶amica hasta el mesenc¶efalo donde es posible que se bifurquen siguiendo dos rutas separadas hasta las ¶areas del hipot¶alamo, determinando as¶³ la liberaci¶on de prolactina, oxitocina y de la
hormona de crecimiento.
Mecanismo lV (Secreci¶
on de inmunoglobulinas): La inmunoglobulina A (IgA) y otras
prote¶³nas se unen a un receptor especi¯co en la
membrana basolateral de la c¶elula. El receptor
y la inmunoglobulina son internados en ves¶³culas endoc¶³ticas pare ser transportadas hacia la
membrana apical y ser excretadas al lumen
alveolar.
S¶³ntesis de leche.
La s¶³ntesis de leche en la gl¶andula mamaria presenta cambios marcados durante el per¶³odo de lac-
Mecanismo V (V¶³a paracelular): se designa como mecanismo paracelular porque involucra el paso entre las c¶elulas epiteliales. A
El origen de los l¶³pidos en la leche. Cecilia Aranda y Martha Del Prado
trav¶es de esta ruta se transportan c¶elulas, prote¶³nas plasm¶aticas y sodio. Durante el embarazo las uniones paracelulares herm¶eticas se encuentran separadas y se cierran progresivamente hacia el ¯nal del embarazo y el principio de la
lactancia.
La s¶³ntesis de leche esta determinada por la capacidad y el n¶
umero de c¶elulas secretoras, el n¶
umero de c¶elulas esta ligado a factores gen¶eticos, mientras que la capacidad de cada una est¶a in°uenciada por factores externos, tales como la alimentaci¶
on
y las condiciones ambientales.
L¶³pidos
Los l¶³pidos son la principal fuente de energ¶³a para
el reci¶en nacido y son esenciales para su crecimiento debido a que proporcionan los ¶acidos grasos necesarios para el desarrollo del cerebro, forman parte integral de las membranas celulares y son el veh¶³culo para transportar vitaminas y hormonas liposolubles (Hamosh, 1984).
Los l¶³pidos son el componente m¶as variable en la leche, su concentraci¶on y composici¶on var¶³a entre individuos, dependiendo de factores como la alimentaci¶
on, la duraci¶on de la gestaci¶on, la etapa de lactancia y la hora del d¶³a. Tambi¶en hay variaciones importantes entre especies (Allen, 1991; Hamosh 1984;
Neville, 1983).
Los l¶³pidos de la leche se encuentran contenidos en
los gl¶
obulos de grasa formados por membranas tridimensionales compuestas por fosfol¶³pidos, colesterol y prote¶³nas. Entre el 97 y el 99 % de los l¶³pidos contenidos en estos gl¶obulos son triacilgliceroles, los cuales proporcionan entre el 40 al 50 % de la
energ¶³a total de la leche (Hamosh, 1984; Neville and
Picciano, 1997).
Los ¶
acidos grasos de los triacilgliceroles de la leche provienen de la circulaci¶on o de la s¶³ntesis en
la gl¶
andula mamaria.
Los ¶
acidos grasos de cadena corta (con menos de 8
carbonos), especialmente but¶³rico y hexanoico, son
tomados directamente de la circulaci¶on o sintetizados a partir de -hidroxibutirato. Los ¶acidos grasos de
cadena media (C8-14) son sintetizados en la gl¶
andula mamaria, a partir de glucosa, acetato, lactato y algunos amino¶
acidos. Los ¶acidos grasos de cadena larga (¸ C16), saturados e insaturados, provienen de la
dieta o de la movilizaci¶on de los ¶acidos grasos del tejido adiposo (Williamson, 1980, 1986; Hachey, 1989;
Neville and Picciano, 1997).
67
Captaci¶
on de los l¶³pidos circulantes.
Durante la lactancia, los ¶
acidos grasos de cadena larga, provenientes de la dieta y de la movilizaci¶on
de los ¶
acidos grasos del tejido adiposo, son transportados hacia la gl¶
andula mamaria en los quilomicrones y en las lipoprote¶³nas de muy baja densidad (VLDL) (Williamson, 1994). La lipoprote¶³na lipasa (LPL) presente en el endotelio capilar mamario, convierte a los triacilgliceroles de las lipoprote¶³nas circulantes en ¶
acidos grasos libres, diacilgliceroles y en 2-monoacilgliceroles, que son transportados hacia el interior del lactocito y convertidos nuevamente en triacilgliceroles.
La lipoprote¶³na lipasa es la enzima que se ha descrito como clave para permitir la captaci¶
on de l¶³pidos por los tejidos. Es una prote¶³na, de 55-kDa, sintetizada por c¶elulas del par¶enquima en varios tejidos y es transportada a su sitio de acci¶
on en el endotelio vascular donde cataliza la hidr¶
olisis de los triacilgliceroles de los quilomicrones y de las VLDL para que los productos de la hidr¶
olisis puedan ser captados por los tejidos (Martin- Hidalgo, 1994, Neville and Picciano, 1997; Tso, 1995). La elevada actividad de la LPL durante la lactancia en la gl¶andula mamaria y su baja actividad en el tejido adiposo, permiten dirigir a los l¶³pidos circulantes preferentemente hacia la gl¶
andula mamaria para la formaci¶
on de leche (Martin- Hidalgo, 1994).
S¶³ntesis de ¶
acidos grasos.
Los ¶
acidos grasos de cadena media presentes en la
leche provienen de la s¶³ntesis en la gl¶
andula mamaria
utilizando a la glucosa como el principal sustrato
lipog¶enico (Fig. 2) (Neville, 1983; Hachey, 1989).
La glucosa dentro de la gl¶
andula mamaria:
1. Es convertida en acetil-coenzima A (CoA) proporcionando las unidades de carbono para la
s¶³ntesis de los ¶
acidos grasos.
2. Es convertida a ribulosa-5-fosfato por la ruta del
acido 6-P-gluc¶
¶
onico, contribuyendo a la formaci¶
on de NADPH necesario para la s¶³ntesis de ¶acidos grasos.
3. Es transformada a gliceraldehido-3-fosfato en la
gluc¶
olisis, el cual puede ser convertido a glicerol3-fosfato y utilizado para la s¶³ntesis de triacilgliceroles.
En el citosol de c¶elulas mamarias, la glucosa es fosforilada y convertida a piruvato en la gluc¶
olisis. Dentro
de la mitocondria el piruvato es oxidado por la enzima piruvato deshidrogenasa a acetil-CoA. La acetilCoA se une con oxalacetato en el primer paso del ciclo de Krebs para formar citrato, el cual es transpor-
68
ContactoS 48, 65{70 (2003)
Figura 1. V¶³as de s¶³ntesis y secreci¶
on de los componentes de la leche por la c¶elula epitelial mamaria; RER: ret¶³culo
endoplasmico rugoso. (En Neville MC, Allen JC. and Watters C. 1983).
tado al citosol y convertido a acetil-CoA por la enzima ATP citrato liasa. La conversi¶on de acetil-CoA
a malonil-CoA es el paso limitante en la bios¶³ntesis de ¶
acidos grasos y es catalizado por la enzima
acetil-CoA carboxilasa en presencia de ATP. Despu¶es de la formaci¶on de malonil-CoA, la sintetasa
de ¶
acidos grasos cataliza una secuencia de siete reacciones, cada reacci¶on adiciona dos carbonos derivados del malonil-CoA para la elongaci¶on de la cadena. Cada ciclo requiere dos mol¶eculas de NADPH
(Neville, 1983; Neville and Picciano, 1997; Williamson, 1994).
En el h¶³gado y en el tejido adiposo, cuando la cadena del ¶
acido graso reci¶en sintetizado, alcanza un tama~
no de 16 carbonos o m¶as, la s¶³ntesis es terminada por la tioesterasa I. Esta parte integral del sistema de la sintetasa de los ¶acidos grasos termina
la s¶³ntesis del ¶
acido graso retirando la mol¶ecula ¯nal de su sitio de uni¶on con la enzima. En el citosol de las c¶elulas epiteliales mamarias en animales no rumiantes, la s¶³ntesis de ¶acidos grasos es detenida por la tioesterasa II, despu¶es de que la cadena de ¶
acido graso alcanza una longitud de 8 a 14 carbonos (Neville, 1997).
La regulaci¶
on a corto plazo de la lipog¶enesis en la
gl¶
andula mamaria es compleja y varia en respuesta
al estado hormonal y nutricional del animal, de manera ¯siol¶
ogica. Entre los m¶
ultiples mecanismos de
control podr¶³a incluirse la regulaci¶
on de la transcripci¶
on del ADN para formar ARN mensajero y la subsecuente traducci¶
on a enzimas espec¶³¯cas de la s¶³ntesis de l¶³pidos.
La acetil-CoA carboxilasa y la piruvato deshidrogenasa (PDH) son enzimas clave y sitios de regulaci¶
on de la lipog¶enesis. La acetil-CoA carboxilasa
existe como un mon¶
omero que presenta muy baja
actividad, y como un pol¶³mero con una elevada actividad. El citrato aumenta marcadamente la polimerazaci¶
on de la enzima causando un aumento en
su actividad. Por otra parte los acil CoA de cadena larga (principalmente palmitoilCoA), productos
de la s¶³ntesis de ¶
acidos grasos, favorecen la disociaci¶
on de la enzima hacia su forma menos activa. La
acetil-CoA carboxilasa es adem¶
as regulada por ciclos de fosforilaci¶
on-defosforilaci¶
on. La fosforilaci¶on
de la enzima causa su inactivaci¶
on.
La piruvato deshidrogenasa tambi¶en es controlada
a trav¶es de su estado de fosforilaci¶
on, se encuentra
en forma activa (defosforilada) e inactiva (fosforilada) (Williamson, 1980, 1994). En la regulaci¶
on de la
piruvato deshidrogenasa cinasa se involucran cuatro
factores: la concentraci¶
on de piruvato y las relacio-
El origen de los l¶³pidos en la leche. Cecilia Aranda y Martha Del Prado
69
Figura 2. S¶³ntesis de l¶³pidos en la gl¶
andula mamaria (En Neville MC. and Picciano MF. 1997).
nes ATP/ADP, acetil-CoA/CoA y NADH/NAD+ .
Elevados valores de estas relaciones activan la cinasa e inhiben a la piruvato deshidrogenasa.
In°uencia de la dieta sobre la composici¶
on de
los l¶³pidos de la leche.
La dieta controla la s¶³ntesis de los ¶acidos grasos y de
los triglic¶eridos de la leche, modulando esta o la actividad de las enzimas que intervienen en ella, aumentando la disponibilidad de cofactores (¶ac. pantot¶enico, biotina) o in°uenciando la concentraci¶
on de hormonas que inducen la s¶³ntesis de algunas de las enzimas lipog¶enicas.
Existen variaciones en la proporci¶on de los ¶
acidos
grasos de la leche, ocasionadas por cambios en el contenido y composici¶on de los l¶³pidos de la dieta (Rasmussen, 1992). Estudios en humanos y animales han
mostrado que al proporcionar una dieta con alto contenido de carbohidratos y un bajo contenido de l¶³pidos, la proporci¶on de ¶acidos grasos de cadena media en la leche aumenta (Hachey, 1989; Williamson,
1986). Cuando se proporciona a la madre, una dieta con elevado contenido de l¶³pidos o se le administra una carga intrag¶astrica de l¶³pidos, que resulta
en la constante liberaci¶on de quilomicrones a la circulaci¶
on, se tiene como resultado una disminuci¶
on
en la lipog¶enesis en la gl¶
andula mamaria que repercute en la disminuci¶
on en la proporci¶
on de los ¶acidos grasos de cadena media en la leche y en el aumento en la proporci¶
on de ¶
acidos grasos de cadena larga provenientes de la dieta (Del Prado, 1999; Hachey, 1989; Souza, 1993; Williamson, 1980, 1986).
Dado que la proporci¶
on de ¶
acidos grasos de cadena media y de cadena larga presentes en la leche
se ve in°uenciada por la dieta materna, es importante considerar el uso metab¶
olico de estos ¶acidos
grasos por el lactante. Se sabe que los ¶
acidos grasos de cadena media son absorbidos m¶
as f¶acilmente por el intestino, son liberados m¶
as r¶
apidamente a la circulaci¶
on y pueden ser totalmente oxidados, mientras que con los ¶
acidos grasos de cadena
larga sucede lo contrario. Tomando en cuenta estas
caracter¶³sticas se ha sugerido que los ¶
acidos grasos
de cadena media se utilizan como una fuente r¶apida de energ¶³a mientras que los ¶
acidos grasos de cadena larga se almacenan preferentemente en los triacilgliceroles del tejido adiposo; mas hay que considerar que entre ellos se encuentran los acidos grasos
esenciales.
Por lo cual es necesario preguntarnos si el aumento en la proporci¶
on de ¶
acidos grasos de cadena larga
70
en la leche, puede repercutir en el crecimiento y desarrollo del lactante o si representa un factor de riesgo para la salud en su vida adulta.
Bibliograf¶³a.
1. Allen, J. C., Keller, R. P., Archer, P. and Neville, M. C. Studies in human lactation: milk composition and daily secretion rates of macronutrients in the ¯rst year of lactation. Am. J. Clin.
Nutr. 54: 69-80.1991.
2. Allen, L., King, J. and LÄonnerdal. Nutrient regulation during pregnancy, lactation, and infant
growth. Volume 352. Plenum Press, New York.
1994. p. 45-70.
3. Del Prado, M., Villalpando, S., Gordillo, J y
Hern¶
andez-Montes H. A high dietary lipid intake during pregnancy and lactation enhaces
mammary gland lipid uptake and lipoprotein lipase activity in rats. J. Nutr. 129: 1574- 1578.
1999.
4. Hachey D. L., Silber G. H., Wong, W. W. and
Garza, C. Human lactation II: endogenous fatty
acid synthesis by the mammary gland. Pediatric. Res. 25: 63-68.1989.
ContactoS 48, 65{70 (2003)
12. Neville, M. C., Allen, J. C. and Watters C. The
mechanism of milk secretion. Lactation: physio_
logy, nutrition and breast-feeding. Neville, MC.
and Neifert, M. R. Plenum Press. New York. pp:
49-102. 1983.
13. Neville, M. C. and Picciano, M. F. Regulation
of milk lipid secretion and composition. Annu.
Rev. Nutr. 17: 159-184. 1997.
14. Rasmussen, K. M. The in°uence of maternal nutrition on lactation. Annu. Rev. Nutr. 12: 103117. 1992.
15. Rodr¶³guez- Palmero, M., Koletzko, B., Kunz, C.
and Jensen, R. Nutritional and biochemical properties of human milk: II. Clin Perinatol. 26 2 :
335-359. 1999.
16. Shenna, D. B. and Peaker, M. Transport of
milk constituent by the mammary gland. Physiol Rev. 80: 925-951. 2000.
17. Souza, P. F. and Williamson, D. H. E®ects of
feeding medium-chain triacylglycerols on maternal lipid metabolism and pup growth in lactanting rats. Br. J. Nutr. 69: 779-787. 1993.
5. Hamosh , M., Bitman, J., Wood, L., Hamosh,
P. and Mehta, N. R. Lipids in milk and the ¯rts
steps in their digestion. Pediatrics. 758 [suppl]:
146-150. 1984.
18. Tso, P., Karlstad, M. D., Bistrian, B. R. and
De Michele, S. J. Intestinal digestion, absorption, and transport of structured triglycerides
and cholesterol in rats. Am. J. Physiol. 268 Gastrointest. Liver Physiol 31 : G568-G577. 1995.
6. Jensen, R. G. Handbook of milk composition.
Academin Press. New York, University of Connecticut, 1995.
19. Villalpando, S. y De Santiago, S. Bases biol¶ogicas de la lactancia materna. Bol. Med. Hosp. Infan. Mex. 50 12 : 889-897. 1993.
7. Mather, I. H. and Keenan, T. W. The cell
biology of milk secretion. Historical notes. J.
Mamm. Gland. Biol. Neop. 3 3 : 227-232. 1998.
8. Martin-Hidalgo A., Holm, C., Belfrage, P., Schotz, M. C. and Herrera, E. Lipoprotein lipase and hormone-sensitive lipase activity and mRNA in rat adipose tissue during pregnancy. Am. J. Physiol. 266: E930-E935. 1994.
9. Neville, M. C. Mammary gland biology and lactation: a short course. Given At The Meeting
Of The International Society For Research In
Human Milk And Lactation. Plymouht, New
Hampshire. October 15. pp: 1-30. 1997.
20. Williamson, D. H. Integration of metabolism in
tissues of the lactanting rat. Febs Letter. 117
supplement : K93- K105. 1980.
21. Williamson, D. H. Regulation Of Metabolism
During Lactation In The Rat. Reprod. Nutr. Develop. 26[2 B]: 597-603. 1986.
22. Williamson, D. H. and Lund, P. Cellular mechanisms for the regulation of adipose tissue
lipid metabolism in pregnancy and lactation.
Nutrient regulation during pregnancy, lactation,
and infant growth. Allen, L., King, J. and
LÄ
onnerdal (Volume 352). Plenum Press, New
York. pp: 45-70. 1994.
10. Neville, M. C. Physiology of lactation. Clin. Perinatol. 26 2 : 251- 279. 1999.
11. Neville, M. C. Physiology and endocrine changes underlying human lactogenesis II. J. Nutr.
13 11 : 3005S- 3008S. 2001.
cs