\ Serie: Salud i 1 z,çz ADRIÁN CANO PROUS MARÍA CONTRERAS CHICOTE, NEUROPSICOLOGÍA DE LA SEXUALIDAD CONCEPTOS PARA E,NTENDER MEJOR LA SEXUALIDAD Lto Ëc TEOLOGIA Valenciå I 2 t'tllvn zgt+ FfI$TffiA MA CfINS;r EDICIONES UNIVERSIDAD DE NAVARRA, S.A PAMPLONA Primera edición: Ab¡il 20 14 Índice @ 2014. Ad¡ián Cano Prous y Marla Contre¡as Chicote Ediciones Universidad de Navarra, S.A. (EUNSA) Plaza de los Sauces, 1 y 2. 31010 Barañáin (Navarra) - España Têléfono: +34 948 25 68 50 - Fax: +34 948 25 68 54 e-mail: [email protected] ISBN: 978-84-3 r3-298r-5 Depósito legal: N A 429 -20 1 4 Queda prohibida, salvo excepción prevista en la le¡ cualquier forma de reproducción, distibución, comunicación pública y transformación, total o parcial, de ata obra sin contar con auto¡iación escrita de los titulares del Copyight, La infracción de los derechos mencionados puede ser constitutiva de delito contra la propiedad intelectual (Ardculos 270 y ss. del Código Penal). I. INrnoouccróN 9 II. DBs¡nnoLLo oNToLócICo DE lA SEXUALIDAD r3 r4 2.1. Nivel genético ..... 2.2. Nivel gonadal 2.2.1. Desarrollo testicular .............. 2.2.2. Diferenciación del ovario 2.3. Nivel somático 2.3.1. Acd6n hormonal ......... 2.3.2. Diferenciación de los genitales internos ...... 2.3.3. Diferenciación de los genitales externos ...... 2.4. Sexo fenotlpico 2.4.1. Sistema reproductor y funcionamiento gona- 17 20 23 24 25 25 )7 28 29 2.4.2. Eje hipotálamo hipofisario gonadal 2.4.3. Caracteres sexuales secundarios .................. Tiatamiento: IToM. 31014 Pamplona Imprime: GR {FIcAs ALZATE, S.L. Pol. Comarca 2. Esparza de Galar (Navarra) Printed in Spain - Impreso en España III. Drr¡neNctnclóN SEXUAL DEL cEREBRo 37 4t 47 3.1. Interacción de las hormonas sexuales en el sistema nervioso cenüal 49 3.1.2. Efecto dual 50 3.1.2. Hormonas sexuales 52 8 Neuropsicologla de la sexualidad I Introducción 3.1.3. Actuación de los esteroides sexuales sobre el sistema neryioso central 3.2. Estructuras cerebrales 3.2.1. Principales núcleos dimórficos en el hipotálamo 3.2.2. Ouos 54 56 62 75 fV NpunorRANsMrsoREs 79 V HrpeRsnxuarm¡o 93 VI. Epfroco 105 MI. Rs¡ensNcrAs BrBLrocRÁrrcAS r13 A 1o largo de la historia ha sido dificil definir el concepto de usexualidado. Se entiende por sexualidad al conjunto de caracterlsticas de cada sexo o a la condición sexual Propia de la espe-' cie humana y de todos los animales y plantas que se reproducen mediante la unión de células específicas y diversas: masculina y femenina. Otras se veces se ha entendido por sexualidad, y ya desde el punto de vista de la especie humana, como el impulso hacia los individuos del sexo opuesto. Impulso que en los animales reviste las caracterlsticas instintivas, pero que en el hombre, Por su racionalidad y por el papel que ejerce la voluntad en la conducta, adquiere otras modalidades. Segun los datos científicos, el ser humano está de tal manera marcado por la sexualidad, que forma parte de los factores que caracteriztnla vida de los hombres. Es el resultado de la interacción de factores biológicos, psicológicos, culturales, éticos, espirituales y esrructuras cerebrales, aclarando que la sexualidad va m¿ís allá de lo puramente biológico y que está bajo la influencia de diversos elementos. 10 Neuropsicología de la sexualidad Este trabajo conriene Lrna revisión bibriográfica acerca de la neuropsicología de la sexualidad humana. par" comenza¡ se ha considerado importante describir el desarrollo ontológico de la sexualidad desde un punto de vista cronológico a ni,rãl genético, gonadal, somárico y fenotípico. Desde ef primer día ãer de- sarrollo embriológico de un individuo hasra su madurez, apa- recen y se desarrollan los caracteres sexuales. Durante los nueve primeros meses del desarrollo embrionario solo se expresa el código genético por una serie de inducciones sucesivas, moduladas y orientadas por las condiciones fisico-químicas del medio interno; los caracteres expresados de esta -"r.r" son los mismos para los individuos del mismo sexo. Después der nacimiento, .l organismo está sometido a la influencia del medio externo al cual debe adaptarse. Desde el punto de vista de la Neuropsicología, la dimen_ sión humana de la sexualidad se basa.n lo, descu-brimienros estructurales y funcionales que intervienen en la conducta sexuar del ser humano. Los investigadores están cada vez más seguros de que el órgano sexual más imporranre es el cerebro y.r"porible explicar la sexualidad humana desde las diferenci".'.rtr,r.trrrales del mismo. Estos dimorfismos enceFálicos dependen de la influencia en la fase remprana de las hormonas que actúan sobre los núcleos cerebrales que median diferentes aspecros de la conducta sexual. H?y en día, gracias a las técnicas de neuroimagen se sabe que . el cerebro es sexualmente dimórfico. Estos conocimientos sobre las diferencias cerebrales de los sexos y los circuitos cerebrales relacionados con la identidad y orientación sexual, van a posibilitar una interpretación de algunas conductas sexuales hu-"n"r. Por ello, se analiza cada una de las estrucruras cerebrales posiblemente involucradas en la sexualidad humana y lo, r.urotransmisores que regulan esta conducta, ya que las hormonas y los Introducción lt neurotransmisores son esenciales en la atracción sexual, la elec_ ción de par€1a, el apego y la conducta sexual. Sin duda, la sexualidad es uno de los grandes rernas de las últimas décadas. Las cifras nacionales revelan una importante necesidad de que este tema sea abordado de manera integral, científico y humano. Hoy en día comienzan a observarse los problemas derivados de una sexualidad poco elaborada y socialmente inadecuada. Con relación a esra cuestión, en las páginas finales se ofrece una explicación de las posibles bases neurobiológicas del trastorno hipersexual (THS). Esre rrastorno se caracteriza por el aumenro en la frecuencia, inrensidad de fanrasías, excitación, impulsos y conductas sexuales no parafilicas, asociadas con impulsividad que provocan malestar significativo, dererioro social y ocupacional. Presenta aJta prevalencia en varones y alta comorbilidad en el Eje I. Aunque esre trasrorno se puede presentar debido a algunos problemas médicos o al consumo de algún medicamento, en la mayofia de los casos la etiología es desconocida. t- II Desarrollo ontológico de la sexualidad El establecimiento del sexo es un proceso secuencial que se puede dividir en tres etapas. La primera etapa se establece a partir de la existencia de los cromosomas sexuales que caracterizan el fenotipo, suceso que ocurre en el momento de la fecundación. El sexo genético determina el sexo gonadal, siendo esta la segunda etapa, en la cual ocurren procesos moleculares que llevan hacia la diferenciación del ovario o testículo a partir de la gónada bipotencial morfológicamente indiferenciada. IJnavez iniciada la morfogénesis de las gónadas, se produce la tercera etapa que incluye la diferenciación sexual somática. La presencia o ausencia de secreciones condiciona la diferenciación de los genitales internos (tracto reproductor): se pueden diferenciar los conductos de tVolffo Müller en conductos masculino o femenino respectivamente, y la diferenciación del seno urogenital que formará los genitales externos de uno u otro sexo. Este último proceso se refuerza en el momento de la pubertad con la aparición de los caracteres sexuales secundarios conformando los fenotipos propios de los varones y mujeres. Por otro lado, las secreciones gonadales durante el proceso influyen en el sistema nervioso en desarrollo con el estableci- t4 Neuropsicología de la sexualidad miento del eje hipotdamo-hopófisis-gónada; diferenciando sexualmente el cerebro, exhibiendo unos mecanismos neuroendocrinos y un repertorio de conductas congruentes con el sexo somático (Fernández, 1996). 2.1. Nvel cENÉTrco En los humanos, la diferenciación sexual primaria es cromosómica y usualmente no está influida por el medio ambiente (Colina y Moncada, 2006). El conjunto de la información genética que define el sexo de un individuo, se conoce como determinación genética del sexo. La capacidad potencial de cada organismo y de cada célula germinal a desarrollarse en uno de los dos sentidos, femenino o masculino, fue expresada por Hartmann (1939) como la uley potencial bisexual de ambos sexos). Esto significa que sin tener en cuenta el sexo genético, todo cigoto puede desarrollarse de una manera o de otra ya que posee la información necesaria. Mtiller (1932) estableció la base para la comprensión del problema y propone la existencia de dos tipos de genes que actúan sobre el fenotipo sexual: genes que deciden el sexo y genes básicos para el sexo. Es en el momento de la fecundación (Czyba, 1978), cuando las moléculas de ADN que contienen la información genética, caracteÍística de la especie humana, se manifiestan en el proceso de las divisiones celulares bajo la forma de 46 cromosomas: 44 autosomas y 2 cromosomas sexuales, designados por las letras )O( en el sexo femenino y XY en el sexo masculino (Czyba, 1978). Por la fusión de los dos gametos se va a formar el cigoto con una carga cromosómica diploide, produciéndose la combinación de los cromosomas sexuales de origen materno y pater- Desarrollo ontológico de la sexualidad 15 no. Esta combinación esrá condicionada por la naturaleza del gonosoma presente en el núcleo del espermatozoide fecundante (Colina E¿ Moncada,2006l' Czyba, 1978). Una vez que se ha producido la fecundación, los gametos masculino y femenino van a reilizar el proceso de meiosis para reducir lacargacromosómica, siendo la misma en todas las células y conteniendo la mitad de la carga cromosómicahumana:22 autosomas y un cromosoma sexual. De esta manera, se determina en la especie humana el sexo genético. La presencia del cromosoma Y implica una determinación testicular con independencia del número de cromosomas X y es considerado genéticamente varón (Fernández, 1996), siendo la formula XY caracterlstica del sexo masculino, que es heterogamético (Colina E¿ Moncada, 2006). En ausencia del cromosoma Y, la gónada primitiva se desarrolla como ovario, y con independencia del número de cromosomas X, se considera genéticamente mujer (Fernández,1996). La formula caracterlstica del sexo femenino es )O(, que es homogamética. El cromosoma X es portador de genes responsables de muchos rasgos, en cambio, el cromosoma Y es más pequeño, tiene una forma diferente y contiene los genes que inician la determinación sexual masculina. Los cromosomas sexuales X e Y difieren en tamaño y en información genética, en cambio, son parcialmente homólogos para poder sufrir entrecruzamientos. La porción de los cromosomas que es homóloga, es pequeña y en ella se encuentran los genes parcialmente ligados al sexo. Cromosoma Y determinación testicular El cromosoma Y actúa como inductor de la masculinidad y determina a la gónada indiferenciada transformándola en testículo (Fernández, 1996). t t6 Neuropsicología de la sexualidad Desarrollo ontológico de la sexualidad 17 En este cromosoma se asienta una enridad genédca llamada nfactor determinante del testículo> o TDF (te*is determiningfac- 2.2. Nrvpr coNADAr tor) (Fená',ndez,1996). Esta región contiene el gen de la determinación sexual (SRÐ que codifica la proteína SRY, siendo esta la inductora del programa que desarrollará aI testículo a parrir de una gónada bipotencial indiferenciada (Sinclair er aL, 1991, Gubbay et al., 1990). Es un péptido de 273 aminoácidos que se liga aI ADN y que se cancteriza por ser un grupo de proteínas de alta movilidad. El SRY es activado por el gen'WTl, que contribuye a la diferenciación sexual (Hernández et â1., 2004), actúa como regulador y e'1erce su función a través de otros genes. El SRY solamente se produce durante un corto período de tiempo en las células somáticas de los cordones gonadales. Durante esta fase se diferencian las células de Sertoli, necesarias para el desarrollo gonadal (Colina & Moncada,2006). Cabe señalar que no solo se expresa en el tejido gonadal embrionario, sino también en el testículo adulto (Capel et al., 1993). Por otro lado, el gen SOXg contribuye a la determinación sexual del testículo. Es un gen autosómico de la familia de SRY. Durante el desarrollo embrionario, las gónadas sufren importantes cambios anatómicos y funcionales que implican diferenciación, maduración, proliferación y atresia de los compo- Cromosoma X, determinación ouárica Las influencias genéticas que determinan aI ovario no están asociadas a inductores especlficos. En ausencia de SRYyTDF se desarrolla una gónada femenina por impulso autosomal (McLa- ren,1990). Los genes presentes en el cromosoma X influyen en la diferenciación sexual tanto de varones como de mujeres y en numerosos caracteres independientes del sexo. En este cromosoma se locaJiza el gen que codifica el recepto r para los andrógenos. nentes celulares. Este proceso de morfogénesis gonadal puede ser dividido en varias fases: - Pregonadal. - Estadio indiferente. - Diferenciación sexual primaria. - Diferenciación sexual secundaria. El proceso es asimétrico cronológica y funcionalmente para el ovario y el testículo y va a presentar un desarrollo embriológico único respecto a los demás órganos del embrión. La gónada va a tener una función doble: producir hormonas y gametos; y un doble origen: la parte somática surge como un par de crestas sobre la porción media de los mesonefros (riñón primitivo) y las células germinales migran dentro de esta estructura desde el saco vitelino (Fernández, 1996). A finales de la cuarta semana de gestación las crestas gonadales emergen como engrosamientos del epitelio celómico en el lado ventromedial del par de mesonefros. El estado pregznadal comienza en el blastocito a los 4 o 5 días con la diferenciación de las células germinales primordiales (CGPÐ. Estas células derivan de las células ectodérmicas de la masa celular interna inicial (Gadner et al., 1985). Durante este período el embrión carece de estructuras gonadales específicas, aunque el desarrollo de las gónadas femenina y masculina está predeterminado por el sexo cromosómico. El período de la ginada indiferente se da desde la mitad de la tercera semana al final de la sexta del desarrollo embrionario. r- Neuropsicologfa de la sexualidad 18 Comienzan a apaÍecer y a colocarse los primeros esbozos de los apâratos genitales (Czþa, 1978). En el interior de la cresta van a tener cabida los esbozos gonadales somáticos. La gónada primitiva se forma en la cara ventromedial del pliegue mesonéfrico, a partir de una proliferación y espesamiento de la cresta genital y su epitelio celómico. Este último recubre la cresta genital y es la sede de multiplicaciones celulares; se condensa y prolifera por su cara interna formando los cordones sexuales primarios (Czþa, I97 8). En esta etapa, las células germinales primordiales que son el origen de los futuros gametos masculinos y femeninos (Czyba, 1978), realizan su migración mediante movimientos ameboides y alcanzan a la gónada primitiva. En el caso del ovario, la presencia de las células germinales es determinante para su formación, ya que en su ausencia no ocurre la diferenciación del mismo. En cambio, la diferenciación morfológica del testículo se lleva a cabo en ausencia de células germinales (Merchant, 1975). A partir de aquí, los gonocitos se mueven desde el epiteleo al parénquima y pierden sus características móviles intercalándose con células somáticas. Las células germinales se desplazan a través del endodermo intestinal, pasando más tarde al mesodermo del mesenterio y terminando en el epitelio celómico de la cresta genital. Los túbulos y glomérulos se localizan en el lado dorsal de la cresta gonadal formando la cresta urogenital. Al inicio del desarrollo, ambos primordios (mesonefros y cresta genital) compar- ten el mismo tejido sin que exista una lámina basal que separe el compârtimento gonadal del mesonéfrico. A las 5 o 6 semanas de la gestación, la gónada indiferente está compuesta por tres tipos celulares: - células germinales (futuros gametos); Desarrollo ontológico de la sexualidad t9 - células de sostén que darán lugar a las células de Sertoli en el testículo o a las células granulosas en el ovario; - células intersticiales, que derivan de la cresta genital y que darán lugar a las células esteroidogénicas: - células de Leydig (testículo) que segregan testosterona; - células tecales en el ovario. En este período la gónada no se distingue todavía de manera histológica como ovario o testlculo (Fernández,1996). Hasta antes de la séptima semana, las gónadas de ambos sexos son morfológicamente iguales, de manera que sus células son potencialmente dimórficas. Al ocurrir el proceso de determinación sexual, las células somáticas mostrarán un comportamiento dimórfico dirigido a la formación del ovario o del testículo. El primer signo de diferenciación sexual primaria en la gónada fetal masculina es la aparición de células de Sertoli (Jost, 1985) y su agregación en forma de cordones sexuales primarios. Las células epiteliales de los cordones están formadas por células somáticas y germinales, precursoras de las células de Sertoli y germinales, respectivamente. Esto tiene lugar en el embrión de 6-7 semanas (Rabinovici y Jaffe, I 990), constituyendo el primer signo de dimorfismo sexual gonadal, mientras que la consolidación de los cordones testiculares ocurre alrededor de la 8-9 semana. Por otro lado, la diferenciación ovárica se realiza en el curso de la octava semana del desarrollo embrionario, momento en que comenzará,las meiosis de los elementos germinales, transformándose las ovogonias en ovocitos (gametos femeninos). Alrededor del sexto mes de vida embrionaria, las células granulosas comienzan a organizarse rodeando a los ovocitos que se encuentran en fase de división y forman los folículos primordiales (Rabinovici y Jaffe, I 990). ) Neuropsicología de la sexualidad 20 Desarrollo ontológico de la sexualidad 2t Este proceso ocurre durante la diferenciación sexual secunda.- Por otro lado, el número de células de Leydig aumenta rápi- ria, en la que la actividad endocrina de las células testiculares, causan la regresión del conducto de Müller, la estabilización del conducto de Wolfi y la diferenciación en el üacto sexual masculino. La ausencia de hormonas testiculares lleva a la regresión del conducto de Mtiller, que desaparece casi por completo, quedando como remanente el conducto de Gartner y diferencián- damente y forman acúmulos que alcanzarán su desarrollo máxi- dose como tracto reproductor femenino. 2.2.L. Desørrollo testicular La diferenciación del testículo comienza hacia la sexta semana de la gestación con: el desarrollo gradual del blastema gonadal en cordones - - testiculares formados por las células de Sertoli (origen de los tubos seminíferos); el desarrollo de las células intersticiales de Leydig (Czy- ba,1978). Las células de Sertoli completan su diferenciación morfológica estableciendo uniones entre las células y formando membranas basales en la parte exterior de los túbulos, de esta manera ofrecen soporte estructural y metabólico a las células durante la espermatogénesis. Además, liberan la hormona inhibidora de los conductos de Müller (Rey et a1.,1995). En el curso del quinto mes,los cordones testiculares se transforman en tubos seminíferos y en el interior los gonocitos inician su maduración y se transforman en esPermatogonias. Estas últimas son células que dan lugar a los espermatozoides mediante el proceso de la espermatogénesis y que liberan mediadores para inhibir el proceso de meiosis de las células germinales' permaneciendo en estado de espermatogonias. mo hacia el tercer y cuarto mes del desarrollo embrionario. Es en este momento cuando tiene lugar la masculinizacióndel tracto genital. lJna vez que se haya acabado la diferenciación masculina del ffacto genital, se produce una regresión del número de células de Leydig en el curso del quinto mes. Por último, en el testículo fetal, que es una glándula endocrina muy activa, la células de Sertoli liberan hormonas bien caracterizadas: testosterona, como principal andrógeno, hormona antimülleriana e inhibinas (Fernández, 1996). Síntesis de esteroides en el testículo La síntesis de testosterona comienzavnavez producida la di- ferenciación histológica de las células de Leydig durante la octava semana de la gestación. El contenido de testosterona en los testículos afcanza un máximo hacia las 12-14 semanas y posteriormente va disminuyendo (Reyes et aL, 7973). La producción de testosterona en el adulto está controlada por hormonas gonadotrópicas hipofisarias: - LH: hormona luteinizante. - FSH: hormona folículo estimulante. - hCG: hormona solo presente en el feto. Es una gonadotropina placentaria que presenta una elevación máxima hacia la semana 10 del embarazo. Se postula que es el regulador ffsico de la esteroidogénesis inicial en el testículo, controlando la síntesis y liberación de testosterona (Voutilainen, 1992). En el testículo fetal humano se encuentran receptores para gonadotropinas desde la 10-11 semanas de la gestación. La ma- Neuropsicología de la sexualidad 22 yor capacidad de unión de estos receptores se sitúa enffe las semanas 10 y 15 (Molsberry et al., 1982). Durante los dos últimos tercios de la gestación' la síntesis de testosterona depende de las gonadotropinas, y por ello, los aspectos del proceso de virilización que tienen lugar durante este período están condicionados por esta hormona. La retroalimentación negativa que existe entre la LH y la testosterona es funcional durante el segundo trimestre, lo que justifica que durante este tiempo los niveles de LH son menores en los fetos masculinos que en los femeninos: estas diferencias sexuales desaparecen durante el tercer trimestre, cuando los niveles de testosterona son similares en ambos sexos (Fernindez,1996). Hormona antimülleriana (AMH) Se trata de una glucoproteína producida por las células de Sertoli. Es responsable de la regresión de la estructura que forma el origen de los genitales internos femeninos' el conducto de Mtiller. El gen responsable de dirigir las síntesis deAMH se sitúa en el brazo corto del cromosoma 19 (Fernández,1996). Las células del conducto de Müller son sensibles a la acción de AMH solamente durante un período crítico. La producción de AMH por el testlculo fetal es máxima durante la fase en la que se produce la regresión del conducto de Mtiller, aunque hay que destacar que también se sintetiza y libera durante el resto de la vida fetal e incluso en el período postnatal (Rey et al., 1995; l}an et aJ.,1987). Por otro lado, cabe destacar que se produce una liberación de esta hormona en el ovario adulto por parte de las células granulosas y su expresión está inhibida por la gonado- tropina (Voutilaine n, 199 2) . Desarrollo ontológico de la sexualidad 23 Inhibinas Son hormonas glucoproteícas liberadas por las células de Sertoli. Su función es regular la FSH hipofisaria. No se conoce todavía el significado biológico de su producción fetal, aunque sí sabemos que los niveles de FSH circulante son menores en los fetos masculinos durante el segundo ffimestre de gestación. Más adelante se hacen similares, cuando la actividad hormonal del testículo fetal disminuye (Fernán dez, 1996) . 2.2.2. Diferenciación del ouario En ausencia de inductores específicos, la gónada indiferenciada tiende a desarrollarse como ovario. En el feto hembra, el estado indiferenciado persiste largo tiempo, sin embargo, existe una proliferación del epitelio celómico y de las células germinales, que aumentan de tamaño y adquieren las características de la ovogonias. El máximo de elementos germinales se produce en- tre las semanas 16-20 y posteriormente se reduce. Este proceso comienza en el embrión y finaliza durante la menopausia (Fernández, 1996). En torno a la semana 72 o 13 se inician las primeras divisiones meióticas, lo que marca el comienzo de la organogénesis ovárica. La formación de folículos primordiales (ovocitos rodeados por una capa de células granulosas) afcanzasu pico m¿íximo hacia la semana 23 coincidiendo con el pico de FSH y comienzan a formarse los folículos primarios (Rabinovici y Jaffe, I 990) . Hacia la semana 25 los ovarios tienen ya las características definitivas. Los ovocitos se mantienen en la fase de la primera división meiótica y permanecen en ese estado hasta su posible ovu- 24 Neuropsicología de la sexualidad lación muchos años más tarde. La división meiótica se completa justo antes de la ovulación. Aunque el inicio de la meiosis parece depender de los factores locales, se desconocen los procesos de regulación de la morfogénesis ovárica. El crecimiento, desarrollo y mantenimiento de la foliculogénesis está influido por las gonadotropinas fetales, en especial, la FSH (Fernández, 1996). Los ovarios fetales no producen grandes cantidades de esteroides durante los dos primeros trimestres, la actividad de las enzimas esteroidogénicas es muy baja en el ovario durante este período (Voutilainen y Mille¡ 1988); es más, el ovario carece de receptores para hCG o LH durante los dos primeros trimestres de gestación (Huhtaniemi et al., 1988). Esto muestra la diferente actividad endocrina de la gónada fetal femenina respecto a la masculina. 2.3. Nrvsl soM,{Trco Tanto los embriones masculinos como femeninos poseen dos pares de conductos genitales y durante la quinta y sexta semana, al igual que en la gónada, los conductos genitales presentan un estado indiferenciado y están presentes en ambos sexo. Desarrollo ontológico de la sexualidad 25 sustancias: andrógenos y hormona antimülleriana (Fernández, 1996). 2.3.1. Acción horrnonal Las hormonas sintetizadas por el testfculo fetal van a consti- tuirse en inductores de la diferenciación somática. Estas hormonas pueden actuar localmente sobre las estructuras próximas a su lugar de producción y, por tanto, de forma independiente de un sistema fetal que las transporte. Estos inductores son vehiculizados por la sangre para llegar a las estructuras diana sobre las que ejercen sus efectos. La testosterona es el principal andrógeno producido por los testículos fetales. En numerosos órganos diana, la acción de este andrógeno está condicionada a su conversión en otros esteroides. La testosterona entra en las células por difusión y sufre diferentes transformaciones que son específicas para cada tejido. Por una parte, la aromatización a estrógenos es el mecanismo general de acción en el sistema nervioso; por otra, la 5-alfa-reductasa actúa sobre los genitales externos e internos. Los esteroides formados interaccionan con receptores nucleares y desencadenan sus efectos biológicos (Fernández, 1996). .Wolff y los conEn el embrión coexisten los conductos de ductos de Mtiller, precursores del tracto reproductor masculino y femenino, respectivamente. Si no se producen alteraciones se va a reafizat una morfogénesis que consiste en la regresión de uno y la persistencia y diferenciación del otro. Este desarrollo o involución tiene lugar durante el tercer mes de vida fetal. El desarrollo femenino es independiente de las secreciones gonadales, mientras que la diferenciación masculina requiere dos 2.3.2. Diferenciación de los genitales internos Patrón masculino El primer signo de diferenciación masculina del ffacto urogenital es la degeneración del conducto de Müller. Este proceso tiene lugar al poco tiempo de formarse los cordones espermáticos y se exdende a todos los conductos que desa- 26 Neuropsicología de la sexualidad parecen. Al mismo tiempo, los conductos de\Wolffse desarrollan y adquieren las características del sistema eyaculador masculino. La porción que se encuentra adyacente al testículo adopta una forma característica de arrollamiento para formar el epidídimo, la parte central se convierte en el conducto deferente, y las vesículas seminales se desarrollan a partir de las porciones inferiores del conducto. El desarrollo de estas glándulas depende de las interacciones del mesénquima y el epitelio, reguladas por la acción de los andrógenos. El proceso de interacción se inicia en las células mesenquimáticas poseedoras de los receptores a andrógenos. Patrónfemenino El tracto reproductor femenino se deriva del conducto de Mtiller y la ausencia de testosterona lleva a la regresión del conducto de'Wolfl desapareciendo casi por completo y persistiendo solo de forma rudimentaria. Las porciones ceFálicas del conducto dan lugar a las trompas de Falopio, y las porciones caudales se funden para formar el útero y el tercio superior de la vagina. Este desarrollo femenino no requiere la presencia de inductores específicos, y se obtiene incluso, en ausencia de gónadas. El contacto de los conductos de Mtiller con el seno urogenital da lugar a una proliferación endodérmica intensa que origina la placa útero-vaginal que se localiza enffe los conductos de Mtiller y el seno urogenital (O'Rahill¡ 1983) y que tiene algún componente de los conductos de \Øolff. Esta placa, en su proliferación, incrementa la distancia entre el útero y el seno urogenital y se canaliza paraformar la luz de la vagina (Fernández, 1996). Desarrollo ontológico de la sexualidad 1'7 2.3.3. Diferenciación de los genitales externos Antes de la octava semana los genitales externos son idénticos y bipotenciales en su capacidad de diferenciación en ambos sexos. Las estructuras que se diferencian son el seno urogenital, los pliegues uretrales, la hendidura urogenital y la protuberancia genital (Fernández, 1996). Desørrollo møsculino En contraste con lo que sucede en el conducto de \Wolf[ el seno urogenital y el tubérculo genital presentan actividad de 5-alfa-reductasa, incluso antes de que el testículo fetal haya desarrollado su capacidad para sintetizar testosterona. La testosterona actúa como una prohormona sobre estas estructuras, transformándose en dihidrotestosterona (DHT), que se une al receptor nuclear desencadenando el Proceso de viriliza- ción de los genitales externos. El estímulo androgénico debe ser de intensidad suficiente para inducir un desarrollo completo de los genitales externos (Fernández , 1996) y así la testosterona llegue a todas las estructuras a través de la circulación. Los genitales externos masculinos comienzan a diferenciarse poco después de la virilización del conducto de Volff; el seno urogenital da lugar a la próstatay ala uretra prostática; los pliegues uretrales se fusionan para formar el pene, y el tubérculo ge- nital forma el glande y los cuerpos cavernosos. Las protuberancias se sitúan por detrás del tubérculo genital ¡ al fusionarse, los pliegues uretrales forman el escroto. Estos procesos tienen lugar durante el primer trimestre de vida fetal. La masculinización de los genitales externos por la DHT requiere niveles adecuados y se produce durante un período crítico 28 Neuropsicología de la sexualidad de la vida fetal: debe ser anterior a la décima o undécima sema- na, que es cuando comienza a diferenciarse la vagina. La DHT es la responsable de inhibir el desarrollo de la vagina. D ife r e n c i a c i ó n fe m e n i n a En ausencia de inductores, los genitales externos se diferencian en sentido femenino. El seno urogenital forma la uretra y los dos tercios inferiores de la vagina, los pliegues uretrales forman los labios menores, y las protuberancias genitales, los labios mayores. El tubérculo genital sufre un crecimiento limitado y forma el clítoris (Fernández, 1996). En conclusión, la biología del desarrollo, la genética y la endocrinología, con sus diversas metodologías, han contribuido al conocimiento actual sobre los procesos de sexuación prenatales. El sexo genético determina la morfogénesis gonadal a rravés de mecanismos complejos que implican genes de diferentes cromosomas, con un proceso de regulación que está actualmente en fase de estudio. El sexo gonadal, unavezestablecido, condiciona la diferenciación de los genitales internos y exrernos a través de la presencia o ausencia de secreciones endocrinas. La diferenciación genital depende de la acción de agentes morfogénicos que deben tener una concentración adecuada y actuar en un tiempo preciso del desarrollo embrionario. Además, la estructura diana debe ser sensible a la acción de estos agentes inductores. 2.4. Spxo FENoríPrco La mayor parte de las características sexuales -primarias o secundarias- se desarrollan a partir de precursores embriológi- ?c) Desarrollo ontológico de [a sexualidad cos comunes o bipotenciales. Esto posibilita un amplio espectro de diferenciación en cada nivel de la organización sexual entre hombres y mujeres. Sobre las estructuras iniciales, la acción de los inductores de masculinización provoca el proceso de la morfogénesis en un tiempo preciso del desarrollo ontogénico como resultado, la ¡ morfología fenotípica del varón y la mujer. 2.41. Sistema reproductor y funcionamiento gonadal Las gónadas de ambos sexos, testlculo y ovario, ejercen una doble función produciendo hormonas sexuales y gametos e in- fluyendo en el comportamiento. Así, la función gonadal provoca diferencias en la morfología y en el funcionamiento. Después del nacimiento, el desarrollo sexual permanece detenido hasta la pubertad. En este momento los órganos reproductores se activan por estimulación de las gonadotropinas incrementadas, debido a la maduración del eje hipotálamo-hipofisario. Sisterna reproductor mas culino Los órganos reproductores primarios o gónadas son los testículos o glándula sexual masculina, los cuales poseen dos tipos de funciones separadas, pero a lavez muy relacionadas: - la producción y almacenamiento de células germinales masculinas (espermatozoides). Estos son producidos por los túbulos seminíferos y son transportados por un sistema de canales para llevar a cabo el proceso de fertilización. - la biosíntesis y secreción de hormonas sexuales masculinas (andrógenos) (Fernández, L996). Los andrógenos, son 30 Neuropsicología de la sexualidad producidos por las células intersticiales de Leydig. Se encargan de la producción de hormonas sexuales masculinas que son las responsables de algunas funciones atribuidas a los varones (Bardin, 1936). Los testículos tienen forma ovoidea y esrán cubiertos por una fibra de la que penetran hacia el interior del tejido resricular tabiques que lo dividen en unos lobulillos de forma piramidal y con base en la periferia. En el polo poste ior del testículo, lu r de entrada y salida de los vasos testiculares, está situado el epidídimo, que es una estructura con forma de capuchón que desemboca en un conducto único, el conducto deferente (Tiesguerres, 1989). a) Espermatogénesis Este proceso comienza en la pubertad en respuesta a las gonadotropinas. Consiste en la formación de los espermatozoides en los túbulos seminíferos que presenran un tejido formado por cuatro a ocho filas de dos tipos de células: las germinales y las de Sertoli. Las células germinales constiruyen la mayor parte del tubo seminífero: - las que se encuentran sobre la membrana son las espermatogonias, que presentan frecuentes mitosis y forman células dando lugar a los espermatocitos primarios; - los espermatocitos primarios sufren una división meiótica, dando lugar a dos células con la mitad de cromosomas, denominados espermatocitos secundarios; - tras una nueva división se forman otras células de menor tamaño, las espermátides, que no se dividen de nuevo, sino que sufren un proceso de metamorfosis y maduración, formando cada una de ellas un espermatozoide. Desarrollo ontológico de la sexualidad 3t Las células de Sertoli ofrecen sostén a las células germinales y participan en el control de la espermatogénesis (Bardin, 1986). Estas células tienen receptores parala FSH, hormona hipofisiaria que controla la espermatogénesis, y bqo el estímulo de esta se producen ciertas sustancias que pasarán a las células germinales para que éstas puedan llevar a cabo su metabolismo. Además, también mantienen a estas células aisladas del resto del organismo mediante la llamada barrera hematotesticular. Por otro lado, la testosterona, hormona masculina que peneffa en el túbulo seminífero para completar la formación de los espermatozoides, lo hace gracias a una proteína transportadora producida en la célula de Sertoli, llamadaABP La FSH también actúa sobre las células de Sertoli, estimulando la espermatogénesis a través de la producción de ABP y de la formación de sustancias energéticas que son necesarias para el metabolismo normal de las células germinales. Finalmente, las células de Sertoli también intervienen en la modulación de la espermatogénsis mediante la inhibina, que actúa sobre la hipófisis disminuyendo los niveles de FSH (Rivier et al., 1986). b) Biosíntesis y secreción de andrógenos Se produce en las células intersticiales de Leydig localizadas entre los túbulos seminíferos y en contacto con los vasos sanguíneos y linfiíticos testisculares permitiendo que sus secreciones pasen rápidamente a la sangre. Estas células producen: - testostefona; - en pequeñas cantidades el estradiol-l7 beta, hormona femenina que deriva de la anterior. La testosterona causa amplios y variados efectos en el organismo, tanto sobre los órganos sexuales internos como externos' \ 32 Neuropsicología de la sexualidad La acción de este tipo de andrógenos se encuentra bajo el control de la LH hipofisaria y acttr mediante la interacción de receptores localizados en las membranas celulares. Son receptores espe- cíficos localizados en el interior del núcleo celular del tejido correspondiente, tras su transformación en un metabolito activo: la 5-alfa-DHT (Tiesguerres, 1989), responsable de mantener los caracteres sexuales secundarios masculinos. La transformación de testosterona en 5-alfa-DHT ocurre mediante la acción de la enzim a 5-alfa-reductasa y es imprescindible para su actuación sobre la mayoría de los tejidos. La ausencia de la enzima capaz de transformar la testosterona en 5-alfaDHT bloquearía las acciones de la que es responsable y tendría efectos sobre el fenotipo. En el sistema nervioso central (SNC) no es necesaÅala 5-alfa-DHT (Tiesguerres, 1996). De esta manera se regula la síntesis de testosterona que se produce de manera pulsátil y con ritmo circadiano (Moreno Esteban et al., 1983). La 5-alfa-DHT es la responsable de la rnayoría de las acciones androgénicas en el varón adulto. Este paso es importante porque a veces falta la enzimay no se produce acción hormonal masculina a pesar de que el testículo produzca testosterona de manera normal. Los andrógenos necesitan unirse a un recePtor androgénico intranuclear para poder ejercer su acción. La presencia de mayores o menores cantidades de este receptor va a determinar el grado de sensibilidad del tejido a andrógenos. Como segundo paso obligado, ocurre un aumento de la síntesis de RNA que da lugar ala síntesis proteica, por lo que se producen sustancias que modificarán el metabolismo celulat y por tanto, la acción de las hormonas, hasta tal punto que la ausencia de receptor da lugar a la imposibilidad de acción hormonal. La importancia de este hecho radica en que independientemente del sexo cromosómico y gonadal, con presencia abundante de testosterona, los posi- :r) Desarrollo ontológico de Ia sexualidad bles efectos de esta sobre los caracteres sexuales secundarios mas- culinos van a depender de que existan receptores, ya que sin su presencia no hay fenotipo de varón ni ciertos comportamientos masculinos. S is tema repro duc to r femenino Las gónadas femeninas son los ovarios. Son dos cuerpos ovalados que se encuentran alojados en la pelvis, a la enffada de las trompas de Falopio y cercanos al útero (Fernández, 1996) formando así el tracto reproductor femenino. En el ovario destacan estructuras fundamentales, como los folículos, cuya organización y componentes van a sufrir una serie de cambios coincidiendo con el grado de desarrollo de los ovocitos contenidos en su interior. Estos cambios están relacionados con la doble función de los ovarios: - por una parte, son los responsables de la secreción de las hormonas femeninas; - por otra, son los encargados de proporcionar los gametos femeninos, los óvulos, para su futura fecundación (Sánchez Criado,I992;Tiesguerres, 1989; Yen yJaffe, l99l) A partir de la pubertad, se inicia la maduración folicular. Los ovocitos, rodeados de una serie de células, surgen del folículo ovárico donde se alojaban durante la ovulación, y penetran en las trompas de Falopio (S¿ínchez Criado, 1992; Tlesguerres,1989). Finalmente, si el óvulo es fecundado por el espermatozoide se formará el óvulo maduro y se implantará.en el útero. /-) .') 34 Neuropsicología de la sexualidad a) Ovulación El proceso de maduración de los oocitos ocurre dentro del ovario, en unas estructuras especiales denominadas folículos que van a ser también las encargadas de la producción hormonal. El folículo está formado por una o varias capas de células granulosas alargadas que rodean el oocito. El complejo oocito-células granulosas, rodeado por una lámina basal, se denomina folículo primordial y da lugar a los folículos primarios. Cuando estos crecen, pueden observarse cambios en las células por fuera de la lámina basal, que provocan la aparición de una serie de capas concéntricas de células alargadas denominadas células tecales (Dizeregay Hodgen, 1981). A medida que se va produciendo el crecimiento y proliferación de las células granulosas y tecales, el folículo va aumentando de tamaño y aparecen acúmulos de líquidos entre las células granulosas, que van confluyendo a la vez que aumentan de tamaño, dando lugar a una cavidad llena de líquido, denominada antro. Esta formación transforma el folículo primario en un folículo de Graafl en el que el oocito se encuentra rodeado de dos o tres capas de células granulosas que constituyen el <cúmulo oóforor, el cual está unido al resto de las células granulosas por uno de sus lados. En la mujer adulta ocurren mensualmente distintos cambios hormonales que culminan con la liberación por parte del ovario de un óvulo fecundable en lo que constituye el ciclo menstrual ovulatorio normal (Goodman y Hodgen, 1933). En los ovarios, en el momento del nacimiento, hay aproximadamente dos millones de folículos primordiales. Entre la época del nacimiento y la pubertad gran parte de estos folículos sufren un proceso de atrofia, de forma que solo 400.000 gametos Desarrollo ontológico de la sexualidad 35 están presentes en el ovario de la mujer que comienza su vida fertil. De estos, solo 400 van a tener la oportunidad de madurar completamente y pasar a las úompas de Falopio para ser fecundados. El resto sufre un proceso de atresia. El folículo que cada mes consigue completar el proceso de maduración será el que, al romperse, libere al oocito en la cavidad abdominal. De allí será captado por la trompa de Falopio y transportado al interior del útero. A partir de los restos foliculares que quedan en el ovario, se va a producir una transformación de las células, formándose el cuerpo lúteo, cuya función será la secreción hormonal en la segunda fase del ciclo (Tiesguerres,1989). Thas la ruptura del folículo, las células granulosas transformadas, más las células tecales y los vasos, se entremezclan para dar lugar al cuerpo lúteo, que será el responsable de la secreción de las hormonas sexuales durante la fase postovulatoria del ciclo. b) Biosíntesis y secreción de estrógenos Las hormonas sexuales femeninas producidas en el ovario son fundamentalmente el estradiol y la progesterona, y varias hormonas no esteroideas como la inhibina, la relaxina y algunos factores locales. Todos los esteroides ováricos se producen fundamentalmente en las estructuras foliculares y en el cuerpo lúteo, que poseen el sistema enzimâtico complementario requerido para la forma- ción de hormonas esteroideas. El estrógeno más importante secretados por el ovario es el l7-beta-estradiol siendo variable a lo largo del ciclo menstrual. Así, a lo largo del proceso de maduración folicular y paralelamente al mismo, se sintetiza el estradiol de forma creciente hasta el momento de la ovulación. De esta manera disminuye los ni- 36 Neuropsicologla de la sexuaÌidad veles en el momento de la misma y vuelve a elevarse hasta niveles pârecidos a los preovulatorios gracias a la contribución del cuerpo lúteo en la segunda fase del ciclo, en la cual, los niveles de es- tradiol se acompañan de un incremento de niveles de progeste¡ona. Además se producen también ciertos niveles de estrógenos (Sánchez Criado, L992;Tiesguerres, 1989; Ben E¿Jaffe, 1991). Por otro lado, la progesterona es un esteroide que se produce a lo largo de todo el ciclo menstrual tanto en el folículo como en el cuerpo lúteo y en las glándulas suprarrenales. A partir de la ovulación el cuerpo lúteo es su productor principal y el incremento comienzaya a observarse poco antes de la ovulación, siendo máximo cuando el cuerpo lúteo está en plena acción con células granulosas perfectamente luteinizadas (Thesguerres, 1994). Además presenta acciones encaminadas a facilitar la gestación. Actúa sobre el endometrio, que ha proliferado por la acción de los estrógenos, y lo transforma preparándolo para la posible nidación del embrión (Tiesguerres, 1994). Los esteroides ováricos actúan en los tejidos diana de diferentes partes del organismo) a través de su unión a receptores específicos intracelulares. El complejo estrógeno-receptor modifica la transcripción genéticay da lugar a diversos fenómenos bioquímicos que conducirán aI efecto biológico. Son responsables del desarrollo y mantenimiento de los caracteres sexuales secundarios femeninos. Cabe tener en cuenta que, a lo largo del desarrollo folicular primero y del cuerpo lúteo después, los niveles hormonales presentan una serie de oscilaciones con periocidad mensual. Por otro lado, producen ciertos cambios en el resto del organismo de la mujer que siguen un patrón característico acompañando a las hormonas femeninas. Estos cambios incluyen alteraciones psicológicas, pues los estrógenos actúan también a nivel del SNC. Desarrollo ontológico de la sexualidad )/ 2.4.2. Eje hipotálamo hipofisario gonadal A comienzos del siglo pasado, se intuyó por primera vez el papel de la hipófisis en la reproducción y a mediados de los años veinte, dos grupos de investigadores, de manera independiente, descubren las hormonas responsables: las gonadotropinas hipofisarias LH y FSH, y se dedican a estudiar sus efectos. El cono- cimiento sobre esta tema ha aumentado de manera considerable en los últimos años gracias a la disponibilidad de nuevas metodologías parala identificación de los diferentes procesos celulares y moleculares que controlan el desarrollo puberal (CastroFeijoo & Pombo, 2006). Así, múltiples estudios han permitido la identificación y lacaracterización de una gran parte de los mecanismos fisiológicos que regulan la función del eje hipotálamohipófiso-gonadal durante el desarrollo sexual (figura 1) (Ojeda, 2002; Philip &. Lazar, 2005) . Los cambios fisiológicos más importantes asociados al proceso de maduración sexual se encuentran bajo la regulación del eje hipot:ílamo-hipófiso-gonadal. Este eje depende de una serie de cambios que se producen en el cerebro y que dan lugar a su activación: la hormona liberadora de gonadotropinas (LHRH o GnRH) es vertida a la circulación estimulando la secreción pulsátil de las gonadotropinas adenohipofisarias LH y FSH que se producen en células de la hipófisis anterior y entre ambas controlan la función de las gónadas tanto masculinas como femeninas, estimulando: - la maduracióny liberación de los correspondientes gametos; . - la producción y secreción de las hormonas sexuales. Neuropsicología de la sexualidad 38 Figura 1 Representación esquemática del eie hipotálamo-hipófiso-gonadal Itpo¡C¡rr Cådo.dd. ìhrsrø i Hlpófbb ADB{ffi tH , FsH Gó¡oà E*oldr¡ y pffidos 39 tein) queconduce aI incremento de la biosíntesis de andrógenos a través de la puesta en marcha de varias enzimas, como la hidroxilasa o beta-hidroxilasa, que conducirán a la transformación del colesterol en varias sustancias, hasta llegar a la testosterona' que será vertida en sangre. A su vez, el nivel de testosterona afcanza' do ejerce una acción de retroalimentación negativa sobre la LH. El regulador de FSH es la inhibina, que se produce en las células de Sertoli en conexión con las células germinales para que' GnRH & I Desarrollo ontológico de la sexualidad Ê iol¡ Coræl¡ro ¡cxuqls¡ I Fuente: Castro-Feijoó & Manuel Pombo, 2006 Regulación testicular El eje hipot:ílamo-hipófisis controla las dos funciones de los testículos: - espermatogénesis; - secreción de testosterona. La FSH activa la espermatogénesis y la función de las células de Sertoli, uniéndose especlficamente a los receptores, localizados en las membranas de las células de Sertoli. Así, a través de la producción de varias sustancias, regula la proliferación y maduración de las células germinales (Bardin, 1986). La LH es la responsable del funcionamiento de las células intersticiales de Leydig. Se une a los receptores de su membrena, activando la proteína SIAR (Srera id.ogenic acute regulatoryr Pro- a más espermatozoides producidos, más cantidad de inhibina secretada, ajustando los niveles de FSH a la espermatogénesis (Burger, 1987; Rivier et al., 1986). La secreción de las gonadotropinas se encuentra a su vez regulada por la secreción de una hormona hipotalámica' la GnRH o LHRH (hormona liberadora de gonadotropinas) que alcanza la hipófisis anterior estimulando la síntesis y secreción de LH y FSH. La GnRH se produce en neuronas localizadas en el núcleo ârcuato hipotalámico. Tiene una vida media de muy pocos minutos, por lo cual el control que ejerce sobre el eje-hipotálamohipófiso-testicular depende de su secreción pulsátil (Rodríguez Poyo-Guerrero et â1., 1989) y de esta manera regula la secreción de la LH y la FSH. Esta pulsatilidad es necesaria para que estas ultimas se sinteticen adecuadamente. Si se administra continuamente LHRH ocurre un fenómeno de desensibilización y la LH y la FSH disminuyen (Conn et al., 1987); sin embargo, la magnitud de la respuesta es proporcional al ambiente hormonal de esteroides sexuales y a los niveles de inhibina. Así, la testosterona y la inhibina son capaces de regular la función hipofisaria, por acción directa sobre las células gonadotropas o de manera indirecta, a través de la modificación de la frecuencia de secreción de la GnRH. La GnRH no solo estimula la secreción de la LH y la FSH, sino también su síntesis. su mecanismo de acción sobre las célu- Neuropsicologla de la sexualidad 40 las adenohipofisarias se ejerce a rfavés de receptores de membra- na localizados en las células gonadotroPas (Burge¡ 1987)' Los mecanismos fundamentales de regulación entre las gosecreción testicunadotropinas yl lar siguen el prin bajan los niveles Desarrollo ontológico de la sexualidad 4r células luteinazadas constituyentes del cuerpo lúteo, que a su vez asumirá la secreción hormonal durante la segunda fase del ciclo. A su vez, la secreción de LH y FSH está dirigida a través de sistemas de retroalimentación por la propia secreción de esteroi- negetiva' Cuando des ováricos (Dizmegay Hodgen, 1981). Segun van incrementan- efecto inhibitorio sobre el hipotálamo, y aumenta la secreción de GnRH, que a su do los niveles de estradiol al ir madurando el folículo, van a disminuir los niveles de gonadotropinas, fundamentalmente la FSH. Por otro lado, la GnRH o LHRH es la hormona hipotalámica que regula la secreción de LH y FSH, a través de la modificación de su frecuencia de pulsos. La secreción pulsátil de LHRH a nivel hipotalámico es capaz de desencadenar, en las células de la hipófisis, la liberación de LH y FSH. Sin embargo, la magnitud de la respuesta es proporcional al ambiente estrogénico (Knobil, 1980; Rodríguez Poyo-Guerrero et al., 1989). La interrelación hipófisis-ovario es el lugar de génesis de la mayorírde los cambios hormonales que dan lugar al ciclo menstrual, desempeñando el hipotálamo un papel secundario en su vez estimula las gonadroffopinas. La LH aumentada producirá una elevación de los niveles de testosterona y viceversa' La secreción de FSH hipofisaria se regula fundamentalmenre por el sistema inhibina-activina que tiene su acción selectiva d.rrtro del nivel hipofisario. La inhibina disminuye los niveles de FSH tanto basales como tras el esdmulo de GnRH, sin modificar los niveles de LH, y la activina, ejerciendo un efecto contra- rio (Rivier er al., 1986). Regulación ouárica El control de la función ovárica se lleva a cabo por las hormonas gonadotrópicas hipofisarias que incluyen a la LH y la FSH. En condiciones normales tanto la LH como la FSH presentan una secreción pulsátil en la mujer, al igual que en el varón, con picos cada hora y media aproximadamente (Knobil, 1930). Además, ambas gonadotropinas cuentan con un perfil cíclico mensual con valores de FSH más elevados al final de la fase lútea y comienzo de la fase folicular y con un pico marcado durante la etapa ovulatoria. La LH presenta valores estables a lo largo del ciclo, con un pico periovulatorio mayor que la FSH' La FSH estimula la maduración de los folículos ováricos, junto con el incremento de la producción de estradiol 'La"LH, a .,.r,0.r, contribuye con la FSH a la maduración folicular y será la responsable de la transformación de las células foliculares en las desarrollo. La interacción de todos los componentes mencionados hasta ahora, a través de los circuitos de información positivos y negativos, da lugar a la instauración del ciclo menstrual (Tiesguerres,1989). 2.4.3. Caracteres sexuales secundarios Los caracteres sexuales secundarios son signos fisicos y fisiológicos de madurez sexual, que se observan en los dos sexos pero que no afectan al sistema reproductor. Los cambios gonadales se producen durante la adolescencia y son los que van a propiciar este tipo de modificaciones, incluyendo cambios corporales que acentúan el dimorfismo sexual aparente (figura'2). 42 NeLrropsicología de la sexualidad Vartin Coincidiendo con el incremento del volumen testicular, tiene lugar la pubarquia. Este proceso flsico puberal que se da en los dos sexos) indica y es consecuencia del incremento de andrógenos adrenales. Se obserua el crecimiento del vello axilar y facial, y más tar- de, el vello corporal se extiende hacia las extremidades, pecho y espalda. Las glándulas sebáceas de la cara, hombros y pechos son especialmente sensibles a la estimulación androgénica, lo que da como resultado la producción de sebo y grasa que consriruye el típico acné. El incremento del desarrollo mamario se puede atribuir a una baja relación de andrógenos/estrógenos como consecuencia de la activación del metabolismo periferico de andrógenos a estrógenos o de escasa producción adrenal de andrógenos. Por último, la laringe se amplía y las cuerdas vocales se engruesan, lo que produce un tono devoz más grave. 43 Desarrollo ontológico cle Ia sexualidad ne lugar el estirón puberal. Le sucede una rápida desaceleración del crecimiento, que alcanza su valor máximo con la aparición de las primeras reglas, después de las cuales las chicas continúan creciendo durante un tiempo. El estirón puberal se acompaña de importantes cambios en la composición corporal relacionados con el aumento de la masa muscular y el tejido adiposo. El ensanchamiento de la cintura pélvica y la distribución periferica del panículo adiposo de las mamas, los glúteos y los muslos alcanzan su máxima amplitud al final de la pubertad, acentuando de esta forma el dimorfismo sexual. Figura 2 Ca racte res sexu a les secu nd a ri os Carnbio en la vor voz fnås gróve D,¡¡¿rtollo de las glåndulas m¿mari¿s Mujer El primer signo que se da es la telarquia o desarrollo mamario que finaliza después de la menarquia. La completa maduración de la mama requiere el concurso de otras hormonas como la progesterona, GH, prolactina y esteroides suprarrenales. La pubarquia sigue de inmediato a la telarquia, que depende de los andrógenos adrenales, como en los varones, y se completa en las últimas fases de la pubertad. La aparición del vello axilar no tiene lugar hasta estadios más avanzados de la pubertad y progresa con mayor rapidez que en el varón. Depende de las secreciones ováricas y coincide en el tiempo con el aumento de la actividad de las glándulas sudoríparas, de manera que el olor corporal cambia sensiblemente. Precediendo o acompañando los primeros signos clínicos y siempre anres que la menarquia, tie- Murculatu¡a ¡nas der¡l.roåTada Desarrollo y rnadur¿cíón de los érganos sexuales Acunrui¡cién de gras.rs en algilnas zonas 44 Neuropsicología de la sexualidad En resumen, el desarrollo ontológico de la sexualidad comienza con el sexo cromosómico. Las moléculas de ADN que contienen el código genético característico de la especie humana se ponen de manifiesto en el curso de las divisiones celulares bajo la forma de 46 cromosomas: 44 autosomas y 2 cromosomas sexuales designados por las letras )O( en el sexo femenino y XY en el sexo masculino. Los gonosomas constituyen los caracteres sexuales primordiales. Así, de la mitad de la tercera semana al final de la sexta del desarrollo embrionario aparecen y se colocan en su lugar los primeros esbozos de los apararos genitales. En la cuarta y quinta semana se individualizala gónada indiferenciada como la emigración de las células germinales hacia un blastema común. A partir de esta gónada se coloca un sisrema de vías genitales indiferenciadas: conducto de \Øolffy conducto de Mi.iller, que servirán para el desarrollo de los futuros gameros. Finalmente, en la extremidad caudal del embrión se individualizan los esbozos indiferenciados de los órganos genitales exrernos. Al principio de la séptima semana del desarrollo embrionario se realiza la diferenciación testicular y las células del blastema común se diferencian en cordones tesriculares y en células interticiales de Leydig, estas últimas son las responsables de la diferenciación masculina de las vías y órganos genitales externos en el curso del tercer y cuarto mes. Por otro lado, la diferenciación femenina se realiza en el curso de la octava semana del desarrollo, las células del blastema común proliferan en forma de cordones celulares que terminan formando los folículos primordiales. En cuanto a los genitales externos e internos, la diferenciación tiene lugar durante el tercer y cuarro mes del desarrollo embrionario, en ausencia de secreciones ováricas. La última etapa de la ontogénesis corresponde al período de la pubertad, que se inicia hacia los 1 1 años y la madurez sexual se al.canzahacia los 16 años. En ambos sexos) este período se carac- Desar¡ollo ontológico de [a sexualidad 45 teriza porque se encuentra bajo la influencia del eje hipotálamohipofisario, las gónadas secretan suficientes hormonas sexuales para provocar el crecimiento acelerado de los órganos genitales y la aparición de los caracteres sexuales secundarios. Estas transformaciones conducen a la puesta en marcha de los aparatos genitales y, como consecuencia, se acentúan las diferencias entre los caracteres genitales y fenotípicos del hombre y la mujer, incluyendo las estructuras sexualmente dimórficas. Los esteroides sexuales, especialmente los estrógenos, producen múltiples efectos en diferentes tejidos. En el sistema nervio- so central, ejercen gran cantidad de efectos organizadores, participando en el establecimiento de las estructuras sexualmente dimórficas que vamos aveÍ a continuación. III Diferenciación sexual del cerebro En el sistema nervioso central de varones y mujeres se encuentran diferencias tanto estructurales como funcionales. Estas diferencias involucran el número de neuronas, su morfología e interconexiones que han sido asociadas con cambios en la fisiología neuronal y en la conducta. Las diferencias morfológicas, bioquímicas, histológicas, fisiológicas y de conducta entre varones y mujeres se denominan dimorfismo sexual. Las causas que lo producen son múltiples: genética, hormonal y ambiental que al interaccionar dan como resultado este dimorfismo. La determinación del sexo se inicia con la fertilización del óvulo y el período de diferenciación gonadal, antecede a la diferenciación del sistema nervioso central (SNC), evento reconocido como dimorfismo sexual cerebral. Más adelante, las secreciones hormonales por parte de las gónadas durante el período crítico y las interacciones que se producen entre sí en condiciones normales, hacen que se establezcan estructuras sexualmente dimórficas en etapas tempranas del desarrollo embrionario. Numerosas investigaciones han presentado evidencia de que existe dimorfismo sexual en la estructuray funcionalidad del ce- 48 Neuropsicología de la sexualidad rebro, señalando la presencia de diferencias morfológicas, neuroquímicas y fisiológicas, así como la existencia de circuitos neuronales sexualmente específicos que van a formar parte de las bases somáticas que explican la importancia de la función cerebral en la conducta reproductiva de las especies. La literatura sobre las diferencias macroscópicas del cerebro humano plantea una mezcla de observaciones científicas y sesgos culturales. A lo largo de la evolución han surgido varias estrategias para diferenciar el cerebro en dos fenotipos, y el hombre ha intentado explicar cómo se originan estas diferencias. Hoy en día se disponen de teorías globales bien fundamentadas conceptual y metodológicamente que explican el dimorfismo sexual cerebral en animales. La evidencia experimental, donde los modelos murinos constituyen la herramienta experimental más preferible, indica que existe un dimorfismo sexual en las estructuras neurales. Pero hay que mencionar que su significado y sus posibles consecuencias, aún no son bien conocidas. Diferenciación sexual del cerebro 49 experimentación en humanos, obliga a utilizar como modelos de estudio exclusivamente a aquellos individuos en los que por accidentes naturales se han producido alteraciones en el proceso de la diferenciación sexual. Otro aspecto que dificulta el estudio de la sexualidad del cerebro es el hecho de que la bibliografía especializada se presenta influida por prejuicios y de este modo se establecen dos corrientes enfrentadas: - un grupo de investigadores que se basan en el determinismo de las variables biológicas; - otro grupo de investigadores que se basan en la influencia cultural y social por encima del impacto de las señales fisiológicas. Muchas de las observaciones que se han realizado son controvertidas, sin embargo, los hallazgos más aceptados muestran diferencias en el cerebro masculino y femenino, por ejemplo, en el tamaño de algunas áreas y núcleos, distinta morfología neuronal y glial y en los patrones sinápticos de determinadas áreas. Este tipo de estudios animales garantiza un control riguroso de las variables y de las condiciones experimentales, sin imponer restricciones bioéticas, pero los resultados son muy variables entre las distintas especies debido a los diferentes patrones de respuesta hormonal. Por otro lado, se corre el riesgo de ofrecer una genenlización simplista de conclusiones obtenidas con modelos animales a procesos psicológicos equiparables en los humanos' En cuanto a la especie humana, se ha empleado con menos frecuencia ya que existen barreras éticas obvias. Existen dos aspectos básicos que limitan nuestro conocimiento sobre el posible determinismo de las variables fisiológicas en la consecución del sexo neural. Por una parte, es imposi- ble cuantificar las influencias socioambientales y su interacción con los sustratos neurales y endocrinos. Por otra, la ausencia de 3.1. INr¡neccróN DE r"cs HORMONAS SEXUATES EN EL SISTEMA NERVIOSO CENTRAI El cerebro se puede considerar como el órgano endocrino más importante del cuerpo si nos atenemos al número de hormonas que se sintetizan en é1. Las hormonas sexuales son las que más afectan al desarrollo y a la función cerebral. Durante el período prenatal las hormonas sexuales organizanla asignación del sexo y más tarde, tras la determinación del sexo gonadal, las hormonas circulantes producidas por la gónada provocarán la diferenciación de los genitales externos e internos. Después, las hormonas gonadales junto con las hormonas 50 Neuropsicología de la sexualidad Diferenciación sexual del ce¡ebro 5t del eje hipotálamo-hipófisis-gónada conducirán el proceso de Se refiere a la capacidad de las hormonas de influir en la ci- diferenciación de los caracteres sexuales secundarios en vafones y mujeres. Por lo tanto, durante el período prenatal las hormonas sexuales organizan la asignación de un sexo' y durante la pu- toarquitectura y estructura del cerebro de manera permanente durante el desarrollo, desde el período fetal hasta el final de la pubertad. Entre 1960 y 1970, gran número de trabajos permitieron definir el tiempo en el cual ocurre este proceso y en el que el SNC es más susceptible al efecto de las hormonas esteroides para organizar el sustrato y la función del mismo. Gracias a estos trabajos se dieron a conocer cuáles eran los esteroides más efectivos y la forma en que actúan las hormonas esteroides (Gorski, bertad, median la aparición de caracrerísticas secundarias y nos proporcionan la capacidad de reproducción. Al mismo tiempo, alrededor de la séptima semana de gestación, el cerebro, que es un importante órgano blanco para la acción de las hormonas, también sufre una diferenciación sexual, donde el resultado es la formación de encéfalos femeninos y masculinos con aspectos diferentes. La acción hormonal eierce sus efectos permanenres en el sNC en desarrollo y efectos transitorios en el adulto. De esta manera, se considera que la mayoría' de las diferencias dimórficas en los encéfalos surgen por las acciones hormonales reguladas por la expresión genética, específicamente Por los esteroides sexuales, provocando además respuestas endocrinas y comportamentales que son congruentes con el sexo genético y fenotípico. 3.1.2. Efecto dual En el inicio de los años cuarenta se obtuvieron las primeras pruebas del efecto ejercido por las hormonas sexuales sobre la diferenciación neural y se obtuvo que estas hormonas, en su condición de factores epigenéticos, influyen en la conducta mediante los procesos de plasticidad, y provocan dos efectos principales: organizador y activador. Lø acción organizad'ora de los esteroides sexuales tiene lugar durante una fase del desarrollo llamada período crltico y se caracterizapor ser irreversible (Fernández, 1996). 1985); por ejemplo, confirmaron que los andrógenos, por medio del estradiol, son responsables de controlar la conducta copulatoria propia del macho. En la literatura científica existen evidencias que demuestran que los esteroides sexuales durante este período, desempeñan un papel importante sobre la manifestación de diferencias sexuales en el comportamiento. Por ejemplo, se ha demostrado que tanto en los hombres como en las mujeres, los niveles altos de andrógenos durante el período crítico del desarrollo embrionario se asocian con la manifestación de la conducta masculina. Por otro lado, los resultados obtenidos en humanos permite inferir que el período crítico se extiende desde el cuarto al séptimo mes de vida intauterina (Abramovich, 1987). Este período comienza con el incremento de la secreción hormonal gonadal y concluye con la desaparición de la matriz celular alrededor del tercer ventrículo. Durante esta fase parecen coincidir un proceso de plasticidady maduración neural con un aumento de sensibilidad a los esteroides sexuales, incrementando sus receptores en diferentes estructuras neurales. El efecto actiuador, de carácter transitorio o reversible, se presenta a lo largo de la vida del individuo y se relaciona con la acti- Neuropsicologla de la sexualidad 52 Diferenciación sexual del ce¡ebro 53 vación de las células diana para facilitar y potenciar condl.lctas en - Estrógenos, producidos por los ovarios, por la placenta contextos específicos. Las hormonas activan respuestas endocrinas o comportamentales actuando sobre un cerebro previamen- durante el embarazo y en menor cantidad por la glándula suprarrenal. Derivan de los andrógenos, siendo el más importante el estradiol. - Progesterona, cuya fuente principal son los ovarios y la placenta, se desarrolla en la pubertad y adolescencia en el sexo femenino. lo A largo de estos últimos años, se ha acumulado una arnplia evidencia que muestre que los esteroides sexuales son importantes en funciones como la conducta afectiva y sexual. El conocimiento actual, que se ha obtenido principalmente de modelos experimentales de roedores o primates, demuestra que desde el período embrionario, los esteroides sexuales denen efectos mitógenos sobre el SNC en desarrollo. Estas hormonas influyen en la citoarquitectura del SNC actuando sobre las neuronas, las sinapsis y las células gliales y regulando la supervivencia, diferenciación y conectividad de grupos neuronales específicos. Tâmbién intervienen en la diferenciación funcional de las áreas cerebrales, induciendo: - Los procesos de crecimiento axonal y dendrítico de las neuronas dianas (modificando el volumen de diferentes estructuras y el número de ellas); - la diferenciación neuronal; - la formación de conexiones sinápticas; - la iniciación de la expresión de sus receptores específicos, muy esenciales en la vida adulta; - la organizacióny estabilidad de las sinapsis. Además, se ha demostrado que participan en la bioquímica cerebral regulando la expresión de prácticamente todos los neurotransmisores y neuromoduladores, y regulando efectos neuroprotectores. te organizado; el resultado es que un gran número de estas respuestas activadoras son dimórficas (Gorski, 1985). Los principales estudios experimentales con animales y humanos sobre el efecto activador de las hormonas en la conducta sexual han aportado diferentes hallazgos: - la conducta sexual del macho se basa en componentes - motivacionales y consumatorios; en mamíferos, la conducta sexual del macho es activada por la secreción de testosterona por las células de Leydig en el testículo; - El área preóptica del hipotálarno es el centro intregador que sirve de mediador de los efectos de la testosterona en la ejecución de la conducta sexual; - el complejo amigdaliano es un componente importante para organizar la conducta sexual masculina, mediante in- puts dopaminérgicos; - el sistema dopaminérgico se piensa que está involucrado en el componente motivacional de la conducta sexual. 3.1.2. Horrnonas sexuales Las hormonas sexuales son esteroides derivados del colesterol. La estructura básica es la mólecula de perhidrociclopentafenantereno, compuesta por tres anillos de seis carbonos y un anillo de cinco carbonos. Son producidos por las gónadas, la corteza suprarrenal y la placenta. Se pueden clasificar en tres tiPos: - Andrógenos, corresponden a la testosterona y son producidos por los testículos. Neuropsicologla de la sexualidad 54 3.1.3. Actuación de los esteroides sexuales sobre el Sistema neruioso centrãl Los esteroides sexuales, adem¿ís de ejercer efectos específicos sobre el sistema nervioso, sufren biotransformaciones. Parala diferenciación cerebral sexual es esencial que se dé un proceso denominado arumatización, en el cual tiene lugar la conversión de andrógenos (esteroides testiculares) a estrógenos (esteroides ováricos) para que se produzca la masculinización cerebral mediante laenzimaaromatasa P-450. Existen zonas claves para la expresión del dimorfismo biomorfológico y funcional, que coinciden con una gran actividad de la aromatasa. Se ha evidenciado una alta actividad de esta enzimaen áreas límbicas e hipotalámicas durante el período del desarrollo neural. Es un proceso muy intenso y amplio en el SNC en desarrollo, pero durante la vida adulta determinadas áreas pierden esta capacidad de transformar andrógenos en estrógenos. Estos organizan el sustrato neural induciendo cambios tanto bioquímica como estructuralmente' generando fenotipos neuronales tanto con relación a la capacidad secretora de las neuronas como a la sensibilidad respecto a estímulos eferentes. Los andrógenos que no son suscePdbles de aromatización o conversión en estrógenos, como la dihidrotestosterona' no parecen desempeñar un papel relevante en el proceso de la diferen- ciación sexual. Mecanismos de acción El mecanismo clásico de acción de las hormonas sexuales en el sistema nervioso es a través de la interacción con sus receptores intracelulares: los receptores de estrógeno alfa y beta, el receptor de andrógenos y el receptor de progesterona. Diferenciación sexual del cerebro 55 Los dos tipos de receptores para los estrógenos (alfa y beta) presentan estructuras muy similares y tienen una alta homología en la secuencia de aminoácidos entre ellos (Kuiper et al., 1996; Klinge, 2000). Las neuronas del cerebro en desarrollo presentan estos receptores, funcionalmente similares a los de los adultos, pero su concentración y distribución es más amplia que en el SNC adulto. Estos receptores están agrupados en distintas regiones de la cotteza cerebral, en el cerebro anterior, el hipotálamo y la amígdala, así como en las subdivisiones de estas regiones, sobre todo en aquellas partes o núcleo, que son necesarios para llevar a cabo la conducta reproductiva (Gutiérrez, 2005). La participación esencial que tienen los estrógenos en el proceso de la diferenciación sexual se ha comprobado por diversos autores con el uso de antiestrógenos como el tamoxifén, paradeterminar si este tiene efecto sobre el neurodesarrollo del SNC. Se ha demostrado que el tamoxifén se une a receptores estrogénicos intracelulares y evita que estos sean ocupados por el estradiol. Impide que se forme el complejo receptor-estrógeno, y de esta manera, en presencia del fármaco, los estrógenos no serán capaces de tener ninguna actividad transcripcional. Además de los receptores nucleares, se ha descrito también la existencia de receptores para esteroides en la membrana celular. Existen varios sitios de unión a la membrana que incluyen la progesterona, la testosterona y los estrógenos. Los efectos que desencadenan se han denominado no transcripcionales, ya que la unión del esteroide con su receptor no provoca una modificación directa en la expresión génica, sino que supone una respuesta celular más rápida y que parece involucrar la intervención de offos receptores de membrana ylo vias de señalización intracelular. Neuropsicología de la sexualidad 56 Por tanto, los efectos de un esteroide son de amplio rango y abarcan efectos genómicos y no genómicos: por un lado, se ligan a receptores nucleares actuando - 57 Diferenciación sexual del cerebro Figura3 Aspectos relacíonados con las diferencias dimórficas sexua/es como factores de transcripción para controlar la diferenciación sexual del cerebro y la activación de conductas se- Las diferencias dimórficas pueden ser de xuales especlficas; - por otro, se ha comprobado que son capaces de actuar so- bre la función cerebral más rápidamente que para lo que se postulan mecanismos no genómicos. En conclusión, el proceso de diferenciación sexual cerebral es el resultado de una interacción compleja de eventos que en su mayoríason motivo de intensa investigación. 3.2. EsrnucruRas cEREBRATES El macho y la hembra de la Bran màyoría de las especies son dimórficos, es decir, muestran diferencias sexuales cualitativas, cuantitativas o ambas en las estructuras del SNC (Guillamón Ec Segovia, 2007). Participan en el control de las funciones endocrinas y conductuales, las cuales son parte de los procesos reproductivos e incluyen patrones esenciales durante la conducta sexual. Como ya hemos visto,las diferencias sexuales en el SNC son el resultado de la secreción de hormonas por parte de las gónadas durante el período crltico (Gutiérrez eta1.,2005). Estas diferencias se refieren principalmente al tamaño del núcleo cerebral o del circuito cerebral. En cuanto al tamaño, se pueden deber a diferencias en el volumen o al área que el núcleo ocupe y no se evidencian hasta el período de la pubertad (figura 3). Esto implica que factores postnatales pueden tener un rol importante en el establecimiento de la naturaleza dimórfica en circuitos neurales que median conductas sexuales. Densidad Area Volumen Fuente: Boragarelli, 2007 Algunas de estas diferencias dimórficas correlacionan con sexo, preferencia sexual, o identidad de género. fuí pues, el cerebro presenta modificaciones específicas que guardan correlación con la identidad y orientación sexual (figura 4). Figura 4 Esquema que explica las diferencias dimorficas asociadas a la conducta sexual, orientación e identidad Las diferencias dimórficas tienen que ver con SEXO Preferencia sexual ldentidad de género Fuente: Boragarelli, 2007 Pero el dimorfismo sexual del cerebro se establece en redes y sistemas neuronales; ningún núcleo por sí solo da cuenta de la función de la red en su conjunto. Así pues, en general, se puede decir que la mayoría de las regiones sexualmente dimórficas del Neuropsicología de la sexualidad 58 cerebro pertenecen al sistema límbico. Se le considera un <sistemâ) ya que los circuitos o núcleos que lo componen están altamente conectados entre sí, de manera que la actividad eléctrica y química de un núcleo puede influir en la actividad de otro -nazonaabultada hacia abajo que se denomina eminencia media del tuber cinereum. Figura 5 Principales medios dimorficos del cerebro núcleo. Por otro lado, la mayoríade los circuitos dimórficos del cerebro son parte del hipotálamo o están íntimamente relacionados con él (Gutiérrez, 2005). Los principales núcleos dimórficos del cerebro son: ' Hipotálamo: - Á"." preóptica (APO). - Núcleo ventromedial (NVM)' - Núcleo supraquiasmático. - Amígdala. - Stria terminalis (estría terminal). . Otros: - Núcleo medular bulbocavernoso. - Núcleo arcuato. - Núcleo supraóptico. - Cuerpos mamilares. El hipotálamo corresponde al 0,3o/o del total del cerebro' Es la región ventral del diencéfalo que rodea a la cavidad del tercer ventrículo. El límite anterior del hipotálamo es un plano vertical que pasa por delante del quiasma óptico. El límite posterior está dado por un plano vertical que pasa inmediatamente por detrás de los cuerpos mamilares, y el límite lateral está representado por el plano imaginario o anteroposterior que pasa Por fuera de los pilares anteriores del fornix. Ventralmente está delimitado por una delgada pared en forma de embudo, el tuber cinereum, que se prolonga hacia abajo en el tallo hipofisiario y el proceso infundibular de la neurohipófisis. Entre el infundíbulo y el tallo hipofisiario existe una peque- 59 l)i ferenciación sexual del cerebro tüo Pl¡rû of tuoÀlrl Anlül0r tr¡ì åru Antèríor iogtfrrûlrú bûdv hypothdàm¡caær Ncl4{r3 5utræptL û¡Jdtùô Rodiu3 Por otro lado, posee la mayor concentración de recePtores de hormonas sexuales (andrógenos, estrógenos y progesterona) (Plaff E¿ Schwartz-Giblin, 19S8) y está constituido por células neuroendocrinas, representando el nexo entre el sistema nervioso central y el sistema endocrino. Las principales diferencias sexuales estrtlcturales entre los cerebros masculino y femenino se han encontrado, los últimos años, en el hipotálamo (tablas 1 y 2). Neuropsicología de la sexualidad 60 Tabla 1 Principales núcleos dimórficos del hipotálamo Área Nucleo Núcleo supra- preóptica ventromedial quiasmático La actividad Facilita la conducta sexual en las hembras. Presenta mayor densidad sináptica en los machos Difiere en su forma en el hombre y en la mujer de estos núcleos está aso- ciada con la iniciación del acto sexual, la erección y el período refractario en el varón y con la iniciación del acto sexual en la hembra Amígdala Estría terminal Es el centro de Es la principal las emociones. eferencia de la amígdala. Presenta una Cumple un importante rol en la conducta sexual (motivación-excitación). Presenta una importante importante diferencia de volumen entre el hombre y la mujer 6r Dilerenciación sexual del ce¡ebro Los núcleos de esta estructura se encuentran conectados con las rutas sensoriales límbico-hipotalámicas que modulan la conducta reproductora y, algunas de ellas, entre sistema llmbico e hipotálamo, permiten que, en la conducta sexual, intervengan impresiones olfativas, cuyas conexiones al hipotálamo están más desarrolladas en los organismos masculinos (Simerl¡ 2002). Figura 6 Vision tridi mension al I í mbico-hi pota I á m ico diferencia de tamaño entre el hombre y la muJer Fuente: Boragarelli, 2007 Tabla 2 Principales diferencias estructurales dimórficas entre hombre y mujer Hombres Mujeres Es 2,5 veces más grande NIHA 3 Tiene más células sensibles a los andrógenos que cualquier otra parte del cuerpo NIHA 1 Doble cantidad de células Núcleo ventromedial > Densidad sináptica Núcleo supraquiasmático Esférico Mitad del número de células Mitad de células Amígdala 85% > volumen Estría terminal Porción basal9T% más grande Área preóptica > Desarrollo dendrítico Fuente: Boragarelli, 2007 Alargado El hipotálamo desempeña el papel crítico de coordinar la expresión de la conducta reproductiva integrando respuestas fisiológicas y sugerencias ambientales. La relación fisiológica y anatómica con la glándula hipófisis es un medio efectivo para coordinar procesos homeostáticos a través de la regulación neuroendocrina de secreciones hormonales, controlando patrones de hambre, sed, sueño, temperatura y conducta sexual. Neuropsicologla de la sexualidad 62 con la rePero el hipotálamo también establece conexiones coordinar límbica, hipocampo y amlgdala' Por ello puede gión que rede la conducta sexual' La estructura del ärprr.rr". .r.trro.tdo"rinas con sugesdones sensoriales gr,rl"., las motivaciones neuroana- f,ipot,íI"rno es compleja' tanto desde el punto de vista en cuatro retómico como neuroquímico' Se puede subdividir a criterios anatómigiones y en más ¿, iS núcleos de acuerdo cos y funcionales. 3.2.t. Principales núcleos dimórfi'cos en el hipotá'larno ,4.rea pretiPtica (AP O) por Hace más de 30 años, varios laboratorios se interesaron se centrael dimorfismo sexual cerebral. Los primeros trabajos á''rea preópron en el continuum formado por el hipotálamo y el y el hìpotática. Por entonces' se suponía que el área preóptica de una lamo constituían la región neuroendocrina' tratándose de los andrógenos' Además' también región sensible l" " "..ió.,del área preóptica-hipotálamo afecta,."s"bía que las lesiones (Guillamón & Segoban a la expresión de la conducta sexual via,2007). la existenEn 1968 Gunter Dörner y su equipo demostraron neuronales. En concia de dimorfismo sexual en dichas regiones del núcleo hicrero descubrieron que en las hembras el volumen años después' potalámico ., rn.no, que el de los machos' unos in l973,Raisman y Field encontraron que las hembras presenmachos y en 1978' taban un área preóptica menor que la de los sexualmente Gorski y r.r.qìipo, describieron este núcleo como más dimórfico, hoy.r, día continúa siendo uno de los ejemplos laboratorio como en claros de dimorfismo' tanto en animales de Diferenciación sexual deI cerebro 63 humanos (Gorski et a1.,7978; Swaab E¿ Fliers, 1985). Desde entonces, el âreapreóptica se ha convertido en el modelo sobre el que se centró la investigación para estudiar los posibles mecanismos hormonales responsables de la diferenciación sexual del cerebro. Es considerada como el árrea donde es probable que resida el mecanismo de regulación diferencial de las caracterlsticas di- mórficas relacionadas con el sexo, incluyendo: - control de la conducta sexual; - mecanismo necesario para la ovulación (Gorski, 1980; Montano et al., 1995). Además, es un área densamente poblada de neuronas. La densidad neuronal en esta región es varias veces más grande que en otras regiones. Sobre esta base, en 1980 Gorski y colaboradores Ia definieron como el núcleo dimórfico sexual del área preóptica. Originalmente se le designó como el nnúcleo sexualmente dimórfico del área preóptico. El nombre de la región vatía' dependiendo de la especie: en ratas eI fuea se la conoce como nla porción medial del área preópticau, y en humanos, como uel núcleo intersticial del hipotálamo anterior-1u (NIFIA-1) (Allen et al., 1989). Los modelos animales han sido útiles para determinar qué conductas conffolan el ârea preóptica. En este tipo de experimentos la estrategia consiste en destruir el circuito neuronal de interés y luego observar la conducta del animal en comparación con animales intactos. Ya que teóricamente no existe una correlación lineal entre el número de neuronas de un circuito y su capacidad funcional, se desconoce si diferencias en el tamaño reflejan diferencias cualitativas o cuantitativas en términos de función neural (Byne et al., 2000). La estrategia experimental más utilizada para conocer la participación de un núcleo es la Neuropsicología de la sexualidad 64 lesión quirúrgica o química (Parra Gimez,2009) y de esta manera se ha demostrado que en las ratas macho, si se destruye el área preóptica, no expresen conducta sexual en presencia de una hembra; en las ratas hembras se considera el irea'preóptica como un circuito inhibidor de receptividad sexual (Pf"tr E¿ Schwartz- Giblin, 1988). Tfas varios años de estudio en ratas sobre el dimorfismo sexual a nivel morfológico, las principales evidencias son: - el tamaño del núcleo dimórfico sexual del área preópti- ca es seis veces mayor en machos que en hembras (Gors- ki, 1g7g); - el número y el tamaño de las neuronas en todo el hipo- tálamo es mayor en el macho (Arnold 8¿ Gorski, 1984); - existe un mayor desarrollo dendrítico del área preóptica en el macho (Greenough et al., 1977); - el número y tipo de sinapsis dendríticas del ártea preóptica del hipotálamo es menor en la hembra (Raisman 8c Field,1973). Tâmbién se ha determinado el rol del área preóptica en conductas sexuales de primates. En macacos machos, el AÌO está asociado a la iniciación del acto sexual, erección y el perlodo re- frectorio después del acto sexual. En las hembras, el APO está asociado con la iniciación del acto sexual (Oomura et al., 1989). Cuando se estudia el área preóptica del cerebro humano, el cuadro es mucho más complejo que en roedores' Además del núcleo que previamente hemos mencionado, se encuentran otros tres núcleos en la región medial del APO. Se ha descrito NIHA-I al4 como regiones dimórficas. Sin embargo, debido a la proximidad anatómica de estos grupos de neuronas, se debate si NIFIA-l-4 representan grupos neuronales anatómica y funcionalmente independientes unos de otros. Diferenciación sexual del ce¡ebro 65 El núcleo INAH-I fue descrito a mediados de la década de los ochenta por Swaab y Fliers (1985). Presentaron los primeros resultados con relación a la presencia de un grupo de células en el hipotfamo anterior de humanos, que es mayor en varones que en mujeres. Sobre la base de su localización y dimorfismo sexual, sugieren que es un grupo de células comparable con el área preóptica de la rata. Las diferencias estructurales se van modificando con la edad. Los cambios parecen deberse tanto a la edad como a modificaciones en los niveles de las hormonas sexuales (Pátrrez Gámez, 2009). Estos autores demuestran que tanto niños como niñas poseen el mismo número de células al nacer y que este número aumenta exponencialmente hasta los 4 o 5 años (forge-Rivera, I 998). Posteriormente, el dimorfismo sexual de este núcleo se establece postnatalmente entre los 4 y l0 años de edad y su tamaño, para un sexo dado, no varía con la orientación sexual. La cantidad de células permanece constante hasta la pubertad, pero en el caso de las niñas un alto porcentaje de las células muere (Swaab et al., 1988), lo que hace que el dimorfismo sexual no sea aparente hasta la pubertad. El dimorfismo se expresa con más magnitud en adultos jóvenes y el número de células de este núcleo declina en los varones hacia los 50 años, mientras que en las mujeres, a partir de los 70 años, tiene lugar una involución mucho más gradual. Asl, se podría considerar que hay estadios a lo largo de la vida en los cuales este núcleo es monomórfico (niños), otros en los que es modestamente dimórfico (60-80 años), y otros estadios (adultos jóvenes y ancianos mayores de 80 años) en los que el núcleo expresa el dimorfismo máximo, siendo dos veces mayor en varones que en mujeres (Swaab et al., 1992). Los núcleos INAH 2 y 4 no muestran dimorfismo sexual. Levay (1991) confirmó el dimorfismo sexual en el NIHA-3, siendo de mayor tamaño en el 66 Neuropsicología de la sexualidad varón que en la mujer. Tämbién confirmó que existía dimorfismo con relación a la orientación sexual. Por otro lado, los astrocitos también exhiben dimorfismo sexual ya que son muy sensibles a la acción de esteroides gonadales perinatales. Estudios en ratas muestran diferencias gliales importantes, sobre todo del área preóptica, con prolongaciones más grandes y numerosas en el macho que en la hembra. Se observó que la relación entre los astrocitos y la morfología neuronal durante el desarrollo es distinta en diferentes regiones del cerebro, y que esto sirve de base para la diferenciación sexual de determinadas áreas cerebrales (McCarthy et il.,2003) Utilizando marcadores para las proteínas fibrilares, se estudió la morfología de los astrocitos del APO de ratas machos y hembras en el día del nacimiento y en el tercer día, y se vio que ya desde el nacimiento los astrocitos del área preóptica mostraban diferencias significadvas entre los machos y las hembras: los machos tenían prolongaciones más numerosas y más largas, y se comprobó que estas diferencias se mantenían en el examen practicado al tercer día (Bocchino, 2006). Núcleo uentromedial (NVM) El caso de dimorfismo sexual en el NVM es interesante, ya que este no se debe a diferencias en cuanto al tamaño o densidad del núcleo, sino a la complejidad estructural de las neuronas (Gutiérrez, 2005). Este núcleo regula y facilita la conducta se- xual de las hembras (Pfaff& Sakuma, 1979). En ratas, las hembras expresan receptividad sexual por medio de una respuesta reflqa que se conoce como lordosis. Esta conducta de receptividad sexual se ve mediada por altos niveles de estrógenos y progesterona; de lo contrario, la hembra rechaza los acercamientos sexuales del macho. Diferenciación sexual del cerebro 67 En los machos la densidad sináptica en este núcleo es mayor que en las hembras (Miller E¿ Aoki, l99l), y el número de contactos sinápticos en la hemb ravaría a través del ciclo estral, existiendo un mayor número de sinapsis y de espinas dendríticas (Cohen E¿ Pfaf[ 1992). En estudios realizados en neuronas pertenecientes al núcleo ventromedial del hipot:íIamo, se observó que estas respondieron a la estimulación con estrógenos, mostrando un aumento significativo en todos los componentes celulares, núcleo, nucleolo y soma celular. Tämbién se constató un incremento notable en la cantidad de retículo endoplásmico y en el número de polirribosomas, indicadores de una aceleración en el metabolismo celular y particularmente en la síntesis de proteínas (Carrer & Aoki, 1982). Las hormonas esteroides parecen participar en el crecimiento neurítico y en la sinaptogénesis de las regiones cerebrales que muestran receptores específicos para estas hormonas, sobre todo en la etapa prenatal, aunque también se ha referido esta actividad remodeladora de la sinapsis en algunas regiones del cerebro adulto, en especial en el núcleo ventromedial del hipotálamo. Por lo tanto, el dimorfismo del NVM está claramente controlado por las hormonas sexuales, y en el caso de las hembras es regulado a lo largo de los ciclos hormonales (Jorge-Rivera, 1998). Núc leo supraquiasmático (NS Q) El NSQ regula los ritmos cíclicos o ritmos circadianos del cuerpo. Este núcleo genera y coordina ritmos hormonales, fisiológicos y conductuales con una periodicidad de 24 horas. El NSQdel hipotálamo es otra de las estructuras cuyo tamaño varía con relación a la orientación sexual (Swaab 8¿ Hofman, 1990). En humanos, al nacimiento, el NSQ contiene solo un 1320o/o de las células que se encuentran en el adulto. Se produce un Neuropsicología de la sexualidad 68 desarrollo r:þido postnatal, alcanzando un máximo aI año y medio de edad (Swaab et aJ.., 1992). Por otro lado, se ha encontrado que el NSQ es morfológisunente diferente en hombres y mujeres; esferico en hombres y aJaryado en mujeres (Swaab et al., 1985). El impacto funcional de este tipo de dimorfismo no se ha determinado. Se ha sugerido que la forma del circuito neuronal pueda depender del tipo de conexiones, aferentes o eferentes, que este establezca con otros circuitos. Además, se ha demostrado que existe una diferencia clara en el volumen y número de células del NSQentre hombres heterosexuales y hombres homosexuales; así, Swaab y Hoffrnan (1990) encontraron que tanto el volumen como el número de neuronas es el doble en el NSQde homosexuales que en el de heterosexuales. En el caso de animales de laboratorio se ha establecido que el NSQ también está asociado con conductas sexuales. Se ha determinado que la amplitud de los ritmos circadianos en la conducta 69 l)iFerenciación sexual del cerebro Núcleo de la estría terminal (ST) La estría terminal es la conexión eferente principal de la amígdala (\Weller E¿ Smith, 1982). Por su ubicación y conexiones, actúa como puente entre el sistema límbico y el hipotálamo. Desempeña un papel importante en el comportamiento sexual masculino y en la regulación de descarga de gonadotropinas (Zhou et al., 7995). La ST es una estructura de materia gris formada por cuatro partes principales: - división medial; - división lateral; - división ventral; - división intermedia. Figura 7 Localización tridimensional de la estría terminal sexual es mayor en ratas tratadas con antiestrógenos durante el pe- ríodo neonatal. De igual modo, si se administran antiestrógenos durante el período prenatal y postnatal, se encuentra un aumento en un 59o/o de las neuronas que contienen vasopresina en el NSQy conducta bisexual en ratâs. Si el tratamiento con antiestrógenos se limita al periodo prenatal no se observan estos cambios. En estos momentos no se ha encontrado evidencia que demuestre el rol del NSQ en el establecimiento de la preferencia sexual, a pesar de que el dimorfismo es claro entre hombres con diferente orientación sexual. Al igual que en el área preóptica, el dimorfismo del NSQ se observa enúe los 4 y 12 años de edad. Este resultado implica que el dimorfismo del NSQ está bajo el control de las hormonas sexuales, y que factores posnatales contribuyen de manera importante en este dimorfismo, aI igual que en el área preóptica. Fuente: http://www.wikipedia.com 70 Neuropsicología de la sexualidad A principio de los noventa se describió el dimorfismo sexual en el.núcleo de la estría terminal de la rata y se demostró que la lesión en la división medial posterior de este núcleo producía déficit copulatorio en el macho (Gutiérrez et a1.,2005). Allen y Gorski (1990) describieron que la porción posteromedial del núcleo es2,5 veces mayor en varones que en mujeres, lo que no se debe a una diferencia de talla del cerebro en ambos sexos, ya que, mientras el peso cerebral en varones eta el l3o/o mayor que el de las mujeres, el volumen de esta porción del núcleo era un |47o/o mayor en varones que en mujeres. Tämbién se observó que la base de la estría terminal (BNST) es 97o/o mas grande en volumen en hombres que en mujeres (Hines et al., 1992). La implicación de este núcleo en conductas sexuales y reProductoras en humanos, deriva de los datos experimentales que demuestran su relevancia en el comportamiento sexual de los roedores (Fernández, 1996). En ratas, este núcleo (BNST) realiza una destacada función en el control de la conducta y de los aspectos reproductivos. La estimulación de este núcleo en algún momento de la etapa estrogénica adelanta el pico de la secreción de la hormona luteinizante en las hembras. Asimismo, la lesión de este núcleo en ratas, hace que estas muestren alteraciones en aspectos del repertorio sexual (Claro et al., 1995).En otro estudio se demostró que si se provocaban lesiones, las ratas no lograban tener estimulación sexual (Liu et aJ., 1997). En humanos, se desconoce qué papel desempeña el BNST en conductas sexuales. Sin embargo, se ha encontrado dimorfismo en la subdivisión central de este núcleo, que está de acuerdo con las diferencias sexuales previamente descritas en una parte más caudal del BST, indicando que este núcleo puede estar involucrado en las funciones humanas sexuales o reproductivas Diferenciación sexual del cerebro 71 (Allen E¿ Gorski, 1995). Además de las conexiones con el sistema límbico y el hipotálamo, este núcleo posee receptores parâ estrógenos y andrógenos, y constituye un lugar de intensa actividad aromàtizarrte en el cerebro de la rata en desarrollo (Abril etal,.,1987). Mediante la utilización de métodos histoquímicos, también se ha demostrado una distribución dimórfica en dicho núcleo de diferentes neuropéptidos, como puede ser el caso de la vaso presina. Núcleo parauennicular (NPV) Se ubica por debajo del tercer ventrículo, sobre el quiasma y las cintillas ópticas. Formado por neuronas grandes y redondas, muy similares a las halladas en el núcleo supraóptico. Consta de varios grupos celulares, uno medial o parvocelular y uno lateral o magnocelular. Estas células producen sustancias neurohormonales como la oxitocina y la vasopresina, muy involucradas en el establecimiento de lazos afectivos entre hombre y mujer. Estas sustancias son enviadas a través de sus axones hacia la neurohipófisis. Estas células también envían proyecciones al ffonco encefálico y a la médula espinal, siendo más grande en hembras que en machos (Raisman et aI., 1973). Amígdala De todas las estructuras subcorticales, la amlgdala es la que se ha relacionado de un modo más consistente con la emoción, tanto en animales como en humanos, y es considerada como el centro de las emociones, regulando las reacciones de agresividad, miedo y ansiedad. El complejo amigdalino se localiza en la pro- fundidad de los lóbulos temporales, formando el sistema límbico. Está integrado por una serie de núcleos con distintos atribu- 7) Neuropsicología de Ia sexualidad tos funcionales y su papel pfincipal es dar signifìcado emocional a los estímulos que recibe. Se subdivide en tres subregiones por criterios anatómicos y funcionales (Amunts et al., 2005)' Grupo laterobasal (LB): controla las respuestas motoras ante un esdmulo que provoca miedo, y procesa la memoria emocional, incluyendo los núcleos: lateral, basolateral, basomedial y paralaminar. Grupo superficial (SF): rcc1be información del sistema olfatorio e involucra el procesamiento afectivo' Este grupo incluye el áreaanterior amigdaloidea, el área amigdalapiriforme, el âtea amígdala hipocampal y el núcleo cortical central y medial. Diferenciación sexual del cerebro 73 otras señales sexuales como a la capacidad de percibir a Ia rata hembra, implicando a esta estructura en el apetito sexual (Kondo et al., 1997).Por otro lado, en ratas se ha observado que la porción anterior de la amígdala medial aumenta la receptividad en ratas hembras, mientras que la porción posterior del núcleo lateral reduce la receptividad (Masco 8¿ Carrer, 1980). Figura 8 Sistema límbico Grupo centromedial (CM): conüola las respuestas endocrinas y modula las respuestas emocionales. Dentro de este grupo se incluye el núcleo medial que ha sido estudiado en animales y se ha demostrado que recibe inputs desde el sistema olfato- rio, el cual es esencial para la conducta de la reproducción sexual (Amunts et al., 2005). Este grupo está formado por el núcleo central y medial. Desde hace tiempo se sabe que la amígdala también desempeña un papel importante en la conducta sexual. Esto se ha demostrado a partir de los resultados obtenidos en estudios en animales y en humanos, como la evidencia clínica que relaciona convulsiones en el lóbulo temporal cerca de la amígdala con la conducta sexual (Leutmezer et al., 1999). Los estudios con ratas, a las cuales se les lesionaba la amíg' dala, han puesto de manifiesto una distinción entre el rol de la amlgdala en el macho en cuanto a la conducta y apetito sexual. El apetito sexual en las ratas involucra la motivación para obte- T.mporal lobe Fuente: http:/iwww brainconnection.com En humanos, la amígdala se diferencia entre hombres y mu- ner recompensa sexual (deseo o querer), mientras que la conducta sexual de consumación incluye una recompensa (copulación). jeres en función del desarrollo del cerebro y en términos estruc- Everitt (1990) estudió que las lesiones en la amlgdala medial afectan a la capacidad del sistema olfativo en la rata macho y a diferencia sexual del tamaño de la amígdala se desconocen, las diferencias estructurales en la anatomía del cerebro a menudo se turales. Aunque las consecuencias que se producen, respecto a la 74 Neuropsicologla de la sexualidad asocian a diferencias en la respuesta y en la función del cerebro (Hamann, 2005). Esto se ha podido observar en estudios de resonancia magnética funcional (fMzu). Una de las mayores diferencias enüe los sexos hace referencia al tamaño de la amígdala. Se demostró que la amígdala medial es 85o/o mayor en volumen en los hombres que en las mujeres, te- niendo en cuenta también el tamaño total del cerebro (Golstein et al., 2001). Por otro lado, los estudios de neuroimagen han encontrado que la amígdala se desarrolla estructuralmente a diferente velocidad en hombres y mujeres. Otra diferencia estructural en áreas que reciben conexiones neuronales por parte de la amígdala, como por ejemplo el hipotálamo el cual es más grande en hombres que en mujeres, puede contribuir a la diferencia cerebral entre sexos que involucran a la amígdala. Los métodos para evaluar el contorno de la forma y tamaño de la amígdala fueron desarrollados y aplicados con éxito para detectar pequeños cambios en la amígdala de pacientes con trastornos psiquiátricos o neurológicos. Actualmente, para el análisis de la forma y tamaño de la amígdala se ha desarrollado un nuevo método que mide el radio medio de cada subregión. Este método emplea un mapa estadístico de las subregiones de la amlgdala para calcular la probabilidad de que aparezcacadapar te de la amlgdala (Amunts et al., 2005) Tämbién es importante el nivel de circulación de las hor- monas sexuales en sangre, yâ que constituye una influencia en la respuesta de la amígdala a través de la acción de los receptores (Hamann, 2005).Las regiones del cerebro que difieren en tamaño entre varones y mujeres, también tienden a que esas regiones contengan grandes concentraciones de receptores de hormonas sexuales (Goldstein et al., 2001) estrógenos y andrógenos, lo cual sugiere que las hormonas en ambos sexos desempeñan un papel fundamental para determinar el tamaño de la estructura Dife¡enciación sexual del cerebro 75 durante el desarrollo cerebral (Parra Gimez,2009). Esto se puede observar en el núcleo cortical, perteneciente al núcleo superficial de la amígdala de las ratas, mientras que en la amígdala de los humanos, la gran expresión de receptores para estrógenos y andrógenos, se observó principalmente en el núcleo cortical y en el áreaamígdala-hipocampal (Österlund et al., 2000). Por otro lado, en estudios con resonancia magnética, se ha podido demostrar que la amígdala medial y el hipotálamo se encuentran más activados en los hombres que en las mujeres ante estímulos sexuales (Hamann et aJ.,2004). Se ha concluido que la amígdala media en la diferencia de género en cuanto al apetito sexual y que la amígdala humana puede también influir en el papel de los estímulos sexuales en el comportamiento sexual masculino, de modo paralelo a los resultados previos en animales (Symos, 1979). Finalmente, sería necesario que en futuros trabajos se explicase la relación que existe entre los diferentes aspectos: estructura, desarrollo y función de la amígdala. 3.2.2. Otros Núcleo medular bulbocauernoso (NMB) Se trata de un grupo de motoneuronas que inervan los músculos bulbocavernosos y otros músculos perineales en varones y mujeres. En los varones, el músculo bulbocavernoso está más desarrollado que en las mujeres, situándose alrededor de la base del pene y contribuyendo a la eyaculación (Fernández, 1996). El tamaño del núcleo es mayor en los machos de diferentes especies animales y asimismo en el varón, siendo así que este Neuropsicología de la sexualidad 76 mayor tamaño se debe a un incremento en el número de motoneuronas. Parece probable que, como ocurre en animales de experimentación, en el varón los andrógenos prenatales induzcan la diferencia del NMB: el período embrionario durante el cual se produce degeneración de las motoneuronas se completa hacia la semana 26 de la gestación en humanos, y este período se solapa con la producción de testosterona por los testículos fetales y con la diferenciación de los genitales externos (Forger & Breedlove, 1986). En las ratas de ambos sexos, las motoneuronas establecen conexiones con el músculo bulbocavernoso en períodos tempranos del desarrollo y ambos presentan en el músculo receptores a andrógenos. En machos, la testosterona se une a dichos receptores, activando el músculo y liberando factores tróficos, los cuales son recogidos por las motoneuronas. En cambio, en las hembras no se produce esta unión a los receptores de andrógenos, por lo que hace que no se active dicho músculo, no se liberen factores tróficos y el NMB muera. Núcleo ãrcuato En las ratas hembra adultas se ha podido comprobar que la astroglia participa en la remodelación fásica de las sinapsis que tiene lugar durante el ciclo menstrual en el núcleo arcuato del hipot:ílamo, bajo la influencia del estradiol, teniendo un importante papel en la génesis de diferencias sexuales en las sinapsis (García Segura et aI., 1995). El patrón sináptico de algunos núcleos hipotalámicos como el arcuato, representa uno de los mayores exponentes de dimorfismo sexual cerebral adulto. Se ha observado que en el núcleo arcuato los machos tienen el doble de axones y la mitad de espinas dendríticas que las hembras (Mong et al., 1999). Diferenciación sexual del cerebro 77 Núcleo supraóptico (NSO) Este núcleo se encarga de producir vasopresina (Parra-Gá- me2,2009). Aunque el número de neuronas que producen vasopresina no difiere entre ambos sexos, sí que existen diferencias en cuanto a los niveles producidos (Swab et al., 2001). Existe una mayor actividad en las neuronas del NSO en hombres jóvenes comparada con la de mujeres (Share et al., 1988; Van Londen et al., 1997). El progresivo aumento en el tamaño de las neuronas del NSO parece estar correlacionado con el incremento del peso corporal; así los hombres con cuerpos más pesados requieren mayor canti- dad de hormonas que aquellos con cuerpos más pequeños (Swaab et al., 2001). A lo largo de la vida, la actividad neuronal en NSO se incrementa gradualmente en mujeres, mientras que en hombres permanece estable. Este es un ejemplo del sistema hipotalámico que no presenta diferencias estructurales sino puramente funcionales (Byne et al., 2000). Cuerpos mamilares (CMB) Otro ejemplo de dimorfismo sexual, basado en el efecto activador de las hormonas sexuales, fue encontrado en los cuerpos mamilares (CMB), que muestran receptores para andrógenos con mayor avidez por los mismos en hombres que en mujeres (Fernández-Guasti, 2000). La estimulación eléctrica de esta área en monos induce la erección. En estudios recientes se encontró que esta diferencia se debe por completo a la cantidad de andrógenos circulantes en la edad madura. Estos datos apoyan la idea de que el dimorfismo se relaciona con los niveles circulantes de hormonas sexuales (IGuijver et al., 2001). 78 Neuropsicologla de la sexualidad En resurnen, las causas que producen el dimorfismo sexual son múldples, interaccionando la genética, las hormonas y el ambiente, lo cual da como resultado las diferencias morfológicas histológicas, bioquímicas y fisiológicas cerebrales. Pero se consi- ry Neurotransmisores dera que las acciones hormonales reguladas por Ia genética, específicamente los esteroides sexuales, son los mayores responsables de las diferencias morfológicas sexuales en el cerebro, actuando de manera organizadora y activadora durante el período crltico. Las diferencias morfológicas se refieren principalmente al tama- ño del núcleo o circuito cerebral, localizandose principalmente en el hipotálamo y en el sistema con el que establece conexiones, el sistema límbico. Desde los años sesenta se han estudiado, principalmente en modelos animales, estas estructuras sexuales, considerándose que varios núcleos intervienen en el control de la conducta sexual, siendo el ireapreóptica el núcleo mejor estudiado y definido. Estas áreas cerebrales liberan sustancias químicas, neurotransmisores, para regular la función sexual en la que interviene cada grupo de neuronas que pertenecen a estos núcleos dimórficos. Desde el punto de vista biológico, existen reacciones químicas que subyacen y motivan las sensaciones que se desencadenan en nuestro organismo durante la respuesta sexual humana. En la cascada de reacciones que ocasionan las emociones se producen descargas de pequeño voltaje en las neuronas para comunicarse entre ellas, conectarse unos sistemas con otros y así coordinar las respuestas a estlmulos. A Iavez,las hormonas secretadas por las glándulas, viajan a través de la sangre para participar en la co- municación entre los órganos y las células. Las hormonas gonadales activan la conducta sexual mediante un efecto local en las estructuras del sistema nervioso. Esta acción se realiza como parte del funcionamiento normal de las neuronas y modula su interacción. Esto supone la interacción de las hormonas gonadales y los neurotransmisores que actúan como mediadores químicos en la comunicación entre neuronas (Gutiérre2,2005). La cascada de neurotransmisores se inicia en el cerebro en la fase del deseo y en el SNC, periferico y en los genitales durante la fase de excitación y orgasmo (Acuña, 2008). En 1905, \X/illiam H"tdy propuso el término nhormonan que significa uexcitar una actividadr. Con el progreso de la Neu- Neuropsicologla de la sexualidad 80 rología y de la Endocrinologla, aunadas en la denominada Neuroendocrinología, el descubrimiento de nuevos componentes bioquímicos que esrimulan diversos órganos y funciones nació el término (neurotransmisor> (NT). En esta revisión se describirân cadauno de los neurotransmisores que intervienen en la regulación de la respuesta sexual hu- mana (RSH). Se hará referencia a algunas biomoléculas y compuestos químicos que abundan en el organismo y que intervienen ã^ h, .-o.ione, y sensaciones involucradas en la sexualidad. Neu¡otransmisores 8l En NO penetra en la célula muscular lisa y activa una enzima llamada guanilato ciclasa. Esta enzima convierte la molécula guanosln trifosfato (GTP) en guanosln monofosfato clclico (GMPc) (Hedlund, 2001), produciéndose la relajación del músculo liso, permitiendo la entrada de sangre dentro del pene y, por lo tanto, dando lugar a la erección. Al disminuir el nivel de GMPc dentro de la célula, el músculo liso se contrae y se produce la erección. En la mujer, el NO desempeña un papel similar al del hombre. Se encuentra tanto en el SNC, SNAy en los órganos sexuales femeninos, especialmente en la vagina y el clítoris, esta última Oxido nítrico (NO) Anteriormente se reconocla al NO como un gas tóxico, pero recientes investigaciones han descubierto que ejerce efecto de neurotransmisor. Se le considera como el motor de todo el proceso supra e infraespinal que interviene en la respuesta sexual humana (RSH), regulando el proceso de los neurotransmisores tanto en el sNC, periferico y a nivel genital, en consonancia con la dopamina (Do), oxitocina (O") y testosterona (T)' Se ha evidenciado que es un importante neurotransmisor de neufonas no adrenérgicas-no colinérgicas (NANC), las cuales inervan el músculo liso en varios tejidos, enrre ellos, el del ffacto genitourinario (Moncada et al., 1991) y producen la respuesia eréctil en el hombre. Para que pueda inciarse la erección, los estímulos recibidos del cerebro, especlficamenre, desde el núcleo paraventricular del hipotálamo, son integrados en la médul" .rpittrl desde donde se envían señales al pene a través de los nervios parasimpáticos. Estos liberan acetilcolina, que actúa sobre los vasos sanguíneos del pene y sobre células nerviosas de los cuefpos cavernosos panlaproducción de No. El adecuado ni',r.l á. T es indispensable para manrener la disponibilidad del NO en el cuerpo cavernoso. responsable de la erección femenina. Las células nerviosas localizadas en el clítoris liberan NO, produciéndose el proceso simi- lar de la erección en el varón. Testosterona (T) La testosterona se ha demostrado que tanto en el hombre como en la muje¡ desempeña una acción central así como perlférica en el curso de la respuesta sexual humana, controlando las funciones sexuales como la libido, la potencia y la fertilidad. La testosterona y los andrógenos se encuentran en altas concentraciones en el hipotálamo y el fuea preóptica media (Rako, 1996), con concentraciones más altas que el estradiol; se debe tener en cuenta que estas á¡eas también corresponden a la actividad de la aromatasa, de manera que es probable que en esas regiones se produzca laaromatización, convirtiéndose la testosterona en estradiol. Como ya hemos visto, los andrógenos no solo controlan la homeostasis hormonal y el desarrollo de la función reproductiva sino que también son los precursores inmediatos de la biosíntesis de los estrógenos, tienen acción en el deseo sexual, los esta- dos de ¿ínimo y humor y la distribución del contenido adiposo. Neuropsicología de la sexualìdad Neurotransmisores Los efectos androgénicos de la testosterona se relacionan también con los genitales, su crecimiento y nutrición, la parte reproductiva y el desarrollo de los caracteres sexuales secundarios. El adecuado nivel de testosterona facilita la acción del NO en los cuerpos cavernosos (Penson, 1996). Como todas las hor- Estnigenos 82 monas esteroides, la testosterona estimula la síntesis de proteínas específicas al cruzar la membrana celular y unirse con un receptor en el núcleo, activando genes específicos. La testosterona fa- cilita la acción de la dopamina, inhibe la acción de la serotonina (S.) y de la prolactica (Prol) e inhibe también el sistema monoamino oxidasa (MAO), que se encarga de la oxidación de las monoaminas, muy importantes en el control de la función sexual (Giuliano & Rampin, 2000). La deprivación androgénica causa una reducción significativa del músculo liso trabecular, con aumento del te.iido conectivo y pérdidade la función erectiva, aunque en los animales casüados se recobra con la administración de testosterona (Tiaish et al., 1999). El papel de la testosterona en la mujer ha sido motivo de estudios recientes; Kaplan en 7979 difundió su participación en el deseo sexual femenino y se ha confirmado que también en la mujer la acción de la testosterona es tanto central como pertférica (Bixo, 1995). 83 La evidencia en modelos animales y humanos indica que los estrógenos modulan el flujo sanguíneo vaginal al regular la acti- vidad neuronal y endotelial, el papel nutritivo sobre la vagina, la preservación de la elasticidad y demás carâcterísticas vitales del órgano. Se han encontrado receptores estrogénicos en la células endoteliales y en el músculo liso de los cuerpos cavernosos humanos (Reiter, 2001). Por otro lado, BrayyYork(L997) afirman que los hombres producen estradiol y corticoesteroides de manera constante a lo largo de la vida. Monoaminas La dopamina, noradrenalinay serotonina son monoaminas, grupo de neurotransmisores más importantes en el sistema nervioso, implicadas en el control de la conducta sexual de ambos sexos. El efecto de cada amina es aumentar o inhibir, dependiendo del tipo de receptor, de cada conducta y del medio endocrino. a) Dopamina D e h idro ep iandros tero na (D H EA) Es la catecolamina más importante. Las neuronas que responden a este transmisor se denominan dopaminérgicas y la Se produce en la glándula suprarrenal sin depender del eje mayoría de ellas se encuentran en la sustancia negra. Además hipotálamo-hipófisis, considerándole como el precursor de las hormonas sexuales, andrógenos y estrógenos. Los m¿íximos valores de este andrógeno se alcanzan entre los 20 y 30 años de edad, y posteriormente decrecen de manera progresiva: un 25o/o por década en los hombres y tn 44o/o en las muje- se localizan en la eminencia media del hipot:ílamo, influyendo res (Saad, 2004). en la regulación de liberación de prolactina y hormona del cre- cimiento. Modula la expresión de la conducta sexual actuando principalmente sobre el receptor D2. En modelos animales se ha demostrado que el aumento de las señales dopaminérgicas influye 84 Neuropsicología de la sexualidad en el incremento de la conducta sexual. En estudios humanos, el efecto en varones es estimulador mientras que en mujeres el efecto es dual, estimulando e inhibiendo, involucrándose en la conducta motora y el proceso motivacional. El núcleo accumbens recibe aferencias de las células dopaminérgicas conteniendo altas concentraciones de dopamina. Este .rú.leo está implicado en el proceso de la recompensa y se ha demostrado que es importante en la regulación del refuerzo en la conducta sexual (Robins Ec Everit, 1996). Así pues, se puede afirmar, que esta área particiPa en la regulación de la dopamina en .l d.r.o y motivación de la conducta sexual y puede que sea una explicación por la cual la dopamina afecta a la motivación sexuai probablemenre acruando en mayor medida sobre el núcleo paraventricular (NP\0 del hipotálamo: en los receptores Dl, estimulando la erección, y en el receptor D2, estimulando el proceso de la emisión seminal. Otras áreas del cerebro también contienen dopamina y Pueden desempeñar un rol importante en el control de la conducta sexual. Por ejemplo, el núcleo estriado dorsal, implicado en el control motor y en la coordinación sensomotora durante el 85 Neurotransmiso¡es b) Noradrenalina La noradrenalina es un neurotransmisor que regula el estado de aleray vigilia, el nivel de conciencia y el sueño, el apetito y la conducta sexual. Se localizan en el locus ceruleus y el irea tegmental ventral y desde estas zonas envían proyecciones al: tálamo, amígadala, hipocampo, hipotálamo y neocórtex. Con la edad disminuye la actividad noradrenérgicay la MAO, la enzima que metaboliza la norepinefrina. Actúa sobre dos clases de receptores: alfa (1 y 2) y beta (1, 2y 3) y diferentes estudios farmacológicos revelan que la noradrenalina provoca distintas respuestas, según el receptor activado. La noradrenalina tiene un efecto estimulador, actuando sobre el receptor alfa I involucrándose en los mecanismos de atención y procesamiento sensorial durante el acto sexual. Este proceso modula el efecto de los estímulos entrantes que son relevantes para incluir una respuesta sexual. Por otro lado, la activación del receptor aJfa2 inhibe la conducta sexual en varones pero no en mujeres. En cambio, a nivel medular el recepto r alfa 2 regula el proceso de la erección. Por último, este neurotransmisor figura entre los neuropéptidos que se encuentran durante el orgasmo en la sangre periferica. Se ha podido observar que durante el orgasmo aumenta la NE, así como la prolactina y la testosterona (Exton, 2000). acto sexual. El efecto de la dopamina en la conducta sexual femenina parece estar mediada por el receptor D2 enel núcleo paraventricular y en el hipotálamo ventromedial por los receptores de progesterona y D5, un suptipo de los receptores D 1. c) Serotonina (St) La serotonina o 5-hidroxitriptamina (5-HT) se sintetiza fundamentalmente en los núcleos del rafe del tronco del encéfalo, desde donde se envlan proyecciones sertoninérgicas a diversas regiones del SNC y de la médula espinal, regulando el humor, el apetito, el sueño, la sed y la conducta sexual. Neuropsicologla de la sexualidad 86 Hay siete subtipos de receptores serotoninérgicos (5HTl, 5HT2, 5HT3, 5HT4, 5HT5, 5lHT6 y 5HT7), que a su vez se dividen en diecisiete. Se considera que el proceso serotoninérgico tiene un efecto inhibidor en la conducta sexual, aunque no toda acción serotoninérgica es inhibidora ya que presenta acciones facilitadoras de procesos dependiendo de la acción de la amina en los diferentes subtipos de receptores, localizados en diversos sitios del SNC y también a nivel medular. Los receptores emplean distintos sis- 87 Neurotransmisores drotestosterona que es el principal metabolito que produce los efectos testosterónicos en los centros cerebrales (Emori, 1991). La serotoninay la prolactina tienen un papel negativo en la respuesta sexual. Sin embargo, se citan a fin de comprender me- jor el balance positivo y negativo entre varios de los principales neurotransmisores. Se trata de dos neurotransmisores que pueden obstaculizarlaRSH por tanto, no intervienen en el proce- ¡ so fisiológico de la respuesta sexual. Sin embargo, suelen ser im- portantes en la fisiopatología sexual. temas efectores en diferentes células, de manera que hay efectos sexuales antagonistas pero también efectos agonistas y, por tan- to, así es como deprime algunos aspectos y estimula otros de la función sexual. Las investigaciones indican que el receptor 5-HT1 tiene efecto inhibitorio en la conducta sexual. En modelos animales se ha observado que la serotonina disminuye el deseo sexual, la ansiedad y la agresividad. En humanos el aumento de serotonina en ambos sexos inhibe la conducta sexual. Este efecto está mediado por el receptor 5HTlBl2C en varones y el receptor 5HTIA en mujeres. Los receptores que median la conducta, estimulándola, son el5HT1A en varones y 5HTlBl2C en mujeres. A nivel central puede hacer que el núcleo paragigantocelular (NPGc) inhiba la acción de la eyaculación, ya que se estima que el NPGc es serotoninérgico. Prolactinø (ProD Debilita la liberación del LH, lo que resulta en una disminución de la producción de testosterona. Se reducen los niveles de 5-alfa-reductasa y, como consecuencia, se disminuye la dihi- Feniletilamina (FEA) La FEA es una amina semejante a las anfetaminas y se la considera como un neuromodulador cerebral. Es un estimulante natural, similar a una anfetamina y se propone que esta molécula orgánica es la responsable de la excitación que sienten las personas enamoradas. La secreción de FEA inicia una cadena de reacciones en el cerebro, estimulando en primer lugar la secre- ción de dopamina y de oxitocina. Se ha postulado que la FEA realiza una función en la capacidad de enamorarse pues en numerosas publicaciones se describen sus acciones como productores de nlos slntomasu de la atracción. Ejerce una acción coordinada en el momento de la cantidades significativas en el cerebro me^parcceen dio en circunstancias de atracción por otra persona. Puede causar taquicardia, polipnea y elevación de la presión arterial. atracción y GABA El sistema de neurotransmisión GABA es el más predominante en los circuitos deAPO (|"tryet al., l99l), N\rM (Makara et al., 1975), BNST (Visden et al., 1992) y amígdala (Ni- Neuropsicología de la sexualidad 88 Neurotransmisores 89 techka E¿ Frotscher, 1939). La función principal del GABA es inhibir la actividad eléctrica del cerebro. El sistema de GABA provee el mecanismo principal para controlar la excitabilidad de La oxitocina facilita la atracción, la sensación táctil, las caricias y aumenta la sensibilidad genital. Con las caricias se libera oxitocina y durante el coito y el orgasmo causa contraccio- las neuronas. nes uterinas. En términos de conducta sexual se ha demostrado que tanto los niveles de GABA en el cerebro (Herbison 8¿ Fénelon, 1995) como el número de receptores de GABA, son sensibles a hormonas sexuales (Schumacher et al., 1989). Se ha confirmado que la transmisón GABA en el NVM del hipotálamo es necesaria parala expresión de la conducta sexual en hembras (McCar- thy et aJ.,1995). Oxitocina y uasopresina La hipófisis posterior libera dos hormonas, oxitocina y vasopresina, producidas por los núcleos supraóptico y paraventricu- lar del tálamo. Estos núcleos hipotalámicos contienen grandes células cuyos axones se dirigen a través de la eminencia media y el tallo hipofisario hasta la neurohipófisis. La neurohipófisis es una red especializada que recibe las hormonas del hipotálamo y las libera a la circulación general. Lavasopresinay la oxitocina son péptidos formados por nueve aminoácidos, con una estructura muy semejante, diferenciándose solo en dos aminoácidos. Se sintetizan como prohormonas en los somas de las neuronas magnocelulares y son transportâdas en vesículas a lo largo de los axones hasta la neurohipófisis donde van a ser liberadas. El procesamiento de las prohormonas en vasopresina y oxitocina se produce durante el transporte a lo largo del axón. La liberación de estas hormonas en la circulación general, tiene lugar cuando los potenciales de acción producidos en las propias células neurosecretoras llegan hasta el terminal axónico. Si bien las neuronas dopaminérgicas se sitúan principalmen- te a nivel del NPV y del ârea preóptica medial, la transmisión espinal la efectúan a través de fibras mediadoras de la oxitocina que finalmente liberan el NO. Las neuronas dopaminérgicas tienen eferencias oxitocinérgicas que constituyen la interneurona mediadora entre el sistema cerebral y las neuronas espinales, demostr¿indose que se trabaja con la dopamina, pero el efecto final es el de la oxitocina. La oxitocina está involucrada fundamentalmente en la fecundación, en el parto y en la lactancia. La estimulación sexual de la mujer durante el coito aumentâ la secreción de oxitocina, la cual interviene en las contracciones uterinas que ocurren durante el orgasmo. Estas contracciones uterinas facilitan la fertilización del óvulo al propulsar el esperma hacia las trompas de Falopio. En el caso de los hombres facilita la circulación del esperma y la contracción del tejido musctllar liso de los órganos reproductores. Además, la oxitocin a act:ú¿_ como neuromodulador en el cerebro, donde interviene en diferentes procesos conductuales. Por tanto, sus efectos sobre la conducta se deben a su intervención en diferentes circuitos del SNC, confirmándose la existencia de receptores para oxitocina en la amígdala, el hipotálamo ventromedial y el septum. Por otro lado, parece ser que es la hormona implicada en la formación de vínculos entre individuos. Es sensible al tacto y al contacto físico y está demostrado que favorece la conducta maternal. También está implicada en el amor romántico, en el deseo y en la receptividad sexual: se han detectado mayores niveles de oxitonina Neuropsicología de la sexualidad 90 en personas que dicen estar enamorándose. Al parece¡ el aumento de los niveles de oxitocina en el cerebro de la hembra durante la actividad sexual es fundamental para el establecimiento de lazos afectivos con su pareia. Se podría decir que la oxitocina es la molécula que se encuentra en la mayoría de las vinculaciones afectivas. Por otro lado, debido a su capacidad para provocar la contracción del útero, la oxitocina participa en el parto, desempeñando un papel fundamental en la intensidad y frecuencia de las contracciones uterinas. Thmbién se la considera como la hormona de la lactancia,ya que produce la eyección de leche al provocar la contracción de las células mioepiteliales que rodean los alveólos de las glándulas mamarias (Abril Alonso, et aJ,., 2007). La uasopresina actua sobre las neuronas del núcleo paraventricular y supraóptico que segregan hormonas, inhibiendo la secreción. Al igual que la oxitocina en el caso de las hembras, parece que está implicada en que los machos establezcan lazos emocionales, apego y vinculación de pareja. Tämbién interviene en la defensa y vigilancia de las crías. Por otro lado, actúa sobre la erección y eyaculación. Se ha observado en una especie de roedores que cuando un macho y una hembra copulan, permanecen ya juntos toda la vida. Esta monogamia se debe a la vasopresina ya que, si se inhibe su liberación, el macho se vuelve polígamo. Se ha descubierto que las variaciones en un gen que codifica el receptor de la vasopresina, están implicadas directamente con la monogamia y la poligamia en esta especie. De este mismo gen existen en el ser humano variantes que, en estudios recientes, se han relacionado con diferencias en la fidelidad de parqa que muestran los hombres (Abril Alonso, et al., 2009). Neurotransmisores 91 En resurnen, las sustancias qulmicas liberadas en el cerebro provocan reacciones que hacen que se desencadenen sensaciones durante la respuesta sexual. Al igual que en las estructuras cerebrales, la mayoría de estudios sobre la actuación de estas sustancias de estas sustancias se han realizado en modelos animales, siendo el estudio en humanos muy escaso. Las monoaminas son el tipo de neurotransmisores más importantes en la regulación de la conducta sexual, incluyendo la dopamina, que es el neurotransmisor principal del placer y se dirige hasta la cortezafrontal, provocando sentimiento de euforia y aumento del deseo sexual. Simultáneamente, el organismo segrega noradrenalina, responsable de la excitación sexual, y serotonina, que provoca la inhibición de la conducta sexual. Hace apenas unas décadas, se planteó el estudio del amor como un proceso bioqulmico regulado por dos sustancias qulmicas: oxitocina y vasopresina. Se ha confirmado también que el cerebro de una persona enamorada contiene grandes cantidades de feniletilamina, responsable de las sensaciones y modificaciones fisiológicas del enamoramiento. Con motivo del estudio de los neurotransmisores' se ha llevado a cabo Ia alternativa de realizar estudios sobre nla adicción aI sexor. Actualmente, la definición de este diagnóstico está re- sultando muy polémica: ha sido clasificado como trastorno hipersexual, pero todavía no se ha llegado a un acuerdo sobre su fisiopatología, por lo que es importante hacer una breve revisión sobre los factores neurobiológicos. V Hipersexualidad I I La hipeisexualidad es una alteración psicopatológica frecuentemente asociada a diversos cuadros psiquiátricos y neurológicos (Iftueger Ec Kaplan, 2000; Frías et al., 2012). Existe un notable desacuerdo en torno a la conceptualización de la hipersexualidad primaria (Kaplan Ec I(ruege¡ 2010), ha- Neuropsicología de la sexualidad 94 biendo sido formulada como una disfunción del deseo sexual (Kafka,2010), una patología del espectro obsesivo (Coleman, 1990), una adicción conductual (Goodman, 2001) o un trâstorno del control de impulsos (Bancroft 8¿ Vukadinovic, 2004). Esta falta de consenso epistemológico ha conllevado que su nosologíahaya sido irrelevante desde que fue incluida entre los (trastornos psicosexuales no especificados, del DSM-III. La DSM-V propone el trastorno hipersexual como un nuevo diagnóstico, caracterizado por el aumento en la frecuencia e intensidad, excitación, impulsos y conductas sexuales no parafílicas, asociadas a un componente de impulsividad que provocan un malestar significativo o un deterioro social o ocupacional. Existe un relativo consenso en torno a la mayor prevalencia de la hipersexualidad primaria en hombres (5:1) (Frías & Vázquez,2012). Los síntomas principales de este trastorno son los siguientes: - las fantaslas, impulsos y conductas sexuales interfieren de manera repetitiva en las actividades y obligaciones diarias; - responder de manera repetitiva a fantasías, impulsos y conductas sexuales con estados de ánimo: ansiedad, depresión o irritabilidad; - tener fantasías sexuales, impulsos y conductas sexuales en respuesta al estrés de la vida diaria; - el sujeto se esfuerza para controlar y reducir esos impulsos, deseos y conductas sexuales; - involucrarse de manera repetitiva en los impulsos, deseos y conductas sexuales, sin darse cuenta de los efectos fisicos o emocionales que se hace a sí mismo o a los demás. Estas fantasías, impulsos y conductas no están directamente relacionadas con efectos psicológicos producidos por sustancias como drogas o medicación. Hipersexualidad 95 Se debe especificar si presentan las siguientes conducras: masturbación, ver pornografia, utilizar cibersexo o ir a clubs de striptease (Kafka, 20 10). Respecto a su etiopatogenia, la mayoría de los estudios son transversales o retrospectivos, sin grupo control comparativo, por lo que los hallazgos que se han obtenido hay que tomarlos con precaución (Frías k Yázquez, 2012). Se han reaJizado pocos estudios en cuanto a la psicobiología de la hipersexualidad. Sin embargo, la literatura y los estudios en sujetos con este trastorno, evidencian la existencia de sistemas neuroquímicos y anatómicos en el cerebro que desempeñan un papel importante, aportando factores biológicos que dan explicación a este trastorno. La mayoría de los trabajos se han hecho con población sin presentar patología, examinando únicamente la excitación sexual mediante estímulos visuales en pacientes sanos. Sería interesante reaJizar estudios de neuroimagen en pacientes hipersexuales. Una falta de consenso con respecto a la fisiopatología de la hipersexualidad, agravada por la falta de datos empíricos, obsttculiza la caracterización de este trastorno sexual no paraftlico asociándolo con diferentes clases de ffastornos (Bancroft Ec Vukadinovic,2004). Mientras que algunos teóricos dudan sobre la validez de establecer una categoría diagnósticaparalos ffastornos de la conducta sexual no parafilica (Giles, 2006),las investigaciones y la literatura de diferentes perspectivas teóricas han planteado si la hipersexualidad es un trastorno primario de la sexualidad (Kafka, 2007), una adicción (Goodman, 1997), un trastorno del espectro obsesivo compulsivo (Black, 1998), un trastorno del control de impulsos (Mick & Hollander,2006) o una excesiva conducta sexual que se encuentra fuera de control (Bancroft E¿ Vukadin ovic, 2004). Neuropsicologfa de la sexudidad 96 97 Esta formulación puede ser aplicada al trastorno hipersexual, Tiastorno drl deseo sexual Los estudios con neurotransmisores en los comportamientos sexuales se han cenüado en las monoaminas (serotonina, dopamina y norepinefrina) utilizando modelos animales. La investigación en población cllnica es escasa. El funcionamiento sexual necesita de estas monoaminas, como se evidencia por la induc- ción de disfunción sexual tras la administración de inhibidores selectivos de la recaptación de serotonina (ISRS) y el aumento de la sexualidad cuando se aplican estimulantes. En la literatura, se ha descrito el deseo sexual como la presencia de fantasías sexuales, impulsos o actividades, y la motivación para participar en la conducta sexual en respuesta a señales internas o externas (Bancroft, 2009). Una explicación neurobiológica sobre la alteración del deseo sexual puede ser el modelo presentado como la ohipótesis de monoaminas, aplicado a los üastornos parafilicos (Kafka, 2003) elaborada por primera vez por Kafka en 1997 . Figura 9 Modelo de la <Hipótesis de monoaminas>> f)npamina Hipersexualidad Noradrenalina Serotonina si este se basa en intensas y frecuentes fantasías sexuales e impulsos y actividades sexuales con graves consecuencias. Este mode- lo se realizó en el laboratorio, demostrando que el cerebro tiene receptores monoaminérgicos que interactúan con hormonas sexuales, testosterona y neuromoduladores, proponiendo una base biológica para el deseo sexual y la conducta copulatoria en los ratones (Meston E¿ Frolich, 2000). Se ha observado, en los estudios publicados en los que se eva- luaba la conducta sexual de los ratones, que la mejora de la dopamina estaba asociada a la excitación sexual, mientras que el aumento de la serotonina se relacionaba con la inhibición de la conducta sexual. La alteración de la dopamina y la serotonina afecta a la motivación sexual y provoca desinhibición e hipersexualidad en animales. Por otro lado, también se ha demostrado que el áreapreóptica es esencial en la conducta consumatoria de la conducta sexual. Las lesiones y manipulaciones qulmicas en esta área influyen, por ejemplo, en la eyaculación y promueven conductas de intromisión, sin alterar el deseo sexual. En cambio, el núcleo ventral esüiado, involucrado en el mecanismo de la dopamina, afecta al deseo sexual pero no altera la conducta copulatoria. Por último, se sabe que la testosterona está muy implicada en la expresión de la conducta sexual. El rol exacto que puede ejercer la testosterona en la conducta hipersexual actualmente se desconoce. En ratones adolescentes, se ha podido observar correlación entre testosterona y frecuencia de conducta sexual (Mick E¿ Hollande¡2006). Sin embargo, se ha encontrado que los agentes que disminuyen los niveles de testosterona han sido efectivos para disminuir los impulsos sexuales en las parafilias ¡ por lo tanto, podríamos aplicarlo al úastorno hipersexual. Neuropsicologla de la sexualidad 98 Hipersexualidad y patología en el Eje I El ffastorno hipersexual está relacionado con una elevada comorbilidad en el Eje I, siendo más frecuente en aquellos ffastornos que presentan alteración en los neurotransmisores monoaminérgicos, englobando dimensiones psicopatológicas como la impulsividad, la compulsión o la depresión (Kafka, 1997). Los trastornos más frecuentes son: trastornos del estado de ánimo, como la depresión y la ansiedad, en las cuales se produce una disminución de los - niveles de serotonina; - trastorno bipolar, en el que se encuentra alterado el gen transportador de serotonina (5HTT); - trastorno por déficit de atención e hiperactividad, en el cual aumentan los niveles de dopamina y norepinefrina. Esto puede proporcionar un puente entre las bases neurológicas, patologla en el Eje I, hormonas sexuales masculinas' nellrotransmisores monoaminérgicos e hipersexualidad (Kafka, 2010). Hipersexualidad 99 (Stein, 2008), pero no ha sido validado en población clínica. En este modelo la amígdala provoca la disregulación emocional, la conducta de recompensa está regulada por el núcleo accumbens y el circuito estriado ventral, mientras que el proceso cognitivo lo controla lacortezaprefrontal (Samenow, 2010). Esto explica la vía mesolímbica, también denominada vía de la recompensa, siendo una de las vías dopa-minérgicas del cerebro. Incluye el área tegmental ventral que se encuentra en el mesencéfalo, enviando sus proyecciones dopaminérgicas al núcleo accumbens, conectando asl con el sistema límbico. Los elementos neuroquímicos en estas estructuras se asocian a la dependencia de sustancias psicoactivas e incluyen una disminución de la dopamina y serotonina en el núcleo estriado ventral. Figura 10 Vias de acción de la dopamina Adicción Recientemente, algunas investigaciones han utilizado la expresión nadicción conductual) para referirse a conductas como la ludopatía, la cleptomanía, el uso del móvil e internet, tricotilomanía o la conducta sexual compulsiva o hipersexualidad (Pallantini, 2006),las cuales tienen caracterlsticas comunes con la adicción de sustancias. Se ha hipotetizado que la conducta sexual repetitiva, acompaiada de una gran emoción, frecuencia e intensidad, puede provocar cambio en los circuitos cerebrales que ayuden a perpetuar la conducta (Mick & Hollander,2006). El modelo ,,Amygdala Behauioural Cognitiuø, ha sido propuesto como hipótesis para dar explicación a la hipersexualidad Fuente: http://www drugabuse. gov T Neuropsicología de la sexualidad 100 En humanos, la coÍteza. prefrontal y la corteza cingulada anterior están asociadas funcionalmente con la motivación y la recompensa y, por otro lado, estudios de neuroimagen funcional han demostrado que estas dos áreas se activan durante la respuesta sexual humana (Ress, 2007).La altencion en estos circuitos y su relación con las estructuras del sistema límbico, en especial la amígdala, han sido detectadas con fMRI y procedimientos de neuroimagen, como con evaluaciones neuropsicológicas que determinaron el abuso de sustancias y conductas adictivas. Algunos investigadores sostienen que las regiones de la vía de la recompensa y el placer son moduladas por hormonas sexuales, para asl mejorar y facilitar la respuesta a las relaciones y deseos sexuales. Estos estudios neurobiológicos pueden ser aplicables a la adicción sexual si la neurobiología y los circuitos neuronales Hipersexualidad 101 Pacientes con lesiones y tumores en el lóbulo frontal pueden provocar la expresión del comportamiento sexual desinhibi- do, lo que puede explicar el aumento de la actividad sexual junto con la disminución del control de la propia conducra. Pero sobre este asunto se necesita más investigación ya que hay sujetos que no experimentan conducta sexual desinhibida (Reid et a1.,2010). Mientras que las lesiones frontales pueden coincidir con desinhibición, incluyendo respuestas hipersexuales a estímulos, las lesiones en el circuito estriatal van acompañadas de activación de patrones de respuesta interna (deseo o motivación sexual). Por otro lado, las lesiones en el circuito temporal límbico, específicamente en la amígdala, provocan el síndrome de Klüver-Bucy, acompañado de perturbaciones en el deseo sexual, incluyendo desviaciones en el impulso sexual. son similares a los mecanismos que regulan el abuso de sustancias o la ludopatía (Kafka, 2010). Se podría concluir que el üastorno hipersexual es un conjun- to de trastornos afectivos, provocados por el estrés, la preocupación y la motivación de obtener una recomPensa sexual, terminando con una conducta repetitiva a pesar de las consecuencias negativas (Samenow, 2010). Lesiones La conducta hipersexual se ha podido observar en individuos que han sufrido lesiones traumáticas cerebrales. Los estudios de estas áreas cerebrales han aportado ideas sobre la localizacióncerebral que regulan este tipo de conductas. Por ejemplo, se ha observado conducta hipersexual en pacientes con infartos talámicos (Spinella, 2004). Enfe rmedades neuro higica s Los trastornos neuronales frecuentemente alteran la respues- ta sexual (Anderson, 2004) y pueden cambiar el procesamiento de los estímulos sexuales, incrementando el deseo sexual (Rees, 2007). Aquellos sujetos con enfermedades neurológicas que implican a los lóbulos temporales y áreas del cerebro medio, como trastornos convulsivos, enfermedad de Huntigton y demencias (frontotemporal) muestran síntomas de hipersexualidad. En los pacientes que sufren epilepsia, en los lóbulos temporales se ha evidenciado que aquellos que están tomando lamotrigina, un fä.rmaco que reduce los ataques epilépticos disminuyendo los niveles de glutamato, manifestaban un incremento del deseo sexual típico del trastorno hipersexual. f Neuropsicología de la sexualidad 102 Hipersexuaìidad 103 el tratamiento más común para tratar la enfermedad de Parkinson, manifiestan conductas hipersexuales debido al aumento de Figura 11 Treating parkinson's disease ???? la dopaminay la activación de sus receptores. corp\¡s 5lrìâl¡¡m St¡lril.rntrr l*içr.ì c 1 e Fuente: Lippincott, W & Wikins (2001) Dopamine L-Dopa Corpus Striatum Substantia Nigra Blood Brain Barrier [L-Dopa Therapy]. http://www smartdraw.com La hipersexualidad también se ha demostrado que está inducida por medicamentos en la enfermedad de Parkinson, ya que implican a los sistemas de dopamina tan involucrados en el comportamiento sexual. Se ha observado que una de las casusas es la inhibición de la prolactina, una hormona que desempeña un papel importante en la regulación de las hormonas sexuales y que, al reprimir el efecto de la dopamina, puede inducir el trastorno hipersexual. Tâmbién indica el nivel de satisfacción sexual: altos niveles de prolactina se asocian con la impotencia, una pérdida de la libido y disminución de la hormona sexual. Así que si un paciente con enfermedad de Parkinson consume un fármaco que inhibe la prolactina, los niveles bajos pueden aumentar la libido y que dé explicación a un comportamiento hipersexual. Tâmbién se ha descrito, que los pacientes que toman L-Dopa, En resumen, el DSM-V propone el úastorno hipersexual como nuevo diagnóstico, caracterizado por el aumento en la frecuencia, intensidad, excitación, impulsos y conductas sexuales no parafilicas, asociadas con impulsividad, que provocan malestar y deterrioro social y ocupacional. Este trastorno se comprende desde diversas perspectivas fisiopatológicas como un comportamiento compulsivo, impulsivo o adictivo. Se han reaJizado pocos estudios sobre la etiopatogenia y correlatos biológicos sobre este trastorno. La explicación neurobiológica de la hipersexualidad se ha centrado principalmente en la hipótesis de las monoaminas, por la cual, la alteración de la dopamina y la serotonina afecta al deseo y a la motivación de la conducta sexual. Por otro lado, si entendemos la hipersexualidad como una nadicción>, la alteración de la vía mesolímbica y en relación con el sistema límbico, puede ser el factor biológico que dé explicación a este trâstorno. Además, la descripción de casos clínicos en los que el sujeto ha sufrido una lesión cerebral o una enfermedad neurológica; o la aplicación de medicamentos, está ayudando a entender los mecanismos neurológicos del trastorno hipersexual. VI Epllogo Decla Gregorio Marañón que nel cerebro es el órgano sexual más importante del ser humanor. La cuestión general que se puede plantear es si existe un sexo cerebral, es decir, una sexualización del cerebro, y en el supuesto de que hubiese tlg"" tipo de diferencia en la organizaciín del cerebro de varones y de mujeres, determinar qué conductas sexuales regula y si estas estructuras están afectadas en trastornos sexuales. La manera como se ha ido estudiando la conducta sexual ha sido a través de modelos animales en el laboratorio, sobre todo con ratas y monos, lesionando los circuitos cerebrales de estos animales y administrando Pármacos que aumentasen o disminuyesen los niveles de neurotransmisores involucrados en este tipo de conductas. En humanos ha sido posible el conocimiento de las estructuras cerebrales que regulan la función sexual mediante la observación de casos cllnicos en individuos que han sufrido una lesión traumátic¿ o enfermedad neurológica, especialmente en epilepsia y enfermedad de Parkinson. Estas enfermedades frecuentemente alteran la respuesta sexual y producen cambios en el procesamiento de los estlmulos sexuales. Desde el a-ño 2000, gracias a los estudios realizados con neuroimagen funcional y tomogra- -T 106 Neuropsicología de la sexualidad fía por emisión de positrones, se han corroborado qué regiones cerebrales son activadas durante la respuesta sexual (figuras 13 y 14), siendo las más importantes las áreas localizadas en el sistema límbico, las áreas parietales y el lóbulo frontal. Epílogo 107 Se necesitan más estudios utilizando fMRI para determinar de manera más concreta qué áreas cerebrales regulan la conducta sexual y de qué mânera establecen conexiones y redes neuronales. Es necesario contrastar grupos de controles sânos con pacien- tes que padezcan algún trastorno, como el hipersexual descrito Figura 13 Regiones cerebrales que controlan la funcion sexual en esta revisión. Habría que prestar atención a las siguientes estructuras posiblemente involucradas en este trastorno: - El área preóptica, ya que es una estructura que participa (lqlùl¡t. gyÀrj l^f¿riur p¡ríctal TlFlàm6 0rbilo ftunt¡l (ol1q ln6riôr fronLrl gyrut Scptum 5$lrl n(¡(í{b{hind) Hypôll!rl¡¡lic nwl.í Hipp(¡mI¡l gyru! lfgM^lum (h(hrftl) Fuente: Ress et al., 2007 Figura 14 lmagen que muestra las regiones que se activan durante la respuesta sexual obtenidas mediante fMRl Fuente: Ress et al , 2007 en la expresión de la conducta sexual. En macacos se ha observado que está asociada con la erección y el acto sexual. En humanos el ârea preóptica es el núcleo intersticial del hipótalamo. El núcleo INAH-I se ha determinado que es el que controla la conducta sexual. - El núcleo ventromedial regula y facilitala conducta sexual en las hembras. Un posible aumento de tamaño o una gran actividad de este núcleo en mujeres que sufren hipersexualidad sería otro factor importante a examinar. - El estudio del núcleo de la estría terminal sería interesânte para establecer un factor biológico en la conducta hipersexual. Es importante en la función sexual, ya que establece conexiones entre el sistema llmbico y el hipotálamo. Desempeña un papel importante en el comportamiento sexual masculino y en la secreción de gonadotropinas. - Respecto al estudio de la estructura de la amígdala, ya se están realizando estudios con resonancia magnética para observar su funcionamiento en la regulación de la conducta sexual. Se ha observado que se activa más en hombres que en mujeres ante estímulos sexuales visuales. Es necesario que se observe en pacientes con conducta hipersexual, específicamente en la parte superficial y centromedial de la amígadala, ya que son las regiones que regulan T 108 Neuropsicologla de la sexualidad 109 Epílogo los procesamientos afectivos y conúolan las respuestas en- docrinas y emocionales. hay una mayor secreción, si las áreas que la producen son más grandes que en individuos sanos o si las proyeccio- Se sabe además que la amígdala establece conexiones con nes dopaminérgicas de la vía mesolímbica al lóbulo fron- el hipotálamo. Es posible que en pacientes con trastorno tal son mayores en personas hipersexuales. - En los hipersexuales, puede haber un aumento de los niveles de noradrenalina, ya que esta monoamina es la responsable de la excitación sexual y del procesamiento de los estímulos entrantes que se incluyen en este tipo de res- hipersexual esta conexión sea mayor. - Cabe señalar que el núcleo medular bulbocavernoso es más grande en los varones y controla el proceso de la eyaculación. Comparando un grupo de hombres sanos y otros que hayan sido diagnosticados de hipersexualidad, se podría observar si en estos el núcleo bulbocavernoso es puesta. - Pueden existir niveles bajos de serotonina, ya que es la res- ponsable de la inhibición de la conducta sexual. La disminución de este neurotransmisor puede hacer que no inhi- mayof. Por otro lado, los procesos relacionados con los neurotransmisores involucrados en la función sexual humana deberían ser un factor importante a estudiar. Ha habido autores que han estudiado la hipótesis de las monoaminas, posible explicación biológica de los trastornos sexuales parafilicos y que se podría aplicar al üastorno hipersexual. Tias hacer la revisión de las sustancias químicas responsables de la conducta sexual, es conveniente profundizar en el estudio de los siguientes neurotransmisores: - El óxido nítrico es un gas que interviene en el cerebro ejecutando la función de neurotransmisor. Es principalmente el responsable de la respuesta de la erección en el hombre. Quizás un aumento de esta sustancia puede ser un factor influyente en el desarrollo de la hipersexualidad. La - testosterona, hormona sexual masculina responsable del aumento de la libido y de la potencia sexual, podría presentar en este tipo de pacientes un nivel sanguíneo mayor. - La dopamina es el neurotransmisor que más se ha estudiado en las adicciones sexuales y trastornos parafllicos sexuales. Se necesitan estudios que se centren en el modo de actuar de esta sustancia química, observando por ejemplo si ba correctamente la conducta sexual, hipersexualizando al - paciente que padece este trastorno. La feniletilamina puede estar aumentadaya que es la responsable de aumentar la liberación de la dopaminay así dar explicación aI aumento de los deseos sexuales. Por último, puede haber un aumento de la oxitocina, ya que actúa en la atracción sexual humana, y una disminución de la vasopresina, responsable de los lazos emocionales. Esto podría explicar la promiscuidad de los pacientes que sufren trastorno hipersexual. Por último, respecto al trastorno hipersexual se deben realizar estudios más profundos paru aclarar la psicopatología subyacente y así poder discriminar entre síntomas o síndrome hi- persexual que acompañen a otras patologías, por un lado, o trastorno hipersexual con entidad propia, por otro. Futuras líneas de investigación deben determinar los correlatos biológicos que dan explicación a este trastorno y asl permitir un mejor abordaje terapéutico. Sería necesaria la utilización de modelos animales y el estudio con fMRI con un grupo control y otro que haya sido diagnosticado de üastorno hipersexual. _- ll0 Neuropsicología de la sexualidad Conviene recordar que el esûdio con seres humanos es más complejo que el esftrdio con animales, ya que hay que tener en cuenta las circunstancias familiares, ambientales, culturales, sociales y experiencias de vida que ejercen su influencia más allá de la neurobiología. CoNcr-usroNes 1. En el desarrollo ontológico de la sexualidad, el cigoto pasa por cierto número de estadios ontogénicos sucesivos, con dos fases críticas. Estas fases críticas son las siguientes, estableciéndose por orden cronológico: - la diferenciación de los câracteres sexuales primarios y de los esbozos indiferenciados de los conductos genitales in- tefnos y externos; la - diferenciación endocrina, cíclica para la mujer y continúa para el hombre mediante el sistema hipotálamo-hipófiso-gonadal; - la diferenciación de los caracteres sexuales secundarios, estableciéndose el fenotipo en la etapa de la pubertad. 2. Los estudios realizados sobre la organización sexual del cerebro en modelos animales y en la especie humana han permitido concluir que en el cerebro hay regiones diferentes responsables del comportamiento sexual en varones y mujeres. Estas estructuras sexualmente dimórficas, se localizan principalmente en el hipotálamo y en el sistema límbico. 3. Los efectos de las hormonas sexuales sobre el desarrollo del cerebro están modulados, al menos en parte, por los neurotransmisores, siendo estos los organizadores y responsables en el control de la función sexual humana. Los neurotransmisores más involucrados en la respuesta sexual humana son las monoaminas: dopamina, serotonina y noradrenalina. Epílogo 111 4. Los correlatos neurobiológicos conocidos que plantean la hipótesis de que pueden ser el factor biológico del trastorno hipersexual, serían, por un lado, la alteración de las monoaminas, principalmente la dopamina y serotoni a, y por otro, una al- teración en la vía dopaminérgica mesolímbica. Además, los estudios basados en sujetos con lesiones frontales o que se les ha diagnosticado de Parkinson o de epilpesia en el lóbulo temporal, han aportado conocimiento sobre la etiología del trastorno hipersexual. VII Referencias bib liográficas Abramovich, D. R., Davidson, I.A.., Longsraff, A. & pearson, C. K. (1987), Sexual differentiation of the huma¡r mid- uimester brain. EuropeanJournal of Obxenic Crynaecotngic and Reprodactiue Biology,25,7-14. Abril Alonso, -Á,. et aJ. (2007), Fund¿mentos biológicos d¿ h conducta. Madrid: Sanz yTones. Abril, .A., Segovia, S. Ec Guillamon, A. (I987),The bed nucleus of the stria terminalis in the rac regional sex differences conuolled by gonadal steroids earþ after birth. Deu. Brain Res., 32,295-300. Acuña, A. (2008), La casceda de neurotransmisores en la función sexueJ.. Reuista Urologia Colombiana, 17, l0T-120. Allen, L. S. & Gorski, R. A. (2002). Sex difference in the bed nucleus of the stria terminalis of the human brain, en G. Einstein (ed.), Sex and the brøin (pp.299-309). Cambridge, MA US: MIT press. Allen, L. S., Hines, M., Shryne,l. E. & Gorski, R. A. (1989), Two sexually dimorphic cell groups in the human brun. J. Neuroscience,9, 497-506. Allen, L. S., E¿ Gorski, R. A. (1990), Sex difference in the bed nucleus of the stria terminalis of the human brun. J. Cornp Neurol., 302,697-706. ,{.munts, K. (2005), Cytoarchitectonic mapping of the human arrrygdala, hippocampal region and entorhinal correx intersubject va- Y 1t4 Neuropsicología de la sexualidad riability and probability maps, Anatomy and Embryology,2l0, 343-352. Arnold, A. P. S¿ Gorski, R. A. (1984), Gonadal steroid induction of structural sex differences in the CNS. Annu. Reu. Neurosci,T (4), t3- 442. Bancrof, J. (2009), Human sexudlity and its problems (3'o ed), Oxford, England: Elseviver. Bancroft, J. &Vukadinovic,Z. (2004), Sexual addiction, sexual compulsivity, sexual impulsivity or what? Toward a theoretical model. Journal of Sex Researclt, 4I,225-2234. Bardin, C. W: (1936), Pituitary- testicular axis. Reproductiue endocri' nologlt: Phyiology, pathopfusiology and clinical mdndgement, Flladelgia: Saunders. Black, D. \ø (199S), Compulsive sexual behavior: A review. Journal of Practical Psychiany and Behauioural Heøhh, 4,217-229. Bocchino, S. (2006), Bases psiconeuroendocrinas del dimorfismo sexual cerebral. Reuista dr Psiquiatria drl Uruguøy,70 (2), 167-177. Borgarelli, M. P. (2007), Aporte para el conocimiento anatomo-funcional del órgano vomeronasal humano y su probable relación con la conducta sexual. Reuista Argentina de Clínica Neuropsiquid- trica, 14,5-48. Bra¡ G. A. E¿ York, D. A. (1997), Leptine and clinical medicine a new piece in the puzzie of obesiryl Clin. Endocrinol. Metab., 82, 277r-2776. Burge¡ H. G. (1987), Reproductive Endocrinology, en H. G. Burger (ed.), Clinical endocrologlt ønd metabolism (vol.l), Londres: Bailliere Tindall. Byne, \Ø, Lasco, M. S., Kemether, E., Shinwari,,{., Jones, L. E¿Tobet, S. (2000), The interstitial nuclei of the human anterior hypothalamus: Assessment for sexual variation in volume and neuronal size, density and number. Brain Res., 856,254-258. Capel, 8., Albrecht, K., \Øashburn, L. E¿ Eicher, E. (n. d), Migration of mesonephric cells into the mammalian gonad depends on Sry. Mechønisms of Deuelopment, B4(l-2), I27-I3L Referencias bibliográficas 115 Carrer, H. F. E¿ Aoki A. (I982),lJltrastructural changes in the hypothalamic ventromedial nucleus of ovariectomized rats after estrogen rrearmenr. Brain Res., 240, 221-233. Castro-Feijoo, L. E¿ Pombo, M. (2006), Pubertad Precoz. An. Pediatr Contin., 4(2), 79-87 . Claro, F., Segovia, S., Guillamon, A. & Del Abril, A. (1995), Lesions in the medial posterior region of the BSI impair sexual behavior in sexually experienced and inexperienced mate rates. Brain Rel Bull.,36,I-10. Coleman, E. (1990), The obsessive-compulsive model for describing compulsive sexual behaviour. American Journal of Preuentiue Psychiøtry y't Neurology, 2,9-I4. Colina Chirinos, A. E¿ Moncada Rodríguez, C. A. (2007), Determinación sexual primaria o sexo genético. Revisión. (Spanish), Mr- duk, 16(2), 67. Conn P M., Huckle, W: R., Andrews, W: V & Mcfudle, C. A. (1987), The molecular mechanism of action of gonadotropin releasing hormone (GnRH), in the pituitary. Recent Progress in Hormones Researclt, 43, 29-61. Czyba, J. C. et al,. (1978), Ontogénesis d¿ l¿ sexuølidad humana: desø- rrollo dz los aparøtos genitales y dzl comportarniento sexual. Barcelona: Eunibar, 1978. Dizarega, G. G. & Hodgen, G. D. (1981), Folliculogenesis in the primate ovarian a¡cIe. Endocrine Reuiews, 2,27. Dörner, G. & Staudt, J. (1968), Structural changes in the preoptic anterior hypothalamic area of the male rat. Following neonatal castration and androgen treatment. Endocrinologlt, 3, 136-140. Emori, T (1991), Cellular mechanism of endothelin-l release of angiotensin and vasopressin. Hipertension, IB, 165-71. Everitt, B. J. (1990), Sexual motivation: a neural and behavioral analysis of the mechanisms underþng appetitive and copulatory responses of male rats. Neurosci. Biobehau. Reu., 14,2I7-32. Exton, M. (2000), Cardiovascular and endocrine alterations after masturbation-induced orgasm in women. Sexual Dffunction in Medicine,l(3), 80. Neuropsicologfa de la sexualidad 116 Fernández, J. 0996), Varones y rf,ujeres: dzsørrollo dß k dtblr realidâ¿ dzl sexo 1, dzl género / Juan Fernández (coordinador), Madrid: Pi- râmide,1996. Fernández-Guasti,,4.., Kruijver, F., Fodor, M. E¿ Swaab, D. (2000), Sex differences in the distribution of androgen receptors in the human hypothalamus. The Journøl of Comparatiue Neurologlt, 425(3),422_435. Frías, A. (2012), Hipersexualidad primaria: estatus nosológico, etiopatogenia y tratamiento. Reuista de Psiquiatía y Salud Mental, 5 (3),205-207. Gadner, R. L., Lyon, M. F., Evans, E. P. & Burtenshaw, M. D. (1985), Clonal analysis of X chromosome inactination and the origin of the germe lime in the mouse. Embriology and Experimental Morpholngy, 88,349-363. Gámez, L., García Hidalgo., Yánquez, S., Sámano, D., Reyes, J., B"surto Acevedo, N. & Rivas Bucio, R. (2009), Las diferencias ana- tómicas cerebrales que implican diferecias funcionales (I y 2a de dos partes), (Spanish), Reuista de l¿ Facubad d.e Medicina d¿ la UNAM, 52(5),2t2-218. Garcia-Segura, L. M., Duenas, M., Busiguina, S., Naftolin, F. & Chowen, l. A. (1995), Gonadal hormone regulation of neuro- nal-glial interactions in the developing neuroendocrine hypothalamus.f Steroid. Biochem. Mol. Biol., 53,293-298. Giles, J. (2006), No such thing as axcessive sexual behavior [ætter to the Editorl, Archiues of Sexual Behauiour 35,641-642. Giuliano, F. & Rampin, O. (2000), The control of the penile erection by the central nervous system. Sex. Disf ofMed.,l(3),70-73. Goldstein, J. M., Seidman, J. L., Horton, N. J., Mal¡ris, N., Kennedy, D. N. ð¿ Caviness, V S. (2001), Normal sexual dimorphism of the adult human brain assessed by in vivo magnetic resonance imaging. Cen Ctx., Il, 490-497. Goodman, A. (1997), Sexual addiction, en J. H. Lowenson, P Ruiz, R. B. Millman, Ec J. G. Langrod (eds.), Substance abuse: A com- Referencias bibliográfi cas lt7 prehensiue textbooþ (3rd ed., pp. 340-354), Baltimore: \Øilliams &'$Øilkins. Goodman, A. (2001), \X/hatt in a name? Terminology for designating a syndrome of driven sexual behavior. Sexuøl Adàiction ò Com- pukiuity, B,l9l-213. Goodman, A. L. & Hodgen, G. D. (1983), The ovarian triad of the primate menstrual grcle. Recent Progress in Horrnone Research,39, r_73. Gorski, R. ,A., Girdon, J. H., Shryne, l. E. & Southman, A. M. (1978), Evidence for a morphological sex difference within the medial preoptic area and the lateral hupothalamic area on sexual behavior in the rat. Brain Research, 148,333-346. Gorski, R.A.., Harlan, R. 8., Jacobson, C. D. & Southam, A. M. Evidence for the existence of a sexually dimorphic nucleus in the preoptic area of the rat.I. Com?l. Neuro., 193,529-539. Gorski, R. (1985), Sexual dimorphism of the brain. J. Anim. Science, 61, 1001-1004. Greenough, \M, Carter, C. S., Steerman, C. ð¿ DeVoogd, T J. 0977), Sex difference in dendritic patterns in hamster preoptic area. Brain Res., 126,63-72. Gubbay J, Collignon J, Koopman P et al. (1990), A gene mapping to the sex determining region of the mouse Y chromosome is a member of a novel family of embryonically expressed genes. À/øture,346,245- 250. Guillamón, A. & Segovia, S. (2007), El sexo del cerebro. Mente 1t Cerebro,25,68-77. Gutiérrez, H., Onofre, M., Rosado-García, A. E¿ Rosales Torres, A. (2005), Sexual differentiation in the central nervous system. Wterinaria México, 36(3), 339-360. Hamann, S. (2005), Sex Differences in the Responses of the Human Amygdala. Neuroscientist, Il(4), 288-293. Hamann, S., Herman, R., Nolan, C. &\Øallen,K. (2004), Men and women differin amygdala response to visual sexual stimuli. -A/¿r. Neurosci.,7,4ll-6. ll8 Neuropsicología de la sexualidad Hedlund, H. (2001), Receptors functions in penile erection. Int Imp. Rel,13,4. Hengjun, J. K, Namkug, K., Sehyun, K., Seokjun, H., K¡rngmo, P, Sabina, L. & Gyungo, C. (n. d), Sex differences in amygdale subregions: Evidence from subregional shape analysis. Neuroimage, 60,2054-206r. Herbison, A. E. & Fénelon, V. S. (1995), Estrogen regulation of GABAA receptor subunit expression in preoptic area and bed nucleus of the stria terminalis of female rat brain. J. Neurosci., 15 (3),2328-2337. Hernández Herrera, A. et al. (2004), WTl [Recurso electninicoJ: sexo, uida y muerte. Monterrey (México): Universidad Autónoma de Nuevo León,2004. Hines, M., Allen, L. S. & Gorski, R. A. (1992), Sex difference in subregions of the medial nucleus of the amygdala and the bed nucleusof the stria terminalis of the rat. Brdin Resclt.,579,321-326. Huhteniemi, I. T., Yamamoto, M. & Ranta, T. (1988), FSH receptors appear earlier in the primate fetal tetis than in the ovary. Journal of Clinical Endocrinologlt and Metabolism, 65, l2I0-I214. Jarry, H., Leonhardt, S.,'Wuttke, \ø ( 1 99 1 ), Gamma-aminobutyric acid neurons in the preoptic/anterior hypothalamic area synchro- nize the phasic activity of the gonadotropin-releasing hormone pulse generator in ovariectomized rats. Neuroendocrinologlt, 53, 262-267. Jorge-Rivera, J. C. (1998), Dimorfismo sexual en el cerebro. Ciencia øl Día,2(1), 1-10. Jost, A. (1985), Initial stages of gonadal development: Theories and methods. Archiues ofAnatorny, 68, 123-131. Kafka, M. P. (1997a), A monoamine hypothesis for the pathophysiology of paraphilic disorders. Archiues of Sexual Behauiour 26, 337-352. Kafka, M. P (1997b), Hypersexual desire in males: An operational definition and clinical implications for men with paraphilias and paraphilia-related disorders. Arc hiues of Sexuøl B ehauior, 26, 505-526. Referencias bibliográfi cas 119 Kafka, M.P. (2007), Paraphilia-related disorders: The evaluation and treatment of nonparaphilic hypersexuality, en S. Leiblum (ed.), Principles ønd practice of sex therapy (4th ed, pp. 442-476), New York Guilford Press. Kafka, M. P (2010), H¡rersexual Disorder: A Proposed Diagnosis for DSM-V Archiues of Sexual Behauiour, 39,377-400. Kaplan, M. S¿ lGueger, R. (2010), Diagnosis, Assesments, and Tfeatment of hipersexualiry. Journal of Sex Reseørch, 47, 181-198. Keegan, J. (2001), The Neurobiology, Neuropharmacology and Pharmacological. Tfeatment of the Paraphilias and Compulsive Sexual Behavior. Canadiøn Journal of Psychiatry, 46 (l),26-34. Klinge, M. (2000), Estrogen receptor interactions with co-activators and co-repre ssor. Steroids, 65, 227 -25I. Knobil, E. (1980), The Neuroendocrine control of menstrual cycle. Recent Progress Hormone Research, 36, 53-88. IGueger, R. B. & Kaplan, M. S. (2000), Disorders of sexual impulse control in neuropsychiatric conditions. Serninar in Clinical Neuropsychiany, 5, 266-27 4. Kruijver, F. P M., Zhou, J. N., Pool, C.-Sí:, Hofman, M. A'., Gooren, L. J. G. S¿ Swaab, D. F. (2000), Male to Female Thanssexuals Have Female Neuron Numbers in a Limbic Nucleus. J. Clinical Endncrino logy and Metabo lism, 85 (5), 2034-2O4L Kuiper, G., Enmark, E., Peito-Huikko, M., Nilsson, S. & Gustaßson, I 0996), Cloning of a novel receptor expressed in rat prostate and ovary. Proc. Natl. Acad. Sci. USA,93,5925-5930. Lui, Y., Salamone, J. & Sachs B.D. (1997), Lesions in medial preoptic area and bed nucleus of stria terminalis: differential effects on copulatory behavior and erection in mate rates. J. Neuroscience, t7, 5245-5253. Makara, G. B., Rappay, G., Stark, E. (1975), Autoradiographic localization of 3H-gamma-aminobutyric acid in the medial hypothalamus. Exp. Brain Res., 22, 449-455. Masco, D. H. & Carrer, H. F. (19S0), Sexual receptivity in female rats after lesions or stimulation in different amygdaloid nuclei. Phy' siol. Behau., 24, 1073-1080. t20 Neuropsicología de la sexualidad McCarth¡ N. M., Todd, B. J. &Amateu, S. K. (2003), Estradiol modulation of astrocytes and the establishment ofsex difference in the bratn. Annak of the Neut York Acadtmy of Sciences, 1007,283-297. McCarth¡ M. M. (1995), Functional significance of steroid modulation of GABAergic neurotransmission: analysis at the behavioral, cellular, and molecular levels. Hormones and Behøuior,29, l3Ir40. -What makes a men a man? Nahtre, 346,216. McClaren, A.( 1990), Merchant, H. (1975), Rat gonadal and ovarian organogenesis with and without germ cells. Deuelopmental Biololgy,44,l-21. Meston, C. M. E¿ Frolich, P F. (2000), The neurobiolory of sexual functioning. Archiues of General Psychiatry, 57, 1012-1032. Mick, T. M. E¿ Hollander, E. (2006), Impulsive-compulsive sexual behavior. CNS Specmlmt lI, 944-955. Molsberry, R. L., Carr, B. R. & Mendelson, C. R. (1982), Human chronic gonadotropin binding to human fetal tests as a function of gestational age. Journal of Clinical Endouinologt and Metabo' lism, 55,79I-794. Mong, J.,4.., Glaser, E. E¿ McCarthy M. M. (1999), Gonadal steroids promote glial differentiation and alter neuronal morphology in the developing hypothalamus in a regionally specific manner. f Neurosci., 19, 1464-147 2. Montano, M. M.,'Welshons,'W:V S¿ Saal, F. S. (1995), Free stradiol in serum and brain uptake of estradiol during fetal and neonatal sexual differentiation in female rats. BioL Reprod.,53,1198-1207. Moreno Esteban, 8., Pérez- Méndez, L., Pato-Castel, I. E¿ Tresguerres, J. A.F. (1983), Respuesta de glucoronato de testosterona a di- ferente dosis de estimulación con HCG. End.orinologla,30, 129. O'Rahilly, R. (1983), The timming and sequence of events in the development of the human reproductive system during the embryonic period proper. Anøtomy and Embriologlt, 166,247-261. Ojeda, S. R. (2002), Pubertad normal, en Pombo, M. (ed.), Ti'atado Referencias bibliogr:íûcas t2r Oomura, Y., Aon, S., Koyama, Y., Fujita, I. E¿ Yoshimatsu, H. (1989), Central control of sexual behavior. Brain Rel Bull.,20,863-870. Osterlund, M. (2000), Targeted destabilization of HY5 during lightregulated development of Arabido psis. Narure, 40 5 (67 85), 462. Penson, D. F. (1996), Androgen and pituitary control of penile ni- tric oxide synthase and erectile function in the rat. BioL Reprod., 55,567-74. Peter, M. R., Clare, J. F. & Cornelis, P M. (n. d), Series: Sexual function in men andwomenwith neurological disorder. The Lancet, 369, 512-525. Pf"fi O. & Schwartz-Giblin, S. (1988), Cellular mechanisms of female reproducdve behaviors, en E. Knobil, & J. Neil (ed.), Zúø Plrysio log of Reproduction. Ptra4 O.'W & Sakuma,Y. (1979), Facilitation of the lordosis reflex of female rats from the ventrolmedial nucleus of the hypothalamus. J. Physiol., 288, 189-202. Phillip, M. &Lazar, L. (2005), Precocious puberty: growth and genetics. Hormone Researclt, 64 Suppl256-61. Rabinovici, J. E¿J"ff., R. B. (1990), Development and regulation of Growth and differentiated functiom in human and subhuman primate fetal gonads. Endncrine Reuieus, ll, 532-557. Raisman, G. & Field, P M. (1973), Sexual dimorphism in the neuro- pil area ofthe rat and its dependence on neonatal androgen. Brain Res.,54,I-29. Rako, S. (1996), La hormona del deseo: Cómo mantener mantener la líbido femenina más allá de la menopausia. Girona: Tikal Ed. Rees, P, Fowler, C. E¿ Maas, C. (2007), Sexual function in men and women with neurological disorders. La.ncet, 369 (9560), 512525. Reid, R., Karim, R., McCory W: & Bruce, N. (2010), Carpenter SelÊReported Differences on Measures of ExecutiveFunction and de Endocrinologla Pediáticø (3ra ed., pp. 669-80), Madrid: Mc- Hypersexual Behavior in a Patient and Community Sample of Graw-Hill, Interamericana. Men. International Journal of Neuroscience, 120, 120-127 _....-r t22 Neuropsicologfa de la sexualidad Reiter, \ø J. (2001), Evidence of estroigen receptors on human corpus cavernosus, Int. I. I*P. Res.,13,4. Rey, R., Schleider, G. & Josso, N. (1995), Anti-Müllerian hormone secretion during fetal and postnatal human testicular development. Frontiers in Endocronology, I0,37-47. S.D. Ec Faiman, C. (1973), Studies on human Reyes, F. I., \Winter, J. sexual development. Journal of Clinical Endncrinolngy anà Metabolisrn, 37,74-78. Referencias bibliográfi cas 123 Spinella, M. (2004), Neurobehavioral correlates of impulsivity. Internati onal Journa I of Neuros c ience, I | 4(I), 9 5 -I 0 4. Stein, D., Ives-Deliperi, V & Thomas, K. (n.d), Psychobiology of mindfulness . Cns. Specnams, 13 (9), 7 52-7 56. Swaab, D. F. Ea Hofman, M. A. (1990), An enlarged suprachiasmatic nucleus in homosexual men, en G. Einstein (ed.), Sex ønd the bra- in (pp.713-720), Cambridge, MA US: MIT Press. Swaab, D. F,, Chung,\ø C.J., Kruijver, F. PM., Hofman, M. A. & Is- Rivier, C., Rivier, J. & Vale, \(i. (1936), Inhibin mediated feed-back control of FSH secretion in the female rat. Science, 234,205. Robbins, T & Everitt, B. (1996), Neurobehavioural mechanisms of Swaab, D. F., Fliers, E. & Partiman, T (1985), The suprachiasmatic reward and motivation. Cunent Opinion in Neurobiology, 6(2), nucleus of the human brain in relation to sex, age and dementia. 228-236. Rodrþez Poyo-Guerrero, Pato-Castel, I. & Tlesguerres, J. A.F. Brain Research, 342, 37 -44. Swaab, D. F., Grundke-Iqbal, I.,Iqbal, K., trGemer, H. P H., Ravid, R. E¿ Van de Nes, J.A. P (1992), Tâu and ubiquitin in the hu- (1989), Posibilidades en el diagnóstico de los hipogonodismos hipogonadotró picos. Ana les Medicina Intema., 6, 598-602. hunina, T A. (2001), Structural and functional differences in the human hypothalamus. Hortnones and Behauior, 40,93-98. man hypothalamus in aging and Alzheimer's disease. Brain Re- Saad, F. (2004), Round Täble: The new frontiers in Sexual Medicine: DHEA un mae and female function. f Sex. Med., 1 (1), 14. Swaab, D. F., Hofman, M.,{., Lucassen, P. J., Salehi, A. E¿ Uylings, Samenow, P (2010), A biopsychosocial Model of Hipersexual Disor- H. B. M. (1994), Neuronal atrophy not cell death is the main der/ Sexual Addition. Sexual Addiction Ò Compul"siuity, 17 , 69-8L . Sánchez-Criado, J. E. (1995), Fisiología del ovario, en J. A. F. Tiesguerres (ed.), Fisiología humøna. Madrid: McGraw-Hill/ Intaramericana. Schumacheç M., Corini, H., McEwen, B. S. (1989), Regulation of high affiniry GABAA-receptors in specific brain regions by ovarian hormon es. Neuro endo crino logy, 50, 3 | 5 -32O. Simerþ R. B. (2002),'Wired for reproduction: Organization and Development of Sexually Dimorphic Circuits in the Mammalia¡r Forebrain. Annual Reuiew of Neuroscience,25 (l),501. Sinclair, A. S., Berta, P, Palmet, M. S., Hawkins, J. R. & Foster, J. \ø. (1991), A gene from the human sex-determining region encodes a protein with homology to a conserved DNA- binding motif . Nature, 346, 240-244. searclt, 590,239-249. hallmark of Alzheimer's disease. Neurobiol Aging, 15, 369-371 . Symons, D. (1979), The euolution of human sexuølity. Oxford, UK: Oxford University Press. Tiaish, A. M., Park, K., Dhir, V., Kim, N. N., Moreland, R. B. E¿ Goldstein, I. (1999), Effects of castration and androgen replacement on erectile function I a rabbit model. End.ocrinologt, 140, 1861-1868. Tian, D., Picard, J.Y. S¿ Campargue, J.0987),Inmmunocitochemical detection of anti-Mtillerian hormone in Sertoli cells of various mammalian species including human. Joumøl of Histochernisny and Cytochemistri, 35, 7 33-7 43. Tfesguerres, J. A.F. (1989), Reproducción I. Eje hipotálamo hipofiso testicular, en J. A. F. Ti:esguerres (ed), Fisiologia endocrina. Ma- drid: Eudema. r24 Neuropsicología de la sexualidad Astrolabio Tiesguerres, J. A.F. (1989), Reproducción II. Eje hipotálamo hipofiso ovárico, en J. A. F. Tiesguerres (ed), Fisiologia endncrina. Ma- drid: Eudema. Vountilainen, R. E{ Miller, \ø L.(1988), Human Müllerian inhibitory factor messenger ribunocleic acid is hormonally regulated in the fetal testis and in adult granulose cells. Molecular Endncrinolog!, 1,604-608. Voutilainen, R. (1992), Differentation on the fetal gonad. Hormone Research, 38, 66-7l. W'eller, K. L. S¿ Smith, D. A. (1982), Afferent connections to the bed nucleus of the stria terminalis. Brain Research,232,255-270. 'Wisden, W, Laurie, D. J., Monyer, H., Seeburg, P H. (1992), The distribution of 13 GABAA receptor subunit mRNAs in the rat brain. I. Têlencephalon, diencephalon, mesencephalon.l Neurosci., 12, 1040-1062. Yen, S. S.C. &Jatre, R. B. (1991), Reproductiue endocrinologlt Filadelfia: Saunders. Zhou,J., Hofman, M. & Swaab, D. (1995), No changes in the number of vasoactive intestinal poþeptide (MP)-expressing neurons in the suprachiasmatic nucleus of homosexual men; comparison with vasopressin-expressing neurons. Brain Research, 672(l-2), 285-288. SALTJDY MEDICINÄ Deporte para todos / Jörg Stäuble Conozca su diabetes (3.'edición) / Emilio Moncada Lorenzo La enfermedad epiléptica i Francisco Abad Alegría Dormir mejor. Causas y tratamiento del insomnio / Luis María Gonzalo El buen hacer médico / David Mendel Comentarios al Código de Ética y Deontología Médica (2.' edición) / Gon zaloHenanz Psicoterapia brisica / Richard Parry Muerte cerebral. Biología y ética / Jesús Colomo Gómez SIDA: Àspectos ético-médicos / Juan Moya y Femando Mora Reflexoterapia: Bases neurológicas / Luis Maía Gonzalo Homosexualidad y esperanza. Terapia y curación en la experiencia de un psicólogo (5." edición) / Gerard van den Aardweg Antropología del dolor, Sombras que son luz / Johan¡es Vilar i Pla¡as de Famés La verdad sobre los tranquilizantes / Rafael Montoya Sáenz El sueño, los sueños, un mundo miste¡ioso, Los ritmos naturales de la vigilia y del descanso, los más frecuentes trastornos de la noche, las conquistas de la medicina del sueño / Elio Lugares y Luciana Omicini hacer, qué debe evitarse? / Michele Lombardo epto y fundamenûo / José lipez Guzmán y Ángela Aparisi Miralles Cercbro y afectividad / Ma¡ía Gudín Romper el círculo vicioso, Salud intestinal mediantela ilieta.Di¿tas para Ia enfermedad de Crohn, la colitis ulcerosø, b diverticulítìs,Ia enfermedad celíaca,la fibrosis quístíca y la dfuneø crónìca (3." edición) / Elaine Gottschall Teoría elemental de la gastronomía / Juan Cruz Cnrz Intervención dietética en la obesidad ( I .' reimpr.) / Giuseppe Russolillo, Icíar Astiasarán, J. Alfredo Martínez Consejos médicos para la tercera edad / Eduardo Alegría, Luis María Gonzalo, Juan Luis Guijarro, Jesús Ibríñez, Emilio Quinønilla, Jesús Repáraz,Ricado Tapata Cefaleas / Pablo Irimia Sieira, Edua¡do Ma¡tínez Vila La ansiedad. Un enemigo sin rostro / Javier Schlatter Navarro Comer bien a cualquier edad / J. Alfredo Martínez, Susana Santiago, M. Iosune Zubieta Depresión y enfermedad bipolar en niños y adolescentes / César Soutullo Esperón Ejercicio y calidad de vida. Claves para mantener la salud mental y fisica / Luis María Gonzalo (Coord.). ¿Sabemos realmente qué cornemos? Alimentos transgénicos, ecológicos y funcionales / Amelia A. Marti del Moral y J. Alfredo Martínez Hemández (Eds.) Cómo prevenir y curar lesiones deportivas / Alfonso del Corral, Francisco Forriol Campos, Javier Vaquero Martín Comprendiendo Ia homosexualidad (3." ed.) / Jokin de Irala Sin miedo. Cómo afrontar la enfermedad y el linal de la vida (2." edición) / Miguel Angel Monge Esquizofrenia / Felipe Ortuño Aborto y contracepción / Guillermo lópez Health Institutions at the Origin of the Welfare Systems in Europe / Edited by Pilar l-eón Sanz Elementos de dieúoterapia / Aranøa Ruiz de las Heras y J. Alfredo Martínez Ingestas Dietéticas de Referencia QDR) para la población española / Federación Española de Sociedades de Nutrición, Alimentación y Dietética (FESNAD) Ser felices sin ser perfectos. Estrategias de cambio para un anancástico (2." edición) / Javier Schlatter NavaÍo Alimentación, ejercicio fisico y salud / Diana Ansorena y J. Alfredo Martínez Aìimentación y deporte / Javier Ibáñez Santos e Icia¡ Astiasarán Anchía
 0
0
Anuncio
Documentos relacionados



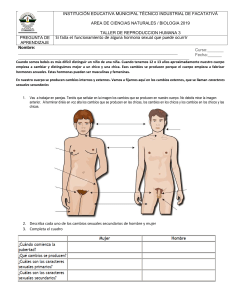




Descargar
Anuncio
Añadir este documento a la recogida (s)
Puede agregar este documento a su colección de estudio (s)
Iniciar sesión Disponible sólo para usuarios autorizadosAñadir a este documento guardado
Puede agregar este documento a su lista guardada
Iniciar sesión Disponible sólo para usuarios autorizados