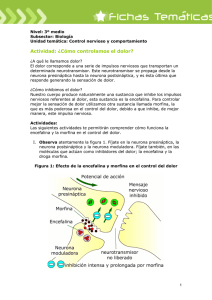
1. INTRODUCCIÓN El término sinapsis fue introducido por Sherrington en 1897 a partir del griego sinapsis, “cerrar”, curiosamente antes de que se conociera con precisión la base estructural de la interacción entre neurona-neurona (debate entre Golgi y Cajal). Sherrington se posicionó a favor de Cajal, y explicó que, aunque la neurona es visible con una apariencia continua, en el punto en que esta se une a otra neurona no es posible demostrar esa continuidad, pudiendo haber ahí otro tipo de transmisión: la sinapsis. La sinapsis es la unión funcional entre neuronas. El proceso de comunicación entre neurona presináptica y la postsináptica. Sherrington recibió el premio nobel de medicina y fisiología en 1932. Introdujo el término de sinapsis. Comprendió que una característica fundamental del sistema nervioso reside en su función integradora (el propio sistema «integra» la información disponible y la «discrimina» de acuerdo con sus necesidades, para así emitir la respuesta adecuada). Además, se percató de que no todas las sinapsis son de naturaleza excitadora, sino que la mayoría exhibe cualidades inhibidoras, frenando la iniciación de los potenciales de acción Se podría decir que Cajal descubrió cómo era el sistema nervioso y que Sherrington explicó cómo funciona. 2. LA SINAPSIS - Definición. La sinapsis es una zona estructural y funcionalmente especializada de la neurona a través de la cual la información pasa de una neurona a otra, o de una neurona a una célula efectora. La sinapsis es, físicamente, la zona de interacción entre una primera neurona y una segunda neurona. Por su parte, la transmisión sináptica es el proceso de comunicación entre neuronas o entre células. Normalmente, la información en las sinapsis se transfiere en un único sentido, lo que hace que haya una neurona presináptica (que envía la información) y una neurona postsináptica (que es la que recibe la información). Entre la neurona presináptica y la postsináptica suele haber un espacio físico más o menos grande que se denomina espacio sináptico o hendidura sináptica. Las sinapsis permiten que las neuronas se activen unas a otras, que se inhiban y, en suma, posibilitan que la actividad neuronal pueda ser modulada. Es decir, si hay un exceso, inhibiendo y si no existe excitación, excitando. Es conveniente poner de relieve que la modificación de la función biológica mediante fármacos y drogas actúan en la sinapsis, modificando la neurotransmisión química entre neuronas en el SNC o SNP. Por ejemplo, la depresión se produce por un déficit de serotonina. Los neurotransmisores van a hacer que haya más serotonina en la sinapsis, modulando así la actividad neuronal. La cocaína, por ejemplo, tiene un exceso de dopamina en la sinapsis. Se calcula que cada neurona establece, aproximadamente, unas 1000 conexiones sinápticas con otras neuronas, o hasta 100.000 conexiones. Estas conexiones son la base de nuestras capacidades cognitivas, de que podamos aprender, memorizar, hablar, etc. Además, son uno de los principales factores que contribuye a las diferencias interindividuales en relación con esas capacidades. Por ejemplo, las diferencias de conexiones entre personas celosas y no celosas debido a su experiencia, aprendizaje, educación, etc. Es decir, nuestro cerebro es plástico y podemos modificarlo. Las drogas, conexiones y la neurogénesis son los tres tipos de cambios en la plasticidad cerebral. El cambio es duradero cuando se produce a nivel de conexiones, por ejemplo, llevar muchos años sin estudiar, le cuesta volver a estudiar. Puesto que cada neurona establece sinapsis con muchas otras neuronas, la transmisión de la información puede seguir un proceso convergente o divergente. • La divergencia ocurre cuando un único botón terminal establece conexiones con más de una neurona. Esta organización sináptica está diseñada para que la información recogida, por ejemplo, por un único receptor sensorial, se transmita y distribuya a amplias regiones del cerebro. • El diseño convergente se produce cuando una neurona recibe información de varios botones terminales, es decir, de varias neuronas. Esta organización sináptica se produce cuando es necesario que una determinada célula reciba la suma de la información de un gran número de neuronas. La información también puede transmitirse de manera simple: una neurona envía información a otra neurona. *Cuando el cerebro envía diferentes acciones a diferentes órganos se genera una transmisión de corriente divergente. * Cuando al cerebro le llegan respuestas de estímulos se genera una transmisión de corriente convergente. - TIPOS DE SINAPSIS Hasta la primera mitad del siglo XX se creía erróneamente que la comunicación entre neuronas se basaba exclusivamente en señales eléctricas. Sin embargo, la mayoría de nuestras sinapsis es de tipo químico (neurotransmisores). La primera evidencia de la existencia de moléculas transmisoras fue aportada por Otto Loewi en 1921, cuando demostró en las ranas que la inhibición de la actividad cardíaca por estimulación de nervio vago estaba mediada por la liberación de una sustancia transmisora, la acetilcolina. Primera clasificación atendiendo a criterios funcionales generales: - Sinapsis eléctrica: los iones y pequeñas moléculas fluyen directamente de la neurona presináptica a la neurona postsináptica a través de unas proteínas, canales intercelulares, que dan lugar a una unión física, formando uniones en hendidura (gap junctions). Al pasar de una neurona a otra a través de la sinapsis eléctrica, la señal se puede atenuar ligeramente, pero es capaz de continuar y afectar de una forma u otra a la neurona postsináptica. Como la corriente eléctrica pasa directamente desde la célula presináptica a la postsináptica, sin ningún paso intermedio, la transmisión se produce con menos retraso que en la química. Dada su mayor velocidad de transmisión, es apropiada cuando se requiere la sincronización de la actividad de amplios grupos de células nerviosas o para la transmisión rápida de señales como en el musculo cardiaco. Sin embargo, no puede ser modulada ni regulada. – Sinapsis química: Cuando llega un impulso nervioso, se libera un neurotransmisor que viaja por la hendidura sináptica (no hay unión física) y se une a un receptor (transporte activo dependiente de ligando). En el sistema nervioso solo hay sinapsis de tipo químico. La sinapsis química ofrece mayor flexibilidad que la transmisión eléctrica al permitir una modulación o regulación de la señal, ya que puede ser ampliada o inhibida. Las sinapsis también se pueden clasificar, siguiendo criterios funcionales: A) De acuerdo con el tipo de neuronas implicadas en la sinapsis: - Neurona - neurona: son las sinapsis típicas del SNC. Ambas células son neuronas. - Neurona - célula muscular: son las sinapsis típicas de la unión neuromuscular (contracción o relajación de los músculos). En el sistema nervioso periférico. Una neurona inerva a una célula muscular. - Neurona - célula secretora: sinapsis típicas del sistema neuroendocrino. Una neurona sinapta con un tipo celular específico (célula postsináptica) encargado de la liberación de alguna sustancia química, por ejemplo, una hormona. B) De acuerdo con los efectos postsinápticos generados: - Sinapsis excitadoras: la transmisión de la señal desde la neurona presináptica a la postsináptica produce una despolarización en la neurona postsináptica (mayor facilidad para generar un potencial de acción). - Sinapsis inhibidoras: la transmisión de la señal desde la neurona presináptica a la postsináptica produce una hiperpolarización de la neurona postsináptica (dificultad para generar un potencial de acción). Según su morfología: Se pueden establecer los siguientes tipos de sinapsis según la localización específica del contacto sináptico: I. II. III. SINAPSIS AXODENDRÍTICA. un botón terminal de un axón establece contacto sináptico con una o más dendritas de una neurona postsináptica. Este es el tipo más común de sinapsis en el SNC y es de naturaleza excitadora. SINAPSIS AXOSOMÁTICA. cuando el contacto se establece entre el botón terminal de un axón de una neurona presináptica y el soma de otra neurona postsináptica. Son de naturaleza inhibidora. SINAPSIS AXOAXÓNICA: el axón de una neurona presináptica establece sinapsis con un axón postsináptico. Es de naturaleza moduladora. Los astrocitos también ejercen un papel importante en los procesos sinápticos. De hecho, estas células gliales son capaces de modular la transmisión sináptica, regulando la excitabilidad neuronal, entre otras formas, por medio de la liberación de glutamato. Precisamente por la acción “a tres bandas” entre la neurona presináptica, la postsináptica y los propios astrocitos se habla de sinapsis tripartita. Además, esta acción tripartita se ha encontrado también en el sistema nervioso periférico, donde la acción “a tres bandas” la ejercen las células de Schwann. 3. RESPUESTA POSTSINÁPTICA La respuesta postsináptica en las sinapsis químicas se origina cuando un potencial de acción llega a un terminal presináptico, y desde este terminal se vierten a la hendidura sináptica las moléculas del neurotransmisor, las cuales interactúan con receptores presentes en la membrana de la neurona postsináptica, a partir de cuya interacción se abren canales iónicos (sensibles a ligando) en la membrana postsináptica, lo que conlleva una determinada corriente iónica en ella, que es la responsable de la respuesta postsináptica. El tipo particular de respuesta postsináptica depende de la naturaleza del efecto que provoque el neurotransmisor al interactuar con los receptores postsinápticos. Esta interacción “neurotransmisor-receptores postsinápticos” puede generar dos tipos de señales: excitadoras-PEP (provoca despolarización y mayor probabilidad de generarse un potencial de acción) o inhibitorias-PIP (provoca hiperpolarización y dificulta la generación de un potencial de acción). Es decir, los neurotransmisores por sí solos no son excitatorios ni inhibidores. Es realmente la unión con el receptor lo que produce que estos sean excitadores o inhibidores. 3.1. POTENCIAL EXCITADOR POSTSINÁPTICO. La interacción entre “neurotransmisores-receptores” puede implicar la apertura de canales de Na+, de modo que este entraría al interior celular y se generaría una cierta despolarización en la membrana postsináptica. A esta despolarización se le denomina Potencial Excitador Postsináptico (PEP). Este potencial es un tipo particular de Potencial Local Graduado y se propaga a lo largo de la membrana en dirección al cono axónico de forma electrotónica, donde si conserva la suficiente intensidad se dispara un potencial de acción. Si no hay intensidad suficiente para alcanzar el umbral de excitación, no se da el potencial de acción. Es un tipo de Potencial Local Graduado. 3.2.POTENCIAL INHIBIDOR POSTSINÁPTICO. - La inhibición postsináptica (PIP). La interacción entre el neurotransmisor y los receptores postsinápticos puede conllevar la apertura de canales de Cl(entrada al interior celular) o K+ (salida al exterior celular) que en cualquier caso se genera una hiperpolarización de la membrana. Esto es lo que se conoce como Potencial Inhibidor Postsináptico (PIP). También son un tipo especial de Potencial Local Graduado y se propagan electrotónicamente hasta desaparecer. Su amplitud es también proporcional a la intensidad del estímulo que lo provoca. - La inhibición presináptica (PIP). La actividad de la neurona presináptica es inhibida por una tercera neurona que establece sinapsis de tipo axoaxónica con ella. Esto provoca un descenso en la despolarización de la neurona presináptica. Como consecuencia de ello se libera menos neurotransmisor desde el terminal presináptico, lo que genera un efecto menor a nivel postsináptico. Es decir, provoca una disminución de la magnitud o amplitud del PEP en la neurona presináptica (no genera un PIP). Su acción es más duradera que la inhibición postsináptica. 3.3. INTEGRACIÓN SINÁPTICA. Un potencial de acción se produce normalmente por una suma de PEP y PIP en el cono axónico. Esto es lo que se conoce como integración sináptica. Hablamos de que un solo PEP no puede producir una despolarización lo suficientemente grande como para producir un potencial de acción y como la neurona establece miles de conexiones sinápticas lo más común es que este potencial surja por una integración y sumación de toda la información excitatoria e inhibitoria en el cono axónico. Esta integración sigue el siguiente esquema: DESPOLARIZACIÓN FINAL = PEPs (pep1+pep2+pep3+...) - PIPs (pip1+pip2+pip3+...) Esta integración sináptica puede ser de dos tipos: espacial o temporal. -En la sumación espacial el cono axónico de una neurona está recibiendo PEPs de sinapsis diferentes, que provienen de neuronas diferentes, al mismo tiempo. A partir de esta sumación espacial de PEPs en el cono axónico, y de su despolarización final resultante, se incrementa o se reduce el número de potenciales de acción que se disparan. Cuando el valor de la despolarización es alto, los potenciales de acción se disparan a modo de trenes o ráfagas. -En la sumación temporal, solamente tenemos una sinapsis, es decir, una única neurona nos está enviando información en tiempos diferentes en el mismo punto. Se produce un fenómeno de cabalgamiento, que es que se va cabalgando un potencial sobre otro, y se van agregando entre ellos. Un segundo PEP tiende a agregarse al primero. En este sentido, cuanto más breve sea el intervalo entre ambos PEPs mayor será el nivel de “cabalgamiento” del segundo PEP sobre el primero. Este fenómeno de agregación de un PEP sobre otro puede llegar a integrarse de tal forma que, finalmente, se aumente el valor final de la despolarización, de tal forma que aumente el número de potenciales de acción generados bajo ese tipo de estimulación. 4. MECANISMOS DE NEUROTRANSMISIÓN QUÍMICA La dinámica general de la neurotransmisión en la sinapsis química es la siguiente: la llegada de un potencial de acción al terminal sináptico hace que se liberen las moléculas del neurotransmisor (almacenadas en el botón terminal) a la hendidura sináptica. Tras liberarse al espacio sináptico, las moléculas del neurotransmisor interactúan con receptores de membrana ubicados en la superficie de la neurona postsináptica y, según esa interacción, se abren canales iónicos de ligando (unión del neurotransmisor). Dependiendo del tipo de neurotransmisor liberado y del tipo de receptor, se abrirán canales para una especie iónica u otra, que generará un PEP o un PIP en la neurona postsináptica. Es decir, lo que pasa a las neuronas postsinápticas son siempre iones, nunca pasa el neurotransmisor. 4.1. ALMACENAMIENTO Y LIBERACIÓN DEL NEUROTRANSMISOR Los neurotransmisores se encuentran almacenados en vesículas que se encuentran en el terminal sináptico. En la superficie de estas vesículas hay una proteína transportadora que capta e introduce al neurotransmisor, previamente sintetizado, dentro de esta. Las vesículas no están distribuidas de manera uniforme por el interior del terminal presináptico, sino que tienden a agruparse en las zonas más activas del mismo, preparadas para ser liberadas. Estas vesículas también se encuentran enganchadas en el citoesqueleto, en los microtúbulos y microfilamentos que, recordemos permitían el paso de moléculas, gracias a la proteína llamada sinapsinas I. La liberación del neurotransmisor requiere que las vesículas se fundan con la membrana presináptica por exocitosis, por medio de un poro de fusión, lo que lleva a que el contenido de la vesícula se vierta al exterior. Pero, previamente, las vesículas han de soltarse del citoesqueleto. Este proceso de fusión es dependiente de los canales iónicos de calcio, que se encuentra mayor concentrado en el exterior celular. Cuando llega el potencial de acción al botón terminal se produce una despolarización y se abren los canales de calcio (sensibles al voltaje) que entra masivamente al interior impulsado por el gradiente electroquímico, provocando una fosforilación de las proteínas sinapsinas I. Esto permite que la sinapsina I suelte las vesículas que estaban unidas en el citoesqueleto y se fusionen a la membrana mediante un proceso de exocitosis. Como consecuencia, se vierte el neurotransmisor al exterior y la superficie de la membrana aumenta. A medida que se funden más vesículas, aumenta la superficie de la membrana. Este incremento es corregido mediante un proceso de endocitosis, volviendo al botón terminal. Tras este proceso las vesículas vuelven a ser de nuevo funcionales, pudiendo contener de nuevo un neurotransmisor o pueden degradarse. Antes de soltarse del citoesqueleto las vesículas se llaman inmaduras, cuando ya se sueltan, maduras. A mayor número de potencial de acción esté llegando al terminal sináptico, mayor cantidad de calcio que entra al terminal sináptico, mayor número de vesículas que se fusionan con la membrana y mayor cantidad de neurotransmisor liberado y mayor es la magnitud de la respuesta postsináptica. 4.2.INACTIVACIÓN DEL NEUROTRANSMISOR Una vez que el neurotransmisor liberado llega al receptor, éste reconoce al neurotransmisor e interactúa con él, acoplándolo. Debido a esta interacción, se abren canales iónicos dependientes de ligando y pasan los iones a la neurona postsináptica. Una vez el neurotransmisor haya realizado su función con el receptor es necesario que se desacople de este y sea retirado del espacio sináptico, puesto que, si no, podría volver a interaccionar con el receptor, provocando otra estimulación. Para retirar e inactivar al neurotransmisor existen dos mecanismos: - Degradación enzimática: consiste en la destrucción física de las moléculas del neurotransmisor, un proceso llevado a cabo por enzimas específicas. Estas se ubican en el espacio sináptico o bien en el interior del terminal presináptico. - Recaptación: este es un proceso de transporte activo (consume energía) llevado a cabo por proteínas transportadores ubicadas en la membrana del terminal presináptico, que recaptan e introducen los neurotransmisores en el interior del botón. Una vez dentro, el neurotransmisor puede ser reintroducido en vesículas y volver a cumplir su función, es decir, se ha reciclado; o degradado. 4.3.RECEPTORES POSTSINÁPTICOS Los receptores postsinápticos son las proteínas que nos encontramos en la parte externa de la membrana de la neurona postsináptica y el lugar de interacción con el neurotransmisor. El tipo de canales que se abren no sólo depende del neurotransmisor, sino también del tipo de receptor. Es un transporte activo de ligando. Tenemos dos tipos de receptores postsinápticos: inotrópicos o metabotrópicos - Los receptores inotrópicos. Estos receptores son aquellos en los que el receptor está acoplado directamente a un canal iónico, formando un complejo receptor-canal, lo que hace que la unión del neurotransmisor al receptor suponga la inmediata apertura del canal. - Los receptores metabotrópicos. Estos receptores son aquellos que están acoplados a sistemas de segundos mensajeros, es decir, no están directamente acoplados a un canal iónico, sino de manera indirecta a través de un dispositivo iónico que genera el segundo mensajero. En estos receptores, el neurotransmisor actúa de primer mensajero y otra molécula (AMPc o IP3) hace la función de segundo mensajero. Pero para que este segundo mensajero “nazca” deben de producirse previamente una serie de cambios. Los receptores metabotrópicos están acoplados a la proteína G, compuesta por tres subunidades (alfa, beta y gamma), además de una molécula de GDP asociada a la unidad alfa. Una vez que el ligando se une al receptor se produce un cambio en la estructura de la proteína G, de tal forma que la subunidad alfa se desvincula de las otras dos unidades y se une y activa a una proteína reguladora (adenilato ciclasa) que se encuentra insertada en la membrana celular. A partir de esa unión (subunidad alfa y adenilato ciclasa) se ponen en marcha una serie de reacciones bioquímicas en el interior celular que desembocará en la liberación de un segundo mensajero. ➢ Receptor metabotrópico asociado al AMPc. En primer lugar, se ha de mencionar todos los ingredientes que intervienen en este proceso: receptor acoplado a una proteína G (con tres subunidades alfa, beta y gamma siempre presentes), Guanosín Difosfato (GDP) (porque tiene dos grupos fosfatos) acoplado a la subunidad alfa, Guanosín Trifosfato (GTP), Proteína Adenilato Ciclasa (incrustada en la membrana), Adenosín Trifosfato (ATP) y Proteína Quinasa A. La transducción de señales mediada por AMP cíclico a partir de un neurotransmisor es de la siguiente manera: una vez que el neurotransmisor se une al receptor correspondiente, se activa la subunidad alfa de la Proteína G, de manera que el Guanosín Difosfato (GDP) acoplado a la subunidad alfa y se cambia por un Guanosín Trifosfato (GTP) (se añade un grupo fosfato). Este guanosín trifosfato junto con la subunidad alfa viaja hacia a la Proteína Adenilato Ciclasa y la activa. Esta activación convierte una molécula de ATP en Adenosín Monofosfato (AMPc). El AMPc interactúa con la Proteína Quinasa que, a su vez, interactúa con los canales iónicos y ésta los abre. ➢ Receptor metabotrópico asociado a IP3. De igual manera, en primer lugar, mencionamos los ingredientes que intervienen en este proceso: un receptor acoplado a una proteína G (con sus tres subunidades), en la subunidad alfa una molécula de Guanosín Difosfato (GDP) (que se convertirá en Guanosín Trifosfato), una Enzima Fosfolipasa C, Proteína Quinasa C (PKC), un Retículo Endoplasmático (con mucho calcio), y Fosfatidilinositol (IP2). Una vez que el neurotransmisor se une a su receptor, se produce un cambio en la proteína G: la subunidad alfa se disocia de las otras subunidades, igual que en el caso anterior, y el guanosín difosfato se convierte en guanosín trifosfato. Este viaja con la subunidad alfa hasta la enzima fosfolipasa C, a partir de esta unión se activa como una tijera cortando el fosfatidilinositol (IP2) convirtiéndolo en: inositol/IP3 + diacilglicerol (DAG); estas dos moléculas actúan como segundos mensajeros. El IP3, por su parte, viaja hasta al retículo endoplasmático y se une al canal dependiente de calcio que se abre. De este modo, el Ca2+ sale y viaja hacia la proteína quinasa C (PKC). El DAG también es capaz de interactuar con la proteína quinasa C. Esta quinasa C con el calcio y el DAG consigue abrir el canal. 4.4. RECEPTORES PRESINÁPTICOS Los receptores presinápticos son proteínas acopladas en la membrana de la neurona presináptica, que responden ante la presencia de un neurotransmisor específico. Su principal función es modular, es decir, producir cambios en el voltaje, cuando este no está siendo adecuado. Esta modulación es casi siempre de tipo inhibitorio El mecanismo de acción de los receptores presinápticos es muy similar al que muestran los receptores postsinápticos: su activación mediante la interacción con su correspondiente ligando supone la apertura o cierre de determinados canales iónicos, modificando la permeabilidad iónica de la membrana. Además, la activación de los receptores presinápticos puede suponer la liberación de segundos mensajeros. Existen dos tipos: - Autorreceptores: responden al neurotransmisor liberado desde el mismo terminal en el que ellos se encuentran para evitar un gasto excesivo del neurotransmisor en las sinapsis, que podría provocar un agotamiento de sus reservas, y proteger a la membrana postsináptica de la sobreexcitación. Es una retroalimentación negativa. Siempre son inhibitorios, corrigen la sobreexcitación enviando PIPs. - Heterorreceptores: en este caso el control lo ejerce una tercera neurona que establece una sinapsis axoaxónica sobre el propio terminal presináptico, desde donde se libera un neurotransmisor (diferente al que se libera en la sinapsis principal) que activa a los heterorreceptores. Estos pueden actuar generando tanto inhibición como activación: 1) Inhibición presináptica (repolarización: más negativa): cierre de canales de Ca ++ y apertura de canales de K+; apertura de canales de Cl-, bloqueo directo de Ca++ 2) Facilitación presináptica (despolarización, más positiva): incremento en la entrada de Ca++ al terminal con el cierre de canales de K+. Esto mantendría y mejoraría la despolarización de la membrana, a partir de lo cual la entrada de Ca++ al terminal sería más amplia, y esto facilitaría la liberación de neurotransmisor al espacio sináptico