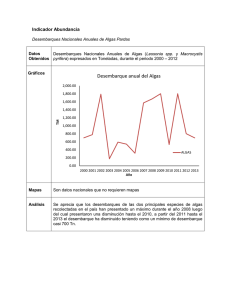
Protistas Wim van Egmond/Visuals Unlimited, Inc. 26 CONCEPTOS CLAVE 26.1 Los protistas son un grupo diverso de organismos eucariotas que varían en plan corporal (unicelular, colonia, cenocítico, multicelular), método de motilidad (seudópodos, cilios, flagelos), tipo de nutrición (autótrofos, heterótrofos) y modo de reproducción (asexual, sexual). 26.2 Mucha incertidumbre rodea la evolución eucariota, pero los organelos eucariotas como los cloroplastos probablemente descienden de células engullidas que sobrevivieron y se convirtieron en organelos. La evidencia científica actual apoya la división de los protistas y otros eucariotas (plantas terrestres, hongos y animales) en cinco “supergrupos” informales. 26.3 Los excavados son protistas unicelulares con mitocondrias atípicas enormemente modificadas; obtienen su nombre porque muchas tienen un surco oral profundo, o excavado. 26.4 Los cromalveolados son un supergrupo de protistas diversos que pudieron originarse como resultado de endosimbiosis secundaria en la que una célula ancestral engulló una alga roja. 26.5 Los rizarios son un supergrupo de células ameboides que con frecuencia tienen testas (conchas) a través de las cuales se extienden proyecciones citoplasmáticas; foraminíferos, actinópodos y ciertas amebas sin concha son rizarios. 26.6 Los arqueplástidos incluyen algas rojas, algas verdes y plantas terrestres, todas las cuales tienen plástidos limitados por membranas externas e internas. 26.7 Los unicontos incluyen amebozoos y coanoflagelados, así como hongos y animales. Micrografía óptica (MO) de una diatomea marina. La diatomea ampliamente difundida Guinardia striata forma colonias compuestas de cadenas de células en espiral. os protistas son un grupo informal de organismos eucariotas principalmente acuáticos con diversas formas corporales, tipos de reproducción, modos de nutrición y estilo de vida. Los protistas, que incluyen algas, oomicetos, mohos mucilaginosos y protozoarios, son organismos unicelulares, coloniales o multicelulares simples que tienen una organización celular eucariota (vea la fotografía). La palabra protista, del griego “lo primerísimo”, refleja la idea de que los protistas fueron los primeros eucariotas en evolucionar. Los protistas son miembros del dominio Eukarya, el tercer dominio en el árbol de la vida. (Recuerde los tres dominios presentados en la figura 23-2). Las células eucariotas son características de los protistas, así como de los complejos organismos multicelulares que pertenecen a los reinos Fungi, Animalia y Plantae. Sin embargo, el tener una estructura celular eucariota diferencia con claridad los protistas de los miembros de los dominios Bacteria y Archaea. Recuerde del capítulo 4 que, a diferencia de las células procariotas, las células eucariotas tienen núcleos y otros organelos encerrados en membrana, como mitocondrias y plástidos, 9 + 2 flagelos, y múltiples cromosomas en los que ADN y proteínas forman un complejo llamado cromatina. La reproducción sexual, la meiosis y la mitosis también son características de los eucariotas. Hasta recientemente, todos los protistas se clasificaron como el reino Protista, pero esta clasificación fue por defecto: los protistas incluían a cualquier eucariota que no fuera planta terrestre, hongo o animal. Principalmente los datos moleculares recientes, han clarificado muchas relaciones evolutivas entre eucariotas, el reino Protista ya no se reconoce como clado. Aunque se han logrado grandes avances en la comprensión de las relaciones evolutivas entre protistas, muchas relaciones siguen siendo inciertas. L 537 26_Cap_26_SOLOMON.indd 537 17/12/12 10:21 En la actualidad, los biólogos reconocen docenas de taxones protista, que, junto con otros eucariotas, se clasifican en cinco “supergrupos”. La consideración de todos los taxones protista está más allá del ámbito de este libro, pero se estudiarán varios ejemplos representativos de cada supergrupo y se proporcionarán elementos de comprensión acerca de cómo pudieron evolucionar tan diversos eucariotas. 26.1 DIVERSIDAD EN LOS PROTISTAS OBJETIVO DE APRENDIZAJE 1 Discutir en términos generales la diversidad inherente en protistas, incluidos medios de locomoción, modos de nutrición, interacciones con otros organismos, hábitats y modos de reproducción. Debido a su enorme número, los protistas son cruciales para el equilibrio natural del mundo viviente. Los protistas son una fuente importante de alimento para otros organismos, y los protistas fotosintéticos suministran oxígeno a los ecosistemas acuático y terrestre. Ciertos protistas son económicamente importantes, y otros producen enfermedades devastadoras como la malaria. El plan corporal varía considerablemente entre protistas. La mayoría de los protistas son unicelulares, y cada célula forma un organismo completo capaz de realizar todas las funciones características de la vida. Algunos protistas forman colonias, grupos de células débilmente conectadas; algunas son cenocíticas, y consisten en una masa multinucleada de citoplasma; y algunas son multicelulares, compuestas de muchas células. A diferencia de los animales, plantas y muchos hongos eucariotas, la mayoría de los protistas multicelulares tienen formas corporales relativamente simples sin tejidos especializados. El tamaño y la complejidad estructural no son las únicas características variables de los protistas. Durante el curso de su larga historia evolutiva, los protistas evolucionaron diversidad en sus medios de locomoción, formas de obtener nutrientes, interacciones con otros organismos, hábitats y modos de reproducción. Los protistas, cuya mayoría son mótiles en algún punto de su ciclo de vida, tienen varios medios de locomoción. Algunos se mueven al empujar extensiones citoplásmicas (seudópodos) a lo largo del borde frontal y retraer el citoplasma que arrastran detrás, como hace una ameba. Otros protistas se mueven al flexionar células individuales; al deslizarse sobre superficies; al ondular cilios, cortos organelos parecidos a cabellos; o al azotar flagelos, largos organelos parecidos a látigos. Algunos protistas tienen dos o más medios de locomoción; por ejemplo, tanto flagelos como seudópodos. Los métodos de obtención de nutrientes difieren ampliamente entre los protistas. La mayoría de las algas son autótrofas y fotosintéticas como las plantas (FIGURA 26-1a). Algunos protistas heterótrofos obtienen sus nutrientes mediante absorción, como los hongos, mientras que otros parecen animales en cuanto a que ingieren alimento. Algunos protistas cambian sus modos de nutrición y son autótrofos en ciertas épocas y heterótrofos en otras. Aunque muchos protistas son de vida libre, otros forman asociaciones simbióticas estables con organismos no relacionados. Estas asociaciones íntimas varían desde el mutualismo, una asociación más o menos igualitaria donde ambos participantes se benefician; al comensalismo, donde un participante se beneficia y el otro no resulta afectado; hasta el parasitismo, donde un participante (el parásito) vive sobre o en otro (el huésped) y depende metabólicamente de él (vea el capítulo 54). Algunos protistas parásitos son importantes patógenos (agentes causantes de enfermedades) de plantas o animales. A lo largo de este capítulo se describirán ejemplos específicos de asociaciones simbióticas que involucran protistas. La mayoría de los protistas son acuáticos y viven en el océano o en corrientes de agua dulce, lagos y estanques (FIGURA 26-1b). Constituyen la mayor parte del plancton, los organismos flotantes, con frecuencia Amoeba Seudópodos Volvox Scenedesmus Chlamydomonas Pediastrum Flagelos Flagelos Euglena Spirogyra Vorticella Didinium Pared celular Flagelo Cilios Paramecium Núcleo Cloroplasto Cilios Stentor Cilios Gránulo de almidón (a) Chlamydomonas es un organismo fotosintético con dos flagelos y un cloroplasto con forma de copa. (b) Los protistas en una gota de agua en un estanque. Se muestran varios modos de locomoción. FIGURA 26-1 Protistas 538 Capítulo 26 26_Cap_26_SOLOMON.indd 538 17/12/12 10:21 microscópicos, que habitan en aguas superficiales y son la base de la red alimenticia en los ecosistemas acuáticos. Otros protistas acuáticos se fijan a rocas u otras superficies en el agua. Incluso los protistas parásitos son acuáticos, porque viven en los ambientes acuosos de los fluidos corporales de otros organismos. Los protistas terrestres están restringidos a lugares húmedos como suelo, grietas en la corteza y mantillo de hojas. La reproducción es variada entre protistas. Casi todos se reproducen asexualmente, y muchos también se reproducen sexualmente. Sin embargo, la mayoría de los protistas no desarrollan órganos reproductores multicelulares, ni forman embriones de la forma en que hacen organismos más complejos. Repaso ■ ■ ¿Cómo varían los protistas en sus medios para obtener nutrientes? ¿Cuáles son algunas de las formas como los protistas interactúan con otros organismos? 26.2 ¿CÓMO EVOLUCIONARON LOS EUCARIOTAS? OBJETIVOS DE APRENDIZAJE 2 3 Discutir la hipótesis de endosimbiosis serial y explicar brevemente parte de la evidencia que la apoya. Describir los tipos de datos que usan los biólogos para clasificar eucariotas. Durante muchos años los biólogos han planteado la hipótesis de que los protistas fueron las primeras células eucariotas y que evolucionaron a partir de procariotas ancestrales. Sin embargo, mientras más se estudian los orígenes eucariotas, más incertidumbre existe. Algo que se puede decir con absoluta certeza es que la evolución de los eucariotas fue un proceso complejo. Los eucariotas quizá aparecieron en el registro fósil tan temprano como hace 2.2 mil millones de años. Aparte de algunos protistas con conchas duras, como diatomeas y foraminíferos, la mayoría de los protistas antiguos no dejaron muchos fósiles porque sus cuerpos eran demasiado blandos para dejar trazas permanentes. Los estudios evolutivos de los protistas se enfocan principalmente en comparaciones moleculares y estructurales de organismos actuales, que contienen muchas pistas acerca de su historia evolutiva. Mitocondrias y cloroplastos probablemente se originaron a partir de endosimbiontes A lo largo de la historia evolutiva, un organismo engulle a otro para beneficio de ambos. De acuerdo con la hipótesis de endosimbiosis serial, ciertos organelos eucariotas, particularmente mitocondrias y cloroplastos, surgieron a partir de relaciones simbióticas entre células más grandes y bacterias más pequeñas que se incorporaron y vivieron dentro de ellas. (Puede repasar la endosimbiosis serial en la figura 21-8). Los biólogos celulares plantean la hipótesis de que las mitocondrias se originaron a partir de bacterias aerobias. Estudios de ADN mitocondrial sugieren que se trata de un resto del pasado de la mitocondria, cuando era un organismo independiente. Secuencias de ARN ribosómico (ARNr) de mitocondrias coinciden estrechamente con los ARNr encontrados en bacterias púrpuras, lo que sugiere que las antiguas bacterias púrpuras fueron los ancestros de las mitocondrias. La evolución de los cloroplastos es más compleja, dado que probablemente hubo varios eventos endosimbióticos (FIGURA 26-2). La evidencia molecular apoya la visión de que la incorporación de una antigua cianobacteria dentro de una célula huésped, conocida como endosimbiosis primaria, resultó en los cloroplastos de las algas rojas, algas verdes y plantas terrestres de hoy. (La célula huésped era eucariota porque las mitocondrias casi seguramente evolucionaron antes que los cloroplastos). Los biólogos plantean la hipótesis de que estos cloroplastos, que están encerrados por dos membranas externas, más tarde proporcionaron a otros eucariotas sus cloroplastos durante endosimbiosis secundaria. En la evolución eucariota ocurrió con frecuencia endosimbiosis secundaria, como evidencia la presencia de membranas de cloroplasto adicionales. Por ejemplo, tres membranas envuelven los cloroplastos de euglenoides y dinoflagelados, y cuatro membranas rodean los cloroplastos de diatomeas, algas doradas y algas pardas. Comprender cómo se originaron estas membranas es un aspecto esencial de la endosimbiosis serial, y muchos investigadores estudian el origen de los cloroplastos en diferentes organismos. Incluso los protistas no fotosintéticos pueden contener reliquias de cloroplasto de eventos endosimbióticos secundarios. Los apicomplejos (protistas como el Plasmodium, que causa malaria) tienen un cloroplasto no funcional rodeado por cuatro membranas. Algunos investigadores plantean la hipótesis de que los medicamentos que inhiben los cloroplastos también pueden matar Plasmodium. Este cloroplasto no funcional, que probablemente derivó de un dinoflagelado, puede tener cierto potencial médico en el tratamiento de la malaria. Comienza a surgir un consenso acerca de la clasificación de eucariotas Los científicos revaloran las relaciones evolutivas entre los eucariotas conforme se vuelve disponible evidencia adicional. Dos tipos de investigación moderna, análisis molecular y estudios ultraestructurales, contribuyen sustancialmente al entendimiento científico de las relaciones filogenéticas entre protistas. Inicialmente, se obtuvieron datos moleculares para el gen que codifica la pequeña subunidad de ARN ribosómico en diferentes eucariotas (SSU ARNr; vea el capítulo 25). Más recientemente, los biólogos compararon otros genes nucleares, muchos de los cuales codifican proteínas, en diferentes taxones de protista. La ultraestructura se refiere a los detalles finos de la estructura celular revelados por microscopia electrónica. En muchos casos, los datos ultraestructurales completan los datos moleculares. La microscopia electrónica revela patrones estructurales similares entre aquellos taxones de protista que la evidencia molecular comparativa sugiere que son monofiléticos; esto es, evolucionaron a partir de un ancestro común (véase el capítulo 23). Por ejemplo, los datos moleculares y ultraestructurales sugieren que los oomicetos, diatomeas, algas doradas y algas pardas, son taxones protista que a primera vista parecen compartir pocas características, son un grupo monofilético. Dada la diversidad en datos ultraestructurales y moleculares de protistas, los biólogos consideran a los protistas como un grupo parafilético; esto es, los protistas contienen algunos de los descendientes de un ancestro eucariota común. Los análisis moleculares y ultraestructurales siguen ayudando a los biólogos a clarificar las relaciones entre los varios filos de protistas y entre los protistas y los otros reinos eucariotas. Los biólogos usan estos datos para desarrollar varios esquemas de clasificación. Un esquema actual, que se adoptó en esta edición, divide los protistas y otros eucariotas (plantas terrestres, hongos y animales) en cinco supergrupos informales (FIGURA 26-3 y TABLA 26-1). Protistas 26_Cap_26_SOLOMON.indd 539 539 17/12/12 10:21 PUNTO CLAVE Los científicos plantean la hipótesis de que los plástidos evolucionaron por endosimbiosis tanto primaria como secundaria. Mitocondria Núcleo Célula eucariota con mitocondria ADN bacterial (a) Endosimbiosis Cianobacteria primaria (ancestro de cloroplasto) Célula eucariota con mitocondrias (b) En un evento endosimbiótico secundario, una célula eucariota heterótrofa (con mitocondrias) engulló una célula eucariota con cloroplastos (se muestra un alga roja). El alga roja sobrevivió y evolucionó en un cloroplasto rodeado por tres membranas (se muestra un cloroplasto dinoflagelado). Otros eventos endosimbióticos secundarios resultaron en estructuras de membrana de cloroplasto más complejas. (a) En la endosimbiosis primaria, una célula eucariota primitiva engulló una cianobacteria, que sobrevivió y evolucionó en un cloroplasto. La célula primitiva se muestra como una célula eucariota porque las mitocondrias casi seguramente evolucionaron antes que los cloroplastos. ADN de cloroplasto (b) Endosimbiosis secundaria Cloroplasto con dos membranas Célula eucariota con mitocondrias y cloroplastos (alga roja) Cloroplasto con tres membranas Célula eucariota con mitocondrias y cloroplastos (dinoflagelado) FIGURA 26-2 Animada Evolución de cloroplastos mediante endosimbiosis primaria y secundaria Este esquema de clasificación es muy diferente de la clasificación eucariota usada en la última edición, y seguramente en futuras ediciones se reportarán más cambios. La ciencia es una labor continua, y el conocimiento actual siempre debe revalorarse a la luz de los nuevos descubrimientos. Ahora que tiene una comprensión básica de la evolución de los protistas y otros eucariotas, examine los protistas representativos dentro de los cinco supergrupos. Repaso ■ ■ ¿Cómo explica la endosimbiosis serial el origen de los cloroplastos? ¿Qué tipos de evidencia científica apoyan la hipótesis de que los protistas son un grupo parafilético? 540 26.3 EXCAVADOS OBJETIVO DE APRENDIZAJE 4 Resumir las características básicas de los excavados y distinguir entre diplomonados, parabasálidos y euglenoides. Los excavados son un grupo diverso de protistas unicelulares con flagelos. Estos protistas se llaman así porque muchos tienen un profundo surco oral, o excavado. A diferencia de otros protistas, los escavados tienen mitocondrias atípicas, enormemente modificadas. Muchos excavados son endosimbiontes y viven en ambientes anóxicos (sin oxígeno). Estos excavados no realizan respiración aeróbica; obtienen energía por la vía anaeróbica de glucólisis (presuntamente mediante fermentación). Capítulo 26 26_Cap_26_SOLOMON.indd 540 17/12/12 10:21 PUNTO CLAVE Revelar las relaciones filogenéticas entre los protistas ha sido difícil, pero se realizan avances. En la actualidad, muchos biólogos clasifican los eucariotas en cinco grupos principales. Euglenoides Excavados Diplomonados Dinoflagelados Apicomplejos Alveolados Oomicetos Diatomeas Cromalveolados Ciliados Estramenópilos Algas pardas B Algas doradas Foraminíferos A Actinópodos Algas verdes Plantas terrestres Arqueplástidos Algas rojas Rizarios Eucariota ancestral Amebas Mohos mucilaginosos plasmodiales Opistocontos Hongos Unicontos Mohos mucilaginosos celulares Coanoflagelados Animales FIGURA 26-3 Relaciones evolutivas entre eucariotas Este árbol evolutivo presenta una interpretación de las relaciones entre eucariotas, en el que los eucariotas se colocan en cinco “supergrupos”. Los organismos dentro de los óvalos representan los reinos eucariotas separados que se estudian en capítulos posteriores. Todos los otros eucariotas se conocen de manera informal como protistas, un grupo parafilético. En el nodo A, los cinco supergrupos se ramifican a partir del mismo punto, lo que indica que el orden de ramificación no está resuelto en este momento. Lo mismo es cierto en el nodo B para las diatomeas, algas pardas y algas doradas. (Con base en Keeling, P. J., “Deep Questions in the Tree of Life,” Science, vol. 317, 28 de septiembre de 2007). Protistas 26_Cap_26_SOLOMON.indd 541 541 17/12/12 10:21 TABLA 26-1 Clados “protista” Características principales Ejemplo específico Excavados Protistas unicelulares con mitocondrias atípicas enormemente modificadas; bicontos Diplomonados y parabasálidos Dos o más flagelos; surco oral ventral (alimentación) Trichomonas vaginalis, un parabasálido Euglenoides y tripanosomas Algunos con plástidos; barra cristalina en flagelos Cromalveolados Protistas diversos que pudieron originarse como resultado de endosimbiosis secundaria en la que una célula ancestral engulló una alga roja; bicontos Alveolados Dinoflagelados, ciliados, apicomplejos Alvéolos (vesículas aplanadas) adentro de la membrana plasmática Estramenópilos Oomicetos, diatomeas, algas pardas, algas doradas La mayoría tienen dos flagelos, uno con pelos; algunos sin flagelos Foraminíferos Testas porosas (conchas duras) a través de las cuales se extienden proyecciones citoplasmáticas (seudópodos) Actinópodos Endoesqueletos (conchas internas) a través de las cuales se extienden axópodos (seudópodos filamentosos) Algas rojas Pigmentos cloroplastos incluyen ficoeritrina (pigmento rojo) y ficocianina (pigmento azul) Algas verdes Pigmentos cloroplastos idénticos a los de las plantas terrestres Unicontos Células que tienen un solo flagelo o son amebas sin flagelos; tienen una fusión genética triple que está ausente en otras eucariotas; incluye animales y hongos Amebozoos Amebas, mohos mucilaginosos plasmodiales, mohos mucilaginosos celulares Amebas desnudas (sin testas) con seudópodos parecidos a lóbulos Opistocontos Coanoflagelados Sin flagelos o único flagelo posterior en células mótiles En la actualidad, los excavados incluyen diplomonados, parabasálidos, euglenoides y tripanosomas. La inclusión de estos organismos en una sola superfamilia está en controversia ya que sus relaciones mutuas son inciertas. Se necesitarán estudios adicionales para determinar si los excavados como se presentan actualmente son un grupo monofilético. Los diplomonados son pequeños flagelados principalmente parásitos Los diplomonados son excavados que tienen uno o dos núcleos, sin mitocondrias funcionales ni complejo de Golgi, y hasta ocho flagelos. Giardia es un parásito diplomonado (FIGURA 26-4a). De manera interesante, Giardia tiene dos núcleos haploides, cada uno de los cuales contiene una copia completa de genoma de Giardia. Giardia carece de mitocondrias funcionales, aunque contiene ciertos genes que codifican proteínas asociadas con mitocondrias en otros organismos. Giardia también tiene estructuras reducidas un poco semejantes a las mitocondrias. Esta infor542 © Wim van Egmond/ Visuals Unlimited, Inc. MO de cuerpo oscuro de Cibicides lobatula, un foraminífero © Wim van Egmond/ Visuals Unlimited, Inc. Arqueplástidos Plástidos rodeados por membranas exterior e interior; incluyen plantas terrestres; bicontos MO de cuerpo oscuro de Ceratium hirundinella, un dinoflagelado MO de cuerpo oscuro de Micrasterias thomasiana, una alga verde © Michael Abbey/ Visuals Unlimited, Inc. Rizarios Células ameboides que con frecuencia tienen testas (conchas); bicontos © Wim van Egmond/ Visuals Unlimited, Inc. Clados “protista” representativos “Supergrupo” eucariota MO de cuerpo oscuro de Amoeba proteus, una ameba mación sugiere a algunos biólogos que un temprano ancestro eucariota de Giardia pudo poseer mitocondrias, que de alguna manera se perdieron o redujeron en una época posterior durante su historia evolutiva. Giardia intestinalis se transporta en el agua y es la mayor causante de diarrea en el mundo. Giardia se elimina como quiste resistente en las heces de muchos animales vertebrados. Estos quistes son un contaminante común en el agua potable no tratada. En una infección severa, gran parte de la pared del intestino delgado está recubierta con estos flagelados, que interfieren con la absorción de nutrientes digeridos y causa pérdida de peso, dolor abdominal y diarrea. Los parabasálidos son endosimbiontes anaerobios que viven en animales Los parabasálidos son excavados anaerobios flagelados que con frecuencia viven en animales. Triconinfas y tricomonas son ejemplos de parabásalidos. Las triconinfas, que tienen cientos de flagelos, viven en Capítulo 26 26_Cap_26_SOLOMON.indd 542 17/12/12 10:21 Unicontos © E. White/Visuals Unlimited, Inc. Arqueplástidos Rizarios Cromalveolados Excavados Núcleos Eucariota ancestral 2 μm (a) Diplomonado. Esta MET coloreada de Giardia intestinalis, un diplomonado parásito, revela dos núcleos. xual (ETS) curable en humanos (vea la MEB en la tabla 26-1). La tricomoniasis afecta a hombres y mujeres, aunque los síntomas son más obvios en las mujeres. De acuerdo con los Centros para el Control y la Prevención de Enfermedades, alrededor de 7.4 millones de nuevos casos ocurren en Estados Unidos cada año. Euglenoides y tripanosomas tienen especies tanto de vida libre como parásitas Euglenoides y tripanosomas se caracterizan por un flagelo inusual: además del arreglo 9 + 2 de los microtúbulos característicos de todos los flagelos eucariotas, estos excavados tienen una barra cristalina en sus flagelos; se desconoce la función de esta barra. Como otros excavados, euglenoides y tripanosomas también tienen mitocondrias atípicas. Flagelos La mayoría de los euglenoides son flagelados unicelulares, y alrededor de un tercio de ellos son fotosintéticos (FIGURAS 26-5a y b). Por lo general tienen dos flagelos: uno largo y con forma de látigo, y otro que frecuentemente es tan corto que no se extiende fuera de la célula. Algunos euglenoides, como Euglena, cambian de forma de manera continua conforme se mueven a través del agua, porque sus películas, o cubiertas exteriores, son flexibles. 50 μm Los euglenoides autótrofos tienen los mismos pigmentos fotosin(b) Parabasálido. MO de Trichonympha, un parabasálido téticos que tienen las algas verdes y plantas, pero los euglenoides no esque vive en el intestino de las termitas y cucarachas tán cercanamente emparentados con alguno de estos grupos, como se comedoras de madera. Trichonympha tiene cientos de muestra en la figura 26-3. Algunos euglenoides fotosintéticos pierden su flagelos. clorofila cuando crecen en la oscuridad, y obtienen sus nutrientes de manera heterótrofa, al ingerir materia orgánica. Otros euglenoides siempre FIGURA 26-4 Diplomonados y parabasálidos carecen de color y son heterótrofos. Algunas especies heterótrofas absorben compuestos orgánicos del agua circundante, mientras que otros engullen bacterias y protistas mediante fagocitosis, y digieren la presa los intestinos de termitas y cucarachas comedoras de madera (FIGURA dentro de vacuolas alimenticias. Los tripanosomas son excavados con una sola mitocondria que 26-4b). Las triconinfas ingieren trozos de madera que las termitas o cucarachas comen. Las triconinfas se apoyan en bacterias endosimbióticas tiene un depósito organizado de ADN llamado cinetoplasto. Los tripara digerir la celulosa de la madera. Los insectos, triconinfas y bacterias panosomas no tienen color y muchos son parásitos y causan enfermedaobtienen sus nutrientes de esta fuente. Éste es un excelente ejemplo de des. En los vertebrados, incluidos los humanos, los tripanosomas viven en la sangre. Por ejemplo, Trypanosoma brucei es un parásito humano mutualismo. La tricomona mejor conocida probablemente sea Trichomonas que causa la enfermedad del sueño (tripanosomiasis africana) (FIGURA vaginalis, que causa tricomoniasis, una enfermedad de transmisión se- 26-5c). Ésta se transmite por la mordedura de moscas tse-tse infectadas. © Michael Abbey/Visuals Unlimited, Inc. Núcleo Protistas 26_Cap_26_SOLOMON.indd 543 543 17/12/12 10:21 Flagelo para locomoción Mancha ocular (estigma) ▲ FIGURA 26-5 Euglenoides y tripanosomas Flagelo no emergente (indistinguible en micrografía) Mancha ocular Mitocondrias (indistinguibles en micrografía) Vacuola contráctil Repaso ■ ■ ¿Qué organelo celular es atípico en los excavados? Ofrezca un ejemplo de enfermedad humana causada por cada uno de los siguientes: diplomonados, parabasálidos y tripanosomas. Cloroplasto Nucleolo Núcleo © Biophoto Associates/Photo Researchers, Inc. Los primeros síntomas incluyen ataques recurrentes de fiebre. Más tarde, cuando los tripanosomas invaden el sistema nervioso central, las personas infectadas están letárgicas y tienen dificultad para hablar o caminar. Al menos 50,000 nuevos casos ocurren cada año; si no se trata, la enfermedad del sueño puede causar la muerte. Cromatina Cuerpo de paramylon (alimento almacenado) Película 25 μm (a) MO de Euglena gracilis, un euglenoide flagelado unicelular. (b) La película de Euglena es flexible y cambia de forma fácilmente. La mancha ocular (estigma) puede blindar un detector de luz en la base del flagelo largo, lo que en consecuencia ayuda a Euglena a moverse hacia la luz con una intensidad adecuada. 26.4 CROMALVEOLADOS OBJETIVOS DE APRENDIZAJE 6 7 Contrastar los dos grupos principales de cromalveolados: alveolados y estramenópilos. Distinguir entre los alveolados: dinoflagelados, apicomplejos y ciliados. Distinguir entre los estramenópilos: oomicetos, diatomeas, algas doradas y algas pardas. Los cromalveolados son un supergrupo compuesto de protistas extremadamente diversos con pocas características compartidas. Los cromalveolados probablemente se originaron como resultado de endosimbiosis secundaria en la que una célula ancestral engulló una alga roja (que en sí misma fue resultado de endosimbiosis primaria). La mayoría de los cromalveolados son fotosintéticos, y la evidencia sugiere que los cromalveolados heterótrofos, como los oomicetos y los ciliados, descendieron de ancestros autótrofos. La clasificación de los cromalveolados como supergrupo monofilético es controversial, porque algunos datos de secuencias de ADN indican que no son monofiléticos. Los cromalveolados se dividen en dos grupos principales, alveolados y estramenópilos. Las características que unifican a los protistas clasificados como alveolados incluyen secuencias de ADN ribosómico similar y alvéolos, que son vesículas aplanadas ubicadas en el interior de la membrana plasmática. En algunos alveolados, las vesículas contienen placas de celulosa. Los alveolados incluyen dinoflagelados, apicomplejos y ciliados. Los estramenópilos incluyen oomicetos, diatomeas, algas doradas y algas pardas. A primera vista, los estramenópilos parecen muy diversos como para clasificarse juntos. Sin embargo, la mayoría de los estramenópilos tienen células mótiles con dos flagelos, uno de los cuales tiene pequeñas proyecciones parecidas a cabellos que se extienden desde el eje. 544 Eritrocitos Tripanosoma con membrana ondulatoria © Eye of Science/Photo Researchers, Inc. 5 Flagelo 10 μm (c) MEB del flagelado Trypanosoma brucei entre eritrocitos humanos. Trypanosoma brucei causa la enfermedad del sueño en los humanos. Capítulo 26 26_Cap_26_SOLOMON.indd 544 17/12/12 10:21 La mayoría de los dinoflagelados son parte del plancton marino Unicontos Arqueplástidos Rizarios Cromalveolados Excavados Los dinoflagelados por lo general son unicelulares, aunque algunos son coloniales. Sus alvéolos contienen placas de celulosa interconectadas impregnadas con silicatos. El dinoflagelado típico tiene dos flagelos. Un flagelo se enreda alrededor de un surco transversal en el centro de la célula como un cinturón, y el otro yace en un surco longitudinal (perpendicular al surco transversal), y se proyecta detrás de la célula (FIGURA 26-6a). La ondulación de estos flagelos impulsa al dinoflagelado a través del agua como una peonza giratoria. De hecho, el nombre se deriva del griego dinos, que significa “girar”. Muchos dinoflagelados marinos son bioluminiscentes. Muchos dinoflagelados son fotosintéticos (vea MO en la tabla 26-1), pero otros son heterótrofos e ingieren otros microorganismos para alimentarse. Algunos dinoflagelados son endosimbiontes que viven en los cuerpos de invertebrados marinos como moluscos, medusas y corales (vea la figura 54-12). Estos dinoflagelados simbióticos, llamados zooxantelas, fotosintetizan y proporcionan carbohidratos a sus compañeros invertebrados. Las zooxantelas contribuyen de manera sustancial a la productividad de los arrecifes de coral. Otros dinoflagelados que son endosimbiontes carecen de pigmentos y son parásitos que viven de sus huéspedes. Ecológicamente, los dinoflagelados son importantes productores en los ecosistemas marinos. Se sabe que algunos dinoflagelados tienen explosiones poblacionales ocasionales, o florecimiento. Estos florecimientos, conocidos como mareas rojas, con frecuencia colorean las aguas costeras de anaranjado, rojo o café (FIGURA 26-6b). Los florecimientos de dinoflagelados son particularmente comunes en agua cálidas ricas en nutrientes. Algunas especies de dinoflagelados que forman mareas rojas producen una toxina que ataca el sistema nervioso de los peces, lo que conduce a matanzas de peces. En ocasiones, algunas aves mueren después de comer peces contaminados. La investigación también vincula las mareas rojas con las muertes de manatíes y delfines en Florida. Los apicomplejos son formadores de esporas que parasitan animales Los apicomplejos son un gran grupo de alveolados parásitos formadores de esporas, algunos de los cuales causan enfermedades severas en los humanos. Contienen los restos no pigmentados de un cloroplasto y acaso evolucionaron a partir de dinoflagelados parásitos que vivieron en los intestinos de invertebrados marinos. Los apicomplejos carecen de Eucariota ancestral Kevin Schafer /Peter Arnold, Inc. /Alamy Dr. David Phillips /Visuals Unlimited, Inc. Flagelos 20 μm (a) MEB de Gymnodinium. Observe los dos flagelos que se ubican en surcos. (b) Marea roja en México. Miles de millones de dinoflagelados producen la turbulencia anaranjada en el agua. FIGURA 26-6 Dinoflagelados Protistas 26_Cap_26_SOLOMON.indd 545 545 17/12/12 10:21 estructuras específicas para locomoción (cilios, flagelos o seudópodos) y se mueven mediante flexión. Los apicomplejos tienen un complejo apical de microtúbulos que fijan el parásito a su célula huésped; el complejo apical es visible sólo usando microscopia electrónica. En alguna etapa en su ciclo de vida, los apicomplejos producen esporozoítos, pequeños agentes infecciosos transmitidos a su siguiente huésped. Muchos apicomplejos pasan parte de su complejo ciclo de vida en especies huésped y parte en una especie huésped diferente. La malaria es causada por un apicomplejo (FIGURA 26-7). De acuerdo con la Organización Mundial de la Salud, aproximadamente 250 millones de personas en la actualidad tienen malaria, y alrededor de 800,000 personas, sobre todo niños de países en desarrollo, mueren de la enfermedad cada año. Aunque durante siglos los escritos chinos, griegos, árabes y romanos describieron la enfermedad, la causa y modo de trans- PUNTO CLAVE misión no se identificaron sino hasta finales del siglo xix. El científico británico Ronald Ross recibió el Premio Nobel en 1902 por su papel en explicar el ciclo de vida de Plasmodium, el apicomplejo que causa malaria. La malaria de hoy es recurrente en muchos países donde estaba bajo control durante décadas. Los métodos de control anteriores, medicamentos antimalaria y pesticidas, han perdido mucha de su efectividad. La cloroquina y muchos otros medicamentos antimalaria se toman de manera profiláctica para evitar la enfermedad, pero Plasmodium adquirió resistencia a muchos de ellos. Más aún: los mosquitos adquirieron resistencia a muchos pesticidas. En la actualidad, investigadores ponen a prueba nuevos medicamentos antimalaria y muchas vacunas posibles contra la enfermedad. La secuenciación del genoma de P. falciparum y el mosquito Anopheles puede conducir a nuevos diagnósticos, medicamentos y vacunas. Plasmodium, un apicomplejo que es el causante de la malaria, vive en dos huéspedes, mosquitos y humanos. 1 El mosquito Anopheles hembra infectado muerde al humano no infectado y transmite esporozoítos de Plasmodium a la sangre Mosquito humana. Anopheles Célula hepática Hígado Esporozoítos (n) Meiosis 2 Los esporozoítos entran Merozoito liberado 6 El cigoto se incrusta DIPLOIDE (2n) HAPLOIDE (n) Eritrocitos Cigoto (2n) Fecundación 3 En las células sanguíneas, los merozoitos se dividen para formar más merozoitos, que infectan a más eritrocitos. Algunos merozoitos forman gametocitos. Mosquito Anopheles Gametos © Dr. Gopal Murti/Photo Researchers, Inc. en el recubrimiento del estómago del mosquito y produce esporozoítos (esporas), que se liberan y migran hacia las glándulas salivales. a las células hepáticas y se dividen para producir merozoitos. Los merozoitos liberados de las células hepáticas infectan eritrocitos. Gametocitos 5 En el aparato digestivo del mosquito, los gametocitos se desarrollan en gametos y ocurre la fecundación. 4 El mosquito Anopheles hembra no infectado pica a una persona infectada y obtiene gametocitos de Plasmodium. Merozoitos 3 μm Eritrocitos FIGURA 26-7 Animada Ciclo de vida de Plasmodium, el agente causante de la malaria La figura muestra una MET de un eritrocito humano lleno con merozoitos. 546 Capítulo 26 26_Cap_26_SOLOMON.indd 546 17/12/12 10:21 Los ciliados usan cilios para locomoción Los ciliados están entre las células eucariotas más complejas. Estos alveolados unicelulares tienen una película que les brinda una forma definitiva pero variable. En Paramecium, la superficie de las células está cubierta con muchos miles de finos cilios cortos con forma de cabello que se extienden a través de poros en la película para facilitar el movimiento (FIGURAS 26-8a y b; vea también la figura 4-4c). Los cilios se baten con una coordinación tan precisa que el organismo puede retroceder y dar la vuelta, así como moverse hacia adelante. No todos los ciliados son mótiles. Algunas formas sésiles tienen tallos, y otros, aunque son capaces de nadar un poco, es más probable que permanezcan unidos a una roca u otra superficie en un punto. Sus cilios generan corrientes de agua que les atraen alimentos. Los ciliados difieren de otros protistas en que tienen dos tipos de núcleos: uno o más pequeños micronúcleos diploides que funcionan en la reproducción; y un macronúcleo poliploide más largo que controla el metabolismo y el crecimiento celulares. La mayoría de los ciliados son capaces de un proceso sexual llamado conjugación, en el que dos individuos se acercan e intercambian material genético Cilios Vacuolas alimenticias Alimento Micronúcleo Surco oral © Robert Brons / Biologcal Photo Service Macronúcleo Vacuola contráctil Vacuola contráctil Poro anal Vacuola alimenticia 50 μm (a) Observe la compleja estructura celular en esta MO de contraste de fase de Paramecium, un ciliado de agua dulce. Como muchos ciliados, Paramecium tiene múltiples núcleos: un macronúcleo y uno o más micronúcleos más pequeños. Macronúcleos que desintegran Macronúcleos sexualmente compatibles se unen en superficies orales. 2 Primera Núcleos diploides (2n) Micronúcleos que desintegran Micronúcleos (2n) 1 Dos individuos (b) Partículas de alimento se barren hacia el surco oral ciliado de Paramecium y se incorporan en vacuolas alimenticias. Lisosomas se fusionan con las vacuolas alimenticias, y el alimento se digiere y absorbe; los desechos no digeridos se eliminan a través del poro anal. 3 Segunda división meiótica en cada célula. división meiótica en cada célula. 4 Un micronúcleo 5 Cada célula en haploide se divide por mitosis; otros se desintegran. conjugación intercambia un micronúcleo. 6 Fusión de micronúcleos haploides. 7 Células separadas. (c) Conjugación en Paramecium caudatum. FIGURA 26-8 Ciliados Protistas 26_Cap_26_SOLOMON.indd 547 547 17/12/12 10:21 (FIGURA 26-8c). La conjugación resulta en dos células “nuevas” que son genéticamente idénticas entre ellas, pero diferentes de lo que eran antes de la conjugación. Después de ésta no es necesario que sigan inmediatamente mitosis y división celular. Por lo general, los ciliados se dividen de modo perpendicular a su eje longitudinal. Los oomicetos producen células reproductoras biflageladas Los oomicetos alguna vez se clasificaron como hongos debido a su parecido superficial. Tanto oomicetos como hongos tienen un cuerpo llamado micelio, que crece sobre material orgánico, al que digieren y luego absorben los nutrientes predigeridos (FIGURA 26-9). Las hifas con forma de hilo que constituyen el micelio en los oomicetos son cenocíti- cos, lo que significa que no tienen paredes cruzadas, y el cuerpo de estos estramenópilos consiste en una sola célula multinucleada. Las paredes celulares de los oomicetos están compuestos de celulosa (como en las plantas), quitina (como en los hongos) o ambos. Cuando el alimento es abundante y las condiciones ambientales son favorables, los oomicetos se reproducen asexualmente. Una punta hifal se hincha y se forma una pared transversal, que separa la punta hifal del resto del micelio. Dentro de esta estructura, llamada zoosporangio, se forman zoosporas biflageladas, cada una de las cuales se balancea, aterriza y enquista, y eventualmente se desarrolla en un nuevo micelio. Cuando las condiciones ambientales empeoran, los oomicetos inician reproducción sexual. Después de la fusión de los núcleos macho y hembra, a partir de las oosferas (gametos femeninos) se desarrollan oosporas de pared gruesa. Con frecuencia, los oomicetos pasan el invierno como oosporas. 2 La meiosis resulta en núcleos de esperma haploide dentro de los anteridios y oosferas haploides (óvulos) dentro de los oogonios. Anteridio (estructura reproductora masculina) Núcleo de oosfera Núcleo de esperma haploide 3 Núcleos de esperma se mueven hacia las oosferas. Meiosis Fecundación GENERACIÓN HAPLOIDE (n) 1 Saprolegnia se Oosporas reproduce asexualmente mediante anteridios y oogonios. 4 REPRODUCCIÓN SEXUAL Oogonio (estructura reproductora femenina) GENERACIÓN DIPLOIDE (2n) Germinación de oospora Micelio Germinación de zoospora Zoospora secundaria enquistada © Collpicto/Shutterstock Zoospora secundaria (forma de frijol) Después de la fecundación, se desarrollan oosporas a partir de las oosferas fertilizadas. Cada oospora puede desarrollarse en un nuevo micelio. Zoosporangio REPRODUCCIÓN ASEXUAL (por mitosis) Zoosporas 5 Zoospora primaria enquistada Saprolegnia se reproduce asexualmente al formar zoosporas dentro de los zoosporangios. Zoospora primaria (forma de pera) Micelio. FIGURA 26-9 Oomicetos Ciclo de vida de Saprolegnia, un oomiceto. Inserto: Un micelio de Saprolegnia radia desde un insecto muerto. 548 Capítulo 26 26_Cap_26_SOLOMON.indd 548 17/12/12 10:21 © Gazanie/Dreamstime Algunos oomicetos tienen un importante papel en la historia humana. Por ejemplo, la hambruna irlandesa de las papas del siglo XIX fue precipitada por el oomiceto Phytophthora infestans, que causa tizón tardío (mildiú) de las papas. (El género Phytophthora se llama así por las palabras griegas que significan “destrucción de planta”). Durante muchos veranos lluviosos y templados en Irlanda durante la década de 1840, el oomiceto se multiplicó sin control, e hizo que los tubérculos de la papa se pudrieran en los campos. Puesto que las papas eran el artículo básico en la dieta de los campesinos irlandeses, hasta un millón de personas murió de hambre. La hambruna desencadenó una migración masiva desde Irlanda hacia Estados Unidos y otros países. Un pariente cercano del oomiceto de mildiú, P. ramorum, provoca muerte súbita de robles, que está aniquilando bosques de roble en varios estados occidentales. Los patólogos de plantas están preocupados de que la enfermedad pueda extenderse hacia los bosques del medio oeste y del este. Este oomiceto particular también ataca secuoyas, abetos de Douglas, laureles, arces y muchas otras especies vegetales, pero la mayoría sólo tienen infecciones en ramas y hojas, no la rápida muerte que se observa en los robles. (a) MEB de una diatomea unicelular. Observe la sorprendente concha que contiene sílice. Las diatomeas tienen conchas compuestas de dos partes La mayoría de las diatomeas son estramenópilos unicelulares, aunque algunas existen como colonias (véase la figura de apertura del capítulo). La pared celular de cada diatomea consiste en dos conchas que traslapan donde encajan, en forma muy parecida a una caja de Petri. En la concha se (b) Reproducción asexual en diatomeas. Después de la división celular, cada nueva célula conserva la mitad de la concha original. La mitad recientemente deposita sílice, y este material parecido al vidrio se deposita sintetizada de la concha siempre encaja adentro de la mitad original. Como en patrones intrincados (FIGURA 26-10a). Existen dos gruresultado, una de las nuevas células es ligeramente menor. pos básicos de diatomeas: las que tienen simetría radial (con forma de rueda) y las que tienen simetría bilateral (con forma FIGURA 26-10 Diatomeas de bote o de aguja). Aunque algunas diatomeas son parte del plancton flotante, otras viven en rocas y sedimentos, donde se mueven mediante deslizamiento. Este movimiento de deslizamiento se facilita mediante la secreción de un material viscoso desde expuso algunos de estos depósitos en tierra. Llamada tierra de diatomeas (o diatomita) estos depósitos se minan y usan para materiales de filtrado, un pequeño surco a lo largo de la concha. Las diatomeas con mucha frecuencia se reproducen asexualmente aislamiento e insonorización. Como agente de filtrado, la diatomita se usa mediante mitosis. Cuando una diatomea se divide, las dos mitades de su para refinar azúcar bruta y para procesar aceites vegetales. Debido a sus concha se separan, y cada una se convierte en la mitad más grande de propiedades abrasivas, la diatomita es un ingrediente común en polvos la concha de una nueva diatomea (FIGURA 26-10b). Dado que la concha para limpieza y pulidores de metales; ya no se agrega a la mayoría de los vítrea no puede crecer, algunas células de diatomea progresivamente se dentífricos porque es muy abrasivo para el esmalte dental. Las conchas de vuelven más pequeñas con cada generación sucesiva. Cuando una diato- diatomea intrincadamente detalladas se usan con frecuencia para poner a mea alcanza una fracción de su tamaño original, ocurre reproducción se- prueba resoluciones de microscopio inferiores a 1 μm. xual, con la producción de gametos sin concha. La reproducción sexual restaura la diatomea a su tamaño original porque el cigoto resultante, una Las algas pardas son estramenópilos multicelulares célula 2n que es fruto de la fusión de n gametos, crece de manera sustanLas algas pardas son las más largas y más complejas de todas las algas cial antes de producir una nueva concha. Las diatomeas son comunes en agua dulce, pero son abundantes de conocidas comúnmente como macroalgas. Todas las algas pardas son manera especial en agua oceánica relativamente fresca. Tienen impor- multicelulares y varían en tamaño de algunos centímetros (alrededor tancia ecológica pues son grandes productoras en los ecosistemas acuá- de 2.5) hasta 75 m. Sus formas corporales son filamentos ramificados; ticos. Al menos una especie es tóxica y se vincula con el envenenamiento penachos; “sogas” carnosas; o gruesas ramas aplanadas. Las algas pardas de mariscos, encalle de mamíferos marinos y la muerte de leones mari- más grandes, llamadas kelp, tienen una apariencia dura y coriácea. Muchos kelp tienen láminas en las que ocurre la mayor parte de la fonos a lo largo de la costa central de California. Cuando las diatomeas mueren, sus conchas caen en pedazos al tosíntesis, estipes parecidos a tallos y rizoides de anclaje parecidas a raíces fondo marino y se acumulan en capas que finalmente se convierten en (FIGURA 26-11a). Con frecuencia tienen vejigas llenas con gas que ofrecen roca sedimentaria. Después de millones de años, el alzamiento geológico flotabilidad. (Láminas, estipes y rizoides de las algas pardas no son homóloProtistas 26_Cap_26_SOLOMON.indd 549 549 17/12/12 10:21 © Gregory Ochocki/Photo Researchers, Inc. Estipe Rizoide (a) La Laminaria está ampliamente distribuida sobre las costas rocosas de mares templados y polares. Crece hasta 2 m. FIGURA 26-11 Algas pardas (b) Un lecho de kelp (Macrocystis pyrifera) es ecológicamente importante para los organismos acuáticos, incluido el león marino que aquí se muestra. Fotografiado en la costa de California. © Jadedme/Dreamstime gas a las hojas, tallos y raíces de las plantas. Algas pardas y plantas surgieron versidad de vida que soportan los lechos de kelp rivaliza con la encontrada en los arrecifes de coral. de diferentes ancestros unicelulares, como se muestra en la figura 26-3). La reproducción es variada y compleja en las algas pardas. Sus células reproductoras, tanto zoosporas asexuales como gametos sexuales, La mayoría de las algas doradas por lo general son biflagelados. La mayoría tiene un ciclo de vida que son biflagelados unicelulares muestra alternancia de generaciones, en la que pasan parte de su vida como organismos haploides multicelulares y parte como organismos di- Las algas doradas se encuentran en ambientes tanto de agua dulce como ploides multicelulares (vea la figura 10-19c). marinos. La mayoría de las especies son organismos unicelulares biflageLas algas pardas tienen importancia comercial por varias razones. lados, aunque algunos son coloniales (FIGURA 26-12a). Algunos de estos Sus paredes celulares contienen un polisacárido llamado algina que se cosecha de kelp como Macrocystis y se usa como agente espesante y estabilizador en helados, dentífricos, crema de afeitar, aerosol para el cabello y loción de manos. Las algas pardas son un importante alimento humano, en particular en el este de Asia, y son una rica fuente de ciertas vitaminas y minerales, como el yodo. Las algas pardas son comunes en aguas marinas más frías, en especial a lo largo de las costas rocosas, donde viven principalmente en la zona entre mareas o aguas relativamente poco profundas. El kelp forma extensos “bosques” subma1 μm 10 μm rinos, o lechos de kelp (FIGURA 26-11b). Son esenciales en dicho ecosistema como (a) MO de una colonia de algas doradas (Synura) (b) MEB de un cocolitofórido (Emiliania huxleyi). encontrada en lagos de agua dulce y estanques. importantes productores de alimentos, y Observe el traslape de escamas de carbonato de calcio. ofrecen hábitat para muchos invertebrados marinos, peces y mamíferos. La diFIGURA 26-12 Algas doradas 550 Dr. Elizabeth Venrick/Scripps Institution of Oceanography © J. R. Waaland/Biological Photo Service Lámina Capítulo 26 26_Cap_26_SOLOMON.indd 550 17/12/12 10:21 Unicontos Arqueplústidos Rizarios Cromalveolados Excavados estramenópilos carecen de flagelos y son similares en apariencia a las amebas, excepto en que las algas doradas contienen cloroplastos. Pequeñas escamas o de sílice o de carbonato de calcio pueden cubrir las células. La reproducción en las algas doradas es principalmente asexual e involucra la producción de esporas mótiles biflageladas llamadas zoosporas. La mayoría de las algas doradas son fotosintéticas, y la composición de pigmento de las algas doradas les brinda un color dorado o café dorado. Algunas especies ingieren bacterias y otras partículas para alimentarse. Ecológicamente, las algas doradas son importantes productores en ambientes marinos. Componen una porción significativa del nanoplancton del océano, algas extremadamente diminutas (2 μm a 10 μm) que son grandes productores debido a su gran abundancia. Existe controversia en torno a la clasificación de las algas doradas. Algunos biólogos colocan diatomeas y algas doradas en un solo filo, mientras que otros clasifican ambos grupos como algas pardas. En el otro extremo, algunos biólogos dividen las algas doradas en dos filos al colocar muchas de las especies marinas, como los cocolitofóridos (FIGURA 26-12b), en un filo separado. Repaso ■ ■ ■ ■ ¿Cuáles son los dos grupos principales de cromalveolados? ¿Cuáles son sus características distintivas? ¿Por qué algunos biólogos consideran que los apicomplejos descendieron de los dinoflagelados? ¿Cuál oomiceto influyó en la historia humana? Explique su respuesta. ¿Cuál es la importancia ecológica de las diatomeas? ¿De las algas pardas? 26.5 RIZARIOS OBJETIVO DE APRENDIZAJE 8 Describir los foraminíferos y los actinópodos, y explicar por qué muchos biólogos los clasifican en el supergrupo rizario. Los rizarios son un supergrupo diverso de células ameboides que con frecuencia tienen conchas externas duras, llamadas testas, a través de las cuales se extienden proyecciones citoplásmicas. Las proyecciones citoplásmicas parecidas a hilos sugieren el nombre rizario, del griego rhiza, que significa “raíz”. Foraminíferos y actinópodos son rizarios, así como ciertas amebas sin concha. Sin embargo, no todas las amebas son rizarios, y muchas especies de amebas están más cercanamente emparentadas con otros clados eucariotas. La evidencia molecular actual indica que el supergrupo rizario es monofilético. Los foraminíferos extienden proyecciones citoplásmicas que forman una red interconectada con forma de hilo Casi todos los foraminíferos son rizarios marinos que producen testas elaboradas (FIGURA 26-13a; vea también la MO en la tabla 26-1). El océano contiene enorme cantidad de foraminíferos, que secretan testas calcáreas de muchas cámaras, con poros a través de los cuales se extienden proyecciones citoplásmicas. (Foraminífera se deriva del latín para “que tiene aberturas”). Las proyecciones citoplásmicas forman una pegajosa red interconectada que atrapa a las presas. Muchos foraminíferos contienen algas unicelulares endosimbiontes (algas verdes, algas rojas o diato- Biophoto Associates © Cortesía M. Reyes-Santos Laboratorio de Protozoología, Facultad de Ciencias, UNAM/México Eucariota ancestral 500 μm (a) MEB de una testa foraminífera. Observe los poros a través de los cuales se proyecta citoplasma. FIGURA 26-13 Rizarios (b) MO de un actinópodo viviente no identificado proveniente del mar Rojo. Muchos axópodos delgados se proyectan de la célula. La concha no es visible porque es un endoesqueleto y el citoplasma lo cubre por todas partes. Protistas 26_Cap_26_SOLOMON.indd 551 551 17/12/12 10:21 meas) que proporcionan alimento mediante fotosíntesis. Muchas especies foraminíferas viven en el fondo marino, pero otras son parte del plancton. Los foraminíferos muertos se acumulan en el fondo del océano, donde sus testas forman un légamo grisáceo que gradualmente se transforma en tiza. Con el alzamiento geológico, estas formaciones de tiza se vuelven parte del terreno, por ejemplo, los Riscos Blancos de Dover, en Inglaterra. (Los Riscos Blancos de Dover son los restos de varios organismos carbonatados, no sólo foraminíferos). Puesto que las testas de los foraminíferos con frecuencia aparecen en capas de roca que cubren depósitos de petróleo, los geólogos que exploran por petróleo buscan testas foraminíferas en estratos de roca. Los foraminíferos están bien preservados en el registro fósil, y los biólogos usan algunos como fósiles índice, marcadores para ayudar a identificar antiguas capas de roca sedimentaria (vea el capítulo 18). Los actinópodos proyectan axópodos delgados Unicontos Arqueplástidos Rizarios Cromalveolados Excavados Los actinópodos son principalmente rizarios de plancton marino con largas proyecciones citoplásmicas filamentosas llamadas axópodos que sobresalen a través de poros en sus conchas (FIGURA 26-13b). Un cúmulo de microtúbulos da fortaleza a cada axópodo. Las algas unicelulares y otras presas quedan atrapadas en estos axópodos y se engullen afuera del cuerpo principal del actinópodo; chorros citoplásmicos llevan la presa adentro del cuerpo. Muchos actinópodos contienen algas endosimbiontes que les brindan los productos de la fotosíntesis. Algunos actinópodos, llamados radiolarios, segregan elaboradas y hermosas conchas vidriosas hechas de sílice. Los radiolarios son un importante constituyente del plancton marino. Cuando los radiolarios y otros actinópodos mueren, sus conchas se asientan y convierten en cieno (sedimento) que puede tener varios metros de espesor sobre el fondo marino. Repaso ■ ■ ¿Por qué foraminíferos y actinópodos se clasifican como rizarios? ¿Qué característica indica el término rizario acerca de foraminíferos y actinópodos? 26.6 ARQUEPLÁSTIDOS OBJETIVO DE APRENDIZAJE 9 Describir la evidencia que apoya la hipótesis de que las algas rojas y las algas verdes deben incluirse en un grupo monofilético con las plantas terrestres. En el esquema de clasificación adoptado en este texto, el grupo monofilético de arqueplástidos incluye algas rojas y algas verdes, que se estudian aquí, y las plantas terrestres, que están en un reino separado (vea los capítulos 27 y 28). Los biólogos clasifican juntos estos grupos basados en datos moleculares y en la presencia de cloroplastos acotados por membranas externas e internas, lo que sugiere que se desarrollaron directamente a partir de cianobacterias endosimbiontes. Todos los protistas fotosintéticos distintos a los arqueplástidos tienen plástidos rodeados por tres o cuatro membranas. Las algas rojas no producen células mótiles La gran mayoría de las algas rojas son organismos multicelulares, aunque existen algunas especies unicelulares. La forma corporal multicelular de las algas rojas por lo general consiste en complejos filamentos entretejidos que son delicados y plumosos (FIGURA 26-14a); algunas (a) Polysiphonia, que está ampliamente distribuida a lo largo del mundo, tiene un cuerpo enormemente ramificado de filamentos entretejidos. © foodfolio /Alamy © Philip Sze/Visuals Unlimited, Inc. © D. Gotshall/ Visuals Unlimited, Inc. Eucariota ancestral (b) Bossiella es un alga roja coralina incrustada con carbonato de calcio. Vive en el océano Pacífico. (c) Nori, hojas secas y comprimidas del alga roja Porphyra, rodea al sushi. FIGURA 26-14 Animada Algas rojas 552 Capítulo 26 26_Cap_26_SOLOMON.indd 552 17/12/12 10:21 © Tom Adams / Visuals Unlimited, Inc. algas rojas son hojas de células aplanadas. La mayoría de las algas rojas multicelulares se adhieren a rocas u otros sustratos mediante un rizoide basal. La reproducción en las algas rojas es notablemente compleja, con alternancia de etapas sexual y asexual. Durante el ciclo de vida no se desarrollan células flageladas. Las algas rojas viven principalmente en aguas oceánicas tropicales cálidas, aunque hay algunas especies en agua dulce y en el suelo. Algunas algas rojas, conocidas como algas coralinas, incorporan en sus paredes celulares carbonato de calcio del agua oceánica (FIGURA 26-14b). El duro carbonato de calcio puede proteger las algas coralinas de los rigores de la acción de las olas. Estas algas coralinas construyen arrecifes “de coral” y acaso son tan cruciales como los animales coralinos en este proceso. Las paredes celulares de las algas rojas con frecuencia contienen gruesos polisacáridos pegajosos que tienen valor comercial. Por ejemplo, el agar es un polisacárido que se extrae de ciertas algas rojas y se utiliza como espesante alimenticio y como medio de cultivo, un sustrato en el que crecen microorganismos y se propagan algunas plantas, como las 50 μm Las algas verdes comparten muchas semejanzas con las plantas terrestres Las algas verdes tienen pigmentos, productos de reserva energética y paredes celulares que son químicamente idénticos a los de las plantas terrestres. Las algas verdes son fotosintéticas, con cloroplastos de una gran variedad de formas. La mayoría de las algas verdes tienen paredes celulares con celulosa, aunque algunas carecen de paredes. Debido a estas y otras semejanzas, los biólogos generalmente aceptan que las plantas terrestres surgieron a partir de algas verdes ancestrales (vea la figura 26-3). Con base en datos moleculares y ultraestructurales recientes, algunos biólogos clasifican este grupo diverso en el reino vegetal. Las algas verdes presentan varios tipos de cuerpos, desde células individuales y colonias, a algas cenocíticas (multinucleadas), hasta filamentos y hojas multicelulares (FIGURA 26-15; vea también el MO en la tabla 26-1). Las formas multicelulares no tienen células diferenciadas en tejidos, una característica que las separa de las plantas terrestres. La mayoría de las algas verdes tienen, o producen, células flageladas durante su ciclo de vida, aunque algunas son totalmente sésiles. La reproducción en las algas verdes es tan variada como su forma corporal, con reproducción tanto sexual como asexual. Muchas algas verdes tienen ciclos de vida con alternancia de generaciones haploide y diploide. La reproducción asexual es por mitosis y división celular en células individuales, o por fragmentación en formas multicelulares. Muchas algas verdes producen esporas asexualmente mediante mitosis; si estas esporas tienen flagelos y son mótiles, se llaman zoosporas (FIGURA 26-16). La reproducción sexual en las algas verdes involucra 100 μm (b) MO de dos colonias Volvox, cada una compuesta de hasta 50,000 células. Las nuevas colonias están adentro de colonias progenitoras, que a la larga se separan. James W. Perry © J. Robert Waaland /Biological Photo Service J. M. Kingsbury in memory of William T. Keeton (a) MO de Spirogyra, una alga verde filamentosa con cloroplastos espirales. orquídeas. Otro polisacárido extraído de las algas rojas, la carragenina, es un aditivo alimenticio que se usa para estabilizar la leche con chocolate y brindar una textura espesa y cremosa al helado y a otros alimentos blandos procesados. La carragenina también se utiliza para estabilizar plantas y cosméticos. Las algas rojas son fuente de vitaminas (en particular A y C) y minerales, especialmente en Japón y otros países del este asiático, donde las personas comen algas rojas frescas, secas o tostadas en alimentos tradicionales como sushi y nori (FIGURA 26-14c). (c) La delgada forma parecida a hoja de la Ulva sugiere su nombre común: lechuga de mar. (d) Chara, alga verde comúnmente llamada asprella, está cercanamente relacionada con las plantas terrestres. Chara está ampliamente distribuida en agua dulce. FIGURA 26-15 Algas verdes Protistas 26_Cap_26_SOLOMON.indd 553 553 17/12/12 10:21 – 5 Ambos tipos de unión se reproducen asexualmente mediante mitosis; sólo se muestra la cepa (−). – REPRODUCCIÓN – ASEXUAL (por mitosis) Zoosporas – – 4 Surgen cuatro células haploides, dos (+) y dos (−). 1 Se producen gametos por mitosis. + – – – REPRODUCCIÓN SEXUAL + – GENERACIÓN HAPLOIDE (n) + – de una cepa diferente GENERACIÓN DIPLOIDE (2n) + Meiosis Fecundación 2 Fusión de gametos (+) y (−), lo que forma un cigoto diploide. 3 Ocurre meiosis. Cigoto (2n) FIGURA 26-16 Animada Ciclo de vida de Chlamydomonas Chlamydomonas es un alga verde haploide unicelular con dos tipos de unión: (+) y (−). formación de gametos en gametangios unicelulares, estructura reproductora donde se producen gametos. Las algas verdes se encuentran en ambientes acuáticos y terrestres. Las algas verdes acuáticas habitan principalmente agua dulce, aunque existen muchas especies marinas. Las algas verdes terrestres están restringidas a suelo húmedo, grietas en corteza de árbol y otros lugares húmedos. Muchas algas verdes son simbiontes con otros organismos; algunas viven como endosimbiontes en células corporales de invertebrados, y algunas crecen junto con hongos como “organismos duales” llamados líquenes (que se estudian en el capítulo 29). Sin importar dónde vivan, las algas verdes tienen importancia ecológica como productores. Repaso ■ ¿Por qué tantos biólogos clasifican a las algas rojas y verdes con las plantas terrestres? 554 26.7 UNICONTOS OBJETIVO DE APRENDIZAJE 10 Describir brevemente y comparar los siguientes unicontos: amebas, mohos mucilaginosos plasmodiales, mohos mucilaginosos celulares y coanoflagelados. Los unicontos son un supergrupo compuesto de ciertas amebas, mohos mucilaginosos plasmodiales, mohos mucilaginosos celulares, coanoflagelados, hongos (que se estudian en el capítulo 29) y animales (que se estudian en los capítulos del 30 al 32). Los unicontos comparten un solo flagelo posterior en células flageladas como espermatozoides y esporas mótiles, aunque algunos organismos existentes en este supergrupo perdieron el flagelo. Muchos unicontos también tienen un solo centriolo. Examine nuevamente la figura 26-3 y observe que los unicontos están Capítulo 26 26_Cap_26_SOLOMON.indd 554 17/12/12 10:21 divididos en dos clados. Uno de éstos es el de los opistocontos, que consiste en hongos, coanoflagelados y animales. En los capítulos 29 y 30 aprenderá más acerca de los opistocontos. Los unicontos también tienen una fusión génica triple que tiene gran importancia evolutiva. En una fusión génica triple, tres genes separados se fusionan en una sola unidad temprano en el curso de la evolución eucariota; el gen fusionado codifica una proteína multienzima. El lector recordará que, en la figura 26-3, los cinco supergrupos se ramificaban desde el mismo punto, lo que indica que el orden de ramificación sigue sin resolverse. Sin embargo, la fusión génica triple ofrece evidencia de una bifurcación, o ramificación de los eucariotas en dos clados principales, cerca de la raíz del ancestro común de todos los eucariotas (FIGURA 26-17). Las dos ramas consisten en (1) unicontos, que tienen un ancestro común con un flagelo posterior individual, y (2) todos los otros eucariotas, llamados de manera colectiva bicontos, que tenían un ancestro común con dos flagelos. E X P E R I M E N TO C L AV E PREGUNTA: ¿Cómo se deducen detalles de la evolución eucariota cerca de la raíz del árbol evolutivo? HIPÓTESIS: Las fusiones genéticas derivadas, que ocurrieron raramente durante el curso de la evolución, pueden proporcionar comprensión acerca de las relaciones evolutivas eucariotas. EXPERIMENTO: Alexandra Stechmann y Thomas Cavalier-Smith, de la Universidad de Oxford, secuenciaron una fusión genética doble que codifica dos enzimas, dihidrofolato reductasa (DHFR) y timidilato sintasa (TS). En un segundo experimento, secuenciaron un gen de fusión triple involucrado en la síntesis enzimática de nucleótidos de pirimidina. Pusieron a prueba la presencia de estas dos fusiones genéticas en varios grupos eucariotas, así como en bacterias y arqueas. Bicontos Excavados Cromalveolados Rizarios Arqueplástidos Unicontos Fusión genética doble (DHFR-TS) Fusión génica triple (síntesis nucleótido pirimidina) Eucariota ancestral RESULTADOS Y CONCLUSIONES: La fusión genética doble está presente en todos los eucariotas examinados, excepto los unicontos; bacterias y arqueas también carecen de fusión genética doble. Los unicontos tienen la fusión genética triple que falta en los otros eucariotas, bacterias y arqueas. Estas fusiones genéticas sugieren una división fundamental de los eucariotas en dos clados principales: los unicontos y todos los demás eucariotas, llamados colectivamente bicontos. Fuente: Stechmann, A. y T. Cavalier-Smith. “The Root of the Eukaryote Tree Pinpointed.” Current Biology, vol. 3, 2 de septiembre de 2003. FIGURA 26-17 Investigación de Stechmann y CavalierSmith acerca de la evolución de protistas Los amebozoos son unicontos con seudópodos lobosos La mayoría de los amebozoos producen proyecciones citoplásmicas temporales llamadas seudópodos (que significa “falso pie”) en algún punto en su ciclo de vida. Los seudópodos de los amebozoos son lobosos (esto es: redondeados y anchos), en oposición a las delgadas proyecciones citoplásmicas características de los rizarios. Los biólogos actualmente clasifican amebas, mohos mucilaginosos plasmodiales y mohos mucilaginosos celulares como amebozoos. Las amebas se mueven mediante la formación de seudópodos Las amebas son amebozoos unicelulares que se encuentran en el suelo, agua dulce, océano y otros organismos (como parásitos). Dada la extrema flexibilidad de su membrana plasmática exterior, muchos miembros de este grupo tienen una forma corporal asimétrica y continuamente cambian de forma conforme se mueven. (La palabra ameba se deriva de una palabra griega que significa “cambio”). Una ameba se mueve al empujar los seudópodos lobosos desde la superficie de la célula (véase la MO en la tabla 26-1). Más citoplasma fluye hacia los seudópodos, lo que los alarga hasta que entra todo el citoplasma y se mueve el organismo como un todo. Los seudópodos también capturan y engullen alimento al rodearlo y formar una vacuola a su alrededor (FIGURA 26-18). Las partículas de alimento se digieren cuando la vacuola alimenticia se fusiona con un lisosoma que contiene enzimas digestivas. Los materiales digeridos se absorben de la vacuola alimenticia, que gradualmente se encoge conforme se vacía. Las amebas se reproducen asexualmente, al dividirse en dos partes iguales después de división mitótica del núcleo; no se ha observado reproducción sexual. Las amebas parásitas incluyen Entamoeba histolytica, que causa disentería amebiana, una seria enfermedad intestinal humana caracterizada por diarrea severa, heces sangrantes y úlceras en la pared intestinal. En casos especialmente severos, el organismo se difunde desde el intestino delgado y causa abscesos en hígado, pulmones o cerebro. La Entamoeba histolytica se transmite como quistes en agua de beber contaminada. Un quiste es una etapa latente, de pared gruesa y resistente, en el ciclo de vida de algunos protistas. Otras amebas, como Acanthamoeba, por lo general son de vida libre, pero producen infecciones oportunistas como infecciones oculares en los usuarios de lentes de contacto. Los mohos mucilaginosos plasmodiales se alimentan como plasmodios multinucleados La etapa de alimentación de un moho mucilaginoso plasmodial es un plasmodio, una masa multinucleada de citoplasma que puede crecer hasta 30 cm de diámetro (FIGURA 26-19a). El plasmodio mucilaginoso fluye sobre troncos en descomposición y basura de hojas húmedos, y con frecuencia forma una red de canales que cubren una gran área superficial. Conforme avanza lentamente, ingiere bacterias, levaduras, esporas y materia orgánica en descomposición. Cuando el suministro de alimento se agota o no hay suficiente humedad, el plasmodio se arrastra hasta una superficie expuesta y comienza a reproducirse. Por lo general, a partir del plasmodio seco se forman estructuras con tallos, de intrincada complejidad y belleza (FIGURA 26-19b). Dentro de dichas estructuras, llamadas esporangios, por meiosis se producen esporas haploides que son extremadamente resistentes a condiciones ambientales adversas. Cuando las condiciones se vuelven favorables, germinan las esporas, y de cada una surge una célula reproductora haploide. Esta célula haploide es o una célula enjambre biflagelada o una myxameba ameboide, Protistas 26_Cap_26_SOLOMON.indd 555 555 17/12/12 10:21 Unicontos © Michael Abbey/ Photo Researchers, Inc. Arqueplástidos Rizarios Cromalveolados Excavados Eucariota ancestral Alga verde Seudópodos 100 μm FIGURA 26-18 Animada Ameba MO de una ameba gigante (Chaos carolinense). Este protista unicelular, que se mueve y alimenta mediante seudópodos, rodea e ingiere una colonia de algas verdes. Las amebas Chaos por lo general son recolectoras que se alimentan de desechos en hábitats de agua dulce, pero ingieren organismos vivos cuando surge la oportunidad. dependiendo de la humedad disponible; en condiciones húmedas forman células flageladas. Las células enjambre y las myxamebas actúan como gametos, que se fusionan para formar un cigoto con un núcleo diploide. El núcleo diploide resultante se divide muchas veces por mitosis, pero el citoplasma no se divide, de modo que el resultado es un plasmodio multinucleado. El moho mucilaginoso plasmodial Physarum polycephalum es un organismo modelo que los investigadores usan para estudiar muchos procesos biológicos fundamentales, como crecimiento, ciclosis y la función del citoesqueleto. Los mohos mucilaginosos celulares se alimentan como células ameboides individuales R. Calentine/CBR Images R. Calentine/CBR Images Los mohos mucilaginosos celulares son amebozoos con cercanas afinidades a amebas y mohos mucilaginosos plasmodiales. Durante su etapa de alimentación, cada moho mucilaginoso celular es una célula ameboide individual que se comporta como un organismo separado solitario (FIGURA 26-20). Cada célula se arrastra sobre troncos en putrefacción y el suelo, o nada en agua dulce, e ingiere bacterias y otras partículas de alimento conforme avanza. Cada célula ameboide tiene un núcleo haploide y se reproduce por mitosis, como hace una ameba verdadera. 250 μm (a) El plasmodio brillantemente coloreado, que se muestra sobre un tronco muerto, se alimenta de bacterias y otros microorganismos. (b) Las estructuras reproductoras son esporangios sobre tallos. FIGURA 26-19 El moho mucilaginoso plasmodial Physarum polycephalum 556 Capítulo 26 26_Cap_26_SOLOMON.indd 556 17/12/12 10:21 © Cabisco / Visuals Unlimited, Inc. © Cabisco / Visuals Unlimited, Inc. 5 Cuerpo fructífero 50 μm Esporas 100 μm 6 Agregación Cuerpo fructífero 1 Amebas moho mucilaginoso 4 Babosa diferenciada Agregación © Cabisco /Visuals Unlimited, Inc. 3 2 Se forma babosa multicelular FIGURA 26-20 Animada El moho mucilaginoso celular Dictyostelium discoideum 50 μm Babosa Las amebas moho mucilaginoso ingieren alimento, crecen y se reproducen mediante división celular. Después de agotarse su alimento, cientos de células ameboides fluyen en conjunto (inserto en superior derecha) y forman un agregado migratorio con forma de babosa (inserto en inferior izquierda). Cuando deja de migrar, el agregado forma un cuerpo fructífero sobre un tallo (inserto en superior izquierda). El cuerpo fructífero libera esporas, cada una de las cuales se abre en un ambiente favorable para liberar una célula ameboide. Cuando la humedad o el alimento se vuelven inadecuados, ciertas células emiten una señal química, adenosín monofosfato cíclico (AMPc), que hace que se agreguen por cientos o miles. Durante esta etapa las células se arrastran durante distancias cortas como un solo agregado multicelular, o babosa. Cada célula de la babosa conserva su membrana plasmática e identidad individual. Finalmente, la babosa se asienta y reorganiza, y forma un cuerpo fructífero con tallo que contiene esporas. Después de liberarse, cada espora se abre y surge una sola cé- lula ameboide haploide: la etapa de alimentación. El ciclo reproductivo de formación de esporas es asexual, aunque ocasionalmente se observa reproducción sexual. Los ciclos de vida de la mayoría de los mohos mucilaginosos celulares carecen de una etapa flagelada. El moho mucilaginoso celular Dictyostelium discoideum es un organismo modelo para el estudio de la diferenciación celular, la comunicación celular y la motilidad y adhesión celulares. Su biología se ha estudiado de manera intensa, particularmente en lo relacionado con las moléculas de Protistas 26_Cap_26_SOLOMON.indd 557 557 17/12/12 10:21 señalización celular, como AMPc, que se encuentra en muchos organismos además de los mohos mucilaginosos celulares (véase el capítulo 6). Flagelo Collar de microvellos Los coanoflagelados están cercanamente emparentados con los animales Los coanoflagelados son flagelados con collar. Estos pequeños unicontos no visibles se encuentran globalmente en ambientes de agua dulce y marinos. Los coanoflagelados incluyen especies tanto de nado libre como sésiles que están permanentemente adheridos mediante un delgado tallo a detritos ricos en bacterias. Su flagelo único está rodeado en la base por un delicado collar de microvellos que atrapan alimento (FIGURA 26-21). Los coanoflagelados son de especial interés debido a su sorprendente parecido con las células de collar en las esponjas (vea la figura 31-1b). Otros filos animales, como los cnidarios, gusanos planos y equinodermos, también contienen células parecidas a coanoflagelados, pero no se ha observado que ningún otro grupo de protistas posea estas células. La evidencia que apoya la cercana relación entre coanoflagelados y animales incluye datos comparativos de secuencias de ADN de genes mitocondriales y nucleares. Dadas las semejanzas en estructura y genómica molecular acumulada en años recientes, muchos biólogos plantean la hipótesis de que los coanoflagelados son los parientes vivientes no animales más cercanos de los animales. Por ende, los coanoflagelados y animales vivientes probablemente comparten un ancestro común parecido a coanoflagelado. Repaso ¿Por qué las amebas que tienen seudópodos lobosos no se clasifican con los rizarios? ¿Cuál grupo de unicontos es estructural y genéticamente más similar a los animales? ■ ■ ■ ■ Célula Lorica (cubierta protectora) Tallo FIGURA 26-21 Coanoflagelados Los coanoflagelados son flagelados de vida libre que obtienen alimento al ondular sus flagelos, lo que hace que corrientes de agua transporten bacterias y otras pequeñas partículas de alimento hacia el collar de microvellos. Aquí se muestra una forma colonial. Cada célula mide de 5 a 10 μm de largo, sin incluir el flagelo. R E S U M E N : E N F O Q U E E N LOS O B J E T I VOS D E A P R E N D I Z A J E 26.1 (página 538) ■ De acuerdo con la hipótesis de endosimbiosis serial, mitocondrias y cloroplastos surgieron de relaciones simbióticas entre células más grandes y bacterias más pequeñas que se incorporaron y vivieron dentro de ellas. ■ Los cloroplastos de algas rojas, algas verdes y plantas probablemente surgieron en un solo evento endosimbiótico primario en el que una cianobacteria se incorporó en una célula. Múltiples endosimbiosis secundarias condujeron a cloroplastos en euglenoides, dinoflagelados, diatomeas, algas doradas y algas pardas, y a los cloroplastos no funcionales en apicomplejos. 1 Discutir en términos generales la diversidad inherente en protistas, incluidos medios de locomoción, modos de nutrición, interacciones con otros organismos, hábitats y modos de reproducción. ■ Los protistas tienen varios medios de locomoción, incluidos seudópodos, flagelos y cilios; algunos son sésiles. 3 Describir los tipos de datos que usan los biólogos para clasificar eucariotas. ■ ■ Los protistas obtienen sus nutrientes de manera autótrofa o heterótrofa. Los protistas son de vida libre o simbiótica, y las relaciones simbióticas varían del mutualismo al parasitismo. ■ La mayoría de los protistas viven en el océano o en estanques, lagos y corrientes de agua dulce. Los protistas parásitos viven en los fluidos corporales o células de sus huéspedes. ■ Muchos protistas se reproducen tanto sexual como asexualmente; otros se reproducen sólo asexualmente. 26.2 (página 539) 2 Discutir la hipótesis de endosimbiosis serial y explicar brevemente parte de la evidencia que la apoya. 558 Las relaciones entre protistas están determinadas principalmente por ultraestructuras, que son los detalles finos de la estructura celular revelados mediante microscopia electrónica, y por datos moleculares comparativos. Los biólogos compararon genes nucleares, muchos de los cuales codifican proteínas, en diferentes taxones de protistas. 26.3 (página 540) 4 Resumir las características básicas de los excavados y distinguir entre diplomonados, parabasálidos y euglenoides. ■ Los excavados son un grupo diverso de protistas unicelulares con flagelos, un surco oral excavado y atípicas mitocondrias enormemente modificadas. La inclusión de diplomonados, parabasálidos y euglenoides en la superfamilia excavada es controversial. ■ Los diplomonados son excavados, organismos con un profundo, o excavado, surco oral. Los diplomonados tienen uno o dos núcleos, mitocondrias no funcionales, sin complejo de Golgi y hasta ocho flagelos. Capítulo 26 26_Cap_26_SOLOMON.indd 558 17/12/12 10:21 ■ Los parabasálidos son excavados anaerobios flagelados, que con frecuencia viven en animales. Triconinfas y tricomonas son ejemplos de parabasálidos. ■ Los euglenoides son unicelulares y flagelados. Algunos euglenoides son fotosintéticos. 26.4 (página 544) (kelps) poseen láminas parecidas a hojas, estipes parecidas a tallos, rizoides para anclarse y vejigas llenas de gas para flotabilidad. 26.5 (página 551) 8 Describir los foraminíferos y los actinópodos, y explicar por qué muchos biólogos los clasifican en el supergrupo rizario. ■ Los rizarios son células ameboides que con frecuencia tienen conchas externas duras llamadas testas, a través de las cuales se extienden proyecciones citoplásmicas; la evidencia molecular indica que este grupo es monofilético. ■ Los foraminíferos secretan testas de muchas cámaras con poros a través de los cuales se extienden proyecciones citoplásmicas para moverse y obtener alimento. ■ Los actinópodos son en su mayoría plancton marino que obtienen alimento mediante axópodos, delgadas proyecciones citoplásmicas que se extienden a través de poros en sus conchas. 5 Contrastar los dos grupos principales de cromalveolados: alveolados y estramenópilos. ■ ■ Los cromalveolados probablemente se originaron como resultado de endosimbiosis secundaria en la que una célula ancestral engulló una alga roja. Algunos datos de secuencias de ADN sugieren que los cromalveolados no son monofiléticos. Los alveolados tienen secuencias de ADN ribosómico y alvéolos similares, vesículas aplanadas ubicadas justo adentro de la membrana plasmática. La mayoría de los estramenópilos tienen células mótiles con dos flagelos, uno de los cuales tiene pequeñas proyecciones parecidas a cabellos que salen del eje. 6 Distinguir entre los alveolados: dinoflagelados, apicomplejos y ciliados. ■ ■ Los dinoflagelados son alveolados principalmente unicelulares, biflagelados, fotosintéticos de gran importancia ecológica como productores en ecosistemas marinos. Sus alvéolos, vesículas aplanadas bajo la membrana plasmática, con frecuencia contienen placas de celulosa impregnadas con silicatos. Algunos dinoflagelados producen blooms tóxicos conocidos como mareas rojas. Los apicomplejos son parásitos que producen esporozoítos y no son mótiles. Un complejo apical de microtúbulos fijan al apicomplejo a su célula huésped. El apicomplejo Plasmodium causa malaria. 26.6 (página 552) 9 Describir la evidencia que apoya la hipótesis de que las algas rojas y las algas verdes deben incluirse en un grupo monofilético con las plantas terrestres. ■ Algas rojas, algas verdes y plantas terrestres, llamadas de manera colectiva arqueplástidos, se consideran un grupo monofilético con base en datos moleculares y en la presencia de cloroplastos acotados por membranas exterior e interior. ■ Las algas rojas, que son principalmente macroalgas multicelulares, son de importancia ecológica en aguas oceánicas tropicales cálidas. ■ Las algas verdes presentan una gran diversidad de tamaños, complejidad estructural y reproducción. Los botánicos plantean la hipótesis de que las algas verdes ancestrales dieron origen a las plantas terrestres. Observe el ciclo de vida del parásito de la malaria haciendo clic sobre la figura en CengageNOW. ■ Los ciliados son alveolados que se mueven mediante cilios parecidos a cabellos, tienen micronúcleos (para reproducción sexual) y macronúcleos (para controlar el metabolismo y el crecimiento celulares), y experimentan un proceso sexual llamado conjugación. Observe la conjugación haciendo clic sobre la figura en CengageNOW. 7 Distinguir entre los estramenópilos: oomicetos, diatomeas, algas doradas y Observe los ciclos de vida de una alga roja y una alga verde haciendo clic sobre las figuras en CengageNOW. 26.7 (página 554) 10 Describir brevemente y comparar los siguientes unicontos: amebas, mohos mucilaginosos plasmodiales, mohos mucilaginosos celulares y coanoflagelados. ■ Los unicontos, que incluyen hongos y animales, comparten un solo flagelo posterior en células flageladas. ■ Las amebas se mueven y obtienen alimento mediante fagocitosis, usando extensiones citoplásmicas llamadas seudópodos. algas pardas. ■ Los oomicetos tienen un micelio cenocítico. Se reproducen asexualmente al formar zoosporas biflageladas y de manera sexual mediante la formación de oosporas. ■ La etapa de alimentación de los mohos mucilaginosos plasmodiales es un plasmodio multinucleado. La reproducción es mediante esporas haploides producidas dentro de esporangios. ■ Las diatomeas son principalmente unicelulares, con conchas que contienen sílice. Algunas diatomeas son parte del plancton flotante, y otras viven en rocas y sedimentos donde se mueven mediante deslizamiento. ■ Los mohos mucilaginosos celulares se alimentan como células ameboides individuales. Se reproducen al reunirse en un agregado (babosa), y luego forman esporas asexuales. ■ Las algas doradas son principalmente estramenópilos unicelulares biflagelados de agua dulce y marinos que son de importancia ecológica como componente del extremadamente diminuto nanoplancton del océano. ■ Las algas pardas son estramenópilos multicelulares que son de importancia ecológica en aguas oceánicas más frías. Las algas pardas más grandes Observe el ciclo de vida de un moho mucilaginoso celular haciendo clic sobre las figuras en CengageNOW. ■ Los coanoflagelados son unicontos que probablemente sean los parientes vivientes no animales más cercanos de los animales. Un collar de microvellos rodea su único flagelo en la base. E VA L Ú E S U CO M P R E N S I Ó N 1. ¿Cuál de los siguientes no es cierto de los protistas? (a) son unicelulares, coloniales, cenocíticos u organismos multicelulares simples (b) sus cilios y flagelos tienen un arreglo 9 + 2 de microtúbulos (c) son procariotas, como las bacterias y arqueas (d) algunos son de vida libre, y algunos son endosimbiontes (e) la mayoría son acuáticos y viven en el océano o en estanques de agua dulce 2. La evidencia molecular apoya la visión de que todos los plástidos evolucionaron a partir de un(a) antiguo(a) (a) cianobacteria (b) arquea (c) diplomonada (e) apicomplejo (e) uniconto 3. Los foraminíferos (a) son endosimbiontes que viven en muchos invertebrados marinos (b) fueron responsables de la hambruna de la papa irlandesa en el siglo xix (c) secretan testas de muchas cámaras Protistas 26_Cap_26_SOLOMON.indd 559 559 17/12/12 10:21 con poros a través de los cuales se proyectan extensiones citoplásmicas (d) tienen numerosos axópodos que pueden ayudar a capturar y retener presas (e) tienen numerosos cilios para dirigir el alimento hacia el surco oral 12. Ponga etiquetas a los diagramas. Use las figuras 26-5b y 26-8b para comprobar sus respuestas. 4. Paramecium y otros ciliados con frecuencia muestran un fenómeno sexual llamado (a) oogamia (b) conjugación (c) endosimbiosis serial (d) marea roja (e) alternancia de generaciones 5. Los alveolados parásitos que forman esporas en alguna etapa de su vida pertenecen, ¿a cuál grupo? (a) actinópodos (b) ciliados (c) cocolitofóridos (d) apicomplejos (e) dinoflagelados 6. La malaria (a) se transmite por la mordedura de una mosca tse-tse hembra (b) es causada por un flagelado parásito, Giardia intestinalis (c) es una forma severa de disentería amebiana causada por Entamoeba histolytica (d) es causada por un apicomplejo que pasa parte de su ciclo de vida en el mosquito Anopheles y parte en humanos (e) se transmite cuando las personas beben agua teñida por una marea roja 7. Los protistas fotosintéticos con conchas compuestos de dos mitades que encajan como una caja de Petri son (a) algas doradas (b) diatomeas (c) euglenoides (d) algas pardas (e) foraminíferos 8. Los pigmentos, productos de reserva energética y paredes celulares que se encuentran en las plantas terrestres también son características de (a) algas verdes (b) algas pardas (c) algas doradas (d) diatomeas (e) euglenoides y diatomeas 9. Los kelps son ______________ con cuerpos multicelulares diferenciados en láminas, estipes, rizoides y vegijas llenas de gas. (a) algas doradas (b) diatomeas (c) euglenoides (d) algas pardas (e) algas rojas 10. Los mohos mucilaginosos celulares (a) incluyen Physarum y Phytophtora (b) están más cercanamente emparentados con las bacterias que cualquier otro protista (c) son responsables del tizón tardío de la papa, que condujo a hambruna en Irlanda en la década de 1840 (d) tienen fusión genética doble (e) forman una babosa cuando las células se agregan en respuesta a AMP cíclico 11. Los oomicetos se reproducen asexualmente al formar __________ y asexualmente al formar _________. (a) oosporas; rizoides (b) zoosporas; zooxantelas (c) zoosporas; oosporas (d) rizoides; oosporas (e) oosporas; zoosporas P E N S A M I E N TO C R Í T I CO Nicole King Lab, University of California, Berkeley 1. VÍNCULO CON LA EVOLUCIÓN. ¿Por qué los protistas se consideran parafiléticos? Use la figura 26-3 como auxiliar para explicar su respuesta. 2. ANÁLISIS DE DATOS. En la figura 26-3, ¿dónde colocaría al ancestro común de algas pardas y oomicetos? ¿Al ancestro común de animales y hongos? 3. ANÁLISIS DE DATOS. En la MO de campo oscuro de coanoflagelados unicelulares (a la derecha), la actina se tiñó de rojo, la tubulina de verde y el ADN de azul. Identifique las partes celulares que contengan cada uno de estos materiales químicos. 4. CIENCIA, TECNOLOGÍA Y SOCIEDAD. ¿Por qué en Plasmodium surgió resistencia a los medicamentos tradicionales contra la malaria? 560 Preguntas adicionales están disponibles en CengageNOW en www.cengage.com/login. Capítulo 26 26_Cap_26_SOLOMON.indd 560 17/12/12 10:21