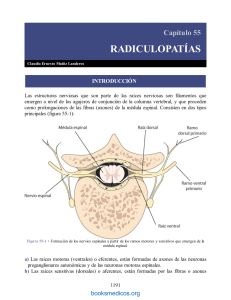
")
Purves • Augustine • Fitzpatrick • Hall LaMantia • White Neurociencia 5ª EDICIÓN Neurociencia Neurociencia 5.a EDICIÓN DIRECTORES Dale Purves • George J. Augustine David Fitzpatrick • William C. Hall Anthony-Samuel LaMantia • Leonard E. White Directores asociados Richard D. Mooney • Michael L. Platt BUENOS AIRES - BOGOTÁ - CARACAS - MADRID - MÉXICO - PORTO ALEGRE e-mail: [email protected] www.medicapanamericana.com Título del original en inglés NEUROSCIENCE. FIFTH EDITION Copyright © 2012 by Sinauer Associates, Inc. All rights reserved. Imagen de la cubierta. La ilustración muestra una porción de los dos hemisferios del encéfalo humano, observada desde arriba en la línea media. (Cortesía de Mark Williams, Dale Purves y Len White). © Gestora de Derechos Autorales, S.L. Madrid, España Traducción de EDITORIAL MÉDICA PANAMERICANA, S.A. Efectuada por Diana Klajn Revisión científica de la traducción: Juan Argüelles Profesor Titular de Fisiología Departamento de Biología Funcional Área de Fisiología Universidad de Oviedo, España 5.a edición (versión impresa) España, septiembre 2015. Los editores han hecho todos los esfuerzos para localizar a los poseedores del copyright del material fuente utilizado. Si inadvertidamente hubieran omitido alguno, con gusto harán los arreglos necesarios en la primera oportunidad que se les presente para tal fin. Gracias por comprar el original. Este libro es producto del esfuerzo de profesionales como usted, o de sus profesores, si usted es estudiante. Tenga en cuenta que fotocopiarlo es una falta de respeto hacia ellos y un robo de sus derechos intelectuales. Las ciencias de la salud están en permanente cambio. A medida que las nuevas investigaciones y la experiencia clínica amplían nuestro conocimiento, se requieren modificaciones en las modalidades terapéuticas y en los tratamientos farmacológicos. Los autores de esta obra han verificado toda la información con fuentes confiables para asegurarse de que ésta sea completa y acorde con los estándares aceptados en el momento de la publicación. Sin embargo, en vista de la posibilidad de un error humano o de cambios en las ciencias de la salud, ni los autores, ni la editorial o cualquier otra persona implicada en la preparación o la publicación de este trabajo, garantizan que la totalidad de la información aquí contenida sea exacta o completa y no se responsabilizan por errores u omisiones o por los resultados obtenidos del uso de esta información. Se aconseja a los lectores confirmarla con otras fuentes. Por ejemplo, y en particular, se recomienda a los lectores revisar el prospecto de cada fármaco que planean administrar para cerciorarse de que la información contenida en este libro sea correcta y que no se hayan producido cambios en las dosis sugeridas o en las contraindicaciones para su administración. Esta recomendación cobra especial importancia con relación a fármacos nuevos o de uso infrecuente. Visite nuestra página web: http://www.medicapanamericana.com ARGENTINA Marcelo T. de Alvear 2145 (C1122AAG) Buenos Aires, Argentina Tel.: (54-11) 4821-5520 / 2066 / Fax (54-11) 4821-1214 e-mail: [email protected] COLOMBIA Carrera 7a A Nº 69-19 - Bogotá D.C., Colombia Tel.: (57-1) 345-4508 / 314-5014 / Fax: (57-1) 314-5015 / 345-0019 e-mail: [email protected] ESPAÑA Quintanapalla Nº 8, Planta 4a (28050) - Madrid, España Tel.: (34-91) 1317800 / Fax: (34-91) 4570919 e-mail: [email protected] MÉXICO Hegel Nº 141, 2° piso Colonia Polanco V Sección Delegación Miguel Hidalgo - C.P. 11560 - México, Distrito Federal Tel.: (52-55) 5250-0664 / 5262-9470 / Fax: (52-55) 2624-2827 e-mail: [email protected] VENEZUELA Edificio Polar, Torre Oeste, Piso 6, Of. 6 C Plaza Venezuela, Urbanización Los Caobos, Parroquia El Recreo, Municipio Libertador, Caracas Depto. Capital, Venezuela Tel.: (58-212) 793-2857/6906/5985/1666 Fax: (58-212) 793-5885 e-mail: [email protected] ISBN: 978-84-9835-983-1 (versión electrónica) ISBN: 978-84-9835-754-7 (versión impresa) Todos los derechos reservados. Este libro o cualquiera de sus partes no podrán ser reproducidos ni archivados en sistemas recuperables, ni transmitidos en ninguna forma o por ningún medio, ya sean mecánicos o electrónicos, fotocopiadoras, grabaciones o cualquier otro, sin el permiso previo de Editorial Médica Panamericana S.A. de C.V. © 2016. EDITORIAL MÉDICA PANAMERICANA S.A. Quintanapalla, 8 - 4ª planta - 28050 Madrid - España La versión electrónica de esta 5ª edición se publicó en el mes de septiembre de 2015. ÍndiCe abreviado 1. Estudio del sistema nervioso 1 Unidad I Señalización neural 2. 3. 4. 5. 6. 7. 8. Señales eléctricas de las células nerviosas 25 Permeabilidad de la membrana dependiente de voltaje 41 Canales y transportadores 57 Transmisión sináptica 77 Neurotransmisores y sus receptores 109 Señalización molecular en el interior de las neuronas 141 Plasticidad sináptica 163 Unidad II Sensibilidad y procesamiento sensitivo 9. 10. 11. 12. 13. 14. 15. El sistema somatosensitivo: tacto y propiocepción 189 Dolor 209 Visión: el ojo 229 Vías visuales centrales 257 Sistema auditivo 277 Sistema vestibular 303 Sentidos químicos 321 Unidad III El movimiento y su control central 16. 17. 18. 19. 20. 21. Circuitos de la neurona motora inferior y su control motor 353 Control del tronco del encéfalo y la médula espinal por la neurona motora superior 375 Modulación del movimiento por los ganglios basales 399 Modulación del movimiento por el cerebelo 417 Movimientos oculares e integración sensitivomotora 435 Sistema motor visceral 451 Unidad IV El encéfalo cambiante 22. 23. 24. 25. Desarrollo encefálico en la fase temprana 477 Construcción de los circuitos neurales 507 Modificación de los circuitos encefálicos como resultado de la experiencia 537 Reparación y regeneración en el sistema nervioso 559 Unidad V Funciones encefálicas complejas 26. 27. 28. 29. 30. 31. Cortezas de asociación y cognición 587 Palabra y lenguaje 607 Sueño y vigilia 625 Las emociones 647 Sexo, sexualidad y el encéfalo 669 La memoria 695 Apéndice: Resumen de la neuroanatomía humana 717 Atlas: El sistema nervioso central humano 745 Colaboradores George J. Augustine, PhD David Fitzpatrick, PhD William C. Hall, PhD Anthony-Samuel LaMantia, PhD James O. McNamara, MD Richard D. Mooney, PhD Michael L. Platt, PhD Dale Purves, MD Sidney A. Simon, PhD Leonard E. White, PhD direCtores de las unidades UNIDAD I: George J. Augustine UNIDAD II: David Fitzpatrick y Richard D. Mooney UNIDAD III: Leonard E. White y William C. Hall UNIDAD IV: Anthony-Samuel LaMantia UNIDAD V: Dale Purves y Michael L. Platt PrefaCio Sea que se lo evalúe desde el punto de vista molecular, celular, sistémico, conductual o cognitivo, el sistema nervioso humano es una maquinaria biológica increíble. Dados sus logros (por ejemplo, todos los productos de la cultura humana), existen buenas razones para querer saber cómo funcionan el encéfalo y el resto del sistema nervioso. Los efectos debilitantes y económicamente onerosos de las enfermedades neurológicas y psiquiátricas incrementan la necesidad urgente de responder a estos interrogantes. El objetivo de este libro es destacar los desafíos e intereses intelectuales (y también las incertidumbres) de lo que muchos consideran la última gran frontera de la ciencia biológica. La información presentada sirve como punto de partida para los estudiantes de grado, los estudiantes de medicina, los graduados que estudian neurociencias y otros lectores que desean conocer cómo funciona el sistema nervioso humano. Al igual que otros muchos retos importantes, la neurociencia tiene controversias y disenso, y también es muy entretenida. La quinta edición de este libro incluye todos estos ingredientes; esperamos que así lo perciban los lectores de todos los niveles. agradeCimientos Agradecemos a los numerosos colegas que aportaron críticas y sugerencias útiles en esta edición y en ediciones anteriores. Queremos agradecer en particular a Paul Adams, Ralph Adolphs, David Amaral, Dora Angelaki, Eva Anton, Gary Banker, el difunto Bob Barlow, Marlene Behrmann, Ursula Bellugi, Carlos Belmonte, Dan Blazer, Bob Burke, Roberto Cabeza, Jim Cavanaugh, Jean-Pierre Changeux, John Chapin, Milt Charlton, Michael Davis, Rob Deaner, Bob Desimone, Allison Doupe, Sasha du Lac, Jen Eilers, Anne Fausto-Sterling, Howard Fields, Elizabeth Finch, Nancy Forger, Jannon Fuchs, Michela Gallagher, Dana Garcia, Steve George, la difunta Patricia Goldman-Rakic, Josh Gooley, Jennifer Groh, Mike Haglund, Zach Hall, Kristen Harris, Bill Henson, John Heuser, Bertil Hille, Miguel Holmgren, Jonathan Horton, Ron Hoy, Alan Humphrey, Jon Kaas, Kai Kaila, Jagmeet Kanwal, Herb Killackey, Len Kitzes, Marc Klein, Chieko Koike, Andrew Krystal, Arthur Lander, Story Landis, Simon LeVay, Darrel Lewis, Jeff Lichtman, Alan Light, Steve Lisberger, John Lisman, Arthur Loewy, Ron Mangun, Eve Marder, Robert McCarley, Greg McCarthy, Jim McIlwain, Daniel Merfeld, Steve Mitroff, Chris Muly, Vic Nadler, Ron Oppenheim, Larysa Pevny, Franck Polleux, Scott Pomeroy, Rodney Radtke, Louis Reichardt, Sidarta Ribiero, Marnie Riddle, Jamie Roitman, Steve Roper, John Rubenstein, Ben Rubin, David Rubin, Josh Sanes, Cliff Saper, Lynn Selemon, Paul Selvin, Carla Shatz, Bill Snider, Larry Squire, John Staddon, Peter Strick, Warren Strittmatter, Joe Takahashi, Stephen Traynelis, Christopher Walsh, Xiaoqin Wang, Richard Weinberg, Jonathan Weiner, Christina Williams, S. Mark Williams, Joel Winston y Ryohei Yasuda. Es obvio que si hubiera algún error este no es atribuible de ninguna forma a nuestros correctores y asesores. También queremos agradecer a nuestros colegas Nell Cant, Dona M. Chikaraishi, Michael D. Ehlers, Gillian Einstein, Erich Jarvis, el difunto Lawrence C. Katz, Julie Kauer, Donald Lo, Miguel A. L. Nicolelis, Peter H. Reinhart, J. H. Pate Skene, James Voyvodic y Fulton Wong por sus contribuciones en ediciones previas. Agradecemos a los estudiantes de las distintas universidades en las que los directores han trabajado, así como a muchos otros estudiantes y colegas que sugirieron mejoras y correcciones. Por último, debemos agradecer especialmente a Andy Sinauer, Graig Donini y Sydney Carroll, Carol Wigg, Christopher Small, Jefferson Johnson, Janice Holabird, Marie Scavotto y el resto del personal de Sinauer Associates por el sobresaliente trabajo y los altos estándares que han mantenido a lo largo de las cinco ediciones de este libro. ÍndiCe CAPÍTULO 1 Estudio del sistema nervioso Aspectos generales Genética, genómica y encéfalo 1 1 1 RECUADRO 1A ORGANISMOS MODELO EN NEUROCIENCIA 2 Los componentes celulares del sistema nervioso Neuronas Células neurogliales Diversidad celular en el sistema nervioso Circuitos neurales 4 6 7 10 10 La organización del sistema nervioso humano Sistemas neurales Análisis estructural de los sistemas neurales Análisis funcional de los sistemas neurales Recuadro 1B Técnicas de imágenes encefálicas Análisis de la conducta compleja Resumen RECUADRO 1B TÉCNICAS POR LA IMAGEN ENCEFÁLICAS Análisis de la conducta compleja Resumen Lecturas adicionales 13 15 15 16 18 21 21 18 21 21 21 UNIDAD I señalizaCión neural 23 CAPÍTULO 3 Permeabilidad de la membrana dependiente de voltaje Aspectos generales Corrientes iónicas a través de las membranas de las células nerviosas CAPÍTULO 2 Señales eléctricas de las células nerviosas RECUADRO 3A MÉTODO DE PINZAMIENTO DE VOLTAJE 25 Aspectos generales Transmisión a larga distancia de las señales eléctricas De qué modo los movimientos iónicos producen señales eléctricas 25 27 RECUADRO 2A PROPIEDADES DE MEMBRANA PASIVA 30 Las fuerzas que crean los potenciales de membrana Equilibrio electroquímico en un medioambiente con más de un ion permeable Base iónica del potencial de membrana de reposo RECUADRO 3B UMBRAL 29 33 33 35 RECUADRO 2B LAS NOTABLES NEURONAS GIGANTES DEL CALAMAR 36 Base iónica de los potenciales de acción Resumen RECUADRO 2C FORMA Y NOMENCLATURA DE LOS POTENCIALES DE ACCIÓN Lecturas adicionales Dos tipos de corriente iónica dependientes de voltaje Dos conductancias de membrana dependientes de voltaje Reconstrucción del potencial de acción Señalización a larga distancia por medio de los potenciales de acción Aumento de la velocidad de conducción como resultado de la mielinización Resumen RECUADRO 3C ESCLEROSIS MÚLTIPLE Lecturas adicionales 41 41 41 42 43 45 47 49 51 51 53 54 55 37 38 CAPÍTULO 4 39 Canales iónicos y transportadores 57 40 Aspectos generales 57 XII ÍNDICE Canales iónicos que participan en los potenciales de acción RECUADRO 4A MÉTODO DE PINZAMIENTO ZONAL DE MEMBRANA 57 59 Categorías de neurotransmisores Acetilcolina RECUADRO 6A NEUROTOXINAS QUE ACTÚAN SOBRE LOS RECEPTORES POSTSINÁPTICOS Glutamato 109 111 114 116 RECUADRO 4B TOXINAS QUE ENVENENAN LOS CANALES IÓNICOS 62 DIVERSIDAD DE LOS CANALES IÓNICOS 63 RECUADRO 4C EXPRESIÓN DE LOS CANALES IÓNICOS EN LOS OVOCITOS DE XENOPUS RECUADRO 6B MIASTENIA GRAVE: UNA ENFERMEDAD AUTOINMUNITARIA DE LAS SINAPSIS NEUROMUSCULARES 117 63 RECUADRO 6C EXCITOTOXICIDAD EN LA LESIÓN ENCEFÁLICA AGUDA 121 Canales iónicos con puerta de voltaje Canales iónicos con puerta de ligando Canales activados por el estiramiento y el calor Estructura molecular de los canales iónicos Los transportadores activos crean y mantienen los gradientes iónicos 64 66 66 66 69 RECUADRO 4C ENFERMEDADES CAUSADAS POR ALTERACIÓN DE LOS CANALES IÓNICOS 70 Propiedades funcionales de la bomba Na+/K+ Estructura molecular de las bombas de Na+/K+ Resumen Lecturas adicionales 72 72 74 75 77 Aspectos generales Sinapsis eléctricas Transmisión de señales en las sinapsis químicas Propiedades de los neurotransmisores 77 78 80 81 RECUADRO 5A CRITERIOS QUE DEFINEN UN NEUROTRANSMISOR 84 Liberación cuántica de los neurotransmisores Liberación de transmisores de las vesículas sinápticas Reciclado local de las vesículas sinápticas Papel del calcio en la secreción de transmisores Mecanismos moleculares del ciclo de las vesículas sinápticas RECUADRO 5B ENFERMEDADES QUE AFECTAN LA TERMINACIÓN PRESINÁPTICA Receptores de neurotransmisores Cambios en la permeabilidad de la membrana postsináptica durante la transmisión sináptica Potenciales presinápticos excitadores e inhibidores Sumación de potenciales sinápticos RECUADRO 5C LA SINAPSIS “TRIPARTITA” Resumen Lecturas adicionales 84 86 87 88 90 93 96 97 102 103 104 106 106 Aspectos generales Aminas biógenas 109 109 122 124 125 RECUADRO 6E LAS AMINAS BIÓGENAS NEUROTRANSMISORAS Y LOS TRASTORNOS PSIQUIÁTRICOS 126 RECUADRO 6F ADICCIÓN El ATP y otras purinas Neurotransmisores peptídicos Neurotransmisores no convencionales RECUADRO 6G LA MARIHUANA Y EL ENCÉFALO 128 131 132 135 137 138 139 CAPÍTULO 7 Señalización molecular en el interior de las neuronas 141 Aspectos generales Estrategias de señalización molecular Activación de las vías de señalización Tipos de receptores Proteínas G y sus puntos diana moleculares Segundos mensajeros 141 141 143 144 145 147 RECUADRO 7A IMÁGENES DINÁMICAS DE SEÑALIZACIÓN INTRACELULAR 149 Los blancos de los segundos mensajeros: proteincinasas y fosfatasas Señalización nuclear RECUADRO 7B ESPINAS DENDRÍTICAS Ejemplos de transducción de señales neuronales Resumen Lecturas adicionales 151 153 154 157 159 160 CAPÍTULO 8 Plasticidad sináptica CAPÍTULO 6 Neurotransmisores y sus receptores RECUADRO 6C ACCIONES EXCITADORAS DEL GABA EN EL ENCÉFALO EN DESARROLLO Resumen Lecturas adicionales CAPÍTULO 5 Transmisión sináptica El GABA y la glicina Aspectos generales Plasticidad sináptica a corto plazo La plasticidad sináptica a largo plazo subyace a la modificación del comportamiento en Aplysia Potenciación a largo plazo en una sinapsis del hipocampo 163 163 163 166 169 ÍNDICE RECUADRO 8A GENÉTICA DEL APRENDIZAJE Y DE LA MEMORIA EN LA MOSCA DE LA FRUTA Mecanismos que subyacen a la plasticidad a largo plazo RECUADRO 8B SINAPSIS SILENTES Mecanismos que subyacen a la depresión a largo plazo 170 173 175 Plasticidad dependiente del momento de la espiga RECUADRO 8C EPILEPSIA: EFECTO DE LA ACTIVIDAD PATOLÓGICA SOBRE EL CIRCUITO NEURALE Resumen Lecturas adicionales XIII 182 182 184 184 178 UNIDAD II sensibilidad y ProCesamiento sensitivo 187 CAPÍTULO 10 Dolor Aspectos generales Nociceptores Transducción y transmisión de señales nociceptivas CAPÍTULO 9 El sistema somatosensitivo: tacto y propiocepción 189 Aspectos generales Las fibras aferentes transmiten la información somatosensitiva al sistema nervioso central Las aferencias somatosensitivas muestran distintas propiedades funcionales RECUADRO 9A DERMATOMAS Mecanorreceptores especializados en información táctil receptiva Mecanorreceptores especializados en la propiocepción Vías centrales que transmiten información táctil del cuerpo: sistema columna dorsal-lemnisco medial Vías centrales que transmiten información táctil desde el rostro: sistema trigeminotalámico Vías centrales que transmiten información propioceptiva del cuerpo Vías centrales que transmiten información propioceptiva del rostro Componentes somatosensitivos del tálamo Corteza somatosensitiva primaria Componentes somatosensitivos del tálamo Corteza somatosensitiva primaria 189 189 191 191 194 196 198 200 200 201 201 202 201 202 RECUADRO 9B MPATRONES DE ORGANIZACIÓN DENTRO DE LAS CORTEZAS SENSITIVAS: MÓDULOS ENCEFÁLICOS 203 Más allá de SI: vías corticorticales y descendentes Plasticidad en la corteza cerebral del adulto Resumen Lecturas adicionales 205 206 207 207 RECUADRO 10A CAPSAICINA Las vías centrales del dolor son distintas de las vías mecanosensitivas RECUADRO 10B DOLOR REFERIDO RECUADRO 10C VÍA DE LAS COLUMNAS DORSALES PARA EL DOLOR VISCERAL Vías paralelas del dolor Vías del dolor y la temperatura para el rostro Otras modalidades mediadas por el sistema anterolateral Sensibilización RECUADRO 10D MIEMBROS FANTASMA Y DOLOR FANTASMA Control descendente de la percepción del dolor El efecto placebo Bases fisiológicas de la modulación del dolor Resumen Lecturas adicionales 209 209 209 211 212 213 214 215 217 218 220 220 222 223 223 224 226 226 CAPÍTULO 11 Visión: el ojo Aspectos generales Anatomía del ojo Formación de las imágenes en la retina RECUADRO 11A LA MIOPÍA Y OTROS DEFECTOS DE REFRACCIÓN La superficie de la retina Circuito retiniano 229 229 229 230 231 233 233 RECUADRO 11B EL PUNTO CIEGO 234 RECUADRO 11C DEGENERACIÓN MACULAR 235 XIV ÍNDICE RECUADRO 11D RETINITIS PIGMENTARIA 237 Epitelio pigmentario retiniano Fototransducción Especialización funcional de los sistemas de bastones y conos Distribución anatómica de bastones y conos Conos y visión de colores 238 238 242 244 245 RECUADRO 11E LA IMPORTANCIA DEL CONTEXTO EN LA PERCEPCIÓN DEL COLOR 246 Circuitos retinianos para detectar los cambios de luminancia 249 RECUADRO 11F LA PERCEPCIÓN DE LA INTENSIDAD DE LA LUZ 250 Contribución de los circuitos retinianos a la adaptación a la luz Resumen Lecturas adicionales 253 256 256 257 Aspectos generales Proyecciones centrales de las células ganglionares de la retina Representación retinotópica del campo visual Defectos del campo visual Propiedades de regulación sincronización espaciotemporal de las neuronas en la corteza visual primaria Arquitectura de la corteza visual primaria Combinación de las aferencias de los dos ojos 257 257 259 261 263 266 267 270 División del trabajo dentro de la vía visual primaria Organización funcional de las áreas visuales extraestriadas Resumen Lecturas adicionales 271 273 275 275 CAPÍTULO 13 Aspectos generales Sonido Espectro audible RECUADRO 13A CUATRO CAUSAS DE PÉRDIDA ADQUIRIDA AUDITIVA 277 277 277 278 279 Sinopsis de la función auditiva 279 RECUADRO 13B LA MÚSICA 280 Oído externo Oído medio RECUADRO 13C PÉRDIDA AUDITIVA NEUROSENSORIAL E IMPLANTES COCLEARES Oído interno Células ciliadas y transducción mecanoeléctrica de las ondas sonoras De qué modo la información proveniente de la cóclea alcanza los puntos diana en el tronco del encéfalo Integración de la información proveniente de los dos oídos Vías monoaurales desde el núcleo coclear hasta el lemnisco lateral Integración en el colículo inferior Tálamo auditivo Corteza auditiva RECUADRO 13E REPRESENTACIÓN DE LOS SONIDOS COMPLEJOS EN EL ENCÉFALO DE LOS MURCIÉLAGOS Y LOS SERES HUMANOS Sistema vestibular Aspectos generales Laberinto vestibular Células ciliadas vestibulares RECUADRO 14B ADAPTACIÓN Y MODULACIÓN DE LAS CÉLULAS CILIADAS VESTIBULARES Órganos con otolitos: utrículo y sáculo De qué modo las neuronas de los órganos con otolitos perciben las inclinaciones y las aceleraciones lineales Conductos semicirculares De qué modo los conductos semicirculares detectan las aceleraciones angulares Vías centrales para estabilizar la mirada, la cabeza y la postura RECUADRO 14C EVALUACIÓN CLÍNICA DEL SISTEMA VESTIBULAR RECUADRO 14D LAS CÉLULAS DE MAUTHNER EN LOS PECES Vías vestibulares hasta el tálamo y la corteza Percepción de la orientación espacial e integración multisensorial Resumen Lecturas adicionales 281 282 CAPÍTULO 15 283 Sentidos químicos 284 Aspectos generales Organización del sistema olfatorio Percepción olfatoria en los seres humanos 286 289 291 291 292 292 293 297 297 297 298 299 301 301 CAPÍTULO 14 RECUADRO 14A TERMINOLOGÍA BÁSICA RECUADRO 12A ESTEREOGRAMAS DE PUNTOS ALEATORIOS Y PASATIEMPOS RELACIONADOS Sistema auditivo RECUADRO 13D EL DULCE SONIDO DE LA DISTORSIÓN Resumen Lecturas adicionales CAPÍTULO 12 Vías visuales centrales Base iónica de la mecanotransducción en las células ciliadas El amplificador coclear Sintonización y cronometrado en el nervio auditivo 303 303 303 304 305 306 307 309 310 312 312 314 316 318 319 320 320 321 321 321 323 ÍNDICE Evaluación de la función olfatoria en el laboratorio o en la clínica 324 RECUADRO 15A USTED TIENE SOLO UNA NARIZ 326 Respuestas fisiológicas y conductuales a las sustancias odoríferas RECUADRO 15B FEROMONAS, REPRODUCCIÓN Y SISTEMA VOMERONASAL Epitelio olfatorio y neuronas receptoras olfatorias Transducción de los olores y proteínas receptoras de sustancias odoríferas RECUADRO 15C EL “DOCTOR PERRO” 327 328 329 332 333 Mecanismos fisiológicos de la transducción de los olores El bulbo olfatorio Procesamiento cortical de la información transmitida desde el bulbo olfatorio Organización del sistema del gusto Percepción del gusto en los seres humanos Proteínas receptoras gustativas y transducción Codificación neural en el sistema gustativo Quimiorrecepción trigeminal Resumen Lecturas adicionales XV 335 337 340 341 343 345 347 349 349 350 UNIDAD III el movimiento y su Control Central 351 CAPÍTULO 17 Control del tronco del encéfalo y la médula espinal por la neurona motora superior 375 CAPÍTULO 16 Circuitos de la neurona motora inferior y su control motor Aspectos generales Centros neurales responsables del movimiento Relaciones neurona motora-músculo La unidad motora Regulación de la fuerza muscular RECUADRO 16A PLASTICIDAD DE LA UNIDAD MOTORA Circuitos de la médula espinal que subyacen a los reflejos de estiramiento muscular Influencia de la actividad sensitiva sobre la conducta motora Otra retroalimentación sensitiva que afecta el rendimiento motor Vías reflejas de la flexión RECUADRO 16B LOCOMOCIÓN EN LA SANGUIJUELA Y LA LAMPREA Circuito de la médula espinal y locomoción 353 353 353 355 357 358 359 362 364 365 367 368 369 RECUADRO 16C AUTONOMÍA DE LOS GENERADORES CENTRALES DE PATRONES: EVIDENCIA PROVENIENTE DEL GANGLIO SOMATOGÁSTRICO DE LA LANGOSTA 370 Síndrome de la neurona motora inferior RECUADRO 16D ESCLEROSIS LATERAL A MIOTRÓFICA Resumen Lecturas adicionales Aspectos generales Organización del control motor descendente Las vías corticoespinales y corticobulbares RECUADRO 17A PATRONES DE DEBILIDAD FACIAL Y SU IMPORTANCIA EN LA LOCALIZACIÓN DE LA LESIÓN NEUROLÓGICA 379 Organización funcional de la corteza motora primaria 380 RECUADRO 17B ¿QUÉ REPRESENTAN LOS MAPAS MOTORES? 381 RECUADRO 17C TALENTOS SENSITIVOMOTORES Y ESPACIO CORTICAL 384 La corteza premotora Centros motores corticales en el tronco encefálico: neuronas motoras superiores que mantienen el equilibrio, gobiernan la postura y orientan la mirada RECUADRO 17D LA FORMACIÓN RETICULAR Daño de las vías motoras descendentes: síndrome de la neurona motora superior RECUADRO 17E TONO MUSCULAR Resumen Lecturas adicionales 372 CAPÍTULO 18 373 Modulación del movimiento por los ganglios basales 373 374 375 375 377 Aspectos generales 386 389 391 395 397 397 398 399 399 XVI ÍNDICE Proyecciones hacia los ganglios basales Proyecciones desde los ganglios basales hacia otras regiones encefálicas Evidencia proveniente de estudios de los movimientos oculares Circuitos dentro del sistema de los ganglios basales La dopamina modula los circuitos de los ganglios basales Trastornos hipocinéticos e hipercinéticos del movimiento 399 CAPÍTULO 20 402 Movimientos oculares e integración sensitivomotora 435 404 405 407 Aspectos generales Qué logran los movimientos oculares Acciones e inervación de los músculos extraoculares 435 435 436 408 RECUADRO 20A PERCEPCIÓN DE LAS IMÁGENES RETINIANAS ESTABILIZADAS 437 RECUADRO 18A ENFERMEDAD DE PARKINSON: UNA OPORTUNIDAD PARA NUEVOS ENFOQUES TERAPÉUTICOS 410 RECUADRO 18B ENFERMEDAD DE HUNTINGTON 411 RECUADRO 18C ESTIMULACIÓN CEREBRAL PROFUNDA RECUADRO 20B INTEGRACIÓN SENSITIVOMOTORA EN EL COLÍCULO SUPERIOR 412 RECUADRO 18DCIRCUITOS DE LOS GANGLIOS BASALES Y FUNCIONES ENCEFÁLICAS NO MOTORAS RECUADRO 20C DE CÓDIGOS DE LUGAR A CÓDIGOS DE FRECUENCIA 414 Resumen Lecturas adicionales 416 416 CAPÍTULO 19 Modulación del movimiento por el cerebelo Control neural de los movimientos de seguimiento suave Control neural de los movimientos de vergencia Resumen Lecturas adicionales 438 440 443 445 449 449 450 450 CAPÍTULO 21 417 Aspectos generales Organización del cerebelo Proyecciones hacia el cerebelo Proyecciones desde el cerebelo Circuitos en el interior del cerebelo Circuito cerebeloso y coordinación del movimiento en curso 417 417 419 421 423 RECUADRO 19A ENFERMEDADES POR PRIONES 427 OTRAS CONSECUENCIAS DE LAS LESIONES CEREBELOSAS 429 RECUADRO 19B ANÁLISIS GENÉTICO DE LA FUNCIÓN CEREBELOSA 431 Resumen Lecturas adicionales Tipos de movimientos oculares y sus funciones Control neural de los movimientos oculares sacádicos 426 433 433 Sistema motor visceral Aspectos generales Primeros estudios del sistema motor visceral Características distintas del sistema motor visceral División simpática del sistema motor visceral RECUADRO 21A EL HIPOTÁLAMO División parasimpática del sistema motor visceral Sistema nervioso entérico Componentes sensitivos del sistema motor visceral Control central de las funciones motoras viscerales RECUADRO 21B SÍNDROME DE HORNER Neurotransmisión en el sistema motor visceral RECUADRO 21C OBESIDAD Y ENCÉFALO Regulación autónoma de la función cardiovascular Regulación autonóma de la vejiga Regulación autonóma de la función sexual Resumen Lecturas adicionales 451 451 451 454 454 456 459 461 461 463 465 466 466 468 470 472 473 474 ÍNDICE XVII UNIDAD IV el enCéfalo Cambiante 475 Base molecular de la motilidad del cono de crecimiento CAPÍTULO 22 Desarrollo encefálico temprano Aspectos generales 477 Formación inicial del sistema nervioso: gastrulación y neurulación 477 RECUADRO 22A CÉLULAS MADRE: PROMESA Y PELIGROS Formación de las subdivisiones encefálicas principales RECUADRO 22B ROMBÓMEROS Base molecular de la inducción neural 479 481 484 485 RECUADRO 22C ÁCIDO RETINOICO: TERATÓGENO Y SEÑAL INDUCTIVA 488 Las señales inductivas integradas establecen la identidad neuronal Diferenciación inicial de neuronas y glía RECUADRO 22D NEUROGÉNESIS: DÓNDE, CUÁNDO Y QUÉ 489 490 492 Regulación molecular de la neurogénesis 493 La generación de la diversidad neuronal 495 Interrupciones moleculares y genéticas del desarrollo neural temprano 496 RECUADRO 22E TRIPLE RIESGO: ENFERMEDADES ASOCIADAS CON EL “ERIZO SÓNICO” Migración neuronal en el sistema nervioso periférico RECUADRO 22F PREPARACIÓN: MIGRACIÓN NEURONAL A LARGAS DISTANCIAS Migración neuronal en el sistema nervioso central Mecanismos moleculares de la migración neuronal y trastornos de la migración cortical Resumen Lecturas adicionales 497 499 501 503 503 505 505 CAPÍTULO 23 Construcción de circuitos neurales507 Aspectos generales Polarización neuronal: el primer paso en la formación de los circuitos neurales Cono de crecimiento axónico 512 SEÑALES NO DIFUSIBLES PARA LA GUÍA DEL AXÓN 514 RECUADRO 23B TRASTORNOS HUMANOS DE LA GUÍA DEL AXÓN 477 507 507 509 510 RECUADRO 23A ELECCIÓN DE LADOS: GUÍA DEL AXÓN EN EL QUIASMA ÓPTICO Quimioatracción y quimiorrepulsión Formación de mapas topográficos Formación de sinapsis selectivas Regulación de las conexiones neuronales por las interacciones tróficas Interacciones competitivas y formación de conexiones neuronales 515 518 521 521 525 527 RECUADRO 23C ¿POR QUÉ LAS NEURONAS TIENEN DENDRITAS? 528 RECUADRO 23D EL DESCUBRIMIENTO DE BDNF Y LA FAMILIA DE LAS NEUROTROFINAS 530 Base molecular de las interacciones tróficas Señalización de las neurotrofinas Resumen Lecturas adicionales 532 534 535 536 CAPÍTULO 24 Modificación de los circuitos neurales como resultado de la experiencia 537 Aspectos generales Actividad neural y desarrollo encefálico Períodos críticos 537 537 539 RECUADRO 24A COMPORTAMIENTOS INNATOS 540 RECUADRO 24B EL CANTO DE LAS AVES 541 Correlaciones celulares y moleculares de la plasticidad dependiente de la actividad durante los períodos críticos RECUADRO 24C MARCAJE TRANSNEURONAL CON AMINOÁCIDOS RADIACTIVOS Períodos críticos en el desarrollo del sistema visual Efectos de la privación visual sobre la dominancia ocular RECUADRO 24D CORRELACIÓN COMO CAUSALIDAD: LECCIONES OBTENIDAS DE UNA RANA DE TRES OJOS Manipulación de la competencia Ambliopía, estrabismo y períodos críticos para la visión humana Evidencia de períodos críticos en otros sistemas sensitivos 542 544 544 545 549 551 551 552 XVIII ÍNDICE Desarrollo del lenguaje: un período crítico para un comportamiento característicamente humano Desarrollo del encéfalo humano, plasticidad dependiente de la actividad y períodos críticos Resumen Lecturas adicionales 553 556 557 557 CAPÍTULO 25 Reparación y regeneración del sistema nervioso Aspectos generales El encéfalo dañado Reorganización funcional sin reparación Tres tipos de reparación neuronal Regeneración de nervios periféricos Base celular y molecular de la reparación del nervio periférico Regeneración de las sinapsis periféricas 559 559 559 560 562 563 564 567 RECUADRO 25A REGENERACIÓN ESPECÍFICA DE CONEXIONES SINÁPTICAS EN LOS GANGLIOS AUTÓNOMOS Regeneración después de la lesión del sistema nervioso central RECUADRO 25B VÍCTIMAS EN LA GUERRA Y EN LOS DEPORTES Respuestas celulares y moleculares a la lesión Crecimiento axónico después de la lesión encefálica Neurogénesis en el sistema nervioso central maduro Neurogénesis del adulto en vertebrados no mamíferos Neurogénesis en el encéfalo de mamíferos adultos Mecanismos celulares y moleculares de la neurogénesis del adulto RECUADRO 25C ARMAS NUCLEARES Y NEUROGÉNESIS Neurogénesis del adulto, células madre y reparación encefálica en los seres humanos Resumen Lecturas adicionales 568 570 571 573 575 576 577 579 579 581 583 583 584 UNIDAD V funCiones enCefáliCas ComPlejas 585 “Neuronas de la planificación” en la corteza frontal del mono Resumen Lecturas adicionales CAPÍTULO 27 CAPÍTULO 26 Cortezas de asociación Aspectos generales Cortezas de asociación Aspectos generales de la estructura cortical Características específicas de las cortezas de asociación RECUADRO 26A LAMINACIÓN CORTICAL La corteza de asociación cortical media la atención “Neuronas de la atención” en la corteza parietal del mono La corteza de asociación temporal media el reconocimiento “Neuronas del reconocimiento” en la corteza temporal del mono RECUADRO 26C PRUEBAS NEUROPSICOLÓGICAS La corteza de asociación frontal media la planificación y la toma de decisiones RECUADRO 26B PSICOCIRUGÍA 604 605 606 587 587 587 588 589 590 591 594 596 599 600 601 602 Habla y lenguaje 607 Aspectos generales El lenguaje está localizado y lateralizado en el encéfalo 607 RECUADRO 27A LA GENERACIÓN DEL HABLA 608 Afasias 607 610 RECUADRO 27B ¿OTROS ANIMALES TIENEN LENGUAJE? 611 RECUADRO 27C PALABRAS Y SIGNIFICADOS 613 Una notable confirmación de la lateralización del lenguaje y otros conceptos RECUADRO 27D LENGUAJE Y LATERALIDAD MANUAL Búsqueda de diferencias anatómicas entre los hemisferios derecho e izquierdo Mapeo de las funciones del lenguaje El papel del hemisferio derecho Lenguaje de signos Resumen Lecturas adicionales 615 617 618 619 622 623 623 623 XIX ÍNDICE CAPÍTULO 28 Sueño y vigilia Aspectos generales ¿Por qué los seres humanos (y muchos otros animales) duermen? RECUADRO 28A ESTILOS DE SUEÑO EN LAS DIFERENTES ESPECIES El ciclo circadiano de sueño y vigilia RECUADRO 28B MECANISMOS MOLECULARES DEL RELOJ BIOLÓGICO Estadios del sueño CAPÍTULO 30 625 625 625 627 628 629 631 RECUADRO 28C ,ELECTROENCEFALOGRAFÍA 632 Cambios fisiológicos en los estadios del sueño Otras posibles funciones del sueño REM y la actividad onírica Circuitos neurales que gobiernan el sueño 635 636 637 RECUADRO 28D LA CONCIENCIA 638 RECUADRO 28E FÁRMACOS Y SUEÑO 641 Interacciones talamocorticales en el sueño Trastornos del sueño RECUADRO 28F SÍNDROME DE FATIGA CRÓNICA Resumen Lecturas adicionales 643 644 644 645 645 CAPÍTULO 29 Las emociones Aspectos generales Cambios fisiológicos asociados con la emoción6 RECUADRO 29A EXPRESIONES FACIALES: CONTRIBUCIONES PIRAMIDALES Y EXTRAPIRAMIDALES La integración de la conducta emocional El sistema límbico Importancia de la amígdala 647 647 47 648 650 652 653 RECUADRO 29B ANATOMÍA DE LA AMÍGDALA 654 RECUADRO 29C EL RAZONAMIENTO SUBYACENTE A UN DESCUBRIMIENTO IMPORTANTE 655 RECUADRO 29D EL MIEDO Y LA AMÍGDALA HUMANA: ESTUDIO DE UN CASO 657 RECUADRO 29E TRASTORNOS AFECTIVOS 659 Relación entre neocorteza y amígdala Lateralización cortical de las funciones emocionales Emoción, razón y conducta social Refuerzo emocional y adicción Resumen Lecturas adicionales 660 661 662 664 667 667 Sexo, sexualidad y encéfalo 669 Aspectos generales Dimorfismos sexuales y conducta sexualmente dimorfa Sexo, gónadas, cuerpos y encéfalos 669 669 671 RECUADRO 30A LA CIENCIA DEL AMOR (O EL AMOR ES UNA DROGA) 672 Influencias hormonales sobre el dimorfismo sexual Dimorfismos sexuales primarios en el encéfalo Dimorfismos encefálicos y control de las conductas reproductivas Dimorfismos estructurales y funcionales para la preñez y la paternidad 675 677 678 680 RECUADRO 30B LA BUENA MADRE 683 Base celular y molecular de las estructuras y comportamientos sexualmente dimórficos Receptores de esteroides y respuestas en el encéfalo del adulto Trastornos genéticos humanos del sexo genotípico y fenotípico Orientación sexual y encéfalo: análisis molecular y genético Orientación sexual y estructura del encéfalo humano Diferencias de las funciones cognitivas basadas en el sexo Resumen Lecturas adicionales 684 686 688 688 691 692 693 693 CAPÍTULO 31 La memoria 695 Aspectos generales Categorías cualitativas de la memoria humana Categorías temporales de la memoria 695 695 696 RECUADRO 31A MEMORIA FILOGENÉTICA 697 Consolidación de la memoria e impronta La importancia de la asociación en el almacenamiento de información RECUADRO 31B SÍNDROME DE SAVANT Aprendizaje condicionado El olvido RECUADRO 31C CASOS CLÍNICOS QUE PONEN EN EVIDENCIA EL SUSTRATO ANATÓMICO DE LA MEMORIA DECLARATIVA Sistemas encefálicos que subyacen a la adquisición y al almacenamiento de la memoria declarativa Sistemas encefálicos que subyacen a la adquisición y al almacenamiento de la memoria no declarativa Memoria y envejecimiento RECUADRO 31D ENFERMEDAD DE ALZHEIMER Resumen Lecturas adicionales 698 699 700 699 702 703 706 711 712 713 715 715 XX ÍNDICE APÉNDICE ATLAS Resumen de la neuroanatomía humana 717 El sistema nervioso central humano Aspectos generales Terminología neuroanatómica Subdivisiones básicas del sistema nervioso central Anatomía externa de la médula espinal Anatomía interna de la médula espinal Tronco del encéfalo y los nervios craneanos Superficie lateral del encéfalo Superficies dorsal y ventral del encéfalo Superficie mediosagital del encéfalo Anatomía interna del prosencéfalo RECUADRO A TÁLAMO Y RELACIONES TALAMOCORTICALES La irrigación del encéfalo y la médula espinal RECUADRO B ACCIDENTE CEREBROVASCULAR La barrera hematoencefálica Las meninges El sistema ventricular Bibliografía 717 717 718 Lámina 1: Superficie del encéfalo Lámina 2: RM coronal Lámina 3: RM axial Lámina 4: RM sagital Lámina 5: Tronco encéfalico Lámina 6: Médula espinal 745 746 748 750 754 756 758 720 720 722 728 728 729 731 GLOSARIO G-1 REFERENCIAS DE LAS ILUSTRACIONES R-1 732 ÍNDICE ANALÍTICO I-1 734 735 740 741 741 744 materiales ComPlementarios en el sitio web El sitio web complementario de esta nueva edición de Neurociencia, Quinta edición, ofrece, a estudiantes y docentes, valiosos materiales para el aprendizaje y la enseñanza de esta asignatura que se describen a continuación: • Glosario. La terminología completa de la obra. Estudiantes • Tarjetas de aprendizaje (flashcards). Preguntas y respuestas en formato de tarjeta digital para memorización y evaluación. • Animaciones. Un conjunto extenso de animaciones detalladas. • Preguntas de autoevaluación. Preguntas y respuestas en español de cada capítulo. Docentes • Figuras y cuadros. Todas las figuras y cuadros del libro para preparar sus clases y trabajar en proyectos Los docentes tienen acceso también a todos los materiales para los estudiantes. El acceso a este sitio es mediante registro y requiere la validación de la actividad docente en una institución educativa (véase la retiración de cubierta). 1 ESTUDIO DEL SISTEMA NERVIOSO Aspectos generales LA NEUROCIENCIA ENGLOBA UNA AMPLIA GAMA de interrogantes acerca de cómo se organizan los sistemas nerviosos de los seres humanos y otros animales, cómo se desarrollan y cómo funcionan para generar la conducta. Estas preguntas pueden explorarse usando las herramientas analíticas de la genética y la genómica, la biología molecular y celular, la anatomía y la fisiología de los aparatos y sistemas, la biología conductual y la psicología. El desafío principal para un estudiante de neurociencia es integrar el conocimiento diverso derivado de estos distintos niveles de análisis en un conocimiento más o menos coherente de la estructura y la función encefálicas. Muchas de las cuestiones exploradas con éxito se vinculan con el modo en que las células principales de todo sistema nervioso (neuronas y glía) realizan sus funciones básicas en términos anatómicos, electrofisiológicos, celulares y moleculares. Diversos subgrupos de neuronas están reunidos en conjuntos llamados circuitos neurales, y estos circuitos constituyen los componentes primarios de los sistemas nerviosos que procesan tipos específicos de información. Los sistemas neurales cumplen una de tres funciones generales. Los sistemas sensitivos representan la información acerca del estado del organismo y su entorno, los sistemas motores organizan y generan acciones y los sistemas asociativos vinculan los aspectos sensitivos y motores del sistema nervioso y aportan las bases para las funciones “de orden superior” como la percepción, la atención, la cognición, las emociones, el lenguaje y el pensamiento racional. Estas notables capacidades subyacen en el centro de la comprensión de los seres humanos, su historia y, tal vez, su futuro. Genética, genómica y encéfalo El encéfalo, como todos los otros órganos, es el producto de la expresión genética iniciada durante la embriogénesis y mantenida durante toda la vida. Un gen comprende tanto secuencias de DNA codificantes (exones) que constituyen el molde para el RNA mensajero (mRNA) que finalmente se traducirá en proteínas, como secuencias de DNA reguladoras (promotores e intrones) que controlarán si el gen se expresará y en qué cantidad lo hará (es decir, transcrito en mRNA y después traducido en una proteína funcional) en un tipo celular dado. Por lo tanto, el análisis genético es fundamental para conocer la estructura, la función y el desarrollo de los órganos y sistemas orgánicos, incluidos el encéfalo y el sistema nervioso. La llegada de la genómica, que se centra en el análisis de secuencias completas de DNA, tanto codificantes 2 CAPÍTULO 1 RECUADRO 1A Organismos modelo en neurociencia Gran parte de la neurociencia moderna se centra en el conocimiento de la organización y la función del sistema nervioso humano y en las bases anatomopatológicas de las enfermedades neurológicas y psiquiátricas. Sin embargo, a menudo es difícil resolver estas cuestiones con el estudio del encéfalo humano; por consiguiente, los neurocientíficos se basaron en los sistemas nerviosos de otros animales para guiar sus estudios. En los dos últimos siglos, se ha obtenido información crítica sobre la anatomía detallada, la bioquímica, la fisiología, la biología celular y la genética del sistema nervioso mediante el estudio de una amplia variedad de especies. Con frecuencia, la elección del modelo animal analizado refleja la existencia de una mayor capacidad funcional en esa especie; por ejemplo, desde la década de 1950 a la década de 1970, los gatos fueron sometidos a los primeros estudios de la función visual porque eran animales altamente “visuales” y por lo tanto podría esperarse que tuvieran regiones encefálicas bien desarrolladas dedicadas a la visión (regiones similares a las halladas en los primates, incluidos los seres humanos). Gran parte de lo que se conoce en la actualidad sobre la visión humana se basa en estudios que se llevaron a cabo en gatos. Investigaciones en invertebrados como el calamar y la babosa de mar Aplysia californica aportaron ideas de importancia crítica sobre la biología celular básica de las neuronas, la transmisión sináptica y la plasticidad sináptica (la base de aprendizaje y memoria). Tanto el calamar como la babosa de mar fueron elegidos devido a la presencia de algunas células nerviosas excepcionalmente grandes con identidad estereotípica y conexiones que eran idealmente apropiadas para las mediciones físicas. En cada caso, las ventajas que ofrecían estas células excepcionales permitieron realizar experimentos que ayudaron a responder preguntas clave sobre la neurociencia. Siguen realizándose estudios bioquímicos, celulares, anatómicos, fisiológicos y conductuales en una amplia gama de animales. Sin embargo, la secuenciación completa de los genomas de especies de invertebrados, vertebrados y mamíferos ha conducido a que muchos neurocientíficos adopten de manera informal los cuatro organismos “modelo” sobre la base de la capacidad para realizar análisis genético y la manipulación en cada una de estas especies. La mayoría de los genes del genoma humano se expresan en el encéfalo en desarrollo y adulto. Lo mismo sucede con el vermes nematodo Caenorhabditis elegans, la mosca de la fruta Drosophila melanogaster, el pez cebra Daniorerio y el ratón Mus musculus (cuatro especies utilizadas comúnmente en la genética moderna y usadas cada vez más en neurociencias). A pesar de ciertas limitaciones en cada una de estas especies, la disponibilidad de su secuencia genómica completa facilita la investigación en una amplia gama de preguntas a nivel molecular, celular, anatómico y fisiológico. Una ventaja de estas especies modelo es que la abundancia de información genética y genómica de cada una permite una manipulación sofisticada de la expresión y la función de los genes. Por lo tanto, una vez que se identifica un gen crítico para el desarrollo o la función del encéfalo, es posible manipularlo específicamente en el gusano, la mosca, el pez o el ratón. En el gusano, la mosca y el pez, las evaluaciones a gran escala de animales mutantes cuyos genomas han sido modificados al azar por mutágenos químicos permite a los investigadores buscar cambios de la estructura y la función de desarrollo normal (el “fenotipo”) e identificar genes que son importantes para aspectos específicos de la arquitectura o la función encefálica. Algunos esfuerzos similares, aunque de alcance más limitado, han identificado en el ratón mutaciones espontáneas o inducidas que interrumpen el desarrollo o la función encefálica. Además, las manipulaciones que conducen a los denominados animales transgénicos permiten introducir genes en el genoma, o producir su deleción o su mutación (“desactivación”) utilizando la notable capacidad de los genomas de dividirse en nuevas secuencias similares a los genes endógenos. Esta capacidad, denominada recombinación homóloga, permite insertar constructos de DNA que interrumpen o alteran la expresión de genes específicos en el lugar del gen normal en la especie huésped. Estos enfoques permiten una evaluación amplia de las consecuencias de eliminar o alterar la función génica. Por supuesto, los neurocientíficos estudian también los sistemas nerviosos y los comportamientos de otras especies. Crustáceos como el cangrejo y la langosta, e insectos como el saltamontes y la cucaracha han sido útiles para discernir reglas básicas que gobiernan la función de los circuitos neurales. Aves y anfibios, en forma de pollos y ranas, siguen siendo de particular utilidad para estudiar el desarrollo neural temprano, y mamíferos como la rata se utilizan ampliamente para estudios neurofarmacológicos y conductuales de la función encefálica del adulto. Por último, primates no humanos (el mono Rhesus en particular) proporcionan oportunidades para estudiar funciones complejas que se aproximan estrechamente a las que se llevan a cabo en el encéfalo humano. Sin Ser humano ~20 000 Ratón ~25 000 Tamaños estimados de los genomas de seres D. rerio (pez cebra) ~24 000 humanos y varias especies “modelo”. Obsérvese que la cantidad de genes en el genoma de un organismo no se correlaciona con la complejidad de la célula ni del organismo; por ejemplo, el simple nematodo Caenorhabditis elegans tiene casi la misma cantidad de genes que un ser humano. Mucha actividad genética depende de factores de transcripción que regulan cuándo y en qué grado se expresa un gen dado. D. melanogaster (mosca de la fruta) ~15 000 C. elegans (nematodo) ~19 000 0 10 000 20 000 Número de genes 30 000 ESTUDIO DEL SISTEMA NERVIOSO 3 RECUADRO 1A (continúa) embargo, ninguna de estas especies permite realizar manipulaciones genéticas y genómicas como las cuatro especies que describimos antes. Cada sistema modelo ha supuesto contribuciones importantes a nuestro conocimiento actual de la estructura, la función y el desarrollo del encéfalo humano. El uso de animales especializados para funciones clave como la visión ha permitido conocer funciones encefálicas humanas paralelas, mientras que el uso de modelos animales manipulados genéticamente ha permitido el descubrimiento de genes clave para el desarrollo y la función del encéfalo, así como explorar la función biológica celular de los genes asociados a enfermedades del encéfalo humano. Bibliografía Bockamp, E. and 7 others (2002) Of mice and models: Improved animal models for biomedical research. Physiol. Genomics 11:115-132. como reguladoras, identificadas para una especie o un individuo, ha permitido conocer de qué modo el DNA nuclear proporciona instrucciones acerca del ensamblado y las funciones del encéfalo. Sobre la base de estimaciones actuales, el genoma humano contiene aproximadamente 20 000 genes, de los cuales se expresan unos 14 000 en el encéfalo en desarrollo o maduro (Fig. 1.1A). De este subgrupo, unos 8 000 se expresan en todas las células y los tejidos. Por lo tanto, mucha información genética Muquit, M. M. .and M. B. Feany (2002) Modelling neurodegenerative diseases in Drosophila: A fruitful approach? Nature Rev. Neurosci. 3: 237-243. Sengupta, P. and A. D. Samuel (2009) Caenorhabditis elegans: A model system for systems neuroscience. Curr. Opin. Neurobiol. 19: 637643. Rinkwitz, S., P. Mourrain and T. S. Becker (2011) Zebrafish: An integrative system for neurogenomics and neurosciences. Prog. Neurobiol. 93: 231-243. “específica del encéfalo” reside en los intrones y las secuencias reguladoras que controlan oportunidad, cantidad, variabilidad y especificidad celular de la expresión genética. A pesar de la cantidad de genes que comparten el encéfalo y otros tejidos, los genes individuales varían en el nivel y la localización de la expresión en regiones y células encefálicas específicas (es decir, en la cantidad de mRNA expresado de una región a otra; Fig. 1.1B). Estas diferencias constituyen el fundamento de la diversidad y la complejidad de las funciones encefálicas. (A) FIGURA 1.1 El genoma y el encéfalo. (A) En este diagrama de Venn del genoma humano, las regiones azul y violeta representan aquellos genes que son expresados ~6 000 genes expresados en el encéfalo ~8 000 genes ubicuos expresados en todas las células y tipos de tejido ~6 000 genes no expresados en el encéfalo Aproximadamente 14 000 de los 20 000 genes humanos se expresan en el encéfalo en desarrollo o adulto (B) selectivamente en el encéfalo junto con los que son expresados de forma ubicua, y que por lo tanto se encuentran en el encéfalo y en todos los demás tejidos. (B) Localizaciones y niveles cuantitativos de expresión de un gen único en el encéfalo humano. Los puntos indican las regiones encefálicas donde puede hallarse el mRNA para este gen particular, mientras que su color (azul a anaranjado) indica el nivel relativo (menor a mayor) de mRNA detectado en cada lugar. (C) Consecuencia de una mutación de gen único para el desarrollo encefálico. Un único gen, ASPM (“abnormal spindle-like microcephaly-associated protein”, proteína asociada a la microcefalia del tipo huso anormal), afecta la función de una proteína asociada con los husos mitóticos y conduce a microcefalia. En un paciente portador de la mutación ASPM (izquierda), el tamaño global del encéfaalo está espectacularmente reducido y la organización anatómica está distorsionada en comparación con el encéfalo de un control normal (derecha) de edad y sexo similar. (A, datos de Ramsköld y cols., 2009; B, cortesía del Allen Brain Atlas; C, de Bond y cols., 2002.) (C) 4 CAPÍTULO 1 Una de las consecuencias más prometedoras de la secuenciación del genoma humano fue el reconocimiento de que uno o algunos genes, cuando se alteran (mutan), pueden explicar ciertos aspectos de las enfermedades neurológicas y psiquiátricas. Mediante el uso de las herramientas de genética y genómica, se han identificado mutaciones de gen único que conducen a cambios raros pero devastadores en el desarrollo y la función del encéfalo. Por ejemplo, una mutación en un único gen que regula la mitosis puede conducir a una microcefalia, trastorno en el cual el encéfalo y la cabeza dejan de crecer y la función encefálica disminuye drásticamente (Fig. 1.1C). Además de los genes que interrumpen el desarrollo encefálico, se han observado genes mutantes que pueden producir trastornos como las enfermedades de Huntington, Parkinson y Alzheimer. El uso de la genética y la genómica para comprender las enfermedades del desarrollo y el sistema nervioso adulto permite un conocimiento más profundo de la patología y plantea la esperanza de la aparición de nuevas opciones a la hora de diseñar terapias racionales. Pero la relación entre genotipo y fenotipo no es sólo el resultado (A) Célula piramidal cortical (B) Célula bipolar retiniana Dendritas de seguir las instrucciones genéticas, y la información genómica aislada no puede explicar de qué modo opera el encéfalo en los individuos normales, o de qué manera los procesos patológicos interrumpen las funciones encefálicas normales. Para comprender cómo funciona el encéfalo y el resto del sistema nervioso en la salud y la enfermedad, debemos conocer la biología celular, la anatomía y la fisiología de las células componentes, así como los circuitos que la forman. Los componentes celulares del sistema nervioso A comienzos del siglo xix, la célula se reconocía como la unidad fundamental de todos los organismos vivos. Sin embargo, no fue hasta bien entrado el siglo xx cuando los neurocientíficos acordaron en que el tejido nervioso, al igual que todos los otros órganos, está formado por estas unidades fundamentales. La razón principal fue que la primera generación de neurobiólo- (D) Célula amacrina retiniana (C) Célula ganglionar retiniana Dendritas Dendritas Cuerpo celular Cuerpo celular Dendritas Axón Axón Cuerpo celular * (F) Células de Purkinje del cerebelo (E) Neuronas del núcleo mesencefálico del nervio craneal V Cuerpo celular Cuerpos celulares Dendritas Axón * Axones * FIGURA 1.2 Ejemplos de la gran variedad de morfologías de las células nerviosas halladas en el sistema nervioso humano. Los trazados provienen de células nerviosas reales teñidas por impregnación con sales de plata (la denominada técnica de Golgi, método utilizado en los estudios clásicos de Golgi y Ramón y Cajal). Los asteriscos indican que el axón recorre mucho más de lo que se muestra. Obsérvese que algunas células, como la bipolar de la retina, tienen un axón muy corto, y que otras, como la célula amacrina retiniana, carecen de axón. Los dibujos no están todos en la misma escala. Cuerpo celular Axón * ESTUDIO DEL SISTEMA NERVIOSO gos “modernos” del siglo xix tuvo dificultades para distinguir la naturaleza unitaria de las células nerviosas con los microscopios y técnicas de tinción celular disponibles entonces. Esta deficiencia se vio agravada por las configuraciones extraordinariamente complejas y las ramificaciones extensas de las células nerviosas individuales, todas las cuales se encuentran empaquetadas juntas y, por lo tanto, es difícil distinguirlas, lo que oscurecía aún más su semejanza con las células de configuración más sencillas de otros tejidos (Fig. 1.2). Algunos biólogos de esa era llegaron incluso a la conclusión de que cada célula nerviosa estaba conectada con sus vecinas por nexos protoplasmáticos que formaban una red continua de células nerviosas, o retículo. La “teoría reticular” de la comunicación de las células nerviosas fue propuesta por el neuropatólogo italiano Camillo Golgi. Este autor hizo muchas contribuciones importantes a la ciencia médica, que incluyen la identificación del orgánulo celular que finalmente se denominó aparato de Golgi, la técnica de tinción celular de importancia crítica que lleva su nombre (véanse Figs. 1.2 y 1.6) y el conocimiento de la fisiopatología del paludismo. Por último, su teoría reticular del sistema nervioso perdió apoyo y fue reemplazada por la que llegó a conocerse como la “doctrina de la neurona”. Los autores principales de esta nueva perspectiva fueron el neuroanatomista español Santiago Ramón y Cajal y el fisiólogo británico Charles Sherrington. Los puntos de vista opuestos representados por Golgi y Cajal generaron a comienzos del siglo xx un debate encendido que sentó las bases de la neurociencia moderna. Fundamentado en el examen del tejido nervioso teñido con sales de plata con el microscopio óptico según un método propuesto por Golgi, Cajal argumentó persuasivamente que las células nerviosas son entidades separadas y que se comunican entre ellas por medio de contactos especializados que no son sitios de continuidad entre las células. Sherrington, que había estado trabajando en la transferencia aparente de señales eléctricas a través de las vías reflejas, denominó a estos contactos especializados sinapsis. A pesar del éxito final d la hipótesis de Cajal sobre la de Golgi, ambos recibieron el premio Nobel de Fisiología o Medicina en 1906 por sus contribuciones esenciales al conocimiento de la organización del encéfalo; del mismo modo, en 1932 Sherrington fue reconocido por sus contribuciones. El trabajo posterior de Sherrington y cols., que demostró la transferencia de señales eléctricas entre las uniones sinápticas de las células nerviosas, brindaba apoyo firme a la doctrina neuronal, pero quedaba el desafío de explicar la autonomía de las neuronas individuales. No fue hasta la aparición de la microscopia electrónica en la década de 1950 cuando se resolvieron todas las dudas acerca de la separación de las neuronas. Las imágenes de alta amplificación y alta resolución obtenidas con el microscopio electrónico (Fig. 1.3) establecieron con claridad que las células nerviosas constituyen unidades funcionalmente independientes; estas micrografías también permitieron identificar las uniones celulares especializadas que Sherrington había denominado sinapsis. Sin embargo, tal vez como consuelo tardío para Golgi, los estudios de microscopia electrónica también de- 5 mostraron continuidades intercelulares especializadas (aunque relativamente poco frecuentes) entre algunas neuronas. Estas continuidades o uniones en hendidura o en brecha, son similares a las halladas entre las células en epitelios como el pulmón y el intestino. En realidad, las uniones en hendidura permiten la continuidad citoplasmática y la transferencia directa de señales eléctricas y químicas entre células del sistema nervioso. Los estudios histológicos de Ramón y Cajal, Golgi y un conjunto de sucesores condujeron a un mayor consenso respecto de que las células del sistema nervioso pueden dividirse en dos categorías amplias: células nerviosas (o neuronas) y células de sostén llamadas neuroglía (o simplemente glía). Las células nerviosas están especializadas en la señalización eléctrica en largas distancias. El conocimiento de este proceso representa uno de los éxitos más espectaculares de la biología moderna y es el tema de la Unidad I de este libro. Al contrario de las células nerviosas, las de sostén mantienen las señales eléctricas en lugar de generarlas. También cumplen otras funciones en el encéfalo en desarrollo y adulto. Tal vez la más importante es que la neuroglía contribuye esencialmente a la reparación del sistema nervioso dañado, ya que actúa como células madre en algunas regiones encefálicas, promueve el nuevo crecimiento de las neuronas dañadas en regiones donde puede ocurrir una regeneración útil e impide la regeneración en otras regiones donde el nuevo crecimiento descontrolado podría producir más daños que beneficios (véase luego y Unidad IV). Las neuronas y la glía comparten el conjunto de orgánulos hallado en todas las células, en el que se incluye el retículo endoplasmático y el aparato de Golgi, las mitocondrias y distintas estructuras vesiculares. Sin embargo, en las neuronas y la glía estos orgánulos a menudo son más destacables en distintas regiones de la célula. Por ejemplo, las mitocondrias suelen estar concentradas en las sinapsis de las neuronas, mientras que los orgánulos que sintetizan proteínas, como el retículo endoplasmático, están excluidos en gran parte de los axones y las dendritas. Además de la distribución de los orgánulos y los componentes subcelulares, en cierta medida las neuronas y la glía son diferentes de otras células en las proteínas fibrilares o tubulares especializadas que constituyen el citoesqueleto (véase Fig. 1.4). Aunque muchas de estas proteínas (isoformas de actina, tubulina y miosina, así como varias otras) se encuentran en otras células, su organización distinta en las neuronas es fundamental para la estabilidad y la función de las prolongaciones neuronales y las uniones sinápticas. Otras proteínas filamentosas caracterizan a las células de la glía y contribuyen con sus funciones. Los distintos filamentos, los túbulos, los motores subcelulares y las proteínas de andamiaje del citoesqueleto neuronal y glial cumplen muchas funciones, incluidos la migración de las células nerviosas, el crecimiento de axones y dendritas; el tráfico y el posicionamiento apropiado de los componentes de la membrana, orgánulos y vesículas, y los procesos activos de exocitosis y endocitosis que subyacen a la comunicación sináptica. El conocimiento de las formas en que se utilizan estos componentes moleculares para asegurar el correcto desarrollo y funcionamiento de las neuronas y la glía aún es un enfoque fundamental para la neurobiología moderna. 6 CAPÍTULO 1 FIGURA 1.3 Características principales de las neuronas al observarlas con microscopio óptico y microscopio electrónico. (A) Diagrama de células nerviosas y sus partes componentes. (B) Segmento inicial del axón (azul) que entra en la vaina de mielina (dorado). (C) Botones terminales (azul) cargados con vesículas sinápticas (puntas de flecha) que forman sinapsis (flechas) con una dendrita (púrpura). (D) Corte transversal de axones (azul) envainados por las prolongaciones de los oligodendrocitos (dorado). (E) Dendritas atípicas (púrpura) de las células piramidales corticales. (F) Cuerpos de células nerviosas (púrpura) ocupados por grandes núcleos redondeados. (G) Porción del axón mielínico (azul) que ilustra los intervalos entre segmentos adyacentes de mielina (dorado) denominados nodos de Ranvier (flechas). (Microfotografías de Peters y cols., 1991.) (B) Axón (A) Retículo endoplasmático Mitocondria Neuronas Las neuronas se distinguen por especializarse en la comunicación intercelular y en la señalización eléctrica momento a momento. Estos atributos se manifiestan en su morfología general, en la organización específica de los componentes de la membrana para la señalización eléctrica y en la complejidad de las estructuras y funciones de los contactos sinápticos entre las neuronas (véase Fig. 1.3C). El signo más evidente de especialización neuronal para la comunicación a través de la señalización eléctrica es la ramificación extensa de las neuronas. Los dos aspectos más sobresalientes de esta ramificación en las células nerviosas típicas son el axón y la arborización compleja de (C) Terminaciones sinápticas (botones terminales) , Núcleo , Soma Dendrite Dendrita Aparato de Golgi (D) Axones mielínicos * ) Ribosomas Axones . + (E) Dendritas (F) Cuerpo celular (soma) (G) Axón mielínico y nodo de Ranvier ESTUDIO DEL SISTEMA NERVIOSO dendritas que surgen del cuerpo de la célula neuronal en forma de ramas dendríticas (o prolongaciones dendríticas; véase Fig. 1.3E). Las dendritas son la diana primaria de las aferencias sinápticas desde otras neuronas y se distinguen por su alto contenido de ribosomas y proteínas específicas del citoesqueleto. Algunas neuronas carecen de dendritas, mientras que otras presentan arborizaciones dendríticas que rivalizan en complejidad con un árbol maduro (véase Fig. 1.2). El número de aferencias que recibe una neurona particular depende de la complejidad de su arborización dendrítica: las células nerviosas que carecen de dendritas están inervadas sólo por una o algunas otras células nerviosas, mientras que las que presentan dendritas cada vez más elaboradas están inervadas por una cantidad comparablemente mayor de otras neuronas. El número de aferencias para una sola neurona refleja el grado de convergencia, mientras que el número de sitios diana inervados por cualquier neurona representa su divergencia. Los contactos sinápticos que se hacen sobre las dendritas (y, menos a menudo, sobre los cuerpos de las células neuronales) comprenden una elaboración especial del aparato secretor que se encuentra en la mayoría de las células epiteliales polarizadas. En condiciones típicas, la neurona presináptica es inmediatamente adyacente a una región especializada de los receptores postsinápticos de la célula diana. En la mayoría de las sinapsis, no hay continuidad física entre estos elementos presinápticos y postsinápticos. En cambio, los componentes presinápticos y postsinápticos se comunican a través de la secreción de moléculas desde la terminación presináptica que se unen a receptores en la célula postsináptica. Estas moléculas deben atravesar un intervalo de espacio extracelular entre los elementos presináptico y postsináptico llamado hendidura sináptica. Sin embargo, esta hendidura no es simplemente un espacio vacío; más bien, es el lugar donde las proteínas extracelulares que influyen en la difusión, la unión y la degradación de las moléculas secretadas por la terminación presináptica (véase Cap. 5). El número de aferencias sinápticas que recibe cada célula nerviosa en el sistema nervioso humano varía entre 1 y 100 000. Este rango refleja un propósito fundamental de las células nerviosas: transmitir e integrar la información proveniente de otras neuronas en un circuito neural. Por lo tanto, el número de contactos sinápticos provenientes de distintas neuronas presinápticas en cualquier célula particular es un determinante de especial importancia en la función neuronal. La información transmitida por las sinapsis a las dendritas neuronales es integrada y “leída” en el origen del axón, parte de la célula nerviosa especializada en la transmisión de señales eléctricas (véase Fig. 1.3B). El axón es una extensión singular del cuerpo de las células neuronales que puede viajar algunos cientos de micrómetros (μm) o mucho más lejos, según el tipo de neurona y el tamaño del animal (algunos axones en los animales grandes pueden tener metros de longitud). Además, el axón posee un citoesqueleto característico cuyos elementos son fundamentales para su integridad funcional (Fig. 1.4). Muchas células nerviosas del encéfalo humano tienen axones cuya longitud no es superior a unos milímetros, y algunas no tienen ningún axón. 7 Los axones relativamente cortos son característicos de las neuronas de circuito local o interneuronas en todo el encéfalo. Sin embargo, los axones de las neuronas de proyección se extienden hasta dianas distantes. Por ejemplo, los axones que discurren desde la médula espinal humana hasta el pie tienen una longitud aproximada de un metro. El acontecimiento que transporta señales por estas distancias es una onda de actividad eléctrica que se autorregenera denominada potencial de acción, un cambio todo o nada del potencial eléctrico (voltaje) a través de la membrana de la célula nerviosa que transmite información de un punto a otro del sistema nervioso (véase Cap. 2). Un potencial de acción se propaga desde su punto de inicio en el cuerpo celular (llamado botón axónico) hacia la terminación del axón, donde ocurren los contactos sinápticos. El proceso químico y eléctrico por el cual la información codificada por los potenciales de acción se transmite en los contactos sinápticos hacia la célula diana se denomina transmisión sináptica (véase Cap. 5). Típicamente, las terminaciones presinápticas (también llamadas terminaciones sinápticas, terminaciones axónicas o botones terminales; véase Fig. 1.3C) y sus especializaciones postsinápticas son sinapsis químicas, el tipo más abundante de sinapsis del sistema nervioso. Otro tipo, la sinapsis eléctrica (facilitada por las uniones en hendidura o en brecha mencionadas antes), es relativamente rara y tiene funciones especiales. Los orgánulos secretores en la terminación presináptica de las sinapsis químicas son vesículas sinápticas, estructuras esféricas llenas de moléculas de neurotransmisor (o simplemente, neurotransmisores). La colocación de las vesículas sinápticas en la membrana presináptica y su fusión para iniciar la liberación del neurotransmisor están regulados por algunas proteínas (incluidas varias del citoesqueleto) que se encuentran dentro de la vesícula o se asocian con ella. Los neurotransmisores liberados en las vesículas sinápticas modifican las propiedades eléctricas de la célula diana al unirse a receptores, que se ubican fundamentalmente en las especializaciones postsinápticas. La actividad compleja y coordinada de neurotransmisores, receptores, elementos relacionados del citoesqueleto y moléculas de transducción de señales conforma la base para que las células nerviosas se comuniquen entre ellas y con las células efectoras en músculos y glándulas. Células neurogliales Las células neurogliales (también denominadas más simplemente glía) son muy diferentes de las células nerviosas. Son más numerosas que las neuronas en el encéfalo, a las que superan en número en una relación, tal vez, de 3 a 1. La diferencia principal es que las células neurogliales no participan directamente en las interacciones sinápticas y en la señalización eléctrica, aunque sus funciones de sostén ayudan a definir contactos sinápticos y a mantener la capacidad de señalización de las neuronas. Aunque las células neurogliales también tienen prolongaciones complejas que se extienden desde sus cuerpos celulares, por lo general son menos sobresalientes que las ramificaciones neuronales, y 8 CAPÍTULO 1 (A) (B) (C) FIGURA 1.4 Disposición característica de los elementos del citoesqueleto en las neuronas. (A) El cuerpo celular, los axones y las dendritas se distinguen por la distribución de tubulina (verde a través de la célula) a diferencia de otros elementos del citoesqueleto (en este caso, Tau [rojo], una proteína fijadora de microtúbulos que se (D) encuentra sólo en los axones). (B) Se muestra aquí la localización notablemente distinta de la actina (rojo) en los extremos en crecimiento de las prolongaciones axónicas y dendríticas en una neurona cultivada tomada del hipocampo. (C) Por el contrario, en una célula epitelial cultivada, la actina (rojo) está distribuida en fibrillas que ocupan la mayor parte del cuerpo celular. (D) En células astrogliales en cultivo, la actina (rojo) también se ve en haces fibrilares. (E) Se observa tubulina (verde) en todo el cuerpo celular y las dendritas de las neuronas. (F) Aunque la tubulina es un componente importante de las dendritas, que se extiende en las espinas, la cabeza de la espina es rica en actina (rojo). (G) El componente de tubulina del citoesqueleto en células no neuronales está dispuesto en redes filamentosas. (H-K) Las sinapsis (E) (G) tienen una disposición distinta de elementos del citoesqueleto, receptores y proteínas de andamiaje. (H) Se observan dos axones (verde, tubulina) de neuronas motoras que emiten cada uno dos ramas a cuatro fibras musculares. El rojo muestra el agrupamiento de receptores postsinápticos (en este caso, para el neurotransmisor acetilcolina). (I) Una imagen a mayor aumento de una sola neurona motora muestra la relación entre el axón (verde) y los receptores postsinápticos (rojo). (J) Se muestra en verde el espacio extracelular entre el axón y su músculo diana. (K) Se muestra en verde el agrupamiento de proteínas de andamiaje (en este caso, distrofina) que localizan receptores y los conectan con otros elementos del citoesqueleto. (A, cortesía de Y.N. Jan; B, cortesía de E. Dent y F. Gertler; C, cortesía de D. Arneman y C. Otey; D, cortesía de A. Gonzales y R. Cheney; E, tomado de Sheng, (F) 2003; F, tomado de Matus, 2000; G, cortesía de T. Salmon y cols.; H-K, cortesía de R. Sealock.) (H) (J) (I) (K) no cumplen los mismos propósitos que los axones y las dendritas. Las células con características gliales son las únicas células madre aparentes que el encéfalo maduro retiene y son capaces de dar origen a nuevas células neurogliales y, en algunos casos, a nuevas neuronas. El término glía (de la palabra griega que significa “pegamento”) refleja la suposición existente en el siglo xix de que estas células de alguna forma mantienen unido el sistema nervioso. El término sobrevivió a pesar de la falta evidencias de que las células neurogliales realmente unan las células nerviosas en su conjunto. Las funciones que están bien establecidas de las células gliales son: mantener el medio iónico de las células nerviosas, modular la velocidad de propagación de las señales nerviosas, modular la acción sináptica al controlar la captación de neurotransmisores en la hendidura sináptica o cerca de ella, proporcionar un andamiaje para ciertos aspectos del desarrollo neural y ayudar en la recuperación de la lesión neural (o, en algunos casos, impedirla). Hay tres tipos de células neurogliales en el sistema nervioso central maduro: astrocitos, oligodendrocitos y células microgliales. Los astrocitos, que están limitados al sistema nervioso ESTUDIO DEL SISTEMA NERVIOSO central (es decir, al encéfalo y la médula espinal), tienen prolongaciones locales elaboradas que aportan a estas células un aspecto estrellado (de ahí el prefijo “astro”) (Figs. 1.5A, F). Una función importante de los astrocitos es mantener, de distintas formas, un entorno químico apropiado para la señalización neuronal. Además, algunas observaciones recientes sugieren que un subgrupo de astrocitos del encéfalo del adulto retiene las características de la célula madre (Fig. 1.5D), es decir, la capacidad de entrar en las mitosis y generar todas las clases de células que se encuentran en el tejido nervioso. Los oligodendrocitos, también restringidos al sistema nervioso central, aportan una envoltura laminada y rica en lípidos llamada mielina alrededor de algunos axones, pero no de todos (Figs. 1.5B, G, H). Además, la mielina tiene efectos importantes sobre la velocidad de transmisión de señales eléctricas (véase Cap. 3). En el sistema periférico, las células que elaboran mielina se denominan células de Schwann. En el sistema nervioso maduro, subgrupos de células oligodendrogliales y de células de Schwann retienen las propiedades de las células madre neurales y pueden generar nuevos oligodendrocitos y células de Schwann en respuesta al traumatismo o la enfermedad (Fig. 1.5E). Por último, las células microgliales derivan en mayor medida de células precursoras hematopoyéticas (aunque algunas pue(A) Astrocito FIGURA 1.5 Variedades de células neurogliales. (A-C) Representaciones de células gliales diferenciadas en el sistema nervioso maduro visualizadas utilizando el método de Golgi incluyen un astrocito (A), un oligodendrocito (B) y una célula microglial (C) Las tres imágenes se encuentran aproximadamente en la misma escala. (D) Las células madre gliales del sistema nervioso maduro incluyen células madre con propiedades de astrocitos que pueden dar origen a neuronas, astrocitos y oligodendrocitos. (E) Otra clase de célula madre glial, el precursor oligodendroglial, tiene un potencial más limitado y da origen principalmente a oligodendrocitos diferenciados. (F) Astrocitos en cultivo tisular, marcados (rojo) con un anticuerpo contra una proteína específica del astrocito. (G) Células oligodendrogliales (verdes) en cultivo tisular marcado con un anticuerpo contra una proteína específica de la oligodendroglia. (H) Los axones periféricos están envainados por mielina (rojo marcado) excepto en los nodos de Ranvier (véase Fig. 1.3G). La marca verde indica canales iónicos (véase Cap.4) concentrados en el nodo; la marca azul indica una región que presenta diferencias moleculares llamada paranodo. (I) Células microgliales de la médula espinal, marcadas con un anticuerpo específico de tipo celular. Recuadro: imagen con mayor amplificación de una sola célula microglial teñida con un marcador selectivo de los macrófagos. (A-C, tomados de Jones y Cowan, 1983; D, E, tomados de Nishiyama y cols., 2009; F, G, cortesía de A.-S. LaMantia; H, cortesía de M. Bhat; I, cortesía de A. Light; recuadro, cortesía de G. Matsushima.) (C) Célula microglial (B) Oligodendrocito Cuerpo celular Prolongaciones gliales (E) Precursor del oligodendrocito (D) Célula madre glial Neuronas existentes Neuronas nuevas Célula madre glial Oligodendrocito mielinizador Vaso sanguíneo (F) (G) (H) (I) 9 10 CAPÍTULO 1 den hacerlo directamente de células precursoras neurales). Estas células comparten muchas propiedades con los macrófagos que se encuentran en otros tejidos, y son fundamentalmente células limpiadoras que eliminan los restos celulares de sitios de lesión o de recambio celular normal. Además, la microglía, como sus análogos los macrófagos, secreta moléculas de señalización (sobre todo una amplia gama de citocinas producidas también por células del sistema inmunitario) que pueden modular la inflamación local e influir en la supervivencia o la muerte celular. En efecto, algunos neurobiólogos prefieren categorizar la microglía como un tipo de macrófago. Tras el daño encefálico, la cantidad de células microgliales en el sitio de lesión aumenta de forma espectacular. Algunas de estas células proliferan a partir de la microglía existente en el encéfalo, mientras que otras provienen de macrófagos que migran hacia el área lesionada y entran en el encéfalo a través de interrupciones locales en la vascularización cerebral. Además de las tres clases de glía diferenciada, también se encuentran en todo el encéfalo del adulto células madre gliales. Estas células retienen la capacidad de proliferar y generar precursores adicionales o glía diferenciada y, en algunos casos, neuronas. Las células madre gliales del encéfalo maduro pueden dividirse en dos categorías: astrocitos, que se encuentran principalmente cerca de los ventrículos en una región denominada zona subventricular o adyacentes a los vasos sanguíneos de la zona ventricular (Fig. 1.5D), y precursores oligodendrogliales, dispersos en toda la sustancia blanca, y que a veces se denominan polidendrocitos (Fig. 1.5E). Tanto in vivo como in vitro, los astrocitos de la zona subventricular in vitro pueden dar origen a más células madre, neuronas y astrocitos y oligodendrocitos maduros. Por lo tanto, poseen las propiedades fundamentales de todas las células madre: proliferación, autorrenovación y capacidad de formar todas las clases celulares de un tejido particular. Los precursores oligodendrogliales tienen una capacidad más limitada. En el encéfalo de un ser vivo, originan principalmente células oligodendrogliales maduras y algunos astrocitos, aunque se ha observado que generan neuronas en ciertas condiciones in vitro. No queda clara la importancia de las células madre gliales activas en el encéfalo maduro. Ellas pueden reflejar la identidad glial como “defecto” de cualquier célula proliferativa derivada de los precursores embrionarios del sistema nervioso o evidenciar distinciones en el estado diferenciado de las neuronas y la glía que sólo permiten la proliferación en células con características gliales. celulares distintos (ya sea categorizados por morfología, identidad molecular o actividad fisiológica) que cualquier otro sistema orgánico (un acontecimiento que se presume puede explicar por qué en el sistema nervioso se expresan tantos genes diferentes; véase antes). Sin duda, la diversidad celular de cualquier sistema nervioso subyace a la capacidad del sistema para formar redes cada vez más complicadas que median conductas cada vez más sofisticadas. Durante gran parte del siglo xx, los neurocientíficos se basaron en el mismo conjunto de técnicas desarrolladas por Ramón y Cajal y Golgi y otros pioneros de la histología (el análisis microscópico de células y tejidos) y en la anatomía patológica para describir y categorizar la diversidad de tipos celulares en el sistema nervioso. El método de tinción denominado de Golgi permitió visualizar células nerviosas individuales y sus prolongaciones que habían sido impregnadas, al parecer aleatoriamente, con sales de plata (Fig. 1.6A, B). Como análogo moderno, los colorantes de fluoresceína y otras moléculas solubles inyectadas en neuronas aisladas, a menudo después del registro fisiológico para identificar la función de la célula, proporcionan un enfoque alternativo para visualizar células nerviosas aisladas y sus prolongaciones (Figura 1.6C, D). Como complemento de estos enfoques (que proporcionan una muestra aleatoria que sólo contiene algunas neuronas y glía), se utilizan otras tinciones para demostrar la distribución de todos los cuerpos celulares (pero no sus prolongaciones ni conexiones) en el tejido nervioso. El método de Nissl, ampliamente utilizado, es uno de ellos; esta técnica tiñe el nucléolo y otras estructuras (p. ej., ribosomas) donde hay DNA o RNA (Fig. 1.6E). Estas tinciones demuestran que el tamaño, la densidad y la distribución de la población total de células nerviosas no son uniformes dentro del encéfalo. En algunas regiones, como la corteza cerebral, las células están dispersas en capas (Figura 1-6F, G), y cada capa se define por diferencias características de la densidad celular. Algunas estructuras, como el bulbo olfatorio, muestran incluso organizaciones más complicadas de los cuerpos celulares (Fig. 1.6H). Otros enfoques, detallados más adelante en el capítulo, han proporcionado una mayor definición de las diferencias entre las células nerviosas de una región a otra. Éstos incluyen la identificación del modo en que se conectan entre sí los subgrupos de neuronas y de qué modo las diferencias moleculares distinguen mejor las clases de células nerviosas en distintas regiones del encéfalo (véase Fig. 1.11). Circuitos neurales Diversidad celular en el sistema nervioso Aunque los componentes celulares del sistema nervioso central son similares en varios aspectos a los de otros órganos, resultan inusuales por su número extraordinario: se estima que el encéfalo humano contiene 100 000 millones de neuronas y varias veces esa cantidad en células de sostén. Lo que es más importante, el sistema nervioso tiene un rango mayor de tipos Las neuronas nunca funcionan de forma aislada; están organizadas en conjuntos o circuitos neurales que procesan tipos específicos de información y aportan las bases para la sensación, la percepción y la conducta. Típicamente, las conexiones sinápticas que definen estos circuitos se realizan en una maraña densa de dendritas, terminaciones axónicas y prolongaciones de células gliales que en conjunto constituyen lo que se denomina neurópilo (el sufijo -pilo proviene de la palabra griega pilos, que significa “sentido”; véase Fig. 1.3C). Por lo tanto, el neurópilo ESTUDIO DEL SISTEMA NERVIOSO (A) (B) (C) (D) (E) (F) (G) (H) 11 FIGURA 1.6 Visualización de las células nerviosas. (A) Neuronas corticales teñidas con técnica de Golgi (impregnación con sales de plata). (B) Células de Purkinje cerebelosas marcadas con técnica de Golgi. Estas células tienen una sola dendrita apical sumamente ramificada. (C) La inyección intracelular de colorante fluorescente marca dos neuronas retinianas que varían mucho en el tamaño y en la extensión de sus arborizaciones dendríticas. (D) La inyección intracelular de una enzima marca una neurona en un ganglio del sistema nervioso autónomo (involuntario). (E) El colorante violeta de cresilo tiñe el RNA de todas las células de un tejido y marca el nucléolo (pero no el núcleo) así como el retículo endoplasmático, rico en ribosomas. Las dendritas y los axones no son marcados, lo que explica los espacios “vacíos” entre estas neuronas. (F) Un corte de la corteza cerebral teñido con técnica de Nissl demuestra laminación –cuerpos celulares dispuestos en capas de diferentes densidades–. Las diferentes densidades laminares definen límites entre áreas corticales con distintas funciones. (G) Amplificación mayor de la corteza visual primaria que se ve en el lado izquierdo del panel (F). Claras diferencias en la densidad celular definen las láminas de la corteza visual primaria y diferencian esta región de otras áreas corticales cerebrales. (H) La coloración de Nissl de los bulbos olfatorios muestra una distribución característica de los cuerpos celulares, especialmente aquellos dispuestos en anillos sobre la superficie externa del bulbo. Estas estructuras, junto con el tejido pobre en células ubicado dentro de cada anillo, se denominan glomérulos. (C, cortesía de C. J. Shatz; todas las demás, cortesía de A.-S. LaMantia y D. Purves.) es la región entre los cuerpos de las células nerviosas donde se produce la mayor parte de la conectividad sináptica. Aunque la disposición de los circuitos neurales varía mucho según la función que cumplen, algunos elementos son característicos de estos conjuntos. Es importante la dirección del flujo de información en todo circuito particular, lo que, como es evi- dente, es esencial para conocer su propósito. Las células nerviosas que transportan información hacia el encéfalo o la médula espinal (o de modo más central, dentro de la médula espinal o el encéfalo) se denominan neuronas aferentes; las células nerviosas que transportan información lejos del encéfalo o la médula espinal (o lejos del circuito en cuestión) se denominan neuronas eferentes. Las interneuronas (o las neuronas de circuito local; véase luego) sólo participan en los aspectos locales de un circuito, sobre la base de las distancias cortas en las cuales se extienden sus axones. Estas tres clases funcionales –neuronas aferentes, neuronas eferentes e interneuronas– son los componentes básicos de todos los circuitos neurales. Un ejemplo simple de un circuito neural es un conjunto de células que corresponden al reflejo espinal miotático (el reflejo “patelar”; Fig. 1.7). Las neuronas aferentes del reflejo son neuronas sensitivas cuyos cuerpos celulares se ubican en los ganglios de las raíces dorsales y cuyos axones periféricos finalizan en terminaciones sensitivas en los músculos esqueléticos (los ganglios que cumplen esta misma función en gran parte de la cabeza y el cuello se denominan ganglios de los nervios craneales; véase el Apéndice). Los axones centrales de estas neuronas sensitivas aferentes entran en la médula espinal, donde terminan sobre distintas neuronas centrales vinculadas con la regulación del tono muscular, sobre todo las neuronas motoras, que determinan la actividad de los músculos relacionados. Éstas son las neuronas eferentes. Un grupo de estas neuronas eferentes en el asta ventral de la médula espinal se proyecta hacia los músculos flexores de la extremidad y el otro hacia los músculos extensores. Las interneuronas de la médula espinal son el tercer elemento de este circuito. Las interneuronas reciben contactos 12 CAPÍTULO 1 Axón sensitivo (aferente) Receptor sensitivo del músculo Corte transversal de la médula espinal 3A Músculo extensor 2B 2A 1 Músculo flexor 4 1 El golpe del martillo estira el tendón, lo que a su vez estira los receptores sensitivos en el músculo extensor de la pierna 3B Axones motores (eferentes) 2A La neurona sensitiva hace sinapsis con una neurona motora de la médula espinal y la excita 3A 2B La neurona sensitiva también excita la interneurona espinal 2C La sinapsis de la interneurona 2C Interneurona La neurona motora conduce el potencial de acción hasta las sinapsis sobre las fibras del músculo extensor y produce su contracción 4 La pierna se extiende 3B El músculo flexor se relaja inhibe la neurona motora de los músculos flexores FIGURA 1.7 Un circuito reflejo simple, la “respuesta patelar”. Técnicamente conocida como reflejo miotático, esta respuesta muestra varios puntos clave acerca de la organización funcional de los circuitos nerviosos. La estimulación de los sensores periféricos (en este caso, un receptor de estiramiento muscular) inicia los potenciales del receptor que disparan potenciales de acción que discurren centralmente a lo largo de los axones aferentes de las neuronas sensitivas. Esta información estimula las neuronas motoras espinales por medio de contactos sinápticos. Los potenciales de acción generados por el potencial sináptico en las neuronas motoras viajan de manera periférica en los axones eferentes y dan origen a la contracción muscular y una respuesta conductual. Uno de los propósitos de este reflejo particular es ayudar a mantener una posición erecta frente a los cambios inesperados (como al tropezar). sinápticos de neuronas aferentes sensitivas y hacen sinapsis sobre las neuronas motoras eferentes que se proyectan hacia los músculos flexores; por lo tanto, son capaces de modular la conexión aferencia-eferencia. Las conexiones sinápticas excitadoras entre los aferentes sensitivos y las neuronas motoras eferentes extensoras producen la contracción de los músculos extensores; a la vez, las interneuronas activadas por los aferentes son inhibidoras y su activación disminuye la acción eléctrica en las neuronas motoras eferentes flexoras y permite que los músculos flexores se vuelvan menos activos. El resultado es una activación y una inactivación complementaria de los músculos sinergistas y antagonistas que controlan la posición de la pierna. debido a que la actividad de sus neuronas motoras se inhibió Un cuadro más detallado de los acontecimientos que subyacen al circuito miotático o a cualquier otro puede obtenerse mediante el registro electrofisiológico, que mide la actividad eléctrica de una célula nerviosa. Hay dos enfoques básicos de este método: el registro extracelular, donde se coloca un electrodo cerca de la célula nerviosa de interés para detectar su actividad, y el registro intracelular, en el que se coloca un electrodo dentro de la célula. El registro extracelular es en particular útil para detectar patrones temporales de actividad de potenciales de acción y relacionar estos patrones con la estimulación por otras aferencias o con episodios conductuales específicos. Los registros intracelulares pueden detectar los cambios graduados más pequeños del potencial que disparan potenciales de acción y permiten así un análisis más detallado en la comunicación entre las neuronas dentro de un circuito. Estos potenciales que se disparan de forma graduada pueden surgir en receptores sensitivos o sinapsis, y se denominan potenciales de receptor o potenciales sinápticos, respectivamente. Para el circuito miotático, es posible medir la actividad eléctrica fuera de la célula y dentro de ella y definir así las relaciones funcionales entre las neuronas del circuito. Con los electrodos colocados cerca de las células individuales pero aún fuera de ellas, el patrón de actividad de potenciales de acción puede medirse para cada elemento del circuito (aferentes, eferentes e interneuronas) antes, durante y después de un estímulo (Fig. 1.8). Si se comparan el inicio, la duración y la frecuencia de la actividad de los potenciales de acción en cada célula, surge ESTUDIO DEL SISTEMA NERVIOSO un cuadro funcional del circuito. Por medio de un registro intracelular, es posible observar directamente los cambios de potencial subyacentes a las conexiones sinápticas del circuito del reflejo miotático (Fig. 1.9). Golpe de martillo Axón sensitivo (aferente) Neurona sensitiva Neurona motora (extensora) La organización del sistema nervioso humano Interneurona Axones motores (eferentes) Interneurona Neurona motora (flexora) La pierna se extiende FIGURA 1.8 El registro extracelular muestra la frecuencia relativa de potenciales de acción en el reflejo miotático. Los potenciales de acción están indicados por las líneas verticales individuales. Como resultado del estímulo, la neurona sensitiva es inducida a disparar con mayor frecuencia (es decir, más potenciales de acción por unidad de tiempo). Este incremento desencadena mayor frecuencia de potenciales de acción tanto en las neuronas motoras extensoras como en las interneuronas. Simultáneamente, las sinapsis inhibidoras que forman las interneuronas en las neuronas motoras flexoras disminuyen la frecuencia de los potenciales de acción en estas células (efecto modulador). Neurona sensitiva Registro Interneurona Potencial de membrana (mV) Registro (A) Neurona sensitiva Microelectrodo para medir el potencial de membrana Potencial de acción FIGURA 1.9 Respuestas intracelulares registradas que subyacen al reflejo miotático. (A) Potencial de acción medido en una neurona sensitiva. (B) Potencial disparador postsináptico registrado en una neurona motora extensora. (C) Potencial disparador postsináptico en una interneurona. (D) Potencial inhibidor postsináptico en una neurona motora flexora. Estos registros intracelulares son la base para conocer los mecanismos celulares de generación de los potenciales de acción y los potenciales de receptor sensitivo y sinápticos que desencadenan estas señales conducidas. (B) Neurona motora (extensora) Potencial Potencial sináptico de acción Sinapsis excitadora activa (C) Interneurona Potencial de membrana (mV) Neurona motora (flexora) Potencial de acción Potencial sináptico Sinapsis excitadora activa (D) Neurona motora (flexora) Potencial de membrana (mV) Registro Potencial de membrana (mV) Registro Neurona motora (extensora) 13 Sinapsis inhibitoria activa Tiempo (ms) Los circuitos que procesan tipos similares de información comprenden sistemas neurales que desempeñan propósitos conductuales más amplios. La distinción funcional más general divide estas colecciones en sistemas sensitivos que adquieren y procesan información del entorno (p. ej., el sistema visual o el auditivo; véase Unidad II) y sistemas motores que responden a esta información generando movimientos y otras conductas (véase Unidad III). Sin embargo, hay gran cantidad de células y circuitos que se ubican entre estos sistemas aferente y eferente relativamente bien definidos. En conjunto, se denominan sistemas de asociación y median las funciones encefálicas más complejas y menos caracterizadas (véase Unidad V). Además de estas distinciones funcionales amplias, los neurocientíficos y los neurólogos hicieron una división convencional del sistema nervioso de los vertebrados desde el punto de vista anatómico en los componentes central y periférico (Fig. 1.10). El sistema nervioso central (SNC) comprende el encéfalo (hemisferios cerebrales, diencéfalo, cerebelo y tronco del encéfalo) y la médula espinal (para mayor información sobre las características anatómicas macroscópicas del SNC, véase Apéndice A). El sistema nervioso periférico (SNP) incluye las neuronas sensitivas que conectan los receptores sensitivos sobre la superficie del cuerpo o más profundo dentro de él con circuitos de procesamiento relevantes en el sistema nervioso central. A su vez, la porción motora del sistema nervioso periférico presenta dos componentes. Los axones motores que conectan el encéfalo y la médula espinal con los músculos esqueléticos constituyen la división motora somática del sistema nervioso periférico, mientras que las células y los axones que inervan los CAPÍTULO 1 (A) Sistema nervioso central Sistema nervioso periférico Encéfalo Nervios craneales Médula espinal Nervios espinales (B) Hemisferios cerebrales, diencéfalo, cerebelo, tronco del encéfalo y médula espinal (análisis e integración de la información sensitivomotora) Componentes sensitivos Receptores sensitivos (en la superficie y el interior del cuerpo) Medioambiente interno y externo músculos lisos, el músculo cardíaco y las glándulas forman la división motora visceral o autónoma. Los cuerpos de las células nerviosas en el sistema nervioso periférico se localizan en los ganglios, que son simplemente acumulaciones de cuerpos de células nerviosas y células de sostén. Los axones periféricos se reúnen en haces llamados nervios, muchos de los cuales están envueltos por las células gliales del sistema nervioso periférico, como mencionamos antes, y éstas se denominan células de Schwann. En el sistema nervioso central, las células nerviosas están organizadas de dos formas distintas. Los núcleos son acumulaciones locales de neuronas que tienen conexiones y funciones más o menos similares; estos grupos se encuentran distribuidos en el cerebro, el tronco del encéfalo y la médula espinal. Por el contrario, la corteza describe disposiciones laminares de células nerviosas (para obtener información adicional e ilustraciones consúltese Apéndice A). Las cortezas de los hemisferios cerebrales y el cerebelo proporcionan el ejemplo más claro de este principio de organización. Los axones del sistema nervioso central se reúnen en tractos que son más o menos análogos a los nervios de la periferia. Los tractos que cruzan la línea media del encéfalo se denominan comisuras. Dos términos histológicos amplios distinguen las regiones ricas en cuerpos de células nerviosas versus las regiones ricas en axones. La sustancia gris se refiere a cualquier acumulación de cuerpos celulares y neurópilo del encéfalo y la médula espinal (p. ej., núcleos o cortezas), mientras que la sustancia blanca, llamada así por su aspecto relativamente claro como re- Componentes motores Sistema motor visceral (divisiones simpática, parasimpática y entérica) Sistema motor somático Nervios motores Sistema nervioso periférico Ganglios y nervios sensitivos Sistema nervioso central 14 Ganglios y nervios autónomos Efectores Músculos lisos, músculos cardíacos y glándulas Músculos esqueléticos (estriados) FIGURA 1.10 Principales componentes del sistema nervioso y sus relaciones funcionales. (A) SNC (encéfalo y médula espinal) y SNP (nervios espinales y craneales). (B) Diagrama de los componentes principales de los sistemas nerviosos central y periférico y sus relaciones funcionales. Los estímulos provenientes del medioambiente transmiten información a los circuitos procesadores en el interior del encéfalo y la médula espinal, que a su vez interpretan su significación y envían señales a los efectores periféricos que mueven el cuerpo y adaptan los funcionamientos de sus órganos internos. sultado del contenido de lípidos de la mielina, se refiere a los tractos axónicos y las comisuras. La organización de la división motora visceral del sistema nervioso periférico (las células nerviosas que controlan las funciones de los órganos viscerales, incluidos el corazón, los pulmones, el tubo digestivo y los genitales) es un poco más complicada (véase Cap. 21). Las neuronas motoras viscerales del tronco del encéfalo y la médula espinal, denominadas neuronas preganglionares, forman sinapsis con neuronas motoras periféricas situadas en los ganglios autónomos. Las neuronas motoras periféricas de los ganglios autónomos inervan el músculo liso, las glándulas y el músculo cardíaco, y controlan así la mayoría de las conductas involuntarias (visceral). En la división simpática del sistema motor autónomo, los ganglios se sitúan a lo largo de la columna vertebral o por delante de ella y envían sus axones a distintos sitios diana periféricos. En la división parasimpática, los ganglios se sitúan dentro de los órganos que inervan o adyacentes a ellos. Otro componente del sistema motor visceral, llamado sistema entérico, está forma- ESTUDIO DEL SISTEMA NERVIOSO do por ganglios pequeños y neuronas individuales dispersas por toda la pared del intestino. Éstas influyen en la motilidad y la secreción gástricas. En el Apéndice de este libro pueden hallarse detalles de las estructuras físicas y la anatomía general del sistema nervioso humano, y hay también un pequeño atlas del sistema nervioso central a continuación del Apéndice. Sistemas neurales Varias características distinguen los sistemas neurales dentro del conjunto complejo de componentes anatómicos que conforman cualquier sistema nervioso; las principales son la unidad funcional, la representación de información específica y la subdivisión en subsistemas para la transmisión y el procesamiento de la información en paralelo. La más importante es la unidad funcional que es evidente en un conjunto interconectado de neuronas. Por ejemplo, el sistema visual es definido por todas las neuronas y conexiones dedicadas a la visión, el sistema auditivo por aquellas dedicadas a la audición, el sistema somatosensitivo por las dedicadas al sentido del tacto, el sistema motor piramidal por las neuronas y conexiones destinadas al movimiento voluntario, y así para muchos otros sistemas identificables. En muchos casos, los componentes de un sistema están distribuidos por todo el cuerpo y el encéfalo. Así, los sistemas sensitivos incluyen las especializaciones sensitivas periféricas del ojo, el oído, la piel, la nariz y la lengua, mientras que los sistemas motores incluyen los nervios motores periféricos y los músculos diana necesarios para realizar distintas acciones. Tanto los sistemas sensitivo como motor poseen vías nerviosas que conectan la periferia con los núcleos en la médula espinal, el tronco del encéfalo, el tálamo y las áreas relevantes de la corteza cerebral. Otras dos características de muchos sistemas neurales son la representación ordenada de la información a distintos niveles y una división de la función del sistema en submodalidades o cualidades sensitivas que típicamente son transportadas y procesadas en vías paralelas, donde se procesa la información de cada una de las submodalidades (p. ej., la frecuencia y el volumen de la señal auditiva). Para los sistemas como la visión y el tacto –los sistemas que funcionan para distinguir las diferencias entre puntos vecinos del campo visual o de la superficie corporal–, la representación de la información es topográfica. Estas representaciones o mapas topográficos reflejan una correspondencia punto a punto entre la periferia sensitiva (el campo visual y la superficie corporal) y las neuronas vecinas en el interior de los componentes centrales del sistema (en la médula espinal y el encéfalo). En los sistemas neurales donde la representación de la información no implica discriminar los puntos vecinos en un campo, como el olfato y el gusto, los mapas computacionales comparan, evalúan e integran múltiples atributos del estímulo en una representación ordenada que facilita la extracción y el procesamiento de la información esencial (p. ej., el número y la configuración de las moléculas odoríferas para determinar el origen y la naturaleza de un olor). Los sistemas motores comprenden representaciones ordenadas de 15 movimientos, aunque aquí la dirección del flujo de información es desde el sistema nervioso central hasta la periferia. Las vías paralelas surgen porque casi todos los sistemas neurales tienen subsistemas identificables. Por ejemplo, el sistema visual humano tiene subsistemas que destacan las características del estímulo como color, forma o movimiento, y, en cierta medida, cada clase de información se transmite y procesa de forma separada. La segregación similar de los subtipos de información en vías paralelas es aparente en otros sistemas sensitivos y motores. Por último, es importante destacar que no todos los sistemas neurales están tan bien organizados como los sistemas sensitivo y motor primarios. Los sistemas que controlan el lenguaje, la atención, la memoria y la emoción se reconocen por su unidad funcional y su localización anatómica, pero no se conoce tan bien de qué modo se representa la información, sus subsistemas ni sus aferencias y eferencias (véase Unidad V). Análisis estructural de los sistemas neurales Los primeros investigadores de los sistemas neurales realizaron inferencias sobre la localización funcional (es decir, qué región del sistema nervioso controla cada función) al correlacionar el daño macroscópico observado post mórtem de las estructuras encefálicas con los déficits funcionales registrados durante la vida del individuo. Este uso de la estructura para inferir la función fue adoptado para la experimentación, y gran parte de la neurociencia descansa en observaciones que se realizaron al dañar intencionalmente una región encefálica, un nervio o un tracto definido en un animal de experimentación, y observar y documentar la pérdida posterior de función en regiones únicas o interconectadas. Estos estudios de lesiones proporcionan la base de gran parte del conocimiento actual de la anatomía de los sistemas neurales. El punto de vista moderno más detallado de la neuroanatomía que define a los sistemas neurales surgió solamente después del descubrimiento de técnicas para rastrear las conexiones neurales desde su origen hasta su terminación (anterógrado) o viceversa, desde su terminación hasta su origen (retrógrado). Estos enfoques permiten la evaluación detallada de las conexiones entre distintas regiones del sistema nervioso y facilitan así el “mapeo” de las conexiones entre las neuronas en una estructura (por ejemplo, el ojo) y sus sitios diana en el encéfalo. En un inicio, estas técnicas se basaban en inyectar moléculas visualizables en el encéfalo que eran captadas por los cuerpos celulares locales y transportadas hasta las terminaciones axónicas, o captadas por los axones y las terminaciones locales y transportadas hacia atrás hasta el cuerpo de la célula madre (Figura 1.11A, B). Otros marcadores pueden demostrar una red completa de proyecciones axónicas que parten de las células nerviosas expuestas al marcador (Fig. 1.11C). En conjunto, estos enfoques permiten evaluar la extensión de las conexiones de una única población de células nerviosas hacia sus sitios diana en todo el sistema nervioso. 16 (A) CAPÍTULO 1 (B) FIGURA 1.11 Enfoques celular y molecular para estudiar la conectividad y la identidad molecular de las células nerviosas. (A-C) Rastreo de conexiones y vías en el encéfalo. (B) Los aminoácidos radiactivos pueden ser captados por una población de células nerviosas (en este caso, la inyección en un ojo de un aminoácido radiactivo) y transportados hasta las terminaciones axónicas de esas células en la región encefálica diana. (B) Las moléculas fluo- (C) (D) rescentes inyectadas en el tejido nervioso son captadas por las terminaciones axónicas en el sitio de la inyección. Después, las moléculas son transportadas y se marcan los cuerpos celulares y las dendritas de las células nerviosas que proyectan hacia el sitio de la inyección. (C) Los marcadores de los axones pueden poner en evidencia vías complejas en el sistema nervioso. En este caso, se ha inyectado un ganglio de la raíz dorsal, que muestra la variedad de vías axónicas desde el ganglio hasta la médula espinal. (D-G) Diferencias moleculares entre las células nerviosas. (D) Se ha marcado un único glomérulo del bulbo olfatorio (véase Figura 1-6H) con un anticuerpo contra el neurotransmisor inhibidor GABA. El marcador muestra una tinción roja, que pone en evidencia que el GABA se localiza en subgrupos de neuronas alrededor del glomérulo y en terminaciones sinápticas en el neurópilo del glomérulo. (E) El cerebelo ha sido marcado con una sonda (azul) para un gen específico que es expresado sólo (E) (F) por las células de Purkinje. (A, cortesía de P. Rakic; B, cortesía de B. Schofield; C, cortesía de W. D. Snider y J. Lichtman; D-F, cortesía de A.-S. LaMantia, D. Meechan y T. Maynard.) Los análisis de conectividad han sido ampliados mediante técnicas histoquímicas de base molecular que demuestran distinciones bioquímicas y genéticas entre las células nerviosas y sus prolongaciones. Mientras los métodos generales de tinción celular muestran principalmente diferencias en el tamaño y la distribución de las células, las tinciones con anticuerpos pueden reconocer proteínas específicas en distintas regiones de una célula nerviosa o diferencias moleculares en clases de células nerviosas. Estos enfoques han aclarado la distribución funcional de sinapsis, dendritas y otras características moleculares definidas de los tipos de células nerviosas en distintas regiones encefálicas (Figura 1.11D, E). Además, se han utilizado con fines similares anticuerpos contra distintas proteínas y sondas para transcritos de mRNA específicos (que detectan la expresión genética en células relevantes) (Fig. 1.11F). Más recientemente aún, se han combinado métodos de genética molecular y neuroanatómicos para visualizar la expresión de moléculas fluorescentes o de otras moléculas marcadoras bajo el control de secuencias reguladoras de los genes neurales (Fig. 1.12). Este enfoque ilumina con un detalle notable células individuales en el tejido fijado o vivo, lo que permite identificar las células nerviosas o sus prolongaciones por su estado transcripcional (es decir, por lo genes que se transcriben en esa célula) así como por su estructura y conexiones. Por lo tanto, en la actualidad distintos enfoques moleculares y de ingeniería genética posibilitan a los investigadores rastrear las conexiones entre poblaciones definidas de neuronas y sus sitios diana. Actualmente, se utilizan de forma sistemática varios enfoques (rastreo de vías, análisis de la identidad molecular de las células nerviosas y enfoques genéticos para identificar identidades celulares y conexiones) en el estudio continuo del modo en que las neuronas y la glía se organizan en circuitos neurales funcionales y sistemas neurales. Análisis funcional de los sistemas neurales Los análisis funcionales proporcionan otra forma poderosa de conocer la organización del sistema nervioso. En la actualidad, existen diversos métodos fisiológicos para evaluar la actividad eléctrica y metabólica de los circuitos neurales que conforman un sistema neural. Dos enfoques, el registro electrofisiológico y las imágenes encefálicas funcionales, han sido particularmente útiles para definir de qué modo los sistemas neurales representan la información. El método electrofisiológico más utilizado ha sido el registro electrofisiológico de una célula única o de una unidad única con microelectrodos, como explicamos antes (véanse Figs. 1.8 y 1.9). Muchas veces, este método registra varias células cercanas además de la seleccionada, y proporciona más información útil. El uso de microelectrodos para registrar la actividad de los potenciales de acción permite el análisis intercelular de la organización de mapas topográficos y puede dar ideas específicas sobre el tipo de estímulo al que está “afinada” la neurona (es decir, ESTUDIO DEL SISTEMA NERVIOSO Promotor FIGURA 1-12 Ingeniería genética utilizada Gen reportero Axón central 17 Segmento de médula espinal para mostrar vías dentro del sistema nervioso. Un “gen reportero” que codifica para alguna sustancia visualizable (p. ej., proteína fluorescente verde, GFP) es insertado en el genoma bajo el control de un promotor específico del tipo celular (una secuencia de DNA que “enciende” el gen en tipos específicos de tejidos y células). El reportero es expresado solamente en aquellos tipos celulares, y pone en evidencia los cuerpos celulares, los axones y las dendritas de todas las células en el sistema Cuerpo celular Terminaciones sensitivas en la piel Axón periférico Ganglio de la raíz dorsal Terminaciones axónicas en la médula espinal nervioso que expresan el gen. Aquí, el gen reportero se encuentra bajo el control de una secuencia promotora de DNA que es activada solamente en un subgrupo de neuronas del ganglio de la raíz dorsal. Las fotografías muestran que el reportero marca los cuerpos de las células neuronales; los axones que proyectan hacia la piel como terminaciones nerviosas libres y el axón que proyecta hacia la raíz dorsal de la médula espinal para transmitir información sensitiva desde la piel hacia el encéfalo. (Fotografías de Zylka y cols., 2005). el estímulo que produce un cambio máximo en la actividad del potencial de acción desde el estado basal). Esta afinación define el campo receptivo de una neurona, o sea la región del espacio sensitivo (p. ej., la superficie corporal o una estructura especializada como la retina) dentro de la cual un estímulo específico produce la respuesta del potencial de acción (Fig. 1.13). Stephen Kuffler y Vernon Mountcastle fueron pioneros en esta forma de conocimiento de los sistemas neurales a comienzos de la década de 1950, y desde entonces ha sido utilizada por varias generaciones de neurocientíficos para evaluar la relación entre los estímulos y las respuestas neuronales en los sistemas sensitivo y motor. Las técnicas de registro eléctrico a nivel de células aisladas se han extendido y refinado para incluir el análisis de células individuales y el análisis simultáneo de múltiples células en animales que realizan tareas cognitivas complejas, los registros intra- Actividad de la neurona cortical (A) Corteza somatosensitiva (B) Campo receptivo (centro) El tacto en el centro del campo receptivo aumenta la descarga celular El tacto en la periferia del campo receptivo disminuye la descarga celular Registro Surco central Circunvolución poscentral Campo receptivo (periferia) El tacto fuera del campo receptivo no tiene efecto alguno Período de estimulación FIGURA 1.13 Registro electrofisiológico de unidad única en un mono. Este ejemplo es de una neurona piramidal cortical, y muestra el patrón de descarga en respuesta a un estímulo periférico específico. (A) Disposición experimental típica, en la cual se inserta un electrodo de registro en el encéfalo. (B) Definición de los campos receptivos neuronales. 18 CAPÍTULO 1 celulares en animales intactos y el uso de electrodos en parches para detectar y controlar la actividad de moléculas de membrana individuales que subyacen finalmente a la señalización neural (véanse Cap. 2 y 3). La segunda área en la que se han realizado adelantos técnicos notables es en el análisis de imágenes encefálicas funcionales (Recuadro 1B). En las últimas tres décadas, el uso de técnicas de imágenes encefálicas funcionales en seres humanos (y, en menor grado, en animales de experimentación) ha revolucionado nues- tro conocimiento de los sistemas neurales y su capacidad para describir y diagnosticar anomalías funcionales. Al contrario de los métodos electrofisiológicos para registrar la actividad neural, las imágenes funcionales se obtienen por métodos no invasivos y, por lo tanto, pueden aplicarse a pacientes clínicos y a individuos sanos. Si bien las imágenes funcionales no pueden resolver la actividad eléctrica de las neuronas individuales, registran la actividad metabólica local dentro de pequeños volúmenes del tejido ence- RECUADRO 1B Técnicas por la imagen encefálicas Hasta fines del último siglo, gran parte de nuestro conocimiento de la estructura y la función del encéfalo humano sano o enfermo se discernía a partir del estudio de muestras humanas post mórtem o se infería por estudios en animales. Aunque estos enfoques eran informativos, existía la preocupación de que esta información no transmitiera la capacidad funcional completa del encéfalo humano, sobre todo al realizar tareas complejas como la comunicación utilizando el lenguaje. La aparición de varias técnicas nuevas para obtener imágenes de los detalles anatómicos y funcionales del encéfalo humano confirmó muchas de las conclusiones extraídas de los estudios post mórtem y en animales, pero también ha abierto nuevas vías para conocer la forma en la cual funciona el encéfalo en los sujetos sanos y en pacientes clínicos. Tomografía computarizada (TC) En la década de 1970, la tomografía computarizada o TC abrió una nueva era en las imágenes no invasoras al introducir Fuente de rayos X Detector de rayos X el uso de la tecnología de procesamiento computarizado para ayudar a sondear el encéfalo viviente. Antes de la TC, la única técnica por imágenes encefálicas disponible era la radiografía convencional, que tenía escaso contraste de tejidos blandos y comprendía una exposición a la radiación relativamente alta. La técnica de la TC utiliza un haz de rayos X estrecho y una hilera de detectores muy sensibles colocados en lados opuestos de la cabeza para sondear sólo una pequeña porción de tejido a la vez con exposición limitada a la radiación (Fig. A). Para formar una imagen, el tubo de rayos X y los detectores rotan alrededor de la cabeza para recoger información de radiodensidad de todas las orientaciones que rodean un corte estrecho. Las técnicas de procesamiento computarizado calculan luego la radiodensidad de cada punto dentro del plano de corte y producen una imagen tomográfica (en griego, tomo significa “corte” o “trozo”). Si el paciente es movilizado lentamente a través del tomó- grafo mientras el tubo de rayos X rota de esta forma, es posible crear una matriz de radiodensidad tridimensional que permite computarizar las imágenes para cualquier plano a través del encéfalo. Las TC posibilitan distinguir con facilidad la sustancia blanca de la gris, diferenciar muy bien los ventrículos y mostrar muchas otras estructuras encefálicas con una resolución espacial de varios milímetros. Resonancia magnética (RM) Las imágenes encefálicas dieron otro gran paso hacia adelante en la década de 1980 con el desarrollo de la resonancia magnética (RM). La RM se basa en que los núcleos de algunos átomos actúan como imanes que giran y que, si son colocados en un campo magnético fuerte, cubrirán el campo y girarán a una frecuencia que depende de la fuerza del campo. Si ellos reciben entonces un pulso breve de radiofrecuencia ajustado a su frecuencia de giro, se alejan de su alineación con el campo y luego emiten energía en forma oscilatoria a medida que se realinean gradualmente con el campo. La fuerza de la señal emitida depende de la cantidad de núcleos que participan en este proceso. En la RM, el campo magnético se distorsiona ligeramente imponiendo gradientes magnéticos a lo largo de tres ejes espaciales diferentes de modo que sólo los núcleos en ciertas localizaciones están ajustados a la frecuencia del detector en cualquier momento dado. Casi todos los resonadores utilizan detectores ajusta- (A) En la tomografía computarizada, la fuente de rayos X y los detectores se mueven alrededor de la cabeza del paciente. El recuadro muestra un corte horizontal de TC del encéfalo de un adulto normal. ESTUDIO DEL SISTEMA NERVIOSO 19 RECUADRO 1B (continúa) (B) En la RM, la cabeza se coloca en el centro de un gran imán. Una antena espiral de radiofrecuencia se coloca alrededor de la cabeza para excitar y registrar la señal de resonancia magnética. En la RMf, los estímulos pueden presentarse utilizando gafas de video de realidad virtual y audífonos estereofónicos mientras el paciente está en el interior del resonador. Imágenes encefálicas funcionales dos a las radiofrecuencias de los núcleos de hidrógeno giratorios en moléculas de agua, y crean así imágenes basadas en la distribución del agua en diferentes tejidos. Una manipulación cuidadosa de los gradientes de campo magnético y los pulsos de radiofrecuencia hace posible construir imágenes extraordinariamente detalladas del encéfalo en cualquier localización y orientación, con una resolución submilimétrica (Fig. B). La seguridad y la versatilidad han convertido a la RM en la técnica de elección para obtener imágenes de la estructura encefálica en la mayoría de las aplicaciones. El campo magnético fuerte y los pulsos de radiofrecuencia utilizados en la RM son inocuos, lo que convierte a esta técnica en completamente no invasiva (aunque los objetos metálicos que se encuentran dentro del resonador o cerca de él constituyen una preocupación para la seguridad). La RM también es en extremo versátil porque, al modificar los parámetros del resonador, se pueden generar imágenes basadas en una amplia variedad de diferentes mecanismos de contraste. Por ejemplo, las imágenes convencionales de RM aprovechan que el hidrógeno en distintos tipos de tejido (p. ej., sustancia gris, sustancia blanca y líquido cefalorraquídeo) tiene velocidades de realineación ligeramente distintos, lo que indica que el contraste de los tejidos blandos puede manipularse simplemente ajustando el momento en que se mide la señal de hidrógeno que se realinea. También pueden utilizarse diferentes regulaciones de los parámetros para generar imágenes en las cuales las sustancias gris y blanca son invisibles, pero en las que sobresale la vascularización encefálica con gran detalle. La seguridad y la versatilidad convirtieron la RM en la técnica de elección para obtener imágenes de la estructura encefálica en la mayoría de las aplicaciones. La alineación de los campos magnéticos de las moléculas de agua en los tractos axónicos hizo posible visualizar las vías axónicas utilizando una variante de RM denominada imágenes de tensor de difusión, que se observa en la imagen que abre este capítulo. Estas imágenes pueden establecer diferencias en la conectividad de la vía axónica, lo que permite estudiar a los pacientes con trastornos genéticos que conducen a alteraciones mayores de las proyecciones axónicas (véase Recuadro 23B). Otros ajustes de los parámetros pueden generar imágenes en las cuales la sustancia gris y blanca son relativamente invisibles pero la vascularización encefálica destaca con mucho detalle. También se hizo posible la obtención de imágenes de las variaciones funcionales en el encéfalo viviente con el desarrollo reciente de técnicas para detectar pequeños cambios localizados en el metabolismo o en el flujo sanguíneo cerebral. Para conservar energía, el encéfalo regula su flujo sanguíneo de modo que las neuronas activas con demandas metabólicas relativamente altas reciben más sangre que neuronas relativamente inactivas. La detección y el trazado de estos cambios locales en el flujo sanguíneo cerebral forma la base para tres técnicas de imágenes encefálicas funcionales muy utilizadas: la tomografía por emisión de positrones (PET), la tomografía computarizada por emisión de fotón único (SPECT) y la resonancia magnética funcional (RMf). En la PET, se incorporan isótopos emisores de positrones inestables en diferentes reactivos (incluidos agua, moléculas precursoras de neurotransmisores específicos o glucosa) y se los inyecta en el torrente sanguíneo. El oxígeno y la glucosa marcados se acumulan con rapidez en las áreas con mayor actividad metabólica y las sondas de transmisores marcados se captan selectivamente en regiones apropiadas. A medida que el isótopo inestable se desintegra, conduce a la emisión de dos positrones que se mueven en direcciones opuestas. Los detectores de rayos gamma colocados alrededor de la cabeza registran un “golpe” sólo cuando dos detectores separados por 180º reaccionan en forma simultánea. Pueden generarse entonces imágenes de la densidad del isótopo en los tejidos (en forma muy similar a la manera en que se calculan las imágenes por TC) que muestran la localización de las regio- (continúa en la página siguiente) 20 CAPÍTULO 1 RECUADRO 1B (continúa) Derecha Izquierda Tumor (C) Imágenes por RM de un paciente adulto con un tumor encefálico, con actividad de RMf durante una labor de movimiento de las manos superpuesta (la actividad de la mano izquierda se muestra en amarillo; de la mano derecha, en verde). A la derecha, hay una vista reconstruida de la superficie tridimensional de los mismos datos. nes activas con una resolución espacial de unos 4 mm. Según la sonda inyectada, las imágenes de PET pueden utilizarse para visualizar cambios dependientes de la actividad en flujo sanguíneo, metabolismo tisular o actividad bioquímica. Las imágenes de SPECT son similares a las de PET, ya que comprenden la inyección o la inhalación de un compuesto radiomarcado (p. ej., 133Xe o yodoanfetamina marcada con 123I), que producen protones detectados por una cámara gamma que se mueve rápidamente alrededor la cabeza. En la actualidad la RM funcional, variante de la RM, ofrece el mejor enfoque para visualizar la función encefálica basada en el metabolismo local. La RMf se basa en que la hemoglobina en la sangre distorsiona ligeramente las propiedades de resonancia magnética de los núcleos de hidrógeno en su vecindad y el grado de distorsión magnética cambia en función de que la hemoglobina tenga oxígeno ligado a ella. Cuando un área encefálica es activada por una tarea especial, comienza a utilizar más oxígeno y en pocos segundos la microvasculatura encefálica responde aumentando el flujo de sangre rica en oxígeno en el área activa. Estos cambios en la concentración de oxígeno y el flujo sanguíneo conducen a alteraciones locali- zadas dependientes del nivel de la oxigenación sanguínea en la señal de resonancia magnética. Estas fluctuaciones se detectan por medio de técnicas estadísticas de procesamiento de imágenes para producir mapas de función específica dependientes de la tarea (Fig. C). Como la RMf utiliza las señales intrínsecas al encéfalo sin radiactividad alguna, es posible realizar observaciones repetidas en el mismo individuo, una ventaja importante sobre los métodos por imágenes como PET. La resolución espacial (2 a 3 mm) y la resolución temporal (algunos segundos) de la RMf también son superiores a la PET y la SPECT, y pueden combinarse fácilmente con imágenes estructurales generadas por RM. Por lo tanto, la RMf surgió como la tecnología de elección para sondear la estructura y la función del encéfalo humano viviente. La magnetoencefalografía (MEG) proporciona una forma de localizar la función encefálica con mejor resolución temporal. Así, a veces la MEG se utiliza para trazar cambios en la función encefálica con una resolución de milisegundos en lugar de la resolución temporal inferior de segundos que aporta la RMf. La MEG, como su nombre sugiere, registra las consecuencias magnéticas de la actividad eléctrica encefálica más que las señales eléctricas propiamente dichas. Así, al contrario de su pariente cercano la electroencefalografía (EEG), la MEG detecta fuentes independientes de flujo de corriente sin referencia a otras corrientes. Las señales magnéticas que detectan los registros de MEG son muy locales, y existe una resolución espacial relativamente buena (0,1-1 cm) de la señal original; es posible detectar así la actividad eléctrica dinámica en el encéfalo con resolución temporal que se aproxima a los eventos clave en la señalización eléctrica neuronal –es decir, potenciales de acción y potenciales sinápticos– y registran la localización del origen de esa actividad. Para obtener un registro de MEG, se coloca un conjunto de dispositivos detectores individuales denominados SQUID (“superconducting quantum interference device”, dispositivo de interferencia de cuántos superconductores) como un casco que se ajusta al sujeto. A continuación, el individuo es colocado en un biomagnetómetro que amplifica las señales detectadas por el conjunto SQUID. Estas señales, al igual que las del EEG, proporcionan un mapa de las fuentes de corrientes a través del encéfalo utilizando puntos de referencia (habitualmente los oídos y la nariz) para crear un espacio tridimensio- ESTUDIO DEL SISTEMA NERVIOSO 21 RECUADRO 1B (continúa) nal. Como estos mapas carecen de detalle anatómico, a menudo la MEG se combina con una RM, combinación denominada imágenes de fuente magnética. El uso de los métodos de imágenes estructurales y funcionales modernos ha revolucionado la neurociencia humana. En la actualidad, es posible obtener imágenes del encéfalo viviente en acción y evaluar los cambios en la actividad encefálica tanto en individuos sanos como en pacientes con trastornos que varían desde el autismo hasta la enfermedad de Alzheimer. Bibliografía Huettel, S. A., A. W. Song and G. McCarthy (2009) Functional Magnetic Resonance Imaging, 2nd Ed. Sunderland, MA: Sinauer Associates. fálico. En consecuencia, este enfoque permite la evaluación simultánea de múltiples estructuras encefálicas (lo que es posible pero difícil con los métodos de registro eléctrico). Análisis de la conducta compleja Muchos de los adelantos más anunciados en la neurociencia moderna involucraron una reducción de la complejidad del encéfalo hasta los componentes que se analizan más fácilmente, esto es, genes, moléculas o células. No obstante, el estudio de las funciones encefálicas más complejas como percepción, lenguaje, emoción, memoria y conciencia aún es un desafío central para los neurocientíficos contemporáneos. Reconociendo este desafío, en los últimos 25 años más o menos surgió un campo llamado neurociencia cognitiva, dedicado específicamente a comprender estas cuestiones (véase Unidad V). Esta evolución también rejuveneció el campo de la neuroetología (dedicado a observar conductas complejas de animales en sus entornos naturales, p. ej., la comunicación social en las aves y los primates no humanos) y estimuló el desarrollo de tareas para evaluar mejor la génesis de las conductas complejas en los seres humanos. Cuando se usan combinadas con imágenes funcionales, las tareas conductuales bien diseñadas pueden facilitar la identificación de los sistemas encefálicos (o de redes si no existen pruebas suficientes de una función unitaria; véase antes) dedicados a funciones complejas específicas, entre las que se incluyen habilidades de lenguaje, capacidad matemática y musical, respuestas emocionales, juicios estéticos y pensamiento abstracto. También es posible utilizar tareas conductuales cuidadosamente construidas para estudiar la patología de las enfermedades encefálicas complejas que comprometen la cognición, como la enfermedad de Alzheimer, la esquizofrenia y la depresión. Estos esfuerzos nuevos o revitalizados utilizan técnicas cada vez más poderosas para estudiar las funciones encefálicas “superiores” de los animales y los seres humanos y ofrecen nuevas formas para comenzar a comprender los aspectos más complejos del comportamiento. Resumen El encéfalo puede estudiase mediante métodos que varían desde la genética y la biología molecular hasta pruebas conductua- Wheless, J. W. and 6 others (2004) Magnetoencephalography (MEG) and magnetic source imaging (MSI). Neurologist 10: 138-153. Oldendorf, W. and W. Oldendorf Jr. (1988) Basics of Magnetic Resonance Imaging. Boston: Kluwer Academic Publishers. Raichle, M. E. (1994) Images of the mind: Studies with modern imaging techniques. Annu. Rev, Psychol 45: 333-356. Schild, H. (1990) MRIMade Easy (...Well, Almost). Berlin: H. Heineman. les en sujetos normales. Además de un conjunto cada vez mayor de conocimientos acerca de la organización anatómica del sistema nervioso, muchos de los éxitos más brillantes de la neurociencia moderna han provenido del conocimiento de las células nerviosas como unidad estructural y funcional del sistema nervioso. Algunos estudios de la arquitectura celular y los componentes moleculares distintos de las neuronas y la glía pusieron en evidencia gran parte de sus funciones individuales a un nivel detallado y proporcionaron una base para saber de qué modo se organizan las células nerviosas en circuitos y éstos en sistemas que procesan tipos específicos de información pertinente al procesamiento sensorial y las respuestas conductuales. Los objetivos pendientes son comprender de qué modo los fenómenos genéticos moleculares básicos están ligados a las funciones celulares, de los circuitos y los sistemas, saber cómo fallan estos procesos en las enfermedades neurológicas y psiquiátricas y comenzar a comprender las funciones especialmente complejas del encéfalo que nos convierten en seres humanos. Lecturas adicionales Brodal, P. (2010) The Central Nervous System: Structure and Function, 4th Ed. New York: Oxford University Press. Gibson, G. and S. Muse (2009) A Primer of Genome Science, 3rd Ed. Sunderland, MA: Sinauer Associates. Nature Vol. 409, No. 6822 (2001) Issue of February 16. Special issue on the human genome. Peters, A., S. L. Palay and H. de F. Webster (1991) The Tine Structure of the Nervous System: Neurons and Their Supporting Cells, 3rd Ed. New York: Oxford University Press. Posner, M. I. and M. E. Raichle (1997) Images of Mind, 2nd Ed. New York: W. H. Freeman & Co. Ramon y Cajal, S. (1984) The Neuron and the Glial Cell. (Transl. by J. de la Torre and W. C. Gibson.) Springfield, IL: Charles C. Thomas. Ramón y Cajal, S. (1990) New Ideas on the Structure of the Nervous System in Man and Vertebrates. (Transl. by N. Swanson and L. W. Swanson.) Cambridge, MA: MIT Press. Ropper, A. and N. Samuels (2009) Adams and Victor‘s Principles of Neurology, 9th Ed. New York: McGraw-Hill Medical. Science Vol. 291, No. 5507 (2001) Issue of February 16. Special issue on the human genome. Schoonover, C. (2010) Portraits of the Mind: Visualizing the Brain from Antiquity to the 21st Century. New York: Abrams. Shepherd, G. M. (1991) Foundations of the Neuron Doctrine. History of Neuroscience Series, No. 6. Oxford: Oxford University Press. UNIDAD I Señalización neural EL ENCÉFALO TIENE LA EXTRAORDINARIA CAPACIDAD de adquirir, coordinar y diseminar la información CAPÍTULO 2 Señales eléctricas de las células nerviosas CAPÍTULO 3 Permeabilidad de la membrana dependiente de voltaje CAPÍTULO 4 Canales iónicos y transportadores CAPÍTULO 5 Transmisión sináptica CAPÍTULO 6 Neurotransmisores y sus receptores CAPÍTULO 7 Señalización molecular en el interior de las neuronas CAPÍTULO 8 Plasticidad sináptica acerca del cuerpo y su ambiente. Esta información debe procesarse en algunos milisegundos, aunque también puede ser almacenada en forma de recuerdos que perduran durante años. Las neuronas realizan estas funciones generando señales eléctricas y químicas sofisticadas. En esta unidad se describen estas señales y el modo en que se producen. Se explica cómo un tipo de señal eléctrica, el potencial de acción, permite que la información viaje a lo largo de una célula nerviosa. También se expone el modo en que otros tipos de señales (eléctricas y químicas) se generan en las conexiones sinápticas entre las células nerviosas. Las sinapsis permiten la transferencia de información mediante la interconexión de numerosas neuronas para formar el circuito del que depende el procesamiento neural. Por último, se describen los episodios intrincados de señalización bioquímica que tienen lugar en el interior de las neuronas y el modo en que esta señalización puede producir cambios dependientes de la actividad en la comunicación sináptica. Analizar estas formas fundamentales de señalización neuronal aporta las bases para la comprensión de las funciones de nivel superior consideradas en el resto del libro. Los mecanismos celulares y moleculares que brindan a las neuronas su capacidad única de señalización también constituyen elementos diana para los procesos patológicos que pueden afectar las funciones del sistema nervioso. Por lo tanto, es fundamental un conocimiento activo de la biología celular y molecular de las neuronas para comprender distintas patologías encefálicas, y desarrollar enfoques nuevos para el diagnóstico y el tratamiento de estos problemas tan frecuentes. En la página anterior: Estructura molecular de los receptores de glutamato tipo AMPA. Esta proteína es importante para la señalización excitatoria sináptica entre las neuronas (imagen de David Mc Intyre según Sobolevsky y cols. 2009 y Heller y cols., 1993). 2 SeñaleS eléctricaS de laS célulaS nervioSaS Aspectos generales LAS CÉLULAS NERVIOSAS PRODUCEN distintas señales eléctricas que transmiten y almacenan información. Si bien las neuronas no son por sí solas buenas conductoras de la electricidad, desarrollaron mecanismos complejos para generar señales eléctricas basadas en el flujo de iones a través de sus membranas plasmáticas. En general, las neuronas originan un potencial negativo, denominado potencial de membrana de reposo, que puede medirse mediante el registro de la diferencia de voltaje entre el interior y el exterior de las células nerviosas. El potencial de acción produce una abolición transitoria del potencial de reposo negativo y lo convierte en potencial transmembrana positivo. Los potenciales de acción se propagan a lo largo de los axones y constituyen la señal fundamental que transmite información de un lugar a otro en el sistema nervioso. Se producen otros tipos de señales eléctricas por la activación de contactos sinápticos entre las neuronas o por las acciones de formas externas de energía sobre las neuronas sensitivas. Todas estas señales eléctricas surgen por el flujo de iones producidos por permeabilidad selectiva de las membranas de las células nerviosas a diferentes iones y por la distribución no uniforme de estos iones a través de la membrana. Señales eléctricas de las células nerviosas Las neuronas emplean diferentes tipos de señal eléctrica para codificar y transferir información. La mejor forma de observar estas señales es utilizar un microelectrodo intracelular para medir el potencial eléctrico a través de la membrana plasmática neuronal. Un microelectrodo típico es un trozo de tubo de vidrio muy fino (con un orificio de diámetro inferior a 1 μm) y lleno con un buen conductor eléctrico, como una solución concentrada de sal. Este conductor puede conectarse entonces a un voltímetro, normalmente un ordenador, para registrar el voltaje transmembrana de la célula nerviosa. El primer tipo de fenómeno eléctrico puede observarse tan pronto como se inserta un microelectrodo a través de la membrana de la neurona. Al ingresar en la célula, el microelectrodo registra un potencial negativo, que indica que las neuronas tienen un medio para generar un voltaje constante a través de sus membranas cuando están en reposo. Este voltaje, llamado potencial de membrana de reposo, depende del tipo de neurona que se examine, pero siempre es una fracción de un voltio (en los casos típicos, −40 a −90 mV). 26 CAPÍTULO 2 Registro Potencial de membrana (mV) (A) Potencial de receptor FIGURA 2.1 Tipos de señales eléctricas neuronales. En todos los casos se utilizan microelectrodos para medir los cambios en el Tacto de la piel –50 potencial de membrana de reposo durante las –55 –60 0 20 30 Tiempo (ms) 40 50 de acción en una neurona motora espinal. –60 Registro Potencial de membrana (mV) (B) Potencial sináptico 10 señales indicadas. (A) Un estímulo táctil breve produce un potencial de receptor en un corpúsculo de Pacini en la piel. (B) La activación de un contacto sináptico en una neurona piramidal del hipocampo produce un potencial sináptico. (C) La estimulación de un reflejo espinal produce un potencial –65 Sinapsis activada Potencial de membrana (mV) membrana de reposo en la neurona piramidal. Los potenciales sinápticos sirven –70 como medio para intercambiar información en circuitos neurales complejos de 0 5 10 15 20 25 los sistemas nerviosos central y periférico Tiempo (ms) (véase Cap. 5). (C) Potencial de acción +40 Por último, las neuronas generan un Registro tipo especial de señal eléctrica que viaja a lo largo de sus largos axones. Las señales eléctricas se llaman potenciales –10 Neurona de acción (también se denominan “espimotora activada gas” o “impulsos”). En la Figura 2.1C se muestra un ejemplo de potencial de ac–60 Estímulo ción registrado en el axón de una neurona 0 2 4 6 8 10 motora espinal. Los potenciales de acción Tiempo (ms) son responsables de la transmisión de largo alcance de la información dentro del Las señales eléctricas producidas por las neuronas se originan sistema nervioso y le permiten transmitir dicha información a a partir de las respuestas a los estímulos, que entonces cambian sus órganos diana, como el músculo. el potencial de membrana de reposo. Los potenciales del reUn modo de obtener un potencial de acción es hacer pasar una ceptor se deben a la activación de neuronas sensitivas por estí- corriente eléctrica a través de la membrana de la neurona. En mulos externos, como luz, sonido o calor. Por ejemplo, el tacto circunstancias normales, esta corriente sería generada por potende la piel activa los corpúsculos de Pacini, neuronas receptoras ciales de receptor o potenciales postsinápticos. Sin embargo, en que detectan alteraciones mecánicas de la piel. Estas neuronas el laboratorio es fácil producir una corriente eléctrica apropiada responden al tacto generando un potencial del receptor que cam- al insertar un segundo microelectrodo en la misma neurona y bia el potencial de reposo durante una fracción de segundo (Fig. conectar luego el electrodo a una batería (Fig. 2.2A). Si la co2.1A). Estas alteraciones transitorias en el potencial son el pri- rriente así aplicada vuelve más negativo el potencial de memmer paso para generar la sensación de vibraciones (o “cosqui- brana (hiperpolarización), no se observa nada espectacular. El llas”) de la piel en el sistema somatosensitivo (véase Cap. 9). potencial de membrana simplemente cambia en proporción a Se observan tipos similares de potenciales del receptor en todas la magnitud de la corriente inyectada (parte central de la Fig. las demás neuronas sensitivas durante la transducción de señales 2.2B). Estas respuestas de hiperpolarización no necesitan una sensitivas (véase Unidad II). propiedad singular de las neuronas y, por lo tanto, se denominan Otro tipo de señal eléctrica se asocia con la comunicación en- respuestas eléctricas pasivas. Se observa un fenómeno mucho tre las neuronas en los contactos sinápticos. La activación de es- más interesante si se aplica la corriente de polaridad opuesta, de tas sinapsis genera potenciales sinápticos, que permiten trans- modo que el potencial de membrana de la célula nerviosa se hace mitir la información de una neurona a otra. En la Figura 2.1B se más positivo que el de reposo (despolarización). En este caso, en muestra un ejemplo de dicha señal. En este caso, la activación cierto nivel del potencial de membrana, llamado potencial umde una terminación sináptica que inerva una neurona piramidal bral, se desarrolla un potencial de acción (véase el lado derecho del hipocampo produce un cambio muy breve en el potencial de de la Fig. 2.2B). Estímulo SEÑALES ELÉCTRICAS DE LAS CÉLULAS NERVIOSAS (A) Microelectrodo para administrar corriente Neurona Registro Microelectrodo para medir el potencial de membrana +2 0 –2 Potenciales de acción +40 Potencial de membrana (mV) Estímulo Corriente (nA) (B) 27 Insertar microelectrodo 0 Despolarización Respuestas pasivas –50 Umbral –65 Potencial de reposo –100 Hiperpolarización FIGURA 2.2 Registro de las señales eléctricas pasivas y activas en una célula nerviosa. (A) Se insertan dos microelectrodos en una neurona; uno mide el potencial de membrana mientras el otro transmite corriente a la neurona. (B) La inserción del microelectrodo medidor de voltaje en la neurona permite registrar un potencial negativo, el potencial de membrana de reposo. La transmisión de corriente a través del microelectrodo para el paso de corriente altera el potencial de membrana neuronal. Los pulsos de corriente hiperpolarizantes solo producen cambios pasivos en el potencial de membrana. Aunque las pequeñas corrientes despolarizantes también producen solo respuestas pasivas, las despolarizaciones que hacen que el potencial de membrana llegue al umbral o lo exceda además evocan potenciales de acción. Los potenciales de acción son respuestas activas en el sentido de que se generan debido a cambios en la permeabilidad de la membrana neuronal. El potencial de acción es una respuesta activa generada por la neurona y, típicamente, es un cambio breve (de alrededor de 1 ms) de negativo a positivo en el potencial transmembrana. Es importante destacar que la amplitud del potencial de acción es independiente de la magnitud de la corriente utilizada para evocarlo; o sea, las corrientes más grandes no producen potenciales de acción mayores. Por lo tanto, se afirma que los potenciales de acción de una neurona dada son todo o nada, pues se desarrollan por completo o no lo hacen. Si se aumenta lo suficiente la amplitud o la duración de la corriente de estímulo, se producen numerosos potenciales de acción, como puede observarse en las respuestas a tres intensidades diferentes de corriente que se muestran en la Figura 2.2B (lado derecho). En consecuencia, la intensidad de un estímulo está codificada en la frecuencia de potenciales de acción y no en su amplitud. Esta organización difiere espectacularmente de los potenciales de receptor, cuyas Tiempo amplitudes están graduadas en proporción a la magnitud del estímulo sensitivo o de los potenciales sinápticos, cuya amplitud varía según el número de sinapsis activadas, la fuerza de cada sinapsis y la cantidad previa de actividad sináptica. Transmisión a larga distancia de las señales eléctricas El uso de las señales eléctricas (como ocurre al trasmitir electricidad por cables eléctricos para proporcionar potencia o información) presenta una serie de problemas en ingeniería eléctrica. Un problema fundamental para las neuronas es que sus axones, que pueden ser muy largos (recuerde que una neurona motora espinal puede extenderse un metro o más), no son buenos conductores eléctricos. Aunque tanto las neuronas como los cables son capaces de conducir pasivamente la electricidad, las propiedades eléctricas de las neuronas no son comparables con las de un cable eléctrico común. Es posible observar esto midiendo las propiedades eléctricas pasivas del axón de una célula nerviosa al determinar el cambio de voltaje resultante de un pulso de corriente que pasa a través de la membrana axónica (Fig. 2.3A; véase también Recuadro 2A). Si este pulso de corriente se encuentra por debajo del umbral para generar un potencial de acción, la magnitud del cambio de potencial resultante disminuirá al aumentar la distancia desde el sitio de inyección de la corriente (Fig. 2.3B). En condiciones típicas, el potencial cae hasta una pequeña fracción de su valor inicial a una distancia de no más de un par de milímetros desde el sitio de la inyección (Fig. 2.3C). En comparación, un cable eléctrico típicamente permitiría el flu- 28 CAPÍTULO 2 La conducción pasiva disminuye con la distancia Estímulo Electrodo para administrar corriente (A) Axón Electrodos para el registro del potencial Registro Potencial de membrana (mV) (B) Re Registro cord Potencial de membrana (mV) Re Registro cord Registro Registro Registro Registro –59 –62 –65 0 10 20 30 40 (C) 1 mm –50 0 10 20 30 40 0 10 20 30 40 0 10 20 30 40 Tiempo (ms) 0 10 20 30 40 0 10 20 30 40 0 10 20 30 40 Umbral –55 –60 –65 Potencial de reposo –0,5 0 0,5 1,0 1,5 Distancia a lo largo del axón (mm) 2,0 2,5 La conducción activa es constante con la distancia Estímulo Electrodo para administrar la corriente (D) Axón Electrodos para el registro del potencial Registro Potencial de membrana (mV) (E) Registro Record 1 mm Registro Record Registro Record Registro Record Registro Record Registro Record 0 –50 –65 0 2 4 6 8 0 2 4 6 8 0 2 4 6 8 0 2 4 6 8 Tiempo (ms) 0 2 4 6 8 0 2 4 6 8 0 (F) Potencial de membrana (mV) 25 0 –25 –50 –65 –0,5 Umbral Potencial de reposo 0 0,5 1,0 1,5 Distancia a lo largo del axón (mm) 2,0 2,5 2 4 6 8 SEÑALES ELÉCTRICAS DE LAS CÉLULAS NERVIOSAS ▼ FIGURA 2.3 Flujo de corriente pasivo y activo en un axón. (A) Organización experimental para examinar el flujo pasivo de corriente eléctrica en un axón. Un electrodo para el paso de corriente produce una corriente que genera un cambio subumbral en el potencial de membrana, que se propaga pasivamente a lo largo del axón. (B) Respuestas del potencial registradas en las posiciones indicadas por los microelectrodos. Con una distancia creciente desde el sitio de inyección de la corriente, se atenúa la amplitud del cambio en el potencial a medida que la corriente escapa del axón. (C) Relación entre la amplitud de las respuestas del potencial y la distancia. (D) Si se repite el experimento que se muestra en (A) con una corriente supraumbral, se evoca una repuesta activa, el potencial de acción. (E) Potenciales de acción registrados en las posiciones indicadas por los microelectrodos. La amplitud del potencial de acción es constante a lo largo del axón, aunque el tiempo de aparición del potencial de acción se retarda con la distancia creciente. (F) Amplitud constante de un potencial de acción (línea negra sólida) a diferentes distancias. (Tomado de Hodgkin y Rushton, 1938.) jo de corriente pasiva en distancias miles de veces más largas. La disminución progresiva de la amplitud del cambio de potencial inducido se produce porque la corriente inyectada se va perdiendo a través de la membrana axónica; en consecuencia, cuanto más lejos se está a lo largo del axón, existe menos corriente cambie el potencial de membrana. Esta pérdida de la membrana axónica impide una conducción pasiva efectiva de señales eléctricas en todos los axones salvo en los más cortos (los que tienen 1 mm de longitud o menos). Para compensar esta deficiencia, los potenciales de acción sirven como un “sistema de refuerzo” que permite a las neuronas conducir señales eléctricas en grandes distancias a pesar de las pobres propiedades eléctricas pasivas de los axones. La capacidad de los potenciales de acción para reforzar la propagación espacial de señales eléctricas se puede observar si se repite el experimento de la Figura 2.3A con un pulso de corriente despolarizante que sea suficientemente grande como para producir un potencial de acción (Fig. 2.3D). En este caso, el resultado es muy distinto. Ahora se observa un potencial de acción de amplitud constante a lo largo de toda la longitud del axón (Fig. 2.3E). El hecho de que la señalización eléctrica ocurra ahora sin ningún decremento (Figura 2.3F) indica que la conducción 29 activa a través de los potenciales de acción constituye una forma muy eficaz de superar la pérdida intrínseca de las neuronas. Como los potenciales de acción son la base de la transferencia de información en el sistema nervioso, es esencial comprender el modo en que surgen estas y otras señales eléctricas neuronales. Cabe destacar que todas las señales eléctricas neuronales antes descritas se producen por mecanismos similares que se basan en el movimiento de iones a través de la membrana neuronal. El resto de este capítulo, se centra en la pregunta de cómo las células nerviosas utilizan iones para generar potenciales eléctricos. En el Capítulo 3 se explora más específicamente el medio por el cual se producen los potenciales de acción y el modo en que estas señales resuelven el problema de la conducción eléctrica a larga distancia en el interior de las células nerviosas. En el Capítulo 4 se examinan las propiedades de las moléculas de la membrana responsables de producir señalización eléctrica. Por último, en los Capítulos 5-8 se considera cómo se transmiten las señales eléctricas entre las células nerviosas en los contactos sinápticos. De qué modo los movimientos iónicos producen señales eléctricas Los potenciales eléctricos se generan a través de las membranas de las neuronas (y en realidad de todas las células) porque 1) hay diferencias en las concentraciones de iones específicos a través de las membranas de las células nerviosas, y 2) las membranas son selectivamente permeables a algunos de estos iones. A su vez, estos dos hechos dependen de dos tipos diferentes de proteínas en la membrana celular (Fig. 2.4). Los gradientes de concentración de los iones son establecidos por proteínas conocidas como transportadores activos, que, como su nombre sugiere, mueven de manera activa los iones hacia el interior o el exterior de las células en contra de sus gradientes de concentración. La permeabilidad selectiva de las membranas se debe en gran parte a los canales iónicos, proteínas que permiten que solo ciertos tipos de iones atraviesen la membrana en la dirección de sus gradientes de concentración. Por lo tanto, los canales y los Iones Exterior 1 El ion se une 2 El ion se transporta a través de la membrana Transportadores de iones • Mueven activamente iones seleccionados en contra de un gradiente de concentración • Crean gradientes de concentración de iones FIGURA 2.4 Los transportadores activos y los canales iónicos son responsables de los Membrana neuronal movimientos de iones a través de las membranas neuronales. Los transportadores crean Interior diferencias en las concentraciones de los iones al transportar en forma activa los iones en contra de sus gradientes químicos. Los canales aprovechan El ion se difunde a través del canal Canales iónicos • Permiten que los iones se difundan a favor del gradiente de concentración • Producen una permeabilidad selectiva a ciertos iones estos gradientes de concentración y permiten el movimiento de iones seleccionados mediante la difusión, de manera que los gradientes químicos disminuyen. 30 CAPÍTULO 2 RECUADRO 2A Propiedades de membrana pasiva donde Vx es la respuesta de voltaje a cualquier distancia x a lo largo del axón, V0 es el cambio de voltaje en el punto en que se inyecta la corriente en el axón, e es la base de los logaritmos naturales (aproximadamente 2,7) y λ es la constante de longitud del axón. Como se aprecia en esta relación, la constante de longitud es la distancia donde la respuesta inicial de voltaje (V0) disminuye hasta 1/e (o el 37%) de su valor. Por lo tanto, la constante de longitud es una forma de caracterizar lo lejos que se propaga el flujo de corriente pasiva antes de salir del axón, y los axones con mayor (A) Respuestas superpuestas a un pulso de corriente, medido a las distancias indicadas a lo rm h ro ri Por lo tanto, para mejorar el flujo pasivo de corriente a lo largo de un axón, la resistencia de la membrana plasmática debe ser tan alta como sea posible, y las resistencias del axoplasma y del medio extracelular deben ser bajas. Otra consecuencia importante de las propiedades pasivas de las neuronas es que las corrientes que fluyen a través de una membrana no cambian inmediatamente el potencial de membrana. Por ejemplo, cuando se inyecta un pulso de corriente rectangular en el axón que se muestra en el experimento ilustrado en la Figura 2.3A, el potencial de membrana se despo(A) Corriente (nA) Vx = V0 e−x/λ pérdida tienen constantes de longitud más pequeñas. La constante de longitud depende de las propiedades físicas del axón, en particular de las resistencias relativas de la membrana plasmática (rm), el axoplasma intracelular (ri) y el medio extracelular (ro). La relación entre estos parámetros es: lariza lentamente en algunos milisegundos y después se repolariza en un curso temporal similar cuando termina el pulso de corriente (véase Fig. A). Estas demoras en el cambio del potencial de membrana se deben al hecho de que la membrana plasmática se comporta como un condensador, almacenando la carga inicial que fluye al comienzo y al final del pulso de corriente. Para el caso de una célula cuyo potencial de membrana es espacialmente uniforme, el cambio del potencial de membrana en cualquier momento, Vt, después del inicio del pulso de corriente (Fig. C) también puede describirse como una relación exponencial: Vt = V∞ (1 – e –t/τ) donde V∞ es el valor del cambio del potencial de membrana en estado de equilibrio, t es el tiempo después de que se inicia el pulso de corriente y τ es la constante de tiempo de la membrana. Por lo tanto, la constante de tiempo se define como el tiempo cuando la respuesta de voltaje (Vt) se eleva hasta 1 – (1/e) +1 0 –1 Distancia desde la inyección de la corriente (mm) –60 0 Potencial de membrana (mV) El flujo pasivo de la corriente eléctrica desempeña un papel central en la propagación del potencial de acción, la transmisión sináptica y todas las otras formas de señalización eléctrica en las células nerviosas. Por lo tanto, vale la pena conocer en términos cuantitativos cómo varía el flujo de corriente pasiva con la distancia a lo largo de una neurona. Para el caso de un axón cilíndrico, como en el que se muestra en la Figura 2.3, la corriente subumbral inyectada en una porción del axón se extiende pasivamente a lo largo de éste hasta que se disipa por pérdida a través de la membrana axónica. Como consecuencia de esta pérdida, los resultados medidos a lo largo del axón se hacen más pequeños según aumenta la distancia desde el sitio en que se inyecta la corriente (Fig. A). Se puede determinar el decremento del flujo de corriente con la distancia representando la reducción relativa de la amplitud de los potenciales con la distancia (Fig. B). Esta relación se describe mediante una función exponencial simple: 0,5 1,0 1,5 2,0 –65 0 largo de un axón. transportadores funcionan básicamente en contra unos de otros, y al hacerlo generan el potencial de membrana de reposo, los potenciales de acción y los potenciales sinápticos y de receptor que desencadenan potenciales de acción. En el Capítulo 4 se descri- 10 2,5 20 30 40 Tiempo (ms) ben la estructura y la función de estos canales y transportadores. Para apreciar el papel de los gradientes iónicos y la permeabilidad selectiva en la generación de un potencial de membrana, considérese un sistema simple en el que una membrana artificial SEÑALES ELÉCTRICAS DE LAS CÉLULAS NERVIOSAS 31 RECUADRO 2A (continúa) (B) 1,0 Vx = V0e 0,8 (B) Disminución espacial del potencial de membrana a lo largo de un axón cilíndrico. Un pulso de corriente inyectado en un punto en el axón (0 mm) –x/h produce respuestas de voltaje (Vc) que disminuyen exponencialmente con la distancia. La distancia Vx/V0 0,6 0,4 donde la respuesta de voltaje es 1/e de su valor inicial (V0) es la constante de longitud, λ. 37% 0,2 0,0 –5 –4 –3 –2 –1 0 1 2 3 4 5 h h Distancia desde la inyección de la corriente (mm) (o 63%) de V∞. Después de que termina el pulso de corriente, también disminuye exponencialmente el cambio del potencial de membrana de acuerdo con la relación: resistencia (rm) y de la capacitancia (cm) de la membrana plasmática como tal: Vt = V∞ e–t/τ Los valores de rm y de cm dependen en parte del tamaño de la neurona, y las células más grandes tienen resistencias más bajas y capacitancias más grandes. En general, las células nerviosas pequeñas suelen tener constante de tiempo prolongadas y las células grandes, constantes de tiempo breves. (C) Corriente (nA) Durante esta disminución, el potencial de membrana retorna a 1/e de V∞ en un tiempo igual a τ. En las células con geometrías más complejas que el axón de la Figura 2.3A, los cursos temporales de los cambios en el potencial de membrana no son exponenciales simples, pero no obstante dependen de la constante de tiempo de la membrana. Por lo tanto, la constante de tiempo caracteriza lo rápido que el flujo de corriente cambia el potencial de membrana. La constante de tiempo de la membrana también depende de las propiedades físicas de la célula nerviosa, específicamente de la τ = rm c m Bibliografía Hodgkin, A. L. and W. A. H. Rushton (1938) The electrical constants of a crustacean nerve fibre. Proc. R. Soc. Lond. 133: 444-479. Johnston, D. and S. M.-S. Wu (1995) Foundations of Cellular Neurophysiology. Cambridge, MA: MIT Press. Rall, W. (1977) Core conductor theory and cable properties of neurons. In Handbook of Physiology, Section 1: The Nervous System, Vol. 1: Cellular Biology of Neurons. E. R. Kandel (ed.). Bethesda, MD: American Physiological Society, pp. 39-98. 1 0 –1 1,0 0,80 63% (C) Curso temporal de los cambios de potencial producidos en una célula espacialmente uniforme por un pulso de corriente. La elevación y la caída del potencial de membrana (V∞τ) pueden 0,60 describirse como funciones exponenciales, y la constante de tiempo que define el tiempo requerido para la respuesta hasta la elevación a 1 – (1/e) del valor en estado de equilibrio (V∞) o hasta la declinación a 1/e de V∞. 0,20 0,40 0,00 separa dos compartimientos que contienen soluciones de iones. Para comparar este hecho con la situación en las neuronas, nos referiremos al compartimiento izquierdo como el interior y denominaremos al compartimiento derecho exterior. En este siste- 37% 0 5 10 15 20 o Tiempo (ms) 25 30 35 40 o ma es posible determinar la composición de las dos soluciones y controlar así los gradientes iónicos a través de la membrana. Por ejemplo, tómese el caso de una membrana que es permeable solo a iones potasio (K+). Si la concentración de K+ a cada 32 CAPÍTULO 2 (A) (C) (B) Condiciones iniciales Voltímetro V=0 En equilibrio Inicialmente V=0 Vinterior-exterior = –58 mV [K+]in (mM) Potencial de membrana V1–2 (mV) 100 Dentro Fuera Dentro Fuera Dentro Fuera 1 mM KCl 1 mM KCl 10 mM KCl 1 mM KCl 10 mM KCl 1 mM KCl Permeable al K+ Sin flujo neto de K+ Flujo neto de K+ desde dentro hasta fuera Flujo de K+ desde dentro hasta fuera equilibrado por el potencial de membrana opuesto FIGURA 2.5 Equilibrio electroquímico. (A) Una membrana permeable solo al K+ (esferas amarillas) separa los compartimientos interior y exterior, que contienen las concentraciones indicadas de KCl. (B) El incremento de la concentración de KCl en el compartimiento interior hasta 10 mM produce primero un movimiento pequeño de K+ hacia el compartimiento exterior (condiciones iniciales) hasta que la fuerza electromotriz que actúa sobre el K+ equilibra el gradiente de concentración, y el movimiento neto de K+ se vuelve cero (en equilibrio). (C) Relación entre el gradiente de concentración transmembrana ([K+]ext./[K+]int.) y el potencial transmembrana. Según lo que predice la ecuación de Nernst, esta relación es lineal cuando se representa en coordenadas semilogarítmicas, con una pendiente de 58 mV por diferencia de diez veces en el gradiente de concentración. lado de esta membrana es igual, entonces no se medirá potencial eléctrico alguno a través de ella (Fig. 2.5A). No obstante, si la concentración de K+ no es igual en ambos lados, se generará un potencial eléctrico. Por ejemplo, si la concentración de K+ en el compartimiento interior es 10 veces mayor que la concentración de K+ en el compartimiento exterior, entonces el potencial eléctrico del interior será negativo respecto del exterior (Fig. 2.5B). Esta diferencia del potencial eléctrico se genera porque los iones potasio fluyen a favor de su gradiente de concentración y trasladan su carga eléctrica (una carga positiva por ion) con ellos a medida que avanzan. Dado que las membranas neuronales contienen bombas que acumulan K+ en el citoplasma celular y puesto que los canales permeables al potasio en la membrana plasmática permiten un flujo de K+ transmembrana, hay una situación análoga en las células nerviosas vivas. Por lo tanto, un eflujo en reposo continuo de K+ es el responsable del potencial de membrana de reposo. En la situación imaginaria que se acaba de describir, el equilibrio se alcanzará con rapidez. A medida que el K+ se mueva del compartimiento interior al exterior (las condiciones inicia- 10 1 0 –58 Pendiente = 58 mV por cambio de 10 veces en el gradiente de K+ –116 –2 –1 0 [K+] log + fuera [K ]dentro les; éstas se muestran a la izquierda de la Fig. 2.5B), se genera el potencial que tiende a impedir un mayor flujo de K+. Este impedimento es el resultado del hecho de que el gradiente de potencial a través de la membrana tiende a rechazar los iones potasio positivos a medida que intentan atravesar la membrana. O sea que a medida que el exterior se vuelve más positivo respecto del interior, la positividad creciente hace al exterior menos atrayente al K+ con carga positiva. El movimiento (o flujo) neto de potasio se detendrá en el punto (“en el equilibrio”, a la derecha de la Fig. 2.5B) donde el cambio de potencial a través de la membrana (la positividad relativa del compartimiento exterior) supere exactamente el gradiente de concentración (un exceso de 10 veces de K+ en el compartimiento interior). En este equilibrio electroquímico hay un balance exacto entre dos fuerzas opuestas: 1) el gradiente de concentración que provoca el movimiento de K+ desde el interior hacia el exterior, trasladando consigo la carga positiva, y 2) un gradiente eléctrico opuesto que tiende a detener cada vez más el movimiento de K+ a través de la membrana (véase Fig. 2.5B). La cantidad de iones que deben fluir para generar este potencial eléctrico es muy pequeña (aproximadamente 10−12 moles de K+ por cm2 de membrana o 1012 iones K+). Este último hecho es importante de dos formas. Primero, indica que las concentraciones de iones permeables a cada lado de la membrana siguen siendo esencialmente constantes, aun después de que el flujo de iones ha generado el potencial. Segundo, los pequeños flujos de iones necesarios para establecer el potencial de membrana no interrumpen la electroneutralidad química porque cada ion tiene otro contrario de carga opuesta (iones cloruro, en la Fig. 2.5) para mantener la neutralidad de las soluciones a cada lado de la membrana. La concentración de K+ sigue siendo igual a la de Cl− en las soluciones de ambos comportamientos, de modo que la separación de la carga que crea la diferencia de potencial está limitada a la proximidad inmediata de la membrana. SEÑALES ELÉCTRICAS DE LAS CÉLULAS NERVIOSAS Las fuerzas que crean los potenciales de membrana El potencial eléctrico generado a través de la membrana en equilibrio electroquímico (potencial de equilibrio) puede predecirse mediante una fórmula sencilla denominada ecuación de Nernst. Por lo general, esta relación se expresa como: EX RT zF ln Xexterior X interio r donde EX es el potencial de equilibrio para cualquier ion X, R es la constante de los gases, T es la temperatura absoluta (en grados en la escala Kelvin), z es la valencia (carga eléctrica) del ion permeable y F es la constante Faraday (la cantidad de carga eléctrica contenida en un mol de un ion equivalente). Los corchetes indican las concentraciones del ion X a cada lado de la membrana, y el símbolo ln indica el logaritmo natural del gradiente de concentración. Como es más fácil realizar cálculos utilizando logaritmos en base 10 a temperatura ambiente, esta relación suele simplificarse a: EX 58 X exterior log X interio r z donde log indica el logaritmo en base 10 del cociente de las concentraciones. Por lo tanto, para el ejemplo de la Figura 2-5B, el potencial a través de la membrana del equilibrio electroquímico es EK K exterior 58 log 1 –58 mV 58 log z 10 K interior De manera convencional, el potencial de equilibrio se define en términos de la diferencia de potencial entre el comportamiento exterior e interior. Por lo tanto, cuando la concentración de K+ es mayor en el interior que en el exterior, se mide un potencial interior negativo a través de la membrana neuronal permeable al K+. Para un sistema hipotético simple que solo tiene una especie de ion permeable, la ecuación de Nernst permite predecir con exactitud el potencial eléctrico a través de la membrana en equilibrio. Por ejemplo, si la concentración de K+ del lado interior se aumenta hasta 100 mM, el potencial de membrana será de −116 mV. De modo más general, si se representa el potencial de membrana contra el logaritmo del gradiente de concentración de K+ ([K+]exterior/[K+]interior), la ecuación de Nernst predice una relación lineal con una pendiente de 58 mV (en realidad, 58/z) por cambio de 10 veces en el gradiente de K+ (véase Fig. 2.5C). Para reforzar y ampliar el concepto de equilibrio electroquímico, considérense algunos experimentos adicionales sobre la influencia de las especies iónicas y la permeabilidad iónica que podría realizarse en el sistema de modelo simple de la Figura 2.5. ¿Qué sucedería con el potencial eléctrico a través de la membrana (es decir, el potencial del interior respecto del exterior) si el K+ en el interior se reemplazara por 1 mM de Na+? No se generaría potencial, dado que no podría fluir el Na+ a través de la membrana (que se definió como solo permeable al K+). Sin embargo, si 33 en estas condiciones iónicas (10 veces más Na+ en el exterior) la membrana permeable al K+ se reemplazara mágicamente por una membrana permeable solo al Na+, se mediría un potencial de +58 mV en equilibrio. Si se presentaran 10 mM de calcio (Ca2+) en el exterior y 1 mM de Ca2+ en el interior y una membrana selectiva al Ca2+ separara los dos lados, ¿qué sucedería con el potencial de membrana? Se desarrollaría un potencial de +29 mV, la mitad del observado para el Na+, porque la valencia del calcio es +2. Por último, ¿qué sucedería con el potencial de membrana si se presentaran 10 mM de Cl− en el exterior, con ambos lados separados por una membrana permeable al Cl−? Como la valencia de este anión es −1, el potencial sería de +58 mV. El balance de las fuerzas físicas y químicas en equilibrio significa que el potencial eléctrico puede determinar los flujos iónicos a través de la membrana, al igual que el gradiente iónico puede determinar el potencial de membrana. Para examinar la influencia del potencial de membrana sobre el flujo iónico, imagínese conectar una batería a través de ambos lados de la membrana para controlar el potencial eléctrico sin modificar la distribución de iones a ambos lados (Fig. 2.6). Mientras la batería no funcione, las cosas estarán igual que en la Figura 2.4, y el flujo de K+ desde el interior hacia el exterior producirá un potencial de membrana negativo (Fig. 2.6A, izquierda). Sin embargo, si se utiliza la batería para volver el interior inicialmente más negativo que el exterior, habrá menor flujo de K+, dado que el potencial negativo tiende a mantener el K+ en el compartimiento interno. ¿Qué grado de negatividad debe tener el compartimiento interno antes de que no haya flujo neto de K+? La respuesta es −58 mV, el voltaje necesario para contrarrestar la diferencia de 10 veces en las concentraciones de K+ a ambos lados de la membrana (Fig. 2.6A, centro). Si en un inicio el interior se hace más negativo que −58 mV, entonces el K+ realmente fluirá desde el exterior (compartimiento 2, a la derecha) hacia el interior (compartimiento 1, a la izquierda) dado que los iones positivos serán atraídos por el potencial más negativo del interior (Fig. 2.6A, derecha). En este ejemplo se demuestra que tanto la dirección como la magnitud del flujo dependen del potencial de membrana. Por lo tanto, en ciertas circunstancias el potencial eléctrico puede superar un gradiente de concentración iónico. La capacidad para alterar experimentalmente el flujo iónico para modificar el potencial impuesto sobre la membrana (Fig. 2.6B) o el gradiente de concentración transmembrana para un ion (véase Fig. 2.5C) proporciona herramientas convenientes para estudiar los flujos de iones a través de las membranas plasmáticas de las neuronas, como se observará en muchos de los experimentos descritos en los capítulos siguientes. Equilibrio electroquímico en un medioambiente con más de un ion permeable Consideremos ahora una situación más compleja en la que Na+ y K+ están distribuidos en forma desigual a través de la Batería conectada Batería conectada Vdentro-fuera = 0 mV Vdentro-fuera = –58 mV Vdentro-fuera = –116 mV Batería Batería Batería (B) Flujo neto de K+ desde dentro hasta fuera fuera Batería desconectada Dentro (A) 0 Fuera Dentro CAPÍTULO 2 Flujo neto de K+ 34 –116 –58 0 Sin flujo neto de K+ Flujo neto de K+ desde fuera hasta dentro Potencial de membrana Vadentro-afuera (mV) Dentro Fuera Dentro Fuera Dentro Fuera 10 mM KCl 1 mM KCl 10 mM KCl 1 mM KCl 10 mM KCl 1 mM KCl Flujo neto de K+ desde dentro hasta fuera Sin flujo neto de K+ Flujo neto de K+ desde fuera hasta dentro FIGURA 2.6 El potencial de membrana influye en los flujos de iones. (A) La conexión de una batería a través de la membrana permeable al K+ permite el control directo del potencial de membrana. Cuando la batería no funciona (izquierda), los iones K+ (amarillo) fluyen simplemente según su gradiente de concentración. La regulación del potencial de membrana inicial (Vinterior-exterior) en el potencial de equilibrio para el K+ (centro) no arroja un flujo neto de K+, mientras que si el potencial de membrana se vuelve más negativo que el potencial de equilibrio del K+ (derecha), el K+ influye en contra de su gradiente de concentración. (B) Relación entre el potencial de membrana y la dirección del flujo de K+. membrana, como aparece en la Figura 2.7A. ¿Qué sucedería si se presentara K+ 10 mM y Na+ 1 mM en el interior, y K+ 1 mM y Na+ 10 mM en el exterior? Si la membrana fuera permeable solo a K+, el potencial de membrana sería −58 mV; si la membrana fuera permeable solo a Na+, el potencial sería +58 mV. No obstante, ¿cuál sería el potencial si la membrana fuera permeable tanto a K+ como a Na+? En este caso, el potencial dependería de la permeabilidad relativa de la membrana a K+ y Na+. Si es más permeable al K+, el potencial se aproxima a −58 mV, y si es más permeable a Na+, el potencial se aproxima a +58 mV. Dado que no hay un término de permeabilidad en la ecuación de Nernst, que considera el caso simple de una sola especie de ion permeable, se necesita una ecuación más elaborada para tomar en cuenta ambos gradientes de los iones permeables y la permeabilidad relativa de la membrana a cada especie permeable. Esta ecuación fue desarrollada por David Goldman en 1943. Para el caso más relevante de las neuronas, en las que Na+, K+ y Cl− son los iones permeables primarios, la ecuación de Goldman se escribe V 58 log PK K ext PNa Na ext PCl Cl int PK K int PNa Na int PCl Cl ext donde V es el voltaje a través de la membrana (de nuevo, el compartimiento interior respecto del compartimiento exterior de referencia) y P indica la permeabilidad de la membrana a cada ion de interés. Por lo tanto, la de Goldman es una ecuación ampliada de la ecuación de Nernst, que toma en cuenta las permeabilidades relativas de cada uno de los iones involucrados. La relación entre las dos ecuaciones se hace obvia en la situación en la cual la membrana es permeable solo a un ion, digamos K+; en este caso, la expresión de Goldman cae hasta la ecuación de Nernst más sencilla. En este contexto, es importante señalar que el factor de valencia (z) en la ecuación de Nernst ha sido eliminado; a esto se debe que las concentraciones de los iones cloruro, Cl−, se invirtieran respecto de las concentraciones de los iones con carga positiva [recuerde que –log (A/B) = log (B/A)]. Si la membrana en la Figura 2.7A es permeable solo a K+ y Na+, los términos que involucran a Cl− desaparecen porque PCl es 0. En este caso, el resultado de la ecuación de Goldman arroja un potencial de −58 mV cuando solo es permeable a K+, de +58 mV cuando solo es permeable a Na+ y algún valor intermedio si es permeable a ambos iones. Por ejemplo, si K+ y Na+ fueran igualmente permeables, el potencial sería 0 mV. Con respecto a la señalización neural, es en particular pertinente preguntarse qué sucedería si la membrana comenzara por ser permeable al K+ y luego cambiara transitoriamente para volverse más permeable al Na+. En esta circunstancia, el potencial de membrana comenzaría con un nivel negativo, se volvería positivo mientras la permeabilidad al Na+ se mantuviera alta y luego caería hasta un nivel negativo a medida que la permeabilidad al Na+ cayera otra vez. A medida que se produce, este último caso describe esencialmente lo que se observa en una neurona durante la generación de un potencial de acción. En el estado de reposo, la PK de la membrana neuronal es mucho más alta que la PNa; dado que como resultado de la acción de los transportadores de iones hay siempre más SEÑALES ELÉCTRICAS DE LAS CÉLULAS NERVIOSAS (A) Voltímetro 35 FIGURA 2.7 Los potenciales de reposo y de acción implican permeabilidades a diferentes iones. (A) Situación hipotética en la cual una membrana variablemente permeable a Na+ (rojo) y K+ (amarillo) separa dos compartimientos que contienen ambos iones. Para simplificar, en el diagrama no se muestran los iones Cl−. (B) Esquema de las permeabilidades iónicas de la membrana asociadas con los potenciales de reposo y acción. En reposo, las membranas neuronales son más permeables a K+ (amarillo) que a Na+ (rojo); en consecuencia, el potencial de membrana de reposo es negativo y se aproxima al potencial de equilibrio para K+, EK. Durante un potencial de acción, la membrana se vuelve muy permeable al Na+ (rojo); de este modo, el potencial de membrana se vuelve positivo y se acerca al potencial de equilibrio para el Na+, ENa. Sin embargo, la elevación en la permeabilidad del Na+ es transitoria, de modo que la membrana se vuelve principalmente permeable al K+ y hace que el potencial retorne a su valor de reposo negativo. Obsérvese que en el potencial de equilibrio para un valor dado no hay flujo neto de ese ion a través de la membrana. Dentro 10 mM KCl 1 mM NaCl Fuera 1 mM KCl 10 mM NaCl Permeabilidad variable a Na +y K + (B) Potencial de membrana Permeable al Na+ Permeable al K+ PNa>> PK ENa PNa PNa 0 Potencial de reposo Potencial de acción PK>>PNa Repolarización PK>>PNa EK Tiempo K+ en el interior de la célula que en el exterior (véase Cuadro 2.1), el potencial de reposo es negativo (Fig. 2.7B). A medida que el potencial de membrana se despolariza (por ejemplo, por acción sináptica), aumenta la PNa. El incremento transitorio en la permeabilidad al Na+ hace que el potencial de membrana se vuelva incluso más positivo (región roja de Fig. 2.7B) porque el Na+ ingresa (hay mucho más Na+ en el interior de una neurona que en el exterior, otra vez como resultado de las bombas iónicas). Debido a este circuito de retroalimentación positiva, se desarrolla un potencial de acción. No obstante, este aumento de la permeabilidad al Na+ durante el potencial de acción es transitorio; a medida que se restablece la permeabilidad al Na+, el potencial de membrana retorna rápidamente a su nivel de reposo. Al conocer algunos principios electroquímicos, será mucho más fácil comprender el siguiente informe, más detallado, de cómo las neuronas generan los potenciales de reposo y acción. Base iónica del potencial de membrana de reposo La acción de los transportadores crea gradientes transmembrana sustanciales para la mayoría de los iones. En el Cuadro 2.1 se resumen las concentraciones de los iones medidas en una célula nerviosa excepcionalmente grande hallada en el sistema nervioso del calamar (Recuadro 2B). Estas mediciones son la base para afirmar que hay mucho más K+ en el interior de la neurona que en el exterior, y mucho más Na+ en el exterior de la célula que en el interior. Se desarrollan gradientes de concentración similares en la mayoría de las células de los animales, incluidos los seres humanos. Sin embargo, dado que la potencia iónica de la sangre de los mamíferos es menor que la de los animales que viven en el mar, como el calamar, en los mamíferos las concentraciones de cada ion son varias veces menores. Estos gradientes de concentración dependientes de transportadores constituyen indirectamente la fuente del potencial de membrana neuronal de reposo y del potencial de acción. Una vez que se conocen los gradientes de concentración de los iones a través de distintas membranas neuronales, es posible utilizar la ecuación de Nernst para calcular el potencial de equilibrio para el K+ y de otros iones importantes. Puesto que el potencial de membrana de reposo de la neurona del calamar es de alrededor de −65 mV, el K+ es el ion que está más próximo al equilibrio electroquímico cuando la célula se encuentra en reposo. Esto implica que la membrana en reposo es más permeable al K+ que a los otros iones mencionados en el Cuadro 2.1 y que esta permeabilidad es el origen de los potenciales de reposo. CUADRO 2.1 Concentraciones extracelulares e intracelulares de iones CONCENTRACIÓN (MM) ION INTRACELULAR INTRACELULAR 400 50 40-150 0,0001 20 440 560 10 140 5-15 4-30 0,0001 5 145 110 1-2 Neurona del calamar Potasio (K+) Sodio (Na+) Cloruro (Cl-) Calcio (Ca2+) Neurona de mamífero Potasio (K+) Sodio (Na+) Cloruro (Cl-) Calcio (Ca2+) 36 CAPÍTULO 2 RECUADRO 2B Las notables neuronas gigantes del calamar Muchos de los conceptos iniciales acerca de cómo los gradientes de concentración iónicos y los cambios en la permeabilidad de la membrana producen señales eléctricas provienen de experimentos realizados sobre las neuronas extraordinariamente grandes del calamar. Los axones de estas células nerviosas pueden tener hasta 1 mm de diámetro (100 a 1 000 veces más grandes que los axones de los mamíferos). Por lo tanto, los axones del calamar tienen el tamaño suficiente como para que puedan efectuarse experimentos que serían imposibles en la mayoría de las otras células nerviosas. Por ejemplo, no es difícil insertar electrodos simples de cable en el interior de estos axones gigantes y efectuar mediciones eléctricas fiables. La facilidad relativa de este enfoque arrojó los primeros registros intracelulares de potenciales de acción de células nerviosas y, como se explicará en el próximo capítulo, las primeras mediciones experimentales de las corrientes iónicas que producen potenciales de acción. También resulta práctico extraer el citoplasma de los axones gigantes y medir su composición iónica (véase Cuadro 2-1). Además, algunas neuronas gigantes forman contactos sinápticos con otras y producen sinapsis muy grandes, que fueron de utilidad para comprender los mecanismos fundamentales de la transmisión sináptica (véase Cap. 5). Las neuronas gigantes evidentemente se desarrollaron en el calamar dado que aumentaban la supervivencia del animal. Estas neuronas participan en un circuito neural simple que activa la contracción del músculo del manto y produce un efecto de propulsión de chorro que permite alejarse de los predadores a una velocidad extraordinaria. Como se explica en el Capítulo 3, el diámetro axónico más grande permite una conducción más rápida de los potenciales de acción. Por lo tanto, se supone que el calamar tiene estas neuronas enormes para escapar de manera exitosa de sus abundantes enemigos. En la actualidad (casi 70 años después de que John Z. Young en el University College de Londres las descubriera) las células nerviosas gigantes del calamar aún son sistemas experimentales útiles para estudiar las funciones neuronales básicas. Bibliografía Llinás, R. (1999) The Squid Synapse: A Model for Chemical Transmission. Oxford: Oxford University Press. Young, J. Z. (1939) Fused neurons and synaptic contacts in the giant nerve fibres of cephalopods. Phil. Trans. R. Soc. Lond. 229(B): 465-503. (A) Esquema de un calamar, que muestra la localización de sus células nerviosas gigantes. Los diferentes colores indican los componentes neuronales del circuito de escape. Las neuronas de primer orden y de segundo orden se originan en el cerebro, mientras que las de tercer orden están en el ganglio estrellado e inervan las células musculares del manto. (B). Sinapsis gigantes en el interior del ganglio estrellado. La neurona de segundo orden forma una serie de prolongaciones digitiformes, cada una de las cuales realiza una sinapsis extraordinariamente grande con una única neurona de tercer orden. (C) Estructura de un axón gigante de una neurona de tercer orden que se localiza en el interior de su nervio. Se muestra abajo la diferencia enorme en los diámetros de un axón gigante de calamar y uno de mamífero. (A) (B) (C) Axón gigante Encéfalo Neurona de primer orden Nervio estrellado Axones más pequeños Presináptico (segundo orden) Neurona de segundo orden Ganglio estrellado Neurona de tercer orden Nervio estrellado con axón gigante Postsináptico (tercer orden) Corte transversal 1 mm 1 mm Axón gigante del calamar = 800 µm de diámetro Axón de mamífero = 2 µm de diámetro SEÑALES ELÉCTRICAS DE LAS CÉLULAS NERVIOSAS Base iónica de los potenciales de acción ¿Por qué el potencial de membrana de una neurona se despolariza durante un potencial de acción? Si bien se ha dado una res- Potencial de membrana de reposo (mV) (A) 200 mM K+ 0 –20 –40 3,5 mM K+ –60 –80 10 mM K+ 0 20 mM K+ 50 mM K+ 450 mM K+ 5 Tiempo (min) 10 (B) Potencial de membrana de reposo (mV) Es posible probar esta hipótesis, como lo hicieron Alan Hodgkin y Bernard Katz en 1949, al preguntarse qué le sucede al potencial de membrana de reposo a medida que se altera la concentración de K+ fuera de la neurona. Si la membrana en reposo es permeable solo al K+, entonces la ecuación de Goldman (o incluso la de Nernst, más simple) predice que el potencial de membrana variará en forma proporcional al logaritmo del gradiente de concentración del K+ a través de la membrana. Si se acepta que la concentración interna de K+ no se modifica durante el procedimiento, un gráfico que relacione el potencial de membrana con el logaritmo de la concentración externa de K+ arrojará una línea recta con una pendiente de 58 mV por cambio de 10 veces en la concentración externa de K+ a temperatura ambiente (véase Fig. 2.5C). (La pendiente se vuelve de alrededor de 61 mV a las temperaturas corporales de los mamíferos.) Cuando Hodgkin y Katz realizaron este experimento en la neurona de un calamar vivo, observaron que el potencial de membrana de reposo realmente cambiaba cuando se modificaba la concentración externa de K+ para volverse menos negativo a medida que ésta se elevaba (Fig. 2.8A). Cuando la concentración externa de K+ se elevaba lo suficiente como para igualar a la concentración de K+ en el interior de la neurona y volverse así el potencial de equilibrio del K+ de 0 mV, el potencial de membrana de reposo también era aproximadamente de 0 mV. En resumen, el potencial de membrana de reposo variaba como se predijo con el logaritmo de la concentración de K+, con una pendiente que se aproximaba a los 58 mV por cambio de diez veces en la concentración de K+ (Fig. 2.8B). El valor obtenido no era exactamente de 58 mV porque otros iones, como Cl− y Na+, también son levemente permeables y, por lo tanto, influyen en el potencial de reposo en bajo grado. La contribución de estos otros iones es en particular evidente con bajos niveles externos de K+, de nuevo como predice la ecuación de Goldman. Sin embargo, en general la manipulación de las concentraciones externas de estos otros iones solo tiene un efecto pequeño (véase Fig. 2.9E), lo que destaca que la permeabilidad del K+ es la fuente primaria del potencial de membrana de reposo. En resumen, Hodgkin y Katz mostraron que el potencial de reposo de interior negativo surge porque: 1) la membrana de la neurona en reposo es más permeable al K+ que a cualquiera de los otros iones presentes y 2) hay más K+ en el interior de las neuronas que en el exterior. La permeabilidad selectiva al K+ es causada por los canales de membrana permeables al K+ que están abiertos en las neuronas en reposo, como señalamos, y el gran gradiente de concentración del K+ es producido por transportadores de membrana que acumulan selectivamente K+ en el interior de las neuronas. En muchos estudios ulteriores, se confirmó la validez general de estos principios. 37 0 –20 –40 Pendiente = 58 mV por cambio de 10 veces en el gradiente del K+ –60 –80 2 5 10 20 50 [K+]exterior (mM) 100 200 500 FIGURA 2.8 El potencial de membrana de reposo del axón gigante de calamar está determinado por el gradiente de concentración de K+ a través de la membrana. (A) El incremento de la concentración externa de K+ hace más positivo el potencial de membrana de reposo. (B) Relación entre el potencial de membrana de reposo y la concentración externa de K+, representada en una escala semilogarítmica. La línea recta representa una pendiente de 58 mV por cambio de diez veces en la concentración, según lo demuestra la ecuación de Nernst. (De Hosgkin y Katz, 1949.) puesta general a esta pregunta (aumento de la permeabilidad al Na+), sería conveniente razonarla con mayor detalle. De acuerdo con los datos presentados en el Cuadro 2.1, es posible utilizar la ecuación de Nernst para calcular que el potencial de equilibrio para el Na+ (ENa) en las neuronas, y en realidad en la mayoría de las células, es positivo. Por lo tanto, si la membrana se volviera muy permeable al Na+, el potencial de membrana se aproximaría al ENa. Sobre la base de estas consideraciones, Hodgkin y Katz postularon la hipótesis de que el potencial de acción surge porque la membrana neuronal se convierte transitoriamente permeable al Na+. Hodgkin y Katz, aprovechando el mismo estilo de experimento de sustitución de iones que utilizaron al evaluar el potencial de reposo, analizaron el papel del Na+ en la generación del potencial de acción al preguntarse qué le sucede al potencial de acción cuando se elimina Na+ del medio externo. Estos autores observaron que la disminución de la concentración externa de Na+ reduce tanto el ritmo de elevación del potencial de acción como su amplitud máxima (Fig. 2.9A-C). En efecto, cuando examinaron esta dependencia del Na+ cuantitativamente, observaron una relación más o menos lineal entre la amplitud del potencial de acción y el logaritmo de la concentración externa de Na+ (Fig. 2.9D). La pendiente CAPÍTULO 2 Potencial de membrana (mV) (A) (D) 100 +40 0 –40 –80 0 1 2 Tiempo (ms) 3 Pendiente = 58 mV por cambio de 10 veces en el gradiente de Na+ 40 20 50 0 100 200 500 [Na+]exterior (mM) 1000 (E) –40 0 1 +40 2 Tiempo (ms) 3 Recuperación 0 –20 –40 0 1 2 Tiempo (ms) centraciones iónicas normales en el interior y el exterior de la célula. (B) La amplitud y la velocidad de elevación del potencial de acción disminuyen cuando la concentración externa de sodio está reducida a un tercio del valor normal, pero (C) se recupera cuando se repone el Na+. (D,E) Aunque la amplitud del potencial de acción es muy sensible a la concentración externa de Na+, el potencial de membrana de reposo (E) se afecta poco por los cambios de concentración de este ion. (De Hodgkin y Katz, 1949.) –40 –60 –80 0 –80 (A) Potencial de acción evocado con las con- 60 [Na+] baja (C) Potencial de membrana (mV) 80 +40 Potencial de membrana de reposo (mV) Potencial de membrana (mV) (B) –80 FIGURA 2.9 Papel del sodio en la generación de un potencial de acción en el axón gigante de un calamar. Control Amplitud del potencial de acción (mV) 38 50 500 100 200 [Na+]exterior (mM) 1000 3 de esta relación se aproximaba a un valor de 58 mV por cambio de 10 veces en la concentración de Na+, como era esperable para una membrana selectivamente permeable al Na+. Por el contrario, la reducción de la concentración de Na+ tuvo muy poco efecto sobre el potencial de membrana de reposo (Fig. 2.9E). Por lo tanto, aunque la membrana neuronal en reposo es solo algo permeable al Na+, se hace extraordinariamente permeable a este compuesto durante la fase de ascenso y la fase de exceso (overshoot) del potencial de acción. (Véase en el Recuadro 2C una explicación de la nomenclatura del potencial de acción.) Este incremento transitorio de la permeabilidad al sodio es el resultado de la apertura de los canales selectivos para el Na+ que en esencia están cerrados en el estado de reposo. Las bombas de membrana mantienen un gradiente electroquímico para el Na+, que se encuentra en una concentración mucho mayor por fuera de la neurona y hace que el potencial de membrana se despolarice y se aproxime al ENa. El tiempo durante el cual el potencial de membrana persiste cerca del ENa (aproximadamente +58 mV) durante la fase de exceso de un potencial de acción es breve, dado que el aumento de la permeabilidad al Na+ en sí es breve. El potencial de membrana se repolariza con rapidez hasta los niveles de reposo, y en realidad le sigue una repolarización exagerada (undershoot) transitoria. Como se indica en el Capítulo 3, estos últimos acontecimientos en el ciclo del potencial de acción se deben a una inactivación de la permeabilidad al Na+ y un aumento de la permeabilidad de la membrana al K+. Durante la repolarización exagerada, el potencial de membrana está hiperpolarizado transitoriamente, pues la permeabilidad al K+ se hace incluso mayor que en reposo. El potencial de acción termina cuando esta fase de aumento de la permeabilidad al K+ cede y el potencial de membrana retorna a su nivel de reposo normal. Los experimentos de sustitución de iones llevados a cabo por Hodgkin y Katz brindan indicios convincentes de que 1) el potencial de membrana de reposo es resultado de una permeabilidad elevada de la membrana en reposo al K+ y 2) que la despolarización durante un potencial de acción es consecuencia de un aumento transitorio de la permeabilidad de la membrana al Na+. Aunque en estos experimentos se identificaron los iones que fluyen durante un potencial de acción, no se estableció de qué modo la membrana neuronal puede modificar su permeabilidad iónica para generar el potencial de acción o qué mecanismos desencadenan este cambio crítico. En el capítulo siguiente se encara esta cuestión, con la conclusión sorprendente de que el propio potencial de membrana neuronal afecta la permeabilidad de la membrana. Resumen Las células nerviosas generan señales eléctricas para transmitir información a lo largo de distancias considerables y enviarla a otras células por medio de las conexiones sinápticas. Estas señales finalmente dependen de los cambios en el potencial eléctrico de reposo a través de la membrana neuronal. Se produce un potencial de reposo porque las membranas de las células nervio- SEÑALES ELÉCTRICAS DE LAS CÉLULAS NERVIOSAS 39 RECUADRO 2C Forma y nomenclatura de los potenciales de acción El potencial de acción del axón gigante del calamar tiene una forma de onda característica, con algunas fases diferentes (Fig. A). Durante la fase creciente, el potencial de membrana se despolariza con rapidez. De hecho, los potenciales de acción hacen que el potencial de membrana de despolarice tanto que el potencial de membrana se vuelve transitoriamente positivo respecto del medio externo, lo que produce una fase de exceso (overshoot). La fase de exceso del potencial de acción deja el camino a una fase de caída en la cual el potencial de membrana se repolariza con rapidez. La repolarización lleva el potencial de membrana hasta niveles incluso más negativos que el potencial de membrana de reposo por un período breve; este instante de hiperpolarización se denomina repolarización exagerada (undershoot). Si bien la forma de la onda del potencial de acción del calamar es típica, los detalles de la forma varían ampliamente de una neurona a otra en diferentes animales. En los axones mielínicos de las neuronas motoras de los vertebrados (Fig. B), el potencial de acción es casi indistinguible del correspondiente al axón del calamar. Sin embargo, el potencial de acción registrado en el cuerpo celular de esta misma neurona motora (Fig. C) se ve algo diferente. Por lo tanto, la forma de la onda del potencial de acción puede variar incluso dentro de la misma neurona. Se observan potenciales de acción más complejos en otras neuronas centrales. Por ejemplo, los potenciales de acción registrados desde los cuerpos celulares de las neuronas en la oliva inferior de los mamíferos (una región del tronco del encéfalo que participa en el control motor) duran décimas de milisegundos (Fig. D). Estos potenciales de acción muestran una meseta pronunciada durante su fase de caída, y su repolarización exagerada dura aún más que la de la neurona motora. Uno de los tipos más espectaculares de potenciales de acción se desarrolla en los cuerpos celulares de las neuronas de Purkinje del cerebelo (Fig. E). Estos potenciales tienen varias fases complejas que son resultado de la suma de múltiples potenciales de acción separados. La variedad de las formas de las ondas de los potenciales de acción podría indicar que cada tipo de neurona tiene un mecanismo diferente de producción del potencial de acción. Sin embargo, por fortuna todas estas formas de ondas diversas son resultado de variaciones relativamente menores en el esquema utilizado por el axón gigante del calamar. Por ejemplo, las mesetas en la fase de repolarización son el resultado de la presencia de canales iónicos permeables al Ca2+ y de repolarizaciones exageradas prolongadas resultado de la presencia de tipos extra de canales de K+. El potencial de acción complejo de la célula de Purkinje es consecuencia de estas características extra, a lo que se suma que los diferentes tipos de potenciales de acción se generan en distintas partes de la neurona de Purkinje (cuerpo celular, dendritas y axones) y se suman en los registros del cuerpo celular. Entonces, las lecciones que aprendimos del axón del calamar son aplicables al conocimiento de la generación del potencial de acción en todas las neuronas y en realidad son esenciales para ello. Bibliografía Barrett, E. F. and J. N. Barrett (1976) Separation of two voltage-sensitive potassium currents, and demonstration of a tetrodotoxin-resistant calcium current in frog motoneurones. J. Physiol. (Lond.) 255: 737-774. Dodge, F. A. and B. Frankenhaeuser (1958) Membrane currents in isolated frog nerve fibre under voltage clamp conditions. J. Physiol. (Lond.) 143: 76-90. Hodgkin, A. L. and A. F. Huxley (1939) Action potentials recorded from inside a nerve fibre. Nature 144: 710-711. Llinás, R. and M. Sugimori (1980) Electrophysiological properties of in vitro Purkinje cell dendrites in mammalian cerebellar slices. J. Physiol. (Lond.) 305: 197-213. Llinás, R. and Y. Yarom (1981) Electrophysiology of mammalian inferior olivary neurones in vitro. Different types of voltage-dependent ionic conductances. J. Physiol. (Lond.) 315: 549-567. (A) Fases de un potencial de acción del axón del calamar gigante. (B) Potencial de acción registrado de un axón mielínico de una neurona motora de la rana. (C) Potencial de acción registrado desde el cuerpo celular de la neurona motora de la rana. El potencial de acción es más pequeño y la repolarización exagerada es prolongada en comparación con el potencial de acción registrado desde el axón de esta misma neurona (B). (D) Potencial de acción registrado desde el cuerpo celular de una neurona de Purkinje en el cerebelo de un cobayo (A, de Hodgkin y Huxley, 1939; B, de Dodge y Frankenhauser, 1958; C, de Barrett y Barrett, 1976; D, de Llinás y Yarom, 1981; E, de Llinás y Sugimori, 1980.) (B) (A) (C) (D) (E) Potencial de membrana (mV) 40 0 Fase de elevación Fase de exceso (overshoot) Fase de caída Fase de repolarización exagerada (undershoot) <40 <80 0 2 4 6 8 0 1 2 3 4 0 2 4 Tiempo (ms) 6 8 0 10 20 30 40 0 50 100 150 40 CAPÍTULO 2 sas son permeables a una o más especies de iones sometidos a un gradiente electroquímico. Más específicamente, un potencial de membrana negativo en reposo es resultado de la salida neta de K+ a través de las membranas neuronales que son en mayor medida permeables al K+. Por el contrario, un potencial de acción se desarrolla cuando un aumento transitorio en la permeabilidad al Na+ permite un flujo neto de Na+ en la dirección opuesta a través de la membrana que es ahora predominantemente permeable al Na+. La elevación breve en la permeabilidad de la membrana al Na+ es seguida por una elevación transitoria secundaria en la permeabilidad de la membrana al K+, que repolariza la membrana neuronal y produce una repolarización exagerada breve del potencial de acción. Como resultado de estos procesos, la membrana se despolariza al estilo de todo o nada durante un potencial de acción. Cuando estos cambios activos de la permeabilidad ceden, el potencial de membrana retorna a su nivel de reposo debido a la permeabilidad elevada de la membrana en reposo al K+. Lecturas adicionales Revisiones Hodgkin, A. L. (1951) The ionic basis of electrical activity in nerve and muscle. Biol. Rev. 26: 339-409. Hodgkin, A. L. (1958) The Croonian Lecture: Ionic movements and electrical activity in giant nerve fibres. Proc. R. Soc. Lond. (B) 148: 1-37. Artículos originales importantes Baker, P. F., A. L. Hodgkin and T. I. Shaw (1962) Replacement of the axoplasm of giant nerve fibres with artificial solutions. J. Physiol. (London) 164: 330354. Cole, K. S. and H. J. Curtis (1939) Electric impedence of the squid giant axon during activity. J. Gen. Physiol. 22: 649-670. Goldman, D. E. (1943) Potential, impedence, and rectification in membranes. J. Gen. Physiol. 27: 37-60. Hodgkin, A. L. and P. Horowicz (1959) The influence of potassium and chloride ions on the membrane potential of single muscle fibres. J. Physiol. (London) 148: 127-160. Hodgkin, A. L. and B. Katz (1949) The effect of sodium ions on the electrical activity of the giant axon of the squid. J. Physiol. (London) 108: 37-77. Hodgkin, A. L. and R. D. Keynes (1953) The mobility and diffusion coefficient of potassium in giant axons from Sepia. J. Physiol. (London) 119: 513-528. Hodgkin, A. L. and W. A. H. Rushton (1938) The electrical constants of a crustacean nerve fibre. Proc. R. Soc. Lond. 133: 444-479. Keynes, R. D. (1951) The ionic movements during nervous activity. J. Physiol. (London) 114: 119-150. Libros Hodgkin, A. L. (1967) The Conduction of the Nervous Impulse. Springfield, IL: Charles C. Thomas. Hodgkin, A. L. (1992) Chance and Design. Cambridge: Cambridge University Press. Junge, D. (1992) Nerve and Muscle Excitation, 3rd Ed. Sunderland, MA: Sinauer Associates. Katz, B. (1966) Nerve, Muscle, and Synapse. New York: McGraw-Hill. 3 Permeabilidad de la membrana dePendiente de voltaje Aspectos generales EL POTENCIAL DE ACCIÓN ES LA SEÑAL ELÉCTRICA primaria generada por las células nerviosas y se origina por cambios en la permeabilidad de membrana a iones específicos. El conocimiento actual de estos cambios en la permeabilidad iónica se basa en las pruebas obtenidas por la técnica de pinzamiento de voltaje, que permiten una caracterización detallada de los cambios de permeabilidad en función del potencial de membrana y del tiempo. En la mayoría de los tipos de axones, estos cambios implican una elevación rápida y transitoria en la permeabilidad al sodio (Na+), seguida por una elevación más lenta pero más sostenida en la permeabilidad al potasio (K+). Ambas permeabilidades son dependientes del voltaje, y aumentan a medida que el potencial de membrana se despolariza. La cinética y la dependencia del voltaje de las permeabilidades al Na+ y al K+ brindan una explicación completa de la generación del potencial de acción. La despolarización del potencial de membrana hasta el nivel umbral produce un aumento rápido y autosostenido en la permeabilidad al Na+ que produce la fase de elevación del potencial de acción; sin embargo, el aumento de la permeabilidad al Na+ es breve y es seguido por un incremento más lento de la permeabilidad al K+ que restablece el potencial de membrana hasta su nivel de reposo negativo habitual. Un modelo matemático que describe el comportamiento de estas permeabilidades iónicas predice casi todas las propiedades observadas de los potenciales de acción. Es importante destacar que este mismo mecanismo iónico permite la propagación de los potenciales de acción a lo largo de los axones neuronales, lo que explica cómo se transmiten las señales eléctricas en todo el sistema nervioso. Corrientes iónicas a través de las membranas de las células nerviosas El capítulo anterior introdujo la idea de que las células nerviosas generan señales eléctricas en virtud de una membrana que es diferencialmente permeable a distintas especies de iones. En particular, un incremento transitorio en la permeabilidad de la membrana neuronal al Na+ inicia el potencial de acción. En este capítulo se considera con exactitud cómo se desarrolla este aumento en la permeabilidad al Na+. Una clave para comprender este fenómeno es la observación de que los potenciales de acción se inician solo cuando el potencial de membrana neuronal se vuelve más positivo que cierto nivel umbral. Esta observación sugiere que el mecanismo responsable del 42 CAPÍTULO 3 incremento en la permeabilidad al Na+ es sensible al potencial de membrana. Por lo tanto, si pudiera comprenderse cómo un cambio en el potencial de membrana activa la permeabilidad al Na+, debería ser posible explicar cómo se generan los potenciales de acción. El hecho de que la permeabilidad al Na+ que genera el cambio en el potencial de membrana es en sí misma sensible al potencial de membrana presenta obstáculos tanto conceptuales como prácticos para estudiar el mecanismo del potencial de acción. Un problema práctico es la dificultad para variar en forma sistemática el potencial de membrana para estudiar el cambio de permeabilidad, porque estas alteraciones en el potencial de membrana producen el potencial de acción, que genera cambios nuevos no controlados. Históricamente, no fue posible comprender entonces los potenciales de acción hasta que se desarrolló una técnica que permitió que los investigadores controlen el potencial de membrana y midan de manera simultánea los cambios de permeabilidad subyacentes. Esta técnica, el método de pinzamiento de voltaje (Recuadro 3A), brinda toda la información necesaria para definir la permeabilidad iónica de la membrana en cualquier nivel del potencial de membrana. A fines de la década de 1940, Alan Hodgkin y Andrew Huxley utilizaron la técnica de pinzamiento de voltaje para investigar los cambios de permeabilidad subyacentes al potencial de acción. Una vez más, estos autores decidieron utilizar la neurona gigante del RECUADRO 3A Método de pinzamiento de voltaje Los descubrimientos en la investigación científica a menudo se basan en el desarrollo de tecnologías nuevas. En el caso del potencial de acción, solo se obtuvo un conocimiento detallado después de que Kenneth Cole, en la década de 1940, inventó la técnica de pinzamiento de voltaje. Este dispositivo se denomina así porque controla o pinza el potencial de membrana (o voltaje) en cualquier nivel que el experimentador desee. El método mide el potencial de membrana con un microelectrodo (u otro tipo de electrodo) colocado en el interior de la célula (1), y compara electrónicamente este voltaje con el que se desea mantener (denominado voltaje comando) (2). A continuación, el circuito de pinzamiento pasa una corriente retrógrada a la célula a través de otro electrodo in- tracelular (3). Este circuito electrónico de retroalimentación mantiene el potencial de membrana en el nivel deseado, aun frente a cambios en la permeabilidad que en condiciones normales alterarían el potencial de membrana (como los generados durante el potencial de acción). Lo que es más importante, el dispositivo permite la medición simultánea de la corriente necesaria para mantener la célula en un voltaje dado (4) . Esta corriente es exactamente igual a la cantidad de corriente que fluye a través de la membrana neuronal, lo que permite la medición directa de estas corrientes de membrana. Por lo tanto, la técnica de pinzamiento de voltaje puede indicar cómo fluye el potencial de membrana en el flujo de corrientes iónicas a través de la membrana. Esta información brindó a Hodgkin 2 El amplificador de pinzamiento 1 Un electrodo interno mide el potencial de membrana (Vm) y está conectado al amplificador de pinzamiento de voltaje. de voltaje compara el potencial de membrana con el potencial deseado (comando). Medidor Vm Electrodo de referencia Voltaje comando Electrodo de registro Bibliografía Cole, K. S. (1968) Membranes, Ions and Impulses: A Chapter of Classical Biophysics. Berkeley, CA: University of California Press. es diferente del potencial 3 Cuando el Vm comando, el amplificador de pinzamiento inyecta corriente en el axón a través de un segundo electrodo. Este sistema de retroalimentación hace que el potencial de membrana se vuelva igual al potencial comando. Amplificador de pinzamiento de voltaje 4 La corriente fluye Medidor de corriente Solución salina Axón del calamar y a Huxley los conocimientos clave que condujeron a su modelo para la generar el potencial de acción. En la actualidad, el método del pinzamiento de voltaje se utiliza ampliamente para estudiar las corrientes iónicas en las neuronas y otras células. La versión contemporánea más popular de este enfoque es la técnica de pinzamiento en parche, método que puede aplicarse casi a cualquier célula y tiene una resolución suficientemente alta como para medir corrientes eléctricas diminutas que fluyen a través de canales iónicos aislados (véase Recuadro 4A). Electrodo de transmisión de corriente hacia atrás en el axón, y por lo tanto a través de su membrana puede medirse aquí. Técnica de pinzamiento de voltaje para estudiar las corrientes de membrana del axón de un calamar. PERMEABILIDAD DE LA MEMBRANA DEPENDIENTE DE VOLTAJE (B) Potencial de membrana (mV) (A) Corriente de membrana (mA/cm2) 43 0 0 –65 –65 Despolarización de 65 mV Hiperpolarización de 65 mV –130 –130 +1 +1 Exterior Exterior 0 Interior Corriente de capacitancia 0 Interior Corriente de capacitancia –1 Corriente transitoria hacia adentro Corriente tardía hacia afuera –1 0 1 2 Tiempo (ms) 3 4 calamar, dado que su gran tamaño (hasta 1 mm de diámetro; véase Recuadro 2B) permitía la inserción de los electrodos necesarios para el pinzamiento de voltaje. Estos investigadores fueron los primeros en evaluar directamente la hipótesis de que los cambios de permeabilidad al Na+ y al K+ sensibles al potencial son tanto necesarios como suficientes para la producción de potenciales de acción. El primer objetivo de Hodgkin y Huxley fue determinar si, de hecho, la membrana neuronal tiene permeabilidad dependiente de voltaje. Para encarar esta cuestión, estos investigadores se preguntaron si las corrientes iónicas fluyen a través de la membrana cuando su potencial se modifica. En la Figura 3.1 se muestra el resultado de uno de estos experimentos. En la Figura 3.1A se exhiben las corrientes producidas por el axón de un calamar cuando su potencial de membrana, Vm, se hiperpolariza desde el nivel de reposo de −65 mV a −130 mV. La respuesta inicial del axón es resultado de la redistribución de la carga a través de la membrana axónica. Esta corriente de capacitancia es casi instantánea, y termina dentro de una fracción de un milisegundo. Además de este acontecimiento breve, fluye muy poca corriente cuando la membrana está hiperpolarizada. Sin embargo, si el potencial de membrana se despolariza desde −65 hasta 0 mV, la respuesta es muy diferente (Figura 3.1B). Tras la corriente de capacitancia, el axón produce una corriente iónica hacia el interior que crece rápidamente (hacia el interior se refiere a una carga positiva que ingresa en la célula, o sea, cationes dentro o aniones fuera), lo que deja el paso a una corriente tardía hacia el exterior y de crecimiento más lento. La generación de estas corrientes iónicas por la despolarización de la membrana establece que la permeabilidad de la membrana de los axones es, en efecto, dependiente de voltaje. Dos tipos de corriente iónica dependientes de voltaje Los resultados que aparecen en la Figura 3.1 demuestran que la permeabilidad iónica de la membrana neuronal es sensible al voltaje, pero los experimentos no identifican cuántos tipos de permeabilidad hay o cuáles son los iones involucrados. Como se explicó en el Capítulo 2 (véase Figura 2.6), la variación del poten- 0 1 2 Tiempo (ms) 3 4 FIGURA 3.1 Flujo de corriente a través de la membrana del axón de un calamar durante un experimento de pinzamiento de voltaje. (A) Una hiperpolarización de 65 mV del potencial de membrana produce solo una corriente de capacitancia muy breve. (B) Una despolarización de 65 mV del potencial de membrana también produce una corriente de capacitancia breve, pero le sigue una frase más prolongada, aunque transitoria, de corriente hacia dentro y una corriente tardía pero sostenida hacia fuera. (De Hodgkin y cols., 1952.) cial a través de una membrana hace posible deducir el potencial de equilibrio para los flujos iónicos a través de la membrana y, por lo tanto, identificar los iones que fluyen. Dado que el método de pinzamiento de voltaje permite modificar el potencial de membrana mientras se miden las corrientes iónicas, para Hodgkin y Huxley era un asunto fácil determinar la permeabilidad iónica mediante el examen de cómo se modificaban las propiedades de las corrientes temprana hacia el interior y tardía hacia el exterior a medida que variaba el potencial de membrana (Figura 3.2). Como ya se señaló, no fluye corriente iónica apreciable alguna en los potenciales de membrana más con valores negativos que los del potencial de reposo. Sin embargo, con potenciales más positivos, las corrientes no solo fluyen sino que cambian de magnitud.La corriente temprana tiene una dependencia con forma de U del potencial de membrana, y aumenta en un intervalo de despolarizaciones hasta alrededor de 0 mV, pero disminuyen a medida que el potencial se despolariza más. Por el contrario, la corriente tardía se incrementa de manera uniforme con los potenciales de membrana crecientemente positivos. Estas respuestas diferentes al potencial de membrana pueden observarse con más claridad cuando las magnitudes de los dos componentes se representan en correlación con el potencial de membrana, como en la Figura 3.3. La sensibilidad al voltaje de la corriente temprana hacia el interior aporta un indicio importante acerca de la naturaleza de los iones que transmiten la corriente, esto es, que no fluye corriente cuando el potencial de membrana se pinza en +52 mV. En las neuronas del calamar, que estudiaron Hodgkin y Huxley, la concentración externa de Na+ es 440 mM y la concentración interna de Na+ es 50 mM. Para este gradiente de concentración, la ecua- Potencial de membrana (mV) 44 CAPÍTULO 3 +65 75 50 +52 +26 25 0 0 –26 –25 –50 Corriente de membrana (mA/cm2) 6 4 2 0 –2 0 2 4 6 8 0 2 4 6 8 0 FIGURA 3.2 Corrientes producidas por despolarizaciones de membrana a varios potenciales diferentes. La corriente temprana al principio aumenta, luego su magnitud disminuye a medida que se incrementa la despolarización; obsérvese que esta corriente se invierte en polaridad con potenciales más positivos que alrededor de +55 mV. La corriente más tardía aumenta de forma monótona con la despolarización creciente. (De Hodgkin y cols., 1952.) ción de Nernst predice que el potencial de equilibrio para el Na+ debe ser +55 mV. Recuérdese también del Capítulo 2 que en el potencial de equilibrio del Na+ no hay flujo neto de Na+ a través de la membrana, aun cuando la membrana sea muy permeable al Na+. Por lo tanto, la observación experimental de que no fluye corriente en el potencial de membrana donde no puede fluir Na+ es una indicación firme de que la corriente temprana hacia el interior es transportada por el ingreso de Na+ en el axón. Una manera incluso más firme de evaluar si el Na+ transmite la corriente temprana hacia el interior es examinar el comporta- Corriente de membrana (mA/cm2) 3,0 Tardía 2,0 1,0 0 Temprana –100 –50 50 0 Potencial de membrana (mV) 2 4 6 Tiempo (ms) 8 0 2 4 6 8 0 2 4 6 8 miento de esta corriente después de eliminar el Na+ externo. La eliminación del Na+ en el exterior del axón convierte en negativo el ENa; si en estas condiciones se aumenta la permeabilidad al Na+, la corriente debe fluir hacia afuera a medida que el Na+ abandona la neurona, debido al gradiente electroquímico invertido. Cuando Hodgkin y Huxley realizaron este experimento, observaron que la eliminación del Na+ exterior ocasionó que la corriente temprana hacia el interior invirtiera su polaridad y se convirtiera en una corriente hacia el exterior con un potencial de membrana que dio origen a una corriente hacia el interior cuando había Na+ en el exterior (Figura 3.4). Este resultado demuestra convincentemente que la corriente temprana hacia el interior medida cuando está presente el Na+ en el medio externo debe ser consecuencia del ingreso de Na+ hacia la neurona. En el experimento que se muestra en la Figura 3.4, la eliminación del Na+ en el exterior tiene poco efecto sobre la corriente hacia el exterior que fluye después de que la neurona se mantuvo en un voltaje de membrana despolarizado durante varios milisegundos. Este nuevo resultado muestra que la corriente tardía hacia el exterior se debe al flujo de un ion distinto de Na+. Varias evidencias presentadas por Hodgkin, Huxley y cols. mostraron que esta corriente hacia el exterior es causada por el K+ que sale de la neurona. Tal vez la demostración más firme de la participación del K+ es que la cantidad del eflujo de K+ desde la neurona, medida por la carga de la neurona con K+ radiactivo, se correlaciona íntimamente con la magnitud de la corriente tardía hacia el exterior. Tomados en conjunto, estos experimentos muestran que la modificación del potencial de membrana hasta un nivel más positivo que el potencial de reposo produce dos efectos: un influjo temprano de Na+ hacia la neurona, seguido por un eflujo tar- FIGURA 3.3 Relación entre la amplitud de corriente y el potencial de membrana. Experimentos como el que se muestra en la Figura 3.2 indican que la corriente tardía hacia el exterior aumenta abruptamente con la despolarización creciente, mientras que la corriente temprana hacia el interior primero aumenta de magnitud pero luego disminuye y se invierte a una corriente hacia el exterior en unos +55 mV (potencial de equilibrio para el sodio). (De Hodgkin y cols., 1952.) PERMEABILIDAD DE LA MEMBRANA DEPENDIENTE DE VOLTAJE FIGURA 3.4 Dependencia del sodio de la corriente temprana hacia dentro. (A) En presencia de concentraciones externas normales de Na+, la despolarización del axón de un calamar 25 Potencial de membrana (mV) 45 0 hasta 0 mV (parte superior) produce una corriente inicial hacia el interior. (B) La eliminación del Na+ externo hace que la corriente inicial hacia dentro se vuelva hacia fuera, efecto que se revierte (C) por el restablecimiento del Na+ externo. (De Hodgkin y Huxley, 1952a). –25 –50 –75 de vías de permeabilidad independientes. Como se explica en el Capítulo 4, en la actualidad se sabe que estas vías son canales iónicos selectivamente permeables a Na+ o K+. De hecho, la tetrodotoxina, el tetraetilamonio y otros fármacos que interactúan con tipos específicos de canales iónicos fueron herramientas de utilidad extraordinaria para caracterizar estas moléculas de los canales (véase Capítulo 4). (A) +1 460 mM Na+ 0 Corriente temprana hacia el interior 2 Corriente de membrana (mA/cm ) –1 (B) Dos conductancias de membrana dependientes de voltaje +1 Libre de Na+ El objetivo siguiente que Hodgkin y Huxley se propusieron fue describir matemáticamente los cambios de permeabilidad al Na+ y al K+. Para hacerlo, aceptaron que las corrientes iónicas se deben a un cambio en la conductancia de la membrana, definida como la recíproca de la resistencia de la membrana. Por lo tanto, la conductancia de la membrana se relaciona en gran medida con su permeabilidad, aunque no es idéntica a ella. Cuando se evalúan los movimientos iónicos desde un punto de vista eléctrico, es conveniente describirlos en términos de conductancias iónicas y no de las permeabilidades iónicas. Para los fines actuales, permeabilidad y conductancia pueden considerarse sinónimos. Si la conductancia (g) de la membrana obedece la 0 La corriente temprana es hacia el exterior –1 (C) +1 460 mM Na+ 0 La corriente temprana es nuevamente hacia el interior 2 6 4 Tiempo (ms) 8 dío de K . El influjo temprano de Na produce una corriente transitoria hacia el interior, mientras que el eflujo tardío de K+ ocasiona una corriente sostenida hacia el exterior. Las diferencias en el curso temporal y la selectividad iónica de los dos flujos sugieren que dos mecanismos diferentes de permeabilidad iónica se activan por cambios en el potencial de membrana. La confirmación de que en realidad existen dos mecanismos distintos proviene de estudios farmacológicos de agentes que afectan específicamente estas dos corrientes (Figura 3.5). La tetrodotoxina, una neurotoxina alcaloide hallada en algunos peces globo, ranas tropicales y salamandras, bloquea la corriente de Na+ sin afectar la corriente de K+. Por el contrario, los iones de tetraetilamonio bloquean las corrientes de K+ sin afectar las de Na+. La sensibilidad diferencial de las corrientes de Na+ y K+ proporciona evidencia adicional firme de que el Na+ y el K+ fluyen a través + + Potencial de membrana (mV) 0 Corriente de membrana (mA/cm2 ) –1 25 0 -25 -50 -75 +1 (2) Corriente de Na+ bloqueada Agregar tetrodotoxina (1) 0 0 Agregar tetraetilamonio -1 0 5 Tiempo (ms) 5 10 Corriente de K+ bloqueada (3) 10 0 5 Tiempo (ms) 10 FIGURA 3.5 Separación farmacológica de las corrientes de Na+ y de K+ en los componentes sodio y potasio. En el panel (1) se observa la corriente que fluye cuando el potencial de membrana del axón de un calamar se despolariza hasta 0 mV en condiciones de control. (2) El tratamiento con tetrodotoxina hace que desaparezcan las corrientes tempranas de Na+ pero respeta las corrientes tardías de K+. (3) El tratamiento de tetraetilamonio bloquea las corrientes de K+ sin afectar las corrientes de Na+. (De Moore y cols., 1967, y Armstrong y Binstock, 1965.) 46 CAPÍTULO 3 ley de Ohm (que establece que el voltaje es igual al producto de corriente por resistencia), entonces la corriente iónica que fluye durante un aumento de la conductancia de la membrana está dada por: Iion = gion (Vm – Eion) donde Iion es la corriente iónica, Vm es el potencial de membrana y Eion es el potencial de equilibrio para el ion que fluye a través de la conductancia, gion. La diferencia entre Vm y Eion es la fuerza impulsora electroquímica sobre el ion. Hodgkin y Huxley utilizaron esta relación sencilla para calcular la dependencia de las conductancias de Na+ y K+ sobre el tiempo y el potencial de membrana. Estos autores conocían Vm, que determinaron a partir de su dispositivo de pinzamiento de voltaje (Figura 3.6A), y pudieron determinar ENa y EK a partir de las concentraciones iónicas a ambos lados de la membrana axónica (véase Cuadro 2.1). Las corrientes transmitidas por Na+ y K+ (INa e IK) pudieron determinarse por separado a partir de los registros de las corrientes de membrana resultantes de la despolarización (Figura 3.6B) midiendo las diferencias entre las corrientes registradas en presencia y ausencia de Na+ externo (como se muestra en la Figura 3.4). A partir de estas medicio- nes, Hodgkin y Huxley pudieron calcular gNa y gK (Figura 3.6C, D), de las cuales extrajeron dos conclusiones fundamentales. La primera es que las conductancias al Na+ y al K+ cambian con el tiempo. Por ejemplo, tanto la conductancia al Na+ como al K+ necesitan cierto tiempo para activarse o encenderse. En particular, la conductancia al K+ tiene un retardo pronunciado, que necesita varios milisegundos para alcanzar su nivel máximo (Figura 3.6D), mientras que la conductancia al Na+ alcanza su nivel máximo más rápidamente (Figura 3.6C). La activación más rápida de la conductancia al Na+ permite que la corriente resultante de Na+ hacia el interior preceda a la corriente de K+ hacia el exterior retardada (Figura 3.6B). Si bien la conductancia al Na+ FIGURA 3.6 Los cambios en la conductancia de membrana subyacentes al potencial de acción son dependientes de tiempo y de voltaje. Las despolarizaciones hasta distintos potencial de membrana (A) producen diferentes corrientes de membrana (B). Debajo se muestran las conductancias al Na+ (C) y al K+ (D) calculadas a partir de estas corrientes. Tanto la conductancia pico al Na+ como la conductancia en estado de equilibrio al K+ aumentan a medida que el potencial de membrana se vuelve más positivo. Además, la activación de ambas conductancias, así como la velocidad de inactivación de la conductancia al Na+, se producen más rápidamente con despolarizaciones más grandes. (De Hodgkin y Huxley, 1952b.) Conductancia al Na+ (mSiemens/cm2) Corriente de membrana (mA/cm2) Potencial de membrana (mV) (A) 50 25 0 –25 –50 –75 –2 –27 –39 (B) 6 4 2 0 –2 30 (C) 20 10 0 60 Conductancia al K+ (mSiemens/cm2) + 44 + 23 (D) 40 20 0 0 2 4 6 8 0 2 4 6 8 0 2 4 6 Tiempo (ms) 8 0 2 4 6 8 0 2 4 6 8 PERMEABILIDAD DE LA MEMBRANA DEPENDIENTE DE VOLTAJE FIGURA 3.7 La despolarización aumenta las conductancias al Na+ y al K+ del axón gigan- 40 Na+ 15 10 5 0 –80 –60 –40 –20 0 20 40 Potencial de membrana (mV) Conductancia (mSiemens/cm2 ) Conductancia (mSiemens/cm2 ) 20 47 K+ 30 20 te del calamar. La magnitud pico de la conductancia al Na+ y el valor en estado de equilibrio de la conductancia al K+ aumentan abruptamente a medida que el potencial de membrana se despolariza. (De Hodgkin y Huxley, 1952b). 10 0 –80 20 40 –60 –40 –20 0 Potencial de membrana (mV) se eleva con rapidez, declina con igual celeridad, aun cuando el potencial de membrana se mantenga en un nivel despolarizado. Esto muestra que la despolarización no solo hace que se active la conductancia al Na+, sino también que disminuya con el tiempo o se inactive. La conductancia al K+ del axón del calamar no se inactiva de esta forma; por lo tanto, aunque las conductancias al Na+ y al K+ comparten la propiedad de activación dependiente del tiempo, sólo la conductancia al Na+ se inactiva. (A partir de este hecho, se descubrió la inactivación de las conductancias al K+ en otros tipos de células nerviosas; véase Cap. 4). Los cursos temporales de las conductancias al Na+ y al K+ son dependientes de voltaje, y la velocidad tanto de la activación como de la inactivación aumenta con los potenciales más despolarizados. Este hallazgo explica los cursos más rápidos de las corrientes de las membranas medidos en potenciales más despolarizados. La segunda conclusión derivada de los cálculos de Hodgkin y Huxley es que la conductancia tanto al Na+ como al K+ es dependiente del K+, o sea que ambas conductancias aumentan progresivamente a medida que la neurona se despolariza. En la Figura 3.7 se muestra esto mediante el gráfico de la relación entre el valor pico de las conductancias (tomado de Figura 3.6C, D) y el potencial de membrana. Obsérvese la dependencia similar del voltaje para cada conductancia; ambas conductancias son muy pequeñas en los potenciales negativos, máximas en los potenciales muy positivos y sensiblemente dependientes del voltaje de la membrana en los potenciales intermedios. La observación de que estas conductancias son sensibles a los cambios del potencial de membrana muestra que el mecanismo subyacente a las conductancias “detecta” el voltaje a través de la membrana. De acuerdo con todo esto, los experimentos de pinzamiento de voltaje llevados a cabo por Hodgkin y Huxley mostraron que las corrientes iónicas que fluyen cuando la membrana neuronal se despolariza se deben a tres procesos diferentes sensibles al voltaje: 1) activación de la conductancia al Na+, 2) activación de la conductancia al K+ y 3) inactivación de la conductancia al Na+. Reconstrucción del potencial de acción A partir de sus observaciones experimentales, Hodgkin y Huxley pudieron construir un modelo matemático detallado de los cambios en las conductancias al Na+ y al K+. El objetivo de estos esfuerzos de modelado es determinar si los cambios al Na+ y al K+ aislados eran suficientes para producir un potencial de acción. Con esta información, de hecho los autores pudieron generar la forma y el curso temporal del potencial de acción con una precisión notable (Figura 3.8A). El modelo de Hodgkin-Huxley pudo simular muchas otras características del comportamiento del potencial de acción en el axón del calamar. Por ejemplo, es bien sabido que después de un potencial de acción, el axón se vuelve refractario a una mayor excitación durante un breve período, denominado período refractario (Figura 3.8B). El modelo fue capaz de imitar estrechamente este comportamiento (Figura 3.8C). El modelo de Hodgkin y Huxley también proporcionó muchas ideas sobre el modo de generación del potencial de acción. La Figura 3.8A muestra un potencial de acción reconstruido, junto con los cursos temporales de las conductancias al Na+ y al K+ subyacentes. La coincidencia del aumento inicial en la conductancia al Na+ con la fase creciente rápida del potencial de acción demuestra que un incremento selectivo en la conductancia al Na+ es responsable de la iniciación del potencial de acción. El aumento de la conductancia al Na+ hace que éste ingrese en la neurona y despolarice así el potencial de membrana, que se aproxima a ENa. Con posterioridad, la velocidad de despolarización cae porque la fuerza impulsora electroquímica sobre el Na+ disminuye y la conductancia al Na+ se inactiva. Al mismo tiempo, la despolarización activa con lentitud la conductancia al K+ dependiente de voltaje y hace que el K+ abandone la célula y repolarice el potencial de membrana hacia EK. Dado que la conductancia al K+ es transitoriamente mayor que en la condición de reposo, el potencial de membrana se vuelve brevemente más negativo que el potencial de reposo normal (repolarización exagerada). La hiperpolarización del potencial de membrana hace que la conductancia al K+ dependiente de voltaje (y cualquier conductancia al Na+ no inactivada) se inactive, lo que permite que el potencial de membrana retorne a su nivel de reposo. El curso temporal relativamente lento de la inactivación de la conductancia al K+, así como la persistencia de la inactivación de la conductancia al Na+, es responsable del período refractario (véase también Figura 3.10). Este mecanismo de generación del potencial de acción representa un circuito de retroalimentación positiva: la activación de la conductancia al Na+ dependiente de voltaje aumenta el ingreso del Na+ en la neurona, lo que hace despolarizar el potencial de CAPÍTULO 3 35 15 0 –15 –50 –65 –20 –40 –60 Potencial de membrana (mV) Conductancia 2 (mSiemens/cm ) –80 30 Na+ 20 10 K+ 0 0 1 2 3 Tiempo (ms) mulando un axón con dos pulsos de corriente que están separadas por intervalos variables. Aunque el primer estímulo evoca de manera fiable un potencial de acción, durante el período refractario el segundo estímulo solamente generará un pequeño potencial de acción o ninguna respuesta en absoluto. (C) El modelo matemático predice con exactitud Amplitud del potencial de acción tencial de acción. (A) Simulación de un potencial mentos de pinzamiento de voltaje. (B) Puede observarse el período refractario esti- 0s 9 ms –15 –50 –65 FIGURA 3.8 Simulación matemática del pode acción (curva negra) junto con los cambios subyacentes en la conductancia al Na+ (curva roja) y al K+ (curva dorada). El tamaño y el curso temporal del potencial de acción se calcularon utilizando solo las propiedades de g(Na) y gK medidas en experi- 6 ms –15 –50 –65 0s 0,8 0,6 0,4 0,2 4 6 8 10 Intervalo entre los estímulos (ms) respuestas del axón durante el período refractario. (De Hodgkin y Huxley, 1952d.) membrana y conduce aún más a la activación de la conductancia al Na+, a un ingreso mayor de Na+ y a la despolarización incluso mayor (Figura 3.9). La retroalimentación positiva no desaparece hasta que la inactivación de la conductancia al Na+ y la activación de la conductancia al K+ restablecen el potencial de membrana hasta el nivel de reposo. Debido a que este circuito de retroalimentación positiva, una vez iniciado, es sostenido por las propiedades intrínsecas de la neurona –a saber, las depenFIGURA 3.9 Ciclos de retroalimentación responsables de los cambios del potencial de membrana durante un potencial de acción. La despolarización de la membrana activa con rapidez un ciclo de retroalimentación positiva impulsado por la activación dependiente de voltaje de la conductancia al Na+. Este fenómeno es seguido por la activación más lenta de un circuito de retroalimentación negativa a medida que la despolarización activa una conductancia al K+, que ayuda a repolarizar el potencial de membrana y terminar el potencial de acción. –65 35 15 0 –15 –50 –65 35 15 0 –15 –50 –65 1,0 0,0 90 0 5 ms –15 –50 0s 35 15 0 Modelo matemático basado en las conductancias al Na+ y al K+ 35 15 0 4,5 ms 35 15 0 4 Corriente estímulo 0 90 0 Potencial de membrana (mV) +20 Amplitud del potencial de acción Potencial de membrana (mV) +40 (C) Potenciales de acción del axón de calamar 0s 6 ms 0s 8 ms 0s 1,0 0,8 0,6 0,4 0,2 0,0 4 6 8 10 Intervalo entre los estímulos (ms) Aumento de la corriente de Na+ po lar iza des Se Corriente estímulo (B) Ciclo positivo rápido Canales de Na+ abiertos Despolariza el potencial de membrana lari za (A) Se hi pe rpo 48 Ciclo negativo lento Aumento de la corriente de K+ Canales de K+ abiertos PERMEABILIDAD DE LA MEMBRANA DEPENDIENTE DE VOLTAJE 49 RECUADRO 3B Umbral Una propiedad importante, posiblemente asombrosa, del potencial de acción es su iniciación apartir de un potencial particular de la membrana, denominado umbral. En efecto, los potenciales de acción nunca se desarrollan sin un estímulo despolarizante que lleve a la membrana a este nivel. El “gatillo” despolarizante puede ser uno de los siguientes acontecimientos: una aferencia sináptica, un potencial de receptor generado por órganos receptores especializados, la actividad de los marcapasos endógenos de las células que generan potenciales de acción en forma espontánea o la corriente local que media la propagación del potencial de acción a través del axón. (A) Escapa algo de calor Las reacciones exotérmicas producen calor adicional Fuente de calor (B) Es posible comprender por qué el potencial de acción “despega” en un nivel particular de despolarización al comparar los hechos subyacentes con una explosión química (Figura A). El calor exógeno (análogo a la despolarización inicial del potencial de membrana) estimula una reacción química exotérmica, que aumenta más la reacción (Figura B). Como resultado de este circuito de retroalimentación positiva, la velocidad de la reacción crece de manera exponencial: la definición de una explosión. Sin embargo, en cualquiera de estos dos procesos hay un umbral, o sea, un punto hasta el cual se puede aportar calor sin que se produzca una explosión. El umbral para la explosión química representada aquí es el punto en el cual la cantidad de calor aportado en forma exógena es solo igual a la cantidad de calor que puede disiparse por las circunstancias de la reacción (como el escape de calor de la jarra). En principio, el umbral de iniciación del potencial de acción es similar (Figura C). Hay una despolarización “subumbral”, donde el ritmo de aumento de ingreso de sodio es menor que el ritmo de salida de potasio (recuérdese que la membrana en reposo es altamente permeable al K+, que por lo tanto fluye hacia fuera a medida que la membrana se despolariza). El punto en el cual el influjo de Na+ iguala el eflujo de (C) Aumento en la velocidad de la reacción K+ representa un equilibrio inestable análogo al punto de la ignición de una mezcla explosiva. El comportamiento de la membrana en el umbral refleja esta inestabilidad: el potencial de membrana puede continuar en el nivel umbral durante un período variable antes de que retorne al nivel de reposo o crezca hasta un potencial de acción completo. Al menos en teoría, si hay ganancia interna neta de un solo ion sodio, se desarrolla un potencial de acción; por el contrario, la pérdida neta de un solo ion potasio conduce a la despolarización. Por lo tanto, una definición más precisa de umbral es ese valor del potencial de membrana en el que la corriente transmitida por el Na+ que ingresa en la neurona es exactamente igual a la corriente de K+ que fluye hacia fuera. Una vez que el acontecimiento gatillo despolariza la membrana más allá de este punto, el circuito de retroalimentación positiva de la entrada de Na+ sobre el potencial de membrana se cierra y el potencial de acción “se dispara”. Como las conductancias al Na+ y al + K cambian dinámicamente con el tiempo, el potencial umbral para producir un potencial de acción también varía como consecuencia de la actividad previa de la neurona. Por ejemplo, después de un potencial de acción, la membrana se vuelve transitoriamente refractaria a una mayor excitación, porque el umbral para disparar un potencial de acción se eleva de manera transitoria. Por lo tanto, no hay un valor específico del potencial de membrana que defina el umbral para una célula nerviosa dada en todas las circunstancias. Entrada de Na+ Reacción exotérmica Explosión química Aumento de la permeabilidad al Na+ Potencial de acción Despolarización de la membrana Calor El escape de calor hace más lenta la reacción La pérdida de K+ repolariza el potencial de membrana El circuito de retroalimentación positiva subyacente al potencial de acción explica el fenómeno de umbral. 50 CAPÍTULO 3 + 1 Los canales del Na se abren Estimular localmente en respuesta al estímulo, y generan un potencial de acción aquí. 2 Algo de corriente despolarizante fluye pasivamente por el axón. Na+ Membrana Canal del Na+ Canal del K+ t=1 Axón Na+ Punto A Punto B Punto C local hace que los canales 3 La despolarización + vecinos se abran y genera un del Na potencial de acción aquí. K+ Na+ t=2 K+ Na+ Punto B Punto A Punto C + corriente arriba se inactivan, 4 Los canales del Na + mientras que los canales del K se abren. El potencial de membrana se repolariza y el axón es refractario aquí. 5 El proceso se repite y propaga el potencial de acción a lo largo del axón. K+ Na+ t=3 Punto A t=1 t=2 Punto B Punto C Na+ t=3 0 mV Punto A K+ Umbral –65 0 Punto B FIGURA 3.10 La conducción del potencial de acción requiere un flujo tanto activo como pasivo de corriente. La despolarización abre los canales del Na+ localmente y produce un potencial de acción en el punto A del axón (tiempo t = 1). La corriente resultante hacia el interior fluye pasivamente a lo largo del axón y despolariza la región adyacente (punto B) del axón. En un momento posterior (t = 2), la despolarización de la membrana adyacente ha abierto los canales de Na+ en el punto B, lo que provoca la iniciación del potencial de acción en este sitio y una corriente adicional hacia el interior que otra vez se propaga en forma pasiva –65 hasta un punto adyacente (punto C) más alejado a lo largo del axón. En un tiempo posterior (t = 3), el potencial de acción se propaga incluso más lejos. Este ciclo continúa a lo largo de 0 todo el axón. Obsérvese que a medida que el potencial de acción se propaga, el potencial de membrana se repolariza a causa de la apertura de los canales de K+ y a la inactivación de los Punto C –65 canales de Na+, lo cual deja una “estela” de refractariedad por detrás del potencial de acción que impide su propagación retrógrada (panel 4). El panel final muestra el curso temporal de los cambios en el potencial de membrana en los puntos indicados. PERMEABILIDAD DE LA MEMBRANA DEPENDIENTE DE VOLTAJE dencias de voltaje de las conductancias iónicas– el potencial de acción es autosostenido o regenerativo. Esta cualidad regenerativa explica por qué los potenciales de acción muestran un comportamiento todo o nada (véase Figura 2.1) y por qué tienen un umbral (Recuadro 3B). La activación retardada de la conductancia al K+ representa un circuito de retroalimentación negativo que finalmente restablece la membrana a su estado de reposo. La reconstrucción que hicieron Hodgkin y Huxley del potencial de acción y de todas sus características muestra que las propiedades de las conductancias al Na+ y al K+ sensibles al voltaje, junto con las fuerzas impulsoras electroquímicas creadas por los transportadores iónicos, son suficientes para explicar los potenciales de acción. El empleo de métodos tanto empíricos como teóricos condujo a un nivel de rigor sin precedentes para un problema tan antiguo, y estableció un estándar de verificación pocas veces logrado en la investigación biológica. Señalización a larga distancia por medio de los potenciales de acción Los mecanismos de generación del potencial de acción dependiente de voltaje también explican la transmisión a larga distancia de estas señales eléctricas. Recuérdese del Capítulo 2 que las neuronas son relativamente malas conductoras pasivas de la electricidad, al menos en comparación con un cable. No obstante, los potenciales de acción pueden atravesar grandes distancias a pesar de las escasas propiedades pasivas. ¿Cómo ocurre? El mecanismo de propagación del potencial de acción es fácil de comprender una vez que se conoce el modo en que se generan los potenciales de acción y cómo la corriente fluye pasivamente a lo largo de un axón. Un estímulo despolarizante –por lo general, una señal sináptica o un potencial de receptor en una neurona intacta o un pulso de corriente inyectado en un experimento como el que se muestra en la Figura 3.10– despolariza localmente el axón, y abre así los canales del Na+ sensibles al voltaje en esa región. La apertura de los canales del Na+ produce un movimiento hacia el interior de Na+, y la despolarización resultante del potencial de membrana genera un potencial de acción en ese sitio. Algo de la corriente local generada por el potencial de acción fluirá entonces pasivamente por el axón, de la misma forma en que las corrientes subumbrales se propagan a lo largo de un axón (véase Figura 2.3). Obsérvese que este flujo pasivo de corriente no requiere el movimiento de Na+ a lo largo del axón, sino que en cambio se produce por un traslado de carga, algo similar a lo que sucede cuando los cables conducen pasivamente electricidad por transmisión de la carga de electrones. Este flujo pasivo de corriente despolariza el potencial de membrana en la región adyacente del axón y abre así los canales del sodio en la membrana vecina. La despolarización local desencadena un potencial de acción en esta región, que se propaga entonces de nuevo en un ciclo continuo hasta que el potencial de acción alcanza el extremo axónico. Por lo tanto, la propagación del potencial de acción requiere la actividad coordinada de dos formas de flujo de corriente: el flujo pasivo de corriente 51 así como las corrientes activas que fluyen a través de los canales iónicos dependientes de voltaje. Las propiedades regenerativas de la apertura de los canales del Na+ permiten que los potenciales de acción se propaguen a todo o nada al actuar como refuerzo en cada punto de la longitud del axón, con lo cual aseguran la transmisión a largas distancias de las señales eléctricas. Recuérdese que los axones son refractarios después de un potencial de acción: la generación de un potencial de acción por un tiempo corto hace más difícil que el axón produzca potenciales de acción posteriores (véase Figura 3.8B). La refractariedad limita la cantidad de potenciales de acción que una neurona puede producir por unidad de tiempo, y los diferentes tipos de neuronas tienen distintas velocidades máximas de disparo del potencial de acción debido a diferentes tipos y densidades de canales iónicos. Como se describió en la sección anterior, el período refractario se origina porque la despolarización que produce la apertura de los canales del Na+ también causa una activación retardada de los canales del K+ e inactivación de los canales de Na+, lo que transitoriamente hace más difícil para el axón producir otro potencial de acción. Esta refractariedad también tiene importantes consecuencias para la conducción del potencial de acción a lo largo de los axones. A medida que el potencial de acción se desplaza a lo largo de un axón, en su camino deja inactivados los canales de Na+ y activados los de K+ por un breve período. La refractariedad resultante de la región de la membrana donde se ha generado un potencial de acción impide la reexcitación posterior de esta membrana a medida que se generan los potenciales de acción en regiones adyacentes del axón (véase Figura 3.10). Esta importante característica impide la propagación retrógrada del potencial de acción hacia su punto de iniciación a medida que viajan por el axón. Por lo tanto, el comportamiento refractario asegura la propagación polarizada de los potenciales de acción desde su punto habitual de iniciación cerca del cuerpo de la célula neuronal hacia las terminaciones sinápticas en el extremo distal del axón. Aumento de la velocidad de conducción como resultado de la mielinización Como consecuencia de su mecanismo de propagación, los potenciales de acción se producen cada vez más tarde a mayores distancias a lo largo del axón (Figura 3.10, abajo a la izquierda). Así, el potencial de acción tiene un ritmo mensurable de transmisión denominado velocidad de conducción, que es un parámetro importante porque define el tiempo necesario para que la información eléctrica viaje del extremo de una neurona a otra y, de ese modo, limite el flujo de información dentro del sistema nervioso. No es sorprendente entonces que se desarrollaran varios mecanismos para optimizar la propagación de los potenciales de acción a lo largo de los axones. Como la conducción del potencial de acción requiere el flujo pasivo y el flujo activo de corriente, la velocidad de propagación del potencial de acción es determinada por los dos fenómenos. Una forma de mejorar el 52 CAPÍTULO 3 Oligodendrocito (A) Axón mielínico (B) Nodo de Ranvier Canales del Na+ Vaina de mielina (C) Propagación del potencial de acción Na+ t=1 Axon Na+ Punto B Punto A K+ P unto C Na+ t = 1,5 Na+ K+ Punto B Punto A K+ P unto C Na+ t=2 Na+ K+ Punto B Punto A t=1 Punto A t = 1,5 t=2 P unto C 0 mV FIGURA 3.11 Conducción saltatoria del potencial de acción a lo largo de un axón mielínico. (A) Diagrama de un axón mielínico. (B) Localización de los canales de Na con –65 puerta de voltaje (rojo) en un nodo de Ranvier en un axón mielínico del nervio óptico. El verde indica la proteína Caspr, que se localiza adyacente al nodo de Ranvier. (C) La corriente local en Umbral 0 respuesta a la iniciación del potencial de acción en un sitio particular fluye localmente, como se describe en la Figura 3.10. Sin embargo, la presencia de mielina impide que la corriente local escape a través de la membrana intermodal; por lo tanto, fluye más allá a lo largo del axón –65 de lo que haría en ausencia de mielina. Más aún, los canales del Na+ con puerta de voltaje solo están presentes en los nodos de Ranvier (los canales del K+ con puerta de voltaje están 0 presentes en los nodos de algunas neuronas, pero no en otras). Esta disposición indica que la Punto B Punto C –65 generación de corrientes activas de Na+ con puerta de voltaje solo debe desarrollarse en estas regiones amielínicas. El resultado es un aumento muy grande de la velocidad de conducción del potencial de acción. El panel final representa el curso temporal de los cambios en el potencial de membrana en los puntos indicados. (B de Chen y cols., 2004.) PERMEABILIDAD DE LA MEMBRANA DEPENDIENTE DE VOLTAJE 53 Otra estrategia para mejorar el flujo pasivo de corriente eléctrica es aislar la membrana axónica, lo que reduce la capacidad de la corriente para escapar del axón y aumenta así la distancia a lo largo del axón que puede fluir pasivamente una corriente local dada. Esta estrategia es evidente en la mielinización de los Axón mielínico axones, proceso por el cual los oligodendrocitos en el sistema nervioso central (y las células de Schwann en el sistema nervioso periférico) envuelven el axón en mielina, que está formada por numerosas capas de membrana glial en íntima aposición (Figura 3.11A; véase también Capítulo 1). Al actuar como aislante eléctrico, la mielina acelera mucho la conducción del potencial de acción (Figura t=2 3.12). Por ejemplo, mientras las velocidades de conducción de los axones amielínicos varían entre unos 0,5 y 10 m/s, los axones mielínicos pueden conducir a velocidades de hasta 150 metros por segundo. La razón principal subyacente a este incremento pronunciado de la velocidad es que el proceso prolongado de generación del potencial de acción se desarrolla solamente en puntos específicos a lo largo del axón, denominados nodos de Ranvier, donde existe una brecha en la envoltura de mielina (véase Figura 1.3G). Si se aislara toda la envoltura de un axón, no habría lugar para que la corriente fluyera fuera del axón y no t=3 podrían generarse los potenciales de acción. Por lo tanto, los canales de Na+ con puerta de voltaje necesarios para los potenciales de acción se encuentran solo en estos nodos de Ranvier (Figura 3.11B). Un potencial de acción generado en un nodo de Ranvier produce una corriente que fluye pasivamente dentro del segmento mielínico hasta que se alcanza el siguiente nodo. Este flujo local de corriente genera entonces un potencial de acción en el segmento vecino, y el ciclo se repite a lo largo FIGURA 3.12 La mielina aumenta la velocidad de conducción del potencial de acción. El del axón. Dado que la corriente fluye a través diagrama compara la velocidad de conducción del potencial de acción en axones amielínicos (panel de la membrana neuronal solo en los nodos superior de cada par) y mielínicos (paneles inferiores). Para aclarar, solo se muestra la conducción (Figura 3.11C), este tipo de propagación se pasiva (flechas) para la dirección de propagación del potencial de acción. denomina saltatoria, lo que indica que el potencial de acción salta de nodo a nodo. No es sorprendente que la pérdida de mielina, como se observa en flujo pasivo de corriente es aumentar el diámetro de un axón, lo enfermedades como la esclerosis múltiple, produzca distintos proque disminuye eficazmente la resistencia interna al flujo pasivo blemas neurológicos graves (Recuadro 3C). de corriente. Se presume que el aumento consiguiente en la velocidad de conducción del potencial de acción explica por qué los Resumen axones gigantes evolucionaron en invertebrados como el calamar El potencial de acción y todas sus propiedades complejas puey por qué los axones de conducción rápida en todos los animales den explicarse por cambios dependientes del tiempo y del voltatienden a ser más grandes que los de conducción lenta. t=1 Axón amielínico 54 CAPÍTULO 3 RECUADRO 3C Esclerosis múltiple La esclerosis múltiple (EM) es una enfermedad del sistema nervioso central caracterizada por distintos problemas clínicos que surgen por la desmielinización y la inflamación a lo largo de las vías axónicas. En general, el trastorno comienza entre los 20 y los 40 años; se caracteriza por el inicio brusco de déficits neurológicos que en los casos típicos persisten durante días o semanas y luego remiten. El curso clínico varía desde los pacientes que no tienen ninguna pérdida neurológica persistente, algunos de los cuales experimentan solo exacerbaciones ocasionales con ulterioridad, hasta otros que se deterioran en forma progresiva como resultado de la afectación extensa e incesante del sistema nervioso central. Los signos y síntomas de la EM están determinados por la localización de las regiones afectadas. Son en particular frecuentes la ceguera monocular (debida a lesiones del nervio óptico), la debilidad motora o la parálisis (debidas a lesiones de los tractos corticoespinales), sensaciones somáticas anormales (debidas a lesiones de las vías somatosensitivas, a menudo en los cordones posteriores), diplopía (debida a lesiones del fascículo longitudinal medial) y mareos (por lesiones de las vías vestibulares). A menudo se presentan anomalías en el líquido cefalorraquídeo, que suele contener una cantidad anormal de células asociada con inflamación y un aumento del contenido de anticuerpos (signo de una respuesta inmunitaria alterada). El diagnóstico de EM es difícil y generalmente se basa en la presencia de un problema neurológico que remite y luego retorna en un sitio no relacionado. A veces, la confirmación puede obtenerse mediante una resonancia magnética (RM) o por la evidencia funcional de lesiones en una vía particular en los potenciales evocados anormales. El distintivo histológico de la EM en el examen post mórtem son las numerosas lesiones en diferentes sitios que muestran pérdida de mielina asociada con infiltración de células inflamatorias y, en algunos casos, pérdida de los propios axones. El concepto de EM como enfermedad desmielinizante está profundamente introducido en la literatura clínica, aunque no se sabe con precisión de qué modo la desmielinización se traduce en déficits funcionales. Es indudable que la pérdida de la vaina de mielina que rodea muchos axones compromete la conducción de los potenciales de acción, y los patrones anormales de conducción nerviosa que presumiblemente aparecen producen la mayor parte de los déficits clínicos en la enfermedad. Sin embargo, la EM puede tener efectos que se extienden más allá de la pérdida de la vaina de mielina. Está claro que algunos axones en realidad están destruidos, quizá como resultado de procesos inflamatorios en la mielina suprayacente, pérdida de apoyo trófico del axón por los oligodendrocitos o ambos. Por lo tanto, la pérdida axónica también contribuye a los déficits funcionales en la EM, en especial en las formas progresivas y crónicas de la enfermedad. La causa final de la EM aún está poco clara. Es indudable que el sistema inmunitario contribuye con el daño y las nuevas terapias inmunorreguladoras proporcionan beneficios sustanciales a muchos pacientes. No se sabe con precisión de qué modo el sistema inmunitario se activa para producir la lesión. La hipótesis más popular es que la EM es una enfermedad autoinmunitaria (esto es, una enfermedad en la que el sistema inmunitario ataca los propios componentes del cuerpo). La posibilidad de que la inmunización de animales de experimentación con cualquiera de varios componentes moleculares de la vaina de mielina induzca una enfermedad desmielinizante (llamada encefalomielitis alérgica experimental) muestra que un ataque autoinmunitario sobre la membrana de la mielina es suficiente como para producir un cuadro similar a la EM. Las pruebas muy recientes sugieren que una diana importante del ataque autoinmunitrio es la contactina-2, una proteína que se encuentra en la glía y los axones en el nodo de Ranvier. Una explicación posible de la enfermedad humana es que un individuo con susceptibilidad genética se infecta transitoriamente (p. ej., por una enfermedad viral menor) por un microorganismo que expresa una molécula estructuralmente similar a la contactina-2 o a algún otro componente de la mielina. Se crea una respuesta inmunitaria a este antígeno para atacar al invasor, pero la incapacidad del sistema inmunitario a la hora de discriminar entre la proteína extraña y la propia conduce a la destrucción de la mielina por otra parte normal, un escenario que se observa en ratones infectados por el virus de Theiler. Una hipótesis alternativa es que la EM está causada por una infección persistente por un virus u otro microorganismo. Bajo esta interpretación, los esfuerzos en curso del sistema inmunitario para desembarazarse del patógeno producen el daño de la mielina. La paraparesia espástica tropical proporciona un precedente para esta idea. Este trastorno es una enfermedad caracterizada por la progresión gradual de debilidad de las piernas y deterioro del control de la función vesical asociados con hiperreflexia osteotendinosa y signo de Babinski positivo (véase Figura 17.16). Este cuadro clínico es similar al de la EM rápidamente evolutiva y se sabe que la paraparesia espástica tropical se produce por la infección persistente por un retrovirus (virus linfocitotrópico T humano-1). No obstante este precedente, la hipótesis de la infección viral persistente para la EM requiere una demostración clara de la presencia de un virus. A pesar de las comunicaciones periódicas de un virus asociado con EM, aún no hay evidencia convincente. En resumen, a pesar de los beneficios para algunos pacientes de las terapias inmunomoduladoras, el diagnóstico y el tratamiento de la esclerosis múltiple todavía representa un desafío clínico desalentador. Bibliografía Bhat, R. and L. Steinman (2009) Innate and adaptive autoimmunity directed to the central nervous system. Neuron 64: 123-132. Derfuss T, and 18 others. (2009) Contactin-2/ TAG-l-directed autoimmunity is identified in multiple sclerosis patients and mediates gray matter pathology in animals. Proc. Natl. Acad. Sci. USA 106: 8302-8307. Ransohoff, R. M. (2007) Natalizumab for multiple sclerosis. Neio Engl. J. Med. 356: 26222629. Waxman, S. G. (2006) Ions, energy, and axo-nal injury: Towards a molecular neurology of multiple sclerosis. Trends Mol. Med. 12: 192-195. Zanvil, S. S. and L. Steinman (2003) Diverse targets for intervention during inflammatory and neurodegenerative phases of multiple sclerosis. Neuron 38: 685-688. PERMEABILIDAD DE LA MEMBRANA DEPENDIENTE DE VOLTAJE je en las permeabilidades al Na+ y al K+ de las membranas neuronales. Esta conclusión deriva fundamentalmente de las pruebas obtenidas mediante un dispositivo denominado pinzamiento de voltaje. Esta técnica es un método de retroalimentación electrónica que permite el control del potencial de membrana neuronal y, simultáneamente, la medición directa de los flujos de Na+ y de K+ dependientes de voltaje que producen el potencial de acción. Los experimentos de pinzamiento de voltaje muestran que una elevación transitoria en la conductancia al Na+ se activa con rapidez y luego se inactiva durante la despolarización sostenida del potencial de membrana. Estos experimentos también demuestran una elevación en la conductancia al K+ que se activa de forma tardía y, al contrario de la conductancia al Na+, no se inactiva. Los modelos matemáticos de las propiedades de estas conductancias indican que ellos solos son responsables de la producción de potenciales de acción todo o nada en el axón del calamar. Los potenciales de acción se propagan a lo largo de los axones de las células nerviosas iniciados por el gradiente de voltaje entre las regiones activas e inactivas del axón en virtud del flujo local de corriente. De esta forma, los potenciales de acción compensan las propiedades eléctricas pasivas relativamente malas de las células nerviosas y permiten el señalamiento neural en largas distancias. Estos hallazgos electrofisiológicos clásicos proporcionan una base sólida para considerar las variaciones funcionales y finalmente moleculares del señalamiento neural que se analizan en el capítulo siguiente. 55 Lecturas adicionales Revisiones Armstrong, C. M. and B. Hille (1998) Voltage-gated ion channels and electrical excitability. Neuron 20: 371-380. Salzer, J. L. (2003) Polarized domains of myelinated axons. Neuron 40: 297-318. Artículos originales importantes Armstrong, C. M. and L. Binstock (1965) Anomalous rectification in the squid giant axon injected with tetraethylammonium chloride. J. Gen, Physiol. 48: 859-872. Chen, C. and 17 others (2004) Mice lacking sodium channel beta1 subunits display defects in neuronal excitability, sodium channel expression, and nodal architecture. J. Neurosci. 24: 4030-4042. Hodgkin, A. L. and A. F. Huxley (1952a) Currents carried by sodium and potassium ions through the membrane of the giant axon of Loligo. J. Physiol. 116: 449-472. Hodgkin, A. L. and A. F. Huxley (1952b) The components of membrane conductance in the giant axon of Loligo. J. Physiol. 116: 473-496. Hodgkin, A. L. and A. F. Huxley (1952c) The dual effect of membrane potential on sodium conductance in the giant axon of Loligo. J. Physiol. 116: 497-506. Hodgkin, A. L. and A. F. Huxley (1952d) A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 116: 507-544. Hodgkin, A. L., A. F. Huxley and B. Katz (1952) Measurements of currentvoltage relations in the membrane of the giant axon of Loligo. J. Physiol. 116: 424-448. Hodgkin, A. L. and W. A. H. Rushton (1938) The electrical constants of a crustacean nerve fibre. Proc. R. Soc. Lond. 133: 444-479. Moore, J. W., M. P. Blaustein, N. C. Anderson and T. Narahashi (1967) Basis of tetrodotoxin‘s selectivity in blockage of squid axons. J. Gen. Physiol. 50: 1401-1411. Libros Aidley, D. J. and P. R. Stanfield (1996) Ion Channels: Molecules in Action. Cambridge: Cambridge University Press. Hille, B. (2001) Ion Channels of Excitable Membranes, 3rd Ed. Sunderland, MA: Sinauer Associates. Johnston, D. and S. M.-S. Wu (1995) Foundations of Cellular Neurophysiology. Cambridge, MA: MIT Press. Junge, D. (1992) Nerve and Muscle Excitation, 3rd Ed. Sunderland, MA: Sinauer Associates. Matthews, G. G. (2003) Cellular Physiology of Nerve and Muscle, 4th Ed. Maiden, MA: Blackwell Publishing. 4 Canales ióniCos y transportadores Aspectos generales LA GENERACIÓN DE SEÑALES ELÉCTRICAS en las neuronas requiere que las membranas plasmáticas establezcan gradientes de concentración para iones específicos y que estas membranas sufran cambios rápidos y selectivos en su permeabilidad a estos iones. Las proteínas de la membrana que crean y mantienen los gradientes iónicos se denominan transportadores activos, mientras que otras proteínas llamadas canales iónicos dan origen a cambios selectivos en la permeabilidad iónica. Los canales iónicos son proteínas transmembrana que contienen una estructura especializada, llamada poro, que permite que iones particulares crucen la membrana neuronal. Algunos canales iónicos también contienen estructuras que son sensibles al potencial eléctrico a través de la membrana. Estos canales con puerta de voltaje se abren o se cierran en proporción a la magnitud del potencial de membrana, lo que permite que la permeabilidad de la membrana esté regulada por los cambios de este potencial. Otros tipos de canales iónicos son abiertos por señales químicas, ya sea señales extracelulares como los neurotransmisores o por señales intracelulares como los segundos mensajeros. Otros incluso responden a estímulos mecánicos, a cambios de temperatura o a una combinación de estas señales. Muchos tipos de canales iónicos han sido caracterizados a nivel tanto genético como proteico, lo que condujo a la identificación de gran cantidad de subtipos de canales que se expresan de modo diferente en las células neuronales y no neuronales. El patrón de expresión específico de los canales iónicos en los diferentes tipos celulares puede generar un amplio espectro de características eléctricas. Al contrario de los canales iónicos, los transportadores activos son proteínas de membrana que producen y mantienen los gradientes de concentración de los iones. La más importante es la bomba de Na+, que hidroliza el ATP para regular las concentraciones intracelulares tanto de Na+ como de K+. Otros transportadores activos producen gradientes de concentración para la gama completa de iones fisiológicamente importantes, que incluyen Cl−, Ca2+ y H+. Desde la perspectiva de la señalización eléctrica, los transportadores activos y los canales iónicos son complementarios: los transportadores crean los gradientes de concentración que ayudan a impulsar flujos iónicos a través de los canales iónicos abiertos, y generan así señales eléctricas. Canales iónicos que participan en los potenciales de acción Si bien Hodgkin y Huxley no tenían ningún conocimiento de la naturaleza física de los mecanismos conductivos subyacentes a los potenciales de acción, propusieron la idea de que las 58 CAPÍTULO 4 (A) Cerrado Abierto 0 –40 –80 0 5 10 Tiempo (ms) 15 0 5 10 Tiempo (ms) 15 0 5 10 Tiempo (ms) 15 0 5 10 Tiempo (ms) 15 2 pA (C) INa microscópicas promediadas (pA) INa microscópicas (B) Potencial de membrana (mV) membranas de las células nerviosas poseen canales que permiten el paso selectivo de los iones de un lado al otro de la membrana (véase Capítulo 3). Sobre la base de las conductancias y las corrientes iónicas medidas a través de los experimentos de pinzamiento de voltaje, los canales postulados debían tener varias propiedades. Primero, como las corrientes iónicas son muy grandes, los canales debían ser capaces de permitir que los iones atravesaran las mem- (D) 0,4 0 –0,4 –0,8 INa microscópicas (pA) 200 0 –200 –400 –600 –800 (E) 0,8 Probabilidad de apertura de los canales del Na+ branas con velocidades altas. Segundo, como las corrientes iónicas dependen del gradiente electroquímico a través de la membrana, los canales deberían utilizar estos gradientes. Tercero, dado que el Na+ y el K+ fluyen a través de la membrana en forma independiente uno del otro, los diferentes tipos de canales deberían ser capaces de discriminar entre Na+ y K+, permitiendo que sólo uno de estos iones fluya a través de la membrana en las condiciones relevantes. Por último, dado que las conductancias iónicas son dependientes de voltaje, los canales deberían ser capaces de sentir el voltaje a través de la membrana y abrirse sólo cuando éste alcanza niveles apropiados. Aunque en la década de 1950 este concepto de los canales era altamente especulativo, las investigaciones ulteriores establecieron que, más allá de ninguna duda, las proteínas transmembrana llamadas canales iónicos sensibles al voltaje en efecto existen y son responsables de los potenciales de acción y de todas las otras señales eléctricas descritas en los Capítulos 2 y 3. La primera prueba directa de la presencia de canales selectivos para iones y sensibles al voltaje en las membranas de las células nerviosas provino de mediciones de las corrientes iónicas que fluyen a través de los canales iónicos individuales. El poder de resolución del aparato de pinzamiento de voltaje utilizado por Hodgkin y Huxley sólo alcanzaba para medir la corriente agregada resultante del flujo de iones a través de muchos miles de canales. En 1976, Erwin Neher y Bert Sakmann diseñaron una técnica capaz de medir las corrientes que fluyen a través de canales únicos en el Max Planck Institute en Alemania. Esta técnica notable, llamada pinzamiento zonal (patch clamp), revolucionó el estudio de las corrientes de membrana (Recuadro 4A). En particular, el método de pinzamiento zonal proporcionó el medio para evaluar directamente las hipótesis de Hodgkin y Huxley sobre las características de los canales iónicos. Las corrientes que fluyen a través de los canales de Na+ se examinan mejor en circunstancias experimentales que previenen el flujo de corrientes a través de otros tipos de canales presentes en la membrana (p. ej., canales del K+). En estas condiciones, la despolarización de una zona de la membrana del axón gigante del calamar produce el flujo de diminutas corrientes hacia el interior, pero sólo en ocasiones (Figura 4.1). La intensidad de estas corrientes es minúscula, aproximadamente 1-2 pA (es decir, 10-12 amperios), pero es estereotipada, y alcanza valores discretos que sugieren el 0,6 0,4 0,2 0 –80 –60 –40 –20 0 20 40 60 Potencial de membrana (mV) FIGURA 4.1 Mediciones con el método de pinzamiento zonal de membrana de las corrientes iónicas que fluyen a través de los canales únicos del Na+ en un axón gigante del calamar. En estos experimentos, se aplicó Cs+ al axón para bloquear los canales del K+ con puerta de voltaje. Los pulsos de voltaje despolarizantes (A) aplicados en un parche de membrana que contenía un único canal del Na+ dan por resultado corrientes breves (B, deflexiones hacia abajo) en los siete registros sucesivos de corriente de membrana (INa). (C) El promedio de muchos de estos registros de corriente muestra que la mayoría de los canales se abren en 1-2 milisegundos iniciales tras la despolarización de la membrana, después de lo cual la probabilidad de la apertura de canales disminuye debido a la inactivación de los canales. (D) Una corriente macroscópica medida desde otro axón muestra la estrecha correlación entre los cursos temporales de las corrientes macroscópicas y microscópicas de Na+. (E) La probabilidad de apertura de un canal del Na+ depende del potencial de membrana, y aumenta a medida que la membrana se despolariza. (B, C, de Bezanilla y Correa, 1995; D, de Vandenberg y Bezanilla, 1991; E, de Correa y Bezanilla, 1994.) CANALES IÓNICOS Y TRANSPORTADORES 59 RECUADRO 4A Método de pinzamiento zonal de membrana Como alternativa, si se separa la pipeA raíz de la invención del método de pin- de vidrio, pequeños trozos de membrana zamiento zonal, surgió gran cantidad de pueden ser separados de la célula sin in- ta mientras se encuentra en la preparainformación acerca de los canales iónicos. terrumpir el sellado; esto proporciona una ción de célula completa, se produce una Esta técnica se basa en una idea muy sim- preparación que está libre de las complica- zona de membrana que tiene su superficie ple. Se utiliza una pipeta de vidrio con un ciones impuestas por el resto de la célula. extracelular expuesta. Esta disposición, orificio muy pequeño para lograr un contac- Si simplemente se separa la pipeta que se denominada configuración de registro con to estrecho con un área pequeña, o zona, de utiliza en la preparación con fijación de la el exterior hacia afuera, es óptima para membrana neuronal. Después de succionar célula, se produce una pequeña vesícula estudiar cómo la actividad de los canales con suavidad a través de la pipeta, la cone- de membrana que queda fijada a la pipeta. es influida por las señales químicas exxión entre la pipeta y la membrana se torna Al exponer la punta de la pipeta al aire, tracelulares, como los neurotransmisores tan fuerte que ningún ion puede filtrarse la vesícula se abre para dar una pequeña (véase Capítulo 5). Esta gama de posibles entre ambas. Por lo tanto, todos los iones zona de la membrana con su (ex) superfi- configuraciones convierte al método de que fluyen cuando se abre un único canal cie intracelular expuesta. Esta disposición, pinzamiento zonal de membrana en una iónico deben ingresar en la pipeta. La di- denominada configuración de registro zo- técnica extraordinariamente versátil para minuta corriente eléctrica resultante puede nal con el interior hacia afuera, permite el estudio de la función de los canales medirse con un amplificador electrónico ul- el cambio del medio al cual está expuesta iónicos. Las versiones robotizadas de las trasensible conectado a la pipeta. Esta dis- la superficie intracelular de la membra- técnicas de pinzamiento zonal están enposición se denomina método de registro de na. Por lo tanto, la configuración con el trando uniformemente en la industria, ya pinzamiento zonal con fijación a la célula. interior hacia afuera es particularmente que sirven como medio muy sensible y ráComo sucede con el método convencional útil cuando se estudia la influencia de las pido para evaluar agentes terapéuticos que de pinzamiento de voltaje, el de pinzamien- moléculas intracelulares sobre la función actúan sobre los canales iónicos. to zonal permite el control experimental de los canales iónicos. del potencial de membrana para caracterizar la dependencia de voltaje de las corrientes de la Registro con fijación celular Bibliografía membrana. Dunlop, J., M. Bowlby, R. Peri, D. Algunas modificaciones técniSucción Pipeta Vasilyev and R. Arias (2008) Highcas menores permiten otras confileve de registro throughput electrophysiology: An guraciones de registro. Por ejememerging paradigm for ion-channel screening and physiology. Nature plo, si se interrumpe la zona de Rev. Drug Discov. 7: 358-368. la membrana en el interior de la Hamill, O. P., A. Marty, E. Neher, B. Contacto estrecho entre pipeta aplicando brevemente una Sakmann and F. J. Sigworth (1981) la pipeta y la membrana succión fuerte, se produce una coImproved patch-clamp techniques for high-resolution current recording municación entre el interior de la Registro de célula completa Registro con el interior hacia afuera from cells and cell-free membrane pipeta y el citoplasma de la célupatches. Pflügers Arch. 391: 85-100. la. Esta disposición permite efecExponer Levis, R. A. and J. L. Rae (1998) al aire Low-noise patch-clamp techniques. tuar mediciones de potenciales Pulso fuerte de Meth. Enzym. 293: 218-266. eléctricos y corrientes desde toda succión Sakmann, B. and E. Neher (1995) la célula y, por lo tanto, se denoSingle-Channel Recording, 2nd Ed. mina método de registro de célula New York: Plenum Press. completa. La configuración de Dominio citoplasmático célula completa también permite El citoplasma se comunica accesible con el interior de la pipeta el intercambio por difusión entre la pipeta y el citoplasma, lo que Registro con el exterior hacia afuera produce una forma conveniente de inyectar de manera “sectorial” Separar Los bordes sustancias en el interior de una la pipeta de la célula. membrana se unen Otras dos variantes del método de pinzamiento zonal se oriCuatro configuraciones en las ginan en el hallazgo de que, una Dominio extracelular mediciones del pinzamiento zonal vez formado un sellado ajustado accesible de membrana de las corrientes entre la membrana y la pipeta iónicas. 60 CAPÍTULO 4 flujo iónico a través de canales iónicos abiertos individuales. Estas corrientes son varios órdenes de magnitud más pequeñas que las corrientes de Na+ medidas mediante pinzamiento de voltaje de todo el axón. Las corrientes que fluyen a través de canales únicos se denominan corrientes microscópicas para distinguirlas de las corrientes macroscópicas que fluyen a través de gran cantidad de canales distribuidos en una región mucho más extensa de la membrana de superficie. Aunque, por cierto, las corrientes microscópicas son pequeñas, una corriente de 1 pA refleja el flujo de miles Potencial de membrana (mV) (A) 50 0 –50 –100 0 10 20 30 Tiempo (ms) 40 0 10 20 30 Tiempo (ms) 40 0 10 20 30 Tiempo (ms) 40 0 10 20 30 Tiempo (ms) 40 (B) (C) IK microscópicas promediadas (pA) IK microscópicas Abierto Cerrado 2 pA 1 0 3 IK macroscópicas (mA/cm2) (D) Pro babilidad de apertura de los canales del K+ (E) 2 1 0 FIGURA 4.2 Mediciones con el método de pinzamiento zonal de membrana de las corrientes iónicas que fluyen a través de canales únicos del K+ en un axón gigante del 0,6 calamar. En estos experimentos, se aplicó tetrodoxotina al axón para bloquear los canales del Na+ con puerta de voltaje. Los pulsos de voltaje despolarizantes (A) aplicados en una zona de la membrana que contenía un único canal del K+ dan por resultado corrientes breves (B, deflexiones hacia arriba) siempre que el canal se abre. (C) El promedio de estos registros de corriente muestra que la mayoría de los 0,4 0,2 0 –80 –60 –40 –20 0 20 40 Potencial de membrana (mV) de iones por milisegundo. Por lo tanto, según se predijo, un canal único puede dejar pasar muchos iones a través de la membrana en un período muy corto. Además, varias observaciones prueban que las corrientes microscópicas que se muestran en la Figura 4.1B se deben a la apertura de canales únicos de Na+ activados por voltaje. Primero, las corrientes son transmitidas por el Na+; por tanto, están dirigidas hacia el interior cuando el potencial de membrana es más negativo que el ENa, invierten su polaridad en el ENa y se dirigen hacia afuera con potenciales más positivos, y se reducen en tamaño cuando la concentración de Na+ del medio externo está disminuida. Este comportamiento es exactamente paralelo al de las corrientes macroscópicas de Na+ descritas en el Capítulo 3. Segundo, los canales tienen un curso temporal de apertura, cierre e inactivación que se equipara a la cinética de las corrientes macroscópicas de Na+. Esta correspondencia es difícil de apreciar en la medición de las corrientes microscópicas que fluyen a través de un canal abierto único, porque los canales individuales se abren y se cierran de manera estocástica (aleatoria), como puede observarse en los trazados individuales de la Figura 4.1B. Sin embargo, la despolarización repetida del potencial de membrana hace que cada canal de Na+ se abra y se cierre muchas veces. Cuando se promedian todas las respuestas de corriente a gran cantidad de estos estímulos, la respuesta colectiva tiene un curso temporal que se parece mucho a la corriente macroscópica de Na+ (Figura 4.1C). En particular, los canales se abren principalmente al inicio de una despolarización prolongada, lo que muestra que con posterioridad se inactivan, como se predice a partir de la corriente macroscópica de Na+ (compárense las Figuras 4.1C y 4.1D). Tercero, tanto la apertura como el cierre de los canales son dependientes del voltaje; por lo tanto, los canales están cerrados en −80 mV pero se abren cuando el potencial de membrana está despolarizado. De hecho, la probabilidad de que cualquier canal dado se abra varía con el potencial de membrana (Figura 4.1E), nuevamente como se predice a partir de la conductancia macroscópica al Na+ (véase Figura 3.7). Por último, la tetrodotoxina, que bloquea la corriente macroscópica de Na+ (véase Recuadro 4B), también bloquea las corrientes microscópicas de Na+. Tomados en conjunto, estos resultados muestran que la corriente macroscópica de Na+ medida por Hodgkin y Huxley surge efectivamente de la sumatoria de muchos miles de corrientes microscópicas de Na+, donde cada una representa la apertura de un canal único de Na+ sensible al voltaje. 60 canales se abren con una demora, pero se mantienen abiertos mientras dure la despolarización. (D) Una corriente macroscópica medida desde otro axón muestra la correlación entre los cursos temporales de las corrientes microscópicas y macroscópicas de K+. (E) La probabilidad de apertura de un canal del K+ depende del potencial de membrana, y aumenta a medida que la membrana se despolariza. (B y C, de Augustine y Bezanilla, en Hille, 1992; D, de Augustine y Bezanilla, 1990; E, de Perozo y cols., 1991.) CANALES IÓNICOS Y TRANSPORTADORES Potencial de membrana (mV) Los experimentos de pinzamiento zonal de membrana también han puesto en evidencia las propiedades de los canales responsables de las corrientes macroscópicas de K+ asociadas a los potenciales de acción. Cuando el potencial de membrana se despolariza (Figura 4.2A), las corrientes microscópicas hacia el exterior (Figura 4.2C) pueden ser observadas en condiciones que bloqueen los canales del Na+. Estas corrientes microscópicas hacia el exterior muestran todas las características esperadas para las corrientes que fluyen a través de los canales del K+ relacionadas con los potenciales de acción. Por lo tanto, ni las corrientes microscópicas (Figura 4.2C) ni sus análogas macroscópicas (Figura 4.2D) se inactivan durante las despolarizaciones breves. Además, estas corrientes de canal único son sensibles a las manipulaciones iónicas y, al igual que las corrientes macroscópicas de K+, son dependientes del voltaje (Figura 4.2E). Estas pruebas y otras demuestran que las corrientes macroscópicas de K+ asociadas con la repolarización del potencial de acción se originan en la apertura de muchos canales de K+ sensibles al voltaje. En resumen, el pinzamiento zonal de membrana ha permitido la observación directa de las corrientes iónicas microscópicas que fluyen a través de canales iónicos únicos, lo que confirma que los canales del Na+ y del K+ sensibles al voltaje son responsables de las conductancias macroscópicas y las corrientes macroscópicas que subyacen al potencial de acción. Las mediciones del comportamiento de canales iónicos únicos también han proporcionado algunas ideas sobre los atributos moleculares de estos canales. Por ejemplo, los estudios de los canales únicos 61 muestran que la membrana del axón del calamar contiene por lo menos dos tipos de canales: uno selectivamente permeable al Na+ y otro selectivamente permeable al K+. Esta selectividad iónica significa que estos canales son capaces de discriminar entre Na+ y K+. Los dos canales tienen una compuerta de voltaje, lo que indica que su apertura está condicionada por el potencial de membrana. En ambos, la despolarización aumenta la probabilidad de apertura de los canales, mientras que la hiperpolarización los cierra (véase Figuras 4.1E y 4.2E). Por lo tanto, ambos tipos de canales deben tener un sensor de voltaje que detecte el potencial a través de la membrana (Figura 4.3). Sin embargo, estos canales difieren en aspectos importantes. Además de sus diferentes selectividades iónicas, la despolarización también inactiva el canal del Na+ pero no el canal del K+ y hacen que los canales del Na+ pasen a un estado no conductor. Por lo tanto, el canal del Na+ debe tener otro mecanismo molecular responsable de la inactivación. Y según se espera a partir del comportamiento macroscópico de las corrientes del Na+ y del K+ descritas en el Capítulo 3, difieren las propiedades cinéticas de las puertas de los dos canales. Por último, estas proteínas de los canales proporcionan sitios de unión singulares para los fármacos y para distintas neurotoxinas que se sabe bloquean subclases específicas de canales iónicos (Recuadro 4B). Esta información sobre la fisiología de los canales únicos establece el camino para estudios ulteriores de la diversidad molecular de los canales iónicos en distintos tipos celulares y de sus características funcionales detalladas. 50 0 –50 –100 0 5 10 15 Tiempo (ms) Canal del Na+ Na+ Na+ Cerrado Abierto En inactivación Inactivado Cerrado FIGURA 4.3 Estados funcionales de los Canal del K+ canales del Na+ y del K+ con puerta de voltaje. Las puertas de ambos canales están cerradas cuando la membrana está hiperpolarizada. Cuando el potencial está despolarizado, los sensores de voltaje (indicados con +) permiten K+ Cerrado Cerrado Abierto K+ Abierto Cerrado que se abran las puertas del canal, primero los canales del Na+ y luego los del K+. Los canales del Na+ también se inactivan durante la despolarización prolongada, mientras que muchos tipos de canales del K+ no lo hacen. 62 CAPÍTULO 4 RECUADRO 4B Toxinas que envenenan los canales iónicos (A) (1) Corriente de Na+ (nA/cm2 ) Conductancia al Na+ 0 <40 <80 0 20 40 60 80 0 20 Tiempo (ms) (2) 40 60 80 Tratado con toxina de escorpión Control Potencial de membrana (mV) 50 puntos diana de organismos que producen toxinas. Las toxinas peptídicas que afectan los canales del K+ incluyen la dendrotoxina, de las avispas; la apamina, de las abejas y la caribdotoxina, también producida por los escorpiones. Todas estas toxinas bloquean los canales del K+ como acción primaria; no se conoce ninguna toxina que afecte su activación o inactivación, aunque es posible que estos agentes no hayan sido descubiertos aún. Bibliografía Cahalan, M. (1975) Modification of sodium channel gating in frog myelinated nerve fibers by Centruroides sculpturatus scorpion venom. J. Physiol. (Lond.) 244: 511-534. Catterall, W. A., S. Cestele, V. Yarov-Yarovoy, F. H. Yu, K. Konoki and T. Scheuer (2007) Voltage-gated ion channels and gating modifier toxins. Toxicon 49: 124-141. Dutertre, S. and R. J. Lewis (2010) Use of venom peptides to probe ion channel structure and function. J. Biol. Chem. 285: 13315-13320. Narahashi, T. (2008) Tetrodotoxin: A brief history. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 84: 147-154. Schmidt, O. and H. Schmidt (1972) Influence of calcium ions on the ionic currents of nodes of Ranvier treated with scorpion venom. Pflügers Arch. 333: 51-61. (B) Tratado con toxina de escorpión Control Potencial de membrana (mV) peptídicas que también afectan los canales iónicos. Entre estas se encuentran las α-toxinas, que retardan la inactivación de los canales del Na+ (Figura A1); la exposición de las neuronas a las α-toxinas prolonga el potencial de acción (Figura A2), y se mezcla así la información en el interior del sistema nervioso de la víctima que está pronta a ser devorada. Otros péptidos del veneno del escorpión, denominadas β-toxinas, desplazan la dependencia del voltaje de la activación del canal del Na+ (Figura B). Estas toxinas hacen que los canales del Na+ se abran con potenciales mucho más negativos de lo normal, lo que induce una descarga descontrolada de potenciales de acción. Algunas toxinas alcaloides combinan estas acciones, ya que eliminan la inactivación y desplazan la activación de los canales del Na+. Una de estas toxinas es la batracotoxina, producida por una especie de rana (algunas tribus de indios sudamericanos utilizan este veneno en las puntas de sus flechas). Algunas plantas producen toxinas similares, que incluyen la aconitina, del botón de oro; la veratridina, de las lilas, y algunas toxinas insecticidas (piretrinas) producidas por plantas como crisantemos y rodondendros. Los canales del K+ también han sido normalizada Dada la importancia de los canales del Na+ y del K+ para la excitación neuronal, no es sorprendente que hayan evolucionado toxinas específicas de los canales en algunos organismos como mecanismos de autodefensa o para capturar presas. Una abundante colección de toxinas naturales tiene como diana selectiva los canales iónicos de neuronas y otras células. Estas toxinas no sólo son útiles para la supervivencia, sino también para el estudio de los canales iónicos celulares. La toxina de los canales mejor conocida es la tetrodotoxina, producida por algunos peces globo y otros animales. La tetrodotoxina produce una obstrucción potente y específica de los canales del Na+ responsables de la generación del potencial de acción, y paralizan así a los animales que son lo suficientemente desafortunados como para ingerirla. La saxitoxina, homólogo químico de la tetrodotoxina producida por dinoflagelados, tiene una acción similar sobre los canales del Na+. Los efectos potencialmente letales de comer mariscos que han ingerido estos dinoflagelados de la “marea roja” se deben a las potentes acciones neuronales de la saxitoxina. Los escorpiones paralizan a su presa inyectando una mezcla potente de toxinas Control Tratado con toxina de escorpión <120 <80 <40 0 40 Potencial de membrana (mV) (A) Efectos del tratamiento con toxina sobre los axones de la rana. (1) La α-toxina del escorpión Leiurus quinquestriatus prolonga las corrientes de Na+ registradas con el método de pinzamiento de voltaje. (2) Como resultado del aumento de la corriente de Na+, la α-toxina prolonga mucho la duración del potencial de acción axónico. 25 0 Obsérvese el cambio en la escala temporal tras el tratamiento con toxina. (B) El tratamiento del axón de una rana con β-toxina de otro escorpión, Centruroides sculpturatus, desplaza la activación de los canales del Na+, de modo que la conductancia al <25 <50 <75 2 4 0 Tiempo (ms) 6 0 8 4 Tiempo (ms) 10 Na+ comienza a aumentar con potenciales mucho más negativos de lo habitual. (A, de Schmidt y Schmidt, 1972; B, de Calahan, 1975.) CANALES IÓNICOS Y TRANSPORTADORES Diversidad de los canales iónicos Los estudios de genética molecular, junto con el método de pinzamiento zonal de membrana y otras técnicas, han conducido a muchos adelantos adicionales en la comprensión de los canales iónicos. Se han identificado y clonado algunos genes que codifican los canales del Na+ y del K+, así como otros tipos de canales. Un hecho sorprendente que ha surgido a partir de estos estudios moleculares es la cantidad y la diversidad de los genes que codifican estos canales iónicos. En la actualidad, se han descubierto muchos más de 100 genes de canales iónicos –cantidad que no pudo ser anticipada a partir de los primeros estudios de la función de los canales iónicos–. Para comprender la importancia funcional de estos múltiples genes de canales iónicos, los canales pueden ser expresados selectivamente en sistemas experimentales bien definidos, como células cultivadas u ovocitos de ranas (Recuadro 4C) y luego estudiados con pinzamiento zonal de membrana y otras técnicas fisiológicas. También es posible RECUADRO 4C Expresión de los canales iónicos en los ovocitos de Xenopus nucleótido) en la parte del gen del canal (A) que codifica una estructura de interés; las proteínas de canal resultantes son luego expresadas en ovocitos para evaluar las consecuencias funcionales de la mutación. La capacidad para combinar los métodos moleculares y fisiológicos en un único sistema celular ha convertido a los ovocitos de Xenopus en una herramienta experimental poderosa. En efecto, este sistema ha sido tan valioso para los estudios contemporáneos de los canales iónicos con puerta de voltaje como lo fue el axón del calamar para esos estudios en las décadas de 1950 y 1960. (B) Bibliografía Stühmer, W. (1998) Electrophysiological recordings from Xenopus oocytes. Meth. Enzym. 293: 280-300. Sumikawa, K., M. Houghton, J. S. Emtage, B. M. (C) Richards and E. A. Barnard (1981) Active multisubunit ACh receptor assembled by translation of heterologous mRNA in Xenopus oocytes. Nature 292: 862-864. 1 mm 50 Potencial de membrana (mV) Gundersen, C. B., R. Miledi and I. Parker (1984) Slowly inactivating potassium channels induced in Xenopus oocytes by messenger ribonucleic acid from Torpedo brain. J. Physiol (Lond.) 353: 231-248. Gurdon, J. B., C. D. Lane, H. R. Woodland and G. Marbaix (1971) Use of frog eggs and oocytes for the study of messenger RNA and its translation in living cells. Nature 233: 177-182. 0 <50 <100 (A) Rana africana de uñas, Xenopus laevis. (B) Varios ovocitos de Xenopus que destacan la coloración oscura del polo animal y la coloración más clara del polo vegetal. (Cortesía de P. Reinhart.) (C) Resultados de un experimento de pinzamiento de voltaje que muestran las corrientes de K+ producidas tras una inyección de mRNA del canal de K+ en un ovocito. (De Gundersen y cols., 1984.) Corriente de K+ (+A) La transición entre la secuencia del DNA del gen de un canal iónico hasta el conocimiento de la función del canal supone todo un desafío. Para abordarlo es esencial contar con un sistema experimental en el cual el producto genético pueda expresarse eficientemente la función del canal resultante pueda ser estudiada con métodos como la técnica de pinzamiento zonal de membrana. En condiciones ideales, el vehículo para la expresión debe hallarse fácilmente disponible, debe tener varios canales endógenos y ser lo suficientemente grande como para permitir que el mRNA y el DNA sean microinyectados con facilidad. Los ovocitos (huevos inmaduros) de la rana africana de uñas, Xenopus laevis (Figura A), cumplen todas estas exigencias. Estas células enormes (de aproximadamente 1 mm de diámetro; Figura B) se obtienen con facilidad de Xenopus hembra. La investigación realizada en la década de 1970 por John Gurdon, un biólogo del desarrollo, mostró que la inyección de mRNA exógeno en los ovocitos de la rana hacía que sintetizaran proteínas en cantidades asombrosas. A comienzos de la década de 1980, Ricardo Miledi, Eric Barnard y otros neurobiólogos demostraron que los ovocitos de Xenopus podían expresar canales iónicos exógenos y que los métodos fisiológicos podían ser utilizados para estudiar las corrientes iónicas generadas por los canales recién sintetizados (Figura C). Como resultado de estos estudios pioneros, los experimentos de expresión heteróloga se han convertido actualmente en una forma estándar para estudiar los canales iónicos. El enfoque ha sido de especial utilidad para descifrar la relación entre la estructura y la función de los canales. En estos experimentos, se efectúan mutaciones precisas (que a menudo afectan un único 63 4 3 2 1 0 0 0,2 0,4 0,6 0,8 1,0 Tiempo (s) 64 CAPÍTULO 4 producir la deleción de los genes de los canales en organismos genéticamente tratables, como los ratones o las moscas de la fruta, para determinar los papeles que desempeñan estos canales en el organismo intacto. Estos estudios han observado muchos canales con puerta de voltaje que responden al potencial de membrana de forma muy similar a los canales del Na+ y del K+ que subyacen al potencial de acción. Sin embargo, otros canales están regulados por señales químicas que se unen a los dominios intracelulares o extracelulares en estas proteínas y no son sensibles al voltaje de la membrana. Incluso otros son sensibles al desplazamiento mecánico o a los cambios en la temperatura. Para aumentar esta diversidad de los canales iónicos, existen algunos mecanismos que pueden producir tipos funcionalmente distintos de canales iónicos a partir de un solo gen. Los genes de los canales iónicos contienen algunas regiones codificadoras que pueden ser cortadas y empalmadas de diferentes formas, y dan origen a proteínas de canal que pueden tener propiedades funcionales notablemente distintas. También es posible montar RNA que codifica canales iónicos mediante el cambio de la composición de bases después de la transcripción a partir del gen. Por ejemplo, el montaje del RNA que codifica algunos receptores para el neurotransmisor glutamato (Capítulo 6) cambia un solo aminoácido en el interior del receptor, lo que a su vez da origen a canales que difieren en su selectividad a los cationes y en su conductancia. Las proteínas de los canales también pueden sufrir modificaciones postraduccionales, como la fosforilación por proteincinasas (véase Capítulo 7), lo que puede modificar más sus características funcionales. Por lo tanto, aunque las señales eléctricas básicas del sistema nervioso son relativamente estereotipadas, las proteínas responsables de generar estas señales son muy diversas y confieren propiedades de señalización especializada a muchos de los tipos de las células neuronales que pueblan el sistema nervioso. Estos canales también participan en una amplia gama de enfermedades neurológicas (véase Recuadro 4D). Canales iónicos con puerta de voltaje Ahora se han descrito canales iónicos con puerta de voltaje que son selectivamente permeables a cada uno de los principales iones fisiológicos: Na+, K+, Ca2+ y Cl− (Figura 4.4A-D). En efecCanales con puerta de voltaje (A) Canal del Na+ Na+ (B) Canal + del Ca2 to, se han descubierto muchos genes diferentes para cada tipo de canal con puerta de voltaje; un ejemplo es la identificación de 10 genes de los canales del Na+ humano. Este hallazgo fue inesperado porque los canales del Na+ de muchos tipos celulares diferentes tienen propiedades funcionales similares, compatibles con su origen a partir de un gen único. Sin embargo, ahora está claro que todos estos genes de los canales del Na+ (llamados genes SCN) producen proteínas que difieren en su estructura, función y distribución en tejidos específicos. Por ejemplo, además de los canales del Na+ de inactivación rápida descubiertos por Hodgkin y Huxley en el axón del calamar, se ha identificado un canal del Na+ sensible al voltaje que no se inactiva en los axones de mamíferos. Este canal da origen a una corriente de Na+ “persistente” que ayuda a regular el umbral del potencial de acción y la descarga repetitiva. Este canal sirve como diana para los anestésicos locales como la benzocaína y la lidocaína. Otras respuestas eléctricas en las neuronas implican la activación de canales del Ca2+ con puerta de voltaje (Figura 4.4B). En algunas neuronas, los canales del Ca+2 con puerta de voltaje dan origen a potenciales de acción de una forma muy similar a los canales del Na+ sensibles al voltaje. En otras neuronas, los canales del Ca2+ controlan la forma de los potenciales de acción generados fundamentalmente por cambios en la conductancia al Na+. De forma más general, al afectar las concentraciones intracelulares de Ca2+, la actividad de los canales del Ca2+ regula una gama enorme de procesos de señalamiento bioquímico en el interior de las células (véase Capítulo 7). Tal vez el proceso encefálico más importante regulado por canales del Ca2+ sensibles al voltaje sea la liberación de neurotransmisores en las sinapsis (véase Capítulo 5). Dadas estas funciones cruciales, quizás no sea sorprendente que se hayan identificado 10 genes diferentes para los canales del Ca2+ (denominados genes CACNA). Al FIGURA 4.4 Tipos de canales iónicos con puerta de voltaje. Los ejemplos de canales con puerta de voltaje incluyen aquellos con permeabilidad selectiva a Na+ (A), Ca2+ (B), K+ (C) y Cl− (D). Los canales iónicos con puerta de ligando incluyen aquellos que son activados por la presencia extracelular de neurotransmisores, como glutamato (E). Otros canales con puerta de ligando son activados por segundos mensajeros intracelulares, como Ca2+ (F) o nucleótidos cíclicos, cAPM y cGMP (G). Canales con puerta de ligando (C) Canal del K+ Ca2+ (D) Canal del Cl– Cl– (E) Receptor de neurotransmisor Na+ (F) Canal del K+ activado por Ca2+ (G) Canal con puerta de nucleótido cíclico Na+ Glutamato Exterior cAMP Sensor de voltaje K+ K+ Ca2+ K+ cGMP cAMP Interior K+ 50 30 0 –30 –60 –90 – 120 (C) HERG –120 mV +50 mV Conductancia al K+ Corriente de K+ (µA) Corriente de K+ (µA) (B) KV4.1 300 +50 mV +50 mV Shaw Conductancia al K+ 10 mM Ca2+ +50 mV 1 mM Ca2+ +50 mV –120 mV pH 8 +50 mV pH 6 +50 mV 100 200 Tiempo (ms) 300 Conductancia al K+ Corriente de K+ (µA) –120 mV 0 –100 0 100 Potencial de membrana (mV) 1 Conductancia al K+ Corriente de K+ (µA) (F) 2–P +50 mV Corriente de K+ (µA) (E) Activado por calcio 0 1 –120 mV (D) Rectificador hacia adentro 1 0 –100 0 100 Potencial de membrana (mV) –120 mV Corriente de K+ (µA) (A) KV2.1 100 200 Tiempo (ms) Conductancia al K+ 0 Conductancia al K+ Potencial de membrana (mV) CANALES IÓNICOS Y TRANSPORTADORES 0 –100 0 100 Potencial de membrana (mV) 65 igual que los canales del Na+, los del Ca2+ difieren en sus propiedades de activación e inactivación, lo que permite variaciones sutiles en los procesos de señalamiento eléctrico y químico mediados por Ca2+. En consecuencia, los fármacos que bloquean los canales del Ca2+ con puerta de voltaje son especialmente útiles en el tratamiento de distintos trastornos que varían desde la cardiopatía hasta los trastornos de ansiedad. Como mucho, la clase más grande y más diversa de los canales iónicos con puerta de voltaje son los canales del K+ (Figura 4.4C). En la actualidad se conocen casi 100 genes de los canales del K+ y pertenecen a varios grupos distintos que difieren sustancialmente en sus propiedades de activación, apertura e inactivación. Algunos tardan minutos en inactivarse, como el caso de los canales del K+ del axón del calamar (Figura 4.5A). Otros se inactivan en milisegundos, como es típico de los canales del Na+ con puerta de voltaje (Figura 4.5B) y algunos son incluso más rápidos (Figura 4.5C). Estas propiedades influyen en la duración y la frecuencia de descarga de los potenciales de acción, con importantes consecuencias para la conducción axónica y la transmisión sináptica. Incluso otros canales del K+ responden a la hiperpolarización de la membrana (Figura 4.5D) o a las concentraciones intracelulares de Ca2+ (Figura 4.5E). Los canales del K+ 2-P, que son responsables primariamente del potencial de reposo de las neuronas (véase Capítulo 2), tienen poca puerta dependiente del voltaje pero son sensibles a las señales químicas (Figura 4.5F). 1 FIGURA 4.5 Propiedades diversas de los canales del K+. Se expresaron diferentes tipos de canales del K+ en ovoci0 –100 0 100 Potencial de membrana (mV) 1 10 +M Ca2+ 1 +M Ca2+ 0 –100 0 100 Potencial de membrana (mV) 1 pH 8 pH 6 0 –100 0 100 Potencial de membrana (mV) tos de Xenopus (véase Recuadro 4C); se utilizó el método de pinzamiento de voltaje para modificar el potencial de membrana (arriba) y medir las corrientes resultantes que fluyen a través de cada tipo de canal. Estos canales del K+ varían mucho en sus propiedades de compuerta, como se aprecia en sus corrientes (izquierda) y conductancias (derecha). (A) Los canales KV2.1 tienen poca inactivación y están estrechamente relacionados con los canales del K+ de rectificador diferido que participan de la repolarización del potencial de acción. (B) Los canales KV4.1 se inactivan durante una despolarización. (C) Los canales HERG se inactivan tan rápidamente que la corriente sólo fluye cuando se elimina con rapidez la inactivación al final de una despolarización. (D) Los canales del K+ de rectificación hacia el interior permiten que fluya más corriente de K+ en potenciales hiperpolarizados que en despolarizados. (E) Los canales del K+ activados por Ca2+ se abren en respuesta a los iones Ca2+ intracelulares y, en algunos casos, a la despolarización de la membrana. (F) Los canales del K+ con dos poros habitualmente responden a señales químicas, como pH, más que a cambios en el potencial de membrana. En el caso de los canales TASK que se muestran aquí, los cambios en el pH extracelular regulan la apertura de los canales, mientras que los cambios en el potencial de membrana no lo hacen. CAPÍTULO 4 Por último, se han identificado varios tipos de canales del Cl− con puerta de voltaje (véase Figura 4.4D). Estos canales están presentes en todos los tipos de neuronas, donde controlan la excitabilidad, contribuyen al potencial de membrana de reposo y ayudan a regular el volumen celular. Canales iónicos con puerta de ligando Muchos tipos de canales iónicos responden a señales químicas (ligandos) más que a cambios en el potencial de membrana (Figura 4.4E-G). El más importante de estos canales iónicos con puerta de ligando en el sistema nervioso es la clase activada por la fijación de neurotransmisores (Figura 4.4E). Estos canales son esenciales para la transmisión sináptica y otras formas de fenómenos de señalamiento célula a célula analizados en los Capítulos 5-7. Aunque típicamente los canales iónicos con puerta de voltaje que subyacen al potencial de acción sólo permiten la permeabilidad a un ion, los canales activados por ligandos extracelulares suelen ser menos activos y, a menudo, permiten el flujo de dos o más tipos de iones. Otros canales con puerta de ligando son sensibles a las señales químicas que se originan en el citoplasma de las neuronas (véase Capítulo 7) y pueden ser selectivos para iones específicos como K+ o Cl−, o permeables a todos los cationes fisiológicos. Estos canales se distinguen por dominios de fijación del ligando en sus superficies intracelulares que interactúan con segundos mensajeros como Ca2+, los nucleótidos cíclicos cAMP o cGMP o protones. Los ejemplos de canales que responden a las señales intracelulares incluyen los canales del K+ activados por Ca2+ (Figura 4.4F), el canal de cationes con puerta de nucleótidos cíclicos (Figura 4.4G) y los canales iónicos que detectan ácidos (acid-sensing ion channels, ASIC). La función principal de estos canales es convertir las señales químicas intracelulares en información eléctrica. Este proceso es de particular importancia en la transducción sensitiva, donde las señales de los segundos mensajeros intracelulares asociadas a los estímulos sensitivos –tales como los olores y la luz– son traducidas en señales eléctricas por los canales con puerta de nucleótidos cíclicos. Aunque muchos de estos canales iónicos con puerta de ligando se localizan en la membrana de la superficie celular, otros están en membranas de los orgánulos intracelulares como las mitocondrias o el retículo endoplasmático. Algunos de estos últimos canales son selectivamente permeables al Ca2+ y regulan su liberación desde el interior del retículo endoplasmático hacia el citoplasma, donde este segundo mensajero puede desencadenar entonces un espectro de respuestas intracelulares como las descritas en el Capítulo 7. Canales activados por el estiramiento y el calor Otros canales iónicos responden al calor o a la deformación de la membrana. Los canales iónicos activados por el calor, como algunos miembros de la familia de genes potenciales de recep- tores transitorios (TRP), contribuyen a las sensaciones de dolor y temperatura y ayudan a mediar la inflamación (véase Capítulo 10). A menudo, estos canales se especializan en detectar cambios específicos de la temperatura y algunos son activados incluso por el frío. Otros canales iónicos responden a la distorsión mecánica de la membrana plasmática y son la base de los receptores de estiramiento y los reflejos de estiramiento neuromuscular (véanse Capítulos 9, 16 y 17). Al parecer, una forma especializada de estos canales permite la audición al posibilitar que las células ciliadas auditivas respondan a las ondas sonoras (véase Capítulo 13). En resumen, esta extraordinaria variedad de canales iónicos permite que las neuronas generen señales eléctricas en respuesta a los cambios del potencial de membrana, las aferencias sinápticas, los segundos mensajeros intracelulares, la luz, los olores, el calor, el sonido, el tacto y muchos otros estímulos. Estructura molecular de los canales iónicos El conocimiento de la estructura física de los canales iónicos es de obvia importancia para discernir el modo en que realmente funcionan. Hasta época reciente, la mayor parte de la información acerca de la estructura de los canales iónicos se obtenía de manera indirecta de estudios de la composición de aminoácidos y las propiedades fisiológicas de estas proteínas. Por ejemplo, se ha aprendido mucho al explorar las funciones de aminoácidos particulares en el interior de las proteínas utilizando la mutagénesis y la expresión de las proteínas de los canales mutantes en células no neuronales, como ovocitos de Xenopus (véase Recuadro 4C). Estos estudios han descubierto una arquitectura transmembrana general común a todas las principales familias de canales iónicos. Estas moléculas son todas las proteínas integrales de la membrana que atraviesan repetidas veces la membrana plasmática. Las proteínas de los canales del Na+ y del Ca2+ consisten en segmentos repetidos de seis regiones que atraviesan la membrana y que se repiten 4 veces, con un total de 24 regiones transmembrana (Figura 4.6A, B). Los canales del Na+ o del Ca2+ están constituidos por una sola de estas proteínas, aunque otras proteínas accesorias, denominadas subunidades β, pueden regular la función de estos canales. Típicamente, las proteínas de los canales del K+ atraviesan la membrana seis veces (Figura 4.6C), aunque existen algunos canales del K+ que lo hacen sólo dos veces (Figura 4.6D) y otros que la atraviesan cuatro (Figura 4.6F) o siete veces (Figura 4.6E). Cada una de estas proteínas de los canales del K+ sirve como una subunidad del canal y, característicamente, cuatro de estas subundidades se agregan para forFIGURA 4.6 Topología de las principales subunidades de canales del Na+, Ca2+, K+ y Cl− con puerta de voltaje. Los segmentos repetidos ▼ 66 de los canales del Na+ (A) y Ca2+ (B) están marcados I, II, III y IV; (C-F) los canales del K+ son más diversos. En todos los casos, se combinan cuatro subunidades para formar un canal funcional. (G) Los canales de cloruro son estructuralmente distintos de todos los otros canales con puerta de voltaje. CANALES IÓNICOS Y TRANSPORTADORES 67 poro es muy apropiada para conducir iones K+; la porción más estrecha está cerca de la boca externa del canal y está tan contraída que sólo un ion K+ no hidratado puede ajustarse a través del cuello de botella (Figura 4.7C). Los cationes más grandes, como Cs+, no pueden atravesar esta región del poro y los cationes más pequeños, como el Na+, no pueden entrar en el poro porque las “paredes” de este están muy separadas como para estabilizar un ion Na+ deshidratado. Esta parte del complejo del canal es responsable de la permeabilidad selectiva al K+ y, por lo tanto, se denomina filtro de selectividad. A mayor profundidad, dentro del canal hay una cavidad llena de agua que conecta con el interior de la célula. Evidentemente, esta cavidad recoge K+ del citoplasma y, utilizando cargas negativas de la proteína, permite que los iones K+ sean deshidratados a medida que ingresan en el filtro de selectividad. Entonces, estos iones “desnudos” son capaces de atravesar cuatro sitios de fijación de K+ dentro del filtro de selectividad para alcanzar finalmente el espacio extracelular (recuérdese que el gradiente de concentración normal impulsa el K+ fuera de células). La presencia de múltiples (hasta cuatro) iones K+ en el interior del filtro de selectividad produce la repulsión electrostática entre los iones, que ayuda a acelerar su tránsito a través del filtro de selectividad y permite así el flujo rápido de iones a través del canal. mar un único canal iónico funcional. La mayoría de los canales iónicos con puerta de voltaje también contienen un tipo distinto de hélice transmembrana que posee algunos aminoácidos con carga positiva y sirve como sensor de voltaje que detecta cambios en el potencial eléctrico a través de la membrana (estructuras amarillas de la Figura 4.6). Rod MacKinnon, en la Rockefeller University, adquirió información directa acerca de las características estructurales de los canales iónicos de estudios de cristalografía de rayos X de canales del K+ de las bacterias (Figura 4.7). La estructura del poro del canal del K+ fue demostrada en estudios de un canal del K+ de las bacterias, el cual fue elegido para el análisis porque la gran cantidad de proteínas del canal necesaria para la cristalografía podía obtenerse haciendo proliferar gran cantidad de bacterias. Los resultados de MacKinnon demostraron que el canal está formado por subunidades que atraviesan dos veces la membrana plasmática; entre estas dos estructuras helicoidales que atraviesan la membrana, existe un asa del poro que se inserta en la membrana plasmática (Figura 4.7A). Se reúnen cuatro de estas subunidades para formar un canal (Figura 4.7B). En el centro del canal montado, es posible observar el poro del canal como un túnel estrecho a través de la proteína que permite que el K+ fluya por la proteína y atraviese así la membrana. La estructura del (A) Canal del Na+ (B) Canal del Ca2+ I II III IV I Subunidad ` N II III IV N C C Subunidad ` C N C N ` Subunidad N C Canales del K+ (E) Activados por Ca2+ (D) Rectificador hacia adentro (C) Kv y HERG (G) Canal del Cl– (F) 2-P N N C C N C N C N N C Subunidad ` C 68 CAPÍTULO 4 (A) (B) Filtro de selectividad Asa del poro Ion K+ en el poro Hélice del poro Hélice externa Hélice externa Hélice interna Hélice interna (C) Filtro de selectividad Iones K+ FIGURA 4.7 Estructura de un canal del K+ bacteriano simple determinada por cristalografía. (A) Estructura de una subunidad del canal, que consiste en los dominios de expansión de la membrana y un asa del poro que se inserta en esta. (B) Disposición tridimensional de cuatro subunidades (cada una de un color diferente) en el interior del poro del canal. (C) La vía de permeabilidad del canal de K+ consiste en una cavidad acuosa grande conectada a un filtro de selectividad estrecho. Los dominios helicoidales del canal señalan cargas negativas (rojo) hacia esa cavidad, lo que permite que los iones K+ (verde) se deshidraten y luego atraviesen el filtro de selectividad. (De Doyle y cols., 1998). Cavidad llena de agua Poro Hélice del poro con carga negativa Otros estudios cristalográficos de un canal del K+ con puerta de voltaje del mamífero han arrojado muchas ideas estructurales sobre cómo ocurre la apertura dependiente del voltaje de los canales iónicos. Como sucede con el canal del K+ de las bacterias descrito antes, cuatro subunidades se reúnen para formar el canal del K+ con puerta del voltaje (Figura 4.8A). La región del poro central de este canal es muy similar a la de los canales del K+ de las bacterias y consiste en dos estructuras que atraviesan la membrana y asas de poro a las que contribuyen cada una de las cuatro subunidades (Figura 4.8B). Este canal con puerta de voltaje tiene estructuras adicionales en su cara citoplasmática, como una subunidad β y un dominio T1 que liga la subunidad β con el canal (véase Figura 4.8A). Aun más importante, cada subunidad de canal tiene cuatro estructuras de membrana adicionales que forman los sensores de voltaje de este canal. Estos sensores de voltaje pueden observarse como dominios separados que se extienden en la membrana plasmática y están vinculados con el poro central de canal (véase Figura 4.8B). Las cargas positivas en el interior de estos sensores de voltaje permiten el movimiento dentro de la membrana en respuesta a los cambios en el potencial de membrana: la despolarización empuja los sensores hacia afuera, mientras que la hiperpolarización tracciona hacia adentro. Estos movimientos de los sensores ejercen entonces fuerza sobre los vinculadores helicoidales que conectan los sensores al poro del canal, traccionando el poro del canal para abrirlo o empujándolo para cerrarlo (Figura 4.8C). Por lo tanto, la puerta dependiente de voltaje de los canales iónicos ahora puede comprenderse en términos estructurales. Los movimientos precisos del sensor de voltaje que ocurren durante la despolarización de la membrana todavía no están claros y son tema de un considerable debate. Una hipótesis es que la despolarización ocasiona que el sensor con forma de silla de montar gire de un lado de la membrana al otro (Figura 4.8D). En resumen, los canales iónicos son proteínas integrales de la membrana con estructuras características que les permiten conducir iones, detectar el potencial transmembrana, inactivarse y unirse a distintas neurotoxinas. La caracterización detallada de la estructura de los canales iónicos ha proporcionado un considerable conocimiento acerca del modo en que los iones son conducidos desde un lado de la membrana plasmática al otro, el modo en que un canal puede ser selectivamente permeable a un ion, cómo son capaces de detectar cambios en el voltaje de la membrana y cómo regulan la apertura de sus poros. Es probable que otros tipos de canales iónicos sean similares en su arquitectura funcional. Este tipo de investigación también ha aclarado el modo en que las CANALES IÓNICOS Y TRANSPORTADORES 69 (B) (A) Transmembrana Poro K+ Dominio T1 Sensor de voltaje Sensores de voltaje 2,5 nm Su bunidad ` 2,5 nm Región del poro central mutaciones en los genes de los canales iónicos pueden conducir a distintos trastornos neurológicos (Recuadro 4D). (C) Los transportadores activos crean y mantienen los gradientes iónicos Hiperpolarizado Despolarizado Ligador helicoidal Poro cerrado (D) Hiperpolarizado Poro abierto Despolarizado Sensor de voltaje Dentro de la célula Poro cerrado Poro abierto Hasta este punto, la explicación de la base molecular del señalamiento eléctrico ha dado por supuesto el hecho de que las células nerviosas mantienen gradientes de concentración iónica a través de sus membranas de superficie. Sin embargo, ninguno de los iones de importancia fisiológica (Na+, K+, Cl− y Ca2+) se encuentra en equilibrio electroquímico. Dado que los canales producen efectos eléctricos al permitir que uno o más de estos iones difundan a favor de sus gradientes electroquímicos, existiría una disipación gradual de los gradientes de concentración a menos que las células nerviosas pudieran restablecer los iones desplazados durante el flujo de corriente que ocurre como resultado del señalamiento nervioso y el escape iónico continuo que ocurre en reposo. Las tareas de generar y mantener gradientes de concentración iónica para iones particulares son llevadas a cabo por un grupo de proteínas de la membrana plasmática conocido como transportadores activos. Los transportadores activos realizan esta tarea formando complejos con los iones que están translocando. Típicamente, el proceso de fijación y liberación de los iones para su transporte requiere varios milisegundos. En consecuencia, la translocación de los iones por los transportadores activos es mucho más lenta que FIGURA 4.8 Estructura de un canal del K+ con puerta del voltaje de un mamífero. (A) El canal incluye cuatro subunidades (en diferentes colores), donde cada una posee un dominio transmembrana y un dominio T1. Hay una subunidad β fijada a cada dominio T1. (B) Cuando el dominio transmembrana se visualiza desde arriba, puede observarse que tiene dominios separados para la detección del voltaje y para formar el poro conductor del K+ (K+ indicado por las esferas negras en el centro del poro). (C) Modelo para puerta dependiente de voltaje del canal del K+. Superior: estructura del dominio del poro central del canal del K+ con puerta de voltaje en estado abierto (despolarizado). Inferior: la despolarización empuja el sensor de voltaje hacia la superficie extracelular de la membrana, traccionando del ligador (rojo) y abriendo así el poro del canal (azul). Por el contrario, la hiperpolarización tira del sensor hacia abajo, empujando sobre este ligador y cerrando el poro del canal. (D) Modelo para movimiento de sensor de voltaje. La despolarización hace que el sensor de voltaje similar a una silla de montar se mueva hacia la superficie extracelular de la membrana, mientras que la hiperpolarización hace que se mueva hacia la superficie intracelular. (A, B, de Long y cols., 2005; C, de Tao y cols., 2010; D, de Lee, 2006.) 70 CAPÍTULO 4 RECUADRO 4D Enfermedades causadas por alteración de los canales iónicos Varias enfermedades genéticas, llamadas en conjunto canalopatías, son el resultado de alteraciones pequeñas pero críticas en los genes de los canales iónicos (Figura A). Entre estas enfermedades, las que mejor se han descrito son las que afectan las células del músculo esquelético. En estos trastornos, las alteraciones en las proteínas de los canales iónicos producen miotonía (rigidez muscular debida a una excitabilidad eléctrica excesiva) o parálisis (debida a excitabilidad muscular insuficiente). Otros trastornos surgen por defectos de los canales iónicos en corazón, riñón y oído interno. Es mucho más difícil estudiar las canalopatías asociadas con los canales iónicos localizados en el encéfalo. No obstante, los canales del Ca2+ con puerta de voltaje han sido involucrados recientemente en una gama de enfermedades neurológicas que incluyen ataxia episódica, degeneración espinocerebelosa, ceguera nocturna y cefaleas migrañosas. La migraña hemipléjica familiar se caracteriza por crisis de migraña que característicamente duran entre uno y tres días. Durante estos episodios, los pacientes experimentan cefaleas intensas y vómitos. Se han identificado varias mutaciones en un canal de Ca2+ humano en familias con migraña hemipléjica familiar y cada una de ellas tiene diferentes síntomas clínicos. Por ejemplo, una mutación en la región formada por poros del canal produce migraña hemipléjica con ataxia cerebelosa progresiva, mientras que otras mutaciones producen sólo los síntomas habituales de la migraña hemipléjica familiar. Se desconoce de qué modo las propiedades alteradas de los canales del Ca2+ conducen a las crisis de migraña. La ataxia episódica de tipo 2 es un trastorno neurológico en el cual los individuos afectados sufren crisis recurrentes de movimientos anormales de las extremidades y ataxia grave. Estos problemas a veces se acompañan de vértigo, náuseas y cefalea. En general, las crisis son precipitadas por estrés emocional, ejercicio o alcohol y duran algunas horas. Las mutaciones en la ataxia episódica tipo 2 hacen que los canales del Ca2+ se vean truncados en varios sitios, lo cual puede producir las manifes- taciones clínicas de la enfermedad al impedir el ensamblaje normal de los canales del Ca2+ en la membrana. La ceguera nocturna estacionaria congénita ligada al X es un trastorno retiniano progresivo que produce ceguera nocturna, disminución de la agudeza visual, mio- (A) Canal del Ca2+ I II III IV C N FHM EA2 CSNB Parálisis Canal del Na+ I II III IV N N C C Subunidad ` C N GEFS Miotonía Parálisis Canal del Cl– Canal del K+ (A) Mutaciones genéticas en canales del Ca2+, canales del Na+, canales del K+ y canales del Cl− que dan por resultado enfermedades. Los símbolos en rojo indican los sitios y las patologías específicas de estas mutaciones. (De Lehmann-Horn y Jurkat-Kott, 1999.) C N EA1 BFNC Miotonía C CANALES IÓNICOS Y TRANSPORTADORES 71 RECUADRO 4D (continúa) pía, nistagmo y estrabismo. La ceguera nocturna estacionaria congénita completa hace que los bastones, fotorreceptores retinianos sean no funcionantes. La forma incompleta produce un funcionamiento subnormal (pero mensurable) de conos y bastones. Al igual que la ataxia episódica tipo 2, el tipo incompleto de ceguera nocturna estacionaria congénita es causado por mutaciones que truncan los canales del Ca2+. La función retiniana anormal puede ser el resultado de una disminución de las corrientes de Ca2+ y de la liberación 40 0 <40 <80 Tipo salvaje Corriente del Na+ (nA) Potencial de membrana (mV) (B) Mutantes de los canales del Na 0 5 Tiempo (ms) (B) Las mutaciones en los canales del Na+ retardan la velocidad de inactivación de las corrientes de Na+. (De Barchi, 1995.) 10 del neurotransmisor desde los fotorreceptores (véase Capítulo 11). Un defecto en los canales del Na+ del encéfalo produce la epilepsia generalizada con convulsiones febriles que comienza en el lactante y habitualmente continúa hasta los primeros años de la pubertad. Este defecto ha sido mapeado a 2 mutaciones: una en el cromosoma 2 que codifica una subunidad β para un canal del Na+ con puerta de voltaje y la otra en el cromosoma 19 que codifica una subunidad β del canal del Na+. Estas mutaciones retardan la inactivación del canal del Na+ (Figura B), lo cual puede explicar la hiperexcitabilidad neuronal subyacente a la epilepsia generalizada con convulsiones febriles. Otro tipo de convulsiones, la convulsión neonatal familiar benigna, se debe a mutaciones en los canales del K+. Esta enfermedad se caracteriza por convulsiones breves y frecuentes que comienzan en la primera semana de vida y desaparecen espontáneamente en algunos meses. La mutación se ha mapeado por lo menos a dos genes de los canales del K+ con puerta de voltaje. Una reducción en el flujo de corriente de K+ a través de los canales mutados probablemente explica la hiperexcitabilidad asociada con este defecto. Una enfermedad relacionada, la ataxia episódica tipo 1, ha sido relacionada con un defecto en otro tipo de canal del K+ con puerta de voltaje. La ataxia episódica el movimiento iónico a través de los canales (recuérdese que los canales iónicos pueden conducir miles de iones a través de una membrana cada milisegundo). En resumen, los transportadores activos almacenan energía gradualmente en forma de gradientes de concentración de los iones, mientras que la apertura de los canales iónicos disipa con rapidez esta energía almacenada durante eventos relativamente breves de señalamiento eléctrico. Se han identificado varios tipos de transportadores activos (Figura 4.9). Aunque las tareas específicas de estos transportadores difieren, todos deben translocar iones en contra de sus gradientes electroquímicos. Esta movilización de iones cuesta arriba requiere el consumo de energía, y los transportadores neuronales pueden ser clasificados en dos clases sobre la base de su fuente de energía. Las bombas ATPasas adquieren energía directamente por la hidrólisis del ATP. El ejemplo más sobresaliente de una bomba ATPasa es la bomba de Na+ (o más correctamente, la bomba ATPasa de Na+/K+), que es responsable de mantener los gra- tipo 1 se caracteriza por episodios breves de ataxia. Los canales mutantes inhiben la función de otros canales de K+ no mutantes y pueden producir síntomas clínicos al deteriorar la repolarización del potencial de acción. Las mutaciones en los canales del K+ del músculo cardíaco son responsables del latido cardíaco irregular de los pacientes con síndrome Q-T prolongado. Muchos trastornos genéticos afectan los canales con puerta de voltaje del músculo esquelético y son responsables de múltiples enfermedades musculares que producen debilidad muscular (parálisis) o contracción muscular (miotonía). Bibliografía Ashcroft, F. M. (2000) Ion Channels and Disease. Boston: Academic Press. Barchi, R. L. (1995) Molecular pathology of the skeletal muscle sodium channel. Annu. Rev. Physiol. 57: 355-385. Escayg, A., and A. L. Goldin (2010) Sodium channel SCN1A and epilepsy: Mutations and mechanisms. Epilepsia 51: 1650-1658. Jurkat-Rott, K., H. Lerche, Y. Weber and F. Lehmann-Horn (2010) Hereditary channelopathies in neurology. Adv. Exp. Med. Biol. 686: 305-334. Khosravani, H. and G. W. Zamponi (2006) Voltage-gated calcium channels and idiopathic generalized epilepsies. Physiol. Rev. 86: 941-966. Lehmann-Horn, F. and K. Jurkat-Rott (1999) Voltage-gated ion channels and hereditary disease. Physiol. Rev. 79: 1317-1372. Pietrobon, D. (2010) CaV2.1 channelopathies. Pflügers Arch. 460: 375-393. dientes de concentración transmembrana de Na+ y de K+ (Figura 4.9A). Otra es la bomba de Ca2+, que proporciona los principales mecanismos para eliminar el Ca2+ de las células (Figura 4.9B). Se han identificado dos tipos diferentes de bomba de Ca2+. Uno, denominado PMCA, se encuentra sobre la membrana plasmática; el otro, denominado SERCA, es utilizado para almacenar Ca2+ en el interior del retículo endoplasmático (véase Capítulo 7). Una segunda clase de transportador activo no utiliza directamente ATP, sino que, en cambio, depende de los gradientes electroquímicos de otros iones como fuente de energía. Este tipo de transportador transporta uno o más iones en contra de su gradiente electroquímico mientras toma simultáneamente otro ion (sobre todo, Na+) a favor de su gradiente. A su vez, estos transportadores pueden ser subdivididos en dos tipos según la dirección del movimiento de iones. Como lo indica el nombre del primer tipo, los intercambiadores de iones intercambian iones intracelulares y extracelulares. Un ejemplo de uno de es- 72 CAPÍTULO 4 Bombas ATPasas Intercambiadores de iones (A) Bomba de Na+/K+ (B) Bomba de Ca 2+ (C) Intercambiador (D) Intercambia- (E) Cotransportador Na+/Ca2+ Na+/K+/Cl– dor Na+/H+ H+ K+ Cotransportadores Na+ Na Na+ K+ (F) Cotransportador (G) Cotransportador Na+/neuroK+/Cl< transmisores Na+ Cl< GABA, Dopamina Exterior Interior ADP Na+ ATP ADP Ca2+ Ca2+ H+ K+ Cl< ATP FIGURA 4.9 Ejemplos de transportadores activos. Algunos transportadores (A,B) son impulsados por la hidrólisis de ATP (bombas ATPasas), mientras que otros utilizan los gradientes electroquímicos de los iones cotransportados como fuente de energía. Estos últimos pueden ser subdivididos a su vez en intercambiadores de iones que intercambian iones a ambos lados de la membrana (C, D) y cotransportadores que transportan múltiples iones en la misma dirección (E-G). tos transportadores es el intercambiador Na+/Ca2+, que comparte con la bomba de Ca2+ la importante tarea de mantener bajas las concentraciones intracelulares de Ca2+ (Figura 4.9C). Otro intercambiador de iones, el intercambiador Na+/H+, regula el pH intracelular (Figura 4.9D). Los transportadores del segundo tipo, los cotransportadores, trabajan transportando múltiples iones en la misma dirección. Dos de estos cotransportadores regulan la concentración intracelular de Cl− al translocar Cl− junto con el Na+ o el K+ extracelular; son el cotransportador Na+/K+/Cl−, que transporta Cl− junto con Na+ y K+ en las células (Figura 4.9E), y el cotransportador K+/Cl−, que intercambia Cl− intracelular por K+ extracelular (Figura 4.9F). Como veremos en el Capítulo 6, los neurotransmisores son transportados en las terminaciones sinápticas y células gliales por medio de otros cotransportadores (Figura 4.9G). Aunque el gradiente electroquímico del Na+ (o de otros iones contrarios) es la fuente próxima de energía para los intercambiadores de iones y los cotransportadores, estos gradientes por último dependen de la hidrólisis de ATP por las bombas ATPasas, como la bomba Na+/K+ATPasa. Propiedades funcionales de la bomba Na+/K+ El transportador activo mejor conocido es la bomba Na+/K+. Su importancia fundamental para la función encefálica es evidente a partir del hecho de que este transportador representa el 20-40% del consumo de energía del encéfalo. La bomba de Na+ fue descubierta en las neuronas en la década de 1950, cuando Richard Keynes, en la Cambridge University, utilizó Na+ radiactivo para demostrar el flujo hacia el exterior (eflujo) de Na+ bomba-dependiente desde los axones gigantes del calamar. Keynes y cols. observaron que este eflujo cesaba cuando se interrumpía el aporte de ATP en el axón por el tratamiento con venenos metabólicos (Figura 4.10A, punto 4). Otras condiciones que reducen el ATP intracelular también impiden el eflujo de Na+. Los experimentos demostraron que la eliminación del Na+ intracelular necesita del metabolismo celular. Otros estudios con K+ radiactivo demostraron que el eflujo de Na+ se asocia con el influjo simultáneo de K+ ATP-dependiente. Estos flujos opuestos de Na+ y de K+ son operativamente separables: la eliminación del K+ externo reduce mucho el eflujo de Na+ (Figura 4.10, punto 2) y viceversa. Estos movimientos de Na+ y de K+ dependientes de la energía implicaban una bomba de Na+/ K+ que hidrolizaba ATP en la generación de los gradientes transmembrana de Na+ y K+. Se considera que la bomba envía alternativamente los iones Na+ y K+ a través de la membrana en un ciclo impulsado por la transferencia de un grupo fosfato del ATP a la proteína de la bomba, lo que cambia la bomba desde eliminar el Na+ intracelular hasta acumular K+ intracelular (Figura 4.10B). Algunos estudios cuantitativos de los movimientos de Na+ y K+ indican que los dos iones son bombeados a velocidades idénticas. La velocidad de influjo de K+ es sólo aproximadamente dos tercios de la velocidad de eflujo de Na+. Por lo tanto, la bomba transporta 2 K+ hacia el interior de la célula por cada 3 Na+ extraídos (véase Figura 4.10B). Esta estequiometría produce una pérdida neta de un ion con carga positiva desde el interior de la célula con cada ronda de bombeado, lo que indica que la bomba genera una corriente eléctrica que puede hiperpolarizar el potencial de membrana. Por esta razón, se dice que la bomba de Na+/K+ es electrogénica. Sin embargo, puesto que las bombas actúan mucho más lentamente que los canales iónicos, la corriente producida por la bomba de Na+/K+ es muy pequeña. Por ejemplo, en el axón del calamar, la corriente eléctrica generada por la bomba es menos del 1% de la corriente que fluye a través de los canales del Na+ con puerta de voltaje y afecta el potencial de membrana de reposo en sólo un milivoltio o menos. Estructura molecular de las bombas ATPasas Estas observaciones implican que la bomba de Na+/K+ debe mostrar varias propiedades moleculares. Primero, debe fijar CANALES IÓNICOS Y TRANSPORTADORES FIGURA 4.10. Movimientos iónicos debi- (A) 1 Eflujo de Na+ 3 Recuperación cuando 5 Recuperación cuando se restablece el K+ 2 Eflujo de Na+ reducido por la eliminación del K+ externo se restablece el ATP 4 Eflujo disminuido por los inhibidores metabólicos, como dinitrofenol, que bloquean la síntesis de ATP dos a la bomba de Na+/K+. (A) Medición del eflujo de Na+ radiactivo del axón gigante de un calamar. Este eflujo depende del K+ externo y el ATP intracelular. (B) Modelo para el movimiento de los iones por la bomba de Na+/K+. Los movimientos activos del Na+ y el K+ son impulsados por el ATP, el cual fosforila Eflujo de Na+ (escala logarítmica) la bomba. La fosforilación cambia la bomba de exportar Na+ a importar K+. Estos flujos de iones son asimétricos, con 3 Na+ llevados hacia afuera y 2 K+ hacia adentro. (A, de Hodgkin y Keynes, 1955; B, de Lingrel y cols., 1994.) 0 (B) 73 50 100 150 Tiempo (min) 200 250 300 K+ que explica estas tres propiedades. Esta bomba es una proteína grande e integral de la membrana, 1 Fijación de Na+ 2 Fosforilación formada por lo menos por dos subunidades, denominadas α y β. La secuencia primaria muestra que Exterior ADP la subunidad α atraviesa 10 veces la membrana, y ATP la mayor parte de la molécula se encuentra sobre el lado citoplasmático, mientras que la subunidad β atraviesa la membrana sólo una vez y es predominantemente extracelular. El análisis de su secuencia Interior de aminoácidos ha identificado estructuras involuNa+ cradas en algunas de las funciones de la bomba (Figura 4.11B). Un dominio intracelular, el dominio de fijación de nucleótidos, es necesario para la fijación y la hidrólisis del ATP, y se ha identificado el ami+ + noácido fosforilado por el ATP. Un dominio extraK Na celular representa el sitio de fijación de la ouabaína. El análisis cristalográfico de la bomba de Na+/K+ muestra estas estructuras con mucho mayor detalle (Figura 4.11C). Lo que es más importante es que estos análisis ponen en evidencia que dos K+ se unen en el interior de la bomba, dentro del dominio de expansión de la membrana. Presumiblemente, ello K+ refleja la conformación de la bomba cuando está translocando K+ al interior de la célula. 4 El cambio conformacional 3 El cambio conformacional inducido por la desfosforilación produce liberación de Na+ Se han obtenido otros conocimientos sobre la conduce a la liberación de K+ y fijación de K+ base estructural de la translocación de iones para una Ca2+ATPasa que está estrechamente relacionada con la bomba de Na+/K+. Esta bomba utiliza la + + tanto Na como K ; segundo, debe poseer sitios que fijen ATP hidrólisis del ATP para impulsar la translocación del Ca2+ desde y reciban un grupo fosfato de este ATP; finalmente, debe fijar el citoplasma a través de la membrana del retículo sarcoplasmáouabaína, la toxina que bloquea esta bomba (Figura 4.11A). Dis- tico, una organela del músculo de almacenamiento intracelular tintos estudios han identificado la estructura de la bomba de Na+/ de Ca2+ que es análoga al retículo endoplasmático. La bomba 74 CAPÍTULO 4 (A) (B) Sitio de fijación de la ouabaína (C) Sitio de fijación de la ouabaína 2 K+ Exterior Fijación de Na+ y K+ C Exterior Subunidad ` Exterior Membrana Membrana Membrana Interior Interior ATP ADP C N + 3 Na+ Subunidad _ Sitio de fosforilación Sitio de fijación de ATP Interior Subunidad ` N Subunidad _ P FIGURA 4.11 Propiedades moleculares de la bomba de Na+/K+. (A) Características generales de la bomba. (B) La subunidad α atraviesa 10 veces la membrana y contiene los aminoácidos importantes para la fijación de ATP, K+ y ouabaína. (C) Estructura de la bomba de Na+/K+. Se observan los dominios responsables de la fijación de nucleótidos (NB), fosforilación (P) y dominio actuador (AD). En esta conformación, el ADP ocupa el dominio NB de la bomba y pueden observarse dos K+ (cuadrado interior) en el dominio transmembrana. Un tercer K+ está unido al dominio P (flecha). (B, de Lingrel y cols., 1994; C, de Shinoda y cols., 2009.) de Ca2+, al igual que la de Na+/K+, es una proteína de membrana muy grande que atraviesa 10 veces la membrana y consiste en varios dominios (Figura 4.12A). De forma muy similar a la bomba de Na+/K+, la de Ca2+ posee un dominio de fijación de nucleótidos que fija ADP/ATP, mientras que otros dominios participan en la fosforilación de la bomba y la translocación de iones. Al igual que la bomba de Na+/K+, la fosforilación de la bomba de Ca2+ impulsa un ciclo de cambios conformacionales. Al examinar la estructura de la bomba de Ca2+ en diferentes etapas de este ciclo, se ha aclarado el mecanismo de translocación de Ca2+. El Ca2+ se une primero en el lado citoplasmático de la bomba y luego es impulsado a través de la membrana como resultado de los cambios conformacionales en los dominios de expansión de la membrana inducidos por la fosforilación, lo que finalmente conduce a que el Ca2+ sea liberado del otro lado de la membrana (Figura 4.12B). Al contrario de los canales iónicos, donde la translocación de iones ocurre por un movimiento basado en la difusión a través de un poro acuoso, la translocación de Ca2+ por la bomba ocurre por el secuestro del ion profundamente en el interior de la proteína, lejos del medio acuoso. Esto explica cómo la bomba es capaz de mover el Ca2+ presente a través de la membrana y, presumiblemente, también explica cómo la bomba de Na+/K+ genera un movimiento cuesta arriba de Na+ y K+. Resumen Los canales iónicos y los transportadores activos tienen funciones complementarias. El propósito primario de los transportadores es generar gradientes de concentración transmembrana, los cuales son explotados luego por los canales iónicos para generar señales eléctricas. Los canales iónicos son responsables de las conductancias dependientes de voltaje de las membra- AD ADP NB nas de las células nerviosas. Los canales que subyacen al potencial de acción son proteínas integrales de la membrana que abren o cierran poros selectivos para iones en respuesta al potencial de membrana, lo que permite la difusión de iones específicos a través de la membrana. El flujo de iones a través de canales abiertos únicos puede ser detectado como pequeñas corrientes eléctricas, y la apertura sincrónica de muchos de estos canales genera las corrientes macroscópicas que producen potenciales de acción. Algunos estudios moleculares muestran que estos canales con puerta de voltaje tienen estructuras altamente conservadas responsables de características, como la permeabilidad a los iones, y la sensibilidad al voltaje, así como las características que especifican la selectividad de iones y la sensibilidad a las toxinas. Otros tipos de canales son sensibles a las señales químicas, como los neurotransmisores o los segundos mensajeros, o al calor y la deformación de la membrana. La gran cantidad de genes para los canales iónicos crea canales con una amplia gama de características funcionales, lo cual permite que diferentes tipos de neuronas tengan un espectro notable de propiedades eléctricas. Las proteínas transportadoras activas son muy diferentes en estructura y función. La energía necesaria para mover los iones en contra de un gradiente de concentración (p. ej., para mantener el potencial de reposo) es proporcionada por la hidrólisis del ATP o por el gradiente electroquímico de los iones cotransportados. La bomba de Na+/K+ produce y mantiene los gradientes transmembrana del Na+ y del K+, mientras que otros transportadores son responsables de los gradientes electroquímicos para otros iones de importancia fisiológica, que incluyen Cl−, Ca2+ y H+. En conjunto, transportadores y canales proporcionan una explicación molecular razonablemente completa de la capacidad de las neuronas de generar señales eléctricas. CANALES IÓNICOS Y TRANSPORTADORES 75 (B) (A) Fosforilación Fijación de ATP Luz del retículo sarcoplasmático 2 Ca2+ Citoplasma P P NB TA NB TA ATP ADP FIGURA 4.12 Estructura molecular de la bomba de Ca2+. (A) Estructura de la bomba de Ca2+. Están indicados los dominios ATP Mg2+ ADP 2 H+ 2 Ca2 responsables de la fijación de nucleótidos (NB), la fosforilación (P) y la actividad de translocación de iones (TA). Se muestra la estructura de la bomba cuando está unida al ADP; en + 2 H+ este estado, dos Ca2+ (círculos violetas) son secuestrados en el interior de las regiones de la bomba que atraviesa la membrana. Obsérvese la similitud entre esta estructura y aquella de la bomba del Na+/K+ que se muestra en la Figura 2 Ca2 + 4.11C. (B) Secuencia hipotética de los cambios estructurales asociados con la translocación del Ca2+ por la bomba de Ca2+. Análogo a la secuencia de acontecimientos involucrados en la función de la bomba de Na+/K+ (véase Figura 4.10B), la bomba de Ca2+ sufre un ciclo de fosforilación y desfosforilación que produce cambios conformacionales que impulsan el Ca2+ a través de la membrana. (De Toyoshima y cols., 2004.) P Mg2+ Fijación de Ca2+ Lecturas adicionales Revisiones Armstrong, C. M. and B. Hille (1998) Voltage-gated ion channels and electrical excitability. Neuron 20: 371-380. Bezanilla, F. (2008) How membrane proteins sense voltage. Nature Rev. Mol. Cell Biol. 9: 323-332. Bezanilla, F. and A. M. Correa (1995) Single-channel proper-sating of Na+ and K+ channels in the squid giant axon. In Cephalopod Neurobiology, N. J. Abbott, R. Williamson and L. Maddock (eds.). New York: Oxford University Press, pp. 131-151. H2O El cambio conformacional produce liberación de Ca2+ Catterall, W. A. (2010) Ion channel voltage sensors: Structure, function, and pathophysiology. Neuron 67: 915-928. Gouaux, E. and R. Mackinnon (2005) Principles of selective ion transport in channels and pumps. Science 310: 1461-1465. Jentsch, T. J., M. Poet, J. C. Fuhrmann and A. A. Zdebik (2005) Physiological functions of CLC Cl- channels gleaned from human genetic disease and mouse models. Annu. Rev. Physiol. 67: 779-807. Kaplan, J. H. (2002) Biochemistry of Na, K-ATPase. Annu. Rev. Biochem. 71: 511-535. Lee, A. G. (2006) Ion channels: A paddle in oil. Nature 444: 697. Lingrel, J. B., J. van Huysse, W. O’Brien, E. Jewell-Motz, R. Askew and P. Schultheis (1994) Structure-function studies of the Xa, K-ATPase. Kidney Internal 45: S32-S39. 76 CAPÍTULO 4 Patapoutian, A., A. M. Peier, G. M. Story and V. Viswanath (2003). ThermoTRP channels and beyond: Mechanisms of Temperature sensation. Nature Rev. Neurosci. 4: 529-539. Swartz, K. J. (2008) Sensing voltage across lipid membranes. Nature 456: 891-897. Artículos originales importantes Boulter, J. and 6 others (1990) Molecular cloning and func-tional expression of glutamate receptor subunit genes. Science 249:1033-1037. Caterina, M. J., M. A. Schumacher, M. Tominaga, T. A. Rosen, J. D. Levine and D. Julius (1997) The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 389: 816-824. Chanda, B., O. K. Asamoah, R. Blunck, B. Roux and F. Bezanilla 2005) Gating charge displacement in voltage-gated ion channels involves limited transmembrane movement. Nature 436: 852-856. Cuello, L. G., D. M. Cortes and E. Perozo (2004) Molecular architecture of the KvAP voltage-dependent K+ channel in a lipid bilayer. Science 306: 491-495. Doyle, D. A. and 7 others (1998) The structure of the potassium channel: Molecular basis of K+ conduction and selectivity. Science 280: 69-77. Fahlke, C, H. T. Yu, C. L. Beck, T. H. Rhodes and A. L. George Jr. (1997) Poreforming segments in voltage-gated chloride channels. Nature 390: 529-532. Ho, K. and 6 others (1993) Cloning and expression of an inwardly rectifying ATP-regulated potassium channel. Nature 362: 31-38. Hodgkin, A. L. and R. D. Keynes (1955) Active transport of cations in giant axons from Sepia and Loligo. J. Physiol. 128: 28-60. Hoshi, T., W. N. Zagotta and R. W. Aldrich (1990) Biophysical and molecular mechanisms of Shaker potassium channel inacti-vation. Science 250: 533-538. Llano, I., C. K. Webb and F. Bezantlla (1988) Potassium conductance of squid giant axon. Single-channel studies. J. Gen. Physiol. 92:179-196. Long, S. B„ E. B. Campbell and R. MacKinnon (2005) Crystal structure of a mammalian voltage-dependent Shaker family K+ channel. Science 309: 897903. Morth, J. P., H. Poulsen, M. S. Toustrup-Jensen, V. R. Schack, J. Egebjerg, J. Andersen, B. Vilsen and P. Nissen (2009) The structure of the Na+,K+-ATPase and mapping of isoform differences and disease-related mutations. Philos. Trans. Roy. Soc. Land. B 364: 217-227. Noda, M. and 6 others (1986) Expression of functional sodium channels from cloned cDNA. Nature 322: 826-828. Nowycky, M. C, A. P. Fox and R. W. Tsien (1985) Three types of neuronal calcium channel with different calcium agonist sensitivity. Nature 316: 440443. Papazian, D. M., T. L. Schwarz, B. L. Tempel, Y. N. Jan and L. Y. Jan (1987) Cloning of genomic and complementary DNA from Shaker, a putative potassium channel gene from Drosophila. Science 237: 749-753. Shinoda, T., H. Ogawa, F. Cornelius and C. Toyoshima (2009) Crystal structure of the sodium-potassium pump at 2.4 A resolution. Nature 459: 446-450. Sigworth, F. J. .and E. Neher (1980) Single Na+ channel currents observed in cultured rat muscle cells. Nature 287: 447-449. Tao, X., A. Lee, W. Limapichat, D. A. Dougherty and R. MacKinnon (2010) A gating charge transfer center in voltage sensors. Science 328: 67-73. Thomas, R. C. (1969) Membrane current and intracellular sodium changes in a snail neurone during extrusion of injected sodium. J. Physiol. 201: 495-514. Toyoshima, C, H. Nomura and T. Tsuda (2004) Luminal gating mechanism revealed in calcium pump crystal structures with phosphate analogues. Nature 432: 361-368. Vanderberg, C. A. and F. Bezantlla (1991) A sodium channel model based on single channel, macroscopic ionic, and gating currents in the squid giant axon. Biophys. J. 60: 1511-1533. Waldmann, R., G. Champigny, F. Bassilana, C. Heurteaux and M. Lazdunski (1997) A proton-gated cation channel involved in acid-sensing. Nature 386:173-177. Wei, A. M., A. Covarrubias, A. Butler, K. Baker, M. Pak and L. Salkoff (1990) K+ current diversity is produced by an extended gene family conserved in Drosophila and mouse. Science 248: 599-603. Libros Aidley, D. J. and P. R. Stanfield (1996) Ion Channels: Molecules in Action. Cambridge: Cambridge University Press. Ashcroft, F. M. (2000) Ion Channels and Disease. Boston: Academic Press. Gribkoff, V. K. and L. K. Kaczmarek (2009) Structure, Function and Modulation of Neuronal Voltage-Gated Ion Channels. New York, NY: Wiley. Hille, B. (2001) Ion Channels of Excitable Membranes, 3rd Ed. Sunderland, MA: Sinauer Associates. Junge, D. (1992) Nerve and Muscle Excitation, 3rd Ed. Sunderland, MA: Sinauer Associates. 5 Transmisión sinápTica Aspectos generales EL ENCÉFALO HUMANO CONTIENE POR LO MENOS 100 000 MILLONES DE NEURONAS, cada una con la capacidad de influir en muchas otras células. Sin duda, se requieren mecanismos complejos y altamente eficientes para hacer posible la comunicación entre este número astronómico de elementos. Esta comunicación se logra con las sinapsis, que son los contactos funcionales entre las neuronas. Es posible distinguir dos tipos diferentes de sinapsis (eléctricas y químicas) sobre la base de su mecanismo de transmisión. En las sinapsis eléctricas, la corriente fluye a través de las uniones en hendidura, que son canales de membrana especializados que conectan dos células. En cambio, las sinapsis químicas permiten la comunicación intercelular a través de la secreción de neurotransmisores; estos agentes químicos liberados por las neuronas presinápticas producen flujo de corriente en las neuronas postsinápticas al activar moléculas receptoras específicas. El número total de neurotransmisores no se conoce, pero es muy superior a 100. Casi todos los neurotransmisores sufren un ciclo similar de uso: síntesis y empaquetamiento en vesículas sinápticas; liberación desde la célula presináptica; fijación a receptores postsinápticos y, por último, una rápida eliminación, degradación o ambas. La secreción de neurotransmisores es desencadenada por el influjo de Ca2+ a través de los canales con puerta de voltaje, que dan origen a un aumento transitorio en la concentración de Ca2+ en el interior de la terminación presináptica. La elevación en la concentración de Ca2+ hace que las vesículas presinápticas se fusionen con la membrana plasmática presináptica y liberen su contenido en el espacio entre las células presinápticas y postsinápticas. Si bien aún no se conoce exactamente de qué modo el Ca2+ desencadena la exocitosis, las proteínas sobre la superficie de las vesículas sinápticas y en otros sitios de la terminación presináptica median este proceso. Los neurotransmisores provocan respuestas eléctricas postsinápticas al fijarse a miembros de un grupo diverso de receptores de neurotransmisores. Existen dos clases principales de receptores: aquellos en los cuales la molécula receptora también es un canal iónico, y aquellos en los cuales receptor y canal iónico son moléculas separadas. Estos receptores dan origen a señales eléctricas por la apertura o el cierre de los canales iónicos inducidos por los neurotransmisores. El hecho de que las acciones postsinápticas de un neurotransmisor particular sean excitadoras o inhibidoras está determinado por la permeabilidad iónica del canal iónico afectado por el transmisor y por el gradiente electroquímico para los iones permeables. 78 CAPÍTULO 5 Sinapsis eléctricas Aunque existen muchos tipos de sinapsis en el interior del encéfalo humano pueden dividirse en dos clases generales: sinapsis eléctricas y sinapsis químicas. Si bien constituyen una minoría definida, las sinapsis eléctricas se encuentran en todos los sistemas nerviosos y permiten el flujo pasivo y directo de corriente eléctrica de una neurona a otra. La estructura de una sinapsis eléctrica se muestra esquemáticamente en la Figura 5.1. La neurona que se encuentra “corriente arriba” (proximal), origen de la corriente, se denomina elemento presináptico, y la neurona que se encuentra “corriente abajo” (distal) hacia la cual fluye esta corriente se denomina postsináptica. Las membranas de las dos neuronas comunicantes se aproximan mucho en la sinapsis y en realidad se conectan por una especialización intercelular llamada unión en hendidura o unión gap. Las uniones en hendidura contienen canales apareados y alineados con precisión, denominados conexo- (A) nes, que se presentan en la membrana de las neuronas presinápticas y postsinápticas; seis conexinas presinápticas se alinean con seis conexinas postsinápticas para formar un poro (Figura 5.1C). El poro de un canal conexón es mucho más grande que el poro de los canales iónicos con puerta de voltaje descritos en el capítulo anterior. En consecuencia, distintas sustancias pueden difundir FIGURA 5.1 Estructura de las sinapsis eléctricas. (A) En las sinapsis eléctricas, ocurren uniones en hendidura entre las membranas presináptica y postsináptica. (B) Las uniones en hendidura consisten en canales intercelulares que permiten que la corriente fluya pasivamente desde la célula presináptica a la postsináptica. (C) Las uniones en hendidura consisten en comploejos hexaméricos formados por la unión de subunidades denominadas conexones, que están presentes tanto en la membrana presináptica como en la postsináptica. Los poros de los canales se conectan y crean una continuidad eléctrica entre las dos células. (D) Los conexones consisten en la proteína integral de la membrana conexina. (B) Membrana presináptica Microtúbulos Neurona presináptica La corriente iónica fluye a través de los canales de los conexones Citoplasma Mitocondria Extracelular Unión en hendidura Membrana postsináptica Neurona postsináptica Membrana presináptica (C) (D) Conexones Extracelular Conexina Conexones Membrana postsináptica 3,5 3.5 nm 20 nm Poros que conectan el citoplasma de dos neuronas N Citoplasma C TRANSMISIÓN SINÁPTICA simplemente entre el citoplasma de las neuronas presinápticas y postsinápticas. Además de los iones, las sustancias que difunden a través de los poros de la unión en hendidura incluyen moléculas con pesos moleculares de hasta varios cientos de daltons. Esto permite que el ATP y otros metabolitos intracelulares importantes, como los segundos mensajeros (véase Cap. 7), sean transferidos entre las neuronas. Los conexones están compuestos por una familia especial de proteínas de canales iónicos, las conexinas (Figura 5.1D). Existen varios tipos diferentes de conexinas, halladas en distintos tipos celulares y que proporcionan uniones en hendidura con diversas propiedades fisiológicas. (A) Neurona presináptica 25 Potencial de membrana (mV) 0 <25 <50 25 Neurona postsináptica 0 <25 <50 Retardo sináptico breve (alrededor de 0,1 ms) 0 1 2 Tiempo (ms) 3 4 (B) 0 Potencial de membrana (mV) <25 <50 Neurona 1 * * * * * * 79 Las sinapsis eléctricas funcionan así permitiendo que la corriente iónica fluya pasivamente a través de los poros de la unión en hendidura desde una neurona a la otra. La fuente habitual de corriente es la diferencia de potencial generada localmente por el potencial de acción (véase Cap. 3). Esta disposición tiene algunas consecuencias interesantes. Una es que la transmisión puede ser bidireccional; esto es, la corriente puede fluir en cualquier dirección a través de la unión en hendidura, dependiendo de qué miembro de la pareja acoplada sea invadido por el potencial de acción (aunque algunos tipos de uniones en hendiduratienen características especiales que hacen su transmisión unidireccional). Otro rasgo importante de la sinapsis eléctrica es que la transmisión es extraordinariamente rápida: dado que el flujo pasivo de corriente a través de la unión en hendidura es casi instantáneo, la comunicación puede ocurrir sin la demora característica de las sinapsis químicas. Estas características son evidentes en la operación de la primera sinapsis eléctrica descubierta, localiza en el sistema nervioso del cangrejo de río. Se observa una señal eléctrica postsináptica en esta sinapsis en una fracción de milisegundo después de la generación de un potencial de acción presináptico (Figura 5.2A). De hecho, al menos parte de esta breve demora sináptica es causada por la propagación del potencial de acción en la terminación presináptica, de modo que esencialmente es posible que no exista ninguna demora en la transmisión de señales eléctricas a través de la sinapsis. Estas sinapsis interconectan muchas de las neuronas en el circuito que permite al cangrejo de río escapar de sus depredadores, y minimizan así el tiempo entre la presencia de un estímulo amenazante y una respuesta motora que tal vez le salve la vida. Un propósito más general de las sinapsis eléctricas es sincronizar la actividad eléctrica entre poblaciones de neuronas. Por ejemplo, las neuronas del tronco encefálico que generan la actividad eléctrica rítmica que subyace a la respiración están sincronizadas por sinapsis eléctricas, al igual que las poblaciones de interneuronas en la corteza cerebral, tálamo, cerebelo y otras regiones encefálicas (Figura 5.2 B). La transmisión eléctrica entre ciertas neuronas hormonosecretantes del hipotálamo de los mamíferos asegura que todas las células disparen potenciales de acción aproximadamente al mismo tiempo y faciliten así una explosión de secreción hormonal en la circulaFIGURA 5.2 Función de las uniones en hendidura en las sinapsis eléctricas. (A) Transmisión rápida de señales en una * 0 sinapsis eléctrica en el cangrejo de río. Un potencial de acción en la neurona presináptica hace que la neurona postsináptica se <25 <50 Neurona 2 0 100 200 Tiempo (ms) 300 400 despolarice en la fracción de un milisegundo. (B) Las sinapsis eléctricas permiten la sincronización de la actividad eléctrica en las interneuronas del hipocampo. En un par de interneuronas conectadas por sinapsis eléctricas, la generación de un potencial de acción en una neurona a menudo conduce a la descarga sincronizada de un potencial de acción en otra neurona. (A, de Furshpan y Potter, 1959; B, de Beierlein y cols., 2000.) 80 CAPÍTULO 5 Transmisión de señales en las sinapsis químicas las sinapsis químicas que en las eléctricas y se denomina hendidura sináptica. Sin embargo, la característica clave de todas las sinapsis químicas es la presencia de pequeños orgánulos limitados por membranas llamados vesículas sinápticas en el interior de la terminación presináptica. Estos orgánulos esféricos están llenos de uno o más neurotransmisores, las señales químicas secretadas desde la neurona presináptica, y son estos agentes químicos que actúan como mensajeros entre las neuronas comunicantes los que proporcionan su nombre a este tipo de sinapsis. La transmisión en las sinapsis químicas se basa en la secuencia de acontecimientos que se detalla en la Figura 5.3. El proceso se inicia cuando un potencial de acción invade la termina- La estructura general de una sinapsis química se muestra en forma esquemática en la Figura 5.3. El espacio entre las neuronas presinápticas y postsinápticas es sustancialmente mayor en FIGURA 5.3 Secuencia de acontecimientos involucrados en la transmisión en una sinapsis química típica. ción. El hecho de que los poros de la unión en hendidura sean lo suficientemente grandes como para permitir que moléculas como ATP y segundos mensajeros difundan hacia el interior de las células también permite que las sinapsis eléctricas coordinen la señalización intracelular y el metabolismo de las células acopladas. Esta propiedad puede ser de particular importancia para las células gliales, que forman grandes redes de señalización intracelular a través de sus uniones en hendidura. Mielina 2 Un potencial de acción invade la terminación presináptica transmisor es sinteti1 El zado y luego almacenado en las vesículas 3 La despolarización de la terminación presináptica produce la apertura de los canales del Ca2+ con puerta de voltaje 2+ 4 Influjo de Ca a través de los canales Vesícula sináptica 11 Recuperación de la membrana vesicular desde la membrana plasmática 2+ 5 El Ca hace que las vesículas se fusionen con la membrana presináptica Moléculas del transmisor Ca2+ 6 El transmisor es liberado en la hendidura sináptica a través de la exocitosis Célula glial Moléculas del transmisor Iones Receptor del transmisor Flujo de corriente postsináptica 7 El transmisor se une a las moléculas receptoras en la membrana postsináptica 10 Remoción del neuro- transmisor por captación glial o degradación enzimática A través de la dendrita 9 La corriente postsináptica produce un potencial postsináptico excitador o inhibidor que cambia la excitabilidad de la célula postináptica 8 Apertura o cierre de los canales postinápticos TRANSMISIÓN SINÁPTICA ción de la neurona presináptica. El cambio en el potencial de membrana causado por la llegada del potencial de acción produce la apertura de los canales del calcio con puerta de voltaje en la membrana presináptica. Debido al acentuado gradiente de concentración de Ca2+ a través de la membrana presináptica (la concentración externa de Ca2+ es aproximadamente de 10-3M, mientras que la concentración interna de Ca2+ es de alrededor de 10-7M), la apertura de estos canales produce un influjo rápido de Ca2+ en la terminación presináptica, con el resultado de que la concentración de Ca2+ del citoplasma en la terminación se eleva transitoriamente hasta un valor mucho más alto. La elevación de la concentración presináptica de Ca2+ permite que las vesículas sinápticas se fusionen con la membrana plasmática de la neurona presináptica. La fusión Ca2+-dependiente de las vesículas sinápticas con la membrana de la terminación hace que su contenido, principalmente los neurotransmisores, sea liberado en la hendidura sináptica. Tras la exocitosis, los transmisores difunden a través de la hendidura sináptica y se unen a receptores específicos sobre la membrana de la neurona postsináptica. La fijación del neurotransmisor a los receptores abre los canales de la membrana postsináptica (o a veces los cierra), lo que altera así la capacidad de los iones de ingresar (o salir) en las células postsinápticas. El flujo de corriente resultante inducido por el neurotransmisor altera la conductancia y (habitualmente) el potencial de membrana de la neurona postsináptica, lo cual aumenta o disminuye la probabilidad de que la neurona dispare un potencial de acción. De esta forma, la información es transmitida de una neurona a otra. 81 el tema de un intenso debate durante la primera mitad del siglo Un experimento clave que apoyó esta idea fue realizado en 1926 por el fisiólogo alemán Otto Loewi. Trabajando sobre la idea que presuntamente se le presentó en la mitad de la noche, Loewi probó que la estimulación eléctrica del nervio vago retarda el latido cardíaco al liberar una señal química. El científico aisló y perfundió los corazones de dos ranas, controlando las frecuencias con las que latían (Figura 5.4). Cuando se estimuló el nervio vago del primer corazón, el latido de este corazón se hizo más lento. Notablemente, aun cuando el nervio vago del segundo corazón no había sido estimulado, su latido también se hizo más lento cuando estuvo expuesto al líquido de perfusión del primer corazón. Este resultado mostró que el nervio vago regula la frecuencia cardíaca al liberar una sustancia química que se acumula en el perfundido. Denominada originariamente “sustancia del vago”, más tarde se demostró que la sustancia era acetilcolina (ACh). En la actualidad, se sabe que la ACh es un neurotransmisor que no sólo actúa sobre el corazón, sino en distintas dianas postsinápticas en los sistemas nerviosos central y periférico, predominantemente en la unión neuromuscular de los músculos estriados y en el sistema motor visceral (véanse Caps. 6 y 21). Con los años, fueron surgiendo algunos criterios formales que identifican de forma definitiva a una sustancia como neurotransmisor (Recuadro 5A). Estos criterios condujeron a la identificación de más de 100 neurotransmisores diferentes, los que pueden clasificarse en dos categorías amplias: neurotransmisores de xx. Propiedades de los neurotransmisores La idea de que la información eléctrica puede transmitirse de una neurona a la siguiente por medio de señales químicas fue lugar en que se estimulaba el nervio vago del corazón aislado de la rana, la frecuencia cardíaca disminuía (panel superior). Si se transfería el líquido de perfusión del corazón estimulado a un segundo corazón, su frecuencia Estímulo del nervio disminuía también (panel inferior). vago del cor azón 1 (B) Estímulo Nervio vago Solución transferida al corazón 2 Fuerza de contracción (A) FIGURA 5.4 Experimento de Loewi que demuestra la neurotransmisión química. (A) Disposición experimental de Loewi. (B) En el Corazón 1 Latido cardíaco lento Tiempo (s) Fuerza de contracción Corazón 1 Corazón 2 Corazón 2 Efecto inhibidor del vago transferido Tiempo (s) CAPÍTULO 5 molécula pequeña y neuropéptidos (véase Cap. 6). Contar con más de un transmisor diversifica el repertorio fisiológico de las sinapsis. Múltiples neurotransmisores pueden producir diferentes tipos de respuestas en las células postsinápticas individuales. Por ejemplo, una neurona puede ser excitada por un tipo de neurotransmisor e inhibida por otro. La velocidad de las respuestas postsinápticas producidas por diferentes transmisores también difiere, lo que permite el control de la señalización eléctrica en distintas escalas temporales. En general, los neurotransmisores de molécula pequeña median acciones sinápticas rápidas, mientras que los neuropéptidos tienden a modular funciones sinápticas en curso y más lentas. Hasta hace relativamente poco, se pensaba que una neurona específica producía sólo un único tipo de neurotransmisor. Sin embargo, ahora está claro que muchos tipos de neuronas sintetizan y liberan dos o más neurotransmisores diferentes. Cuando se presenta más de un neurotransmisor en el interior de una terminación nerviosa, las moléculas se denominan cotransmisores. Como diferentes tipos de transmisores pueden ser empaquetados en distintas poblaciones de vesículas sinápticas, los cotransmisores no necesariamente son liberados de manera simultánea. Cuando los neurotransmisores peptídicos y de molécula pequeña actúan como cotransmisores en la misma sinapsis, son liberados de modo diferente según el patrón de actividad sináptica: a menudo, la actividad de baja frecuencia sólo libera neurotransmisores pequeños, mientras que la actividad de alta frecuencia es necesaria para liberar neuropéptidos de las mismas terminaciones presinápticas. En consecuencia, las propiedades de señalización química de estas sinapsis cambian según la velocidad de la actividad. Una transmisión sináptica eficaz requiere un control riguroso de la concentración de neurotransmisores en el interior de la hendidura sináptica. Por lo tanto, las neuronas han desarrollado una capacidad muy compleja de regular la síntesis, el empaquetamiento, la liberación y la degradación (o eliminación) de neurotransmisores para lograr los niveles deseados de moléculas de transmisor. La síntesis de neurotransmisores de molécula pequeña ocurre localmente en el interior de las terminaciones presinápticas (Figura 5.5A). Las enzimas necesarias para sintetizar estos transmisores se producen en el cuerpo de las neuronas y son transportadas hasta el citoplasma de la terminación nerviosa a una velocidad de 0,5-5,0 milímetros por día por un mecanismo denominado transporte axónico lento. Habitualmente, las moléculas precursoras necesarias para formar nuevas moléculas de neurotransmisor son captadas en la terminación nerviosa por transportadores que se encuentran en la membrana plasmática de la terminación. Las enzimas sintetizan neurotransmisores en el citoplasma de la terminación presináptica y luego los transmisores son cargados en vesículas sinápticas mediante transportadores en la membrana vesicular (véase Cap. 4). Para algunos neurotransmisores de molécula pequeña, los pasos finales de la síntesis ocurren en el interior de las vesículas sinápticas. La mayoría de los neurotransmisores de molécula pequeña son empaquetados en vesículas de 40 a 60 nm de diámetro, cuyos centros FIGURA 5.5 Metabolismo de los transmisores de molécula pequeña y los transmisores peptídicos. (A) Los neurotransmisores ▼ 82 de molécula pequeña son sintetizados en las terminaciones nerviosas. Las enzimas necesarias para la síntesis de los neurotransmisores se forman en el cuerpo celular de la célula presináptica (1) y son transportadas por el axón a través del transporte axónico lento (2). Los precursores son captados en las terminaciones por transportadores específicos, y la síntesis y el empaquetamiento de los neurotransmisores tienen lugar en el interior de las terminaciones nerviosas (3). Después de la fusión y la liberación de las vesículas (4), el neurotransmisor puede ser degradado por vía enzimática. La recaptación del neurotransmisor (o sus metabolitos) comienza otro ciclo de síntesis, empaquetamiento, liberación y eliminación (5). (B) Vesículas pequeñas de centro claro entre una terminación presináptica y una espina dendrítica en el sistema nervioso central. (C) Los neurotransmisores peptídicos, y las enzimas que modifican sus precursores, son sintetizados en el cuerpo celular (1). Las enzimas y los propéptidos son empaquetados en vesículas en el aparato de Golgi. Durante el transporte axónico rápido de estas vesículas hasta las terminaciones nerviosas (2), las enzimas modifican los propéptidos para producir uno o más péptidos neurotransmisores (3). Después de la fusión y la exocitosis de las vesículas, los péptidos difunden alejándose y son degradados por enzimas proteolíticas (4). (D) Vesículas grandes de centro denso en una terminación presináptica central que hacen sinapsis en una dendrita. En los casos típicos estas vesículas contienen neuropéptidos o, en ocasiones, aminas biógenas. (B y D, de Peters, Palay y Webster, 1991.) aparecen claros en micrografías electrónicas; en consecuencia, estas vesículas se denominan vesículas pequeñas de centro claro (Figura 5.5B). Los neuropéptidos son sintetizados en el cuerpo celular de una neurona, lo que significa que el péptido es producido en un lugar distante de su sitio de secreción (Figura 5.5C). Para resolver este problema, vesículas llenas de péptidos son transportadas a lo largo de un axón y por la terminación sináptica mediante el transporte axónico rápido. Este proceso lleva las vesículas a velocidades de hasta 400 mm/día a lo largo de elementos del citoesqueleto llamados microtúbulos (al contrario del transporte axónico lento de las enzimas que sintetizan transmisores de molécula pequeña). Los microtúbulos son filamentos cilíndricos largos de 25 nm de diámetro, presentes en todas las neuronas y en otras células. Las vesículas que contienen péptidos son movilizadas a lo largo de estas “pistas” de microtúbulos por proteínas “motores” que requieren ATP como cinesina. Los neuropéptidos son empaquetados en vesículas sinápticas de un diámetro que varía entre 90 y 250 nm. Estas vesículas son electrodensas en las electromicrografías, de ahí que se las denomine vesículas grandes de centro denso (Figura 5.5D). Una vez que un neurotransmisor ha sido secretado en la hendidura sináptica, debe ser eliminado para permitir que la célula postsináptica participe en otro ciclo de transmisión sináptica. La eliminación de los neurotransmisores comprende la difusión lejos de los receptores postsinápticos, combinada con recaptación en las terminaciones nerviosas o las células gliales circundantes, degradación por enzimas específicas o una combinación de estos mecanismos. Las proteínas transportadoras específicas eliminan la mayor parte de los neurotransmisores de molécula pequeña (o TRANSMISIÓN SINÁPTICA (A) Transmisor de molécula pequeña (C) Neurotransmisores peptídicos 1 Síntesis de 1 Síntesis de Núcleo precursores de nuerotransmisores y enzimas enzimas en el cuerpo celular RER Aparato de Golgi Microtúbulos 2 Transporte de las enzi- mas y los precursores peptídicos a través de las pistas de microtúbulos 2 Transporte axónico lento de las enzimas Axón 5 Transporte de precursores en la terminación Terminación Precursor Enzimas 3 Síntesis y empaquetamiento del neurotransmisor 4 El neurotransmisor difunde y es degradado por enzimas proteolíticas 3 Las enzimas modifican los precursores para producir un neurotransmisor peptídico 4 Liberación y difusión del neurotransmisor Neurotransmisor (D) (B) Terminaciones presinápticas Vesículas Dendritas 0.5 μm 83 84 CAPÍTULO 5 RECUADRO 5A Criterios que definen un neurotransmisor Para confirmar que una molécula actúa como neurotransmisor en una sinapsis química determinada se utilizan tres criterios primarios: 1. La sustancia debe estar presente en el interior de la neurona presináptica. Indudablemente, una sustancia química no puede ser secretada desde una neurona presináptica a menos que esté presente allí. Dado que se necesitan vías bioquímicas complejas para producir neurotransmisores, la demostración de que las enzimas y los precursores necesarios para sintetizar la sustancia están presentes en las neuronas presinápticas brinda pruebas adicionales de que la sustancia es utilizada como neurotransmisor. Sin embargo, obsérvese que, como los transmisores glutamato, glicina y asparato también son necesarios para la síntesis proteica y otras reaccio(1) nes metabólicas en todas las neuronas, su presencia no es prueba suficiente para establecerlos como neurotransmisores. 2. La sustancia debe ser liberada en respuesta a la despolarización presináptica, la cual debe ocurrir en forma Ca2+-dependiente. Otro criterio esencial para identificar a un neurotransmisor es demostrar que es liberado de la neurona presináptica en respuesta a la actividad eléctrica presináptica, y que esta liberación exige el influjo de Ca2+ en la terminación presináptica. Cumplir este criterio es un desafío técnico, no solo porque puede ser difícil estimular selectivamente las neuronas presinápticas, sino también porque las enzimas y los transportadores eliminan eficientemente los neurotransmisores secretados. (2) (3) 1 Neurotransmisor Potencial de acción presente Terminación presináptica Ca2+ Célula postsináptica Acción de agonistas o antagonistas del neurotransmisor Ca2+ del 2 Liberación neurotransmisor 3 sus metabolitos) de la hendidura sináptica, y finalmente vuelven a entregarlos a la terminación presináptica para su reutilización (véase Cap. 6). Liberación cuántica de los neurotransmisores Gran parte de las pruebas que condujeron al conocimiento actual de la transmisión en las sinapsis químicas se obtuvo de experimentos que examinaron la liberación de ACh en las uniones neuromusculares. Estas sinapsis entre las neuronas motoras espinales y las células del músculo esquelético son sencillas, grandes y de localización periférica, lo que las hace particularmente indicadas para el análisis Activación de los receptores del neurotransmisor 3. Se deben presentar receptores específicos para la sustancia en la célula postsináptica. Un neurotransmisor no puede actuar sobre su diana a menos que se presenten receptores específicos para el transmisor en la membrana postsináptica. Una forma de probar que están los receptores es mostrando que la aplicación del transmisor exógeno imita el efecto postsináptico de la estimulación presináptica. Otra forma más rigurosa de hacerlo es demostrar que los agonistas y los antagonistas que alteran la respuesta postsináptica normal tienen el mismo efecto cuando la sustancia en cuestión se aplica exógenamente. También se pueden utilizar métodos histológicos de alta resolución para mostrar que los receptores específicos están presentes en la membrana postsináptica (por detección de anticuerpos receptores marcados radiactivamente, por ejemplo). El cumplimiento de estos criterios establece inequívocamente que una sustancia dada es utilizada como transmisor en una sinapsis. Sin embargo, algunas dificultades prácticas han impedido la aplicación de estos estándares en muchos tipos de sinapsis. Por esta razón, tantas sustancias deben ser denominadas neurotransmisores “putativos”. La identidad de un neurotransmisor en una sinapsis se demuestra con (1) su presencia, (2) su liberación y (3) la presencia postsináptica de los receptores específicos. experimental. Estas sinapsis ocurren en unas zonas especializadas llamadas placas terminales por el aspecto de platillo que presenta las zonas de la fibra muscular en donde el axón presináptico proyecta sus terminaciones (Figura 5.6A). La mayor parte de las primeras investigaciones sobre la transmisión neuromuscular fue realizada por Bernard Katz y cols. en el University College, Londres, durante las décadas de 1950 y 1960, y alcanzando Katz un gran reconocimiento por sus notables contribuciones en el conocimiento de la transmisión sináptica. Aunque este autor investigó fundamentalmente la unión neuromuscular de la rana, muchos experimentos posteriores han confirmado la aplicabilidad de sus observaciones a la transmisión de todas las sinapsis químicas. Cuando se utiliza un microelectrodo intracelular para registrar el potencial de membrana de una célula muscular, es posi- TRANSMISIÓN SINÁPTICA (A) Estímulo FIGURA 5.6 Transmisión sináptica en la unión neuromuscular. (A) Disposición experimental: Estimular el axón Registro Registro del potencial de membrana postsináptica Axón 85 el axón de la neurona motora que inerva la fibra muscular es estimulado con un electrodo extracelular, mientras se inserta un microelectrodo intracelular en la célula muscular postsináptica para registrar sus respuestas eléctricas. (B) Los potenciales de placa terminal (PPT) provocados por la estimulación de una neurona motora se encuentran normalmente por encima del umbral y, por lo tanto, producen un potencial de acción en la célula muscular postsináptica. (C) Los PPT en miniatura (PPTM) espontáneos se desarrollan en ausencia de estimulación presináptica. (D) Cuando la unión neuromuscular es bañada en una solución que tiene baja concentración de Ca2+, la estimulación de la neurona motora evoca PPT cuyas amplitudes están reducidas hasta el tamaño aproximado de los PPTM. (De Fatt y Katz, 1952.) Célula muscular 50 Potencial de acción 0 Umbral <50 <100 0 Potencial de placa terminal (PPT) 2 4 6 Tiempo (ms) (D) (C) 1 mV Potencial de membrana postsináptica (mV) Estimular el axón motor Potencial de membrana postsináptica (mV) Potencial de membrana postsináptica (mV) (B) MEPP 0 200 400 Tiempo (ms) ble observar que un potencial de acción en la neurona motora presináptica provoca una despolarización transitoria de la fibra muscular postsináptica. Este cambio en el potencial de membrana, llamado potencial de placa terminal (PPT), por lo habitual suficientemente grande como para elevar el potencial de membrana de la fibra muscular muy por encima del umbral para producir un potencial de acción postsináptico (Figura 5.6B). El potencial de acción postsináptico desencadenado por el PPT hace que la fibra muscular se contraiga. Al contrario de las sinapsis eléctricas, existe una demora acentuada entre el momento en que la neurona motora presináptica es estimulada y el momento en que ocurre el PPT en la célula muscular postsináptica. Esta demora es característica de todas las sinapsis químicas. Uno de los hallazgos fundamentales de Katz, en estudios que realizó de manera conjunta con Paul Fatt en 1951, fue que los cambios espontáneos en el potencial de membrana de la célula muscular ocurren incluso en ausencia de estimulación de la neurona motora presináptica (Figura 5.6C). Estos cambios tienen la misma forma que los PPT, pero son mucho más pequeños (típicamente, una amplitud inferior a 1 mV, comparada con el PPT de más de 50 mV). Tanto los PPT como estos fenómenos espontáneos pequeños son sensibles a los agentes farmacológicos que bloquean los receptores colinérgicos postsinápticos, como el curare (véase Recuadro 6A). Estos paralelismos y otros entre los PPT y las despolarizaciones espontáneas condujeron a Katz y cols. a denominar los hechos espontáneos potencial de placa terminal en miniatura o PPTM. La relación entre el potencial de placa terminal completo y los PPTM fue aclarada mediante un análisis cuidadoso de los PPT. La magnitud de los PPT proporciona un ensayo eléctrico conveniente de la secreción de los neurotransmisores desde la terminación de la Estímulo del axón motor PPT subumbral 1 mV PPTM espontáneo 0 20 40 60 80 100 Tiempo (ms) neurona motora; sin embargo, medirlo es complicado por la necesidad de evitar que la contracción muscular desaloje el microelectrodo. El método habitual para eliminar las contracciones musculares es reducir la concentración de Ca2+ en el medio extracelular o bloquear parcialmente los receptores colinérgicos postsinápticos con el agente curare. Como es de esperar a partir del esquema que se muestra en la Figura 5.3, la reducción de la concentración de Ca2+ reduce la secreción del neurotransmisor y desciende así la magnitud del PPT por debajo del umbral para la producción del potencial de acción postsináptico, lo que permite medirlo con mayor precisión. En estas condiciones, la estimulación de la neurona motora produce PPT muy pequeños que fluctúan en amplitud de un ensayo a otro (Figura 5.6D). Estas fluctuaciones brindan un conocimiento considerable acerca de los mecanismos responsables de la liberación del neurotransmisor. En particular, la respuesta provocada con bajo Ca2+ parece ser el resultado de la liberación de cantidades unitarias de ACh por la terminación nerviosa presináptica. Por lo tanto, la amplitud de la respuesta provocada más pequeña es notablemente similar al tamaño de los PPTM aislados (compárense Figuras 5.6C y D). De acuerdo con esta similitud, los incrementos en la respuesta del PPT (Figura 5.7A) ocurren en unidades que tienen aproximadamente el tamaño de PPTM aislados (Figura 5.7B). Estas fluctuaciones “cuánticas” en la amplitud de los PPT indicaron a Katz y a su colaborador José del Castillo que éstos estaban formados por unidades individuales, cada una equivalente a un PPTM. La idea de que los PPT representan la liberación simultánea de muchas unidades similares a PPTM puede ser evaluada estadísticamente. Un método de análisis estadístico basado en la ocurrencia independiente de eventos unitarios (llamado estadístico de Poisson) predice que la distribución de las amplitudes 86 (A) CAPÍTULO 5 Cantidad de PPTM Cantidad de PPT dura sináptica. Aproximadamente en la época en que Katz y cols. descubrían la liberación cuántica de neurotransmisor, la microscopia electrónica Predicción del 20 modelo estadístico puso en evidencia, por primera vez, la presencia de vesículas sinápticas en las terminaciones presinápticas. Uniendo estos dos descubrimientos, 15 Katz y otros dos autores propusieron que las vesículas sinápticas cargadas con neurotransmisor constituían la fuente de los cuantos. Algunos es10 tudios bioquímicos posteriores mostraron que las vesículas sinápticas eran los repositorios de los transmisores. Estos estudios han mostrado que 5 la acetilcolina está altamente concentrada en las vesículas sinápticas de las neuronas motoras, donde se presenta en una concentración de unos 0 100 mM. Dado el diámetro de una vesícula sinápti0,4 0,8 1,2 1,6 2,0 2,4 2,8 0 ca pequeña de centro claro (~50 nm), una vesícula Amplitud del PPT (mV) sináptica contiene aproximadamente 10 000 moFIGURA 5.7 Distribución en cuantos de las am(B) léculas de neurotransmisor. Este número se coplitudes de los PPT provocados en una solución rresponde muy bien con la cantidad de ACh que con baja concentración de Ca2+. Los picos de las debe aplicarse en una unión neuromuscular para 30 amplitudes de los PPT (A) tienden a desarrollarse en imitar un PPTM, lo que proporciona otro apoyo a múltiplos enteros de la amplitud media de PPTM, cuya la idea de que los cuantos surgen de la descarga distribución de amplitudes se muestra en (B). La barra del contenido de una sola vesícula sináptica. 20 más a la izquierda en la distribución de las amplitudes Para probar que los cuantos son causados por la en los PPT muestra ensayos en los cuales la estimulafusión de vesículas sinápticas individuales con la ción presináptica no pudo obtener un PPT en la célula membrana plasmática, es necesario mostrar que muscular. La curva roja indica la predicción de un 10 cada vesícula fusionada produce el registro postsimodelo estadístico basado en la presunción de que los náptico de un único evento cuántico. Este desafío PPT son el resultado de la liberación independiente de fue logrado a fines de la década de 1970, cuando múltiples cuantos similares a PPTM. La equiparación 0 John Heuser, Tom Reese y cols. correlacionaron las observada, que incluye el número predicho de fracasos, 0 0,4 0,8 mediciones de la fusión vesicular con el contenido apoya esta interpretación. (De Boyad y Martin, 1955). Amplitud de los PPTM (mV) cuántico de los PPT en la unión neuromuscular. Estos autores utilizaron la microscopia electrónica de los PPT debe ser similar durante gran cantidad de ensayos de para determinar el número de vesículas que se fusionaron con la estimulación de la neurona motora, bajo la presunción de que los membrana plasmática presináptica en las terminaciones que haPPT están formados por eventos unitarios como PPTM (véase bían sido tratadas con una droga (4-aminopiridina, o 4-AP) que Fig. 5.7B). La distribución de las amplitudes de los PPT deter- aumentaba la cantidad de eventos de fusión de vesículas produciminada de manera experimental es compatible con lo esperado dos por los potenciales de acción aislados (Figura 5.8A). Se efecsi la liberación del transmisor de la neurona motora es efecti- tuaron mediciones eléctricas paralelas del contenido cuántico de vamente cuántica (la curva roja en Figura 5.7A). Estos análisis los PPT así obtenidos. La comparación de la cantidad de fusiones confirmaron la idea de que la liberación de acetilcolina ocurre de vesículas sinápticas observadas con microscopia electrónica y en paquetes separados, cada uno equivalente a un PPTM. Por la cantidad de cuantos liberados en la sinapsis mostró que la correlo tanto, un potencial de acción presináptico produce un PPT lación entre las dos medidas era buena (Figura 5.8B). Estos resulpostsináptico porque sincroniza la liberación de muchos cuantos tados aún son una de las líneas más firmes de apoyo para la idea de que un cuanto de liberación de transmisor se debe a la fusión de transmisor. de una vesícula sináptica con la membrana presináptica. La evidencia posterior, basada sobre otro método para medir la fusión de Liberación de transmisores de las las vesículas, no ha dejado ninguna duda acerca de la validez de esta interpretación general de la transmisión de las sinapsis químicas. vesículas sinápticas Investigaciones muy recientes han identificado estructuras en el El descubrimiento de la liberación cuántica de paquetes de interior de la terminación presináptica que conectan las vesículas neurotransmisor planteó inmediatamente el interrogante de con la membrana plasmática y pueden estar involucradas en la cómo se forman esos cuantos y cómo se descargan en la hendi- fusión de la membrana (Figura 5.8C). Sin PPT en respuesta a la estimulación TRANSMISIÓN SINÁPTICA (A) 87 (C) No estimulada Estimulada Vesículas sinápticas Fusión de las vesículas con la membrana plasmática Vesículas fusionadas Canales del Ca2+ Cantidad de vesículas que se fusionan (B) 5000 Concentración de 4-AP: 10<3M representan la fusión de vesículas sinápticas con la membrana presináptica. La imagen es como si se observaran sobre los sitios de liberación desde el exterior de la terminación presináptica. (B) Comparación del número de fusiones de vesículas observadas con el número de cuantos liberados por un potencial de acción presináptico. Se varió la liberación del transmisor utilizando una droga (4-AP) que 3000 10<4M 10<5M 1000 0 FIGURA 5.8 Relación entre la exocitosis de vesículas sinápticas y la liberación cuántica de transmisores. (A) Se utilizó una técnica especial de microscopia electrónica denominada microscopia de criofractura para visualizar la función de vesículas sinápticas en las terminaciones presinápticas de neuronas motoras de rana. Izquierda: imagen de la membrana plasmática de una terminación presináptica no estimulada. Derecha: imagen de la membrana plasmática de una terminación estimulada por un potencial de acción; la estimulación produce la aparición de estructuras similares a hoyuelos que 0 3000 5000 1000 Cantidad de cuantos liberados afecta la duración del potencial de acción presináptico, cambiando así la cantidad de calcio que ingresa durante el potencial de acción. La línea diagonal es la relación 1:1 esperada si cada vesícula que se abriera liberara un único cuanto de transmisor. (C) Estructura fina de los sitios de fusión de las vesículas de las terminaciones presinápticas de la rana. Las vesículas sinápticas están dispuestas en hileras y están conectadas entre sí y con la membrana plasmática por distintas estructuras proteináceas (azul). Se cree que las estructuras verdes en la membrana presináptica, que corresponden a las hileras de partículas observadas en (A), son canales del Ca2+. (A y B, de Heuser y cols., 1979; C, de Harlow y cols., 2001.) Reciclado local de las vesículas sinápticas La fusión de las vesículas sinápticas hace que se agregue nueva membrana a la membrana plasmática de la terminación presináptica, pero el agregado no es permanente. Si bien una tanda de exocitosis puede aumentar espectacularmente el área de superficie de las terminaciones presinápticas, esta membrana en exceso es eliminada en algunos minutos. Heuser y Reese realizaron otro conjunto importante de experimentos que mostraron que la membrana de la vesícula fusionada es recuperada y captada nuevamente en el citoplasma de la terminación nerviosa (proceso denominado endocitosis). Los experimentos, que volvieron a realizarse en la unión neuromuscular de la rana, se basaron en el llenado de la hendidura sináptica con peroxidasa de rábano picante (PRP), una enzima que produce un producto de reacción denso que puede observarse con el microscopio electrónico. En condiciones experimentales apropiadas, es posible visualizar la endocitosis por la captación de la peroxidasa en la terminación nerviosa (Figura 5.9). Para activar la endocitosis, se estimuló la terminación presináptica con un tren de potenciales de acción y se siguió el destino posterior de la PRP mediante microscopia electrónica. Inmediatamente después de la estimulación, se encontró peroxidasa de rábano picante en orgánulos endocitósicos especiales denominados vesículas con cubierta (Figura 5.9A, B). Sin embargo, algunos minutos más tarde, las vesículas con cubierta habían desaparecido y la PRP se encontraba en un orgánulo diferente, el endosoma (Figura 5.9C). Por último, aproximadamente una hora después de estimular la terminación, apareció el producto de reacción de la PRP en el interior de las vesículas sinápticas (Figura 5.9D). Estas observaciones indican que la membrana de las vesículas sinápticas es reciclada en el interior de la terminación presináptica a través de la secuencia resumida en la Figura 5.9E. En este proceso, llamado ciclo de las vesículas sinápticas, la membrana vesicular reciclada atraviesa diversos compartimientos intracelulares –vesículas con cubiertas y endosomas– y finalmente es utilizada para formar nuevas vesículas sinápticas. Las vesículas recién formadas se almacenan en un pool de reserva, anclado en la membrana plasmática presináptica, y son preparados para participar nuevamente en la liberación de un neurotransmisor. Algunos experimentos más recientes, que emplearon un marcador fluorescente en lugar de PRP, han determinado el curso temporal del reciclado de las vesículas sinápticas. Estos estudios indican que la totalidad del ciclo vesicular requiere aproximadamente 1 minuto, y la gemación de la membrana durante la endocitosis requiere 10-20 segundos. Como puede observarse a partir de la demora de 88 CAPÍTULO 5 (A) (B) (C) (D) Lavar la PRP extracelular; esperar 5 minutos Estimular brevemente la terminación presináptica 1 hora más tarde 3 El endosoma 2 contiene PRP Las fositas con cubierta y las vesículas con cubierta contienen PRP 1 Las vesículas Peroxidasa de rábano picante (PRP) 4 Las vesículas sinápticas se fusionan sinápticas contienen PRP (E) FIGURA 5.9 Reciclado local de vesículas sinápticas en las Endosoma terminaciones presinápticas. (A) La peroxidasa de rábano picante (PRP) introducida en la hendidura sináptica fue utilizada para seguir Endocitosis el destino de la membrana recuperada desde la membrana plasmática presináptica. La estimulación de endocitosis por potenciales de acción presinápticos hace que la PRP sea captada en las terminaciones presinápticas a través de una vía que incluye (B) vesículas con cubierta y (C) endosomas. (D) Finalmente, la PRP se encuentra en las vesículas Gemación 1 min 10–20 s sinápticas recién formadas. (E) Interpretación de los resultados que se muestran en A-D. La fusión de las vesículas con la membrana presináptica regulada por el calcio es seguida por la recuperación endocitósica de la membrana vesicular a través de las vesículas con cubierta y los endosomas, y la nueva formación posterior de nuevas vesículas sinápticas. (De Heuser y Reese, 1973.) 1 milisegundo en la transmisión que siguió a la excitación de la terminación presináptica (véase Fig. 5.6B), la fusión de la membrana durante la exocitosis es mucho más rápida que la gemación durante la endocitosis. Por lo tanto, todos los pasos de reciclado interpuestos entre la gemación de la membrana y la refusión posterior de una vesícula se completan en menos de un minuto. Los precursores de las vesículas sinápticas se producen originariamente en el retículo endoplasmático y el aparato de Golgi en el cuerpo de las células neuronales. Debido a la larga distancia entre el cuerpo celular y la terminación presináptica en la mayoría de las neuronas, el transporte de las vesículas desde el soma no permitiría el rápido relleno de las vesículas sinápticas durante la actividad neural continua. Por lo tanto, el reciclado local es muy apropiado para la anatomía peculiar de las neuronas, lo que brinda a las terminaciones nerviosas el medio para proporcionar el aporte continuo de vesículas sinápticas. Papel del calcio en la secreción de transmisores Como se observó en los experimentos de Katz y otros descritos en las secciones precedentes, la reducción de la concentración de Ca2+ en el exterior de una terminación nerviosa motora Gemación Anclaje Preparación Fusión Exocitosis Ca2+ 1 ms presináptica reduce el tamaño del PPT (compárense las Figuras 5.6B y D). Además, la medición de la cantidad de cuantos de transmisores liberados en esas condiciones muestra que la razón para que el PPT se haga más pequeño es que la reducción de la concentración de Ca2+ disminuye la cantidad de vesículas que se fusionan con la membrana plasmática de la terminación. Un paso importante en el conocimiento acerca de cómo el Ca2+ regula la fusión de las vesículas sinápticas fue el descubrimiento de que las terminaciones presinápticas tienen canales del Ca2+ con puerta de voltaje en sus membranas plasmáticas (véase Cap. 4). La primera indicación de los canales del Ca2+ presinápticos derivó de Katz y Ricardo Miledi. Estos autores observaron que las terminaciones presinápticas tratadas con tetrodotoxina (que bloquea los canales del Na+; véase Cap. 3) podían seguir produciendo un potencial de acción. La explicación para este hallazgo sorprendente fue que la corriente seguía fluyendo a través de los canales del Ca2+, que sustituían la corriente transportada comúnmente por los canales del Na+ bloqueados. Los experimentos posterior de pinzamiento de voltaje, realizados por Rodolfo Llinás y otros autores en una terminación presináptica gigante de calamar (Figura 5.10A), confirmaron la presencia de canales del Ca2+ con puerta de voltaje en la terminación presináptica (Figura 5.10B). Estos experimentos mostraron que la cantidad de neurotransmisor liberada es muy sensible a la cantidad exac- TRANSMISIÓN SINÁPTICA FIGURA 5.10 La entrada de Ca2+ a través de los canales del calcio con puerta de voltaje presináptica produce la liberación del transmisor. Registro (A) Potencial de membrana postsináptica Neurona presináptica Vpre Pinzamiento de voltaje 89 (A) Disposición experimental que utiliza una sinapsis extraordinariamente grande del calamar. El método de pinzamiento de voltaje detecta corrientes que fluyen a través de la membrana presináptica cuando el potencial de membrana es despolarizado. (B) Los agentes farmacológicos que bloquean las corrientes que fluyen a través de los canales del Na+ y del K+ ponen en evidencia una corriente remanente hacia el Neurona postsináptica interior que fluye a través de los canales del Ca2+. Este influjo de calcio desencadena la secreción del transmisor, según lo que indica un cambio en el potencial de membrana postsináptico. El tratamiento del mismo terminal presináptico con cadmio, un Ipre bloqueante de los canales del calcio, elimina tanto la corriente presináptica de calcio como la respuesta postsináptica. (De Augustine y Eckert, 1984.) (B) Potencial de membrana presináptica (mV) Corriente presináptica de calcio (+A/cm2) 0 –25 –50 –75 Bloqueador del canal del Ca2+ Control 0 –200 0 Potencial de membrana postsináptica (mV) FIGURA 5.11 Prueba de que una elevación en la concentración presináptica de Ca2+ desencadena la liberación del transmisor de las terminaciones presinápticas. (A) Mediciones con microscopia de fluorescencia de la concentración presináptica de –25 –50 –75 –3 0 3 6 9 12 –3 0 Tiempo (ms) 3 6 9 Ca2+ en la sinapsis gigante del calamar (véase Fig. 5.10A). Un tren de potenciales de acción presinápticos produce una elevación en la concentración de Ca2+, según lo demuestra un colorante (denomi- 12 nado fura-2) que da fluorescencia muy intensa cuando aumenta la concentración de Ca2+. (B) La microinyección de Ca2+ en la terminación presináptica gigante del calamar desencadena la liberación del transmisor, medida como una despolarización del potencial de mem- (A) brana postsináptico. (C) La microinyección de BAPTA, un quelante del Ca2+, en una terminación presináptica gigante de calamar impide 50 +m Ca2+ la liberación del transmisor. (A, de Smith y cols., 1993; B, de Miledi, 1971; C, de Adler y cols., 1991.) Inyección de Ca2+ Po tencial de membrana postsináptica (mV) Po tencial de membrana postsináptica (mV) (B) Potencial de membrana presináptica (mV) (C) < 64 < 65 0 1 2 Tiempo (s) 3 4 25 Inyección de quelador de Ca2 Control 0 <25 <50 <75 25 0 <25 <50 <75 0 1 2 3 4 5 0 Tiempo (ms) 1 2 3 4 5 90 CAPÍTULO 5 Neurotransmisor de molécula pequeña en vesículas pequeñas de centro claro FIGURA 5.12 Liberación diferencial de cotransmisores neuropeptídicos y de molécula pequeña. La estimulación de baja frecuencia eleva preferentemente la concentración de Ca2+ cerca de la membrana, lo que favorece la liberación del transmisor de las vesículas pequeñas de centro claro acopladas a especializaciones presinápticas. La estimulación de alta frecuencia conduce a un aumento más general en el Ca2+ que produce la liberación de neurotransmisores peptídicos desde vesículas grandes de centro denso, y neurotransmisores de molécula pequeña desde vesículas pequeñas de centro claro. Neuropéptido en vesículas grandes de centro denso Aumento localizado en la concentración de Ca2+ Estimulación de baja frecuencia Liberación preferencial de neurotransmisores de molécula pequeña Aumento más difuso en la concentración de Ca2+ ta de Ca2+ que ingresa. Además, el bloqueo de estos canales del Ca2+ con fármacos también inhibe la liberación de transmisores (Figura 5.10B, derecha). Todas estas observaciones confirman que los canales del Ca2+ con puerta de voltaje están involucrados directamente en la neurotransmisión. Por lo tanto, los potenciales de acción presinápticos abren los canales del Ca2+ con puerta de voltaje, con un influjo de Ca2+ resultante. El hecho de que el ingreso de Ca2+ en las terminaciones presinápticas eleve la concentración de Ca2+ dentro de la terminación ha sido documentado mediante imágenes microscópicas de terminaciones llenas con colorantes fluorescentes sensibles al Ca2+ (Figura 5.11A). Las consecuencias de que se eleve la concentración presináptica de Ca2+ para la liberación de los neurotransmisores se ha demostrado de dos modos. Primero, la microinyección de Ca2+ en las terminaciones presinápticas desencadena la liberación de transmisores en ausencia de potenciales de acción presinápticos (Figura 5.11B). Segundo, la microinyección presináptica de quelantes del calcio (sustancias químicas que fijan Ca2+ y mantienen amortiguada su concentración en bajos niveles) impide que los potenciales de acción presinápticos produzcan secreción del transmisor (Figura 5.11C). Sin duda alguna, estos resultados prueban que una elevación en la concentración presináptica de Ca2+ es necesaria y suficiente para la liberación de los neurotransmisores. Por lo tanto, como sucede con muchas otras formas de señalización neuronal (véase Cap. 7), el Ca2+ sirve como segundo mensajero durante la liberación del transmisor. Aunque el Ca2+ es un desencadenante universal para la liberación de transmisores, no todos los transmisores son liberados con la misma velocidad. Por ejemplo, aunque la secreción de ACh de las neuronas motoras sólo requiere una fracción de un milisegundo (véase Fig. 5.6), la liberación de neuropéptidos requiere descargas de alta frecuencia de potenciales de acción durante muchos segundos. Estas diferencias en la velocidad de liberación probablemente surgen de diferencias en la disposición espacial de las vesículas en relación con los canales del Ca2+presinápticos. Estimulación de alta frecuencia Liberación de ambos tipos de transmisores Tal vez esto sea más evidente en los casos en que las moléculas pequeñas y los péptidos sirven como cotransmisores (Figura 5.12). Aunque típicamente las vesículas pequeñas de centro claro que contienen transmisores de molécula pequeña se encuentran acopladas en la membrana plasmática antes del ingreso de Ca2+, las vesículas grandes de centro denso que contienen transmisores peptídicos están más alejadas de la membrana plasmática (véase Fig. 5.5D). Con bajas frecuencias de descarga, la concentración de Ca2+ puede aumentar sólo de modo local en la membrana plasmática presináptica, en la vecindad de los canales del Ca2+ abiertos, lo que limita la liberación a transmisores de molécula pequeña desde las pequeñas vesículas de centro claro acopladas. La estimulación prolongada de alta frecuencia aumenta la concentración de Ca2+ en toda la terminación presináptica, lo que induce la liberación más lenta de neuropéptidos. Mecanismos moleculares del ciclo de las vesículas sinápticas No se sabe exactamente cómo un aumento en la concentración presináptica de Ca2+ continúa hasta desencadenar la fusión de las vesículas y la liberación del neurotransmisor. Sin embargo, se han obtenido muchos indicios importantes de estudios moleculares que han identificado y caracterizado las proteínas que se encuentran en las vesículas sinápticas (Figura 5.13A) y sus patrones de fijación sobre la membrana plasmática presináptica y el citoplasma. La mayoría de estas proteínas, sino todas ellas, actúan en uno o más pasos en el ciclo de las vesículas sinápticas. TRANSMISIÓN SINÁPTICA (A) SNAP25 Vti1a Sinaptobrevina FIGURA 5.13 Proteínas presinápticas y sus funciones en el ciclo de las vesículas sinápticas. Sinaptotagmina ATPasa V (A) Modelo de la organización molecular de una vesícula sináptica. La superficie citoplasmática de la membrana CIC3 vesicular se encuentra densamente cubierta por proteínas, de las cuales sólo se muestra aquí el 70%. (B) El ciclo del tráfico de vesículas que se muestra en la Figura 5.9E se sabe ahora que está mediado por algunas proteínas presinápticas, que incluyen algunas de las que se muestran en (A), y diferentes proteínas participan en Sinaptofisina CSP SNAP29 SV2 VAMP4 SCAMP Sintaxina diferentes reacciones. (A, de Takamori y cols., 2006.) Sinapsina (m)unc18 VGLUT Rab3A GTPasatrimérica Otros transportadores (B) Carga de transmisores Bomba de protones de los transportadores de transmisores Pérdida de la cubierta Clatrina Auxilina Hsc-70 Gemación Endofilina Sinaptojanina Gemación Movilización Dinamina Anfifisina Endofilina Sinaptojanina Sinapsinas Actina Pool de reserva Clatrina Actina Sindapina WASP Sinapsinas Actina Anclaje Cubierta Proteína fijadora de ATP SNARE Clatrina AP-2 AP-180 Sinaptotagminas Sinaptobrevina Ca2+ Fusión Preparación SNARE (m)unc13 (m)unc18/nSec1 NSF SNAP Complexina Tomosina CAPS SV2 Snapina Sintafilina Rab3a RIM Doc2 Sinaptotagmina 1 SNARE Epsina Eps-15 Endofilina Stoned NSF SNAP Sintafilina SNIP 91 92 CAPÍTULO 5 (B) (1) (A) Membrana de la vesícula sináptica Sinaptotagmina Sinaptotagmina Sinaptobrevina Vesícula Canal del Ca2+ Sinaptobrevina Ca2+ Sintaxina SNAP-25 (2) Se forman complejos SNARE para reunir las membranas SNAP-25 Sintaxina Membrana plasmática presináptica (3) El Ca2+ que ingresa se une a la sinaptotagmina FIGURA 5.14 Mecanismos moleculares de exocitosis durante la liberación del neurotransmisor. (A) Estructura del complejo SNARE. El SNARE vesicular, la sinaptobrevina (azul), forma un complejo helicoidal con los SNARE de la membrana plasmática sintaxina (rojo) y SNAP-25 (verde). Se muestra también la estructura de la sinaptotagmina, una proteína vesicular fijadora de Ca2+, con el Ca2+ ligado indicado por esferas. (B) Modelo para la fusión vesicular desencadenada por Ca2+. Las proteínas SNARE sobre la vesícula sináptica y la membrana plasmática forman un complejo (como sucede en A) que aproxima las dos membranas. El Ca2+ se une entonces a sinaptotagmina, lo que hace que la región citoplasmática de esta proteína catalice la fusión de la membrana al unirse a SNARE e insertarlos en la membrana plasmática. (A, de Chapman, 2002.) Aunque falta todavía un cuadro molecular completo de la liberación de los neurotransmisores, se han deducido los papeles de varias proteínas involucradas en el ciclo de las vesículas (Figura 5.13B). Varias líneas de evidencia indican que la proteína sinapsina, que se une de forma reversible a las vesículas sinápticas, puede mantener fijadas estas vesículas dentro del pool de reserva uniendo a las vesículas con enlaces cruzados entre sí y a filamentos de actina en el citoesqueleto. La movilización de estas vesículas desde el pool de reserva es causada por la fosforilación de la sinapsina por proteincinasas, principalmente la proteincinasa dependiente de Ca2+/calmodulina, tipo II (CaMKII; véase Cap. 7), que permite que la sinapsina se disocie de las vesículas. Una vez que las vesículas se encuentran libres de sus fijaciones del pool de reserva, se dirigen hacia la membrana plasmática y se fijan luego a esta membrana mediante reacciones de anclaje poco conocidas. Una serie de reacciones de impresión prepara entonces las membranas vesicular y plasmática para la fusión. Una gran cantidad de proteínas participan en la preparación e incluyen a algunas que también participan en otros tipos de acontecimientos de fusión de la membrana comunes a todas ellas (véase Fig. 5.13B). Por ejemplo, dos proteínas originariamente consideradas importantes para la fusión de las vesículas con la Ca2+ (4) La sinaptotagmina ligada al Ca2+ cataliza la fusión al unirse a SNARE y a la membrana plasmática membrana del aparato de Golgi, la ATPasaNFS (NEM-sensitive fusion protein, proteína de fusión sensible al NEM) y SNAP (soluble NFS-attachment proteins, proteínas solubles de fijación al NFS), también participan en la preparación de las vesículas sinápticas para la fusión. Estas dos proteínas funcionan regulando la reunión de otras proteínas que se denominan SNARE (SNAP receptores, receptores del SNAP). Muchas de las otras proteínas que participan en la preparación –como munc-13, nSec-1, complexina, snapina, sintafilina y tomosina– también interactúan con los SNARE. Uno de los propósitos principales de la preparación parece ser organizar las proteínas SNARE en la conformación correcta para la fusión de la membrana. Una de las proteínas SNARE, la sinaptobrevina, se encuentra en la membrana de las vesículas sinápticas, mientras que otras dos proteínas SNARE lla- TRANSMISIÓN SINÁPTICA 93 RECUADRO 5B Enfermedades que afectan la terminación presináptica Distintos pasos en la exocitosis y la endocitosis de las vesículas sinápticas son puntos diana de algunas enfermedades neurológicas raras pero debilitantes. Síndromes miasténicos En los síndromes miasténicos, la transmisión anormal en las sinapsis neuromusculares conduce a debilidad y fatigabilidad de los músculos esqueléticos (véase Recuadro 7B). Uno de los ejemplos mejor conocidos de estos trastornos es el síndrome miasteniforme de Eaton-Lambert, complicación ocasional de pacientes con ciertos tipos de cáncer. Biopsias de tejido muscular tomadas de pacientes con síndrome miasteniforme de Eaton-Lambert permiten efectuar registros intracelulares idénticos a los que se muestran en la Figura 5.6. Estos registros han demostrado que, cuando se estimula una neurona motora, se reduce mucho el número de cuantos contenidos en los PPT individuales, aunque la amplitud de los PPTM espontáneos es normal. Por lo tanto, el síndrome miasteniforme de Eaton-Lambert deteriora la liberación provocada por el neurotransmisor, pero no afecta el tamaño de los cuantos individuales.Varias líneas de evidencia indican que esta reducción en la liberación del neurotransmisor se debe a una pérdida de canales del Ca2+ con puerta de voltaje en la terminación presináptica de las neuronas motoras (véase Fig. A); tal vez sean más notables los estudios anatómicos que ponen en evidencia una densidad reducida de proteínas de los canales de Ca2+ en la membrana plasmática presináptica. La pérdida de canales del Ca2+ presinápticos en los pacientes con síndrome miasteniforme de Eaton-Lambert aparentemente surge por un trastorno autoinmunitario. La sangre de estos pacientes tiene una concentración muy alta de anticuerpos que se unen a los canales del Ca2+ y parece probable que estos anticuerpos sean la causa primaria del síndrome miasteniforme de Eaton-Lambert. Por ejemplo, la eliminación de los anticuerpos contra los canales del Ca2+ de la sangre de pacientes con síndrome miasteniforme de Eaton-Lambert mediante plasmaféresis reduce la debilidad muscular. Asimismo, los agentes inmunosupresores también pueden aliviar los síntomas del síndrome. sinápticas en el interior de la terminación Tal vez lo más contundente sea que la inyec- de la neurona motora. Las sinapsis neuroción de estos anticuerpos en animales de ex- musculares en algunos de estos pacientes perimentación produce debilidad muscular tienen PPT con un contenido cuántico rey una transmisión neuromuscular anormal. ducido, déficit especialmente prominente No está claro por qué el sistema inmunita- cuando la sinapsis es estimulada repetidas rio genera anticuerpos contra los canales veces. La microscopia electrónica muestra del Ca2+. La mayoría de los pacientes con que las terminaciones nerviosas motoras síndrome miasteniforme de Eaton-Lambert presinápticas tienen una cantidad muy retienen un carcinoma de células pequeñas, ducida de vesículas sinápticas. El defecto una forma de cáncer de pulmón que puede en la liberación del neurotransmisor eviiniciar de algún modo la respuesta inmuni- dentemente es el resultado de una cantitaria a los canales del Ca2+. Cualquiera sea dad insuficiente de vesículas sinápticas el origen, la unión de los anticuerpos a los disponibles para la liberación durante la canales del Ca2+ produce una reducción en actividad presináptica sostenida. Los orílas corrientes de los canales del Ca2+. Es este genes de esta escasez de vesículas no están defecto inducido por los anticuerpos en la claros, pero podría ser un resultado de un entrada presináptica de Ca2+, lo que explica deterioro en la endocitosis en la terminala debilidad muscular asociada con el sín- ción nerviosa (véase Fig. A) o por un aporte reducido de vesículas desde el cuerpo drome miasteniforme de Eaton-Lambert. Los síndromes miasténicos congénitos celular de una neurona motora. Otros pacientes que padecen miastenia son trastornos genéticos que, al igual que el síndrome miasteniforme de Eaton-Lam- familiar del lactante parecen tener una bert, también producen debilidad muscu- debilidad neuromuscular que surge por lar al afectar la transmisión neuromuscu- reducciones en el tamaño de los cuantos lar. Algunos de estos síndromes afectan la individuales, más que en la cantidad de acetilcolinesterasa que degrada la acetilco- cuantos liberados. Las terminaciones nerlina en la hendidura sináptica, mientras que viosas motoras de estos pacientes tienen otros surgen por el (Continúa en la página siguiente) ataque autoinmuniLos síndromes miasténicos congénitos co nducen a un deterioro del re ciclado tario de los recepde vesículas tores colinérgicos (véase Recuadro Endosoma 6B). Sin embargo, algunos síndromes miasténicos congénitos surgen por defectos en la liberación de acetilcolina debido a una alteración Gemación del tráfico de vesículas Gemación (A) Puntos diana Anclaje Fusión presinápticos de distintos trastornos neurológicos. La latrotoxina desencadena la fusión de la vesícula Preparación Ca2+ El síndrome miasteniforme de Eaton-Lambert ataca los canales del Ca2+ presinápticos Los trastornos cognitivos deterioran la señalización transináptica Las toxinas botulínica y tetánica afectan las proteínas SNARE involucradas en la fusión de las vesículas 94 CAPÍTULO 5 RECUADRO 5B (continúa) vesículas sinápticas en cantidad normal, pero de tamaño más pequeño. Este hallazgo sugiere un tipo diferente de lesión genética que altera de alguna forma la formación de nuevas vesículas sinápticas tras la endocitosis y deja así menos acetilcolina en cada vesícula. Botulismo y tétanos El deterioro de la liberación sináptica del transmisor es el resultado del envenenamiento por bacterias anaerobias Clostridium. Este género de microorganismos produce algunas de las toxinas más potentes conocidas, que incluyen varias toxinas botulínicas y la toxina tetánica. Tanto el botulismo como el tétanos son trastornos potencialmente fatales. El botulismo se puede desarrollar por consumir alimentos que contienen bacterias Clostridium o por la infección de heridas con las esporas de estos organismos ubicuos. En cualquier caso, la presencia de la toxina puede producir parálisis de las sinapsis neuromusculares periféricas debido a la abolición de la liberación de neurotransmisores. Esta interferencia con la transmisión neuromuscular hace que el músculo esquelético se debilite, y en los casos extremos produce parálisis del diafragma y otros músculos necesarios para la respiración. Las toxinas botulínicas también bloquean las sinapsis que inervan los músculos lisos de varios órganos, y dan origen a disfunción motora visceral. En los casos típicos, el tétanos es el resultado de la contaminación de heridas punzantes por bacterias Clostridium que producen toxina tetánica. Al contrario del botulismo, la intoxicación tetánica bloquea la liberación de transmisores inhibidores desde las interneuronas en la médula espinal. Este efecto produce una pérdida de inhibición sináptica sobre las neuronas motoras espinales, lo que genera hiperexcitación del músculo esquelético y contracciones tetánicas en los músculos afectados (de ahí el nombre de la enfermedad). Aunque sus consecuencias clínicas son espectacularmente diferentes, las toxinas de los clostridios poseen un mecanismo de acción común: son proteasas altamente específicas que inhiben la liberación de neurotransmisores al dividir las proteínas SNARE que participan en la fusión de las vesículas sinápticas con la membrana plasmática presináptica (Fig. B). La toxina tetánica y las toxinas del botulismo tipos B, D, F y G dividen específicamente la proteína SNARE sinaptobrevina de las vesículas. Otras toxinas botulínicas dividen a la sintaxina (tipo C) y SNAP-25 (tipos A y E), proteínas SNARE halladas sobre la membrana plasmática presináptica. La destrucción de estas proteínas presinápticas es la base para las acciones inhibidoras de las toxinas de los clostridios sobre la liberación de los neurotransmisores. En apariencia, las diferentes acciones de estas toxinas sobre la transmisión sináptica en las sinapsis motoras excitadoras versus inhibidoras es el resultado del hecho de que estas toxinas son captadas por diferentes tipos de neuronas: mientras las toxinas botulínicas son captadas por neuronas motoras, la toxina tetánica está dirigida preferencialmente hacia las interneuronas. Se presume que la captación diferencial de las toxinas surge de la presencia de diferentes tipos de receptores de toxina sobre dos tipos de neuronas. Conocimientos a partir de la α-latrotoxina El lactrodectismo se refiere al dolor muscular intenso y los calambres experimentados por los pacientes mordidos por la araña viuda negra, Latrodectus. Estos síntomas surgen por una neurotoxina presináptica llamada α-latrotoxina, presente en el veneno de la araña viuda negra; esta toxina produce una descarga masiva de neurotransmisor desde las terminaciones presinápticas. Aunque la liberación de los neurotransmisores habitualmente requiere Ca2+, la α-latrotoxina es capaz de liberar neurotransmisor incluso cuando el Ca2+ está ausente del medio extracelular. Aunque aún no está claro el modo en que la toxina desencadena la exocitosis independiente de Ca2+, sabemos que la α-latrotoxina se une a dos tipos diferentes de proteínas presinápticas que pueden mediar sus acciones. Un grupo de compañeros de unión para la α-latrotoxina son las neurexinas, grupo de proteínas integrales de la membrana hallado en las terminaciones presinápticas. Las neurexinas se unen a las sinaptotagmina, el sensor presináptico de Ca2+, y esta interacción puede permitir a la α-latrotoxina evitar el requerimiento habitual de Ca2+ para desencadenar la fusión de las vesículas. Otro tipo de proteína presináptica que puede Membrana de la vesícula sináptica Sinaptobrevina BoTX-G TeTX BoTX-B BoTX-D BoTX-F BoTX-A BoTX-C SNAP-25 BoTX-E (B) División de las proteínas SNARE por las toxinas de los clostridios. Se indican los sitios de proteólisis de la toxina tetánica (TeTX) y distintos tipos de toxina botulínica (BoTX). (De Sutton y cols., 1998.) Sintaxina Membrana plasmática presináptica TRANSMISIÓN SINÁPTICA 95 RECUADRO 5B (continúa) unirse a la α-latrotoxina se denomina CL1 (sobre la base de sus nombres anteriores, receptor independiente de Ca2+ para latrotoxina y latrofilina-1). CL1 es un pariente de los receptores acoplados a la proteína G que median las acciones de neurotransmisores y otras señales químicas extracelulares (véase Cap. 7). Por lo tanto, la unión de α-latrotoxina a CL1 podría activar una cascada de transducción de señales intracelulares involucrada en las acciones Ca2+independientes de la α-latrotoxina. Aunque se necesitan otras investigaciones para establecer los papeles definitivos de las neurexinas y CL1 en la acción de α-latrotoxina, es probable que estas dos proteínas constituyan la base para la potente actividad presináptica de la toxina. Además de su posible papel en el latrodectismo, las neurexinas han sido vinculadas a distintos trastornos cognitivos. Se han identificado mutaciones en el gen de la neurexina en algunos pacientes que padecen esquizofrenia, una enfermedad psiquiátrica que produce delirios, alucinaciones y pérdida de expresión emocional (véase Recuadro 18D). También se sabe que las neurexinas desempeñan un papel importante en la señalización a través de la hendidura sináptica al unirse a una familia de proteínas de membrana postsinápticas denominadas neuroliginas. Es importante señalar que mutaciones en las neuroliginas se han asociado con autismo, distintos trastornos psiquiátricos caracterizados por deterioro de la interacción social, problemas de la comunicación y otros trastornos de la conducta. Aunque no se ha establecido aún una conexión directa entre neurexinas/neuroliginas y estos trastornos psiquiátricos, parece probable que defectos en estos patrones de señalización transináptica puedan servir como mecanismo central subyacente a algunos trastornos de conducta. En resumen, no sólo la investigación sobre el tráfico de vesículas sinápticas ilumina las causas de algunos trastornos neurológicos y psiquiátricos, sino que a su vez la investigación sobre estas enfermedades ha proporcionado herramientas, como las toxinas de los clostridios y la α-latrotoxina, que han probado ser útiles para dilucidar los mecanismos básicos de tráfico de vesículas sinápticas. Bibliografía Engel, A. G. (1991) Review of evidence for loss of motor nerve terminal calcium channels in Lambert-Eaton myasthenic syndrome. Ann. N.Y. Acad. Sci. 635: 246-258. Engel, A. G. (1994) Congenital myasthenic syndromes. Neurol. Clin. 12: 401-437. Grumelli, C., C. Verderio, D. Pozzi, O. Rossetto, C. Montecucco and M. Matteoli (2005) Internalization and mechanism of action of clostridial toxins in neurons. Neurotoxicology 26: 761-767. Humeau, Y., F. Doussau, N. J. Grant and B. Poulain (2000) How botulinum and tetanus neurotoxins block neurotransmitter release. Biochimie 82: 427-446. Maselli, R. A. (1998) Pathogenesis of human botulism. Ann. N.Y. Acad. Sci. 841: 122-139. Silva, J. P., J. Suckling and Y. Ushkaryov (2009) Penelope‘s web: Using α-latrotoxin to untangle the mysteries of exocytosis. J. Neurochem. 111: 275-290. Südhof, T. C. (2008) Neuroligins and neurexins link synaptic function to cognitive disease. Nature 455: 903-911. Sutton, R. B., D. Fasshauer, R. Jahn and A. T. Brünger (1998) Crystal structure of a SNARE complex involved in synaptic exocytosis at 2.4 Å resolution. Nature 395: 347-353. Vincent, A. (2010) Autoimmune channelopathies: Well-established and emerging immunotherapy-responsive diseases of the peripheral and central nervous systems. J. Clin. Immunol. 30 (Suppl. 1): S97-S102. Dolly, J. O. and K. R. Aoki (2006) The structure and mode of action of different botulinum toxins. Eur. J Neurol. Suppl 4: 1-9. madas sintaxina y SNAP-25 se encuentran fundamentalmente en la membrana plasmática. Estas proteínas SNARE pueden formar un complejo macromolecular que se extiende por las dos membranas, colocándolas en estrecha aposición (Figura 5.14A). Esta disposición es muy apropiada para promover la fusión de las dos membranas, y según varias líneas de evidencia es lo que ocurre realmente. Una observación importante es que las toxinas que separan las proteínas SNARE bloquean la liberación de neurotransmisor (Recuadro 5B). Además, colocar las proteínas SNARE en membranas lipídicas artificiales y permitir que estas proteínas formen complejos entre ellas hace que las membranas se fusionen. Como las proteínas SNARE no fijan Ca2+, otras moléculas deben ser responsables de la regulación de la liberación de los neurotransmisores. Varias proteínas presinápticas, que incluyen calmodulina, CAPS y munc-13, son capaces de fijar Ca2+. Sin embargo, el candidato principal para regular la liberación de neurotransmisores por el Ca23+ parece ser la sinaptotagmina, proteína hallada en la membrana de las vesículas sinápticas (véase Fig. 5.14A). La sinaptotagmina se une al Ca2+ en con- centraciones similares a las requeridas para desencadenar la fusión vesicular en el interior de la terminación presináptica y esta propiedad le permite a la sinaptotagmina actuar como sensor del Ca2+, señalando la elevación del Ca2+ en el interior de la elevación para desencadenar así la fusión vesicular. En apoyo de esta idea, la interrupción de la sinaptotagmina en las terminaciones presinápticas de ratones, moscas de la fruta, calamar y otros animales de experimentación deteriora la liberación de neurotransmisores Ca2+-dependiente. De hecho, la deleción de sólo uno de los 19 genes de sinaptotagmina de los ratones es una mutación letal, que hace que los ratones mueran poco después del nacimiento. No está claro el modo en que la fijación del Ca2+ a la sinaptotagmina podría conducir a exocitosis. Se sabe que el Ca2+ cambia las propiedades químicas de la sinaptotagmina y permite que se inserte en las membranas y se una a otras proteínas SNARE. Un modelo admisible es que las proteínas SNARE aproximan mucho las dos membranas y que los cambios inducidos por el Ca2+ en la sinaptotagmina producen entonces la fusión final de estas membranas (Figura 5.14B). 96 (A) CAPÍTULO 5 FIGURA 5.15 Mecanismos moleculares de endocitosis que siguen a la liberación de neurotransmisores. (A) Las trisquelias individuales de clatrina (izquierda) se reúnen para Trisquelia de clatrina formar cubiertas de membrana (derecha) que participan en la gemación de la membrana durante la endocitosis. (B) Modelo de la gemación de la membrana durante la endocitosis. Después del agregado de la membrana de la vesícula sináptica durante la exocitosis, las trisquelias de clatrina se unen a la membrana vesicular. Proteínas adaptadoras, como AP-2 y AP180, ayudan a su fijación. La polimerización de la clatrina hace que la membrana se curve, lo que permite que la dinamina separe la vesícula con cubierta. La pérdida posterior de la cubierta de la vesícula, por Hsc-70 y auxilina, da una vesícula sináptica. (A, de Marsh y McMahon, 1999.) Cubierta de clatrina (B) Citosol Actina Clatrina Hsc-70/auxilina Exterior de la célula 1 Proteínas adaptadoras conectan la clatrina a la membrana vesicular Dinamina 2 Trisquelias de clatrina se reúnen en una cubierta que cubre la membrana 3 Se forma el anillo de dinamina que se separa de la membrana Aun otras proteínas parecen participar en los pasos posterior del ciclo de las vesículas sinápticas (Figura 5.15). La proteína más importante involucrada en la gemación endocitósica de vesículas desde la membrana plasmática es la clatrina. Esta proteína tiene una estructura singular denominada trisquelia porque tiene un aspecto de tres patas (Figura 5.15A). Durante la endocitosis, las trisquelias de la clatrina se unen a la membrana vesicular que va a ser recuperada (Figura 5.15B). Algunas proteínas, como AP2 y AP180, conectan la clatrina a las proteínas y los lípidos de la membrana. Estas proteínas adaptadoras, así como otras proteínas como la anfifisina, la epsina y Eps-15, ayudan a reunir las trisquelias individuales en estructuras que se asemejan a cúpulas geodésicas (véase Figura 5.15A). Estas estructuras cupuliformes forman fositas con cubierta que inician la gemación de la membrana y aumentan la curvatura de la membrana en gemación hasta que forma una estructura similar a una vesícula con cubierta. Otra proteína, llamada dinamina, produce la separación final de la membrana, que completa la producción de las vesículas con cubierta. Las cubiertas de la clatrina son eliminados entonces por una ATPasa, Hsc70, con otra proteína llamada auxilina, que sirve como cofactor que recluta Hsc70 para la vesícula con cubierta. Otras proteínas, como sinaptojanina, también son importantes para la pérdida de revestimiento de las vesículas. Las vesículas sin cubierta pueden continuar entonces su viaje a través del proceso de reciclado, para ser rellenadas finalmente con neurotransmisor debido a las acciones de los transportadores de neurotransmisores en la membrana vesicular. Estos transportadores intercambian protones en el interior de la vesícula por neurotransmisor; el interior ácido de la vesícula es producido por una bomba de protones que también se localiza en la membrana vesicular. 4 Vesícula con cubierta translocada por filamentos de actina 5 Hsc-70 y auxilina eliminan la cubierta de la vesícula En resumen, una cascada compleja de proteínas que actúan en un orden temporal y espacial definido permite que las neuronas secreten transmisores. Aunque los mecanismos moleculares responsables de la secreción de transmisores no están completamente claros, en la actualidad se ha identificado a la mayoría de las proteínas involucradas en este proceso. Receptores de neurotransmisores La generación de señales eléctricas postsinápticas también se conoce en considerable profundidad. Estos estudios comenzaron en 1907, cuando el fisiólogo británico John N. Langley introdujo el concepto de moléculas receptoras para explicar las acciones específicas y potentes de ciertas sustancias químicas sobre las células musculares y nerviosas. En la actualidad, sabemos que los receptores de los neurotransmisores son proteínas introducidas en la membrana plasmática de las células postsinápticas y tienen un sitio fijador para los neurotransmisores extracelulares que detecta la presencia de neurotransmisores en la hendidura sináptica. Existen dos familias amplias de proteínas receptoras que difieren en su mecanismo de traducir la unión de los transmisores en las respuestas postsinápticas. Los receptores de una familia contienen un dominio que atraviesa la membrana y forma un canal iónico (Figura 5.16A). Estos receptores combinan funciones de unión a transmisores y canales en una sola entidad molecular y, por lo tanto, se denominan receptores inotrópicos (el griego tropos significa moverse en respuesta a un estímulo) o canales iónicos con puerta de ligando. La segunda familia de receptores de neurotransmisores son los receptores metabotrópicos, denominados así porque el movi- TRANSMISIÓN SINÁPTICA 97 (B) Receptores acoplados a proteína G (A) Canales iónicos con puerta de ligando Se une el Iones 1 neurotransmisor 1 Se une el neurotransmisor Neurotransmisor Receptor 2 El canal se abre Proteína efectora Exterior de la célula 5 Los iones fluyen a través de la membrana Interior de la célula ` _ Proteína G 3 Los iones fluyen a través de la membrana 2 La proteína G FIGURA 5.16 Dos tipos diferentes de receptor de neurotransmisores. (A) Los canales iónicos con puerta de ligando combinan las funciones de receptor y de canal en un único complejo proteico. (B) Los receptores metabotrópicos suelen activar proteínas G, que modulan los canales iónicos directa o indirectamente a través de enzimas efectoras intracelulares y segundos mensajeros. miento eventual de los iones a través de un canal depende de los pasos metabólicos interpuestos. Estos receptores no poseen canales iónicos como parte de su estructura; en cambio, tienen un dominio intracelular que afecta indirectamente los canales a través de la activación de moléculas intermedias denominadas proteínas G (Figura 5.16B). La unión del neurotransmisor a estos receptores activa proteínas G, las que entonces se disocian del receptor e interactúan directamente con los canales iónicos o se unen a otras proteínas efectoras, como las enzimas, para formar mensajeros intracelulares que abren o cierran canales iónicos. Por esta razón, los receptores metabotrópicos también son denominados receptores acoplados a la proteína G. Los acontecimientos de señalización postsináptica iniciados por los receptores metabotrópicos se tratan en detalle en el Capítulo 7. Estas dos familias de receptores postsinápticos dan origen a acciones postsinápticas que varían desde menos de un milisegundo hasta minutos, horas o incluso días. Por lo general, los receptores ionotrópicos median efectos postsinápticos rápidos. Son ejemplos los PPT producidos en las sinapsis neuromusculares por la ACh (véase Fig. 5.6B), así como las respuesta postsinápticas producidas en ciertas sinapsis glutaminérgicas y GABAérgicas (véase Fig. 5.21B). En estos casos, los potenciales postsinápticos se originan dentro del milisegundo o dos de un potencial de acción que invade la terminación presináptica es activada a _ Me ns aj er os i ntr ace lul ar e s 3 Subunidades de proteína G o mensajeros intracelulares modulan los canales iónicos 4 Se abre el canal iónico Iones y duran sólo algunas decenas de milisegundos o menos. Por el contrario, típicamente la activación de los receptores metabotrópicos produce respuestas mucho más lentas, que varían desde cientos de milisegundos hasta minutos o incluso más. La lentitud comparativa de las acciones de los receptores metabotrópicos refleja el hecho de que es necesaria la unión de múltiples proteínas entre sí de manera secuencial para producir la respuesta fisiológica final. Es importante señalar que un transmisor dado puede activar tanto receptores ionotrópicos como metabotrópicos para producir potenciales postsinápticos rápidos y lentos en la misma sinapsis. Cambios en la permeabilidad de la membrana postsináptica durante la transmisión sináptica Al igual que los estudios de la sinapsis neuromuscular marcaron el camino para conocer los mecanismos de liberación de los neurotransmisores, esta sinapsis periférica ha sido igualmente útil para conocer los mecanismos que permiten que los receptores de neurotransmisores generen señales postsinápticas. La unión de ACh a los receptores postsinápticos abre los canales iónicos en la membrana de la fibra muscular. Este efecto puede ser demostrado directamente utilizando el método de pinzamiento zonal de membrana (véase Recuadro 4A) para medir las corrientes postsinápticas diminutas que fluyen cuando las dos moléculas de la ACh individual se unen a receptores, como Erwin Ehner y Bert Sakmann hicieron por primera vez en 1976. La exposición de la superficie extracelular de un parche de membrana postsináptica a ACh ocasiona que fluyan corrientes de canal único durante algunos milisegundos (Figura 5.17A). Esto muestra 98 CAPÍTULO 5 (A) Medición por pinzamiento de membrana de una única corriente de receptor de ACh (B) Corrientes producidas por: Micropipeta Receptor de ACh 2 4 (2) POCOS CANALES ABIERTOS Na+ Número de canales abiertos 2 +M de acetilcolina 0 Canal cerrado 2 Canal abierto 0 0 10 20 (3) TODOS LOS CANALES ABIERTOS I (pA) 0 0 1 2 2 4 6 8 10 12 Tiempo (ms) FIGURA 5.17 Activación de los receptores colinérgicos en las sinapsis neuromusculares. (A) Medición de corrientes de receptor colinérgico único con el método de pinzamiento zonal extracelular de una porción de membrana extraída de la célula muscular postsináptica. Cuando se aplica ACh a la superficie extracelular de la membrana pinzada en voltajes negativos, la apertura breve repetida de un único canal puede observarse como corrientes hacia el interior (es decir, iones positivos que fluyen en la célula). (B) Apertura sincronizada de muchos canales activados por ACh en una sinapsis con pinzamiento de voltajes negativos. (1) Si se examina un solo canal durante la liberación de ACh desde la terminación presináptica, el canal se abre transitoriamente. (2) Si se examinan juntos algunos canales, la liberación de ACh abre los canales casi sincrónicamente. (3) La apertura de muchos canales postsinápticos se suma para producir una corriente de placa terminal macroscópica. (C) En una célula muscular normal (es decir, sin pinzamiento de voltaje), la corriente de placa terminal hacia dentro despolariza la célula muscular postsináptica y da origen a un PPT. En el caso típico, esta despolarización genera un potencial de acción (no se muestra). que la unión de ACh a sus receptores abre canales iónicos con puerta de ligando, de forma muy similar a la manera en que los cambios en el potencial de membrana abren los canales iónicos con puerta de voltaje (véase Cap. 4). Las acciones eléctricas de la ACh se multiplican cuando un potencial de acción en una neurona motora presináptica produce la liberación de millones de moléculas de ACh en la hendidura sináptica. En este caso más fisiológico, las moléculas transmisoras se unen a muchos miles de receptores de ACh empaquetados en un denso conjunto sobre la membrana postsináptica, que abren transitoriamente una cantidad muy grande de canales iónicos postsinápticos. Aunque los receptores de ACh individuales sólo se abren brevemente (Fig. 5.17B1), la apertura de una gran cantidad de canales está sincronizada por la breve duración durante la cual se secreta ACh desde las terminaciones presinápticas (Figura 5.17B2, 3). La corriente macroscópica resultante de la apertura sumada de muchos canales iónicos se denomina corriente de placa terminal. Como el flujo de corriente durante la corriente de placa terminal normalmente es hacia el interior, el 0 Número de canales abiertos 0 Corriente de membrana (pA) Zona de membrana con el exterior hacia fuera ACh 0 Número de canales abiertos (1) CANAL ABIERTO ÚNICO Liberación de ACh por la estimulación de neurona motora 200.000 600.000 300.000 –2 0 2 4 6 8 Tiempo (ms) 10 12 14 (C) Cambio en el potencial postsináptico (PPS) producido por la corriente de placa terminal <70 Potencial de <80 membrana <90 (mV) <0 –2 0 2 4 6 8 Tiempo (ms) 10 12 14 potencial de membrana postsináptico se despolariza. Este cambio despolarizante en el potencial es el PPT (Figura 5.175C), el cual de manera típica desencadena un potencial de acción postsináptico al abrir canales del Na+ y del K+ con puerta de voltaje (véase Fig. 5.6B). La identidad de iones que fluyen durante la corriente de placa terminal puede determinarse mediante los mismos enfoques utilizados para identificar los papeles de los flujos de Na+ y K+ en las corrientes subyacentes a los potenciales de acción (véase Cap. 3). La clave para este análisis es la identificación del potencial de membrana en el cual no fluye ninguna corriente durante la acción de los transmisores. Cuando el potencial de la célula muscular postsináptica se controla mediante el método de pinzamiento de voltaje (Figura 5.18A), la magnitud del potencial de membrana afecta claramente la amplitud y la polaridad de las corrientes de placa terminal (Figura 5.18B). Por lo tanto, cuando el potencial de membrana postsináptico se vuelve más negativo que el potencial de reposo, la amplitud de la corriente de placa terminal se vuelve más grande, mientras que la corriente está reducida cuando el potencial de membrana se vuelve más positivo. Aproximadamente en 0 mV, no se detecta ninguna corriente de placa terminal, e incluso con potenciales más positivos, la corriente invierte su polaridad y se vuelve hacia el exterior en lugar de hacerlo hacia el interior (Figura 5.18C). El potencial donde 99 TRANSMISIÓN SINÁPTICA la corriente de placa terminal se invierte se denomina potencial de reversión. Como sucedió con las corrientes que fluían a través de los canales iónicos con puerta de voltaje (véase Cap. 3), la magnitud de la corriente de placa terminal en cualquier potencial de membrana está dada por el producto de la conductancia iónica activada por ACh (gACh) y la fuerza impulsora electroquímica sobre los iones que fluyen a través de canales con puerta de ligando. Por lo tanto, el valor de la corriente de placa terminal está dado por la relación donde Erev es el potencial de reversión para la corriente de placa terminal. Esta relación predice que la de placa terminal será una corriente hacia el interior con potenciales más negativos que Erev porque la fuerza impulsora electroquímica, Vm − E , es un rev FIGURA 5.18 Influencia del potencial de membrana postsináptico sobre las corrientes de placa terminal. (A) Se realiza un pinzamiento de voltaje en una fibra muscular postsináptica mediante utilización de dos electrodos, mientras que la neurona presináptica es estimulada eléctricamente para producir la liberación de ACh de las terminaciones presinápticas. Esta disposición experimental permite el registro de las corrientes de placa terminal producidas por la ACh. (B) La amplitud y el curso temporal de las CPT = gACh (Vm − E ) rev corrientes de placa terminal generadas por la estimulación de la neurona motora presináptica mientras se realiza el pinzamiento zonal de voltaje de la célula postsináptica en cuatro potenciales de membrana diferentes. (C) La relación entre la amplitud pico de las corrientes de placa terminal y el potencial de membrana postsináptico es casi lineal, con un potencial de reversión (voltaje en el cual la dirección de la corriente cambia de dentro hacia fuera) próximo a 0 mV. Sobre este gráfico también están indicados (A) Esquema de pinzamiento de voltaje de una fibra muscular postsináptica Estímulo Axón de neurona motora postsináptica Amplificador de pinzamiento de voltaje los potenciales de equilibrio de los iones Na+, K+ y Cl−. (D) La identidad de los iones permeables a los receptores postsinápticos se pone en evidencia por el potencial invertido (Erev). La activación de los canales postsinápticos que sólo son permeables K+ (negro) conduce a corrientes que se revierten en Ek, cerca de −100 mV, mientras que la activación de los canales del Na+ postsinápticos conduce a corrientes que se revierten en ENa, cerca de +70 mV (rojo). Las corrientes selectivas al Cl− se revierten en Ecl, cerca de −50 Electrodo para el pasaje de corriente Fibra muscular postsináptica Electrodo para medir el voltaje Terminaciones presinápticas mV (amarillo). (A-C, de Takeuchi y Takeuchi, 1960.) Corri ente de placa terminal (nA) (B) Efecto del voltaje de membrana sobre las corrientes de placa terminal postsinápticas –110 mV –60 mV Estimular el axón presináptico Estimular el axón presináptico 200 100 0 mV +70 mV Estimular el axón presináptico Estimular el axón presináptico 0 <100 <200 <300 0 2 4 6 0 2 (D) (C) EK ECl Amplitud de la corriente de placa terminal (nA) 4 ENa 6 0 Tiempo (ms) 2 Se abren los canales sólo selectivos para K+ 4 Se abren los canales sólo selectivos al Cl– 300 200 ERev = EK Potencial de reversión 100 ERev = ENa 0 <100 ERev = ECl <200 Se abren los canales sólo selectivos al Na+ <300 <110 <60 0 +70 Potencial de membrana postsináptica (mV) <150 <100 <50 0 Potencial de membrana 50 100 6 0 2 4 6 100 CAPÍTULO 5 Amplitud de la corriente de placa terminal (nA) (A) La concentración menor de Na+ en el exterior desplaza el potencial de reversión hacia la izquierda (B) La concentración mayor de K+ en el exterior desplaza el potencial de reversión hacia la derecha 200 100 0 FIGURA 5.19 El potencial de reversión de la corriente de placa terminal cambia cuando cambian los gradientes iónicos. (A) La reducción de la concentración externa de Na+ hace que las corrientes de placa terminal se reviertan con potenciales más negativos. (B) La elevación de la concentración externa de K+ hace que el potencial de reversión sea más positivo. (De Takeuchi y Takeuchi, 1960). <100 <200 <300 <110 <60 0 +70 <110 <60 Potencial de membrana postsináptica (mV) número negativo. Además, la corriente de placa terminal se hace más pequeña con potenciales que se aproximan a Erev porque la fuerza impulsora se reduce. Con potenciales más positivos que Erev, la corriente de placa terminal es hacia fuera porque la fuerza impulsora tiene una dirección invertida (es decir, positiva). Dado que los canales abiertos para la ACh no son sensibles al voltaje de la membrana, gACh debe depender sólo de la cantidad de canales abiertos por la ACh, lo cual depende a su vez de la concentración de Ach en la hendidura sináptica. Por lo tanto, la magnitud y la polaridad del potencial de membrana postsináptica determinan la dirección y la amplitud de la corriente de placa terminal sólo al alterar la fuerza impulsora sobre los iones que fluyen a través de los canales receptores abiertos por la ACh. Cuando Vm está en el potencial de reversión, Vm − E es igual rev a 0 y no existe ninguna fuerza impulsora neta sobre los iones que pueden atravesar el canal activado por el receptor. En consecuencia, la identidad de los iones que fluyen durante la corriente de placa terminal puede deducirse observando el modo en que el potencial de reversión de la corriente de placa terminal se compara con el potencial de equilibrio para distintas especies de iones (Figura 5.18D). Por ejemplo, si la ACh fuera a abrir un canal sólo permeable al K+, entonces el potencial de reversión de la corriente de placa terminal estaría en el potencial de equilibrio para el K+, el cual para una célula muscular está próximo a −100 mV. Si los canales activados por la ACh fueran permeables sólo al Na+, el potencial de reversión de la corriente sería aproximadamente de +70 mV, el potencial de equilibrio del Na+ de las células musculares; si estos canales fueran permeables sólo al Cl−, entonces el potencial de reversión sería aproximadamente de −50 mV. Por este razonamiento, los canales activados por la ACh no pueden ser permeables sólo a uno de estos iones, porque el potencial de reversión de la corriente de la placa terminal no está cerca del potencial de equilibrio para ninguno de ellos (véase Fig. 5.18C). Sin embargo, si estos canales fueran aproximadamente permeables a Na+ y K+, entonces el potencial de inversión de la corriente de placa terminal estaría entre +70 mV y −100 mV. Por lo tanto, el hecho de que las corrientes de placa terminal se inviertan aproximadamente a 0 mV es compatible con la idea 0 +70 de que los canales iónicos activados por la ACh son permeables tanto al Na+ como al K+. Esta implicación ha sido evaluada en 1960 por el equipo de esposos de Akira y Noriko Takeuchi mediante experimentos en los cuales alteraron la concentración extracelular de estos dos iones. Como se esperaba, la magnitud y el potencial de reversión de la corriente de placa terminal se modificaron al alterar el gradiente de concentración de cada ion. La reducción de la concentración externa de Na+, lo cual vuelve más negativo el Erev, produjo un desplazamiento negativo en el Erev (Figura 5.19A), mientras que la elevación de la concentración externa de K+, que hace más positivo el EK, hace que el Erev se desplace hacia un potencial positivo (Figura 5.19B). Estos experimentos confirman que los canales iónicos activados por ACh son, de hecho, permeables tanto al Na+ como al K+. Aun cuando los canales abiertos para la fijación de ACh a sus receptores sean permeables tanto al Na+ como al K+, la corriente de placa terminal es generada fundamentalmente por el influjo de Na+ (Figura 5.20). Si el potencial de membrana se mantiene en el EK, la corriente de placa terminal surge totalmente de un influjo de Na+ porque en este potencial no existe ninguna fuerza impulsora sobre K+ (Figura 5.20A). En el potencial de membrana de reposo habitual de las fibras musculares de −90 mV, existe una fuerza impulsora pequeña sobre el K+, pero una mucho mayor sobre el Na+. Por lo tanto, durante la corriente de placa terminal fluye mucho más Na+ hacia la célula muscular que K+ hacia afuer (Figura 5.20B) a; este influjo neto de Na+ cargado positivamente constituye la corriente hacia el interior medida como corriente de placa terminal. En el potencial de reversión de 0 mV, el influjo de Na+ y el eflujo de K+ están exactamente equilibrados, de modo que no fluye corriente durante la apertura de los canales por la fijación de ACh (Figura 5.20D). Con potenciales más positivos que el Erev, el equilibrio se invierte; por ejemplo, en el ENa no hay influjo de Na+ y existe un gran eflujo de K+ debido a la fuerza impulsora sobre el K+ (Figura 5.20D). Potenciales aún más positivos producen eflujo de Na+ y generan una corriente de placa terminal hacia el exterior más grande. Si fuera posible medir el PPT al mismo tiempo que la corriente de placa terminal (por supuesto, la técnica de pinzamiento de voltaje impide mantener constante el potencial de membra- TRANSMISIÓN SINÁPTICA Corrientes de placa terminal Flujos iónicos netos (A) Potencial de membrana postsináptica Exterior de la célula Na+ PPT ACh <100 mV (EK) Canal activado por ACh Interior de la célula (B) Na+ <90 mV K+ (C) Na+ 0 mV (Erev) K+ (D) +70 mV (ENa) (F) Amplitud pico de la corriente de placa terminal (nA) Amplitud pico del PPT (mV) K+ (E) Hacia fuera ENa EK Hacia dentro <100 <90 0 +70 Potencial de membrana postsináptica Despolarizante EK 101 ENa Hiperpolarizante <100 <90 0 +70 Potencial de membrana postsináptica FIGURA 5.20. Movimientos de Na+ y de K+ durante las corrientes de placa terminal y los PPT. (A-D) Cada uno de los potenciales postsinápticos (Vpost) indicados a la izquierda produce diferentes flujos relativos de Na+ y de K+ (flujos iónicos). Estos flujos iónicos determinan la amplitud y la polaridad de las corrientes de placa terminal, las cuales a su vez determinan los PPT. Obsérvese que aproximadamente a 0 mV el flujo de Na+ es equilibrado exactamente por un flujo opuesto de K+, lo que lleva a la ausencia de flujo neto de corriente y, por ello, no hay cambios en el potencial de membrana. (E) Las corrientes de placa terminal son corrientes hacia el interior con potenciales más negativos que Erev y corrientes hacia el exterior con potenciales más positivos que Erev. (F) Los PPT despolarizan a la célula postsináptica en potenciales más negativos que Erev. A potenciales más positivos que Erev, los PPT hiperpolarizan la célula. na), se vería que el PPT varía en forma paralela a la amplitud y a la polaridad de la corriente de placa terminal (Figura 5.20E, F). En el potencial de membrana de reposo postsináptico habitual de −90 mV, la gran corriente de la placa terminal hacia el interior hace que el potencial de membrana se torne más despolarizado (véase Fig.5.20F). Sin embargo, en 0 mV, el PPT revierte su polaridad y, con potenciales más positivos, el PPT es hiperpolarizante. Por lo tanto, la polaridad y la magnitud de la corriente de placa terminal dependen de la fuerza impulsora electroquímica que determina la polaridad y la magnitud del PPT. Los PPT se despolarizarán cuando el potencial de membrana sea más negativo que el Erev, y se hiperpolarizarán cuando el potencial de membrana sea más positivo que el Erev. Entonces, la regla general es que la acción de un transmisor impulsa el potencial de membrana postsináptico hacia el Erev para que se activen los canales iónicos particulares. Si bien esta explicación se ha focalizado sobre la unión neuromuscular, mecanismos similares generan respuestas postsinápticas en todas las sinapsis. Un principio general que surge es que la fijación del transmisor a los receptores postsinápticos produce un cambio de la conductancia postsináptica a medida que los canales iónicos se abren (o a veces se cierran). La conductancia postsináptica está aumentada si (como sucede en la unión neuromuscular) los canales están abiertos y está disminuida si los canales están cerrados. En los casos típicos, este cambio de conductancia genera una corriente eléctrica, la corriente postsináptica (CPS), la cual, por su parte, modifica el potencial de membrana postsináptico para producir un potencial postsináptico (PSP). Como en el caso específico del PPT en la unión neuromuscular, los potenciales postsinápticos serán despolarizantes si su potencial de reversión es más positivo que el potencial de membrana postsináptico e hiperpolarizantes si su potencial de reversión es más negativo. Los cambios de la conductancia y los potenciales postsinápticos que característicamente los acompañan son el resultado final de la mayor parte de la transmisión sináptica química, y concluyen una secuencia de acontecimientos eléctricos y químicos Potencial de membrana postsináptica (mV) 102 CAPÍTULO 5 ENa +50 (A) (D) (C) (B) Potencial de acción 0 Erev Umbral <40 <50 Vreposo <60 <70 Erev > Umbral = excitador Activar la sinapsis de GABA PPSE <110 Erev < Umbral = inhibidor PPSI PPSI Erev Activar la sinapsis de glutamato EK Erev 0 1 2 3 Activar la sinapsis de GABA 4 0 1 2 que comienza con la invasión de un potencial de acción en las terminaciones de una neurona postsináptica. De muchas formas, los acontecimientos que producen potenciales postsinápticos en las sinapsis son similares a los que generan potenciales de acción en los axones; en ambos casos, los cambios de la conductancia producidos por los canales iónicos conducen al flujo de corriente iónica que modifica el potencial de membrana. Potenciales presinápticos excitadores e inhibidores Los potenciales postasinápticos alteran finalmente la probabilidad de que un potencial de acción se produzca en la célula postsináptica. En la unión neuromuscular, la acción sináptica sólo aumenta la probabilidad de que un potencial de acción se desarrolle en la célula muscular postsináptica; en efecto, la gran amplitud del PPT asegura que siempre se dispare un potencial de acción. En muchas otras sinapsis, los potenciales postsinápticos aumentan de modo similar la probabilidad de que se dispare un potencial de acción postsináptico. Sin embargo, aun otras sinapsis realmente reducirán la probabilidad de que la célula postsináptica genere un potencial de acción. Los potenciales postsinápticos excitadores (PPSE) aumentan la probabilidad de que se desarrolle un potencial de acción postsináptico. Los potenciales postsinápticos inhibidores (PPSI) disminuyen esta probabilidad. Dado que la mayoría de las neuronas reciben aferencias tanto de sinapsis excitadoras como inhibidoras, es importante conocer con mayor precisión los mecanismos que determinan si una sinapsis particular excita o inhibe a su compañera postsináptica. Los principios de la excitación para la unión neuromuscular que acabamos de describir son pertinentes a todas las sinapsis excitadoras. Los principios de la inhibición postsináptica son muy parecidos a los de la excitación, y también son muy generales. En ambos casos, los neurotransmisores que se fijan a los receptores abren o cierran canales iónicos en la célula postsináptica. El hecho de que una respuesta postsináptica sea un PPSE o un PPSI depende del tipo de canal que está acoplado con el receptor y de la concentración de los iones permeables en el interior y el exterior 3 4 0 Tiempo (ms) 1 2 3 4 FIGURA 5.21 Los potenciales de reversión y los potenciales umbral determinan la excitación y la inhibición postsinápticas. (A) Si el potencial de reversión para un potencial postsináptico (0 mV) es más positivo que el umbral del potencial de acción (−40 mV), el efecto de un transmisor es excitador y genera potenciales postsinápticos excitadores (PPSE). (B) Si el potencial de reversión para un potencial postsináptico es más negativo que el umbral del potencial de acción, el transmisor es inhibidor y genera PPSI. (C) No obstante, los PPSI pueden despolarizar la célula postsináptica si su potencial de reversión se encuentra entre el potencial de reposo y el umbral del potencial de acción. (D) La regla general de la acción postsináptica es: si el potencial de reversión es más positivo que el umbral, se produce excitación; cuando el potencial de reversión es más negativo que el umbral, se desarrolla inhibición. de la célula. De hecho, la única distinción entre la excitación y la inhibición postsináptica es el potencial de reversión del potencial postsináptico respecto del voltaje umbral para generar potenciales de acción en la célula postsináptica. Consideremos, por ejemplo, una sinapsis neuronal que utiliza glutamato como transmisor. Muchas de estas sinapsis tienen receptores que, al igual que los receptores para ACh en las uniones neuromusculares, abren canales iónicos que no son selectivamente permeables a los cationes (véase Cap. 6). Cuando estos receptores para el glutamato son activados, fluye tanto Na+ como K+ a través de la membrana postsináptica, lo que arroja un Erev de alrededor de 0 mV para la corriente postsináptica resultante. Si el potencial de reposo de la neurona presináptica es de −60 mV, el PPSE resultante despolarizará el potencial de membrana postsináptico y llevará hasta 0 mV. En la neurona hipotética que se muestra en la Figura 5.21A, el voltaje umbral del potencial de acción es de −40 mV. Por lo tanto, el PPSE aumenta la probabilidad de que esta neurona produzca un potencial de acción y define a la sinapsis como excitadora. Como ejemplo de acción postsináptica inhibidora, consideremos una sinapsis neuronal que utiliza GABA como transmisor. En estas sinapsis, los receptores para el GABA abren canales que son selectivamente permeables al Cl− y la acción del GABA hace que Cl− fluya a través de la membrana postsináptica. Consideremos un caso en el que el ECl es de −70 mV, como es típico para TRANSMISIÓN SINÁPTICA 103 (A) FIGURA 5.22 Suma de los potenciales postsinápticos. Excitadora E1 (A) Un microelectrodo registra los potenciales postsinápticos producidos por la actividad de dos sinapsis excitadoras (E1 y E2) y Registro Potencial de membr ana postsináptica Inhibidora I una sinapsis inhibidora (I). (B) Respuestas eléctricas a la activación sináptica. La estimulación de cualquiera de las sinapsis excitadoras (E1 o E2) produce un PPSE subumbral, mientras que la estimulación de ambas sinapsis al mismo tiempo (E1 + E2) produce un PPSE supraliminal que provoca un potencial de acción postsináptico (que Excitadora E2 se muestra en azul). La activación de la sinapsis inhibidora aislada (I) produce un PPSI hiperpolarizante. La suma de este PPSI (línea roja rayada) con el PPSE (línea amarilla rayada) producidos por una sinapsis excitadora (E1 + I) reduce la amplitud del PPSE (línea naranja), mientras que la suma con el PPSE supraliminal producido Cuerpo celular Axón Dendritas por la activación de las sinapsis E1 y E2 mantiene a la neurona postsináptica por debajo del umbral, de modo que no se produce ningún potencial de acción. (B) PPSE (sinapsis E1 o E2) Potencial de membrana postsináptica (mV) +20 PPSE sumados (Sinapsis E1 + E2) PPSI (Sinapsis I) PPSE + PPSI sumados (Sinapsis E1 + I) PPSE + PPSI sumados (Sinapsis E1 + I + E2) 0 <20 <40 <60 Umbral Vrest E1 or E2 E1 + E2 I E1 + I E1 + E2 + I Tiempo (ms) la mayoría de las neuronas, de modo que el potencial de reposo postsináptico de −60 mV es menos negativo que el ECl. La fuerza impulsora electroquímica positiva resultante (Vm − Erev) hace que el Cl− con carga negativa fluya hacia la célula, lo que genera una PPSI hiperpolarizante (Figura 5.21B). Este PPSI hiperpolarizante aleja a la membrana postsináptica del umbral para el potencial de acción de −40 mV y claramente inhibe a la célula postsináptica. Sorprendentemente, no todas las sinapsis inhibidoras producen PPSI hiperpolarizantes. Por ejemplo, si el ECl fuera de −50 mV en lugar de −70 mV, entonces la fuerza impulsora electroquímica negativa haría que el Cl− fluyera hacia fuera de la célula y produciría un PPSI despolarizante (Figura 5.21C). Sin embargo, la sinapsis sería aún inhibidora: dado que el potencial de reversión del PPSI sigue siendo más negativo que el umbral para el potencial de acción (−40 mV), el PPSI despolarizante es aún más negativo que el umbral para la iniciación del potencial de acción. Otra forma de pensar en esta situación peculiar es que si otra aferencia excitadora sobre esta neurona llevara el potencial de membrana hasta −41 mV, inmediatamente por debajo del umbral para disparar un potencial de acción, entonces el PPSI hiperpolarizaría el potencial de membrana hasta −50 mV y alejaría el potencial del umbral del potencial de acción. Por lo tanto, mientras los PPSE siempre despolarizan a la célula postsináptica, los PPSI pueden hiperpolarizarla o despolarizarla; en realidad, un cambio inhibidor en la conductancia puede no producir ningún cambio de potencial en absoluto y ejerce aún un efecto inhibidor al hacer que sea más difícil para un PPSE provocar un potencial de acción en la célula postsináptica. Si bien las particularidades de la acción postsináptica pueden ser complejas, una regla simple distingue la excitación postsináptica de la inhibición: un PPSE tiene un potencial de reversión más positivo que el umbral del potencial de acción, mientras que un PPSI tiene un potencial de reversión más negativo que el umbral (Figura 5.21D). Intuitivamente, esta regla puede comprenderse reconociendo que un PPSE tiende a despolarizar el potencial de membrana de modo que excede el umbral, mientras que un PPSI siempre actúa manteniendo el potencial de membrana más negativo que el potencial umbral. Sumación de potenciales de acción Los potenciales postsinápticos en la mayoría de las sinapsis del encéfalo son mucho más pequeños que los de la unión neuromuscular; en efecto, los PPSE producidos por sinapsis excita- 104 CAPÍTULO 5 RECUADRO 5C La “sinapsis tripartita” Hasta este punto, nuestra explicación ha considerado la señalización entre las neuronas presinápticas y sus dianas postsinápticos como un diálogo privado entre estas dos células. Sin embargo, algunas investigaciones recientes sugieren que esta conversación sináptica puede involucrar también a células gliales. Como mencionamos en el Capítulo 1, las células gliales sostienen de algunas formas a las neuronas. Por ejemplo, está bien establecido que la glía regula el entorno extracelular al eliminar el K+ que se acumula durante la generación del potencial de acción y al eliminar los neurotransmisores al concluir la transmisión sináptica. De forma compatible con estos papeles, la glía parece ocupar casi todo el volumen no neuronal del encéfalo. Como resultado de esta disposición, la glía se encuentra (A) Imagen de microscopia electrónica de una sinapsis entre una neurona presináptica terminal (pre) y una neurona postsináptica (post) con una célula glial (astrocito, astro) inmediatamente adyacente a la sinapsis. (B) Estructura tridimensional de contacto entre una célula glial (azul) rodeada de dendritas de cuatro neuronas postsinápticas (diferentes colores). (De Witcher y cols., 2007). (A) astro en una asociación muy estrecha con las sinapsis (Fig. A); una sinapsis dada típicamente no está a más de una fracción de micrómetro de una célula glial. Las células gliales forman prolongaciones extremadamente finas que envuelven las sinapsis (Fig. B), asociación íntima que plantea la posibilidad de un papel de señalización para la glía en las sinapsis. El primer apoyo para este papel provino del descubrimiento de que las células gliales responden a la aplicación de neurotransmisores. La lista de los neurotransmisores que producen respuestas en la glía incluye ahora acetilcolina, glutamato, GABA y muchos otros. Estas respuestas están mediadas por los mismos tipos de receptores de neurotransmisores que se emplean en la señalización sináptica, principalmente los receptores metabotrópicos que se acoplan a las cascadas de señalización intracelular (véase Fig. 5.16B). En algunos casos, los neurotransmisores producen cambios en el potencial de membrana de las células gliales. Con mayor frecuencia, estos neurotransmisores producen cambios transitorios en la concentración intracelular de calcio dentro de la célula glial. A menudo se observa que estas señales intracelulares de calcio desencadenan ondas de calcio que se propagan tanto dentro de una única célula glial, como también entre ellas (Fig. C). Estas elevaciones transitorias del calcio intracelular sirven como señales de segundo mensajero (véase Cap. 7) que desencadenan algunas respuestas fisiológicas en la glía. La respuesta más notable es la liberación de varias moléculas, como glutamato, GABA y ATP, consideradas tradicionalmente como neurotransmisores. La liberación de estos “gliotransmisores” ocurre tanto a través de mecanismos de exocitosis desencadenados por el calcio empleado en las terminaciones presinápticas de las neuronas (véase Fig. 5.3) como por mecanismos no convencionales de liberación como la permeabilidad a través de ciertos canales iónicos. La capacidad tanto de responder como de liberar neurotransmisores convierte a las células gliales en participantes potenciales en la señalización sináptica. En efecto, se ha observado que la liberación de neurotransmisores a partir de distintas terminaciones presinápticas produce respuestas en las células gliales. Además, se ha observado que la liberación de “gliotransmisores” regula la transmisión en numerosas sinapsis (Fig. D). En algunos casos, la glía regula la transmisión de transmisores a partir de las terminaciones presinápticas, mientras que en otros casos alteran la capacidad de respuesta postsináptica. Esta capacidad de la glía para participar (C) pre 6s 12 s 18 s 24 s post 200 +m (D) Respuesta sináptica (porcentaje del control) (B) (C) La aplicación de glutamato (en la flecha) aumenta el Ca2+ intracelular (blanco) en células gliales cultivadas. El examen de estas células en diferentes momentos indica que 150 las señales de calcio se propagan como una onda a través de la glía vecina. (De 100 Raise Elevación Ca2+del in glial Ca2+cell en la célula glial 50 -60 0 60 Tiempo (s) 120 Cornell-Bell y cols., 1990). (D) La elevación transitoria de Ca2+ en el interior de una única célula glial (flecha) aumenta la transmisión en una sinapsis excitadora en el hipocampo. (De Perea y Araque, 2007.) TRANSMISIÓN SINÁPTICA 105 RECUADRO 5C (continúa) en la señalización sináptica ha conducido al concepto de la sinapsis tripartita, una unión de tres vías que involucra la terminación presináptica, la prolongación postsináptica y las células gliales vecinas. Aunque se discute aún la importancia fisiológica de estas interacciones neuronas-glía, la capacidad recién descubierta de la glía para liberar neurotransmisores, similar a las terminaciones presinápticas, y de responder a ellos, similar a las neuronas postsinápticas, promete modificar espectacularmente nuestro punto de vista sobre los mecanismos de señalización encefálica. Bibliografía Cornell-Bell, A. H., S. M. Finkbeiner, M. S. Cooper and S. J. Smith (1990) Glutamate induces calcium waves in cultured astrocytes: Long-range glial signaling. Science 247: 470-473. Fiacco, T. A., C. Agulhon and K. D. McCarthy (2009) Sorting out astrocyte physiology from pharmacology. Annu. Rev. Pharmacol. Toxicol. 49: 151-174. Halassa, M. M. and P. G. Haydon (2010) Integrated brain circuits: astrocytic networks modulate neuronal activity and behavior. Annu. Rev. Physiol. 72: 335-355. Hamilton, N. B. axd D. Attwell (2010) Do astrocytes really exocytose neurotransmitters? Nat. Rev. Neurosci. 11: 227-238. Haydon, P. G. and G. Carmignoto (2006) Astrocyte torias individuales pueden representar sólo una fracción de un milivoltio y suelen estar muy por debajo del umbral para generar potenciales de acción postsinápticos. ¿De qué modo entonces pueden estas sinapsis transmitir información si sus potenciales postsinápticos están por debajo del umbral? La respuesta es que, típicamente, las neuronas del sistema nervioso central se encuentran inervadas por miles de sinapsis, y los potenciales postsinápticos producidos por cada sinapsis activa pueden sumarse –en espacio y en tiempo– para determinar el comportamiento de la neurona postsináptica. Consideremos el caso sumamente simplificado de una neurona que es inervada por dos sinapsis excitadoras, y cada una de ellas genera un PPSE subumbral, y una sinapsis inhibitoria que produce un PPSI (Figura 5.22A). Si bien la activación de cualquiera de estas dos sinapsis excitadora sola (E1 o E2 en la Figura 5.22B) produce un PPSE subumbral, la activación de ambas sinapsis excitadoras al mismo tiempo hace que se sumen dos PPSE. Si la suma de los dos PPSE (E1 + E2) despolariza la neurona postsináptica lo suficiente como para alcanzar el potencial umbral, aparece un potencial de acción postsináptico. Por lo tanto, la sumación permite que los PPSE subumbrales influyan en la producción de potenciales de acción. Asimismo, FIGURA 5.23 Acontecimientos desde la liberación del neurotransmisor hasta la excitación o la inhibición postsináptica. La liberación del neurotransmisor en todas las terminaciones presinápticas sobre una célula conduce a la fijación al receptor, la cual produce la apertura o el cierre de canales iónicos específicos. El cambio de conductancia resultante hace que fluya la corriente, lo cual puede modificar el potencial de membrana. La célula postsináptica suma (o integra) todos los PPSE y los PPSI, lo que conduce a un control momento a momento de la generación del potencial de acción. control of synaptic transmission and neurovascular coupling. Physiol. Rev. 86: 1009-1031. Jahromi, B. S., R. Robitaille and M. P. Charlton (1992) Transmitter release increases intracellular calcium in peri-synaptic Schwann cells in situ. Neuron 8: 1069-1077. Lee, S. and 7 others (2010) Channel-mediated tonic GABA release from glia. Science 330: 790-796. Perea, G. .and A. Araque (2007) Astrocytes potentiate transmitter release at single hippocampal synapses. Science 317: 1083-1086. Perea, G., M. Navarrete and A. Araque (2009) Tripartite synapses: astrocytes process and control synaptic information. Trends Neurosci. 32: 421-431. Witcher, M. R., S. A. Kirov axd K. M. Harris (2007) Plasticity of perisynaptic astroglia during syn aptogenesis in the mature rat hippocampus. Glia 55: 13-23. Liberación del neurotransmisor Unión al receptor Los canales iónicos se abren o se cierran El cambio de conductancia produce flujo de corriente El potencial postsináptico cambia Células postsinápticas excitadas o inhibidas La sumación determina si ocurre o no un potencial de acción 106 CAPÍTULO 5 un PPSI generado por una sinapsis inhibitoria (I) puede sumarse (hablando en términos algebraicos) con un PPSE subumbral para reducir su amplitud (E1 + I) o puede sumarse con un PPSE supraumbral para evitar que la neurona postsináptica alcance el umbral (E1 + I + E2). En resumen, la sumación de PPSE y PPSI por una neurona postsináptica permite que una neurona integre la información eléctrica provista por todas las sinapsis inhibitorias y excitadoras que actúan sobre ella en cualquier momento. El hecho de que la suma de las aferencias sinápticas activas resulte en la producción de un potencial de acción depende del equilibrio entre excitación e inhibición. Si la suma de todos los PPSE y PPSI conduce a una despolarización de suficiente amplitud como para elevar el potencial de membrana por encima del umbral, entonces la célula postsináptica producirá un potencial de acción. Por el contrario, si prevalece la inhibición, la célula postsináptica se mantendrá silente. En condiciones normales, el equilibrio entre PPSE y PPSI cambia continuamente con el tiempo, dependiendo de la cantidad de sinapsis excitadoras e inhibidoras activas en un momento dado y de la magnitud de la corriente en cada sinapsis activa. Por lo tanto la sumación es un tira y afloje inducido por el neurotransmisor entre todas las corrientes excitadoras e inhibidoras; el resultado de la batalla determina si una neurona postsináptica dispara o no un potencial de acción y, por lo tanto, si se convierte en un elemento activo en los circuitos nerviosos a los que pertenece (Figura 5.23). La investigación reciente sugiere que las células gliales también pueden contribuir a la señalización sináptica (Recuadro 5C). Resumen Las sinapsis comunican la información transmitida por los potenciales de acción de una neurona a la siguiente en los circuitos nerviosos. Los mecanismos celulares que subyacen a la transmisión sináptica están íntimamente relacionados con aquellos que generan otros tipos de señales eléctricas neuronales, es decir, el flujo de iones a través de los canales de la membrana. En el caso de las sinapsis eléctricas, estos canales son uniones en hendidura; el flujo directo pero pasivo de corriente a través de las uniones en hendidura es la base para la transmisión. En el caso de las sinapsis químicas, los canales con poros más pequeños y más selectivos son activados por la unión de los neurotransmisores a los receptores postsinápticos después de la liberación de los neurotransmisores desde la terminación presináptica. La gran cantidad de neurotransmisores en el sistema nervioso puede dividirse en dos clases amplias: transmisores de molécula pequeña y neuropéptidos. Los neurotransmisores son sintetizados a partir de precursores definidos por vías enzimáticas reguladas, empaquetados en uno de varios tipos de vesículas sinápticas y liberados en la hendidura sináptica de una forma Ca2+-dependiente. Muchas sinapsis liberan más de un tipo de neurotransmisor e incluso se pueden empaquetar múltiples tipos de neurotransmisores dentro de la misma vesícula sináptica. Los agentes neurotransmisores son liberados en la terminación presináptica en unida- des o cuantos, que reflejan su almacenamiento en el interior de las vesículas sinápticas. Las vesículas descargan su contenido en la hendidura sináptica donde la despolarización presináptica generada por la invasión de un potencial de acción abre los canales del calcio con puerta de voltaje, lo que permite que el Ca2+ ingrese en la terminación presináptica. No se ha establecido aún cómo el calcio desencadena la liberación del neurotransmisor, pero están claramente involucradas sinaptotagmina, SNARE y algunas otras proteínas que se encuentran en el interior de la terminación presináptica. Los receptores postsinápticos constituyen un grupo diverso de proteínas que traducen la fijación de neurotransmisores en señales eléctricas al abrir o cerrar los canales iónicos postsinápticos. Se han desarrollado dos familias ampliamente diferentes de receptores de neurotransmisores para llevar a cabo las acciones de señalización postsináptica de los neurotransmisores. Las corrientes postsinápticas producidas por la apertura o el cierre sincrónico de los canales iónicos cambian la conductancia de la célula postsináptica, y así aumentan o disminuyen su excitabilidad. Los cambios en la conductancia que aumentan la probabilidad de generar un potencial de acción son excitadores, mientras que aquellos que disminuyen esa probabilidad son inhibidores. Como las neuronas postsinápticas suelen estar inervadas por muchas aferencias diferentes, el efecto integrado de los cambios en la conductancia subyacentes a todos los PPSE y los PPSI producidos en una célula postsináptica en cualquier momento determina si la célula dispara o no un potencial de acción. Los efectos postsinápticos de los neurotransmisores concluyen con la degradación del transmisor en la hendidura sináptica, el transporte del transmisor nuevamente hacia las células o la difusión fuera de la hendidura sináptica. La respuesta producida en una sinapsis dada depende del tipo de neurotransmisor liberado y del complemento postsináptico de receptores y canales asociados. Lecturas adicionales Revisiones Augustine, G. J. and H. Kasai (2006) Bernard Katz, quantal transmitter release, and the foundations of presynaptic physiology. J. Physiol. (Lond.) 578: 623-625. Brodsky, F. M., C. Y. Chen, C. Knuehl, M. C. Towler and D. E. Wakeham (2001) Biological basket weaving: Formation and function of clathrin-coated vesicles. Annu. Rev. Cell. Dev. Biol. 17: 517-568. Brunger, A. T. (2005) Structure and function of SNARE and SNAREinteracting proteins. Quart. Rev. Biophys. 38: 1-47. Carlsson, A. (1987) Perspectives on the discovery of central monoaminergic neurotransmission. Annu. Rev. Neurosci. 10: 19-40. Chapman, E. R. (2002) Synaptotagmin: A Ca2+ sensor that triggers exocytosis? Nat. Rev. Mol. Cell Biol. 3: 498-508. Connors, B. W. and M. A. Long (2004) Electrical synapses in the mammalian brain. Annu. Rev. Neurosci. 27: 393-418. Emson, P. C. (1979) Peptides as neurotransmitter candidates in the CNS. Prog. Neurobiol. 13: 61-116. Hestrin, S. and M. Galarreta (2005) Electrical synapses define networks of neocortical GABAergic neurons. Trends Neurosci. 28: 304-309. Jackson, M. B. and E. R. Chapman (2006) Fusion pores and fusion machines in Ca2+ -triggered exocytosis. Annu. Rev. Biophys. Biomol. Struct. 35: 135160. TRANSMISIÓN SINÁPTICA Marsh, M. and H. T. McMahon (1999) The structural era of endocytosis. Science 285: 215-220. Pierce, K. L., R. T. Premont and R. J. Lefkowitz (2002) Seven-membrane receptors. Nature Rev. Mol. Cell Biol. 3: 639-650. Rizzoli, S. O. and W. J. Betz (2005) Synaptic vesicle pools. Nature Rev. Neurosci. 6: 57-69. Royle, S. J. and L. Lagnado (2010) Clathrin-mediated endocytosis at the synaptic terminal: Bridging the gap between physiology and molecules. Traffic 11: 1489-1497. Südhof, T. and J. E. Rothman (2009) Membrane fusion: Grappling with SNARE and SM proteins. Science 323: 474-477. Artículos originales importantes Adler, E., M. Adler, G. J. Augustine, M. P. Charlton and S. N. Duffy (1991) Alien intracellular calcium chelators attenuate neurotransmitter release at the squid giant synapse. J. Neurosci. 11: 1496-1507. Augustine, G. J. and R. Eckert (1984) Divalent cations differen-tiallv support transmitter release at the squid giant synapse. J. Physiol. (Lond.) 346: 257-271. Beierlein, M., J. R. Gibson and B. W. Connors (2000) A network of electrically coupled interneurons drives synchronized inhibition in neocortex. Nature Neurosci. 3: 904-910. Boyd, I. A. and A. R. Martin (1955) The end-plate potential in mammalian muscle. J. Physiol. (Lond.) 132: 74-91. Curtis, D. R. J. W. Phillis and J. C. Watkins (1959) Chemical excitation of spinal neurons. Nature 183: 611-612. Dale, H. H., W. Feldberg and M. Vogt (1936) Release of acetylcholine at voluntary motor nerve endings. J. Physiol. 86: 353-380. del Castillo, J. and B. Katz (1954) Quantal components of the end plate potential. J. Physiol. (Lond.) 124: 560-573. Fatt, P. and B. Katz (1951) An analysis of the end plate potential recorded with an intracellular electrode. J. Physiol. (Lond.) 115: 320-370. Fatt, P. and B. Katz (1952) Spontaneous subthreshold activity at motor nerve endings. J. Physiol. (Lond.) 117: 109-128. Furshpan, E. J. and D. D. Potter (1959) Transmission at the giant motor synapses of the crayfish. J. Physiol. (Lond.) 145: 289-325. Geppert, M. and 6 others (1994) Synaptotagmin I: A major Ca2+ sensor for transmitter release at a central synapse. Cell 79: 717-727. Harris, B. A., J. D. Robishaw, S. M. Mumby and A. G. Gilman (1985) Molecular cloning of complementary DNA for the alpha subunit of the G protein that stimulates adenylate cyclase. Science 229: 1274-1277. Heuser, J. E., T. S. Reese, M. J. Dennis, Y. Jan, L. Jan and L. Evans (1979) Synaptic vesicle exocytosis captured by quick freezing and correlated with quantal transmitter release. J. Cell Biol. 81: 275-300. 107 Heuser, J. E. and T. S. Reese (1973) Evidence for recycling of synaptic vesicle membrane during transmitter release at the frog neuromuscular junction. J. Cell Biol. 57: 315-344. Hökfelt, T., O. Johansson, A. Ljungdahl, J. M. Lundberg and M. Schultzberg (1980) Peptidergic neurons. Nature 284: 515-521. Jonas, P., J. Bischofberger and J. Sandkuhler (1998) Co-release of two fast neurotransmitters at a central synapse. Science 281: 419-424. Loewi, O. (1921) Tiber humorale Ubertragbarkeit der Herznerven-wirkung. Pfliigers Arch. 189: 239-242. Miledi, R. (1973) Transmitter release induced by injection of calcium ions into nerve terminals. Proc. R. Soc. Lond. B 183: 421-425. Neher, E. and B. Sakmann (1976) Single-channel currents recorded from membrane of denervated frog muscle fibres. Nature 260: 799-802. Smith, S. J., J. Buchanan, L. R. Osses, M. P. Charlton and G. J. Augustine (1993) The spatial distribution of calcium signals in squid presynaptic terminals. J. Physiol. (Lond.) 472: 573-593. Sutton, R. B., D. Fasshauer, R. Jahn and A. T. Brünger (1998) Crystal structure of a SNARE complex involved in synaptic exo-cytosis at 2.4 A resolution. Nature 395: 347-353. Takamori, S. and 21 others (2006) Molecular anatomy of a trafficking organelle. Cell 127: 831-846. Takeuchi, A. and N. Takeuchi (1960) On the permeability of erid-plate membrane during the action of transmitter. J. Physiol. (Lond.) 154: 52-67. Wickman, K. and 7 others (1994) Recombinant Gβγ activates the muscarinicgated atrial potassium channel 1KACh. Nature 368: 255-257. Libros Cooper, J. R., F. E. Bloom and R. H. Roth (2002) The Biochemical Basis of Neuropharmacology. New York: Oxford University Press. Hall, Z. (1992) An Introduction to Molecular Neurobiology. Sunderland, MA: Sinauer Associates. Katz, B. (1969) The Release of Neural Transmitter Substances. Liverpool: Liverpool University Press. Llinás, R. R. (1999) The Squid Giant Synapse: A Model for Chemical Synaptic Transmission. Oxford: Oxford University Press. Nicholls, D. G. (1994) Proteins, Transmitters, and Synapses. Oxford: Blackwell. Peters, A., S. L. Palay and H. deF. Webster (1991) The Fine Structure of the Nervous System: Neurons and their Supporting Cells, 3rd Ed. Oxford: Oxford University Press. 6 NeurotraNsmisores y sus receptores Aspectos generales LAS NEURONAS DEL ENCÉFALO HUMANO SE COMUNICAN entre sí, en su mayor parte, liberando mensajeros químicos denominados “neurotransmisores”. Se conoce actualmente una gran cantidad de neurotransmisores y aún quedan más por descubrir. El principal neurotransmisor excitador del encéfalo es el aminoácido glutamato, mientras que el principal neurotransmisor inhibidor es el ácido γ-aminobutírico (GABA). Los neurotransmisores provocan respuestas eléctricas postsinápticas al unirse a los receptores de los neurotransmisores y activarlos. La mayoría de los neurotransmisores son capaces de activar varios receptores diferentes y de generar muchos modos posibles de señalizaciones sinápticas. Después de activar sus receptores postsinápticos, los neurotransmisores son eliminados de la hendidura sináptica por los transportadores de los neurotransmisores o por las enzimas degradadoras. Las anomalías en la función de los sistemas de neurotransmisores contribuyen a una amplia gama de trastornos neurológicos y psiquiátricos; por lo tanto, muchas terapias neurofarmacológicas se basan en fármacos que afectan a los neurotransmisores, a sus receptores o a los transportadores responsables de eliminarlos de la hendidura sináptica. Categorías de neurotransmisores Se conocen más de 100 neurotransmisores diferentes. Esta gran cantidad de transmisores permite una gran diversidad en la señalización química entre dos neuronas. Es útil separar esta variedad de transmisores en dos categorías amplias basadas simplemente en el tamaño (Fig. 6.1). Los neuropéptidos son moléculas transmisoras relativamente grandes compuestas por 3 a 36 aminoácidos. Los aminoácidos individuales, como el glutamato y el GABA, así como las transmisoras acetilcolina, serotonina e histamina, son mucho más pequeños que los neuropéptidos y, por lo tanto, se los denomina neurotransmisores de molécula pequeña. Dentro de la categoría de los neurotransmisores de molécula pequeña, las aminas biógenas (dopamina, noradrenalina, adrenalina, serotonina e histamina) se explican a menudo por separado debido a que sus propiedades químicas y sus acciones postsinápticas son similares. Las particularidades de síntesis, empaquetamiento, liberación y eliminación difieren para cada neurotransmisor (Cuadro 6.1). En este capítulo, se describen algunas de las características principales de estos transmisores y sus receptores postsinápticos. 110 CAPÍTULO 6 NEUROTRANSMISORES DE MOLÉCULA PEQUEÑA O + Acetilcolina (CH3)3N AMINOÁCIDOS H3N CH2 O CH3 C CATECOLAMINAS CH2 Dopamina H C + Glutamato CH2 AMINAS BIÓGENAS + CH2 NH3 CH2 NH3 CH2 NH2 COO< HO CH2 OH CH2 OH COOH Aspartato Noradrenalina H C + H3N CH2 + COO< HO CH2 OH COOH OH + GABA H3N CH2 CH2 Adrenalina COO< CH2 CH2 + CH3 HO OH H C + Glicina H3N COO< INDOLAMINA H HO Serotonina (5-HT) PURINAS P O< CH2 NH3 N O O CH2 N NH2 ATP < CH2 O O P O< N O O P IMIDAZOLAMINA O CH2 O N N CH2 Histamina HN O< H + N H OH OH NEUROTRANSMISORES PEPTÍDICOS (más de 100 péptidos, habitualmente de 3-30 aminoácidos de longitud) Ejemplo: metionina encefalina (Tyr–Gly–Gly–Phe–Met) O O + H3N H C CH2 C H N H C H C O H N H C H C O H N H C CH2 C O H N H C C CH2 CH2 S CH3 OH Tyr Gly Gly Phe Met O< + NH3 NEUROTRANSMISORES Y SUS RECEPTORES ▼ FIGURA 6.1 Ejemplos de neurotransmisores de molécula pequeña y de neurotransmisores peptídicos. Los neurotransmisores de molécula pequeña pueden subdividirse en acetilcolina, aminoácidos, purinas y aminas biógenas. Las catecolaminas, denominadas así porque todas comparten la porción catecol (es decir, un anillo benceno hidroxilado), forman un subgrupo distinto dentro de las aminas biógenas. La serotonina y la histamina contienen un anillo indol y un anillo imidazol, respectivamente. Las diferencias de tamaño entre los neurotransmisores de molécula pequeña y los neurotransmisores peptídicos están indicadas por los modelos de esferas interpuestas para glicina, noradrenalina y metionina encefalina. (Los átomos de carbono están en negro; los átomos de hidrógeno, en blanco; los átomos de nitrógeno, en azul; y los átomos de oxígeno, en rojo). Acetilcolina Como mencionamos en el capítulo anterior, la acetilcolina (ACh) fue la primera sustancia identificada como neurotransmisora. Además de la función de la ACh como neurotransmisor en las uniones neuromusculares esqueléticas (véase el Cap. 5), así como en la sinapsis neuromuscular entre el nervio vago y las fibras del músculo cardíaco, la ACh sirve como transmisor en las sinapsis en los ganglios del sistema motor visceral y en distintos lugares dentro del sistema nervioso central. Aunque se sabe mucho sobre la función de la transmisión colinérgica en las uniones neuromusculares y en las sinapsis ganglionares, no se conocen bien las acciones de la ACh en el sistema nervioso central. La acetilcolina es sintetizada en las terminaciones nerviosas de los precursores de la acetilcoenzima A (acetilCoA, que es sintetizada a partir de glucosa) y colina, en una reacción catalizada por la 111 colinaacetiltransferasa (CAT; Figura 6.2). La colina está presente en el plasma en alta concentración (alrededor de 10 mM) y es captada en las neuronas colinérgicas por un cotransportador de colina Na+dependiente de alta afinidad. Después de la síntesis en el citoplasma de la neurona, un transportador vesicular de la ACh carga aproximadamente 10 000 moléculas de ACh en cada vesícula colinérgica. La energía necesaria para concentrar ACh dentro de la vesícula es provista por el pH ácido de la luz vesicular, que permite al transportador vesicular de ACh intercambiar H+ porACh. Al contrario de la mayoría de los neurotransmisores de molécula pequeña, las acciones postsinápticas de la ACh en muchas sinapsis colinérgicas (la unión neuromuscular en particular) no son terminadas por recaptación, sino por una potente enzima hidrolítica, la acetilcolinesterasa (AChE). Esta enzima se encuentra muy concentrada en la hendidura sináptica, lo que asegura una rápida disminución de la concentración de ACh después de su liberación de la terminación presináptica. La AChE tiene una actividad catalítica muy alta (aproximadamente 5 000 moléculas de ACh por molécula de AChE por segundo) e hidroliza la ACh en acetato y colina. La colina producida por la hidrólisis de la ACh es reciclada al ser transportada nuevamente hacia las terminaciones nerviosas, donde es utilizada para sintetizar nuevamente ACh. Entre los muchos fármacos interesantes que interactúan con enzimas colinérgicas están los organofosforados. Este grupo incluye algunos potentes agentes de la guerra química. Uno de estos compuestos es el gas nervioso “sarín”, que adquirió notoriedad en 1995 después de que un grupo de terroristas liberó este gas en el sistema de subterráneos de Tokio. Los organofosfora- CUADRO 6.1. Características funcionales de los principales neurotransmisores EFECTO NEUROTRANSMISOR POSTINÁPTICOa PRECURSOR(ES) PASO LIMITANTE DE LA VELOCIDAD EN LA SÍNTESIS MECANISMO DE ELIMINACIÓN TIPO DE VESÍCULA Pequeña, clara Pequeña, clara Pequeña, clara Pequeña, clara Pequeña, centro denso o centro denso irregular grande Grande, centro denso Grande, centro denso Pequeña, clara ACh Glutamato GABA Glicina Catecolaminas (adrenalina, noradrenalina, dopamina) Serotonina (5-HT) Excitador Excitador Inhibidor Inhibidor Excitador Colina + acetilcoA Glutamina Glutamato Serina Tirosina CAT Glutaminasa GAD Fosfoserina Tirosina hidroxilasa Acetilcolinesterasa Transportadores Transportadores Transportadores Transportadores, MAO, COMT Excitador Triptófano Triptófano hidroxilasa Histamina Excitador Histidina Histidina descarboxilasa Transportadores, MAO Transportadores ATP Excitador ADP Neuropéptidos Excitador e inhibidor Inhibe la inhibición Excitador e inhibidor Aminoácidos (síntesis de proteínas) Lípidos de membrana Fosforilación oxidativa mitocondrial; glucólisis Síntesis y transporte Hidrólisis a AMP y adenosina Proteasas Modificación enzimática de lípidos Óxido nítrico sintasa Hidrólisis por FAAH Grande, centro denso Ninguna Oxidación espontánea Ninguna Endocannabinoides Óxido nítrico Arginina Se indica el efecto postsináptico más frecuente; el mismo transmisor puede producir excitación o inhibición postsináptica según los canales iónicos afectados por la fijación del transmisor (véase el Cap. 5). a 112 CAPÍTULO 6 FIGURA 6.2 Metabolismo de la acetilcolina en las terminaciones nerviosas colinérgicas. La síntesis de la acetilcolina a partir de la colina y de la acetilCoA necesita de la colina acetiltransferasa. La acetilCoA deriva del piruvato generado por glucólisis, mientras que la colina es transportada en las terminaciones mediante un cotranspor- Glucosa Piruvato tador Na+-dependiente (ChT). La acetilcolina es almacenada en vesículas sinápticas mediante un transportador vesicular (VAChT). Después Terminación presináptica de la liberación, la acetilcolina es metabolizada ChT Acetil CoA Na+ Colina Colina O CH3 + CoA S C HO CH2 + N CH2 rápidamente por la acetilcolinesterasa y la colina es transportada nuevamente hasta la terminación mediante el cotransportador Na+-dependiente. (CH3)3 Colina acetiltransferasa Acetilcolina O CH3 C O CH2 CH2 + N (CH3)3 VAChT Acetato Acetilcolina Acetilcolinesterasa CH3 < C OO Colina + HO CH2 CH2 + N (CH3)3 Célula postsináptica Receptores de la acetilcolina dos pueden ser letales porque inhiben la AChE y hacen que se acumule acetilcolina en las sinapsis colinérgicas. Este aumento de la acetilcolina despolariza la célula postsináptica y la hace refractaria a la liberación posterior de ACh, y así provoca parálisis neuromuscular y otros efectos. La alta sensibilidad de los insectos a los inhibidores de la AChE convirtió a los organofosforados en insecticidas populares. Muchas de las acciones postsinápticas de la acetilcolina están mediadas por el receptor colinérgico nicotínico (nAChR), denominado así porque la nicotina, un estimulante del SNC, también se une a estos receptores. El consumo de nicotina produce cierto grado de euforia, relajación y finalmente adicción, efectos que se cree están mediados por los nAChR. Como se describió en el Capítulo 5, los nAChR son canales catiónicos no selectivos que generan respuestas postsinápticas excitadoras. Algunas toxinas se unen específicamente a los receptores nicotínicos y los bloquean (Recuadro 6A). La disponibilidad de estos ligandos altamente específicos –en particular, un componente del veneno de víbora llamado “α-bungarotoxina”– ha proporcionado una forma útil para aislar y purificar el nAChR. En consecuencia, los nAChR representan el tipo ionotrópico de receptores de neurotransmisores mejor estudiado, y el descubrimiento de su organización molecular proporciona conocimientos profundos sobre el funcionamiento de los receptores ionotrópicos. Los receptores nicotínicos son complejos proteicos grandes que consisten en cinco subunidades. En la unión neuromuscular, el nAChR contiene dos subunidades α, cada una de las cuales tiene un sitio de fijación que se une a una sola molécula de acetilcolina. Ambos sitios de fijación de la ACh deben estar ocupados por el receptor para ser activados, de modo que solo concentraciones relativamente altas de acetilcolina activan estos receptores. Estas subunidades también se unen a otros ligandos, como la nicotina y la α-bungarotoxina. Las subunidades α se combinan hasta con otros cuatro tipos de subunidad: β, δ y γ o ε– en la relación 2α:1β:1δ:1γ/ε. Los nAChR neuronales difieren de los del músculo en que carecen de sensibilidad a α-bungarotoxina y solo comprenden dos tipos de subunidades de receptores (α y β), que están presentes en una relación 3α:2β. Cada subunidad del receptor contiene una región extracelular grande (que, en las subunidades α, contiene el sitio de fijación de la ACh) y cuatro dominios que atraviesan la membrana (Figura 6.3A). Los dominios transmembrana de las cinco subunidades individua- NEUROTRANSMISORES Y SUS RECEPTORES (A) (B) Sitio fijador de la acetilcolina (C) FIGURA 6.3 Estructura del receptor nACh. (A) Estructura de la subunidad α del receptor. Sitios fijadores de la acetilcolina Cada subunidad atraviesa cuatro veces la membrana; la subunidad α contiene además un sitio de fijación para la acetilcolina en su dominio extracelular. (B) Cinco subunidades se reúnen para formar un receptor colinérgico nicotínico (AChR) b ` Exterior de la célula Membrana plasmática completo. (C) Vista del AChR desde la perspectiva de la hendidura sináptica. Se aprecia la disposición de las cinco subunidades, donde cada subunidad _ Poro contribuye con una hélice transmembrana que _ a Citoplasma forma el poro del canal. (D) Vista transversal del dominio transmembrana del AChR. Los orificios en cualquiera de los extremos del poro del canal son muy grandes, y el poro se estrecha en la puerta del canal. La esfera turquesa indica la dimensión de un ión sodio (0,3 nm de diámetro). (E) Modelo de la compuerta del AChR. La fijación de ACh a sus sitios (E) (D) Extracelular ACh Poro 113 fijadores sobre las dos subunidades α produce un cambio conformacional en parte del dominio extracelular, que hace que las hélices formadoras de poros se muevan y abran la puerta del poro. (F) Se reúnen distintas subunidades para formar receptores ionotrópicos de neurotransmisores. (A-C de Unwin, 2005; D, E de Miyazawa y cols., 2003.) Puerta Citoplasma (F) Receptor nACh AMPA NMDA Cainato GABA Glicina Serotonina Purinas Subunidades (se requieren combinaciones de 4 o 5 para cada tipo de receptor) _< GluA1 GluN1 GluK1 _< _< 5-HT3A P2X1 ` 5-HT3B P2X2 `< GluA3 GluN2A GluK2 `< a GluA3 GluN2B GluK3 a< 5-HT3C P2X3 b GluA4 GluN2C GluK4 b 5-HT3D P2X4 GluN2D GluK5 ¡ 5-HT3E P2X5 ¡ GluN3A e P2X6 GluN3B d P2X7 l< les unidas forman un canal con un poro central que atraviesa la membrana (Figura 6.3B, C). El ancho de este poro (Figura 6.3D) es sustancialmente más grande que el de los poros de los canales iónicos con puerta de voltaje (véase la Figura 4.7), lo que es compatible con la capacidad relativamente baja de los nAChR para discriminar entre diferentes cationes. En el interior de este poro hay una constricción que puede representar la puerta del receptor. Se cree que la unión de la acetilcolina con las subunidades α produce un cambio de conformación que reorganiza los dominios transmembrana del receptor, que, de este modo, abren la puerta y permiten que los iones difundan a través del poro del canal (Figura 6.3E). En resumen, el nACh es un canal iónico con puerta de ligando. La íntima asociación de los sitios de fijación de la ACh de este receptor con el poro del canal permite la rápida respuesta a la ACh característica de estos receptores. Esta disposición general –varias subunidades receptoras que se reúnen para formar un canal iónico con puerta de ligando– es característica de todos los receptores ionotrópicos en las sinapsis de acción rápida, lo que al mismo tiempo explica su importancia en varias enfermedades neuromusculares (Recuadro 6B). En la Figura 6.3F se resumen las subunidades utilizadas para formar otros tipos de receptores de neurotransmisores ionotrópicos. 114 CAPÍTULO 6 RECUADRO 6A Neurotoxinas que actúan sobre los receptores postsinápticos Las plantas y los animales venenosos se encuentran ampliamente distribuidos en la naturaleza. Las toxinas que sintetizan han sido utilizadas para distintos fines, que incluyen la caza, la curación, la alteración del sensorio y, de forma más reciente, la investigación. Muchas de estas toxinas tienen una acción potente sobre el sistema nervioso y, a menudo, interfieren en la transmisión sináptica al tener por diana los receptores de los neurotransmisores. Los venenos hallados en algunos organismos contienen un único tipo de toxina, mientras que otros contienen una mezcla de decenas o incluso centenas de toxinas. Dado el papel central de los receptores de la acetilcolina en la mediación de la contracción muscular en las uniones neuromusculares de muchas especies, no es sorprendente que gran cantidad de toxinas naturales interfieran en la transmisión en esta sinapsis. De hecho, la clasificación de los receptores de la acetilcolina nicotínicos y muscarínicos se basa sobre la sensibilidad de estos receptores a los alcaloides vegetales tóxicos nicotina y muscarina, los cuales activan los receptores de la acetilcolina nicotínicos y muscarínicos, respectivamente. La nicotina deriva de las hojas desecadas de la planta de tabaco Nicotinia tabacum y la muscarina del hongo rojo venenoso Amanita muscaria. Ambas toxinas son estimulantes que producen náuseas, vómitos, confusión mental y convulsiones. La intoxicación con muscarina también puede conducir al colapso circulatorio, el coma y la muerte. El veneno α-bungarotoxina, uno de los muchos péptidos que en conjunto constituyen el veneno de la víbora krait rayada, Bungarus multicinctus (Figura A), bloquea la transmisión en las uniones neuromusculares y es utilizado por la víbora para paralizar a su presa. Esta toxina de 74 aminoácidos bloquea la transmisión neuromuscular al unirse de forma irreversible a los receptores de la acetilcolina nicotínicos, e impedir así que la ACh abra los canales iónicos postsinápticos. La parálisis se produce porque los músculos esqueléticos ya no pueden ser activados por las neuronas motoras. Como resultado de su especificidad y de su alta afinidad por los receptores nicotínicos, la α-bungarotoxina ha contribuido mucho al conocimiento de la molécula del receptor de la acetilcolina. Otras toxinas de las víboras que bloquean los receptores nicotínicos son la α-neurotoxina de la cobra y el péptido de la víbora de mar erabutoxina. La misma estrategia utilizada por estas víboras para paralizar a su presa fue adoptada por los aborígenes sudamericanos que empleaban curare, una mezcla de toxinas vegetales de Chondrodendron tomentosum, como veneno en la punta de las flechas para inmovilizar a sus presas. El curare también bloquea los receptores nicotínicos; el agente activo es el alcaloide δ-tubocurarina. Otra clase interesante de toxinas animales que bloquean selectivamente los receptores de la acetilcolina y otros receptores incluye los péptidos producidos por los caracoles cónicos marinos cazadores de peces (Figura B). Estos caracoles coloridos matan a los pequeños peces “disparándoles” flechas envenenadas. El veneno contiene cientos de péptidos, conocidos como “conotoxinas”, muchas de las cuales están dirigidas contra proteínas importantes en la transmisión sináptica. Hay péptidos de conotoxina que bloquean los canales del Ca2+, los canales del Na+, los receptores del glutamato y de la acetilcolina. El conjunto de respuestas fisiológicas producidas por estos péptidos sirve para inmovilizar a cualquier presa que desafortunadamente se encuentre con el caracol cónico. Muchos otros organismos, incluidos moluscos, corales, gusanos y ranas, también utilizan toxinas que contienen bloqueantes específicos de receptores de la acetilcolina. Otras toxinas naturales poseen efectos que alteran el sensorio o la conducta y, en algunos casos, han sido utilizadas durante miles de años por los chamanes y, más recientemente, por los médicos. Dos ejemplos son las toxinas alcaloides vegetales que bloquean los receptores muscarínicos: la atropina de la belladona y la escopola- (A) (B) (C) (A) Víbora krait rayada Bungarus multicinctus. (B) Un caracol cónico marino (especie de los Conus) utiliza dardos venenosos para matar a los peces pequeños. (C) Semilla de betel, Areca catechu, que crece en Malasia. (A, Robert Zappalorti/Photo Researchers, Inc.; B, Zoya Maslak y Baldomera Olivera, University of Utah; C, Fletcher y Baylis/Photo Researchers, Inc.) NEUROTRANSMISORES Y SUS RECEPTORES 115 RECUADRO 6A (continúa) mina del beleño. Dado que estas plantas crecen de forma salvaje en muchas partes del mundo, la exposición no es infrecuente y el envenenamiento con cualquiera de las toxinas también puede ser fatal. Otra neurotoxina postsináptica que, al igual que la nicotina, es utilizada como droga social, se encuentra en las semilla del betel, Areca catechu (Figura C). El mascado del betel, si bien se desconoce en los Estados Unidos de América, es practicado hasta por el 25% de la población de la India, Bangladés, Sri Lanka, Malasia y las Filipinas. El mascado de estas semillas produce euforia causada por la arecolina, un alcaloide agonista de los receptores nicotínicos. Al igual que la nicotina, la arecolina es un estimulante del sistema nervioso central que produce adicción. Muchas otras neurotoxinas alteran la transmisión en las sinapsis no colinérgicas. Por ejemplo, los aminoácidos hallados en algunos hongos, algas y semillas son potentes agonistas de los receptores del glutamato. Los aminoácidos excitotóxicos cainato, provenientes del alga roja Digenea simplex y quiscualato, de la semilla de Quisqualis indica, son utilizados para separar dos familias de receptores del glutamato no NMDA (véase el texto). Otros activadores aminoácidos neurotóxicos de los receptores de glutamato incluyen ácido iboténico y ácido acromélico, hallados ambos en hongos, y el domoato, el cual se desarrolla en algas de agua dulce, algas marinas y mejillones. Otro grupo grande de neurotoxinas peptídicas bloquea los receptores del glutamato. Estas incluyen las α-agatoxinas de la araña de red “en embudo”, la NSTX-3 de la araña tejedora de esferas, la jorotoxina de la araña Joro y la β-filantotoxina del veneno de la avispa, así como muchas toxinas del caracol cónico. Todas estas toxinas de las que nos ocupamos hasta ahora se dirigen contra las sinapsis excitadoras. Sin embargo, los receptores del GABA y de la glicina inhibidores no han sido pasados por alto por las exigencias de la supervivencia. La estricnina, un alcaloide extraído de las semillas de Strychnos nux-vomica, es la única droga conocida con acciones específicas sobre la transmisión en las sinapsis glicinérgicas. Puesto que la toxina bloquea los receptores de la glicina, el envenenamiento con estrictina produce hiperactividad en la médula espinal y en el tronco del encéfalo, y conduce a convulsiones. La estricnina se utiliza desde hace mucho tiempo en el comercio como veneno contra los roedores, aunque en la actualidad son más populares otras alternativas como el anticoagulante cumadina porque son más seguros para los seres humanos. Las neurotoxinas que bloquean los receptores del GABAA incluyen los alcaloides vegetales como bicuculina Una segunda clase de receptores colinérgicos es activada por la muscarina: un alcaloide venenoso hallado en algunos hongos (véase, Recuadro 6A); y, por lo tanto, se los denominan receptores colinérgicos muscarínicos (mAChR). Estos receptores son metabotrópicos y median la mayor parte de los efectos de la ACh en el encéfalo. Al igual que otros receptores metabotrópicos, los mAChR tienen siete dominios helicoidales que atraviesan la membrana (Figura 6.4A). La ACh se une en un único sitio de fijación sobre la superficie extracelular del mAChR; este sitio de fijación está formado por asas que conectan varias de las hélices transmembrana (Figura 6.4B). La unión de la acetilcolina en este sitio produce un cambio de conformación que permite a las proteínas G unirse al dominio citoplasmático del mAChR (Figura 6.4A). Se conocen cinco subtipos de mAChR (Figura 6.4C) que están acoplados a diferentes tipos de proteínas G, lo que produce así distintas respuestas postsinápticas lentas. Los receptores colinérgicos muscarínicos tienen una expresión importante en el de la dicentra (Dutchman’s breeches) y la picrotoxina de Anamirta cocculus. El dieldrín, un insecticida comercial, también bloquea estos receptores. Al igual que la estricnina, estos agentes son estimulantes potentes del sistema nervioso central. El muscimol, una toxina de un hongo que es un potente depresor así como un alucinógeno, activa los receptores del GABAA. Un análogo sintético del GABA, el baclofeno, es un agonista de los receptores del GABAB que reduce los PPSE en algunas neuronas del tronco del encéfalo, y es utilizado clínicamente para reducir la frecuencia y la gravedad de los espasmos musculares. La guerra química contra las especies ha dado origen así a un conjunto enorme de moléculas cuyo punto diana son las sinapsis de todo el sistema nervioso. Si bien estas toxinas están ideadas para doblegar la transmisión sináptica normal, también proporcionan un conjunto de herramientas potentes para comprender los mecanismos postsinápticos. Bibliografía Lewis, R. L. and L. Gutmann (2004) Snake venoms and the neuromuscular junction. Seminars Neurol. 24: 175-179. Olivera, B. M. (2006) Conus peptides: Biodiversity-based discovery and exogenomics. J. Biol Chem. 281: 31173-31177. estriado y en otras distintas regiones del encéfalo anterior, donde activan canales del K+ rectificadores hacia el interior o canales del K+ activados por el Ca2+, los cuales ejercen de ese modo una influencia inhibidora sobre los efectos motores mediados por la dopamina. En otras partes del encéfalo, como en el hipocampo, los mAChR son excitadores y actúan cerrando los canales del K+ tipo KCNQ. Estos receptores también se encuentran en los ganglios del sistema nervioso periférico. Por último, los mAChR median las respuestas colinérgicas periféricas de los órganos efectores autónomos como el corazón, el músculo liso y las glándulas exocrinas, y son responsables de la inhibición de la frecuencia cardíaca por el nervio vago. Muchos fármacos actúan como agonistas o antagonistas de los mAChR; los bloqueantes de los receptores colinérgicos muscarínicos que son útiles desde el punto de vista terapéutico incluyen atropina (utilizada para dilatar la pupila), escopolamina (efectiva para prevenir la enfermedad del movimiento) e ipratropio (útil para el tratamiento contra el asma). 116 CAPÍTULO 6 (A) (B) FIGURA 6.4 Receptores muscarínicos y otros receptores metabotrópicos. Acetylcholine Acetilcolina fijada bound to mAChR al mAChR Extracelular (A) Estructura prevista del mAChR M1 humano. Este receptor atraviesa 7 veces la membrana plasmática y tiene un dominio citoplasmático, que se une a las proteínas G y las activa, así como un dominio extracelular que se une a la Membrana plasmática acetilcolina. (B) Vista de la superficie extracelular del mAChR que muestra la acetilcolina (esferas coloreadas) unida a su sitio de fijación. Citoplasma (C) Variedades de receptores metabotrópicos Sitio fijador de la proteína G de los neurotransmisores. (A, B de Peng y cols., 2006, cortesía de W. Goddard.) (C) Clases de Muscarínicos receptores Subtipos de receptores Histamina Serotonina Purinas Alfa H1 5-HT1A Adenosina D2 _A H2 5-HT1B A1 mGlu D3 _B H3 5-HT1D A2A M Clase II D4 _D H4 5-HT1E A2B M mGlu D5 _A 5-HT1F A3 _B 5-HT2A P2Y Clase III _C 5-HT2B P2Y1 mGlu Beta 5-HT2C P2Y2 mGlu ` 5-HT4 P2Y4 mGlu ` 5-HT5A P2Y6 mGlu ` 5-HT6 P2Y11 5-HT7 P2Y12 Glutamato GABAB M Clase I GABAB1 D1 M mGlu GABAB2 M mGlu Dopamina Adrenérgicos P2Y13 P2Y14 Glutamato El glutamato es el transmisor más importante para la función encefálica normal. Casi todas las neuronas excitadoras del sistema nervioso central son glutamatérgicas y se estima que más del 50% de todas las sinapsis encefálicas libera este neurotransmisor. Durante el traumatismo encefálico, ocurre una liberación excesiva de glutamato que puede producir daño encefálico “excitotóxico” (Recuadro 6C, p. 121). El glutamato es un aminoácido no esencial, que no atraviesa la barrera hematoencefálica y, por lo tanto, debe sintetizarse en las neuronas a partir de precursores locales. El precursor más prevalente de la síntesis de glutamato es la glutamina, que es captada en las terminaciones presinápticas por el transportador 2 del sistema A (SAT2) y metabolizada luego a glutamato por la enzima mitocondrial glutaminasa (Figura 6.5). La glucosa metabolizada por las neuronas también puede ser utilizada para sintetizar glutamato por la transaminación de 2-oxoglutarato, intermediario del ciclo de los ácidos tricarboxílicos (Krebs). El glutamato sintetizado en el citoplasma presináptico es empaquetado en vesículas sinápticas por los transportadores vesiculares de glutamato (VGLUT). Se han identificado por lo menos tres genes para estos transportadores y diferentes transportadores vesiculares de glutamato participan en el empaquetamiento de glutamato en las vesículas en diferentes tipos de terminaciones sinápticas glutamatérgicas. Una vez liberado, el glutamato es eliminado de la hendidura sináptica por los transportadores de aminoácidos excitadores (EAATS). Estos transportadores representan una familia de cinco cotransportadores de glutamato Na+-dependientes diferentes. Algunos transportadores de aminoácidos excitadores están presentes en las células gliales y otros en las terminaciones presinápticas. El glutamato transportado en las células gliales por medio de los transportadores de aminoácidos excitadores es convertido en glutamina por la enzima glutaminasintetasa. La glutamina es transportada luego hacia el exterior de las células gliales por un transportador diferente, el transportador 1 del sistema N (SN1), y transportado en las terminaciones nerviosas por medio NEUROTRANSMISORES Y SUS RECEPTORES 117 RECUADRO 6B Miastenia grave: una enfermedad autoimmunitaria de las sinapsis neuromusculares La miastenia grave, enfermedad que interfiere en la transmisión entre las neuronas motoras y las fibras del músculo esquelético, afecta aproximadamente a 1 de cada 200 000 personas. Descrita originariamente por Thomas Willis en 1685, la característica de esta enfermedad es la debilidad muscular, sobre todo, durante la actividad sostenida (Figura A). Si bien la evolución es variable, la miastenia afecta comúnmente los músculos que controlan los párpados (da por resultado la caída del párpado o ptosis) y los movimientos oculares (da por resultado la visión doble o la diplopía). Otros puntos diana frecuentes de la enfermedad son los músculos que controlan la expresión facial, la masticación, la deglución y la palabra. Un indicio importante de la causa de la miastenia grave provino de la observación clínica de que la debilidad muscular mejora tras el tratamiento con inhibidores de la acetilcolinesterasa, la enzima que normalmente degrada la acetilcolina en la unión neuromuscular (véase la Fig. A). Algunos estudios del músculo obtenidos por biopsia de pacientes miasténicos mostraron que tanto los potenciales de la placa terminal (PPT) como los potenciales de la placa terminal en miniatura (PPTM) son mucho más pequeños que lo normal (Figura B). Ya que tanto la frecuencia de los PPTM como el contenido cuántico de los PPT son normales, al parecer la miastenia grave comprende un trastorno de las células musculares postsinápticas. En efecto, la microscopia electrónica muestra que la estructura de las uniones neuromusculares está alterada; los cambios incluyen ensanchamiento de la hendidura sináptica y reducción aparente en la cantidad de receptores de la acetilcolina en la membrana postsináptica. Una observación fortuita condujo al descubrimiento de la causa subyacente de estos cambios a comienzos de la década de 1970. Jim Patrick y Jon Lindstrom, que trabajaban entonces en el Salk Institute, intentaban obtener anticuerpos contra los receptores de la acetilcolina nicotínicos inmunizando conejos con los receptores. Inesperadamente, los conejos inmunizados desarrollaron debilidad muscular, que mejoró después del tratamiento con inhibidores de la acetilcolinesterasa. La investigación posterior mostró que la sangre de los pacientes miasténicos contiene anticuerpos dirigidos contra el receptor de la acetilcolina y que estos anticuerpos están presentes en las sinapsis neuromusculares. La eliminación de los anticuerpos mediante plasmaféresis mejora la debilidad. Finalmente, la inyección de suero de pacientes miasténicos en ratones produce efectos miasténicos (porque el suero transporta anticuerpos circulantes). Registro electromiográfico de los potenciales de acción musculares (mV) (A) Normal 8 Miastenia grave 4 0 –4 Cantidad de PPTM (B) 0 20 40 60 80 0 20 40 60 80 Tiempo (ms) 15 Miastenia grave 10 Normal Estos hallazgos indican que la miastenia grave es una enfermedad autoinmunitaria dirigida contra los receptores de la acetilcolina nicotínicos. La respuesta inmunitaria reduce la cantidad de receptores funcionales en la unión neuromuscular y, finalmente, los destruye disminuyendo la eficiencia de la transmisión sináptica. La debilidad muscular se desarrolla porque las neuronas motoras son menos capaces de excitar a las células musculares postsinápticas. Esta secuencia causal también explica por qué los inhibidores de la colinesterasa alivian los síntomas. Los inhibidores aumentan la concentración de acetilcolina en la hendidura sináptica, lo que permite una activación más eficaz de los receptores postsinápticos que aún no han sido destruidos por el sistema inmunitario. A pesar de todos estos hallazgos, todavía no está claro qué es lo que lleva al sistema inmunitario a producir una respuesta autoinmunitaria contra los receptores de la acetilcolina. Bibliografía Elmqvist, D., W. W. Hofmann, J. Kugelberg and D. M. J. Quastel (1964) An electrophysiological investigation of neuromuscular transmission in myasthenia gravis. J. Physiol. (Lond.) 174: 41734. Farrugia, M. E. and A. Vincent (2010) Autoimmune mediated neuromuscular junction defects. Curr. Opin. Neurol. 23: 489-495. Harvey, A. M. and J. L. Lilienthal (1941) Miastenia grave después del Observations on the nature of myasthenia gravis: tratamiento con neostigmina The intra-arterial injection of acetylcholine, prostigmine, and adrenaline. Bull. Johns Hopkins Hosp. 69: 566-577. Patrick, J. and J. Lindstrom (1973) Autoimmune response to acetylcholine receptor. Science 180: 871-872. Vincent, A. (2002) Unravelling the pathogenesis of myasthenia gravis. Nature Rev. Immunol. 2: 797-804. 0 20 40 60 80 La miastenia grave reduce la eficiencia de la transmisión neuromuscular. (A) Los electromiogramas muestran respuestas musculares obtenidas por la estimulación de los nervios motores. En los individuos normales, cada estímulo de una sucesión provoca la misma respuesta contráctil. Por el contrario, la transmisión se fatiga rápidamente en los pacientes miasténicos, pero puede ser restablecida parcialmente administrando el inhibidor de la colinesterasa neostigmina. (B) Distribución de las amplitudes de los potenciales de la placa terminal en miniatura (PPTM) en las fibras musculares de los pacientes miasténicos y en controles. El 5 0,05 0,10 0,20 0,50 1 Amplitud de los PPTM (mV) 2 3 tamaño más pequeño de los PPTM en los miasténicos se debe a una cantidad disminuida de receptores postsinápticos. (A, de Harvey y Lilienthal, 1941; B, de Elmqvist y cols., 1964.) 118 CAPÍTULO 6 FIGURA 6.5 La síntesis del glutamato y el ciclo entre las neuronas y la glía. La SN1 SAT2 Célula glial Terminación presináptica Glutamina Glutamina COO< + H3N CH Glutamina sintetasa O CH2 CH2 C NH2 Glutamato Glutaminasa COO< + NH3 CH CH2 CH2 COO Glutamato EAAT VGLUT Célula postsináptica gliales, el glutamato es convertido en glutamina por la glutamina sintetasa y liberado por las células gliales a través del transportador SN1. La glutamina es captada en las terminaciones nerviosas a través de transportadores SAT2 y convertida nuevamente en glutamato por la glutaminasa. Después, el glutamato es cargado < a Gl u ta m acción del glutamato liberado en la hendidura sináptica es terminada por la captación en las células gliales (y las neuronas) circundantes a través de los transportadores de aminoácidos excitadores (EAAT). En el interior de las células en las vesículas sinápticas a través de transportadores vesiculares de glutamato (VGLUT) para completar el ciclo. to Receptores de glutamato del SAT2. Esta secuencia global de acontecimientos se denomina ciclo de glutamato-glutamina (véase la Fig. 6.5). Este ciclo permite que tanto células gliales como terminaciones presinápticas cooperen para mantener un aporte suficiente de glutamato para la transmisión sináptica y terminar la acción postsináptica del glutamato. Existen varios tipos de receptores ionotrópicos de glutamato (véase Fig. 6.3F). Los receptores de AMPA, los receptores de NMDA y los receptores de cainato se denominan según los agonistas que los activan: AMPA (α-amino-3-hidroxil-5-metil4-isoxazol-propionato), NMDA (N-metil-d-aspartato) y ácido caínico, respectivamente. Todos estos receptores son canales catiónicos con puerta de glutamato que permiten el pasaje de Na+ y K+, de modo similar a los nAChR. Por ello, la activación de los receptores de AMPA, cainato y NMDA siempre produce respuestas postsinápticas excitadoras. La mayoría de las sinapsis centrales posee tanto receptores de AMPA como de NMDA. A menudo, se utilizan fármacos antagonistas que bloquean selectivamente estos receptores para identificar las respuestas sinápticas mediadas por cada tipo de receptor. Estos experimentos muestran que los PPSE producidos por los receptores de NMDA son más lentos y duran más que los producidos por los receptores de AMPA (Figura 6.6A). Los PPSE generados por los receptores de AMPA habitualmente son mucho más grandes que los producidos por otros tipos de receptores ionotrópicos del glutamato, de modo que los receptores de AMPA representan los mediadores primarios de la transmisión excitadora en el encéfalo. Las funciones fisiológicas de los receptores de cainato no están tan bien definidas; en algunos casos, estos receptores se encuentran en las terminaciones presinápticas y sirven como mecanismos de retroalimentación para regular la liberación del glutamato. Cuando se encuentran sobre las células postsinápticas, los receptores del cainato generan PPSE, que se elevan rápidamente pero disminuyen más lentamente que los mediados por los receptores de AMPA (Figura 6.6B). Los receptores de NMDA tienen propiedades fisiológicas que los separan de los otros receptores ionotrópicos de glutamato. Tal vez el hecho más importante sea que el poro del canal del receptor de NMDA permite el ingreso de Ca2+ además de Na+ y de K+. En consecuencia, los PPSE producidos por los receptores de NMDA aumentan la concentración de Ca2+ en el interior de la neurona postsináptica, y el Ca2+ actúa entonces como segundo mensajero para activar las cascadas de señalizaciones intracelulares (véase el Cap. 7). Otra propiedad clave es que el Mg2+ bloquea el poro de este canal en los potenciales de membrana hiperpolarizados, mientras que la despolarización empuja el Mg2+ fuera del poro (Figura 6.6). Esto implica una dependencia peculiar del voltaje al flujo desde la corriente a través del receptor (línea roja en la Figura 6.6D); la eliminación del Mg2+ extracelular evita el comportamiento (línea azul), lo que demuestra que el Mg2+ confiere la dependencia del voltaje. A causa de esta propiedad, los receptores de NMDA dejan pasar cationes (principalmente Ca2+) solo cuando el potencial de membrana postsináptico se encuentra despolarizado, como durante la activación de aferencias excitadoras fuentes o durante la descarga del potencial de acción en la célula postsináptica. Este requerimiento de la presencia coincidente tanto de glutamato como la de la despolarización postsináptica para abrir los receptores de NMDA NEUROTRANSMISORES Y SUS RECEPTORES (A) (B) 0 Amplitud del PPSE (pA) 119 0 NMDA –25 –5 AMPA –75 –10 Cainato –20 AMPA –30 0 25 (C) 50 Tiempo (ms) 75 100 Hiperpolarizado, el Mg2+ lo bloquea Poro del canal 0 100 200 Tiempo (ms) FIGURA 6.6 Respuestas postsinápticas mediadas por los re- Despolarizado, sin el bloqueo de Mg2+ ceptores del glutamato ionotrópicos. (A) Contribuciones de los Mg Na Ca Glutamato Mg K (D) 150 PPSE (pA) 100 0 Glutamato + Mg2 <100 Glutamato, sin Mg2 <150 <100 <50 0 50 Potencial de membrana (mV) receptores del AMPA y del NMDA a los PPSE en una sinapsis entre una célula piramidal presináptica y una interneurona postsináptica en la corteza visual. El bloqueo de los receptores del NMDA pone en evidencia un PPSE grande y rápido mediado por los receptores del AMPA, mientras que el bloqueo de estos últimos pone en evidencia un componente más lento del PPSE mediado por los receptores del NMDA. (B) Contribuciones de los receptores del AMPA y del cainato a los PPSE en miniatura en las sinapsis excitadoras formadas entre las fibras musgosas y las células piramidales CA3 en el hipocampo. Los antagonistas farmacológicos muestran que el componente de los PPSE mediado por los receptores del AMPA es más grande y más rápido que el mediado por los receptores del cainato. (C) Bloqueo dependiente del voltaje del poro del receptor del NMDA por Mg2+. En potenciales hiperpolarizados, el Mg2+ reside en el interior del poro del canal y lo bloquea (izquierda). La despolarización del potencial de membrana empuja el Mg2+ fuera del poro, de modo que la corriente puede fluir a través de los receptores de los NMDA activados por glutamato. En presencia de Mg2+ (rojo), el bloqueo del Mg2+ del poro del canal impide el flujo de corriente en los potenciales de membrana hiperpolarizados. Si se elimina el Mg2+ extracelular, no hay bloqueo del poro del canal. (A, de Watanabe y cols., 2005; B, de Mott y cols., 2008.) 50 <50 300 100 es ampliamente considerado subyacente a ciertas formas de almacenamiento de la información sináptica, como la plasticidad sináptica a largo plazo (véase el Cap. 8). Otra característica poco habitual es que la apertura de la puerta de los receptores de NMDA requiere un coagonista: el aminoácido glicina, que está presente en el medioambiente extracelular del encéfalo. Al igual que otros receptores ionotrópicos, los receptores de AMPA están compuestos por múltiples subunidades. Las cuatro subunidades diferentes del receptor de AMPA se designan de GluA1 a GluA4 (véase Fig. 6.3F). Cada subunidad tiene varios dominios diferentes, que incluye un dominio extracelular para fijar el ligando que es responsable de la fijación del ligando, y un dominio transmembrana que forma parte del canal iónico (Figura 6.7A). Estas subunidades están organizadas en la estructura tetramérica que se muestra en las Figuras 6.7B-D. Al contrario de los nAChR, la estructura extracelular de los receptores de AMPA es asimétrica y, por lo tanto, se ve diferente cuando se la visualiza desde las superficies frontal y lateral (Figura 6.7B, C). El receptor de AMPA tiene forma de Y (Figura 6.7B), y los dominios extracelulares grandes de las subunidades se estrechan hacia abajo a medida que el receptor atraviesa la membrana plasmática (Figura 6.7D). Los dominios extracelulares para fijar el ligando tienen una forman característica en “concha de almeja”, donde el glutamato y otros ligandos se unen en el interior de la apertura de la concha (círculo de la Figura 6.7C). El dominio transmembrana consiste en unas hélices que forman tanto el poro del canal como una puerta que ocluye el poro cuando el glutamato no está unido al receptor. La unión del glutamato hace 120 CAPÍTULO 6 (A) (B) N Dominio de amino terminal (ATD) Dominio del ligando (LBD) Glutamato (C) (D) ATD Receptor antagonista del AMPA LBD Extracelular Dominio de la transmembrana (TMD) Membrana plasmática C Dominio del carboxilo terminal (CTD) TMD Poro Citoplasma (E) (F) Cerrado Glutamato GluN1 GluN2A Abierto Glicina GluN2A Glutamato Puerta Poro FIGURA 6.7 Estructura del receptor del AMPA. (A) Estructura de dominio de una subunidad del receptor del AMPA. La parte más grande de cada subunidad es extracelular y consiste en dos dominios: el dominio aminoácido (ATD) y el dominio de fijación del ligando (LBD). Además, el dominio transmembrana (TMD) forma parte del poro del canal iónico, y un dominio carboxiterminal (CTD) conecta el receptor con las proteínas intracelulares. (B-D) Estructura cristalográfica del receptor del AMPA. Cada una de las cuatro subunidades está indicada en un color diferente. (B) Desde esta perspectiva, se aprecia la forma en Y del receptor del AMPA. (C) Después de rotar 90 grados el receptor, se observan las dimensiones asimétricas. Se aprecia un dominio de fijación del ligando que está ocupado por un fármaco antagonista (en círculos). (D) Las vistas transversales del receptor del AMPA en dos posiciones diferentes (flechas grises) muestran las relaciones que la estructura de la concha se “cierre”; este movimiento hace entonces que las hélices el interior del dominio transmembrana se muevan y abran así el poro del canal (Figura 6.7E). Los receptores de NMDA también son conjuntos tetraméricos de subunidades. Existen tres grupos de subunidades del receptor de NMDA (GluN1, GluN2 y GluN3), con un total de siete tipos diferentes de subunidades (véase Fig. 6.3F). Mientras las subunidades GluN2 fijan el glutamato, las subunidades GluN1 y GluN3 fijan la glicina. Los tetrámeros del receptor de NMDA típicamente están compuestos por dos subunidades que fijan glutamato (GluN2) y dos subunidades que fijan glici- espaciales entre las subunidades y también demuestran los cambios en la forma que ocurren a lo largo del receptor. (E) Modelo para la apertura con puerta del receptor del AMPA por el glutamato. Se muestra el dominio transmembrana (hélices azules) y parte del dominio fijador de ligando extracelular. La fijación del glutamato cierra la estructura en forma de concha de almeja del dominio fijador del ligando, lo que conduce al movimiento de las hélices de la puerta (flechas laterales) que abre el poro del canal. (F) Modelo de la estructura de un receptor del NMDA que consiste en las subunidades GluN1 y GluN2A. El dominio fijador de ligando de GluN2A se une al glutamato, mientras que el dominio fijador de ligando de GluN1 se une a la coagonista, la glicina. (A, E de Traynelis y cols., 2010; B-D y F de Sobolevsky y cols., 2009.) na (GluN1). En algunos casos, la GluN3 reemplaza a algunas subunidades GluN2. Esta mezcla de subunidades asegura que el receptor se una tanto al glutamato liberado de las terminaciones presinápticas como al coagonista glicina del ambiente. Se cree que las subunidades del receptor de NMDA forman una estructura que se asemeja mucho a los receptores de AMPA (Figura 6.7F). Además de estos receptores ionotrópicos del glutamato, hay tres tipos de receptores del glutamato metabotrópicos (mGluR) (véase Fig. 6.4C). Estos receptores, que modulan de manera indirecta los canales iónicos postsinápticos, difieren de los NEUROTRANSMISORES Y SUS RECEPTORES 121 RECUADRO 6C Excitotoxicidad en la lesión encefálica aguda La excitotoxicidad se refiere a la capacidad del glutamato y de los compuestos afines para destruir neuronas a través de la transmisión sináptica prolongada. Normalmente, la concentración de glutamato liberado en la hendidura sináptica se eleva hasta niveles altos (aproximadamente, 1 mM), pero se mantiene en esta concentración solo durante algunos milisegundos. Si se acumulan concentraciones anormalmente elevadas en la hendidura, la activación excesiva de los receptores neuronales del glutamato puede, literalmente, excitar a las neuronas hasta producir la muerte. El fenómeno de la excitotoxicidad fue descubierto en 1957, cuando D. R. Lucas y J. P. Newhouse observaron de manera accidental que, si se alimenta a ratones lactantes con glutamato de sodio, se destruyen las neuronas de la retina. Alrededor de una década después, J. Olney en la Washington University amplió este descubrimiento al demostrar que, en el encéfalo, se desarrollan regiones de pérdida neuronal inducida por el glutamato. El daño está limitado evidentemente a las células postsinápticas –las dendritas de las neuronas diana estaban groseramente tumefactas–, mientras que las terminaciones presinápticas estaban conservadas. Olney también examinó la potencia relativa de los análogos del glutamato y observó que sus acciones neurotóxicas corrían paralelas a su capacidad para activar los receptores postsinápticos del glutamato. Además, los antagonistas de los receptores del glutamato eran eficaces para bloquear los efectos neurotóxicos del glutamato. A la luz de esta evidencia, Olney postuló que el glutamato destruye las neuronas por un mecanismo similar a la transmisión en las sinapsis glutamatérgicas excitadoras, y acuñó el término excitotóxico para referirse a este efecto. Las pruebas de que la excitotoxicidad es una causa importante de daño neuronal después de la lesión encefálica se obtuvieron fundamentalmente del estudio de las consecuencias de la reducción del flujo sanguíneo. La causa más frecuente de reducción del flujo sanguíneo encefálico (isquemia) es la oclusión de un vaso sanguíneo cerebral (es decir, un accidente cerebrovascular; véase el Recuadro B del Apéndice). La idea de que la actividad sináptica excesiva contribuye a la lesión isquémica surgió de la observación de que las concentraciones de glutamato y de aspartato en el espacio extracelular que rodea a las neuronas aumentan durante la isquemia. Además, la microinyección de antagonistas de los receptores de glutamato en animales de experimentación protege a las neuronas del daño inducido por la isquemia. En conjunto, estos hallazgos sugieren que la acumulación extracelular de glutamato durante la isquemia activa de manera excesiva los receptores de glutamato, y que, de alguna forma, esto desencadena una cadena de acontecimientos que conduce a la muerte neuronal. Presumiblemente, la disminución del aporte de oxígeno y glucosa eleva los niveles extracelulares de glutamato al hacer más lenta la eliminación de glutamato dependiente de energía en las sinapsis. En la actualidad, los mecanismos excitotóxicos han sido involucrados en otras formas agudas de lesión neuronal, que incluyen hipoglucemia, lesión traumática y convulsiones intensas repetidas (denominadas estado de mal epiléptico). Por lo tanto, el conocimiento de la excitotoxicidad tiene importantes consecuencias en el tratamiento contra distintos trastornos neurológicos. Por ejemplo, el bloqueo de ionotrópicos en su acoplamiento a las vías de transducción de señales intracelulares (véase el Cap. 7) y en su sensibilidad a los agentes farmacológicos. La activación de muchos de estos receptores conduce a la inhibición de los canales postsinápticos del Ca2+ y del Na+. A diferencia de los receptores glumatatérgicos ionotrópicos excitadores, los mGluR producen respuestas postsinápticas más lentas que pueden excitar o inhibir las células postsinápticas. En consecuencia, los papeles fisiológicos de los mGluR son muy variados. Aunque no se conoce de modo com- los receptores de glutamato podría, en principio, proteger a las neuronas del daño debido a un accidente cerebrovascular, traumatismo u otras causas. Lamentablemente, los ensayos clínicos de antagonistas de los receptores de glutamato no han permitido mejorar mucho el resultado del accidente cerebrovascular. Es probable que la ineficacia de este tratamiento muy lógico se deba a varios factores, uno de los cuales es el desarrollo de una lesión excitotóxica sustancial muy poco después de la isquemia, antes de la iniciación típica de tratamiento. También es probable que la excitotoxicidad sea solamente uno de los varios mecanismos por los cuales la isquemia daña las neuronas; otra causa podría ser el daño secundario a la inflamación. Con todo, las intervenciones farmacológicas dirigidas contra todos estos mecanismos se muestran muy promisorias para minimizar la lesión encefálica después de un accidente cerebrovascular y otras causas. Bibliografía Lau, A. and M. Tymianski (2010) Glutamate receptors, neurotoxicity and neurodegenera-tion. Pflügers Arch. 460: 525-542. Lucas, D. R. and J. P. Newhouse (1957) The toxic effects of sodium L-glutamate on the inner layers of the retina. Arch. Opthalmol. 58: 193-201. Olney, J. W. (1969) Brain lesions, obesity and other disturbances in mice treated with monosodium glutamate. Science 164: 719-721. Olney, J. W. (1971) Glutamate-induced neuronal necrosis in the infant mouse hypothalamus: An electron microscopic study. J. Neuropathol. Exp. Neurol. 30: 75-90. Rothman, S. M. (1983) Synaptic activity mediates death of hypoxic neurons. Science 220: 536-537. Syntichaki, P. and N. Tavernarakis (2003) The biochemistry of neuronal necrosis: Rogue biology? Nature Neurosci. Rev. 4: 672-684. pleto la estructura de los mGluR, se sabe que sus dominios que fijan glutamato son muy similares a los dominios con forma de concha que fijan los ligandos en los receptores ionotrópicos del glutamato (véanse la Figuras 6.7A, B). Se cree que la organización general de los mGluR es muy similar a la de otros receptores metabotrópicos, como el mAChR (véase la Figura 6.4A) –es decir, siete dominios helicoidales que atraviesan la membrana y conectan el dominio extracelular de fijación del ligando al dominio intracelular que activa las proteínas G–. 122 CAPÍTULO 6 El GABA y la glicina La mayoría de las sinapsis inhibidoras en el encéfalo o en la médula espinal emplea el ácido γ-aminobutírico o la glicina como neurotransmisores. Al igual que el glutamato, el GABA fue identificado en el tejido encefálico durante la década de 1950. Los detalles de su síntesis y degradación fueron descubiertos poco después gracias a la investigación de Ernst Florey y Eugene Roberts. En la misma época, David Curtis y Jeffrey Watkins mostraron por primera vez que el GABA puede inhibir la capacidad de las neuronas de los mamíferos para disparar potenciales de acción. Los estudios posteriores de Edward Kravitz y cols. establecieron que el GABA es un neurotransmisor inhibidor en las sinapsis neuromusculares de la langosta. En la actualidad, se sabe que hasta un tercio de las sinapsis del encéfalo utiliza el GABA como neurotransmisor inhibidor. El GABA se halla más comúnmente en las interneuronas del circuito local, aunque las células de Purkinje del cerebelo brindan un ejemplo de neuronas de proyección GABAérgicas (véase el Cap. 19). El precursor predominante en la síntesis del GABA es la glucosa, que es metabolizada a glutamato por las enzimas del ciclo de los ácidos tricarboxílicos. (El piruvato y el glutamato también pueden actuar como precursores del GABA). La enzima ácido glutámico descarboxilasa (GAD), la cual se encuentra casi exclusivamente en las neuronas GABAérgicas, cataliza la conversión de glutamato a GABA (Figura 6.8A). La enzima requiere un cofactor, el fosfato de piridoxal, para realizar su actividad. Como el fosfato de piridoxal deriva de la vitamina B6, una deficiencia de esta vitamina en la dieta puede conducir a una reducción de la síntesis del GABA. La importancia de este hecho quedó aclarada tras relacionar una trágica serie de muertes de lactantes con la omisión de vitamina B6 en las fórmulas para su alimentación. La falta de vitamina B6 redujo mucho el contenido de GABA del encéfalo, y la pérdida posterior de inhibición sináptica produjo convulsiones que, en algunos casos, fueron fatales. Una vez sintetizado el GABA, es transportado hacia las vesículas sinápticas a través del transportador vesicular de aminoácidos inhibidores (vesicular inhibitory amino acid transporter, VIAAT). El mecanismo de eliminación del GABA es similar al del glutamato: tanto las neuronas como las células gliales contienen cotransportadores de Na+-dependientes de alta afinidad para el GABA. Estos cotransportadores se denominan “GAT”, y se han identificado varias formas. La mayor parte del GABA es convertida finalmente en succinato, el cual es metabolizado además en el ciclo de los ácidos tricarboxílicos, que media la síntesis del ATP celular. Se requieren dos enzimas mitocondriales para su degradación: la GABA transaminasa y la succínico semialdehído deshidrogenasa. También existen otras vías para la degradación del GABA, la más notable de estas conduce a la producción de γ-hidroxibutirato, un derivado del GABA que fue utilizado como droga “date rape” (drogas que facilitan el abuso sexual en las citas). La administración oral de γ-hidroxibutirato puede producir euforia, déficit de memoria e inconciencia. Presumiblemente, estos efectos surgen por acción sobre las sinapsis GABAérgicas (A) Degradación del GABA Célula glial Glucosa Terminación presináptica Na+ GAT Glutamato COO< + NH3 CH CH2 CH2 Na+ COO< Ácido glutámico descarboxilasa + fosfato de piridoxal GABA + H3N CH2 CH2 CH2 COO< GABA VIAAT Célula postsináptica Receptores del GABA (B) Terminación presináptica Glucosa Transportador de glicina Célula glial Na+ Serina + H3N H C COO< COOH Na+ Serina hidroximetiltransferasa Glicina + H3 N H C COO< H Glicina Célula postsináptica VIAAT Receptores de glicina FIGURA 6.8. Síntesis, liberación y recaptación de los neurotransmisores inhibidores del GABA y de la glicina. (A) El GABA es sintetizado a partir de glutamato por la enzima ácido glutámico descarboxilasa (GAT), la cual requiere fosfato de piridoxal. (B) La glicina puede ser sintetizada por algunas vías metabólicas; en el encéfalo, la principal precursora es la serina. Los transportadores de alta afinidad terminan las acciones de estos transmisores y devuelven GABA o glicina hacia las terminaciones sinápticas para reutilizarlos; ambos transmisores son cargados en las vesículas sinápticas mediante el transportador vesicular de aminoácidos inhibidores (VIATT). NEUROTRANSMISORES Y SUS RECEPTORES 123 en el sistema nervioso central. La inhibición de la degradación del GABA produce un aumento del contenido tisular de GABA y un incremento de la actividad inhibidora sináptica. Las sinapsis GABAérgicas emplean tres tipos de receptores postsinápticos denominados GABAA, GABAB y GABAC. Los GABAA y GABAC son receptores ionotrópicos, mientras que el GABAB es metabotrópico. Al igual que otros receptores ionotrópicos, GABAA Y GABAC son pentámeros reunidos a partir de una combinación de muchos tipos de subunidades (véase Fig. 6.3F). Como consecuencia de esta diversidad de subunidades, la composición y la función de los receptores del GABAA difieren mucho entre los tipos neuronales. Aunque aún no se ha resuelto la estructura de los receptores del GABAA, es probable que su estructura sea similar a los nAChR (véase la Figura 6.3). Los receptores ionotrópicos del GABA son canales iónicos con puerta de GABA, y el Cl- es el principal ión permeable bajo condiciones fisiológicas (Figura 6.9A). El potencial de reversión para el Cl- es más negativo que el umbral para la descarga neuronal (véase Fig. 5.21) debido a la acción del cotransportador K+/Cl- (véase Fig. 4.9F), que mantiene baja la concentración intracelular de Cl-. Por lo tanto, la activación de los receptores GABAérgicos produce el influjo de Cl- con carga negativa que inhibe a las células postsinápticas (Figura 6.9B). En los casos en que la concentración postsináptica de Cl- es elevada –por ejemplo, en las neuronas en desarrollo–, los receptores del GABAA pueden excitar a sus puntos diana postsinápticos (Recuadro 6D). Los fármacos que actúan como agonistas o moduladores de los receptores del GABA, como las benzodiacepinas y los barbitúricos, se utilizan clínicamente para el tratamiento de la epilepsia y son sedantes y anestésicos eficaces. Todos los sitios de unión para el GABA, los barbitúricos, los corticosteroides y la picrotoxina se encuentran en el interior del dominio del poro del canal (Figura 6.9B). Otro sitio, llamado “sitio de fijación de las benzodiacepinas”, se ubica en el exterior del poro y modula la actividad del canal. Las benzodiacepinas, como diazepam (Valium) y clordiazepóxido (Librium) son fármacos que reducen la ansiedad y aumentan la transmisión GABAérgica al unirse a las subunidades α y β de algunos receptores GABAérgicos y se utilizan, desde el punto de vista terapéutico, para la anestesia y el control de la epilepsia. Otra sustancia que puede alterar la actividad de los circuitos inhibidores mediados por el GABA es el alcohol; al menos algunos aspectos del comportamiento del bebedor son causados por las alteraciones mediadas por el alcohol sobre los receptores ionotrópicos del GABA. Los receptores metabotrópicos del GABAB también se encuentran distribuidos de modo amplio en el encéfalo. Al igual que los receptores ionotrópicos del GABA, los receptores del GABAB son inhibidores. Sin embargo, en lugar de activar canales selectivos para Cl-, la inhibición mediada por el GABAB se debe, a menudo, a la activación de los canales del K+. Un segundo mecanismo para la inhibición mediada por el GABAB es el bloqueo de los canales del Ca2+, que también hiperpolariza las células postsinápticas. Se cree que la estructura de los receptores del GABAB es similar a la de los otros receptores metabotrópicos, aunque los receptores del GABAB parecen reunirse como heterodímeros de subunidades B1 y B2. La distribución del aminoácido neutro glicina en el sistema nervioso central es más localizada que la del GABA. Aproximadamente el 50% de las sinapsis inhibidoras en la médula espinal utiliza glicina; la mayor parte de las otras sinapsis inhibidoras (A) (B) Benzodiazepina Subunidad _ Poro del canal Potencial de membrana (mV) GABA 0 Estímulo sobre la neurona presináptica –50 Barbitúricos 0 50 100 150 200 250 Tiempo (ms) 300 350 400 FIGURA 6.9 Receptores ionotrópicos del GABA. (A) Los receptores ioCorticosteroides Picrotoxina Iones cloruro notrópicos del GABA contienen dos sitios de fijación para el GABA y muchos sitios que se unen a los fármacos y modulan estos receptores. Los iones cloruro (esferas verdes) fluyen a través del poro formado en el centro del receptor; dependiendo del gradiente electroquímico del cloruro, el flujo neto de cloruro puede dirigirse hacia el interior o el exterior de la célula. (B) La estimulación de una interneurona GABAérgica presináptica, en el momento indicado por la flecha, produce una inhibición transitoria del disparo del potencial de acción en su blanco postsináptico. Esta respuesta inhibidora es causada por la activación de los receptores postsinápticos del GABAA. (B, de Chavas y Marty, 2003.) 124 CAPÍTULO 6 RECUADRO 6D Acciones excitadoras del GABA en el encéfalo en desarrollo Aunque el GABA normalmente funciona como un neurotransmisor excitador en el encéfalo maduro, en el encéfalo en desarrollo excita a sus células diana. Esta inversión notable de la acción surge de los cambios evolutivos en la homeostasis del Cl- intracelular. Los mecanismos involucrados en este cambio fueron estudiados más extensamente en neuronas corticales. En las neuronas jóvenes, la concentración intracelular de Cl- está controlada principalmente por el cotransportador de Na+/K+/Cl-, que bombea cloro hacia el interior de las neuronas y produce una concentración intracelular elevada de Cl− (Figura A, izquierda). A medida que las neuronas siguen desarrollándose, comienzan a expresar el cotransportador K+/ Cl-, que bombea Cl- fuera de las neuronas y desciende su concentración intracelular (Figura A, derecha). Estos desplazamientos en la homeostasis del Cl- pueden hacer que su concentración intracelular caiga varias veces en la primera o dos primeras semanas de desarrollo posnatal (Figura B). Como los receptores ionotrópicos del GABA son canales impermeables al Cl-, el flujo de iones a través de estos receptores varía según la fuerza impulsora electromagnética sobre el Cl-. En las neuronas jóvenes, donde la concentración intracelular de Cles alta, el ECl es más positivo que el potencial de reposo. En consecuencia, el GABA despolariza estas neuronas. Además, el ECl a menudo es más positivo que el umbral, de modo que el GABA puede excitar estas neuronas para que disparen potenciales de acción (Figura C). Como se describe en el texto, la menor concentración intracelular de Cl- de las neuronas maduras hace que el ECl sea más negativo que el umbral del potencial de acción (y, a menudo, más negativo que el potencial de reposo), lo que conduce a respuestas inhibidoras al ácido γ-aminobutírico. ¿Por qué sufre el GABA este cambio en sus acciones postsinápticas? Aunque la lógica de este fenómeno todavía no es completamente clara, al parecer las respuestas despolarizantes al GABA producen actividad eléctrica que controla la proliferación, migración, crecimiento y maduración de las neuronas, y determina la conectividad sináptica. Una vez que estos procesos del desarrollo completaron, el circuito nervio- (A) El cambio en el desarrollo de la expresión de los transportadores del Cl- reduce su concentración intracelular, y revierte así la dirección del flujo de Cl- a través de los receptores del GABA. (B) Las imágenes de la concentración intracelular de cloruro entre los días 5 y 20 después del nacimiento (izquierda) demuestran una reducción progresiva de la concentración intracelular de Cl- (derecha). (C) Los cambios del desarrollo en la concentración intracelular de cloruro hacen que las respuestas del GABA se desplacen desde la despolarización en las neuronas jóvenes (6 días de vida) (izquierda) hacia la hiperpolarización en las neuronas más viejas (10 días de vida) (derecha) cultivadas a partir de la médula espinal del pollo. (B, de Berglund y cols., 2006; C, de Obata y cols., 1978.) 2 Cl– Respuestas despolarizantes Receptor del GABA Exterior de la célula Respuestas hiperpolarizantes Cl– Cl– K+ Concentración elevada de [Cl–] Cl– Concentración baja de [Cl–] Interior de la célula Interior de la célula Bibliografía Berglund, K. and 8 others (2006) Imaging synaptic inhibition in transgenic mice expressing the chloride indicator, Clomeleon. Brain Cell Biol. 35: 207-228. Cherubini, E., J. L. Gaiarsa and Y. Ben-Ari (1991) GABA: An excitatory transmitter in early postnatal life. Trends Neurosci. 14: 515-519. Obata, K., M. Oide and H. Tanaka (1978) Excitatory and inhibitory actions of GABA and glycine on embryonic chick spinal neurons in culture. Brain Res. 144: 179-184. Owens, D. F. and A. R. Kriegstein (2002) Is there more to GABA than synaptic inhibition? Nature Rev. Neurosci. 3: 715-727. Payne, J. A., C. Rivera, J. Voipio and K. Kaila (2003) Cation-chloride co-transporters in neuronal communication, development and trauma. Trends Neurosci. 26: 199-206. Rivera, C. and 8 others (1999) The K+/Cl- cotransporter KCC, renders GABA hyperpolarizing during neuronal maturation. Nature 397: 251-255. Neurona madura Neurona inmadura Exterior de la célula Na+ K+ (C) Potencial de membrana (mV) (A) so resultante requiere la transmisión inhibidora que también puede ser provista por el ácido γ-aminobutírico. Se necesitan nuevas investigaciones para apreciar el significado completo de las acciones excitadoras del GABA, así como para comprender los mecanismos subyacentes a la expresión del cotransportador K+/Cl-, los cuales hacen finalizar la breve carrera del GABA como neurotransmisor excitador. 6 días –10 –30 –50 GABA –70 GABA 0 [Cl–] (mM) (B) 10 días 25 60 120 0 40 mM 20 15 – [Cl ] 10 5 0 0 0 5 10 15 20 25 Días posnatales 5 días 10 días 20 días 60 120 NEUROTRANSMISORES Y SUS RECEPTORES utiliza GABA. La glicina es sintetizada a partir de la serina por la isoforma mitocondrial de serina hidroximetiltransferasa (véase Fig. 6.8B) y es transportada hacia las vesículas sinápticas por medio del mismo transportador vesicular de aminoácidos inhibidores que carga GABA en las vesículas. Una vez liberada de la célula presináptica, la glicina es rápidamente eliminada de la hendidura sináptica por los transportadores de glicina en la COO< Tirosina CH2 CH + NH3 125 membrana plasmática. Las mutaciones en los genes que codifican algunas de estos transportadores conducen a hiperglicinemia, enfermedad neonatal devastadora caracterizada por letargia, convulsiones y retardo mental. Los receptores de la glicina también son canales del Cl- con puerta de ligando y su estructura general imita a la de los receptores del GABAA. Los receptores de glicina son pentámeros que consisten en mezclas de los cuatro tipos de subunidades α, junto con la subunidad β accesoria (véase Fig. 6.3F). La estricnina produce un bloqueo potente sobre los receptores de la glicina, lo cual puede explicar las propiedades tóxicas de ese alcaloide vegetal (véase Recuadro 6A). HO Aminas biógenas O2 Tirosina hidroxilasa COO< Dihidroxifenilalanina (DOPA) CH2 CH + NH3 H CH + NH3 H CH + NH3 H CH + NH2 HO OH CO2 DOPA descarboxilasa Dopamina CH2 HO OH O2 Dopamina-` hidroxilasa OH Noradrenalina CH HO OH RCH3 Feniletanolamina-Nmetiltransferasa R OH Adrenalina CH CH3 HO OH Los transmisores aminas biógenas regulan muchas funciones encefálicas y también se encuentran en estado activo en el sistema nervioso periférico. Como las aminas biógenas están implicadas en una gama tan amplia de comportamientos (que varían desde funciones homeostáticas centrales hasta fenómenos cognitivos como la atención), no es sorprendente que los defectos en la función de las aminas biógenas estén implicados en la mayoría de los trastornos psiquiátricos. La farmacología de las sinapsis aminérgicas tiene una importancia crítica para la psicoterapia, y los fármacos que afectan la síntesis, la unión a los receptores o el catabolismo de estos neurotransmisores son algunos de los agentes más importantes en el arsenal con que cuenta la farmacología moderna (Recuadro 6E). Muchas drogas de abuso también actúan sobre las vías de las aminas biógenas. Existen cinco aminas biógenas neurotransmisoras bien definidas: las tres catecolaminas –dopamina, noradrenalina (norepinefrina) y adrenalina (epinefrina)–, histamina y serotonina (véase la Fig. 6.1). Todas las catecolaminas (llamadas así porque comparten la porción catecol) derivan de un precursor común, el aminoácido tirosina (Figura 6.10). El primer paso en la síntesis de las catecolaminas es catalizado por la tirosina hidroxilasa en una reacción que requiere oxígeno como cosustrato y tetrahidrobiopterina como cofactor para sintetizar dihidroxifenilalanina (DOPA). La histamina y la serotonina son sintetizadas a través de otras vías, como se describe más adelante. • La dopamina está presente en varias regiones encefálicas (Figura 6.11A), aunque la principal área del encéfalo que contiene dopamina es el cuerpo estriado, que recibe las principales aferencias de la sustancia nigra y desempeña un papel esencial en la coordinación de los movimientos corporales. En la enfermedad de Parkinson, por ejemplo, las neuronas dopaminérgicas de la sustancia nigra degeneran, lo que conduce a una disfunción motora característica (véase Recuadro 18A). También se cree que la dopamina está involucrada en la motivación, la recompenFIGURA 6.10 Ruta biosintética para los neurotransmisores de las catecolaminas. El aminoácido tirosina es el precursor de las tres catecolaminas. El primer paso en esta vía de reacciones, catalizada por la tirosina hidroxilasa, es limitante de la velocidad. 126 CAPÍTULO 6 RECUADRO 6E Las aminas biógenas neurotransmisoras y los trastornos psiquiátricos La regulación de los neurotransmisores del tipo aminas biógenas está alterada en distintos trastornos psiquiátricos. En efecto, la mayoría de los agentes psicotrópicos (definidos como los fármacos que alteran el comportamiento, el estado de ánimo o la percepción) afecta de manera selectiva uno o más pasos en la síntesis, el empaquetamiento o la degradación de las aminas biógenas. Determinar cómo funcionan estos fármacos ha sido extremadamente útil para comenzar a comprender los mecanismos moleculares que subyacen a algunas de estas enfermedades. Sobre la base de sus efectos en los seres humanos, los agentes psicoterapéuticos pueden ser divididos en varias categorías amplias: antipsicóticos, ansiolíticos, antidepresivos y estimulantes. El primer agente antipsicótico utilizado para mejorar trastornos como la esquizofrenia fue la reserpina, que se desarrolló en la década de 1950 y fue utilizada inicialmente como agente antihipertensivo; la reserpina bloquea la captación de la noradrenalina en las vesículas sinápticas y, por lo tanto, produce la depleción del transmisor en las terminaciones aminérgicas y la disminución de la capacidad de la división simpática del sistema motor visceral para producir la vasoconstricción (véase el Cap. 21).A partir de un efecto colateral importante en los pacientes hipertensos tratados con reserpina, la depresión, sugirió la posibilidad de utilizarlo como agente antipsicótico en pacientes con agitación y ansiedad patológica (su capacidad para producir depresión en individuos mentalmente sanos también sugirió que los transmisores aminérgicos están involucrados en los trastornos del estado de ánimo; véase Recuadro 29E). Si bien la reserpina ya no se utiliza como agente antipsicótico, su éxito inicial estimuló el desarrollo de los agentes antipsicóticos como clorpromazina, haloperidol y benperidol, los cuales, en las últimas décadas, han cambiado radicalmente el enfoque del tratamiento contra los trastornos psicóticos. Antes del descubrimiento de estos fármacos, los pacientes psicóticos eran hospitalizados durante períodos prolongados y, a veces, indefinidamente, y, en la década de 1940, eran sometidos a medidas extremas como la lobotomía frontal (véase Recuadro 26C). Ahora, los agentes antipsicóticos modernos permiten que la mayoría de los pacientes sean tratados de forma ambulatoria después de una breve estancia hospitalaria. Es importante señalar que la eficacia clínica de estos fármacos se correlaciona con su capacidad para bloquear los receptores de la dopamina encefálicos, lo que implica que la activación de los receptores de la dopamina contribuye a ciertos tipos de enfermedad psicótica. Se siguen realizando muchos esfuerzos para desarrollar agentes antipsicóticos con menos efectos colaterales y para descubrir el mecanismo y el sitio de acción de estas medicaciones. La segunda categoría de fármacos psicoterapéuticos es la de los agentes ansiolíticos. Se estima que los trastornos de ansiedad afectan del 10 al 35% de la población, lo cual los convierte en los trastornos psiquiátricos más frecuentes. Las dos formas principales de ansiedad patológica –las crisis de angustia y el trastorno de ansiedad generalizada– responden a los fármacos que afectan la transmisión aminérgica. Los agentes utilizados para tratar los trastornos de angustia incluyen (1) los inhibidores de la enzima monoaminooxidasa (inhibidores de la MAO o IMAO) necesarios para el catabolismo de las aminas neurotransmisoras, y (2) los bloqueantes de los receptores de la serotonina. Los agentes más eficaces para el tratamiento contra el trastorno de la ansiedad generalizada han sido las benzodiacepinas, como clordiazepóxido (Librium®) y diazepam (Valium®). Al contrario de la mayoría de los otros agentes psicoterapéuticos, estos agentes aumentan la eficacia de la transmisión en las sinapsis de GABAA, en lugar de actuar en las sinapsis aminérgicas. Los antidepresivos y los estimulantes también afectan la transmisión aminérgica. Gran cantidad de fármacos se utilizan clínicamente para tratar los trastornos depresivos. Las tres clases principales de antidepresivos –inhibidores de la MAO, antidepresivos tricíclicos y bloqueantes de la recaptación de la serotonina, como fluoxetina (Prozac®) y trazodona– influyen en distintos aspectos de la transmisión aminérgica. Los inhibidores de la MAO, como la fenelzina, bloquean la degradación de las aminas, mientras que los antidepresivos tricíclicos, como desipramina, bloquean la recaptación de la noradrenalina y de otras aminas. El antidepresivo extraordinariamente popular fluoxetina (Prozac®) bloquea de manera selectiva la recaptación de las catecolaminas. También se han utilizado estimulantes como la anfetamina para tratar algunos trastornos depresivos. La anfetamina estimula la liberación de noradrenalina de las terminaciones nerviosas; el “vuelo” transitorio como resultado del consumo de anfetamina puede reflejar el opuesto emocional de la depresión que sigue a veces a la depleción de la noradrenalina inducida por la reserpina. A pesar de la cantidad relativamente pequeña de neuronas aminérgicas en el encéfalo, esta larga enumeración de las acciones farmacológicas destaca la importancia crítica de estas neuronas en el mantenimiento de la salud mental. Bibliografía Karam, C. S. and 8 others (2010) Signaling pathways in schizophrenia: Emerging targets and therapeutic strategies. Trends Pharmacol. Sri. 31: 381-390. Krishnan, V. and E. J. Nestler (2008). The molecular neurobiology of depression. Nature 457: 894-902. Margolis, R. L. (2009) Neuropsychiatric disorders: The choice of antipsychotics in schizophrenia. Nature Rev. Neurosci. 5:308-310. Ross, C.A., R. L. Margolis, S. A. Reading, M. Pletnkov and J.T. Coyle (2006). Neurobiology of schizophrenia. Neuron 52:139-153. Sharp, T. and P. J. Cowen (2011) 5-HT and depression: Is the glass half-full? Curr. Opin. Pharmacol. 11: 45-51. Shin, J. K., D. T. Malone, I. T. Crosby and B. Capuano (2011) Schizophrenia: A systematic review of the disease state, current therapeutics, and their molecular mechanisms of action. Curr. Med. Chem. 18: 1380-1404. NEUROTRANSMISORES Y SUS RECEPTORES (A) Dopamina Cuerpo calloso Corteza cerebral Al estriado Tálamo Cerebelo Sustancia nigra y área tegmental ventral Protuberancia Bulbo raquídeo (B) Noradrenalina A la médula espinal Cuerpo calloso Corteza cerebral Tálamo Cerebelo Locus cerúleo ProtubeA la médula rancia Bulbo espinal raquídeo (C) Adrenalina Cuerpo calloso Corteza cerebral Tálamo Hipotálamo Protuberancia Cerebelo Neuronas adrenérgicas bulbares Bulbo raquídeo A la médula espinal FIGURA 6.11 Distribución en el encéfalo humano de las neuronas que contienen los neurotransmisores de las catecolaminas. Se muestran las neuronas y sus proyecciones (flechas) que contienen los neurotransmisores de las catecolaminas. Las flechas curvas a lo largo del perímetro de la corteza indican la inervación de las regiones corticales laterales que no se muestran en este plano mediosagital de corte. 127 sa y el refuerzo, y muchas drogas de abuso funcionan afectando las sinapsis dopaminérgicas en el SNC (Recuadro 6F). Además de estas funciones en el SNC, la dopamina también desempeña otras funciones poco conocidas en algunos ganglios simpáticos. La dopamina es producida por la acción de la DOPA descarboxilasa sobre la DOPA (véase Fig. 6.10). Tras su síntesis en el citoplasma de las terminaciones presinápticas, la dopamina es almacenada en las vesículas sinápticas por medio de un transportador vesicular de monoaminas (vesicular monoamine transporter, VMAT). La acción de la dopamina en la hendidura sináptica concluye con la recaptación de dopamina en las terminaciones nerviosas o en las células gliales circundantes por un cotransportador de dopamina Na+-dependiente, denominado “DAT”. Al parecer, la cocaína produce sus efectos psicotrópicos uniéndose al DAT e inhibiéndolo, lo que arroja un aumento neto en la liberación de dopamina desde áreas encefálicas específicas. La anfetamina, otra droga adictiva, también inhibe el DAT y el transportador para noradrenalina (véase más adelante). Las dos enzimas principales involucradas en el catabolismo de la dopamina son la monoaminooxidasa (MAO) y la catecol O-metil-transferasa (COMT). Tanto las neuronas como la glía contienen MAO mitocondrial y COMT citoplasmática. Los inhibidores de estas enzimas, como fenelzina y tranilcipromina, son utilizados en la práctica clínica como antidepresivos (véase Recuadro 6E). Una vez liberada, la dopamina actúa exclusivamente activando los receptores acoplados a la proteína G. Uno de estos, el receptor dopaminérgico D3, se muestra en la Figura 6.12A. La estructura de este receptor se asemeja mucho a la de otros receptores metabotrópicos, como el receptor mACh (véase la Figura 6.4A), excepto porque su sitio de fijación al ligando es optimizado por la fijación a la dopamina. La mayoría de los subtipos de receptores de la dopamina (véase Fig. 6.4C) actúa activando o inhibiendo la adenililciclasa (véase el Cap. 7). En general, la activación de estos receptores contribuye a comportamientos complejos: por ejemplo, la administración de agonistas de los receptores de dopamina produce hiperactividad y comportamiento estereotipado repetitivo en animales de laboratorio. La activación de otro tipo de receptor de dopamina en el bulbo raquídeo inhibe los vómitos. Por lo tanto, los antagonistas de estos receptores se utilizan como eméticos para inducir el vómito después del envenenamiento o de la sobredosis de una droga. Los antagonistas de los receptores de dopamina también pueden producir catalepsia, un estado en el cual es difícil iniciar los movimientos motores voluntarios, lo que sugiere una base para esta característica presente en algunas psicosis. • La noradrenalina (también llamada “norepinefrina”) es utilizada como neurotransmisora en el locus cerúleo, un núcleo del tronco del encéfalo que se proyecta de forma difusa a distintos puntos diana o blanco del encéfalo anterior (Figura 6.11B) e influye en el sueño y la vigilia, en la atención y en la conducta alimentaria. Tal vez las neuronas noradrenérgicas más sobresalientes sean las células ganglionares simpáticas, que emplean 128 CAPÍTULO 6 RECUADRO 6F Adicción La adicción a las drogas es una enfermedad crónica y recurrente con consecuencias médicas, sociales y políticas evidentes. La adicción (también denominada “farmacodependencia”) es un trastorno persistente de la función encefálica en el cual se desarrolla un consumo compulsivo de drogas a pesar de las serias consecuencias negativas para el individuo afectado. El manual diagnóstico de la American Psychiatric Association define “adicción” en términos de dependencia física y de dependencia psicológica (en la que un individuo persiste en el consumo de drogas a pesar de las consecuencias adversas). La gama de sustancias que pueden generar este tipo de dependencia es amplia; los agentes primarios de abuso actualmente son opioides, cocaína, anfetaminas, marihuana, alcohol y nicotina. La adicción a agentes más “socialmente aceptables”, como el alcohol y la nicotina, a veces se considera menos problemática; pero, de hecho, involucran consecuencias médicas y conductuales tan importantes como las de las drogas de abuso consideradas más peligrosas. Es importante destacar que el fenómeno de la adicción no está limitado al comportamiento humano, sino que se puede demostrar en animales de laboratorio. La mayor parte de estos mismos agentes es autoadministrada si se brinda a los primates, roedores u otras especies la oportunidad de hacerlo. Además de la compulsión para obtener el agente de abuso, una característica importante de la adicción a muchas drogas es la aparición de una constelación de características fisiológicas y emocionales negativas –denominadas, en general, “síndrome de abstinencia”– cuando no se ingiere la droga. Los signos y síntomas de la abstinencia son diferentes para cada agente de abuso, pero, en general, se caracterizan por aspectos opuestos a los de la experiencia positiva inducida por la droga propiamente dicha. Consideremos, por ejemplo, la cocaína, droga que se estima era consumida regularmente por 5 a 6 millones de estadounidenses durante la década de 1990 con unos 600 000 consumidores regulares adictos o en alto riesgo de adicción. El efecto positivo de la droga fumada o inhalada como polvo en forma de base libre de alcaloide es un “vuelo” casi inmediato, pero que generalmente dura unos minutos y conduce a un deseo de más droga tan solo 10 minutos a media hora después. El “vuelo” es descrito como una sensación de bienestar, autoconfianza y satisfacción. Por el contrario, cuando no cuentan con la droga, los consumidores habituales experimentan depresión, somnolencia, fatiga, un deseo irresistible de consumo y una sensación general de malestar. Otro aspecto de la adición a la cocaína o a otros agentes es la tolerancia, definida como una reducción en la respuesta a la droga con la administración repetida. La tolerancia se desarrolla como consecuencia del consumo persistente de algunas drogas, pero es particularmente importante en los casos de adicción, ya que la dosis necesaria para experimentar el efecto deseado debe aumentarse de manera progresiva. Aunque es correcto decir que la neurobiología de la adicción no se conoce de forma completa, para la cocaína y muchos otros agentes de abuso, los efectos adictivos involucran la activación de los receptores de la dopamina en regiones encefálicas críticas que participan en la motivación y en el refuerzo condicional (véase Cap. 29). Las más importantes de estas áreas son el sistema dopaminérgico del mesencéfalo, especialmente, sus proyecciones desde el área ventrotegmental hacia el núcleo accumbens. Agentes como la cocaína parecen actuar elevando las concentraciones de dopamina en estas áreas, lo que hace que este transmisor se encuentre más disponible para los receptores al interferir en la recaptación de la dopamina liberada en la sinapsis mediante el transportador de dopamina. Se cree que el refuerzo y la motivación de los comportamientos de consumo de la droga están relacionados con las proyecciones hacia el núcleo accumbens. La droga opioide de uso más frecuente es la heroína. La heroína es un derivado de la amapola del opio y no se encuentra legalmente disponible para fines clínicos en los Estados Unidos de América. Se estima que el número de adictos a la heroína en los Estados Unidos es entre 750 000 y un millón de individuos. Las sensaciones positivas producidas por la heroína, descritas en general como “el rush”, a menudo se comparan con la sensación del orgasmo sexual y comienzan en menos de un minuto después de la inyección intravenosa. Luego hay una sensación de bienestar general (“on the nod”, estar volando) que dura alrededor de una hora. Los síntomas de abstinencia pueden ser intensos: inquietud, irritabilidad, náuseas, dolor muscular, depresión, insomnio, y sensación de ansiedad y de malestar. Los aspectos reforzadores de la droga se relacionan con el mismo circuito dopaminérgico en el área ventrotegmental y en el núcleo accumbens que el de la cocaína; aunque, por cierto, participan otras áreas, sobre todo los sitios de receptores opioides descritos en el Capítulo 10. Es interesante destacar que la adicción a la heroína o a cualquier otro agente no es una consecuencia inevitable del consumo de la droga, sino que depende fundamentalmente del entorno. Por ejemplo, los veteranos de Vietnam que se habían vuelto adictos a la heroína allí, en general, perdieron su adicción al volver a los Estados Unidos. Asimismo, los pacientes que reciben otros opioides (p. ej., morfina) contra afecciones dolorosas pocas veces se vuelven adictos. El tratamiento contra cualquier forma de adicción es difícil y debe ajustarse a las circunstancias del individuo. Además de tratar los problemas agudos de la abstinencia y la “destoxificación ” deben modificarse los patrones de comportamiento, lo cual puede llevar meses o años. La adicción es, por lo tanto, un estado patológico crónico que requiere una monitorización continua de los individuos susceptibles durante toda la vida. Bibliografía American Psychiatric Association (2000) Diagnostic and Statistical Manual of Mental Disorders, 4th Edition (DSM IV). Washington, D.C. Hyman, S. E. and R. C. Malenka (2001) Addiction and the brain: The neurobiology of compulsion and its persistence. Nature Rev. Neurosci. 2: 695-703. Kauer, J. S. and R. C. Malenka (2007) Synaptic plasticity and addiction. Nature Rev. Neurosci. 8:844-858. Laakso, A., A. R. Mohn, R. R. Gainetdinov and M. G. Caron (2002) Experimental genetic approaches to addiction. Neuron 36: 213-228. Madras, B. K., C. M. Colvis, J. D. Pollock, J. L. Rutter, D. Shurtleff and M. von Zastrow (2006) Cell Biology of Addiction. Cold Spring Harbor: Cold Spring Harbor Laboratory Press. O‘Brien, C. P. (2006) Goodman and Gilman’s The Pharmaceutical Basis of Therapeutics, 11th Edition. New York: McGraw-Hill, Chapter 23, pp. 607-627. Schultz, W. (2010) Dopamine signals for reward value and risk: Basic and recent data. Behav. Brain Funct. 6: 24. NEUROTRANSMISORES Y SUS RECEPTORES FIGURA 6.12 Receptores metabotrópicos para los neurotransmisores de las catecolaminas. (A) Estructura del receptor de la dopamina D3. Al igual que todos los receptores metabotrópicos, el receptor D3 atraviesa 7 veces la membrana plasmática y tiene un dominio citoplasmático que se une a las proteínas G y las activa, así como un dominio extracelular que se une a la dopamina. (B) Estructura del receptor adrenérgico β2 y de su proteína G asociada. (Izquierda) En ausencia de ligando, el dominio citoplasmático del receptor β2 no está unido a la proteína G (subunidades α, β y γ). (Derecha) La unión del ligando (agonista β indicado por las (A) 129 (B) Sitio de fijación de la dopamina Sitio de fijación de noradrenalina/adrenalina Inactivo Activo Extracelular Membrana plasmática Citoplasma _ Sitio de fijación de la proteína G esferas de colores) al sitio de fijación extracelular para noradrenalina (NE) y adrenalina (Epi) hace que el receptor β2 se una a la subunidad α de la proteína G, lo que a su vez induce un cambio espectacular en la estructura de esta subunidad. ` a ` a _ (A, de Chien y cols., 2010; B, de Wall y cols., 1995, Rasmussen y cols., 2007 y Rasmussen y cols., 2011.) noradrenalina como principal transmisor periférico en esta división del sistema motor visceral (véase Cap. 21). La síntesis de noradrenalina requiere dopamina β-hidroxilasa, la cual cataliza la producción de noradrenalina a partir de la dopamina (véase Fig. 6.10). La noradrenalina es almacenada luego en las vesículas sinápticas mediante el mismo VMAT que participa en el transporte vesicular de la dopamina. La noradrenalina es eliminada de la hendidura sináptica por el transportador de noradrenalina (norepinephrine transporter, NET), un cotransportador Na+-dependiente que también es capaz de captar dopamina. Como mencionamos antes, el NET sirve como punto diana molecular de la anfetamina, la cual actúa como estimulante al producir un aumento neto en la liberación de noradrenalina y de dopamina. Una mutación en el gen NET es una causa de intolerancia al ortostatismo, trastorno que produce mareos al incorporarse. Al igual que la dopamina, la noradrenalina es degradada por la MAO y la COMT. La noradrenalina, al igual que la adrenalina, actúa sobre los receptores α- y β-adrenérgicos (véase Fig. 6.4C). Ambos tipos de receptores están acoplados a la proteína G; de hecho, el receptor β-adrenérgico fue el primer receptor metabotrópico de neurotransmisores que se ha identificado. Como se muestra en la Figura 6.12B, la estructura de este receptor es muy similar a la de otros receptores metabotrópicos (como el receptor de la dopamina de la Figura 6.12A). La fijación de noradrenalina o de adrenalina produce pequeños cambios en la estructura de este receptor, lo que permite la unión de la proteína G (Figura 6.12, derecha). Esto, a su vez, produce cambios mayores en la forma de la subunidad α de la proteína G, el primer paso en una serie de reacciones que permiten a la proteína G regular las cascadas de señalización intracelular (véase Cap. 7). Actualmente, se han identificado dos subclases de receptores α-adrenérgicos. En general, la activación de los receptores α1 produce una lenta despolarización relacionada con la inhibición de los canales del K+, mientras que la activación de los receptores α2 produce una lenta hiperpolarización debido a la activación de un tipo diferente de canal del K+. Existen tres subtipos de receptores β-adrenérgicos, dos de los cuales se expresan en muchos tipos de neuronas. Los agonistas y los antagonistas de los receptores adrenérgicos, como el betabloqueante propranolol (Inderol®), son utilizados en la clínica para distintos trastornos que varían desde arritmias cardíacas hasta cefaleas migrañosas. Sin embargo, la mayor parte de las acciones de estos fármacos recae sobre los receptores del músculo liso, sobre todo, en los aparatos cardiovascular y respiratorio (véase Cap. 21). • La adrenalina (también denominada “epinefrina”) se halla en el encéfalo en niveles mucho menores que cualquier otra catecolamina y también se presenta en menos neuronas encefálicas que otras catecolaminas. Las neuronas del sistema nervioso central que contienen adrenalina están principalmente en el sistema tegmental lateral y en el bulbo raquídeo, y proyectan hacia el hipotálamo y el tálamo (Figura 6.11C). No se conoce la función de estas neuronas que secretan adrenalina. La enzima que sintetiza adrenalina, la feniletanolamina-N-metiltransferasa (véase la Figura 6.10), solo está presente en neuronas que secretan adrenalina. Por otra parte, el metabolismo de la adrenalina es muy similar al de la noradrenalina. La adrenalina es almacenada en vesículas a través del transportador vesicular de monoaminas. No se identificó ningún transportador de la membrana plasmática específico para adrenalina, aunque el NET es capaz de transportarla. Como ya mencionamos, la adrenalina actúa tanto sobre los receptores α-adrenérgicos como los β-adrenérgicos. 130 CAPÍTULO 6 (A) Histamina (B) Serotonina Cuerpo calloso Corteza cerebral Tálamo Cerebelo Núcleo tuberomamilar del hipotálamo Cuerpo calloso Corteza cerebral Protuberancia Bulbo raquídeo A la médula espinal Tálamo Cerebelo ProtubeA la médula Núcleos del rafe rancia Bulbo espinal raquídeo FIGURA 6.13 Distribución en el encéfalo humano de las neuronas que contienen histamina o serotonina. Los diagramas muestran la distribución de neuronas y sus proyecciones (flechas) que contienen histamina (A) o serotonina (B). Las flechas curvas a lo largo del perímetro de la corteza indican la inervación de las regiones corticales laterales que no se muestran en este plano mediosagital de corte. • La histamina se encuentra en las neuronas del hipotálamo que envían proyecciones escasas pero difusas a casi todas las regiones del encéfalo y de la médula espinal (Figura 6.13A). Las proyecciones histamínicas centrales median el despertar y la atención, de modo similar a las proyecciones colinérgicas y noradrenérgicas centrales. La histamina también controla la reactividad del sistema vestibular. Las reacciones alérgicas o el daño tisular producen liberación de histamina de los mastocitos en el torrente sanguíneo. La estrecha proximidad de los mastocitos a los vasos sanguíneos junto con las acciones potentes de la histamina sobre los vasos sanguíneos también plantean la posibilidad de que la histamina pueda influir en el flujo sanguíneo encefálico. La histamina es producida a partir del aminoácido histidina por una histidina descarboxilasa (Figura 6.14A) y es transportada en vesículas mediante el mismo VMAT que las catecolaminas. No se identificó aún ningún transportador de la membrana plasmática para la histamina. La histamina es degradada por las acciones combinadas de la histamina metiltransferasa y la monoaminooxidasa. Los tres tipos conocidos de receptores de la histamina son todos receptores metabotrópicos (véase Fig. 6.4C). Debido a la importancia de los receptores de la histamina en la mediación de las respuestas alérgicas, se desarrollaron muchos antagonistas de los receptores de la histamina como agentes antihistamínicos. Los antihistamínicos que atraviesan la barrera hematoencefálica, como la difenhidramina (Benadryl®), actúan como sedantes al interferir en las funciones de la histamina sobre el despertar del sistema nervioso central. Los antagonistas del receptor H1 también se utilizan para prevenir la enfermedad del movimiento, tal vez debido al papel de la histamina para controlar la función vestibular. Los receptores H2 controlan la secreción de ácido gástrico en el siste- ma digestivo, lo que permite que los antagonistas de este receptor sean utilizados en el tratamiento contra distintos trastornos gastrointestinales (p. ej., úlceras pépticas). • La serotonina, o 5-hidroxitriptamina (5-HT), fue inicialmente considerada una sustancia que aumentaba el tono vascular en virtud de su presencia en el suero (de ahí el nombre de “serotonina”). La serotonina se encuentra fundamentalmente en grupos de neuronas en la región del rafe de la protuberancia y del tronco del encéfalo superior, las cuales tienen amplias proyecciones con el encéfalo anterior (Figura 6.13B) y regulan el sueño y la vigilia (véase Cap. 28). La 5-HT ocupa un lugar prominente en la neurofarmacología porque gran cantidad de agentes antipsicóticos que son útiles en el tratamiento contra la depresión y la ansiedad actúan sobre las vías serotoninérgicas (véase Recuadro 6E). La 5-HT es sintetizada a partir del aminoácido triptófano, el cual es un requerimiento esencial de la dieta. El triptófano es captado en las neuronas por un transportador de la membrana plasmática e hidroxilado en una reacción catalizada por la enzima triptófano-5-hidroxilasa (Figura 6.14B), el paso limitante de la velocidad para la síntesis de 5-hidroxitriptamina. El almacenamiento de 5-HT en las vesículas sinápticas es realizada por el VMAT, que también es responsable del almacén de otras monoaminas en las vesículas sinápticas. Los efectos sinápticos de la serotonina concluyen con el transporte retrógrado hacia las terminaciones nerviosas a través de un transportador específico de serotonina (SERT). Muchos agentes antidepresivos son inhibidores selectivos de la recaptación de serotonina (ISRS) que inhiben el transporte de 5-HT mediante el transportador específico de serotonina. Tal vez el ejemplo mejor conocido del ISRS sea la fluoxetina (Prozac®) (véase Recuadro 6E). La vía catabólica primaria para 5-HT está mediada por la monoaminooxidasa Se han identificado gran cantidad de receptores serotoninérgicos. La mayoría de estos receptores son metabotrópicos (véase Fig. 6.4C). Estos fueron relacionados con determinados comportamientos, que incluyen las emociones, el ritmo circadiano, la conducta motora y el estado de alerta mental. El deterioro de la función de estos receptores se considera responsable de mu- NEUROTRANSMISORES Y SUS RECEPTORES (A) COO< Histidina CH2 HN CH + NH3 N Histidina descarboxilasa CH2 HN FIGURA 6.14 Síntesis de histamina y de serotonina. (A) La histamina es sintetizada a partir del aminoácido histidina. (B) La serotonina deriva del aminoácido triptófano por un proceso de dos pasos que necesita las enzimas triptófano-5-hidroxilasa y una descarboxilasa. CO2 Histamina H CH + NH3 COO< Triptófano CH2 CH + NH3 N O2 Triptófano 5-hidroxilasa COO< 5-Hidroxitriptófano CH2 CH + NH3 H CH + NH3 N L-aminoácidos aromáticos descarboxilasa CO2 Serotonina (5-hidroxitriptamina) HO diana para una amplia variedad de agentes terapéuticos que incluyen ondansetrón (Zofran®) y granisetrón (Kytril®), utilizados en la prevención de náuseas posoperatorias y emesis inducida por quimioterapia. El ATP y otras purinas N (B) HO 131 CH2 N chos trastornos psiquiátricos, como depresión, trastornos de ansiedad y esquizofrenia (véase el Capítulo 29) y los fármacos que actúan sobre los receptores serotoninérgicos constituyen tratamientos eficaces para algunos de estos trastornos. La activación de los receptores de la serotonina también media la saciedad y el menor consumo de alimento, razón por la cual algunos agentes a veces son útiles para el tratamiento de los trastornos de la conducta alimentaria. Solo un grupo de receptores de la serotonina, denominados “receptores 5-HT3”, son canales iónicos con puerta de ligando (véase la Figura 6.3F). Se trata de canales catiónicos no selectivos y, por lo tanto, median respuestas postsinápticas excitadoras. Su estructura general, con canales funcionales formados por la unión de múltiples subunidades, es similar a la de otros receptores ionotrópicos descritos en el capítulo. Se conocen dos tipos de subunidades 5-HT3, y forman canales funcionales al unirse como un heteromultímero. Los receptores serotoninérgicos son puntos Todas las vesículas sinápticas contienen ATP, que es liberado junto con uno o más neurotransmisores “clásicos”. Esta observación plantea la posibilidad de que el ATP actúe como cotransmisor. Desde la década de 1920, se sabe que la aplicación extracelular de ATP (o sus productos de degradación AMP y adenosina) pueden producir respuestas eléctricas en las neuronas. La idea de que algunas “purinas” (denominadas así debido al anillo purínico; véase Fig. 6.1) también son neurotransmisoras ha recibido ahora considerable respaldo experimental. El ATP actúa como neurotransmisor excitador en las neuronas motoras de la médula espinal, y en los ganglios sensitivos y autónomos. También se demostraron acciones postsinápticas del ATP en el sistema nervioso central, específicamente, en las neuronas del asta dorsal y en un subgrupo de neuronas del hipocampo. Las enzimas extracelulares degradan el ATP liberado a adenosina, que posee su propio conjunto de acciones de señalización. Por lo tanto, la adenosina no puede ser considerada un neurotransmisor clásico porque no es almacenada en vesículas sinápticas ni liberado en la forma de Ca2+-dependiente. Algunas enzimas, como la apirasa y la ecto-5’ nucleotidasa, así como los transportadores de nucleósidos participan en el rápido catabolismo y en la eliminación de las purinas de las localizaciones extracelulares. Los receptores de ATP y adenosina se encuentran ampliamente distribuidos en el sistema nervioso y en muchos otros tejidos. Actualmente, se conocen tres clases de estos receptores purinérgicos. Una de estas clases consiste en receptores ionotrópicos denominados receptores P2X (véase Fig. 6.3F). La estructura de estos receptores es algo singular entre los receptores ionotrópicos porque cada subunidad tiene solo dos dominios transmembrana (Figura 6.15A). Además, solo se requieren tres de estas subunidades para formar un receptor trimérico (Figura 6.15B). Al igual que todos los receptores ionotrópicos, un poro se localiza en el centro del receptor P2X y forma un canal catiónico no selectivo. Por lo tanto, los receptores P2X median respuestas postsinápticas excitadoras. Los receptores purinérgicos ionotrópicos se encuentran ampliamente distribuidos en las neuronas centrales y periféricas. En los nervios sensitivos, desempeñan evidentemente un papel en la mecanosensibilidad y en el dolor; sin embargo, no se conoce su función en la mayoría de las otras células. Las otras dos clases de receptores purinérgicos son receptores metabotrópicos acoplados a la proteína G (véase Fig. 6.4C). Las dos clases difieren en su sensibilidad de los agonistas: un tipo 132 CAPÍTULO 6 (A) (C) (B) Cuerpo Cabeza Aleta derecha Sitio de fijación de ATP Aleta izquierda Poro Aleta dorsal Extracelular Uña Membrana plasmática Citoplasma N C (D) Sitio de fijación de la adenosina Extracelular FIGURA 6.15 Receptores purinérgicos. (A) Subunidad de un receptor ionotrópico P2X4. Cada subunidad tiene un dominio transmembrana que consiste en dos estructuras helicoidales que forman parte de un canal, así como un dominio extracelular grande que incluye el sitio de fijación del ATP. La forma de la subunidad recuerda a un delfín, con las estructuras codificadas por color como se indica en el recuadro. (B) Vista lateral de un receptor P2X4; este receptor es un trímero de tres subunidades, y cada subunidad se Membrana plasmática muestra con un color diferente. Se propone que el sitio de fijación del ATP está en el centro del dominio extracelular. (C) Vista superior del receptor P2X4, que indica el poro del canal de localización central. (D) Estructura de un receptor metabotrópico A2A de adenosina. Este receptor tiene la estructura de dominio que atraviesa 7 veces la membrana característica Citoplasma Sitio de fijación de la proteína G es estimulado preferencialmente por la adenosina, mientras que el otro es activado preferencialmente por el ATP. En la Figura 6.15D, se muestra un ejemplo del primero: el receptor de la adenosina A2A. Ambos tipos de receptores se encuentran en todo el encéfalo, y en los tejidos periféricos como el del corazón, del tejido adiposo y del riñón. Las xantinas como la cafeína y la teofilina bloquean los receptores de la adenosina, y se considera que esta actividad es responsable de los efectos estimulantes de estos agentes. Neurotransmisores peptídicos Muchos péptidos conocidos como hormonas también actúan como neurotransmisores. Algunos transmisores peptídicos han sido involucrados en la modulación de las emociones (véase Cap. 29). Otros, como la sustancia P y los péptidos opioides, participan en la percepción del dolor (véase Cap. 10). Otros péptidos, como las hormonas melanocitoestimulante, adreno- de los receptores metabotrópicos y se muestra con un agente antagonista ocupando el sitio de fijación de la adenosina. (A-C, de Kawate y cols., 2009; D, de Jaakola e Ijzerman, 2010.) corticotrofina y β-endorfina, regulan respuestas complejas al estrés. Los mecanismos responsables de la síntesis y del empaquetamiento de los transmisores peptídicos son fundamentalmente diferentes de los utilizados para los neurotransmisores de molécula pequeña y son muy similares a la síntesis de proteínas secretadas desde células distintas de las neuronas (enzimas pancreáticas, por ejemplo). Las neuronas que secretan péptidos generalmente sintetizan polipéptidos que son mucho más grandes que el péptido “maduro” final. El procesamiento de estos polipéptidos, que se denominan prepropéptidos (o preproproteínas), tiene lugar por una secuencia de reacciones en varios orgánulos intracelulares. Los prepropéptidos son sintetizados en el retículo endoplasmático rugoso, donde se elimina la secuencia señal de aminoácidos, es decir, la secuencia que indica que el péptido va a ser secretado. El polipéptido restante, denominado propéptido (o proproteína), atraviesa luego el aparato de Golgi y es empaquetado en vesículas en la red trans-Golgi. NEUROTRANSMISORES Y SUS RECEPTORES (A) Secuencia señal FIGURA 6.16 Procesamiento proteolítico de los prepropéptidos. Se muestra acá la preproo- Preproopiomelanocortina Prepropéptido piomelanocortina (A) y la preproencefalina A (B). Por cada prepropéptido, la secuencia señal está indicada en color naranja a la izquierda; las localizaciones de los productos peptídicos activos están indicadas por diferentes colores. La maduración de Proopiomelanocortina Propéptido ACTH 133 `-lipotropina Péptido activo los prepropéptidos comprende la división de la secuencia señal y otro procesamiento proteolítico. Este procesamiento puede producir algunos péptidos neuroactivos diferentes como ACTH, γ-lipotropina a-lipotropina `-endorfina Péptidos activos (B) Secuencia señal y β-endorfina (A) o múltiples copias del mismo péptido, como met-encefalina (B). Pre-proencefalina A Prepropéptido Proencefalina A Propéptido Péptidos activos Met-encefalina Met-encefalina Leu-encefalina Los estadios finales del procesamiento de los neurotransmisores peptídicos se desarrollan después del empaquetamiento en vesículas y comprenden la división proteolítica, la modificación de los extremos del péptido, la glucosilación, la fosforilación y la formación de enlaces disulfuro. Los precursores propéptidos son típicamente más grandes que sus productos peptídicos activos y pueden dar origen a más de una especie de neuropéptido (Figura 6-16). Eso significa que se pueden liberar múltiples péptidos neuroactivos de una única vesícula. Además, los neuropéptidos a menudo son liberados de forma conjunta con los neurotransmisores de molécula pequeña. Por lo tanto, las sinapsis peptidérgicas a menudo producen respuestas postsinápticas complejas. Los péptidos son catabolizados en fragmentos de aminoácidos inactivos por enzimas llamadas “peptidasas”, que habitualmente se localizan sobre la superficie extracelular de la membrana plasmática. La actividad biológica de los neurotransmisores peptídicos depende de su secuencia de aminoácidos (Figura 6.17). Sobre la base de sus secuencias de aminoácidos, los transmisores neuropéptidos han sido agrupados de manera no muy estricta en cinco categorías: péptidos de encéfalo/intestino, péptidos opioides, péptidos hipofisarios, hormonas liberadoras hipotalámicas y una categoría general que contiene otros péptidos que no se clasifican con facilidad. El estudio de los neuropéptidos comenzó hace más de 60 años con el descubrimiento accidental de la sustancia P (Figura 6.17A): un agente hipotensor potente y un ejemplo de la primera categoría de péptido. (El nombre peculiar deriva del hecho de que esta molécula era un componente no identificado de extractos secos en polvo de encéfalo e intestino). Este péptido de 11 aminoácidos está presente en altas concentraciones en el hipocampo, en la neocorteza y también en el tracto gastrointestinal humanos; de ahí su clasificación como péptido de encéfalo/intestino. También es liberado de las fibras C (véase Cuadro 9.1), las ramas aferentes de diámetro pequeño de los nervios periféricos que transmiten información acerca del dolor y de la temperatura (y señales autónomas posganglionares). La sustancia P es un neurotransmisor sensitivo de la médula espinal, donde su liberación puede ser inhibida por péptidos opioides que provienen de las interneuronas de la médula espinal, lo que conduce a la supresión del dolor (véase Cap. 10). La diversidad de los neuropéptidos es destacada por el hallazgo de que el gen que codifica la sustancia P codifica algunos otros péptidos neuroactivos que incluyen la neurocinina A, el neuropéptido K y el neuropéptido γ. Una categoría especialmente importante de neurotransmisores peptídicos es la familia de los “opioides” (Figura 6.17B). Estos péptidos se denominan así porque se unen a los mismos receptores postsinápticos activados por el opio. La amapola del opio es cultivada por lo menos desde hace 5 000 años y sus derivados utilizan como analgésico, al menos desde el Renacimiento. Los ingredientes activos del opio son una variedad de alcaloides vegetales, predominantemente morfina. La “morfina”, denominada así en honor a Morfeo, dios griego de los sueños, sigue siendo uno de los analgésicos más eficaces en uso actualmente, a pesar de su potencial adictivo (véase Recuadro 6F). Los opioides sintéticos como meperidina y metadona también se usan como analgésicos, y el fentanilo, droga con una potencia analgésica 80 veces superior a la de la morfina, es utilizado ampliamente en la anestesiología clínica. 134 CAPÍTULO 6 (A) Péptidos de encéfalo/intestino Sustancia P (YN 7YV 3`Z 7YV .SU .SU 7OL 7OL .S` 3L\ 4L[ Octapéptido de colecistocinina (CCK-8) (ZW ;`Y 4L[ .S` ;YW 4L[ (ZW 7OL Péptido intestinal /PZ (ZW (SH =HS 7OL ;OY (ZW (ZU ;`Y ;OY (YN 3L\ (YN 3`Z .SU 4L[ (SH =HS 3`Z 3`Z ;`Y 3L\ (ZU :LY 0SL 3L\ (ZU vasoactivo (VIP) (B) Péptidos opioides Propiedades de los aminoácidos Leucina encefalina ;`Y .S` .S` 7OL 3L\ Hidrofóbicos _-endorfina ;`Y .S` .S` 7OL 4L[ ;OY :LY .S\ 3`Z :LY .SU ;OY 7YV 3L\ =HS ;OY Dinorfina A ;`Y .S` .S` 7OL 3L\ (YN (YN 0SL (YN 7YV 3`Z 3L\ 3`Z ;YW (ZW (ZU .SU Polares, sin carga Ácidos Básicos (C) Péptidos hipofisarios Vasopresina *`Z ;`Y 7OL .SU (ZU *`Z 7YV (YN 3`Z .S` Oxitocina *`Z ;`Y 0SL .SU (ZU *`Z 7YV 3L\ .S` Hormona adrenocorticotrófica :LY ;`Y :LY 4L[ .S\ /PZ 7OL (YN ;YW .S` 3`Z 7YV =HS .S` 3`Z 3`Z (YN (YN 7YV =HS 3`Z =HS ;`Y 7YV (ACTH) (D) Péptidos liberadores hipotalámicos Hormona liberadora de tirotrofina (TRH) .S\ /PZ 7YV Hormona liberadora de hormona luteinizante (LHRH) .S\ /PZ ;YW :LY ;`Y .S` 3L\ (YN 7YV .S` Somatostatina 14 (SH .S` *`Z 3`Z (ZU 7OL 7OL ;YW 3`Z ;OY 7OL ;OY :LY *`Z (E) Péptidos diversos Angiotensina II (ZW (YN =HS ;`Y 0SL /PZ 7YV 7OL Neuropéptido Y ;`Y 7YV :LY 3`Z 7YV (ZW (ZU 7YV .S` .S\ (ZW (SH 7YV (SH .S\ (ZW 3L\ (SH (YN ;`Y ;`Y :LY (SH 3L\ (YN /PZ ;`Y 0SL (ZU 3L\ 0SL ;OY (YN .SU (YN ;`Y Neurotensina .S\ 3L\ ;`Y .S\ (ZU 3`Z 7YV (YN (YN 7YV 0SL 3L\ FIGURA 6.17 Secuencia de aminoácidos de los neuropéptidos. Los neuroendopéptidos tienen longitud variable, pero habitualmente contienen entre 3 y 36 aminoácidos. La secuencia de aminoácidos determina la actividad biológica de cada péptido. Los péptidos opioides fueron descubiertos en la década de 1970 durante la búsqueda de endorfinas, compuestos endógenos que imitan las acciones de la morfina. Se tenía la esperanza de que estos compuestos fueran analgésicos y de que su comportamiento arrojara luz sobre la adicción a las drogas. En la actualidad, se han identificado los ligandos endógenos de los receptores de opioides como una familia de más de 20 péptidos opioides agrupados en tres clases: las endorfinas, las encefalinas y las dinorfinas (Cuadro 6.2). Cada una de estas clases se libera a partir de un prepropéptido inactivo (preproopiomelanocortina, preproencefalina A y preprodinorfina), derivado de distintos genes (véase Fig. 6.16). El procesamiento de los precursores opioides es efectuado por enzimas procesadoras específicas de los tejidos que son empaquetadas en vesículas, junto con el péptido precursor, en el aparato de Golgi. Los péptidos opioides están ampliamente distribuidos en todo el encéfalo y a menudo se localizan junto con otros neurotransmisores de molécula pequeña, como el GABA y la 5-hidroxitriptamina. En general, estos péptidos tienden a ser depresores. Cuando se los inyecta de forma intracerebral en animales de experimentación, actúan como analgésicos; sobre la base de esta prueba y de otras, es probable que estén involucrados en los mecanismos que subyacen a la analgesia inducida por la acupuntura. Los opioides también participan en comportamientos complejos como la atracción sexual y en las conductas agresivas o sumisas. Asimismo tendrían un papel en algunos trastornos psiquiátricos como la esquizofrenia y el autismo, aunque las pruebas de ello son tema de debate. Lamentablemente, la administración repetida de opioides conduce a la tolerancia y a la adicción. Casi todos los neuropéptidos inician sus efectos activando receptores acoplados a las proteínas G. El estudio de estos receptores metabotrópicos de péptidos en el encéfalo ha sido difícil porque se conocen pocos agonistas y antagonistas específicos. Los péptidos activan sus receptores con bajas concentraciones (nM a μM) en comparación con las concentraciones requeridas para activar los receptores de los neurotransmisores de molécula pequeña. Estas propiedades permiten a los puntos diana postsinápticos de los péptidos localizarse alejados de las terminaciones presinápticas y modular las propiedades eléctricas de las neuronas que simplemente se encuentran en la vecindad del sitio de liberación del péptido. La activación de los receptores de los neuropéptidos es especialmente importante para regular NEUROTRANSMISORES Y SUS RECEPTORES 135 CUADRO 2. Péptidos opioides endógenos NOMBRE SECUENCIA DE AMINOÁCIDOSa Endorfinas α-Endorfina α-Neoendorfina β-Endorfina γ-Endorfina Tyr-Gly-Gly-Phe-Met-Thr-Ser-Glu-Lys-Ser-Gln-Thr-Pro-Leu-Val-Thr Tyr-Gly-Gly-Phe-Leu-Arg-Lys-Tyr-Pro-Lys Tyr-Gly-Gly-Phe-Met-Thr-Ser-Glu-Lys-Ser-Gln-Thr-Pro-Leu-Val-Thr-Leu-Phe-Lys-Asn-Ala-Ile-Val-Lys-Asn-AlaHis-Lys-Gly-Gln Tyr-Gly-Gly-Phe-Met-Thr-Ser-Glu-Lys-Ser-Gln-Thr-Pro-Leu-Val-Thr-Leu Encefalinas Leu-encefalina Met-encefalina Tyr-Gly-Gly-Phe-Leu Tyr-Gly-Gly-Phe-Met Dinorfinas Dinorfina A Dinorfina B a Tyr-Gly-Gly-Phe-Leu-Arg-Arg-Ile-Arg-Pro-Lys-Leu-Lys-Trp-Asp-Asn-Gln Tyr-Gly-Gly-Phe-Leu-Arg-Arg-Gln-Phe-Lys-Val-Val-Thr Obsérvese la homología inicial, indicada por las letras en bastardilla la eferencia posganglionar desde los ganglios simpáticos y la actividad del intestino (véase Cap. 21). Los receptores de neuropéptidos –sobre todo, el receptor del neuropéptido Y– también están involucrados en la iniciación y en el mantenimiento de la conducta alimentaria que conduce a la saciedad o la obesidad. Otros comportamientos adjudicados a la activación de los receptores de péptidos incluyen la ansiedad y las crisis de angustia, y los antagonistas de los receptores de la colecistocinina son útiles desde el punto de vista clínico en el tratamiento de estas afecciones. Se han desarrollado otros fármacos útiles que tienen como diana los receptores opioides. Tres subtipos bien definidos de receptores de opioides (μ, δ y κ) desempeñan un papel en los mecanismos de recompensa y adicción. El receptor de opioides μ ha sido identificado específicamente como sitio primario para la recompensa de la droga mediada por las drogas opioides. Neurotransmisores no convencionales Además de los neurotransmisores convencionales ya descritos, también se utilizan algunas moléculas poco comunes para la señalización entre las neuronas y sus puntos diana. Estas señales químicas pueden considerarse neurotransmisores debido a sus funciones en la señalización interneuronal y porque su liberación desde las neuronas está regulada por el Ca2+. Sin embargo, en comparación con otros neurotransmisores, no son convencionales, porque no son almacenados en vesículas sinápticas ni son liberados de las terminaciones presinápticas a través de mecanismos de exocitosis. De hecho, estos neurotransmisores no convencionales no necesitan ser liberados desde las terminaciones presinápticas y, a menudo, se asocian con señalización retrógrada (es decir, desde las células postsinápticas nuevamente hacia las terminaciones presinápticas). • Los endocannabinoides constituyen una familia de señales endógenas relacionadas que interactúan con los receptores de cannabinoides. Estos receptores son las diana moleculares del ∆9-tetrahidrocannabinol, el componente psicoactivo de la planta de marihuana, Cannabis (Recuadro 6G). Aunque no fueron definidos aún, algunos miembros de este grupo emergente de señales químicas, la anandamida y el 2-araquidonilglicerol (2-AG) son considerados endocannabinoides. Estas señales son grupos de ácidos grasos no saturado con cabeza polar y se producen por la degradación enzimática de los lípidos de membrana (Figura 6.18A, B). La producción de endocannabinoides es estimulada por una señal de segundo mensajero en el interior de las neuronas postsinápticas; en los casos típicos, una elevación en la concentración de Ca2+ postsináptica. Aunque el mecanismo de liberación de los endocannabinoides no es bien conocido, es probable que estas señales hidrofóbicas difundan a través de la membrana postsináptica para alcanzar los receptores de cannabinoides en otras células cercanas. La acción de los endocannabinoides concluye con el transporte, mediado por transportadores, de estas señales nuevamente al interior de la neurona postsináptica. Allí son hidrolizados por la hidrolasa de ácidos grasos (FAAH). Se han identificado por lo menos dos tipos de receptores de cannabinoides y la mayor parte de las acciones de los endocannabinoides en el SNC está mediada por el tipo denominado “CB1”. El CB1 es un receptor acoplado a la proteína G que se relaciona con los receptores metabotrópicos para la ACh, el glutamato y los otros neurotransmisores convencionales. Se sintetizaron varios compuestos relacionados estructuralmente con los endocannabinoides que se unen al receptor CB1 (Figura 6.18C). Estos compuestos actúan como agonistas o antagonistas del receptor CB1 y sirven como herramientas para dilucidar las funciones fisiológicas de los endocannabinoides y como elementos diana para desarrollar agentes útiles desde el punto de vista terapéutico. Los endocannabinoides participan en varias formas de regulación sináptica. La acción mejor documentada de estos agentes es la inhibición de la comunicación entre las células diana postsinápticas y sus aferencias presinápticas. En el hipocampo y en el cerebelo, además de otras regiones encefálicas, los endocannabinoides sirven como señales retrógradas para regular la liberación del GABA en ciertas terminaciones inhibidoras. En estas 136 CAPÍTULO 6 (A) Alkyl O C O O Acyl C O Fosfatidiletanolamina CH2 FIGURA 6.18 Señales de endocannabinoides involucradas en la transmisión sináptica. Posible mecanismo de producción de los endocan- CH nabinoides. (A) anandamida y (B) 2-AG. (C) Estructura del agonista del receptor de endocannabinoides WIN 55,212-2 y el antagonista rimonabante. (A, B, de CH2 Freund y cols., 2003; C, de Iversen, 2003.) O –O N-Aciltransferasa P O (C) O O NH3 O C O O C O CH2 CH N-arquidonoil fosfatidiletanolamina N CH2 CH3 WIN 55,212–2 O O –O Fosfolipasa D P O N O O O NH HO N O O N NH2 N Anandamida N Cl Cl (B) Acilo Araquidonilo Rimonabant O C O O C O Cl CH CH2 O Fosfatidilinositol Fosfolipasa C CH2 –O OH O O 1,2-Diacilglicerol (1,2-DAG) C O O C O Fosfolipasa A1 O P OH OH HO OH Inositol CH2 HO O CH C CH O CH2 CH2 O Lisofosfatidilinositol OH CH2 –O P O O 1,2-Diacilglicerol lipasa HO O C CH2 CH O CH2 OH 2-Araquidonilglicerol (2-AG) OH OH OH HO OH Lisofosfolipasa C NEUROTRANSMISORES Y SUS RECEPTORES 137 RECUADRO 6G La marihuana y el encéfalo El uso medicinal de la planta de la marihuana, Cannabis sativa (Figura A), se remonta a miles de años. Las antiguas civilizaciones –que incluyen tanto la sociedad griega como la romana en Europa, y las culturas hindú y china en Asia– sabían que esta planta era capaz de producir relajación, euforia y algunas otras acciones farmacológicas. En épocas más recientes, el uso medicinal de la marihuana en su mayor parte ha desaparecido (aunque sigue siendo útil para aliviar los síntomas de los pacientes oncológicos terminales); el uso recreativo de la marihuana (Figura B) se ha vuelto tan popular que algunas sociedades despenalizaron su uso. La comprensión de los mecanismos encefálicos que subyacen a las acciones de la marihuana progresó con el descubrimiento de que un cannabinoide, el ∆9-tetrahidrocannabinol (THC; Figura C), es el componente activo de la marihuana. El hallazgo condujo al desarrollo de derivados sintéticos, como WIN55, 212-2 y rimonabante (véase Fig. 6.18), que han servido como herramientas útiles para sondear las acciones encefálicas del ∆9-tetrahidrocan(C) (A) CH3 nabinol. Es de particular interés el hecho de que los receptores para estos cannabinoides existan en el encéfalo, muestren variaciones regionales pronunciadas en su distribución y sean especialmente ricas las áreas encefálicas –como la sustancia nigra y el caudado putamen– que fueron relacionadas con el abuso de drogas (Figura D). La presencia de estos receptores encefálicos para cannabinoides condujo, a su vez, a una investigación de los compuestos cannabinoides endógenos en el encéfalo, que culminó en el descubrimiento de los endocannabinoides como el 2-AG y la anandamida (véase Fig. 6.18). Este descubrimiento fue paralelo a la identificación de los péptidos opioides endógenos, que fueron el resultado de la búsqueda de compuestos endógenos similares a la morfina en el encéfalo (véase el texto y el Cuadro 6.2). Dado que el THC interactúa con los receptores encefálicos de endocannabinoides –sobre todo, el receptor CB1– es probable que estas acciones sean responsables de las consecuencias conductuales del consumo de marihuana. En efecto, 69-Tetrahidrocannabinol (THC) OH H 3C O Cannabis sativa (B) CH2 CH2 CH2 CH2 CH3 CH3 (D) Caudado putamen Hipocampo Cerebelo muchos de los efectos bien documentados de la marihuana son compatibles con la distribución y con las acciones de los receptores CB1 en el encéfalo. Por ejemplo, los efectos de la marihuana sobre la percepción podrían deberse a los receptores CB1 en la neocorteza; los efectos sobre el control psicomotor, a los receptores de endocannabinoides en los ganglios basales y el cerebelo; los efectos sobre la memoria a corto plazo, a los receptores de cannabinoides en el hipocampo; y los efectos bien conocidos de la marihuana como estimulante del apetito, a las acciones hipotalámicas. Aunque aún se está investigando las conexiones formales entre estas consecuencias conductuales de la marihuana y los mecanismos encefálicos subyacentes, algunos estudios de las acciones de esta droga han arrojado una comprensión fundamental de los mecanismos sinápticos básicos que prometen dilucidar mejor el modo de acción de una de las drogas más populares en todo el mundo. Bibliografía Adams, A. R. (1941) Marihuana. Harvey Lecture 37: 168-168. Freund, T. F., I. Katona and D. Piomelli (2003) Role of endogenous cannabinoids in synaptic signaling. Physiol. Rev. 83: 1017-1066. Gerdeman, G. L., J. G. Partridge, C. R. Lupica and D. M. Loytnger (2003) It could be habit forming: Drugs of abuse and striatal synaptic plasticity. Trends Neurosci. 26: 184-192. Howlett, A. C. (2005) Cannabinoid receptor signaling. Handbook Exp. Pharmacol. 168: 53-79. Iversen, L. (2003) Cannabis and the brain. Brain 126: 1252-1270. Mechoulam, R. (1970) Marihuana chemistry. Science 168: 1159-1166. Onaivi, E. S. (2009) Cannabinoid receptors in brain: Pharmacogenetics, neuropharmacology, neurotoxicology, and potential therapeutic applications. Int. Rev. Neurobiol. 88: 335-369. Sustancia nigra (A) Hoja de Cannabis sativa, planta de marihuana. (B) Fumar hojas de Cannabis cultivada es un método popular para lograr los efectos euforizantes de la marihuana. (C) Estructura del THC (∆9-tetrahidrocannabinol), el ingrediente activo de la marihuana. (D) Distribución de los receptores encefálicos CB1, visualizados por examen de la fijación de CP-55,940, un ligando del receptor CB1. (B, fotografía © Henry Diltz/Corbis; C, de Iversen, 2003; D, cortesía de M. Herkenham, NIMH). 138 CAPÍTULO 6 (A) Interneurona inhibidora (B) (C) Despolarización Después de la despolarización Tratado con rimonabante Amplitud del CPSI (%) Neurona piramidal Registro Estímulo CPSI (pA) 0 – Control – – Tiempo (ms) FIGURA 6.19 Control retrógrado de la liberación del GABA mediado por endocannabinoides. (A) Disposición experimental. La estimulación de una interneurona presináptica produce liberación de GABA en una neurona piramidal postsináptica. (B) Las corrientes postsinápticas inhibidoras (CPSI) producidas por la sinapsis inhibidora (control) tienen una amplitud reducida tras una despolarización breve de la neurona postsinápti- sinapsis, la despolarización de la neurona postsináptica produce una reducción transitoria en las respuestas postsinápticas inhibidoras (Figura 6.19). La despolarización reduce la transmisión sináptica al elevar la concentración de Ca2+ en el interior de la neurona postsináptica. Esta elevación en el Ca2+ desencadena la síntesis y liberación de endocannabinoides desde las células postsinápticas. Los endocannabinoides se dirigen luego hacia las terminaciones presinápticas y se unen a los receptores CB1 sobre estas terminaciones. La activación de los receptores CB1 inhibe la cantidad de GABA liberado en respuesta a los potenciales de acción presinápticos, y que reduce así la transmisión inhibidora. Los mecanismos responsables de la reducción en la liberación del GABA no se conocen bien, pero es probable que involucren efectos sobre los canales del Ca2+ o en los canales del K+ con puerta de voltaje en las neuronas presinápticas. • El óxido nítrico (NO) es una señal química poco común pero especialmente interesante. El NO es un gas producido por la acción de la óxido nítrico sintasa, enzima que convierte el aminoácido arginina en un metabolito (citrulina) y simultáneamente genera NO (Figura 6.20). En el interior de las neuronas, la NO sintasa neuronal está regulada por la unión del Ca2+ a la proteína sensora de Ca2+ calmodulina (véase Cap. 7). Una vez producido, el NO puede atravesar la membrana plasmática, lo que indica que el NO generado en el interior de una célula puede viajar a través del medio extracelular y actuar en el interior de células cercanas. Por lo tanto, esta señal gaseosa tiene una gama de influencias que se extiende mucho más allá de la célula de origen, y difunde algunas decenas de micrómetros de su sitio de producción antes de ser degradada. Esta propiedad convierte al NO en un agente potencialmente útil para coordinar las actividades de múltiples células en una región muy localizada y puede mediar ciertas formas de plasticidad sináptica que se propagan en el interior de pequeñas redes de neuronas. Todas las acciones conocidas del NO están mediadas en el 100 75 50 Control 25 0 –20 0 20 40 Tiempo (ms) 60 ca. Esta reducción en la corriente postsináptica inhibidora se debe a que se libera menos GABA desde la neurona presináptica. (C) La reducción en la amplitud de la corriente postsináptica inhibidora producida por la despolarización postsináptica dura algunos segundos y está mediada por endocannabinoides, porque es impedida por el antagonista de los receptores de endocannabinoides rimonabante. (B, C, de Ohno-Shosaku y cols., 2001.) interior de sus células diana; por esta razón, a menudo se lo considera un segundo mensajero más que un neurotransmisor. Algunas de estas acciones del NO se deben a la activación de la enzima guanilil ciclasa, la cual luego produce el segundo mensajero cGMP en el interior de las células diana (véase el Cap. 7). Otras acciones del NO son el resultado de la modificación covalente de las proteínas diana a través de la nitrosilación: el agregado de un grupo nitrilo a aminoácidos seleccionados en el interior de proteínas. El NO se degrada espontáneamente al reaccionar con oxígeno para producir óxidos de nitrógeno inactivos. Como resultado, las señales de NO duran solo un período breve, del orden de segundos o menos. La señalización del NO evidentemente regula distintas sinapsis que también emplean neurotransmisores convencionales; hasta ahora, las terminaciones presinápticas que liberan glutamato constituyen el punto diana mejor estudiado del NO en el sistema nervioso central. El NO también puede estar involucrado en algunas enfermedades neurológicas. Por ejemplo, se ha propuesto que el desequilibrio entre la generación de óxido nítrico y superóxido subyace a algunas enfermedades neurodegenerativas. Resumen Las complejas operaciones sinápticas que ocurren en los circuitos neurales de todo el encéfalo surgen por la acción de gran cantidad de neurotransmisores, los cuales actúan sobre una cantidad incluso mayor de receptores postsinápticos de neurotransmisores. El glutamato es el principal neurotransmisor excitador del encéfalo, mientras que el GABA y la glicina son los principales neurotransmisores inhibidores. Las acciones de estos neurotransmisores de molécula pequeña en los casos típicos son más rápidas que las de los neuropéptidos. Por lo tanto, los neurotransmisores de molécula pequeña median la transmisión sináptica cuando es esencial una respuesta rápida, mientras que NEUROTRANSMISORES Y SUS RECEPTORES Otras células diana Óxido nítrico Guanilil ciclasa Arginina GTP COO< + NH3 CH NO sintasa NH CH2 139 CH2 CH2 NH C cGMP O2 Óxido nítrico NH2 Citrulina Distintos óxidos de nitrógeno + Proteincinasa G COO< + NH3 CH O CH2 CH2 CH2 NH C NH2 FIGURA 6.20 Síntesis, liberación y terminación del NO. los transmisores neuropeptídicos, así como las aminas biógenas y algunos neurotransmisores de molécula pequeña, tienden a modular la actividad en curso en el encéfalo o en los tejidos diana periféricos de una forma más gradual y continua. Se han desarrollado dos familias bien diferentes de receptores de neurotransmisores que llevan a cabo las acciones de señalización postsináptica de los neurotransmisores. Los canales iónicos ionotrópicos o con puerta de ligando combinan el receptor para los neurotransmisores y el canal iónico en una entidad molecular y, por lo tanto, dan origen a respuestas eléctricas postsinápticas rápidas. Los receptores metabotrópicos regulan la actividad de los canales iónicos postsinápticos de modo indirecto, habitualmente a través de proteínas G, e inducen respuestas eléctricas más lentas y más duraderas. Los receptores metabotrópicos son especialmente importantes para regular el comportamiento, y los fármacos dirigidos a estos receptores han sido valiosos en la clínica para el tratamiento de una amplia gama de trastornos conductuales. La respuesta postsináptica en una sinapsis dada está determinada por la combinación de subtipos de receptores, de subtipos de proteínas G y de canales iónicos que son expresados en la célula postsináptica. Dado que cada una de estas características puede variar tanto en cada neurona como entre ellas, es posible una gran diversidad de efectos mediados por transmisores. Los fármacos que influyen en las acciones de los transmisores tienen una enorme importancia en el tratamiento de los trastornos neurológicos y psiquiátricos, y en un amplio espectro de otros problemas médicos. Lecturas adicionales Revisiones Beaulieu, J. M. and R. R. Gainetdinov (2011) The physiology, signaling, and pharmacology of dopamine receptors. Pharmacol Rev. 63: 182-217. Brown, D. A. and G. M. Passmore (2009) Neural KCNQ (Kv7) channels. Brit. J. Pharmacol. 156: 1185-1195. Carlsson, A. (1987) Perspectives on the discovery of central monoaminergic neurotransmission. Annu. Rev. Neurosci. 10: 19-40. Freund, T. F., I. Katona and D. Piomelli (2003) Role of endogenous cannabinoids in synaptic signaling. Physiol Rev. 83: 1017-1066. Hökfelt, T., O. Johansson, A. Ljungdahl, J. M. Lundberg and M. Schultzberg (1980) Peptidergic neurons. Nature 284: 515-521. Hyland, K. (1999) Neurochemistry and defects of biogenic amine neurotransmitter metabolism. J. Inher. Metab. Dis. 22: 353-363. Iversen, L. (2003) Cannabis and the brain. Brain 126: 1252-1270. Jaakola, V. P. and A. P. Ijzerman (2010) The crystallographic structure of the human adenosine A2A receptor in a high-affinity antagonist-bound state: Implications for GPCR drug screening and design. Curr. Opin. Struct. Biol. 20: 401-414. Koob, G. F., P. P. Sanna and F. E. Bloom (1998) Neuroscience of addiction. Neuron 21: 467-476. Laube, B., G. Maksay, R. Schemm and H. Betz (2002) Modulation of glycine receptor function: A novel approach for therapeutic intervention at inhibitory synapses? Trends Pharmacol. Sci. 23: 519-527. Nakanishi, S. (1992) Molecular diversity of glutamate receptors and implication for brain function. Science 258: 597-603. Pierce, K. L., R. T. Premont and R. J. Lefkowitz (2002) Seven-transmembrane receptors. Nature Rev. Mol. Cell Biol. 3: 639-650. Rosenbaum, D. M., S. G. Rasmussen and B. K. Kobilka (2009) The structure and function of G-protein-coupled receptors. Nature 459: 356-363. Schwartz, M. W., S. C. Woods, D. Porte Jr., R. J. Seeley and D. G. Baskin (2000) Central nervous system control of food intake. Nature 404: 661-671. Traynelis, S. F. and 9 others (2010) Glutamate receptor ion channels: structure, regulation, and function. Pharmacol Rev. 62: 405-496. Wilson, R. I. and R. A. Nicoll (2002) Endocannabinoid signaling in the brain. Science 296: 678-682. Artículos originales importantes Charpak, S., B. H. Gähwiler, K. Q. Do and T. Knöpfel (1990) Potassium conductances in hippocampal neurons blocked by excitatory amino-acid transmitters. Nature 347: 765-767. Chavas, J. and A. Marty (2003) Coexistence of excitatory and inhibitory GABA synapses in the cerebellar interneuron network. J. Neurosci. 23: 2019-2031. Chien, E. Y. and 10 others (2010) Structure of the human dopamine D3 receptor in complex with a D2/D3 selective antagonist. Science 330: 10911095. Curtis, D. R., J. W. Phillis and J. C. Watkins (1959) Chemical excitation of spinal neurons. Nature 183: 611-612. 140 CAPÍTULO 6 Dale, H. H., W. Feldberg and M. Vogt (1936) Release of acetylcholine at voluntary motor nerve endings. J. Physiol. 86: 353-380. Gomeza, J. and 6 others (2003) Inactivation of the glycine transporter 1 gene discloses vital role of glial glycine uptake in gly-cinergic inhibition. Neuron 40: 785-796. Hughes, J., T. W. Smith, H. W. Kosterlitz, L. A. Fothergill, B. A. Morgan and H. R. Morris (1975) Identification of two related pentapeptides from the brain with potent opiate agonist activity. Nature 258: 577-580. Kawate, T., J. C. Michel, W. T. Birdsong and E. Gouaux (2009) Crystal structure of the ATP-gated P2X4 ion channel in the closed state. Nature 460: 592-598. Kaupmann, K. and 10 others (1997) Expression cloning of GABAβ receptors uncovers similarity to metabotropic glutamate receptors. Nature 386: 239-246. Ledent, C. and 9 others (1997) Aggressiveness, hypoalgesia and high blood pressure in mice lacking the adenosine A2a receptor. Nature 388: 674-678. Miyazawa, A., Y. Fujiyoshi and N. Unwin (2003) Structure and gating mechanism of the acetylcholine receptor pore. Nature 423: 949-955. Mott, D. D., M. Benveniste and R. J. Dingledine (2008) pH-dependent inhibition of kainate receptors by zinc. J. Neurosci. 28: 1659-1671. Ohno-Shosaku, T., T. Maejima and M. Kano (2001) Endogenous cannabinoids mediate retrograde signals from depolarized postsynaptic neurons to presynaptic terminals. Neuron 29: 729-738. Peng, J. Y., N. Vaidehi, S. E. Hall and W. A. Goddard (2006) The predicted 3D structures of the human Ml muscarinic acetylcholine receptor with agonist or antagonist bound. Chem. Med. Chem. 1: 878-890. Rasmussen, S. G. and 12 others (2007) Crystal structure of the human beta2 adrenergic G-protein-coupled receptor. Nature 450: 383-387. Rasmussen, S. G. and 19 others (2011) Crystal structure of the α2 adrenergic receptor-Gs protein complex. Nature (in press). Rosenmund, C., Y. Stern-Bach and C. F. Stevens (1998) The tet-rameric structure of a glutamate receptor channel. Science 280: 1596-1599. Sobolevsky, A. I., M. P. Rosconi and E. Gouaux (2009) X-ray structure, symmetry and mechanism of an AMPA-subtype glutamate receptor. Nature 462: 745-756. Thomas, S. A. and R. D. Palmites (1995) Targeted disruption of the tyrosine hydroxylase gene reveals that catecholamines are required for mouse fetal development. Nature 374: 640-643. Unwin , N. (2005) Refined structure of the nicotinic acetylcholine receptor at 4 A resolution. J. Mol. Biol. 346: 967-989. Wang, Y. M. and 8 others (1997) Knockout of the vesicular monoamine transporter 2 gene results in neonatal death and supersensitivity to cocaine and amphetamine. Neuron 19: 1285-1296. Wall, M. A. and 6 others (1995) The structure of the G protein heterotrimer Giα1β1γ2. Cell 83: 1047-1058. Watanabe, J., A. Rozov .and L. P. Wollmuth (2005) Target-specific regulation of synaptic amplitudes in the neocortex. J. Neurosci. 25: 1024-1033. Libros Balazs, R., R.J. Bridges and C. W. Cotman (2006) Excitatory Amino Acid Transmission in Health and Disease. New York: Oxford University Press. Cooper, J. R., F. E. Bloom and R. H. Roth (2003) The Biochemical Basis of Neuropharmacology. New York: Oxford University Press. Feldman, R. S., J. S. Meyer and L. F. Quenzer (1997) Principles of Neuropharmacology, 2nd Edition. Sunderland, MA: Sinauer Associates. Hille, B. (2002) Ion Channels of Excitable Membranes, 3rd Edition. Sunderland, MA: Sinauer Associates. Howland, R. D., M. J. Maycek, R. A. Harvey and P. C. Champe (2005) Pharmacology, 3rd Edition. Philadelphia: Lippincott, Williams and Wilkins. Nestler, E., S. Hyman and R. Malenka (2009) Molecular Neuropharmacology: A Foundation for Clinical Neuroscience. New York: McGraw-Hill Medical. Siegel, G. J., R. W. Albers, S. Brady and D. Price (2006) Basic Neurochemistry: Molecular, Cellular and Medical Aspects. Burlington, MA: Elsevier Academic Press. Van Dongen, A. M. (2009) Biology of the NMDA Receptor. Boca Raton, FL: CRC Press. 141 CAPÍTULO 2 7 Señalización molecular en el interior de laS neuronaS Aspectos generales LOS MECANISMOS DE SEÑALIZACIÓN ELÉCTRICOS Y QUÍMICOS hacen posible que una célula nerviosa reciba información y la transmita a la otra. Este capítulo enfoca los acontecimientos que ocurren en el interior de las neuronas y otras células, los cuales son desencadenados por la interacción de una señal química con su receptor. En general, el procesamiento intracelular comienza cuando las señales químicas extracelulares, como los neurotransmisores, las hormonas y los factores tróficos, se unen a receptores específicos localizados sobre la superficie o en el interior del citoplasma o del núcleo de las células diana. Esta unión activa los receptores y, al hacerlo, estimula cascadas de reacciones intracelulares que comprenden proteínas fijadoras de GTP, moléculas de segundos mensajeros, proteincinasas, canales iónicos y muchas otras proteínas efectoras cuya modulación modifica transitoriamente el estado fisiológico de la célula diana. Estas mismas vías de transducción de señales intracelulares también pueden producir cambios duraderos al alterar la transcripción de genes, y afectar así la composición proteica de las células diana de forma más o menos permanente. La gran cantidad de componentes involucrados en las vías de señalización intracelular posibilita un control temporal y espacial preciso sobre la función de las neuronas individuales y, por esa vía, la coordinación de la actividad eléctrica y química en las poblaciones de neuronas relacionadas que comprenden circuitos y sistemas neurales. Estrategias de señalización molecular La comunicación química coordina el comportamiento de las células nerviosas y gliales individuales en los procesos fisiológicos que varían desde la diferenciación neural hasta el aprendizaje y la memoria. En efecto, la señalización molecular finalmente media y modula todas las funciones encefálicas. Para llevar a cabo esta comunicación, se ha desarrollado una serie de vías de señalización química extraordinariamente diversas y complejas. Los capítulos anteriores han descrito con cierto detalle los mecanismos de señalización eléctrica por medio de los cuales las neuronas generan potenciales de acción para conducir la información. Esos capítulos también describieron la transmisión sináptica, una forma especial de señalización química que transfiere información de una neurona a otra. Sin embargo, la señalización química no está limitada a las sinapsis. Otras formas bien caracterizadas de comunicación química son la señalización paracrina, cuya acción es más amplia que la transmisión sináptica y compren- 142 CAPÍTULO 7 de la secreción de señales químicas en un grupo de células diana cercanas, y la señalización endocrina, que se refiere a la secreción de hormonas en el torrente sanguíneo, desde donde pueden alcanzar puntos diana de todo el cuerpo (Figura 7.1). La señalización química de cualquier tipo requiere tres componentes: una señal molecular que transmite información de una célula a otra, una molécula receptora que transduce la información proporcionada por la señal y una molécula diana que media la respuesta celular (véase Figura 7.1A). La parte de este proceso que tiene lugar dentro de los límites de la célula diana se denomina transducción de señales intracelulares. Un buen ejemplo de transducción en el contexto de la comunicación intercelular es la secuencia de acontecimientos desencadenada por la transmisión sináptica química (véanse Fig. 7.1B y Cap. 5): los neurotransmisores son la señal, los receptores del neurotransmisor son el receptor transductor y la molécula diana es un canal iónico que se altera y produce la respuesta eléctrica de la célula postsináptica. En muchos casos, la transmisión sináptica activa vías intracelulares adicionales que tienen distintas consecuencias funcionales. Por ejemplo, la unión del neurotransmisor noradrenalina a su receptor activa proteínas fijadoras de GTP, lo que produce segundos mensajeros en el interior de la célula diana postsináptica, activa cascadas enzimáticas y finalmente (A) Célula de señalización FIGURA 7.1 Señalización química. Señal (A) Los componentes esenciales de la señalización química son: células que inician el proceso al liberar moléculas Receptor de señalización; receptores específicos sobre las células diana; moléculas diana segundos mensajeros y respuestas Molécula diana celulares posteriores. (B) Las formas de comunicación química incluyen transmisión sináptica, señalización paracrina y señalización endocrina. Respuesta (B) modifica las propiedades químicas de numerosas moléculas diana en el interior de la célula afectada. Una ventaja general de la señalización química en los contextos intercelular e intracelular es la amplificación de señales. La amplificación se desarrolla porque las reacciones de señalización individuales pueden generar un número mucho mayor de productos moleculares que el de moléculas que inician la reacción. En el caso de la señalización por noradrenalina, por ejemplo, una sola molécula de noradrenalina que se une a su receptor puede generar muchos miles de moléculas de segundo mensajero (como AMP cíclico), lo que ocasiona una amplificación de decenas de miles de fosfatos transferidos a las proteínas diana (Figura 7.2). Una amplificación similar tiene lugar en todas las vías de transducción de señales. Como los procesos de transducción a menudo están mediados por un conjunto secuencial de reacciones enzimáticas, cada una con su propio factor de amplificación, una pequeña cantidad de moléculas de señal finalmente puede activar una cantidad muy grande de moléculas diana. Esta amplificación garantiza que la respuesta fisiológica se produzca frente a otras influencias que potencialmente la contrarrestan. Otra razón para estos esquemas de transducción de señales complejas es la posibilidad de control preciso del comportamiento celular en una amplia gama de circunstancias. Algunas interacciones moleculares transfieren la información rápidamente, mientras que otras son más lentas y duran más. Por ejemplo, las cascadas de señalización asociadas con la transmisión sináptica en las uniones neuromusculares le permiten a una persona responder con rapidez a las señales cambiantes, como la trayectoria de una pelota arrojada, mientras que las respuestas más lentas desencadenadas por las hormonas de la médula suprarrenal (adrenalina y noradrenalina) secretadas durante un juego de competencia produce efectos más lentos (y más duraderos) sobre el metabolismo muscular (véase Cap. 21) y el estado emocional (véase Cap. 29). Para codificar información que varía con tanta amplitud en el tiempo, la concentración de las moléculas de señalización pertinentes debe estar cuidadosamente controlada. Por un lado, la concentración de toda molécula de señalización Sináptica Paracrina Endocrina Capilar Moléculas diana en células distantes Receptores activados Moléculas diana Moléculas diana Señales transportadas en el torrente sanguíneo SEÑALIZACIÓN MOLECULAR EN EL INTERIOR DE LAS NEURONAS Receptor Proteínas G Adenililciclasa Proteincinasas AMP cíclico Fosfatos transferidos a proteínas diana 143 FIGURA 7.2 Amplificación en las vías de transducción de señales. La activación de un único receptor por una molécula de señalización, como el neurotransmisor noradrenalina, puede conducir a la activación de numerosas proteínas G en el interior de las células. Estas proteínas activadas pueden unirse a otras moléculas de señalización, como la enzima adenilil-ciclasa. Cada molécula de enzima activada genera gran cantidad de moléculas de cAMP. El cAMP se une a otra familia de enzimas, las proteincinasas, y las activa. Estas enzimas pueden entonces fosforilar muchas proteínas diana. Aunque no todos los pasos de esta vía de señalización comprenden una Amplificación Sin amplificación Amplificación Sin amplificación en el interior de la cascada de señalización debe retornar a los valores subliminales antes de la llegada del otro estímulo. Por otro lado, el mantenimiento de los intermediarios en una vía de señalización activada es fundamental para una respuesta sostenida. La presencia de múltiples niveles de interacciones moleculares facilita el cronometrado intrincado de estos acontecimientos. Activación de las vías de señalización Los componentes moleculares de estas vías de transducción de señales siempre son activados por una molécula de señalización química. Estas moléculas de señalización pueden agruparse en tres clases: moléculas de señalización impermeables a la célula, permeables a la célula y asociadas a la célula (Figura (A) M oléculas impermeables a la célula Moléculas de señalización (B) Moléculas permeables a la célula Amplificación amplificación, globalmente la cascada produce un aumento tremendo en la potencia de la señal inicial. 7.3). Las dos primeras clases son moléculas secretadas y, por lo tanto, pueden actuar sobre células diana alejadas del sitio de síntesis o liberación de la señal. Las moléculas de señalización impermeables a la célula se caracterizan por unirse a receptores asociados con las membranas celulares. En la actualidad, se han identificado cientos de moléculas secretadas, que incluyen los neurotransmisores analizados en el Capítulo 6, proteínas como factores neurotróficos (véase Cap. 23) y hormonas peptídicas como glucagón, insulina y distintas hormonas de la reproducción. En general, estas moléculas de señalización son de vida corta, porque son metabolizadas rápidamente o porque son internalizadas mediante endocitosis una vez unidas a sus receptores. Las moléculas de señalización permeables a la célula pueden atravesar la membrana plasmática para actuar directamente so- (C) Moléculas asociadas a la célula Moléculas de señalización Moléculas de señalización FIGURA 7.3 Tres clases de moléculas de señalización. (A) Las moléculas impermeables a la célula, como neurotransmisores, no pueden atravesar fácilmente la membrana plasmática de la célula diana y deben unirse a la porción extracelular de las proteínas receptoras transmembrana. (B) Las moléculas permeables a la célula pueden atravesar la membrana plasmática Receptores transmembrana Receptor Receptor intracelular Núcleo y unirse a receptores en el citoplasma o el núcleo de las células diana. (C) Las moléculas asociadas a células se presentan sobre la superficie extracelular de la membrana plasmática. Estas señales activan receptores sobre las células diana solo si están directamente adyacentes a la célula de señalización. 144 CAPÍTULO 7 bre receptores que se encuentran en el interior de la célula. Los ejemplos incluyen muchas hormonas esteroides (glucocorticoides, estradiol y testosterona) y tiroideas (tiroxina), y los retinoides. Estas moléculas de señalización son relativamente insolubles en soluciones acuosas y a menudo son transportadas en la sangre y otros líquidos extracelulares unidas a proteínas transportadoras específicas. De esta forma, pueden persistir en el torrente sanguíneo durante horas o incluso días. Las moléculas del tercer grupo, de señalización asociadas a la célula, se disponen sobre la superficie extracelular de la membrana plasmática. En consecuencia, solo actúan sobre otras células cuando estas se encuentran en contacto físico con la célula que transporta estas señales. Los ejemplos incluyen proteínas como las integrinas y moléculas de adhesión a células nerviosas (NCAM), que influyen en el crecimiento axónico (véase Cap. 23). Las moléculas de señalización ligadas a la membrana ofrecen más dificultades para su estudio, pero indudablemente son importantes para el desarrollo neuronal y otras circunstancias en 1 La señal se une 2 El canal se abre Canal cerrado 3 Los iones fluyen a través de la membrana (C) Receptores acoplados a proteína G 1 La señal se une Receptor 1 La señal _ _ 2 La proteína G se une ` Tipos de receptores Cualquiera sea la naturaleza de la señal de iniciación, las respuestas celulares están determinadas por la presencia de receptores a los que se unen específicamente las moléculas de señalización. La unión de las moléculas señal produce un cambio conformacional en el receptor, el que entonces desencadena la cascada posterior de señalización en el interior de la célula afectada. Dado que las señales químicas pueden actuar en la membrana plasmática o en el citoplasma (o el núcleo) de la célula diana, no es sorprendente que los receptores se encuentren en realidad a ambos lados de la membrana plasmática. Los receptores para las moléculas de señalización impermeables son proteínas de expansión de la membrana. El dominio extracelular de estos receptores contiene el sitio de unión para la señal, mientras que el dominio intracelular activa las cascadas de señalización intracelulares una vez que se une la señal. Se ha identificado una (B) Receptores ligados a la enzima gran cantidad de estos receptores, los cuales han sido agrupados en familias definidas 1 La señal se une por el mecanismo utilizado para transducir Enzima la unión de señales en una respuesta celular 2 activada (Figura 7.4). Los receptores ligados a los canales (también denominados canales iónicos con puerta de ligando) tienen funciones de receptor y transductor como parte de la misma molécula proteica. La interacción entre la señal química y el sitio de fijación del receptor produce la apertura o el cierre de un Enzima inactiva Sustrato Producto poro del canal iónico en parte de la misma molécula. El flujo iónico resultante cambia 3 La enzima genera un el potencial de membrana de la célula diana producto y, en algunos casos, también puede conducir a la entrada de iones Ca2+ que sirven como (D) Receptores intracelulares señal de segundo mensajero en el interior de la célula. Son buenos ejemplos los receptoMolécula res ionotrópicos de los neurotransmisores de señalización descritos en el Capítulo 6. Los receptores ligados a enzimas tamEl re ceptor activado 2 regula la transcripción bién tienen un sitio de fijación extracelular para las señales químicas. El dominio intra- (A) Receptores ligados a los canales Iones las cuales el contacto físico entre las células proporciona información acerca de la identidad celular. ` se une Proteína G 3 Proteína G activada Receptor FIGURA 7.4 Categorías de receptores celulares. Las moléculas de señalización impermeables a la membrana pueden unirse a receptores ligados a los canales (A), receptores ligados a enzimas (B) o receptores acoplados a proteínas G (C) y activarlos. Las moléculas de señalización permeables a la membrana activan receptores intracelulares (D). SEÑALIZACIÓN MOLECULAR EN EL INTERIOR DE LAS NEURONAS celular de estos receptores es una enzima cuya actividad catalítica está regulada por la unión de una señal extracelular. La gran mayoría de estos receptores son proteincinasas, a menudo tirosincinasas, que fosforilan las proteínas diana intracelulares y modifican así la función fisiológica de las células diana. Son miembros destacables de este grupo la familia Trk de receptores de neurotrofinas (véase Cap. 23) y otros receptores para factores de crecimiento. Los receptores acoplados a la proteína G regulan reacciones intracelulares por un mecanismo indirecto que comprende una molécula transductora intermedia, denominada proteínas de fijación de GTP (o proteínas G). Como todos estos receptores comparten la característica estructural de atravesar la membrana plasmática siete veces, también se los denomina receptores 7-transmembrana (o receptores metabotrópicos; véase Cap. 5). Se han identificado cientos de receptores ligados a la proteína G diferentes. Ejemplos bien conocidos incluyen el receptor β-adrenérgico, el tipo muscarínico de receptor colinérgico, receptores metabotrópicos para glutamato, receptores para sustancias odoríferas en el sistema olfatorio y muchos tipos de receptores para hormonas peptídicas. La rodopsina, una proteína β-transmembrana sensible a la luz de los fotorreceptores retinianos, es otra forma de receptor ligado a la proteína G (véase Fig. 11.9). Los receptores intracelulares son activados por moléculas de señalización permeables a la célula o lipofílicas (Figura 7.4D). Muchos de estos receptores conducen a la activación de cascadas de señalización que producen nuevo mRNA y proteínas en el interior de la célula diana. A menudo, estos receptores comprenden una proteína receptora ligada a un complejo proteico inhibidor. Cuando la molécula de señalización se une al receptor, el complejo inhibidor se disocia para exponer un dominio de fijación de DNA sobre el receptor. Entonces, esta forma activada del receptor puede moverse hacia el núcleo e interactuar en forma directa con el DNA nuclear, lo que conduce a una alteración de la transcripción. Algunos receptores intracelulares se localizan fundamentalmente en el citoplasma, mientras que otros están en el núcleo. En cualquier caso, una vez que estos receptores son activados pueden afectar la expresión genética al alterar la transcripción del DNA. Proteínas G y sus puntos diana moleculares Tanto los receptores ligados a la proteína G como los ligados a enzima pueden activar cascadas de reacciones bioquímicas que finalmente modifican la función de las proteínas diana. Para ambos tipos de receptores, el acoplamiento entre la activación del receptor y sus efectos posteriores son las proteínas fijadoras de FIGURA 7.5 Tipos de proteína fijadora de GTP. (A) Las proteínas G heterotriméricas están compuestas por tres subunidades distintas (α, β y γ). La activación de los receptores produce la unión de la proteína G y hace que la subunidad α cambie GDP por GTP, lo que lleva a una disociación de las subunidades α y βγ. Las acciones biológicas de estas proteínas G son terminadas por hidrólisis de GTP, la cual se incrementa por medio de las proteínas activadoras de la GTPasa (GAP). (B) Las proteínas G monoméricas utilizan mecanismos similares para transmitir señales desde los receptores activados de la superficie celular a los puntos diana intracelulares. La unión de GTP estimula las acciones biológicas de estas proteínas G, y su actividad es finalizada por la hidrólisis de GTP, la cual está regulada también por proteínas GAP. (A) Proteínas G heterotriméricas (B) Proteínas G monoméricas Molécula de señalización química Molécula de señalización química Receptor ` _ 145 Receptor a GTP GDP Proteína efectora Proteína G ` GDP GTP _ GDP a GAP Activo Ras Ras GDP GTP Inactivo _ GDP GAP 146 CAPÍTULO 7 GTP. Existen dos clases generales de proteínas fijadoras de GTP (Figura 7.5). Las proteínas G heterotriméricas están compuestas por tres subunidades distintas (α, β y γ). Existen muchas subunidades α, β y γ diferentes, lo que permite una cantidad asombrosa de permutaciones de proteína G. Independientemente de la composición específica de la proteína G heterotrimérica, su subunidad α se fija a nucleótidos de guanina, ya sea GTP o GDP. La fijación de GDP permite entonces que la subunidad α se fije a las subunidades β y γ para formar un trímero inactivo. A su vez, la fijación de una señal extracelular a un receptor acoplado a la proteína G permite que dicha proteína se fije al receptor y hace que el GDP sea reemplazado por GTP (Figura 7.5A). Cuando el GTP está ligado a la proteína G, la subunidad β se disocia del complejo βγ y activa la proteína G. Tras la activación, tanto la subunidad α ligada al GTP como el complejo βγ libre pueden unirse a moléculas efectoras corriente abajo que median distintas respuestas en la célula diana. La segunda clase de proteínas fijadoras de GTP son proteínas G monoméricas (también denominadas proteínas G pequeñas). Estas GTPasas monoméricas también transmiten señales desde receptores de la superficie celular activados hasta puntos diana intracelulares como el citoesqueleto y el aparato de tráfico de vesículas de la célula. La primera proteína G pequeña fue descubierta en un virus que produce tumores sarcomatosos en la rata y por ello se la denominó ras. Ras es una molécula que ayuda a regular la diferenciación y proliferación celular al transmitir señales desde las cinasas de los receptores hasta el núcleo; la forma viral de la ras es defectuosa, lo que explica la capacidad del virus para producir proliferación celular descontrolada que conduce a los tumores. Se sabe que ras está implicada en varias Neurotransmisor Receptor Noradrenalina formas de señalización neuronal, como la potenciación sináptica a largo plazo (véase Cap. 8). Desde el descubrimiento de ras, se han identificado gran cantidad de GTPasas pequeñas que pueden dividirse en cinco subfamilias diferentes con distintas funciones. Por ejemplo, algunas participan en el tráfico de vesículas en la terminación presináptica o en otro sitio de la neurona, mientras que otras desempeñan un papel central en el tráfico de proteínas y RNA hacia el núcleo y desde él. La finalización de la señalización por las proteínas G heterotriméricas y monoméricas está determinada por la hidrólisis de GTP a GDP. La velocidad de la hidrólisis de GTP es una propiedad importante de una proteína G particular que puede ser regulada por otras proteínas, denominadas proteínas activadoras de GTPasa (GAP). Al reemplazar GTP por GDP, las GAP devuelven las proteínas G a su forma inactiva. Las GAP fueron reconocidas primero como reguladoras de proteínas G pequeñas, pero recientemente se ha observado que proteínas simples regulan las subunidades α de las proteínas G heterotriméricas. Por ello, las proteínas G monoméricas y triméricas funcionan como cronómetros moleculares activos en su estado ligado al GTP, y se vuelven inactivas cuando han hidrolizado el GTP ligado a GDP (Figura 7.5B). Las proteínas G activadas alteran la función de muchos efectores corriente abajo. La mayoría de estos efectores son enzimas FIGURA 7.6 Vías efectoras asociadas con receptores acoplados a proteínas G. En los tres ejemplos que se muestran, la unión de un neurotransmisor a uno de estos receptores conduce a la activación de una proteína G y al reclutamiento posterior de vías de segundos mensajeros. Gs, Gq y Gi se refieren a tres tipos diferentes de proteína G heterotrimérica. Glutamato Dopamina mGluR Dopamina D2 `adrenérgico Proteína G Gs Proteína efectora Segundos mensajeros Efectores posteriores Acción diana Adenilil-ciclasa cAMP Proteincinasa A Aumentan la fosforilación de proteínas Gq Fosfolipasa C Diacilglicerol Proteincinasa C Gi Adenilil-ciclasa IP3 Liberación de Ca2+ Aumentan la fosforilación de proteínas y activan proteínas fijadoras de calcio cAMP Proteincinasa A Disminuyen la fosforilación de proteínas SEÑALIZACIÓN MOLECULAR EN EL INTERIOR DE LAS NEURONAS que producen segundos mensajeros intracelulares. Las enzimas efectoras incluyen adenil-ciclasa, guanilil-ciclasa, fosfolipasa C y otras (Figura 7.6). Los segundos mensajeros producidos por estas enzimas desencadenan las cascadas de señalización bioquímica complejas, que se analizan en la sección siguiente. Como cada una de estas cascadas es activada por subunidades de proteínas G específicas, las vías activadas por un receptor particular están determinadas por la identidad específica de las subunidades de proteína G asociadas con él. Además de activar moléculas efectoras, las proteínas G también pueden unirse directamente y activar canales iónicos. Por ejemplo, algunas neuronas, así como las células del músculo cardíaco, tienen receptores acoplados a la proteína G que fijan acetilcolina. Como estos receptores también son activados por el agonista muscarina, habitualmente se los denomina receptores muscarínicos (véanse Caps. 6 y 21). La activación de los receptores muscarínicos puede abrir canales del K+, e inhibir así la frecuencia con la cual la neurona dispara potenciales de acción, o hacer más lento el latido cardíaco de las células musculares cardíacas. Se cree que estas respuestas inhibidoras son el resultado de las subunidades βγ de las proteínas G que se fijan a los canales del K+. La activación de las subunidades α también puede conducir al rápido cierre de los canales del Ca2+ y del Na+ con puerta de voltaje. Como estos canales transportan hacia el interior corrientes involucradas en la generación de los potenciales de acción, su cierre hace más fácil que las células diana disparen (véanse Caps. 3 y 4). En resumen, la fijación de señales químicas a sus receptores activa cascadas de transducción de señales en el citosol de las células diana. En el interior de estas cascadas, las proteínas G cumplen una función fundamental como elementos de transducción molecular que acoplan los receptores de membrana a sus efectores moleculares en el interior de la célula. La diversidad de proteínas G y sus puntos diana corriente abajo conduce a muchos tipos de respuestas fisiológicas. Al regular directamente la compuerta de los canales iónicos, las proteínas G pueden influir en el potencial de membrana de las células diana. Segundos mensajeros Las neuronas utilizan diferentes segundos mensajeros como señales intracelulares. Estos mensajeros difieren en el mecanismo por el cual son producidos y eliminados, así como en sus puntos diana y en sus efectos corriente abajo (Figura 7.7A). Esta sección resume las características de algunos de los principales segundos mensajeros. • Calcio. El ion calcio (Ca2+) tal vez sea el mensajero intracelular más común en las neuronas. En efecto, pocas funciones neuronales son inmunes a la influencia –directa o indirecta– del Ca2+. En todos los casos, la información es transmitida por una elevación transitoria en la concentración citoplasmática de calcio, lo cual favorece la unión del Ca2+ a gran cantidad de proteínas fijadoras de Ca2+ que sirven como puntos diana moleculares. Uno de los puntos diana del Ca2+ mejor estudiados es la calmo- 147 dulina, una proteína fijadora de Ca2+ abundante en el citosol de todas las células. La fijación del Ca2+ a la calmodulina activa esta proteína, la cual luego inicia sus efectos por medio de la unión a otros puntos diana corriente abajo, como las proteincinasas. Comúnmente, la concentración de iones Ca2+ en el citosol es muy baja, por lo general de 50-100 nanomolar (10-9 M). La concentración de iones Ca2+ en el exterior de las neuronas –en el torrente sanguíneo o el líquido cefalorraquídeo, por ejemplo– es de varios órdenes de magnitud más grande; en los casos típicos, 2+ de varios milimoles (10-3 M). El importante gradiente de Ca es mantenido por algunos mecanismos (Figura 7.7B). Lo más importante para este mantenimiento son dos proteínas que traslocan Ca2+ desde el citosol hasta el medio extracelular: una ATPasa denominada bomba de calcio y un intercambiador Na+/Ca2+, el cual es una proteína que reemplaza el Ca2+ intracelular por los iones sodio extracelulares (véase Capítulo 4). Además de estos mecanismos de membrana plasmática, el Ca2+ también es bombeado al retículo endoplasmático y las mitocondrias. Así, estos orgánulos pueden servir como sitios de almacenamiento de iones Ca2+ que más tarde son liberados para participar en procesos de señalización. Por último, las células nerviosas contienen otras proteínas fijadoras de Ca2+ –como calbindina– que sirven como amortiguadores del Ca2+. Estos amortiguadores se unen de forma reversible al Ca2+ y disminuyen así la magnitud y la cinética de las señales del Ca2+ en el interior de las neuronas. Los iones Ca2+ que actúan como señales intracelulares entran en el citosol por medio de uno o más tipos de canales iónicos permeables al Ca2+ (véase Cap. 4). Estos pueden ser canales del Ca2+ con puerta de voltaje o canales con puerta de ligando en la membrana plasmática, y ambos permiten que el Ca2+ fluya a favor de su gradiente y hacia la célula desde el medio extracelular. Además, otros canales hacen que el Ca2+ sea liberado desde el interior del retículo endoplasmático hasta el citosol. Estos canales que liberan Ca2+ intracelular tienen compuertas, de modo que pueden ser abiertos o cerrados en respuesta a distintas señales intracelulares. Uno de estos canales es el receptor de trifosfato de inositol (IP3). Como su nombre lo indica, estos canales están regulados por IP3, un segundo mensajero descrito con cierto detalle más adelante. Un segundo tipo de canal liberador de Ca2+ intracelular es el receptor de rianodina, que recibe este nombre por un fármaco que se une a estos receptores y los abre parcialmente. Entre las señales biológicas que activan los receptores de rianodina se encuentran el Ca2+ citoplasmático y, al menos en las células musculares, la despolarización de la membrana plasmática. Estos distintos mecanismos para elevar y eliminar iones Ca2+ hacen posible el control preciso del momento oportuno y la localización de la señalización del Ca2+ en el interior de las neuronas y, de esa manera, que el Ca2+ controle muchos eventos de señalización diferentes. Por ejemplo, los canales del Ca2+ con puerta de voltaje permiten que las concentraciones de Ca2+ se eleven localmente de forma muy rápida en el interior de las terminaciones presinápticas para desencadenar la liberación del neurotransmisor, como ya se describió en el Capítulo 5. Las elevaciones más 148 CAPÍTULO 7 FIGURA 7.7 Segundos mensajeros neuro- (A) Segundo mensajero Ca2+ nales. (A) Mecanismos responsables de producir y eliminar segundos mensajeros, así como los Fuentes Puntos diana intracelulares Mecanismos de eliminación Membrana plasmática: Canales del Ca2+ con puerta de voltaje Distintos canales con puerta de ligando Calmodulina Proteincinasas Proteinfosfatasas Canales iónicos Sinaptotagmina Muchas otras proteínas fijadoras de Ca2+ Membrana plasmática: Intercambiador Na+/Ca2+ Bomba de Ca2+ Retículo endoplasmático: Receptores de IP3 puntos diana corriente abajo de estos mensajeros. (B) Proteínas involucradas en la entrega Retículo endoplasmático: Bomba de Ca2+ de calcio al citoplasma y la eliminación de calcio del citoplasma. (C) Mecanismos de producción y degradación de nucleótidos cíclicos. (D) Vías Mitocondria involucradas en la producción y la eliminación de diacilglicerol (DAG) e IP3. Receptores de rianodina AMP cíclico La adenilil-ciclasa actúa sobre el ATP Proteincinasa A Canales con puerta de nucleótidos cíclicos Fosfodiesterasa de cAMP GMP cíclico La guanilil-ciclasa actúa sobre el GTP Proteincinasa G Canales con puerta de nucleótidos cíclicos Fosfodiesterasa de cGMP IP3 La fosfolipasa C actúa sobre PIP2 Receptores de IP3 sobre el retículo endoplasmático Fosfatasas Diacilglicerol La fosfolipasa C actúa sobre PIP2 Proteincinasa C Distintas enzimas (C) (B) 2+ 2+ Canal del Ca con puerta de voltaje Canal del Ca con puerta de ligando Adenililciclasa Intercambiador Bomba Na+/Ca2+ de Ca2+ Na+ H+ ATP Canal con puerta de nucleótidos cíclicos ATP Guanililciclasa GTP cAMP Canal con puerta de nucleótidos cíclicos cGMP ADP Retículo endoplasmático PKA Fosfodiesterasa de cAMP Retículo endoplasmático Fosfodiesterasa de cGMP AMP [Ca2+]i Proteínas efectoras fijadoras de Ca2+ Receptor de rianodina Fosfatidilinositol bifosfato (PIP2) ADP Diacilglicerol Fosfolipasa C Bomba de Ca2+ Proteínas amortiguadoras fijadoras de Ca2+ PKC Fosfatasas Inositol Mitocondria GMP (D) [Ca2+]i ATP Receptor de IP3 PKG IP3 Ca2+ Receptores de IP3 SEÑALIZACIÓN MOLECULAR EN EL INTERIOR DE LAS NEURONAS lentas y más difusas en la concentración de Ca2+ regulan una amplia variedad de otras respuestas, que incluyen la expresión genética en el núcleo celular. • Nucleótidos cíclicos. Otro grupo importante de segundos mensajeros son los nucleótidos cíclicos, especialmente el monofosfato cíclico de adenosina (cAMP) y el monofosfato cíclico de guanosina (cGMP) (Figura 7.7C). El AMP cíclico es un derivado de la molécula común de almacenamiento de energía, ATP. El AMP cíclico se produce cuando las proteínas G activan la adenilil-ciclasa en la membrana plasmática. Esta enzima convierte el ATP en cAMP al eliminar dos grupos fosfato del ATP. El GMP cíclico es producido de modo similar a partir de GTP por acción de la guanilil-ciclasa. Una vez que la concentración intracelular de cAMP o de cGMP está elevada, estos nucleótidos pueden unirse a dos clases diferentes de puntos diana. Los puntos diana más frecuentes de la acción de los nucleótidos cíclicos son las proteincinasas, ya sea la proteincinasa cAMP-dependiente (PKA) o la proteincinasa cGMP-dependiente (PKG). Estas enzimas median muchas respuestas fisiológicas al fosforilar proteínas diana, como se describe en la sección siguiente. Además, cAMP y cGMP pueden unirse a ciertos canales iónicos con puerta de ligando, e influir así en la señalización neuronal. Estos canales con puerta de nucleótidos cíclicos son particularmente importantes en la fototransducción y otros procesos de transducción sensitiva, como la olfación. Las señales de los nucleótidos cíclicos son degradadas por fosfodiesterasas, enzimas que dividen los enlaces fosfodiéster y convierten el cAMP en AMP o el cGMP en GMP. • Diacilglicerol e IP3. Hay que destacar que los lípidos de membrana también pueden convertirse en segundos mensaje- 149 ros intracelulares (Figura 7.7D). Los dos mensajeros más importantes de este tipo son producidos a partir de bifosfato de fosfatidilinositol (PIP2). Este componente lipídico es separado por la fosfolipasa C, enzima activada por ciertas proteínas G y por iones calcio. La fosfolipasa C divide el PIP2 en dos moléculas más pequeñas que actúan cada una como segundos mensajeros. Uno de estos mensajeros es diacilglicerol (DAG), molécula que se mantiene dentro de la membrana y activa a la proteincinasa C, la cual fosforila las proteínas sustrato en la membrana plasmática y en otros sitios. El otro mensajero es trifosfato de inositol (IP3), molécula que abandona la membrana celular y difunde en el interior del citosol. El IP3 se une a sus receptores, canales que liberan calcio desde el retículo endoplasmático. Por lo tanto, la acción de IP3 es producir otro segundo mensajero (¡tal vez un tercer mensajero, en este caso!) que desencadena un espectro completo de reacciones en el citosol. Las acciones de DAG e IP3 concluyen cuando ciertas enzimas convierten las dos moléculas en formas inertes que pueden ser recicladas para producir nuevas moléculas de PIP2. La concentración intracelular de estos segundos mensajeros cambia dinámicamente con el tiempo, lo que permite un control muy preciso sobre los puntos diana corriente abajo. Estas señales pueden localizarse también en comportamientos pequeños en el interior de células únicas o propagarse a través de grandes distancias, incluso entre las células a través de las uniones en brecha (véase Cap. 5). El conocimiento de la dinámica temporal y espacial compleja de estas señales de segundo mensajero ha sido muy favorecida por el desarrollo de las técnicas de imágenes que visualizan los segundos mensajeros y otras señales moleculares en el interior de las células (Recuadro 7A). RECUADRO 7A Imágenes dinámicas de señalización intracelular Los adelantos espectaculares en nuestro conocimiento del encéfalo a menudo se basan en el desarrollo de nuevas técnicas experimentales. Esto seguramente ha sido cierto para nuestro conocimiento de la señalización intracelular en las neuronas, que se ha beneficiado mucho de la invención de las técnicas de imágenes que permiten la visualización directa de los procesos de señalización en el interior de las células vivas. El primer adelanto –y muy posiblemente el más importante– provino del desarrollo, por Roger Tsien y cols., del colorante fluorescente fura-2 (Figura A). Los iones calcio se unen al fura-2 y hacen que cambien las propiedades de fluorescencia del colorante. Cuando se introduce fura-2 en el interior de las células y luego se obtienen imágenes de ellas con un microscopio de fluorescencia, este colorante sirve como informador de la concentración intracelular de Ca2+. Las imágenes con fura-2 nos han permitido detectar la dinámica espacial y temporal de las señales del Ca2+ que desencadenan innumerables procesos en el interior de las neuronas y las células gliales; por ejemplo, el fura2 se utilizó para obtener la imagen de la señalización del Ca2+ durante la liberación de neurotransmisores que se muestra en la Figura 5.11A. El refinamiento posterior de la estructura química de fura-2 ha proporcionado muchos otros colorantes fluorescentes indicadores de Ca2+ con diferentes propiedades de fluorescencia y distintas sensibilidades al Ca2+. Uno de estos colorantes es el Calcio Verde, que fue utilizado para obtener imágenes de los cambios dinámicos en la concentración del Ca2+ producidos en el interior de las dendritas de las células de Purkinje del cerebelo por el mensajero intracelular IP3 (Figura B). Otros adelantos han conducido a indicadores para visualizar la dinámica espacial y temporal de otras señales de segundos mensajeros, como cAMP. Otro adelanto notable en las imágenes dinámicas de los procesos de señalización provino del descubrimiento de una proteína fluorescente verde aislada por primera vez de la medusa Aequorea victoria. La proteína fluorescente verde (GFP) es una proteína que, como su nombre lo indica, es brillantemente fluorescente (Figura C). La clonación molecular del gen GFP permite obtener técnicas de imágenes para visualizar la expresión de los productos génicos marcados con fluorescencia GFP. El primer uso de este tipo de la GFP fue en experimentos con el gusano Caenorhabdi(Continúa en la página siguiente) 150 CAPÍTULO 7 RECUADRO 7A (continúa) (A) O O –O C C H 2C O– –O O O C C H 2C CH2 N CH2 N O CH2 CH2 O (A) Estructura química del colorante indicador de Ca2+ fura-2. (B) Imágenes de O– los cambios en la concentración intracelular de Ca2+ producida en una neurona cerebelosa de Purkinje por las acciones del segundo mensajero IP3. (C) Estructura molecular de la proteína fluorescente verde. GFP tiene forma de lata, con la porción fluorescente contenida en el interior de la lata. (D) Expresión de GFP en una neurona piramidal de la corteza cerebral del ratón. (A, de Grynkiewicz y cols., 1985; B, de Finch y Augustine, 1998; C, Armand Tepper, Leiden University; D, de Feng y cols., 2000.) O CH3 N (B) (C) (D) O C O– O tis elegans, en los cuales Martin Chalfie y cols. volvieron fluorescentes las neuronas al inducir la expresión de GFP en estas células. Muchos experimentos posteriores han utilizado la expresión de GFP en el encéfalo de mamíferos para obtener imágenes de la estructura de neuronas individuales (Figura D). Las estrategias de genética molecular hacen posible fijar GFP a casi cualquier proteína, lo que permite así obtener imágenes mediante microscopia de fluorescencia de la distribución espacial de las proteínas marcadas. De esta forma ha sido posible visualizar cambios dinámicos en la localización de las proteínas neuronales durante eventos de señalización. Por ejemplo, la Figura 8.11 muestra el uso de imágenes de GFP para controlar la activación de la proteincinasa Ca2+/calmodulinadependiente tipo II durante la potenciación sináptica prolongada. Como sucedió con fura-2, el refinamiento posterior de la GFP ha conducido a muchos adelantos. Uno de ellos es la producción de proteínas que dan fluorescencia en colores distintos del verde, lo cual permite obtener imágenes simultáneas de múltiples tipos de proteínas o múltiples tipos de neuronas. Otros refinamientos han conducido a técnicas para controlar la actividad enzimática de proteincinasas y otras proteínas de señalización. De la misma forma en que el desarrollo de la técnica de tinción de Golgi abrió nuestros ojos sobre la composición celular del encéfalo (véase Cap. 1), el estudio de la señalización intracelular en el encéfalo ha sido revolucionado por fura-2, GFP y otros métodos dinámicos de imágenes. No existe ningún fin a la vista para el potencial de estos métodos de imágenes de iluminar aspectos nuevos e importantes de la señalización encefálica. Bibliografía Bacskai, B. J. and 6 others (1993) Spatially resolved dynamics of cAMP and protein kinase A subunits in Aplysia sensory neurons. Science 260: 222-226. Chalfie, M., Y. Tu, G. Euskirchen, W. W. Ward and D. C. Prasher (1994) Green fluorescent protein as a marker for gene expression. Science 263: 802-805. Connor, J. A. (1986) Digital imaging of free calcium changes and of spatial gradients in growing processes in single mammalian central nervous system cells. Proc. Natl. Acad. Sci. USA 83: 6179-6183. Feng, G. and 8 Others (2000) Imaging neuronal subsets in transgenic mice expressing multiple spectral variants of GFP. Neuron 28: 41-51. Finch, E. A. and G. J. Augustine (1998) Local calcium signaling by IP3 in Purkinje cell dendrites. Nature 396: 753-756. Giepmans, B. N., S. R. Adams, M. H. Ellisman and R. Y. Tsien (2006) The fluorescent toolbox for assessing protein location and function. Science 312: 217-224. Grynkiewicz, G., M. Poenie and R. Y. Tsien (1985) A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 260: 3440-3450. Meyer, T. and M. N. Teruel (2003) Fluorescence imaging of signaling networks. Trends Cell Biol. 13: 101-106. Miyawaki, A. (2005) Innovations in the imaging of brain functions using fluorescent proteins. Neuron 48:189-199. Okumoto, S. (2010) Imaging approach for monitoring cellular metabolites and ions using genetically encoded biosensors. Curr. Opin. Biotechnol. 21: 45-54. Tsien, R. Y. (1998) The green fluorescent protein Annu. Rev. Biochem. 67: 509-544. SEÑALIZACIÓN MOLECULAR EN EL INTERIOR DE LAS NEURONAS Los blancos de los segundos mensajeros: proteincinasas y fosfatasas 151 (Figura 7.9A). La subunidad catalítica de PKA fosforila los residuos de serina y treonina de muchas proteínas diana diferentes. Aunque esta subunidad es similar a los dominios catalíticos de otras proteincinasas, distintos aminoácidos permiten que la PKA Como ya se mencionó, en general los segundos mensajeros re- se una a proteínas diana específicas, lo cual permite que solo gulan las funciones neuronales al modular el estado de fosforila- esos puntos diana sean fosforilados en respuesta a señales de ción de las proteínas intracelulares (Figura 7.8). La fosforilación cAMP intracelular. • Proteincinasa Ca2+/calmodulina-dependiente tipo II (CaM(agregado de grupos fosfato) cambia de forma rápida y reversible la función de las proteínas. Las proteínas son fosforiladas KII). Los iones Ca2+ que se unen a calmodulina pueden regular por una amplia variedad de proteincinasas; los grupos fosfato la fosforilación/desfosforilación de proteínas. En las neuronas, son eliminados por otras enzimas denominadas proteinfosfata- la proteincinasa Ca2+/calmodulina-dependiente más abundansas. El grado de fosforilación de una proteína diana refleja así un te es la CaMKII, una proteincinasa de Ser/Thr multifuncional. equilibrio entre las acciones competitivas de las proteincinasas CaMKII está compuesta por aproximadamente 14 subunidades, y las fosfatasas, lo que integra un conjunto de vías de señaliza- y en el encéfalo se encuentran los tipos α y β. Cada subunidad ción celular. Los sustratos de proteincinasas y fosfatasas inclu- contiene un dominio catalítico y uno regulador, así como otros yen enzimas, receptores de neurotransmisores, canales iónicos y dominios que permiten que la enzima se oligomerice y se dirija a la región correcta en el interior de la célula. Ca2+/calmodulina proteínas estructurales. Las proteincinasas y las fosfatasas actúan sobre los residuos activa CaMKII por el desplazamiento del dominio inhibidor del de serina y de treonina (Ser/Thr cinasas o fosfatasas) o los resi- sitio catalítico (Figura 7.9B). CaMKII fosforila gran cantidad de duos de tirosina (Tyr cinasas o fosfatasas) de sus sustratos. Al- sustratos, que incluyen canales iónicos y otras proteínas involugunas de estas enzimas actúan específicamente solo sobre una o cradas en la transducción de señales intracelulares. • Proteincinasa C (PKC). Otro grupo importante de proteincipocas proteínas diana, mientras que otras son multifuncionales y tienen una amplia gama de proteínas sustrato. La actividad nasas de Ser/Thr es la proteincinasa C (PKC). PKC son cinasas de proteincinasas y fosfatasas puede ser regulada por segundos monoméricas diversas activadas por los segundos mensajeros mensajeros, como cAMP o Ca2+, o por señales químicas extra- DAG y Ca2+. DAG causa que la PKC se mueva desde el citocelulares, como factores de crecimiento (véase Cap. 23). En ge- sol hasta la membrana plasmática, donde también se une a Ca2+ neral, los segundos mensajeros activan Ser/Thr cinasas, mientras y fosfatildilserina, un fosfolípido de membrana (Figura 7.9C). que las señales extracelulares activan Tyr cinasas. Aunque miles Estos acontecimientos atenúan la autoinhibición y ocasionan de proteincinasas se expresan en el encéfalo, relativamente po- que la PKC fosforile distintos sustratos proteicos. PKC también difunde a sitios distintos de la membrana plasmática –como el cas funcionan como reguladores de señalización neuronal. • Proteincinasa cAMP-dependiente (PKA). El efector prima- citoesqueleto, sitios perinucleares y el núcleo– donde fosforila rio de cAMP es la proteincinasa cAMP-dependiente (PKA). otras proteínas sustrato. La activación prolongada de PKC puede PKA es un complejo tetramérico de dos subunidades catalíticas lograrse con ésteres de forbol, compuestos promotores de tumoy dos subunidades inhibidoras (reguladoras). El cAMP activa a res que activan la PKC imitando a DAG. • Proteínas tirosincinasas. Dos clases de proteincinasas transla PKA por fijación a las subunidades reguladoras y ocasiona que libere subunidades catalíticas activas. Este desplazamien- fieren grupos fosfato a residuos de tirosina sobre las proteínas to de los dominios inhibidores es un mecanismo general para sustrato. Las tirosincinasas del receptor son proteínas transmemla activación de varias proteincinasas por segundos mensajeros brana con un dominio extracelular que se une a los ligandos de proteínas (factores de crecimiento, factores neurotróficos o citosinas) y un dominio catalítico Proteína intracelular que fosforila las proteínas sustratos relevantes. Las tirosincinasas que no son del reATP ceptor son enzimas citoplasmáticas o ligadas a Segundos Segundos la membrana que se activan indirectamente por mensajeros mensajeros señales extracelulares. La fosforilación de tirosina es menos frecuente que la fosforilación de ADP Proteinfosfatasa Ser/Thr y a menudo sirve para reclutar moléculas Proteincinasa de señalización para la proteína fosforilada. Las Fosfoproteína tirosincinasas son particularmente importantes para el crecimiento y la diferenciación celular FIGURA 7.8 Regulación de las proteínas celulares por fosforilación. Las proteincina(véanse Caps. 22 y 23). sas transfieren grupos fosfato (Pi) de ATP a residuos serina, treonina o tirosina sobre las proteí• Proteincinasa activada por mitógeno nas sustrato. Esta fosforilación altera de forma reversible la estructura y función de las proteínas (MAPK). Además de las proteincinasas que son celulares. La eliminación de grupos fosfato es catalizada por proteinfosfatasas. Tanto las cinasas activadas directamente por segundos mensajeros, como las fosfatasas son reguladas por distintos segundos mensajeros intracelulares. 152 CAPÍTULO 7 FIGURA 7.9 Mecanismo de activación de las proteincinasas. Las proteincinasas contienen varios dominios especializados con funciones es- (A) PKA Inactiva pecíficas. Cada una de las cinasas posee dominios catalíticos homólogos responsables de transferir grupos fosfato a proteínas sustrato. Estos dominios catalíticos son mantenidos inactivos por la presen- Dominios catalíticos Dominio regulador cia de un dominio autoinhibidor que ocupa el sitio catalítico. La unión de segundos mensajeros, como cAMP, DAG y Ca2+, al dominio regulador apropiado de la cinasa elimina el dominio autoinhibidor y permite que se active el dominio catalítico. En algunas cinasas, como PKC y CaMKII, los dominios autoinhibidor y catalítico forman parte de la misma molécula. En otras, como PKA, el dominio autoinhibidor es una subunidad separada. Fosforila sustratos cAMP Activa (B) CaMKII Inactiva Fosforila sustratos algunas de estas moléculas pueden ser actiActiva vadas por otras señales, como fosforilación Ca2+/CaM por otra proteincinasa. Son ejemplos importantes de estas las proteincinasas activadas por mitógeno (MAPK), también denominaDominio Dominio das cinasas reguladas por señales extracelucatalítico regulador lares (ERK). Las MAPK fueron identificadas por primera vez como participantes en el control del crecimiento celular, y en la (C) PKC actualidad se sabe que tienen muchas otras funciones de señalización. Por lo general, las MAPK están inactivas en las neuronas pero se activan cuando son fosforiladas por otras cinasas. De hecho, las MAPK Inactiva forman parte de una cascada de cinasas en Activa la cual una proteincinasa fosforila y activa Dominio Dominio regulador a la proteincinasa siguiente en la cascada. catalítico Fosforila DAG A menudo, las señales extracelulares que sustratos Ca2+ desencadenan estas cascadas de cinasas PS son factores de crecimiento extracelulares que se unen a las tirosincinasas del receptor que, por su parte, activan proteínas G monoméricas como ras. Una vez activadas, las MAPK pueden fosforilar factores de transcripción, proteínas sa de Ser/Thr más prevalente en las células de mamíferos. La que regulan la expresión genética. Entre la amplia variedad de actividad de PP1 está regulada por varias proteínas inhibidoras otros sustratos de MAPK existen varias enzimas, que incluyen expresadas en las neuronas. PP2A es una enzima con múltiples otras proteincinasas y proteínas del citoesqueleto. subunidades y una amplia gama de sustratos que se superponen Las proteinfosfatasas mejor caracterizadas son las fosfatasas con PP1. PP2B, o calcineurina, está presente en altas concentrade Ser/Thr PP1, PP2A y PP2B (también llamada calcineurina). ciones en las neuronas. Un rasgo característico de esta fosfataEn general, las proteinfosfatasas muestran menos especificidad sa es su activación por Ca2+/calmodulina. PP2B está compuesta de sustrato que las proteincinasas. Su especificidad limitada pue- por una subunidad catalítica y otra reguladora. Ca2+/calmodulina de surgir del hecho de que las subunidades catalíticas de las tres activa PP2B fundamentalmente por la fijación a una subunidad proteinfosfatasas principales son altamente homólogas, aunque catalítica y desplazamiento del dominio regulador inhibidor. En cada una de ellas aún se asocia con puntos diana o subunidades general, PP2B no tiene los mismos puntos diana moleculares reguladoras específicas. La PP1 desfosforila un amplio conjun- que CaMKII, aun cuando ambas enzimas son activadas por Ca2+/ to de proteínas sustrato y probablemente sea la proteinfosfata- calmodulina. SEÑALIZACIÓN MOLECULAR EN EL INTERIOR DE LAS NEURONAS En resumen, la activación de los receptores de membrana puede producir cascadas complejas de activación enzimática, lo que conduce a la producción de segundos mensajeros y fosforilación o desfosforilación de proteínas. Estas señales citoplasmáticas producen distintas respuestas fisiológicas rápidas al regular transitoriamente la actividad enzimática, los canales iónicos, las proteínas del citoesqueleto y muchos otros procesos celulares. En las sinapsis excitadoras, estos componentes de señalización a menudo están contenidos en el interior de las espinas dendríticas, que parecen servir como compartimientos de señalización especializados en el interior de las neuronas (Recuadro 7B). Además, estas señales pueden propagarse hasta el núcleo para producir cambios duraderos en la expresión genética. Señalización nuclear Los segundos mensajeros producen cambios prolongados en la función neuronal al promover la síntesis de nuevo RNA y proteínas. La acumulación resultante de nuevas proteínas requiere de por lo menos 30-60 minutos, marco temporal acorde con la magnitud más lenta de las respuestas mediadas por los flujos de iones o la fosforilación. Asimismo, la reversión de estos acontecimientos requiere horas a días. En algunos casos, los “cambios” genéticos pueden “producirse” para alterar permanentemente una neurona, como en la diferenciación neuronal (véase Cap.22). La cantidad de proteínas presente en la célula está determinada primariamente por la velocidad de transcripción del DNA Cromosoma 153 a RNA (Figura 7.10). El primer paso en la síntesis de RNA es la descondensación de la estructura de cromatina para proporcionar sitios de fijación para el complejo de la RNA polimerasa y para las proteínas activadoras transcripcionales, también llamadas factores de transcripción. Las proteínas activadoras transcripcionales se unen a los sitios de fijación presentes sobre la molécula de DNA cerca del comienzo de la secuencia del gen diana; ellas también se unen a otras proteínas que promueven el desenrrollamiento del DNA. El resultado neto de estas acciones es permitir que la RNA polimerasa, un complejo enzimático, se una sobre la región promotora del DNA y comience la transcripción. Además de aclarar el promotor para la RNA polimerasa, las proteínas activadoras pueden estimular la transcripción al interactuar con el complejo de la RNA polimerasa o con otras proteínas activadoras que influyen en la polimerasa. Las cascadas de la transducción de señales intracelulares regulan la expresión genética al hacer pasar las proteínas activadoras transcripcionales de un estado inactivo a uno activo, en el cual pueden unirse al DNA. Esta conversión se produce de varias formas. Las proteínas activadoras clave y los mecanismos que les permiten regular la expresión genética en respuesta a eventos de señalización se resumen brevemente en las secciones siguientes. • CREB. La proteína de fijación al elemento de respuesta al cAMP, que habitualmente se denomina CREB (cAMP response element binding), es un activador transcripcional de amplia distribución (Figura 7.11). Por lo general, la CREB se une a su sitio de fijación sobre el DNA (llamado elemento de respuesta al Cromatina en “collar de cuentas” UAS Proteína activadora transcripcional UAS Complejo coactivador UAS Unión de la proteína activadora transcripcional Unión de complejo coactivador la transcripción de DNA a RNA. La cromatina condensada (A) es desconden- Unión de RNA polimerasa RNA polimerasa y factores asociados UAS Cromatina condensada FIGURA 7.10 Pasos involucrados en Sitio de inicio del RNA Comienza la transcripción sada en una matriz de cuentas sobre una cadena de DNA (B) en la cual un sitio activador corriente arriba (upstream activator site, UAS) está libre de proteínas y unido por una proteína activadora transcripcional (factor de transcripción) específico de secuencia. Entonces, la proteína activadora transcripcional se une a complejos coactivadores, lo cual hace que la RNA polimerasa con sus factores asociados se una al sitio de inicio de la transcripción e inicie la síntesis de RNA. 154 CAPÍTULO 7 RECUADRO 7B Espinas dendríticas Muchas sinapsis del encéfalo involucran protrusiones dendríticas microscópicas denominadas espinas (Figura A). Las espinas se distinguen por la presencia de puntas globulares denominadas cabezas de espinas, que sirven como sitio postsináptico de inervación por las terminaciones presinápticas. Las cabezas de las espinas están conectadas a los tallos principales de las dendritas por conexiones estrechas denominadas cuellos de las espinas (Figura B). Inmediatamente por debajo del sitio de contacto entre las terminaciones y las cabezas de la espina están las estructuras intracelulares denominadas densidades postsinápticas (Figura C). El número, el tamaño y la forma de las espinas dendríticas son muy variables y, al menos en algunos casos, pueden cambiar dinámicamente con el tiempo (véase Fig. 8.15). Se ha descrito que las propiedades de las espinas se encuentran alteradas en algunos trastornos neurodegenerativos (como la enfermedad de Alzheimer) y trastornos psiquiátricos (como el autismo y la esquizofrenia). Desde la primera descripción de estas estructuras por Santiago Ramón y Cajal a fines del siglo xix, las espinas dendríticas han fascinado a generaciones de neurocientíficos e inspiraron muchas especulaciones sobre su función. Una de las primeras conjeturas fue que el cuello estrecho de la espina aísla eléctricamente las sinapsis del resto de la neurona. Dado que el tamaño de los cuellos de la espina puede cambiar, este mecanismo podría hacer que el efecto fisiológico de las sinapsis individuales varíe con el tiempo, lo cual aporta un mecanismo celular para formas de plasticidad sináptica como la potenciación prolongada y la depresión prolongada. Sin embargo, algunas mediciones posteriores de las propiedades de los cuellos de las espinas indican que estas estructuras serían relativamente inefectivas para atenuar el flujo de corriente eléctrica entre las cabezas de las espinas y las dendritas. Otra teoría –actualmente el concepto funcional más popular– postula que las espinas crean compartimientos bioquímicos. Esta idea se basa en otra de que el cuello de la espina podría impedir la difusión de las señales bioquímicas desde la cabeza de la espina hasta el resto de la dendrita. Varias (A) 3 +m 50 +m 5 +m (A) Dibujos clásicos de espinas dendríticas de Ramón y Cajal. Izquierda, dendritas de neuronas piramidales corticales. Derecha, imágenes de mayor amplificación de varios tipos diferentes de espinas dendríticas. observaciones resultan compatibles con esta idea. Primero, las mediciones muestran que el cuello de la espina sirve en efecto como barrera contra la difusión, y retarda la velocidad del movimiento molecular por un factor de 100 o más. En segundo lugar, las espinas se encuentran solo en las sinapsis excitadoras, donde se sabe que la transmisión sináptica genera muchas señales difusibles, principalmente el segundo (B) mensajero Ca2+. Por último, las imágenes de fluorescencia muestran que las señales sinápticas de Ca2+ pueden estar restringidas realmente a las espinas dendríticas en ciertas circunstancias (Figura D). No obstante, existen contraargumentos de la hipótesis de que las espinas proporcionan compartimientos bioquímicos relativamente aislados. Por ejemplo, se sabe que otros segundos mensajeros, como IP3, (C) Dendrita Terminación presináptica Densidad postsináptica Cuello de la espina Espina Cabeza de la espina Dendrita 1 +m Densidad postsináptica 1 +m (B) Reconstrucción de alta resolución con el microscopio electrónico de una región pequeña de la dendrita de una neurona piramidal del hipocampo. (C) Microfotografía electrónica de un corte transversal a través de una sinapsis excitadora. (B, de Harris, 1994; C, de Kennedy, 2000.) SEÑALIZACIÓN MOLECULAR EN EL INTERIOR DE LAS NEURONAS 155 RECUADRO 7B (continúa) (D) Señal localizada de Ca2+ (verde) producida en la espina de una neurona piramidal del hipocampo tras la activación de una sinapsis glutamatérgica. (E) (D) Las densidades postsinápticas incluyen docenas de moléculas de transmisión de señales, que incluyen los receptores glutamatérgicos (NMDA-R; mGluR), los re- 1 +m (E) NMDA–R CaMKII PSD–95 Calirina Rac ceptores de tirosinacinasa (RTK) y muchas moléculas de transducción de señales intracelulares, principalmente la proteincinasa CaMKII. (D de Sabatini y cols., 2002; E de Sheng y Kim, 2002.) RTK PSD–95 NMDA–R PSD–95 AKA SPAR Src PKA PYK2 PP2B PP1 SynGAP Rap Ras Raf MEK ERK mGluR PSD–95 PSD–95 nNOS GKAP GKAP H Shank (mango) Densidad postsináptica de la espina H Shank H H H H Retículo endoplasmático así como la proteína G monomérica ras, pueden difundir fuera de la cabeza de la espina y en el interior de la vaina dendrítica. Esta diferencia en la difusión presumiblemente se debe al hecho de que estas señales duran más que las señales de Ca2+, lo que les permite el tiempo suficiente como para superar la barrera de difusión del cuello de la espina. Otro punto relevante es que las señales postsinápticas de Ca2+ son altamente localizadas, incluso en las sinapsis excitadoras que no tienen espinas. Por lo tanto, al menos en algunos casos, las espinas no son necesarias ni suficientes para localizar la señalización sináptica de los segundos mensajeros. Una idea final y menos controvertida es que el propósito de las espinas es servir como reservorios donde pueden concentrarse proteínas de señalización (como los Receptores de IP3 puntos diana moleculares corriente debajo de Ca2+. IP3 y ras). Consistente con esta posibilidad, los receptores de glutamato se encuentran altamente concentrados en las cabezas de las espinas, y la densidad postsináptica comprende docenas de proteínas involucradas en la transducción de señales intracelulares (Figura E). Según este punto de vista, la cabeza de la espina es el destino para las moléculas de señalización durante la reunión de las sinapsis, así como el punto diana de los segundos mensajeros que son producidos por la activación local de receptores glutamatérgicos. Investigaciones recientes indican que las espinas también pueden atrapar moléculas que están difundiendo a lo largo de la dendrita, lo que podría ser un medio de concentrar estas moléculas en el interior de las espinas. Aunque la función de las espinas dendríticas sigue siendo enigmática, Ramón y Cajal indudablemente estaría encantado con el enorme grado de atención que estas pequeñas estructuras sinápticas siguen captando y el progreso real que se ha logrado en el conocimiento de las distintas tareas que son capaces de realizar. Bibliografía Bhatt, D.H., Zhang, S. and W. B. Gan (2009) Dendritic spine dynamics. Ann. Rev. Physiol. 71: 261-282. Goldberg, J. H., G. Tamas, D. Aronov and R. Yuste (2003) Calcium microdomains in aspiny dendrites. Neuron 40: 807-821. Harris, K. M. (1994) Serial electron microscopy as an alternative or complement to confocal microscopy for the study of synapses and dendritic spines in the central nervous system. In Three-Dimensional Confocal Microscopy: Volume Investigation of Biological Specimens, J. K. Stevens et al. (eds.), pp. 441-446. New York: Academic Press. Harris, K. M. and J. K. Stevens (1988) Dendritic spines of rat cerebellar Purkinje cells: serial electron microscopy with reference to their biophysical characteristics. J. Neurosci. 8: 4455-4469. Harvey, C. D., R. Yasuda, H. Zhong and K. Svoboda (2008) The spread of Ras activity triggered by activation of a single dendritic spine. Science 321: 136-140. Kennedy, M. B. (2000) Signal-processing machines at the postsynaptic density. Science 290: 750-754. Miyata, M. and 9 others (2000) Local calcium release in dendritic spines required for long-term synaptic depression. Neuron 28: 233-244. Noguchi, J., M. Matsuzaki, G. C. Ellis-Davies and H. Kasai (2005) Spine-neck geometry determines NMDA receptor-dependent Ca2+ signaling in dendrites. Neuron 46: 609-622. Penzes, P., M. E. Cahill, K. A. Jones, J. E. VanLeeuwen and K. M. Woolfrey (2011) Dendritic spine pathology in neuropsychiat-ric disorders. Nature Neurosci. 14: 285-293. Sabatini, B. L., T. G. Oertner and K. Svoboda (2002) The life cycle of Ca2+ ions in dendritic spines. Neuron 33: 439-452. Santamaria, F., S. Wils, E. De Schutter and G. J. Augustine (2006) Anomalous diffusion in Purkinje cell dendrites caused by spines. Neuron 52: 635-648. Sheng, M. and M. J. Kim (2002) Postsynaptic signaling and plasticity mechanisms. Science 298: 776-780. Yuste, R. and D. W. Tank (1996) Dendritic integration in mammalian neurons, a century after Cajal. Neuron 16: 701-716. 156 CAPÍTULO 7 cAMP, o CRE), ya sea como un homodímero o ligada a otro factor de transcripción estrechamente relacionado. En las células no estimuladas, la CREB no está fosforilada y tiene poca o ninguna actividad transcripcional. Sin embargo, la fosforilación de la CREB potencia mucho la transcripción. Varias vías de señalización pueden hacer que la CREB sea fosforilada. Tanto la PKA como la vía del ras, por ejemplo, pueden fosforilar la CREB. La CREB también puede ser fosforilada en respuesta a un aumento del calcio intracelular, en cuyo caso el sitio CRE también se denomina CaRE (calcium response element, elemento de respuesta al calcio). La fosforilación calcio-dependiente de CREB es causada primariamente por la cinasa Ca2+/calmodulina cinasa IV (de la familia de la CaMKII) y por la MAP cinasa, la cual conduce a una fosforilación prolongada de la CREB. La fosforilación de la CREB debe mantenerse el tiempo suficiente para que se produzca la transcripción, aun cuando la actividad eléctrica neuronal solo eleve transitoriamente la concentración intracelular de calcio. Estas cascadas de señalización pueden potenciar la transcripción mediada por la CREB al inhibir una proteinfosfatasa que la desfosforila. Por lo tanto, la CREB es un ejemplo de la convergencia de múltiples vías de señalización en un único activador transcripcional. Se han identificado muchos genes cuya transcripción está regulada por la CREB. Entre los genes sensibles a la CREB se hallan el gen temprano inmediato, c-fos (véase más adelante), la neurotrofina BDNF (véase Cap. 23), la enzima tirosina hidroxilasa (la cual es importante para la síntesis de neurotransmisores FIGURA 7.11 Regulación transcripcional por la CREB. Múltiples vías de señalización convergen activando cinasas que fosforilan la CREB. Estas incluyen PKA, Ca2+/calmodulina cinasa IV y MAP cinasas. La fosforilación de la CREB permite que se una a coactivadores (no se muestran en la figura), que luego estimulan la RNA polimerasa a comenzar la síntesis de RNA. El RNA es procesado y exportado al citoplasma, donde sirve como mRNA para la traducción en proteínas. Canal del Ca2+ Receptor acoplado a la proteína G Tirosincinasa del receptor Ca2+ Señal eléctrica Exterior celular ` Interior celular _ Adenilatociclasa Proteína G heterotrimérica Ca2+ cAMP ras Proteincinasa A MAP cinasa Ca2+/calmodulina cinasa IV Traducción mRNA CREB mRNA Transcripción DNA CRE/CaRE RNA polimerasa Genes diana Núcleo Proteína recién sintetizada (p. ej., enzima, proteína estructural, canales iónicos) SEÑALIZACIÓN MOLECULAR EN EL INTERIOR DE LAS NEURONAS catecolaminérgicos; véase Cap. 6) y muchos neuropéptidos (que incluyen somatostatina, encefalina y hormona liberadora de corticotrofina). También se considera que la CREB media cambios duraderos en la función encefálica; por ejemplo, aprendizaje espacial, sensibilización conductual, memoria a largo plazo del comportamiento condicionado por sustancias odoríferas y plasticidad sináptica prolongada (véanse Caps. 24 y 25). • Receptores nucleares. Los receptores nucleares para los ligandos permeables a la membrana también son activadores transcripcionales. El receptor para las hormonas glucocorticoideas ilustra un modo de acción de estos receptores. En ausencia de hormonas glucocorticoideas, los receptores se localizan en el citoplasma. La unión de los glucocorticoides ocasiona que el receptor se despliegue y se mueva hasta el núcleo, donde se une a un sitio de reconocimiento específico sobre el DNA. Esta unión del DNA activa el complejo RNA polimerasa pertinente para iniciar la transcripción y la posterior expresión genética. Por lo tanto, un acontecimiento regulador crítico para los receptores de esteroides es su translocación hasta el núcleo para permitir la unión al DNA. Los receptores para la hormona tiroidea (TH) y otros receptores nucleares no esteroides ilustran un segundo modo de regulación. En ausencia de TH, el receptor está unido a DNA y sirve como potente represor de la transcripción. Después de su unión a TH, el receptor sufre un cambio de conformación que finalmente abre el promotor para la unión de la polimerasa. Por ello, con la unión a TH el receptor pasa de ser un represor a ser un activador de la transcripción. Algunas hormonas regulan la expresión genética a través de estos receptores nucleares. • c-fos. Una estrategia diferente de regulación genética se aprecia en la función de la proteína activadora transcripcional, c-fos. En las células en reposo, c-fos está presente en concentración muy baja. Sin embargo, la estimulación de la célula diana hace que se sintetice c-fos, y la cantidad de esta proteína se eleva espectacularmente en 30-60 minutos. Por lo tanto, c-fos es considerado un gen temprano inmediato porque su síntesis es desencadenada directamente por el estímulo. Una vez sintetizada, la proteína c-fos puede actuar como activador transcripcional para inducir la síntesis de genes de segundo orden. Estos se denominan genes de respuesta retardada porque su actividad está retardada por el hecho de que un gen temprano inmediato –c-fos, en este caso– debe ser activado primero. Múltiples señales convergen sobre c-fos y activan diferentes factores de transcripción que se unen por lo menos a tres sitios distintos en la región promotora del gen. La región reguladora del gen c-fos contiene un sitio de fijación que media la inducción transcripcional por citosinas y factor neurotrópico ciliar. Otro sitio es el punto diana de factores de crecimiento, neurotrofinas a partir de ras y cinasa C, y un CRE/CaRE que puede unirse a CREB y responder así a cAMP o a la entrada de calcio resultante de la actividad eléctrica. Además de las interacciones sinérgicas entre estos sitios c-fos, las señales transcripcionales pueden integrarse convergiendo sobre el mismo 157 activador, como CREB. Por lo general, los eventos de señalización nuclear conducen a la generación de un complejo grande y relativamente estable compuesto por una proteína activadora transcripcional funcional, proteínas adicionales que se unen a la proteína activadora, y la RNA polimerasa y proteínas asociadas ligadas en el sitio de inicio de la transcripción. La mayoría de los eventos de señalización relevantes actúan para “sembrar” este complejo al generar una proteína activadora transcripcional activa por fosforilación, al inducir un cambio conformacional en el activador con fijación de ligando, al estimular la localización nuclear, al eliminar un inhibidor o simplemente al formar más proteína activadora. Ejemplos de transducción de señales neuronales El conocimiento de las propiedades generales de los procesos de transducción de señales en la membrana plasmática, en el citosol y en el interior del núcleo hace posible considerar cómo funcionan estos procesos conjuntamente para mediar funciones específicas en el encéfalo. Tres vías importantes de transducción de señales ilustran algunos de los papeles de los procesos intracelulares de transducción de señales en el sistema nervioso. • NGF/TrkA. El primero de estos es la señalización por el factor de crecimiento nervioso (NGF). Esta proteína es miembro de la familia de los factores de crecimiento de las neurotrofinas y es necesaria para la diferenciación, la supervivencia y la conectividad sináptica de las neuronas simpáticas y sensitivas (véase Cap. 23). El NGF funciona uniéndose a un receptor de tirosincinasa de alta afinidad, TrkA, que se encuentra sobre la membrana plasmática de estas células diana (Figura 7.12). La unión de NGF ocasiona que los receptores TrkA se dimericen y entonces la actividad intrínseca de tirosina cinasa de cada receptor fosforila a su receptor compañero. Los receptores TrkA fosforilados desencadenan la cascada ras, que conduce a la activación de múltiples proteincinasas. Algunas de estas cinasas se translocan hasta el núcleo y ponen en funcionamiento activadores de la transcripción como CREB. Este componente basado en ras de la vía de la NGF es el principal responsable de la inducción y el mantenimiento de la diferenciación en las neuronas sensibles al NGF. La fosforilación de TrkA también ocasiona que este receptor estimule la actividad de la fosfolipasa C, lo cual aumenta la producción de IP3 y DAG. El IP3 induce la liberación de Ca2+ desde el retículo endoplasmático, y el diacilglicerol activa la PKC. Estos dos segundos mensajeros parecen tener por puntos diana muchos de los mismos efectores corriente abajo como ras. Por último, la activación de los receptores TrkA también produce la activación de otras proteincinasas (como Akt cinasa) que inhiben la muerte celular. Por lo tanto, esta vía media fundamentalmente la supervivencia NGF-dependiente de las neuronas simpáticas y sensitivas descritas en el Capítulo 23. • Depresión prolongada. Se puede observar un interjuego en- 158 CAPÍTULO 7 Dímero de NGF FIGURA 7.12 Mecanismo de acción de NGF. El NGF se une a un receptor de tirosina cinasa de alta afinidad, TrkA, sobre la TrkA Exterior celular Interior celular membrana plasmática para inducir la fosforilación de TrkA en dos residuos diferentes de tirosina. Estas tirosinas fosforiladas sirven para unir distintas proteínas adaptadoras o fosfolipasa C (PLC), las cuales a su vez activan tres vías de señalización importantes: la vía de PI 3 cinasa que conduce a la activación de Akt cinasa, la vía de ras que conduce a MAP cinasas y la vía de PLC que conduce a la liberación del Ca2+ intracelular y a la activación de PKC. Las vías de ras y de PLC estimulan principalmente los procesos responsables de la diferenciación neuronal, Vía de PI 3 cinasa Vía de ras Vía de PLC Proteínas adaptadoras GEF Fosfolipasa C PI 3 cinasa ras Akt cinasa Cinasas IP3 MAP cinasa Supervivencia celular Liberación de Ca2+ del RE DAG PKC Excrecencia de neuritas y diferenciación neuronal tre varias señales intercelulares en las sinapsis excitadoras que inervan las células de Purkinje en el cerebelo. Estas sinapsis son fundamentales para el flujo de información a través de la corteza cerebelosa, lo que a su vez ayuda a coordinar los movimientos motores (véase Cap. 19). Una de las sinapsis se produce entre las fibras paralelas y sus células de Purkinje diana. La depresión prolongada es una forma de plasticidad sináptica que hace que las sinapsis de las fibras paralelas se vuelvan menos eficaces (véase Cap. 25). Cuando las fibras paralelas están activas, liberan el neurotransmisor glutamato en las dendritas de las células de Purkinje. Esto activa los receptores tipo AMPA, los cuales son canales iónicos con puerta de ligando (véase Cap. 6), y produce un pequeño PPSE que despolariza brevemente la célula de Purkinje. Además de esta señal eléctrica, la transmisión sináptica de las fibras paralelas también genera dos segundos mensajeros en el interior de la célula de Purkinje (Figura 7.13). El glutamato liberado por las fibras paralelas activa los receptores metabotrópicos de glutamato, lo cual estimula a la fosfolipasa C a producir IP3 y DAG. Cuando solo las sinapsis de las fibras paralelas están activas, estas señales intracelulares no son suficientes para abrir los receptores de IP3 o estimular la PKC. La depresión prolongada es inducida cuando las sinapsis de las fibras paralelas son activadas al mismo tiempo que las sinapsis glutamatérgicas de las fibras trepadoras que también inervan las células de Purkinje. Las sinapsis de las fibras trepadoras producen PPSE grandes que despolarizan fuertemente el potencial de mientras que la vía de PI 3 cinasa lo hace en la supervivencia celular. membrana de la célula de Purkinje. Esta despolarización permite que el Ca2+ entre en la célula de Purkinje a través de los canales del Ca2+ con puerta de voltaje. Cuando ambas sinapsis se activan simultáneamente, la elevación en la concentración intracelular de Ca2+ causada por la sinapsis de las fibras trepadoras aumenta la sensibilidad de los receptores de IP3 a la IP3 producida por las fibras de sinapsis paralelas y permite que se abran los receptores de IP3 en el interior de la célula de Purkinje. Esto libera Ca2+ desde el retículo endoplasmático y eleva aun más la concentración de Ca2+ localmente cerca de las sinapsis de las fibras paralelas. Esta elevación mayor en el Ca2+, junto con el DAG producido por las sinapsis de fibras paralelas, activa la PKC. A su vez, esta fosforila algunas proteínas sustrato. Finalmente, estos procesos de señalización cambian los receptores tipo AMPA en la sinapsis de las fibras paralelas, de modo que estos receptores producen señales eléctricas más pequeñas en respuesta al glutamato liberado de las fibras paralelas. Este debilitamiento de la sinapsis de las fibras paralelas es la causa final de la depresión prolongada. En síntesis, la transmisión en las sinapsis de las células de Purkinje produce señales eléctricas y señales químicas breves que duran mucho más. La interacción temporal entre estas señales permite que se produzca la depresión prolongada solo cuando están activas tanto las sinapsis de las fibras paralelas como las de las fibras trepadoras. Las acciones de IP3, DAG y Ca2+ también están restringidas a partes pequeñas de la dendrita de la célula de Purkinje, lo cual es un rango espacial más limitado que los PPSE, que se propagan en la totalidad de la dendrita y el cuerpo celular de la célula de Purkinje. Por lo tanto, al contrario de las señales eléctricas, las señales de segundos mensajeros pueden crear información precisa acerca de la localización de las sinapsis activas y permiten que se desarrolle la depresión prolongada solo en la vecindad de las fibras paralelas activas. • Fosforilación de la tirosina hidroxilasa. Un tercer ejemplo de señalización intracelular en el sistema nervioso es la regulación de la enzima tirosina hidroxilasa. La tirosina hidroxilasa SEÑALIZACIÓN MOLECULAR EN EL INTERIOR DE LAS NEURONAS 159 FIGURA 7.13 Señalización en las sinapsis de las vías paralelas cerebelosas. El glutamato liberado por las fibras paralelas activa los receptores tipo AMPA y metabotrópicos. El último produce IP3 y DAG en el interior de la célula de Purkinje. Cuando se conjuga con Terminación presináptica de fibra paralela una elevación en el Ca2+ asociado con la actividad de las sinapsis de las fibras trepadoras, el IP3 hace que el Na mGluR Glutamato Receptor de AMPA Ca2+ sea liberado del retículo endoplasmático, mientras que el Ca2+ y el DAG juntos activan a la proteincinasa C. Estas señales juntas cambian las propiedades de los receptores AMPA para producir una depresión prolongada. Na Fosfolipasa C Depresión prolongada DAG PKC Espina dendrítica de la célula de Purkinje Ca2+ La fibra trepadora despolariza el VM PIP 2 IP3 Liberación de Ca2+ Ca2+ Retículo endoplasmático gobierna la síntesis de los neurotransmisores catecolaminérgicos: dopamina, noradrenalina y adrenalina (véase Cap. 6). Algunas señales, entre ellas la actividad eléctrica, otros neurotransmisores y NGF, aumentan la velocidad de síntesis de catecolaminas al incrementar la actividad catalítica de la tirosina hidroxilasa (Figura 7.14). El rápido aumento de la actividad de la tirosina hidroxilasa se debe en gran parte a la fosforilación de esta enzima. La tirosina hidroxilasa es un sustrato para varias proteincinasas, como PKA, CaMKII, MAPcinasa y PKC. La fosforilación produce cambios conformacionales que aumentan la actividad catalítica de la tirosina hidroxilasa. Los estímulos que elevan cAMP, Ca2+ o DAG pueden aumentar la actividad de tirosina hidroxilasa e incrementar así la velocidad de biosíntesis de catecolaminas. Esta regulación por varias señales diferentes posibilita el control estrecho de la actividad de la tirosina hidroxilasa, e ilustra el modo en que varias vías diferentes pueden convergen para influir en una enzima clave involucrada en la transmisión sináptica. Resumen Existen distintas vías de transducción de señales en el interior de todas las neuronas. En los casos típicos, la activación de estas vías es iniciada por señales químicas como neurotransmisores y hormonas. Estas moléculas se unen a receptores que incluyen canales iónicos con puerta de ligando, receptores acoplados a la proteína G y receptores de tirosincinasa. Muchos de estos receptores activan proteínas G heterotriméricas o monoméricas que regulan cascadas enzimáticas intracelulares, canales iónicos o ambos. Un resultado frecuente de la activación de estos receptores es la producción de segundos mensajeros, como cAMP, Ca2+ e IP5 que se unen a las enzimas efectoras. Son efectores particularmente importantes las proteincinasas y las fosfatasas, que regulan el estado de fosforilación de los sustratos, y por lo tanto, su función. Estos sustratos pueden ser enzimas metabólicas u otras moléculas de transducción de señales, como canales iónicos, proteincinasas o factores de transcripción que regulan la expresión genética. Los ejemplos de factores de transcripción son CREB, receptores de hormonas esferoides y c-fos. Con esta 160 CAPÍTULO 7 FIGURA 7.14 Regulación de la tirosina hidroxilasa por la fosforilación de proteínas. Esta enzima gobierna la síntesis de los neurotransmisores catecolaminérgicos y es estimulada por algunas señales intracelulares. En el ejemplo que se muestra aquí, la actividad eléctrica neuronal (1) produce influjo de Ca2+ (2). La elevación resultante en la concentración de Ca2+ intracelular (3) activa las proteincinasas (4), las cuales fosforilan a la tirosina hidroxilasa (5) para estimular la síntesis de catecolaminas (6). Esto a su vez aumenta la liberación de catecolaminas (7) y potencia la respuesta postsináptica producida por la sinapsis (8). 1 Potencial de acción Tirosina hidroxilasa 2 Influjo de 4 Activación calcio de proteincinasas Ca2+ 5 Tirosina hidroxilasa fosforilada 3 Activación de segundos mensajeros Ca2+ 6 Aumento Proteincinasa Canal de calcio de la síntesis de catecolaminas 7 Aumento en la liberación del neurotransmisor 8 Aumento de la respuesta postsináptica plétora de componentes moleculares, las vías de transducción de señales intracelulares generan respuestas en una amplia gama de momentos y distancias, que aumentan y refinan mucho la capacidad de procesamiento de la información de los circuitos neurales y, finalmente, de los sistemas encefálicos. Ito, M. (2002) The molecular organization of cerebellar long-term depression. Nature Rev. Neurosci. 3: 896-902. Lecturas adicionales Nishizuka, Y. (1992) Intracellular signaling by hydrolysis of phospholipids and activation of protein kinase C. Science 258: 607-614. Revisiones Reichardt, L. F. (2006) Neurotrophin-regulated signalling pathways. Philos. Trans. R. Soc. London B 361: 1545-1564. Augustine, G. J., F. Santamaria and K. Tanaka (2003) Local calcium signaling in neurons. Neuron 40: 331-346. Rosenbaum, D. M., S. G. Rasmussen and B. K. Kobilka (2009) The structure and function of G-protein-coupled receptors. Nature 459: 356-363. Diesseroth, K., P. G. Mermelstein, H. Xia and R. W. Tsien (2003) Signaling from synapse to nucleus: The logic behind the mechanisms. Curr. Opin. Neurobiol. 13: 354-365. Sheng, M. and M. J. Kim (2002) Postsynaptic signaling and plasticity mechanisms. Science 298: 776-780. Fischer, E. H. (1999) Cell signaling by protein tyrosine phosphorylation. Adv. Enzyme Regul. Rev. 39: 359-369. Gilman, A. G. (1995) G proteins and regulation of adenylyl cyclase. Biosci. Rep. 15: 65-97. Kennedy, M. B., H. C. Beale, H. J. Carlisle and L. R. Washburn (2005) Integration of biochemical signalling in spines. Nature Rev. Neurosci. 6: 423434. Kumer, S. and K. Vrana (1996) Intricate regulation of tyrosine hydroxylase activity and gene expression. J. Neurochem. 67: 443-462. Artículos originales importantes Graves, J. D. and E. G. Krebs (1999) Protein phosphorylation and signal transduction. Pharmacol. Therap. 82: 111-121. Burgess, G. M., P. P. Godfrey, J. S. McKinney, M. J. Berridge, R. F. Irvine and J. W. Putney Jr. (1984) The second messenger inking receptor activation to internal calcium release in liver. Nature 309: 63-66. Greengard, P. (2001) The neurobiology of slow synaptic transmission. Science 294: 1024-1030. De Koninck, P. and H. Schulman (1998) Sensitivity of CaM kinase II to the frequency of Ca2+ oscillations. Science 279: 227-230. Greer, P. L. and M. E. Greenberg (2008) From synapse to nucleus: Calciumdependent gene transcription in the control of syn apse development and function. Neuron 59: 846-860. De Zeeuw, C. I. and 6 others (1998) Expression of a protein kinase C inhibitor in Purkinje cells blocks cerebellar long-term depression and adaptation of the vestibulo-ocular reflex. Neuron 20: 495-508. SEÑALIZACIÓN MOLECULAR EN EL INTERIOR DE LAS NEURONAS Finch, E. A. and G. J. Augustine (1998) Local calcium signaling by IP3 in Purkinje cell dendrites. Nature 396: 753-756. Lee, S. J., Y. Escobedo-Lozoya, E. M. Szatmari and R. Yasuda (2009) Activation of CaMKII in single dendritic spines during long-term potentiation. Nature 458: 299-304. Lindgren, N. and 8 others (2000) Regulation of tyrosine hydroxylase activity and phosphorylation at ser(19) and ser(40) via activation of glutamate NMDA receptors in rat striatum. J. Neurochem. 74: 2470-2477. Miller, S. G. and M. B. Kennedy (1986) Regulation of brain type II Ca2+/ calmodulin-dependent protein kinase by autophosphor-ylation: A Ca2+triggered molecular switch. Cell 44: 861-870. Northup, J. K P. C. Sternweis, M. D. Smigel, L. S. Schleifer, E. M. Ross and A. G. Gilman (1980) Purification of the regulatory component of adenylate cyclase. Proc. Natl. Acad. Sci. USA 77: 6516-6520. Rosenberg, O. S., S. Deindl, R. J. Sung, A. C. Nairn and J. Kuriyan (2005) Structure of the autoinhibited kinase domain of CaMKII and SAXS analysis of the holoenzyme. Cell 123: 849-860. 161 Shifman, J. M., M. H. Choi, S. Mihalas, S. L. Mayo and M. B. Kennedy (2006) Ca2+/calmodulin-dependent protein kinase II is activated by calmodulin with two bound calciums. Proc. Natl. Acad. Sci. USA 103: 13968-13973. Su, Y. and 7 Others (1995) Regulatory subunit of protein kinase A: Structure of deletion mutant with cAMP binding domains. Science 269: 807-813. Tao, X., S. Finkbeiner, D. B. Arnold, A. J. Shaywitz and M. E. Greenberg (1998) Ca2+ influx regulates BDNF transcription by a CREB family transcription factor-dependent mechanism. Neuron 20: 709-726. Tesmer, J. J., R. K. Sunahara, A. G. Gilman and S. R. Sprang (1997) Crystal structure of the catalytic domains of adenylyl cyclase in a complex with GsαGTPγS. Science 278:1907-1916. Libros Alberts, B., A. Johnson, J. Lewis, M. Raff, K. Roberts and P. Walter (2007) Molecular Biology of the Cell, 5th Ed. New York: Garland Science. Carafoli, E. and C. Klee (1999) Calcium as a Cellular Regulator. New York: Oxford University Press. 8 Plasticidad sináPtica Aspectos generales LAS CONEXIONES SINÁPTICAS ENTRE LAS NEURONAS proporcionan el “cableado” básico del circuito encefálico. Sin embargo, al contrario del cableado de un dispositivo electrónico como un ordenador, la conectividad sináptica entre las neuronas es una entidad dinámica que está cambiando constantemente en respuesta a la actividad neural y a otras influencias. Estos cambios en la transmisión sináptica surgen de algunas formas de plasticidad que varían en escala temporal desde milisegundos a años. La mayoría de las formas breves de plasticidad sináptica afecta la cantidad de neurotransmisor liberado desde las terminaciones presinápticas en respuesta a un potencial de acción presináptico. Varias formas de plasticidad sináptica a corto plazo –que incluyen facilitación, amplificación y potenciación– aumentan la liberación del neurotransmisor y son causadas por las acciones persistentes de los iones calcio en el interior de la terminación presináptica. Otra forma de plasticidad a corto plazo, la depresión sináptica, disminuye la cantidad de neurotransmisor liberado y, al parecer, se debe a una depleción de la actividad-dependiente de las vesículas sinápticas que están listas para la exocitosis. Las formas prolongadas de plasticidad sináptica alteran la transmisión sináptica en las escalas temporales de 30 minutos o más. Son ejemplos de esta plasticidad a largo plazo la potenciación prolongada y la depresión prolongada. Estas formas prolongadas de plasticidad sináptica se originan en mecanismos moleculares que varían en el tiempo: los cambios iniciales en la transmisión sináptica se originan en modificaciones postraduccionales de las proteínas existentes, principalmente cambios en el tráfico de los receptores glutamatérgicos, mientras que las fases posteriores de modificación sináptica son el resultado de cambios en la expresión genética, que producen cambios duraderos en la transmisión sináptica, que incluyen la proliferación de sinapsis, que pueden arrojar modificaciones esencialmente permanentes de la función encefálica. Plasticidad sináptica a corto plazo Las sinapsis químicas pueden experimentar cambios plásticos que refuerzan o debilitan la transmisión sináptica. Los mecanismos de la plasticidad sináptica ocurren en escalas temporales que varían desde milisegundos hasta días, semanas o más. Las formas a corto plazo de plasticidad, aquellas que duran algunos minutos o menos, se observan fácilmente durante la activación repetida de cualquier sinapsis química. Existen varias formas de plasticidad sináptica a corto plazo que difieren en su curso temporal y en sus mecanismos subyacentes. CAPÍTULO 8 100 60 40 Potencial de membrana postsináptico (mV) –63 –65 20 10 30 Tiempo (s) Respuesta postsináptica (mV/ms) –65 0 0,5 Cantidad de facilitación Depresión fuerte y rápida 20 0 0,30 [Ca2+] intermedia 0,25 Depresión más lenta mezclada con aumento 0,20 0,15 0,10 0,15 [Ca2+] 0,1 Normal Aumento solo 0,10 0,05 0,4 0,00 0,3 (D) 0,2 0,1 0,0 Normal [Ca2+] 80 Facilitación (B) Estímulo (C) 0 0 50 10 20 30 40 Intervalo entre los estímulos (ms) FIGURA 8.1 Formas de plasticidad sináptica prolongada. (A) Facilitación en la sinapsis gigante del calamar. Un par de potenciales de acción presinápticos produce dos potenciales postsinápticos excitadores (PPSE). A causa de la facilitación, el segundo PPSE es más grande que el primero. (B) Al variar el intervalo temporal entre los pares de potenciales de acción depresión, y deja solo un aumento (abajo). (D) La depresión sináptica en la sinapsis neuromuscular de la rana aumenta en proporción a la cantidad de transmisor liberado desde la terminación presináptica. (E) La aplicación de un estímulo tetánico de alta frecuencia (barra) en los axones presinápticos que inervan la neurona motora espinal produce una potenciación postetánica que persiste por un par de minutos una vez que termina el tétanos. (A, B, de Charlton y Bittner, 1978; C, de Swandulla y cols., 1991; D, de Betz, 1970; E, de Lev-Tov y cols., 1983.) La facilitación sináptica es un aumento rápido en la fuerza sináptica que ocurre cuando dos o más potenciales de acción invaden la terminación presináptica con pocos milisegundos entre sí (Figura 8.1A). Al variar el intervalo temporal entre los potenciales de acción presinápticos, se puede observar que la facilita- 10 15 20 Tiempo (s) 25 30 0,6 0,4 0,2 0 (E) 0 1 2 3 4 5 6 Cantidad relativa de liberación de transmisor 6,0 Respuesta postsináptica (mV) El descenso de la concentración externa de Ca hasta un nivel intermedio reduce la liberación del transmisor y produce una mezcla de depresión y aumento (intermedio). La mayor reducción del Ca2+ externo elimina la 2+ 5 0,8 presinápticos, se puede observar que la facilitación decae en el curso temporal de décimas de milisegundos. (C) En condiciones fisiológicas normales, un estímulo tetánico de alta frecuencia (barra más oscura) produce una depresión pronunciada del PPSE en la sinapsis gigante del calamar (arriba). 0 1,0 Cantidad relativa de depresión (A) Potencial de membrana presináptico (mV) 164 Potenciación postetánica 4,5 3,0 Estímulo 1,5 0 20 40 60 80 100 Tiempo (s) 120 140 160 ción producida por el primer potencial de acción dura decenas de milisegundos (Figura 8.1B). Existe abundante evidencia que indica que la facilitación es el resultado de la elevación prolongada de las concentraciones presinápticas de calcio después de la actividad sináptica. Aunque el ingreso de Ca2+ en el interior PLASTICIDAD SINÁPTICA 165 (A) Estímulos presinápticos de la terminación presináptica ocurre uno o dos milisegundos después de la aparición de un potencial de acción (véase Fig. 5.10B), los mecanismos que devuelven el Ca2+ a los niveles de reposo son mucho más lentos. Por lo tanto, cuando los potenciales de acción aparecen próximos unos a otros en el tiempo, el calcio aumenta en el interior de la terminación y permite que un potencial de acción presináptico posterior libere más neurotransmisor. El punto diana de esta señal de Ca2+ residual no está claro aún: una posibilidad es la ocupación parcial de los sitios de fijación del Ca2+ de la sinaptotagmina, la proteína sensora del Ca2+ que desencadena la liberación de Ca2+. Oponiéndose a la facilitación, se encuentra la depresión sináptica, que hace que se reduzca la liberación del neurotransmisor durante una actividad sináptica sostenida (Figura 8.1C, arriba). Un indicio importante de la causa de la depresión sináptica proviene de observaciones de que la depresión depende de la cantidad de neurotransmisor que ha sido liberado. Por ejemplo, el descenso de la concentración externa de Ca2+, para reducir el número de las cantidades liberadas por cada potencial de acción presináptico, reduce la velocidad de la depresión (Figura 8.1C). Asimismo, la intensidad de la depresión es proporcional a la cantidad de transmisor liberado desde la terminación pre- Potencial de membrana postsináptico (mV) <80 <90 <100 0 100 200 Tiempo (ms) 300 Cantidad relativa de liberación de transmisor (B) Disminución de la facilitación y del aumento Aumento sináptica (Figura 8.1D). Estos resultados condujeron a la idea de que la depresión está causada por la depleción progresiva de un grupo de vesículas sinápticas que se encuentran disponibles para la liberación: cuando las velocidades de liberación son altas, estas vesículas sufren una depleción rápida y producen mucha depresión; la depleción se hace más lenta a medida que la velocidad de liberación se reduce, lo que arroja menos depresión. Según esta hipótesis de la depleción vesicular, la depresión hace que decline la fuerza de la transmisión hasta que este grupo se recupera por la movilización de las vesículas desde el grupo de reserva. Son coherentes con esta explicación las observaciones de que se observa más depresión después de que se reduce el tamaño del grupo de reserva por alteración de la sinapsina, una proteína que mantiene las vesículas en el grupo de reserva (véase Cap. 5). Otras formas de plasticidad sináptica, como la potenciación y el aumento, también son producidas por la actividad sináptica repetida y sirven para aumentar la cantidad de transmisor liberado desde las terminaciones presinápticas. Tanto el aumento como la potenciación incrementan la capacidad de los iones calcio entrantes para desencadenar la fusión de las vesículas sinápticas con la membrana plasmática, pero trabajan en escalas temporales diferentes. Mientras el aumento se eleva y decae en algunos segundos (Figura 8.1C, panel inferior), la potenciación actúa en una escala temporal de decenas de segundos a minutos (Figura 8.1E). Como resultado de este curso temporal lento, la potenciación puede superar mucho al estímulo tetánico que la induce y esto se denomina a menudo potenciación postetánica. Aunque se considera que tanto el aumento como la potenciación surgen de la elevación prolongada de las concentraciones presinápticas de calcio durante la actividad sináptica, los mecanismos responsables de esta forma de plasticidad son poco conocidos. Se ha propuesto que el aumento es el resultado del Ca2+ que refuerza las acciones de la proteína presináptica munc13, mientras que la potenciación puede surgir cuando el Ca2+ activa las proteincinasas presinápticas que con(30 segundos tinúan hasta los sustratos fosforilados (como la sinapdespués) sina) que regulan la liberación de los transmisores. Durante la actividad sináptica repetitiva, estas for- Potenciación postetánica FIGURA 8.2 Plasticidad a corto plazo en la sinapsis neuromuscular. (A) Un tren de estímulos eléctricos (arriba) aplicados en el nervio motor presináptico produce cambios en la amplitud del potencial postexcitador (PPE) (abajo). (B) Cambios dinámicos en la liberación del transmisor causados Depresión Facilitación Potenciación Disminución de la depresión 0 100 200 Tiempo (ms) 300 por el interjuego de varias formas de plasticidad a corto plazo. La facilitación y el aumento del PPE ocurren al inicio del tren de estímulos y son seguidos por una depresión pronunciada del PPE. La potenciación comienza tarde en el tren de estímulos y persiste durante muchos segundos después de terminado el estímulo, lo que conduce a una potenciación postetánica. (A, de Katz, 1966; B, de Malenka y Siegelbaum, 2001.) 166 CAPÍTULO 8 mas de plasticidad a corto plazo pueden interactuar para hacer que la transmisión sináptica cambie de formas complejas. Por ejemplo, en la sinapsis neuromuscular periférica, la actividad repetida produce primero la acumulación de Ca2+ que permite la facilitación y luego el aumento para reforzar la transmisión sináptica (Figura 8.2). La depleción producida en las vesículas sinápticas hace entonces que domine la depresión y debilita la (A) sinapsis. Los potenciales de acción presinápticos que ocurren dentro del minuto o dos de la terminación del estímulo tetánico liberan más neurotransmisor por la persistencia de la potenciación postetánica. Aunque sus contribuciones relativas varían de una sinapsis a otra, estas formas de plasticidad sináptica a corto plazo producen un cambio dinámico en la transmisión de todas las sinapsis químicas como consecuencia de la historia reciente de actividad sináptica. Manto La plasticidad sináptica a largo plazo subyace a la modificación del comportamiento en Aplysia Sifón Cola Cabeza La facilitación, la depresión, el aumento y la potenciación modifican la transmisión sináptica en las escalas temporales de un par de minutos o menos. Aunque probablemente estos mecanismos sean responsables de muchos cambios breves en el circuito encefálico, no pueden proporcionar la base para los cambios en la función encefálica que persisten durante semanas, meses o años. Muchas sinapsis muestran formas prolongadas de plasticidad sináptica que son sustratos plausibles para cambios más permanentes en la función encefálica. Debido a su duración, estas formas de plasticidad sináptica pueden ser correlatos celulares de aprendizaje y memoria. Por lo tanto, se ha realizado un gran esfuerzo por comprender cómo se generan. Un obstáculo obvio para explorar la plasticidad sináptica en los encéfalos de los seres humanos y de otros mamíferos es la enorme cantidad de neuronas y la complejidad de las conexiones si- Branquia (B) Superficie dorsal Conectivo izquierdo Conectivo derecho Cuerpos celulares del ganglio Nervio del sifón Nervio pericardiogenital Ensayo 1 Retracción de la branquia (% de la primera respuesta) Contacto con el sifón 0 Ensayo 6 Contacto con el sifón 4 8 12 Tiempo (s) 0 4 8 12 Tiempo (s) 0 (E) 200 150 Sin descarga 100 Con una descarga en la cola 50 Única descarga en la cola 0 Ensayo 13 –2 0 2 Tiempo (horas) Ensayo 14 Contacto con el sifón Retracción de la branquia (% de la primera respuesta) Magnitud de la contracción de la branquia (C) (D) Nervio branquial Descarga en la cola y contacto en el sifón 4 8 12 Tiempo (s) 1000 0 4 8 12 Tiempo (s) Descargas en la cola 4 trenes diarios, por 4 días 100 4 0 Sin descargas 2 4 6 Tiempo (días) del reflejo de retirada de la branquia en Aplysia. (A) Esquema del animal (comúnmente conocido como babosa de mar). (B) Ganglio abdominal de Aplysia. Los cuerpos celulares de muchas de las neuronas involucradas en la retirada de la branquia pueden reconocerse por su tamaño, forma y posición en el interior del ganglio. (C) Cambios en el comportamiento de retirada de la branquia debido a la habituación y a la sensibilización. La primera vez que se toca el sifón, la branquia se contrae vigorosamente. Los contactos repetidos producen contracciones más pequeñas de la branquia debido a la habituación. Posteriormente, cuando se aparea un contacto con el sifón con una descarga eléctrica en la cola, se restablece la contracción grande y rápida de la branquia, resultado de la sensibilización a corto plazo. (D) Se observa sensibilización a corto plazo de la respuesta de retirada de la branquia después 4 descargas únicas en la cola 4 trenes de descargas en la cola 500 FIGURA 8.3 Sensibilización a corto plazo 8 del apareamiento de un único golpe en la cola con un contacto con el sifón. (E) Las aplicaciones repetidas de descargas en la cola producen una sensibilización prolongada de la respuesta de retirada de la branquia. (De Squire y Kandel, 1999.) PLASTICIDAD SINÁPTICA nápticas. Una forma de resolver este dilema es examinar la plasticidad en sistemas nerviosos mucho más alejados. La presunción de esta estrategia es que la plasticidad es tan esencial que sus fundamentos celulares y moleculares esenciales probablemente estén conservados en los sistemas nerviosos de organismos muy diferentes. Este abordaje resultó exitoso a la hora de identificar varias formas de plasticidad sináptica a largo plazo y para demostrar que estas formas subyacen a formas simples de aprendizaje. Eric Kandel y cols., en la Universidad de Columbia se ocuparon de estas cuestiones utilizando el molusco marino Aplysia californica (Figura 8.3A). Esta babosa marina tiene solo algunas decenas de miles de neuronas, muchas de las cuales son muy grandes (hasta 1 mm de diámetro) y se encuentran en localizaciones estereotipadas en el interior de los ganglios que conforman el sistema nervioso del animal (Figura 8.3B). Estos atributos hacen práctico controlar la actividad eléctrica de células nerviosas identificables específicas y definir así los circuitos sinápticos involucrados en la mediación del repertorio limitado de comportamientos de Aplysia. Aplysia muestra varias formas de plasticidad conductual. Una forma es la habituación, proceso que hace que el animal se vuelve menos reactivo ante la aparición repetida de un estímulo. La habituación se encuentra en muchas otras especies, incluido el ser humano. Por ejemplo, al vestirnos inicialmente experimentamos sensaciones táctiles debido a la ropa que estimula nuestra piel, pero la habituación hace rápidamente que estas sensaciones desaparezcan. Asimismo, un contacto leve en el sifón de Aplysia conduce a la retracción de la branquia, pero la habituación hace que la retracción de la branquia se vuelva más débil durante la estimulación repetida del sifón (Figura 8.3C). La respuesta de retracción de la branquia de Aplysia muestra otra forma de plasticidad denominada sensibilización. La sensibilización es un proceso que permite a un animal generalizar una respuesta contraria que es desencadenada por un estímulo nocivo a otros distintos estímulos no nocivos. En las Aplysia que se habituaron al contacto con el sifón, la sensibilización de la retirada de la branquia se produce al aparear un estímulo eléctrico fuerte en la cola del animal con otro tacto leve del sifón. Este apareamiento hace que el estímulo del sifón nuevamente desencadene la retracción fuerte de la branquia (Figura 8.3C, derecha) porque el estímulo nocivo de la cola sensibiliza al reflejo de retirada de la branquia al tacto leve. Incluso después de un estímulo aislado en la cola, el reflejo de retracción de la branquia sigue potenciado por lo menos por una hora (Figura 8.3D). Esto se puede considerar una forma simple de memoria a corto plazo. Con un apareamiento repetido de los estímulos en la cola y en el sifón, este comportamiento puede alterarse durante días o semanas (Figura 8.3E), y demuestra también una forma simple de memoria a largo plazo. La pequeña cantidad de neuronas en el sistema nervioso de Aplysia hace posible definir los circuitos sinápticos involucrados en la retracción de la branquia y controlar la actividad de las neuronas individuales en estos circuitos. Aunque finalmente cientos de neuronas están involucradas en la producción de este comportamiento simple, las actividades solo de algunos tipos 167 diferentes de neuronas pueden explicar la retracción de la branquia y su plasticidad durante la habituación y la sensibilización. Estas neuronas críticas incluyen neuronas mecanosensibles que inervan el sifón, neuronas motoras que inervan los músculos de la branquia e interneuronas que reciben aferencias de distintas neuronas sensitivas (Figura 8.4A). El contacto con el sifón activa las neuronas mecanosensoriales, que forman sinapsis excitadoras que liberan glutamato tanto en las interneuronas como en las neuronas motoras; por lo tanto, el contacto con el sifón aumenta la probabilidad de que ambos puntos diana postsinápticos produzcan potenciales de acción. Las interneuronas forman sinapsis excitadoras sobre las neuronas motoras, por lo que aumenta más la probabilidad de que las neuronas motoras disparen potenciales de acción en respuesta a la estimulación mecánica del sifón. Cuando las neuronas motoras son activadas por la excitación sináptica sumada de las neuronas sensitivas y las interneuronas, liberan acetilcolina que excita a las células musculares de la branquia, produciendo su retracción. Tanto la habituación como la sensibilización parecen surgir de cambios plásticos en la transmisión sináptica en este circuito. Durante la habituación, la transmisión en la sinapsis de los receptores glutamatérgicos entre las neuronas sensitivas y motoras está deprimida (Figura 8.4B, izquierda). Se considera que esta depresión sináptica es responsable de disminuir la capacidad de los estímulos en el sifón para evocar contracciones de la branquia durante la habituación. Muy similar a la forma a corto plazo de depresión sináptica descrita en la sección precedente, esta depresión es presináptica y se debe a una reducción en el número de vesículas sinápticas disponibles para la liberación. Por el contrario, la sensibilización modifica la función de este circuito al reclutar neuronas adicionales. La descarga en la cola que evoca la sensibilización activa neuronas sensitivas que la inervan. A su vez, estas neuronas sensitivas excitan interneuronas moduladoras que liberan serotonina en las terminaciones presinápticas de las neuronas sensitivas del sifón (véase Fig. 8.4A). La serotonina aumenta la liberación del transmisor desde las terminaciones de las neuronas sensitivas del sifón, y conduce a un aumento de la excitación sináptica de las neuronas motoras (Figura 8.4B). Esta modulación de la sinapsis neurona sensitiva-neurona motora dura aproximadamente una hora (Figura 8.4C), lo que es similar a la duración de la sensibilización a corto plazo de la retracción de la branquia producida por la aplicación de un único estímulo en la cola (Figura 8.3D). Por lo tanto, al parecer, la sensibilización a corto plazo se debe al reclutamiento de elementos sinápticos adicionales que modulan la transmisión sináptica en el circuito de retracción de la branquia. El mecanismo considerado responsable del aumento de la transmisión de los receptores del glutamato durante la sensibilización a corto plazo se muestra en la Figura 8.5A. La serotonina liberada por las interneuronas facilitadoras se une a los receptores acoplados a la proteína G sobre las terminaciones presinápticas de las neuronas sensitivas del sifón (paso 1), lo que estimula la producción del segundo mensajero, cAMP (paso 2). El AMP cíclico se une a las subunidades reguladoras de la proteincinasa 168 CAPÍTULO 8 FIGURA 8.4 Mecanismos sinápticos subyacentes a la sensibilización a corto plazo. (A) Neurona sensitiva Piel del sifón (A) Circuito neural que participa en la sensibilización. Normalmente, en contacto con la piel del sifón activa neuronas sensitivas que excitan interneuronas y neuronas motoras de la branquia, lo que ocasiona una contracción del músculo de Branquia Descarga del estímulo Cola la branquia. Una descarga en la cola del animal estimula neuronas moduladoras que alteran la transmisión sináptica entre las neuronas sensitivas Neurona motora Interneurona del sifón y las neuronas motoras de la branquia, Neurona sensitiva lo que conduce a la sensibilización. (B) Cambios de la eficacia sináptica en la sinapsis sensitivomotora durante la sensibilización a corto plazo. Interneurona moduladora Estímulo en el nervio de la cola (B) 0 min 20 min 50 min 20 s 50 min Potencial de 0 acción de la neurona <25 sensitiva (mV) <50 las neuronas sensitivas en las neuronas motoras, que incrementa el PPSE en las neuronas motoras y hace que estas exciten más intensamente el músculo de la branquia. (C) Curso temporal de la facilitación de la transmisión inducida por la sero- <40 PPSE de la neurona motora (mV) <50 tonina en la sinapsis sensitivomotora. (De Squire y Kandel, 1999.) 0 (C) Antes de la sensibilización, la activación de las neuronas sensitivas del sifón produce un PPSE en las neuronas motoras de la branquia. La activación de las interneuronas moduladoras serotoninérgicas aumenta la liberación de transmisor desde 100 200 300 0 Tiempo (ms) 100 200 300 Tiempo (ms) 0 100 200 300 Tiempo (ms) 0 100 200 300 Tiempo (ms) 500 PPSE de la neurona 300 motora (% del control) 100 0 0 10 20 30 Tiempo (min) 40 50 A (PKA; paso 3), las cuales liberan subunidades catalíticas de PKA que son capaces entonces de fosforilar varias proteínas, incluidos probablemente los canales del K+ (paso 4). El efecto neto de la acción de la PKA es reducir la probabilidad de que los canales de K+ se abran durante un potencial de acción presináptico. Este efecto prolonga el potencial de acción presináptico y abre así más canales de Ca2+ presinápticos (paso 5). Existen pruebas de que la apertura de los canales del Ca2+ presinápticos también es aumentada directamente por la serotonina. Por último, el influjo elevado de Ca2+ en las terminaciones presinápticas aumenta la cantidad de transmisor liberado en las neuronas motoras durante un potencial de acción de las neuronas sensitivas (paso 6). En resumen, una cascada de transducción de señales que involucra neurotransmisores, segundos mensajeros, proteincinasas y canales iónicos media la sensibilización a corto plazo de la retracción de la branquia. Esta cascada finalmente aumenta la transmisión sináptica entre las neuronas sensitivas y motoras en el interior del circuito de retracción de la branquia. Se considera que el mismo incremento de la liberación de glutamato inducido por la serotonina que media la sensibilización a corto plazo también subyace a la sensibilización a largo plazo. Sin embargo, durante la sensibilización a largo plazo, este circuito se ve afectado hasta durante varias semanas. La duración prolongada de esta forma de plasticidad se debe evidentemente a los cambios en la expresión genética y, por lo tanto, a la síntesis proteica (Figura 8.5B). Con el entrenamiento repetido (es decir, descargas adicionales en la cola), la PKA activada por serotonina involucrada en la sensibilización a corto plazo también fosforila ahora –y, por lo tanto, activa– al activador transcripcional CREB. Como se describe en el Capítulo 7, la fijación del CREB a los elementos de respuesta al cAMP (CREs) en las regiones reguladoras del DNA nuclear aumenta la velocidad de transcripción de los genes corriente abajo. Primero, el CREB estimula la síntesis de una enzima, la ubicuitina hidroxilasa, que estimula la degradación de la subunidad reguladora de la PKA. Esto produce un aumento persistente en la cantidad de subunidad catalítica libre, lo que indica que algo de la PKA es persistentemente activa y ya no requiere que se active la serotonina. El CREB también estimula otra proteína activadora transcripcional denominada C/EBP, la que estimula la transcripción de otros genes desconocidos que producen el agregado de terminaciones sinápticas, lo que produce un aumento prolongado de la cantidad de PLASTICIDAD SINÁPTICA FIGURA 8.5 Mecanismo de refuerzo presináptico subyacente a la sensibilización (A) Canal del K+ Interneurona facilitadora Canal del Ca2+ Neurona motora 5 Adenililciclasa Ca2+ K+ 4 2 cAMP 1 Subunidades 3 catalíticas 6 169 conductual. (A) La sensibilización a corto plazo se debe a un refuerzo agudo de la PKA-dependiente de la liberación de glutamato desde las terminaciones presinápticas de las neuronas sensitivas. Véase la explicación en el texto. (B) La sensibilización a largo plazo se debe a cambios en la expresión genética, que conducen a la síntesis de proteínas que cambian la actividad de la PKA y conducen a cambios en el crecimiento de las sinapsis. (De Squire y Kandel, 1999.) ATP los priones (véase Recuadro 19A), lo que podría permitir que se mantenga activo en perpetuidad y, por lo tanto, medie cambios Subunidades reguladoras permanentes en la estructura sináptica. Estos estudios de Aplysia y la investiReceptor del gación relacionada en otros invertebraNeurona glutamato dos, como la mosca de la fruta (Recuadro sensitiva 8A), condujeron a varias generalizaciones sobre la plasticidad sináptica. Primero, la plasticidad sináptica puede conducir claramente a cambios en la función de circui(B) to y, finalmente, a plasticidad conductual. Neurona Esta conclusión desencadenó un intenso CREB Interneurona motora facilitadora interés en los mecanismos de la plasticidad Núcleo sináptica. En segundo lugar, estos cambios plásticos en la función sináptica pueden PKA ser efectos a corto plazo que se basan en persistente una modificación postraduccional de las cAMP proteínas sinápticas existentes, o pueden DNA ser cambios a largo plazo que requieren Ubicuitina cambios en la expresión génica, síntesis de hidrolasa nuevas proteínas y proliferación de nuevas sinapsis (o la eliminación de las existentes). Por lo tanto, al parecer, los cambios a Proteínas no identificadas responsables del crecimiento corto y largo plazo en la función sináptica sináptico Neurona tienen diferentes bases mecánicas. Como sensitiva C/EBP veremos en las secciones siguientes, estas generalizaciones se aplican a la plasticidad sináptica en el encéfalo de los mamíferos sinapsis entre las neuronas sensitivas y motoras. Estos incremen- y, de hecho, han ayudado a guiar nuestro conocimiento de estas tos estructurales no se observan después de la sensibilización a formas de plasticidad sináptica. corto plazo. Pueden representar la causa final del cambio duradero en la fuerza global de las conexiones de los circuitos relevantes que producen un refuerzo de la respuesta de retracción de Potenciación a largo plazo en una la branquia. Otra proteína involucrada en la facilitación sináp- sinapsis del hipocampo tica a largo plazo es una proteína fijadora del elemento de poSe ha identificado plasticidad sináptica a largo plazo en el inliadenilación citoplasmático, cuya denominación algo confusa es CPEB (cytoplasmatic polyadenilation element binding). Esta terior del encéfalo de los mamíferos. Aquí, algunos patrones de proteína activa los mRNA y puede ser importante para el control actividad sináptica producen un aumento duradero de la fuerza local de la síntesis proteica. Lo más importante es que CPEB sináptica, conocida como potenciación a largo plazo (LTP), tiene propiedades de autosustentabilidad, como las proteínas de mientras que otros patrones de actividad producen una disminuReceptor de Proteína G la serotonina Proteincinasa A 170 CAPÍTULO 8 RECUADRO 8A Genética del aprendizaje y de la memoria en la mosca de la fruta Estos estudios condujeron a la identificación de una cantidad siempre creciente de mutaciones de gen único que interrumpen el aprendizaje o la memoria en las moscas. Los estudios conductuales y moleculares de los mutantes (dados los nombres imaginativos pero descriptivos como dunce, rutabaga y amnesiac) sugirieron que una vía central para el aprendizaje y la memoria en la mosca es la transducción de señales mediada por el nucleótido cíclico cAMP. Por lo tanto, los productos génicos de los loci dunce, rutabaga y amnesiac son, respectivamente, una fosfodiesterasa (que degrada cAMP), una adenililciclasa (que convierte el ATP en cAMP) y un transmisor peptídico que estimula adenililciclasa. Esta conclusión sobre la importancia del cAMP fue confirmada por el hallazgo de que la manipulación genética del factor de transcripción CREB también interfiere con el aprendizaje y con la memoria en las moscas normales. Estas observaciones en Drosophila concuerdan con las conclusiones alcanzadas en estudios de Aplysia y en mamíferos (véase el texto), que destacan la importancia del aprendizaje y la memoria mediados por cAMP en una amplia gama de otras especies. Bibliografía Davis, R. L. (2004) Olfactory learning. Neuron 44: 31-48. Quinn, W. G., W. A. Harris and S. Benzer (1974) Conditioned behavior in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 71: 708-712. Tully, T. (1996) Discovery of genes involved with learning and memory: An experimental synthesis of Hirshian and Benzerian perspectives. Proc. Natl. Acad. Sci. USA 93: 13460-13467. Waddell, S. and W. G. Quinn (2001) Flies, genes, and learning. Annu. Rev. Neuroscu 24: 12831309. Weiner, J. (1999) Time, Love, Memory: A Great Biologist and His Quest for the Origins of Behavior. New York: Knopf. 100 Desempeño de las moscas de la fruta de tipo salvaje y mutantes (Drosophila melanogaster) en una tarea de aprendizaje olfatorio. El desempeño de los mutantes dunce y rutabaga en esta tarea está disminuido por lo menos en un 50%. Las moscas mutantes en ambos loci dunce y rutabaga muestran incluso una disminución mayor del desempeño, lo que sugiere que los dos genes interrumpen aspectos diferentes del aprendizaje, pero que están relacionados. (De Tully, 1996.) ción duradera de la fuerza sináptica, conocida como depresión a largo plazo (LTD) por sus siglas en inglés. Potenciación y depresión a largo plazo son términos amplios que solo describen la dirección del cambio en la eficacia sináptica; de hecho, pueden participar diferentes mecanismos celulares y moleculares en la producción de ambas en diferentes sinapsis de todo el encéfalo. En general, la potenciación y la depresión a largo plazo son producidas por diferentes historias de actividad y están mediadas 80 Índice de rendimiento Como parte del renacimiento del análisis genético de los organismos simples a mediados de la década de 1970, varios investigadores reconocieron que la base genética del aprendizaje y de la memoria podría ser estudiada efectivamente en la mosca de la fruta, Drosophila melanogaster. En el cuarto de siglo que ha pasado, este abordaje arrojó algunas ideas fundamentales. Aunque por cierto los mecanismos del aprendizaje y de la memoria fueron uno de los problemas más difíciles que enfrentaron los genetistas de Drosophila, sus esfuerzos resultaron sorprendentemente exitosos. Descubrieron algunas mutaciones genéticas que alteran el aprendizaje y la memoria, y la identificación de estos genes ha proporcionado un marco de trabajo útil para estudiar los mecanismos celulares de estos procesos. El problema inicial en este trabajo fue desarrollar pruebas conductuales para poder identificar el aprendizaje anormal o los defectos mnésicos en poblaciones grandes de moscas. Este desafío fue cubierto por Seymour Benzer y sus colegas Chip Quinn y Bill Harris en el California Institute of Technology, quienes desarrollaron las pruebas de aprendizaje olfatorio y visual que se convirtieron en la base para la mayoría de los análisis posteriores de aprendizaje y memoria en la mosca de la fruta (véase la figura). Paradigmas conductuales que apareaban olores o luz con un estímulo adverso permitieron a Benzer y cols. evaluar el aprendizaje asociativo en las moscas. El diseño de un aparato de prueba ingenioso controló las señales sensoriales no relacionadas con el aprendizaje que previamente habían complicado estas pruebas conductuales. Además, el aparato permitió evaluar gran cantidad de moscas con relativa facilidad y acelerar el análisis de poblaciones con mutagénesis. Drosophila melanogaster 60 40 20 0 Tipo salvaje dunce rutabaga Mutantes de un solo gen dunce, rutabaga Mutantes dobles por diferentes complementos de vías de transducción de señales intracelulares en las células nerviosas involucradas. La plasticidad sináptica a largo plazo ha sido muy estudiada en las sinapsis excitadoras del hipocampo de los mamíferos. El hipocampo es especialmente importante en la formación o en la recuperación de algunas formas de memoria (véase Cap. 31). En los seres humanos, las imágenes funcionales muestran que el hipocampo es activado durante ciertos tipos de tareas de PLASTICIDAD SINÁPTICA 171 FIGURA 8.6 Circuito trisináptico del hipocampo. El corte a través del diafragma del hipocampo de un roedor muestra vías excitadoras y conexiones sinápticas. Se ha observado Hipocampo potenciación a largo plazo (signos más) en cada una de las tres conexiones sinápticas que se muestran aquí. Colaterales de Schaffer Células piramidales CA1 CA1 CA3 Células grano Células piramidales Fibras CA3 de Mossy Circunvolución dentada Vía perforante (desde la corteza entorrinal) memoria y que el daño de esta región del encéfalo conduce a una incapacidad para formar ciertos tipos de nuevas memorias. En los roedores, algunas neuronas del hipocampo solo disparan potenciales de acción cuando un animal se encuentra en ciertas localizaciones. Estas “células de lugar” parecen codificar memorias espaciales, interpretación apoyada por el hecho de que el daño del hipocampo impide que las ratas desarrollen eficiencia en las tareas de aprendizaje espacial (véase Fig. 31.10). Aunque muchas otras áreas encefálicas participan en el proceso complejo de la formación, del almacenamiento y de la recuperación de la memoria, estas observaciones impulsaron a muchos investigadores a estudiar la plasticidad sináptica a largo plazo de las sinapsis del hipocampo. La investigación sobre potenciación a largo plazo comenzó a fines de la década de 1960, cuando Terje Lomo y Timothy Bliss, que trabajaban en el laboratorio de Per Andersen en Oslo, Noruega, descubrieron que algunos segundos de estimulación eléctrica de alta frecuencia pueden aumentar la transmisión sináptica en el hipocampo del conejo durante días o incluso semanas. Sin embargo, más recientemente, el progreso en el conocimiento del mecanismo de la potenciación a largo plazo se basó principalmente en estudios in vitro de cortes de hipocampo vivo. La disposición del hipocampo permite cortarlo de modo que la mayoría de los circuitos relevantes queden intactos. En estos preparados, los cuerpos celulares de las neuronas piramidales se ubican en una capa única densamente empaquetada que es fácilmente evidente (Figura 8.6). Esta capa está dividida en varias regiones distintas, y las principales son CA1 y CA3. “CA” se refiere a cornu Ammonis –el latín para el cuerno de Amón–, el cuerno del carnero que se asemeja a la forma del hipocampo. Las dendritas de las células piramidales en la región CA1 forman una banda gruesa (el estrato radiado), donde reciben sinapsis de las colaterales de Schaffer, los axones de las células piramidales en la región CA3. Gran parte de la investigación sobre potenciación a largo plazo se ha concentrado en las conexiones sinápticas entre las colaterales de Schaffer y las células piramidales CA1. La estimulación eléctrica de las colaterales de Schaffer genera potenciales postsinápticos excitadores (PPSE) en las células CA1 postsinápticas (Figura 8.7A,B). Cuando se estimulan las colaterales de Schaffer solo dos o tres veces por minuto, el tamaño del PPSE evocado en las neuronas CA1 se mantiene constante. Sin embargo, una sucesión breve de estímulos de alta frecuencia en los mismos axones produce potenciación a largo plazo, que se evidencia como un aumento prolongado en la amplitud del PPSE (Figura 8.7B, C). Aunque no se conoce la duración máxima de la potenciación a largo plazo, puede durar más de un año en algunos casos (Figura 8.7D). La duración prolongada de la potenciación a largo plazo muestra que esta forma de plasticidad sináptica es capaz de servir como mecanismo para el almacenamiento prolongado de información. La potenciación a largo plazo ocurre en cada una de las tres sinapsis excitadoras del hipocampo que se muestran en la Figura 8.6, y en sinapsis de distintas regiones encefálicas, incluida la corteza, la amígdala y el cerebelo. La potenciación a largo plazo de la sinapsis de las colaterales de Schaffer muestra varias propiedades que la convierten en un mecanismo atractivo para el almacenamiento de información. Primero, la potenciación a largo plazo requiere una actividad fuerte tanto en las neuronas presinápticas como postsinápticas. Si se aparean potenciales de acción en un pequeño número de colaterales de Schaffer presinápticas –lo que induce la liberación de transmisores que producen PPSE subumbral que normalmente no darían una potenciación a largo plazo– con una despolarización fuerte de la célula CA1 postsináptica, las sinapsis de las colaterales de Schaffer activadas sufren potenciación a largo plazo (Figura 8.8). Este aumento de la transmisión sináptica ocurre solo si las actividades apareadas de las células presinápticas y postsinápticas están estrechamente ligadas en el tiempo, de modo que la despolarización postsináptica intensa ocurre dentro de los 100 ms de la liberación del neurotransmisor desde las colaterales de Schaffer. Este requerimiento de una actividad presináptica y postsináptica coincidente es el postulado central de una teoría de aprendizaje diseñada por Donald Hebb en 1949. Este autor propuso que la actividad coordinada de una terminación presináptica y una neurona postsináptica fortalecería la conexión sináptica entre ellas, precisamente como se observa para la potenciación a largo plazo. Esto indica la participación de un detector de coincidencias que permite que la potenciación a largo plazo ocurra solo cuando tanto las neuronas presináptica y postsináptica están activas. El postulado de Hebb también resultó útil para considerar el papel de la actividad neuronal en otras funciones encefálicas, principalmente el desarrollo de circuitos neurales (véase Cap. 24). 172 CAPÍTULO 8 Neuronas piramidales CA3 (A) FIGURA 8.7 Potenciación a largo plazo de las sinapsis de colaterales de Schaffer-CA1. (A) Disposición para registrar la transmisión Neuronas piramidales CA1 sináptica. Dos electrodos estimuladores (1 y 2) activan poblaciones separadas de colaterales de Schaffer, y proveen las vías sinápticas de prueba y control. (B) Izquierda. Respuestas sinápticas registradas en una neurona CA1 en respuesta a estímulos únicos de la vía sináptica 1, minutos antes y 1 hora después de un tren de estímulos de alta Registro Colaterales de Schaffer frecuencia. El estímulo de alta frecuencia aumenta el tamaño del PPSE inducido por un estímulo único. Derecha. El tamaño del PPSE producido por estimulación de la vía sináptica 2, que no recibió estimulación Estímulo 1 Po tencial de membrana PPSE (mV) (B) de alta frecuencia, no se modifica. (C) Curso temporal de los cambios Estímulo 2 en la amplitud de los PPSE inducidos por la estimulación de las vías 1 y 2. La estimulación de alta frecuencia de la vía 1 (barra más oscura) produce un refuerzo prolongado de los PPSE en esta vía (puntos vio- Vía1 Vía 2 Estímulo <50 Estímulo Después de la tetanización <55 <60 Antes de la tetanización <65 0 25 (C) 50 75 100 0 Tiempo (ms) Después de la tetanización de la vía 1 leta). Esta potenciación a largo plazo (LTP) de la transmisión sináptica en la vía 1 persiste durante varias horas, mientras que la amplitud de los PPSE producidos por la vía 2 (puntos anaranjados) sigue siendo constante. (D) Los registros de PPSE del hipocampo in vivo muestran que la estimulación de alta frecuencia puede producir una potenciación a largo plazo que dura más de un año. (A-C, de Malinow y cols., 1989; Antes de la tetanización D, de Abraham y cols., 2002.) de la vía 1 100 25 50 75 Estimulación de alta frecuencia 300 Amplitud del PPSE (% del control) Vía 1 200 Vía 2 LTP de la vía tetanizada 100 0 <15 (D) Amplitud de PPSE (% del control) 140 15 30 Tiempo (min) 45 60 Estimulación de alta frecuencia 120 100 80 0 60 120 180 Tiempo (días) 240 300 360 Una segunda propiedad que la convierte en un mecanismo nervioso atractivo para el almacenamiento de información es que la potenciación a largo plazo es específica de la aferencia: cuando es inducida por la activación de una sinapsis, no ocurre en otras sinapsis inactivas que hacen contacto con la misma neurona (véase Fig. 8.7C). Por lo tanto, la potenciación a largo plazo está limitada a las sinapsis activadas más que a todas las sinapsis sobre una célula dada (Figura 8.9A). Esta característica de la potenciación a largo plazo es compatible con su participación en la formación de memorias (o, al menos, el almacenamiento selectivo de información en las sinapsis). Si la activación de un conjunto de sinapsis condujera a la potenciación de todas las otras sinapsis –incluso a las inactivas–, sería difícil potenciar de manera selectiva conjuntos particulares de aferencias, como se requiere presumiblemente para almacenar información específica. Otra propiedad importante de la potenciación a largo plazo es la asociatividad (Figura 8.9B). Como señalamos, la estimulación débil de una vía no desencadenará por sí sola la potenciación a largo plazo. Sin embargo, si una vía es activada débilmente al mismo tiempo que una vía vecina en la misma célula es activada intensamente, ambas vías sinápticas experimentan potenciación a largo plazo. Este refuerzo selectivo de conjuntos de aferencias sinápticas activadas de manera conjunta se considera a menudo un análogo celular del aprendizaje asociativo, donde dos estímulos son necesarios para que ocurra el aprendizaje. El tipo de aprendizaje asociativo mejor Neurona piramidal CA3 Colaterales de Schaffer Neurona piramidal CA1 Registro Cambio en la amplitud del PPSE (mV) Pulsos fuertes de despolarización apareados con PPSE Prueba LTP 173 conocido es el condicionamiento clásico (de Pávlov). Más genéricamente, se espera asociatividad en cualquier red de neuronas que vincula un conjunto de información con otra. Aunque claramente existe una brecha entre conocer la potenciación a largo plazo de las sinapsis del hipocampo y conocer aprendizaje, memoria y otros aspectos de la plasticidad conductual en los mamíferos, esta forma de plasticidad sináptica a largo plazo proporciona un mecanismo plausible para los cambios neurales persistentes en una parte del encéfalo que se sabe participa en ciertos tipos de memorias. Estímulo PLASTICIDAD SINÁPTICA Mecanismos que subyacen a la potenciación a largo plazo En la actualidad, se conocen bien los fundamentos moleculares de la potenciación a largo plazo (LTP). Un adelanto clave fue el 0 10 20 30 40 50 60 descubrimiento de que los antagonistas de tipo NMDA del reTiempo (min) ceptor del glutamato impiden la potenciación a largo plazo, pero no tienen ningún efecto sobre la respuesta sináptica evocada por FIGURA 8.8 El apareamiento de la actividad presináptica y postla estimulación de baja frecuencia de las colaterales de Schaffer. sináptica produce potenciación a largo plazo. Los estímulos únicos Aproximadamente al mismo tiempo, se apreciaron por primeaplicados en una aferencia sináptica de la colateral de Schaffer inducen PPSE ra vez las propiedades bioquímicas singulares del receptor de en la neurona CA1 postsináptica. Estos estímulos aislados no producen NMDA. Como se describió en el Capítulo 6, el canal del receptor ningún cambio en la fuerza sináptica. Sin embargo, una polarización breve de NMDA es permeable al Ca2+ pero es bloqueado por Mg2+. Esta del potencial de membrana de la neurona CA1 (por aplicación de pulsos de propiedad proporciona una idea crítica sobre el modo en que la corriente a través del electrodo de registro), juntamente con los estímulos de las colaterales de Schaffer, conduce a un incremento persistente en los PPSE. potenciación a largo plazo es inducida selectivamente por la actiLTP: potenciación a largo plazo. (De Gustafsson y cols., 1987.) vidad de alta frecuencia. Durante la transmisión sináptica de baja frecuencia, el glutamato liberado por las colaterales de Schaffer se une a receptores glutamatérgicos tanto de (A) Especificida d (B) Capacidad de asociación tipo NMDA como de tipo AMPA/cainato. Aunque ambos tipos de receptores se unen a Vía 1: Vía 1: glutamato, cuando la neurona postsináptica activa estimulación fuerte se encuentra en su potencial de membrana de reposo normal, el poro del canal del reSinapsis Sinapsis ceptor del NMDA será bloqueado por iones fortalecida fortalecida Mg2+ y no habrá flujo de corriente (Figura Vía 2: Vía 2: 8.10, izquierda). Bajo estas condiciones, el inactiv a estimulación débil PPSE estará mediado completamente por los receptores de AMPA. Dado que el bloqueo Sinapsis Sinapsis del receptor de NMDA por el Mg2+ es voltajeno fortalecida no fortalecida dependiente, la función de la sinapsis cambia mucho cuando la célula postsináptica se despolariza. Por lo tanto, la estimulación de alta frecuencia (como en la Figura 8.7) producirá la suma de PPSE, que conduce a una despolarización prolongada que expulsa Mg2+ desde FIGURA 8.9 Propiedades de la potenciación a largo plazo en una neurona piramidal el poro del canal de NMDA (Figura 8.10, deCA1 que recibe aferencias sinápticas de dos conjuntos independientes de axones recha). La eliminación del Mg2+ permite que de colaterales de Schaffer. (A) Una actividad fuerte inicia la potenciación a largo plazo en las el Ca2+ entre en la neurona postsináptica, y el sinapsis activas (vía 1) sin iniciar la potenciación a largo plazo en las sinapsis inactivas cercanas (vía incremento resultante en la concentración de 2). (B) La estimulación débil de la vía 2 sola no desencadena la potenciación a largo plazo. Sin Ca2+ en el interior de las espinas dendríticas embargo, cuando el mismo estímulo débil de la vía 2 es activado juntamente con la estimulación de la célula postsináptica se conviertan en el fuerte de la vía 1, ambos conjuntos de sinapsis se fortalecen. desencadenante de la potenciación a largo 174 CAPÍTULO 8 FIGURA 8.10 El canal del receptor de NMDA se puede abrir sólo durante la despolari- En potencial de reposo Durante la despolarización postsináptica Terminación presináptica zación de la neurona postsináptica de su potencial de reposo normal. La despolarización expulsa Mg2+ del canal de NMDA, y permite que la corriente fluya hacia el interior de la célula postsináptica. Esto conduce a la entrada de Ca2+, lo que a su vez desencadena la potenciación a largo plazo (LTP). (De Nicoll y cols., 1988.) Glutamato Na+ Receptor de AMPA Terminación presináptica El Mg2+ bloquea el receptor NMDA Na+ Receptor de AMPA Receptor de NMDA Espina dendrítica de la neurona postsináptica Varios tipos de observaciones confirmaron que una elevación en la concentración de Ca2+ en la neurona CA1 postsináptica –el resultado del ingreso de Ca2+ a través de los receptores de NMDA– sirve como señal de segundo mensajero que induce la potenciación a largo plazo. Los estudios de imágenes mostraron que la activación de los receptores de NMDA produce aumentos en las concentraciones postsinápticas de Ca2+. Además, la inyección de quelantes del Ca2+ bloquea la inducción de potenciación a largo plazo, mientras que la elevación de las concentraciones de Ca2+ en las neuronas postsinápticas potencia la transmisión sináptica. En la neurona postsináptica, el Ca2+ induce una potenciación a largo plazo al activar cascadas complejas de transducción de señales que incluyen, por lo menos, dos proteincinasas activadas por Ca2+: proteincinasa dependiente de Ca2+ y calmodulina (CaMKII) y proteincinasa C (PKC; véase Cap. 7). La CaMKII, que es la proteína postsináptica más abundante en FIGURA 8.11 Actividad de CaMKII en la dendrita de una neurona piramidal CA1 durante la potenciación a largo plazo. (A) El grado de activación de CaMKII (indicado por la escala a seudocolor abajo) en una espina dendrítica aumenta en forma notable durante la estimulación que induce potenciación a largo plazo. (B) Curso temporal de cambios transitorios en la actividad de CaMKII asociado con el potenciación a largo plazo. (De Lee y cols., 2009.) 30 s después de la estimulación Dendrita 1+m Espina Alta Baja CaMKII activity Estímulo (B) Cambio en la CaMKII (% del máximo) Durante la estimulación Na+ Ca2+ LTP plazo. Por lo tanto, el receptor de NMDA se comporta como un detector de coincidencias moleculares: El canal de este receptor se abre (para inducir potenciación a largo plazo) solo cuando ocurren dos eventos de manera simultánea: el glutamato se une al receptor y la célula postsináptica es despolarizada para aliviar el bloqueo por el Mg2+ del poro del canal. Estas propiedades del receptor de NMDA pueden explicar muchas de las características de la potenciación a largo plazo. En primer lugar, el requerimiento de una actividad presináptica y postsináptica coincidente y fuerte para inducir potenciación a largo plazo (véase Fig. 8.8) surge porque la actividad presináptica libera glutamato, mientras que la despolarización postsináptica coincidente alivia el bloqueo de Mg2+ del receptor de NMDA. La especificidad de la potenciación a largo plazo (véase Fig. 8.9A) puede explicarse por el hecho de que los canales de NMDA se abrirán solo con las aferencias sinápticas que están activas y liberan glutamato, que confinan así la potenciación a largo plazo a estos sitios aun cuando los PPSE generados en las sinapsis activas despolaricen la neurona postsináptica. Con respecto a la asociatividad (véase Fig. 8.9B), una aferencia débilmente estimulada libera glutamato, pero no puede despolarizar lo suficiente la célula postsináptica como para aliviar el bloqueo de Mg2+. Sin embargo, cuando se estimulan intensamente las aferencias vecinas, proporcionan la despolarización “asociativa” necesaria para aliviar el bloqueo. Control Receptor de NMDA Na+ Na+ (A) El Mg2+ es expulsado del canal Ca2+ 100 50 0 0 1 Tiempo (min) 2 PLASTICIDAD SINÁPTICA 175 RECUADRO 8B Sinapsis silentes Corriente excitadora postsináptica (pA) (B) Varias observaciones indican que los (A) Estimulación receptores del glutamato postsinápticos 10 +m están regulados dinámicamente en las si<65 mV napsis excitadoras. Las primeras ideas sobre este proceso provinieron del hallazgo de que la estimulación de algunas sinapsis glutamatérgicas no genera ninguna señal eléctrica cuando la célula postsináptica se encuentra en su potencial de membrana 55 mV de reposo normal (Figura A). Sin embargo, una vez que la célula postsináptica es despolarizada, estas “sinapsis silentes” o silenciosas, pueden transmitir respuestas AMPA-R eléctricas postsinápticas robustas. El heNMDA-R 0 5 10 15 20 cho de que la transmisión en estas sinapAMPA-R y NMDA-R Tiempo (ms) sis pueda ser encendida o apagada según el potencial de membrana postsináptico (A) Evidencia electrofisiológica de sinapsis silentes. La estimulación de algunos axones no activa sugiere un medio interesante y simple de las sinapsis cuando la célula postsináptica se mantiene en un potencial negativo (-65 mV, trazado superior). Sin embargo, cuando la célula postsináptica es despolarizada (+55 mV), la estimulamodificar el circuito neural. Las sinapsis silenciosas son especialmen- ción produce una respuesta robusta (trazado inferior). (B) Localización inmunofluorescente de los te prevalentes durante el desarrollo y fueron receptores de NMDA (verde) y de los receptores de AMPA (rojo) en una neurona del hipocampo halladas en muchas regiones encefálicas, cultivada. Muchas espinas dendríticas son positivas para los receptores de NMDA, pero no para los incluido el hipocampo, la corteza cerebral receptores de AMPA, las cuales indican que sólo tienen sinapsis los receptores de NMDA. (A, de Liao y la médula espinal. El “silencio” de estas y cols., 1995; B, cortesía de M. Ehlers.) sinapsis se debe evidentemente al bloqueo dependiente del voltaje de los receptores sinapsis excitadoras que carecen de recep- NMDA no están tan íntimamente ligados de NMDA por el Mg2+ (véanse el texto y tores de AMPA, sino más bien una clase a las sinapsis excitadoras, sino que están Cap. 6). En el potencial de membrana de temprana en la maduración oncogénica de dirigidos a través de mecanismos celulares reposo normal, la liberación presináptica la sinapsis glutamatérgica (Figura D). Evi- independientes. Esta composición de los de glutamato no produce ninguna respuesta dentemente, los receptores de AMPA y de receptores glutamatérgicos específicos de postsináptica en estas sinapsis porque sus (Continúa en la página siguiente) receptores de NMDA están bloqueados por 2+ el Mg . Sin embargo, la despolarización de Receptor de NMDA AMPA-P + NMDA-R (D) la neurona postsináptica desplaza el Mg2+, y (C) NMDA-R de la sinapsis de la sinapsis esto permite que la liberación de glutamato Juvenil silente solamente funcional induzca respuestas postsinápticas mediadas Pre Terminación Maduración por los receptores de NMDA. presináptica Post El glutamato liberado en las sinapsis Hendidura Adulto sináptica silenciosas evidentemente solo se une a Pre receptores de NMDA. Entonces, ¿de qué Post modo la liberación de glutamato evita acNMDA-R tivar a los receptores de AMPA? La expliAMPA-R cación más probable es que estas sinapsis 0.1 +m pueden tener solo receptores de NMDA. Espina postsináptica NMDA-R NMDA-R AMPA-R Algunos experimentos inmunocitoquímicos demuestran que algunas sinapsis excitadoras tienen solo receptores de NMDA (C) Microscopia electrónica de sinapsis excitadoras en el estrato radiado CA1 del hipocampo de (puntos verdes en la Figura B). Estas si- ratas de 10 días de vida (juveniles) o de 5 semanas de vida (adultas) con doble marcación para napsis que solo cuentan con receptores receptores de AMPA y receptores de NMDA. Está indicada la terminación presináptica (pre), la de NMDA son particularmente abundan- hendidura sináptica y la espina postsináptica (post). Los receptores de AMPA son abundantes en la tes al comienzo del desarrollo posnatal y sinapsis del adulto, pero están ausentes en la sinapsis de los más jóvenes. (D) Diagrama de madudisminuyen en los adultos (Figura C). Por ración de la sinapsis glutamatérgica. Al comienzo del desarrollo posnatal, muchas sinapsis excitadolo tanto, al menos algunas sinapsis silen- ras contienen solo receptores de NMDA. A medida que las sinapsis maduran, se reclutan receptores tes no representan una clase separada de de AMPA. (C, de Petralia y cols., 1999.) 176 CAPÍTULO 8 RECUADRO 8B (continúa) la sinapsis implica mecanismos sofisticados para regular la localización de cada tipo de receptor. Los cambios dinámicos en el tráfico de receptores de AMPA y de NMDA pueden fortalecer o debilitar la transmisión sináptica y son importantes en la potenciación a largo plazo y en la depresión a largo plazo, así como en la maduración de las sinapsis glutamatérgicas. Aunque las sinapsis silentes han comenzado a murmurar sus secretos, queda mucho por aprender sobre su importancia fisiológica y sobre los mecanismos moleculares que median el reclutamiento rápido o la eliminación de receptores de AMPA sinápticos. Bibliografía Derkach, V. A., M. C. Oh, E. S. Guire and T. R. Soderling (2007) Regulatory mechanisms of AMPA receptors in synaptic plasticity. Nature Rev. Neurosci. 8:101-113. Gomperts, S. N., A. Rao, A. M. Craig, R. C. Malenka and R. A. Nicoll (1998) Postsynaptically las sinapsis de las colaterales de Schaffer, parece desempeñar un papel especialmente importante. Esta enzima es activada durante los estímulos que inducen potenciación a largo plazo (Figura 8.11), y la inhibición farmacológica o la deleción genética de CaMKII previenen la potenciación a largo plazo. Los puntos diana corriente abajo de estas cinasas, que no se conocen aún completamente, al parecer incluyen receptores de AMPA y muchas otras proteínas de señalamiento. Al parecer el fortalecimiento de la transmisión sináptica durante la potenciación a largo plazo se origina en un incremento en la sensibilidad de la célula postsináptica al glutamato. Varias observaciones recientes indican que las sinapsis excitadoras pueden regular dinámicamente sus receptores postsinápticos de glutamato e incluso pueden agregar nuevos receptores de AMPA a las sinapsis “silentes” que no los tenían antes (Recuadro 8B). La “expresión” o mantenimiento de la potenciación a largo plazo aparentemente se debe a esta inserción de receptores de AMPA en la membrana postsináptica (por oposición a la “inducción” de potenciación a largo plazo, que se basa en la activación de receptores de NMDA). El incremento resultante en la densidad de los receptores FIGURA 8.12 Agregado de receptores de (A) Antes de la LTP Después de la LTP 20 pA Espina 10 Dendrita 5 0 1 µm (B) (C) 180 Estimulación pA 160 40 Antes de la LTP 20 140 0 120 40 pA Respuesta del glutamato (% del control) silent synapses in single neuron cultures. Neuron 21: 1443-1451. Liao, D., N. A. Hessler and R. Malinow (1995) Activation of postsynaptically silent synapses during pairing-induced LIP in CA1 region of hippocampal slice. Nature 375: 400-404. Luscher, C. R. A. Nicoll, R. C. Malenka and D. Muller (2000) Synaptic plasticity and dynamic modulation of the postsynaptic membrane. Nature Neurosci. 3: 545-550. Petralia, R. S. and six others (1999) Selective acquisition of AMPA receptors over postnatal development suggests a molecular basis for silent synapses. Nature Neurosci. 2: 31-36. 100 Induce LTP 80 –20 0 Después de la LTP 20 20 40 Tiempo (min) 60 80 0 0 10 20 30 40 50 60 70 80 90 100 Tiempo (ms) AMPA postsinápticos durante la potenciación a largo plazo. (A) Mapas espaciales de la sensibilidad al glutamato de una dendrita de la neurona del hipocampo antes y después de 120 minutos de la inducción de la potenciación a largo plazo (LTP). La escala en color indica la amplitud de las respuestas a la aplicación de glutamato sumamente localizada. La potenciación a largo plazo produce un aumento de la respuesta de una espina dendrita al glutamato debido a un incremento en la cantidad de receptores de AMPA en la membrana de la espina. (B) Curso temporal de los cambios en la sensibilidad al glutamato de las espinas dendríticas durante la potenciación a largo plazo. La inducción de potenciación a largo plazo en el tiempo = 0 hace que aumente la sensibilidad al glutamato durante más de 60 minutos. (C) La potenciación a largo plazo induce respuestas de los receptores de AMPA en las sinapsis silentes del hipocampo. Antes de inducir potenciación a largo plazo, no se obtienen PPSE en -65 mV en esta sinapsis silente (trazado superior). Después de inducir potenciación a largo plazo, el mismo estímulo produce PPSE que están mediados por receptores de AMPA (trazado inferior). (A, B, de Matsuzaki y cols., 2004; C, de Liao y cols., 1995.) PLASTICIDAD SINÁPTICA Glutamato Terminación presináptica Na+ Receptores de AMPA Ca2+ Na+ Se insertan receptores de AMPA adicionales Ca2+/ calmodulina cinasa II Proteincinasa C Fosforilación del sustrato Espina dendrítica de la neurona postsináptica de AMPA en la espina postsináptica incrementa la respuesta de la célula postsináptica al glutamato liberado (Figura 8.12A), lo que arroja el fortalecimiento de la transmisión sináptica que puede durar mientras se mantenga la potenciación a largo plazo (Figura 8.12B). En las sinapsis silentes, donde la actividad sináptica no genera ninguna respuesta postsináptica en el potencial de reposo normal, la potenciación a largo plazo agrega receptores de AMPA de modo que la sinapsis puede producir respuestas postsinápticas (Figura 8.12C). En resumen, las vías de señalamiento molecular involucradas en la potenciación a largo plazo en la sinapsis colateral de Schaffer-CA1 son bien conocidas. El Ca2+ que ingresa a través de los receptores de NMDA postsinápticos conduce a la activación de proteincinasas que regulan el tráfico de los receptores de AMPA, por lo que aumenta así la respuesta postsináptica al glutamato liberado de la terminación presináptica (Figura 8.13). En algunas (A) (B) Control Amplitud de los PPSE (% del control) 250 de glutamato, el canal de NMDA se abre sólo cuando la célula postsináptica está suficientemente despolarizada. Los iones Ca2+ que entran en la célula a través del canal activan proteincinasas postsinápticas como CaMKII y PKC. Estas cinasas postsinápticas desencadenan una serie de reacciones que conducen a la inser- circunstancias, la potenciación a largo plazo también puede aumentar la capacidad de las terminaciones presinápticas para liberar glutamato. Como la potenciación a largo plazo es desencadenada claramente por las acciones del Ca2+ en el interior de la neurona postsináptica, esta potenciación de la función presináptica requiere la propagación de una señal retrógrada (tal vez NO; véase Cap. 6) desde la espina postsináptica nuevamente hasta las terminaciones presinápticas. Se observan aún otras formas de potenciación a largo plazo en otras sinapsis y, en algunos casos, se basan en mecanismos de señalización diferentes de los involucrados en la potenciación a largo plazo en la sinapsis colateral de Schaffer-CA1. El esquema de la Figura 8.13 puede explicar los cambios en la transmisión sináptica que ocurren en la primera o dos primeras horas después de inducir la potenciación a largo plazo. Sin embargo, también existe una fase más tardía que depende de los cambios en la expresión genética y de la síntesis de nuevas proteínas. Las contribuciones de esta fase tardía pueden observarse tratando sinapsis con agentes que inhiben la síntesis proteica: el bloqueo de la síntesis previene la potenciación a largo plazo medida varias horas después de un estímulo, pero no afecta la medida en los primeros momentos (Figura 8.14). Esta fase tardía de la potenciación a largo plazo parece ser iniciada por la proteincinasa A, que continúa hasta activar los factores de transcripción como CREB, que estimulan la expresión de otras proteínas. Aunque la mayoría de estas proteínas recién sintetizadas no fueron identificadas aún, incluyen otros reguladores transcripcionales, proteincinasas y receptores de AMPA (Figura 8.15A). No se sabe aún el modo en que estas proteínas contribuyen a la fase tardía de la potenciación a largo plazo. Existen pruebas de que el Se agrega inhibidor de la síntesis de proteínas 300 200 FIGURA 8.14 Papel de la síntesis de proteínas en el mantenimiento de la potenciación a largo plazo. 250 (A) La estimulación repetitiva a alta frecuencia (flecha) induce la potenciación a largo plazo que persiste durante muchas horas. (B) El tratamiento con anisomicina, un inhi- 200 150 150 100 100 50 FIGURA 8.13 Mecanismos de señalización que subyacen a la potenciación a largo plazo. Durante la liberación ción de nuevos receptores de AMPA en la espina postsináptica y a un aumento de la sensibilidad de la espina al glutamato. Ca2+ Receptor de NMDA 50 0 4 2 6 Tiempo (horas) 8 177 0 2 4 6 Tiempo (horas) 8 bidor de la síntesis proteica (en la barra), produce la caída de la potenciación a largo plazo algunas horas después de la estimulación de alta frecuencia (flecha). (De Frey y Morris, 1997.) 178 CAPÍTULO 8 Fase inicial (A) FIGURA 8.15 Mecanismos responsables de los cambios duraderos en la transmisión si- Fase tardía Ca2+ Receptor de AMPA náptica durante la potenciación a largo plazo. (A) El componente tardío de la potenciación a largo plazo (LTP) es el resultado de la PKA, la cual activa al regulador transcripcional CREB, que activa la expresión de algunos genes que producen Terminación presináptica Receptor de NMDA Receptor de AMPA Ca2+ Receptor de NMDA Proteínas de crecimiento de la sinapsis Ca2+/ calmodulina Espinas dendríticas de la neurona postsináptica Proteincinasas ATP cAMP Reguladores transcripcionales CREB cambios duraderos en la actividad de PKA y en la estructura de las sinapsis. (B,C) Cambios estructurales asociados con la potenciación a largo plazo en el hipocampo. (B) Se visualizaron las dendritas de una neurona piramidal CA1 mediante el relleno de la célula con un colorante fluorescente. (C) Se pueden observar nuevas espinas dendríticas (flechas blancas) aproximadamente 1 hora después de un estímulo que induce potenciación a largo plazo. La presencia de nuevas espinas plantea la posibilidad de que la potenciación a largo plazo pueda surgir, en parte, de la formación de nuevas sinapsis. (A, de Squire y Kandel, 1999; B y C, de Engert y Bonhoeffer, 1999.) Proteincinasa A (B) (C) Vista superior Vista lateral LTP centes a la sensibilización del comportamiento en Aplysia. Ambos consisten en una fase transitoria temprana que se basa en las proteincinasas para producir cambios postraduccionales en los canales de membrana, y ambos tienen fases prolongadas más tardías que requieren cambios en la expresión genética mediados por CREB. Ambas formas de plasticidad sináptica a largo plazo probablemente estén involucradas en la escasez prolongada de información, aunque no se estableció firmemente el papel de la potenciación a largo plazo en el almacenamiento de la memoria en el hipocampo. Mecanismos subyacentes a la depresión a largo plazo 50 +m número y el tamaño de los contactos sinápticos aumentan durante la potenciación a largo plazo (Figura 8.15B, C). Por lo tanto, es probable que algunas de las proteínas recién sintetizadas durante la fase tardía de potenciación a largo plazo estén involucradas en la construcción de nuevos contactos sinápticos que sirven para volver la potenciación a largo plazo esencialmente permanente. En conclusión, al parecer, la potenciación a largo plazo en el hipocampo de los mamíferos tiene muchos puntos paralelos con los cambios a largo plazo de la transmisión sináptica subya- Si simplemente las sinapsis siguen aumentando en fuerza como resultado de la potenciación a largo plazo, finalmente alcanzarían cierto nivel de eficacia máxima, lo que hace difícil la codificación de nueva información. Por lo tanto, para que el fortalecimiento sináptico sea útil, otros procesos deben debilitar de modo selectivo conjuntos específicos de sinapsis, y la depresión a largo plazo es uno de estos procesos. A fines de la década de 1970, se observó que ocurría depresión a largo plazo (LTD) en las sinapsis entre las colaterales de Schaffer y las células piramidales CA1 en el hipocampo. Mientras la potenciación a largo plazo en estas sinapsis requiere una estimulación breve de alta frecuencia, la depresión a largo plazo ocurre cuando las colaterales de Schaffer son estimuladas a baja frecuencia –aproximadamente, 1 Hz– durante PLASTICIDAD SINÁPTICA (A) 179 Célula piramidal CA3 Colaterales de Schaffer Amplitud de los PPSE (% del control) (B) Célula piramidal CA1 Registro 150 100 LTD Estímulo de 1 Hz 50 0 Estímulo 15 30 45 Tiempo (min) 60 75 FIGURA 8.16 Depresión sináptica a largo plazo en el (C) Glutamato Terminación presináptica hipocampo. (A) Procedimientos electrofisiológicos utilizados para controlar la transmisión en las sinapsis de las colaterales de Schaffer sobre las neuronas piramidales CA1. (B) La estimulación de baja frecuencia (1 por segundo) de los axones de las colaterales de Schaffer produce una depresión a largo plazo (LTD) de la transmisión sináptica. (C) Mecanismos que subyacen a la depresión a largo plazo. Una elevación de baja amplitud en la Na+ Receptores de AMPA Ca2+ Receptor de NMDA Ca2 Internalización de los receptores de AMPA de las colaterales de Schaffer. (B, de Mulkey y cols., 1993.) + Na+ Proteinfosfatasas Espina dendrítica de la neurona postsináptica concentración de Ca2+ en la neurona CA1 postsináptica activa las proteinfosfatasas postsinápticas, que producen la internalización de los receptores de AMPA postsinápticos, por lo que disminuye así la sensibilidad al glutamato liberado desde las terminaciones Defosforilan sustratos períodos prolongados (10-15 minutos). Este patrón de actividad deprime el PPSE durante varias horas y, al igual que la potenciación a largo plazo, es específico de las sinapsis activadas (Figura 8.16A, B). Más aun, la depresión a largo plazo puede borrar el incremento en el tamaño de los PPSE debido a la potenciación a largo plazo y, por el contrario, esta última puede borrar la disminución del tamaño de los PPSE debido a la depresión a largo plazo. Esta complementariedad sugiere que la depresión a largo plazo y la potenciación a largo plazo afectan inversamente la eficiencia sináptica al actuar en un sitio común. En realidad, tanto la potenciación como la depresión a largo plazo en las sinapsis colaterales de Schaffer-CA1 comparten varios elementos clave. Ambas requieren la activación de los receptores de los glutamatos de tipo NMDA y el ingreso resultante del Ca2+ en la célula postsináptica. El principal determinante de que surja una potenciación o una depresión a largo plazo parece ser la naturaleza de la señal de Ca2+ en la célula postsináptica: elevaciones pequeñas y lentas en el Ca2+ conducen a la depresión, mientras que los incrementos grandes y rápidos en el Ca2+ desencadenan la potenciación. Como señalamos antes, la poten- ciación a largo plazo se debe, al menos parcialmente, a la activación de las proteincinasas, que fosforilan sus proteínas diana. Por otra parte, la depresión a largo plazo parece ser el resultado de la activación de fosfatasas Ca2+-dependientes, que dividen los grupos fosfato de sus moléculas diana (véase Cap. 7). La evidencia que apoya esta idea es que los inhibidores de la fosfatasa previenen la depresión a largo plazo, pero no bloquean la potenciación a largo plazo. Los diferentes efectos del Ca2+ durante ambas pueden originarse en la activación selectiva de proteinfosfatasas y cinasas por diferentes tipos de señales de Ca2+ Aunque no se identificaron aún los sustratos de las fosfatasas que son importantes en la depresión a largo plazo, es posible que esta y la potenciación a largo plazo fosforilen y desfosforilen el mismo conjunto de proteínas reguladoras para controlar la eficacia de la transmisión en la sinapsis colateral de Schaffer-CA1. Al igual que la potenciación a largo plazo, en esta sinapsis, se asocia con la inserción de receptores de AMPA, la depresión a largo plazo se asocia a menudo con una pérdida de receptores de AMPA sinápticos. Esta pérdida probablemente surge por la internalización de receptores de AMPA en la célula postsináptica (Figura 8.16C), debido al mismo tipo de mecanismos de endocitosis dependiente de clatrina importantes para el reciclado de las vesículas sinápticas en la terminación presináptica (véase el Cap. 5). Se observa una forma muy diferente de depresión a largo plazo en el cerebelo. La depresión a largo plazo de las aferencias sinápticas sobre las células de Purkinje del cerebelo fue descrita por primera vez por Masao Ito y cols. en Japón a comienzos de la década de 1980. Las neuronas de Purkinje del cerebelo reciben 180 CAPÍTULO 8 (A) Espinas dendríticas Fibras paralelas (C) Fibra trepadora Espina dendrítica de la célula de Purkinje Fibras paralelas Sinapsis debilitada La fibra trepadora se despolariza Vm Axón de la célula grano (D) Terminación presináptica de la fibra paralela Registro Cuerpo de la célula de Purkinje Célula grano Receptores de AMPA Glutamato mGluR Axón Fibra trepadora Estimulación de la fibra trepadora Fibras paralelas Amplitud del PPSE (mV ) (B) Internalización de los receptores de AMPA Depresión a largo plazo PIP2 Fosforilan proteínas sustrato Par fibra paralela y fibra trepadora Fosfolipasa C DAG PKC 7 6 Ca2+ 5 La fibra trepadora se despolariza en V 4 Espina dendrítica de la célula de Purkinje Se libera Ca2+ 3 0 IP3 −20 0 20 40 Tiempo (min) 60 80 Ca2+ Retículo endoplasmático FIGURA 8.17 Depresión a largo plazo en el cerebelo. (A) Disposición experimental. Se registraron las respuestas sinápticas de las células de Purkinje después de la estimulación de las fibras paralelas y de las fibras trepadoras. (B) El apareamiento de la estimulación de las fibras trepadoras (CF) y de las fibras paralelas (PF) produce depresión a largo plazo, que reduce el PPSE de las fibras paralelas. (C) La depresión a largo plazo requiere la despolarización de la célula de Purkinje, producida por la activación de las fibras trepadoras, y señales generadas por las sinapsis activas con las fibras paralelas. (D) Mecanismo subyacente a la depresión a largo plazo cerebelosa. El glutamato liberado por las fibras paralelas activa tanto los receptores de AMPA como los receptores glutamatérgicos metabotrópicos (mGluRs). Los mGluRs activados producen dos segundos mensajeros, DAG e IP3, que interactúan con Ca2+ que entra cuando la actividad de las fibras trepadoras abre los canales de Ca2+ con puerta de voltaje. La liberación resultante de Ca2+ desde el retículo endoplasmático conduce a un mayor incremento de la concentración intracelular de Ca2+ y a la activación de la PKC; esto desencadena la internalización dependiente de clatrina de los receptores de AMPA postsinápticos, lo que debilita las sinapsis de las fibras paralelas. (B, de Sakurai, 1987.) dos tipos distintos de aferencias excitadoras: fibras trepadoras y fibras paralelas (Figura 8.17A; véase Cap. 19). La depresión a largo plazo reduce la fuerza de la transmisión en la sinapsis de las fibras paralelas (Figura 8.17B) y recientemente se demostró que deprime la transmisión también en la sinapsis de las fibras trepadoras. Esta forma de depresión a largo plazo ha sido implicada en el aprendizaje motor que media la coordinación, la adquisición y el almacenamiento de movimientos complejos en el interior del cerebelo. Si bien el papel de la depresión a largo plazo en el aprendizaje motor cerebeloso sigue siendo controvertido, ha sido un sistema modelo útil para comprender los mecanismos moleculares de la plasticidad sináptica prolongada. La depresión a largo plazo cerebelosa es asociativa porque ocurre solo cuando las fibras trepadoras y las fibras paralelas son activadas al mismo tiempo (Figura 8.17C). En este caso, la asociatividad surge por las acciones combinadas de dos vías distintas de transducción de señales que son activadas en la célula de Purkinje postsináptica debido a la actividad de las si- Potencial de membrana postsináptico (mV) +40 0 –60 0 (B) Pre Post 20 40 60 Tiempo (ms) Potencial de membrana postsináptico (mV) Estimulación postsináptica antes de la presináptica +40 0 –60 Post Pre 0 20 40 60 Tiempo (ms) Amplitud de los PPSE (% del control) Estimulación presináptica antes de la postsináptica (A) Amplitud de los PPSE (% del control) PLASTICIDAD SINÁPTICA 180 LTP 100 Pre + post –20 –10 0 10 Tiempo (min) Amplitud de los PPSE (% del control) 20 30 Estímulo potenciación a largo plazo (LTP) del PPSE. (B) La plazo (LTD) del PPSE. (C) La dependencia compleja de la plasticidad sináptica del momento de la espiga depende del intervalo entre la actividad presináptica y la actividad postsináptica. Si la neurona presináptica se activa 40 ms o menos antes que la neurona 140 100 LTD Post + pre 60 un potencial de acción que se sobreagrega al PPSE. El gráfico de la derecha muestra que la aplicación repetitiva de este paradigma de estímulo produce inversión en el orden de la estimulación, de modo que se excite la neurona postsináptica antes que la neurona presináptica, produce depresión a largo 180 –20 –10 0 10 Tiempo (min) 20 30 postsináptica, ocurre la potenciación a largo plazo. Por el contrario, cuando la neurona postsináptica es activada 40 ms o menos antes que la neurona presináptica, ocurre la depresión a largo plazo. Si el intervalo entre los dos eventos es mayor de 40 ms, no se observa plasticidad sináptica dependiente del momento de la espiga. (De Bi y Poo, 1998.) 100 Post antes que pre Pre antes que post 60 40 20 0 –20 –40 –60 nas de hipocampo cultivadas. (A) El gráfico de la izquierda muestra que la estimulación de una neurona presináptica (Pre) produce un PPSE en la neurona postsináptica; la aplicación de un estímulo posterior en la neurona postsináptica (Post) produce 140 (C) 80 FIGURA 8.18 Plasticidad sináptica dependiente del momento de la espiga en neuro- Estímulo 60 181 –80 –40 40 80 0 Intervalo entre la actividad presináptica y la postsináptica (ms) napsis de las fibras trepadoras y paralelas. En la primera vía, el glutamato liberado de las terminaciones de las fibras paralelas se activa en dos tipos de receptores, el tipo AMPA y los receptores glutamatérgicos metabotrópicos (véase Cap. 6). La unión del glutamato al receptor de AMPA conduce a la despolarización de la membrana, mientras que la fijación al receptor metabotrópico produce los segundos mensajeros trifosfato de inositol (IP3) y diacilglicerol (DAG) (véase Cap. 7). La segunda vía de transducción de señales, iniciada por la activación de las fibras trepadoras, produce un influjo de Ca2+ a través de los canales con puerta de voltaje y un incremento posterior en la concentración intracelular de Ca2+. Estos segundos mensajeros funcionan juntos para producir una elevación amplificada de la concentración intracelular de Ca2+, debido a que el IP3 y el Ca2+ actúan juntos sobre los receptores IP3, que desencadena la libe- ración de Ca2+ de los depósitos intracelulares, y la activación sinérgica de PKC por Ca2+ y DAG (Figura 18.7D). Por lo tanto, la propiedad asociativa de la depresión a largo plazo cerebelosa parece surgir tanto de los receptores IP3 como de la PKC que sirven como detectores de coincidencia. Aunque las proteínas sustratos de la PKC corriente abajo no están determinadas todavía, al parecer los receptores de AMPA representan una de las proteínas fosforiladas por la PKC. La consecuencia de la activación de la PKC es producir una internalización de los receptores de AMPA a través de una endocitosis dependiente de clatrina (Figura 8.17D). Esta pérdida de los receptores de AMPA disminuye la respuesta de la célula de Purkinje postsináptica al glutamato liberado desde las terminaciones presinápticas de las fibras paralelas. Por lo tanto, al contrario de la depresión a largo plazo del hipocampo, la depresión a largo plazo cerebelosa necesita la actividad de una proteincinasa, más que una fosfatasa, y no involucra la entrada de Ca2+ a través del tipo de NMDA de receptor glutamatérgico (que no está presente en las células de Purkinje maduras). Sin embargo, el efecto neto es el mismo en ambos casos: la internalización de los receptores de AMPA es un mecanismo frecuente para la menor eficacia de las sinapsis del hipocampo y cerebelosas durante la depresión a largo plazo. Como sucede con la potenciación a largo plazo en la sinapsis de las colaterales de Schaffer del hipocampo, y para la plasticidad sináptica prolongada en Aplysia, el CREB parece ser necesario para una fase tardía de depresión a largo plazo cerebelosa. No se sabe aún qué proteínas son sintetizadas como consecuencia de la activación del CREB. 182 CAPÍTULO 8 Plasticidad dependiente del momento de la espiga tica (Figura 8.18A, B). La relación entre el intervalo temporal y la magnitud del cambio sináptico es una función muy sensible del intervalo temporal, sin que se observen cambios si las actividades presináptica y postsináptica están separadas por 100 milisegundos o más (Figura 8.18C). Como el momento preciso de la actividad presináptica y postsináptica determina la polaridad de estas formas de plasticidad sináptica a largo plazo, se las denomina plasticidad dependiente del momento de la espiga (STDP). Aunque no se conocen bien aún los mecanismos involucrados, al parecer las propiedades de este tipo de plasticidad se originan en las diferencias en las señales de Ca2+ postsinápticas dependientes del tiempo. Cuando específicamente un potencial de acción postsináptico ocurre después de la actividad presináptica, la despolarización resultante aliviará el bloqueo de los receptores de NMDA por el Mg2+ y producirá una cantidad relativamente grande de influjo Las secciones anteriores muestran que la potenciación a largo plazo y la depresión a largo plazo son iniciadas preferentemente por diferentes velocidades de actividad sináptica repetitiva: la potenciación a largo plazo requiere actividad de alta frecuencia y la depresión es inducida por la actividad de baja frecuencia. Sin embargo, la relación temporal precisa entre la actividad en las células presinápticas y postsinápticas también es un determinante importante de la cantidad y de la dirección de la plasticidad sináptica a largo plazo. En una frecuencia dada (baja) de actividad sináptica, la depresión a largo plazo ocurrirá si la actividad presináptica es precedida por un potencial de acción postsináptico, mientras la potenciación a largo plazo ocurre si el potencial de acción postsináptico sigue a la actividad presináp- RECUADRO 8C Epilepsia: efecto de la actividad patológica sobre el circuito neural a los de la epilepsia humana. Las manifestaciones conductuales de las crisis epilépticas en los seres humanos varían desde la contracción leve de una extremidad hasta la pérdida de conocimiento y las convulsiones incontrolables. Aunque muchas personas muy Inicio de la convulsión Fp2 – F4 F 4 – C4 Posición del electrodo de registro La epilepsia es un trastorno encefálico caracterizado por crisis periódicas e impredecibles mediadas por la descarga rítmica de grandes grupos de neuronas. Parece probable que la actividad anormal genera cambios plásticos en el circuito cortical que son críticos para la patogenia de la enfermedad. La importancia de la plasticidad neuronal en la epilepsia está indicada más claramente por un modelo animal de producción de crisis denominado autoencendido (kindling). Para inducirlo, se implanta un electrodo estimulador en el encéfalo, a menudo en la amígdala (componente del sistema límbico que hace y recibe conexiones con la corteza, el tálamo y otras estructuras límbicas, incluido el hipocampo; véase Cap. 29). Al comienzo de este experimento, la estimulación eléctrica leve, en forma de un tren de baja amplitud de impulsos eléctricos, no tiene ningún efecto discernible sobre el comportamiento del animal ni sobre el patrón de actividad eléctrica en el encéfalo (se utilizaron típicamente ratas o ratones de laboratorio para estos estudios). A medida que esta estimulación débil es repetida una vez al día durante varias semanas, comienza a producir indicaciones conductuales y eléctricas de crisis. Este fenómeno es esencialmente permanente; incluso después de un intervalo de un año, el mismo estímulo débil desencadenará nuevamente una crisis. Por lo tanto, la estimulación débil repetitiva produce cambios a largo plazo en la excitabilidad del encéfalo que el tiempo no puede revertir. Por lo tanto, la palabra autoencendido es muy apropiada: un único fósforo puede iniciar un incendio devastador. Los cambios en los patrones eléctricos de la actividad encefálica detectados en los animales con autoencendido se asemejan Electroencefalograma (EEG) registrado durante una convulsión. Los trazados muestran actividad rítmica que persistió mucho más que la duración de este registro. Este patrón anormal refleja la descarga sincrónica de gran cantidad de neuronas corticales (las designaciones son distintas posiciones de los electrodos sobre la cabeza; véase Recuadro 28C). (De Dyro, 1989.) C 4 – P4 P4 – O 2 FP1 – F3 F 3 – C3 C 3 – P3 P3 – O 1 0 1 2 Tiempo (s) 3 4 PLASTICIDAD SINÁPTICA de Ca2+ a través de los receptores de NMDA presinápticos, lo que produce una potenciación a largo plazo. Por el contrario, cuando el potencial de acción postsináptico ocurre antes del potencial de acción presináptico, la despolarización asociada al potencial de acción postsináptico cederá para el momento en que ocurre el PPSE. Esta secuencia de acontecimientos reduce la cantidad de entrada de Ca2+ a través de los receptores de NMDA, lo que conduce a la depresión a largo plazo. Se ha postulado que otras señales, como los endocanabinoides (véase Cap.6), también pueden ser necesarios para la inducción de la depresión a largo plazo durante la plasticidad dependiente del momento de la espiga. El requerimiento de una relación temporal precisa entre la actividad presináptica y postsináptica significa que la plasticidad dependiente del momento de la espiga puede realizar varios tipos nuevos de computación neuronal. Esta plasticidad puede proporcionar un medio para codificar la información 183 sobre causalidad. Por ejemplo, si una sinapsis genera un PPSE supraumbral, el potencial de acción postsináptico resultante seguirá rápidamente a la actividad presináptica y la potenciación a largo plazo resultante codificaría el hecho de que el potencial de acción postsináptico fuera el resultado de la actividad de esa sinapsis. La plasticidad dependiente del momento de la espiga también podría servir como mecanismo para la competencia entre las aferencias sinápticas: es más probable que las aferencias más fuertes generen PPSE supraumbrales y que sean reforzadas por la potenciación a largo plazo resultante, mientras que las aferencias más débiles no generarían potenciales de acción postsinápticos correlacionados con la actividad presináptica. Existen pruebas de que la plasticidad dependiente del momento de la espiga es importante para la función del circuito neural, como la determinación de mapas de orientación en el sistema visual (véase Cap. 12). RECUADRO 8C (continúa) destacadas sufrieron epilepsia (Alejandro Magno, Julio César, Napoleón, Dostoievski y Van Gogh, por nombrar solo algunos), obviamente las crisis de suficiente intensidad y frecuencia pueden interferir en muchos aspectos de la vida cotidiana. Más aún, las convulsiones incontrolables pueden conducir a excitotoxicidad (véase Recuadro 6C). Está afectada hasta un 1% de la población, lo que convierte a la epilepsia en uno de los problemas neurológicos más frecuentes. El pensamiento moderno sobre las causas (y posibles curaciones) de la epilepsia se concentra en dónde se originan las crisis y en los mecanismos que vuelven hiperexcitable la región afectada. La mayor parte de la evidencia sugiere que la actividad anormal en áreas pequeñas de la corteza cerebral (denominados “focos”) proporciona los desencadenantes para una crisis que luego se propaga a otras regiones conectadas a través de sinapsis. Por ejemplo, una crisis que se origina en el área para el pulgar de la corteza motora derecha se apreciará primero como un movimiento descontrolado del pulgar izquierdo que posteriormente se extiende a otros músculos más proximales de la extremidad, mientras que una crisis que se origina en la corteza de asociación visual del hemisferio derecho puede ser anunciada por alucinaciones complejas en el campo visual izquierdo. Por lo tanto, las manifestaciones conductuales de las crisis proporcionan indicios importantes para el neurólogo que busca señalar la región anormal de la corteza cerebral. Las crisis epilépticas pueden ser causadas por distintos factores adquiridos o congénitos, que incluyen el daño cortical por traumatismo, accidente cerebrovascular, tumores, disgenesia cortical congénita (falta de crecimiento apropiado de la corteza) y malformaciones vasculares congénitas. Una forma rara de epilepsia, la encefalitis de Rasmussen, es una enfermedad autoinmunitaria que aparece cuando el sistema inmunitario ataca el encéfalo, y utiliza tanto agentes humorales (es decir, anticuerpos) como celulares (linfocitos y macrófagos) que pueden destruir las neuronas. Algunas formas de epilepsia son hereditarias y se han demostrado más de una docena de genes que subyacen a tipos inusuales de epilepsia. Sin embargo, la mayoría de las formas de epilepsia familiar (como la epilepsia mioclónica juvenil y el pequeño mal) son causadas por la herencia simultánea de más de un gen mutante. No existe prevención ni curación efectiva de la epilepsia. Los tratamientos farmacológicos que inhiben con éxito las crisis se basan en dos estrategias generales. Un abordaje es aumentar la función de las sinapsis inhibidoras que utilizan el neurotransmisor GABA; el otro es limitar la descarga de los potenciales de acción que actúan sobre los canales del Na+ con puerta de voltaje. Las medicaciones anticomiciales de uso habitual incluyen carbamazepina, fenobarbital, fenitoína (Dilantin) y ácido valproico. Estos agentes, que deben ingerirse diariamente, inhiben con éxito las crisis en el 60-70% de los pacientes. En una pequeña fracción de ellos, la región epileptógena puede ser resecada quirúrgicamente. En los casos extremos, los médicos recurren al corte del cuerpo calloso para prevenir la propagación de las crisis (principalmente de los individuos con “encéfalo dividido” descritos en el Capítulo 27 que padecen epilepsia intratable). Una de las principales razones para controlar las crisis epilépticas es prevenir los cambios plásticos más permanentes que aparecen como consecuencia de la actividad neural anormal y excesiva. Bibliografía Engel, J. Jr. and T. A. Pedley et al. (2008) Epilepsy: A Comprehensive Textbook, 2nd Ed. (3 volumes). Philadelphia: Lippincott-Raven McNamara, J. O., Y. Z. Huang and A. S. Leonard (2006) Molecular signaling mechanisms underlying epileptogenesis. Science’s Signal Transduction Knowledge Environment 356: rel2. Scheffer, I. E. and S. F. Berkovic (2003) The genetics of human epilepsy. Trends Pharm. Sci. 24: 428-433. Reid, C. A., S. F. Berkovic and S. Petrou (2009) Mechanisms of human inherited epilepsies. Prog. Neurobiol. 87: 41-57. 184 CAPÍTULO 8 En resumen, las formas de plasticidad sináptica dependientes de la actividad producen cambios en la transmisión sináptica que modifican las conexiones funcionales dentro de los circuitos neurales y entre ellos. Estos cambios de la eficacia y la geometría local de la conectividad sináptica pueden proporcionar una base para el aprendizaje, la memoria y otras formas de plasticidad encefálica. Los cambios en la transmisión sináptica dependientes de la actividad también pueden participar en algunas patologías. Patrones anormales de actividad neuronal, como los que ocurren en la epilepsia, pueden estimular cambios anormales en las conexiones sinápticas que pueden aumentar más la frecuencia y la gravedad de las convulsiones (Recuadro 8C). A pesar de los adelantos sustanciales en el conocimiento de las bases celulares y moleculares de algunas formas de plasticidad, simplemente no se conoce el medio por el cual los cambios selectivos de la fuerza sináptica codifican memorias y otras modificaciones conductuales complejas en el encéfalo del mamífero. Bliss, T. V. P. and G. L. Collingridge (1993) A synaptic model of memory: Long-term potentiation in the hippocampus. Nature 361: 31-39. Dan, Y. and M. M. Poo (2006) Spike timing-dependent plasticity: From synapse to perception. Physiol. Rev. 86: 1033-1048. Hilfiker, S., V. A. Pieribone, A. J. Czernik, H. T. Kao, G. J. Augustine and P. Greengard (1999) Synapsins as regulators of neurotransmitter release. Philos. Trans. Roy. Soc. Lond. B 354: 269-279. Kessels, H. W. and R. Malinow (2009) Synaptic AMPA receptor plasticity and behavior. Neuron 61: 340-350. Malenka, R. C. and S. A. Siegelbaum (2001) Synaptic plasticity: Diverse targets and mechanisms for regulating synaptic efficacy. In Synapses, W. M. Cowan, T. C. Sudhof, and C. F. Stevens (eds.). John Hopkins University Press, Baltimore, MD, Ch. 9: pp. 393-413. Massey, P. V. and Z. I. Bashir (2007) Long-term depression: Multiple forms and implications for brain function. Trends Neurosci. 30: 176-184. Nicoll, R. A. (2003) Expression mechanisms underlying long-term potentiation: A postsynaptic view. Philos. Trans. Roy. Soc. Lond. B 358: 721-726. Pittenger, C. and E. R. Kandel (2003) In search of general mechanisms for long-lasting plasticity: Aplysia and the hippocampus. Philos. Trans. Roy. Soc. Lond. B 358: 757-763. Zucker, R. S. and W. G. Regehr (2002). Short-term synaptic plasticity. Annu. Rev. Physiol. 64: 355-405. Artículos originales importantes Resumen Las sinapsis muestran muchas formas de plasticidad que ocurren en un rango temporal amplio. En los tiempos más cortos (segundos a minutos), la facilitación, el aumento, la potenciación y la depresión proporcionan modificaciones rápidas, pero transitorias, en la transmisión sináptica. Estas formas de plasticidad cambian la cantidad de neurotransmisor liberado de las terminaciones presinápticas y se basan en alteraciones en la señalización del Ca2+ y de los grupos de vesículas sinápticas en las terminaciones recientemente activas. Las formas más prolongadas de plasticidad sináptica, como la potenciación a largo plazo y la depresión a largo plazo, también se basan en el Ca2+ y en otros segundos mensajeros intracelulares. Al menos, algunos de los cambios sinápticos producidos por estas formas prolongadas de plasticidad son postsinápticos, causados por cambios en el tráfico de receptores de neurotransmisores, aunque también pueden ocurrir alteraciones en la liberación de neurotransmisores desde la terminación presináptica. En estas formas más duraderas de plasticidad, la fosforilación de las proteínas y los cambios en la expresión genética superan mucho el período de actividad sináptica y pueden producir cambios en la fuerza sináptica que persisten durante horas, días o incluso más. La plasticidad sináptica prolongada puede servir como mecanismo neural para muchas formas de plasticidad encefálica, como el aprendizaje de nuevos comportamientos o la adquisición de nuevos recuerdos. Lecturas adicionales Revisiones Bailey, C. H. and E. R. Kandel (2008) Synaptic remodeling, synaptic growth, and the storage of long-term memory in Aplysia. Prog. Brain Res. 169: 179198. Abraham, W. C. B. Logan, J. M. Greenwood and M. Dragunow (2002) Induction and experience-dependent consolidation of stable long-term potentiation lasting months in the hippocampus. J. Neurosci. 22: 9626-9634. Ahn, S., D. D. Ginty and D. J. Linden (1999) A late phase of cerebellar long-term depression requires activation of CaMKIV and CREB. Neuron 23: 559-568. Betz, W. J. (1970) Depression of transmitter release at the neuromuscular junction of the frog. J. Physiol. (Lond.) 206: 629-644. Bi, G. Q. and M. M. Poo (1998) Synaptic modifications in cultured hippocampal neurons: Dependence on spike timing, synaptic strength, and postsynaptic cell type. J. Neurosci. 18: 10464-10472. Bliss, T. V. P. and T. Lomo (1973) Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J. Physiol. 232: 331-356. Charlton, M. P. and G. D. Bittner (1978) Presynaptic potentials and facilitation of transmitter release in the squid giant synapse. J. Gen. Physiol. 72: 487-511. Chung, H. J., J. P. Steinberg, R. L. Huganir and D. J. Linden (2003) Requirement of AMPA receptor GluR2 phosphorylation for cerebellar long-term depression. Science 300: 1751-1755. Collingridge, G. L., S. J. Kehl and H. McLennan (1983) Excitatory amino acids in synaptic transmission in the Schaffer collateral-commissural pathway of the rat hippocampus. J. Physiol. 334: 33-46. Engert, F. and T. Bonhoeffer (1999) Dendritic spine changes associated with hippocampal long-term synaptic plasticity. Nature 399: 66-70. Enoki, R., Y. L. Hu, D. Hamilton and A. Fine (2009) Expression of long-term plasticity at individual synapses in hippocampus is graded, bidirectional, and mainly presynaptic: Optical quantal analysis. Neuron 62: 242-253. Frey, U. and R. G. Morris (1997) Synaptic tagging and long-term potentiation. Nature 385: 533-536. Gustafsson, B., H. Wigstrom, W. C. Abraham and Y. Y. Huang (1987) Long-term potentiation in the hippocampus using depolarizing current pulses as the conditioning stimulus to single volley synaptic potentials. J. Neurosci. 7: 774-780. Hayashi, Y., S. H. Shi, J. A. Esteban, A. Piccini, J. C. Poncer and R. Malinow (2000) Driving AMPA receptors into synapses by LTP and CaMKII: Requirement for GluR1 and PDZ domain interaction. Science 287: 2262-2267. Junge, H. J. and 7 others (2004) Calmodulin and Muncl3 form a Ca2+ sensor/ effector complex that controls short-term synaptic plasticity. Cell 118: 389-401. Katz, B. and R. Miledi (1968) The role of calcium in neuromuscular facilitation. J. Physiol. (Lond.) 195: 481-492. Konnerth, A., J. Dreessen and G. J. Augustine (1992) Brief dendritic calcium signals initiate long-lasting synaptic depression in cerebellar Purkinje cells. Proc. Natl. Acad. Sci. USA 89: 7051-7055. Lee, S. J., Y. Escobedo-Lozoya, E. M. Szatmari and R. Yasuda (2009). Activation of CaMKII in single dendritic spines during long-term potentiation. Nature 458: 299-304. PLASTICIDAD SINÁPTICA Lev-Tov, A., M. J. Pinter and R. E. Burke (1983) Posttetanic potentiation of group Ia EPSPs: Possible mechanisms for differential distribution among medial gastrocnemius motoneurons. J. Neurophysiol. 50: 379-398. Liao, D., N. A. Hessler and R. Malinow (1995) Activation of postsynaptically silent synapses during pairing-induced LTP in CA1 region of hippocampal slice. Nature 375: 400-404. Malinow, R., H. Schulman and R. W. Tsien (1989) Inhibition of postsynaptic PKC or CaMKII blocks induction but not expression of LTP. Science 245: 862-866. Matsuzaki, M., N. Honkura, G. C. Ellis-Davies and H. Kasai (2004) Structural basis of long-term potentiation in single dendritic spines. Nature 429: 761-766. Miyata, M. and 9 others (2000) Local calcium release in dendritic spines required for long-term synaptic depression. Neuron 28: 233-244. Mulkey, R. M., C. E. Herron and R. C. Malenka (1993) An essential role for protein phosphatases in hippocampal long-term depression. Science 261: 1051-1055. Sakurai, M. (1987) Synaptic modification of parallel fibre-Purkinje cell transmission in in vitro guinea-pig cerebellar slices. J. Physiol, (bond.) 394: 463-480. 185 Silva, A. J., R. Paylor, J. M. Wehner and S. Tonegawa (1992) Impaired spatial learning in alpha-calcium-calmodulin kinase II mutant mice. Science 257: 206-211. Tanaka, K. and G. J. Augustine (2008) A positive feedback signal transduction loop determines timing of cerebellar long-term depression. Neuron 59: 608620. Libros Bliss, T., G. Collingridge and R. Morris (2004) Long-term Potentiation: Enhancing Neuroscience for 30 Years. Oxford University Press. Kandel, E. R. (2007) In Search of Memory: The Emergence of a New Science of Mind. New York: W. W. Norton & Company. Katz, B. (1966) Nerve, Muscle, and Synapse. New York: McGraw-Hill. Squire, L. R. and E. R. Kandel (1999) Memory: From Mind to Molecules. New York: Scientific American Library. UNIDAD II Sensibilidad y procesamiento sensitivo LA SENSIBILIDAD IMPLICA LA CAPACIDAD CAPÍTULO 9 Sistema somatosensitivo: Tacto y propiocepción CAPÍTULO 10 Dolor CAPÍTULO 11 El ojo CAPÍTULO 12 Vías visuales centrales CAPÍTULO 13 Sistema auditivo CAPÍTULO 14 Sistema vestibular CAPÍTULO 15 Sentidos químicos para traducir, codificar y finalmente percibir la información generada por los estímulos que se originan en el medioambiente externo e interno. Gran parte del encéfalo se dedica a estos aspectos. Aunque los sentidos básicos –la sensibilidad somática, la visión, la audición, la sensación vestibular y los sentidos químicos– son muy diferentes entre sí, algunas reglas fundamentales gobiernan la forma en que el sistema nervioso se ocupa de cada una de estas diversas modalidades. Las células nerviosas altamente especializadas denominadas “receptores” convierten la energía asociada con las fuerzas mecánicas, la luz, las ondas sonoras, las moléculas odoríferas o las sustancias químicas ingeridas en señales neurales –señales sensitivas aferentes– que transmiten información acerca del estímulo al encéfalo. Estas señales sensitivas aferentes activan las neuronas capaces de representar tanto la naturaleza cualitativa como cuantitativa del estímulo (qué es y qué intensidad tiene) y, en algunas modalidades (sensibilidad somática, visión, audición), la localización del estímulo en el espacio (dónde está). La evaluación clínica de los pacientes habitualmente requiere una evaluación de los sistemas sensitivos con el fin de inferir la naturaleza y la localización de posibles problemas neurológicos. Por lo tanto, el conocimiento de dónde y cómo se transducen, transmiten, representan y procesan las diferentes modalidades sensitivas resulta esencial para conocer y tratar una amplia variedad de enfermedades. En consecuencia, estos capítulos que se refieren a la neurobiología de la sensibilidad también introducen algunas de las principales relaciones estructura/función en los componentes sensitivos del sistema nervioso. En la página anterior: Corte vertical de la retina del pollo, visualizado mediante tinción por anticuerpos. En la parte superior, están los fotorreceptores (bastones y conos; gris), que transducen la energía luminosa en señales eléctricas. Las señales resultantes se pasan a las capas intermedias de la retina (amarillo y rojo), que procesan aún más esta información. Las células ganglionares de la retina (anaranjadas) en la región intermedia de la imagen transmiten la información procesada hasta el tálamo. Las regiones azules en la parte inferior de la imagen son las capas de las fibras retinianas. (Microfotografía cortesía de Andy J. Fischer.) 189 CAPÍTULO 2 9 SiStema SomatoSenSitivo: tacto y propiocepción Aspectos generales EL SISTEMA SOMATOSENSITIVO posiblemente sea el más diverso de los sistemas sensitivos, y media una gama de sensaciones –tacto, presión, vibración, posición de las extremidades, calor, frío y dolor– que son traducidas por receptores en el interior de la piel o en los músculos y transmitidas a distintos puntos diana en el sistema nervioso central. No es sorprendente que esta maquinaria neurobiológica compleja pueda dividirse en subsistemas funcionalmente distintos con conjuntos diferentes de receptores periféricos y vías centrales. Un subsistema transmite la información desde los mecanorreceptores cutáneos y media las sensaciones de tacto fino, vibración y presión. Otro se origina en receptores especializados que se asocian con músculos, tendones y articulaciones, y es responsable de la propiocepción: nuestra capacidad para percibir la posición de nuestras propias extremidades y otras partes del cuerpo en el espacio. Un tercer subsistema se origina en receptores que proporcionan información sobre estímulos dolorosos y cambios en la temperatura y el tacto grueso. Este capítulo se enfoca en los subsistemas táctil y propioceptivo; los mecanismos responsables de las sensaciones de dolor, temperatura y tacto grueso se consideran en el capítulo siguiente. Las fibras aferentes transmiten información somatosensitiva al sistema nervioso central La sensación somática se origina en la actividad de las fibras nerviosas aferentes cuyas prolongaciones periféricas se ramifican en el interior de la piel o en el músculo (Figura 9.1 A). Los cuerpos celulares de las fibras aferentes residen en una serie de ganglios que se ubican a lo largo de la médula espinal y en el tronco del encéfalo, y se consideran parte del sistema nervioso periférico. Las neuronas de los ganglios de las raíces dorsales y de los ganglios de los nervios craneales (para el cuerpo y la cabeza, respectivamente) son las conexiones críticas que aportan al sistema nervioso central circuitos con información sobre los acontecimientos sensitivos que ocurren en la periferia. Los potenciales de acción generados en las fibras aferentes por acontecimientos que ocurren en la piel o en el músculo se propagan a lo largo de la fibra y atraviesan el cuerpo celular situado en los ganglios hasta que alcanzan las terminaciones presinápticas de las fibras, que se localizan en distintas estructuras diana del sistema nervioso central (Figura 9.1B). Los componentes 190 CAPÍTULO 9 Corteza cerebral Corteza somatosensitiva (A) (B) Células del ganglio de la raíz dorsal Tálamo Fibra aferente mecanosensitiva Tronco del encéfalo Ganglio trigeminal (receptores sensitivos para la cara) Médula espinal Fibra aferente para el dolor y la temperatura Terminaciones receptoras Ce rvical FIGURA 9.1 Las aferencias somatosensitivas transmiten información desde la superficie cutánea hasta los circuitos centrales. (A) Los cuerpos celulares de las fibras aferentes Receptor sensitivo Torácica Ganglios de la raíz dorsal (receptores sensitivos para el cuerpo) somatosensitivas que transmiten información sobre el cuerpo residen en una serie de ganglios de las raíces dorsales que se sitúan a lo largo de la médula espinal; aquellos que transmiten información sobre la cabeza se encuentran en primaria lugar en los ganglios trigeminales. (B) Las neuronas seudounipolares en los ganglios de las raíces dorsales dan origen a prolongaciones periféricas que se ramifican en el interior de la piel (o del músculo) y a prolongaciones centrales que hacen sinapsis con neuronas localizadas en la médula espinal y en niveles superiores del sistema nervioso. Las prolongaciones periféricas de las aferencias de los mecanorreceptores están encapsuladas por células receptoras especializadas; las aferencias que transmiten información del dolor y de la temperatura (termoalgésica) finalizan como terminaciones libres. Lumbar Fibra aferente encapsulada Sacra (A) Exterior de la fibra aferente periféricos y centrales de las fibras aferentes son continuos, y están unidos al cuerpo celular en los ganglios mediante una sola prolongación. Por esta razón, las neuronas de los ganglios de las raíces dorsales se denominan a menudo seudounipolares. A causa de esta configuración, la conducción de la actividad eléctrica a través de la membrana del cuerpo celular no es un paso obligatorio para transmitir la información a los puntos diana centrales. No obstante, los cuerpos celulares de las aferencias sensitivas desempeñan un papel crítico para mantener la maquinaria celular que media la transducción, la conducción y la transmisión por las fibras aferentes sensitivas. El mecanismo fundamental de la transducción sensitiva –el proceso de convertir la energía de un estímulo en una señal eléctrica– es similar en todos las aferencias somatosensitivas: un estímulo altera la permeabilidad de los canales catiónicos en las terminaciones nerviosas aferentes, que genera una corriente Interior de la fibra aferente (B) Estímulo débil Potencial del receptor Na+ Canales iónicos cerrados Estímulo moderado Potencial del receptor Membrana estirada, canales iónicos abiertos Estímulo fuerte Potencial del receptor Potencial de la espiga Umbral FIGURA 9.2 Transducción en la aferencia de un mecanorreceptor. Se muestra aquí la prolongación de un corpúsculo de Pacini. (A) La deformación de la cápsula conduce a un estiramiento de la membrana de la fibra aferente, lo que aumenta la probabilidad de abrir los canales catiónicos de la membrana sensibles al estiramiento. (B) La apertura de los canales catiónicos conduce a la despolarización de la fibra aferente (potencial del receptor). Si la aferencia está suficientemente despolarizada, se genera un potencial de acción que se propaga hasta los puntos diana centrales. SISTEMA SOMATOSENSITIVO: TACTO Y PROPIOCEPCIÓN despolarizante conocida como potencial del receptor (o del generador) (Figura 9.2). Si el potencial del receptor es de suficiente magnitud, alcanza el umbral para la generación de potenciales de acción en la fibra aferente; la tasa de descarga resultante de potenciales de acción es más o menos proporcional a la magnitud de la despolarización, como se describe en los Capítulos 2 y 3. Las fibras aferentes a menudo están encapsuladas por células receptoras especializadas llamadas mecanorreceptores, que ayudan a sincronizar la fibra aferente a las características particulares de la estimulación somática. Las fibras aferentes que no tienen células receptoras especializadas se denominan terminaciones nerviosas libres y son especialmente importan- 191 tes en la sensibilidad al dolor (véase el Capítulo 10). Las aferencias que poseen terminaciones encapsuladas generalmente tienen umbrales más bajos para la generación de potenciales de acción y, por lo tanto, son más sensibles a la estimulación sensitiva que las terminaciones nerviosas libres. Las aferencias somatosensitivos muestran distintas propiedades funcionales Las aferencias somatosensitivos difieren de forma significativa en sus propiedades de respuesta. Estas diferencias, tomadas RECUADRO 9A Dermatomas Cada ganglio de la raíz dorsal (o sensitivo) y su nervio espinal asociado nace de una serie repetida de masas de tejido embrionario denominadas somitas (véase el Cap. 22). Este hecho del desarrollo explica la disposición segmentaria global de los nervios somáticos (y los puntos diana que inervan) en el adulto. El territorio inervado por cada nervio espinal se denomina dermatoma. En los seres humanos, el área cutánea de cada dermatoma se definió en los pacientes en los que están afectadas las raíces dorsales específicas (como en el C2 C3 C4 C5 Cervicales Torácicos Ramas nerviosas trigeminales S1 La inervación que nace de un solo ganglio de la raíz dorsal y su nervio espinal se denomina “dermatoma”. El conjunto completo de dermatomas sensitivos se muestra aquí para un adulto típico. El conocimiento de esta disposición es particularmente importante para definir la Sacros Lumbares C5 C6 C7 C8 Lumbares lidad algésica proporciona una evaluación más precisa de la lesión de un nervio segmentario que el examen de las respuestas al tacto, la presión o la vibración. Sin embargo, la distribución segmentaria de los propioceptores no sigue un mapa dermatómico, sino que se relaciona de manera más íntima con el patrón de inervación muscular. A pesar de estas limitaciones, el conocimiento de los dermatomas es esencial en la evaluación clínica de los pacientes neurológicos; sobre todo, para determinar el nivel de una lesión medular. localización de las lesiones medulares (y de otro tipo) sospechosas. Los números se refieren a los segmentos espinales por los que cada nervio recibe su nombre. (De Rosenzweig y cols., 2002.) T1 T2 T3 T4 T5 T6 T7 T8 T9 T10 T11 T12 L1 L2 L3 L4 L5 Sacros herpes zóster o “culebrilla”), o después de la interrupción quirúrgica (para aliviar el dolor u otras razones). En estos estudios, se observa que los mapas de los dermatomas varían entre los individuos. Más aún, los dermatomas se superponen de forma sustancial, de modo que la lesión de una raíz dorsal individual no conduce a la pérdida completa de sensibilidad en la región cutánea relevante, y la superposición es más extensa para el tacto, la presión y la vibración que para el dolor y la temperatura. Por lo tanto, el examen de la sensibi- S2 Torácicos Cervicales 192 CAPÍTULO 9 (A) (C) Dedo 4 Lado izquierdo Dedo 3 Lado derecho Dedo 2 Dedo 1 Pulgar Palma Antebrazo a Brazo Hombro c Frente Mejilla Nariz Labio superior Frecuencia de espigas b a b c a b c a b c Mama Espalda (B) Vientre Puntas del compás Muslo Pantorrilla Planta Dedo 0 en conjunto, definen distintas clases de aferencias, cada una de las cuales aporta contribuciones singulares a la sensibilidad somática. El diámetro axónico es un factor que diferencia las clases de aferentes somatosensitivas (Cuadro 9.1). Las aferencias sensitivas de mayor diámetro (designados Ia) son los que inervan los receptores sensitivos de los músculos. La mayor parte de la información correspondiente al tacto es transmitida por fibras de diámetro ligeramente menor (aferentes Aβ) y la información sobre dolor y temperatura es transmitida incluso por fibras de menor diámetro (Aδ y C). El diámetro axónico determina la velocidad de conducción del potencial de acción y es muy consistente con las propiedades de los circuitos centrales y con las distintas demandas conductuales para las que se emplea cada tipo de aferente sensitivo (véase el Cap. 16). Otra característica que distingue a las aferencias sensitivas es el tamaño del campo receptivo: el área de la superficie cutánea cuya estimulación conduce a un cambio importante en la frecuencia de los potenciales de acción (Figura 9.3A). Una región dada de la superficie corporal es inervada por aferentes sensitivos que varían mucho en el tamaño de sus campos receptivos. El tamaño del campo receptivo en gran parte es función de las 5 10 15 20 25 30 35 40 45 Umbral medio de discriminación de dos puntos (mm) 50 FIGURA 9.3 Campos receptivos y umbral de discriminación de dos puntos. (A) Patrones de actividad en tres fibras aferentes mecanosensitivas con campos receptivos a, b y c superpuestos sobre la superficie cutánea. Cuando los estímulos de discriminación de dos puntos están poco separados (puntos e histograma verdes), existe un único foco de actividad neural, y la aferencia b dispara de forma más activa. A medida que los estímulos se separan y alejan (puntos e histograma rojos), la actividad de las aferencias a y c aumenta y la actividad en b disminuye. Con alguna distancia de separación (puntos e histograma azules), la actividad en a y c excede a la de b en tal medida que es posible identificar dos focos separados de estimulación. Este patrón diferencial de actividad forma la base para el umbral de discriminación de dos puntos. (B) El umbral de discriminación de dos puntos en los dedos es mucho más fino que en la muñeca a causa de las diferencias en los tamaños de los campos receptivos aferentes –es decir, la distancia de separación necesaria para producir dos focos distintos de actividad neural en la población de aferentes que inervan la muñeca es mucho mayor que aquella para las aferencias que inervan el pulpejo de los dedos. (C) Diferencias en el umbral de discriminación de dos puntos en la superficie del cuerpo. La agudeza somática es mucho mayor en los dedos de las manos y pies, y en el rostro que en los brazos, piernas o torso. (C, de Weinstein, 1968.) características de ramificación de la aferencia en el interior de la piel; las arborizaciones más pequeñas conducen a campos receptivos más pequeños. Además, existen variaciones regionales SISTEMA SOMATOSENSITIVO: TACTO Y PROPIOCEPCIÓN 193 CUADRO 9.1 Aferentes somatosensitivos que conectan receptores con el sistema nervioso central FUNCIÓN SENSITIVA TIPO DE RECEPTOR Propiocepción Huso muscular TIPO DE AXÓN AFERENTEA Axón Ia, II Tacto Corpúsculos de Merkel, Meissner, Pacini y Ruffini DIÁMETRO DEL AXÓN VELOCIDAD DE CONDUCCIÓN 13-20 μm 80-120 m/s 6-12 μm 35-75 m/s 1-5 μm 5-30 m/s 0,2-1,5 μm 0,5-2 m/s Mielina Aβ Dolor, temperatura Terminaciones nerviosas libres Dolor, temperatura, prurito Terminaciones nerviosas libres (amielínicas) Aδ C Durante las décadas de 1920 y 1930, hubo una virtual artesanía que clasificó los axones de acuerdo con su velocidad de conducción. Se distinguieron tres categorías principales, llamadas A, B y C. La A comprende los axones más grandes y más rápidos y la C, los más pequeños y más lentos. Los axones de los mecanorreceptores generalmente pertenecen a la categoría A. El grupo A se divide, además, en subgrupos denominados α (el más rápido), β y δ (el más lento). Para hacer aún más confusa la cuestión, los axones aferentes de los músculos se clasifican habitualmente en cuatro grupos adicionales –I (el más rápido), II, III y IV (el más lento)– con subgrupos denominados con letras romanas minúsculas. (De Rosenzweig y cols., 2005.) a sistemáticas en el tamaño promedio de los campos receptivos aferentes que reflejan la densidad de fibras aferentes que inervan el área. Los campos receptivos de las regiones que tienen una densa inervación (dedos de manos y pies, labios) son relativamente pequeños comparados con aquellos del antebrazo o del dorso que están inervados por una cantidad menor de fibras aferentes (Figura 9.3B). Las diferencias regionales en el tamaño y en la densidad de inervación del campo receptivo son los principales factores que limitan la exactitud espacial con la cual se pueden percibir los estímulos táctiles. Por lo tanto, las medidas de discriminación de dos puntos –la distancia mínima interestímulo necesaria para percibir dos estímulos aplicados simultáneamente como distintos– varían mucho en la superficie cutánea (Figura 9.3C). En el pulpejo de los dedos, los estímulos (puntos de indentación Estímulo De adaptación lenta FIGURA 9.4 Los mecanorreceptores de adaptación lenta y rápida proporcionan diferente información. Los receptores de adaptación lenta siguen respondiendo De adaptación rápida 0 1 2 Tiempo (s) producidos, por ejemplo, por la punta de un compás) se perciben como diferentes cuando están separados más o menos por 2 mm, pero los mismos estímulos aplicados en el antebrazo no se perciben como distintos hasta que están separados por ¡40 mm! Las aferencias sensitivas se diferencian además por la dinámica temporal de su respuesta a la estimulación sensitiva. Algunos aferentes descargan rápidamente cuando se presenta por primera vez un estímulo, luego quedan silentes en presencia de una estimulación continua; otros generan una descarga sostenida en presencia de un estímulo continuo (Figura 9.4A). Se cree que las aferencias de adaptación rápida (aquellas que quedan quiescentes frente a una estimulación continua) son particularmente efectivos para transmitir información sobre los cambios en la estimulación continua, como los producidos por el movimiento del estímulo. Por el contrario, las aferencias de adaptación lenta son más apropiadas para proveer información sobre los atributos espaciales del estímulo, como el tamaño y la forma. Al menos para algunas clases de fibras aferentes, las características de adaptación son atribuibles a las propiedades de las células receptoras que la encapsulan. Las aferencias de adaptación rápida asociados a los corpúsculos de Pacini (véase luego) se convierten en aferentes de adaptación lenta cuando se elimina el corpúsculo. 3 4 a un estímulo, mientras que los receptores de adaptación rápida responden solo al inicio (y a menudo al final) de la estimulación. Estas diferencias funcionales permiten a los mecanorreceptores proporcionar información tanto sobre las cualidades estáticas (a través de los receptores de adaptación lenta) como dinámicas (a través de los receptores de adaptación rápida) de un estímulo. 194 CAPÍTULO 9 Por último, las aferencias sensitivas responden de modo diferente a las cualidades de la estimulación somatosensitiva. Debido a diferencias en las propiedades de los canales de las aferencias sensitivas o en las propiedades de filtro de las células receptoras especializadas que encapsulan muchas aferencias sensitivas, los potenciales generadores son producidos solo por un conjunto restringido de estímulos que recaen sobre una fibra aferente dada. Por ejemplo, las aferencias encapsuladas en el interior de las células receptoras en la piel responden enérgicamente a la deformación mecánica de la superficie cutánea, pero no a los cambios en la temperatura o a la presencia de fuerzas mecánicas o sustancias químicas que se sabe producen sensaciones dolorosas. Los últimos estímulos son especialmente eficaces para impulsar las respuestas de las aferencias sensitivas conocidas como nociceptores (véase el Cap. 10), que terminan en la piel como terminaciones nerviosas libres. Los otros subtipos de mecanorreceptores y nociceptores se identifican sobre la base de sus respuestas distintas a la estimulación somática. Aunque una aferente sensitiva dada puede dar origen a múltiples ramas periféricas, las propiedades de transducción de todas las ramas de una fibra aislada son idénticas. En consecuencia, las aferencias sensitivas somáticas constituyen vías paralelas, que difieren en la velocidad de conducción, en el tamaño del campo receptivo, en la dinámica y en las características del estímulo efectivo. Como se hará patente, estas vías diferentes se mantienen separadas a través de varios estadios Terminaciones nerviosas libres Corpúsculo de Meissner Complejo neuritacélula de Merkel Corpúsculo de Ruffini de procesamiento central, y su actividad contribuye a la extracción de la información somatosensitiva necesaria para el control apropiado tanto de los movimientos orientados a objetivos como reflexivos. Mecanorreceptores especializados en la información táctil receptiva Conocemos mejor la contribución de las distintas vías aferentes a la sensación cutánea ejercida sobre las porciones glabras (lampiñas) de la mano (es decir, la palma y el pulpejo de los dedos). Estas regiones de la superficie cutánea están especializadas en proporcionar una imagen neural de alta definición de los objetos manipulados. El tacto activo, o háptica, comprende la interpretación de patrones espaciotemporales complejos de estímulos que probablemente activen muchas clases de mecanorreceptores. En efecto, la manipulación de un objeto con la mano puede proporcionar a menudo suficiente información como para identificar el objeto, capacidad denominada estereognosia. Al registrar las respuestas de las aferencias sensitivas individuales en los nervios de seres humanos y primates no humanos, fue posible caracterizar las respuestas de estas aferencias bajo condiciones controladas y obtener nociones de su contribución a la sensibilidad somática. Actualmente consideramos cuatro clases distintas de aferencias mecanorreceptoras que inervan la piel glabra de la mano (Figura 9.5; Cuadro 9.2). Las aferencias de las células de Merkel son fibras de adaptación lenta que representan un 25% de las aferencias mecanosensitivas de la mano. Son especialmente ricas en el pulpejo de los dedos, y representan las únicas aferencias que muestrean la información de las células receptoras localizadas en la epidermis. Los complejos “célula de MerkelEpidermis neurita” se ubican en los extremos de las crestas epidérmicas primarias: extensiones de la epidermis en la dermis subyacente que coinciden con las crestas prominentes (huellas digitales) sobre la superficie de los dedos de la mano. Sigue siendo poco claro el papel exacto de las células de Merkel en la transducción somatosensitiva. Las células de Merkel son células excitables que expresan canales del calcio sensibles al voltaje y moléculas que son requeridas para la liberación de las vesículas Dermis sinápticas. Sin embargo, los potenciales de acción parecen surgir de canales iónicos mecanosensibles en la membrana de la fibra aferente, lo que sugiere FIGURA 9.5 La piel alberga distintos mecanorreceptores con diferente morfología. Este esquema Corpúsculo de Pacini Lecho subcutáneo representa la piel lisa y sin vello (glabra) del pulpejo de un dedo. En el Cuadro 9.2 se resumen las principales características de los distintos tipos de receptores. (De Johansson y Vallbo, 1983.) SISTEMA SOMATOSENSITIVO: TACTO Y PROPIOCEPCIÓN 195 CUADRO 9.2 Sistemas aferentes y sus propiedades CAMPO RECEPTOR PEQUEÑO Localización Diámetro del axón Velocidad de conducción Función sensitiva Estímulo efectivo Área del campo receptivoa Densidad de inervación (pulpejo del dedo) Agudeza espacial Respuesta a la indentación sostenida Rango de frecuencia Sensibilidad pico CAMPO RECEPTOR GRANDE MERKEL MEISSNER PACINI RUFFINI Extremo de las crestas del sudor epidérmicas 7-11 μm 40-65 m/s Percepción de la forma y de la textura Papilas dérmicas (cerca de la superficie de la piel) 6-12 μm 35-70 m/s Detección de movimiento; control del agarre Dermis Bordes, puntas, ángulos, curvatura 9 mm2 Movimiento de la piel Dermis y tejidos más profundos 6-12 μm 35-70 m/s Percepción de eventos distantes a través de vibraciones transmitidas; uso de herramientas Vibración 60 mm2 100/cm2 150/cm2 Totalidad del dedo o de la mano 20/cm2 0,5 mm Sostenida (adaptación lenta) 0-100 Hz 5 Hz 3 mm Ninguna (adaptación rápida) 1-300 Hz 50 Hz 10+ mm Ninguna (adaptación rápida) 5-1000 Hz 200 Hz 7+ mm Sostenida (adaptación lenta) 0-? Hz 0,5 Hz 2 μm 6 μm 0,01 μm 0,08 μm 40 μm 300 μm 22 mm2 6-12 μm 35-70 m/s Fuerza tangencial; forma de la mano; dirección del movimiento Estiramiento de la piel 10/cm2 Umbral para la vibración o indentación rápidas Mejor Promedio 8 μm 30 μm Áreas de campo receptivo según se miden con la indentación rápida de 0,5 mm. (De K. O. Johnson, 2002.) a que las células de Merkel sirven para modular la actividad de las aferencias más que como sitio de transducción. Las aferencias de las células de Merkel tienen la resolución espacial más alta de todos las aferencias sensitivas: las aferencias de Merkel individuales pueden resolver detalles espaciales de 0,5 mm. También son sumamente sensibles a los puntos, a los bordes y a la curvatura, lo que los convierte en idealmente adecuados para procesar la información sobre forma y textura. Las aferencias de Meissner son fibras de adaptación rápida que inervan la piel incluso de manera más densa que las aferencias de Merkel, lo que explica un 40% de la inervación mecanosensitiva de la mano humana. Los corpúsculos de Meissner se ubican en los extremos de las papilas dérmicas adyacentes a las crestas primarias y más próximos a la superficie cutánea (véase la Fig. 9.5). Estos receptores alargados están formados por una cápsula de tejido conectivo que comprende varias laminillas de células de Schwann. El centro de la cápsula contiene de 2 a 6 fibras nerviosas aferentes que terminan como discos entre las láminas de las células de Schwann; se cree que esta configuración contribuye a la respuesta transitoria de estas aferencias a la estimulación somática. Debido –al menos, en parte– a su estrecha proximidad con la superficie cutánea, los aferentes de Meissner son más de cuatro veces más sensibles a la deformación cutánea que las aferencias de Merkel; sin embargo, sus campos receptivos son más grandes que las aferencias de Merkel, y, por lo tanto, transmiten señales con una resolución espacial reducida (véase Cuadro 9.2). Los corpúsculos de Meissner son particularmente eficientes en la transducción de información sobre vibraciones de relativa baja frecuencia (3-40 Hz) que tienen lugar cuando se mueven objetos texturados en contacto con la piel. Existen evidencias que sugieren que la información transmitida por las aferencias de Meissner es responsable de la detección del deslizamiento entre la piel y un objeto que se sostiene en la mano, información esencial de retroalimentación para el control eficiente de la prensión. Las aferencias de Pacini son fibras de adaptación rápida que representan el 10-15% de la inervación mecanosensitiva de la mano. Los corpúsculos de Pacini se localizan en la profundidad de la dermis o en el tejido subcutáneo; su aspecto se asemeja al de una pequeña cebolla, con capas concéntricas de membranas rodeando a una única fibra aferente (véase la Fig.9.5). Esta cápsula laminar actúa como filtro y permite que solo las perturbaciones transitorias en las altas frecuencias (250-350 Hz) activen las terminaciones nerviosas. Los corpúsculos de Pacini se adaptan más rápidamente que los corpúsculos de Meissner y tienen un umbral de respuesta más bajo. Las aferencias de Pacini más sensibles generan potenciales de acción para los desplazamientos de la piel de hasta 10 nanómetros. Como son tan sensibles, los campos receptivos de las aferencias de Pacini a menudo son grandes y es difícil definir sus límites. Las propiedades de las aferencias de Pacini los hacen apropiados para detectar las vibraciones transmitidas a través de los objetos que hacen contacto con la mano o que son tomadas en ella –lo que sin duda tiene 196 CAPÍTULO 9 importancia para el uso hábil de las herramientas (p. ej., utilizar una llave inglesa, cortar el pan con un cuchillo, escribir). Las aferencias de Ruffini son fibras de adaptación lenta y son los mecanorreceptores cutáneos menos conocidos. Las terminaciones de Ruffini son especializaciones capsulares fusiformes alargadas que se hallan localizadas en la profundidad de la piel, y en ligamentos y tendones (véase la Figura 9.5). El eje mayor del corpúsculo suele estar orientado paralelo a las líneas de estiramiento en la piel; por lo tanto, los corpúsculos de Ruffini son particularmente sensibles al estiramiento cutáneo producido por el movimiento de los dedos o de la extremidad; representan alrededor del 20% de los mecanorreceptores en la mano humana. Aunque existen todavía algunas dudas sobre su función, se cree que los corpúsculos de Ruffini responden especialmente a los estímulos generados de manera interna, como el movimiento de los dedos de la mano. La información aportada por las aferencias de Ruffini contribuye, junto con los receptores musculares, a proporcionar una representación exacta de la posición de los dedos de la mano y de su conformación (véase la sección siguiente sobre propiocepción). Los diferentes tipos de información que transmiten las aferencias sensitivas a las estructuras centrales se demuestran muy bien en los experimentos llevados a cabo por K. O. Johnson y cols., que compararon las respuestas de diferentes aferencias según se movía el pulpejo de un dedo a través de una hilera de letras de Braille en relieve (Figura 9.6). Indudablemente, todos los tipos de aferencias son activadas por esta estimulación, pero la información que aporta cada tipo varía enormemente. El patrón de actividad en las aferencias de Merkel es suficiente para reco- nocer los detalles del patrón de Braille, y las aferencias de Merkel aportan una versión ligeramente más grosera de este patrón. Pero estos detalles se pierden en la respuesta de las aferencias de Pacini y de Ruffini; presumiblemente estas respuestas tienen más que ver con rastrear el movimiento y la posición del dedo que con la identidad específica de los caracteres del Braille. Mecanorreceptores especializados en la propiocepción Mientras los mecanorreceptores cutáneos proporcionan información derivada de estímulos externos, otra clase importante de receptores brinda información sobre las fuerzas mecánicas que se originan en el cuerpo propiamente dicho, sobre todo, en el sistema musculoesquelético. El propósito de estos propioceptores (“receptores para lo propio”) es, fundamentalmente, brindar información detallada y continua sobre la posición en el espacio de las extremidades y de otras partes del cuerpo. Los mecanorreceptores de bajo umbral –que incluyen los husos neuromusculares, los órganos tendinosos de Golgi y los receptores articulares– proporcionan este tipo de información sensitiva, esencial para la ejecución exacta de los movimientos complejos. La información sobre la posición y el movimiento de la cabeza resulta particularmente importante; en este caso, los propioceptores están integrados con el sistema vestibular sumamente especializado, que se considera por separado en el Capítulo 14. (Los propioceptores especializados también existen en el corazón y en los vasos sanguíneos mayores para proporcionar información sobre Hilera de receptores en un dedo que se mueve a través de una hilera de letras Braille en relieve “A” “B” “C” FIGURA 9.6 Simulación de los patrones de actividad en diferentes aferencias mecanosensitivas en el pulpejo de un dedo. Cada punto en los registros de respuesta Célula de Merkel representa un potencial de acción registrado de una única fibra aferente mecanosensitiva que inerva el dedo humano Corpúsculo de Meissner cuando se mueve a través de una hilera de tipo Braille. Una línea horizontal de puntos en el gráfico de tipo raster, representa el patrón de actividad enla aferencia como resultado de mover el patrón de izquierda a derecha a través del dedo. Después, se desplazó la posición del patrón (en relación con la punta del dedo) por una pequeña distancia y el patrón se movió nuevamente a través del dedo. La repetición de este Corpúsculo de Ruffini patrón varias veces produce un registro que simula el patrón de actividad que surgiría en una población de aferencias cuyos campos receptivos se sitúan a lo largo de una línea en el pulpejo del dedo (puntos rojos). Solo las aferencias de las células de Merkel de adaptación lenta (panel superior) proporcionan una representación de alta fidelidad del patrón Corpúsculo de Pacini 10 mm Braille –es decir, los puntos Braille individuales pueden distinguirse solo en el patrón de la actividad neural aferente de Merkel. (De Phillips y cols., 1990). SISTEMA SOMATOSENSITIVO: TACTO Y PROPIOCEPCIÓN la presión arterial, pero estas neuronas se consideran parte del sistema motor visceral; véase el Cap. 21). El conocimiento más detallado sobre la propiocepción proviene de estudios de los husos musculares, que se encuentran en todos los músculos estriados (esqueléticos), salvo en algunos. Los husos musculares consisten en cuatro a ocho fibras musculares intrafusales rodeadas por una cápsula de tejido conectivo. Las fibras intrafusales están distribuidas entre las fibras extrafusales del músculo esquelético, que son las verdaderas fibras productoras de fuerza (Figura 9.7A) y en una disposición paralela a ellas. Las aferencias sensitivas se enrollan alrededor de la porción central del huso intrafusal y, cuando el músculo es estirado, la tensión sobre las fibras intrafusales activa canales iónicos con puerta mecánica en las terminaciones nerviosas, lo que desencadena potenciales de acción. La inervación del huso muscular se origina en dos clases de fibras: terminaciones primarias y secundarias. Las terminaciones primarias se originan en los axones sensitivos mielínicos más grandes (aferencias del grupo 197 Ia) y tienen respuestas de adaptación rápida a los cambios en la longitud muscular; por el contrario, las terminaciones secundarias (aferencias del grupo II) producen respuestas sostenidas a longitudes musculares constantes. Se cree que las terminaciones nerviosas transmiten información sobre la dinámica de las extremidades –la velocidad y la dirección del movimiento–, mientras que las terminaciones secundarias brindan información sobre la posición estática de las extremidades. Los cambios en la fuerza muscular no son los únicos factores que impactan en la respuesta de las aferencias fusales. Las fibras intrafusales por sí solas son fibras contráctiles y están controladas por un conjunto separado de neuronas motoras (neuronas motoras γ) que se ubican en el asta ventral de la médula espinal. Aunque las fibras intrafusales no agregan fuerza apreciable a la contracción muscular, los cambios en la tensión de las fibras intrafusales tienen un impacto importante sobre la sensibilidad de las aferencias fusales a los cambios en la longitud muscular. Por lo tanto, para que los circuitos centrales proporcionen un relato (B) Órgano tendinoso de Golgi (A) Huso muscular Axón de neurona motora _ Fibras musculares extrafusales Cápsula Aferente axónico Ib Fibras musculares extrafusales Fibras musculares intrafusales Axón de neurona motora a Axones aferentes del grupo Ia Cápsula (tejido conectivo) que rodea el huso Axones aferentes del grupo II Axón Tendón Fibrillas de colágeno FIGURA 9.7 Propioceptores en el sistema musculoesquelético. Estos “autorreceptores” aportan información acerca de la posición de las extremidades y de otras partes del cuerpo en el espacio. (A) Un huso muscular y varias fibras musculares extrafusales. Las fibras musculares intrafusales especializadas del huso están rodeadas por una cápsula de tejido conectivo. (B) Los órganos de Golgi son mecanorreceptores de bajo umbral que se encuentran en los tendones; aportan información sobre los cambios en la tensión muscular. 198 CAPÍTULO 9 exacto de la posición de las extremidades y el movimiento, se debe tener en cuenta el nivel de actividad del sistema γ. (Para una explicación más detallada de la interacción del sistema γ, así como de la actividad de las aferencias del huso, remítase a los Capítulos 16 y 17). La densidad de los husos en el músculo humano varía. Los músculos grandes generan movimientos gruesos y tienen relativamente pocos husos; por el contrario, los músculos extraoculares y los músculos intrínsecos de la mano y del cuello están ricamente cargados de husos, lo que refleja la importancia de los movimientos oculares precisos, la necesidad de manipular objetos con gran fineza y la demanda continua de una posición precisa de la cabeza. Esta relación entre la densidad de los receptores y el tamaño muscular es compatible con la generalización de que el aparato sensitivo motor en todos los niveles del sistema nervioso es mucho más rico para las manos, la cabeza, los órganos de la palabra y otras partes del cuerpo que se utilizan para realizar tareas especialmente importantes y exigentes. Los husos están ausentes en algunos músculos, como los del oído medio, que no requieren el tipo de retroalimentación que proporcionan estos receptores. Mientras los husos musculares se encuentran especializados detectar cambios en la longitud muscular, los mecanorreceptores de bajo umbral en los tendones informan al sistema nervioso central sobre los cambios en la tensión muscular. Estos mecanorreceptores, denominados órganos tendinosos de Golgi, están formados por ramos de aferencias del grupo Ib distribuidas entre las fibras de colágeno que forman los tendones (Figura 9.7B). Cada órgano tendinoso de Golgi está dispuesto en serie con una pequeña cantidad (10-20) de fibras musculares extrafusales. En conjunto, la población de órganos tendinosos de Golgi para un músculo dado proporciona una muestra exacta de la tensión que existe en el músculo. Sigue siendo un área de investigación activa el modo en que cada una de estas aferencias propioceptivas contribuye a la percepción de la posición, del movimiento y de la fuerza de las extremidades. Los experimentos que utilizaron vibradores para estimular los husos de músculos específicos proporcionaron una evidencia firme de que la actividad de estas aferencias puede dar origen a sensaciones vívidas de movimiento en las extremidades inmovilizadas. Por ejemplo, la vibración del músculo bíceps conduce a la ilusión de que el codo se está moviendo hasta una posición extendida, como si el bíceps fuera estirado. Se han producido ilusiones similares de movimientos en músculos posturales y faciales. En algunos casos, la magnitud del efecto es tan grande que produce la percepción de que es anatómicamente imposible; por ejemplo, cuando se hizo vibrar de modo enérgico un músculo extensor de la muñeca, los sujetos informaron que la mano se hiperextendió hasta el punto en que casi estuvo en contacto con el dorso del antebrazo. En todos estos casos, solo ocurre la ilusión si se ocluye la vista del sujeto para que no pueda ver la posición de la extremidad, lo que demuestra que incluso a través de aferencias propioceptivas aisladas se puede proporcionar indicios sobre la posición de las extremidades: en condiciones normales tanto las señales somáticas como visuales desempeñan papeles importantes. Antes de estos estudios, se consideraba que la fuente primaria de información propioceptiva sobre la posición y el movimiento de las extremidades surgía de los mecanorreceptores en las articulaciones y a su alrededor. Estos receptores articulares se asemejan a muchos de los receptores que se encuentran en la piel, incluidas las terminaciones de Ruffini y los corpúsculos de Pacini. Sin embargo, los individuos que se han sometido a reemplazos articulares artificiales mostraron solo déficit menores para juzgar la posición o el movimiento de las extremidades, y la anestesia de una articulación como la de la rodilla no tuvo ningún efecto sobre el juicio de la posición o el movimiento articular. Aunque contribuyen poco a la propiocepción de las extremidades, los receptores articulares parecen ser importantes a la hora de juzgar la posición de los dedos de las manos. Junto con las señales cutáneas provenientes de las aferencias de Ruffini y de las aferencias de los husos musculares que contribuyen a la representación fina de la posición de los dedos de la mano, los receptores articulares desempeñan un papel protector al señalar las posiciones situadas por debajo de los límites del rango de movimiento articular normal de los dedos. Vías centrales que transmiten información táctil del cuerpo: sistema columna dorsal-lemnisco medial Los axones de las aferencias mecanosensitivas cutáneos entran en la médula espinal a través de las raíces dorsales; la mayoría asciende homolateralmente a través de las columnas dorsales (también llamadas cordones posteriores) de la médula hasta la parte inferior del bulbo raquídeo, donde hacen sinapsis sobre las neuronas de los núcleos de las columnas dorsales (Figura 9.8A). El término columna se refiere al aspecto columnar macroscópico de estas fibras cuando discurren a lo largo de la médula espinal. Estas neuronas de primer orden en la vía pueden tener prolongaciones axónicas muy largas: las neuronas que inervan las extremidades inferiores, por ejemplo, tienen axones que se extienden a través de toda la longitud de la médula. Las columnas dorsales de la médula espinal están organizadas topográficamente de modo que las fibras que transmiten información desde las extremidades inferiores se ubican más mediales y discurren en un haz circunscrito conocido como fascículo grácil (del latín fasciculus, ‘haz’; gracilis, ‘delgado’), mientras que aquellas que transmiten información de las extremidades superiores, del tronco y del cuello se ubican en un haz más lateral conocido como fascículo cuneiforme (“haz con forma de cuña”). A su vez, las fibras de estos dos tractos terminan en diferentes subdivisiones de los núcleos de las columnas dorsales: una subdivisión medial, el núcleo grácil, y una subdivisión lateral, el núcleo cuneiforme. Las neuronas de segundo orden en los núcleos de las columnas dorsales envían sus axones a la porción somatosensitiva del SISTEMA SOMATOSENSITIVO: TACTO Y PROPIOCEPCIÓN (A) 199 (B) Cerebro Corteza somatosensitiva primaria Núcleo ventral posterolateral del tálamo Núcleo ventral posteromedial del tálamo Mesencéfalo Lemnisco trigeminal Lemnisco medial Ganglio trigeminal Protuberancia media Lemnisco medial Lemnisco medial Bulbo raquídeo rostral Fibras arciformes internas Bulbo raquídeo caudal Receptores Núcleo mecanosensitivos principal del complejo desde el rostro trigeminal Núcleo grácil (vías desde la parte inferior del cuerpo) Núcleo cuneiforme (vías desde la parte superior del cuerpo) Tracto grácil Tracto cuneiforme Médula espinal cervical Médula espinal lumbar Receptores mecanosensitivos desde la parte superior del cuerpo Receptores mecanosensitivos desde la parte inferior del cuerpo FIGURA 9.8 Esquema de las vías mecanosensitivas principales. (A) La vía cordonal posterior-lemnisco medial transmite información mecanosensitiva desde el tercio posterior de la cabeza y del resto del cuerpo. (B) La porción trigeminal del sistema mecanosensitivo transmite información similar desde el rostro. 200 CAPÍTULO 9 tálamo. Los axones que salen de los núcleos de las columnas dorsales son identificados como fibras arciformes internas. Las fibras arciformes internas posteriormente cruzan la línea media y luego forman un tracto alargado de modo dorsoventral, conocido como lemnisco medial. La palabra lemnisco significa ‘cinta’; las fibras arciformes internas se cruzan en equis entre sí, por lo que se denomina “decusación del lemnisco medial”, pues remite al numeral romano x o decem (10). En un corte transversal a través de la médula, como el que se muestra en la Figura 9.8A, los axones del lemnisco medial que transmiten información desde las extremidades inferiores se localizan de modo ventral, mientras que aquellos relacionados con las extremidades superiores se localizan en el dorso. A medida que el lemnisco medial asciende a través de la protuberancia y del mesencéfalo, rota 90 grados lateralmente, de modo que las fibras que representan la parte superior del cuerpo finalmente se localizan en la porción medial del tracto y las que representan la parte inferior del cuerpo están en la porción lateral. Los axones del lemnisco medial hacen sinapsis con neuronas talámicas localizadas en el núcleo ventral posterolateral (VPL). Las neuronas de tercer orden en el VPL envían sus axones a través de la cápsula interna para terminar en el giro poscentral de la corteza cerebral, región conocida como corteza somatosensitiva primaria o SI. Las neuronas en el VPL también envían axones a la corteza somatosensitiva secundaria (SII), región más pequeña que se ubica en el margen superior del surco lateral. Vías centrales que transmiten información táctil desde el rostro: sistema trigeminotalámico La información de los mecanorreceptores del rostro es transmitida centralmente por un conjunto separado de neuronas de primer orden que se localizan en el ganglio trigeminal (nervio craneal V) (Figura 9.8B). Las prolongaciones periféricas de estas neuronas forman las tres subdivisiones principales del nervio trigémino (las ramas oftálmica, maxilar y mandibular). Cada rama inerva un territorio bien definido del rostro y de la cabeza, que incluye los dientes y las mucosas de las cavidades oral y nasal. Las prolongaciones centrales de las células del ganglio trigeminal forman las raíces sensitivas del nervio trigémino; entran en el tronco del encéfalo a nivel de la protuberancia para terminar en neuronas del complejo trigeminal del tronco encefálico. El complejo trigeminal tiene dos componentes principales: el núcleo principal y el núcleo espinal. (Más tarde se considerará por separado un tercer componente: el núcleo mesencefálico trigeminal.) La mayoría de las aferencias que transmiten información desde los mecanorreceptores cutáneos de bajo umbral termina en el núcleo principal. En efecto, este núcleo corresponde a los núcleos de la columna dorsal que transmiten información mecanosensitiva del resto del cuerpo. El núcleo espinal contiene neuronas sensibles al dolor, a la temperatura y al tacto grueso, y se explicará con mas detalle en el Capítulo 10. Las neuronas de segundo orden de los núcleos trigeminales del tronco del encéfalo dan axones que atraviesan la línea media y ascienden hasta el núcleo ventral posteromedial (VPM) del tálamo a través del tracto trigeminotalámico (también denominado lemnisco trigeminal). Las neuronas en el VPM envían sus axones a las áreas corticales SI y SII. Vías centrales que transmiten información propioceptiva del cuerpo Al igual que sus análogos para la sensibilidad cutánea, los axones de las aferencias propioceptivas entran en la médula espinal a través de las raíces dorsales y, en gran parte de su recorrido, viajan con los axones que transmiten información cutánea. Sin embargo, algunas diferencias en las vías de la médula espinal para las vías propioceptivas reflejan el papel importante que desempeña la información propioceptiva en la regulación refleja del control motor y en la percepción. Primero, cuando entran en la médula espinal, muchas de las fibras de las aferencias propioceptivas se bifurcan en las ramas ascendentes y descendentes, las que, a su vez, envían ramas colaterales a varios segmentos medulares (Figura 9.9). Algunas ramas colaterales penetran en el asta dorsal de la médula espinal y hacen sinapsis en neuronas situadas allí, así como en neuronas del asta ventral. Estas sinapsis median, entre otras cosas, reflejos segmentarios como el reflejo “rotuliano” o el reflejo miotático descritos en el Capítulo 1; los Capítulos 16 y 17 describen estos reflejos con más detalle. Segundo, la información aportada por las aferencias propioceptivas es importante no solo para nuestra capacidad de percibir la posición de las extremidades; también es esencial para las funciones del cerebelo: una estructura que regula el momento oportuno de las contracciones musculares necesarias para realizar los movimientos voluntarios. En consecuencia, la información propioceptiva alcanza circuitos corticales más altos que las ramas de las vías que también se dirigen al cerebelo, y algunos de estos axones discurren a través de los tractos de la médula espinal cuyos nombres reflejan su asociación con esta estructura. La asociación con las vías cerebelosas es especialmente clara para la vía que transmite información propioceptiva para la parte inferior del cuerpo hasta los núcleos de la columna dorsal. Las aferencias propioceptivas de primer orden que entran en la médula espinal entre los niveles mediolumbar y torácico (L2-T1) hacen sinapsis en las neuronas del núcleo de Clarke, situado en la cara medial del asta dorsal (véase Fig. 9.9, vía roja). Las aferencias que ingresan por debajo de este nivel ascienden a través de la columna dorsal y luego hacen sinapsis con neuronas en el núcleo de Clarke. Las neuronas de segundo orden del núcleo de Clarke envían sus axones a la columna lateral posterior homolateral de la médula espinal, donde viajan hacia arriba hasta el nivel del bulbo raquídeo en el tracto espinocerebeloso dorsal. Estos axones continúan en el cerebelo, pero en su recorrido, dan SISTEMA SOMATOSENSITIVO: TACTO Y PROPIOCEPCIÓN Hacia el cerebelo y los núcleos de la columna dorsal Aferentes de los husos musculares, parte superior del cuerpo Médula espinal lumbar FIGURA 9.9 Vías propioceptivas para la parte superior e inferior del cuerpo. Las aferencias propioceptivas para la parte inferior del cuerpo hacen sinapsis sobre neuronas en el asta dorsal y ventral de la médula espinal y en neuronas del núcleo de Clarke. Estas neuronas envían sus axones a través del tracto espinocerebeloso dorsal hasta los núcleos de la columna dorsal y el cerebelo. Las Médula espinal cervical Médula espinal torácica 201 Tracto espinocerebeloso dorsal aferencias propioceptivas para la parte superior del cuerpo también tienen sinapsis en las astas dorsales y ventrales, pero luego ascienden a través de los núcleos de la columna dorsal. Las neuronas diana propioceptivas de los núcleos de la columna dorsal envían sus axones a través de la línea media y ascienden a través del lemnisco medial hasta el núcleo ventral posterior (véase Fig. 9.8). pos celulares de las neuronas propioceptivas de primer orden para el rostro tienen Aferencias de los una localización inusual: en lugar de resihusos musculares, dir en el ganglio trigeminal, se encuentran parte inferior del dentro del sistema nervioso central, en el cuerpo núcleo mesencefálico del trigémino, un conjunto bien definido de neuronas situadas en la extensión lateral de la región gris central del mesencéfalo. Al igual que sus análogos en los ganglios trigeminal y de las raíces dorsales, estas neuronas seudounipolares tienen prolongaciones periféricas que inervan husos musculares y órganos tendinosos de Golgi asociados con la musculatura facial (sobre todo, de los músculos de la mandíbula) y prolongaciones centrales que incluyen proyecciones hasta los núcleos del tronco encefálico responsables del control reflejo de los músculos faciales. Aunque no está clara la vía exacta, la información proveniente de las aferencias propioceptivas en el núcleo mesencefálico del trigémino también alcanza el tálamo y está representada en la corteza somatosensitiva. Núcleo de Clarke Médula espinal sacra colaterales que hacen sinapsis con neuronas situadas inmediatamente por fuera del núcleo grácil (para este propósito, las “neuronas propioceptivas” de los núcleos de la columna dorsal). Los axones de estas neuronas de tercer orden se decusan, se unen al lemnisco medial, y acompañan a las fibras de los mecanorreceptores cutáneos en su recorrido hasta el VPL del tálamo. Los aferencias propioceptivas de primer orden provenientes de las extremidades superiores tienen un recorrido similar al de los mecanorreceptores cutáneos (véase Fig. 9.9, vía azul). Entran en la médula espinal y discurren a través de la columna dorsal (fascículo cuneiforme) hasta el nivel del bulbo raquídeo, donde hacen sinapsis sobre neuronas propioceptivas en los núcleos de la columna dorsal. Las neuronas de segundo orden envían sus axones a través de la línea media, donde se unen al lemnisco medial y ascienden hasta el VPL del tálamo. Vías centrales que transmiten información propioceptiva del rostro Al igual que la información de los mecanorreceptores cutáneos, la información propioceptiva proveniente del rostro es transmitida a través del nervio trigémino. Sin embargo, los cuer- Componentes somatosensitivos del tálamo Cada una de las distintas vías somatosensitivas ascendentes que se originan en la médula espinal y en el tronco encefálico converge en el complejo ventral posterior del tálamo y terminan de forma organizada (Figura 9.10). Una de las características de organización de este complejo, que ilustra el patrón de las terminaciones aferentes, es una representación del cuerpo y de la cabeza. Como ya mencionamos, el núcleo ventral posterolateral (VPL), situado más lateralmente, recibe proyecciones del lemnisco medial que transmiten información somatosensitiva del cuerpo y de la parte posterior de la cabeza, mientras que el núcleo ventral posteromedial (VPM), situado más me- 202 CAPÍTULO 9 Corteza somatosensitiva primaria (SI) Corteza parietal posterior FIGURA 9.10 Diagrama de las porciones somatosensitivas del tálamo y sus puntos diana correspondientes en la circunvolución poscentral. El complejo nuclear ventral Corteza somatosensitiva 1 4 7 3b posterior comprende el VPM, que transmite la información somatosensitiva transportada por el sistema trigeminal desde el rostro, y el VPL, que transmite esta información desde el resto del cuerpo. En el recuadro superior, se observa la or- 2 5 3a Surco central Circunvolución poscentral Núcleo ventral posteromedial (VPM) ganización de la corteza somatosensitiva primaria en la circunvolución poscentral; aquí, en un corte a través de la circunvolución desde adelante hacia Tálamo atrás. (De Brodal, 1992, y de Jones y cols., 1982.) Corteza somatosensitiva secundaria (SII) Núcleo ventral posterolateral (VPL) Complejo VP dialmente, recibe axones del lemnisco trigeminal que transmiten información somatosensitiva del rostro. Además, las señales que transmiten diferentes tipos de información somatosensitiva –por ejemplo, aquellas que responden a diferentes tipos de mecanorreceptores, a las aferencias del huso muscular o a los órganos tendinosos de Golgi– terminan en poblaciones separadas de células de relevo dentro del complejo ventral posterior. Por lo tanto, la información aportada por diferentes receptores somatosensitivos se mantiene segregada en su pasaje a los circuitos corticales. Corteza somatosensitiva primaria proyecta hasta neuronas corticales situadas en la capa 4 de la corteza somatosensitiva primaria (véase Recuadro 26A para una descripción de la laminación cortical). La corteza somatosensitiva primaria en los seres humanos (también llamada SI) se sitúa en el giro poscentral del lóbulo parietal y comprende cuatro regiones distintas o campos, denominados áreas 3a, 3b, 1 y 2 de Brodmann (Figura 9.11A). Estudios de mapeos en seres humanos y en otros primates muestran además que cada una de estas cuatro áreas corticales contiene una representación separada y (A) Área 1 Corteza somatosensitiva primaria (S1) La mayoría de los axones que se originan en neuronas del complejo ventral posterior del tálamo se Corteza somatosensitiva secundaria (S2) región de la corteza humana a partir de la cual la se registra actividad eléctrica luego de realizar una estimulación mecanosensitiva en diferentes partes del cuerpo. (Los pacientes del (B) Tronco Cuello Cabeza Hombro Brazo Codo Antebrazo estudio fueron sometidos a procedimientos neuroquirúrgicos, los cuales eran necesarios para llevar a cabo este mapeo). Si bien actualmente los métodos por imágenes modernos están refinando estos datos clásicos, el mapa somatotópico humano maria, por las mismas razones (véase Cap. 19). (De Penfield y cols., 1953, y de Corsi,1991.) Área 3a Área 3b FIGURA 9.11 Orden somatotópico en la corteza somatosensitiva primaria humana. (A) Representación de la definido por primera vez en la década de 1930 en general mantuvo su validez. (B) Diagrama que muestra la representación somatotópica de partes del cuerpo de medial a lateral. (C) Dibujo del homúnculo construido sobre la base de ese mapeo. Obsérvese que la cantidad de corteza somatosensitiva dedicada a las manos y al rostro es mucho mayor que la cantidad relativa de superficie corporal en estas regiones. Se observa una desproporción similar en la corteza motora pri- Área 2 Medial Pierna Pies Dedos del pie Genitales Mano Dedos 5 4 3 2 Pulgar Ojos Nariz Rostro Labio superior Labio inferior Mentón Garganta Lengua Lateral Dientes, mandíbula, encías (C) SISTEMA SOMATOSENSITIVO: TACTO Y PROPIOCEPCIÓN completa del cuerpo. En estos mapas somatotópicos, el pie, la pierna, el tronco, los miembros anteriores y la cara están representados en una disposición medial a lateral, como se muestra en la Figura 9.11B. Una característica sobresaliente de los mapas somatotópicos, reconocida poco después de su descubrimiento, es su falta de representación del cuerpo humano en las proporciones reales. Cuando los neurocirujanos determinaron la representación del cuerpo humano en la corteza sensitiva (y motora) primaria, el homúnculo (“hombrecillo”) definido por estos procedimientos de mapeo tenía un rostro y manos macroscópicamente agrandados, en comparación con el torso y las extremidades proximales (Figura 9.11C). Estas anomalías se originan porque la manipulación, la expresión facial y la palabra son extraordinariamente importantes para los seres humanos y requieren muchos circuitos, tanto centrales como periféricos, para gobernarlos. Por lo tanto, en los seres humanos, la médula espinal cervical está agrandada para acomodar los circuitos extras relacionados con la mano y con la extremidad superior, y, como afirmamos antes, la densidad de los receptores es mayor en regiones como las manos y los labios. Estas distorsiones también son evidentes cuando se comparan mapas topográficos entre las especies. Por ejemplo, en el cerebro de la rata, una cantidad desmesurada de corteza somatosensitiva está dedicada a representar sus grandes bigotes componentes clave de la aferencia somatosensitiva para ratas y ratones (Recuadro 9B), mientras que los mapaches tienen una representación exagerada de sus patas y el ornitorrinco de su hocico. En RECUADRO 9B Patrones de organización dentro de las cortezas sensitivas: módulos encefálicos En los últimos 40, años distintas observaciones aclararon que, en muchas partes del encéfalo, hay una subestructura repetida en el interior de los mapas corticales somatosensitivos (y en muchos otros). Esta subestructura adquiere la forma de unidades llamadas módulos, y cada módulo comprende cientos o miles de células nerviosas en patrones repetidos. La ventaja de estos patrones repetidos para la función encefálica aún es, en parte, un misterio; sin embargo, para los neurobiólogos, esos patrones repetidos proporcionan indicios importantes sobre la conectividad cortical y los mecanismos por los que la actividad neural influye en el desarrollo encefálico (véanse Caps. 22 y 23). En la década de 1920, el neuroanatomista español Rafael Lorente de Nó observó por primera vez que la corteza somatosensitiva primaria comprende unidades elementales de células conectadas verticalmente, sobre la base de sus estudios en la rata. Sin embargo, la importancia de la modularidad cortical siguió en gran parte sin explicación hasta la década de 1950, 203 cuando algunos experimentos electrofisiológicos indicaron una disposición de unidades repetidas en los encéfalos de gatos y posteriormente en monos. Vernon Mountcastle, un neurofisiólogo de Johns Hopkins University School of Medicine, observó que la penetración con microelectrodos verticales en la corteza somatosensitiva primaria de estos animales encontraba células que respondían al mismo tipo de estímulo presentado en la misma localización sobre la superficie corporal. Poco después del trabajo pionero de Mountcastle, David Hubel y Torsten Wiesel descubrieron una disposición similar en la corteza visual primaria del gato. Tomadas en conjunto, estas y otras observaciones condujeron a Mountcastle al punto de vista más general de que “el patrón elemental de organización de la corteza cerebral es una columna de orientación vertical o un cilindro de células capaz de cumplir funciones de aferencias-eferencias de complejidad considerable”. Desde estos descubrimientos a fines de la década de 1950 y comienzos de la de 1960, el punto de vista de que los cir- (A) (B) (C) (D) (E) (Continúa en la página siguiente) Ejemplos de subestructuras modulares repetidas en el encéfalo de los mamíferos. (A) Columnas de dominancia ocular en la capa IV de la corteza visual primaria (V1) del mono Rhesus. (B) Unidades repetidas denominadas “blobs” en las capas II y III en V1 de un mono ardilla. (C) Tiras en las capas II y III en V2 de un mono ardilla. (D) Toneles en la capa IV en la corteza somatosensitiva primaria de una rata. (E) Glomérulos del bulbo olfatorio de un ratón. (F) Unidades repetidas denominadas “barreloides” en el tálamo de una rata. Estos y otros ejemplos indican que la organización modular es frecuente en el encéfalo. Estas unidades son del orden de cien a varios cientos de micrones. (De Purves y cols., 1992.) (F) 204 CAPÍTULO 9 RECUADRO 9B (continúa) cuitos modulares representan una característica fundamental de la corteza cerebral de los mamíferos ganó amplia aceptación y muchas de estas entidades se describen ahora en distintas regiones corticales (véase la figura). Este conjunto de indicios para el patrón de los circuitos condujo a muchos neurocientíficos a la conclusión, como Mountcastle, de que los módulos son una característica fundamental de la corteza cerebral, esencial para la percepción, la cognición y, tal vez, incluso la conciencia. A pesar de la prevalencia de los módulos, surgen algunos problemas con el punto de vista de que las unidades modulares son universalmente importantes en la función cortical. Primero, si bien se observan con facilidad circuitos modulares de una clase dada en los encéfalos de algunas especies, no se hallaron en las mismas regiones encefálicas de otros animales, a veces íntimamente relacionados. Segundo, no todas las regiones de la corteza de los mamíferos están organizadas de forma modular. Tercero, no se determinó una función constante de los módulos, a pesar de todo el esfuerzo y la especulación. Por lo tanto, esta característica sobresaliente de la organización de la corteza somatosensitiva y de otras regiones corticales (y algunas subcorticales) es un enigma. Referencias Da Costa, N. M. and K. A. Martin (2010) Whose cortical column would that be? Front. Neuroanat. 4 (May 31): 16. Hortox, J. C. and D. L. Adams (2005) The cortical column: A structure without a function. Philos. Trans. R. Soc. Lond. B 360: 837-862. resumen, las aferencias sensitivas (o las eferencias motoras) que son particularmente importantes para una especie dada tienen relativamente más representación cortical. Aunque la organización topográfica de varias áreas somatosensitivas es similar, las propiedades funcionales de las neuronas Hacia las áreas corticales motora y premotora Hacia la amígdala y el hipocampo Corteza somatosensitiva secundaria 3a 3b 1 Áreas parietales 5, 7 2 Complejo ventral posterior del tálamo FIGURA 9.12 Las conexiones en el interior de la corteza somatosensitiva establecen jerarquías funcionales. Las aferencias desde el complejo ventral posterior del tálamo terminan en las áreas 3a, 3b, 1 y 2, con la máxima densidad de las proyecciones en el área 3b. Esta área a su vez proyecta de forma considerable a las áreas 1 y 2, y las funciones de estas áreas dependen de la actividad del área 3b. Todas las subdivisiones de la corteza somatosensitiva primaria proyectan a la corteza somatosensitiva secundaria; las funciones de SII son dependientes de la actividad de SI. Hubel, D. H. (1988) Eye, Brain, and Vision. Scientific American Library. New York: W. H. Freeman. Lorente de Nó, R. (1949) The structure of the cerebral cortex. Physiology of the Nervous System, 3rd Ed. New York: Oxford University Press. Mountcastle, V. B. (1957) Modality and topographic properties of single neurons of cat’s somatic sensory cortex. J. Neurophysiol. 20: 408-434. Mountcastle, V. B. (1998) Perceptual Neuroscience: The Cerebral Cortex. Cambridge: Harvard University Press. Purves, D., D. Riddle and A. LaMantia (1992) Iterated patterns of brain circuitry (or how the cortex gets its spots). Trends Neurosci. 15: 362369. Woolsey, T. A. and H. Van der Loos (1970) The structural organization of layer IV in the somatosensory region (SI) of mouse cerebral cortex. The description of a cortical field composed of discrete cytoarchitectonic units. Brain Res. 17: 205-242. de cada región son distintas. Los experimentos llevados a cabo en primates no humanos indican que las neuronas de las áreas 3b y 1 responden primariamente a estímulos cutáneos, mientras que las neuronas en el área 3a responden principalmente a la estimulación de los propioceptores; las neuronas del área 2 procesan tanto estímulos táctiles como propioceptivos. Estas diferencias en las propiedades de respuesta reflejan, al menos en parte, conjuntos paralelos de aferencias provenientes de clases de neuronas funcionalmente distintas en el complejo ventral posterior. Además, un rico patrón de conexiones corticocorticales entre las áreas SI contribuye significativamente a la elaboración de las propiedades de respuesta SI. El área 3b recibe la mayor parte de las aferencias del complejo ventral posterior y proporciona una proyección particularmente densa hacia las áreas 1 y 2. Esta disposición de las conexiones establece una jerarquía funcional en la cual el área 3b sirve como primer paso obligatorio en el procesamiento cortical de información somatosensitiva (Figura 9.12). Compatible con este punto de vista, las lesiones del área 3b en primates no humanos producen déficit profundos en todas las formas de sensaciones táctiles mediadas por mecanorreceptores cutáneos, mientras que las lesiones limitadas a las áreas 1 y 2 conducen a déficit parciales y a una incapacidad para utilizar la información táctil para discriminar la textura de los objetos (déficit del área 1) o el tamaño y la forma de los objetos (déficit del área 2). Existen incluso parcelaciones más finas de poblaciones de neuronas funcionalmente distintas dentro de áreas corticales únicas. Sobre la base de sus análisis de penetraciones con electrodos en la corteza somatosensitiva primaria, Vernon Mountcastle fue el primero en sugerir que las neuronas con propiedades similares de respuesta podrían estar agrupadas juntas en “columnas” funcionalmente distintas que atraviesan la profundidad de la corteza. 205 SISTEMA SOMATOSENSITIVO: TACTO Y PROPIOCEPCIÓN 3b (A) 1 (B) 8 Pie DV Pierna posterior 11 K Panículos del pie IV V III II I DI F 3b 1 Brazo DIV DIII DII Mentón Tronco Distal Labio inferior Boca Labio superior 9 H E D C 2 B 5 1 Medio A Proximal (C) Brazo De adaptación lenta K H J L I G C 11 9 10 B 5 8 3 A D 9 6 7 E H Mentón La bio Labio inf. superior De adaptación rápida F DV II I DI 6 0,5 mm 4 3 F J I Panículos de la mano IV III L Foot A. leg Mano DV 10 G 7 1 4 2 FIGURA 9.13 Las neuronas de la corteza somatosensitiva primaria forman columnas funcionalmente distintas. (A) El mapa somatosensitivo primario del mono cara de búho se basó, al igual que para el ser humano de la Figura 9.11, en la reactividad eléctrica de la corteza a la estimulación periférica. La amplificación de la derecha muestra las áreas 3b y 1 de Brodmann, que proyectan la mayor parte de la información mecanosensitiva cutánea. La disposición generalmente es similar a la determinada en los seres humanos. Obsérvese la presencia de regiones que están dedicadas a la representación de los dedos individuales. (B) La organización modular de las respuestas dentro de la representación de un único dedo, que muestra la localización de las penetraciones con electrodos que encontraron res- Más allá de la SI: vías corticocorticales y descendentes La información somatosensitiva se distribuye desde la corteza somatosensitiva primaria hasta los campos corticales de “orden puestas de adaptación rápida (azul claro) y lenta (azul oscuro) dentro de la superior”. Uno de estos centros corticales de orden superior, la representación del cuarto dedo. (C) Distribución de los campos receptivos corteza somatosensitiva secundaria (SII), se ubica en el marde adaptación lenta y adaptación rápida utilizados para obtener el gráfico de gen superior del surco lateral (véanse Figs. 9.10 y 9.11). La SII (B). Aunque los campos receptivos de estas diferentes clases de aferencias se superponen sobre la superficie cutánea, están segregados dentro de la recibe proyecciones convergentes de todas las subdivisiones de representación cortical. (A, de Kaas, 1983; C, de Sur y cols., 1984.) la SI y estas aferencias son necesarias para la función de la SII; las lesiones de SI eliminan las respuestas somatosensitivas de las neuronas SII. El área SII envía proyecciones hacia las estructuras límbicas, como la amígdala y el hipocampo (véanse Cap. 29 Estudios posteriores de penetraciones con electrodos finamente y 31). Se cree que estas últimas vías desempeñan un papel muy espaciados en el área 3b proporcionaron fuerte evidencia en apo- importante en el aprendizaje y la memoria táctil. yo de esta idea, lo que demostró que las neuronas con respuestas Las neuronas en SI también proyectan hacia las áreas pariea mecanorreceptores de adaptación rápida y lenta se agrupaban tales posteriores al área 2, sobre todo, las áreas 5a y 7b. Estas en zonas separadas dentro de la representación de un único dedo áreas reciben proyecciones directas del área 2 y, a su vez, envían (Figura 9.13). Se han descrito grupos similares de neuronas que aferencias a neuronas de las áreas motora y premotora del lóbulo responden preferencialmente a diferentes aferencias somatosen- frontal. Es una vía importante mediante la cual la información sitivas en las áreas 1 y 2. Esta organización modular de las áreas derivada de aferencias propioceptivas que señalan el estado accorticales, característica fundamental de la organización cortical, tual de ganancias en la contracción muscular accede a circuitos es especialmente pronunciada en las áreas corticales visuales que inician movimientos voluntarios. En términos generales, las (véase Cap. 11). Aunque estos patrones reflejan la especificidad proyecciones desde la corteza parietal hasta la corteza motora en los patrones subyacentes de las conexiones talamocorticales y son fundamentales para integrar la información sensitivomotora corticocorticales, la importancia funcional de las columnas sigue (véanse Caps. 20 y 26, que consideran las funciones de las regiosiendo poco clara (véase Recuadro 9B). nes de “asociación” de la corteza cerebral). 206 CAPÍTULO 9 Por último, una característica fundamental, pero a menudo descuidada del sistema somatosensitivo, es la presencia de proyecciones descendentes masivas. Estas vías se originan en campos corticales sensitivos y discurren hasta el tálamo, el tronco del encéfalo y la médula espinal. En efecto, las proyecciones descendentes desde la corteza somatosensitiva superan en número a las vías somatosensitivas ascendentes. Aunque su papel fisiológico no es bien conocido, en general se acepta (con cierto apoyo experimental) que las proyecciones descendentes modulan el flujo ascendente de información sensitiva a nivel del tálamo y del tronco del encéfalo. 3 2 5 1 Representación de la mano Corteza somatosensitiva (C) Representación de la mano dos meses después de la amputación del dedo 3 (B) Representación de la mano normal 5 Rostral El análisis de los mapas de la superficie cerebral en la corteza somatosensitiva primaria y las respuestas a patrones alterados de actividad en las aferencias periféricas resultaron efectivos a la hora de comprender el potencial para la reorganización de los circuitos corticales en los adultos. Jon Kaas y Michael Merzenich fueron los primeros en explorar esta cuestión al examinar el impacto de las lesiones periféricas (p. ej., el corte de un nervio que inerva la mano o la amputación de un dedo) sobre los mapas topográficos de la corteza somatosensitiva. Inmediatamente después de la lesión, se observó que la región correspondiente de la corteza no respondía. Sin embargo, después de algunas semanas, el área que no respondía se volvió reactiva a la estimulación de las regiones vecinas de la piel (Figura 9.14). Por ejemplo, al amputar el tercer dedo, las neuronas corticales que antes respondían a la estimulación de ese dedo respondían ahora a la estimulación de los dedos 2 o 4. Por lo tanto, la representación central de los dedos restantes se había ampliado para abarcar el territorio cortical que había perdido sus aferencias principales. Este “remapeo funcional” también ocurre en los núcleos somatosensitivos del tálamo y del tronco del encéfalo; en efecto, algo de la reorganización de los circuitos corticales puede depender de esta plasticidad subcortical concomitante. Este tipo de adaptación del sistema somatosensitivo pue- 4 1 3b 4 Plasticidad en la corteza cerebral del adulto (A) (A) Cerebro del mono búho 3 3 2 2 1 1 1 FIGURA 9.14 Cambios funcionales en la corteza somatosensitiva después de la amputación de un dedo. (A) Esquema de la corteza somatosensitiva en el mono cara de búho, que muestra la localización aproximada de la representación de la mano. (B) Representación de la mano en el animal antes de la amputación; los números corresponden a diferentes dígitos. (C) Mapa cortical determinado en el mismo animal dos meses después de la amputación del dedo 3. El mapa ha cambiado sustancialmente; las neuronas en el área que antes respondían a la estimulación del dedo 3 ahora responden a la estimulación de los dedos 2 y 4. (De Merzenich y cols., 1984.) de contribuir a la alteración de la sensibilidad de los miembros “fantasma” después de la amputación (véase Recuadro 10D). Se han demostrado cambios plásticos similares en las cortezas visual, auditiva y motora, lo que sugiere que cierta capacidad para reorganizarse después de la privación periférica o de una lesión es una propiedad general de la neocorteza madura. Después de la estimulación diferencial 4 2 Lateral Antes de la estimulación diferencial 4 Caudal 2 (C) 5 4 1 (B) 5 3 5 FIGURA 9.15 Expansión funcional de una representación cortical debido a una tarea conductual repetitiva. (A) Un mono cara de búho fue entrenado en una tarea que requería el uso importante de los dedos 2, 3 y, en ocasiones, 4. (B) Mapa de los dedos en la corteza somatosensitiva primaria antes del entrenamiento. (C) Después de varios meses de “práctica”, una región más grande de la corteza contenía neuronas activadas 1 mm por los dedos utilizados en la tarea. Obsérvese que las disposiciones específicas de las representaciones de los dedos son algo diferentes de aquellas del mono que se muestran en la Figura 9.14, lo que indica la variabilidad de la representación cortical en animales particulares (De Jerkins y cols., 990.) SISTEMA SOMATOSENSITIVO: TACTO Y PROPIOCEPCIÓN También pueden ocurrir cambios apreciables en la representación cortical en respuesta a los cambios fisiológicos en la experiencia sensitiva o motora. Por ejemplo, si un mono es entrenado para utilizar un dedo específico para una tarea particular que se repite muchas veces, la representación funcional de ese dedo, determinada mediante mapeo electrofisiológico, puede expandirse a expensas de los otros dedos (Figura 9.15). De hecho, pueden detectarse cambios importantes en los campos receptivos de las neuronas somatosensitivas cuando se bloquea transitoriamente un nervio periférico con un anestésico local. La pérdida transitoria de aferencias sensitivas desde un área pequeña de piel induce una reorganización reversible de los campos receptivos de las neuronas corticales y subcorticales. Durante este período, las neuronas asumen nuevos campos receptivos que responden a la estimulación táctil de la piel que rodea a la región anestesiada. Una vez que ceden los efectos del anestésico local, los campos receptivos de las neuronas corticales y subcorticales retornan a su tamaño habitual. La experiencia frecuente de un área anestesiada de la piel que se percibe desproporcionadamente grande –como se experimenta, por ejemplo, después de la anestesia dental– puede ser consecuencia de este cambio transitorio. A pesar de estas observaciones intrigantes, no se conoce el mecanismo, el propósito ni la importancia de la reorganización de los mapas sensitivomotores que ocurre en la corteza del adulto. Indudablemente, ocurren cambios en los circuitos corticales del encéfalo del adulto. Sin embargo, siglos de observaciones clínicas indican que estos cambios pueden tener un valor limitado para la recuperación de la función después de la lesión cerebral y es muy probable que puedan conducir a síntomas que impidan hacer nada más que mejorar la calidad de vida después del daño neural. Dado su carácter rápido y reversible, la mayoría de estos cambios en la función cortical probablemente refleje alteraciones en la fuerza de las sinapsis ya presentes. En efecto, encontrar formas para prevenir o redirigir los eventos sinápticos que subyacen a la plasticidad inducida por la lesión podría reducir el impacto a largo plazo del daño encefálico agudo. Resumen Los componentes del sistema somatosensitivo procesan información transmitida por los estímulos mecánicos que inciden sobre la superficie corporal (mecanorrecepción cutánea) o que son generados en el cuerpo propiamente dicho (propiocepción). El procesamiento somatosensitivo es realizado por neuronas distribuidas en varias estructuras encefálicas que están conectadas por vías ascendentes y descendentes. La transmisión de la información mecanosensitiva aferente desde la periferia hasta el encéfalo comienza con distintos tipos de receptores que inician potenciales de acción. Esta actividad es transmitida centralmente a través de cadenas de células nerviosas, denominadas neuronas de primer, de segundo y de tercer orden. Las neuronas de primer orden se localizan en los ganglios de las raíces dorsales y de los nervios craneales. Las neuronas de segundo orden se sitúan en los núcleos del tronco encefálico. Las neuronas de tercer 207 orden se encuentran en el tálamo, desde donde proyectan hacia la corteza cerebral. Estas vías están dispuestas topográficamente en todo el sistema, y la cantidad del espacio cortical y subcortical asignada a las distintas partes del cuerpo es proporcional a la densidad de los receptores periféricos. Estudios en primates no humanos muestran que regiones corticales específicas corresponden a cada submodalidad funcional; por ejemplo, el área 3b procesa la información proveniente de receptores cutáneos de bajo umbral, mientras que el área 3a procesa aferencias provenientes de propioceptores. Por lo tanto, al menos dos criterios amplios operan en la organización del sistema somatosensitivo: modalidad y somatotopía. El resultado final de esta interacción compleja es la representación perceptual unificada del cuerpo y su interacción continua con el entorno. Lecturas adicionales Revisiones Barnes, S. J. and G. T. Finnerty (2010) Sensory experience and cortical rewiring. Neuroscientist 16: 186-198. Chapin, J. K. (1987) Modulation of cutaneous sensory transmission during movement: Possible mechanisms and biological significance. In Higher Brain Function: Recent Explorations of the Brain’s Emergent Properties, S. P. Wise (ed.). New York: John Wiley and Sons, pp. 181-209. Darian-Smith, I. (1982) Touch in primates. Annu. Rev. Psychol. 33: 155-194. Johansson, R. S. and A. B. Vallbo (1983) Tactile sensory coding in the glabrous skin of the human. Trends Neurosci. 6: 27-32. Johnson, K. O. (2002) Neural basis of haptic perception. In Seven’s Handbook of Experimental Psychology, 3rd Ed., H. Pashler and S. Yantis (eds.). Vol 1: Sensation and Perception. New York: Wiley, pp. 537-583. Kaas, J. H. (1990) Somatosensory system. In The Human Nervous System, G Paxinos (ed.). San Diego: Academic Press, pp. 813-844. Kaas, J. H. (1993) The functional organization of somatosensory cortex in primates. Ann. Anat. 175: 509-519. Kaas, J. H. and C. E. Collins (2003) The organization of somatosensory cortex in anthropoid primates. Adv. Neurol. 93: 57-67. Mountcastle, V. B. (1975) The view from within: Pathways to the study of perception. Johns Hopkins Med. J. 136: 109-131. Nicolelis, M. A. and E. E. Fanselow (2002) Thalamocortical optimization of tactile processing according to behavioral state. Nature Neurosci. 5 (6): 517523. Petersen, R. S., S. Panzeri and M. E. Diamond (2002) Population coding in somatosensory cortex. Curr. Opin. Neurobiol. 12: 441-447. Woolsey, C. (1958) Organization of somatic sensory and motor areas of the cerebral cortex. In Biological and Biochemical Bases of Behavior, H. F. Harlow and C. N. Woolsey (eds.). Madison, WI: University of Wisconsin Press, pp. 63-82. Artículos originales importantes Adrian, E. D. and Y. Zotterman (1926) The impulses produced by sensory nerve endings. II. The response of a single end organ. J. Physiol. 61: 151-171. Friedman, R. M., L. M. Chen and A. W. Roe (2004) Modality maps within primate somatosensory cortex. Proc. Natl. Acad. Sci. USA 101: 12724-12729. Johansson, R. S. (1978) Tactile sensibility of the human hand: Receptive field characteristics of mechanoreceptive units in the glabrous skin. J. Physiol. (Lond.) 281: 101-123. Johnson, K. O. and G. D. Lamb (1981) Neural mechanisms of spatial tactile discrimination: Neural patterns evoked by Braillelike dot patterns in the monkey. J. Physiol (Lond.) 310: 117-144. Jones, E. G. and D. P. Friedman (1982) Projection pattern of functional components of thalamic ventrobasal complex on monkey somatosensory cortex. J. Neurophysiol. 48: 521-544. 208 CAPÍTULO 9 Jones, E. G. and T. P. S. Powell (1969) Connexions of the somatic sensory cortex of the rhesus monkey. I. Ipsilateral connexions. Brain 92: 477-502. Moore, C. I., S. B. Nelson and M. Sur (1999) Dynamics of neuronal processing in rat somatosensory cortex. Trends Neurosci. 22: 513-520. Lamotte, R. H. and M. A. Srinivasan (1987) Tactile discrimination of shape: Responses of rapidly adapting mechanoreceptive afferents to a step stroked across the monkey fingerpad. J. Neurosci. 7: 1672-1681. Laubach, M. J. Wessber and M. A. L. Nicolelis (2000) Cortical ensemble activity increasingly predicts behavior outcomes during learning of a motor task. Nature 405: 567-571. Moore, C. I. and S. B. Nelson (1998) Spatiotemporal subthreshold receptive fields in the vibrissa representation of rat primary somatosensory cortex. J. Neurophysiol. 80: 2882-2892. Nicolelis, M. A. L., L. A. Baccala, R. C. S. Lin and J. K. Chapin (1995) Sensorimotor encoding by synchronous neural ensemble activity at multiple levels of the somatosensory system. Science 268: 1353-1359. Sur, M. (1980) Receptive fields of neurons in areas 3b and 1 of somatosensory cortex in monkeys. Brain Res. 198: 465-471. Wall, P. D. and W. Noordenhos (1977) Sensory functions which remain in man after complete transection of dorsal columns. Brain 100: 641-653. Zhu, J. J. and B. Connors (1999) Intrinsic firing patterns and whisker-evoked synaptic responses of neurons in the rat barrel cortex. J. Neurophysiol. 81: 11711183. 209 CAPÍTULO 2 10 Dolor Aspectos generales AUNQUE RESULTA NATURAL SUPONER que las sensaciones asociadas con los estímulos nocivos surgen de la estimulación excesiva de los mismos receptores que generan otras sensaciones somáticas (como las explicadas en el Capítulo 9), no sucede así. La percepción de los estímulos nocivos, denominada nocicepción, depende de receptores y vías específicos. Además, la respuesta de un organismo a los estímulos nocivos es multidimensional, y comprende componentes discriminativos, afectivos y motivacionales. La distribución central de la información nociceptiva es correspondientemente compleja, y comprende múltiples áreas en tronco del encéfalo, tálamo y encéfalo anterior. Dada la importancia fundamental del dolor en la práctica clínica (tanto como diagnóstico y como foco del tratamiento) y los muchos aspectos de la fisiología y la farmacología del dolor que no se conocen aún de forma perfecta convierten a la nocicepción en un área de investigación extremadamente activa. Nociceptores Las terminaciones de las células nerviosas relativamente poco especializadas que inician la sensación de dolor se denominan nociceptores (noci deriva del latín nocere, “herir”). Al igual que otros receptores cutáneos y subcutáneos, transducen distintos estímulos en potenciales del receptor, que a su vez desencadenan potenciales de acción aferentes. Más aun, los nociceptores, al igual que otros receptores somatosensitivos, surgen de los cuerpos celulares en los ganglios de las raíces dorsales (o el ganglio trigeminal) que envían una prolongación axónica a la periferia y la otra a la médula espinal o el tronco encefálico (véase Figura 9.1). Dado que los axones nociceptivos periféricos culminan en “terminaciones libres” no especializadas desde el punto de vista morfológico, por convención se categoriza a los nociceptores según las propiedades de los axones asociados con ellos (véase Cuadro 9.1). Como se describió en el capítulo anterior, los receptores somatosensitivos responsables de la percepción de estímulos mecánicos inocuos se asocian con axones mielínicos que tienen velocidades de conducción relativamente rápidas. Por el contrario, los axones asociados con los nociceptores conducen en forma relativamente lenta, y son solo algo mielínicos o, lo que es más común, amielínicos. En consecuencia, los axones que transmiten información acerca del dolor caen en el grupo Aδ de axones amielínicos, que conducen a alrededor de 5-30 m/s, o en el grupo de fibras C de axones amielínicos, que conducen a velocidades en general inferiores a 2 m/s. 210 CAPÍTULO 10 (A) (B) Registrar Estímulo de calor Nociceptor 45°C Estímulo Termorreceptor no nociceptivo (C) FIGURA 10.1 Base neuronal del dolor. Demostración experimental de que la nocicepción involucra neuronas especializadas, no simplemente una descarga excesiva de las neuronas que responden a las intensidades normales del estímulo. (A) Disposi- Termorreceptor Magnitud de la respuesta aferente (potenciales de acción por segundo) ción para el registro transcutáneo de las fibras nerviosas. (B) En el rango del estímulo doloroso, los axones de los termorreceptores disparan potenciales de acción con la misma frecuencia que en temperaturas inferiores; sin embargo, la cantidad y la frecuencia de los potenciales de acción en la fibra nociceptiva aumentan. (Obsérvese que los 43 ºC son el umbral aproximado para el dolor.) (C) Resumen de resultados. (De Fields, 1987.) Por lo tanto, aun cuando la conducción de toda la información nociceptiva es relativamente lenta, hay vías rápidas y lentas para el dolor. Hace cierto tiempo, en algunos estudios llevados a cabo tanto en seres humanos como en animales de experimentación se demostró que los axones de conducción rápida que controlan la sensación somatosensitiva no participan en la transmisión de dolor. En la Figura 10.1 se muestra un experimento típico de este tipo. Los axones periféricos que responden a estímulos mecánicos o térmicos no dolorosos, no descargan a una mayor frecuencia cuando los estímulos dolorosos se aplican en la misma región de la superficie cutánea. Por otra parte, los axones nociceptivos comienzan a descargar solo cuando la fuerza del estímulo (un estímulo térmico en el ejemplo de la Figura 10.1) alcanza niveles elevados; con la misma intensidad del estímulo, otros termorreceptores descargan con una frecuencia no diferente de la frecuencia máxima ya lograda en el intervalo de temperatura no dolorosa, lo que indica que hay tanto termorreceptores nociceptivos como no nociceptivos. Igualmente importante, una estimulación directa sobre las aferencias somatosensitivas de gran diámetro a cualquier frecuencia, no produce sensaciones descritas como dolorosas. Por el contrario, las fibras Aδ y C de conducción más lenta y menor diámetro son activas cuando se entregan estímulos dolorosos; asimismo, cuando se estimulan en forma directa en los seres humanos producen sensaciones de dolor. Entonces, ¿cómo consiguen estas diferentes clases de nociceptores dar lugar a la percepción del dolor? Como ya mencionamos, una forma para determinar la respuesta es estimular di- Nociceptor 0 40 45 Temperatura (°C) 50 ferentes nociceptores en seres humanos voluntarios mientras se anotan las sensaciones comunicadas. En general, se describieron dos categorías de sensaciones dolorosas: un primer dolor agudo y una sensación más tardía difusa y más duradera que suele llamarse segundo dolor (Figura 10.2A). La estimulación de los axones Aα y Aβ de conducción rápida en los nervios periféricos no produce la sensación de dolor. Sin embargo, cuando se eleva la intensidad del estímulo hasta un nivel que activa un subgrupo de fibras Aδ, se comunica una sensación de hormigueo o, si la estimulación es de intensidad suficiente, una sensación de dolor agudo. Si se incrementa aun más la intensidad del estímulo, de modo que entren en juego los axones de las fibras C de conducción lenta y diámetro pequeño, se experimenta una sensación de dolor más sordo y duradero. También es posible anestesiar de manera selectiva las fibras C y las fibras Aδ; en general, en estos experimentos de bloqueo selectivo se confirmó que las fibras Aδ son responsables del primer dolor y que las fibras C responden por el segundo dolor más sordo y duradero (Figura 10.2B,C). Se sabe ahora que los nociceptores Aδ de conducción más rápida pertenecen a dos clases principales. Las fibras tipo i Aδ responden a una estimulación mecánica y química peligrosamente intensa pero tienen umbrales de calor relativamente altos, mientras que las fibras tipo ii Aδ tienen sensibilidades complementarias –es decir, umbrales para el calor mucho más bajos, pero umbrales muy altos para la estimulación mecánica–. Por lo tanto, el sistema Aδ tiene vías especializadas para la transmisión del calor y los estímulos nociceptivos mecánicos. La mayoría de los nociceptores de fibras C amielínicas de conducción más lenta responden a todas las formas de estímulos nociceptivos –térmicos, mecánicos y químicos– y por lo tanto se dice que son polimodales. Sin embargo, los nociceptores de las fibras C también son heterogéneos, con subgrupos que responden preferentemente al calor o a la estimulación química más que a la estimulación mecánica. Otros subtipos de nociceptores de fibras C responden especialmente a los irritantes químicos, las sustancias ácidas o el frío. En resumen, cada una de las clases principales de aferencias nociceptivas está compuesta por múltiples subtipos con diferentes perfiles de sensibilidad. Intensidad subjetiva del dolor DOLOR (A) Fibra Aδ Fibra C (B) Primer dolor Segundo dolor 211 (C) Tiempo FIGURA 10.2 Primero y segundo dolor. El dolor puede separarse en una percepción temprana de dolor agudo y una sensación más tardía de una cualidad más sorda y quemante. (A) El primero y el segundo dolor, como se denomina a estas sensaciones, son transmitidos por axones diferentes, como puede demostrarse por (B) el bloqueo selectivo de los axones mielínicos de conducción más rápida que transmiten dolor o (C) el bloqueo de las fibras C de conducción más lenta que transmiten la sensación del segundo dolor. (De Fields, 1990.) Transducción y transmisión de las señales nociceptivas Dada la variedad de estímulos (mecánicos, térmicos y químicos) que pueden originar sensaciones dolorosas, la transducción de señales nociceptivas es una tarea compleja. Si bien persisten muchos interrogantes, algunas nociones provienen de la identificación de receptores específicos asociados a la sensación de calor nocivo. El umbral para percibir un estímulo térmico como nocivo es de alrededor de 43 °C (110 °F) y este umbral doloroso corresponde a la sensibilidad de los subtipos de terminaciones nociceptivas de fibras Aδ y C. El receptor que confiere esta sensibilidad al calor también confiere sensibilidad a la capsaicina, el ingrediente de los pimientos chiles responsable de las parestesias o la sensación ardiente producida por los alimentos picantes (Recuadro 10A). El denominado receptor vanilloide (TRPV1), que se encuentra tanto en las fibras C como en las Aδ, es miembro de una familia más grande de canales del potencial transitorio de receptor (TRP), identificada por primera vez en estudios de las vías de fototransducción en las moscas de las fruta que actualmente se sabe que comprenden gran cantidad de receptores sensibles a diferentes intervalos de calor y frío. Desde el punto de vista estructural, los canales TRP se asemejan a los del potasio con puerta de voltaje o a los canales con puerta de nucleótidos cíclicos, con seis dominios transmembrana con un poro entre los dominios 5 y 6. En condiciones de reposo, el poro del canal está cerrado. En el estado activado abierto, estos receptores permiten la entrada de sodio y calcio que inicia la generación de potenciales de acción en las fibras nociceptivas. Como el mismo receptor responde al calor y a la capsaicina, no es sorprendente que muchas personas experimenten que el gusto del chile es “caliente”. Sin embargo, un enigma es por qué el sistema nervioso ha desarrollado receptores que son sensibles a una sustancia química en los pimientos chiles. Como sucede con otros compuestos vegetales que activan selectivamente los receptores neurales (véase la explicación de los opioides, pp. 224-225), parece probable que los receptores TRVP1 detecten sustancias endógenas cuya estructura química se asemeja a la capsaicina. De hecho, algunas pruebas recientes sugieren que los “endovainilloides” son producidos por tejidos periféricos en respuesta a la lesión y que estas sustancias, junto con otros factores, contribuyen a la respuesta nociceptiva a la lesión. Los receptores responsables de la transducción de formas mecánicas y químicas de estimulación nociceptiva no son tan bien conocidos. Se han identificado algunos candidatos diferentes para los mecanotransductores, incluidos otros miembros de la familia TRP (TRPV2 y TRPA1) y algunos miembros de la familia ASIC (canales iónicos sensibles al ácido, “acid-sensing ion channels”). Los canales TRP también parecen ser responsables de la detección de irritantes químicos en el entorno. Se ha demostrado que TRPA1 en particular es sensible a un grupo diverso de irritantes químicos, que incluyen los ingredientes picantes en las plantas de mostaza y ajo, así como irritantes volátiles presentes en el gas de las lágrimas, los gases de escape de los vehículos y los cigarrillos. El subtipo de canal ASIC3 es expresado específicamente en los nociceptores y también está bien representado en las fibras que inervan el músculo esquelético y cardíaco. Se cree que los canales ASIC3 son responsables del dolor muscular y cardíaco resultado de los cambios en el pH asociado con isquemia. La base molecular compleja de la mecanonocicepción y la quimionocicepción es un área de investigación activa fundamental para el conocimiento de los pasos iniciales en las vías neurales que contribuyen al dolor. Los potenciales graduados que se originan en receptores de las ramas distales de las fibras nociceptivas deben transformar- 212 CAPÍTULO 10 RECUADRO 10A Capsaicina La capsaicina, el principal ingrediente responsable de la acritud de los pimientos picantes, es ingerida diariamente por más de un tercio de la población del mundo. La capsaicina activa respuestas en un subgrupo de fibras C nociceptivas (nociceptores polimodales) al abrir canales iónicos con puerta de ligando que permiten el ingreso de Na+ y Ca2+. Uno de estos canales, TRPV1, se clonó y se observó que era activado por capsaicina, ácido y anandamida (un compuesto endógeno que también activa los receptores de cannabanoides) y por el calentamiento del tejido hasta unos 43 °C. En consecuencia, es probable que la anandamida y la temperatura sean los activadores endógenos de estos canales. Los ratones cuyos receptores TRPV-1 fueron eliminados genéticamente beben soluciones de capsaicina como si fueran agua. Se encontraron receptores de capsaicina en los nociceptores polimodales de todos los mamíferos, pero no están presentes en las aves (lo que ha conducido a la producción de alpiste mezclado con capsaicina a prueba de ardillas). Cuando se aplica a las membranas mucosas de la cavidad oral, la capsaicina actúa como irritante y produce reacciones protectoras. Cuando se inyecta en la piel, genera un dolor ardiente e hiperalgesia a los estímulos térmicos y mecánicos. Las aplicaciones repetidas de capsaicina también desensibilizan las fibras dolorosas e impiden que los neuromoduladores como la sustancia P, VIP y somatostatina sean liberados por las terminaciones nerviosas periféricas y centrales. En consecuencia, la capsaicina se utiliza clínicamente como agente analgésico y antiinflamatorio. Suele aplicarse en forma tópica en una crema (al 0,075%) para aliviar el dolor asociado (A) con artritis, neuralgia posherpética, mastectomía y neuralgia del trigémino. Por lo tanto, este notable irritante químico no solo da placer al gusto en gran escala, sino que también es útil para aliviar el dolor. Bibliografía Caterina, M. J., M. A. Schumacher, M. Tominaga, T. A. Rosen, J. D. Levine and D. Julius (1997) The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 389: 816-766. Caterina, M. J. and 8 others (2000) Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 288: 306-313. Szallasi, A. and P. M. Blumberg (1999) Vanilloid (capsaicin) receptors and mechanisms. Pharm. Reviews 51: 159-212. Tominaga, M. and 8 others (1998) The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron 21: 531-543. Zygmunt, P. M. and 7 others (1999) Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature 400: 452-457. (B) Capsaicina CH3O Chile rojo Habanero O N H HO Jalapeño (D) Capsaicina Ca2+ Na+ Calor (C) H+ Exterior Interior (A) Algunos pimientos populares que contienen capsaicina. (B) Estructura química de la capsaicina. (C) Molécula de la capsaicina. (D) Diagrama del Receptor VR-1 canal de receptor de VR-1/capsaicina. Este canal puede ser activado por capsaicina intercelularmente o por calor o protones (H+) en la superficie celular. se en potenciales de acción para ser transmitidos a las sinapsis en el asta dorsal de la médula espinal. Los canales del sodio y del potasio con puerta del voltaje son fundamentales para este proceso (véase Capítulo 4), y un subtipo específico de canal del sodio –Nav1.7– parece ser especialmente importante para la transmisión de la información nociceptiva. La alteración de la actividad de Nav1.7 es responsable de distintos trastornos de dolor humano. Las mutaciones del gen NAV1.7 que conducen a la pérdida de la función de este canal producen incapacidad para detectar estimulación nociva, mientras que las mutaciones que conducen a hiperexcitabilidad del canal se asocian con trastornos dolorosos que causan sensaciones ardientes intensas. El gen NAV1.8 es expresado intensamente por los nociceptores de fibras C y, sobre la base de estudios en ratones, la proteína se ha asociado con la transmisión de información mecánica y térmica nociva. El desarrollo de anestésicos locales específicos de estas DOLOR sustancias de los canales del sodio puede seguir siendo la clave para el tratamiento de distintos síndromes dolorosos intratables. Las vías centrales del dolor son distintas de las vías mecanosensitivas Las vías responsables del dolor se originan con otras neuronas sensitivas en los ganglios de las raíces dorsales y, al igual que otras células nerviosas sensitivas, los axones centrales de las células nerviosas nociceptivas ingresan en la médula espinal a través de las raíces dorsales (Figura 10.3A). Cuando estos axones de proyección central alcanzan el asta dorsal de la médula espinal, se ramifican en colaterales ascendentes y descendentes, los que forman el tracto dorsolateral de Lissauer (llamado así en honor al neurólogo alemán que describió esta vía por primera vez a fines del siglo xix). En los casos típicos, los axones del tracto de Lissauer se dirigen hacia arriba y hacia abajo por uno o dos segmentos de la médula espinal antes de penetrar en la sustancia gris del asta dorsal. Una vez allí, los axones dan origen (A) Izquierda Derecha Axón ascendente del tracto anterolateral Aferencia nociceptivo Ganglio de la raíz dorsal Tracto de Lissauer (B) Fibra C Fibra Ab Zona marginal Sustancia gelatinosa Núcleo propio Base del asta dorsal 1 2 3 4 5 6 Decusación 213 a ramas que hacen contacto con neuronas de segundo orden situadas en las láminas de Rexed 1, 2 y 5. (Las láminas de Rexed son las divisiones descriptivas de la sustancia gris medular en el corte transversal, llamadas así en honor al neuroanatomista que describió estos detalles en la década de 1950; véanse Cuadro A1 y Figura A6 en el Apéndice). Las láminas 1 y 5 contienen neuronas de proyección cuyos axones discurren hasta los puntos diana en el tronco encefálico y el tálamo. La lámina 2 contiene las interneuronas de la médula espinal. Estas terminaciones aferentes están organizadas en una forma específica de las láminas; por ejemplo, las fibras C terminan exclusivamente en las láminas de Rexed 1 y 2, mientras que las fibras Aδ terminan en las láminas 1 y 5. Las aferencias nociceptivas (Aβ) terminan primariamente en la lámina 5, y un subgrupo de las neuronas de la lámina 5 recibe aferencias convergentes desde aferencias nociceptivas y no nociceptivas. Estas neuronas multimodales de la lámina 5 se denominan neuronas de rango dinámico amplio. Algunas de ellas reciben también aferencias sensitivas viscerales, lo que las convierte en un sustrato probable del dolor referido (es decir, el dolor que se origina en el daño de los órganos viscerales pero que es percibido erróneamente como proveniente de una localización somática). El ejemplo clínico más común es la angina, en la cual la escasa perfusión del músculo cardíaco es percibida erróneamente como dolor en la pared torácica, el hombro y el brazo y la mano izquierdos (Recuadro 10B). Los axones de las neuronas de segundo orden en las láminas 1 y 5 del asta dorsal de la médula espinal atraviesan la línea media y ascienden hasta el tronco del encéfalo y el tálamo en el cuadrante anterolateral (también denominado ventrolateral) de la mitad contralateral de la médula espinal (Figura 10.3B). Por esta razón, la vía neural que transmite información sobre dolor y temperatura a los centros superiores se denomina a menudo sistema anterolateral, para distinguirla del sistema de columna dorsal-lemnisco medio que transmite información mecanosensitiva (véase Capítulo 9). Los axones que transmiten información para el sistema anterolateral y el sistema columna dorsal-lemnisco medio discurren en diferentes partes de la sustancia blanca de la médula espinal. Esta diferencia proporciona un signo de relevancia clínica que es útil para definir la localización de una lesión en la médula espinal. Los axones de las neuronas de primer orden para el sistema FIGURA 10.3 Sistema anterolateral. (A) Las aferencias primarias en los ganglios de las raíces dorsales envían sus axones a través de las raíces dorsales para terminar en el asta dorsal de la médula espinal. Las aferencias se ramifican y discurren por varios segmentos hacia arriba y debajo de la médula espinal en el tracto de Lissauer y dan origen a las ramas colaterales que terminan en el asta dorsal. Las neuronas de segundo orden en el asta dorsal envían sus axones (negro) a través de la línea media para ascender hasta niveles superiores en la columna anterolateral de la médula espinal. (B) Las aferencias de las fibras C terminan en las láminas de Rexed 1 y 2 del asta dorsal, mientras que las fibras Aδ terminan en las capas 1 y 5. Los axones de segundo orden en las láminas 1 y 5 cruzan la línea media y ascienden hasta los centros superiores. 214 CAPÍTULO 10 RECUADRO 10B Dolor referido Es sorprendente, pero hay pocas neuronas en el asta dorsal de la médula espinal especializadas solo en la transmisión del dolor visceral (interno). Como es evidente, nosotros reconocemos ese dolor, pero se transmite centralmente a través de las neuronas del asta dorsal que también están involucradas en el dolor cutáneo. Como resultado de esta disposición, la enfermedad de un órgano interno a veces se percibe como dolor cutáneo. Como consecuencia, el paciente puede concurrir a la consulta médica por dolor en un sitio que no es el de origen real; esto se denomina dolor referi- do. El ejemplo clínico más frecuente es el dolor anginoso (el que surge del músculo cardíaco cuando no es perfundido con la sangre suficiente) atribuido a la pared torácica superior, con irradiación al brazo y la mano izquierdos. Otros ejemplos importantes son el dolor vesicular referido a la región escapular, el dolor esofágico referido a la pared torácica, el dolor ureteral (p. ej., por la evacuación de un cálculo renal) referido a la pared abdominal inferior, el dolor vesical referido al periné y el dolor del apéndice inflamado referido a la pared abdominal inferior que rodea el ombligo. El conocimiento del dolor referido puede conducir a un diagnóstico más certero que, de otro modo, podría omitirse. Bibliografía Capps, J. A. and G. H. Coleman (1932) An Experimental and Clinical Study of Pain in the Pleura, Pericardium, and Peritoneum. New York: Macmillan. Head, H. (1893) On disturbances of sensation with special reference to the pain of visceral disease. Brain 16: 1-32. Kellgrew, J. H. (1939-1942) On the distribution of pain arising from deep somatic structures with charts of segmental pain Clin. Sci. 4: 35-46. Corazón Urinario/vesical Uréter izquierd o Próstata Esófago Ejemplos de dolor que surgen de un trastorno visceral referido a una región cutánea (color). columna dorsal-lemnisco medio entran en la médula espinal, giran y ascienden por las columnas dorsales homolaterales en todo el camino en la médula, donde hacen sinapsis sobre las neuronas de los núcleos de la columna dorsal (Figura 10.4A). Los axones de las neuronas de los núcleos de la columna dorsal atraviesan luego la línea media y ascienden hasta el tálamo homolateral. Por el contrario, el punto de cruzamiento para la información transmitida por el sistema anterolateral se ubica en el interior de la médula espinal. Las neuronas de primer orden que constituyen al sistema anterolateral terminan en el asta dorsal, y las neuronas de segundo orden en el asta dorsal envían sus axones a través de la línea media y ascienden en la mitad contralateral de la médula espinal (en el asta anterolateral) hasta sus puntos diana en el tálamo y el tronco encefálico. Debido a esta diferencia anatómica en el sitio de decusación, una lesión unilateral de la médula espinal conduce a síntomas del sistema columna dorsal-lemnisco medio (pérdida de sensibilidad de tacto, presión, vibración y propiocepción) del lado del cuerpo homolateral a la lesión, y síntomas anterolaterales (déficit de percepción de dolor y temperatura) del lado contralateral DOLOR 215 FIGURA 10.4 Vías nociceptivas y mecanosensitivas. Como se muestra aquí, el sistema anterolateral (azul) cruza y asciende por la columna anterolateral de la médula espinal, mientras que el sistema de columnas dorsales-lemnisco medial (rojo) asciende por la columna dorsal homolateral. Una lesión restringida a la mitad izquierda de la médula espinal produce pérdida sensitiva disociada y déficit mecanosensitivo en la mitad izquierda del cuerpo, con la aparición de un déficit termoalgésico del lado derecho. Sensibilidad normal Columna dorsal Lesión Columna anterolateral Zona de pérdida completa de sensibilidad Lesión (torácica inferior) Disminución de la sensibilidad de discriminación de dos puntos, vibración y propiocepción Disminución de la sensibilidad termoalgésica Aferencias nociceptivos Aferencias mecanorreceptores Derecha Izquierda Derecha Izquierda RECUADRO 10C Vía de las columnas dorsales para el dolor visceral En los Capítulos 9 y 10 se presenta un marco de trabajo para considerar las vías neurales centrales que transmiten señales mecanosensitivas inocuas y señales dolorosas desde los orígenes cutáneos y somáticos profundos. Si se consideran solo las señales derivadas del cuerpo por debajo de la cabeza, la información mecanosensitiva discriminativa y propioceptiva viaja hasta el tálamo ventral posterior a través del sistema columna dorsal-lemnisco medial (véase Figura 9.6A), mientras la información nociceptiva se dirige hasta los mismos relevos talámicos (y otros) a través de los sistemas anterolaterales (véase Figura 10.3A). No obstante, ¿de qué modo las señales dolorosas que surgen en los órganos viscerales de la pelvis, el abdomen y el tórax ingresan en el sistema nervioso central y finalmente alcanzan la conciencia? La respuesta se obtuvo a partir de un componente recién descubierto de la vía de columnas dorsales-lemnisco medial que transmite la nocicepción visceral. Aunque en el Capítulo 21 se presentará más información sobre los sistemas que reciben y procesan la información sensitiva visceral, en este punto vale la pena considerar este componente de las vías del dolor y el modo en que esta vía particular comenzó a tener impacto en la medicina clínica. Las aferencias viscerales primarias de las vísceras pelvianas y abdominales entran en la médula espinal y hacen sinapsis sobre neuronas de segundo orden en el asta dorsal de la médula espinal lumbosacra. Como se explica en el Recuadro 10B y en el Capítulo 21, algunas de estas neuronas de segundo orden son células que dan origen a los sistemas anterolaterales y contribuyen a los patrones de dolor visceral referido. Sin embargo, otras neuronas –tal vez fundamentalmente las que dan origen a las señales nociceptivas– hacen sinapsis sobre neuronas en la región gris intermedia de la médula espinal cerca del conducto central. Por su parte, estas neuronas no envían sus axones a través de la sustancia blanca anterolateral de la médula espinal (como podría esperarse para una vía para el dolor), sino por las columnas dorsales en una posición muy cercana a la línea media (véase Figura A). Asimismo, neuronas de segundo orden en la médula espinal torácica que transmiten señales nociceptivas desde las vísceras torácicas envían sus axones rostralmente a través de las columnas dorsales a lo largo del tabique intermedio dorsal, cerca de la división de los fascículos grácil y cuneiforme. Estos axones de segundo orden hacen sinapsis luego en los núcleos de las columnas dorsales en el bulbo raquídeo caudal, donde las neuronas dan origen a las fibras arciformes que forman el lemnisco medial contralateral y finalmente hacen sinapsis sobre las neuronas de proyección talamocorticales en el tálamo ventral posterior. En la actualidad, esta proyección sensitiva visceral de los cordones posteriores parece ser la vía principal por la cual las sensaciones dolorosas originadas en las vísceras son detectadas y discriminadas. Varias observaciones apoyan esta conclusión: 1) las neuronas en el núcleo ventral posterolateral, el núcleo grácil y cerca del conducto central de la médula espinal responden a la estimulación visceral nociva; 2) las respuestas de las neuronas en el núcleo ventral posterolateral y el núcleo grácil a esa estimulación se reducen mucho con las lesiones espinales de las columnas dorsales (Figura B), pero no con lesiones de la sustancia blanca anterolateral, y 3) la infusión de fármacos que bloquean la transmisión sináptica nociceptiva en la región gris intermedia de la médula espinal (Continúa en la página siguiente) 216 CAPÍTULO 10 RECUADRO 10C (continúa) (A) Cerebro (B) Lesión simulada Lesión de columna dorsal Antes de la cirugía 4 meses después de la cirugía (C) Complejo nuclear ventral posterior del tálamo Aguja Corteza insular Columna dorsal Asta dorsal Mesencéfalo Tracto gastrointestinal Núcleo grácil Núcleo cuneiforme Lemnisco medial Bulbo raquídeo Células del ganglio de la raíz dorsal Médula espinal sacra bloquea las respuestas de las neuronas en el núcleo grácil a la estimulación visceral nociva, pero no a la estimulación cutánea inocua. El descubrimiento de este componente sensitivo visceral en el sistema de columnas dorsales-lemnisco medial ayudó a explicar por qué la sección quirúrgica de los axones que se encuentran en la porción medial de las columnas dorsales (procedimiento denominado mielotomía de la línea media) genera un alivio importante del dolor debilitante que aparece como consecuencia de los cánceres viscerales en el abdomen y la pelvis. Aunque (A) Vía para el dolor visceral en el sistema de columnas dorsales-lemnisco medial. Para simplificar, solo se muestran las vías que median el dolor visceral desde la pelvis y el abdomen inferior. Como comparación, también se muestra el componente mecanosensitivo de este sistema para la discriminación de los estímulos táctiles y el sistema anterolateral para la detección de estímulos cutáneos termoalgésicos (véanse también las Figuras 9.6 y 10.3A). (B) La evidencia empírica que apoya la existencia de la vía para el dolor visceral que se muestra en (A). Se observó que un aumento de la actividad neural con las técnicas de RM funcional en el tálamo de monos que fueron sometidos a una distensión nociva del colon y el recto, lo que indica el procesamiento del dolor visceral. Esta actividad fue abolida por la lesión de las columnas dorsales en T10, pero no por una cirugía “simulada”. (C) Arriba, un método de mielotomía puntiforme de la línea media para alivio del dolor visceral grave. Abajo, corte de la médula espinal torácica (T10) con tinción para mielina de un paciente que se sometió a una mielotomía de la línea media para el tratamiento del dolor por cáncer de colon que no se controló con analgésicos. Después de la cirugía, el paciente experimentó alivio del dolor durante los tres meses restantes de su vida. (B, de Willis y cols., 1999; C, de Hirshberg y cols., 1996; dibujo tomado de Nauta y cols., 1997.) DOLOR 217 RECUADRO 10C (continúa) el desarrollo inicial de este procedimiento quirúrgico precedió a la dilucidación de esta vía para el dolor visceral, estos descubrimientos nuevos renovaron el interés en la mielotomía de la línea media como intervención neuroquirúrgica paliativa para los pacientes oncológicos cuyo dolor por otra parte es inmanejable. En realidad, el conocimiento preciso de la vía sensitiva visceral en las columnas dorsales condujo a refinamientos nuevos que permiten un procedimiento quirúrgico mínimamente invasivo (“puntiforme”) que intenta interrumpir los axones de segundo orden de esta vía tan sólo en un segmento medular (en los casos típicos, un nivel torácico medio o inferior; Figura C). Al hacerlo, este procedimiento ofrece cierta esperanza a los pacientes que luchan por mantener una calidad de vida razonable en circunstancias extraordinariamente difíciles. Bibliografía Al-Chaer, E. D., N. B. Lawand, K. N. Westlund and W. D. Willis (1996) Visceral nociceptive input into the ventral posterolateral nucleus of the thalamus: a new function for the dorsal column pathway. J. Neurophysiol. 76: 2661-2674. Al-Chaer, E. D., N. B. Lawand, K. N. Westlund and W. D. Willis (1996) Pelvic visceral input into the nucleus gracilis is largely mediated by the postsynaptic dorsal column pathway. J. Neurophysiol. 76: 2675-2690. del cuerpo (Figura 10.4B). Los déficits se deben a la interrupción de las fibras que ascienden desde los niveles inferiores de la médula; por esta razón, incluyen todas las regiones del cuerpo (del lado contralateral u homolateral) que están inervadas por segmentos medulares que se encuentran por debajo del nivel de la lesión. Este patrón de pérdida sensitiva disociada (dolor y temperatura contralateral, tacto y presión homolateral) es una característica de las lesiones de la médula espinal y, juntamente con los signos dermatómicos locales (véase Recuadro 9A), se pueden utilizar para definir el nivel de la lesión. (Véase Recuadro 10C para una excepción importante a la disociación funcional de los sistemas de columna dorsal-lemnisco medio y anterolateral para el dolor visceral.) Vías paralelas del dolor Las fibras de segundo orden en el sistema anterolateral proyectan hacia algunas estructuras diferentes en el tronco encefálico y el encéfalo anterior, lo que deja claro que el dolor es procesado por una red diversa y distribuida de neuronas. Aunque no está claro el significado pleno de este patrón complejo de conexiones, estos destinos centrales probablemente medien aspectos diferentes de la respuesta sensitiva y conductual ante un estímulo doloroso. Un componente de este sistema media los aspectos discriminativos sensitivos del dolor: localización, intensidad y calidad del estímulo nocivo. Se cree que estos aspectos del dolor dependen de la información transmitida a través del núcleo ventral posterolateral (VPL) hasta las neuronas en la corteza somatosensitiva primaria y secundaria (Figuras 10.5 y 10.6A). (La vía para transmitir la información del rostro al núcleo ventral posteromedial o VPM se considera en la siguiente sección.) Aunque los axones del sistema anterolateral se superponen a los del sistema Becker, R., S. Gatscher, U. Sure and H. Bertalanffy (2001) The punctate midline myelotomy concept for visceral cancer pain control - case reort and review of the literature. Acta Neurochir. [Suppl] 79: 77-78. Hitchcock, E. R. (1970) Stereotactic cervical myelotomy. J. Neurol. Neurosurg. Psychiatry 33: 224-230. Kim, Y. S. and S. J. Kwon (2000) High thoracic midline dorsal column myelotomy for severe visceral pain due to advanced stomach cancer. Neurosurgery 46: 85-90. Nauta, H. and 8 others (2000) Punctate midline myelotomy for the relief of visceral cancer pain. J. Neurosurg. (Spine 2) 92: 125-130. Willis, W. D., E. D. Al- Chaer, M. J. Quast and K. N. Westlund (1999) A visceral pain pathway in the dorsal column of the spinal cord. Proc. Natl. Acad. Sci. USA 96: 7675-7679. de la columna dorsal en los núcleos ventrales posteriores, estos axones tienen conexiones con diferentes clases de neuronas de relevo, de modo que la información nociceptiva se mantiene segregada hasta el nivel de los circuitos corticales. Consistente con la mediación de los aspectos discriminativos del dolor, los registros electrofisiológicos provenientes de las neuronas nociceptivas en S1 muestran que estas neuronas tienen pequeños campos receptivos localizados –propiedades compatibles con las medidas conductuales de localización del dolor–. Otras partes del sistema transfieren información sobre los aspectos afectivos-emocionales del dolor: la sensación desagradable, el miedo y la ansiedad, y la activación autónoma que acompañan a la exposición a un estímulo nocivo (la reacción clásica de “lucha o huida”; véase Capítulo 21). Los puntos diana de estas proyecciones incluyen varias subdivisiones de la formación reticular, las capas profundas del colículo superior, la sustancia gris central, el hipotálamo y la amígdala. Además, un conjunto distinto de núcleos talámicos que se localizan mediales al núcleo ventral posterior, que aquí agrupamos como los núcleos talámicos de la línea media (véase Figura 10.5), se cree que desempeñan un papel importante en la transmisión de señales nociceptivas tanto a la corteza cingular anterior como a la ínsula (la región de la corteza que se localiza en la pared medial de la fisura lateral). Los registros electrofisiológicos en seres humanos, que muestran que las neuronas cingulares responden a los estímulos nocivos, apoyan el papel de la corteza cingular anterior en la percepción del dolor. Más aún, los pacientes que se han sometido a cingulotomías informan una atenuación de la sensación desagradable que acompaña al dolor. Las pruebas provenientes de estudios de imágenes funcionales en seres humanos apoyan el punto de vista de que diferentes regiones encefálicas median los aspectos sensiti- 218 CAPÍTULO 10 Sensitivo-discriminativa FIGURA 10.5 Dos aspectos distintos de la Afectivo-motivacional Corteza somatosensitiva (S1-S2) Corteza cingular anterior Corteza insular experiencia del dolor. El sistema anterolateral transmite información a diferentes partes del tronco del encéfalo y el encéfalo anterior que contribuyen a diferentes aspectos de la experiencia de dolor: Núcleo ventral posterior aquellos que son responsables de la discriminación sensitiva del dolor y las que son responsables de las repuestas afectivas y motivacionales al dolor. Amígdala Hipotálamo Sustancia gris periacueductal Núcleos talámicos de la línea media Colículo superior Formación reticular SISTEMA ANTEROLATERAL vos-discriminativos y afectivos-motivacionales del dolor. La presentación de un estímulo doloroso conduce a la activación tanto de la corteza somatosensitiva primaria como de la corteza cingular anterior; sin embargo, al utilizar la sugerencia hipnótica para aumentar o disminuir selectivamente la sensación desagradable de un estímulo doloroso, ha sido posible separar la respuesta neural a los cambios en la intensidad de un estímulo doloroso de los cambios en su sensación desagradable. Los cambios de intensidad se acompañan por cambios en la actividad de las neuronas en la corteza somatosensitiva, con pocos cambios en la actividad de la corteza cingular, mientras que los cambios en la sensación desagradable están altamente correlacionados con cambios en la actividad de las neuronas en la corteza cingular. A partir de esta descripción, resulta evidente que la experiencia plena del dolor involucra la acción cooperativa de una red extensa de regiones del encéfalo anterior cuyas propiedades solo estamos comenzando a comprender. En efecto, los análisis de imágenes encefálicas a menudo se refieren al amplio conjunto de áreas cuya actividad se asocia con la experiencia del dolor –incluida la corteza somatosensitiva, la corteza insular, la amígdala y la corteza cingular anterior– como matriz del dolor. En resumen, la naturaleza extendida de la representación del dolor no resulta sorprendente dado que el dolor es una experiencia multidimensional con efectos sensitivos, motores, emocionales/ afectivos y cognitivos. Una representación extendida también explica por qué las ablaciones de la corteza somatosensitiva no suelen aliviar el dolor crónico, aun cuando deterioran gravemente la percepción mecanosensitiva contralateral. Vías del dolor y la temperatura para el rostro La información sobre la estimulación nociva y térmica del rostro se origina en las neuronas de primer orden situadas en el ganglio trigeminal y en los ganglios asociados a los nervios craneales vii, ix y x (Figura 10.6B). Después de ingresar en la protuberancia, estas pequeñas fibras trigeminales mielínicas y amielínicas descienden hasta el bulbo raquídeo y forman el tracto trigeminal espinal (o el tracto espinal del nervio craneal v) y terminan en dos subdivisiones del complejo trigeminal espinal: la porción interpolar y la porción caudal. Los axones provenientes de las neuronas de segundo orden en estos dos núcleos trigeminales atraviesan la línea media y terminan en distintos puntos diana del tronco encefálico y el tálamo. Al igual que sus análogos en el asta dorsal de la médula espinal, estos puntos diana pueden ser agrupados en aquellos que median los aspectos discriminativos del dolor y los que median los aspectos afectivos/motivacionales. Se considera que los aspectos discriminativos del dolor facial están mediados por proyecciones hacia el núcleo ventral posteromedial (a través del tracto trigeminotalámico) y proyecciones desde el VPM hasta la corteza somatosensitiva primaria y secundaria. Los aspectos afectivos/motivacionales están mediados por conexiones hacia distintos puntos diana en la formación reticular y el mesencéfalo, y por los núcleos de la línea media del tálamo, que dan aferencias a las regiones cingular e insular de la corteza. Otras modalidades mediadas por el sistema anterolateral Aunque el sistema anterolateral desempeña un papel fundamental en la mediación de la nocicepción, también es responsable de transmitir otra información inocua diferente a los centros superiores. Por ejemplo, en ausencia del sistema de las columnas dorsales, el sistema anterolateral parece ser capaz de mediar lo DOLOR (A) (B) Cerebro Cerebro 219 Corteza somatosensitiva primaria Núcleo ventral posteromedial del tálamo Mesencéfalo Núcleo ventral posterolateral del tálamo Mesencéfalo Tracto trigeminotalámico Tracto espinotalámico Protuberancia media Protuberancia media Información termoalgésica desde el rostro Bulbo raquídeo medio Bulbo raquídeo medio Tracto espinal del nervio trigémino (axones aferentes) Bulbo raquídeo caudal Bulbo raquídeo caudal Sistema anterolateral Médula espinal cervical Información termoalgésica desde la parte superior del cuerpo (excluido el rostro) Médula espinal lumbar Información termoalgésica desde la parte inferior del cuerpo Núcleo espinal del complejo trigeminal FIGURA 10.6 Vías discriminativas del dolor. Comparación de las vías que median los aspectos discriminativos de dolor y temperatura para (A) el cuerpo y (B) el rostro. 220 CAPÍTULO 10 que habitualmente se denomina “tacto no discriminativo”, una forma de sensibilidad táctil que carece de la resolución espacial fina que solo puede ser provista por las columnas dorsales. Por lo tanto, después del daño del sistema columna dorsal-lemnisco medial, queda una forma cruda de sensibilidad táctil, en la cual están aumentados los umbrales de discriminación de dos puntos y está muy afectada la capacidad para identificar los objetos solo a través del tacto (estereognosia). Como ya mencionamos, el sistema anterolateral es responsable de mediar la sensación inocua de la temperatura. Se cree que la sensación de calor y frío corresponde a conjuntos separados de aferencias primarias: las fibras para el calor que responden con un aumento de las frecuencias de descargas a los aumentos de la temperatura, y las fibras para el frío que responden con un aumento de las descargas a las disminuciones de la temperatura. Ninguna de estas aferencias responde a la estimulación mecánica, y se distinguen de las aferencias que responden a temperaturas consideradas dolorosas (calor nocivo, por encima de 43 °C; o frío nocivo, por debajo de −17 °C). La identificación reciente de canales de TRP con sensibilidad a temperaturas en el rango inocuo –TRPV3 y TRPV4 que responden a temperaturas calientes y TRPM8 que responden a temperaturas frías– plantea la posibilidad de líneas marcadas para la transmisión de calor y de frío, comenzando a nivel de la transducción y continuando dentro de las vías centrales. Consistente con esta idea, la información aportada por las aferencias del calor y el frío inocuos es transmitida a los centros superiores por distintas clases de neuronas secundarias que residen en la lámina 1 de la médula espinal. En efecto, el punto de vista emergente sobre la lámina 1 es que consiste en algunas clases distintas de neuronas selectivas de modalidad que transmiten los tipos de información sensitiva nociva e inocua al sistema anterolateral. Estas incluyen clases individuales de neuronas que son sensibles a distintos estímulos: dolor agudo (primero), dolor ardiente (segundo), calor inocuo, frío inocuo, histamina (media la sensación de “prurito”), estimulación mecánica lenta (“tacto sensual”) y una clase de aferencias que inerva músculos y detecta el ácido láctico y otros metabolitos que son liberados durante la contracción muscular. Esta última podría contribuir al “ardor” o dolor que puede acompañar al ejercicio extenuante. ¿Es la lámina 1 simplemente una mezcla ecléctica de células con diferentes propiedades, o un tema unificador podría explicar esta diversidad? Se ha propuesto que el sistema de la lámina 1 funciona como la aferencia sensitiva de una red que es responsable de representar el estado fisiológico del cuerpo –modalidad que ha sido denominada interocepción, para distinguirla de la exterocepción (tacto y presión) y de la propiocepción–. Estas aferencias impulsan los mecanismos homeostáticos que mantienen un estado interno óptimo. Algunos de estos mecanismos son automáticos, y los cambios necesarios para mantener la homeostasis pueden estar mediados por una adaptación reflexiva del sistema nervioso autónomo (véase Capítulo 21). Por ejemplo, los cambios en la temperatura evocan reflejos autónomos (p. ej., sudoración o escalofríos) que contrarrestan un desequilibrio en la temperatura óptima del cuerpo. Las alteraciones homeostáticas a veces son demasiado grandes como para estar mediadas por reflejos autónomos aislados y requieren una adaptación conductual (p. ej., colocarse o sacarse un suéter) para restablecer el equilibrio. En esta concepción, las sensaciones asociadas con la activación del sistema de la lámina 1 –ya sean placenteras o nocivas– motivan la iniciación de conductas apropiadas para mantener la homeostasis fisiológica del cuerpo. Sensibilización Tras un estímulo doloroso asociado con daño tisular (p. ej., cortes, raspones y hematomas), los estímulos en el área de la lesión y en la región circundante, que por lo común se percibirían como poco dolorosos, se perciben como significativamente más dolorosos, fenómeno denominado hiperalgesia. Un buen ejemplo de hiperalgesia es el aumento de sensibilidad a la temperatura que se desarrolla después de una quemadura solar. Este efecto se debe a cambios en la sensibilidad neuronal que ocurren a nivel de los receptores periféricos y en sus puntos diana centrales. La sensibilización periférica es el resultado de la interacción de los nociceptores con la “sopa inflamatoria” de sustancias liberadas cuando se daña el tejido. Estas sustancias se originan en nociceptores activados o en células no neuronales que residen en el interior del área lesionada o migran hacia ella. Los nociceptores liberan péptidos y neurotransmisores como sustancia P, péptido relacionado con el gen de la calcitonina (CGRP) y ATP, todos los cuales contribuyen a la respuesta inflamatoria (vasodilatación, sudoración y liberación de histamina desde los mastocitos). La lista de células no neuronales que contribuyen a esta “sopa inflamatoria” incluye mastocitos, plaquetas, basófilos, macrófagos, neutrófilos, células endoteliales, queratinocitos y fibroblastos. Estas células son responsables de liberar protones extracelulares, ácido araquidónico y otros metabolitos lipídicos, bradicinina, histamina, serotonina, prostaglandinas, nucleótidos, factor de crecimiento nervioso (NGF) y muchas citocinas, (entre las cuales la principal es la interleucina-1β) y el factor de necrosis tumoral α (TNF-α). La mayoría de estas sustancias interactúan directamente con los receptores o los canales iónicos de las fibras nociceptivas, y aumentan su respuesta (Figura 10.7). Por ejemplo, la respuesta del receptor TRVP1 al calor puede ser potenciada por la interacción directa del canal con protones o metabolitos lipídicos. NGF y bradicinina también potencian la actividad de los receptores TRVP1, pero lo hacen de forma indirecta a través de las acciones de receptores separados en la superficie celular (TrkA y receptores de bradicinina, respectivamente) y sus vías de señalamiento intracelular asociadas. Se considera que las prostaglandinas contribuyen a la sensibilización periférica al unirse a receptores acoplados a las proteínas G que aumentan los niveles de AMP cíclico en el interior de los nociceptores. Las prostaglandinas también reducen la despolarización umbral necesaria para generar potenciales de acción a través de la fosforilación de una clase específica de canales del DOLOR 221 FIGURA 10.7 Respuesta inflamatoria al daño tisular. Las sustancias liberadas por los tejidos dañados aumentan la respuesta de las fibras nociceptivas. Además, la activación eléctrica de los nociceptores produce la liberación de péptidos y neurotransmisores Lesión tisular que contribuyen más a la respuesta inflamatoria. Plaquetas Macrófago ATP Bradicinina 5–HT Prostaglandina Histamina Mastocito o neutrófilo H+ IL-1β NGF TNF-α Sustancia P Vaso sanguíneo Sistema anterolateral CGRP, sustancia P Cuerpo celular del ganglio de la raíz dorsal Médula espinal sodio resistentes a la TTX que se expresan en los nociceptores. Las citocinas también pueden impactar en los receptores, pero su principal contribución a la hiperalgesia es el resultado de la potenciación de la respuesta inflamatoria a través de una mayor producción de prostaglandinas, NGF, bradicinina y protones extracelulares. El presunto propósito de la cascada de señalización química compleja que se origina en el daño local no es solo proteger el área lesionada (como resultado de las percepciones dolorosas producidas por estímulos comunes cercanos al sitio del daño), sino también promover la cicatrización y preservar contra la infección por medio de los efectos locales como el aumento del flujo sanguíneo y la migración de los leucocitos hasta el sitio. La identificación de los componentes de la sopa inflamatoria y sus mecanismos de acción es un área fértil de investigación en la búsqueda de analgésicos potenciales (compuestos que reducen la intensidad del dolor). Por ejemplo, los denominados AINE (agentes antiinflamatorios no esteroides), que incluyen aspirina e ibuprofeno, actúan por la inhibición de la ciclooxigenasa (COX), una enzima importante para la biosíntesis de prostaglandinas. La interferencia con la señalización de neurotrofina o de citocinas se ha convertido en una estrategia importante para controlar la enfermedad inflamatoria y el dolor resultante. El bloqueo de la acción de TNF-α con un anticuerpo neutralizan- te ha sido significativamente eficaz para el tratamiento de las enfermedades autoinmunitarias, incluida la artritis reumatoidea, lo que conduce a una reducción espectacular tanto de la destrucción tisular como de la hiperalgesia asociada. Asimismo, se ha demostrado que los anticuerpos anti-NGF previenen y revierten los signos conductuales de hiperalgesia en modelos animales. La sensibilización central se refiere a un aumento dependiente de la actividad y de inicio inmediato en la excitabilidad de las neuronas del asta dorsal de la médula espinal que sigue a los niveles elevados de actividad en las aferencias nociceptivas. En consecuencia, los niveles de actividad en las aferencias nociceptivas que estaban por debajo del umbral antes del acontecimiento sensibilizante se vuelven suficientes como para generar potenciales de acción en las neuronas del asta dorsal, y contribuyen con un aumento de la sensibilidad al dolor. Aunque la sensibilización central se desencadena en las neuronas del asta dorsal por la actividad en los nociceptores, los efectos se generalizan a otras aferencias que se originan en los mecanorreceptores de umbral bajo. Así, los estímulos que en condiciones normales serían inocuos (como el cepillado de la superficie de la piel) activan neuronas de segundo orden en el asta dorsal que reciben aferencias nociceptivas, y dan origen a una sensación de dolor. La inducción de dolor por un estímulo normalmente inocuo se denomina alodinia. En los casos típicos, este fenómeno se desarrolla inmediatamente después del acontecimiento doloroso y puede superar en duración al estímulo por varias horas. Al igual que su análogo periférico, algunos mecanismos diferentes contribuyen a la sensibilización central. Una forma de sensibilización central, denominada “elevación”, comprende un aumento progresivo en la frecuencia de descarga de las neuronas del asta dorsal en respuesta a la activación repetida de baja frecuencia de las aferencias nociceptivas. Una correlación conductual del fenómeno de elevación ha sido estudiada al examinar la intensidad percibida de dolor en respuesta a múltiples presentaciones de un estímulo nocivo. Aunque la intensidad de la estimulación es constante, la intensidad percibida aumenta con cada presentación del estímulo. La elevación solo dura el período de estimulación y se origina en la suma de los potenciales sinápticos lentos evocados en las neuronas del asta dorsal por las aferencias nociceptivas. La despolarización sostenida de las neuronas del asta dorsal es en parte resultado de la activación de canales del calcio dependientes del voltaje de tipo L y en parte de la eliminación del bloqueo de los receptores de NMDA por el Mg. La eliminación del bloqueo de magnesio aumenta la 222 CAPÍTULO 10 RECUADRO 10D Miembros fantasma y dolor fantasma Luego de la amputación de una extremidad, casi todos los pacientes experimentan una ilusión de que el miembro faltante sigue presente. Si bien esta ilusión habitualmente disminuye con el tiempo, persiste en cierto grado durante toda la vida del amputado y a menudo puede reactivarse por la lesión del muñón u otras alteraciones. Estas sensaciones fantasma no están limitadas a las extremidades amputadas; se comunican mamas fantasma luego de la mastectomía, genitales fantasma después de la castración y sensación fantasma de toda la mitad inferior del cuerpo después de la sección de la médula espinal. Esta sensación también es común después del bloqueo nervioso local para cirugía. Durante la recuperación de la anestesia del plexo braquial, por ejemplo, es frecuente que el paciente experimente un brazo fantasma, percibido como entero e intacto, pero desplazado del brazo real. Cuando se visualiza el brazo real, el miembro fantasma parece “saltar” al brazo y puede emerger y volver a entrar de manera intermitente mientras la anestesia desaparece. Estos fantasmas sensitivos demuestran que la maquinaria central para el procesamiento de la información somatosensitiva no está ociosa en ausencia de estímulos periféricos; al parecer, los mapas sensitivos centrales y los sistemas de procesamiento siguen operando en forma independiente de la periferia para organizar estas sensaciones extrañas. Los fantasmas podrían ser simplemente una curiosidad –o un indicio provocativo sobre el procesamiento somatosensitivo de orden superior– si no fuera por el hecho de que una cantidad sustancial de amputados también sufren dolor fantasma. Este problema frecuente suele describirse como una sensación de hormigueo o ardor en la parte faltante. Sin embargo, a veces, la sensación se convierte en un dolor mucho más grave que los pacientes experimentan cada vez como más debilitante. De hecho, el dolor fantasma es una de las causas más frecuentes de síndromes de dolor crónico, trastorno de tratamiento extraordinariamente difícil. Debido a la naturaleza difusa del procesamiento central del dolor, la ablación del tracto espinotalámico, porciones del tálamo o incluso de la corteza somatosensitiva primaria por lo general no alivia el malestar que perciben estos pacientes. Es interesante destacar que se desarrolla una reorganización funcional considerable de los mapas somatotópicos en la corteza somatosensitiva primaria de los amputados. Esta reorganización comienza inmediatamente después de la amputación y tiende a evolucionar durante varios años. Uno de los efectos de este proceso es que las neuronas que perdieron sus aferencias originales (esto es, de la extremidad amputada) responden a la estimulación táctil de otras partes del cuerpo. Una consecuencia sorprendente es que la estimulación del rostro, por ejemplo, puede experimentarse como si se hubiera tocado el miembro faltante. Otros indicios de que el fenómeno del miembro fantasma es resultado de una representación central es la experiencia de los niños nacidos sin extremidades. Estos individuos tienen ricas sensaciones fantasma, a pesar de que nunca desarrollaron una extremidad. Esta observación sugiere que hay una representación completa del cuerpo independientemente de los elementos periféricos que se mapeen. Sobre la base de estos resultados, Ronald Melzack propuso que la pérdida de una extremidad genera un desequilibrio interno entre la representación del cuerpo en el encéfalo y el patrón de aferencias táctiles periféricas que alcanza la neocorteza. La consecuencia sería una sensación ilusoria de que la parte del cuerpo ausente sigue presente y es funcional. Con el tiempo, el encéfalo puede adaptarse a esta pérdida y alterar su representación somática intrínseca para adaptarse mejor a la nueva configuración del cuerpo. Este cambio podría explicar por qué la sensación fantasma aparece casi inmediatamente después de la pérdida de la extremidad, pero su intensidad suele disminuir con el tiempo. Bibliografía Melzack, R. (1989) Phantom limbs, the self, and the brain. The D.O. Hebb Memorial Lecture. Can. Psychol. 30: 1-14. Melzack, R. (1990) Phantom limbs and the concept of a neuromatrix. Trends Neurosci. 13: 88-92. Nashold, B. S., Jr. (1991) Paraplegia and pain. In Deafferentation Pain Syndromes: Pathophysiology and Treatment, B. S. Nashold, Jr. and J. OvelmenLevitt (eds.). New York: Raven Press, pp. 301-319. Ramachandran, V. S. and S. Blakeslee (1998) Phantoms in the Brain. New York: William Morrow & Co. Solonen, K. A. (1962) The phantom phenomenon in amputated Finnish war veterans. Acta. Orthop. Scand. Suppl. 54: 1-37. Dibujos de brazos y piernas fantasma sobre la base de las comunicaciones de los pacientes. El fantasma está indicado por una línea rayada, donde las regiones coloreadas muestran las partes que se experimentan más vívidamente. Obsérvese que algunos fantasmas se encuentran telescopados en el muñón. (De Solomen, 1962.) DOLOR sensibilidad de la neurona del asta dorsal al glutamato, el neurotransmisor en las aferencias nociceptivas. Se cree que otras formas de sensibilización central que duran más tiempo del período de estimulación sensitiva (p. ej., alodinia) involucran un refuerzo de los potenciales postsinápticos similar a la potenciación a largo plazo muy parecido a lo descrito para el hipocampo (véase Capítulo 8). Estos efectos dependen de las elevaciones del Ca2+ en las neuronas de la médula espinal postsinápticas a los nociceptores mediadas por el receptor de NMDA. También se considera que la reducción del nivel de inhibición GABAérgica o glicinérgica contribuye a los síndromes dolorosos persistentes al aumentar la excitabilidad de las neuronas de proyección del asta dorsal. La microglia y los astrocitos también contribuyen al proceso de sensibilización central, sobre todo cuando existe una lesión del nervio. Por ejemplo, las citocinas liberadas de la microglia promueven la transcripción difusa de la enzima COX-2 y la producción consiguiente de prostaglandinas en las neuronas del asta dorsal. Como se describió para las aferencias nociceptivas, el aumento de las concentraciones de prostaglandinas en las neuronas del SNC aumenta la excitabilidad neuronal. Por lo tanto, los efectos analgésicos de los fármacos que inhiben la transcripción de COX-2 se deben a las acciones tanto en la periferia como en el interior del asta dorsal. A medida que el tejido lesionado cicatriza, la sensibilización inducida por mecanismos periféricos y centrales habitualmente disminuye y el umbral para el dolor retorna a los niveles previos a la lesión. Sin embargo, cuando las propias fibras aferentes o las vías centrales se dañan –una complicación frecuente en las condiciones patológicas que incluyen diabetes, zóster, sida, esclerosis múltiple y accidente cerebrovascular– estos procesos pueden persistir. El trastorno resultante se denomina dolor neuropático: una experiencia crónica e intensamente dolorosa difícil de tratar con medicaciones analgésicas convencionales. (Véase Recuadro 10D para una descripción detallada del dolor neuropático asociado con la amputación de una extremidad.) El dolor neuropático puede surgir espontáneamente (es decir, sin ningún estímulo) o puede ser producido por estímulos leves que son comunes a la experiencia cotidiana, como el tacto suave y la presión de la ropa, o las temperaturas cálidas o frías. Los pacientes a menudo describen su experiencia como una sensación ardiente constante interrumpida por episodios de dolor lacerante, pinchazos o descargas eléctricas. Dado que la discapacidad y el estrés psicológico asociados con el dolor neuropático crónico pueden ser intensos, gran parte de la investigación actual está dedicada a conocer mejor los mecanismos de la sensibilización periférica y central con la esperanza de desarrollar terapias más efectivas para este síndrome discapacitante. Control descendente de la percepción del dolor Respecto de la interpretación del dolor, los investigadores han observado desde hace mucho que existe una diferencia entre la 223 realidad objetiva de un estímulo doloroso y la respuesta subjetiva a él. Algunos estudios modernos de esta discrepancia aportaron cierta comprensión acerca del modo en que ciertas circunstancias afectan la percepción del dolor y, acerca de la anatomía y la farmacología del sistema del dolor. Durante la Segunda Guerra Mundial, Henry Beecher y cols., en la Escuela Médica de Harvard, realizaron una observación fundamental. En el primer estudio sistemático de este tipo, observaron que los soldados que sufrían lesiones graves a menudo experimentaban poco o ningún dolor. En efecto, muchos de los heridos expresaban su asombro por esta extraña disociación. Beecher, un anestesista, arribó a la conclusión de que la percepción del dolor depende de su contexto. Por ejemplo, el dolor de un soldado herido en el campo de batalla presumiblemente estaría mitigado por los beneficios imaginarios de alejarse del peligro, mientras que una lesión similar en un contexto doméstico presentaría un entorno de circunstancias muy diferentes que podrían exacerbar el dolor (pérdida de trabajo, problemas económicos, etc.). Estas observaciones, junto con el efecto placebo bien conocido (que se explica a continuación), dejaron claro que la percepción del dolor está sometida a una modulación central; en efecto, todas las sensaciones están sometidas por lo menos a cierto grado de este tipo de modificación. Esta afirmación no debe tomarse como un reconocimiento vago sobre la importancia de las influencias psicológicas o “arriba-abajo” sobre la experiencia sensitiva. Por el contrario, entre los científicos en neurociencia y los neurólogos se desarrolló un conocimiento gradual de estos efectos “psicológicos” como reales e importantes, como cualquier otro fenómeno neural. Esta apreciación ha proporcionado un punto de vista mucho más racional de los problemas psicosomáticos en general y del dolor en particular. El efecto placebo La palabra placebo significa “complaceré”, y el efecto placebo se define como una respuesta fisiológica después de la administración de un “remedio” farmacológicamente inerte. El efecto placebo tiene una larga historia de uso (y abuso) en medicina, pero su realidad es indiscutible. En un estudio clásico, los estudiantes de medicina recibieron una de dos píldoras diferentes, se les indicó que una era sedante y la otra, estimulante. De hecho, ambas píldoras contenían solo ingredientes inertes. De los estudiantes que recibieron el “sedante”, más de dos tercios comunicaron que habían sentido somnolencia, y los que tomaron dos píldoras se sintieron más somnolientos que los que tomaron una. Por el contrario, una gran fracción de los estudiantes que tomó el “estimulante” comunicaron que se sentían menos cansados. Más aún, ¡alrededor de un tercio de todo el grupo comunicó efectos colaterales que variaron desde cefaleas y mareos hasta parestesias en las extremidades y una marcha tambaleante! Solo 3 de los 56 estudiantes del grupo informaron que las píldoras que habían recibido no tuvieron ningún efecto. En otro estudio de este tipo general, el 75% de los estudiantes que sufrieron dolor posoperatorio en las heridas relataron un 224 CAPÍTULO 10 alivio satisfactorio después de una inyección de solución salina estéril. Los investigadores que llevaron a cabo este estudio observaron que aquellos que respondieron eran indistinguibles de los que no lo hicieron, tanto en la gravedad aparente de su dolor como en sus características psicológicas. Más notablemente, el efecto placebo en los pacientes posoperatorios puede ser bloqueado con naloxona, un antagonista competitivo de los receptores de opiáceos, lo que indica una base farmacológica sustancial para el alivio del dolor experimentado (véase la siguiente sección en el texto). Además, algunos estudios de imágenes muestran que la administración de un placebo con la expectativa de que representa un agente analgésico se asocia con la activación de receptores endógenos de opiáceos en regiones encefálicas corticales y subcorticales que forman parte de la matriz del dolor, que incluyen las regiones cingular anterior e insular de la corteza y la amígdala. Una interpretación errónea frecuente sobre el efecto placebo es el punto de vista de que los pacientes que responden a un reactivo sin importancia terapéutica no sufren dolor real, sino solo lo “imaginan”. Ciertamente esto no es así. Entre otras cosas, el efecto placebo probablemente explica la eficacia de la anestesia con acupuntura y la analgesia que a veces puede lograrse mediante la hipnosis. En China, la cirugía a menudo se lleva a cabo bajo el efecto de una aguja (que con frecuencia transmite una corriente eléctrica pequeña) insertada en localizaciones dictadas por antiguos atlas de acupuntura. Antes de la llegada de las técnicas anestésicas modernas, las operaciones como las tiroidectomías para el bocio se realizaban comúnmente sin un malestar extraordinario, sobre todo en las poblaciones donde el estoicismo era la norma cultural. Los mecanismos de alivio del dolor en el campo de batalla, en la anestesia por acupuntura y con la hipnosis supuestamente están relacionados. Aunque solo están comenzando a comprenderse los mecanismos por medio de los cuales el encéfalo afecta la percepción del dolor, el efecto no es mágico ni un signo de un intelecto sugestionable. En resumen, el efecto placebo es muy real. Base fisiológica de la modulación del dolor El conocimiento de la modulación central de la percepción del dolor (sobre la cual supuestamente se basa el efecto placebo) avanzó mucho con el hallazgo de que la estimulación eléctrica o farmacológica de ciertas regiones del mesencéfalo produce alivio del dolor. Este efecto analgésico surge de la activación de las vías descendentes que modulan el dolor que proyectan hasta el asta dorsal de la médula espinal (así como el núcleo trigeminal espinal) y regulan la transmisión de información a los centros superiores. Una de las principales regiones encefálicas que producen este efecto se localiza en la sustancia gris periacueductal del mesencéfalo. La estimulación eléctrica en esta zona en los animales de experimentación no solo produce analgesia por cri- terios conductuales, sino que se demostró que inhibe la actividad de las neuronas de proyección nociceptivas en el asta dorsal de la médula espinal. Otros estudios de las vías descendentes hasta la médula espinal que regulan la transmisión de la información nociceptiva han mostrado que se originan en algunos sitios del tronco encefálico, incluido el núcleo parabraquial, el rafe dorsal y el locus cerúleo, y la formación reticular medular (Figura 10.8A). Los efectos analgésicos de la estimulación de la sustancia gris periacueductal están mediados a través de estos sitios en el tronco encefálico. Estos centros emplean muchos neurotransmisores diferentes (p. ej., noradrenalina, serotonina, dopamina, histamina, acetilcolina) y pueden ejercer tanto efectos facilitadores como inhibidores sobre la actividad de las neuronas en el asta dorsal. La complejidad de estas interacciones se vuelve incluso mayor por el hecho de que las proyecciones descendentes pueden ejercer sus efectos sobre distintos sitios en el asta dorsal, incluidas las terminaciones sinápticas de las aferencias nociceptivas, interneuronas excitadoras e inhibidoras, y las terminaciones sinápticas de otras vías descendentes, así como por el contacto con las neuronas de proyección propiamente dichas. Aunque en principio estas proyecciones descendentes se consideraban como un mecanismo que servía primariamente para inhibir la transmisión de señales nociceptivas, ahora es evidente que estas proyecciones brindan un equilibrio de influencias facilitadoras e inhibidoras que determina, por último, la eficacia de la transmisión nociceptiva. Además de las proyecciones descendentes, las interacciones locales entre las aferencias de los mecanorreceptores y los circuitos neurales en el interior del asta dorsal pueden modular la transmisión de información nociceptiva hasta los centros superiores (Figura 10.8B). Se cree que estas interacciones explican la capacidad para reducir la sensación del dolor agudo al activar los mecanorreceptores de umbral bajo –por ejemplo, si usted se quiebra la tibia o se golpea un dedo del pie, una reacción natural (y efectiva) es frotar enérgicamente el sitio de la lesión durante uno o dos minutos–. Estas observaciones, apoyadas por experimentos en animales, condujeron a Ronald Melzack y Patrick Wall a proponer que el flujo de información nociceptiva a través de la médula espinal es modulado por la activación simultánea de las grandes fibras mielínicas asociadas con los mecanorreceptores de bajo umbral. Aun cuando una más profunda investigación condujo a la modificación de algunas de las propuestas originales en la teoría de la puerta del dolor de Melzack y Wall, esta propuesta estimuló muchas investigaciones sobre la modulación del dolor y ha destacado la importancia de las interacciones sinápticas en el interior del asta dorsal para modular la percepción de la intensidad del dolor. El adelanto más interesante en este esfuerzo por comprender los mecanismos centrales de la regulación del dolor ha sido el descubrimiento de los opioides endógenos. Durante siglos ha sido evidente que los derivados del opio, como la morfina, son analgésicos potentes (en realidad, aún son fundamentales en la terapia analgésica). En la era moderna, algunos estudios en ani- DOLOR (A) 225 (B) Corteza somatosensitiva Amígdala Fibra Aβ (mecanorreceptor) Neurona de circuito local inhibidora Sistema anterolateral Sustancia gris periacueductal Núcleo parabraquial Hacia las columnas dorsales Hipotálamo Formación reticular del bulbo raquídeo Locus cerúleo Fibra C (nociceptor) Neurona de proyección del asta dorsal Núcleos del rafe Hacia el sistema anterolateral Asta dorsal de la médula espinal (C) FIGURA 10.8 Sistemas descendentes que modulan la transmisión de las señales dolorosas ascendentes. (A) Estos sistemas modula- Aferencias descendentes, p. ej., núcleos del rafe dores se originan en la corteza somatosensitiva, el hipotálamo, la sustancia gris periacueductal del mesencéfalo, los núcleos del rafe y otros núcleos del Terminal axónico de neurona de circuito local que contiene encefalina bulbo raquídeo rostroventral. En cada uno de estos sitios, así como en el asta dorsal, se desarrollan efectos moduladores complejos. (B) Teoría de la Neurona de proyección del asta dorsal puerta del dolor. La activación de mecanorreceptores modula la transmisión de la información nociceptiva hacia los centros superiores. (C) Papel de las neuronas de circuitos locales que contienen encefalina en el control descendente de la transmisión de señales nociceptivas. males han mostrado que distintas regiones encefálicas son susceptibles a la acción de los agentes opioides, sobre todo –y significativamente– la sustancia gris periacueductal y otras fuentes de proyecciones descendentes. Además, existen neuronas sensibles a los opioides en el asta dorsal de la médula espinal. En otras palabras, las áreas que producen analgesia cuando son estimuladas también responden a los opioides administrados por vía exógena. Parece probable entonces que los agentes opioides actúan en la mayoría o en todos los sitios que se muestran en la Figura 10.8 para producir sus espectaculares efectos de alivio del dolor. La acción analgésica de los opioides condujo a los investigadores a sospechar la existencia de receptores encefálicos y medulares específicos para estos fármacos mucho antes de las décadas de 1960 y 1970, cuando se identificaron realmente estos receptores. Como es improbable que esos receptores existan con el propósito de responder a la administración exógena de opio y sus derivados, aumentó la convicción de que debía haber compuestos endógenos similares a los opioides para explicar la evolución de los receptores de opioides (véase Capítulo 6). Se han aislado y estudiado intensamente varias categorías Fibra C (nociceptor) de opioides endógenos del encéfalo. Estos agentes se encuentran en las mismas regiones involucradas en la modulación de las aferencias nociceptivas, aunque cada una de las familias de péptidos opioides endógenos tiene una distribución algo diferente. Los tres grupos principales –encefalinas, endorfinas y dinorfinas (véase Cuadro 6.2)– están presentes en la sustancia gris periacueductal. Las encefalinas y las dinorfinas también se hallaron en el bulbo raquídeo ventral y en regiones de la médula espinal involucradas en la regulación del dolor. Uno de los ejemplos más convincentes del mecanismo por el cual los opioides endógenos modulan la transmisión de la información nociceptiva se desarrolla en la primera sinapsis en la vía para el dolor entre las aferencias nociceptivas y las neuronas de proyección en el asta dorsal de la médula espinal (véase Figura 10.8B). Una clase de neuronas de circuito local que contienen encefalina en el asta dorsal hacen sinapsis con las terminaciones axónicas de las aferencias nociceptivas, los que a su vez hacen sinapsis con las neuronas de proyección del asta dorsal. La liberación de encefalina en las terminaciones nociceptivas inhibe su liberación de neurotransmisor en la neurona de proyección, y re- 226 CAPÍTULO 10 ducen así el nivel de actividad que pasa a los centros superiores. Las propias neuronas de circuito local que contienen encefalina son los puntos diana de las proyecciones descendentes, lo que proporciona así un mecanismo poderoso por el cual los centros pueden disminuir la actividad transmitida por las aferencias nociceptivas. De forma similar, los efectos analgésicos de la marihuana (Cannabis) condujeron al descubrimiento de los endocannabinoides. Se sabe que los cannabinoides administrados por vía exógena suprimen las neuronas nociceptivas en el asta dorsal de la médula espinal sin alterar la actividad de las neuronas no nociceptivas. Actualmente, sabemos que los cannabinoides endógenos del SNC actúan como neurotransmisores; son liberados desde las células despolarizadas y viajan hasta las terminaciones presinápticas, donde activan a los receptores de cannabinoides (CB1) a través de un mecanismo de señalamiento retrógrado. Se cree que las acciones de los endocannabinoides disminuyen la liberación de neurotransmisores como GABA y glutamato, y modulan así la excitabilidad neuronal. La evidencia de un efecto directo de los endocannabinoides sobre la transmisión de señales nociceptivas proviene de estudios que muestran que los efectos analgésicos inducidos por la estimulación eléctrica de la sustancia gris periacueductal pueden ser bloqueados si se administran antagonistas de CB1. Además, al parecer la estimulación a los estímulos nocivos aumenta el nivel de los endocannabinoides en la sustancia gris periacueductal, hallazgo que apoya un papel importante para estas moléculas en el control descendente de la transmisión del dolor. La historia de los compuestos antinociceptivos endógenos es impresionante si se analizan las conexiones entre la fisiología, la farmacología y la investigación clínica para arrojar un conocimiento más rico de la modulación intrínseca del dolor. Finalmente, esta información ha comenzado a explicar la variabilidad subjetiva de los estímulos dolorosos y la notable dependencia de la percepción del dolor sobre el contexto de la experiencia. Muchos laboratorios están explorando los mecanismos precisos por los cuales el dolor es modulado, motivados por los beneficios clínicos (y económicos) enormes que surgirían del conocimiento aún más profundo del sistema del dolor y sus características moleculares. Resumen El dolor, desde una perspectiva tanto estructural como funcional, es una modalidad sensitiva extraordinariamente compleja. Dado que el dolor es un medio importante para advertir a un animal acerca de las circunstancias peligrosas, los mecanismos y las vías que regulan la nocicepción son amplios y redundantes. Un conjunto distinto de aferencias para el dolor con receptores de membrana conocidos como nociceptores traduce la información nociva y transmite esta información a las neuronas del asta dorsal de la médula espinal. La vía central principal responsable de transmitir los aspectos discriminativos del dolor (localización, intensidad y calidad) difiere de la vía mecanosensitiva primariamente en que los axones centrales de las células de los ganglios de las raíces dorsales hacen sinapsis en neuronas de segundo orden en el asta dorsal; los axones de las neuronas de segundo orden cruzan luego la línea media en la médula espinal y ascienden hasta los núcleos talámicos que transmiten información hasta la corteza somatosensitiva de la circunvolución poscentral. Otras vías que comprenden algunos centros en el tronco del encéfalo, el tálamo y la corteza median las respuestas afectivas y motivacionales a los estímulos dolorosos. Las vías descendentes interactúan con los circuitos locales en la médula espinal para regular la transmisión de señales nociceptivas hasta los centros superiores. Los investigadores han logrado un progreso enorme en el conocimiento del dolor en los últimos 25 años, y mucho más parece probable, dada la importancia del problema. Ningún paciente está más angustiado ni es más difícil de tratar que aquel con dolor crónico, un subproducto devastador de la función protectora de esta modalidad sensitiva vital. Lecturas adicionales Revisiones Basbaum, A. I., D. M. Bautista, G. Scherrer and D. Julius (2009) Cellular and molecular mechanisms of pain. Cell 139: 267-28’4. Caterina, M. J. and D. Julius (1999) Sense and specificity: A molecular identity for nociceptors. Curr. Opin. Neurobiol. 9: 525-530. Cregg, R., A. Momin, F. Rugiero, J. N. Wood and J. Zhao (2010) Pain channelopathies. J. Physiol. 588: 1897-1904. Di Marzo, V., P. M. Blumberg and A. Szallasi (2002) Endovanilloid signaling in pain. Curr. Opin. Neurobiol. 12: 372-3710. Dubner, R. and M. S. Gold (1999) The neurobiology of pain. Proc. Natl. Acad. Sci. USA 96: 7627-7630. Fields, H. L. and A. I. Basbaum (1978) Brainstem control of spinal pain transmission neurons. Annu. Rev. Physiol. 40: 217-248. Gold, M. S. and G. F. Gebhart (2010) Nociceptor sensitization in pain pathogenesis. Nature Med. 16: 1248-1257. Guindon, J. and A. G. Hohmann (2009) The endocannabinoid system and pain. CNS Neurol. Disord. Drug Targets 8: 403-421. Hunt, S. P. and P. W. Mantyh (2001) The molecular dynamics of pain control. Nature Rev. Neurosci. 2: 83-91. Ji, R. R„ T. Kohno, K. A. Moore and C. J. Woolf (2003) Central sensitization and LTP: Do pain and memory share similar mechanisms? Trends Neurosci. 26: 696-705. Julius, D. and A. I. Basbaum (2001) Molecular mechanisms of nociception. Nature 413: 203-209. Millan, M. J. (2002) Descending control of pain. Prog. Neurobiol. 66: 355-474. Neugebauer, V., V. Galhardo, S. Maione and S. C. Mackey (2009) Forebrain pain mechanisms. Brain Res. Rev. 60: 226-242. Patapoutian, A., A. M. Peier, G. M. Story and V. Viswanath (2003) ThermoTRP channels and beyond: Mechanisms of temperature sensation. Nature Rev. Neurosci. 4: 529-539. Rainville, P. (2002) Brain mechanisms of pain affect and pain modulation. Curr. Opin. Neurobiol. 12: 195-204. Scholz, J. and C. J. Woolf (2002) Can we conquer pain? Nature Rev. Neurosci. 5 (Suppl): 1062-1067. Treede, R. D., D. R. Kenshalo, R. H. Gracely and A. K. Jones (1999) The cortical representation of pain. Pain 79: 105-111. Zubieta, J.-K. and S. Christian (2009) Neurobiological mechanisms of placebo responses. The Year in Cognitive Neuroscience 2009, Ann. NY Acad. Sci. 1156: 198-210. DOLOR Artículos originales importantes Basbaum, A. I. and H. L. Fields (1979) The origin of descending pathways in the dorsolateral funiculus of the spinal cord of the cat and rat: Further studies on the anatomy of pain modulation. J. Comp. Neurol. 187: 513-522. Beecher, H. K. (1946) Pain in men wounded in battle. Ann. Surg. 123: 96. Blackwell, B., S. S. Bloomfield and C. R. Buncher (1972) Demonstration to medical students of placebo response and non-drug factors. Lancet 1: 1279-1282. Caterina, M. J. and 8 others (2000) Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 288: 306-313. Craig, A. D., E. M. Reiman, A. Evans and M. C. Bushnell (1996) Functional imaging of an illusion of pain. Nature 384: 258-260. 227 Levine, J. D., H. L. Fields and A. I. Basbaum (1993) Peptides and the primary afferent nociceptor. J. Neurosci. 13: 2273-2286. Libros Fields, H. L. (1987) Pain. New York: McGraw-Hill. Fields, H. L. (ed.) (1990) Pain Syndromes in Neurology. London: Butterworths. Kolb, L. C. (1954) The Painful Phantom. Springfield, IL: Charles C. Thomas. Skrabanek, P. and J. McCormick (1990) Follies and Fallacies in Medicine. New York: Prometheus Books. Wall, P. D. and R. Melzack (1989) Textbook of Pain. New York: Churchill Livingstone. 229 CAPÍTULO 2 11 El ojo Aspectos generales EL SISTEMA VISUAL HUMANO ES EXTRAORDINARIO debido a la cantidad y calidad de información que aporta acerca del mundo. Una mirada rápida es suficiente para describir la localización, el tamaño, la forma y la textura de los objetos y, si están en movimiento, su dirección y su velocidad. Igualmente notable es la posibilidad de discernir la información visual entre una amplia gama de intensidades de los estímulos, desde la luz débil de las estrellas por la noche hasta la luz solar brillante. En los dos capítulos siguientes, se describen los mecanismos moleculares, celulares y de orden superior que nos permiten ver. Las etapas iniciales del proceso de la visión comprenden la transmisión y la refracción de la luz por la óptica del ojo, la transducción de la energía luminosa en señales eléctricas por los fotorreceptores y la codificación de estas señales por medio de las interacciones sinápticas en el interior de los circuitos neurales de la retina. Anatomía del ojo El ojo es una esfera llena de líquido encerrada por tres capas de tejido (Figura 11.1). Solo la capa más interna, la retina, contiene neuronas que son sensibles a la luz y capaces de transmitir señales visuales hasta los puntos diana centrales. La capa de tejido inmediatamente adyacente presenta tres estructuras distintas pero continuas denominadas en conjunto tracto uveal. El componente mayor del tracto uveal es la coroides, compuesta por un lecho capilar rico (importante para la nutrición de los fotorreceptores de la retina) y por una elevada concentración de un pigmento que absorbe la luz, la melanina. El cuerpo ciliar se extiende desde la coroides hasta cerca de la parte frontal del ojo y es un anillo de tejido que rodea el cristalino y presenta un componente muscular importante para ajustar el poder de refracción del cristalino, y un componente vascular (los denominados procesos ciliares) que produce el líquido que llena la parte anterior del ojo. El componente más anterior del tracto uveal es el iris, la porción coloreada del ojo que puede observarse a través de la córnea. Contiene dos conjuntos de músculos con acciones opuestas, que permiten ajustar el tamaño de la pupila (el orificio de su centro) bajo el control neural. La esclerótica forma la capa tisular más externa del ojo y está compuesta por un tejido fibroso blanco resistente. Sin embargo, en la parte anterior del ojo esta capa externa opaca se transforma en la córnea, un tejido transparente especializado que permite que los rayos de luz entren en el ojo. 230 CAPÍTULO 11 Más allá de la córnea, los rayos de luz atraviesan dos ambientes líquidos distintos antes de impactar en la retina. En la cámara anterior, inmediatamente por detrás de la córnea y por delante del cristalino, se ubica el humor acuoso, un líquido acuoso y claro que aporta nutrientes a ambas estructuras. El humor acuoso es producido por los procesos ciliares en la cámara posterior (la región entre el cristalino y el iris) y fluye en la cámara anterior a través de la pupila. La cantidad de líquido producido por los procesos ciliares es considerable: se estima que todo el volumen de líquido de la cámara anterior se reemplaza 12 veces al día. Por lo tanto, la velocidad de producción del humor acuoso debe equilibrarse con una velocidad de drenaje comparable desde la cámara anterior para asegurar una presión intraocular constante. Una red especializada de células que se ubica en la unión del iris y la córnea es responsable del drenaje acuoso. La falta de un drenaje suficiente conduce a un trastorno conocido como glaucoma, en el que los niveles elevados de presión intraocular pueden reducir la irrigación del ojo y, finalmente, dañar las neuronas retinianas. El espacio entre el dorso del cristalino y la superficie de la retina está lleno de una sustancia gelatinosa y espesa denominada humor vítreo, que representa alrededor del 80% del volumen del ojo. Además de mantener la forma del ojo, el humor vítreo contiene células fagocíticas que eliminan sangre y otros detritos que, de otra forma, podrían interferir con la transmisión de la luz. Sin embargo, la capacidad de limpieza del humor vítreo es limitada, como lo atestigua una gran cantidad de individuos de edad media y ancianos con “flotadores” vítreos. Estos son grupos de desecho demasiado grandes para ser fagocitados que permanecen y arrojan sombras molestas sobre la retina; en los casos típicos, surge cuando la membrana vítrea que envejece se separa del globo ocular, muy alargado en los individuos miopes (Recuadro 11A). Formación de imágenes en la retina La visión normal requiere que los medios ópticos del ojo sean transparentes, y tanto la córnea como el cristalino son ejemplos notables de especializaciones tisulares que logran un nivel de transparencia que compite con el que se encuentra en materiales inorgánicos como el cristal. No sorprende que ciertas alteraciones en la composición de la córnea o el cristalino puedan reducir significativamente su transparencia y tener consecuencias graves para la percepción visual. En efecto, las opacidades del cristalino conocidas como cataratas explican más o menos el 50% de los casos de ceguera en el mundo, y casi todas las personas mayores de 70 años experimentan cierta pérdida de transparencia en el cristalino que termina por degradar la calidad de la experiencia visual. Felizmente, hay tratamientos quirúrgicos satisfactorios para las cataratas que pueden restablecer la visión en la mayoría de los casos. Además, el reconocimiento de que un factor importante en la producción de cataratas es la exposición a la radiación solar ultravioleta (UV) aumentó la conciencia del público acerca de la necesidad de proteger el cristalino (y la retina) reduciendo la exposición a la luz UV mediante uso de lentes para sol. Pupila Iris Córnea Humor acuoso en la cámara anterior Fibras de la zónula Cámara posterior Músculo ciliar Cristalino Coroides Esclerótica Humor vítreo Retina Fóvea Papila óptica Nervio óptico y vasos retinianos FIGURA 11.1 Anatomía del ojo humano. Más allá de transmitir de manera eficiente la energía luminosa, la función principal de los componentes ópticos del ojo es lograr una imagen enfocada sobre la superficie de la retina. La córnea y el cristalino son los principales responsables de la refracción (inclinación) de la luz necesaria para la formación de imágenes enfocadas sobre los fotorreceptores de la retina (Figura 11.2). La córnea es responsable de la mayor parte de la refracción necesaria, como puede apreciarse si se consideran las imágenes turbias y fuera de foco que se experimentan al nadar debajo del Sin acomodación Córnea Humor acuoso Con acomodación Iris Músculo ciliar Cristalino Humor vítreo Fibras de la zónula FIGURA 11.2 Acomodación en el ojo humano. El diagrama que muestra la porción anterior del ojo humano en el estado sin acomodación (izquierda) y con acomodación (derecha). La acomodación para enfocar objetos cercanos comprende la contracción del músculo ciliar, el cual reduce la tensión en las fibras de la zónula y permite que la elasticidad del cristalino aumente su curvatura (flechas amarillas). EL OJO 231 RECUADRO 11A La miopía y otros defectos de refracción Las discrepancias ópticas entre los distintos componentes del ojo hacen que la mayoría de la población humana tenga alguna forma de defecto de refracción, conocido como ametropía. A las personas que no pueden enfocar con claridad un objeto distante se las llama miopes (Figura B). La miopía puede producirse porque la superficie corneana es demasiado curva o porque el globo ocular es demasiado alargado. En cualquiera de los casos, con el cristalino lo más plano posible, la imagen de los objetos distantes se enfocan por delante de la retina y no sobre ella. A los sujetos que no pueden enfocar sobre objetos cercanos se los denomina hipermétropes. La hipermetropía puede presentarse porque el globo ocular es demasiado corto o el sistema de refracción es demasiado débil (Figura C). Aun con el cristalino en su estado más redondeado, la imagen está fuera de foco sobre la superficie retiniana (se enfoca en algún punto por detrás de la retina). Tanto la miopía como la hipermetropía pueden corregirse mediante lentes apropiadas –cóncavas (negativas) y convexas (positivas), respectivamente– o mediante la técnica cada vez más popular de la cirugía corneana. La miopía es, con mucho, la ametropía más frecuente; se estima que el 50% de la población de los Estados Unidos está afectada. Dada la gran cantidad de personas que necesitan gafas, lentes de contacto o cirugía para corregir este defecto de refracción, uno naturalmente se pregunta cómo se arreglaban los individuos miopes antes de que se inventaran las gafas hace solo algunos siglos. A partir de lo que se sabe en el presente acerca de la miopía, la visión de la mayoría de los individuos puede haber sido considerablemente mejor en tiempos antiguos. La base para esta afirmación es el hallazgo sorprendente de que el crecimiento del globo ocular está muy influido por la luz enfocada que cae sobre la retina. Este fenómeno fue descrito por primera vez en 1977 por Torsten Wiesel y Elio Raviola en la Escuela Médica de Harvard Medical School. Ellos estudiaron monos criados con sus párpados suturados (el mismo enfoque utilizado para demostrar los efectos de la privación visual sobre las conexiones corticales en el sistema visual; véase Capítulo 24), procedimiento que priva al ojo de las imágenes retinianas enfocadas. Estos autores observaron que los animales que crecían hasta la madurez en estas condiciones mostraban una elongación del globo ocular. El efecto de la privación de la luz enfocada parece ser local, ya que el crecimiento anormal del ojo se produce en animales de experimentación, aun cuando se corta el nervio óptico. En efecto, si solo se priva de luz enfocada a una porción de la superficie retiniana, solo crece en forma anormal esa región del globo ocular. Aunque el mecanismo de control del globo ocular mediado por la luz no se conoce plenamente, muchos expertos creen ahora que la prevalencia de miopía se debe a cierto aspecto de la civilización moderna –tal vez aprender a leer y escribir a una edad temprana– que interfiere con el control normal por retroalimentación de la visión sobre el desarrollo ocular y conduce a un alargamiento anormal del globo ocular. Un corolario de esta hipótesis es que si los niños (o más probablemente sus padres) quisieran mejorar su visión, podrían lograrlo practicando la visión lejana para equilibrar la “sobrecarga” de trabajo cercano. Por supuesto, en la práctica, la mayoría de los individuos quizá elegirían utilizar gafas o lentes de contacto, o someterse a una cirugía corneana en lugar de participar en la onerosa práctica cotidiana que se presume sería necesaria. Mucho más, no todos están de acuerdo en que este remedio sería eficaz, y algunos investigadores (y laboratorios) exploran la posibilidad de una intervención farmacológica durante la infancia, época en la que se presume que se genera el desarrollo ocular anormal. En cualquier caso, es notable que la privación de luz enfocada sobre la retina produzca un crecimiento compensador del ojo y que este circuito de retroalimentación se altere con tanta facilidad. Aun los individuos con una visión normal (emetrópica) cuando son adultos jóvenes finalmente experimentan dificultad para enfocar sobre objetos cercanos. Una de las muchas consecuencias del envejecimiento es que el cristalino pierde su elasticidad; como resultado, se reduce en forma gradual la curvatura máxima del cristalino que puede lograrse cuando se (A) Emetropía (normal) (B) Miopía (miope) (C) Hipermetropía (hipermétrope) Defectos de refracción. (A) En el ojo normal, con los músculos ciliares relajados, una imagen de un objeto distante se enfoca sobre la retina. (B) En la miopía, los rayos luminosos se enfocan por delante de la retina. (C) En la hipermetropía, las imágenes se enfocan en un punto más allá de la retina. contrae el músculo ciliar. El punto cercano (el más próximo que se puede enfocar con claridad) retrocede y los objetos (como este libro) deben alejarse cada vez más del ojo para poder enfocarlos sobre la retina. En algún momento, por lo general a comienzos de la edad media, la capacidad de acomodación del ojo se encuentra tan reducida que las tareas visuales cercanas como la lectura se vuelven difíciles o imposibles (Figura D). Este trastorno se denomina presbicia y puede corregirse mediante lentes convexas para tareas de visión cercana o mediante lentes bifocales si también se presenta miopía (que requiere una corrección negativa). La corrección bifocal presenta un problema particular para los que prefieren los lentes de contacto. Dado que los lentes de contacto flotan sobre la superficie de la córnea, no funciona contar con la corrección para distancia arriba y la corrección cercana abajo (como sucede con las gafas bifocales convencionales) (aunque recientemente se han utilizado lentes de contacto “polifocales” con cierto éxito). (Continúa en la página siguiente) CAPÍTULO 11 RECUADRO 11A (continúa) (D) (D) Cambios en la capacidad del cristalino para redondearse (acomodarse) con la edad. El gráfico también muestra cómo cambia acomodación, una medida óptima del poder de refracción del cristalino, se expresa en dioptrías. (De Westheimer, 1974.) ¡Una solución sorprendentemente eficaz de este problema para algunos individuos que utilizan lentes de contacto fue colocar un lente para corrección cercana en un ojo y otro para corrección lejana en el otro! El éxito de este enfoque es otro testimonio de la notable capacidad del sistema visual para adaptarse a una amplia variedad de demandas poco comunes. Bibliografía Bock, G. and K. Widdows (1990) Myopia and the Control of Eye Growth. Ciba Foundation Symposium 155. Chichester: Wiley. Coster, D. J. (1994) Physics for Ophthalmologists. Edinburgh: Churchill Livingston. 14 13 12 0,07 11 10 9 8 7 6 0,1 Rango de acomodación 0,2 5 4 3 2 1 0 Punto cercano (m) el punto cercano (el punto más próximo al ojo que se puede enfocar con claridad). La Amplitud de la acomodación (dioptrías) 232 0,5 1,0 15 20 25 30 35 40 45 Edad (años) Kaufman, P. L. and A. Alm (eds.) (2002) Adler’s Physiology of the Eye: Clinical Application, 10th Ed. St. Louis, MO: Mosby Year Book. Sherman, S. M., T. T. Norton and V. A. Casagrande (1977) Myopia in the lid-sutured tree shrew. Brain Res. 124:154-157. Wallman, J., J. Turkel and j. Tractman (1978) Extreme myopia produced by modest changes in agua. El agua, por oposición al aire, tiene un índice de refracción próximo al de la córnea; en consecuencia, la inmersión en agua prácticamente elimina la refracción que se desarrolla en condiciones normales en la interfase aire/córnea; por lo tanto, la imagen ya no está enfocada en la retina. El cristalino tiene un poder de refracción considerablemente menor que la córnea; sin embargo, la refracción aportada por el cristalino es ajustable y permite que los objetos ubicados a diferentes distancias del observador se coloquen en un foco definido. Los cambios dinámicos en el poder de refracción del cristalino se denominan acomodación. Cuando observamos objetos distantes, el cristalino se torna relativamente delgado y plano, y tiene el poder de refracción más bajo. Para la visión cercana, el cristalino se torna más grueso y redondeado, y tiene el poder de refracción mayor (véase Figura 11.2). Estos cambios son resultado de la actividad del músculo ciliar que rodea al cristalino. Este último es mantenido en el lugar por bandas de tejido conectivo de disposición radial llamadas fibras de la zónula y que están fijadas al músculo ciliar. Por lo tanto, la forma del cristalino está determinada por dos fuerzas opuestas: la elasticidad del cristalino, que tiende a mantenerlo redondeado (al extraerlo del ojo, el cristalino se vuelve esférico), y la tensión ejercida por las fibras de la zónula, que tiende a aplanarlo. Cuando se visualizan objetos distantes, la fuerza de las fibras de la zónula es mayor que la elasticidad del cristalino, y este adopta la forma más aplanada, apropiada para visualizar a distancia. Para enfocar objetos más cercanos, es necesario relajar la tensión en las 50 55 60 65 70 early visual experience. Science 201: 1249-1251. Wallman, J. and J. Winawer (2004) Homeostasis of eye growth and the question of myopia. Neuron 43: 447-468. Wiesel, T. N. and E. Raviola (1977) Myopia and eye enlargement after neonatal lid fusion in monkeys. Nature 266: 66-68. fibras de la zónula, lo que permite que la elasticidad intrínseca del cristalino aumente su curvatura. Esta relajación se logra por la contracción similar a un esfínter del músculo ciliar. Dado que este músculo forma un anillo alrededor del cristalino, cuando se contrae, los puntos de inserción de las fibras de la zónula se mueven hacia el eje central del ojo y reducen así la tensión sobre el cristalino. Lamentablemente, los cambios en la forma del cristalino no siempre pueden producir una imagen enfocada sobre la retina, en cuyo caso solo es posible lograr una imagen nítida con ayuda de lentes correctivos adicionales (véase Recuadro 11A). Los ajustes en el tamaño de la pupila también contribuyen a la claridad de las imágenes formadas sobre la retina. Al igual que las imágenes formadas por otros instrumentos ópticos, las generadas por el ojo están afectadas por aberraciones esféricas y cromáticas, que tienden a enturbiar la imagen retiniana. Como estas aberraciones son máximas para los rayos de luz que pasan más alejados del centro del cristalino, el estrechamiento de la pupila reduce tanto la aberración esférica como la cromática, al igual que el cierre del diafragma del iris en el lente de una cámara mejora la nitidez de una imagen fotográfica. La reducción del tamaño de la pupila también aumenta la profundidad del campo, o sea, la distancia dentro de la cual se observan los objetos sin que estén borrosos. Sin embargo, una pupila pequeña también limita la cantidad de luz que alcanza la retina y, en condiciones de iluminación tenue, la agudeza visual se vuelve limitada por la cantidad de fotones disponibles más que por las aberraciones ópticas. Por lo tanto, una pupila ajustable proporciona un medio EL OJO 233 Mácula amarilla FIGURA 11.3 Superficie interna de la retina, vista con un oftal- Fóvea moscopio. El disco óptico es la región donde los axones de las células ganglionares abandonan la retina para formar el nervio óptico; también se caracteriza por la entrada y la salida, respectivamente, de las arterias y venas Disco óptico (papila) Rama de la vena oftálmica Rama de la arteria oftálmica oftálmicas que irrigan la retina. La mácula lútea puede observarse en el centro del eje óptico (el disco óptico tiene una localización nasal); la mácula es la región de la retina que tiene la máxima agudeza visual. La fóvea es una depresión u hoyo, de aproximadamente 1,5 mm de diámetro que se sitúa en el centro de la mácula. fotorreceptores. El daño de esta región de la retina, como ocurre en la degeneración macular relacionada con la edad, tiene un impacto devastador sobre la percepción visual (Recuadro 11C). Circuito retiniano eficaz para reducir las aberraciones ópticas, mientras aumenta al máximo la profundidad del campo en la medida en que lo permiten los diferentes niveles de iluminación. El tamaño de la pupila está controlado por inervación de las divisiones simpática y parasimpática del sistema motor visceral, que a su vez están modulados por varios centros del tronco del encéfalo (véanse Capítulos 20 y 21). A pesar de su localización periférica, la retina, que es la porción neural del ojo, es en realidad parte del sistema nervioso central. Durante el desarrollo, la retina se forma como una evaginación del diencéfalo denominada vesícula óptica, que sufre una invaginación para formar la copa óptica (Figura 11.4; véase también Capítulo 22). La pared interna de la copa óptica da ori- La superficie de la retina FIGURA 11.4 Desarrollo del ojo humano. (A) La retina se desarrolla Mediante el uso de un oftalmoscopio, la superficie interna de la retina o fondo de ojo puede visualizarse a través de la pupila (Figura 11.3). Muchos vasos sanguíneos, arteriales y venosos, se abren en abanico sobre la superficie interna de la retina. Estos vasos sanguíneos se originan en la arteria y la vena oftálmicas, que ingresan en el ojo a través de un área circular blancuzca conocida como disco óptico o papila óptica. El disco óptico también es el sitio donde los axones retinianos abandonan el ojo y discurren a través del nervio óptico para alcanzar las estructuras diana en el tálamo y el mesencéfalo. Esta región de la retina no contiene fotorreceptores y, debido a que es insensible a la luz, produce los fenómenos perceptivos conocidos como punto ciego (Recuadro 11B). Además de ser un reparo retiniano conspicuo, el aspecto del disco óptico es una medida útil de la presión intracraneana. El espacio subaracnoideo que rodea al nervio óptico se continúa con el del encéfalo; en consecuencia, los incrementos en la presión intracraneana –signo de problemas neurológicos graves como las lesiones ocupantes– pueden detectarse como una tumefacción del disco óptico. Otra característica sobresaliente del fondo de ojo es la mácula lútea, una mancha ovalada que contiene pigmento amarillo (xantófilo), de más o menos 3 milímetros de diámetro y situada cerca del centro de la retina. La mácula es la región de la retina que aporta una alta agudeza visual (la capacidad para reconocer los detalles finos). La agudeza es máxima en el centro de la mácula, una pequeña depresión o pozo en la retina denominada fóvea. El pigmento xantófilo tiene un papel protector al filtrar las longitudes de onda ultravioletas que podrían ser nocivas para los como una evaginación del tubo neural, denominada vesícula óptica. (B) La vesícula óptica se invagina para formar la copa óptica. (C, D) La pared interna de la copa óptica se convierte en la retina neural, mientras que la pared externa se convierte en el epitelio pigmentario. (A-C, de Hilfer y Yang, 1980; D, cortesía de K. Tosney.) (A) Embrión de 4 mm (B) Embrión de 4,5 mm Copa óptica Ventrículo Vesícula óptica (C) Embrión de 5 mm (D) Embrión de 7 mm Formación del cristalino Cristalino Retina Epitelio pigmentario 234 CAPÍTULO 11 RECUADRO 11B El punto ciego Parece lógico suponer que un defecto del campo visual, o escotoma, resultaría evidente para cualquier individuo que padeciera de esta patología. Sin embargo, cuando el déficit afecta una región periférica del campo visual, a menudo el escotoma pasa inadvertido hasta que un accidente automovilístico u otro percance pone claramente de manifiesto el déficit sensitivo. De hecho, todos nosotros tenemos un escotoma fisiológico: el “punto ciego”, una brecha en cada campo visual monocular que corresponde a la localización del disco óptico (la región de la retina libre de receptores donde el nervio óptico abandona el ojo; véase Figura 11.1). Para localizar el “punto ciego” de nuestro ojo derecho, sostenga este libro aproximadamente a 30 cm de su rostro, cierre el ojo izquierdo y fije la vista en la X del recuadro dibujado abajo. Tome un lápiz con su mano derecha y, sin alterar la fijación, mueva lentamente la punta hacia la X desde el lado derecho de la página. En algún punto, la punta (en realidad, todo el extremo) del lápiz desaparece. Marque ese punto y siga moviendo el lápiz hacia la izquierda hasta que vuelva a aparecer y luego haga otra marca. Usted puede determinar los bordes del punto ciego en el eje vertical de la misma forma moviendo el lápiz hacia arriba y abajo a lo largo del plano de la página de modo que su camino caiga entre las dos marcas horizontales. Para probar que no se percibe la información del espacio visual delimitado por las marcas, coloque una moneda en el interior del área demarcada. Cuando usted fija la mirada en la X con ambos ojos y cierra el ojo izquierdo, la moneda desaparece. ¿Cómo podemos no tomar conciencia de un defecto tan grande (por lo común de 5-8°) en el campo visual? El disco óptico se localiza en la retina nasal de cada ojo. Con ambos ojos abiertos, la información sobre la región correspondiente del campo visual está disponible desde la retina temporal del otro ojo. Pero esto no explica por qué el punto ciego no es detectada cuando se cierra un ojo. Cuando se visualiza el mundo con un solo ojo, el sistema visual parece “llenar” la parte faltante de la escena con la información aportada por las regiones que rodean al disco óptico. Para observar este fenómeno, observe lo que sucede cuando el lápiz se ubica a través de la representación del disco óptico: notablemente, ¡se ve completo! Aunque algunos registros electrofisiológicos han mostrado que las neuronas en la corteza visual gen a la retina, mientras que la pared externa origina el epitelio pigmentario retiniano. Este epitelio es una estructura delgada que contiene melanina y reduce la dispersión retrógrada de la luz que ingresa en el ojo y, como veremos, desempeña un papel crítico en el mantenimiento de la maquinaria de fototransducción de los fotorreceptores retinianos. Consistente con su estado como porción plena del sistema nervioso central, la retina muestra un circuito neural complejo que convierte la actividad eléctrica graduada de las neuronas fotosensibles especializadas –los fotorreceptores– en potenciales de acción que discurren hasta los puntos diana centrales a través cuyos campos receptivos se ubican en la representación del disco óptico pueden ser activadas por la estimulación de las regiones que rodean al disco óptico del ojo contralateral –lo que sugiere que el relleno del punto ciego se basa en los mecanismos que integran la información de diferentes puntos en el campo visual– el mecanismo de este fenómeno no está claro. Herman von Helmholtz señaló en el siglo xix que puede ser que esta parte del mundo visual sea simplemente ignorada y se completa el lápiz a través del punto ciego porque el resto de la escena “colapsa” a su alrededor. Bibliografía Fiorani, M., M. G. P. Rosa, R. Gattass and C. E. Rocha-Miranda (1992) Dynamic surrounds of receptive fields in striate cortex: A physiological basis for perceptual completion? Proc. Natl. Acad. Sci. USA 89: 8547-8551. Gilbert, C. D. (1992) Horizontal integration and cortical dynamics. Neuron 9: 1-13. Ramachandran, V. S. and T. L. Gregory (1991) Perceptual filling in of artificially induced scotomas in human vision. Nature 350: 699-702. Von Helmholtz, H. (1968). Helmholtz’s Treatise on Physiological Optics, Vol. III, J. P. C. Southall (ed.). New York: Dover Publications. Trans, of 3rd German Ed., 1910. Pp. 204ff. de axones en el nervio óptico. Aunque tiene los mismos tipos de elementos funcionales y neurotransmisores que se encuentran en otras partes del sistema nervioso central, la retina no comprende tantas clases de neuronas, y están organizadas de una forma que ha sido menos difícil de desentrañar que los circuitos de otras áreas del encéfalo. Hay cinco clases básicas de neuronas en la retina: fotorreceptores, células bipolares, células ganglionares, células horizontales y células amacrinas. Los cuerpos celulares y las prolongaciones de estas neuronas están apilados en capas alternantes, y los cuerpos celulares se sitúan en las capas nuclear in- EL OJO 235 RECUADRO 11C Degeneración macular Se estima que seis millones de personas en los Estados Unidos sufren un trastorno conocido como degeneración macular relacionada con la edad (DME), que produce una pérdida progresiva de la visión central. Como la visión central es fundamental para la vista, las enfermedades que afectan la mácula (véase Figura 11.1) limitan gravemente la capacidad para realizar tareas visuales. En efecto, la DME es la causa más frecuente de pérdida de visión en las personas mayores de 55 años y su incidencia aumenta con el porcentaje creciente de ancianos en la población. El problema subyacente, que aún se comprende poco, es la degeneración de los fotorreceptores. En general, los pacientes notan por primera vez una visión central borrosa cuando realizan tareas como la lectura. Las imágenes también pueden aparecer distorsionadas. Se utiliza un papel para gráficos, conocido como rejilla de Amsler, como una prueba sencilla para detectar los signos incipientes de DME. Al enfocar sobre un punto marcado en el centro de la rejilla (que se asemeja a un trozo de papel milimetrado), el paciente puede evaluar si las líneas paralelas y perpendiculares sobre la rejilla parecen borrosas o distorsionadas. A menudo, la visión central borrosa progresa hasta la presencia de puntos ciegos en el interior de la visión central, y en la mayoría de los casos ambos ojos terminan por afectarse. Aunque claramente el riesgo de desarrollar DME aumenta con la edad, no se conocen las causas de la enfermedad. En distintos estudios se ha considerado la influencia de factores hereditarios, enfermedad cardiovascular, factores ambientales como el tabaquismo y la exposición a la luz y causas nutricionales. En efecto, puede ser que todos ellos contribuyan al riesgo de sufrir DME. En términos descriptivos, la degeneración macular se divide ampliamente en dos tipos. En la forma exudativa-neovascular, o DME “húmeda”, que constituye alrededor del 10% de los casos, hay un crecimiento anormal de los vasos sanguíneos debajo de la mácula. Estos pierden líquido y sangre dentro de la retina y producen daño en los fotorreceptores. La DME húmeda tiende a progresar con rapidez y puede producir daño grave; en tan solo unos meses, es posible desarrollar una pérdida rápida de la visión central. El tratamiento para esta forma de la enfermedad es la terapia con láser. Al transferir energía térmica, el haz de láser destruye los vasos sanguíneos con pérdida debajo de la mácula y hace así más lenta la velocidad de pérdida visual. Una desventaja de este enfoque es que la energía térmica elevada transmitida por el haz también destruye el tejido sano cercano. Una mejoría en el tratamiento con láser de la DME comprende un fármaco activado por la luz dirigido hacia los vasos sanguíneos anormales. Una vez que el fármaco se administra, se aplican pulsos de láser de energía relativamente baja en los vasos sanguíneos anormales para estimular el fármaco, que a su vez destruye los vasos sanguíneos anormales con un daño mínimo para el tejido circundante. El 90% restante de los casos de DME son de la forma no exudativa o “seca”. En estos pacientes hay una desaparición gradual del epitelio pigmentario retiniano, que conduce a áreas circunscritas de atrofia. Como la pérdida de fotorreceptores sigue a la desaparición del epitelio pigmentario, las áreas retinianas afectadas tienen poca función visual o ninguna. La pérdida de visión por la DME seca se desarrolla de manera más gradual, en los casos típicos en el curso de muchos años. Estos pacientes suelen retener cierta visión central, aunque la pérdida puede llegar a afectar la realización de las tareas que requieran ver detalles. Lamentablemente, en este momento no hay un tratamiento para la DME seca. Un enfoque nuevo, radical y bastante interesante que ofrece algo promisorio implica la reposición quirúrgica de la retina alejada del área anormal. En ocasiones, la degeneración macular aparece en individuos mucho más jóvenes. Gran parte de estos casos es consecuencia de distintas mutaciones, cada una de ellas con sus propias manifestaciones clínicas y causas genéticas. La forma más frecuente de degeneración macular juvenil se conoce como enfermedad de Stargardt, que se hereda de forma autonómica recesiva. Por lo general, los pacientes reciben el diagnóstico antes de los 20 años de edad. Aunque la progresión de la pérdida visual es variable, la mayoría de estos pacientes se considera legalmente ciega alrededor de los 50 años. Las mutaciones que produce la enfermedad de Stargardt se identificaron con el gen ABCR, que codifica una proteína que transporta retinoides a través de la membrana de los fotorreceptores. Por lo tanto, el ciclo visual de la regeneración de los fotopigmentos puede estar interrumpido en esta forma de degeneración macular, presumiblemente por proteínas disfuncionales codificadas por el gen anormal (véase Figura 11.6). Es interesante destacar que el gen ABCR se expresa solo en los bastones, lo que sugiere que los conos pueden tener sus propias enzimas del ciclo visual. En las formas esporádicas de DME, se ha identificado la variación en la secuencia en dos genes involucrados en la cascada completa –factor B y factor H– como factores de riesgo, lo que implica que la cascada completa es un punto diana terapéutico potencial para algunas formas de DME. Bibliografía Bressler, N. M., S. B. Bressler and S. L. Fine (2006) Neovascular (exudative) age-related macular degeneration. In Retina, 4th Ed., Vol. 2: Medical Retina. S. J. Ryan (ed.-in-chief). Philadelphia: Elsevier Mosby, pp. 1074-1114. Bressler, S. B., N. M. Bressler, S. H. Sarks and J. P. Sarks (2006) Age-related macular degeneration: Nonneovascular early AMD, intermediate AMD, and geographic atrophy. In Retina, 4th Ed., Vol. 2: Medical Retina. S. J. Ryan (ed.-in-chief). Philadelphia: Elsevier Mosby, pp. 1041-1074. Deutman, A. F., C. B. Hoyng and J. J. C. van Lith-Verhoeven (2006) Macular dystrophies. In Retina, 4th Ed., Vol. 2: Medical Retina. S. J. Ryan (ed.-in-chief). Philadelphia: Elsevier Mosby, pp. 1163-1210. Fine, S. L., J. W. Berger, M. G. Maguire and A. C. Ho (2000) Drug therapy: Age-related macular degeneration. New Eng. J. Med. 342: 483-492. The Foundation Fighting Blindness of Hunt Valley, MD, maintains a web site that provides updated information about many forms of retinal degeneration: www.blindness.org RetNet provides updated information, including references to original articles, on genes and mutations associated with retinal diseases: www.sph. uth.tmc.edu/RetNet 236 CAPÍTULO 11 (B) (A) Corte de retina Epitelio pigmentario Cone Cono Cone Cono Bastón Rod Bastón Rod Cone Cono Rod Bastón Segmentos externos de los fotorreceptores Distal Célula Horizontal horizontal cell Laterallateral de Flujo information información flow Luz Célula Bipolar bipolar cell Célula Amacrine amacrina cell Célula Ganglion cell ganglionar Cono Espacio citoplasmático Membrana plasmática óptico optic nerve To nervio Al Segmento externo Cilio Núcleo Vesículas sinápticas Capa de fibras nerviosas FIGURA 11.5 Estructura de la retina. (A) Corte de la retina que muestra la Segmento interno Segmento interno Capa de células ganglionar es Luz Mitocondria Terminación sináptica Capa nuclear interna Bastón Discos Segmento externo Capa plexiforme externa Capa plexiforme interna Proximal (C) Virtical information flow Vertical Capa nuclear externa Terminación sináptica disposición global de las capas retinianas. (B) Diagrama del circuito básico de la retina. Una cadena de tres neuronas –fotorreceptor, célula bipolar, célula ganglionar– proporciona la vía más directa para transmitir la información visual hasta el encéfalo. Las células horizontales y las células amacrinas median las interacciones laterales en las capas plexiforme externa y plexiforme interna, respectivamente, Los términos interna y externa designan distancias relativas desde el centro del ojo (interna, cerca del centro del ojo; externa, lejos del centro del ojo, o hacia el epitelio pigmentario). (C) Diferencias estructurales entre bastones y conos. Aunque generalmente tienen una estructura similar, los bastones y los conos difieren en su tamaño y su forma, y en la disposición de los discos membranosos en sus segmentos externos. terna, nuclear externa y en la capa de células ganglionares, y las prolongaciones y los contactos sinápticos se localizan en las capas plexiforme interna y plexiforme externa (Figura 11.5A, B). La retina contiene dos tipos de fotorreceptores: bastones y conos (Figura 11.5B, C). Ambos tipos tienen un segmento externo (adyacente al epitelio pigmentario), compuesto por discos membranosos que contienen fotopigmento sensible a la luz, y un segmento interno que contiene el núcleo celular y da origen a las terminaciones sinápticas que hacen contacto con células bipolares u horizontales. Una cadena de tres neuronas –célula fotorreceptora a célula bipolar a célula ganglionar– es la vía más directa de flujo de información desde los fotorreceptores hasta el nervio óptico. La absorción de luz por el fotopigmento en el segmento externo de los fotorreceptores inicia una cascada de acontecimientos que cambia el potencial de membrana del receptor y, por lo tanto, la cantidad de neurotransmisor liberado por las terminaciones del fotorreceptor. (Este proceso, denominado fototransducción, se explica en detalle más adelante en el capítulo.) Las sinapsis entre las terminaciones del fotorreceptor y las células bipolares (y EL OJO 237 RECUADRO 11D Retinitis pigmentaria La retinitis pigmentaria (RP) se refiere a un grupo heterogéneo de trastornos oculares hereditarios caracterizados por una pérdida progresiva de la visión debida a una degeneración gradual de los fotorreceptores. Se estima que 100 000 individuos en los Estados Unidos tienen retinitis pigmentaria. A pesar del nombre, la inflamación no es una parte prominente del proceso patológico; en cambio, las células de los fotorreceptores parecen morir por apoptosis (determinada por la presencia de fragmentación del DNA). La clasificación de este grupo de trastornos bajo una denominación se basa en las características clínicas comunes observadas en estos pacientes. Las características de la RP son ceguera nocturna, reducción de la visión periférica, estrechamiento de los vasos retinianos y la migración del pigmento desde el epitelio pigmentario retiniano interrumpido hasta la retina, formando grumos de distintos tamaños, a menudo próximos a los vasos sanguíneos retinianos. En los casos típicos, los pacientes primero notan dificultad para ver por la noche debido a la pérdida de los fotorreceptores de los bastones; entonces, los fotorreceptores conos restantes se convierten en la fuente principal de la función visual. Después de muchos años, los conos también se degeneran, lo que conduce a una pérdida progresiva de la visión. En la mayoría de los pacientes con RP, los defectos campimétricos comienzan en la periferia media, entre los 30° y los 50° desde el punto de fijación de la fóvea. Las regiones defectuosas crecen en forma gradual y dejan islotes de visión en la periferia y un campo central contraído, trastorno conocido como visión en túnel. Cuando el campo visual se contrae hasta 20° o menos, o la visión central es de 20/200 o peor, el paciente se categoriza como legalmente ciego. Los patrones de herencia indican que la RP puede transmitirse en una forma ligada al cromosoma X, autosómica dominante o recesiva. En los Estados Unidos, el porcentaje de estos tipos genéticos se estima en el 9, el 16 y el 41%, respectivamente. Cuando solo un miembro de un árbol genealógico tiene RP, el caso se clasifica como “simple”, lo que representa alrededor de un tercio de todos los casos. Entre los tres tipos genéticos de RP, la forma autosómica dominante es la más leve. A menudo, estos pacientes retienen una buena visión central hasta los 60 años o más. Por el contrario, aquellos con la forma recesiva ligada al cromosoma X de la enfermedad suelen declararse legalmente ciegos alrededor de los 30 a 40 años de edad. Sin embargo, la gravedad y la edad de inicio de estos síntomas varía mucho entre los pacientes con el mismo tipo de RP, e incluso dentro de la misma familia (cuando presumiblemente todos los miembros afectados tienen la misma mutación genética). Hasta el presente, se identificaron mutaciones inductoras de RP en 30 genes. Muchos de estos codifican proteínas específicas de los fotorreceptores, y varios se asocian con la fototransducción en los bastones. Entre los últimos están los genes para rodopsina, subunidades de la fosfodiesterasa del cGMP y el canal con puerta de cGMP. Se observaron numerosas mutaciones en cada uno de estos genes clonados. Por ejemplo, en el caso del gen de la rodopsina, se identificaron 90 mutaciones diferentes entre los pacientes con RP autosómica dominante. La heterogeneidad de la RP en todos los niveles, desde las mutaciones genéticas hasta los síntomas clínicos, tiene implicaciones importantes para comprender la patogenia de la enfermedad y para el diseño de los tratamientos. Dada la etiología molecular compleja de la RP, es poco probable que un solo mecanismo celular explique la enfermedad en todos los casos. Más allá de la mutación o de la secuencia causal específica, la pérdida de visión que es más crítica para los pacientes con RP se debe a la degeneración gradual de los conos. En muchos casos, la mutación que produce la RP afecta a proteínas que no se expresan ni siquiera en los conos; el ejemplo fundamental es la rodopsina, el pigmento visual específico de los bastones. Por lo tanto, la pérdida de conos puede ser un resultado indirecto de una mutación específica de los bastones. La identificación de mecanismos celulares que producen directamente degeneración de los conos debería conducir a un mejor conocimiento de esta patología. Bibliografía Rivolta, C., Sharon, M. M. DeAngelis and T. P. Dryja (2002) Retinitis pigmentosa and allied diseases: Numerous diseases, genes and inheritance patterns. Hum. Molec. Genet. 11: 1219-1227. Weleber, R. G. and K. Gregory-Evans (2006) Retinitis pigmentosa and allied disorders. In Retina, 4th Ed., Vol. 1: Basic Science and Inherited Retinal Diseases. S. J. Ryan (ed.-in-chief). Philadelphia: Elsevier, pp. 395-498. Wright, A. F., C. F. Chakarova, M. M. Abd ElAziz and S. S. Bhattacharya (2010) Photoreceptor degeneration: Genetic and mechanistic dissection of a complex trait. Nature Rev. Genet. 11: 273-284. The Foundation Fighting Blindness of Hunt Valley, MD, maintains a web site that provides updated information about many forms of retinal degeneration: www.blindness.org RetNet provides updated information, including references to original articles, on genes and mutations associated with retinal diseases: www.sph. uth.tmc.edu/RetNet Aspecto característico de la retina en pacientes con retinitis pigmentaria. Obsérvense los grupos oscuros de pigmento que constituyen el sello de este trastorno. 238 CAPÍTULO 11 las células horizontales) ocurren en la capa plexiforme externa; más específicamente, los cuerpos celulares de los fotorreceptores constituyen la capa nuclear externa, mientras que los cuerpos celulares de las células bipolares se sitúan en la capa nuclear interna. A su vez, las prolongaciones axónicas cortas de las células bipolares hacen contactos sinápticos sobre las prolongaciones dendríticas de las células ganglionares en la capa plexiforme interna. Los axones mucho más grandes de las células ganglionares forman el nervio óptico y transmiten información sobre la estimulación retiniana al resto del sistema nervioso central. Los otros dos tipos de neuronas retinianas, células horizontales y células amacrinas, tienen sus cuerpos celulares en la capa nuclear interna y poseen prolongaciones que están limitadas a las capas plexiformes externa e interna, respectivamente (véase Figura 11.5B). Las prolongaciones de las células horizontales permiten interacciones laterales entre los fotorreceptores y las células bipolares que se cree mantienen la sensibilidad del sistema visual al contraste sobre una amplia gama de intensidades de luz o luminancia. Las prolongaciones de las células amacrinas son postsinápticas a las terminaciones de las células bipolares y presinápticas a las dendritas de las células ganglionares. Se considera que diferentes subclases de células amacrinas realizan distintas contribuciones a la función visual. Por ejemplo, un tipo de célula amacrina tiene un papel obligatorio en la vía que transmite información desde los fotorreceptores de los bastones hasta las células ganglionares de la retina. Se cree que otro tipo es fundamental para generar las respuestas selectivas de dirección que muestra un subgrupo especializado de células ganglionares. La variedad de células amacrinas muestra la regla más general: incluso con solo cinco clases básicas de neuronas retinianas puede haber una considerable diversidad dentro de una clase dada. Esta diversidad también es un sello de las células ganglionares de la retina y la base para las vías que transmiten diferentes tipos de información hacia los puntos diana centrales de una forma paralela, tema que se considera con mayor detalle en el Capítulo 12. Epitelio pigmentario de la retina La disposición espacial de las capas retinianas al principio parece contradictoria: los rayos de luz deben atravesar varios elementos no sensibles a la luz y la vasculatura retiniana (que se ramifica extensamente sobre la superficie interna de la retina) antes de alcanzar los segmentos externos de los fotorreceptores, que es donde se absorben los fotones (véase Figura 11.5A, B). La razón para esta característica curiosa de la organización retiniana reside en la relación especial que existe entre los segmentos externos de los fotorreceptores y el epitelio pigmentario. Las células que conforman el epitelio pigmentario de la retina tienen prolongaciones largas que se extienden en la capa de fotorreceptores, rodeando los extremos de los segmentos externos de cada fotorreceptor (Figura 11.6A). El epitelio pigmentario desempeña dos papeles fundamentales para la función de los fotorreceptores de la retina. En primer lugar, los discos membranosos del segmento externo, que albergan el fotopigmento sensible a la luz y otras proteínas involucradas en la fototransducción, tienen un tiempo de vida relativamente limitado de unos 12 días. Se forman de manera continua nuevos discos del segmento externo cerca de la base del segmento, mientras se eliminan o “desprenden” los discos más antiguos en la punta del segmento externo (Figura 11.6B). Durante su vida, los discos se mueven más progresivamente desde la base del segmento externo hasta la punta, donde el epitelio pigmentario desempeña un papel esencial en la eliminación de los discos receptores gastados. El desprendimiento comprende el “pinzamiento” de un conjunto de discos receptores por la membrana del segmento externo del fotorreceptor. Luego, este conjunto de discos encerrado es fagocitado por el epitelio pigmentario (Figura 11.6C). El segundo papel del epitelio, como destacaremos con más detalle en la siguiente sección, es regenerar las moléculas de fotopigmento una vez que han estado expuestas a la luz. El fotopigmento es reciclado continuamente entre el segmento externo del fotorreceptor y el epitelio pigmentario. Estas consideraciones, junto con el hecho de que los capilares en la coroides subyacente al epitelio pigmentario representan la fuente primaria de nutrición para los fotorreceptores retinianos, presumiblemente explica por qué bastones y conos se encuentran en la parte más externa y no en la capa más interna de la retina. En efecto, las interrupciones en esta relación normal entre el epitelio pigmentario y los fotorreceptores retinianos tiene graves consecuencias para la visión (Recuadro 11D, página precedente). Fototransducción En la mayoría de los sistemas sensitivos, la activación de un receptor por el estímulo apropiado causa la despolarización de la membrana celular, lo que finalmente estimula un potencial de acción y la liberación del transmisor en las neuronas con las que hace contacto. Sin embargo, en la retina los fotorreceptores no muestran potenciales de acción; más bien, la activación por la luz produce un cambio graduado en el potencial de membrana y una alteración correspondiente en la velocidad de liberación del transmisor en las neuronas postsinápticas. En efecto, gran parte del procesamiento en el interior de la retina está mediado por potenciales graduados, sobre todo porque los potenciales de acción no son necesarios para transmitir información sobre las distancias relativamente cortas involucradas. Tal vez sea más sorprendente saber que la luz que brilla sobre un fotorreceptor, sea un bastón o un cono, conduce la membrana a la hiperpolarización y no a la despolarización (Figura 11.7). En la oscuridad, el receptor se encuentra en estado despolarizado, con un potencial de membrana de aproximadamente –40 mV (que involucra las porciones de la célula que liberan transmisores). Los aumentos progresivos en la intensidad de la iluminación hacen que el potencial a través de la membrana del receptor se torne más negativo, respuesta que se satura cuando el potencial de la membrana alcanza unos –65 mV. Aunque el signo del cambio de potencial puede parecer extraño, el único requisito lógico para el procesamiento visual ulterior es una relación coherente entre los cambios de luminancia y la veloci- EL OJO (A) Retículo endoplasmático Mitocondria (B) Célula del epitelio pigmentario Pigmento 239 Los discos se desprenden y son fagocitados Segmento externo Aminoácidos marcados Segmento interno 0 3 6 9 Días después de la inyección del radiomarcador 12 (C) Los discos se enrollan Bastones dad de liberación del transmisor desde las terminaciones de los fotorreceptores. Como en otras células nerviosas, la liberación del transmisor desde las terminaciones sinápticas del fotorreceptor depende de los canales de Ca2+ sensibles al voltaje en la membrana del terminal. Por lo tanto, en la oscuridad, cuando los fotorreceptores están relativamente despolarizados, la cantidad de canales del Ca2+ abiertos en la terminación presináptica es alta, y le corresponde una velocidad de liberación del transmisor también grande; en la luz, cuando los receptores están hiperpolarizados, la cantidad de canales del Ca2+ abiertos está disminuida, al igual que la velocidad de liberación del transmisor. La razón para esta disposición inusual, comparada con otras células receptores sensitivas, se desconoce. En la oscuridad, fluyen cationes (tanto Na+ como Ca2+) en el segmento externo a través de los canales de membrana con puer- El extremo se torna esférico El extremo se separa del bastón El extremo es engullido por el epitelio pigmentario FIGURA 11.6 Eliminación de los discos de fotorreceptores por el epitelio pigmentario. (A) Las puntas de los segmentos externos de los fotorreceptores están introducidos en el epitelio pigmentario. Las prolongaciones de las células epiteliales se extienden hacia abajo entre los segmentos externos. (B) La duración de la vida de los discos de los fotorreceptores se observa en el movimiento de aminoácidos marcados con radiactividad e inyectados en el segmento interno e incorporados en los discos. Los discos marcados migran desde la porción interna a la porción externa del segmento externo en un período de 12 días. (C) Los discos gastados son eliminados del segmento externo y fagocitados. El fotopigmento proveniente de los discos entra en el epitelio pigmentario, donde sufrirá un ciclo bioquímico nuevamente a discos de fotorreceptores “recién nacidos”. (A, de Oyster, 1999; B, C, de Young, 1971.) Potencial de membrana (mV) Destello de luz Respuesta al destello menos intenso −40 FIGURA 11.7 Hiperpolarización de un fotorreceptor. Este −45 registro intracelular proviene de un único cono estimulado con diferentes cantidades de luz (el cono ha sido tomado de la retina −50 −55 −60 Respuesta al destello más intenso −65 0 100 200 300 Tiempo (ms) 400 500 600 de una tortuga, lo que explica el curso temporal relativamente largo de la respuesta). Cada trazado representa la respuesta a una luz breve cuya intensidad se hizo variar. En los niveles más altos de luz, la amplitud de la respuesta se satura (en unos -65 mV). La respuesta hiperpolarizante es característica de los fotorreceptores de los vertebrados. (De Schnapf y Baylor, 1987.) 240 CAPÍTULO 11 Na+ pero el mismo esquema se aplica a los conos. (A) En la oscuridad, las concentraciones de GMPc en la membrana del segmento externo son altas; el cGMP se une a canales permeables al Na+ en la membrana y los mantiene abiertos permitien- Segmento externo del bastón cGMP cGMP cGMP cGMP Ca2+ Na+ Disminución del cGMP do el ingreso de sodio y otros cationes, lo que despolariza la célula. (B) La absorción de fotones conduce a una disminución en las concentraciones de cGMP, lo que cierra los canales de cationes y conduce a la hiperpolarización de los K+ K+ 0 receptores. 0 Interior Segmento interno del bastón Ca2+ FIGURA 11.8 Canales con puerta de GMP cíclico y cambios inducidos por la luz en la actividad eléctrica de los fotorreceptores. Este esquema simplificado muestra un bastón, (B) Luz (A) Oscuridad Segmento externo del bastón Interior Exterior Entrada de Na+ Salida de K+ Despolarización Segmento interno del bastón ta del nucleótido monofosfato cíclico de guanosina (cGMP) de una forma similar a la de otros sistemas de segundos mensajeros (véase Capítulo 7). A esta corriente hacia el interior se le opone una corriente hacia el exterior mediada por los canales selectivos para el potasio en el segmento interno. Por lo tanto, el estado despolarizado del fotorreceptor en la oscuridad refleja la contribución del influjo de Na+ y de Ca2+, que actúa para despolarizar la célula y la salida de K+, que actúa para hiperpolarizar la célula (Figura 11.8A). La absorción de luz por el fotorreceptor reduce la concentración de cGMP en el segmento externo y conduce a su vez a un cierre de los canales con puerta de cGMP en la membrana del segmento externo y, en consecuencia, a una reducción en el flujo hacia el interior de Na+ y Ca2+. Como resultado, la carga positiva (transportada por el K+) fluye hacia el exterior de la célula más rápidamente de lo que lo hace la carga positiva (transportada por Na+ y Ca2+) hacia el interior y la célula se hiperpolariza (Figura 11.8B). La serie de cambios bioquímicos que por último conduce a una reducción en los niveles de cGMP comienza cuando un fotón es absorbido por el fotopigmento en los discos de los receptores. Este contiene un cromóforo absorbente de luz, el retinal (un aldehído de vitamina A) acoplado a una de varias proteínas posibles llamadas opsinas. Las diferentes opsinas afinan la absorción de luz de la molécula hacia una región particular del espectro; en efecto, es el componente proteico diferente del fotopigmento en los bastones y los conos que contribuye a la especialización funcional de estos dos tipos de receptores. La mayor parte de lo que se conoce acerca de los acontecimientos moleculares de la fototransducción se obtuvo de experimentos en bastones, en los cuales el fotopigmento es rodopsina. Los siete dominios transmembrana de la molécula de opsina atraviesan la membrana de los discos en el segmento externo y forman un bolsillo en el cual reside la molécula de retinal Exterior Disminución de la entrada de Na+ Salida de K+ Hiperpolarización (Figura 11.9A). Cuando el retinal de la rodopsina absorbe un fotón de luz, se rompe uno de los enlaces dobles entre los átomos de carbono de la molécula de retinal y su configuración cambia del isómero 11-cis a retinal todo-trans (Figura 11.9B); esta modificación desencadena entonces una serie de alteraciones en el componente proteico de la molécula. Por su parte, los cambios conducen a la activación de un mensajero intracelular llamado transducina, que activa una fosfodiesterasa que hidroliza cGMP. Todos estos acontecimientos tienen lugar dentro de la membrana del disco. La hidrólisis por la fosfodiesterasa en la membrana del disco reduce la concentración de cGMP en todo el segmento externo, y disminuye así la cantidad de moléculas de cGMP disponibles para unirse a los canales en la superficie de la membrana del segmento externo, lo que conduce a su vez al cierre del canal (Figura 11.9C). Una de las características importantes de esta cascada bioquímica compleja iniciada por la captura de fotones es que proporciona una amplificación enorme de las señales. Se ha estimado que una sola molécula de rodopsina activada por la luz puede activar 800 moléculas de transducina, aproximadamente el 8% de las moléculas de transducina sobre la superficie del disco. Aunque cada molécula de transducina solo activa una molécula de fosfodiesterasa, cada una de estas a su vez es capaz de catalizar la degradación de hasta seis moléculas de cGMP. En consecuencia, la absorción de un fotón único por una molécula de rodopsina conduce al cierre de alrededor de 200 canales iónicos o cerca del 2% de la cantidad de canales en cada bastón que están abiertos en la oscuridad. Esta cantidad de cierres de canales produce un cambio neto en el potencial de membrana de alrededor de 1 mV. Una vez iniciada, otros mecanismos limitan la duración de esta cascada amplificadora y restablecen las distintas moléculas a sus estados inactivados. La rodopsina activada es rápidamente fos- EL OJO (A) (B) C Luz 11 Retinal 11-cis N FIGURA 11.9 Fototransducción en los fotorreceptores de bastones. (A) La rodopsina reside en la membrana del disco del segmento externo de los fotorreceptores. Los siete dominios transmembrana de la molécula de opsina encierran 12 Opsina 241 la molécula de retinal sensible a la luz. (B) La absorción de un fotón de luz por el retinal conduce a un cambio en la configuración desde el 11-cis al isómero todo-trans. (C) Cascada de segundos mensajeros de la fototransducción. El cambio en el isómero retinal activa a la transducina, lo que a su vez activa a una fosfodiesterasa (PDE). Esta fosfodiesterasa hidroliza entonces al cGMP, lo que reduce su concentración en el segmento externo y conduce al cierre de los canales en la membrana del segmento externo. Retinal 11-cis 11 12 Retinal todo-trans (C) cantidad importante de moléculas activas de fotopigmento. GMP GMP La magnitud de esta amplificación varía GMP Na+ con los niveles prevalentes de iluminación, cGMP cGMP cGMP fenómeno conocido como adaptación a la luz. Con bajos niveles de iluminación, los Na+ fotorreceptores son los más sensibles a la Ca2+ luz. A medida que los niveles de iluminación aumentan, la sensibilidad disminuye, lo que impide que los receptores se saturen, Luz Disco Canal y así se incrementa mucho el intervalo de cerrado Rodopsina intensidades de luz sobre el cual operan. La Membrana del disco concentración de Ca2+ en el segmento exterTransducina γ no parece desempeñar un papel clave en la α β GTP α modulación de la sensibilidad de los fotoPDE rreceptores inducida por la luz. Los canales GTP GDP Interior Exterior con puerta de cGMP en el segmento externo de la célula de la célula son permeables tanto al Na+ como al Ca2+ (Figura 11.10B); por lo tanto, el cierre de forilada por la rodopsina cinasa, lo que permite que la proteína estos canales inducido por la luz conduce a una disminución arrestina se una a la rodopsina. La arrestina ligada bloquea la neta en la concentración interna de Ca2+. Esta reducción descapacidad de la rodopsina activada para activar la transducina, y encadena algunas modificaciones en la cascada de la fototransasí trunca efectivamente la cascada de fototransducción. ducción, todas las cuales tienden a reducir la sensibilidad del El restablecimiento del retinal a una forma capaz de señalar la receptor a la luz. Por ejemplo, la disminución en el Ca2+ aucaptura de fotones es un proceso complejo conocido como ciclo menta la actividad de la guanilato ciclasa, la enzima sintetizade retinoides (Figura 11.10A). El retinal todo-trans se disocia dora de cGMP, lo que conduce a un incremento en los niveles de la opsina, se difunde en el citoplasma del segmento externo, de cGMP. Asimismo, la disminución del Ca2+ incrementa la se convierte en retinol todo-trans y se transporta por el epite- actividad de la rodopsina cinasa, lo que permite que se una más lio pigmentario mediante una proteína chaperona, la proteína arrestina a la rodopsina. Por último, la disminución del Ca2+ fijadora de retinoides interreceptores (IRBP, interphotorre- aumenta la afinidad de los canales con puerta de cGMP por ceptor retinoid binding protein; véase Figura 11.10A), donde el cGMP, lo cual reduce el impacto de la disminución de los finalmente enzimas apropiadas lo convierten en retinal 11-cis. niveles de cGMP inducidos por la luz. Los efectos reguladores Una vez que se transporta de nuevo hacia el segmento externo del Ca2+ sobre la cascada de la fototransducción son solo una a través de la IRBP, el retinal 11-cis se recombina con opsina parte del mecanismo que adapta la sensibilidad del retinal a los en los discos receptores. El reciclaje de rodopsina tiene una im- niveles de iluminación de fondo; otra contribución importante portancia fundamental para mantener la sensibilidad a la luz de proviene de las interacciones neurales entre las células horilos fotorreceptores. Aun con niveles intensos de iluminación, la zontales y las terminaciones de los fotorreceptores (véase más velocidad de regeneración es suficiente como para mantener una adelante). Membrana del segmento externo Canal abierto 242 CAPÍTULO 11 (A) Retinol 11-cis Epitelio pigmentario Retinal 11-cis Éster de retinil todo-trans OH Retinal 11-cis FA Retinal todo-trans Retinol todo-trans Especialización funcional de los sistemas de bastones y conos Los dos tipos de fotorreceptores, bastones y conos, se distinguen por la forma (de la que Fotoisomerización deriva su nombre), el tipo de fotopigmento que contienen, su distribución a través de la retina y el patrón de conexiones sinápticas. OH Rh Rh* Estas propiedades reflejan la especialización OH Retinol de los sistemas de bastones y conos (los retodo-trans HO ceptores y sus conexiones en el interior de IRBP IRBP la retina) para diferentes aspectos de la visión. El sistema de los bastones tiene muy Luz baja resolución espacial, pero es en extremo sensible a la luz; por lo tanto, está especia(B) lizado en la sensibilidad a expensas de la resolución. Por el contrario, el sistema de Luz los conos tiene muy alta resolución espacial, pero es relativamente insensible a la luz; por lo tanto, está especializado en la agudeza a Membrana del segmento externo expensas de la sensibilidad. Las propiedades del sistema de los conos también permiten que los seres humanos y muchos otros animales vean el color. Rodopsina Disco En la Figura 11.11 se muestra el intervalo Transducina Membrana de iluminación sobre el que operan los basdel disco Ca2+ tones y los conos. Con los niveles más bajos cGMP Ca2+ PDE de iluminación, los bastones son los únicos cGMP receptores activados. Esta percepción meGMP Na+ diada por los bastones se denomina visión GuanilatoNa+ escotópica. La dificultad para efectuar disRodopsinciclasa Ca2+ cinasa criminaciones visuales finas en condiciones GTP de luz muy baja, donde solo está activo el Ca2+ sistema de los bastones, es una experiencia Rh+-P Interior Exterior frecuente. El problema es fundamentalmende la célula de la célula te la resolución escasa del sistema de los bastones (y, en menor medida, la pérdida FIGURA 11.10 Ciclo de los retinoides y fotoadaptación. de la percepción de color en la luz tenue, porque los conos no (A) Después de la fotoisomerización, el retinal todo-trans es participan en grado importante). Aunque los conos comienzan a convertido en retinol todo-trans y es transportado por la proteína contribuir con la percepción visual aproximadamente a la luz de chaperona IRBP al epitelio pigmentario. Allí, en una serie de pasos, es convertida en 11-cis retinal y transportado nuevamenuna estrella, la discriminación espacial con este nivel de luz aún te al segmento externo (otra vez a través de IRBP), donde se es muy baja. recombina con opsina. (B) Adaptación de los fotorreceptores. El A medida que aumenta la iluminación, los conos se vuelven calcio del segmento externo inhibe la actividad de la guanilato cada vez más dominantes para determinar lo que se ve y son ciclasa y la rodopsina cinasa, y reduce la afinidad de los canales el determinante principal de la percepción en condiciones tales con puerta de cGMP por el cGMP. El cierre inducido por la luz como la iluminación normal de interiores o la luz solar. La conde los canales en la membrana del segmento externo conduce tribución de los bastones a la visión desaparece casi por complea una reducción de la concentración de Ca2+ y a una reducción to en la denominada visión fotópica, dado que su respuesta a la en la inhibición de estos elementos de la cascada mediada por luz se satura, esto es, el potencial de membrana de los bastones Ca2+. En consecuencia, la sensibilidad de los fotorreceptores a la individuales ya no varía en función de la iluminación porque captura de fotones está reducida. todos los canales están cerrados (véase Figura 11.9). La visión mesópica se desarrolla con niveles de luz en los que contribuyen tanto los bastones como los conos (p. ej., al anochecer). A partir OH EL OJO Luz de una estrella Luz de luna Iluminación del interior 243 Luz solar Luminancia del papel blanco en: Sin visión de colores Agudeza deficiente Buena visión de colores Mejor agudeza Escotópica Función visual Mesópica Mesopic Umbral de los conos Umbral absoluto −6 Fotópica −4 Comienza la saturación de los bastones 2 0 Luminancia (log cd/m−2) −2 FIGURA 11.11 Gamas de valores de luminancia sobre las cuales opera el sistema visual. Con los niveles más bajos de iluminación, solo están activados los bastones. Los conos comienzan a contribuir con la percepción aproximadamente al nivel de la luz de una estrella y son los únicos receptores que funcionan en condiciones relativamente brillantes. de estas consideraciones, debería estar claro que la mayor parte de lo que consideramos la “vista” normal está mediado por el sistema de los conos, y que la pérdida de la función de los conos es devastadora, como la que se desarrolla en los individuos ancianos que sufren degeneración macular (véase Recuadro 11.C). Los individuos que perdieron la función de los conos son legalmente ciegos, mientras que los que perdieron la función de los bastones solo experimentan dificultad para ver con niveles bajos de iluminación (ceguera nocturna). Las diferencias en los mecanismos de transducción utilizados por los dos tipos de receptores son un factor importante en la capacidad de bastones y conos para responder a rangos diferentes (B) (A) Estímulo luminoso 15-30 bastones Corriente interior (pA) 0 Bastón Rod –10 –20 –30 Corriente interior (pA) Oftalmoscopia indirecta Mejor agudeza 4 Posible daño 6 8 de intensidad de luz. Por ejemplo, los bastones producen una respuesta fiable ante un solo fotón de luz, mientras que se necesitan más de 100 fotones para producir una respuesta comparable en un cono. Sin embargo, no es que los conos no capturen fotones eficazmente. Más bien, el cambio en la corriente producido por la captura de fotones únicos en los conos es comparativamente pequeño y difícil de distinguir del ruido de fondo. Otra diferencia es que la respuesta de un cono individual no se satura con niveles altos de iluminación constante, como lo hace la respuesta de los bastones. Aunque los bastones y los conos se adaptan para operar en un rango de valores de dominancia, los mecanismos de adaptación de los conos son más eficaces. Esta diferencia en la adaptación es aparente en el curso temporal de la respuesta de los bastones y los conos a destellos de luz. La respuesta de un cono, incluso a un destello de luz brillante que produce el cambio máximo en la corriente del fotorreceptor, se recupera en unos 200 milisegundos, más de cuatro veces más rápido que la recuperación de los bastones (Figura 11.12A). La disposición de los circuitos que transmiten la información hacia las células ganglionares retinianas también contribuye a las diferentes características de la visión escotópica y fotópica. En la mayor parte de la retina, las 1 célula bipolar de bastón FIGURA 11.12 Respuestas diferenciales de los conos y bastones de los primates. (A) Registros con electrodos de succión de la reducción de la corriente hacia el interior producida por destellos de intensidad de luz sucesivamente mayor. Con los destellos moderados a largos, la respuesta de los bastones continuó durante más de 600 ms, mientras que aún con los des- 1 cono tellos evaluados más brillantes, la respuesta de los conos retorna al basal (con una depresión exagerada) aproximadamente a los 0 0 50% de descoloración Cone Cono –10 200 ms. (B) Diferencia en la cantidad de convergencia en la vía –20 1 célula bipolar de cono 0 200 Tiempo (ms) 400 600 de bastones y conos. Cada célula bipolar de los bastones recibe sinapsis de 15-30 bastones. Ocurre convergencia adicional en localizaciones corriente abajo en la vía de los bastones (véase texto). Por el contrario, en el centro de la fóvea, cada célula bipolar recibe sus aferencias de un único cono y hace sinapsis con una única célula ganglionar. (A, de Baylor, 1987.) 244 CAPÍTULO 11 señales de bastones y conos convergen sobre las mismas células ganglionares; o sea, las células ganglionares individuales responden tanto a las aferencias de conos como de bastones, en función del nivel de iluminación. Sin embargo, las primeras etapas de las vías que conectan bastones y conos con las células ganglionares en su mayor parte son independientes. Por ejemplo, la vía desde los bastones hasta las células ganglionares comprende una clase distinta de célula bipolar que, al contrario de las células bipolares de los conos, no hace contacto con las células ganglionares retinianas. En cambio, las células bipolares de los bastones hacen sinapsis con las prolongaciones dendríticas de una clase específica de célula amacrina que forma uniones en hendidura y sinapsis químicas con las terminaciones de las células bipolares de los conos; a su vez, estas prolongaciones hacen contactos sinápticos sobre las dendritas de las células ganglionares en la capa plexiforme interna. En consecuencia, los circuitos que relacionan los bastones y los conos con las células ganglionares de la retina difieren espectacularmente en su grado de convergencia (Figura 11.12B). Cada célula bipolar de un bastón hace contacto con una célula amacrina dada. Por el con- trario, el sistema de los conos es mucho menos convergente. Así, cada célula ganglionar retiniana que domina la visión central (denominada célula ganglionar enana) recibe aferencias solo de una célula bipolar de un cono, que a su vez hace contacto con un solo cono. La convergencia convierte el sistema de los bastones en un mejor detector de la luz, porque señales pequeñas de muchos bastones se acumulan para generar una respuesta mayor en la célula bipolar. Al mismo tiempo, la convergencia reduce la resolución espacial del sistema de los bastones, dado que la fuente de una señal en una célula bipolar de un bastón o en la célula ganglionar retiniana podría haber provenido de cualquier sitio dentro de un área relativamente grande de la superficie retiniana. Por supuesto, la relación uno a uno entre conos y células bipolares y ganglionares es justo lo que se necesita para aumentar la agudeza al máximo. Distribución anatómica de bastones y conos La distribución de bastones y conos por la superficie de la retina también tiene consecuencias importantes para la visión. A pesar de que la percepción con los niveles típicos de luz diurna está dominada por la visión mediada por los conos, el número total de bastones en la retina humana (alrededor de 90 millones) excede en mucho el número de conos (aproximadamente 4,5 millones). En consecuencia, la densidad de los bastones es muy superior a la de los conos en la mayor parte de la retina (Figura 11.13A). Sin embargo, esta relación cambia de manera espectacular en la fóvea, una región altamente especializada de (A) con un pico agudo en el centro de la fóvea (la fovéola). Por el contrario, los bastones están presentes en alta densidad en la mayor parte de la retina, con un descenso brusca en la fóvea; los bastones están ausentes en la fovéola. Los cuadros muestran el aspecto frontal de los cortes a través de 160 140 120 100 Bastones 80 60 Papila óptica 3 Densidad de receptores (mm−2 × 10 ) FIGURA 11.13 Distribución de los fotorreceptores en la retina humana. (A) Los conos están presentes en baja densidad en toda la retina, los segmentos externos de los fotorreceptores en diferentes excentricidades. El aumento de densidad de los conos en la fóvea se acompaña por una reducción notable del diámetro de sus segmentos externos. Obsérvese también la ausencia de receptores en el disco óptico, donde los axones de las Bastones 40 20 0 80 60 Temporal Conos 40 Conos 20 0 20 40 Eccentricity Excentricidad (degrees) (grados) 60 80 Nasal células ganglionares se reúnen para abandonar la retina. (B) Esquema del corte transversal a través de la fóvea. Las capas celulares y los vasos sanguíneos suprayacentes están desplazados de modo que la luz está sometida a un mínimo de dispersión antes de que los fotones golpeen los segmentos externos de los conos en la fovéola. (B) Epitelio pigmentario Conos Capilares Bastones Células bipolares Coroides Capa nuclear externa Capa nuclear interna Capa de células ganglionares Zona avascular Fovéola Fóvea Células ganglionares EL OJO la retina central que mide cerca de 1,2 milímetros de diámetro (véase Figura 11.1). En la fóvea (que literalmente significa “pozo”), la densidad de los conos aumenta casi 200 veces y alcanza, en su centro, la densidad máxima de empaquetamiento de receptores en cualquier otro sitio de la retina. Esta densidad elevada se logra al disminuir el diámetro de los segmentos externos de los conos, de modo que los conos de la fóvea se asemejen en su aspecto a los bastones. El aumento de densidad de los conos en la fóvea se acompaña de una disminución brusca de la densidad de los bastones. De hecho, los 300 μm centrales de la fóvea, denominados la fovéola, están totalmente libres de bastones (Figura 11.13B). La densidad en extremo elevada de receptores de conos en la fóvea, y la relación uno a uno con las células bipolares y las células ganglionares retinianas (véase Figura 11.12), proporciona a este componente del sistema de los conos la capacidad de mediar una gran agudeza visual. A medida que disminuye la densidad de conos con la excentricidad y aumenta el grado de convergencia hacia las células ganglionares de la retina, se reduce mucho la agudeza. Esta disminuye en un 75%, a sólo 6 grados por fuera de la línea de la visión, lo que puede apreciarse con facilidad si se intenta leer las palabras de cualquier línea de esta página que se encuentran alejadas de aquellas que están en la línea de visión. La restricción de la visión de máxima agudeza a una región tan pequeña de la retina es la razón principal por la cual los seres humanos pasan tanto tiempo moviendo sus ojos (y sus cabezas) –para dirigir, en efecto, las fóveas de ambos ojos hacia los objetos de interés (véase Capítulo 20)–. También es la razón por la cual los trastornos que afectan el funcionamiento de la fóvea tienen efectos tan devastadores sobre la visión (véase Recuadro 11C). Por el contrario, la exclusión de los bastones de la fóvea y su presencia en alta densidad lejos de la fóvea explican por qué el umbral para detectar un estímulo luminoso es menor fuera de la región de visión central. Es más fácil observar un objeto de luz tenue (como una estrella débil) mirando un poco lejos de ella, así la luz estimula la región de la retina más rica en bastones (véase Figura 11.13A). Otro rasgo anatómico de la fóvea que contribuye a la agudeza superior del sistema de conos es el desplazamiento de las capas (A) Corta 100 Bastones Intermedia Absorbanci