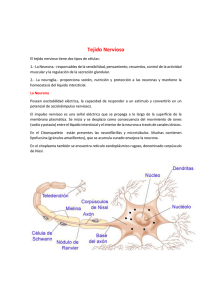
“AÑO DEL FORTALECIMIENTO DE LA SOBERANÍA NACIONAL” UNIVERSIDAD PRIVADA SAN JUAN BAUTISTA FACULTAD DE CIENCIAS DE LA SALUD ESCUELA PROFESIONAL DE MEDICINA HUMANA ASIGNATURA: Terapéutica y Cuidados Paliativos TEMA: Neurotransmisión del Sistema Nervioso Central DOCENTE: Gladys Yanet Diaz Hernández CICLO: V SEDE: Chincha ALUMNA: Melany Francesca Lazaro Berrocal El SNC está formado por varios tipos de células especializadas que se integran fisiológicamente para formar un tejido cerebral funcional complejo. La célula primaria de comunicación es la neurona, que está fuertemente influida y sostenida por una variedad de células de soporte importantes. Las conexiones específicas entre las neuronas, tanto dentro como a través de las macrodivisiones del cerebro, son esenciales para la función neurológica. Mediante patrones de circuitos neuronales, las neuronas individuales forman conjuntos funcionales para regular el flujo de información dentro y entre las regiones del cerebro. Bajo estas pautas, la comprensión actual de la organización celular del SNC se puede ver desde la perspectiva del tamaño, la forma, la ubicación y las interconexiones entre las neuronas NEURONAS: Las neuronas son las células funcionales del sistema nervioso. Las neuronas aferentes (o sensitivas) transmiten información al SNC, mientras que las neuronas eferentes (o motoras) envían la información que sale del SNC. Intercaladas entre las neuronas aferentes y eferentes se halla una red de neuronas interconectadas (llamadas también interneuronas o neuronas intercaladas) que modulan y controlan la respuesta del cuerpo a los estímulos sensitivos tanto de entornos internos como externos. Las neuronas constan de 3 partes: el cuerpo celular, las dendritas y los axones. Estas estructuras forman las conexiones funcionales, o sinapsis, con otras células nerviosas, con células receptoras o con células efectoras. Los procesos axonales están diseñados especialmente para lograr una rápida comunicación con otras neuronas y con las muchas estructuras corporales enervadas por el sistema nervioso. El cuerpo celular (o soma) de una neurona contiene un núcleo vesicular grande con 1 o más núcleos distintos y un retículo endoplásmico rugoso bien desarrollado. El núcleo de una neurona tiene el mismo contenido de ácido desoxirribonucleico (ADN) y código genético que el que se encuentra en otras células del cuerpo. Su nucléolo, que está compuesto por partes de diversos cromosomas, produce ácido ribonucleico (ARN) necesario para la síntesis proteica. El citoplasma contiene grandes masas de ribosomas que son prominentes en la mayoría de las neuronas. Estas masas ácidas de ARN, que participan en la síntesis de proteínas, tiñen de oscuro los cuerpos de Nissl con tintes histológicos básicos Las dendritas son múltiples ramificaciones cortas del cuerpo de la célula nerviosa. Transmiten la información hacia el cuerpo celular y son la fuente principal de información para la neurona. Las dendritas y el cuerpo celular están repletos de terminales sinápticas que se comunican con los axones y las dendritas de otras neuronas. Los axones son largas prolongaciones eferentes que salen del cuerpo celular. La mayoría de las neuronas sólo tienen un axón. Sin embargo, los axones pueden exhibir múltiples ramificaciones que dan como resultado muchas terminales axónicas. El axón de una neurona conduce los impulsos nerviosos desde el cuerpo celular hasta su sinapsis. También proporciona un conducto físico para el transporte de materiales entre el cuerpo celular y las terminales sinápticas del axón. El cuerpo celular de la neurona está preparado para llevar a cabo un alto nivel de actividad metabólica. Esto es necesario porque el cuerpo celular debe sintetizar los elementos citoplásmicos y membranales que se necesitan para mantener la función del cuerpo celular más las muchas proteínas y otros materiales citoplásmicos que el axón y sus terminales sinápticas utilizan CÉLULAS NEUROGLIALES: Las células neurogliales separan a las neuronas en compartimentos metabólicos aislados, lo que es necesario para la función neuronal normal. Algunos tipos de células neurogliales (los astrocitos) ayudan a formar la barrera hematoencefálica que impide que los materiales tóxicos que se encuentran en la sangre entren al encéfalo. Dos tipos de células neurogliales (los oligodendrocitos en el SNC y las células de Schwann en el SNP) producen la mielina que se emplea para aislar los procesos celulares nerviosos y aumentar la velocidad de los impulsos nerviosos. La mielina tiene un alto contenido en lípidos, lo cual le da un color blanquecino y el nombre de sustancia blanca que se le da a las masas de fibras mielinizadas de la médula espinal y el encéfalo. Además de su función para aumentar la velocidad de conducción, la vaina de mielina es esencial para la supervivencia de los procesos neuronales más grandes, quizá por la secreción de los compuestos neurotróficos. Las células neurogliales del SNC están compuestas de oligodendrocitos, astrocitos, microglías y células ependimarias. Los oligodendrocitos forman la mielina del SNC. En lugar de formar una cubierta de mielina para un único axón, estas células llevan a cabo diversos procesos, enrollándose y formando un segmento multicapa de mielina alrededor de varios axones diferentes. Los astrocitos, las células neurogliales más numerosas, son particularmete prominentes en la sustancia gris del SNC. Estas células grandes tienen muchos procesos, algunas llegando a la superficie de los capilares, otras a la superficie de las células nerviosas y otras más que llenan la mayor parte del espacio intercelular dentro del SNC. Los astrocitos mantienen un vínculo importante entre las neuronas (especialmente entre la sinapsis) y el flujo sanguíneo capilar. También ayudan a mantener la concentración correcta de iones de potasio en el espacio extracelular que hay entre las neuronas. Un tercer tipo de célula neuroglial, la microglía, es una pequeña célula fagocítica que aparece para limpiar los desechos que quedan después de un daño o infección celular, o una muerte celular. El cuarto tipo de célula, la célula ependimaria, forma el revestimiento de la cavidad del tubo neural, esto es, el sistema ventricular. En algunas zonas, estas células se combinan con una rica red vascular para formar el plexo coroide, donde tiene lugar la producción del líquido cefalorraquídeo (LCR). Las células satélites y las de Schwann son los 2 tipos de células neurogliales del SNP. Las células satélites son células capsulares planas que segregan una membrana de fondo que protege al cuerpo celular de la difusión de moléculas grandes. Una única capa de células satélite separa cada uno de los cuerpos celulares, y los procesos de los nervios periféricos se separan del marco de tejido conectivo del ganglio. Las células de Schwann son parientes cercanos de las células satélite. La membrana celular y el citoplasma de las células de Schwann rodean los procesos de las neuronas aferentes y eferentes más grandes. Las células de Schwann sucesivas están separadas por pequeñas interrupciones de líquido extracelular, llamadas nódulos de Ranvier, donde falta la mielina y se concentran canales de sodio dependientes del voltaje . Los nódulos de Ranvier aumentan la conducción nerviosa ya que permiten que el impulso salte de un nódulo a otro a través del líquido extracelular en un proceso llamado conducción saltatoria. BARRERA HEMATOENCEFÁLICA: La barrera hematoencefálica (BHE) es una barrera de transporte estructural y bioquímica, altamente especializada, que consta de vasos formados por células endoteliales, conectadas por uniones estrechas y rodeadas por los pies de los astrocitos. La BHE impide el paso de ciertas sustancias al intersticio cerebral con criterio selectivo. Muchos fármacos son útiles en el tratamiento de trastornos sistémicos pero resultan ineficaces con respecto a trastornos similares del sistema nervioso central (SNC), debido a su incapacidad para cruzar la barrera: los neuropéptidos, las proteínas y los antineoplásicos son ejemplos importantes de agentes terapéuticos. La BHE consta de células endoteliales capilares, aunque otras células, como los pericitos, astrocitos y células neuronales, también desempeñan un papel importante en la diferenciación y el mantenimiento de la función de la BHE. POTENCIAL DE ACCIÓN: Las señales nerviosas se transmiten por los potenciales de acción, que son cambios abruptos y por pulsos en el potencial de la membrana que duran 5 mseg aproximadamente. Las membranas celulares del tejido excitable, incluyendo las de las células nerviosas y musculares, contienen canales de iones que son responsables de generar estos potenciales de acción. Los controles dependientes del voltaje, que se abren y cierran cuando el potencial de la membrana cambia, controlan los canales de iones de la membrana Los potenciales de acción se pueden dividir en 3 etapas: el estado de reposo o polarización, despolarización y repolarización • POTENCIAL DE REPOSO DE LA MEMBRANA: El potencial de reposo de la membrana (de aproximadamente −70 mV para las fibras nerviosas grandes) es el período tranquilo del potencial de acción durante el cual el nervio no transmite impulsos. Durante este período, se dice que la membrana está polarizada debido a la gran separación de la carga (es decir, positiva en el exterior y negativa en el interior). La fase de reposo del potencial de membrana continúa hasta que algún suceso hace que esta aumente su permeabilidad al sodio. Un potencial umbral (de aproximadamente −55 mV en las fibras nerviosas grandes) representa el potencial de membrana en el que las neuronas u otros tejidos excitables se estimulan para lanzar una señal6 . Cuando se alcanza el potencial umbral, las estructuras en forma de compuertas de los canales de iones se abren. Por debajo del potencial umbral, estas compuertas permanecen firmemente cerradas. Las compuertas funcionan en una base de todo o nada, es decir, están totalmente abiertas o totalmente cerradas. En circunstancias normales, el estímulo umbral es suficiente para abrir muchos canales de iones, poniendo en marcha la desporalización masiva de la membrana (el potencial de acción). • DESPOLARIZACIÓN: La despolarización se caracteriza por el flujo de iones cargados eléctricamente. Durante la fase de despolarización, la membrana se vuelve de repente permeable a los iones de sodio. La rápida entrada de iones de sodio produce corrientes locales que viajan a través de la membrana adyacente, haciendo que los canales de sodio de esta parte de la membrana se abran. En las neuronas, las compuertas de booksmedicos.org iones de sodio permanecen abiertas durante aproximadamente un cuarto de milisegundo. Durante esta etapa del potencial de acción, la cara interna de la membrana se hace positiva (aproximadamente +30 mV). • REPOLARIZACIÓN: La repolarización es la etapa durante la cual se restablece la polaridad del potencial en reposo de la membrana. Esto se logra con el cierre de los canales de sodio y la apertura de los canales de potasio. La salida de iones de potasio cargados positivamente a través de la membrana celular devuelve el potencial de membrana a la negatividad. La bomba sodio-potasio trifosfatasa de adenosina (Na+ /K+ −ATPasa) restablece gradualmente las concentraciones iónicas de reposo en cada lado de la membrana. Las membranas de las células excitables deben repolarizarse lo suficiente antes de que puedan volver a excitarse. Durante la repolarización, la membrana sigue siendo refractaria hasta que la repolarización esté aproximadamente a un tercio de completarse. Este período, que dura de 0,4 mseg a 4 mseg, se llama período refractorio absoluto. Durante una parte del período de recuperación, la membrana se puede excitar, aunque sólo por un estímulo más fuerte de lo normal. Este período se llama el período refractario relativo. TRANSMISIÓN SINAPTICA: Las neuronas se comunican entre sí mediante estructuras llamadas sinapsis. En el sistema nervioso hay 2 tipos de sinapsis: eléctricas y químicas. Las sinapsis eléctricas permiten el paso de iones portadores de corriente a través de pequeñas aperturas llamadas uniones comunicantes que penetran la unión celular de células adyacentes y permiten que la corriente viaje en cualquier dirección. Las uniones comunicantes permiten que un potencial de acción pase directa y rápidamente de una neurona a otra. Pueden enlazar neuronas que tengan relaciones funcionales cercanas formando circuitos. El tipo más común es la sinapsis química. Las sinapsis químicas implican estructuras de membrana presinápticas y postsinápticas especiales, separadas por una hendidura sináptica. La terminal presináptica segrega una molécula transmisora química y a menudo varias. Los neurotransmisores segregados se difunden en la hendidura sináptica y se unen a los receptores de la membrana postsináptica. A diferencia de la sinapsis eléctrica, la química sirve como rectificador, permitiendo la comunicación en un sólo sentido. Las sinapsis químicas se dividen en 2 tipos: excitatorias e inhibitorias. En las sinapsis excitatorias, la unión del neurotransmisor con el receptor produce la despolarización de la membrana postsináptica. La unión del neurotransmisor con el receptor en una sinapsis inhibitoria reduce la capacidad de la neurona postsináptica de generar un potencial de acción. La mayor parte de los neurotransmisores inhibitorios inducen la hiperpolarización de la membrana postsináptica haciendo que esta sea más permeable al potasio, al cloruro o a ambos TRANSMISORES SINÁPTICOS: En más de 50 sustancias químicas se ha comprobado o se ha propuesto su acción como transmisores sinápticos. Existen dos grupos de transmisores sinápticos. Uno comprende transmisores de acción rápida y molécula pequeña. El otro está configurado por un gran número de neuropéptidos con un tamaño molecular muy superior y que normalmente presentan una acción mucho más lenta • Transmisores de acción rápida y molécula pequeña: En la mayoría de los casos, los tipos de transmisores de molécula pequeña se sintetizan en el citoplasma del terminal presináptico y las numerosas vesículas transmisoras presentes a este nivel los absorben por transporte activo. Acetilcolina: Se segrega por las neuronas situadas en muchas regiones del sistema nervioso, pero específicamente en: 1 ) los terminales de las células piramidales grandes de la corteza motora; 2 ) diversos tipos diferentes de neuronas pertenecientes a los ganglios basales; 3) las motoneuronas que inervan los músculos esqueléticos; 4) las neuronas preganglionares del sistema nervioso autónomo; 5) las neuronas posganglionares del sistema nervioso parasimpático, y 6) parte de las neuronas posganglionares del sistema nervioso simpático. En la mayoría de los casos, la acetilcolina posee un efecto excitador; sin embargo, se sabe que ejerce acciones inhibidoras. En algunas terminaciones nerviosas parasimpáticas periféricas, como la inhibición del corazón a cargo de los nervios vagos. Noradrenalina: Se segrega en los terminales de muchas neuronas cuyos somas están situados en el tronco del encéfalo y el hipotálamo. En concreto, las que están localizadas en e] locus ceruleus de la protuberancia envían fibras nerviosas a amplias regiones del encéfalo que sirven para controlar la actividad global y el estado mental, como por ejemplo aumentar el nivel de vigilia. En la mayoría de estas zonas, la noradrenalina probablemente activa receptores excitadores, pero en unas cuantas, en cambio, estimula los inhibidores. También se segrega en la mayor parte de las neuronas posganglionares del sistema nervioso simpático, donde excita algunos órganos, pero inhibe otros. Dopamina: Se segrega en las neuronas originadas en la sustancia negra. Su terminación se produce básicamente en la región estriada de los ganglios basales. El efecto que ejerce suele ser una inhibición. Glicina: Se segrega sobre todo en las sinapsis de la médula espinal. Se cree que siempre actúa como un transmisor inhibidor. GABA: Se segrega en los terminales nerviosos de la médula espinal, el cerebelo, los ganglios basales y muchas áreas de la corteza. Se piensa que siempre causa una inhibición. Glutamato: Se segrega en los terminales presinápticos de muchas de las vías sensitivas que penetran en el sistema nervioso central, lo mismo que en muchas áreas de la corteza cerebral. Probablemente siempre causa excitación. Serotonina: Se segrega en los núcleos originados en el rafe medio del tronco del encéfalo que proyectan hacia numerosas regiones del cerebro y de la médula espinal, especialmente a las astas dorsales de la médula y al hipotálamo. Actúa en la médula como un inhibidor de las vías del dolor, y se piensa que la acción inhibidora sobre las regiones superiores del sistema nervioso ayuda a controlar el estado de ánimo de una persona, tal vez incluso provocando sueño Óxido Nítrico: Se segrega especialmente en los terminales nerviosos de las regiones encefálicas responsables de la conducta a largo plazo y de la memoria. Por tanto, este sistema transmisor podría esclarecer en el futuro algunas de las funciones correspondientes a estos dos aspectos que hasta ahora han desafiado toda explicación. El óxido nítrico difiere de otros transmisores de molécula pequeña por su mecanismo de producción en el terminal presináptico y por sus acciones sobre la neurona postsináptica. No está formado con antelación y almacenado en vesículas dentro del terminal presináptico como los demás transmisores. En su lugar, se sintetiza casi al instantesegún las necesidades, durante un período de segundos en vez de ser liberado en paquetes vesiculares, y después hacia las neuronas postsinápticas cercanas. En ellas, no suele alterar mucho el potencial de membrana, sino que modifica las funciones metabólicas intracelulares que cambian la excitabilidad neuronal durante segundos, minutos o tal vez incluso más tiempo. • Los neuropéptidos se sintetizan de otro modo y tienen acciones que normalmente son lentas y en otros aspectos bastante diferentes de las que ejercen los transmisores de molécula pequeña. Estas sustancias no se sintetizan en el citoplasma de los terminales presinápticos. Por el contrario, se forman en los ribosomas del soma neuronal ya como porciones íntegras de grandes moléculas proteicas Sustancia P: Es un Neuro-transmisor implicado en el aumento de la respuesta inflamatoria y la sensibilización nociceptiva. Es decir, a nivel periférico actúa provocando: -Vasodilatación -Un aumento de la permeabilidad -La activación fagocitaria de neutrófilos y macrófagos (Sistema Inmune). -La Liberación de mediadores inflamatorios, como por ejemplo un aumento de la histamina. REVISIÓN BIBILIOGRÁFICA • Loch-Neckel G, Koepp J. La barrera hematoencefálica y la administración de medicamentos en el sistema nervioso central. Rev Neurol 2010;51 (03):165-174 • Grossman, S., & Porth, C. M. (2014). Port Fisiopatología: Alteraciones de la salud. Conceptos básicos (9a. ed. --.). Barcelona: Wolters Kluwer. • Brunton, L. L.; Chabner, B. A.; Knollmann, B. C. Goodman & Gilman. Bases farmacológicas de la terapéutica. (2012) México: McGraw-Hill Interamericana. • Hall, J. E., & Guyton, A. C. (2016). Guyton y Hall: Compendio de fisiología médica (13a ed. --.)