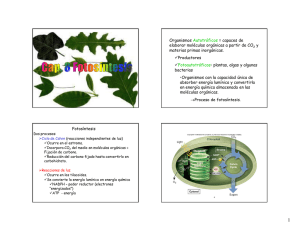
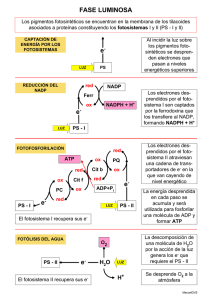
Fotosíntesi s Etimología: proviene del griego: foto=luz y síntesis=producción o ensamblaje Concepto: es uno de los procesos más claros de anabolismo autótrofo, porque apartar de elementos inorgánicos-agua y dióxido de carbono-se sintetiza glucosa, se produce oxígeno y cierta cantidad de ATP, mediante el uso de la energía luminosa. Este proceso es propio de las plantas, las algas y algunas bacterias que poseen moléculas de clorofila, pigmento verde que se encargadecapturarlaenergíasolaralmismotiempoquelesdauncolorcaracterístico. Organeroscitoplasmáticosencargados de realizarla fotosíntesis: Cloroplasto: Son los cloroplastos son los organeros en los cuales se produce el proceso de fotosíntesis. Tienen forma de disco que pueden existir hasta cincuenta por célula .asi, en un milímetro cuadradodehojapuedehaberhastaunoscincomilcloroplastos. La estructura de los cloroplastos consta de dos membranas que rodean un espacio interno llamado estroma. Membrana externa lisa: es la que rodea completamente al organero, aislándole del resto del citoplasma. Membrana interna: se pliega barias beses hacia el espacio interior generando un elaborado sistema de membranas en forma de sacos aplanados, circulares, semejantes a monedas llamados tilacoides. La membrana tilacoide contiene todos los pigmentos principalmente la clorofila y encimas de la fase luminosa de la fotosíntesis. Cada conjunto de tilacoides apilados se conoce como grana. El estroma, es una matriz acuosa que contiene proteínas ADN ARN y pequeños ribosomas responsables de la síntesis de proteínas dentro del cloroplasto. Aquí también se encuentra la mayorpartedelasenzimasdelafaseoscuradelafotosíntesis. Formas de fotosíntesis I. Anoxigenica: - NohayliberacióndeO2sinomásbienazufre. - Presenta como pigmento a la bacterioclorofila. ufáres.delasbacterias -E Alcudrardeoerndebaecletecrtiraosnepsurepsurgaesnseuralflumroesnatesuynnocosmup lfureosstaosd,eadaezm verdes. CO2 + H2S + EL C6 H12 O6 + S II. Oxigenica: - En los vegetales el lugar exclusivo de la fotosíntesis son las hojas. una hoja obtiene co2 para la fotosíntesis a partir del aire; poros ajustables en la epidermis, llamados estomas,seabrenysecierranenelmomentoadecuadoparaadmitiralCO2. - En el interior de la hoja hay unascapas de células queen cojunto, reciben elnombre de mesofilo(que significa “centro de la hoja”). - Las células del mesofilo contienen la mayor parte de los cloroplastos de las hojas en consecuencia la fotosintesis sucede principalmente en estas células. La fotosintesis se resumeenla siguientefotosintesis: CLOROFILA 6H2O +6CO2+LUZSOLAR C6H12O6+6O2 Donde el H2O, CO2, LUZ SOLAR Y LA CLOROFILA son los elementos que participan. En tantolaglucosayO2sonelproductodelproceso. FASES DE LA FOTOSINTESIS A.FASE LUMINOSA: Es la primera etapa o fase de la fotosíntesis, que depende directamente de la luz o energía luminosa para poder obtener energía química en forma de ATP y NADPH, a partir de la disociación de moléculas de agua, formando oxígeno e hidrógeno.1 La energía creada en esta fase,seráutilizadadurantelafaseoscura,paradeestaformacontinuarconlafotosíntesis. Conversión de luz a energía química. -Llamada fase fotodependiente, reacciones de Hill. -Lasreaccionesquedependendelaluzconviertenlaenergíadeluzsolarenenergíaquímica de dos moléculastransportadorasdiferentes: Una molécula transportadora llamada ATP y el transportador de electrones (y protones) +) dinucleótidodeniacina– adenina– fosfatooxidado(NADP . El primer paso de la conversión de la energía luminosa en energía química es la absorción de laluz. ¿QUIEN ABSORBE LA LUZ? Un pigmento es cualquier sustancia que absorbe luz visible. El sol emite energía en un amplio espectro de radiación electromagnética que va desde rayos gama de longitud corta pasando por luz ultravioleta visible y por la luz infrarroja hasta ondas de radio de longitud de onda larga. Como se sabe, la luz y otro tipo de radiación están compuestos de paquetes individuales de energía llamados fotones. La energía de un fotón corresponde a su longitud deonda:losfotonesdelongituddeondacortasonenergéticos,mientrasquelosfotonesde longituddeondalargatienenmenorenergía. Cuando laluzchocaconunobjetodigamosunahoja,puedensucedertresprocesos: 1. Laluzpuede absorberse. 2. Reflejarse(emitir nuevamente). 3. Transmitirse (pasar a través de). La luz que es absorbida puede calentar el objeto o intervenir en proceso biológico (fotosíntesis).Laluzqueesreflejadotransmitidasleda coloralosobjetos. Los fotopigmentos Son sustancias capasesde absorber la luz.Destacan: 1. La clorofila: existen bastantes tipo de clorofilas a, b, c, las cuales difieren entre si solamente en algunos detalles de su estructura molecular. la clorofila “a”, se haya en todos los eucariotas fotosintetisador y en la cianobacterias y se consideran que es esencial para el tipo de fotosíntesis que llevan a cabo los organismos de estos grupos. 2. Los pigmentos accesorios: integrados a las membranas y asociados con proteínas constituyen unidades fotosintéticas llamada cuantosomas. Estos se localizanenlostilacoidesdeloscloroplastos. Cada cuantosoma cuenta con dos fotosistemas I y II. El fotosistemaconstaasuvezdedospartes: a) Un complejo conector de luz, consta de cerca de trecientas moléculas de clorofila y pigmentos accesorios. La clorofila es la encargada de absorber la energía solar (pigmento antena), la clorofila “b”, “c” y los carotenos pasan esta energía hacia una molécula especifica de clorofila llamada centro de reacción. b) Un sistema de transporte de electrones, son una serie de moléculas portadoras de electrones que están metidas en la membrana del tilacoides ahí tenemos: PQ (PLASTOQUINONA), CIT b/f (citocromo b f ),pc (plastocianina) y Fd (ferrodoxina). ¿Cómo funcionan estos fotosistemas? Haciendo una analogía con la recepción de la imagen de TV, sucede lo siguiente: la energía luminosa absorbida por un pigmento antena en cualquier punto del entramado, es transferida de una a otra molécula de pigmento hasta alcanzar el centro de reacción, que está constituido por una forma especial de clorofila “a”. Cuando esto sucede la clorofila absorbe la energía, uno de sus electrones es lanzado a un nivel energético superior y transferido a una molécula receptora para iniciar el flujo de electrones. Así la molécula de clorofila se oxida y queda cargadapositivamente.Enla actualidadseconocendostiposdiferentesdefotosistemas: ➢ , F Po 70to0s,idsotenm deaPI, eesl eclepnitgromdeentoreyacecl iósunbeísnduincea 7fo0r0mdaedsiegncaloeronfilnaa“naó”m deetrnoosmeilnpaidca o de absorción óptima. Fotosistema II, elcentrodereacciónesunaformaespecialde clorofila “a”, pero supicodeabsorción es680nm,llamándoseP680. PASOS DE LA FASE LUMINOSA Comomencionamoselprimerpasoesla absorción de la luz,elpasosiguientees: ❖ ❖ ❖ ❖ ❖ FOTOEXCITACIÓN: cuando un fotón excita a una de las numerosas moléculas de clorofila antena del FSII, uno de los electrones de esta sale orbita molecular y su energíastransferidaalaclorofilaP680localizadaenelcentrodereacción. El nuevo electrón energizado abandona el FSII y pasa al PQ (plastoquinona) que es el primereslabóndelcentrodereacciones. Por otro lado, en el espacio tilacoidal, por un proceso de fotolisis de agua, dos moléculas de H2O se rompen y generan cuatro hidrógenos, cuatro electrones y un molécula de O2 que va a ser liberada al medio ambiente cada uno de estos electrones pasa al centro de reacción del FSII y reemplaza al salido de la P680 transferido a la PQ (plastoquinona).Esdecirelaguaeseldonadorfinaldelectrones. El electrón pasa de la plastoquinona al complejo b-f donde parte de su energía se utiliza para transportar un hidrogeno hacia el espacio tilacoidal en contra de la gradiente, parta que se una a los otros hidrógenos generados por la fotosíntesis del agua. El electrón ahora con menos potencial energético pasa del complejo b-f a la PC (plastocianina)yde estaalFSI. lEanaenlteFnSaIyotsrouefonteórngcía petsurtradnosfexrcidita a unoade loas enluem cteroronseassdm eolaléclou rlaosfildaePc7lo0r0o.fila de ❖ ❖ ❖ El electrón excitado es trasferido a la FD (ferrodoccina) y es remplazado por el electróndebajopotencialprovenientedelaplastocianina. El electrón que paso del P700 a la FD ingresa a la NADP reductasa donde parte de su energía es utilizada para reducir (foto reducción) un NADP+ a NADPH en la cara de la membrana del tilacoide que da al estroma del cloroplasto. En este proceso se utiliza un H+ tomado del estroma. El último paso de las reacciones fotoquímicas corresponde a la fotofosforilación, es deciralaformacióndeATPapartirdeADPyfosfato.EstoocurreenlaATPsintetasa que por su porción F permite el traslado pasivo de los H + desde el espacio tilaco+idal hacia el estroma. Durante este pasaje la energía promotora contenida en los H es cedida a la porción F de la ATP sintetasa que la utiliza para sintetizar ATP (quimiosmosis). FASES OSCURAS Conversión de dióxido de carbono en glucosa llamada reacción termoquímica o ciclo de Calvin-Benson. ¿Cuál es exactamente la manera en el que el CO2 se fija, es decir, como se convierte estas moléculasenazucares,almidón,celulosaytodoslosdemásproductosdel metabolismo. Fotosíntesis C3: ciclo de Calvin. Los primeros experimentos en este campo los realizaron Melvin Calvin, Andrew Benson y sus colegas de la universidad de california. Se llama del C3 porque algunas de las moléculas importantes en el ciclo tienen tres átomos de carbono. Este ciclo necesita los siguientes componentes: 1. CO2(que seobtiene generalmentede aire) 2. Unazúcarque captadióxidodecarbono, laribulosa bifosfato (BPRu) 3. Enzimas quecatalizan todas lasreacciones 4. Energía enforma deATPyNADPH,queproviene delasreacciones luminosas Elciclosehadivididosentrespartes: 1. Fijación del dióxidode carbono, lavía C3seinicia ytermina con azúcar decinco carbonos, el bifosfato de ribulosa (BPRu). El BPRu se combina con el dióxido de carbono atmosférico gracias a la enzima ribulosa difosfato carboxilasa (rubisco), para formar un compuesto de seis átomos de carbono muy inestable. Este compuesto reacciona espontáneamente con el agua para formar dos moléculas y tres carbonos de ácido fosfoglicérido (PGA) o ácido carboxílico de ahí el nombre de vía del carbono C3, como se ve el dióxido de carbono se fija en un compuesto orgánico más o menos estable. El PGA no es el producto más importantequeresultadelafijacióndeCO2 2. Síntesis de fosfogliceraldehido, la energía en forma de APT y NADPH se emplea en la formación de PGA en una molécula que aunque está estrechamente relacionada con ella, es más energética. Esta molécula se llama fosgliceraldehido (PGAL) y como se trata de un azúcar fosfatado de tres carbonostambiénseconocecomo triosafosfato. 3. R reeagcecnioenraecsióqnuedereqbuisieforesfnateonedregíala dreibl uAloTsPa, dioalnétceulaun eAcLom de) 1m 0 em s adesePriG (1p0lexja3C pueden regenerar las seis moléculas de ribulosa bifosfato utilizados al inicio de la fijación del carbono. Todo ciclo de Calvin Benson se lleva a cabo en el estroma delos cloroplastos. Como ya lo hemos mencionado el ciclo del C3 se inicia con la ribulosa bifosfato, se agrega Carbonos a partir de CO2, termina en un ciclo con ribulosa bifosfato otra vez, por la que hay carbono en exceso a partir de CO2 captado. Al utilizar la “cuenta del número de carbono” speagraúnfosrm eainriucinaymtoelrémciunla duenaglvuuceolstad(6eCl)c.iclo con seis moléculas de PGAL (3C) se combina Posteriormente la glucosa puede desdoblarse durante la respiración celular o unirse en cadenas para formar almidón (moléculas de almacenamiento) o Celulosa (el componente principal de las paredes celulares), o modificarse en aminoácidos, lípidos u otros constituyentescelulares. El almidón suele almacenarse en los mismos cloroplastos, pero algunos atraviesan las membranasdeloscloroplastosysealmacenanenlosamiloplastos. Allá por el año 1961 Calvin, gano el premio nobel en química, (único premio nobel otorgado por trabajosdefisiologíasenplantasverdes)elcicloensuhonorsedenominóciclodeCalvin. Faseluminosa Faseoscura Fotólisis delagua FijacióndelCO2 ProduccióndelATPyNADPH2 Consumo del ATPyNADPH2 Sellamaacabosóloenpresencia deluz Noesnecesario lapresencia deluz Tienelugarenlostilacoidesdelcloroplasto Tienelugarenelestromadelcloroplasto Fotosíntesis en bacterias ( Fotosíntesis Anoxigénica) Como hemos mencionado, las cianobacterias antes llamadas algas azul verdosas realizan la misma fotosíntesis que las plantas verdes, sin embargo otro grupo de bacterias como las púrpuras y las verdes su fotosíntesis es de tipo anoxigénica es decir no utilizan agua como dador de electrones y como consecuencia no pueden formar oxígeno. Asimismo presentan las bacterioclorofila en vez de clorofila y además que se encuentran dispuestas en empalizadasdentro delcitoplasma. La fuente de electrones es alguna sustancia reducida del ambiente, por ejemplo, compuestos de azufre reducido( H2S, azufre elemental, tiosulfato, H2 ) o compuestos orgánicos( succinato, malato, butirato) fórmula : 6CO2 +12H2S glucosa + 6H20+ 12S El azufre que se forma se deposita, ya sea fuera de las células en las bacterias autótrofas verdes, yaseadentrodelascélulasenlasbacteriaspúrpuras. Cuadro comparativo entre la fotosíntesis oxigénica y anoxigénica Fotosíntesis Oxigénica Desprende oxígeno( a partir del agua) Pigmento clorofila Eldador deelectrones es elagua Sedaluminosa enplantas,de algas cianobacteriasIII La Fase layfotosíntesis Cadenas de transporte electrónico fotosintético ✓ Fotosíntesis Anoxigénica No desprende oxígeno, libera azufre Pigmento Bacterioclorofila Eldadordeelectronesesgeneralmenteun compuesto de azufre. Sedaenbacteria s púrpuras sulfurosas y nono sulfurosas y bacterias verdes. Las bacterias “púrpuras”. Estas bacterias tienen un fotosistema del tipo PSII. El “par especial” está formado por una molécula de bacterioclorofila a y otra de bacterioclorofila b, y presenta un pico de absorción a 870 nm, en la zona del rojo, por lo que también se denomina P870. Los electrones abandonan el fotosistema, de dos en dos, transportados por una molécula de quinona que los cede a una bomba redox similar al Centro respiratorio III, que cataliza la transferencia de los electrones a dos moléculas de un citocromo soluble, del tipo c, denominado citocromo c2 La reacción es la siguiente: Quinol + 2 Cit c oxidado + nH+ d → Quinona + 2 Cit c red + nH+ f Los electrones vuelven al fotosistema transportados por el citocromo c2: El gradiente de hidrogeniones así creado puede ser empleado por la bacteria para sintetizar ATP a través de la ATPasa F de su membrana. Observe que los electrones abandonan el fotosistema con un potencial redox mayor que el correspondiente al par + NAD/NADH (-0,32 V, en condiciones estándar), por lo que la transferencia de + electronesdesde lacoenzima Qreducida alNAD esun procesoendergónico. ✓ Cadena de transporte electrónico en bacterias “verdes” Estas bacterias tienen un fotosistema del tipo PSI, con un pico de absorción a 840 nm. Puede tener bacterioclorofilas de los tipos c, d, e. Los electrones abandonan PSI unidos a una ferrosulfoproteína soluble, de pequeño tamaño, la ferredoxina, que se desplaza por la cara interna –citosólica- y son transferidos a la coenzima Q por la enzima ferredoxina-coenzima Q oxidorreductasa y,a continuación siguen un camino similar al anterior; en este caso el citocromodetipocquedevuelveelelectrónalfotosistemaeselc 553 . La diferencia fundamental con el caso anterior es que el potencial redox de la ferredoxina es + menor que el del NAD ,por lo que esaferrosulfoproteína puede reducirdirectamente alNAD + + (o al NADP ,quetiene igualpotencialredox). ✓ La reducción del NAD(P) Lascadenasdetransporteelectrónicofotosintéticoquehemosvistohastaahorasirvenpara la síntesis de ATP. Pero el ATP sólo no basta para la autotrofía, esto es, para la síntesis de material celular a partir de CO 2, que es la forma más oxidada del carbono. Para ello se necesita, además de ATP, una fuente de poder reductor utilizable, esto es de NADPH. El problema está que la mayor parte de los posibles sustratos (excepto compuestos orgánicos muy reducidos, como ácidos grasos, claro es) existentes en el medio tienen potenciales redox mas positivos que el par NADP/NADPH. Es decir, la transferencia de electrones desde esos donadores exógenos (sulfuro, tiosulfato, azufre, incluso hierro ferroso en algún caso, es un proceso endergónico. La excepción a lo anterior la forman las bacterias que poseen la enzima hidrogenasa, que emplea directamente el hidrógeno molecular (Eº´-0,42 V aprox. a pH 7), para reducir el NADP. La reacción es: + + H +NADP → H +NADPH 2 Esta enzima está presente en un número bastante elevado de bacterias, incluyendo bacterias púrpuras, y el hidrógeno molecular es frecuente en medios anóxicos como producto de la actividadmicrobiana. Ahora bien, no es precisa la existencia de hidrógeno molecular para poder llevar a cabo la reducción del CO,2ya que se han desarrollado dos estrategias que permiten cabo la + reducción endergónica del NADP a partir de un donador más electropositivo empleando la energía de la luz: 1) Eltransporte inverso deelectrones 2) Lafotosíntesisno cíclica. ✓ El Transporte inverso de electrones El transporte inverso de electrones se basa en la reversibilidad del centro respiratorio I. Consisteenemplearlaenergíaliberadaporloshidrogeniones alentrara +favor desupotencial + electroquímico para llevar a cabo la reducción delNADP (odelNAD )apartirde lacoenzima Q reducida. Los electrones para reducir a la coenzima Q pueden provenir de una fuente orgánica (succinato) o de un compuesto inorgánico, como el sulfuro o el azufre elemental. Esteprocedimientoeselquesiguenlasbacteriaspúrpuras. En este caso la energía del gradiente de hidrogeniones creada por la fotosíntesis se empleatantoparasintetizarATPatravésde laATP sintetasaFcomoparareduciralNAD(P) + atratnrsafveérsir lodes lelcecntrtoronersedspeisrdaetoerilodaI.doSreorrgeáqnuicieoreoni,noardgeám neacQ es. aErniaesl niácos,halastaelanzciomeanszim cpasrao de que el dador de electrones sea el succinato se emplea la misma succinato deshidrogenasa queenel ciclodeKrebs. Note que el transporte de electrones desde el fotosistema hasta el citocromo bf, centro respiratorio III, también lo lleva a cabo la quinona reducida, aunque en la figura se ha representado separadamente para mayor claridad. Si el producto de la reducción fuese el + NADH,éste cede sus electrones al NADP mediantelaenzimatranshidrogenasa. ✓ La fotosíntesis No cíclica En el caso de las bacterias que emplean el PSI los electrones son transportados por la ferredoxina reducida, que tiene un potencial más negativo que el NADP y que por consiguiente puede reducirlo directamente. Para ello se requiere la enzima ferredoxinaNADP oxidoreductasa, que es una enzima de membrana. El problema ahora es doble: en primer lugar, los electrones no llegan al citocromo bf, por lo que NO se crea el gradiente de hidrogeniones;además, se requiere un donador de electrones para reduciralradical catiónico de la clorofila, ya que el electrón la clorofila se ha desviado hacia el NADP. La cadena de transporte electrónico queda delade siguiente manera: Las bacterias “verdes” (Chlorobium y relacionados) pueden emplear diversos donadores de electrones: sulfuro, tiosulfato, azufre elemental y diversos compuestos orgánicos, para llevar a cabo la reducción de la clorofila oxidada. Evidentemente, ajustan el flujo de electrones a través de las vías cíclica (produce sólo ATP sin consumir poder reductor externo) y no cíclica (produce sólo NADPH usando un donador externo de electrones) para maximizar la eficiencia del sistema en función de sus necesidades metabólicas. El esquema de la cadena de transporteelectrónicodeestasbacteriaseselsiguiente: 1: PSI; 2, citocromo bf; 3: Ferredoxina:coQ oxidorreductasa; 4: Ferredoxina:NADP oxidoreductasa.Enrojo,transportenocíclico;enazul. Hace mucho, mucho tiempo (pero No en una galaxia muy lejana) Las estrategias explicadas en los dos apartados anteriores permiten la supervivencia de las bacterias fotosintéticas en medios muy diversos, siempre que dispongan de luz y de una fuente de poder reductor. Sin embargo, tienen una limitación grave: precisamente la fuente de poder reductor; el azufre reducido, o compuestos orgánicos, aunque son relativamente abundantes en muchos ambientes, faltan en otros. Además, dependen de la actividad de otros organismos: bacterias reductoras de sulfato, o productoras de hidrógeno, o fermentadoras. En resumen, las bacterias fotosintéticas que hemos estudiado dependen de la actividad de otras bacterias o están restringidas a nichos ecológicos muy concretos, por la faltade agentesreductoresadecuados. Una molécula muy abundante es el agua; pero con un potencial redox de +0,82 para el par oxigeno/agua, es un donador de electrones muy poco favorable. El empleo del agua no fue posible hasta que las cianobacterias desarrollaron una fotosíntesis en la que acoplaron en serie un fotosistema de tipo II y un fotosistema de tipo I, junto con una proteína especial, el complejo productor de oxígeno o OEC para extraer electrones del agua y emplearlos para reducira la clorofila oxidada: Probablemente la necesidad del empleo de dos fotosistemas viene dada porque el intervalo energético entre un aceptor de electrones mas oxidante que el agua y un donador de electrones más reductor que el NADP es excesivamente grande para ser cubierto por un solo fotosistema. Sea como fuere, el mecanismo empleado inicialmente por las cianobacterias ha supuesto todo un éxito evolutivo. Además, el oxígeno producido como subproducto es el aceptor de electrones más eficiente para la respiración, aunque por otro lado es altamente tóxico en ausencia de enzimas capaces de eliminar los radicales libres que pueden producir un daño oxidativo significativo en macromoléculas y lípidos. En resumen, este tipo de fotosíntesis supuso una auténtica revolución en las condiciones existentes en la Tierra primitiva. ✓ Algo de evolución y sistemática La distribución de los diferentes tipos de fotosíntesis en el mundo eubacteriano es la siguiente: Árbol filogenético eubacteriano basado en datos del ARN 16S . Se señalan exclusivamente los linajes que tienen miembros fotosintéticos. Heliobacter es la única bacteria fotosintética de las rama de las bacterias Gram+ . Las bacterias “púrpuras” pertenecen al grupo de las proteobacterias, que engloba también a muchas bacterias no fotosintéticas, como E.coli, Pseudomonas o Rhizobium. Las bacterias “verdes” como Chlorobium representan un linaje independiente de las restantes eubacterias. Prochloron se consideraenlaactualidadunacianobacteriaespecializada. Eneltexto “Brock Biología de los microorganismos” (ed. Prentice Hall, 8ª edición), de donde se ha tomado este árbol filogenético, a Chloroflexus se la denomina como bacteria “verde no del azufre”; al grupo de Chlorobium “verde del azufre”; y las aquí denominadas “bacterias púrpuras” corresponden a los grupos fisiológicos “rojas del azufre” y “rojas no del azufre”. Chl:Clorofila,delostiposayb. Bchl:bacterioclorofila,quepuedeserdelostiposa,b, c,d,e, yg. Importancia de la fotosíntesis 1. Inicia lacadena alimenticia,alsintetizar elnutriente (glucosa). 2. Oxigenaelmediopermitiendolarespiracióndelosanimalesyfavoreciendola formación del ozono. 3. Disminuye elefecto invernadero, al absorber, alabsorber elCO2.