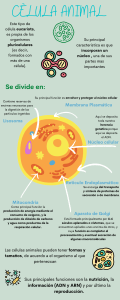
a LANGE medical book Fisiología médica Un enfoque por aparatos y sistemas ERRNVPHGLFRVRUJ Hershel Raff, PhD Professor Departments of Medicine and Physiology Medical College of Wisconsin Endocrine Research Laboratory Aurora St. Luke’s Medical Center Milwaukee, Wisconsin Michael Levitzky, PhD Professor of Physiology and Anesthesiology Louisiana State University Health Sciences Center New Orleans, Louisiana Traducción: Dr. Bernardo Rivera Muñoz Dr. Germán Arias Rebatet MÉXICO • BOGOTÁ • BUENOS AIRES • CARACAS • GUATEMALA MADRID • NUEVA YORK • SAN JUAN • SANTIAGO • SÃO PAULO AUCKLAND • LONDRES • MILÁN • MONTREAL • NUEVA DELHI SAN FRANCISCO • SIDNEY • SINGAPUR • ST. LOUIS • TORONTO Director editorial: Javier de León Fraga Editor de desarrollo: Manuel Bernal Pérez Supervisor de producción: Juan José Manjarrez de la Vega NOTA La medicina es una ciencia en constante desarrollo. Conforme surjan nuevos conocimientos, se requerirán cambios de la terapéutica. El(los) autor(es) y los editores se han esforzado para que los cuadros de dosificación medicamentosa sean precisos y acordes con lo establecido en la fecha de publicación. Sin embargo, ante los posibles errores humanos y cambios en la medicina, ni los editores ni cualquier otra persona que haya participado en la preparación de la obra garantizan que la información contenida en ella sea precisa o completa, tampoco son responsables de errores u omisiones, ni de los resultados que con dicha información se obtengan. Convendría recurrir a otras fuentes de datos, por ejemplo, y de manera particular, habrá que consultar la hoja informativa que se adjunta con cada medicamento, para tener certeza de que la información de esta obra es precisa y no se han introducido cambios en la dosis recomendada o en las contraindicaciones para su administración. Esto es de particular importancia con respecto a fármacos nuevos o de uso no frecuente. También deberá consultarse a los laboratorios para recabar información sobre los valores normales. FISIOLOGÍA MÉDICA. UN ENFOQUE POR APARATOS Y SISTEMAS Prohibida la reproducción total o parcial de esta obra, por cualquier medio, sin autorización escrita del editor. DERECHOS RESERVADOS © 2013, respecto a la primera edición en español por, McGRAW-HILL INTERAMERICANA EDITORES, S.A. de C.V. A subsidiary of the McGraw-Hill Companies, Inc. Prolongación Paseo de la Reforma 1015, Torre A, Piso 17, Col. Desarrollo Santa Fe, Delegación Álvaro Obregón C. P. 01376, México, D. F. Miembro de la Cámara Nacional de la Industria Editorial Mexicana Reg. No. 736 ISBN: 978-607-15-0913-0 Translated from the first English edition of: Medical Physiology: A Systems Approach Copyright © 2011 by McGraw-Hill Companies, Inc. All Rights Reserved ISBN : 978-0-07-162173-1 1234567890 Impreso en México 2456789013 Printed in Mexico A nuestros estudiantes, maestros, colaboradores y familiares. Características clave de Fisiología médica UN ENFOQUE POR APARATOS Y SISTEMAS Un enfoque conciso, con orientación clínica, para aprender importantes conceptos fisiológicos y correlaciones clínicas iv • NUEVA presentación a todo color • Refleja el mayor hincapié de la educación médica en proporcionar a los estudiantes de Medicina contenido con más orientación clínica durante sus dos primeros años de estudios • Se centra en los conceptos esenciales necesarios para entender la fisiopatología • Ofrece una perspectiva general sucinta pero integral de la fisiología, junto con una introducción a principios de ciencias básicas y su importancia para la expresión clínica de enfermedades • Detalla los principales procesos fisiológicos involucrados tanto en la salud como en la enfermedad • Excelente para la revisión del USMLE • Los cuadros y gráficas ilustran conceptos difíciles • Cada capítulo empieza con una lista de objetivos, y finaliza con un resumen y preguntas de estudio diseñados para probar el conocimiento de conceptos importantes cubiertos en ese capítulo • Casi todos los capítulos incluyen correlaciones clínicas que refuerzan los principios fisiológicos importantes cubiertos, y que ilustran su importancia para entender estados de enfermedad v CARACTERÍSTICAS CLAVE 2 SECCIÓN I Introducción Gránulos secretores Aparato de Golgi Nuevo diseño a todo color Centríolos Retículo endoplasmático rugoso Retículo endoplasmático liso Lisosomas Envoltura nuclear CAPÍTULO 1 Conceptos fisiológicos generales Gotitas de lípido Mitocondria los centrosomas, que también tienen importancia en el movimiento de los cromosomas durante la división celular. Por último, el núcleo, también rodeado por una membrana bicapa lipídica llamada la envoltura nuclear, contiene cromatina, la cual está compuesta de DNA que contiene el código de ácidos nucleicos para la diferenciación, la función y la replicación celulares. El DNA contiene los genes que codifican para mRNA que se producen a partir del DNA mediante transcripción. El núcleo también contiene el nucléolo, que es el sitio de síntesis de ribosomas. Como se menciona en varios capítulos de este libro, la membrana celular contiene diferentes tipos de receptores, los cuales detectan señales extracelulares que son transformadas hacia señales intracelulares mediante transducción. Además, hay receptores dentro del citoplasma y el núcleo que responden a señales que entran a la célula. Los ejemplos de esas señales son hormonas esteroides, como los Cabezas globulares Nucléolo FIGURA 11 Diagrama que muestra una célula hipotética en el centro, observada con un microscopio óptico. (Adaptada, con autorización de Fawcett DW et al. The ultrastructure of endocrine glands, Recent Prog Horm Res. 1969;25:315-380.) se muestra la estructura de la bicapa lipídica y las proteínas asociadas de la membrana celular. El interior de la célula está compuesto de citosol, que es un líquido que consta de agua, en la cual están disueltas proteínas, como metabolitos, combustible e iones inorgánicos (que se conocen como electrólitos). Dispersos en el citosol también hay diversas partículas subcelulares y organelos. En conjunto, la combinación de citosol y las estructuras intracelulares se llama el citoplasma. Los orgánulos incluyen el retículo endoplasmático, que es una extensa red de membranas dentro de las cuales hay proteínas y otras sustancias químicas importantes. El retículo endoplasmático tiene importancia en muchas funciones metabólicas y en el empaque de productos secretores. Los ribosomas están involucrados en la traducción, que es la síntesis de proteínas a partir del RNA mensajero (mRNA). Estos ribosomas se asocian con el retículo endoplasmático en una estructura combinada llamada retículo endoplasmático rugoso (RER). El aparato de Golgi se vincula con el retículo endoplasmático, y empaca material sintetizado en el RER. Los lisosomas son estructuras intracelulares, rodeadas por membrana, y contienen enzimas digestivas que se sitúan en gránulos involucrados en el metabolismo intracelular. Los gránulos secretores contienen moléculas que la célula liberará hacia el líquido extracelular mediante exocitosis, en respuesta a estímulos. Algunas células contienen muchas gotitas de lípido, porque la grasa es hidrofóbica y no se disuelve con facilidad en el ambiente acuoso del citosol. Las mitocondrias tienen dos membranas de bicapa lipídica en aposición, y son los organelos que generan energía. A los orgánulos citoplasmáticos los mantienen en su posición filamentos y microtúbulos, los cuales surgen a partir de 3 estrógenos y la testosterona, las cuales son lipofílicas (“que aman la grasa”) y, como resultado, pueden difundirse con facilidad a través de la membrana celular para ejercer una acción intracelular. ESTRUCTURA GENERAL DEL CUERPO En la figura 1-3 se presenta un diagrama del cuerpo humano. Los órganos (por ejemplo, cerebro y corazón) reciben nutrientes y eliminan productos de desecho por medio del sistema circulatorio. El corazón se ilustra en dos partes —derecha e izquierda— como una representación funcional, aun cuando en realidad es un solo órgano. El lado derecho del corazón recibe sangre parcialmente desoxigena- Líquido extracelular Carbohidrato de glucoproteína Proteínas transmembrana Fosfolípidos Sistema nervioso central Canal Nervios aferentes y eferentes Proteínas integrales O2 CO2 Lado derecho Proteína periférica Regiones polares Sangre venosa Regiones no polares Atmósfera O2 CO2 Pulmón Lado izquierdo del corazón del corazón Tejidos Líquido intracelular Sangre arterial Nutrientes FIGURA 12 Organización de la bicapa de fosfolípido y proteínas asociadas en una membrana celular biológica. (Reproducida con autorización de Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology, 11th ed. Productos de desecho McGraw-Hill, 2008.) Glándulas endocrinas Hormonas Hígado Tracto GI Síntesis Nutrientes Metabolismo Bilis Desecho Resorción Riñón FIGURA 13 Organización general de Filtración Desecho Cuadros y gráficas con información crucial 122 CORRELACIÓN CLÍNICA RESUMEN DEL CAPÍTULO ■ ■ Los receptores sensoriales por lo común se clasifican como mecanorreceptores, nociceptores, quimiorreceptores o fotorreceptores. El tacto y la presión son detectados por cuatro tipos de mecanorreceptores: corpúsculos de Meissner (que muestran respuesta a cambios de la textura y vibraciones lentas), las células de Merkel (que responden a presión y tacto sostenidos), los corpúsculos de Orina Heces los principales órganos del cuerpo. Las flechas muestran las direcciones de flujo sanguíneo y flujo de gases, nutrientes, hormonas y productos de desecho. Prácticas ayudas para el aprendizaje que propician retener información esencial SECCIÓN IV Fisiología del sistema nervioso central/neural Una mujer de 55 años de edad, ejecutiva en una corporación grande, presentó una sensación de ardor en la palma de la mano derecha alrededor de seis meses antes. También notó hormigueo y entumecimiento en el pulgar y los dedos índice y medio derechos. Estos síntomas aparecieron después de pasar muchas y largas horas frente a la computadora preparando documentos del reporte anual para la corporación. Al inicio, los síntomas fueron más prominentes por la noche, e interrumpían el sueño de la paciente. El problema se ha intensificado a últimas fechas, y ahora tiene dolor en la muñeca derecha y dificultad para tomar objetos pequeños desde el escritorio. Visitó a su médico porque el trabajo en la computadora se había hecho cada vez más difícil. El médico llevó a cabo varias pruebas diagnósticas sencillas; cuando ejerció presión sobre el nervio mediano en la muñeca, la paciente experimentó una sensación parecida a descarga eléctrica (signo de Tinel). Cuando el médico le pidió que mantuviera los antebrazos hacia arriba al apuntar con los dedos hacia abajo y presionar los dorsos de las manos uno contra otro, en el transcurso de un minuto la mujer sintió hormigueo y entumecimiento creciente en los dedos de la mano (signo de Phalen). Las pruebas de conducción nerviosa indicaron conducción lentificada en el nervio mediano. Se diagnosticó síndrome del túnel carpiano, que se debe a compresión (quizá debido a inflamación) del nervio mediano que pasa por el túnel. Es más prevaleciente en mujeres que en varones, y se diagnostica por lo regular en individuos que usan las muñecas en actividades repetitivas (operadores de computadora, cajeros, músicos, pintores). Alrededor de 3% de las mujeres y 2% de los varones tienen probabilidades de que se les diagnostique este síndrome durante su vida. El nervio mediano proporciona información sensitiva proveniente del pulgar, y de los dedos índice y anular, y los nueve tendones que flexionan los dedos de las manos. El síndrome se caracteriza por dolor, parestesias y debilidad en la distribución del nervio mediano. El dolor en la muñeca o la mano, o el entumecimiento y hormigueo de los dedos de la mano (excepto el dedo meñique, que no está inervado por el nervio mediano) a menudo son los primeros síntomas. Los pacientes a veces informan debilidad en la mano, y una tendencia a dejar caer cosas. Los síntomas a menudo aparecen primero por la noche más que durante la actividad. El mejor tratamiento suele ser la colocación de una férula en la muñeca, AINE o corticosteroides. Si el dolor persiste, puede requerirse intervención quirúrgica. Ingesta de alimentos y agua ■ ■ ■ ■ ■ ■ Ruffini (que muestran respuesta a presión sostenida) y los corpúsculos de Pacini (que responden a presión profunda y vibraciones rápidas). Los nociceptores y termorreceptores son terminaciones nerviosas sas libres sobre fibras no mielinizadas o un poco mielinizadas en piel con pelo y glabra, y tejidos profundos. La hiperalgesia es una respuesta exagerada a un estímulo nocivo; vo; la alodinia es una sensación de dolor en respuesta a un estímulo o inocuo. La conversión de un estímulo de receptor en una sensación reconocible se denomina codificación sensorial. Todos los sistemas sensoriales codifican para cuatro atributos elementales es de un estímulo: modalidad, ubicación, intensidad y duración. El tacto discriminativo, la propiocepción y las sensaciones vibratorias son transmitidos por medio de la vía de la columnaa dorsal (lemnisco medial) a SI. Las sensaciones de dolor y temperatura son mediadas por medio del tracto espinotalámico o ventrolateral a SI. Las vías descendentes desde la PAG mesencefálica inhiben la transmisión en vías nociceptivas. Esta vía descendente incluye una sinapsis en el núcleo del rafe y la liberación de opiáceos endógenos. La morfina es un agente antinociceptivo eficaz que se une a receptores opiáceos endógenos en el mesencéfalo, el tallo encefálico y la médula espinal. PREGUNTAS DE ESTUDIO 1. Los corpúsculos de Pacini: A) son un tipo de termorreceptor. B) por lo general están inervados por fibras nerviosas Aδ. C) son receptores de tacto que se adaptan con rapidez. D) son receptores de tacto que se adaptan de manera lenta. E) son nociceptores. 2. La adaptación a un estímulo sensorial produce: A) una sensación disminuida cuando se suspenden otros tipos de estímulos sensoriales. B) una sensación más intensa cuando un estímulo dado se aplica ca de forma repetitiva. C) una sensación localizada a la mano cuando se estimulan los nervios del plexo braquial. D) una sensación disminuida cuando un estímulo dado se aplica ca repetidas veces con el tiempo. E) una tasa de activación disminuida en el nervio sensorial del receptor cuando se dirige la atención a otro tema. 3. ¿Los sistemas sensoriales codifican para cuál de los atributos que ue siguen de un estímulo? A) modalidad, ubicación, intensidad y duración B) umbral, campo receptivo, adaptación y discriminación C) tacto, gusto, audición y olfato D) umbral, lateralidad, sensación y duración E) sensibilización, discriminación, energía y proyección 4. Los termorreceptores: A) sólo son activados por frío o calor intenso. B) están ubicados sobre capas superficiales de la piel C) son un subtipo de nociceptor. D) están en terminaciones dendríticas de fibras Aδ y fibras C. E) todas las anteriores. 14 Reflejos espinales C A P Í T U L O Susan M. Barman O B J E T I V O S ■ ■ ■ ■ ■ Definir los componentes de un arco reflejo. Describir los husos musculares y su función en el reflejo de estiramiento. Explicar las funciones de los órganos tendinosos de Golgi como parte de un sistema de retroacción que mantiene la fuerza muscular. Definir la inervación recíproca, el reflejo de estiramiento inverso, y el clono. Describir los efectos a corto y largo plazo de la lesión de la médula espinal sobre los reflejos espinales. INTRODUCCIÓN La unidad básica de la actividad refleja integrada es el arco reflejo. Este arco consta de un órgano de sentido, una neurona aferente, sinapsis dentro de una estación integradora central, una neurona eferente, y un órgano efector. Las neuronas aferentes entran al sistema nervioso central (SNC) por medio de las raíces dorsales espinales o los nervios craneales, y tienen su cuerpo celular en los ganglios de la raíz dorsal o en los ganglios homólogos para los nervios craneales. Las fibras eferentes salen del SNC por medio de las raíces ventrales espinales o nervios craneales motores correspondientes. La actividad en el arco reflejo empieza en un receptor sensorial con un potencial generador cuya magnitud es proporcional a la fuerza del estímulo (figura 14-1). Esto genera potenciales de acción de todo o nada en el nervio aferente; el número de potenciales de acción es proporcional al tamaño del potencial generador. En el SNC, las respuestas de nuevo son graduadas en términos de potenciales postsinápticos excitadores (EPSP) y potenciales postsinápticos inhibidores (IPSP) en las uniones sinápticas (capítulo 7). Se generan respuestas de todo o nada en el nervio eferente; cuando éstas llegan al órgano efector, de nuevo establecen una respuesta graduada. Cuando el efector es músculo liso, las respuestas se suman para producir potenciales de acción en el músculo liso, pero cuando el efector es músculo esquelético, la respuesta graduada es adecuada para producir potenciales de acción que desencadenan contracción muscular. La actividad dentro del arco reflejo es modificada por las múltiples aferencias que convergen en las neuronas eferentes o en cualquier estación sináptica dentro del asa refleja. El arco reflejo más simple es el que tiene una sinapsis única entre las neuronas aferentes y eferentes. Dichos arcos son monosinápticos, y los reflejos que ocurren en ellos se llaman reflejos monosinápticos. Los arcos reflejos en los cuales hay una o más interneuronas interpuestas entre las neuronas aferentes y eferentes se llaman reflejos polisinápticos. Puede haber desde dos, hasta cientos de sinapsis en un arco reflejo polisináptico. Como es evidente a partir de la descripción que se presenta más adelante, la actividad refleja se estereotipa y especifica en términos tanto del estímulo como de la respuesta; un estímulo particular desencadena una respuesta particular. El hecho de que las respuestas reflejas se estereotipen no excluye la posibilidad de que sean modificadas por la experiencia. Los reflejos son adaptables y pueden cambiarse para realizar tareas motoras y mantener el equilibrio. Las aferencias descendentes que provienen de regiones más altas del cerebro desempeñan un papel importante en la modulación de reflejos espinales y la adaptación de los mismos. REFLEJO MONOSINÁPTICO: EL REFLEJO DE ESTIRAMIENTO Cuando un músculo esquelético con inervación intacta es estirado, se contrae. Esta respuesta se llama reflejo de estiramiento. El estímulo que inicia el reflejo es estiramiento del músculo, y la respuesta es contracción del mismo músculo. El órgano de sentido (receptor) es una pequeña estructura que se parece a huso o fusiforme encapsulada que se llama el huso muscular, el cual se ubica dentro de la parte 125 Acerca de los autores Hershel Raff Michael Levitzky Hershel Raff recibió su Ph.D. in Environmental Physiology en la Johns Hopkins University en 1981, y realizó adiestramiento posdoctoral en Endocrinology en la University of California en San Francisco. En 1983 ingresó al profesorado del Medical College of Wisconsin, y en 1991 ascendió al rango de Professor of Medicine (Endocrinology, Metabolism, and Clinical Nutrition) and Physiology. También es Director of the Endocrine Research Laboratory en Aurora St. Luke’s Medical Center. En el Medical College of Wisconsin, imparte fisiología y farmacología a estudiantes de medicina y graduados. Tiene una admisión inaugural a la Society of Teaching Scholars, recibió el Beckman Basic Science Teaching Award y el Outstanding Teacher Award, y ha sido uno de los MCW’s Outstanding Medical Student Teachers cada año que se ha otorgado el premio. El Dr. Raff fue electo para la Alpha Omega Alpha (AOA) Honor Medical Society como un maestro del profesorado en 2005. También es Adjunct Professor of Biomedical Sciences at Marquette University. Es Associate Editor of Advances in Physiology Education. Fue Secretary-Treasurer de la Endocrine Society y en la actualidad es Chair del Publications Committee de la American Physiological Society. En 2005 fue electo Fellow de la American Association for the Advancement of Science. La investigación básica del Dr. Raff se enfoca en la adaptación a oxígeno bajo (hipoxia). Su interés clínico se centra en enfermedades hipofisarias y suprarrenales, con un enfoque especial en el diagnóstico de síndrome de Cushing. El Dr. Raff también es coautor de Vander’s Human Physiology (McGraw-Hill) actualmente en su 12ª edición, y de Physiology Secrets, en la actualidad en su 2ª edición. Michael Levitzky es Professor of Physiology and Anesthesiology en el Louisiana State University Health Sciences Center, y Director of Basic Science Curriculum en la LSU School of Medicine en New Orleans. En 1969 recibió un B.A. en la University of Pennsylvania, y en 1975 un Ph.D. en Physiology en el Albany Medical College. Ingresó al profesorado de la LSU School of Medicine en 1975, y ascendió al rango de Professor en 1985. También ha sido Adjunct Professor of Physiology en la Tulane University School of Medicine desde 1991. El Dr. Levitzky imparte fisiología a estudiantes de medicina, residentes, becarios y estudiantes graduados. Ha recibido muchos premios de enseñanza por parte de organizaciones estudiantiles tanto en la LSU como en la Tulane University. En 1997 recibió el primer LSUHSC Allen A. Copping Award for Excellence in Teaching in the Basic Sciences, y en 1998 el American Physiological Society’s Arthur C. Guyton Teacher of the Year Award. Fue electo para la Alpha Omega Alpha (AOA) Honor Medical Society como un maestro del profesorado en 2006. El Dr. Levitzky ha prestado servicio en la American Physiological Society como miembro del Education Committee y del Steering Committee of the Teaching Section. De 2007 a 2011 prestó servicio como miembro del National Board of Medical Examiners United States Medical Licensing Examination (USMLE) Step 1 Physiology Test Material Development Committee. Es autor o coautor de varios otros libros, uno de los cuales, Pulmonary Physiology (Lange/McGraw-Hill), en la actualidad está en su 7ª edición. vi Contenido Colaboradores Prefacio xiii S E C C I Ó N xi I INTRODUCCIÓN Capítulo 1. Capítulo 10. Estructura y función del músculo cardiaco 93 1 Kathleen H. McDonough Capítulo 11. Estructura y función del músculo liso 99 Conceptos fisiológicos generales 1 Hershel Raff y Michael Levitzky S E C C I Ó N II FISIOLOGÍA CELULAR Capítulo 2. Kathleen H. McDonough S E C C I Ó N FISIOLOGÍA DEL SISTEMA NERVIOSO CENTRAL/NEURAL 105 9 Células y mecanismos celulares 9 Capítulo 12. Introducción al sistema nervioso 105 David Landowne Capítulo 3. Membranas celulares y mecanismos de transporte 15 Susan M. Barman Capítulo 13. Sistemas sensoriales generales: tacto, dolor y temperatura 115 David Landowne Capítulo 4. Canales y control del potencial de membrana 33 Susan M. Barman Capítulo 14. Reflejos espinales 125 David Landowne Capítulo 5. Susan M. Barman Potenciales generadores sensoriales 43 Capítulo 15. Sentidos especiales I: visión 133 Susan M. Barman David Landowne Capítulo 6. Potenciales de acción Capítulo 16. Sentidos especiales II: audición y equilibrio 147 47 David Landowne Capítulo 7. IV Susan M. Barman Sinapsis 59 Capítulo 17. Sentidos especiales III: olfato y gusto 159 David Landowne Susan M. Barman S E C C I Ó N III FISIOLOGÍA MUSCULAR Capítulo 8. Capítulo 18. Control de la postura y el movimiento 167 Susan M. Barman 79 Capítulo 19. Sistema nervioso autónomo 177 Susan M. Barman Perspectiva general de la función muscular 79 Capítulo 20. Actividad eléctrica del cerebro, estados de sueño-vigilia y ritmos circadianos 185 Kathleen H. McDonough Capítulo 9. Estructura y función del músculo esquelético 83 Susan M. Barman Capítulo 21. Aprendizaje, memoria, lenguaje y habla 191 Kathleen H. McDonough Susan M. Barman vii viii CONTENIDO S E C C I Ó N V FISIOLOGÍA CARDIOVASCULAR 199 Capítulo 22. Perspectiva general del sistema cardiovascular 199 Capítulo 37. Regulación acido-básica y causas de hipoxia 375 Michael Levitzky Capítulo 38. Control de la respiración 385 Michael Levitzky Lois Jane Heller y David E. Mohrman Capítulo 23. Células de músculo cardiaco 211 Lois Jane Heller y David E. Mohrman Capítulo 24. La bomba cardiaca 223 Lois Jane Heller y David E. Mohrman Capítulo 25. Evaluaciones de la función cardiaca 235 Lois Jane Heller y David E. Mohrman Capítulo 26. Sistema vascular periférico 251 David E. Mohrman y Lois Jane Heller S E C C I Ó N FISIOLOGÍA RENAL Capítulo 28. Retorno venoso y gasto cardiaco 275 Douglas C. Eaton y John P. Pooler Capítulo 40. Flujo sanguíneo renal y filtración glomerular 409 Douglas C. Eaton y John P. Pooler Capítulo 41. Depuración 417 Douglas C. Eaton y John P. Pooler Capítulo 42. Mecanismos de transporte tubular 423 Douglas C. Eaton y John P. Pooler David E. Mohrman y Lois Jane Heller Capítulo 29. Regulación de la presión arterial 285 David E. Mohrman y Lois Jane Heller Capítulo 30. Respuestas cardiovasculares al estrés fisiológico 295 Lois Jane Heller y David E. Mohrman 397 Capítulo 39. Funciones, procesos básicos y anatomía renales 397 Capítulo 27. Control vascular 263 David E. Mohrman y Lois Jane Heller VII Capítulo 43. Manejo renal de sustancias orgánicas 429 Douglas C. Eaton y John P. Pooler Capítulo 44. Procesos renales básicos para sodio, cloruro y agua 437 Douglas C. Eaton y John P. Pooler S E C C I Ó N Capítulo 45. Regulación de la excreción de sodio y agua 449 VI FISIOLOGÍA PULMONAR 305 Capítulo 31. Función y estructura del sistema respiratorio 305 Michael Levitzky Capítulo 32. Mecánica del sistema respiratorio 313 Michael Levitzky Capítulo 33. Ventilación alveolar 331 Douglas C. Eaton y John P. Pooler Capítulo 46. Regulación del equilibrio de potasio 463 Douglas C. Eaton y John P. Pooler Capítulo 47. Regulación del equilibrio acidobásico 471 Douglas C. Eaton y John P. Pooler Capítulo 48. Regulación del equilibrio de calcio y fosfato 485 Douglas C. Eaton y John P. Pooler Michael Levitzky Capítulo 34. Perfusión pulmonar 341 Michael Levitzky Capítulo 35. Relaciones ventilaciónperfusión e intercambio de gases respiratorio 353 Michael Levitzky Capítulo 36. Transporte de oxígeno y dióxido de carbono 363 Michael Levitzky S E C C I Ó N VIII FISIOLOGÍA GASTROINTESTINAL 491 Capítulo 49. Perspectiva general del sistema gastrointestinal: anatomía funcional y regulación 491 Kim E. Barrett Capítulo 50. Secreción gástrica 507 Kim E. Barrett CONTENIDO Capítulo 51. Secreciones pancreática y salival 517 Capítulo 62. Adenohipófisis 623 Kim E. Barrett Capítulo 52. Absorción y secreción de agua y electrólitos 527 Kim E. Barrett Capítulo 53. Aspectos inmunitarios y ecológicos de la mucosa intestinal 535 Kim E. Barrett Patricia E. Molina Capítulo 63. Glándula tiroides 633 Patricia E. Molina Capítulo 64. Glándula paratiroides y regulación del calcio y fosfato 643 Patricia E. Molina Capítulo 65. Glándula suprarrenal 655 Capítulo 54. Motilidad intestinal 543 Kim E. Barrett Capítulo 55. Anatomía funcional del hígado y el sistema biliar 559 Kim E. Barrett Capítulo 56. Formación, secreción y almacenamiento de bilis 565 Patricia E. Molina Capítulo 66. Páncreas endocrino 671 Patricia E. Molina Capítulo 67. Aparato reproductor masculino 683 Patricia E. Molina Capítulo 68. Aparato reproductor femenino 695 Patricia E. Molina Kim E. Barrett Capítulo 57. Manejo de la bilirrubina y el amoniaco por el hígado 575 Capítulo 69. Integración endocrina del equilibrio energético y electrolítico 715 Patricia E. Molina Kim E. Barrett Capítulo 58. Digestión y absorción de carbohidratos, proteínas y vitaminas hidrosolubles 583 Kim E. Barrett Capítulo 59. Asimilación de lípidos 593 Kim E. Barrett ix S E C C I Ó N X FISIOLOGÍA INTEGRATIVA 729 Capítulo 70. Control de la temperatura corporal 729 Hershel Raff y Michael Levitzky S E C C I Ó N IX ENDOCRINOLOGÍA Y FISIOLOGÍA METABÓLICA 601 Capítulo 60. Principios generales de fisiología endocrina 601 Capítulo 71. Hipoxia e hiperbaria 735 Michael Levitzky y Hershel Raff Capítulo 72. Ejercicio 745 Michael Levitzky y Kathleen H. McDonough Capítulo 73. Envejecimiento 753 Hershel Raff Patricia E. Molina Capítulo 61. Hipotálamo y adenohipófisis 613 Patricia E. Molina Respuestas a las preguntas de estudio Índice 761 757 Colaboradores Susan M. Barman, PhD Professor Department of Pharmacology & Toxicology and Neuroscience Program Michigan State University East Lansing, Michigan Kathleen H. McDonough, PhD Professor Department of Physiology Associate Dean, School of Graduate Studies Louisiana State University Health Sciences Center New Orleans, Louisiana Kim E. Barrett, PhD Professor of Medicine and Dean of Graduate Studies University of California, San Diego La Jolla, California Patricia E. Molina, MD, PhD Richard Ashman, PhD Professor and Head of Physiology Department of Physiology Louisiana State University Health Sciences Center New Orleans, Louisiana Douglas C. Eaton, PhD Distinguished Professor and Chair of Physiology and Professor of Pediatrics Department of Physiology and Center for Cell & Molecular Signaling Emory University School of Medicine Atlanta, Georgia Lois Jane Heller, PhD Professor Department of Physiology and Pharmacology University of Minnesota Medical School Duluth, Minnesota David Landowne, PhD Professor Department of Physiology and Biophysics University of Miami, Miller School of Medicine Miami, Florida David E. Mohrman, PhD Associate Professor, Emeritus Department of Physiology and Pharmacology University of Minnesota Medical School Duluth, Minnesota John P. Pooler, PhD Professor of Physiology Emeritus Emory University School of Medicine Atlanta, Georgia Hershel Raff, PhD Professor Departments of Medicine and Physiology Medical College of Wisconsin Endocrine Research Laboratory Aurora St. Luke’s Medical Center Milwaukee, Wisconsin Michael Levitzky, PhD Professor of Physiology and Anesthesiology Louisiana State University Health Sciences Center New Orleans, Louisiana xi Asesora para la revisión científica de la edición en español Dr. med. Nancy Esthela Fernández Garza Jefe del Departamento de Fisiología Facultad de Medicina Universidad Autónoma de Nuevo León xii Prefacio como las secciones de fisiología muscular y de fisiología integrativa son nuevas. Cada capítulo empieza con una lista de objetivos y concluye con un resumen. Casi todos los capítulos también finalizan con una correlación clínica que refuerza los principios fisiológicos principales que acaban de aprenderse, e ilustra su importancia para entender estados de enfermedad. Cada capítulo termina con preguntas de opción múltiple diseñadas para probar el conocimiento de algunos de los conceptos importantes cubiertos. Los autores estamos en deuda con nuestros mentores, quienes nos proporcionaron un fundamento para los avances en educación fisiológica durante el siglo XXI. También agradecemos a nuestros estudiantes por proporcionarnos una caja de resonancia para los métodos pedagógicos explotados en este libro. Los autores agradecen a Michael Weitz, Karen Davis y Brian Kearns de McGraw-Hill por su sobresaliente ayuda editorial. Finalmente, un agradecimiento especial a nuestras familias: Judy y Jonathan; y Elizabeth, Edward y Sarah. Fisiología médica: un enfoque por aparatos y sistemas tiene el propósito de proporcionar a estudiantes de primer año de medicina y graduados, y estudiantes de pregrado avanzados la base de los principales procesos fisiológicos necesarios para entender tanto la salud como la enfermedad. Los planes de estudios de muchas escuelas de medicina están cambiando: casi todas ellas han pasado por una transición desde un enfoque de bloque, con un curso propio para cada disciplina, hacia una estructura integrada verticalmente, o se encuentran en dicha transición. Uno de los objetivos de un plan de estudios integrado es la presentación de mucho más material clínico durante los primeros dos años de estudios de medicina, así como el reforzamiento de conceptos básicos durante los dos años principalmente clínicos. Como resultado, hay un enfoque creciente en los conceptos esenciales necesarios para entender fisiopatología. En consecuencia, este libro es considerablemente más corto que el tratado de fisiología estándar completo. Se enfoca en conceptos fisiológicos y correlaciones clínicas importantes, y deja los detalles específicos para libros de mayor tamaño. Casi todo este libro evolucionó a partir de las series de monografías Lange Physiology Series. La sección sobre el sistema nervioso central surgió a partir de la 23ª edición de Ganong. Fisiología médica. Finalmente, la introducción, así Hershel Raff Michael Levitzky xiii SECCIÓN I INTRODUCCIÓN Conceptos fisiológicos generales C A P Í T U L O 1 Hershel Raff y Michael Levitzky O B J E T I V O S ■ ■ ■ ■ ■ ■ ■ Entender las propiedades generales de una célula eucarionte. Explicar la organización general de los órganos internos del cuerpo. Comparar y contrastar la composición del líquido extracelular con la del intracelular. Describir los diferentes tipos de transporte de membrana. Entender los conceptos generales de presión, flujo, resistencia y adaptabilidad. Explicar el equilibrio de masas. Definir la retroalimentación negativa y positiva. INTRODUCCIÓN LA CÉLULA La fisiología es la ciencia que estudia la función de los organismos. El objeto de la fisiología es explicar cómo los sistemas, las células, e incluso las moléculas, interactúan para mantener una función normal. La característica distintiva de la fisiología es el concepto de la homeostasis, que es el mantenimiento de un ambiente interno normal ante perturbaciones externas o internas, de modo que se mantengan las funciones de las células y los sistemas de cuerpo. Esto se logra sobre todo por medio de sistemas de retroalimentación, de modo que cuando un sistema queda alterado, varias respuestas locales, reflejos sistémicos (reacciones rápidas, automáticas, a estímulos) y ajustes a largo plazo se activan para regresar el sistema a su valor establecido normal. Al entender cómo funcionan las cosas en condiciones normales, es posible apreciar cuándo hay un mal funcionamiento y por qué. Esto se llama fisiopatología —una alteración duradera de la función normal causada por enfermedad o lesión. Por ende, la fisiología es uno de los fundamentos de las ciencias de la salud. La célula es el bloque de construcción básico de los órganos del cuerpo. Los detalles de la fisiología celular se cubren en la sección II. En la figura 1-1 se muestra la estructura general de una célula nucleada (eucariota); está rodeada por una membrana celular compuesta de una bicapa lipídica, proteínas de membrana, y carbohidratos en asociación con lípidos (glucolípidos) o proteínas (glucoproteínas). La membrana celular es el portero para cualquier cosa que entre o salga de la célula, y es una barrera que ayuda a mantener la composición interna de la célula. Algunas proteínas y glucoproteínas de membrana funcionan como sensores, o receptores, que detectan el ambiente y señales químicas externos, y después emiten señales al interior de la célula, por lo general mediante sustancias químicas que actúan como segundos mensajeros, o por medio de cambios de la actividad eléctrica de la membrana. Otras proteínas de membrana funcionan como transportadores, que regulan la entrada o la salida de sustancias hacia la célula o hacia afuera de la célula. En la figura 1-2 1 2 SECCIÓN I Introducción Gránulos secretores Aparato de Golgi Centríolos Retículo endoplasmático rugoso Retículo endoplasmático liso Lisosomas Envoltura nuclear Gotitas de lípido Mitocondria Cabezas globulares Nucléolo FIGURA 11 Diagrama que muestra una célula hipotética en el centro, observada con un microscopio óptico. (Adaptada, con autorización de Fawcett DW et al. The ultrastructure of endocrine glands, Recent Prog Horm Res. 1969;25:315-380.) se muestra la estructura de la bicapa lipídica y las proteínas asociadas de la membrana celular. El interior de la célula está compuesto de citosol, que es un líquido que consta de agua, en la cual están disueltas proteínas, como metabolitos, combustible e iones inorgánicos (que se conocen como electrólitos). Dispersos en el citosol también hay diversas partículas subcelulares y organelos. En conjunto, la combinación de citosol y las estructuras intracelulares se llama el citoplasma. Los orgánulos incluyen el retículo endoplasmático, que es una extensa red de membranas dentro de las cuales hay proteínas y otras sustancias químicas importantes. El retículo endoplasmático tiene importancia en muchas funciones metabólicas y en el empaque de productos secretores. Los ribosomas están involucrados en la traducción, que es la síntesis de proteínas a partir del RNA mensajero (mRNA). Estos ribosomas se asocian con el retículo endoplasmático en una estructura combinada llamada retículo endoplasmático rugoso (RER). El aparato de Golgi se vincula con el retículo endoplasmático, y empaca material sintetizado en el RER. Los lisosomas son estructuras intracelulares, rodeadas por membrana, y contienen enzimas digestivas que se sitúan en gránulos involucrados en el metabolismo intracelular. Los gránulos secretores contienen moléculas que la célula liberará hacia el líquido extracelular mediante exocitosis, en respuesta a estímulos. Algunas células contienen muchas gotitas de lípido, porque la grasa es hidrofóbica y no se disuelve con facilidad en el ambiente acuoso del citosol. Las mitocondrias tienen dos membranas de bicapa lipídica en aposición, y son los organelos que generan energía. A los orgánulos citoplasmáticos los mantienen en su posición filamentos y microtúbulos, los cuales surgen a partir de Líquido extracelular Carbohidrato de glucoproteína Proteínas transmembrana Fosfolípidos Canal Proteínas integrales Proteína periférica Regiones polares Regiones no polares Líquido intracelular FIGURA 12 Organización de la bicapa de fosfolípido y proteínas asociadas en una membrana celular biológica. (Reproducida con autorización de Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology, 11th ed. McGraw-Hill, 2008.) CAPÍTULO 1 Conceptos fisiológicos generales los centrosomas, que también tienen importancia en el movimiento de los cromosomas durante la división celular. Por último, el núcleo, también rodeado por una membrana bicapa lipídica llamada la envoltura nuclear, contiene cromatina, la cual está compuesta de DNA que contiene el código de ácidos nucleicos para la diferenciación, la función y la replicación celulares. El DNA contiene los genes que codifican para mRNA que se producen a partir del DNA mediante transcripción. El núcleo también contiene el nucléolo, que es el sitio de síntesis de ribosomas. Como se menciona en varios capítulos de este libro, la membrana celular contiene diferentes tipos de receptores, los cuales detectan señales extracelulares que son transformadas hacia señales intracelulares mediante transducción. Además, hay receptores dentro del citoplasma y el núcleo que responden a señales que entran a la célula. Los ejemplos de esas señales son hormonas esteroides, como los 3 estrógenos y la testosterona, las cuales son lipofílicas (“que aman la grasa”) y, como resultado, pueden difundirse con facilidad a través de la membrana celular para ejercer una acción intracelular. ESTRUCTURA GENERAL DEL CUERPO En la figura 1-3 se presenta un diagrama del cuerpo humano. Los órganos (por ejemplo, cerebro y corazón) reciben nutrientes y eliminan productos de desecho por medio del sistema circulatorio. El corazón se ilustra en dos partes —derecha e izquierda— como una representación funcional, aun cuando en realidad es un solo órgano. El lado derecho del corazón recibe sangre parcialmente desoxigena- Sistema nervioso central Nervios aferentes y eferentes O2 CO2 Lado derecho Sangre venosa Atmósfera O2 CO2 Pulmón Lado izquierdo del corazón del corazón Tejidos Sangre arterial Nutrientes Productos de desecho Glándulas endocrinas Hormonas Hígado Tracto GI Síntesis Nutrientes Metabolismo Riñón Bilis Ingesta de alimentos y agua Desecho Resorción FIGURA 13 Organización general de Filtración Desecho Orina Heces los principales órganos del cuerpo. Las flechas muestran las direcciones de flujo sanguíneo y flujo de gases, nutrientes, hormonas y productos de desecho. 4 SECCIÓN I Introducción da que regresa desde los tejidos, y bombea sangre hacia los pulmones. En los pulmones, el oxígeno se difunde hacia la sangre desde la fase gaseosa para uso en la respiración celular en el cuerpo, y el dióxido de carbono, un producto de desecho de la respiración celular, se elimina mediante difusión desde la sangre hacia la fase gaseosa. El lado izquierdo del corazón recibe sangre oxigenada desde los pulmones, y bombea la sangre hacia el árbol arterial para regar los órganos del cuerpo. Los nutrientes, minerales, vitaminas, y el agua, son introducidos mediante la ingestión de alimentos y líquidos, y absorción en el tracto gastrointestinal (GI). El hígado, que por lo general se considera parte del sistema GI, procesa sustancias absorbidas hacia la sangre desde el tracto GI, y sintetiza también nuevas moléculas, como glucosa a partir de precursores. Los productos de desecho metabólicos se eliminan a través del sistema GI en las heces, y por los riñones en la orina. Los dos controladores integrativos principales del ambiente interno son los sistemas nervioso y endocrino. Cerebro, médula espinal, sistemas sensoriales y nervios conforman el sistema nervioso. El sistema endocrino está constituido de glándulas sin conductos y células secretoras dispersas que se distribuyen en todo el cuerpo, las cuales liberan hormonas hacia la sangre en respuesta a señales metabólicas, hormonales y nerviosas. La función de los sistemas nervioso y endocrino es coordinar la conducta y las interacciones de los sistemas descritos en todo el libro. El agua es la molécula más abundante en el cuerpo; constituye alrededor de 50 a 60% del peso corporal total. Todas las células y órganos existen en un ambiente acuoso. El agua intracelular es el principal componente del citosol. El agua también es el principal componente del líquido extracelular. Este último incluye el líquido intersticial, que baña las células del organismo; el plasma sanguíneo, es el componente líquido de la sangre; el líquido cefalorraquídeo sólo se encuentra en el sistema nervioso central; el líquido sinovial, se encuentra en articulaciones como la rodilla, y la linfa es un líquido que se forma a partir del líquido intersticial, el cual fluye de regreso hacia el sistema circulatorio por medio del sistema linfático. Hay diferencias importantes en la composición de los líquidos intracelular y extracelular, que tienen importancia en varios aspectos de la función celular (cuadro 1-1). FACTORES Y CONCEPTOS FÍSICOS GENERALES No es un accidente que las palabras “fisiología” y “física” provengan de la misma palabra griega physis (“naturaleza”). Es importante que los estudiantes de fisiología entiendan las fuerzas y los factores físicos que rigen la función del cuerpo. CUADRO 1-1 Composición de los líquidos extracelular e intracelular. Concentración extracelular (mM) Concentración intracelular (mM) Na+ 140 12 K+ 5 150 Ca2+ 1 0.0001 Mg2+ 1.5 12 100 7 HCO3 24 10 Aminoácidos 2 8 Glucosa 4.7 1 Proteína 0.2 4 Cl– – Las concentraciones intracelulares son un poco diferentes para distintos tejidos. Las concentraciones de Ca2+ mostradas son los iones libres, biológicamente activos, no unidos a proteínas. El Ca2+ total (unido más libre) es considerablemente más alto en los líquidos extracelular (2.5 mM) e intracelular (1.5 mM). Reproducido con autorización de Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology, 11th ed. McGraw-Hill, 2008. con la cual cada molécula puede pasar por la membrana celular (permeabilidad); no se requiere de manera directa gasto de energía para la difusión, razón por la cual a veces se llama difusión pasiva. También hay transportadores proteínicos, los cuales se ubican en la membrana celular que media la difusión facilitada de moléculas que son demasiado grandes o hidrofílicas como para permear la membrana mediante difusión simple. La difusión facilitada no requiere energía, y mueve moléculas en favor de un gradiente de concentración. En contraste, el transporte activo es un proceso que consta de movimiento de moléculas a través de una membrana celular contra un gradiente de concentración; puede considerarse una bomba que usa energía para hacer trabajo. El movimiento de moléculas de agua a través de la membrana celular también ocurre mediante difusión desde una “concentración” de agua más alta hacia una más baja. Esto se llama ósmosis; el agua se mueve desde un compartimiento con menos partículas osmóticamente activas (concentración más alta de agua) hacia un compartimiento con más partículas osmóticamente activas (concentración más baja de agua). Los ejemplos de partículas osmóticamente activas son iones como sodio, potasio y cloruro, y moléculas orgánicas como glucosa y aminoácidos. AMORTIGUACIÓN Y pH TRANSPORTE DE MEMBRANA Hay varios mecanismos con los cuales las moléculas cruzan la membrana celular, sea para entrar a la célula o para salir de ella. Todos ellos se describen con detalle en la Sección II. El más simple es la difusión, en la que la tasa a la cual una molécula cruza la membrana celular está regida por el gradiente de concentración y la facilidad Una de las variables más controlada en el organismo es la concentración del ion hidrógeno de los líquidos intracelular y extracelular. Esto se debe a que casi todas las proteínas tienen función óptima dentro de un rango de pH muy estrecho. Recuérdese que el pH es el logaritmo negativo (base 10) de la concentración de ion hidrógeno en unidades molares —cuando el pH es bajo, el líquido es ácido; cuando el pH es alto, el líquido es alcalino—. El cuerpo tiene varios mecanismos para mantener un pH normal; éstos se explican en las Secciones CAPÍTULO 1 Conceptos fisiológicos generales VI y VII. El organismo puede deshacerse de ácido al aumentar la eliminación de dióxido de carbono desde los pulmones, lo cual se debe a que el dióxido de carbono y el ion hidrógeno están enlazados por medio de reacciones químicas al bicarbonato, uno de los principales amortiguadores en el cuerpo. Un amortiguador es un compuesto iónico que atenúa cambios del pH al combinarse con iones hidrógeno o liberarlos. Los riñones también pueden eliminar ion hidrógeno del organismo por medio de los complejos procesos involucrados en la producción de orina. Por último, los cambios del pH intracelular y extracelular pueden evitarse con diversos amortiguadores además del bicarbonato. FUERZAS HIDROSTÁTICAS Y PRESIÓN, RESISTENCIA Y ADAPTABILIDAD Presión se define como fuerza por unidad de área. La presión en el fondo de una columna de líquido aumenta con la altura de la columna y depende también de la densidad del líquido y de la gravedad. La presión en cualquier punto en una columna de líquido se llama presión hidrostática, y es la diferencia de presión entre ese punto y la parte superior de la columna. Las diferencias de presión hidrostática tienen muchas consecuencias fisiológicas importantes, en particular en los vasos sanguíneos (Sección V). El flujo de un fluido (un líquido o gas) se cuantifica como el volumen de fluido que se mueve a través de un conducto por unidad de tiempo. Las relaciones entre presión, flujo y la resistencia ofrecida por los conductos a través de los cuales fluye un fluido pueden ser complejas, pero se simplifican como sigue. La tasa de flujo de líquido a través de un tubo es proporcional a la diferencia de presión entre los dos extremos del tubo, e inversamente proporcional a la resistencia al flujo a través del tubo. La resistencia no puede determinarse de manera directa, sino que se calcula a partir de la presión y el flujo. En ausencia de cambio de la resistencia, el incremento de la desigualdad de presión a través de un tubo aumentará el flujo. Si la diferencia de presión de un extremo del tubo al otro no cambia, el incremento de la resistencia disminuirá el flujo. Si el flujo a través del tubo no cambia, GANANCIA NETA HACIA EL CUERPO Alimento Tracto GI Aire Pulmones aumentar la resistencia incrementará la desigualdad de presión entre los extremos del tubo. La diferencia de presión entre los dos extremos del tubo representa la conversión de energía en calor por la fricción interna del fluido con sí mismo y con la pared del conducto. El lector notará que la relación entre presión, flujo y resistencia para líquido que fluye a través de un tubo es análoga a la ley de Ohms para la electricidad, en la cual la disminución de voltaje a través de un circuito (análogo a una caída de presión en el tubo con líquido fluyendo a través de él) es proporcional al producto de la corriente (análoga al flujo) y la resistencia. Casi todos los vasos o cavidades en el cuerpo se distenderán de manera pasiva si la diferencia de presión a través de sus paredes aumenta; esto da lugar a incremento del volumen del vaso. Tal capacidad para distenderse en respuesta a un aumento en la diferencia de presión transmural (a través de la pared) aumentada se llama adaptabilidad. Un término menos específico para adaptabilidad es distensibilidad. El inverso de la adaptabilidad es la elasticidad, por ende, la complianza puede considerarse la resistencia a la distensión cuando la diferencia de presión transmural aumenta, o como la capacidad de un vaso para volver a su volumen original después de que se elimina la diferencia de presión transmural aumentada. Esto se relaciona de manera directa con la ley de Hooke de la elasticidad para resortes mecánicos. EQUILIBRIO DE MASAS Y METABOLISMO A fin de alcanzar el estado estable que define la homeostasis, la cantidad de cualquier sustancia que ingrese al cuerpo debe ser casi igual a la cantidad de la sustancia que sale del cuerpo, más la eliminada por el metabolismo (figura 1-4). El flujo de entrada de una sustancia es la suma de la captación en los pulmones, la absorción en el tracto GI, la síntesis en el organismo (p. ej., síntesis hepática de glucosa a partir de precursores moleculares), y liberación a partir de células (p. ej., liberación de ácidos grasos a partir de tejido adiposo). El flujo de salida de una sustancia es la suma del metabolismo, la captación DISTRIBUCIÓN DENTRO DEL CUERPO PÉRDIDA NETA DESDE EL CUERPO Depósitos de almacenamiento Metabolismo Síntesis en el cuerpo 5 FONDO COMÚN Incorporación reversible hacia otras moléculas Excreción desde el cuerpo por medio de los pulmones, el tracto GI, los riñones, piel, flujo menstrual FIGURA 14 Concepto de equilibrio de masas. El compartimiento central por lo general es líquido extracelular (que incluye plasma sanguíneo); recibe sustancias por ingestión, síntesis y liberación desde células, y pierde sustancias por excreción, metabolismo y captación hacia células. En el estado estable, cuando se dice que una sustancia está “en equilibrio”, la ingesta y la excreción son casi iguales. (Reproducida con autorización de Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology, 11th ed. McGraw-Hill, 2008.) 6 SECCIÓN I Introducción hacia células, las pérdidas por medio del tracto GI, el sistema respiratorio, sudor y excreción urinaria. En el estado estable, la diferencia entre los flujos de entrada y de salida totales debe ser muy cercana a cero. De un minuto a otro, hay grandes diferencias entre el flujo de entrada y el flujo de salida, pero después de días a semanas, cuando la sustancia, por lo general, está en equilibrio, la desigualdad debe estar cerca de cero. Los ejemplos de esto son el equilibrio de sodio descrito en la SecciónVII, y los equilibrios de calcio y fosfato descritos en la Sección IX. EXCITABILIDAD Las células vivas tienen una diferencia de carga eléctrica a través de la membrana celular, creada por diferencias de la concentración de iones y el movimiento de los mismos entre el exterior y el interior de la célula (Secciones II a IV; cuadro 1-1). Como resultado, las membranas tienen un potencial eléctrico en reposo que puede ser cambiado por diversas entradas. Los cambios notorios del flujo de iones a través de la membrana celular llevan a grandes modificaciones del potencial eléctrico que pueden dar por resultado respuestas celulares importantes. Por ejemplo, la contracción muscular descrita en la Sección III se produce por la despolarización de la membrana celular muscular que es transducida hacia una señal química dentro de la célula, la cual lleva a la generación de fuerza y movimiento. SISTEMAS DE CONTROL El enfoque principal de la fisiología es el entendimiento de los mecanismos con los cuales las células, los órganos y los sistemas mantienen la homeostasis. Esto se logra por medio de retroalimentación negativa. El concepto general es que el organismo trata de aumentar una variable cuando está por debajo de su nivel óptimo (lo cual se denomina valor establecido), y disminuye una variable cuando está por arriba de su nivel óptimo. Esto es análogo al termostato que controla la temperatura ambiente al ajustar el calentamiento, el enfriamiento, o ambos, de la habitación. Por ejemplo, si se abre una ventana en un día frío, la temperatura ambiente disminuye desde el valor establecido del termostato; lo anterior se llama perturbación. El termostato contiene un sensor que detecta la diferencia entre la temperatura ambiente y el valor establecido. El termostato emite señales al aparato calefactor para que genere calor, y la temperatura ambiente es devuelta a lo normal. La diferencia entre el punto bajo de la temperatura ambiente y la temperatura ambiente final a estado estable se llama corrección. Puesto que la ventana se deja abierta en este ejemplo, la temperatura ambiente no vuelve por completo al valor establecido; la diferencia restante entre la temperatura ambiente final y su valor establecido termostático se llama error. La capacidad del sistema de control para restituir el sistema a su valor establecido se llama ganancia, que es representada con la ecuación que sigue: Ganancia = INTERACCIONES ENTRE UNA CÉLULA Y OTRA Las células interactúan entre sí localmente (Secciones II a IV, VIII y IX). Un mecanismo es mediante contacto directo entre células a través de uniones intercelulares estrechas (zonas de oclusión) y uniones intercelulares comunicantes (conexiones comunicantes). Otro es la sinapsis, en la cual las neuronas pueden liberar sustancias químicas que se llaman neurotransmisores para alterar la función de una célula vecina. Por último, hay diversas señales químicas mediante las cuales las células pueden comunicarse con células vecinas por medio de difusión. Un ejemplo de esto es la comunicación paracrina con la cual una célula libera factores humorales que se difunden a través del líquido intersticial y se unen a un receptor en una célula vecina dentro del mismo tejido. Punto más bajo FIGURA 15 Hemorragia moderada como un ejemplo de la ganancia de un sistema de control por retroacción. Mientras más alta es la ganancia de un sistema, más capaz es de restituir una variable controlada a su valor establecido en respuesta a una perturbación. Error restante (1) Un ejemplo clásico se presenta en la figura 1-5, que muestra la respuesta del sistema cardiovascular a la pérdida rápida de sangre (hemorragia). En este ejemplo, la pérdida rápida de 1 L de sangre lleva a un decremento de la presión arterial media desde el valor establecido de 100 a 75 mmHg. El sistema cardiovascular tiene sensores que se conocen como barorreceptores, los cuales detectan la presión arterial (capítulo 29). Estos sensores cambian sus aferencias nerviosas hacia el cerebro para activar reflejos sistémicos para restituir la presión arterial a lo normal. En este ejemplo, estos reflejos restituyen la presión arterial a 95 mmHg; por consiguiente, la corrección es de 20 mmHg, y el error restante es de 5 mmHg. Al usar la ecuación (1), se proporciona una ganancia de alrededor de 4. Aunque los médicos por lo general no calculan la ganancia cuando atienden a pacientes, es una manera conveniente de pensar acerca de la capacidad de los Pérdida rápida de sangre Presión arterial final 100 95 100 – 95 = 5 Error restante Perturbación 95 – 75 = 20 Corrección original debida a reflejos 75 Presión arterial media (mmHg) Valor establecido Corrección Ganancia = Corrección Error restante = 20 = 4 5 Tiempo (min) CAPÍTULO 1 Conceptos fisiológicos generales reflejos para restituir a lo normal, por medio de retroalimentación negativa, un sistema perturbado. Mientras mayor es la ganancia, más alta es la proporción entre la corrección y el error restante, y mejor es el sistema de control para restituir el sistema a su valor establecido; así, por ejemplo, el control de la temperatura corporal tiene una ganancia muy alta (capítulo 70). Muchos sistemas de retroalimentación incluyen un cambio de conducta. Por ejemplo, beber agua extra cuando el volumen sanguíneo está disminuido ayuda a restituir el volumen plasmático. Ponerse ropa caliente y acurrucarse ayuda a minimizar la pérdida de calor en un ambiente frío; por último, los valores establecidos de sistemas de control pueden cambiar. Los ejemplos de esto son reajustar el valor establecido del barorreceptor durante aumentos crónicos de la presión arterial (hipertensión) (capítulo 29), y durante la aclimatación a la baja de oxígeno ambiente, característica de la altitud elevada (hipoxia) (capítulo 71). Aunque casi todos los sistemas de control del organismo son retroalimentación negativa, hay algunos ejemplos de retroalimentación positiva, que son asas de retroalimentación que se amplifican por sí mismas. En el capítulo 68 se presentan varios ejemplos de esto. Uno es la estimulación de la hormona de la parte anterior del hipófisis, la hormona luteinizante (LH), por estrógeno justo antes de la ovulación, que causa un aumento grande de la LH, que después estimula más liberación de estrógeno, y así sucesivamente. Otro ejemplo es el parto, durante el cual la distensión del cuello del útero estimula la liberación de oxitocina desde la parte posterior de la hipófisis que, a su vez, estimula contracciones uterinas más fuertes. Esto causa distensión cervical adicional, más liberación de oxitocina, y mayores contracciones del útero. La retroalimentación positiva también causa efectos perjudiciales en el cuerpo. Un ejemplo es la insuficiencia cardiaca, durante la cual el bombeo del corazón se reduce debido a, por ejemplo, una infección del músculo cardiaco. La disminución resultante de la presión arterial lleva a reflejos que estimulan al corazón para que bombee más fuerte en un esfuerzo por aumentar la presión arterial. Tal estrés adicional sobre el corazón en realidad hace que funcione menos bien, y la insuficiencia cardiaca se alimenta por sí misma. Otro concepto importante en el control homeostático es la potenciación, lo cual tiene lugar cuando una sustancia incrementa la respuesta a otra sustancia, aun cuando la primera sustancia no ejerce una respuesta importante por sí misma. Un ejemplo de esto es la liberación de las hormonas GI desde el intestino en respuesta a una comida (capítulos 49 y 66). Dichas hormonas pueden potenciar la respuesta pancreática de insulina a la glucosa absorbida; ese es un ejemplo de potenciación por anteroacción, porque estas hormonas GI “anuncian” el aumento inminente de la glucosa en sangre antes de que en realidad ocurra la absorción de glucosa en el intestino delgado. Cuando la glucosa llega por medio del torrente sanguíneo al páncreas, hay una respuesta de insulina potenciada, de modo que se evita la hiperglucemia. RESUMEN DEL CAPÍTULO ■ ■ La célula está rodeada por una membrana que regula la composición intracelular y el flujo de moléculas hacia adentro de la célula y hacia afuera de la misma. El agua es la molécula más abundante en el cuerpo, y su concentración y equilibrio están altamente regulados. ■ ■ ■ ■ ■ 7 Hay gradientes de concentración importantes entre los líquidos intracelular y extracelular para sodio, potasio, calcio, magnesio, cloruro y bicarbonato, así como para compuestos orgánicos. Las moléculas pueden entrar a la célula mediante difusión pasiva, y por medio de transportadores que no usan energía celular (transporte facilitado) y que usan de manera directa dicha energía (transporte activo). La tasa de flujo de un líquido a través de un tubo está determinada por la diferencia de presión entre el flujo de entrada y el flujo de salida, y por la resistencia del tubo al flujo. Las sustancias más importantes en el cuerpo están en equilibrio; los flujos de entrada y de salida son casi iguales con el tiempo. Casi todos los sistemas están controlados mediante retroalimentación; la variable controlada es capaz de suspender su propia liberación de modo muy parecido a la manera en que un termostato controla la temperatura ambiente. PREGUNTAS DE ESTUDIO 1. ¿Cuál de los organelos que siguen es el principal responsable de la generación de energía? A) aparato de Golgi B) mitocondrias C) lisosomas D) ribosomas 2. ¿Cuál de los siguientes tiene una concentración mucho más alta en el líquido intracelular que en el extracelular? A) ion sodio B) ion cloruro C) glucosa D) ion potasio 3. ¿Cuál de los siguientes daría lugar a un incremento del flujo de un líquido a través de un tubo? A) aumento de la resistencia B) aumento de la presión en el extremo de salida del tubo C) aumento de la presión en el extremo del flujo de entrada del tubo D) aumento de la longitud del tubo 4. ¿Cuál de los siguientes tiene la ganancia de retroacción más alta? A) presión arterial inicial = 100; punto bajo de presión arterial = 70; presión arterial final después de corrección por retroalimentación = 90 B) temperatura corporal inicial = 37.2 °C; punto alto de temperatura corporal = 38.9 °C; temperatura corporal final después de corrección por retroalimentación = 37.4 °C C) glucosa sanguínea inicial = 80 mg/dL; punto alto de glucosa en sangre = 110 mg/dL; glucosa en sangre final después de corrección por retroacción = 85 mg/dL D) osmolalidad plasmática inicial = 280 mOsm/kg; punto bajo de osmolalidad plasmática = 270 mOsm/kg; osmolalidad final después de corrección por retroacción = 278 mOsm/kg SECCIÓN II FISIOLOGÍA CELULAR Células y mecanismos celulares C A P Í T U L O 2 David Landowne O B J E T I V O S ■ ■ ■ Reconocer los tipos de eventos electrofisiológicos, y describirlos. Describir los tipos de canales de membrana y sus funciones. Describir sistemas fisiológicos de control. entradas, procesos y salidas (figura 2-1). Los procesos complejos pueden fragmentarse en procesos más simples; las salidas de uno o más procesos se convierten en las entradas del siguiente. Para estudiar los procesos que se comentan aquí, es útil considerar un modelo de tres células del cuerpo. En la figura 2-2 se muestran una neurona (o célula nerviosa) sensorial, una neurona motora, y una célula de músculo esquelético. Estas células representan el soporte físico que el cuerpo usa para llevar a cabo los mecanismos celulares dinámicos descritos en el párrafo previo. Las células tienen porciones especializadas para los diferentes procesos. Empezando desde la izquierda, la célula sensorial tiene un extremo que se especializa en la transducción de un estímulo hacia una señal celular. Los diversos sentidos tienen diferentes especialidades para lograr esta transducción. Además de los cinco sentidos clásicos (tacto, audición, visión, gusto y olfato), hay sensores o propioceptores dentro del organismo que detectan parámetros internos —por ejemplo, la temperatura corporal, la presión arterial, la concentración de oxígeno en sangre, o la longitud de diversos músculos. Si es suficientemente grande, la señal inicial hace que otra señal se propague por el axón (la porción cilíndrica larga de la célula nerviosa) hasta que llega al otro extremo, donde la neurona sensorial hace una conexión sináptica con dendritas de la neurona motora, que se ubica en el sistema nervioso central (SNC). El mensaje se INTRODUCCIÓN La vida es celular, y las células son las unidades fundamentales de la vida. Sin células, no habría seres vivos. Todas las células de un individuo dado se derivan de un huevo fecundado único. Casi todas las células de organismos multicelulares residen dentro de sus tejidos y órganos. Este capítulo se centra en los mecanismos celulares, y deja la discusión de su organización superior a capítulos dedicados a los diversos sistemas. Se introducen fármacos, toxinas y enfermedades para ilustrar los mecanismos celulares. COMUNICACIÓN Mecanismos celulares dinámicos apoyan la percepción sensorial del ambiente, la comunicación y la integración de información dentro de células y entre las mismas, así como su expresión, o acciones sobre el ambiente. Se trata de los procesos que permiten a la célula contribuir al funcionamiento de tejidos, órganos e individuos. Tales mecanismos constituyen uno de los fenómenos de las células —la excitabilidad—. Los otros, la reproducción y el metabolismo, no se cubren a profundidad aquí. La percepción, integración y expresión pueden considerarse mejores eventos fisiológicos en términos de 9 10 SECCIÓN II Fisiología celular Entrada Salida Proceso FIGURA 21 El marco estructural entrada-proceso-salida es una especificación de relación causal en un sistema. Los sistemas complejos pueden considerarse compuestos de unidades simples. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) transmite de la célula presináptica a la célula postsináptica, donde se integra o se evalúa junto con otros mensajes provenientes de otras neuronas que hacen sinapsis en la misma neurona motora. En el organismo completo, esta integración y comparación ocurren en muchas células a distintos niveles dentro del SNC, de modo que la decisión de moverse o no moverse se puede tomar al considerar más de una aferencia, y cualquier cosa que el organismo haya aprendido en el pasado. Si la neurona motora es excitada lo suficiente, enviará otro mensaje a lo largo del axón que va a una sinapsis en una célula muscular. En personas sanas, esta sinapsis neuromuscular lleva a una señal que se propaga a todo lo largo de la célula muscular y activa la contracción, que puede actuar sobre el ambiente. Otras acciones sobre el ambiente son efectuadas por las secreciones de diversas glándulas; éstas también pueden estar controladas por conexiones sinápticas. Estos músculos y glándulas pueden actuar internamente (p. ej., para controlar la frecuencia cardiaca o la presión arterial) o externamente (para locomoción o comunicación con otras personas). Todas estas señales son eléctricas; representan cambios de la diferencia de potencial eléctrico a través de las diversas membranas celulares. Cada célula viva tiene una membrana de superficie que separa sus espacios intracelular y extracelular. Todas las células, no sólo las de nervios y músculos, son eléctricamente negativas en su interior respecto del exterior. Esto se llama potencial de membrana. Cuando las células están “en reposo” —es decir, no emiten señales— su potencial de membrana se llama potencial de reposo. En el capítulo 4 se abordan los orígenes del potencial de reposo. Aun cuando las señales antes descritas son cambios de potencial, por lo general se denominan potenciales nombrados. En la parte izquierda de la figura 2-2 se muestra un potencial generador sensorial, que tiene dos propiedades para distinguirlo de la siguiente señal, el potencial de acción. El potencial generador sensorial es local; sólo ocurre dentro de algunos milímetros de la terminación sensorial. El potencial de acción es propagado; viaja de la terminación sensorial a la terminal presináptica, quizás a más de 1 m de distancia. El potencial generador sensorial también es graduado; un estímulo de mayor amplitud produce un potencial generador sensorial de mayor amplitud. En contraste, el potencial de acción tiene amplitud y duración estereotipadas; es una respuesta de todo o nada. La información acerca del estímulo está codificada en el número de potenciales de acción, o el número por segundo. Un estímulo de mayor amplitud dará lugar a una frecuencia más alta de potenciales de acción, cada uno con la misma amplitud estereotipada. Dado que la característica de todo o nada de las neuronas es similar a la característica de cierto o falso de las proposiciones lógicas, los especialistas en cibernética (quienes estudian el control y la comunicación en el animal y en la máquina) consideran que los eventos neurales y las relaciones entre ellos pueden tratarse mediante lógica proposicional. Los capítulos 5 y 6 se dedican a potenciales generadores sensoriales y potenciales de acción, respectivamente. Soporte físico Terminación sensorial Axón Sinapsis Axón Músculo Señales (potenciales) Generador sensorial Acción Sináptico Acción Placa terminal Local Propagado Local Propagado Local Graduado Todo o nada Graduado Todo o nada Graduado Canales Mecanosensible Sensible a voltaje Quimiosensible Sensible a voltaje Quimiosensible Cibernética Entrada Transmisión Proceso Transmisión Salida FIGURA 22 Los mecanismos celulares de un organismo de tres células hipotético. Diferentes tipos de canales sustentan distintos mecanismos fisiológicos que apoyan las funciones de entrada-proceso-salida, de animales, incluso seres humanos. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) CAPÍTULO 2 Células y mecanismos celulares Las terminales presinápticas contienen un mecanismo para liberar el contenido de vesículas que incluyen transmisores químicos que se difunden a través de la hendidura sináptica estrecha y reaccionan con la célula postsináptica para producir un potencial postsináptico, éste también es local y graduado. Sólo se observa dentro de algunos milímetros del sitio de la terminación presináptica, y su amplitud depende de qué tanto transmisor se libera. Hay potenciales postsinápticos excitadores (EPSP) y potenciales postsinápticos inhibidores (IPSP), dependiendo de si el potencial postsináptico hace que la célula tenga más o menos probabilidades de iniciar un potencial de acción. Si hay suficiente excitación como para abrumar cualquier inhibición que pueda estar ocurriendo, se iniciará un potencial de acción en la célula postsináptica. Hay muchas células presinápticas que terminan en cada neurona postsináptica, así como diferentes neurotransmisores en distintas sinapsis. Estos transmisores, el mecanismo de liberación, y los potenciales postsinápticos resultantes, se comentan en el capítulo 7. El potencial de acción en la neurona motora y la sinapsis con el músculo son similares a la sinapsis entre una neurona y otra antes comentada. Al microscopio óptico, la unión neuromuscular tiene el aspecto de una pequeña placa; de ahí que la unión a menudo se denomine una placa terminal, y el potencial postsináptico un potencial de placa terminal. La unión neuromuscular difiere de casi todas las otras sinapsis porque sólo hay una célula presináptica, su efecto siempre es excitador y —en personas sanas— el potencial de placa terminal siempre es suficientemente grande como para iniciar un potencial de acción en la célula muscular. El potencial de acción muscular se propaga a lo largo de la célula y hacia el interior mediante túbulos transversos pequeños, cuyas membranas son continuas con la membrana celular. La excitación del potencial de acción está acoplada a la contracción muscular mediante procesos descritos en el capítulo 10, donde también se comenta el control de las células de músculo cardiaco y liso. El potencial de reposo, los potenciales generadores sensoriales, los potenciales de acción y los potenciales sinápticos ocurren mediante la abertura y el cierre de canales en las membranas celulares. Estos canales están hechos de proteínas, las cuales están embebidas en la membrana que conecta los espacios intracelular y extracelular, y que la abarcan. Cada una tiene un pequeño poro en la parte central que se puede abrir o cerrar, y que es lo bastante grande como para permitir que iones específicos fluyan a través de ellos, al tiempo que es lo bastante pequeño como para evitar que metabolitos y proteínas fluyan hacia afuera de la célula. Hay muchos canales, y buena parte del capítulo 3 se dedica a su descripción. En general, se denominan por el ion que pasa a través de ellos o por el agente que hace que se abran. Hay tres clases de canales que actúan para producir los cambios de potencial descritos en la figura 2-2. Todos estos canales se comentarán de manera individual en el capítulo 3, y después de nuevo en el contexto de los diversos potenciales en capítulos subsiguientes. Los canales sensibles a la deformación mecánica ayudan a las sensaciones de tacto y audición, y a los muchos propioceptores que proporcionan información sobre la longitud de músculos, la tensión de músculos, la posición de articulaciones, la orientación y la aceleración angular de la cabeza, y la presión arterial. Estos canales se abren cuando la membrana de la terminación sensorial está distendida, fluyen iones sodio a través de los canales, y el potencial de membrana cambia. 11 Los canales sensibles a voltaje sustentan los potenciales de acción. Se abren en respuesta a un cambio del potencial de membrana. Cuando están abiertos, fluyen iones a través de ellos, y esto también cambia el potencial de membrana. El potencial generador o los potenciales sinápticos activan estos canales, y después ellos abren los canales restantes sensibles a voltaje adyacentes. Esto explica la propagación y la calidad estereotipada, de todo o nada, de los potenciales de acción. Los potenciales de acción de nervios y de músculo esquelético los produce la activación sucesiva de canales de sodio sensibles a voltaje, seguidos por canales de potasio sensibles a voltaje. También hay canales de calcio sensibles a voltaje en las terminaciones nerviosas presinápticas. Cuando el potencial de acción llega a la terminal presináptica, estos canales de calcio se abren y permiten que entre calcio a la célula. El calcio se une a componentes intracelulares e inicia la liberación de transmisores sinápticos. Los potenciales sinápticos dependen de canales quimiosensitivos. Los transmisores se unen a estos canales, lo que hace que se abran. Hay diferentes canales para distintos transmisores, y diferentes canales para EPSP e IPSP. Los canales quimiosensitivos también ayudan a los sentidos químicos del olfato y el gusto. Asimismo, hay canales que se abren o se cierran en respuesta a sustancias químicas intracelulares como el trifosfato de adenosina (ATP) o los nucleótidos cíclicos, monofosfato de adenosina cíclico (cAMP) o monofosfato de guanosina cíclico (cGMP). La visión es apoyada por una serie de reacciones por las cuales la absorción de luz lleva a un decremento de cGMP, que produce un cierre de canales sensibles a nucleótido cíclico (quimiosensitivos). Cuando iones sodio dejan de fluir por estos canales, el potencial de membrana cambia. Desde un punto de vista cibernético, la figura 2-2 indica que el cuerpo tiene mecanismos para dar entrada a información, transmitirla dentro del cuerpo, procesarla y proporcionar salida; este tipo de análisis aparece con frecuencia en fisiología. Gran parte de lo que el lector aprenderá puede fraccionarse en diversos pasos, donde la salida de un proceso se torna en la entrada para el siguiente. Por ejemplo, los potenciales generadores sensoriales son una entrada para el proceso de generación de potencial de acción, y el potencial de acción es la entrada para el canal de calcio sensible a voltaje, que permite que entre calcio a la terminal presináptica. Este calcio es la entrada para el proceso de liberación de transmisor, y así sucesivamente. CONTROL Si bien casi todo este libro se centra en aislar los diferentes procesos de modo que sea más fácil analizarlos, un entendimiento del valor y la importancia verdadera de cada cualidad fisiológica debe referirse al organismo entero. Un tema recurrente en toda la fisiología es el mantenimiento de un ambiente interno estable por medio de homeostasis. Muchas propiedades internas (p. ej., la temperatura corporal o la concentración de glucosa en sangre) están controladas de manera homeostática dentro de límites estrechos por medio de sistemas de control por retroacción. La homeostasis es una propiedad de muchos sistemas abiertos complejos. El control por retroalimentación es la característica fundamental de la actividad organizada. Un sistema homeostático (p. ej., una célula, el cuerpo, un ecosistema) es un sistema abierto que se mantiene a sí mismo al controlar muchos equilibrios dinámicos. El sistema mantiene su equilibrio interno al reaccionar a cambios en 12 SECCIÓN II Fisiología celular Valor deseado Comparador Efector Parámetro controlado Sensor A Desde centros superiores Motoneurona Músculo Neurona sensorial Huso muscular B FIGURA 23 Homeostasis y control por retroalimentación. A y B) Al tener entradas que detectan la salida y alimentan información de regreso al controlador, las máquinas y los seres humanos pueden tener control de sus condiciones de operación. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) el ambiente con respuestas de dirección opuesta a las que crearon la alteración. El equilibrio se mantiene por medio de retroalimentación negativa. Quizá el sistema de control por retroalimentación negativa más familiar es el termostato, que controla la temperatura de una habitación (capítulo 1). Los pasos básicos (figura 2-3A) en el control por retroalimentación negativa de cualquier parámetro medible son la medición con un sensor, comunicación de esa medición a un comparador, hacer la comparación, y comunicar la comparación a un efector que cambia el parámetro de interés. La retroalimentación se llama negativa porque la señal al efector va en la dirección opuesta a cualquier alteración, y reduce la diferencia entre el valor medido y el valor deseado. Las tres células en la figura 2-2, dispuestas como un asa de retroalimentación negativa (figura 2-3B), representan el proceso que se usa para controlar la longitud de los músculos tanto para mantener la postura como para lograr movimiento en respuesta a señales que provienen del cerebro. Esta asa de retroalimentación puede demostrarse por medio del reflejo de estiramiento —es decir, el reflejo rotuliano (capítulo 14)—. Cuando se estira un músculo, se abren canales sensibles a la deformación mecánica en órganos sensoriales, lo cual cambia potenciales de membrana en las terminaciones sensoriales que inducen la propagación de potenciales de acción hacia la médula espinal. Se libera transmisor, que excita el nervio que va de regreso al músculo, donde el proceso sináptico se repite y el músculo se acorta para compensar el estiramiento inicial. Hay algunos sistemas de retroalimentación positiva que tienen importancia fisiológica. Un sistema de retroalimentación positiva es inestable; la señal que proviene del sensor aumenta el efecto, lo que incrementa la señal desde el sensor en un “círculo vicioso”, que sólo se limita por la disponibilidad de recursos. La propiedad de todo o nada de los potenciales de acción se debe a un asa de retroalimentación positiva. En el capítulo 1 se describieron algunos ejemplos de retroalimentación positiva. RESUMEN DEL CAPÍTULO ■ ■ La comunicación en células excitables ocurre por medio de señales eléctricas dentro de las células y mediante señales químicas en sinapsis entre las células. Hay dos clases de señales eléctricas: las que son locales y graduadas y las que son propagadas y estereotipadas, o de todo o nada. CAPÍTULO 2 Células y mecanismos celulares ■ ■ ■ ■ Los transmisores químicos son liberados en el espacio presináptico, y producen una señal eléctrica en la célula postsináptica. Tres clases de canales iónicos producen las señales eléctricas: canales sensibles a la deformación mecánica, quimiosensitivos y sensibles a voltaje. La homeostasis mediante control por retroacción negativa es una característica importante de los sistemas vivos. Un asa de retroalimentación negativa tiene tres elementos básicos: un sensor, un comparador, un efector y dos enlaces de comunicación que los conectan. PREGUNTAS DE ESTUDIO 1. ¿Cuál de los cambios que siguen en el potencial eléctrico requiere canales sensibles a voltaje? A) potenciales sinápticos excitadores B) potenciales generadores sensoriales mecánicos C) potenciales de acción propagados 13 D) potenciales generadores sensoriales a la luz E) potenciales sinápticos inhibidores 2. Los sistemas de control por retroalimentación negativa no A) mejoran la confiabilidad del control. B) requieren la detección o medición del proceso controlado. C) requieren comunicación entre partes separadas del sistema. D) regulan la presión arterial y la temperatura corporal. E) causan la propiedad de todo o nada del potencial de acción. Membranas celulares y mecanismos de transporte C A P Í T U L O 3 David Landowne O B J E T I V O S ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ Comprender la composición molecular de membranas biológicas. Definir las propiedades funcionales biofísicas de membranas biológicas. Describir clases de canales iónicos, su estructura molecular y sus propiedades biofísicas. Detallar la organización molecular, las propiedades, el control y los papeles funcionales de canales célula-célula. Explicar el movimiento y transporte de sustancias a través de membranas biológicas mediante procesos pasivos. Describir el movimiento y transporte de sustancias a través de membranas biológicas por medio de procesos activos. Entender la importancia fisiológica de dos ejemplos de transporte activo y dos ejemplos de transporte pasivo. Definir presión osmótica. Calcular la osmolaridad de soluciones simples. Estimar los cambios de la osmolaridad en compartimientos corporales causados por beber diversas soluciones simples. Describir los mecanismos fisiológicos para regular la osmolaridad. Toda célula viva tiene una membrana de superficie que define sus límites y la conectividad de los compartimientos intracelular y extracelular. Las membranas celulares miden alrededor de 10 nm de grosor, y constan de una bicapa lipídica de 3 a 4 nm de grosor, con diversas proteínas embebidas que pueden sobresalir hacia uno u otro compartimiento (véase figura 1-2). Las membranas también delimitan organelos intracelulares, incluso la envoltura nuclear, el aparato de Golgi, el retículo endoplasmático (ER), las mitocondrias, y diversas vesículas (véase figura 1-1). La bicapa lipídica es impermeable a sustancias cargadas o polares. Las proteínas manejan el transporte de iones o moléculas específicos a través de las membranas y, así, controlan la composición de diferentes soluciones a ambos lados. Apoyan la comunicación a través de las membranas y a lo largo de la superficie de la célula, y proporcionan acoplamiento mecánico entre células. LÍPIDOS Casi todos los lípidos de membrana son glicerofosfolípidos, que tienen un esqueleto glicerol con dos de sus tres grupos —OH esterificados por ácidos grasos, y el tercero esterificado a un grupo fosfato que, a su vez, es esterificado a una molécula pequeña que da su nombre a toda la molécula (figura 3-1). Los glicerofosfolípidos más comunes son la fosfatidilcolina (PC), la fosfatidiletanolamina (PE), y la fosfatidilserina (PS). Las membranas también contienen fosfatidilinositol (PI), que desempeña un papel importante en la emisión de señales dentro del citoplasma. Nótese que la PS y el PI tienen una carga negativa neta. Las membranas de células animales también pueden contener esfingolípidos, incluso fosfoesfingolípido, esfingomielina, que tienen dos cadenas acilo y una cabeza de colina enlazada a fosfato, unida a un esqueleto de serina, y glucoes- 15 16 SECCIÓN II Fisiología celular R O O P O– O + N(CH3)3 HCH Fosfatidilcolina (PC) HCH O O C=O C=O + NH3 HCH Fosfatidiletanolamina (PE) HCH + NH3 COO– Fosfatidilserina (PS) HCH OH OH OH HO OH Fosfatidilinositol (PI) FIGURA 31 Glicerofosfolípidos. Junto con el colesterol, forman la bicapa que separa el interior de las células, y apoya las proteínas de membrana embebidas. (Modificada con autorización de Landowne D: Cell Physiology, New York: Lange Medical Books/McGraw-Hill, 2006.) fingolípidos, los cuales tienen azúcares en el grupo cabeza. Las membranas también contienen colesterol, mismo que tiene una estructura en anillo esteroide. Todos estos lípidos son anfipáticos porque tienen grupos cabeza hidrofílicos, que “aman el agua”, y colas acilo hidrofóbicas, que “temen al agua”. El grupo —OH del colesterol es hidrofílico, y el resto es hidrofóbico. Un efecto hidrofóbico surge por la falta de interacciones de hidrocarburos con agua y la fuerte atracción del agua por sí misma. Así, cuando se colocan en un ambiente acuoso, estos lípidos se montan de manera espontánea hacia vesículas de membrana bicapa cerradas. Los lípidos están relativamente libres para difundirse en dirección lateral dentro del plano de las membranas, pero —con la excepción del colesterol— tienen pocas probabilidades de bascular (flip-flop) desde una mitad de la bicapa hacia la otra debido a la hidrofilicidad de los grupos cabeza. La bicapa es asimétrica; los fosfolípidos que contienen colina, PC y esfingomielina están en la mitad externa, y los que contienen amino, PE y PS, en la mitad interna. Además, los glucoesfingolípidos están en la mitad no citoplasmática, y el PI está mirando hacia el citoplasma. La disposición asimétrica se produce a medida que las membranas son ensambladas en el ER. Los fosfolípidos son sintetizados e insertados en el lado citoplasmático de la membrana; a continuación un translocador de fosfolípidos o “flipasa” transfiere PC al lado no citoplasmático. La esfingomielina y los glucoesfingolípidos son producidos en el aparato de Golgi en el lado no citoplasmático. La facilidad de difusión lateral, o fluidez de membrana, aumenta por la presencia de insaturación o dobles enlaces en las colas de hidrocarburo. Esto forma un codo en la cola y, por ende, empaque menos apretado. A las concentraciones que por lo general se encuentran en membranas biológicas, el colesterol reduce la fluidez debido a su estructura en anillo rígida. Los grupos cabeza glucoesfingolípidos tienden a asociarse uno con otro y reducen la fluidez. Las interacciones de lípidos y proteínas también pueden reducir la fluidez. Hay microdominios colesterol-esfingolípido, o “balsas de lípido” involucrados en el tráfico intracelular de proteínas y lípidos. PROTEÍNAS Las proteínas intrínsecas de la membrana apoyan el movimiento selectivo de iones y moléculas pequeñas desde un lado de la membrana hacia el otro, detectan un ligando en un lado de la membrana y transmiten una señal al otro lado, y proporcionan enlace mecánico para otras proteínas a ambos lados de la membrana. Las proteínas que mueven materiales a través de la membrana pueden dividirse, desde el punto de vista funcional, en canales, bombas y transportadores. Los canales pueden ser específicos y se pueden abrir y cerrar, pero, cuando están abiertos, facilitan el movimiento de materiales sólo en favor de sus gradientes electroquímicos. Los canales iónicos controlan el flujo de corriente eléctrica a través de la membrana. Las bombas mueven iones en contra de su gradiente electroquímico a CAPÍTULO 3 Membranas celulares y mecanismos de transporte expensas del consumo de ATP. Las bombas mantienen los gradientes que permiten a los canales y los transportadores hacer su trabajo. Los transportadores pueden enlazar el movimiento de dos (o más) sustancias, y pueden mover una de ellas contra su gradiente a expensas de mover la otra en favor de su gradiente. Una proteína es el producto de la traducción de un gen; es una secuencia enlazada y plegada de aminoácidos α elegidos a partir de una paleta con 20 posibles cadenas laterales diferentes. El enlace peptídico entre aminoácidos —CO—NH— tiene una transconformación planar; el plegamiento ocurre de acuerdo con los ángulos de torsión entre el grupo amino y el carbono α (Φ), y entre el carbono α y el grupo carboxilo (ψ). El eje de la hélice α y la hoja β son estructuras secundarias, con ángulos de torsión particulares, que se encuentran en proteínas. La conformación o estructura terciaria de toda la proteína es la relación tridimensional de todos sus átomos. Las proteínas tienen regiones de diversas estructuras secundarias conectadas por enlazadores con estructura menos fácil de caracterizar. Casi todas las proteínas que se comentan en este libro tienen más de una conformación. Así, por ejemplo, un canal puede estar abierto o cerrado. Las estructuras secundarias locales no cambian mucho durante estos cambios conformacionales; más bien el cambio ocurre en la relación entre porciones de mayor tamaño de la molécula. También hay un nivel de organización supramolecular o cuaternario. Algunos canales están hechos de una cadena polipeptídica única, mientras que otros de 4 a 6 cadenas. Muchos canales también tienen proteínas accesorias que modulan su función. Además, la matriz lipídica impone restricciones estructurales sobre las proteínas embebidas. En general, las proteínas son anfipáticas y tienen regiones que son más hidrofóbicas o hidrofílicas, dependiendo de la naturaleza de las cadenas laterales. Las proteínas de membrana ya citadas tienen uno o más segmentos helicoidales α transmembrana (TM) con cadenas laterales hidrofóbicas en contacto con el hidrocarburo del lípido. Si hay más de una hélice involucrada, es posible que tenga residuos hidrofóbicos que miran hacia el lípido y otros grupos que se miran uno a otro en las partes más interiores de la proteína. El patrón general es que la proteína cruce varias veces la membrana, con asas intracelulares y extracelulares entre los segmentos TM. También hay una región N terminal antes del primer segmento y una región C terminal después del último; en la figura 3-3 se muestra un ejemplo. La región N terminal puede estar en uno u otro lado, pero la región C terminal, por lo general, es citoplasmática. Una u otra región terminal, o ambas, pueden ser bastante grandes en comparación con las regiones TM. El plegado TM ocurre conforme la proteína es sintetizada en el ER. Las porciones no citoplasmáticas de la proteína pueden ser glucosiladas en el aparato de Golgi antes de que sean insertadas en la membrana celular. El montaje de la subunidad también puede ocurrir en el ER o en el aparato de Golgi. Para casi todas las proteínas de membrana, sólo se conoce la secuencia primaria. La estructura secundaria puede predecirse mediante análisis de secuencia. La presencia de hélices hidrofóbicas putativas de suficiente longitud se toma como una sugerencia de un segmento TM. Es posible predecir propiedades topológicas o patrón de asas y segmentos TM; esa predicción se ha probado para muchas proteínas al preparar anticuerpos para las porciones extracelulares putativas. Análisis de secuencia de genomas enteros sugieren que 17 alrededor de 20% de las proteínas contiene uno o más segmentos TM y, así, son proteínas de membrana. Sólo algunas proteínas de membrana se cristalizaron y sujetaron a análisis de difracción de rayos X. Estos cristales deben incluir moléculas de lípido o detergente para satisfacer las necesidades hidrofóbicas de los segmentos TM. Casi todas las estructuras resueltas son de proteínas bacterianas que se modificaron genéticamente para aumentar la cristalización. Una homología de secuencia fuerte entre la molécula cristalizada y parte de la proteína del ser humano se toma para indicar que tienen estructuras similares. Los canales, las bombas, los transportadores, receptores y moléculas de adhesión celular vienen en muchas variedades para desempeñar varias funciones. En las cinco secciones que siguen se describirán una clasificación taxonómica y las características anatómicas de ejemplos de cada clase funcional. Quizá sea útil volver a esta sección mientras se lee la última parte de este capítulo y las partes del resto del libro en donde se describe la función de estas moléculas en procesos fisiológicos. CANALES En el capítulo previo, los canales se distinguieron por el mecanismo mediante el cual se abren. Hay canales mecanosensitivos involucrados en procesos sensoriales, canales sensibles a voltaje que participan en la propagación de potenciales de acción, y canales quimiosensitivos comprendidos en la transmisión sináptica. También hay canales que por lo general están abiertos, como los canales que mantienen el potencial de reposo, los canales de agua y canales célula-célula especializados que conectan el citoplasma de una célula con el de otra. En esta sección se describen algunos canales que apoyan diversos mecanismos celulares que se comentarán más adelante en el libro. Esto no es exhaustivo; hay muchos canales y clases de canales que no se mencionan. La actual es una “edad de oro” en el estudio de canales iónicos. La electrofisiología y las biologías molecular y estructural revelan algunas proteínas de membrana asombrosas. Muchos canales iónicos son selectivos y se nombran de acuerdo con el ion que pasa a través de ellos. El primer canal que se cristalizó es el de potasio de potencial de reposo, también conocido como rectificador interno o hacia adentro, o Kir. La razón de este nombre se comenta en el capítulo siguiente, junto con su función. El Kir es un tetrámero con cuatro subunidades idénticas, dispuestas con simetría radial, y un poro que permite el flujo iónico en el eje (figura 3-2A). Cada monómero tiene dos segmentos TM con un asa P extracelular en medio (figura 3-2B; figura 3-4, segmentos 5 y 6). Las cuatro asas P se vuelven a sumergir en la membrana, y juntas forman el revestimiento de un poro que entra hasta alrededor de una tercera parte de la membrana. Este poro se vacía hacia una cavidad intramembranosa de mayor tamaño que se comunica con el espacio citoplasmático. Las ocho hélices forman una pared para la cavidad y rodean también las asas P insertadas. Las hélices TM forman una estructura cónica con la punta hacia el citoplasma. La selectividad del poro para iones potasio depende de los aminoácidos específicos que forman el revestimiento. VGYGD es la secuencia característica del canal de K (figura 3-2C); se encontró en canales de K de más de 200 organismos. Esta porción de la molécula es el filtro de selectividad porque acepta iones K+ y excluye otros iones. El poro se reviste con los grupos oxígeno carbonilo; éstos 18 SECCIÓN II Fisiología celular A B C FIGURA 32 La estructura cristalina de un canal de K rectificador interno (Kir). A) Vista superior de una representación de estructura de cintas, con varillas y esferas para las secuencias GYG. B) Vista lateral con eliminación de dos monómeros; la secuencia GYG es una representación de llenado de espacio. C) Acercamiento de dos secuencias VGYGD y un ion. (Modificada con autorización de Landowne D: Cell Physiology, New York: Lange Medical Books/ McGraw-Hill, 2006.) están en la misma relación entre sí que el oxígeno de las moléculas de agua que coordinan iones K+ en solución debido a su carga positiva y la electronegatividad del oxígeno. En la figura 3-2 se observan dos de los oxígenos coordinadores provenientes de glicinas, justo por debajo de las tirosinas. Iones con cargas o radios diferentes coordinarán agua de manera distinta y, así, tendrán menos probabilidades que los iones K+ de abandonar el agua y entrar al canal de K. En la figura 3-2 se muestra una estructura que se cree que representa un canal de Kir cerrado. Se ha resuelto la estructura de otro canal 2-TM procarionte; sus hélices internas están flexionadas y separadas, lo que crea una vía de entrada amplia. Este segundo canal Kir responde a Ca2+ en su lado intracelular al aumentar su probabilidad de abrirse. El Ca2+ se une al dominio regulador de conductancia de K (RCK) en la parte C terminal de la proteína, que no se muestra en la figura 3-2, lo que induce un cambio conformacional que separa las hélices internas. El Ca2+ y los nucleótidos cíclicos aumentan la probabilidad de abrirse de algunos otros canales 2-TM y 6-TM mediante un mecanismo similar. Hay ocho subfamilias de canales Kir 2-TM en el genoma del ser humano. Varias son importantes en la electrofisiología cardiaca. El Kir2 (o IK1) es el rectificador interno original descubierto en el músculo cardiaco; se encarga de mantener el potencial en reposo. Los canales Kir3 se abren por medio de receptores acoplados a proteína G (GPCR); en el corazón, se denominan KACh. Los canales Kir 6 se abren cuando la proporción ADP/ATP aumenta. En el corazón, se denominan KATP . CANALES SENSIBLES A LA DEFORMACIÓN MECÁNICA Los canales mecanosensitivos son una clase diversa de canales no relacionados, desde el punto de vista estructural, que desempeñan muchas funciones diferentes en distintas células. La mecanosensación es importante para el tacto y la audición, y para detectar presión arterial y para la propiocepción, al proporcionar información acerca CAPÍTULO 3 Membranas celulares y mecanismos de transporte G Y G P S1 S2 + + S4 + + S3 S5 19 S6 C N FIGURA 33 Características topológicas de un monómero de canales de K dependientes de voltaje (KV). Las seis hélices transmembrana (S1-S6) son características de todos los canales iónicos dependientes de voltaje. (Modificada con autorización de Landowne D: Cell Physiology, New York: Lange Medical Books/McGraw-Hill, 2006.) de la posición, orientación, velocidad y aceleración del cuerpo y sus partes. Los canales se asocian con moléculas accesorias y estructuras celulares que aumentan sus funciones particulares. Las células somáticas no sensoriales también responden a tensión mecánica sin informar al sistema nervioso —por ejemplo, para compensar la hinchazón osmótica o modular la secreción o la contracción. Muchos canales mecanosensitivos son canales catiónicos no selectivos. Algunos son de gran tamaño y permiten que electrólitos y metabolitos pequeños, no así proteínas, crucen la membrana. Las dos estructuras que se resolvieron son bacterianas. Una es un homopentámero, y cada subunidad contiene dos hélices TM. La otra es un heptámero, y cada subunidad contiene tres hélices TM, éstas son estructuras hermosas, pero no aclaran las otras formas de canales mecanosensitivos. CANALES SENSIBLES A VOLTAJE Los canales de K sensibles a voltaje (KV) se encargan del regreso al estado de reposo, al terminar un potencial de acción. El KV tiene una estructura central similar a la del Kir, y otras cuatro hélices TM en cada subunidad (figura 3-3). El cuarto segmento TM (S4) se distingue porque tiene entre 4 y 8 cadenas laterales con carga positiva (Arg o Lis). S4 es una característica distintiva de canales sensibles a volta- P S1 N S2 S3 + + S4 + + S5 S6 je. Se cree que es el sensor de voltaje que se mueve hacia la superficie extracelular cuando el potencial de membrana cambia y causa los cambios conformacionales que llevan a la abertura del canal. Hay nueve subfamilias de canales KV y varias subfamilias más de canales 6-TM, incluso los canales de K activados por Ca; los canales que se activan por hiperpolarización, importantes para la actividad de marcapasos en el corazón, y los canales sensibles a nucleótido cíclico. Las dos últimas familias son canales catiónicos no selectivos. Los canales de Na sensibles a voltaje (Nav) se encargan de la deflexión ascendente del potencial de acción, y apoyan su propagación. Los canales de Ca2+ sensibles a voltaje (Cav) acoplan cambios del potencial de membrana con un incremento de la concentración intracelular de Ca2+, que actúa como un segundo mensajero para controlar diversos procesos intracelulares. Los canales Nav y Cav tienen una estructura similar a los canales KV, excepto porque son moléculas de mayor tamaño únicas que incorporan cuatro dominios, cada uno con segmentos 6-TM un poco diferentes (figura 3-4). Los filtros de selectividad tienen cuatro paredes distintas. El canal Cav tiene cuatro glutamatos característicos (EEEE) en su revestimiento de poro, uno en cada dominio. El canal Nav tiene un patrón DEKA en las cuatro paredes de su poro. Estas cadenas laterales deben estar expuestas a la luz del poro. Las cargas que exponen a la luz y el tamaño del poro determinan la selectividad del canal. P S1 S2 S3 + + S4 + + S5 S6 P S1 S2 S3 + + S4 + + S5 S6 P S1 S2 S3 S4 S5 S6 C FIGURA 34 Características topológicas de los canales de Na dependientes de voltaje (Nav). Cuatro dominios un poco diferentes están enlazados juntos en una proteína. (Modificada con autorización de Landowne D: Cell Physiology, New York: Lange Medical Books/McGraw-Hill, 2006.) 20 SECCIÓN II Fisiología celular CANALES QUIMIOSENSITIVOS Hay muchos canales quimiosensitivos o sensibles a ligando, los cuales controlan el flujo de iones y, así, generan señales eléctricas en respuesta a sustancias químicas específicas, como acetilcolina (ACh), glutamato, o ATP. Pueden agruparse en tres superfamilias distintas de acuerdo con las características estequiométricas, y las características topológicas de membrana, de sus subunidades. Muchas de éstas se descubrieron por vez primera en farmacología al notar que ciertos compuestos, llamados agonistas, producían corrientes de membrana o alteraban la actividad eléctrica de células y otros compuestos; los antagonistas podían bloquear estos efectos. Para algunas corrientes inducidas por agonista, el ligando se une a la misma molécula que contiene el canal; son los canales sensibles a ligando, que a veces se llaman receptores de ligando ionotrópicos para distinguirlos de los receptores de ligando metabotrópicos, donde el ligando se une a un GPCR y desencadena una cascada química que puede incluir abertura de otros canales, por ejemplo, KACh, antes descritos. Los canales receptores de ACh (AChR) se denominan AChR nicotínicos, o nAChR. El término “nicotínico” indica que a estos receptores se une la nicotina, que también abre los canales. Los nAChR se distinguen de los AChR muscarínicos, que no son canales sino GPCR. Los nAChR se encuentran en las membranas postsinápticas en uniones neuromusculares esqueléticas y en los sistemas nerviosos autónomo y central. Los nAChR mejor estudiados son pentámeros heteroméricos (figura 3-5). Los monómeros tienen cuatro segmentos TM cada uno, y una región N terminal extracelular grande. En la unión neuromuscular, el nAChR tiene dos subunidades α, con sitios de unión a ACh en la interfaz entre subunidades y lejos de la membrana lipídica. La unión a ACh induce un cambio conformacional que abre el poro formado en el nivel de la membrana lipídica y revestido por el segundo segmento TM de cada una de las cinco subunidades del monómero. Los canales abiertos son muy permeables tanto a Na+ como a K+, un poco permeables a Ca2+, y no permeables a aniones. No son tan selectivos como los canales Kir o sensibles a voltaje. Desde el punto de vista funcional, la permeabilidad al Na+ es más importante (capítulo 7). Los receptores postsinápticos del SNC para glicina (glyR), ácido gammaminobutírico (GABAAR), y serotonina (5HT3R) tienen estructura pentamérica similar, aunque algunos son homoméricos, como lo son algunos nAChR. Los glyR y los GABAAR son selectivamente permeables a aniones y producen potenciales postsinápticos inhibidores (IPSP). Los 5HT3R son selectivos a catión, similares a nAChR, y producen potenciales postsinápticos excitadores (EPSP). Los canales de EPSP del SNC más comunes son los receptores de glutamato (gluR), los cuales tienen una estructura (figura 3-6) que recuerda la de una molécula de Kir invertida, con segmentos TM extra. Los gluR son tetrámeros heteroméricos con tres TM por cada subunidad. Tienen una región extracelular grande con cuatro sitios de unión a glutamato y un asa P que mira hacia el citoplasma; varios gluR diferentes desde el punto de vista funcional se comentan con mayor detalle en el capítulo 7. Todos son selectivos para catión; algunos permiten la entrada de Ca2+ y otros no. N N C Afuera Afuera Adentro Adentro C β α γ α δ FIGURA 35 Características topológicas de un monómero de FIGURA 36 Características topológicas de un monómero de canales receptores de acetilcolina nicotínicos (nAChR), con una vista desde arriba que muestra la disposición de los cinco monómeros. canales de receptor de glutamato (gluR), con una vista desde arriba que muestra la disposición de los cuatro monómeros. (Modificada con (Modificada con autorización de Landowne D: Cell Physiology, New York: Lange Medical autorización de Landowne D: Cell Physiology, New York: Lange Medical Books/ Books/McGraw-Hill, 2006.) McGraw-Hill, 2006.) CAPÍTULO 3 Membranas celulares y mecanismos de transporte Los canales sensibles a ATP o receptores P2X (P2XR) tienen dos TM por cada subunidad y tres subunidades por cada canal. “P” se refiere a la sensibilidad a purina; la adenina es una purina. P2 los distingue de los receptores P1, que son sensibles a adenosina y actúan por medio de la adenilil ciclasa (AC). Los receptores P1 a menudo se denominan receptores A (A de adenosina); son GPCR. La cafeína es un antagonista de algunos de los receptores A. Los receptores P2 prefieren ADP o ATP en lugar de adenosina. Los P2XR son canales y los P2YR son GPCR. Los receptores purinérgicos se conocen mejor como reguladores del flujo sanguíneo en los tejidos; también quedaron implicados en varios procesos sensoriales. Otras dos familias de canales tienen miembros quimiosensitivos, pero también cuentan con miembros importantes sin ligandos conocidos; son la familia del canal de sodio epitelial (ENaC) y la familia de receptor de inositol trifosfato (IP3) (IP3R). Los ENaC son importantes en la resorción de sodio a partir de la orina que se forma en los túbulos de la nefrona. Se cree que son tetrámeros heteroméricos, cada uno con dos segmentos TM; estos no son dependientes de voltaje. Se sabe que son regulados mediante control de su inserción y eliminación de la membrana, y puede haber un ligando desconocido para este canal. Hay canales estructuralmente relacionados en invertebrados que tienen ligandos conocidos. Los IP3R y los receptores de rianodina (RyR) relacionados se encuentran en la membrana del ER. Cuando están abiertos, permiten la liberación de Ca2+ desde el ER. El IP3 es un segundo mensajero producido por la acción de la fosfolipasa C (PLC) sobre el lípido de membrana fosfatidilinositol, que con anterioridad fue fosforilado para que sea PIP2. Los RyR también controlan la liberación de calcio, sobre todo en el músculo, a partir del retículo sarcoplasmático. Rianodina se refiere a una toxina que abre de manera parcial estos canales. Los RyR son abiertos por interacción directa con un canal Cav modificado en el músculo esquelético, y por Ca2+ intracelular en el músculo cardiaco. Las funciones de los IP3R y de los RyR se comentan con mayor detalle en los capítulos 9 y 10. Los RyR son homotetrámeros con una enorme región N terminal citoplasmática de 20 nm de diámetro. El peso molecular total para el tetrámero es de alrededor de 2 000 kDa, un tamaño alrededor de 10 veces mayor que el de los canales Nav o KV. Los canales IP3R también son homotetrámeros de alrededor de la mitad del tamaño de los RyR. Se ha predicho que los IP3R tienen seis segmentos TM por cada monómero, y que los RyR tienen de 4 a 12. CANALES DE AGUA Algunas células requieren más permeabilidad al agua que la que proporciona la bicapa lipídica. Los eritrocitos, que deben cambiar con rapidez de forma para pasar a través de capilares estrechos, y muchas células epiteliales, entre las que destacan las de los riñones, tienen canales de agua especializados o acuaporinas (AQP), que permiten el paso de agua, pero que excluyen iones. Las AQP se encuentran como tetrámeros con cuatro poros funcionales, uno en cada subunidad. Las subunidades tienen seis segmentos TM y dos regiones similares al asa P de canales KV. Una de las asas mira hacia la superficie extracelular, y la otra hacia el citoplasma, y se reúnen en la parte media de la membrana. Las funciones de las AQP y de los ENaC se comentan hacia el final de este capítulo. 21 CANALES CÉLULACÉLULA En casi todos los tejidos hay canales que conectan el citoplasma de una célula con el citoplasma de su vecina. Las excepciones son células que flotan con libertad en la sangre y en las células del músculo esquelético. Casi todos estos canales unen células del mismo tipo, pero hay algunas células de diferente tipo con uniones entre ellas. Estos canales se detectaron eléctricamente al mostrar que podía pasar corriente de una célula a otra a través de una sinapsis eléctrica. Más tarde se asociaron con una estructura anatómica que se conoce como unión intercelular comunicante (conexión comunicante), llamada así por su aspecto en micrografías electrónicas. En realidad esta brecha es abarcada por disposiciones de proteínas que coinciden, provenientes de cada célula, con hasta miles de canales célulacélula por cada unión intercelular comunicante. Cada canal célula-célula está formado de dos hemicanales, uno que proviene de cada célula (figura 3-7). También se llaman conexones. Un hemicanal es un hexámero homomérico o heteromérico de proteínas conocidas como conexinas. Hay más de 15 conexinas diferentes con peso molecular entre 25 y 50 kDa. Todas tienen cuatro segmentos TM y dos asas extracelulares, y sus N y C terminales están en el citoplasma. Algunas conexinas, más no todas, pueden formar canales híbridos que unen distintos hemicanales sobre las dos células. Afuera Adentro N C Adentro Afuera Adentro FIGURA 37 Características topológicas de la conexina, un monómero de canales célula-célula; vista desde arriba que muestra la disposición de seis monómeros en un hemicanal, y vista lateral que muestra dos membranas celulares con hemicanales alineados. (Modificada con autorización de Landowne D: Cell Physiology, New York: Lange Medical Books/McGraw-Hill, 2006.) 22 SECCIÓN II Fisiología celular El poro es de tamaño mucho mayor que los canales iónicos antes descritos. Mide alrededor de 1.2 nm y es permeable a aniones, cationes y metabolitos pequeños, así como a segundos mensajeros, como ATP, cAMP o IP3, no así a proteínas. Experimentalmente, el poro es permeable a moléculas con peso molecular de menos de 1 kDa. Los canales célula-célula permiten que las células en un tejido funcionen de una manera coordinada. Si una célula queda dañada, puede cerrar sus canales célula-célula que van hacia sus vecinas y, así, evitar la pérdida de moléculas pequeñas desde el tejido entero. Este mecanismo de compuerta está controlado por el Ca2+, H+ o voltaje transunión, intracelular. Diferentes conexones tienen sensibilidad distinta a estos tres cambios. El mecanismo de compuerta también puede ser inducido por octanol y anestésicos como halotano. En algunas situaciones, conexones no pareados, o los panexones relacionados, pueden abrirse y permitir que moléculas pequeñas se muevan desde el citoplasma hacia el espacio extracelular. Se insinúa que los panexones tienen un papel en la inflamación y en la respuesta a isquemia al permitir la liberación de ATP para emitir señales a células cercanas a un sitio de fenómeno adverso tisular. Afuera 2Ko 3Nao E2 3Na 2K Adentro P P P Afuera Ocluido Adentro 3Na 2K P ATP ADP BOMBAS Los iones se mueven a través de membranas celulares por medio de canales, bombas y transportadores. Estos son tres mecanismos distintos, y el estudiante debe tener cuidado de no confundirlos. Las bombas crean gradientes iónicos y los mantienen; mueven iones contra el gradiente a expensas de ATP. Los canales usan estos gradientes para producir las diversas señales eléctricas. Los transportadores usan uno o más gradientes; el movimiento de un ion (a menudo Na+) en favor del gradiente está acoplado al movimiento de otra sustancia en contra del gradiente. Dado que consumen ATP, las bombas a menudo se denominan ATPasas. Se describirán cinco bombas con detalle: la bomba de Na/K, la bomba de Ca, y tres tipos de bombas de protones. El primer tipo de éstas se llama bombas tipo P, porque son autofosforiladas durante el ciclo de reacción, o bombas E1-E2, porque tienen dos estados conformacionales principales. BOMBA DE Na/K La bomba de Na/K, que a menudo se conoce como bomba de Na, mueve tres iones de Na+ hacia afuera de la célula y dos iones de K+ hacia la célula en un ciclo que convierte una molécula de ATP en ADP + Pi. A velocidad máxima, la bomba completa alrededor de 100 ciclos por segundo (cps), lo que significa que el movimiento de iones por cada molécula es mucho menor que el de un canal Nav, lo cual puede permitir el flujo de 1 000 iones/ms hacia la célula. Los canales Nav sólo se abren brevemente cuando la célula está activa; la bomba funciona de manera continua para recuperarse luego de la actividad. La actividad de la bomba se incrementa cuando el Na+ intracelular o el K+ extracelular aumenta, y la bomba actúa de manera homeostática para restituir las concentraciones originales. La bomba de Na es un heterodímero con una subunidad α que tienen los sitios de unión a Na+, K+ y ATP y una subunidad β que se cree que es importante para la inserción en la membrana. La subunidad β tiene un segmento TM; la subunidad α probablemente tiene Afuera E1 Adentro 2Ki 3Na ATP 3Nai 2K ATP FIGURA 38 El ciclo de la bomba de Na/K. La operación de la bomba en la dirección de las manecillas de la carátula del reloj mueve tres Na+ hacia afuera, y después mueve dos K+ a expensas de convertir un ATP en ADP. (Modificada con autorización de Landowne D: Cell Physiology, New York: Lange Medical Books/McGraw-Hill, 2006.) 10. El Na+ y el ATP intracelulares se unen a la forma E1 de la subunidad α, que a continuación es fosforilada y se convierte en la forma E2 (figura 3-8, parte inferior izquierda, avanzando en la dirección de las manecillas del reloj). La forma E2 libera el Na+ hacia el espacio extracelular y se une al K+ extracelular. La estructura cristalina de la forma E2-2K+—Pi hace poco se resolvió. La estructura general de dominios y hélices es similar a la de la bomba de Ca descrita más adelante, pero hay diferencias relacionadas con las funciones específicas de cada bomba. El ciclo continúa cuando el fosfato es separado de la proteína por hidrólisis; la proteína cambia de regreso a la forma E1, libera el K+ dentro de la célula, y después se une a la siguiente carga de Na. A medida que el Na+ y el K+ se mueven de manera alternativa a través de la membrana, la bomba pasa por un estado ocluido en el cual los iones no son accesibles a una u otra solución. Los digitálicos y la ouabaína, un glucósido cardiaco relacionado, suspenden la acción de la bomba al unirse fuera de la célula a la forma E2 cerca del sitio de unión a K+. Los digitálicos se usan para tratar diversas afecciones cardiacas. Es un fármaco peligroso, y debe CAPÍTULO 3 Membranas celulares y mecanismos de transporte 23 usarse con precaución, de modo que sólo se bloqueen algunas de las moléculas de la bomba y se dejen otras funcionales. El peligro se complica porque el K+ extracelular antagoniza la unión de digitálicos al impulsar la bomba hacia la forma E1; el médico prudente vigilará la concentración de potasio en sangre durante el tratamiento con digital. La bomba de Na es electrogénica, porque cada ciclo mueve una carga neta hacia afuera de la célula. Esta corriente sólo tiene un pequeño efecto sobre el potencial de membrana, en comparación con el flujo de iones a través de canales, que se comenta en el capítulo siguiente. El movimiento neto de Na+ hacia afuera de la célula evita la acumulación de NaCl en la célula. Si la bomba es bloqueada con glucósidos cardiacos, la célula se hinchará debido al flujo de entrada osmótico de agua siguiendo el NaCl. CUADRO 3-1 Ubicación de bombas de membrana. BOMBA DE CA La bomba tiene ocho subunidades diferentes y más de 20 cadenas polipeptídicas. La porción F0 abarca la membrana y porta los iones H; la F1 se extiende hacia la matriz mitocondrial. Parte del complejo rota alrededor de un eje perpendicular al plano de la membrana, similar a una turbina, a medida que los iones H fluyen a través de ésta. Otra porción, el estator, permanece fija en su posición, y la interacción entre el rotor y el estator produce una secuencia de estados conformacionales que favorece la síntesis de ATP. En presencia de ATP alto, ADP bajo y gradiente de protón nulo, el proceso puede revertirse para bombear H+. Hay dos bombas de Ca importantes, una que bombea Ca2+ desde el citoplasma hacia el espacio extracelular, y otra, la bomba SERCA, que bombea Ca2+ desde el citoplasma hacia la luz del retículo sarcoplasmático o el ER. Se cree que tienen mecanismos similares; ambas son bombas E1-E2 tipo P que mueven dos iones Ca2+ hacia afuera del citoplasma y 2 o 3 iones H+ hacia el citoplasma por cada ATP consumido. La estructura de la bomba SERCA se ha resuelto en varios estados diferentes. Es una molécula alta, de alrededor de 15 nm de alto y 8 nm de grosor, que se extiende en su mayor parte fuera de la membrana sobre el lado citoplasmático. Hay 10 segmentos TM. La pieza de la cabeza citoplasmática consta de los dominios actuador (A), de unión a nucleótido (N) y de fosforilación (P). Los tres dominios citoplasmáticos están divididos en el estado E1 • 2Ca, pero se reúnen para formar una pieza de la cabeza compacta en los otros estados. Este movimiento se transmite hacia la porción de la membrana a través de las hélices 1 a 3, fija al dominio A, y 4 y 5, fijas al dominio P, para permitir que el Ca2+ se libere en el lado no citoplasmático. La distancia entre los sitios de unión a Ca2+ y el sitio de fosforilación es de más de 5 nm. Bomba Tipo de célula Membrana Inhibidor Na/K Todas Superficie Digital Ca Todas Superficie y ER Tapsigargina H/K Intestino, riñón Superficie Omeprazol H tipo F Todas Mitocondrias Oligomicina H tipo V Todas Superficie y vesículas Bafilomicina BOMBAS DE H TIPO V Las bombas de H tipo V también son complejos proteínicos de hasta 14 subunidades con rotores y estatores. Mueven protones hacia vacuolas y otros orgánulos intracelulares, como lisosomas, el aparato de Golgi, y vesículas secretoras. El gradiente de H+ que se produce a través de membranas de vesículas sinápticas se usa para impulsar el empaque de neurotransmisores (véanse figuras 7-3 a 7-5). Estas bombas se encargan del H+ que es secretado por osteoclastos para disolver huesos, y de la secreción de H+ en el riñón y el epidídimo (cuadro 3-1). BOMBA DE H/K La bomba de H/K secreta ácido hacia el estómago al bombear dos iones H+ fuera de las células parietales de las glándulas gástricas y dos iones K+ hacia la célula, mientras divide una molécula de ATP. Bombas similares también operan en las células epiteliales en el intestino y el riñón; esta es una bomba tipo P E1-E2, y tiene una subunidad β, similar a la bomba de Na/K. La bomba de H/K es inhibida por el omeprazol y otros fármacos similares que se usan en el tratamiento de pirosis frecuente. BOMBAS DE H TIPO F La bomba de H tipo F más importante, por lo general, corre en reversa como la F0-F1 ATP sintasa que se encuentra en las mitocondrias. Este complejo proteínico permite que fluyan protones en favor de su gradiente electroquímico, y convierte el flujo de 10 protones para formar tres ATP a partir de ADP. Los gradientes de hidrógeno se producen mediante metabolismo oxidativo en mitocondrias. TRANSPORTADORES Los transportadores mueven iones y otras moléculas pequeñas a través de la membrana, y no son canales ni bombas. A veces la palabra transportador se usa en el sentido general para incluir todos los mecanismos de transporte, y transportador secundario se utiliza para distinguir este grupo. Los transportadores pasan por un cambio conformacional según transportan; en este aspecto son similares a las bombas y distintos de un canal abierto. A diferencia de una bomba, no consumen ATP. Se cree que casi todos los transportadores tienen 12 segmentos TM en dos grupos de seis, con un asa citoplasmática de mayor tamaño entre ellos. Algunos tienen seudosimetría doble y asas P que miran hacia ambas superficies. Hay tres categorías generales de transportadores: uniportadores, simportadores o cotransportadores, y antiportadores o intercambiadores (figura 3-9). El transportador de glucosa (GLUT) es un uniportador que facilita la difusión de glucosa en favor de su gradiente de concentración hacia muchas células que consumen glucosa. También facilita el 24 SECCIÓN II Fisiología celular El intercambiador de Cl/HCO3, que también se conoce como el intercambiador aniónico (AE), es importante para mover CO2 desde los tejidos hacia los pulmones. El CO2, producido por el metabolismo en las células se convierte en bicarbonato por la anhidrasa carbónica en los eritrocitos, y el HCO3– se mueve hacia el plasma en intercambio por cloruro por medio de AE. El proceso se revierte a medida que la sangre pasa por los pulmones y el CO2 se mueve hacia el aire para ser exhalado. Este proceso se analiza en el capítulo 37. Uniportador Simportador Antiportador FIGURA 39 Tres tipos de transportadores. (Modificada con autorización de Landowne D: Cell Physiology, New York: Lange Medical Books/McGraw-Hill, 2006.) movimiento de glucosa desde las células que liberan glucosa al desintegrar glucógeno, y desde superficies basales de células epiteliales que revisten los intestinos y los túbulos renales (figura 3-14). El cotransportador de Na-glucosa (SGLT) es un simporte que transporta glucosa hacia células epiteliales intestinales y renales a través de sus superficies apicales contra el gradiente de concentración de glucosa. La energía que se requiere para este transporte proviene del movimiento de uno o dos iones sodio en favor de su gradiente electroquímico por cada molécula de glucosa transportada. Un cotransportador de Na/glutamato recupera glutamato que se usa como un neurotransmisor en sinapsis en el SNC (véase figura 7-4). Acopla el movimiento corriente abajo de tres iones Na+ y un ion K+ al transporte corriente arriba de un glutamato. Hace poco se resolvió la estructura de un transportador de glutamato bacteriano, que se cree que es similar al de seres humanos. Tiene ocho segmentos TM y dos asas P, una que mira el citoplasma y otra el exterior. Se cree que movimientos relativamente pequeños de la proteína pueden transferir el glutamato desde un asa P a la otra y, así, a través de la membrana. Hay un antiportador de H/glutamato que usa el gradiente de H+, establecido por una bomba tipo V, a través de la membrana que rodea vesículas sinápticas para concentrar glutamato dentro de la vesícula (véase figura 7-4). Hay muchos cotransportadores impulsados por Na para mover otras moléculas pequeñas hacia las células, y transportadores impulsados por H para mover algunos materiales hacia vesículas. Algunos de estos transportadores son blancos para intervención farmacológica; por ejemplo, la fluoxetina actúa sobre un cotransportador de Na/serotonina. Otros se comentan más en el capítulo 7. Algunos aniones son cotransportados con sodio; por ejemplo, el simportador Na/I concentra yodo hacia las células foliculares tiroideas. El intercambiador de Na/Ca (NCX) es un importante regulador de la concentración intracelular de Ca2+. Tres iones sodio que se mueven en favor de su gradiente electroquímico hacia la célula pueden mover un ion calcio hacia afuera, o viceversa; todos los intercambiadores pueden correr en una u otra dirección dependiendo de los gradientes relativos. El efecto de la digital sobre el músculo cardiaco es aumentar el Na intracelular al inhibir la bomba de Na/K. El Nai+ alto significa que hay menos gradiente hacia adentro para el Na+ y, por ende, menos flujo de salida de Ca2+ por medio de NCX. Esto aumenta el Cai2+ y produce una contracción más fuerte (capítulo 23). TRANSPORTADORES ABC Este grupo mixto de proteínas de transporte de 12 TM contiene una secuencia de aminoácidos secuencia de unión a ATP (ABC) característica y, en ausencia de información más específica, se supone que consume ATP mientras transporta algún material a través de la membrana. Dos transportadores ABC merecen mención aquí, el transportador de resistencia a múltiples fármacos (MDR), que es una bomba, y el regulador transmembrana de la fibrosis quística (CFTR), que es un canal. El MDR1 produce extrusión de fármacos hidrofóbicos a través de la membrana celular. Se cree que actúa de algún modo como la flipasa y produce extrusión de los fármacos sin mucha especificidad. Una amplia variedad de células en el tracto GI, el hígado y el riñón, expresan proteínas MDR; éstas pueden frustrar al médico que intenta proporcionar fármacos para tratar cáncer entre estas células. El CFTR es una proteína que, cuando está mutada, lleva a fibrosis quística. La proteína natural es un canal de cloruro que requiere fosforilación por la proteína cinasa A (PKA) e hidrólisis de ATP adicional por la proteína CFTR activada para abrirse. El Cl– se mueve en favor de su gradiente electroquímico. La fibrosis quística ocurre debido a la falta de transporte de Cl– en el conducto pancreático (y por ende del cístico). El Cl– disminuido lleva a decremento del agua, y la secreción rica en proteína se hace espesa y puede bloquear los conductos que después se hacen fibróticos. Antes del desarrollo de terapia de reemplazo por vía oral para las enzimas pancreáticas faltantes, muchos pacientes con CF morían por complicaciones de malnutrición. Ahora el principal problema es el espesamiento del moco en los pulmones debido a secreción insuficiente de líquido. RECEPTORES DE MEMBRANA La palabra receptor proviene de estudios farmacológicos, en los cuales designa el sitio de acción o la molécula sobre la cual actúa una pequeña molécula de interés, quizás una hormona o neurotransmisor. Aquí se usa en un sentido más restrictivo para significar moléculas que abarcan la membrana, sobre las cuales actúa la molécula pequeña en la superficie externa, y que desencadenan alguna acción dentro de la célula cuando la molécula pequeña está presente. También hay receptores intracelulares, por ejemplo, el receptor de hormona esteroide. Las hormonas esteroides y fármacos relacionados pueden cruzar la bicapa lipídica y unirse a estas proteínas intracelulares. También se excluyen los canales quimiosensitivos, aunque algunos farmacólogos les gusta llamarlos receptores ionotrópicos. Hay dos categorías principales de estos receptores de membrana: los GPCR y los receptores ligados a enzima o catalíticos. CAPÍTULO 3 Membranas celulares y mecanismos de transporte Agonista AC Efector GPCR P β/γ Proteína G 25 α ATP cAMP PKA FIGURA 310 La vía de emisión de señales Gαs. La unión de agonista al receptor acoplado a proteína G causa la disociación de la subunidad α, lo cual hace que la adenilil ciclasa aumente la concentración de cAMP. Esto, a su vez, hace que la proteína cinasa A fosforile una proteína efectora (en este caso un canal). (Modificada con autorización de Landowne D: Cell Physiology, New York: Lange Medical Books/McGraw-Hill, 2006.) RECEPTORES ACOPLADOS A PROTEÍNA G Los GPCR tienen siete segmentos TM con una N terminal extracelular. Están acoplados a una proteína de unión a GTP trimérica compleja. Cuando una hormona o un neurotransmisor interactúa con un GPCR, induce una conformación en el receptor que activa una proteína G heterotrimérica en la superficie de la membrana interna de la célula (figura 3-10). En el estado heterotrimérico inactivo, el GDP está unido a la subunidad Gα. En el momento de la activación se libera el GDP, se une GTP a Gα, y después Gα-GTP se disocia de Gβγ y del receptor. Tanto Gα-GTP como Gβγ a continuación están libres para activar otras proteínas de membrana. Casi toda la Gα y la Gγ están lipidadas; tienen un ancla de lípido fija de manera covalente en la bicapa de la membrana. La duración de la señal de la proteína G está determinada por la tasa de hidrólisis de GTP intrínseca de la subunidad Gα y la reasociación subsiguiente de Gα-GDP con Gβγ. Hay más de 2 000 GPCR predichos en el genoma del ser humano, más de 5% de todos los genes. Más de 800 son receptores olfatorios; otros detectan casi todos los neurotransmisores y muchas hormonas. Los GPCR también detectan luz en el ojo (figura 5-2). Distintas células tienen diferentes paletas de GPCR acoplados a distintas proteínas G que controlan diferentes grupos de reacciones intracelulares. Sólo hay alrededor de 16 subunidades de Gα distintas, y aún menos Gβγ. Tres clases de subunidades de Gα inician casi todos los eventos subsiguientes descritos en este libro. Gαs estimula la adenilil ciclasa (AC), Gαi inhibe la AC, y su βγ asociada activa de manera directa canales KACh, y Gαq estimula una fosfolipasa (PLCβ). La AC produce cAMP que puede influir de manera directa sobre algunos canales. El cAMP también activa la proteína cinasa A (PKA) al disociar las subunidades reguladoras de las subunidades catalíticas. La PKA fosforila muchas proteínas, lo que altera la actividad de las células. La PLCβ divide el fosfolípido de membrana PI para producir IP3 y diacilglicerol (DAG). Como se describió, IP3 se une a los canales IP3R, lo que aumenta el Cai, que a su vez desencadena diversas reacciones. En capítulos subsiguientes se describen con mayor detalle varios ejemplos de cascadas iniciadas por GPCR. Las toxinas que están detrás de dos enfermedades infecciosas, el cólera y la tosferina, ADP-ribosilan subunidades Gα, lo que lleva a activación constitutiva. En el cólera, la Gα activada en el tejido epitelial intestinal estimula la AC, las concentraciones de cAMP aumentan, y los canales de cloruro CFTR se abren, lo que lleva a diarrea acuosa. Las personas con fibrosis quística pueden ser resistentes al cólera porque tienen menos canales de cloruro funcionales. No está clara la patogenia celular de la tosferina. RECEPTORES LIGADOS A ENZIMA Muchos receptores ligados a enzima son receptor tirosina cinasas (RTK) y actúan al fosforilar cadenas laterales de tirosina en otras proteínas, que a su vez pueden fosforilar otras proteínas. Algunos receptores ligados a enzima no son cinasas por sí mismos, sino que están acoplados a una proteína asociada que fosforila otras proteínas. Algunos receptores ligados a enzima son guanilil ciclasas, tirosina fosfatasas, o serina cinasas. Casi todos los factores de crecimiento y de diferenciación actúan al unir RTK específicos. El receptor de insulina es un RTK que fosforila una familia de sustratos que se conocen como sustratos receptores de insulina; éstos estimulan cambios del metabolismo de glucosa, proteína y lípido, y desencadenan también la vía de emisión de señales Ras, lo que activa factores de transcripción que promueven el crecimiento. Los receptores de interferón y las moléculas CD4 y CD8 sobre la superficie de linfocitos T, son ejemplos de receptores que están acoplados a tirosina cinasas citoplasmáticas. 26 SECCIÓN II Fisiología celular MOLÉCULAS DE ADHESIÓN CELULAR Casi todas las células, excepto los eritrocitos, tienen proteínas de membrana integrales que se fijan a la matriz extracelular o con moléculas de adhesión sobre células vecinas; estas moléculas se mantienen junto el tejido, y pueden permitir la transmisión de fuerzas mecánicas de una célula a otra. Pueden actuar como señales durante el desarrollo, de modo que una célula puede reconocer otra. También pueden trabajar como receptores, al informar al interior de la célula que tienen algo unido. Algunos son controlados desde el interior, y sólo se unen cuando se recibe alguna señal. Las integrinas son ejemplos de moléculas de adhesión de la matriz celular. Tienen un segmento TM único y enlazan células a fibronectina o laminina en la matriz extracelular. Las cadherinas son moléculas de adhesión célula-célula dependientes de Ca; son glucoproteínas con un segmento TM único, y se cree que se unen de manera homofílica (a otra copia de la misma molécula) a cadherinas en la otra célula. Se han encontrado cadherinas en muchas sinapsis neurona-neurona. Hay una familia grande de moléculas de adhesión celular, de las cuales las N-CAM son las que se entienden mejor. Las N-CAM se encuentran sobre diversos tipos de células y en casi todas las células nerviosas. Al igual que las cadherinas, las N-CAM tienen un segmento TM único y se unen de manera homofílica, pero difieren en que no requieren Ca2+ para unirse. Las moléculas de adhesión intercelular (ICAM) son una clase relacionada que se expresa sobre la superficie de células endoteliales capilares que fueron activadas por una infección en el tejido circundante. Se unen de manera heterofílica (a una molécula diferente) a integrinas sobre leucocitos y los ayudan a moverse hacia el sitio de infección. Las selectinas son proteínas de unión a carbohidrato sobre la membrana de células endoteliales, que reconocen azúcares sobre la superficie del leucocito, y forman el enlace inicial, que es fortalecido por las ICAM. TRANSPORTE A TRAVÉS DE MEMBRANAS CELULARES Desde un punto de vista funcional, la exposición sobre el transporte de materiales a través de membranas celulares puede dividirse en transporte pasivo, en el cual los materiales se mueven en favor de su Flujo gradiente de concentración, y transporte activo, que crea estos gradientes o los mantiene. TRANSPORTE PASIVO Difusión simple Algunos materiales pueden moverse en favor de su gradiente de concentración mediante difusión simple a través de la bicapa lipídica. Moléculas pequeñas, sin carga, como O2, CO2, NH3, NO, H2O, esteroides y fármacos lipofílicos pueden entrar a las células o salir de las mismas mediante difusión simple. El flujo neto de estos compuestos a través de la membrana es proporcional a la diferencia de sus concentraciones en los dos lados, o como se expresa en la ecuación que sigue: J1→2 = –P(C2 – C1) = –PΔC (1) Con el sistema de unidades centímetro-gramo-segundo (CGS), J1→2 es el número de moles que se mueven a través de 1 cm² de membrana desde el lado 1 hacia el lado 2 cada segundo, y C1 y C2 son los números de moles del material por centímetro cúbico de solución en los dos lados. P, la constante de proporcionalidad, se llama permeabilidad de la membrana a este material en centímetros por segundo. La ecuación se escribe con el signo de menos, al principio, para ayudar a recordar que el flujo se mueve en favor del gradiente de concentración. Esta relación se muestra gráficamente en la figura 3-11. La ecuación (1) es la primera ley de Fick; puede usarse para describir el flujo de sustancias simples sin carga a través de cualquier membrana. Por ejemplo, es útil para describir el movimiento de oxígeno desde el aire hacia los alvéolos de los pulmones y hacia la sangre, a través de las células del epitelio alveolar y el endotelio capilar. Una sustancia con carga también estará influida por la diferencia de potencial eléctrico a través de la membrana de una manera que se comentará en el siguiente capítulo. Si no hay diferencia de potencial a través de la membrana, la ley de Fick también es aplicable a sustancias cargadas. La permeabilidad describe una propiedad de una membrana particular en relación con una sustancia particular. La membrana se considera permeable, mientras que se dice que las sustancias son permeantes o que permean. La permeabilidad será proporcional a Flujo Vmáx Concentración Km Concentración FIGURA 311 La dependencia de concentración de la difusión simple (izquierda) y de la difusión facilitada (derecha). (Reproducida con autorización de Landowne D: Cell Physiology, New York: Lange Medical Books/McGraw-Hill, 2006.) CAPÍTULO 3 Membranas celulares y mecanismos de transporte la capacidad de la sustancia para dividirse hacia la membrana, y para difundirse dentro de la membrana. La permeabilidad será inversamente proporcional al grosor de la membrana. Por lo general, no es fácil o necesario conocer estos tres factores por separado, pero es necesario apreciar que el engrosamiento de la membrana compleja entre el alvéolo y la sangre capilar pulmonar reducirá el movimiento de oxígeno hacia la sangre. A veces es conveniente pensar en la ley de Fick al expresar que el flujo neto de una sustancia es igual al flujo de entrada unidireccional (PC0) menos el flujo de salida unidireccional (PCi). La permeabilidad es una medida de la facilidad con la cual un soluto cruza una membrana. Las bicapas lipídicas simples son permeables a moléculas pequeñas, sin carga; la permeabilidad al agua es de alrededor de 10–3 cm/s. Así, el agua se equilibra a través de una membrana celular en algunos segundos. La urea es moderadamente permeable (P = 10–6 cm/s), y su tiempo de equilibrio es de algunos minutos. Las moléculas orgánicas pequeñas hidrofílicas, como la glucosa o aminoácidos no cargados, son menos permeables, con P = 10–7 y tiempos de equilibrio de horas; los iones son, en esencia, impermeables, con P = 10–12 cm/s y tiempos de equilibrio de muchos años. Difusión facilitada Muchas sustancias, como la glucosa o la urea, entran con facilidad a las células a pesar del hecho de que la bicapa lipídica es impermeable a ellas. El flujo de estos materiales es descrito por la ley de Fick sólo para concentraciones bajas. A concentraciones más altas, el flujo se satura a un valor máximo (figura 3-11). Esta conducta puede describirse mediante la ecuación de Michaelis-Menten, que también se usa para describir cinética enzimática. El flujo unidireccional es dado por la ecuación que sigue: JmáxC J = _____ C+K (2) m donde Jmáx es el flujo máximo y Km es la afinidad o la concentración a la cual el flujo es de la mitad de su valor máximo. Esta propiedad saturable del flujo sugiere que hay un número fijo de sitios en los cuales el flujo puede tener lugar. Asimismo, como en el caso de las enzimas, puede ser posible demostrar la competencia de diferentes sustancias por el mismo sitio, o inhibición no competitiva de los sitios de transporte. Los sitios son selectivos para una sustancia particular o un grupo de sustancias que transportarán o que permitirán la competencia por el transporte. La selectividad, afinidad y Jmáx son tres cualidades independientes de los sitios; se encontrarán con distintos valores en diferentes sistemas. La difusión facilitada ahora se entiende en términos de canales o transportadores. Casi todos los canales tienen afinidad baja o valores de Km altos; no están saturados en condiciones fisiológicas normales. Tres uniportadores de glucosa, GLUT1, GLUT3 y GLUT4 (que están regulados por insulina), se encuentran en muchos tejidos, y tienen afinidad alta por la glucosa; están saturados a todas las concentraciones fisiológicas. GLUT2, que se encuentra en tejidos que transportan grandes flujos de glucosa (como el intestino, el riñón y el hígado), tiene afinidad baja por la glucosa, y el flujo de entrada a través de transportadores GLUT2 aumenta conforme se incrementa la concentración de glucosa. 27 TRANSPORTE ACTIVO Las bombas proporcionan transporte activo primario, que mueve materiales contra sus gradientes electroquímicos a expensas de ATP. La bomba de Na/K mueve Na+ hacia afuera de la célula y K+ hacia la célula; ambos iones se mueven contra sus gradientes respectivos. La bomba SERCA mueve Ca2+ contra su gradiente desde el citoplasma hacia la luz del ER. Los cotransportadores e intercambiadores pueden proporcionar transporte activo secundario, usando un gradiente producido por el transporte activo primario para mover otro material contra su gradiente. Muchos transportadores acoplan el movimiento de Na+ o H+ en favor de su gradiente electroquímico con el movimiento de otra sustancia contra su gradiente. El NaGLUT y el antiportador H/glutamato son dos ejemplos de mecanismos de transporte secundario. El flujo mediante bombas y transportadores puede describirse con ecuaciones similares a la ecuación (2), modificadas para incluir la afinidad por cada sustancia y por ATP. Cuando más de un ion a la vez está involucrado en las reacciones en una molécula de bomba o de transportador, la ecuación también debe modificarse para reflejar esta cooperatividad. Así, el flujo de salida de sodio a través de la bomba de Na/K tiene una relación sigmoide con la concentración interna de Na+. ÓSMOSIS La vida está asociada con el movimiento de agua. El cuerpo del ser humano está conformado, en su mayor parte, por agua, y depende de manera vital de su aporte. El agua es una molécula pequeña, pero abundante. No es mucho más grande que un átomo de oxígeno; mide alrededor de 0.2 nm de diámetro transversal —lo bastante pequeña como para intercalarse entre otras moléculas, incluso en algunos cristales—. Un mol de agua contiene 18 mL; así, el agua pura es 55 mol/L. Esta concentración es varios cientos de veces más alta que las concentraciones de Na+, K+ o Cl– en el organismo, que son las siguientes más altas. Más de 99% de las moléculas en el cuerpo es H2O. Puesto que las moléculas son pequeñas, se mueven con facilidad; dado que son tan abundantes, sus movimientos tienen importancia para la salud del ser humano. Hay 2 o 3 mecanismos separados para el movimiento de agua: flujo masivo, difusión molecular y, quizá, bombeo molecular. Cuando se quita el tapón de una tina de baño o el corazón late, hay flujo masivo de líquido en respuesta a una fuerza mecánica externa —un empuje o un tirón. La fuerza impulsora para el flujo masivo es la presión mecánica que por lo regular se produce por empuje o por gravedad. La difusión molecular u ósmosis es un proceso pasivo mediante el cual el agua se difunde desde áreas de concentración alta de agua hasta aquellas de concentración baja. Hay una concentración alta de agua donde hay concentración baja de solutos, y viceversa. El agua puede difundirse a través de casi todas las membranas celulares de manera directa a través de la bicapa lipídica o al viajar a través de AQP. Muchas células producen AQP, porque la difusión simple no permite suficiente flujo de agua. Algunas células renales insertan AQP en respuesta a hormona antidiurética (ADH), de modo que aumente el flujo de agua desde la orina que se forma, de regreso hacia la sangre, lo que conserva agua. Este tipo pasivo de movimiento de agua se 28 SECCIÓN II Fisiología celular llama flujo osmótico, y la fuerza impulsora asociada es el gradiente de concentración del agua. El agua también puede transportarse a través de membranas a expensas de energía por el cotransportador de Na-glucosa (SGLT). El transporte TM de dos iones Na+ y una molécula de azúcar se asocia con el flujo de entrada de 210 moléculas de agua, independiente del gradiente osmótico. La energía podría provenir de permitir que el Na+ se mueva en favor de su gradiente de concentración. Este bombeo molecular sería un mecanismo de transporte activo secundario, y podría explicar casi la mitad de la captación diaria de agua a partir del intestino delgado. La presión osmótica es la presión mecánica necesaria para producir un flujo de agua igual y opuesto al flujo osmótico producido por un gradiente de concentración de agua. En células animales, esta presión no se desarrolla a través de la membrana celular porque las células cambiarán su volumen en respuesta al flujo osmótico. El concepto de presión osmótica es similar a (e históricamente precedido por) el potencial de equilibrio de Nernst, un potencial eléctrico que produce un flujo de iones igual y opuesto a un flujo producido por un gradiente de concentración. El potencial de Nernst se comenta en el capítulo 4. Si dos soluciones diferentes están en contacto, la presión osmótica, π, entre ellas es: (3) π = RTΔc donde R es la constante de gases molar (número de Avogadro multiplicado por la constante de Boltzmann), T la temperatura absoluta, y Δc la diferencia de concentración de todos los solutos impermeables. La diferencia de concentración se refiere a la concentración molar sumada de todas las partículas que se crean cuando el soluto está disuelto en agua. Se mide como la osmolaridad, es decir, la suma de los moles de cada componente de la solución. Una solución 2 mM de MgCl2 contiene 6 miliosmoles (mOsm) por litro de solución, dos para el Mg2+ y dos para cada Cl–. La osmolaridad de esta solución es de 6 mOsm. Una solución de NaCl 3 mM y una de urea 6 mM tienen la misma osmolaridad porque tienen el mismo número de partículas por litro de solución. Se dice que son isoosmóticas. La osmolalidad de una solución puede medirse por el cambio que produce en el punto de congelación o presión de vapor. La osmolalidad se refiere a moles de soluto por kilogramo de solvente, mientras que la osmolaridad se refiere a moles de soluto por litro de solución. Dado que 1 L de cualquier líquido corporal contiene una cifra muy cercana a 1 kg de agua, la distinción es de interés académico en situa- 300 mM NaCl 150 mM NaCl 300 mM urea Volumen celular 150 mM NaCl ciones clínicas, y el lector tal vez escuche los términos usados como sinónimos. Asimismo, las presiones reales rara vez se comentan; más bien, se mencionan los osmoles. La tonicidad es un concepto que se relaciona con la osmolaridad, pero es un caso especial para las células. Se dice que una solución es isotónica si no causa ni disminución del volumen de células ni tumefacción de las mismas. Una solución de NaCl 150 mM (9 g/L o 0.9%) es isotónica para las células de mamífero, e isoosmótica para el contenido de la célula. Una solución de urea 300 mM también es isoosmótica para el contenido de la célula, pero una célula colocada en esta solución se hinchará y por último mostrará lisis o estallará (figura 3-12). La solución de urea es hipotónica; tiene insuficiente tonicidad para evitar que la célula se hinche. Difiere de la solución de NaCl porque la urea puede cruzar la membrana celular. La adición de un material permeable a una solución aumenta su osmolaridad, pero no su tonicidad. La adición de más solutos impermeables hace una solución hipertónica; una solución de NaCl 300 mM es hipertónica y hará que las células disminuyan de volumen. Si se añade un soluto permeable a una solución isotónica (p. ej., urea 300 mM + NaCl 150 mM), las células disminuirán de volumen de manera transitoria y después volverán a su volumen original (figura 3-13). La tasa a la cual disminuyen de volumen es proporcional a la permeabilidad de la membrana al agua; la tasa a la cual se recupera el volumen es proporcional a la permeabilidad de la urea. Si se vuelve a añadir la solución de NaCl 150 mM original, ocurrirán los efectos opuestos. Las células se hincharán a medida que entra agua con rapidez, y después vuelven a su volumen original conforme la urea (y el agua) sale de la célula. En algunos casos, es conveniente considerar un coeficiente de reflexión como una descripción de la permeabilidad de solutos. El movimiento de agua a través de paredes capilares depende de la diferencia de presión mecánica o hidrostática, y de la diferencia de presión coloidosmótica debido a diferencias de la concentración de proteína en el plasma y el líquido intersticial. Si la pared capilar es impermeable por completo a las proteínas, se dice que tiene un coeficiente de reflexión de 1.0. Si las paredes empiezan a permitir escape, el coeficiente de reflexión disminuye, entran proteínas al espacio intersticial, y van seguidas por agua. El movimiento de agua en todo el cuerpo se relaciona con dos compartimientos: intracelular y extracelular. El compartimiento extracelular tiene dos subcompartimientos: el líquido plasmático en los vasos sanguíneos y el líquido intersticial, que baña el resto de las células. El plasma y el líquido intersticial están separados por las Tiempo FIGURA 312 Las células disminuyen de volumen en soluciones hipertónicas, y se hinchan en soluciones hipotónicas. (Modificada con autorización de Landowne D: Cell Physiology, New York: Lange Medical Books/McGraw-Hill, 2006.) CAPÍTULO 3 Membranas celulares y mecanismos de transporte 150 mM NaCl + 300 mM urea 150 mM NaCl Volumen celular 150 mM NaCl 29 Tiempo FIGURA 313 La adición de urea causa disminución de volumen transitoria, pero no cambia la tonicidad de estado estable. (Modificada con autorización de Landowne D: Cell Physiology, New York: Lange Medical Books/McGraw-Hill, 2006.) paredes capilares, que son libremente permeables a todas las moléculas pequeñas e iones, pero en circunstancias normales evitan que las proteínas plasmáticas entren al líquido intersticial. Las proteínas tienen una carga negativa neta general al pH de la sangre. El equilibrio que surge con proteínas impermeables e iones libremente permeables se llama equilibrio de Gibbs-Donnan. Este efecto produce pequeños gradientes de concentración de iones (<3%) y un potencial pequeño (algunos milivoltios, negativo en la luz) a través de las paredes capilares. Para casi todos los propósitos clínicos, puede hacerse caso omiso de éstos, y puede considerarse que las concentraciones iónicas en el plasma, que se miden con facilidad, representan el líquido extracelular en general. Las proteínas plasmáticas son importantes desde el punto de vista osmótico; tienden a mantener el agua en los vasos sanguíneos. El equilibrio entre la presión hidrostática y esta presión osmótica “coloide” es descrito mediante la hipótesis de Starling (capítulo 26). Regula el flujo de agua a través de endotelios capilares. El edema representa la pérdida de este equilibrio. Las proteínas intracelulares promueven la entrada de agua a las células que es contrarrestada por la extrusión de iones sodio por bombas de Na/K. Las membranas celulares no son libremente permeables a iones, y el compartimiento intracelular no está en equilibrio de Gibbs-Donnan con el espacio extracelular. Los cambios a corto plazo en los volúmenes y la osmolalidad de compartimientos corporales pueden calcularse con los cuatro principios que a continuación se presentan: en cada compartimiento el volumen por la osmolaridad da el número total de osmoles. El agua se moverá entre los compartimientos para hacer igual la osmolaridad en todos ellos. La cantidad total de agua y el número total de osmoles es la suma de las cantidades en los compartimientos. Cualquier sustancia osmóticamente activa, que no puede pasar a través de una membrana semipermeable, añadida, permanecerá fuera de la célula; el agua añadida se distribuirá entre los compartimientos de acuerdo con los primeros tres principios. Estos cálculos no incluyen los efectos del sistema renal (capítulo 45) que actuará para restituir los volúmenes y las osmolaridades originales. Por ejemplo, una persona tiene 70 kg de peso con 16 L de líquido extracelular y 24 L de líquido intracelular, para un total de 40 L. Si la osmolaridad de estos compartimientos es de 300 mOsm, el compartimiento extracelular contiene 16 × 0.3 = 4.8 osm, y el intracelular contiene 7.2 osm, para un total de 12 osm. Si este estudiante ingiriera 1 L de agua de mar que contuviera 1 osm de sales, en su mayor parte NaCl, el agua total aumenta a 41 L, y el número total de osmoles aumenta a 13, de modo que la nueva osmolaridad es de 13/41 = 317 mOsm. El compartimiento extracelular tendrá 5.8 osm, de modo que su nuevo volumen será de 5.8/0.317 = 18.3 L. El nuevo volumen intracelular será de 7.2/0.317 = 22.7 L. Note que el agua salió de las células para diluir el agua de mar. TRANSPORTE A TRAVÉS DE CÉLULAS EPITELIALES Muchas capas de células epiteliales separan funcionalmente dos soluciones con distintas composiciones, y actúan de una manera coordinada para transportar de manera selectiva solutos y agua a través de la capa. Esto se logra al tener uniones intercelulares herméticas (zonas de oclusión) entre los lados de las células epiteliales, de modo que la capa de células sea impermeable a sustancias que no pueden pasar a través de las membranas celulares, y al incorporar bombas y canales selectivos apropiadamente en las dos superficies de la capa. Los dos lados pueden tener distintos nombres en diferentes epitelios. La membrana apical mira hacia la luz o hacia el exterior del cuerpo; puede conocerse como la membrana luminal o mucosa, o como el borde en cepillo por el aspecto de sus microvellosidades. La membrana basolateral que mira hacia el interior del cuerpo se puede conocer como la membrana serosa o peritubular. En la figura 3-14 se muestran vías para el transporte de Na+ y glucosa a través de capas de células epiteliales. Las bombas de Na/K en la membrana basolateral mantienen bajo el Na+ intracelular al moverlo hacia el líquido extracelular. El Na+ puede entrar a la célula al moverse en favor de su gradiente de concentración a través de canales ENaC en la membrana apical y salir por medio de la bomba en el otro lado. La glucosa se puede introducir a la célula a través de la membrana apical, contra su gradiente de concentración por el SGLT, y después moverse en favor de su gradiente de concentración a través del uniportador de glucosa (GLUT) en la superficie basolateral. Cuando se mueven solutos a través de membranas epiteliales, el agua suele fluir de manera osmótica, “siguiendo” el soluto. Este efecto tiene importancia para la terapia de rehidratación para combatir la pérdida de agua propia de la diarrea. Añadir glucosa y sal al agua que bebe estimulará el SGLT para que mueva Na+, glucosa y agua hacia la célula. La bomba de Na/K y el transportador GLUT a continuación moverán los solutos hacia el cuerpo, y el agua los seguirá. 30 SECCIÓN II Fisiología celular Unión intercelular hermética (zona de oclusión) ENaC Bomba de Na/K Na Na K Mucosa apical luminal ATP Serosa basal ADP Na Glucosa SGLT Glucosa GLUT FIGURA 314 El sodio y la glucosa son transportados a través de capas de células epiteliales mediante una combinación de bombas, canales y transportadores. (Modificada con autorización de Landowne D: Cell Physiology, New York: Lange Medical Books/McGraw-Hill, 2006.) El agua y el material hidrosoluble pueden moverse a través de epitelios mediante transcitosis, o por medio de endocitosis mediada por receptor. La sustancia es captada hacia vesículas mediante endocitosis en una superficie, y es liberada sin cambios por medio de exocitosis en la otra superficie, o se desintegra en endosomas y los productos son liberados por transportadores. La transcitosis ocurre a través de endotelios capilares; la endocitosis mediada por receptor es importante en el riñón y el hígado. CORRELACIÓN CLÍNICA Cuando pasó delante de su entrenador y sus amigos en la marca del kilómetro 24 del maratón de Boston, la corredora de 28 años de edad sonrió y agitó la mano alegremente. Se veía bien conforme pasó Heartbreak Hill, a unos 9.5 km de la meta. Unos 3 km más adelante, se detuvo para beber un vaso de líquido. Otro corredor la recuerda diciendo que se sentía mareada y desorientada. Empezó a tambalearse, y dijo a una amiga que estaba corriendo junto a ella que sentía débiles las piernas, y después se derrumbó en el pavimento. Cuando llegó al hospital, carecía de capacidad de respuesta y tenía signos vitales estables. Después de intubación endotraqueal, se encontró que el valor de sodio sérico en su sangre era muy bajo, de 113 mmol/L. La tomografía computarizada del cerebro y la radiografía del tórax mostraron edemas cerebral y pulmonar difusos. Se le administró solución salina isotónica por vía intravenosa (150 mmol/L), pero nunca recuperó el conocimiento. En el examen post mortem se encontró edema cerebral difuso. Algunos días más tarde, los diarios reportaron que murió por una enfermedad que se conoce como encefalopatía hiponatrémica. La hiponatremia, definida por una concentración de sodio en sangre de menos de 135 mmol/L, puede llevar a encefalopatía hipotónica con edema cerebral mortal. De 488 corredores en el maratón de Boston del 2002 que proporcionaron una muestra de sangre utilizable en la línea de meta, 13% tuvo hiponatremia y 0.6% tuvo hiponatremia crítica (120 mmol/L o menos). En el estudio se concluyó que ocurre hiponatremia en una fracción considerable de corredores de maratón que no pertenecen a la élite de esta especialidad, y que puede ser grave. Por lo general, se origina por beber grandes cantidades de líquido que exceden la capacidad de los riñones para excretar agua durante el ejercicio. El aumento considerable de peso durante la carrera, un tiempo de carrera prolongado, y extremos de índice de masa corporal se asociaron con hiponatremia, no así el sexo femenino, la composición de los líquidos ingeridos, y el uso de antiinflamatorios no esteroideos. Los casos leves pueden manejarse mediante restricción de líquidos hasta el inicio de la producción de orina. Las manifestaciones de encefalopatía hiponatrémica indican la necesidad de tratamiento urgente con soluciones hipertónicas, como solución salina a 3% (513 mmol/L). El Na+ y el Cl– explican casi toda la fuerza osmótica del suero. Si la concentración de Na+ es baja, el suero será hipotónico y el agua se moverá hacia las células del cuerpo y hará que se hinchen (edema). Esto puede tener efectos graves en el cerebro porque está en el espacio cerrado del cráneo, y el tejido tumefacto restringirá el flujo sanguíneo y, por ende, el aporte de oxígeno. La tumefacción puede causar hernia del cerebro a través del tentorium y el agujero occipital, lo cual comprime el tallo encefálico y causa paro respiratorio. La concentración de sodio en sangre está influida por la ingestión de sal y agua, la sudoración y la secreción en la orina, y está regulada por el sistema endocrino. La recomendación para corredores de maratón es que sólo beban líquidos cuando tengan sed. CAPÍTULO 3 Membranas celulares y mecanismos de transporte ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ Una membrana de superficie de bicapa lipídica con proteínas embebidas separa las células y las conecta con el ambiente extracelular circundante. Las moléculas de lípido de la membrana celular son anfipáticas, con grupos hidrofóbicos que miran hacia el interior de la membrana y grupos hidrofílicos que ven ambas superficies acuosas. Las proteínas de la membrana celular llevan a cabo funciones específicas al actuar como canales, bombas, transportadores, receptores o moléculas de adhesión celular. Las proteínas de membrana celular son anfipáticas, por lo general con una o más hélices TM hidrofóbica. Los canales iónicos son proteínas de membrana con un poro que selecciona para el tipo de ion o iones que pasan por el canal en favor de su gradiente electroquímico. Los canales mecanosensitivos, por lo general, son selectivos para catión y tienen estructuras diversas. Los canales sensibles a voltaje tienen una simetría cuádruple. Cada una de las partes tiene seis hélices transmembrana, una de las cuales porta múltiples aminoácidos con carga positiva. Hay muchas familias diferentes de canales quimiosensitivos o sensibles a ligando que corresponden a distintas sustancias químicas. Los canales célula-célula conectan el interior de una célula con el interior de una célula adyacente por medio de una vía acuosa que permite el paso de iones y otras moléculas pequeñas. Las bombas mueven iones y otras moléculas contra sus gradientes a expensas de ATP. Los transportadores mueven algunos iones y otras moléculas contra sus gradientes a expensas del movimiento de otros iones en favor de sus gradientes. Los GPCR inician cascadas de proteína G intracelulares bajo el control de activadores extracelulares. Algunos materiales pueden sólo difundirse con su gradiente de concentración a través de los lípidos de membrana. Muchas sustancias tienen mecanismos de difusión facilitada específicos que se caracterizan por una afinidad y una velocidad de transporte máxima. La presión osmótica es proporcional a la osmolaridad o la concentración total de todos los solutos. La tonicidad describe la capacidad de una solución para evitar la disminución de volumen de células o la tumefacción de las mismas. Las sustancias pueden ser transportadas a través de capas de células epiteliales mediante combinaciones de bombas y transportadores dispuestos en lados opuestos de las células. PREGUNTAS DE ESTUDIO 1. Las membranas celulares A) constan casi por completo de moléculas de proteína. B) son impermeables a sustancias liposolubles. C) contienen moléculas de fosfolípido anfipáticas. D) son libremente permeables a electrólitos, pero no a proteínas. E) tienen una composición estable durante toda la vida de la célula. 2. En una célula epitelial intestinal, el transporte de glucosa desde la luz del intestino hacia la sangre comprende, ¿cuál de los procesos que siguen? A) transporte activo secundario B) difusión facilitada C) transporte activo D) transporte activo secundario y difusión facilitada E) transporte activo y transporte activo secundario 3. Se prepara una solución al añadir 10 g de NaCl (peso de la fórmula = 58.5) a 1 L de agua destilada. Una solución isotónica es 300 mOsm. La solución preparada es A) muy hipotónica (con menos de 50% de la tonicidad normal). B) un poco hipotónica (alrededor de 10% baja). C) isotónica (dentro de 1%). D) un poco hipertónica (alrededor de 10% alta). E) muy hipertónica (más de dos veces la tonicidad normal). 4. Beber solución salina isotónica disminuirá A) el volumen extracelular. B) la osmolaridad extracelular. C) el volumen intracelular. D) la osmolaridad intracelular. E) ninguna de las anteriores. Tasa de transporte RESUMEN DEL CAPÍTULO 31 Gradiente de concentración 5. El diagrama anterior es típico para la dependencia del gradiente de concentración de A) la tasa de transporte activo secundario. B) la tasa de transporte activo primario. C) la tasa de transporte mediante difusión pasiva. D) la tasa de transporte mediante difusión facilitada. 6. ¿Cuál de los que siguen tiene menos probabilidades de afectar la actividad de la bomba de Na/K? A) glucósidos cardiacos B) segundos mensajeros (p. ej., cAMP y diacilglicerol) C) concentración intracelular DNA D) concentración extracelular de Mg2+ E) concentración extracelular de K+ Canales y control del potencial de membrana David Landowne C A P Í T U L O 4 O B J E T I V O S ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ Describir cómo los potenciales de membrana se miden y proporcionan valores típicos para diferentes células. Comentar la relación entre la separación de carga a través de la membrana y el potencial de membrana. Listar las concentraciones aproximadas de los principales iones en los compartimientos intracelular y extracelular. Describir los tres factores que controlan el movimiento de iones a través de membranas. Determinar si un ion se moverá hacia adentro o hacia afuera de la célula dado el potencial de membrana y el gradiente de concentración del ion. Comentar cómo el potencial de membrana cambia cuando fluyen iones a través de las membranas celulares. Explicar los pasos que ocurren durante la generación de un potencial de Nernst. Explicar los pasos que suceden durante la generación de un potencial de membrana en reposo. Comentar por qué el flujo neto de carga es de cero en el estado en reposo aun cuando se están moviendo iones a través de la membrana. Comentar la función de la bomba de Na/K en la generación del potencial de membrana. Definir el registro de canal aislado y describir el flujo de corriente a través de canales de K aislados. Describir los dos tipos de diseminación de información eléctrica en células nerviosas y musculares. Comentar por qué la membrana celular actúa como un capacitor y qué propiedades confiere esto a las células nerviosas y musculares. Discutir la diferencia entre constantes de longitud (espacio) y tiempo, y las relaciones de éstas con la conducción nerviosa. Explicar el estado estable y las propiedades de cable transitorias de células nerviosas y musculares. por alrededor de 0.01 a 0.1 V o 10 a 100 mV. Los gradientes de concentración de iones a través de la membrana son la fuente inmediata de energía para crear el potencial de reposo y mantenerlo. Este potencial de reposo es necesario para la excitabilidad eléctrica de células nerviosas y musculares, la recepción sensorial, el cálculo en el SNC y para ayudar a regular la transferencia de iones a través de la membrana. INTRODUCCIÓN Todas las células vivas tienen una diferencia de potencial eléctrico a través de sus membranas de superficie. Las células actúan como baterías en miniatura; la celda de la pila se nombra así por la célula biológica. En reposo, el interior de las células es negativo respecto al exterior 33 34 SECCIÓN II Fisiología celular 0 mV Microelectrodo Afuera Afuera Adentro Adentro FIGURA 41 Los potenciales de membrana se –90 mV Osciloscopio miden con microelectrodos llenos con soluciones electrolíticas. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) Músculo MEDICIÓN DE POTENCIALES DE MEMBRANA En la figura 4-1 se muestra cómo se miden los potenciales de reposo. Se fija un músculo al fondo de una placa que está llena con una solución salina isotónica con una composición iónica similar a la de la sangre. Un microelectrodo con una punta fina hecha de vidrio y que se llena con KCl 3-M, se coloca sobre una de las células musculares. Un alambre con cloruro de plata en el microelectrodo se fija a una terminal de un dispositivo de medición de voltaje que despliega un trazo de voltaje contra tiempo. La otra terminal se fija a otro alambre con cloruro de plata colocado en la placa; éste se llama el alambre de tierra. Cuando el microelectrodo se encuentra en la solución, está al mismo potencial que el alambre de tierra, y el osciloscopio marca 0 mV. Cuando se hace avanzar el microelectrodo algunos micrómetros hacia la célula muscular, el trazo en el osciloscopio salta de manera repentina alrededor de –90 mV, y permanece ahí en tanto el microelectrodo está colocado. Cuando se retira el electrodo, el trazo regresa a 0 mV. El experimento se puede repetir. Si se inserta un segundo microelectrodo, mide el mismo potencial, lo que indica que los electrodos no están creando el potencial de alguna manera. Cuando el microelectrodo está dentro de la célula, el KCl permanece en contacto con el citoplasma que está en contacto con la membrana. El alambre de tierra se encuentra en contacto con la solución externa, que está en contacto con el exterior de la membrana. La diferencia de potencial es a través de la membrana; se llama el potencial de membrana. El potencial de membrana particular medido cuando la célula está en reposo —es decir, no activa— también se llama el potencial de reposo. Diferentes células tienen distintos potenciales de reposo. Las células de músculo esquelético y cardiaco tienen un potencial de reposo de alrededor de –90 mV; las neuronas sensoriales y motoras, uno de casi –70 mV; las células de músculo liso, uno de alrededor de –60 mV, y los eritrocitos, uno de aproximadamente –10 mV. positiva y negativa exactamente equilibradas excepto por la capa dentro de alrededor de 1 nm desde la superficie de la membrana. Las soluciones masivas a ambos lados son eléctricamente neutras. Las cargas excesivas de signo opuesto experimentan una fuerza de atracción entre sí, pero se evita que lleguen una a la otra porque no pueden abandonar con facilidad las soluciones acuosas y entrar en la membrana lípida oleosa. Cualquier carga dentro de la membrana también experimenta esta fuerza, que tiende a tirar de las cargas positivas hacia adentro y empujar las cargas negativas hacia afuera. El voltaje a través de la membrana es la medición eléctrica de esta fuerza electromotriz o este potencial para el movimiento de cargas si sucede que están dentro de la membrana. El voltaje es directamente proporcional a la cantidad de carga (Q) que está separada. La proporción entre carga separada y el voltaje se llama capacitancia de membrana: Q C = __ V (1) La carga eléctrica se mide en términos de coulomb (C); hay 96 484 C/mol de carga (constante de Faraday). La unidad de capacitancia es el farad (F); 1 C/1 V es 1 F. La capacitancia es la capacidad para almacenar cargas separadas. Muchas computadoras pequeñas usan un capacitor para almacenar suficiente carga para permitir que permanezca algo de función mínima durante un corto tiempo mien- SEPARACIÓN DE LA CARGA El potencial de membrana en reposo es un reflejo de la separación de las cargas a través de la membrana. Existe un poco de exceso de cargas negativas (alrededor de 1 pmol/cm2) en la superficie interna, y el mismo número de cargas positivas excesivas en la superficie externa (figura 4-2). Las soluciones en los dos lados contienen aproximadamente 150 mmol/L de cationes y aniones (cuadro 4-1) con cargas FIGURA 42 La separación de carga. Izquierda: una capa única de cargas separadas por la membrana. Derecha: añadidura de una representación de las cargas móviles en las soluciones masivas. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) CAPÍTULO 4 Canales y control del potencial de membrana CUADRO 4-1 Concentraciones de algunos iones de importancia a través de una membrana de célula muscular.a Concentración extracelular (mM) Ion Concentración intracelular Eion (mV) Cationes Na+ 145 12 mM +65 K+ 4 155 mM −95 Ca2+ 2.5 100 nM +132 132 4 mM −90 ~0 155 mM 22 8 mM Aniones Cl− A− − HCO3 −26 35 10% en diferentes personas o diferentes músculos para los valores de Na+, K+ y Cl–. En circunstancias normales, el Ca2+ externo es de alrededor de 2.5 mM, pero el Ca2+ interno puede cambiar de manera notoria con la actividad; aumenta por arriba de 1 μM cuando el músculo se está contrayendo. La membrana celular es permeable a todos los iones listados en el cuadro 4-1 excepto A–; pueden moverse a través de la membrana mediante diversos canales. El transporte activo de Na+, K+ y Ca2+ mantiene los gradientes. Si la membrana celular queda dañada o no hay aporte adecuado de ATP, la célula ganará Na+ y Ca2+, y perderá K+. La membrana no es muy permeable a iones; en comparación con el agua, su permeabilidad es insignificante. Sin embargo, es el control de esta permeabilidad iónica lo que regula el potencial de membrana y los movimientos pequeños (por estándares químicos) de iones que cambian el potencial de membrana. aA– representa aniones que carecen de la capacidad para cruzar una membrana semipermeable, dentro de la célula. Muchos son polivalentes; en conjunto, contribuyen con menos de 155 mOsm a la presión osmótica. También hay otros osmolitos sin carga en la célula. tras se está cambiando la pila. La membrana almacena las cargas opuestas al mantenerlas separadas. La capacitancia de membrana es de alrededor de 1 μF/cm2. Un pmol de iones univalentes transporta alrededor de 100 nC de carga. Colocar estos valores en la ecuación (1) significa que separar 1 pmol/ cm2 de iones univalentes producirá un potencial de membrana de 100 mV. GENERACIÓN DEL POTENCIAL DE REPOSO La membrana separa dos soluciones con composiciones iónicas bastante diferentes. Se cree que las concentraciones extracelulares representan la concentración de estos iones en el agua de mar en el momento en que el ancestro antiguo abandonó el mar. Son alrededor de una tercera parte de la concentración actual en el agua de mar. La generación del potencial de reposo y todos los cambios de potencial (como el potencial de acción y los potenciales sinápticos) depende de los gradientes de concentración de iones a través de la membrana celular. En el cuadro 4-1 se presentan algunos valores típicos para un músculo esquelético. Ambos lados son eléctricamente neutros; la suma de cargas positiva y negativa es igual. La solución externa tiene concentraciones relativamente altas de Na+ y Cl–, y concentraciones modestas de K+ y Ca2+, mientras que la solución interna es alta en K+ y baja en Na+ y Cl–, y muy baja en Ca2+; tiene una concentración alta de otros aniones (A–), aminoácidos con carga negativa en las proteínas y como grupos fosfato en las proteínas o ácidos nucleicos. Hay un gradiente de concentración hacia adentro para Na+ y Cl–, y un gradiente de concentración hacia afuera para K+. El gradiente de sodio es de alrededor de 10 veces; el de Cl–, de casi 30 veces, y el de K+, de alrededor de 40 veces. En el cuadro 4-1 se indica un gradiente de concentración hacia adentro de 25 000 veces para el Ca2+. En este cuadro se proporcionan números exactos para facilitar el cálculo de ejemplos más adelante en este capítulo. Existe una variación normal de casi FACTORES QUE CONTROLAN LOS MOVIMIENTOS DE IONES El movimiento de iones a través de la membrana es proporcional a la fuerza impulsora neta sobre ellos, que es el gradiente electroquímico o la diferencia entre la fuerza impulsora debida al gradiente de concentración, y la fuerza debida al gradiente de voltaje o potencial de membrana. El movimiento de partículas cargadas es una corriente eléctrica. La corriente, I, transportada por un ion particular, X, se relaciona con la fuerza impulsora mediante la expresión que sigue: Ix = gx (V − Ex) (2) donde Ex es la fuerza impulsora química para el ion x expresada como un potencial eléctrico; esto se describe con mayor detalle más adelante. V es el potencial de membrana y (V – Ex) es la fuerza impulsora sobre el ion x. La conductancia de membrana para el ion x es gx. La conductancia de membrana general para el ion x es proporcional al número de canales para ese ion, N; la probabilidad de que un canal esté abierto, Po, y la conductancia de un canal abierto único, γ, o: gx = NPoγ (3) La conductancia es proporcional a la permeabilidad de la membrana (la facilidad con la cual se mueven iones a través de ella). También la conductancia es proporcional a la concentración del o los iones que se están conduciendo. En ausencia de iones de sodio, un canal de sodio puede ser permeable (si está abierto), pero no conducirá corriente alguna. El gradiente de voltaje empuja un ion o tira de éste porque el ion está cargado. El gradiente de concentración es una fuerza conjugada; los iones tienden a moverse desde una concentración alta hacia una concentración baja. Más iones llegarán a un canal abierto desde el lado con concentración más alta que desde el lado con concentración más baja, de modo que habrá un flujo a favor del gradiente de concentración en proporción al gradiente. Para determinar el flujo neto de un ion a través de la membrana es necesario conocer el gradiente de concentración, el gradiente de voltaje (el potencial de membrana), y la conductancia para el ion. A menos que se conozcan los tres factores, es imposible predecir el flujo del ion. Las dos fuerzas sobre el ion provenientes de los gra- 36 SECCIÓN II Fisiología celular Na+ ClC V 60 mV 132 log –1 4 ECI = –90 mV C V Cl– –90 mV ECI = K+ C V Na+ –90 mV ENa = 60 mV 145 log +1 12 ENa = +65 mV K+ –90 mV EK = 60 mV 4 log +1 155 EK = –95 mV FIGURA 43 La fuerza impulsora sobre iones que cruzan a través de la membrana, gradientes de voltaje (V), y gradientes de concentración (C) para los tres iones más comunes en las soluciones en los líquidos intracelular y extracelular. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) dientes de voltaje y de concentración pueden actuar en la misma dirección o en direcciones opuestas. EL POTENCIAL DE EQUILIBRIO DE NERNST Para cualquier gradiente de concentración particular, es posible tomar un gradiente de voltaje que es igual y opuesto, de modo que el término que aparece entre paréntesis en la ecuación (2) es cero, y no hay corriente neta. Esto se llama potencial de equilibrio electroquímico, o potencial de Nernst, y es dado por: C Ci RT __o Ex = ___ ln Fz e (4) donde Ex es el potencial de Nernst (o el potencial de equilibrio o potencial de difusión) para el ion, Co y Ci las concentraciones en el exterior y el interior de la célula, z la carga del ion o la valencia, R la constante de gas molar, T la temperatura absoluta y F la constante de Faraday. RT es la energía térmica del material a la temperatura T, y RT/F es esta energía expresada en unidades eléctricas. A temperatura ambiente, RT/F es de alrededor de 25 mV. A la temperatura corporal, 37 °C, la ecuación puede simplificarse a: C Ci 60mV __o Ex = _____ z log10 (5) con z = +1 para Na+ o K+, +2 para Ca2+, –1 para Cl–, y así sucesivamente. El potencial de equilibrio para un ion es el potencial al cual el flujo neto es cero. Suele calcularse de manera teórica usando la ecuación (5) sin conocimiento del potencial de membrana real. Es una manera de expresar el gradiente de concentración en términos eléctricos, de modo que el gradiente de concentración se puede comparar con el gradiente de voltaje. Los potenciales de Nernst para los diversos iones que aparecen en el cuadro 4-1 se listan en la última columna. En la figura 4-3 se comparan tres de estos potenciales de equilibrio con un potencial de reposo de –90 mV. Para el cloruro, el gradiente de concentración es hacia adentro; el Cl– se movería hacia la célula porque existe una concentración más alta afuera. El potencial de reposo de –90 mV ejerce una fuerza hacia afuera sobre los iones cloruro que tienen carga negativa. Estos dos son iguales y opuestos, es decir (V – ECl) = –90 – (–90) = 0 mV, y los iones cloruro están en equilibrio electroquímico. Para el sodio, el gradiente de concentración también es hacia adentro, pero el potencial de membrana negativo ejerce una fuerza hacia adentro sobre el Na+ que tiene carga positiva. Ambas fuerzas son hacia adentro y los iones de sodio están lejos de encontrarse en equilibrio, es decir (V – ENa) = –90 – (+65) = –155 mV. Si la membrana fuera permeable a Na+, éste entraría con facilidad. Para el potasio, el gradiente de concentración es hacia afuera, mientras que la fuerza proveniente del gradiente de voltaje es hacia adentro. La magnitud del gradiente de concentración es un poco más grande que la del gradiente de voltaje, esto es (V – EK) = –90 – (–95) = +5 mV. Los iones potasio no están en equilibrio; tienen una tendencia a abandonar la célula. El cloruro es el único ion que se encuentra en equilibrio de los que se presentan en el cuadro 4-1. Los Cl– están distribuidos en equilibrio o muy cerca del equilibrio en células de músculo esquelético, pero no en la mayoría de las células nerviosas. GENERACIÓN DEL POTENCIAL DE NERNST El potencial en reposo tiene su valor particular debido a los gradientes de K+ y Na+, y porque la membrana en reposo es mucho más permeable a K+ que a Na+. Es más fácil entender esto al considerar primero una membrana que está separando el mismo gradiente, que sólo es permeable a K+; esa membrana podría construirse al reconstituir canales de K biológicos hacia una bicapa lipídica artificial (figura 4-4). Cuando se añaden por vez primera las soluciones a los compartimientos, hay potencial de membrana de cero. El K+ empezará a moverse a favor de su gradiente de concentración y, así, moverá carga positiva desde el compartimiento B hacia el A, dejando un exceso de carga negativa en el lado B, y produciendo un exceso de carga CAPÍTULO 4 Canales y control del potencial de membrana A K+ cero electrónicamente y después se liberara. Esa condición puede arreglarse con el aparato de pinzamiento de voltaje descrito en el capítulo 6. Para entender el proceso, es necesario conocer los gradientes de concentración listados en el cuadro 4-1, y que la permeabilidad de la membrana al K+ es 50 a 100 veces mayor que su permeabilidad al Na+. Empezando a potencial de membrana de 0 mV, el K+ iniciará a desplazarse hacia afuera de la célula, mientras que el Na+ empezará a moverse hacia adentro, ambos moviéndose a favor de sus gradientes de concentración. Sin embargo, se moverá más K+ que Na+ porque la permeabilidad al K+ es mucho mayor que la del Na+, de modo que una carga positiva neta se moverá hacia afuera de la célula, lo cual hará el interior de la célula negativo respecto al exterior. El potencial de membrana negativo que se está desarrollando se opone al flujo de salida adicional de K+, y actúa para elevar el flujo de entrada de Na+; esta tendencia continuará; el potencial de membrana se hará cada vez más negativo hasta que tres Na+ estén entrando a través de los canales de Na por cada dos K+ que están saliendo a través de los canales de K. En este punto se llegará a un estado estable porque la bomba de Na/K está extrudiendo tres Na+ y captando dos K+ por cada ciclo que consume ATP. No hay flujo neto en este estado estable, de modo que el potencial de membrana no cambiará en tanto el aporte de ATP sea adecuado (figura 4-5). Es importante percatarse de que la principal función de la bomba es indirecta; la bomba es muy importante para mantener los gradientes, pero sólo contribuye con algunos milivoltios directamente al potencial de membrana. Si el experimento “empezando en 0 mV” se repitiera con la bomba bloqueada mediante ouabaína (un glucósido cardiaco similar a la digital) o la falta de ATP, los procesos iniciales serían los mismos, y el proceso continuaría hasta que el flujo de entrada de sodio fuera igual al flujo de salida de potasio. En este punto el potencial de membrana dejaría de hacerse más negativo y, entonces, muy lentamente, empezaría a moverse de regreso hacia 0 mV puesto que las concentraciones en ambos lados de la membrana cambiaron durante varias horas. Empleando las cifras que aparecen en el cuadro 4-1, es posible estimar la diferencia inmediata en el potencial de membrana que puede atribuirse a la bomba que está trabajando. Cuando el potencial de membrana es de –90 mV, existe una fuerza impulsora neta de 5 mV sobre el K+. Si el potencial de membrana se hizo 2.5 mV menos negativo a –87.5 mV, la fuerza impulsora sobre el K+ estaría aumentada a 50%, de modo que saldrían tres K+ por cada dos que salían a B 4 mM 155 mM K+ Cl– Cl– FIGURA 44 Flujo de K+ a favor de su gradiente de concentración a través de una bicapa artificial que sólo es permeable a K+. El Cl– carece de la capacidad para cruzar una membrana semipermeable, y se forma una separación de carga. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) positiva en el lado A. Esta separación de carga significa que ahora hay un potencial de membrana con el lado B negativo al lado A (o, de manera equivalente, el lado A positivo respecto al lado B). Conforme el lado B se hace más negativo, se reducirá el flujo neto adicional de K de B a A, hasta que finalmente suficiente carga habrá sido separada de modo que el flujo debido a la tracción eléctrica creciente es igual y opuesto al flujo debido al gradiente de concentración. En este punto se habrá alcanzado el equilibrio electroquímico, y el potencial de membrana será igual al potencial de Nernst —en el ejemplo, –95 mV con el lado B negativo al lado A—, lo cual también podría expresarse al decir que el potencial de Nernst es de +95 mV, con el lado A positivo al lado B. Esta es una propiedad de la ecuación (5), porque log A/B = –log B/A. Note que se necesita menos de 1 pmol/cm2 de flujo de carga positiva para establecer el potencial de membrana. Las concentraciones masivas de K+ en los dos lados de la membrana no han cambiado de manera significativa. El cambio de las concentraciones es indetectable mediante experimentos químicos ordinarios. POTENCIAL DE REPOSO El ejemplo anterior puede extenderse para explicar el potencial de reposo en una célula muscular al considerar la situación que ocurriría si el potencial de membrana se mantuviera de forma artificial en Na K 37 3 Na 2 K FIGURA 45 Iones que fluyen a favor de su gradiente de concentración a través de canales y que son transportados de manera activa contra su gradiente de concentración por bombas. (Modificada con autorización de Landowne D: Cell ATP ADP Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) 38 SECCIÓN II Fisiología celular –90 mV. Habría un decremento de 2.5 mV de la fuerza impulsora sobre el Na+, pero esto es menos de 2% de la fuerza impulsora de 155 mV, de modo que haría un cambio insignificante del flujo de entrada de Na+, y entrarían tres Na+ por cada tres K+ que salieran. Así, alrededor de 87.5 mV del potencial de reposo proviene de los gradientes, y otros 2.5 mV provienen directamente de la bomba. Si se conocen las concentraciones y las conductancias iónicas, el potencial de membrana puede calcularse usando la ecuación (4) para encontrar los potenciales de Nernst y la ecuación (2) para ubicar las corrientes. Cuando el potencial de membrana no está cambiando, no existe corriente neta. Si la bomba no está funcionando y la membrana sólo conduce Na+ y K+, INa = –IK o gNa(V – ENa) = –gK(V – EK), que puede reordenarse para resolver V: gNaENa + gKEK V = _________ gNa + gK (6) El potencial de membrana es el promedio ponderado de los potenciales de equilibrio; la ponderación es por sus conductancias respectivas. Si gK >> gNa, el potencial de membrana estará cerca de EK; si gNa >> gK, estará cerca de ENa, y si son iguales, estará a la mitad entre ambos. Si la membrana sólo es permeable a estos dos iones, y no hay una fuente externa de corriente eléctrica, el potencial de membrana siempre estará entre EK y ENa. Estos conceptos se harán más útiles cuando las conductancias cambien, como se muestra en los tres capítulos siguientes. Cuando la membrana en reposo es preferentemente permeable a potasio, el potencial en reposo es sensible a la concentración externa de potasio (figura 4-6). Aumentar el K externo llevará el potencial de membrana más cerca a cero, o despolarizará la membrana. La membrana en reposo en su ambiente iónico normal se considera polarizada. Un cambio de potencial en la dirección positiva, hacia 0 mV, es una despolarización. Un cambio en la otra dirección, que hace el potencial de membrana más negativo, es una hiperpolarización. El Ko aumentado despolariza membranas porque reduce el gradiente de K+ a través de la membrana y hace que EK esté más cerca de cero. Esto reduce la tendencia del K+ a salir de la célula, de modo que el equilibrio se alcanza a un potencial menos negativo. El Ko+ aumentado es un estado peligroso, en potencia mortal, porque las células excitables requieren el potencial de reposo normal para permanecer excitables. Quizá la duplicación de la concentración de K+ en sangre (hiperpotasemia) altere la función del músculo cardiaco. LOS CANALES KIr APOYAN EL POTENCIAL DE REPOSO Algunas células, entre las que destacan las de músculo cardiaco y esquelético, tienen canales Kir que están abiertos y, así, conduciendo, al potencial de reposo, y se cree que son el principal contribuyente a la conductancia de K en reposo. Estos se nombraron rectificadores internos o hacia adentro cuando se demostró en experimentos que la corriente hacia adentro a través de ellos, cuando el potencial de membrana estuvo hiperpolarizado más allá de EK, fue de mayor tamaño que la corriente hacia afuera observada cuando la membrana estuvo despolarizada. Tal vez es un nombre poco afortunado porque, en la vida normal, las membranas nunca experimentan una hiperpolarización tan grande. Los aspectos importantes de la función de este canal son estar abierto para el movimiento hacia afuera de potasio cerca del potencial de reposo, y después hacerse no conductor cuando la célula está despolarizada. Se observará que este bloqueo en el estado despolarizado es importante para los potenciales de acción del músculo cardiaco (capítulo 6). El Kir no es un canal sensible a voltaje. El bloqueo surge porque el Mg2+ u otros cationes polivalentes en el citoplasma intentan pasar por el canal cuando están despolarizados y quedan atorados, lo que evita que el K+ use el canal. Si se estudian los canales en condiciones sin cationes polivalentes, conducen K+ igual de bien en ambas direcciones. 0 Potencial de membrana (mV) –20 –40 –60 –80 –100 EK = 60 mV log [K]o/155 –120 –140 –160 1 10 [K]o mM (note la escala logarítmica) 100 FIGURA 46 El potencial de membrana observado en función de la concentración externa de K+. La línea continua es la predicción teórica para una membrana que sólo es permeable a K+. Advierta la escala de concentración logarítmica. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) 39 CAPÍTULO 4 Canales y control del potencial de membrana ECUACIÓN DE GOLDMAN-HODGKIN-KATZ Si se conocen las permeabilidades, más que las conductancias, se puede calcular un potencial de membrana usando la ecuación teórica de Goldman-Hodgkin-Katz (GHK) o de campo constante: P Na + + P K + + P Cl − PNaNai + PKKi + PClClo Na o K o Cl i V = 60 mV log10 ___________________ + + − (7) Al igual que en la ecuación (6), la ecuación de GHK se simplifica a la ecuación de Nernst si sólo una permeabilidad es mayor de cero. La ecuación de GHK ha sido útil para describir resultados experimentales cuando algunas de las concentraciones se ajustan a cero, lo que hace que los potenciales de Nernst en la ecuación (6) carezcan de sentido. La relación entre permeabilidad y conductancia puede ajustarse de una manera cuantitativa al considerar la conducción cuando el potencial de membrana es de cero y, más tarde, tras multiplicar el flujo químico por la constante de Faraday, igualar las ecuaciones (3-1) y (4-2) para obtener la corriente eléctrica. Así, se tiene: gxEx = PxF ΔCx (8) CAMBIOS DEL POTENCIAL DE MEMBRANA El potencial de membrana cambiará si se inyecta corriente en la célula al abrir canales que permiten que fluyan iones a favor de sus gradientes electroquímicos. Se requiere tiempo para cambiar el potencial de membrana; no saltará de forma instantánea a un nuevo valor. Muchas células nerviosas y musculares son bastante largas, de más de 1 m para algunas células nerviosas. El efecto de una corriente localizada se propagará de manera pasiva desde el sitio de inyección, pero puede no cambiar el potencial de toda la célula. Estos efectos temporales y espaciales son compartidos por los cables eléctricos, y se denominan las propiedades de cable. Pueden entenderse al considerar la capacitancia de membrana, la resistencia de membrana y la resistencia citoplasmática longitudinal entre diferentes partes de la célula. La propagación pasiva mediante propiedades de cable debe distinguirse de la propagación activa mediante potenciales de acción. Los efectos pasivos ocurren sin cambio alguno del número de canales abiertos. Si entra suficiente corriente a un axón nervioso y lo despolariza por arriba del umbral, se desencadenará un potencial de acción y se propagará sin pérdida de amplitud en toda la longitud de la célula. El potencial de acción es regenerado conforme se propaga. A medida que la onda de abertura de canales de sodio se mueve, se proporciona energía al proceso desde el gradiente de Na+ a todo lo largo del axón. En contraste, una despolarización de menor tamaño o una hiperpolarización que no abre canales de Na, sólo se propagará algunos milímetros, y disminuirá progresivamente conforme se aleja del sitio del estímulo. La capacitancia de membrana es la proporción entre la carga separada y el potencial de membrana —ecuación (1)—. La capacitancia se relaciona con las características geométricas de membrana mediante la ecuación que sigue: K × área C = ________ Grosor (9) donde K es una constante que describe la composición material de la membrana. Si el área es de mayor tamaño, requerirá una cantidad elevada de carga para cambiar el potencial. Mientras más delgada sea la membrana, más cerca están las cargas una de otra, y más cargas tendrán que moverse para cambiar el potencial. La capacitancia de una membrana típica es de alrededor de 1 μF/cm2; este valor a menudo se usa para estimar el tamaño de una célula al medir su capacitancia. La resistencia de la membrana es el recíproco de la conductancia de la membrana: 1 Rm = __ g m (10) La resistencia longitudinal es proporcional a la longitud, e inversamente proporcional al área de corte transversal: ρ × longitud Área Rl = __________ (11) donde ρ es la resistividad del contenido de la célula. PROPIEDADES PASIVAS DE UNA CÉLULA REDONDA PEQUEÑA Cuando se inyecta un pulso de corriente en una célula redonda pequeña (que puede suponerse que tiene el mismo potencial de membrana en toda su superficie), el potencial de membrana no cambia de manera instantánea. En lugar de eso, lo realiza con una evolución temporal exponencial, con una constante de tiempo (τ) característica, el tiempo que se requiere para descargar el cambio de voltaje a 1/e = 37 por ciento de su valor (o el tiempo que se requiere para cambiar a 63% de su valor final) (figura 4-7). De inicio, las cargas inyectadas se están sumando a las cargas almacenadas que crearon el potencial de membrana original. Más tarde, cuando el potencial de membrana ha alcanzado un nuevo estado estable, que es cuando una corriente igual a la corriente inyectada está escapando de regreso hacia afuera a través de canales de membrana. Cuando se termina el impulso, la carga almacenada excesiva escapa a través de los canales, y el potencial de membrana declina de manera exponencial a su valor original. La constante de tiempo de estos cambios exponenciales es el producto de la resistencia y la capacitancia de la membrana de la célula. Muchas células tienen constantes de tiempo dentro del rango de 1 a 20 ms. Estas constantes de tiempo limitan la rapidez con la cual el potencial de membrana puede cambiar, y permite la suma temporal de eventos sinápticos en el sistema nervioso central (capítulo 7). PROPIEDADES PASIVAS DE UNA CÉLULA CILÍNDRICA LARGA Una célula extendida o un tejido con células que están eléctricamente conectadas mediante uniones intercelulares comunicantes (conexiones comunicantes) puede tener diferentes potenciales de membrana en distintas ubicaciones. Si existe un cambio local de permeabilidad, fluirá corriente hacia adentro o hacia afuera de la célula, y el potencial de membrana cambiará en ese sitio y, en menor grado, en lugares cercanos. Con una corriente estable prolongada, que dura mucho 40 SECCIÓN II Fisiología celular I V Adentro C I R V Afuera A B I ΔV = IR[1 −exp(−t/τ)] V 63% 37% τ ΔV = IR exp(−t /τ) τ C FIGURA 47 Una célula esférica (A), su circuito equivalente (B) y la respuesta de voltaje a un pulso de corriente inyectado (C). (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/ McGraw-Hill, 2006.) más tiempo que la constante de tiempo descrita en la sección previa, habrá un cambio estable del potencial que es de mayor magnitud en el punto de entrada de la corriente, y disminuye de manera exponen- I cial con la distancia, con una constante de longitud (λ) o constante de espacio característica, que es la distancia que se requiere para que el potencial disminuya a 37% de su valor en el sitio de inyección (figura 4-8). Las constantes de longitud típicas para células nerviosas y musculares son de 0.1 a 2.0 mm. Una célula de 10 μm es aproximadamente isopotencial, pero una célula de 150 cm de largo requiere un mecanismo de propagación activo para ser capaz de comunicar actividad eléctrica de un extremo a otro. El cambio de voltaje declina porque parte de la corriente inyectada escapa de la célula y no está disponible para despolarizar las regiones adyacentes. La cantidad que escapa es proporcional al cambio de voltaje, de modo que la declinación es exponencial. La constante de longitud depende de la proporción entre las resistencias de membrana y la axoplasmática longitudinal. A medida que la distancia desde la inyección se incrementa, la amplitud de la respuesta transitoria disminuye, y el tiempo de aumento se hace más prolongado y sigmoideo (figura 4-9). De modo inicial casi toda la carga que entra a la célula va a la membrana inmediatamente adyacente a la fuente; sólo más tarde está lo bastante disponible como para cargar la membrana distal. Cuando se termina el impulso, todas las respuestas declinan a la misma tasa. Las sinapsis están distribuidas en el árbol dendrítico a diferentes distancias desde el cuerpo celular. Las sinapsis más distantes tendrán menos efecto sobre la actividad de la célula; la amplitud del efecto será más baja, y su evolución temporal, más lenta. La propagación pasiva es importante para la propagación del potencial de acción; es el mecanismo de conexión entre la región activa y la región en reposo adyacente. Los potenciales de acción se propagan con mayor rapidez en axones de mayor diámetro porque V1 V2 V3 A RI Adentro I C RI Rm RI Rm Rm RI Rm Afuera B 1 ΔV(x) = ΔVo exp(−x/λ) 2 3 37% λ Distancia x C FIGURA 48 Una célula larga (A), su circuito equivalente (B) y la distribución de estado estable de su potencial de membrana en respuesta a una inyección continua de corriente (C). (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) CAPÍTULO 4 Canales y control del potencial de membrana I V2 V1 41 V3 I 1 2 V 3 FIGURA 49 Las respuestas de voltaje transitorias a tres distancias desde el sitio de inyección de un pulso de corriente. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) tienen resistencia longitudinal más baja y constantes de longitud más largas. Las propiedades pasivas, la capacitancia de membrana, la resistencia de membrana y la resistencia longitudinal se denominan propiedades de cable porque determinan la capacidad de cables subacuáticos para transmitir señales. La constante de longitud para cables submarinos es de varios kilómetros; para axones nerviosos varía de alrededor de 0.1 a 20.0 mm, dependiendo del diámetro. Los cables submarinos dependen de amplificadores repetidores para distancias más grandes; los nervios usan canales de sodio dependientes de voltaje (capítulo 6). Cuando las uniones célula-célula unen células, éstas pueden operar eléctricamente como si todas fueran una célula. Muchas de las células en el corazón están acopladas, y los potenciales de acción se propagan de una célula a otra apoyados por la propagación pasiva de la despolarización por medio de las uniones célula-célula. Además hay uniones célula-célula entre algunas neuronas en el SNC. Para algunas personas es útil visualizar una analogía hidráulica de estos fenómenos eléctricos. El voltaje eléctrico es análogo a la presión de agua, y la corriente eléctrica, al flujo de solución. La célula larga es similar a una manguera que tiene escapes; la resistencia de membrana más baja corresponde a más escapes, y la resistencia longitudinal más baja corresponde a un mayor diámetro de la manguera. RESUMEN DEL CAPÍTULO ■ ■ ■ Un potencial de membrana eléctrico es directamente proporcional a la separación de cargas positiva y negativa a través de la membrana celular. La proporción entre carga separada y voltaje es la capacitancia de la membrana. Las membranas celulares separan soluciones con composiciones iónicas bastante distintas. El movimiento de iones es directamente proporcional a la fuerza impulsora neta sobre los iones. La fuerza impulsora neta es el gradiente electroquímico o la diferencia entre el efecto del potencial de membrana y el efecto del gradiente químico. ■ ■ ■ ■ ■ ■ ■ El efecto del gradiente químico puede expresarse mediante el potencial de equilibrio de Nernst. Sólo un número muy pequeño de iones debe separarse para producir el potencial de membrana. Esto es insignificante en comparación con las concentraciones disponibles en ambos lados. El potencial de membrana en reposo es un estado estable con movimiento de iones a favor de su gradiente electroquímico a través de canales y bombeo de un número igual contra su gradiente electroquímico a expensas de ATP. La ecuación de GHK puede usarse para calcular el potencial de membrana si se conocen las permeabilidades a los diversos iones y sus concentraciones. Cuando fluye corriente a través de la membrana, el potencial de membrana cambia en tiempo y espacio, regido por las “propiedades de cable”. Cuando se inyecta un pulso de corriente en una célula, hay un tiempo característico que se requiere para que el potencial de membrana cambie. Cuando se inyecta una corriente estable en una célula larga, el cambio de potencial es mayor en el sitio de inyección y disminuye de manera característica lejos del sitio. PREGUNTAS DE ESTUDIO 1. Si todas las bombas de Na/K en la membrana de una célula muscular se detuvieran, se esperarían todos los cambios que siguen para la célula muscular, excepto: A) pérdida inmediata de la capacidad de la célula para transportar potenciales de acción. B) disminución gradual de la concentración interna de K+. C) aumento gradual de la concentración interna de Na+. D) disminución gradual del potencial de membrana en reposo (el potencial sería menos negativo). E) incremento gradual de la concentración interna de Cl–. 42 SECCIÓN II Fisiología celular 2. Si la concentración de ion potasio en el exterior de una célula de músculo esquelético en reposo se duplica a dos veces el valor normal al añadir K+ y Cl– en cantidades iguales, ¿cuál sería el mejor estimado del efecto sobre el potencial de membrana en reposo? A) hiperpolarizar alrededor de 100 Mv B) despolarizar alrededor de 5 mV C) hiperpolarizar alrededor de 15 mV D) despolarizar alrededor de 20 mV E) ningún efecto medible 3. La célula que sigue en un organismo llamado el piojo Europa se recuperó a partir de una luna de Júpiter con una sonda espacial. Las concentraciones intracelulares y extracelulares de todos los iones se dan como sigue: Extracelular Intracelular Rb+ = 100 mM Rb+ = 1 mM SO42– = 50 mM SO42– = 0.5 mM La membrana celular es permeable a Rb+ y no a SO42– o agua. ¿Cuál es el potencial de membrana en reposo? (El signo se refiere al potencial dentro de la célula.) A) +30 mV. B) +60 mV. C) +120 mV. D) –30 mV. E) –60 mV. 4. Un científico está registrando a partir del cuerpo de una neurona con un microelectrodo intracelular para estudiar las aferencias sinápticas en las dendritas. Las letras a, b y c en los trazos que aparecen a continuación indican los potenciales sinápticos registrados a partir de tres aferencias sinápticas diferentes. Para aferencias sinápticas idénticas a las dendritas, ¿cuál potencial sináptico fue generado por la sinapsis en un sitio de las dendritas más cercano al cuerpo? A B C Potenciales generadores sensoriales David Landowne C A P Í T U L O 5 O B J E T I V O S ■ ■ ■ ■ Listar ocho sensaciones y los nombres de las células receptoras sensoriales especializadas que se encargan de generarlas. Describir la adaptación sensorial en estos receptores. Dibujar un cartel esquemático de a) un corpúsculo de Pacini y su célula ganglionar sensorial (incluso el cuerpo celular y la prolongación central); b) una célula pilosa coclear y sus sinapsis, y c) un fotorreceptor y sus sinapsis. Numerar tres o más diferencias entre canales iónicos que sustentan potenciales de acción, potenciales de reposo y potenciales de receptor. También, cada célula tiene un campo receptivo que es la región en el espacio del estímulo que evoca una respuesta en esa célula. El campo receptivo de un fotorreceptor en la retina es un sitio particular en el espacio visual enfrente del ojo y una gama de colores a los cuales ese receptor es sensible. El campo receptivo para un nervio somatosensorial en la piel es el área de piel que desencadena una respuesta. El campo receptivo para una neurona olfatoria es la gama de sustancias químicas que puede detectar. Las células en el SNC que se relacionan con información sensorial también tienen campos receptivos. La información sensorial proveniente de los pies es manejada por diferentes células de las que manejan la información sensorial proveniente de las manos. La información entrante llega por “líneas marcadas”; los procesadores del SNC saben de dónde proviene. Existen varias ubicaciones en el cerebro que tienen campos receptivos, incluso la misma ubicación en el espacio visual. Los campos receptivos de estas células de orden superior son más complejos, puesto que ha ocurrido un procesamiento de señal que compara las salidas de una célula de orden inferior con las de otras. La transducción mecanosensorial es directa, mediante canales mecanosensitivos en la membrana. La célula sensorial a menudo tiene moléculas o estructuras para enfocar la energía mecánica o filtrar alteraciones mecánicas no deseadas, y puede haber un órgano complejo —como el que comprende los oídos externo, medio e interno— para suministrar la energía mecánica deseada a la célula apropiada. Al final, se abre un canal catiónico relativamente inespecífico, y tanto Na+ como K+ se mueven a favor de sus gradientes de concentración. En mecanorreceptores cutáneos, como el corpúsculo de Pacini (véase más adelante), hay una gran fuerza impulsora sobre el Na+, de Los animales han desarrollado una gran variedad de órganos sensoriales capaces de vigilar sustancias químicas, luz, sonido y otros eventos mecánicos en los ambientes externo e interno. En todos estos órganos existen mecanismos para convertir la información acerca del ambiente en señales eléctricas dentro del sistema nervioso; este capítulo se refiere al proceso de conversión y algunas propiedades generales de todos los receptores. En la Sección IV: capítulos 13, 15, 16 y 17 se abordan más detalles acerca de los órganos sensoriales y los sistemas que procesan las señales nerviosas. Los transductores pueden convertir un tipo de energía en otro. Las células o porciones de las células que realizan el paso inicial de transducción sensorial convierten la energía luminosa o mecánica, o la presencia de condiciones químicas específicas, en un cambio en el potencial de membrana llamado potencial receptor o potencial generador sensorial. En células sensoriales pequeñas, este potencial generador controla de manera directa el proceso de liberación sináptica (capítulo 7). En células más largas, el potencial generador iniciará un potencial de acción que se propaga hacia una terminación presináptica distante y después desencadena el proceso de liberación. La información acerca de la energía del estímulo que fue transducida en un potencial generador a continuación es codificada en la frecuencia de potenciales de acción. Cada célula sensorial tiene un estímulo apropiado, llamado su estímulo adecuado. El sistema nervioso central (SNC) interpreta señales que provienen de esta célula en términos de su estímulo adecuado el cual, para los fotorreceptores en el ojo, es la luz visible. Si se aplica una descarga eléctrica o suficiente presión en el ojo, una persona reportará destellos, incluso si la habitación está oscura. 43 44 SECCIÓN II Fisiología celular FIGURA 51 Los cambios del potencial de membrana de una terminación nerviosa mecanosensorial a estímulos de tres diferentes amplitudes. (Modificada con autorización de Landowne D: Cell Physiology, New York: Lange Medical Books/McGraw-Hill, 2006.) modo que se mueve más Na+ que K+, y la célula se despolariza. El número de canales mecanosensitivos que se abren es proporcional a la magnitud de distensión de la membrana por el estímulo. Un estímulo mayor abrirá más canales y producirá una despolarización de mayor magnitud (figura 5-1). Si la despolarización es suficientemente grande, se inician potenciales de acción y se propagan hacia el SNC. La situación es más compleja en el oído, donde las células sensoriales (llamadas células pilosas sensoriales, porque los cilios modificados en su superficie apical tienen aspecto de pelo) forman parte de un epitelio que separa dos soluciones diferentes. Sin embargo, la alteración mecánica de estas células por el sonido apropiado tam- bién lleva a corriente hacia adentro transportada por K+ a través de canales mecanosensitivos en los cilios, y despolariza la célula. Las células pilosas sensoriales son cortas y hacen sinapsis con células del nervio auditivo en el oído. Las células pilosas no tienen potenciales de acción; son cortas en comparación con su constante de longitud, de modo que pueden depender de la propagación pasiva para abrir canales Cav para liberar transmisores. Parte de la quimiosensación del gusto es apoyada de forma directa por canales quimiosensitivos, como los receptores de glutamato para el sabor umami (el sabor distintivo del glutamato); éstos son canales catiónicos relativamente no selectivos que despolarizan las células. Otros usan canales de manera aún más directa; el Na+ que se mueve a través de canales de sodio epiteliales (ENaC) despolariza células para proporcionar la sensación de sabor salado. Los olores se detectan mediante receptores acoplados a proteína G (GPCR) cuyas proteínas G activan la adenilil ciclasa, lo que aumenta la concentración de monofosfato de adenosina cíclico (cAMP). El cAMP abre un canal catiónico inespecífico sensible a nucleótido, cíclico (CNG) que despolariza la célula. Los canales CNG son tetrámeros con seis segmentos TM, y son estructuralmente similares a canales KV, pero carecen de la selectividad extrema de estos últimos por iones K y la sensibilidad a voltaje. La transducción de luz también involucra GPCR con siete segmentos TM: rodopsina en los bastones, y otras tres opsinas en los conos afinadas para longitudes de onda corta, mediana y larga (o azul, verde y roja). El cromóforo que absorbe la luz es el 11-cis retinal (MW 284). La absorción de un fotón desencadena la conversión del retinal en el isómero holo-trans, que causa un cambio conformacio- Membrana de disco Rodopsina (GPCR) Fosfodiesterasa β/γ Transducina (proteína G) α cGMP GMP hν cGMP Na FIGURA 52 Los procesos que enlazan la absorción de luz por rodopsina y el cierre de los canales sensibles a nucleótido cíclico. La luz induce un cambio conformacional de la rodopsina que hace que las subunidades de transducina se disocien. La subunidad α estimula una fosfodiesterasa que degrada cGMP. En ausencia de cGMP un canal que estaba permitiendo la entrada de Na+ se cierra, y la célula se hiperpolariza. (Modificada con autorización de Landowne D: Cell Physiology, New York: Lange Medical Books/McGraw-Hill, 2006.) CAPÍTULO 5 Potenciales generadores sensoriales nal en la proteína opsina que informa a la proteína G que ha tenido lugar un evento (figura 5-2). La proteína G se llama transducina, fue la primera que se identificó, y se nombró antes de que la familia se conociera bien. La transducina activa una fosfodiesterasa que hidroliza monofosfato de guanosina cíclico (cGMP). En la oscuridad, hay un canal CNG que está abierto y transportando corriente hacia adentro. El canal se cierra cuando la concentración de cGMP disminuye; cuando la luz está encendida, la corriente que existe en la oscuridad disminuye y la célula se hiperpolariza. Hay amplificación a lo largo de esta vía química, de modo que un fotón lleva al cierre de muchos canales CNG. La hiperpolarización reduce una salida estable de vesículas sinápticas para transmitir el mensaje hacia la siguiente célula en la vía al cerebro. La sensación de temperatura cutánea incómodamente caliente se ha enlazado con la activación directa de un canal VR1, que se llama así por receptor de vanilloide. Además, se conoce como el receptor de capsaicina porque puede ser activado por el vanilloide capsaicina, el principal ingrediente picante en los chiles. VR1 es un miembro de la familia de canales de potencial de receptor transitorio (TRP); tiene una estructura 6-TM, y es permeable a cationes. El aumento de la temperatura dentro del rango de 42 °C (107.6 °F) que muchos observadores humanos identifican como dolorosamente caliente, abre este canal, despolariza la terminación sensorial, e inicia una serie de potenciales de acción. Otros miembros de la familia TRP se han relacionado con la sensación de temperatura y otras funciones, aunque no todos con dolor. La experiencia cotidiana de los sentidos no es una representación directa de los estímulos sino más bien el resultado del procesamiento que ocurre en el sistema nervioso. El humano no ve el mundo como destellos en diferentes posiciones de sus campos visuales, sino más bien como objetos y lo que los rodea. El dolor es una experiencia que puede surgir a partir de una amplia variedad de estímulos sin decir necesariamente algo definido acerca del estímulo. Se han identificado algunos nociceptores específicos, pero también hay muchos otros receptores que pueden asociarse con dolor. La elevación del K+ proveniente de células dañadas, o el corte directo de una célula nerviosa, pueden inducir potenciales de acción que suelen interpretarse como dolor. Los canales iónicos que detectan ácido (ASIC) en la familia ENaC muestran respuesta al ácido láctico liberado en el corazón, y despolarizan nervios que proporcionan la vía sensorial para la experiencia dolorosa de angina. Los canales receptores P2X3, que pueden ser activados por trifosfato de adenosina (ATP) liberado por células dañadas, se han vinculado con dolor por sobredistensión de la vejiga, y los receptores P2X4 se han asociado con un dolor neuropático generado dentro del sistema nervioso sin estímulos externos obvios. ADAPTACIÓN SENSORIAL Todos los sentidos, excepto el dolor, se adaptan; si se presentan con un estímulo mantenido, la respuesta disminuirá con el tiempo. El corpúsculo de Pacini se adapta con rapidez y muestra respuesta a un estímulo sostenido con sólo 1 o 2 potenciales de acción al principio (figura 5-3). Cuando se libera el estímulo, hay una respuesta de apagado, y se inicia otro potencial de acción. Casi toda esta adaptación ocurre en la cápsula en forma de cebolla de células accesorias que rodean la terminación nerviosa. Cuando el estímulo deforma un 45 FIGURA 53 Adaptación sensorial rápida y lenta. Las barras a color indican un nivel estable de estimulación. El receptor que se adapta con rapidez a la izquierda se adapta por completo después de que han ocurrido dos impulsos. En el receptor de la derecha, que se adapta de forma lenta, la tasa de activación declina con menos velocidad. (Modificada con autorización de Landowne D: Cell Physiology, New York: Lange Medical Books/McGraw-Hill, 2006.) lado de la cápsula, al principio la deformación es transmitida a la terminación nerviosa y el nervio se despolariza. A continuación la cápsula se abomba hacia los lados, las fuerzas en el nervio se alivian, y el nervio deja de activarse. Cuando se elimina el estímulo, la cápsula rebota a su forma original, lo que empuja de manera transitoria los lados del nervio en el proceso. El corpúsculo de Pacini está afinado para proporcionar información máxima acerca de estímulos vibratorios, y para hacer caso omiso de presión constante. Los órganos del huso muscular son estructuras sensoriales embebidas en músculos esqueléticos, que proporcionan información acerca de la longitud del músculo al SNC (véase figura 2-3 y capítulo 14). Los husos musculares se adaptan con rapidez a cambios de longitud, pero también siguen activándose durante un estímulo sostenido. La tasa de activación sólo disminuye durante el estímulo; se dice que los husos musculares son de adaptación lenta (figura 5-3). El sistema nervioso muestra respuesta a cambios en el ambiente, y al reducir los mensajes que indican que un estímulo aún está presente, puede darse más atención a cualesquier cambios. La adaptación se presenta en muchos niveles —el tejido accesorio antes del potencial de receptor, el potencial de receptor en sí, el mecanismo codificador que inicia potenciales de acción y en muchas sinapsis más altas donde el mensaje que está llegando es integrado con otras señales—. La adaptación a la luz ocurre mediante constricción de las pupilas, fotoblanqueamiento de los pigmentos y regulación por retroacción de los pasos en la cascada bioquímica. Muchos sentidos tienen alguna forma de control eferente. El sistema nervioso simpático puede liberar norepinefrina hacia el corpúsculo de Pacini, lo cual aumentará su sensibilidad a estímulos mecánicos. Los órganos del huso muscular (capítulo 14) tienen nervios eferentes (nervios motores γ) que establecen el rango de longitudes a las cuales el nervio sensorial es más sensible. También existen células pilosas motoras en el oído que pueden incrementar de manera selectiva la sensibilidad de células pilosas sensoriales a sonidos particulares (capítulo 16). Hay muchos controles en el ojo para asegurar que el objeto de interés se enfoque de manera idónea en una porción apropiada de la retina incluso conforme la cabeza cambia su posición en el espacio (capítulo 15). RESUMEN DEL CAPÍTULO ■ ■ Cada célula sensorial tiene un estímulo adecuado. El tacto, la audición y otras mecanosensaciones ocurren por medio de canales mecanosensitivos. 46 ■ ■ ■ ■ SECCIÓN II Fisiología celular El gusto está mediado por canales quimiosensitivos y el olfato por GPCR y canales CNG. También la vista está mediada por GPCR —por ejemplo, rodopsina— y canales CNG. El dolor está mediado por ASIC y canales activados por purina. Todos los sentidos, excepto el dolor, se adaptan. 3. Las células pilosas son las células receptoras sensoriales en la cóclea. Son excitadas por la vibración del haz piloso. ¿La vibración del haz piloso causa cuál de los eventos que a continuación se presentan? A) flujo de entrada de K+ a través de canales catiónicos sensibles a la deformación mecánica en la punta de los cilios B) flujo de entrada de Ca2+ a través de canales sensibles a nucleótido cíclico (CNG) en la punta de los cilios C) hiperpolarización duradera de la célula pilosa PREGUNTAS DE ESTUDIO 10 Respuesta pulsos/s Respuesta pulsos/s 10 5 5 30 50 Segundos A) Respuesta pulsos/s Respuesta pulsos/s 10 10 10 30 50 Segundos B) 10 30 50 Segundos D) 10 5 10 30 50 Segundos C) 2. ¿Cuál de las células sensoriales que siguen tiene un potencial generador hiperpolarizante en respuesta a su estímulo adecuado? A) terminación nerviosa de corpúsculo de Pacini B) nervio del huso muscular C) célula de papila gustativa D) célula de cono retiniano E) terminación nerviosa olfatoria 5 1. Los gráficos indican la frecuencia de potenciales de acción (eje y) registrados a partir de una fibra aferente sensorial primaria durante estimulación sensorial. ¿Cuál de éstos muestra la respuesta de una fibra sensorial típica (excluyendo fibras de dolor) a un estímulo mantenido constante aplicado empezando en 10 s y que dura de principio a fin del registro (es decir, hasta 50 s)? D) una serie de potenciales de acción propagados desde los cilios hacia el cuerpo celular de la célula pilosa Potenciales de acción C A P Í T U L O 6 David Landowne O B J E T I V O S ■ ■ ■ ■ ■ ■ ■ ■ ■ Describir la activación de potenciales de acción. Explicar la propagación de potenciales de acción. Describir las corrientes de membrana que subyacen potenciales de acción. Describir la actividad de canales que producen potenciales de acción. Explicar las características de la membrana que determina el umbral del potencial de acción y el periodo refractario. Explicar las acciones del calcio, los anestésicos locales y las neurotoxinas sobre los potenciales de acción. Describir la relación entre la actividad de canal y la contracción del músculo cardiaco. Describir las características de la membrana de los marcapasos cardiacos intrínsecos. Describir los efectos de la acetilcolina y la norepinefrina sobre potenciales de acción cardiacos. de reposo. Su duración en los nervios y los músculos esqueléticos es del orden de 1 ms; en las células musculares ventriculares cardiacas, su duración es de varios cientos de milisegundos. En nervios y músculos esqueléticos, los cambios de permeabilidad subyacentes son un aumento transitorio de la permeabilidad al sodio seguido, después de un retraso, por un incremento de la permeabilidad al potasio, por la activación de canales de sodio y potasio, respectivamente (figura 6-1). FUNCIÓN DE LOS CANALES DE SODIO SENSIBLES A VOLTAJE Los potenciales de acción son cambios del potencial de membrana que se propagan a lo largo de la superficie de células excitables. Se conocen mejor en las células nerviosas y musculares, pero también ocurren en otras células, entre ellas las células huevo asociadas con la fecundación. A diferencia de otros cambios del potencial de membrana, los potenciales de acción se caracterizan por ser de “todo o nada”; tienen un umbral para excitación y una duración estereotipada. Inmediatamente después de un potencial de acción, la célula excitable tiene un periodo refractario durante el cual es más difícil o imposible desencadenar un segundo potencial de acción. Al igual que casi todos los cambios del potencial de membrana, los potenciales de acción son el resultado de cambios de la permeabilidad de membrana por la actividad de canales, o proteínas embebidas en la membrana de bicapa lipídica que facilitan el movimiento pasivo de iones específicos en favor de sus gradientes electroquímicos. Un potencial de acción es un cambio del potencial de membrana desde un potencial de reposo de alrededor de –70 mV (el interior de la célula es negativo) hasta alrededor de +30 mV y después de regreso al potencial 25 mV 10 mS/cm2 1 ms FIGURA 61 Un potencial de acción (trazo rojo) y los cambios subyacentes de la conductancia de membrana para Na+ (trazo azul) y K+ (trazo beige). (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) 47 48 SECCIÓN II Fisiología celular Los potenciales de acción cardiacos son más complejos e involucran también la activación de canales de calcio. Los potenciales de acción son de todo o nada y se propagan porque los canales de sodio son sensibles a voltaje. La despolarización, la reducción del potencial de membrana, de –70 a 0 mV, induce un cambio conformacional en el transcurso de algunos cientos de microsegundos en la proteína del canal de sodio, lo cual lleva a un aumento de la permeabilidad a iones sodio. Los iones sodio entran con rapidez a la célula a través de estos canales de Na dependientes de voltaje (Nav), y llevan carga positiva con ellos, lo que despolariza más la célula, y abre más canales Nav (figura 6-2). Esta asa de retroacción positiva persiste hasta que todos los canales de sodio están abiertos. Una vez que se inicia el asa, continúa hasta completarse. La despolarización se disemina de manera pasiva hacia regiones adyacentes de la membrana y activa canales de sodio cercanos. Esta onda de cambio conformacional molecular y actividad eléctrica se propaga a lo largo de la superficie de la célula a velocidades de hasta 120 ms. La energía potencial que se almacena en el gradiente de concentración de sodio se utiliza de manera secuencial a lo largo de la vía de propagación. La velocidad de propagación la determina la tasa de cambio molecular y las propiedades eléctricas de la célula que controlan la diseminación de cambios de potencial (propiedades de cable). Alrededor de 1 milisegundo más tarde, los canales de sodio pasan por un segundo cambio conformacional y se desactivan. En esta tercera conformación, están cerrados y el sodio ya no pasa a través de ellos. Además, los canales Nav son incapaces de abrirse de nuevo Cambio de conformación de canal Despolarización del potencial de membrana Aumento de la permeabilidad al sodio Entrada de sodio a la célula FIGURA 62 El ciclo de retroacción positiva del potencial de acción. El ciclo es iniciado por una despolarización, y continúa hasta que todos los canales de sodio se han activado. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) sino hasta que la membrana se repolariza hasta el potencial de reposo durante algunos milisegundos para permitir la recuperación luego de la desactivación (figura 6-3). Este cierre automático de los canales de sodio limita la duración de los potenciales de acción de nervios y de músculo esquelético. La pérdida de la capacidad para abrirse de nuevo produce el periodo refractario. Abierto Activado Cerrado en reposo Cerrado desactivado FIGURA 6-3 Los canales de sodio pueden estar en estados funcionales diferentes. Una despolarización primero hace que el canal cambie desde el estado en reposo hacia los estados activado y abierto, y más tarde al estado desactivado. Se requiere repolarización para ir desde el estado desactivado de regreso hacia el estado en reposo. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) CAPÍTULO 6 Potenciales de acción Vi Vm 49 Axón Alambre axial Vo Vc I FIGURA 64 Un circuito de pinza de voltaje simplificado para un axón gigante de calamar. El potencial de membrana, Vm, es detectado como la diferencia entre el potencial interno, Vi, y el potencial externo, Vo. Vm se compara con el potencial de comando, Vc y si son diferentes fluye una corriente a través del alambre axial y la membrana celular para hacer Vm igual a Vc. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) El movimiento hacia afuera de K+ que transporta carga positiva afuera de la célula produce la repolarización (la fase de disminución del potencial de acción). En algunas células, los canales de K dependientes de voltaje (KV) —cuya activación es más lenta que la de los canales de sodio— facilitan la repolarización. En axones mielinizados de mamífero, la corriente de repolarización pasa a través de los canales de sodio (no sensibles a voltaje) que producen el potencial de reposo. Los axones parecen ser una excepción; las terminales nerviosas presinápticas y los cuerpos celulares de casi todas las neuronas tienen canales KV. PINZA DE VOLTAJE Este entendimiento del mecanismo de potencial de acción proviene de la investigación de Alan Hodgkin y Andrew Huxley hace unos 50 años. Al trabajar con axones nerviosos gigantes aislados a partir del calamar, lograron romper el asa de retroacción positiva y medir el efecto de un cambio del potencial de membrana en las permeabilidades iónicas sin cambio alguno del potencial de membrana debido al movimiento de iones. Su técnica fue incluir la membrana nerviosa en un circuito de retroacción negativa (figura 6-4). Un par de electrodos mide el potencial de membrana; esto a continuación se compara con un potencial de comando deseado. Si un potencial de membrana es diferente del potencial de comando, se hace que fluya una corriente a través de la membrana en una dirección que reduce la diferencia. Así, el voltaje a través de la membrana está pinzado a un valor deseado. Cuando el voltaje controlado es un impulso desde el potencial en reposo hasta 0 mV, pueden identificarse cuatro clases distintas de corriente (figura 6-5). La primera es el movimiento de carga necesario para cambiar el potencial o cambiar la carga sobre la capacitancia de membrana. En segundo lugar, hay una pequeña corriente hacia afuera conocida como corriente de compuerta. A continuación hay una corriente hacia adentro que es reemplazada en algunos milisegundos por una corriente hacia afuera, que dura tanto como el impulso. 0 mV Potencial de membrana con “pinza de voltaje” –70 mV Ic Ig IK INa Hacia afuera Hacia adentro Corriente con iones tanto potasio como sodio FIGURA 65 Las corrientes de membrana (trazo inferior) en respuesta a un impulso de pinza de voltaje (trazo superior). Ic, corriente de capacidad; Ig, corriente de compuerta; INa, corriente de sodio; IK, corriente de potasio. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) 50 SECCIÓN II Fisiología celular 0 mV Potencial de membrana con “pinza de voltaje” -70 mV IK Hacia afuera Corriente sin iones sodio Hacia adentro Corriente sin iones potasio INa 1 ms Ig Corriente sin iones sodio ni potasio FIGURA 66 La separación de corrientes al cambiar las soluciones. Las abreviaturas significan lo mismo que las de la figura 6-5. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) Es posible reemplazar el contenido de un segmento de axón de calamar con una solución salina simple y mantener canales funcionando. Al cambiar las soluciones que bañan ambos lados de la membrana, es posible separar las corrientes transportadas por Na+ (INa) y K+ (IK), y ver también la corriente de compuerta (Ig) aún presente en ausencia de ambos iones (figura 6-6). Note que a 0 mV, la corriente de Na es hacia adentro y la corriente de K hacia afuera. La corriente de Na se activa o aumenta con mayor rapidez que la corriente de K. Se desactiva o disminuye durante el impulso, aun cuando el potencial de membrana se mantiene en 0 mV, mientras que la corriente de K permanece durante el tiempo que dura el impulso. Si el potencial es pulsado a otros potenciales despolarizados, los cuatro componentes de la corriente están presentes, aunque su amplitud y evolución temporal y, en el caso de las INa, la dirección, pueden cambiar (figura 6-7). La corriente de Na se hace más hacia adentro entre el potencial de reposo y alrededor de 0 mV. Los impulsos de mayor tamaño producen menos corriente de Na hacia aden- tro hasta que, alrededor de +60 mV, no pasa corriente neta a través de los canales de Na. Los impulsos de magnitud aún mayor pueden impulsar la corriente de Na hacia afuera a través de los canales de Na. La reversión de la corriente ocurre en el potencial de equilibrio de sodio, ENa. Si se cambia la proporción de las concentraciones de sodio que bañan ambos lados de la membrana, este potencial de reversión también cambia. Con despolarizaciones modestas, la corriente hacia adentro aumenta porque impulsos de mayor magnitud abren más canales de sodio. Sin embargo, el potencial menos negativo disminuye la fuerza impulsora hacia adentro sobre los iones de sodio; después de que casi todos los canales Nav han sido abiertos, despolarizaciones de magnitud aún mayor disminuyen la corriente de Na. Cuando el potencial de membrana excede el potencial de equilibrio de sodio, el Na es forzado hacia afuera de la célula a través de los canales Nav abiertos. En un potencial de acción que corre libre, el potencial de membrana nunca excede el potencial de equilibrio de sodio, y siempre hay una entrada neta de Na a la célula. +80 mV +60 1 mA/cm2 1 ms +40 0 –20 –70 +80 mV +60 +40 0 –20 FIGURA 67 Las respuestas de corriente (trazos superiores) a pasos de voltaje de amplitud variable (trazos inferiores). No se muestran estados transitorios de corriente de capacidad. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) CAPÍTULO 6 Potenciales de acción 1 mA/cm2 10 ms 51 impulsora, se usa para describir casi todos los fenómenos electrofisiológicos restantes en todas las células y tejidos. Las ecuaciones de Hodgkin-Huxley están disponibles en un programa de computación comercial que se conoce como Neuron. El sitio web (http://nerve.bsd.uchicago.edu/nerve1.html) tiene una interpretación en JavaScript que permite manipular las ecuaciones con casi todos los navegadores web modernos. 0 mV –70 FIGURA 68 La recuperación luego de desactivación mostrada mediante un experimento de dos impulsos con cantidades diferentes de tiempo al potencial de reposo entre impulsos. No se muestran estados transitorios de corriente de capacidad. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/ McGraw-Hill, 2006.) La corriente de Na se activa y desactiva con mayor rapidez a medida que se incrementa la magnitud del impulso. Si se da un segundo impulso inmediatamente después del primero, la corriente de compuerta y la corriente de sodio durante el segundo impulso son de menor magnitud que durante el primer impulso (figura 6-8). Ambas se recuperan en paralelo a medida que se aumenta la duración entre los impulsos. La tasa de recuperación luego de desactivación también es dependiente del voltaje, puesto que los canales se recuperan con mayor rapidez a potenciales más hiperpolarizados. La corriente de K se incrementa y se hace más rápida conforme aumenta el potencial de membrana. Por arriba de alrededor de +20 mV, el incremento de la amplitud se hace proporcional al cambio de potencial, lo que indica que todos los canales están abiertos y que sólo la fuerza impulsora aumenta. La corriente de compuerta es un signo directo de los cambios conformacionales en las proteínas del canal Nav. Estas moléculas contienen grupos y dipolos cargados que se mueven o reorientan cuando el campo eléctrico cambia, de manera específica las hélices TM S4 (véanse figuras 3-3 y 3-4). Este movimiento puede medirse como la corriente de compuerta. A medida que el impulso se hace progresivamente más positivo, y más canales de sodio se abren, la amplitud de la corriente de compuerta aumenta, y las corrientes se hacen más rápidas. Por arriba de alrededor de +20 mV, estos dos cambios son complementarios, y el área bajo el trazo de corriente de compuerta es constante, lo que indica que todos los canales pasan por cambios conformacionales, y lo hacen con mayor rapidez a potenciales más positivos. La corriente de capacitancia aumenta de modo lineal con el tamaño del impulso porque requiere más carga para cambiar más el voltaje. Hodgkin y Huxley separaron las corrientes y mostraron cómo las corrientes iónicas fueron proporcionales a la fuerza impulsora sobre los iones. Crearon ecuaciones matemáticas que emularon la amplitud y la evolución temporal de los cambios de permeabilidad, y mostraron que estas ecuaciones podrían predecir la amplitud de potenciales de acción, y la evolución temporal de los mismos, así como su umbral, velocidad de conducción, periodo refractario y otras características. Su concepto de describir la corriente iónica como el producto de tantas veces la conductancia como la fuerza UMBRAL El umbral surge porque las despolarizaciones pequeñas tienen dos efectos diferentes. Por un lado, la despolarización aumentará la probabilidad de que los canales Nav se abran y permitan corriente hacia adentro, lo que llevará a despolarización adicional; por otro lado, la despolarización aleja más el potencial de membrana del potencial de equilibrio de potasio, lo que aumenta la fuerza impulsora neta sobre los iones de potasio y, así, produce una corriente hacia afuera a través de los canales de potasio de potencial de reposo, lo que llevará a repolarización. Si un número suficiente de canales de sodio se abren, de modo que la corriente de sodio hacia adentro excede la corriente de potasio hacia afuera, la célula ha excedido el umbral y seguirá despolimerizándose hasta que todos los canales de sodio disponibles se hayan abierto. Los tratamientos que reducen la corriente de sodio —por ejemplo, la reducción de la concentración extracelular de sodio o la reducción del número de canales Nav— aumentarán el umbral. PERIODOS REFRACTARIOS Durante un potencial de acción, casi todos los canales Nav se activan o abren y después se desactivan y cierran hacia un estado que difiere de su estado antes del potencial de acción. Para recuperarse luego de desactivación y estar disponibles para abrirse de nuevo, los canales Nav deben pasar cierto tiempo con el potencial de membrana cerca del potencial de reposo. No se recuperarán si la membrana permanece despolarizada. Durante esta recuperación, se dice que el axón es refractario porque es resistente a la estimulación. El periodo refractario se divide en dos segmentos: un periodo refractario absoluto cuando ningún estímulo, independientemente de cuán grande sea, puede desencadenar un segundo potencial de acción, seguido por un periodo refractario relativo, cuando el axón puede estimularse de nuevo, pero requiere un estímulo de mayor magnitud para desencadenar la segunda respuesta que el que se necesitó para la primera (figura 6-9). Durante el periodo refractario absoluto, se recuperaron tan pocos canales Nav que aun si todos los canales recuperados se abrieran, la corriente de sodio sería insuficiente para exceder la corriente de potasio hacia afuera, lo que tiende a restituir el potencial de reposo y a mantenerlo. Durante el periodo refractario relativo, se requiere una despolarización de mayor magnitud porque una fracción más grande de los canales Nav disponibles debe ser abierta para obtener el mismo número de canales abiertos en el primer estímulo. Además, en muchas células nerviosas y musculares hay más canales de potasio abiertos después de un potencial de acción, lo cual hace que la célula sea más difícil de excitar una segunda vez. 52 SECCIÓN II Fisiología celular 3 Umbral relativo Absoluto Relativo 2 1 0 0 2 4 ms 6 FIGURA 69 Los periodos refractarios absoluto y relativo. El eje del tiempo empieza con un potencial de acción. Durante el periodo refractario absoluto, ningún estímulo, independientemente de qué tan grande sea, puede desencadenar un segundo potencial de acción. Durante el periodo refractario relativo puede desencadenarse un segundo potencial de acción, pero requiere un estímulo de mayor magnitud que el que se requiere en el estado de reposo. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) MIELINIZACIÓN ENFERMEDADES Los sistemas nerviosos de vertebrados presentan una especialización de la función nerviosa que no se observa en invertebrados, a saber, la mielinización (figura 6-10). Células accesorias envuelven axones nerviosos con muchas capas de su membrana externa, lo que aísla eléctricamente casi toda la célula. Los canales Nav se agrupan en las regiones entre estas envolturas, en los nodos de Ranvier. La corriente de Na sólo entra a la célula en estos nodos; la excitación “salta” de un nodo a otro en lo que se llama conducción saltatoria. La diseminación entre nodos es la misma diseminación pasiva que se observa en células nerviosas no mielinizadas, pero es más eficaz, es decir, produce una velocidad de conducción más rápida. Las envolturas de mielina aumentan la resistencia entre el axoplasma y los medios circundantes, lo cual, a su vez, incrementa la constante de longitud para la diseminación pasiva. La mielina también aumenta el grosor efectivo, lo cual disminuye la capacitancia efectiva y reduce la cantidad de carga que se requiere para cambiar el potencial. Ambos efectos aceleran la conducción. Hay muchas enfermedades caracterizadas por una excitación reducida o excesiva de células. Quizá la más familiar es la conducción de información de dolor agudo, que a menudo se trata con anestésicos locales; éstos actúan al bloquear los canales Nav. Algunas formas de epilepsia y algunas arritmias cardiacas también se tratan con bloqueadores de los canales Nav. Un tipo de síndrome de QT largo (LQT), una arritmia cardiaca, se ha enlazado con una mutación en uno de los genes que codifican para el canal de Na+, y una parálisis periódica hiperpotasémica (hiperKPPP) se ha enlazado con otro. Otros síndromes de LQT se asociaron con los canales KV. La hipocalcemia se relaciona con excitabilidad aumentada de nervios y músculo esquelético, y puede producir contracción muscular incontrolable (tetania). La hipercalcemia hace a los nervios y músculos menos excitables. El calcio se une a la membrana cerca del sensor de voltaje S4 (véase figura 3-4) del canal Nav, y tiene un efecto similar a la hiperpolarización. La carga positiva en el ion calcio repele la hélice S4 que tiene carga positiva, lo que hace más difícil que el S4 se FIGURA 610 El efecto de la mielinización sobre la diseminación longitudinal de corriente. En el diagrama superior se muestra el Na+ entrando (flecha a color) en un nodo de Ranvier, y las asas de corriente asociadas se muestran en negro. En un nervio no mielinizado (diagrama inferior) ocurren las mismas asas de corriente, pero en una distancia más corta; por ende, el potencial de acción se propaga con mayor lentitud. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) CAPÍTULO 6 Potenciales de acción mueva hacia afuera y abra el canal. El resultado es que, en condiciones de calcio bajo, el canal de sodio se abre en respuesta a un estímulo de menor magnitud o incluso de modo espontáneo al potencial de reposo. La unión de calcio no cambia el potencial de reposo según se mide con electrodos en los compartimientos masivos en ambos lados de la membrana. Hay enfermedades desmielinizantes, como la esclerosis múltiple (MS), en la cual se pierde la mielina y la conducción puede hacerse más lenta o fracasar. La MS es una enfermedad autoinmunitaria y, por lo general, se trata con corticosteroides sintéticos, como la prednisona. Los síntomas pueden aliviarse al proporcionar aire acondicionado o mudarse hacia un clima más frío. El enfriamiento ayuda, de modo un poco paradójico, porque si bien lentifica la abertura de los canales de sodio y, así, lentifica la velocidad de propagación, también lentifica la desactivación de canales Nav y aumenta la duración de los potenciales de acción; así, el mayor flujo de entrada de Na+ hace más fiable la propagación. La fiabilidad a menudo se comenta en términos de factor de seguridad para la propagación. En individuos sanos, el potencial de acción de 100 mV que llega al siguiente nodo de Ranvier es de tamaño unas cinco veces mayor que la repolarización de 20 mV que se requiere para iniciar un nuevo impulso en ese nodo. En pacientes con MS, el potencial de acción que llega al siguiente nodo puede estar disminuido a cerca del tamaño necesario para reiniciar el impulso, o por debajo de dicho tamaño. Un efecto del enfriamiento de nervios es aumentar el factor de seguridad para la propagación. 1 1 2 2 3 3 53 FÁRMACOS Y TOXINAS Después de la identificación de estas conductancias de Na+ y K+ específicas, se mostró que están separadas en el ámbito molecular porque difieren en sus propiedades farmacológicas y muestran respuesta diferente a diversos medicamentos. La tetrodoxina (TTX), un veneno que se encuentra en los órganos internos del pez globo, bloquea de manera selectiva canales Nav de nervios a concentraciones nanomolares. Los anestésicos locales, como la lidocaína o benzocaína, también bloquean los canales Nav. Hay mayor diversidad entre los canales KV y entre los fármacos que los bloquean. Los iones tetraetil amonio (TEA) y 4-aminopiridina figuran entre los bloqueadores de canal KV. También hay compuestos que activan de manera crónica canales Nav, como la veratridina, insecticidas piretroides, y brevetoxina, una de las toxinas de la marea roja. REGISTROS EXTRACELULARES: POTENCIALES DE ACCIÓN COMPUESTOS Los potenciales de acción pueden registrarse con un par de electrodos colocados sobre la superficie de un fascículo nervioso, separados alrededor de 1 cm. Cuando un impulso nervioso pasa por estos electrodos, se observa un potencial de acción bifásico en el monitor (figura 6-11); éste es un registro diferencial del mismo impulso ner- 4 FIGURA 611 Potenciales de acción 5 2 1 2 3 5 1 3 registrados externamente. Izquierda: potencial de acción bifásico registrado a partir de un axón intacto. Derecha: potencial de acción monofásico apuntado cerca del sitio de una lesión por aplastamiento. El potencial se mide entre los dos círculos que se muestran arriba de cada diagrama. Los números en los trazos indican la cronología del diagrama asociado arriba. La región en color dentro de la célula nerviosa se propaga de izquierda a derecha. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/ 4 McGraw-Hill, 2006.) 54 SECCIÓN II Fisiología celular α δ 1 mV 10 ms β B 1 μV 50 ms C γ δ FIGURA 612 Un potencial de acción compuesto. Izquierda: rapidez de barrido alta. Derecha: rapidez de barrido más baja, ganancia vertical más alta. Las letras se refieren a grupos de axones específicos dentro del nervio. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) vioso que aparecería como en la figura 6-1 si el registro se hiciera con un microelectrodo intracelular. Una deflexión ocurre a medida que el impulso pasa por el primer electrodo, y la segunda sucede conforme pasa por el segundo electrodo. Van en direcciones opuestas porque los dos electrodos llevan a entradas opuestas para el monitor. Si el nervio es aplastado entre los electrodos, de modo que el impulso no llega al segundo electrodo, la respuesta se hace monofásica. Este tipo de registro con electrodos externos se usa en clínica para probar la integridad de nervios. Un fascículo nervioso también se puede estimular con otro par de electrodos en un tramo remoto del mismo fascículo. Con equipo apropiado, pueden realizarse estimulación y registro a través de la piel sin disecar el fascículo nervioso. Cuando un fascículo nervioso se estimula, puede excitarse más de un axón. El registro eléctrico de la combinación de potenciales de acción producidos se llama potencial de acción compuesto; este último también es bifásico si el nervio está intacto entre los electrodos de registro. Además de ser bifásicos, hay muchas diferencias entre potenciales de acción compuestos registrados con electrodos externos y el potencial de acción de célula única registrado con un electrodo dentro de la célula y un electrodo de referencia fuera de la célula. Los potenciales de acción compuestos son de tamaño mucho menor, del orden de 1 mV, y no hay signo del potencial de reposo porque ambos electrodos están fuera del nervio. El potencial de acción compuesto no es de todo o nada porque un estímulo de mayor magnitud llevará más axones individuales por arriba del umbral, y la amplitud del potencial de acción compuesto es proporcional al número de axones que se activan. El potencial de acción compuesto se hace de menor tamaño y más largo a distancias mayores desde los electrodos estimulantes porque la velocidad de conducción de los diversos axones no es exactamente la misma, y los potenciales de acción se dispersan conforme viajan en dirección contraria al sitio de estimulación. El umbral y la velocidad de conducción de los diversos axones dentro de un fascículo nervioso varían con el diámetro de los axones. Los axones grandes tienen un umbral más bajo a la estimulación por electrodos externos (por supuesto, in vivo, por lo general, son estimulados de manera más selectiva por un receptor o aferencia sináptica específica). Las fibras de mayor diámetro tienen un umbral más bajo; una cantidad mayor de la corriente estimulante fluye a través de ellas porque tienen una resistencia interna más baja. Los axones de mayor tamaño también tienen una velocidad de conducción más rápida, de nuevo por su resistencia interna más baja. Los axones periféricos de vertebrados se clasifican por su diámetro (o velocidad de conducción o umbral a la estimulación externa). Hay grupos de fibras nerviosas con diámetros similares. Los grupos de diferentes diámetros pueden distinguirse como elevaciones separadas en el potencial de acción compuesto (figura 6-12). Hay cierta correlación de la función con el diámetro; por ejemplo, las motoneuronas mielinizadas grandes que van a músculos esqueléticos son fibras Aα, y las fibras no mielinizadas pequeñas que llevan información sobre dolor son fibras C. Las fibras de mayor tamaño tienen velocidades de conducción más rápidas y umbrales más bajos a estímulos eléctricos externos. POTENCIALES DE ACCIÓN CARDIACOS El corazón es una bomba hecha de células musculares excitables. La actividad eléctrica de estas células controla su contracción. La función de estas células se comentará más en el contexto de la función del corazón en el capítulo 23. El control general del patrón de contracción del corazón se efectúa con la diseminación de potenciales de acción a través de un sistema conductor especial de células musculares cardiacas modificadas (fibras de Purkinje) y a través de las células musculares auriculares y ventriculares mismas (figura 23-3). Hay dos tipos de potenciales de acción en el corazón que se distinguen por su tasa de despolarización y su velocidad de conducción. Los potenciales de acción rápidos, con una tasa de despolarización rápida y una velocidad de propagación rápida, se encuentran en células musculares auriculares y ventriculares, y en fibras de Purkinje. Los potenciales de acción lentos se encuentran en el nodo sinoauricular (SA) y el nodo auriculoventricular (AV). POTENCIALES DE ACCIÓN DEL MÚSCULO CARDIACO En los potenciales de acción del músculo cardiaco, corriente que proviene de células adyacentes despolariza la célula hasta una magnitud en la cual los canales Nav dependientes de voltaje, rápidos, se abren, y despolarizan con rapidez la membrana hacia el potencial de equilibrio de sodio (fase 0 en la figura 6-13). Estos canales son similares a los canales de sodio de nervios y de músculo esquelético; se abren en respuesta a la despolarización. También son bloqueados por anestésicos locales. Después de abrirse, se desactivan con rapidez y el potencial de membrana empieza a regresar. Sin embargo, la despolarización también abre canales Cav tipo L activados por voltaje que no se desactivan. Esto mantiene el potencial de acción en la fase de meseta (fase 2). La reducción de la concentración externa de CAPÍTULO 6 Potenciales de acción 1 2 Vm 50 mV 0 3 50 ms 4 IK ICa 1 mA/cm2 INa FIGURA 613 Un potencial de acción de célula muscular ventricular (trazo superior) y sus corrientes iónicas subyacentes. Las corrientes INa e ICa son hacia adentro, y la corriente IK es hacia afuera. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) Ca2+ al añadir fármacos que bloquean los canales de calcio reducirá esta última fase y la fuerza de la contracción muscular. La contracción del músculo cardiaco, a diferencia de la del músculo esquelético, requiere Ca2+ externo (figura 6-13). Las células del músculo cardiaco también difieren de los nervios y del músculo esquelético porque carecen de canal KV rápido para repolarización rápida. El sistema de conductancia de potasio del corazón es más bien complejo; se han identificado, al menos, cinco componentes distintos con base en su cinética y dependencia de voltaje. Dos de éstos son importantes para entender la fase de meseta. Durante esta última fase, la conductancia es menor que durante la diástole, el periodo entre potenciales de acción. Esto se debe al canal rectificador interno (hacia adentro) (Kir), que se encarga de mantener el potencial de reposo y tiene una conductancia alta, cerca del potencial de reposo y por debajo del mismo (a potenciales más negativos); no conduce durante la fase de meseta cuando la membrana está despolarizada. El canal Kir rectifica, lo que permite que la corriente fluya y mantenga el potencial de reposo, pero no permite que mucha corriente fluya hacia afuera durante la despolarización. La rectificación es causada por Mg2+ u otros cationes polivalentes provenientes de la solución interna que se mueven hacia el canal y lo taponan cuando la célula está despolarizada. La conductancia baja para K+ durante la fase de meseta significa que la conductancia modesta al Ca2+ a través de canales Cav mantiene el potencial de membrana a cifras despolarizadas durante la meseta. Los canales KV lentos se abren con mucha lentitud durante el potencial de acción, y la pendiente descendente durante la fase de meseta depende de ellos. Cuando el potencial de membrana cae por debajo de una cierta magnitud, los canales Cav se cierran y la repolarización hacia el potencial de equilibrio de potasio se acelera (fase 3). Puesto que la membrana ya no está despolarizada, los canales KV se cierran. La descripción anterior es una vista simplificada de los potenciales 55 de acción del músculo cardiaco. La historia completa revela más canales de K, y debe explicar diferencias entre los potenciales de acción musculares en distintas regiones del corazón, así como cambios relacionados con la edad. Hay dos canales KV que se abren de manera transitoria justo después de los canales Nav y producen la repolarización parcial inicial (fase 1) desde el máximo hasta la meseta (IKto). Hay al menos dos canales de K dependientes de voltaje lentos diferentes, con cinética similar, pero con propiedades farmacológicas distintas (IKR e IKS). Algunas células musculares cardiacas tienen canales de calcio tipo T. En todas las células cardiacas, algo de corriente es transportada por el intercambiador de sodio-calcio y por la bomba de Na/K. Las diferencias regionales y de edad en los potenciales de acción tienen importancia funcional y clínica. Los potenciales de acción del músculo ventricular cerca de la superficie endocárdica (interna) son de mayor duración que los que están cerca de la superficie epicárdica (externa). Las fibras internas realizan más trabajo, y tienen más probabilidades de quedar dañadas en un infarto de miocardio. Estas diferencias deben surgir debido a un equilibrio distinto de actividades de los canales de Na, Ca y K. Las interacciones entre los efectos de diferentes canales son complejas, y se exploran mejor con modelos de computadora. Está claro que se necesita más información para entender los detalles. POTENCIALES DE ACCIÓN DE LOS NODOS SA Y AV En circunstancias normales, el control general del patrón de contracción del corazón es iniciado por potenciales de acción que surgen de manera espontánea 60 a 80 veces/min a partir de células musculares modificadas en el nodo SA. También se observan potenciales de acción similares en el nodo AV, donde regulan la activación de los ventrículos. En ausencia de estimulación desde las aurículas, las células del nodo AV producen de manera espontánea alrededor de 40 potenciales de acción/min; sin embargo, en corazones sanos, las células auriculares los impulsan a la tasa establecida por el nodo SA. Los potenciales de acción en los nodos carecen de la deflexión positiva rápida, y no tienen una fase de meseta tan pronunciada como los potenciales de acción del músculo cardiaco. Además, se caracterizan por la despolarización lenta entre potenciales de acción: el potencial de marcapaso; estas células se activan de manera rítmica; nunca están en reposo y no tienen potencial de reposo verdadero. La deflexión positiva del potencial de acción es producida por una corriente hacia adentro lenta transportada por Ca2+ (figura 6-14). Hay una fase inicial a través de canales Cav tipo T y una fase mayor a través de canales Cav tipo L. Los canales tipo T son transitorios y tienen un umbral bajo para abertura, de cerca de –60 mV. Los canales tipo L son duraderos y tienen un umbral más alto, de cerca de –30 mV. Los canales tipo L son similares a los canales Cav y mantienen la meseta de los potenciales de acción del músculo cardiaco; son bloqueados por dihidropiridinas. Los canales tipo T tienen propiedades farmacológicas diferentes. La reducción de la concentración externa de Ca2+ o la adición de bloqueadores de canales de Ca2+ reduce la amplitud de los potenciales de acción del nodo. La corriente de K+ hacia afuera reemplaza de manera gradual la corriente hacia adentro lenta, y las células se repolarizan hacia el EK. Conforme el 56 SECCIÓN II Fisiología celular Vm 50 mV 50 ms IK If 1 mA/cm2 ICa FIGURA 614 Los potenciales de acción del nodo SA (trazo superior) y sus corrientes subyacentes. Las corrientes If e ICa son hacia adentro, y la corriente IK es hacia afuera. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) potencial pasa de –50 mV, una corriente activada por hiperpolarización, If, hacia adentro, aparece, compite con IK y, finalmente, empieza a despolarizar la célula de nuevo. La If es transportada principalmente por iones sodio. Cuando el potencial pasa de nuevo de –60 mV, los canales Cav se activan otra vez y el ciclo se repite. EFECTOS DE LAS INERVACIONES SIMPÁTICA Y PARASIMPÁTICA El corazón puede latir de manera espontánea sin aferencias neurales; sin embargo, en individuos sanos, el sistema nervioso autónomo y las concentraciones de hormonas circulantes regulan la frecuencia cardiaca y la fuerza de contracción del corazón. El sistema nervioso autónomo controla muchos órganos internos por medio de sus dos divisiones, los sistemas nerviosos simpático y parasimpático; éstos liberan norepinefrina (NE) y acetilcolina (ACh), hacia el corazón. El sistema nervioso autónomo también puede hacer que la médula suprarrenal libere epinefrina hacia la sangre. La epinefrina tiene efectos sobre el corazón similares a los de la NE. En el capítulo siguiente se describen algunos de los detalles acerca de las sinapsis del sistema nervioso autónomo y sus propiedades farmacológicas. Las células en los nodos SA y AV tienen GPCR que producen una estimulación (por medio de Gαs) o inhibición (mediante Gαi) de la adenilil ciclasa que, a su vez, aumenta o disminuye la concentración de cAMP en respuesta a NE y ACh. El cAMP incrementa la actividad de los canales de If. El resultado final es que la NE aumenta la If y, así, despolariza las células con mayor rapidez e incrementa la frecuencia cardiaca. La ACh aminora la If, lentifica la frecuencia de despolarización, y reduce la frecuencia cardiaca (figura 23-4). El cambio de la If también lleva a una aceleración o lentificación de la conducción a través del nodo AV. Estos efectos se comentan más en términos de la función cardiaca en el capítulo 23. La concentración alta de ACh lleva a la abertura de otro canal de potasio (KACh). (Es un canal GIRK rectificador interno activado por proteína G.) Este canal reduce más la tendencia a despolarizarse entre potenciales de acción, y puede detener temporalmente el corazón. LA NOREPINEFRINA TAMBIÉN AUMENTA LA CONTRACTILIDAD En presencia de NE, la meseta de los potenciales de acción musculares está alta, y tiene una duración más breve (figura 6-15); este acortamiento del potencial de acción acorta la duración de la contracción muscular, que tiene importancia funcional para el corazón. A frecuencias cardiacas altas, el tiempo requerido para que vuelva a llenarse el corazón limita su rendimiento. Al disminuir el tiempo durante el cual se genera fuerza muscular (sístole), se deja más tiempo para el llenado (diástole). El acortamiento de los potenciales de acción ventriculares puede observarse en el ECG como un acortamiento del intervalo QT. La NE aumenta la amplitud de la meseta al hacer que el potencial de acción abra más canales de Ca2+ de tipo L; esto impulsa la membrana más cerca al potencial de equilibrio de Ca. El flujo aumentado de Ca hacia adentro lleva a una mayor fuerza de contracción mediante un mecanismo que se describe en el capítulo 10. La NE acorta la duración al hacer que los canales KV se abran con más rapidez. Los efectos sobre los canales de K y Ca2+ están mediados por cAMP, que actúa como un segundo mensajero, lo que estimula la proteína cinasa A (PKA) y fosforila los canales; esta vía también aumenta el La NE eleva la meseta La NE acorta la duración 50 mV 50 ms FIGURA 615 Los efectos de la norepinefrina sobre potenciales de acción de células musculares ventriculares. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/ McGraw-Hill, 2006.) CAPÍTULO 6 Potenciales de acción mecanismo de recaptación de calcio al fosforilar el fosfolambán. Esto acelera la relajación muscular. LA ACETILCOLINA REDUCE LA CONTRACTILIDAD AURICULAR El canal de K activado por ACh (KACh) permanece abierto durante los potenciales de acción; en el músculo auricular y las fibras de Purkinje, hace la fase de meseta más corta y más baja. Las contracciones auriculares son más débiles. Los receptores de ACh son escasos en células musculares ventriculares. 57 puede causar debilidad de músculo esquelético al despolarizar las células, lo que desactiva canales Nav normales, que entonces son incapaces de generar potenciales de acción. Los pacientes con hiperKPP tienen riesgo aumentado de hipertermia maligna inducida por anestesia durante una intervención quirúrgica. Otras mutaciones del canal Nav en el músculo cardiaco están enlazadas con síndromes de muerte súbita. Los ataques pueden suspenderse al ingerir una carga de azúcar alta o mediante diuréticos tipo tiazida, ambos de los cuales reducen el potasio extracelular. Pueden evitarse con una dieta baja en potasio y alta en carbohidratos, y con tiazidas. La enfermedad dura toda la vida. CORRELACIÓN CLÍNICA Desde principios de su niñez, una mujer de 42 años de edad experimentó rigidez muscular, en particular al aflojar la mano después de empuñarla apretadamente, o al empezar a caminar. La exposición al frío exacerbaba estos síntomas. Al permanecer al aire libre en un día frío y ventoso, presentaba rigidez de la cara que producía una mueca, y no podía abrir los ojos ni moverlos de un lado a otro; estos síntomas desaparecían algunos minutos después de entrar a una habitación más caliente. Cuando comía helado, presentaba rigidez de garganta e imposibilidad para deglutir. Desde los 16 años de edad, también había presentado ataques de debilidad generalizada no relacionados con el frío. A veces se despertaba por la noche gravemente paralizada. La tendencia a presentar un ataque era mayor cuando tenía hambre. Durante el embarazo tuvo ataques diarios de debilidad; mejoró en el transcurso de algunos días después del parto. Un neurólogo realizó pruebas diagnósticas. Se administraron a la paciente 60 mEq de potasio por vía oral con una mezcla de aniones, 45 minutos más tarde, la paciente presentó rigidez de tal magnitud que no podía hacer movimientos rápidos. Alrededor de una hora más tarde, notó debilidad creciente y tuvo que acostarse. El ataque paralítico alcanzó su máximo aproximadamente media hora más tarde. En ese momento, no podía levantar la cabeza, los brazos o las piernas, ni podía mover las extremidades sobre la mesa de exploración. La miotonía (dificultad para relajar músculos) de los músculos faciales y extraoculares fue intensa. Hubo alteración leve de la respiración. Los reflejos no mostraron cambios, y la sensibilidad fue normal. La mejoría empezó media hora más tarde, y fue completa a las 3.5 horas del inicio. Los valores de potasio sérico antes, durante y después fueron: 4.5, 7.3 y 3.9 mEq/L (lo normal es de 3.5 a 4.5 mEq/L). Esta enfermedad también afectó a su hijo, hermana, madre, tía materna y abuelo materno. La herencia se debió a un gen autosómico, dominante, único, quizá con penetrancia completa. La paciente padecía parálisis periódica hiperpotasémica (hiperKPP) familiar, que ocurre en alrededor de una de cada 200 000 personas. Se origina por mutaciones en el canal Nav de músculo esquelético que hace que se desactiven con lentitud; la miotonía se produce por reaperturas anormales de los canales Nav. El aumento moderado del potasio extracelular favorece el mecanismo de compuerta aberrante con reaperturas persistentes y prolongadas. La corriente de Na a través de estos canales RESUMEN DEL CAPÍTULO ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ La despolarización abre canales Nav, lo que permite que entre Na+ con rapidez y produce despolarización adicional. Esta asa de retroacción positiva es responsable de la respuesta de todo o nada de potenciales de acción y la propagación de los mismos. El K+ que sale de la célula repolariza el potencial de membrana y termina los potenciales de acción. La pinza de voltaje, o control del potencial de membrana mediante retroacción negativa, facilita el entendimiento de las corrientes que subyacen el potencial de acción. La amplitud de la corriente de sodio y la dirección de la misma varían con la amplitud de los pasos de pinza de voltaje en el potencial de membrana. Los pasos despolarizantes primero activan y después desactivan la corriente de Na+. También activan la corriente de K+ luego de un retraso. La corriente de compuerta es un signo directo de los cambios conformacionales en las proteínas del canal de sodio. Hay un umbral para el inicio del potencial de acción. Después de un potencial de acción, las células excitables tienen un periodo refractario absoluto durante el cual no producirán un segundo potencial de acción, y después un periodo refractario relativo durante el cual se requiere un estímulo de mayor magnitud para producir un segundo potencial de acción. La mielinización aumenta la velocidad de conducción al incrementar la constante de longitud. La hipocalcemia (calcio extracelular bajo) hace a las células excitables más excitables. Las enfermedades desmielinizantes lentifican la velocidad de conducción y pueden bloquear la propagación de potenciales de acción. Los potenciales de acción aparecen de manera diferente cuando se registran con un par de electrodos colocados en el exterior de un fascículo nervioso. Los potenciales de acción compuestos, la suma de muchos potenciales de acción registrados externamente, tienen propiedades que difieren de las de potenciales de acción únicos registrados con electrodos intracelulares. En el corazón, los potenciales de acción surgen de manera automática en el nodo SA y después se diseminan de una célula a otra por el corazón mediante uniones intercelulares comunicantes (conexiones comunicantes). 58 ■ ■ ■ ■ SECCIÓN II Fisiología celular Las células del músculo cardiaco tienen canales Kir para mantener el potencial de reposo, canales Nav para la deflexión positiva del potencial de acción, canales Cav para la fase de meseta, y canales KV lentos para la repolarización. Las células del nodo SA usan canales Cav para la deflexión positiva del potencial de acción, canales KV para la repolarización, y un canal de If activado por hiperpolarización para producir la despolarización “marcapaso” lenta entre potenciales de acción. La ACh y la NE lentifican o aceleran la frecuencia cardiaca, por medio de receptores acoplados a proteína G, lo que lleva a un decremento de la If o a un aumento de la misma. La NE incrementa la amplitud de la meseta y disminuye la duración de potenciales de acción de músculo ventricular. PREGUNTAS DE ESTUDIO 1. La hiperpotasemia (concentración extracelular alta de potasio) puede detener el corazón porque: A) los iones potasio se unen a canales de sodio, lo que evita su actividad. B) los iones potasio estimulan la bomba de sodio-potasio y, así, evitan potenciales de acción cardiacos. C) el potencial de membrana de células cardiacas se despolariza y sus canales de sodio se desactivan. D) iones potasio salen con rapidez a través del rectificador interno. E) iones potasio bloquean la interacción actina-miosina en el corazón. 2. La mielinización de axones: A) reduce la velocidad de conducción para proporcionar transmisión más fiable. B) fuerza el impulso nervioso a saltar de un nodo a otro. C) ocurre en exceso en la esclerosis múltiple (MS). D) lleva a un incremento de la capacitancia efectiva de membrana. E) disminuye la constante de longitud para la diseminación pasiva del potencial de membrana. 3. Considere los tres canales que siguen en las células musculares ventriculares: de sodio (Nav), de potasio rectificador interno (Kir) y de calcio (Cav). Elija la respuesta que mejor describa cuál de estos canales está abierto durante la fase de meseta del potencial de acción ventricular. A) los tres. B) sólo Nav y Kir. C) sólo Cav y Kir. D) sólo Kir. E) sólo Cav. 4. Hay una corriente hacia adentro (If ) asociada con la actividad de marcapasos en células del nodo sinoauricular. La estimulación de nervios simpáticos que van al corazón, o la aplicación de norepinefrina, produce: A) disminución de la If, disminución de la frecuencia cardiaca, y aumento de la fuerza de la contracción. B) disminución de la If, aumento de la frecuencia cardiaca, y aumento de la fuerza de contracción. C) aumento de la If, aumento de la frecuencia cardiaca, y aumento de la fuerza de contracción. D) aumento de la If, disminución de la frecuencia cardiaca, y disminución de la fuerza de contracción. E) incremento de la If, aumento de la frecuencia cardiaca, y disminución de la fuerza de contracción. 5. La propagación de un impulso nervioso no requiere: A) cierre de canales de potasio que mantienen el potencial de reposo. B) un cambio conformacional en proteínas de membrana. C) una despolarización de membrana que abre canales de Na+. D) corriente para entrar al axón y fluir dentro del mismo. E) entrada de iones sodio al axón. 6. El potencial de acción compuesto registrado con un par de electrodos extracelulares a partir de un fascículo de fibras nerviosas intacto A) se propaga sin cambio de tamaño o forma. B) es de todo o nada. Si se excede un umbral, el incremento adicional del estímulo no aumenta la respuesta. C) tiene una amplitud de alrededor de 100 mV. D) es bifásico, y muestra deflexiones tanto positiva como negativa desde la basal. E) no es bloqueado por tetrodoxina (TTX). Sinapsis C A P Í T U L O 7 David Landowne O B J E T I V O S ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ Comprender los pasos en la transmisión sináptica química. Describir la biosíntesis y las acciones de la acetilcolina, las catecolaminas (dopamina, norepinefrina, epinefrina), la serotonina, histamina, y aminoácidos excitadores e inhibidores. Detallar la biosíntesis de neuropéptidos y las acciones de los mismos. Definir la estructura de la unión neuromuscular y las funciones de las diversas subestructuras. Describir los pasos involucrados en la transmisión neuromuscular y explicarlos. Explicar las acciones del Ca2+ y del Mg2+ sobre la liberación de transmisor, y explicar los mecanismos para los efectos de los mismos. Entender cómo la acetilcolina interactúa con receptores sobre la membrana postsináptica, y el destino de la acetilcolina. Describir la generación del potencial de placa terminal y los efectos y mecanismos de acción de inhibidores de la acetilcolinesterasa y bloqueadores de los receptores de acetilcolina. Comprender la facilitación y la potenciación postetánica de la liberación de transmisor, y la manera en que estos procesos pueden usarse para explicar ciertas características de la miastenia grave y su recuperación luego de bloqueo de receptor. Explicar las estructuras y las funciones de las diversas partes de las neuronas. Describir el transporte de materiales en dirección ascendente y descendente por axones (transporte axonal ortógrado y retrógrado), incluso mecanismos y materiales. Calcular el tiempo requerido para la regeneración de nervios periféricos. Describir las diferencias y similitudes entre la transmisión sináptica en una sinapsis central y en uniones neuromusculares. Conocer la generación de IPSP y EPSP por receptores ionotrópicos y metabotrópicos. Comprender la integración de información y la activación repetitiva en neuronas, y el concepto de la inhibición presináptica. rona presináptica libera una sustancia transmisora que se difunde a través de la hendidura sináptica y se une a un receptor en la célula postsináptica. El receptor postsináptico puede ser ionotrópico, en cuyo caso abrirá un poro selectivo y permitirá el flujo de iones para producir un potencial postsináptico (PSP), o puede ser metabo- INTRODUCCIÓN Una sinapsis es una región especializada donde una neurona se comunica con una célula blanco: otra neurona, una célula muscular, o una célula glandular. Casi todas las sinapsis son químicas; la neu- 59 60 SECCIÓN II Fisiología celular trópico e informar a una proteína G para que inicie una cascada química, que puede incluir la abertura de canales o el cierre de los mismos. Algunas sinapsis son eléctricas; pasa corriente a través de canales célula-célula de manera directa hacia la célula postsináptica. Las sinapsis químicas ofrecen la posibilidad de amplificación, inversión de señal y efectos persistentes; las sinapsis eléctricas son más rápidas y parecen ser usadas cuando la sincronización es más importante que el cálculo (procesamiento de información). Las sinapsis químicas pueden ser excitadoras o inhibidoras, dependiendo de su efecto sobre la célula postsináptica. En el sistema nervioso central (SNC) las neuronas reciben ambos tipos de sinapsis, e integran la información que llega a ellas antes de enviar el mensaje procesado a otra célula. Las sinapsis químicas son blancos farmacéuticos importantes. PROCESOS PRESINÁPTICOS La terminal presináptica debe hacer posible la síntesis, el empaque y la liberación de los diversos transmisores (figura 7-1). Los transmisores no peptídicos se concentran dentro de vesículas por cotransportadores de H/transmisor específicos. Una bomba de H+ tipo V, en que consume ATP, produce el gradiente de H+. La concentración de transmisor dentro de la vesícula puede ser bastante alta, del orden de 20 000 moléculas en una esfera de 20 nm de radio, o alrededor de 30 mM. Después de liberación, los transmisores se degradan o se transportan de regreso hacia la terminal presináptica para que vuelvan a usarse. Las membranas vesiculares también se reciclan. Algunos transmisores son polipéptidos pequeños que se sintetizan en el retículo endoplasmático rugoso, cerca del núcleo, empacados por el aparato de Golgi, y después transportados en vesículas a lo largo del axón mediante un proceso activo que se llama transporte axoplásmico; este proceso también lleva otras proteínas a las terminales presinápticas. Los neurotransmisores pueden clasificarse desde el punto de vista químico en cinco grupos (figura 7-2). Todos son hidrofílicos y contienen grupos que están cargados a pH fisiológico. Así, no pasan con facilidad por membranas lipídicas, y pueden ser compartimentados según sea necesario. ACETILCOLINA La acetilcolina (ACh) fue el primer transmisor que se reconoció. La usan motoneuronas espinales para excitar músculos esqueléticos; por los nervios parasimpáticos para comunicarse con diversos órganos terminales, incluso el nervio vago, lo que lentifica regiones marcapaso del corazón; en ganglios simpáticos y parasimpáticos, y en diversos sitios en el SNC. Hay dos clases de receptores de ACh (AChR) postsinápticos, que se nombran con base en otros agonistas que también pueden unirse a ellos. Los AChR nicotínicos se encuentran en uniones neuromusculares, ganglios simpáticos y parasimpáticos, y en el SNC. Los AChR nicotínicos son receptores ionotrópicos o pentámeros heteroméricos (véase figura 3-5). Son canales quimiosensitivos que son abiertos por la nicotina y bloqueados por el curare. Los AChR muscarínicos se encuentran en el corazón, músculos lisos, células glandulares y el SNC. Son GPCR 7-TM metabotrópicos que son activados por muscarina y bloqueados por la atropina. Los nAChR tienden a excitar la célula postsináptica; los mAChR pueden ser excitadores o inhibidores. La ACh se sintetiza a partir de acetil-CoA y colina con la enzima colina acetiltransferasa (CAT), que se encuentra en el citoplasma presináptico. La ACh se concentra en vesículas por un cotransporta- Llena Acoplamiento Fusión y liberación Reciclado FIGURA 71 Acoplamiento de vesícula sináptica, liberación del contenido, y reciclado. New York: Lange Medical Books/McGraw-Hill, 2006.) (Modificada con autorización de Landowne D: Cell Physiology. CAPÍTULO 7 Sinapsis Colinérgico Aminas biogénicas O + O N(CH3)3 NH2 HOOC HO Dopamina HO Peptidérgico Aminoácidos COOH HO Acetilcolina 61 NH2 Ácido glutámico NH2 HOOC HO NH2 OH Endorfina Encefalina Dinorfina Péptido relacionado con el gen de la calcitonina (CGRP) Sustancia Y Sustancia P Vasopresina (ADH) Oxitocina Colecistocinina (CCK) Péptido intestinal vasoactivo (VIP) Ácido γ-aminobutírico (GABA) Norepinefrina HO N(CH3) HO HOOC NH2 OH Epinefrina Glicina NH2 HO Purinérgicos ATP NH Adenosina Serotonina NH2 HN FIGURA 72 Los neurotransmisores. (Modificada con autorización de Landowne D: Cell N Physiology. New York: Lange Medical Books/McGraw-Hill, Histamina dor de H/ACh (figura 7-3). Un proceso activado por Ca2+ libera las vesículas. Se describe más adelante, después de que se comenten todos los transmisores. El envenenamiento por toxina botulínica (Botox) bloquea la liberación de ACh y da lugar a falla de la transmisión neuromuscular. Recientemente se han usado inyecciones de Botox para tratar distonía, un trastorno del movimiento que se caracteriza por contracciones musculares involuntarias, y con fines estéticos para bloquear localmente músculos faciales que arrugan la piel. Las dosis excesivas o el suministro de toxina sistémica por contaminación durante el enlatado de alimentos pueden llevar a la muerte. El veneno de araña viuda negra (o parda) (BWSV) también bloquea la transmisión neuromuscular. Hace a las membranas presinápticas permeables al Ca2+, y causa una liberación masiva de vesículas, seguida por falla de la transmisión debido a la falta de ACh almacenada. Después de la liberación, la ACh puede fragmentarse hacia acetato y colina por la acetilcolinesterasa (AChE) en el espacio extracelular. Un cotransportador de Na/colina recapta casi toda la colina; a continuación, la ACh es resintetizada por la CAT y reempacada. Los inhibidores de la ACh o anticolinesterasas se usan para propósitos médicos, como insecticidas, y como gases nerviosos en la guerra química. El efecto es aumentar la cantidad y duración de la interac- 2006.) ción de la ACh con los receptores postsinápticos. Las contramedidas en el campo de batalla son bloquear los receptores postsinápticos en el corazón con atropina. Los gases nerviosos son organofluorofosfatos que se unen de manera irreversible a la AChE, pero pueden ser desplazados por piridina aldoxima metil yoduro (PAM). AMINOÁCIDOS GLUTAMATO El glutamato es el principal neurotransmisor excitador del SNC. Es un aminoácido no esencial pero, dado que no puede cruzar la barrera hematoencefálica, el SNC debe sintetizarlo. Hay varias vías sintéticas, pero ninguna específica para neuronas. Los receptores de glutamato ionotrópicos, gluR, se clasifican como de tipo NMDA si el agonista sintético N-metil-d-aspartato los activa, o como tipo no NMDA si no lo hace. Ambos tipos son tetrámeros heteroméricos (véase figura 3-6) y permiten el paso de Na+ y K+, pero los gluR NMDA también permiten la entrada de Ca2+ a la célula, y tienen un papel especial en la plasticidad sináptica (véase más adelante). También hay gluR metabotrópicos (mgluR). En circunstancias normales, todos éstos son activados por glutamato (figura 7-4). 62 SECCIÓN II Fisiología celular Glía Colinesterasa H+ GPCR muscarínico ATP ACh Canal nicotínico Presináptica ACh ATP Colina Na Colinesterasa Postsináptica K Na FIGURA 73 Una sinapsis de acetilcolina esquemática generalizada. La terminal presináptica tiene transportadores para la recaptación de colina y empaque de ACh. La membrana postsináptica tiene receptores de ACh y colinesterasa. La membrana de célula glial cercana también tiene colinesterasa. La ACh es hidrolizada por las esterasas, y la parte de colina es llevada de regreso por la terminal presináptica. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) El glutamato se elimina del espacio extracelular por el cotransportador de Na/glu, el transportador de aminoácido excitador (EAAT), que también contratransporta K+. Los EAAT se encuentran en la membrana terminal presináptica, y en la membrana postsináp- tica y membranas de células gliales cercanas. Dentro de la glía, el glutamato puede convertirse en glutamina, que es liberada y captada por la terminal presináptica mediante un cotransportador acoplado a Na/Cl, y, por último, reconvertido en glutamato. Na glu K gln Na gln Glía mgluR GPCR H+ ATP Canal ionotrópico Presináptica glu ATP K glu Na Postsináptica K Na FIGURA 74 Una sinapsis de glutamato esquemática generalizada. Además de transportadores de recaptación y empaque, las sinapsis de glutamato tienen transportadores de captación en las membranas de células postsinápticas y gliales. También hay una vía de glutamina (gln) para mover glutamato en la célula glial de regreso a la terminal presináptica. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/ McGraw-Hill, 2006.) CAPÍTULO 7 Sinapsis 63 Na GABA Cl Glía GABAB GPCR H+ ATP Canal GABAA Presináptica GABA ATP Cl GABA Na Postsináptica K Na FIGURA 75 Una sinapsis de GABA esquemática generalizada. Ésta es similar al esquema de glutamato, pero tiene receptores y transportadores de GABA. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) El glutamato extracelular excesivo mata neuronas al permitir que entre un exceso de Ca2+ a las células, lo que puede llevar a necrosis (muerte celular). Se postula que esta neurotoxicidad desempeña un papel en la apoplejía isquémica, la esclerosis lateral amiotrófica (ALS), enfermedad de Huntington, enfermedad de Alzheimer, y quizá, en algunas formas de epilepsia. La isquemia puede aumentar el glutamato extracelular al limitar el metabolismo oxidativo, el ATP y los gradientes de sodio y, así el movimiento de glutamato en dirección contraria a los receptores. GABA Y GLICINA El ácido γ-aminobutírico (GABA) y la glicina son los principales neurotransmisores inhibidores en el SNC. La glutamato descarboxilasa (GAD) convierte el glutamato en GABA en el citoplasma de la terminal presináptica. El GABA es empacado y liberado como otros transmisores (figura 7-5). Hay un cotransportador de Na/ GABA que elimina el GABA de la hendidura sináptica. Los receptores GABAA y los receptores de glicina son heterómeros pentaméricos en la superfamilia de nAChR; son permeables a iones Cl–. Los receptores GABAB son GPCR que activan canales Kir (o GIRK). El SNC opera con una magnitud tónica de inhibición que se puede cambiar con diversos fármacos. El muscimol, del hongo Amanita muscaria, es un potente agonista de GABAAR. Los tranquilizantes comunes, como el diazepam, y los barbitúricos, como el fenobarbital, aumentan la abertura de GABAAR. La picrotoxina, un potente convulsivo, bloquea GABAAR. La estricnina, también un convulsivo, bloquea gliR. La toxina tetánica produce una parálisis espástica al bloquear el mecanismo de liberación para GABA y glicina. AMINAS BIOGÉNICAS Las catecolaminas, serotonina e histamina son aminas biogénicas. Las catecolaminas son la dopamina, norepinefrina (NE) y epinefrina (EPI). Casi todos los efectos que producen estas aminas biogénicas se efectúan por medio de GPCR, a menudo sin producir PSP. Todas son concentradas en vesículas y liberadas por medio de mecanismos similares, pero algunas son liberadas por abultamientos nerviosos, que están en la vecindad de los receptores, pero no en aposición tan estrecha (figura 7-19). Células no nerviosas también liberan EPI e histamina. CATECOLAMINAS La dopamina y la NE se encuentran en el SNC. La NE es el principal transmisor final del sistema nervioso simpático, y a la EPI la sintetiza y la libera la médula suprarrenal. Las tres son sintetizadas mediante la misma vía, empezando con la hidroxilación de tirosina hacia dihidroxifenilalanina (DOPA), que a continuación es descarboxilada para formar dopamina. La adición de un grupo β-hidroxilo forma NE y, en las células de la médula suprarrenal, la transferencia subsiguiente de un grupo N-metilo forma EPI. La tirosina hidroxilasa (TH) es la enzima limitante. La TH y la dopa descarboxilasa están en el citoplasma de la terminal presináptica. La dopamina es concentrada hacia vesículas, donde la dopamina β-hidroxilasa (DBH) la convierte en NE. La NE es captada de regreso hacia la terminal presináptica por medio de un cotransportador acoplado a Na/Cl; ahí, la desintegra la monoaminooxidasa (MAO) en las mitocondrias, y la catecolamina-O-metiltransferasa (COMT) en el citoplasma. 64 SECCIÓN II Fisiología celular Los receptores de catecolamina son GPCR, y se encuentran en el SNC, el músculo liso, y el corazón. Los receptores adrenérgicos muestran respuesta a NE, o a EPI, o ambas. Hay dos categorías de receptores adrenérgicos: los receptores α-adrenérgicos tienen una afinidad más alta por NE, y los receptores β-adrenérgicos tienen una afinidad más alta por EPI. Sin embargo, hay reactividad cruzada, y ambos receptores mostrarán respuesta a concentraciones más altas de ambos agonistas. En el sistema cardiovascular, los receptores α se encuentran en las células de músculo liso que controlan el diámetro de vasos sanguíneos de pequeño calibre; la NE actúa para constreñir estos vasos. Los receptores β están principalmente en el corazón, y pueden hacer que lata más rápido y más fuerte. La relajación muscular por medio de la activación de receptor adrenérgico ocurre en células de músculo liso en el intestino y los pulmones. Algunas de estas funciones se comentan con mayor profundidad en la sección siguiente. La enfermedad de Parkinson se caracteriza por la pérdida de neuronas dopaminérgicas; su tratamiento, a menudo, incluye DOPA, que puede aliviar parcialmente los síntomas. Los fármacos que bloquean receptores de dopamina se han usado para tratar esquizofrenia; a veces inducen temblores que se parecen a la enfermedad de Parkinson. La reserpina, un tranquilizante temprano, inhibe el transporte de dopamina hacia vesículas. La cocaína bloquea la recaptación de catecolaminas, lo cual prolonga las acciones de estas últimas. Muchos de los remedios de venta sin receta para congestión nasal, como la neosinefrina, activan receptores de catecolamina. SEROTONINA La serotonina, o 5-hidroxitriptamina (5-HT), se sintetiza a partir del triptófano mediante hidroxilación y descarboxilación. Los receptores de 5-HT funcionan en el intestino en la secreción y el peristaltismo, median la agregación plaquetaria y la contracción de músculo liso, y están distribuidos en todo el sistema límbico del cerebro. En un inicio la serotonina fue identificada como una sustancia en el suero sanguíneo que constreñía vasos sanguíneos, de ahí su nombre. La triptófano hidroxilasa es el paso limitante de la síntesis de 5-HT; en el SNC la triptófano hidroxilasa sólo está presente en neuronas serotoninérgicas. La 5-HT es desactivada por medio de recaptación, y después desintegrada por la MAO en las mitocondrias. Casi todos los receptores de 5-HT son GPCR; los receptores 5-HT3 son canales iónicos. Los inhibidores selectivos de la recaptación de serotonina, como el clorhidrato de fluoxetina se prescriben, por lo regular, como antidepresivos. La dietilamida del ácido lisérgico (LSD) y la psilocina, el metabolito activo de la psilocibina, son alucinógenos que activan receptores de 5-HT. HISTAMINA Casi toda la histamina en el organismo se libera a partir de mastocitos (células cebadas) (parte del sistema inmunitario) en respuesta a antígenos o lesión tisular. La histamina también es un regulador de la secreción de ácido en el intestino, y actúa como un neurotransmisor en el SNC. La liberación de histamina se asocia con reacciones alérgicas; inicia respuestas inflamatorias, dilata vasos sanguíneos y aumenta la permeabilidad capilar, disminuye la frecuencia cardiaca y contrae músculos lisos en los pulmones. Las células parecidas a enterocromafines en la mucosa gástrica también liberan histamina, lo que promueve la producción de ácido. La histamina se sintetiza a partir de histidina, se almacena en vesículas, y se libera; a continuación la desintegra la histamina N-metil transferasa. Hay cuatro receptores de histamina diferentes, todos los cuales son GPCR. PURINAS El ATP está contenido en vesículas sinápticas y se libera con NE en neuronas vasoconstrictoras simpáticas. Induce constricción cuando se aplica de manera directa a los músculos lisos. Los receptores de ATP P2X son canales iónicos que permiten la entrada de Ca2+, y las células también tienen GPCR P2Y; estos receptores también se encuentran en el cerebro, así como receptores P1 para adenosina. PÉPTIDOS Los neuropéptidos son polipéptidos pequeños que se sintetizan como precursores inactivos de mayor tamaño (propéptidos) y después los dividen endopeptidasas específicas. Puesto que son proteínas, son sintetizados en el cuerpo celular y transportados en vesículas hacia las terminales. No hay mecanismo de recaptación. Los péptidos están menos concentrados que otros neurotransmisores en vesículas, pero tienen afinidad más alta por sus receptores, que son GPCR. Los neuropéptidos son liberados a partir de vesículas grandes de centro denso, mientras que otros neurotransmisores son secretados a partir de vesículas de menor tamaño, más claras. A menudo los neuropéptidos actúan de manera conjunta con neurotransmisores clásicos. No se sabe mucho acerca de la función de casi todos los neuropéptidos en el SNC, excepto los péptidos opiáceos, endorfina, encefalina y dinorfina, que están involucrados en la regulación de la percepción del dolor. Se han identificado tres receptores de opiáceo, al inicio como los sitios que se unen a opiáceos sintéticos, como la morfina. Hay muchos péptidos opioides liberados a partir de neuronas. El péptido que se relaciona con el gen de la calcitonina (CGRP) y la sustancia Y están involucrados en el mantenimiento de la presión arterial. La hormona antidiurética (ADH, que también se conoce como vasopresina) ayuda a controlar la recaptación de agua en los riñones. La oxitocina, la hormona luteinizante (LH) y la hormona estimulante del folículo (FSH) están involucradas en la reproducción. La colecistocinina (CCK), la gastrina y el péptido intestinal vasoactivo (VIP) facilitan la digestión. Todos éstos y muchos más se han identificado como neurotransmisores potenciales en el SNC. LIBERACIÓN SINÁPTICA Los detalles del proceso de liberación sináptica se están investigando de manera activa. Está claro que el proceso se desencadena por un incremento de la concentración citoplasmática de Ca2+. En muchas sinapsis, cuando llega un potencial de acción presináptico, el Ca2+ entra a la terminal a través de canales Cav. En algunas células sensoriales pequeñas no hay potencial de acción, y el potencial generador sensorial abre los canales Cav. Las vesículas sinápticas pasan por un ciclo que consta de almacenamiento de transmisores, acoplamiento en una zona activa o sitio de libe- CAPÍTULO 7 Sinapsis 65 Na Acoplamiento Ca Fusión y liberación Reciclado FIGURA 76 Los canales involucrados en la liberación sináptica. Los canales Nav despolarizan la terminación, y los canales Cav permiten que el flujo de entrada de Ca2+ desencadene la liberación. También hay canales KV que repolarizan la membrana y, así, limitan el flujo de entrada de Ca2+. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) ración, fusión con la membrana de superficie y liberación del contenido, recuperación endocitótica y después al almacenamiento de nuevo. En la figura 7-6, cada paso en el ciclo vesicular se ilustra con un cambio de la posición de la vesícula. Sin embargo, en realidad hay poco movimiento en los estados adjuntos. En otras sinapsis, el sitio de liberación es transversal desde un área postsináptica, la cual contiene los canales que son sensibles al transmisor. En la unión neuromuscular (figura 7-9), los canales Cav son adyacentes al sitio de liberación, de modo que el Ca2+ interno sólo necesita estar alto de manera local para causar liberación. El acoplamiento y la fusión involucran la SNARE o proteína de fijación a factor sensible a N-etilmaleimida (SNAP) —proteína receptora que está presente en ambas membranas antes de la fusión y se asocia con complejos centrales apretados durante la fusión—. En la figura 7-7 se muestra la sinaptobrevina v-SNARE vesicular uniéndose al blanco sintaxina t-SNARE y SNAP-25. La sinaptobrevina es el sustrato de las endopeptidasas contenidas en las toxinas botulínica y tetánica. Sinaptotagmina Sinaptobrevina v-SNARE Ca Sintaxina t-SNARE FIGURA 77 Un posible mecanismo de fusión de vesícula. Las proteínas SNARE acoplan la vesícula, y el Ca2+ se une a la sinaptotagmina para causar fusión. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) 66 SECCIÓN II Fisiología celular La fusión estimulada por Ca requiere, Ca2+, la proteína de unión a sinaptotagmina, que está en la membrana vesicular y se une al Ca2+. Un modelo propuesto sugiere que el Ca2+ permite que la sinaptotagmina se una a la membrana superficial y tire de las dos capas lipídicas una hacia otra. El proceso de reciclado devuelve los lípidos y las proteínas al fondo común de vesícula. La vesícula vuelve a formarse como un hoyuelo cubierto con clatrina. Las moléculas de clatrina tienen la forma de un trisquelión, o tres piernas flexionadas. La clatrina forma una superficie cerrada cubierta con pentágonos, y separa de la superficie “por pellizco” la vesícula recuperada. TRANSPORTE AXOPLÁSMICO Todas las proteínas en la terminal presináptica son sintetizadas en el cuerpo celular, y transportadas quizá 1 m antes de que sean útiles. Además, la neurona tiene mecanismos que transportan algunos materiales en la dirección reversa o retrógrada de regreso hacia el cuerpo celular. Algunos de los mecanismos que se usan para este transporte se utilizan en otras células para llevar material a la periferia de la célula, y para el movimiento de cromosomas durante la mitosis. El transporte axoplásmico se distingue por la dirección en ortógrado y retrógrado. El transporte ortógrado puede subdividirse en rápido (100 a 400 mm/día, o 1 a 5 μm/s) y lento (0.5 a 4 mm/día). El transporte rápido es para vesículas y mitocondrias; el transporte lento es para enzimas solubles y las que constituyen el citoesqueleto. El transporte retrógrado sólo es del tipo rápido. El transporte axoplásmico rápido comprende motores moleculares que hidrolizan ATP y caminan a lo largo de microtúbulos, cilindros huecos largos de 25 nm de diámetro. Se emplean dos clases de motores: cinesinas para el transporte ortógrado, y dineínas para el transporte retrógrado. Los microtúbulos están polarizados, y estos motores pueden detectar la polaridad y moverse en pasos de 8 nm en la dirección apropiada. Los motores tienen dos “pies”, o sitios de interacción con los microtúbulos, y muestran procesividad, o la capacidad para funcionar repetidas veces sin disociarse de su sustrato, el microtúbulo. Las moléculas accesorias se usan para fijar la carga al motor (figura 7-8). Ortógrado Cinesina Microtúbulo Dineína Retrógrado FIGURA 78 Transporte axoplásmico. Los motores de cinesina transportan vesículas hacia las terminales nerviosas. Los motores de dineína transportan diferentes vesículas hacia el cuerpo celular. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) El transporte ortógrado rápido lleva proteínas necesarias a la membrana en la terminal, tanto para las vesículas como para la membrana terminal. Durante el desarrollo, también puede llevar moléculas de adhesión celular que reconocen blancos o los inducen. El transporte retrógrado puede devolver proteínas dañadas para la vía endolítica, y llevar información acerca de eventos de emisión de señales de regreso hacia el cuerpo celular. El transporte retrógrado forma parte de la fisiopatología de varias enfermedades, entre ellas poliomielitis, rabia, tétanos y herpes simple. El virus del herpes entra en terminales nerviosas periféricas y después viaja de regreso hacia el cuerpo celular, donde se replica o entra en latencia. Más tarde puede regresar a la terminación nerviosa mediante transporte ortógrado, y ponerse a sí mismo a disposición para transmisión por contacto a otra persona. La toxina tetánica es transportada de manera retrógrada en motoneuronas hacia las dendritas, y después por vía transináptica a terminales liberadoras de GABA y glicina, donde inhibe la liberación sináptica. El transporte axoplásmico es importante para la regeneración de nervios después de lesión en el sistema nervioso periférico. Por lo general, los nervios en el SNC no se regeneran, aunque investigadores actuales tienen esperanza de que esto cambie en el futuro. Si un axón de nervio periférico se corta o aplasta, la porción distal morirá y pasará por una degeneración walleriana característica conforme el axón es resorbido al cabo de algunas semanas. En el transcurso de algunos días, el cuerpo celular pasa por la reacción de axón, que con frecuencia se llama cromatolisis, debido a un cambio de la tinción en el estudio histológico. El nucléolo se agranda, el retículo endoplasmático rugoso, o ER (sustancia de Nissl), se dispersa, y el núcleo se desplaza; se han activado genes, transcrito RNA y sintetizado proteínas. Mientras mayor es la distancia desde la lesión hasta el cuerpo celular, mayor es la latencia, lo que indica que el transporte retrógrado está involucrado en la emisión de señales para iniciar la reacción axonal. En el sitio de lesión, el extremo que está acoplado al cuerpo celular se volverá a sellar en horas, y aparecerán brotes en un día o dos. La punta cortada se hincha con mitocondrias y ER liso. Los brotes crecen como fibras delgadas. Si la regeneración es exitosa, una de las nuevas fibras encuentra su camino por la vaina del nervio en degeneración distal y reinerva un blanco postsináptico. La fibra a continuación aumentará de diámetro y quedará remielinizada. La tasa de crecimiento de fibra es de alrededor de 1 mm por día, en el rango de transporte axonal lento; ese es el número que debe usarse al estimar tiempos de recuperación. Además de los sistemas basados en microtúbulos, el transporte intracelular también puede ocurrir por medio de motores de miosina que viajan a lo largo de filamentos de actina. La interacción es similar a la descrita en la sección de los capítulos 8 y 9, excepto porque la actina permanece fija y las moléculas de miosina individuales avanzan a lo largo de ella. Hay moléculas adaptadoras que fijan la carga a la miosina. PROCESOS POSTSINÁPTICOS Hay varios receptores postsinápticos diferentes para cada transmisor; se distinguen por sus secuencias de aminoácidos y, en algunos casos, características farmacológicas. Distintas regiones del sistema nervioso tienen receptores característicos; a veces una célula postsi- CAPÍTULO 7 Sinapsis náptica individual tendrá múltiples tipos de receptor. Los receptores ionotrópicos son excitadores o inhibidores de acuerdo con su selectividad iónica. Los receptores metabotrópicos pueden hacer, de manera indirecta, que los canales se abran o se cierren, y pueden también modular la actividad de las células de otras maneras. Los PSP se llaman potenciales postsinápticos excitadores (EPSP), si su efecto es hacer que la célula postsináptica tenga más probabilidades de mostrar respuesta con un potencial de acción, o potenciales postsinápticos inhibidores (IPSP), si hacen que la célula postsináptica tenga menos probabilidades de activar un potencial de acción. Cada canal tiene un patrón de selectividad, y permite que iones diferentes fluyan por él con facilidad que difiere. Esto significa que cada canal tendrá un potencial reverso: un potencial al cual no habrá flujo neto de iones a través del canal. Si el potencial de membrana es más positivo que el potencial reverso, fluirá corriente neta hacia afuera de la célula, y tenderá a hiperpolarizarla. Si la membrana es menos positiva o más negativa, la corriente fluirá hacia adentro y tenderá a despolarizar la célula. La corriente que fluye por canales impulsa el potencial de membrana hacia el potencial reverso para ese canal. Casi todas las neuronas en el SNC reciben aferencias que fluctúan constantemente, provenientes de diversas sinapsis, y su potencial de membrana siempre está cambiando. Si una sinapsis abre canales que tienen un potencial reverso más positivo que el umbral para potenciales de acción, producirá un EPSP. Si el potencial reverso es más negativo que el umbral, el resultado será un IPSP. Si un canal es permeable a un ion único, su potencial reverso es para ese ion el potencial de Nernst (ecuación [4] en el capítulo 4). Si el canal es permeable a múltiples iones, su potencial reverso es el promedio ponderado de los potenciales de Nernst para sus iones (ecuación [6] en el capítulo 4). Los canales nAChR y GluR son casi igual de permeables al Na+ y + K , y su potencial reverso es de alrededor de –10 mV; cuando son activados, hacen EPSP. Los canales GABAAR y gliR son canales de Cl; su potencial reverso es de alrededor de –80 mV. El mAChR cardiaco, por medio de una proteína G, activa un canal Kir (KACh) que tiene un potencial reverso de alrededor de –90 mV. Tanto los canales de Cl– como los de K+ hacen IPSP. Si por alguna razón sucede que la célula sea más negativa que –80 mV, la abertura de canales de Cl despolarizará la célula, pero aún funcionará para evitar que otros canales despolaricen más la célula hasta el umbral. LA UNIÓN NEUROMUSCULAR, UNA SINAPSIS ESPECIALIZADA Debido a su accesibilidad fácil, la unión neuromuscular (o mioneural) (figura 7-9) es la sinapsis mejor estudiada; es la fuente de gran parte de lo que se sabe acerca de sinapsis. En esta sección se describe el funcionamiento de esta sinapsis y se juntan e ilustran muchas de Mielina Axón Célula de Schwann Mitocondrias Vesículas sinápticas Hendidura sináptica 67 Membrana presináptica Membrana postsináptica Núcleo del músculo Pliegue sináptico Miofibrillas FIGURA 79 La unión neuromuscular. Un nervio mielinizado termina en una región especializada de un músculo esquelético (coloreado). (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) 68 SECCIÓN II Fisiología celular las ideas antes introducidas de manera más abstracta. La unión neuromuscular despierta considerable interés clínico. La miastenia grave es una enfermedad que incapacita la unión neuromuscular; hay otras patologías y varios fármacos y toxinas que establecen como blanco la unión. La unión neuromuscular proporciona una valoración conveniente para el anestesiólogo que evalúa la recuperación luego de inmovilización muscular para intervención quirúrgica. Una motoneurona única controla 3 a 1 000 células musculares. Cada célula muscular recibe aferencias que provienen de una motoneurona. Las motoneuronas y todas sus células musculares funcionan juntas como una unidad motora. En personas sanas, un potencial de acción en la motoneurona producirá un EPSP grande en todas sus células musculares, de magnitud suficiente para exceder mucho el umbral de las células musculares y producir potenciales de acción, y contracción. El SNC regula el movimiento al elegir cuáles unidades motoras activar. Las unidades motoras de menor tamaño producen movimientos más finos. En la terminación nerviosa, el axón pierde su mielina y se disemina hacia afuera para formar la placa terminal motora, que se denomina así por su aspecto anatómico. Las terminales nerviosas contienen muchas mitocondrias y vesículas sinápticas de 40 nm de diámetro, que contienen ACh. La terminal nerviosa está separada del músculo por una brecha de 50 nm, la hendidura sináptica, que contiene una lámina basal. La membrana muscular contiene AChR y AChE. En micrografías electrónicas de transmisión, las membranas tanto presináptica como postsináptica aparecen engrosadas, lo que indica la presencia de canales y otras proteínas. La transmisión neuromuscular puede describirse como un proceso de 10 pasos: 1) un potencial de acción entra a la terminal presináptica; 2) la terminal nerviosa es despolarizada; 3) la despolarización abre canales Cav; 4) entra Ca2+ a la célula, y se mueve en favor de su gradiente electroquímico; 5) el Ca2+ actúa sobre un sitio de liberación, quizá sinaptotagmina, lo que hace que vesículas sinápticas se fusionen con la membrana presináptica; 6) alrededor de 200 vesículas liberan su ACh hacia la hendidura sináptica; 7) la ACh en la hendidura sináptica: a) se difunde hacia afuera de la hendidura, b) es hidrolizada por AChE hacia acetato y colina, o c) interactúa con AChR sobre la membrana postsináptica; 8) los AChR activados son muy permeables al Na+ y K+, y un poco permeables al Ca2+; por ende, un flujo de entrada neto de carga positiva hacia la célula muscular despolariza la membrana muscular en la región de la placa terminal; 9) cuando la membrana muscular se despolariza hasta el umbral, se desencadena un potencial de acción, que se propaga en ambas direcciones hasta los extremos de la célula muscular (el enlace entre excitación y contracción musculares se comenta en la sección que sigue), y, por último, 10) la colina se recicla hacia la terminal nerviosa, el Ca2+ es bombeado hacia afuera de la terminal nerviosa, y las vesículas son recicladas y vueltas a llenar. REGISTRO DEL POTENCIAL DE PLACA TERMINAL Si se inserta un microelectrodo en una fibra muscular cerca de la unión neuromuscular, se medirá un potencial de reposo de alrededor de –90 mV. Si se estimula el nervio y se evita que el músculo se contraiga mediante estiramiento extremo, se observará que el potencial de membrana cambia, como se muestra en el trazo continuo en FIGURA 710 Un potencial de placa terminal y potencial de acción en la unión neuromuscular (izquierda) y a 2 cm (derecha). Las líneas discontinuas indican respuestas en Ca2+ bajo (véase el texto). (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) el lado izquierdo de la figura 7-10. Si, en lugar de eso, el electrodo se coloca a varios centímetros de la unión neuromuscular, se observará el potencial que se muestra en el trazo de la derecha. Si se disminuye la concentración de Ca2+ en el baño, o se aumenta la concentración de Mg2+, y se estimula de nuevo el nervio, el potencial en la unión neuromuscular cambiará como se muestra en el trazo discontinuo. En estas condiciones no habrá cambio del potencial de membrana a varios centímetros de la unión. El trazo continuo a la izquierda muestra un potencial de acción superpuesto sobre un potencial de placa terminal (EPP). Hay una despolarización inicial debida a una entrada neta de carga positiva a través de AChR, que fue activado por la ACh liberada. Cuando el potencial alcanzó alrededor de –50 mV, se inició un potencial de acción. En presencia de Ca2+ normal el EPP es de tamaño 2 o 3 veces mayor que el necesario para despolarizar la membrana muscular hasta el umbral. El potencial de acción puro se observa en el trazo de la derecha; puede registrarse al estimular eléctricamente un extremo del músculo o al colocar el electrodo de registro a algunos centímetros de la placa terminal. El trazo discontinuo a la izquierda muestra un EPP con amplitud reducida. El EPP no es visible a algunos centímetros de la placa terminal (derecha). Una reducción del Ca2+ extracelular disminuye la liberación de ACh y, así, aminora el EPP. Un incremento del Mg2+ reduce la liberación de transmisor al disminuir la entrada de Ca2+ a través de canales Cav. Estos efectos que se oponen del Ca2+ y Mg2+ se observan en todas las sinapsis químicas que se examinaron; esto ahora se considera una de las pruebas para identificar una sinapsis química. Las concentraciones de Ca2+ y Mg2+ tienen diferentes efectos sobre la excitabilidad o el umbral para potenciales de acción sobre las células nerviosas y musculares. La reducción de Ca2+ hace a las células más excitables o que tengan un umbral más negativo, o requiere una despolarización de menor magnitud para alcanzar el umbral para un potencial de acción; éste es un efecto sobre los canales Nav; en presencia de Ca2+ bajo, los canales Nav se abrirán a potenciales más negativos. El Ca2+ y Mg2+ tienen una acción sinérgica sobre los canales Nav; tienen acciones que se oponen sobre la transmisión neuromuscular. En clínica, los efectos de la hipocalcemia son hiperexcitabilidad y potenciales de acción espontáneos en nervio y músculo. Estos efectos se observan cuando aún hay suficiente Ca2+ 1 mV CAPÍTULO 7 Sinapsis 10 ms 69 total se liberan alrededor de 200 000 vesículas, lo que es igual al número que se observa mediante el microscopio electrónico en una unión neuromuscular no estimulada. Después de tratamiento con BWSV, no hay vesículas visibles. El BWSV paraliza al agotar las vesículas sinápticas de terminales nerviosas; puede ser mortal si afecta las terminaciones nerviosas que controlan la respiración. INTERACCIÓN TRANSMISORRECEPTOR FIGURA 711 Algunos potenciales de placa terminal miniatura (MEPP) que se observan al estimular cuatro veces una unión neuromuscular bañada en Ca2+ bajo. El segundo MEPP en el trazo inferior fue espontáneo. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) para apoyar suficiente liberación de ACh, de modo que todo potencial de acción nervioso lleve a un potencial de acción muscular. En este caso de Ca2+ bajo y Mg2+ alto, el EPP no es suficientemente grande como para alcanzar umbral y desencadenar un potencial de acción. Los potenciales de acción se propagan de manera activa; el EPP se disemina de manera pasiva y no será visible a algunos centímetros de la unión neuromuscular. Estos dos potenciales son producidos por la actividad de diferentes canales que tienen propiedades farmacológicas que difieren. El curare bloqueará los AChR y el EPP sin afectar el potencial de acción que se observa después de estimulación eléctrica directa del músculo. Una toxina de un caracol cono (conotoxina μ) bloqueará el potencial de acción muscular, pero no el EPP. La conotoxina μ bloquea canales Nav musculares, pero no canales Nav de nervios, que son un producto de gen diferente. Si la proporción Ca2+/Mg2+ es baja, la respuesta a la estimulación aparecerá como en la figura 7-11. Cada trazo representa la respuesta a una estimulación que se repite cada 5 s; tres de los trazos muestran un EPP pequeño; en el tercer intento no hubo respuesta. La primera respuesta tiene alrededor de 1 mV de alto; la segunda y cuarta respuestas son de alrededor de 0.5 mV. Cuando este experimento se repitió muchas veces, se encontró que las respuestas estuvieron cuantizadas con una respuesta de unidad de alrededor de 0.5 mV. Es decir, hubo muchas respuestas de 0.5, 1 y 1.5 mV, pero muy pocas con amplitudes interpuestas. Además, a veces hay respuestas de 0.5 mV espontáneas sin estimulación alguna; una de éstas se captó en el cuarto trazo. Estos potenciales de placa terminal miniatura (MEPP) representan la respuesta postsináptica a la liberación de 1, 2 o 3 cuantos de ACh. Cada cuanto es el contenido de una vesícula única. Es imposible conocer el número exacto de vesículas liberadas en cualquier estimulación particular; sólo puede predecirse el número promedio o el contenido cuántico medio. El EPP en condiciones de Ca2+/Mg2+ normales es la respuesta a alrededor de 200 cuantos. La tasa promedio de MEPP espontáneo es de alrededor de una vesícula/s. En un EPP normal, las 200 vesículas son liberadas en el transcurso de 1 ms, lo que significa que la estimulación aumentó 200 000 veces la tasa de liberación. Si se aplica BWSV a una unión neuromuscular, la frecuencia de MEPP aumenta a algunos cientos por segundo durante alrededor de 30 min y después se suspende. En El AChR nicotínico en la unión neuromuscular tiene cinco subunidades, cada una con cuatro segmentos TM. Dos de las subunidades se llaman subunidades α y se unen a ACh en las interfases α-γ y α-δ cerca de la parte superior de la molécula, a alrededor de 5 nm desde el centro de la membrana. El canal a continuación pasa por un cambio conformacional que se transmite a través de la molécula para abrir el poro, con más probabilidad al hacer que los segmentos TM M2 se alejen del eje del poro, lo que aumenta el tamaño de este último. El poro abierto permite el paso de Na+ y K+ y, en menor grado, de Ca2+. El poro permanece abierto alrededor de 1 ms y pasan alrededor de 20 000 iones a una tasa de 2 × 107/s, lo que equivale aproximadamente de 3 pA. Si se captura un AChR único en una placa de membrana y se mantiene con un potencial de –90 mV, la aplicación de ACh hará que el canal se abra y se cierre varias veces; cada abertura aparece como un pulso de corriente de 3 pA, de duración variable, y promedia alrededor de 1 ms. Un cuanto único abre aproximadamente 2 000 canales; 200 cuantos abren alrededor de 400 000. Una unión neuromuscular tiene muchos más canales, aproximadamente 20 millones; así, sólo una pequeña fracción se usa en cualquier momento. El número de canales abiertos es proporcional a la concentración de ACh al cuadrado, y al número efectivo de receptores. En la figura 7-12 se muestra un esquema cinético para la reacción. El receptor puede abrirse con 1 o 2 moléculas de ACh unidas; permanece abierto unas 10 veces más tiempo con dos unidas. Es la concentración de R • 2ACh la que es proporcional a la concentración de ACh al cuadrado: Número de canales abiertos = k [R][ACh]2 (1) DESENSIBILIZACIÓN Si un AChR queda expuesto a ACh de manera continua durante varios minutos, su respuesta se lentificará y las aberturas se harán menos frecuentes. Si se agrega ACh al baño que contiene una unión neuromuscular, el potencial de membrana muscular se despolariza- R 3r"$I 3r"$I $FSSBEP R* r"$I R* r"$I "CJFSUP FIGURA 712 Un esquema cinético de la reacción entre la acetilcolina y el receptor de acetilcolina nicotínico. El receptor (R) se puede unir a dos moléculas de acetilcolina. Una vez que la ACh se une a los receptores se pueden abrir (R*) y permitir el paso de iones. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/ McGraw-Hill, 2006.) 70 SECCIÓN II Fisiología celular La serpiente Bungarus paraliza su presa con α-bungarotoxina, que se une de manera irreversible a AChR y evita su abertura. La bungarotoxina se ha marcado con fluorescencia y se usa experimentalmente para identificar nAChR y ubicarlos. 10 μM ACh 10 mV 1 min MIASTENIA GRAVE FIGURA 713 Desensibilización de receptores de acetilcolina (AChR). Con la exposición prolongada a ACh, los AChR primero se abren y después entran en un estado desensibilizado y cerrado en el cual ya no muestran respuesta a la ACh. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) rá, pero la respuesta alcanzará un máximo y después declinará (figura 7-13). Esta declinación se llama desensibilización; la molécula de AChR ha entrado a un estado desactivado desde el cual no se abre. Esto es un poco similar desde el punto de vista funcional a la desactivación de canales Nav, excepto porque la evolución temporal, el agente que causa la desactivación, y la base molecular de los canales, son por completo diferentes. La desensibilización quizá no ocurre con el uso normal de uniones neuromusculares, pero puede convertirse en un problema cuando se usan fármacos que bloquean la AChE. Un paciente con AChR sensibilizado puede quedar paralizado y ser incapaz de respirar debido a la falta de AChR funcionales. ALGUNOS FÁRMACOS QUE ACTÚAN EN LA UNIÓN NEUROMUSCULAR El D-tubocurare (o D-tubocurarina) es un bloqueador neuromuscular clásico, descubierto como un veneno de flechas de Sudamérica. El curare se une a AChR de manera reversible y evita que la ACh abra los canales. Después de aplicación de curare, el EPP disminuye de tamaño; si hay suficiente curare, el EPP se hace tan pequeño que ya no desencadena un potencial de acción, de modo similar a la respuesta representada con una línea discontinua en la figura 7-10, y la unión es bloqueada efectivamente. Las dosis más altas de curare pueden eliminar el EPP. El curare reduce el EPP al disminuir el número de receptores disponibles para mostrar respuesta a la ACh. El curare o un fármaco relacionado con frecuencia se usa durante intervención quirúrgica para inmovilizar músculos; también puede facilitar la intubación traqueal y la ventilación mecánica. Las anticolinesterasas, como la neostigmina y la fisostigmina, se combinan con AChE y evitan la hidrólisis de ACh, lo que lleva a un EPP de mayor magnitud. La neostigmina se usa para acelerar la recuperación luego de los efectos del curare, y para reducir los síntomas de miastenia grave. La neostigmina se asocia con peligros; un exceso de ACh puede llevar a desensibilización de los receptores restantes. Asimismo, el organismo usa ACh para lentificar la frecuencia cardiaca y liberar saliva; ambos de estos efectos pueden aumentarse por la fisostigmina. El botulismo es una intoxicación alimentaria en potencia mortal causada por la bacteria anaerobia Clostridium botulinum. Algunas de las toxinas liberadas por este microorganismo son endopeptidasas, que son captadas por células nerviosas y dividen la sinaptobrevina, lo que evita la liberación de transmisor. Las toxinas purificadas se usan en clínica para prevenir transmisión neuromuscular no deseada. La miastenia grave es una enfermedad asociada con debilidad muscular y fatigabilidad con el esfuerzo. Es una enfermedad autoinmunitaria que lleva a la destrucción de AChR. Los pacientes pueden tener sólo 10 a 30% del número de AChR que se encuentran en individuos sanos. El tratamiento con anticolinesterasas aumenta la cantidad de ACh disponible, lo que hace más probable que se activen los AChR restantes (ecuación [1]). Es peligroso administrar demasiada anticolinesterasa, porque puede llevar a desensibilización de los AChR y debilidad adicional. Si esta debilidad se malinterpreta como terapia insuficiente con anticolinesterasa, puede surgir un asa de retroacción positiva trágica que lleva a crisis miasténica. SÍNDROME DE LAMBERTEATON El síndrome de Lambert-Eaton se observa con una enfermedad autoinmunitaria que reduce el número de canales Cav en la terminal presináptica. Con el esfuerzo prolongado, estos pacientes ganan fuerza, lo opuesto de los pacientes miasténicos. La prolongación de los potenciales de acción presinápticos con fármacos que bloquean canales KV, como la diaminopiridina, puede aliviar algunos de los síntomas. La despolarización prolongada abre los canales Cav restantes durante un tiempo más prolongado, lo que permite más entrada de Ca2+ y, por ende, más liberación. Si el experimento que se muestra en la figura 7-11 se efectúa en estas uniones neuromusculares, se encontrará que tienen un contenido cuántico más bajo, es decir, liberan un menor número de vesículas por cada estímulo. Esto contrasta con la miastenia grave, que mostrará el contenido cuántico normal, pero un MEPP de menor magnitud, la despolarización por cada cuanto. ESTIMULACIÓN REPETITIVA La cantidad de transmisor liberada por una sinapsis no es constante de un impulso a otro, sino que depende del antecedente de actividad. Si el nervio que va a una unión neuromuscular es estimulado una vez cada 10 s o a intervalos de tiempo mayores, liberará de manera constante alrededor de 200 vesículas. Si la tasa de estimulación se cambia repentinamente a 50/s, que es aproximadamente la tasa que usa el SNC para causar contracción muscular normal, la cantidad liberada por cada impulso aumentará durante el primer medio segundo y después disminuirá (figura 7-14). El incremento, que se conoce como facilitación, se relaciona con una acumulación de calcio residual en la terminal nerviosa. Se cree que el decremento, llamado depresión, refleja un agotamiento de vesículas en los sitios de liberación. Esta variación no afecta el funcionamiento de la unión neuromuscular en un individuo sano. Cada uno de esos impulsos nerviosos libera ACh para producir un EPP de magnitud suficiente para desencadenar un potencial de acción muscular. Sin embargo, la persona miasténica puede tener transmisión neuromuscular funcio- CAPÍTULO 7 Sinapsis Facilitación X Normal 1 Depresión 40 80 Impulsos 120 FIGURA 714 Facilitación y depresión de la transmisión sináptica en la unión neuromuscular. Con la estimulación repetitiva la cantidad de transmisor liberado por cada estímulo cambia; primero aumenta y más tarde disminuye. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) nal sólo al principio de la tarea, y experimentar debilidad a medida que ocurre depresión con un esfuerzo prolongado y la cantidad de ACh liberada disminuye por debajo de la que se necesita para desencadenar un potencial de acción muscular. Una anticolinesterasa con una duración de acción breve, el cloruro de edrofonio, con frecuencia se utiliza como una prueba para miastenia grave en pacientes que muestran debilitación rápida cuando se les solicita que hagan una contracción sostenida. Cuando se suspende el estímulo de 50/s, hay un incremento de la cantidad de transmisor que puede liberarse con un impulso nervioso único (figura 7-15). El nervio se estimuló una vez cada 30 s antes y después de la estimulación tetánica. Durante el tétanos, la liberación PTP X Normal 1 2 3 4 Minutos FIGURA 715 Potenciación postetánica (PTP) de la transmisión sináptica en la unión neuromuscular. Después del final de un periodo de estimulación repetitiva, la cantidad de transmisor liberado por estímulos poco frecuentes subsiguientes aumenta durante varios minutos. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) aumentó y disminuyó, como en la figura 7-14. Después del tétanos, a medida que la sinapsis se recuperó luego de depresión, se observó una potenciación postetánica (PTP) que duró varios minutos. La PTP también se relaciona con aumento de la concentración residual de Ca2+ en la terminal nerviosa, pero tiene un inicio y una declinación más lentos que la facilitación. La PTP se usa como un procedimiento diagnóstico después de procedimientos quirúrgicos cuando se han usado curare u otros bloqueadores neuromusculares para evitar movimiento no deseado. El anestesiólogo administrará al paciente inhibidores de la colinesterasa, pero desea saber cuándo se ha administrado suficiente inhibidor a fin de evitar administrar demasiado y de sensibilizar los AChR. El anestesiólogo repetirá el experimento que se muestra en la figura 7-15; estimulará la rama tenar del nervio mediano del paciente y palpará la fuerza de la contracción de los músculos tenar. Se administran dos descargas eléctricas antes del tétanos y después una 30 s más tarde. Bajo efectos profundos del curare, ninguna de éstas producirá una contracción palpable. A medida que más ACh queda disponible mediante bloqueo de la esterasa, el estímulo posterior al tétanos dará una respuesta de mayor tamaño que las dos anteriores al tétanos porque será la primera con un EPP suficientemente grande como para excitar el músculo. El punto terminal es cuando se ha administrado bastante esterasa, de modo que las tres respuestas son iguales porque los tres EPP están por arriba del umbral para la activación muscular. SINAPSIS DEL SISTEMA NERVIOSO AUTÓNOMO POTENCIACIÓN POSTETÁNICA 1 71 La sistema nervioso autónomo (SNA) tiene dos divisiones, ambas con dos sinapsis fuera del SNC (figura 7-16). La sinapsis más cercana al SNC se denomina la sinapsis ganglionar; los nervios que entran a los ganglios y salen de los mismos se llaman preganglionares y posganglionares. Los ganglios simpáticos yacen en una cadena adyacente a la columna vertebral; los ganglios parasimpáticos están cerca de los órganos terminales donde ocurre la segunda sinapsis. Las segundas sinapsis están sobre músculos lisos o células cardiacas o células de glándulas. Muchos tejidos reciben inervación tanto simpática como parasimpática. El transmisor primario en la sinapsis ganglionar de ambas divisiones es la ACh; los receptores son nAChR nicotínicos que son pentámeros heteroméricos de productos de gen relacionados, pero diferentes que los nAChR del músculo esquelético. Los receptores ganglionares son menos sensibles al curare y son bloqueados con mayor facilidad con hexametonio. El transmisor posganglionar primario en el sistema nervioso simpático es la NE, y hay dos categorías de GPCR en las células postsinápticas que se llaman receptores α y β-adrenérgicos. El receptor posganglionar primario en la división parasimpática es la AChR, y los receptores son mAChR muscarínicos, que también son GPCR, pero en general con proteínas G distintos de los receptores de NE. Por lo general, se describe que las sinapsis ganglionares se comportan más o menos como la unión neuromuscular. Sin embargo, la situación es más complicada; las neuronas postsinápticas tienen dendritas con más de una terminación nerviosa presináptica en ellas. Al analizar los transmisores peptídicos que están en esas célu- 72 SECCIÓN II Fisiología celular SNC ACh Parasimpática ACh Preganglionar ACh Simpática Posganglionar Blanco NE Blanco ACh Epinefrina Motora ACh Músculo FIGURA 716 Esquema de las fibras eferentes del sistema nervioso autónomo y las motoneuronas. De arriba a abajo: parasimpática, simpática, médula suprarrenal, motoneurona. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) las junto con sus transmisores clásicos, se distinguieron diferentes subpoblaciones de células presinápticas y postsinápticas. Las células posganglionares también tienen mAChR que producen un EPSP lento mediante cierre de un canal de K. En los ganglios también hay células pequeñas, muy fluorescentes, que están inervadas por fibras preganglionares y liberan NE o dopamina. Si se considera todo lo anterior, parece que debe llevarse a cabo algún cálculo en los ganglios, más que el simple circuito de cruce que se observa en la unión neuromuscular. Las sinapsis entre las células posganglionares y los órganos terminales son distintas de las que se encuentran en la unión neuromuscular. Los procesos presinápticos son similares, pero las células posganglionares hacen sinapsis “de paso” sobre blancos tisulares a lo largo del axón. Las vesículas sinápticas se almacenan en varicosidades del nervio, que continúan a otras varicosidades antes de llegar a su terminal. La activación de mAChR por el SNA aumenta el tono y la motilidad GI, incrementa el tono de la vejiga urinaria y su motilidad, aumenta la salivación y la sudoración, constriñe bronquiolos, y disminuye la frecuencia cardiaca y la presión arterial. El SNA activa receptores α, β1 y β2-adrenérgicos, y los receptores α-adrenérgicos incrementan la presión arterial. Los receptores β1-adrenérgicos aumentan la frecuencia cardiaca y la fuerza de contracción y la presión arterial. Los receptores β2-adrenérgicos dilatan los bronquiolos en los pulmones. El mecanismo de los efectos sobre los músculos cardiaco y liso se comenta en la sección siguiente. Se han utilizado muchos fármacos agonistas y antagonistas para controlar estos procesos, algunos con más especificidad que otros. Así, hay agonistas α y bloqueadores β específicos. Las anfetaminas y la cocaína tienen un efecto adrenérgico indirecto al estimular la liberación de NE. Algunos compuestos, como la efedrina, tienen efectos adrenérgicos tanto directos como indirectos. La atropina es el antagonista de mAChR arquetípico; sus efectos son los opuestos a los que se atribuyen a la ACh antes mencionados. En muchos sitios hay una liberación tónica tanto de ACh como de NE desde el SNA, de modo que el bloqueo de un grupo de receptores puede producir efectos similares a la activación del otro. SINAPSIS DEL SISTEMA NERVIOSO CENTRAL El SNC del ser humano tiene miles de millones de neuronas, con billones de sinapsis entre ellas. Una neurona única puede tener miles de aferencias tanto excitadoras como inhibidoras; algunas neuronas de mayor tamaño tienen más de 100 000 terminaciones sobre ellas. Para dar cabida a esta convergencia de aferencias sinápticas, casi todas las neuronas tienen un árbol dendrítico que expande mucho el área disponible para el contacto sináptico. El cuerpo celular (soma) y la región inicial del axón (montículo del axón) integran las señales sinápticas que llegan, y determinan cuándo y qué tan a menudo la neurona activará potenciales de acción (figura 7-17). El axón transporta la eferencia de la neurona al grupo siguiente de neuronas o a células de músculo esquelético si es una motoneurona. Por lo general, sólo sale un axón del cuerpo celular, pero más tarde se ramifica para permitir que la neurona haga sinapsis con otras células. Esta divergencia de información, combinada con la convergencia de muchas aferencias hacia una neurona, imparte al SNC gran parte de su poder de cálculo. Cada neurona en el SNC actúa como una o más computadoras pequeñas. Si bien cada célula realiza sus cálculos en milisegundos, millones de veces más lento que la unidad de procesamiento central de una computadora moderna, los miles de millones de neuronas que operan en paralelo hacen que el SNC descolle en la comparación. El SNC es capaz de crear cada pensamiento en la historia registrada, mientras que al mismo tiempo regula tanto la ambulación como mascar chicle. Las sinapsis hacen posible esto. El aprendizaje y la memoria se logran con la modificación de sinapsis. CAPÍTULO 7 Sinapsis 73 Los axones convergen sobre la neurona Dendritas Las aferencias sinápticas cubren casi toda la superficie del soma y las dendritas Soma Montículo o segmento inicial de axón Axón Las sinapsis divergen FIGURA 717 La convergencia y divergencia de sinapsis en el SNC. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) Hay dos tipos generales de sinapsis en el SNC: eléctricas y químicas. Las sinapsis eléctricas operan mediante flujo directo de corriente eléctrica desde la neurona presináptica hacia la neurona postsináptica a través de canales de unión intercelular comunicante (conexión comunicante) entre las membranas de las dos células (figura 7-18). No hay neurotransmisores involucrados, y las sinapsis eléctricas pueden tener menos retraso sináptico que las sinapsis químicas. Sin embargo, a diferencia de las sinapsis químicas, las sinapsis eléctricas no pueden amplificar la señal, ni revertir la dirección del flujo de corriente. Las uniones intercelulares comunicantes, que funcionan como sinapsis eléctricas y permiten que potenciales de acción fluyan de manera selectiva de una célula a otra, también conectan células en el corazón y algunos tipos de músculo liso. FIGURA 718 Una sinapsis eléctrica. La corriente pasa directamente de la célula presináptica a la postsináptica a través de canales célula-célula especializados. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) Hay dos tipos generales de sinapsis químicas en el SNC: excitadoras e inhibidoras. Las sinapsis excitadoras generan EPSP que despolarizan la membrana hacia el umbral. Las sinapsis inhibidoras generan IPSP que hiperpolarizan la membrana o se resisten a la despolarización hasta el umbral. Cada uno de estos tipos puede subdividirse en canales iónicos quimiosensitivos (o receptores ionotrópicos) y canales iónicos enlazados a proteína G (o receptores metabotrópicos). Los canales iónicos quimiosensitivos típicamente dan lugar a eventos sinápticos rápidos que duran algunos milisegundos; los canales iónicos enlazados a proteína G pueden producir efectos durante cientos de milisegundos. INTEGRACIÓN DE CORRIENTES SINÁPTICAS Las sinapsis excitadoras e inhibidoras inyectan corriente (positiva o negativa) en células. Estas corrientes fluyen hacia el cuerpo celular y se suman. Los IPSP se diseminan de manera pasiva hacia el sitio de inicio de la espiga o la parte de la célula con el umbral más bajo debido a las propiedades de cable de la célula. Las sinapsis más distales estarán disminuidas en comparación con las que están cerca del sitio. La célula produce el sitio de inicio de espiga al controlar la densidad local de canales Nav. A menudo el sitio de inicio de la espiga es el cono axónico cerca del inicio del axón (figura 7-17) o en el primer nodo de Ranvier. Dado que los PSP duran entre varios y muchos milisegundos, pueden sumarse unos con otros aun cuando no ocurren de manera sincrónica; esto se llama suma temporal. Los efectos de sinapsis en diferentes ubicaciones sobre la misma célula postsináptica también pueden sumarse; esto se llama suma espacial. La suma espacial es ponderada de manera inversa por la distancia desde la sinapsis hasta el sitio de inicio del potencial de acción. 74 SECCIÓN II Fisiología celular Hendidura sináptica Espina postsináptica sobre dendrita Axón Terminal nerviosa presináptica Vesículas sinápticas Densidad postsináptica FIGURA 719 Una sinapsis del SNC. Éstas son menos complejas que las sinapsis en la unión neuromuscular (figura 6-9). (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/ McGraw-Hill, 2006.) En la figura 7-19 se presenta un esquema de una sinapsis química en el SNC. La terminal presináptica tiene alrededor de 1 μm de diámetro y contiene mitocondrias y vesículas sinápticas llenas con neurotransmisor. La despolarización de la terminal abre canales Cav, y fluye Ca2+ en favor de su gradiente electroquímico para actuar sobre la sinaptotagmina y desencadenar la fusión de algunas vesículas con la membrana presináptica para producir exocitosis del neurotransmisor. A continuación la membrana se recicla y las vesículas se vuelven a llenar. Los receptores postsinápticos a menudo están sobre protrusiones desde dendritas, llamadas espinas, aunque también se encuentran sinapsis sobre la vaina dendrítica, el cuerpo de la célula neuronal y otras terminaciones sinápticas. Las sinapsis en el SNC comparten muchas características con la unión neuromuscular, pero difieren en varios aspectos importantes. Las sinapsis en el SNC son de tamaño mucho menor y liberan menos vesículas, típicamente menos de cinco por cada impulso, en comparación con 200 en la placa terminal motora. En el SNC, las hendiduras sinápticas son más estrechas, de alrededor de 20 nm, y las cadherinas y otras moléculas de adhesión celular abarcan la brecha. La ACh es el transmisor en la unión neuromuscular; hay una amplia variedad de transmisores en el SNC. El EPP siempre es excitador y suficientemente grande como para llevar la membrana muscular al umbral; las sinapsis en el SNC son excitadoras o inhibidoras, y el umbral se alcanza con la combinación de cientos de EPSP. Hay algunas sinapsis excepcionales en el SNC. En el cerebelo, un axón de fibra trepador puede hacer docenas de sinapsis sobre una célula de Purkinje. En el cáliz de Held, en la vía auditiva, la terminación presináptica forma una cubierta con tallos digitiformes que envuelven la neurona postsináptica, y cubren alrededor de 40% de su soma. En estas dos sinapsis un impulso presináptico único libera cientos de cuantos, y el EPSP que resulta es lo bastante grande como para desencadenar un potencial de acción postsináptico. El glutamato es el principal neurotransmisor excitador en el SNC. Hay varios receptores de glutamato postsinápticos, tanto canales como GPCR. Los canales pueden agruparse en dos tipos principales, canales NMDA y no NMDA, de acuerdo con su sensibilidad al agonista sintético N-metil-d-aspartato. Ambos tipos muestran respuesta al glutamato. Los canales no NMDA pueden llamarse canales de AMPA, quisqualato, o kainato, de acuerdo con cuál de estos agonistas no fisiológicos los abre. Los canales no NMDA generan EPSP rápidos que duran alrededor de 5 ms. Cuando son activados por glutamato, los canales no NMDA permiten que el Na+ y K+ fluyan a través de sus poros. Cada ion se mueve en la dirección que tenderá a llevar el potencial de membrana a su potencial de equilibrio de Nernst. Dado que ambos se mueven, el potencial de membrana tiende a aproximarse al promedio de los dos potenciales de equilibrio, que es de alrededor de –10 mV. Este potencial, donde las dos corrientes iónicas son iguales, se llama potencial reverso para el canal. Cuando estos canales se abren a potenciales más negativos que el potencial reverso, la tendencia a que el Na+ entre a la célula dominará, y la membrana se despolarizará hacia el potencial reverso. Si el potencial inicial fuera más positivo que el potencial reverso, los iones K+ dominarían, y la célula se hiperpolarizaría hacia el potencial reverso. Los canales receptores NMDA generan EPSP que duran cientos de milisegundos. Los canales NMDA abiertos permiten que el Na+ y el K+, y el Ca2+, pasen a través de sus poros. En presencia de glutamato, los canales de NMDA sólo se abren si la célula postsináptica también se despolariza con algún otro medio. Este control doble de la entrada de Ca2+ tiene un papel clave en el aprendizaje (véase más adelante). El GABA es el principal transmisor inhibidor en el cerebro. La glicina es un transmisor inhibidor en el tallo encefálico y la médula espinal. El GABA abre canales GABAA de manera directa, lo cual permite que iones Cl– pasen a través de sus poros. El GABA también puede causar inhibición por medio de receptores GABAB, que son GPCR que llevan a la abertura de canales de K. El potencial reverso para canales GABAA está en el potencial de Nernst para Cl–, a alrededor de –80 mV. Si la membrana es más positiva que ECl, entrará Cl– a la célula y hará más negativo el potencial de membrana, lo cual hará menos probable que inicie un potencial de acción. Las benzodiazepinas, como el diazepam y los barbitúricos, aumentan la probabilidad de abertura de GABAAR activados. Ambos se usaron como sedantes y anticonvulsivos. Los anestésicos generales, como éter, cloroformo y halotano aumentan la duración de IPSP, y disminuyen la amplitud de EPSP y la duración de los mismos. SNC, NEUROTRANSMISORES MODULADORES En el SNC, la ACh, NE, dopamina y serotonina actúan sobre todo como moduladores difusos de actividad; actúan en marcos de tiempo que son largos, en comparación con los potenciales de acción, en lugar de estar involucrados en tareas separadas específicas. Cada uno de estos neurotransmisores tiene su propio juego de neuronas y blancos; algunas de estas neuronas pueden influir sobre más de 100 000 neuronas postsinápticas. Los receptores postsinápticos son metabotrópicos y alteran la capacidad de respuesta de las neuronas postsinápticas por medio de vías de segundo mensajero. También hay nAChR ionotrópicos en el SNC, pero hay 10 a 100 veces más mAChR. Los sistemas moduladores de ACh y NE forman parte del sistema activador reticular ascendente que despierta el prosencéfalo en respuesta a estímulos. De manera general, los sistemas moduladores desempeñan un papel en el SNC similar al papel que desempeña el SNA en el resto del cuerpo. CAPÍTULO 7 Sinapsis 75 Axón B A Neurona C FIGURA 720 La inhibición presináptica puede ocurrir por una sinapsis (A y B) sobre otra terminación sináptica. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) INHIBICIÓN PRESINÁPTICA Algunas sinapsis del SNC actúan de manera directa sobre otras terminaciones sinápticas en lugar de sobre dendritas o cuerpos celulares (figura 7-20). La terminal A libera GABA sobre la terminal B, lo que activa canales de Cl que tienden a hiperpolarizar la terminal B. Si un potencial de acción llega en B mientras los canales de Cl están abiertos, la amplitud del potencial de acción se reducirá, de modo que abrirá menos canales Cav y, por ende, la terminal B liberará menos vesículas, y tendrá un efecto de menor magnitud sobre la neurona C. TRANSMISORES QUÍMICOS LIBREMENTE DIFUSIBLES RETRÓGRADOS Además de los transmisores clásicos que son liberados a partir de vesículas y se unen a receptores, hay mensajeros químicos en el SNC con un modo de operación diferente. El óxido nítrico (NO) no se almacena, sino que más bien se produce cuando es necesario. Puede difundirse con libertad a través de las membranas celulares desde el interior de una célula (un cuerpo celular postsináptico) hacia el interior de otras células (terminaciones presinápticas), donde altera algunas reacciones químicas. El NO puede diseminarse hacia varias terminaciones presinápticas en la vecindad. Es eliminado del tejido por unión a hemoglobina. La anandamida, un cannabinoide endógeno, también se produce según se requiere en células postsinápticas, y llega al espacio extracelular con un proceso no vesicular. Se une a receptores de cannabinoide (CB1) presinápticos, que son GPCR, y pueden alterar la liberación subsiguiente de neurotransmisores tradicionales. ACTIVACIÓN REPETITIVA DE CÉLULAS NERVIOSAS Si un axón o una célula muscular queda sujeto a una despolarización mantenida, mostrará respuesta con un potencial de acción o quizá con dos, y después dejará de activarse porque los canales Nav entran en el estado desactivado y requieren un breve periodo cerca del potencial de reposo para recuperarse. Muchas células del SNC y las terminaciones nerviosas sensoriales de adaptación lenta mostrarán respuesta a una despolarización sostenida con una serie de potenciales de acción a alrededor de 50/s. Los canales Cav y los canales de K activados por Ca hacen posible esto. La despolarización del potencial de acción abre los canales Cav, y el Ca2+ que entra abre los canales de K activados por Ca al unirse a la porción intracelular de la molécula. A continuación, el canal de K activado por Ca permite que el K+ salga y el potencial de membrana se aproxime a EK para una hiperpolarización de larga duración, suficientemente prolongada como para que los canales Nav se recuperen luego de desactivación (figura 7-21). El equilibrio entre el estímulo sostenido y la tasa a la cual se elimina Ca2+ desde los canales de K activados por Ca determina la tasa de activación. APRENDIZAJE, MEMORIA Y PLASTICIDAD SINÁPTICA La base celular del aprendizaje y la memoria es un remodelado funcional de conexiones sinápticas, que con frecuencia se conoce como plasticidad sináptica. Esto incluye memoria tanto explícita o declarativa cuando la persona puede recordar y describir algún hecho o evento pasado, y memoria implícita o procedural, como en una habilidad motora aprendida. La memoria, por lo regular, se subdivi- 76 SECCIÓN II Fisiología celular +40 mV 0 –60 –70 0 200 400 ms Aferencia sináptica excitadora sostenida FIGURA 721 Activación repetitiva de una motoneurona. A diferencia de los axones, muchas células nerviosas muestran respuesta de manera repetitiva a una aferencia sostenida. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) de a corto plazo, de minutos a horas, y a largo plazo, de días a toda la vida. La formación de memoria a corto plazo comprende la modificación de proteínas existentes, a menudo por medio de fosforilación. Los cambios a largo plazo involucran activación de gen, síntesis de proteína y reordenamiento de membrana, incluso la formación, o resorción, o ambas, de terminales presinápticas y espinas postsinápticas. En algunos estudios se ha mostrado que el volumen de corteza cerebral que se dedica a una tarea aumenta con el entrenamiento específico. El fenómeno de aprendizaje celular estudiado de manera más intensiva es la potenciación a largo plazo (LTP) en sinapsis hipocampales. El hipocampo se requiere para la formación de nuevos recuerdos a largo plazo. Si ambos hipocampos están alterados, la persona vivirá en el presente, sin recuerdo de eventos después del daño. En el hipocampo, la LTP ocurre en sinapsis de glutamato entre células CA3 presinápticas y células CA1 postsinápticas. La LTP y la depresión a largo plazo (LTD) relacionada también ocurren en otros sitios en el SNC. El experimento clásico es similar a la demostración de PTP que se muestra en la figura 7-15; la sinapsis se prueba con poca constancia, sujeta a estimulación de alta frecuencia, y des- pués probada con poca continuidad de nuevo. A diferencia de la PTP, que desaparece en algunos minutos, con la LTP la potenciación permanece durante muchas horas o días (figura 7-22). Asimismo, a diferencia de la PTP, la LTP es un evento postsináptico. Es innecesario proporcionar la estimulación de alta frecuencia a las terminales presinápticas; la despolarización simple de la célula postsináptica, emparejada con la estimulación presináptica, inducirá LTP. Esta respuesta de pares de aferencias hace a la LTP una base candidato para el aprendizaje asociativo. Hay dos tipos de receptores de glutamato en las membranas postsinápticas: receptores AMPA (no NMDA) y NMDA. Durante la estimulación no pareada de baja frecuencia, sólo los receptores AMPA están activados; los receptores NMDA están bloqueados por iones Mg2+ externos. Los canales del receptor AMPA son permeables al Na+ y K+; cerca del potencial de reposo, el movimiento de Na+ hacia la célula es favorecido. Cuando la membrana postsináptica se despolariza, sea por aferencia sináptica de alta frecuencia o al inyectar corriente en la célula postsináptica, el Mg2+ es impulsado fuera de los receptores NMDA y muestra respuesta al glutamato y permite que el Na+ y el Ca2+ entren en la célula. El Ca2+ alto activa una serie de eventos bioquímicos que llevan a la inserción de más receptores AMPA en la membrana postsináptica. La LTP se ha asociado con aprendizaje en ratas cuando se usa un laberinto de agua. Las ratas con extirpación quirúrgica del hipocampo no aprenden el laberinto; tampoco lo aprenden las ratas que recibieron tratamiento con un antagonista específico para canales de receptor NMDA. Hay otros ejemplos de plasticidad sináptica en otras regiones del cerebro, y bien puede ser que haya mecanismos adicionales, incluso acción retrógrada del NO o la anandamida. CORRELACIÓN CLÍNICA Aproximadamente un mes antes de acudir al hospital, una mujer de 56 años de edad notó que era incapaz de sostener su bolsa de compras, y que su cabeza caía hacia adelante al arrodillarse para atarse los zapatos. Dos semanas más tarde tuvo que permanecer en cama y tenía dificultad para enderezarse. La mandíbula empezó a caer, tuvo que sostenerla levantada con la mano, y el párpado izquierdo también empezó a caer. Su habla se hizo indistinta cuando estuvo excitada; presentó dificultad 5 4 mV 3 2 1 0 10 20 30 Minutos 40 50 60 FIGURA 722 Potenciación a largo plazo. Un estímulo condicionante breve (barra azul) causa un aumento a largo plazo de la eficacia sináptica. (Modificada con autorización de Landowne D: Cell Physiology. New York: Lange Medical Books/McGraw-Hill, 2006.) CAPÍTULO 7 Sinapsis ■ para deglutir, y a veces regurgitaba líquido por la nariz. Algunos días después de la admisión al hospital, presentó debilidad en los dedos medio y anular de ambas manos, que aumentó con la excitación y disminuyó en reposo. No hubo emaciación muscular y todos los reflejos tendinosos estuvieron presentes. Los músculos maseteros mostraron una respuesta decreciente a la estimulación eléctrica tetánica. En el hospital se le inyectó 1 mg de fisostigmina. Alrededor de una hora más tarde, el párpado izquierdo se elevó, los movimientos del brazo fueron más fuertes, la mandíbula colgó menos, la deglución mejoró, y la paciente reportó sentirse “menos pesada”. El efecto desapareció de manera gradual en 2 a 4 h. Con 1.3 mg la mejoría fue mayor y duró 4 a 5 h. Después de una inyección de 1.5 mg se observaron mejorías aún mayores, que duraron 6 a 7 h, pero la paciente se sintió débil, como si “algo fuera a suceder”. El diagnóstico es miastenia grave. Es una enfermedad autoinmunitaria que afecta a alrededor de una de cada 5 000 personas. El sistema inmunitario produce anticuerpos contra el AChR nicotínico de la unión neuromuscular, y la transmisión neuromuscular está alterada. Con menos receptores los efectos de la depresión de la transmisión sináptica llevan a falla de la transmisión neuromuscular durante esfuerzo sostenido. La fatiga es una característica de la enfermedad. La fisostigmina es un inhibidor de la AChE. En su presencia una mayor cantidad de la ACh liberada puede interactuar con el receptor, y la transmisión es más confiable. Se usan inhibidores de la AChE para aliviar los síntomas de miastenia grave. Los inmunosupresores, por ejemplo, el corticosteroide sintético prednisona, se usan para reducir la producción de anticuerpos. En algunos casos se practica timectomía (extirpación quirúrgica del timo) para suprimir el sistema inmunitario. El cloruro de edrofonio es un inhibidor de la AChE de acción breve que se ha usado para ayudar en el diagnóstico. La estimulación eléctrica y las pruebas para anticuerpos circulantes también se usan con fines diagnósticos. RESUMEN DEL CAPÍTULO ■ ■ ■ ■ ■ Las sinapsis pueden ser químicas o eléctricas. Las sinapsis químicas pueden ser excitadoras o inhibidoras. En las sinapsis químicas, la terminal presináptica almacena un neurotransmisor en vesículas. Cuando la sinapsis es activada, se libera el contenido de las vesículas y a continuación un proceso de reciclado recupera parte del transmisor y los componentes vesiculares liberados. La ACh es el neurotransmisor en la unión neuromuscular. También es un componente de importancia de las sinapsis de los sistemas nerviosos autónomo y central. El glutamato es el principal neurotransmisor excitador en el SNC. El GABA y la glicina son los principales neurotransmisores inhibidores en el SNC. ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ 77 Varias aminas biogénicas son neurotransmisores importantes. La NE es liberada por nervios simpáticos para controlar los músculos lisos cardiaco y vascular. Los neuropéptidos son proteínas pequeñas liberadas como neurotransmisores. La liberación sináptica involucra muchas proteínas y está controlada por canales Cav, que son abiertos cuando un potencial de acción invade la terminal presináptica. Los axones tienen un sistema de transporte que se basa en microtúbulos para mover materiales desde el cuerpo celular hacia la terminal presináptica (transporte ortógrado) y en la otra dirección (transporte retrógrado). Los PSP son excitadores (EPSP) si hacen que la célula postsináptica tenga más probabilidades de iniciar un potencial de acción, e inhibidores (IPSP) si hacen que eso sea menos probable. La transmisión neuromuscular es un ejemplo bien estudiado de transmisión sináptica. La hipocalcemia reduce el número de vesículas que se liberan cuando un potencial de acción invade la terminal presináptica. En la unión neuromuscular, el número de canales abiertos es proporcional a la concentración de ACh al cuadrado multiplicada por el número efectivo de canales de AChR. Varios fármacos importantes en clínica actúan en la unión neuromuscular. El número de vesículas liberadas por cada potencial de acción depende de la frecuencia y el patrón de llegada de los potenciales de acción. El SNA tiene dos sinapsis fuera del sistema nervioso central. La primera es colinérgica; la segunda es adrenérgica o colinérgica. En general, las sinapsis del SNC son similares a la unión neuromuscular, pero difieren en muchos aspectos importantes. En el SNC, varios transmisores actúan por medio de receptores acoplados a proteína G para modular la actividad del cerebro. Para activarse de manera repetitiva las células nerviosas usan canales de K activados por Ca para hiperpolarizar la célula y permitir que los canales Nav se recuperen de su desactivación. El aprendizaje y la memoria comprenden cambios de la eficacia sináptica. PREGUNTAS DE ESTUDIO 1. Se necesitan iones Ca2+ en la solución extracelular para la transmisión sináptica porque: A) entran iones Ca2+ a la terminal nerviosa presináptica con la despolarización y desencadenan la liberación del contenido de vesículas sinápticas hacia la hendidura sináptica. B) se requieren iones Ca2+ para activar el metabolismo del glucógeno en la célula presináptica. C) los iones de Ca2+ deben entrar en la célula postsináptica para despolarizarla. D) los iones Ca2+ evitan que los iones Mg2+ liberen el transmisor en ausencia de impulsos nerviosos. E) los iones Ca2+ inhiben la acetilcolinesterasa, lo que permite que la acetilcolina liberada llegue a la membrana postsináptica. 78 SECCIÓN II Fisiología celular 2. Los potenciales postsinápticos inhibidores pueden surgir a partir de todos los que siguen, excepto: A) permeabilidad aumentada de la membrana de nervio al ion Cl–. B) aplicación directa de GABA a neuronas. C) permeabilidad aumentada de la membrana de nervio a ion K+. D) permeabilidad aumentada de la membrana celular a ion Na+. 3. Las sinapsis eléctricas y químicas difieren en que: A) las sinapsis eléctricas tienen un retraso sináptico más prolongado que las químicas. B) las sinapsis químicas pueden amplificar una señal, no así las eléctricas. C) las sinapsis eléctricas no tienen una hendidura sináptica, mientras que las eléctricas la tienen. D) las sinapsis eléctricas usan canales activados por agonista, no así las químicas. E) las sinapsis eléctricas sólo se encuentran en animales invertebrados, mientras que las químicas se hallan en todos los animales. 4. ¿Cuál de los que siguen no contribuye a la integración de potenciales sinápticos por neuronas? A) convergencia de muchas aferencias sinápticas en una neurona, lo que permite suma espacial B) la presencia de EPSP que tiene amplitudes que exceden el umbral para la generación de un potencial de acción en la neurona C) suma temporal de potenciales sinápticos en neuronas debido a la constante de tiempo de las neuronas D) el flujo de corrientes desde las regiones distales de las dendritas hacia el soma debido a las constantes de longitud de las dendritas E) aferencias sinápticas inhibidoras 5. ¿Cuál de los iones que siguen es contratransportado para dar energía al transporte de neurotransmisor hacia vesículas presinápticas? A) Na+ B) K+ C) H+ D) Cl– E) Ca2+ 6. Una rama del nervio cubital de un varón de 26 años de edad fue aplastada en su antebrazo izquierdo, y cortó axones en un punto a una distancia de 15 cm (6 pulgadas) de la parte medial de la palma, donde se perdió la sensibilidad cutánea. ¿Aproximadamente cuánto tiempo es probable que transcurra antes de que el paciente empiece a sentir estímulos en esa parte de la palma? A) un día B) 10 días C) 100 días D) 1 000 días E) nunca, porque los axones periféricos no se regeneran. 7. ¿Los tratamientos para envenenamiento por gas nervioso se dirigen a cuál de las proteínas que siguen? A) acetilcolinesterasa (AChE) y colina acetiltransferasa (CAT) B) AChE y receptores de acetilcolina nicotínicos C) receptores de acetilcolina muscarínicos y nicotínicos D) receptores de acetilcolina muscarínicos y AChE E) CAT y transportadores de colina sinápticos SECCIÓN III FISIOLOGÍA MUSCULAR Perspectiva general de la función muscular Kathleen H. McDonough C A P Í T U L O 8 O B J E T I V O S ■ ■ ■ ■ Explicar las diferencias en los músculos esquelético, cardiaco y liso en lo que se refiere a aspecto, proteínas contráctiles, proteínas de unión a calcio y aferencias nerviosas. Describir cómo el sarcolema y el retículo sarcoplasmático están involucrados en la contracción muscular. Explicar la fuente de energía para la contracción. Describir el papel de las proteínas que siguen: actina, miosina, receptores de dihidropiridina, receptores de rianodina, calmodulina y troponina. El requerimiento de calcio para iniciar la contracción es uniforme en todo el músculo. Los mecanismos para aumentar el calcio para la contracción pueden variar de un tipo de músculo a otro; la eliminación de calcio para la relajación también cambia. Sin embargo, el unificador predominante es que la concentración citosólica de calcio debe aumentar para que ocurra contracción, y disminuir para que haya relajación. Puesto que en el músculo liso por lo general hay algo de contracción moderada, el calcio citosólico debe aumentar para que la fuerza de la contracción se incremente, y disminuir para que dicha fuerza se reduzca. Así, los organelos como el sarcolema (SL) y el retículo sarcoplasmático (SR, del inglés sarcoplasmic reticulum) que contienen proteínas, las cuales efectúan flujos de calcio, están organizados y son muy eficientes en el músculo. Por ejemplo, la concentración de calcio en el músculo cardiaco en reposo es de sólo 0.1 μM, y puede aumentar 100 veces durante excitación; por ende, la eliminación de calcio es crucial para que haya relajación. Los procesos involucrados en el movimiento del calcio son importantes en el entendimiento de la contracción muscular en los tres tipos de músculo. COMPARACIÓN GENERAL El músculo es el sistema de mayor tamaño del organismo. Consta de tres tipos diferentes con base en factores como características morfológicas, vías de emisión de señales celulares, maneras de alterar la fuerza de la contracción, patrón de contracción (cíclico en contraposición con gradado), y la función del sistema nervioso en la función muscular. Los tres tipos de músculo son esquelético, cardiaco y liso. En su mayor parte, el músculo esquelético está fijo a huesos, y representa alrededor de 40% de la masa corporal de una persona sana típica. El músculo cardiaco es el principal componente del corazón, y se contrae de una manera cíclica durante toda la vida del individuo. El músculo liso es el principal componente de los órganos del tracto gastrointestinal, la vejiga, el útero, las vías respiratorias y los vasos sanguíneos —en general, el músculo liso constituye las paredes de estructuras huecas en el cuerpo, excepto el corazón. 79 80 SECCIÓN III Fisiología muscular La contracción muscular es apoyada por hidrólisis de trifosfato de adenosina (ATP). El ATP es producido por fosforilación oxidativa mitocondrial a partir de sustratos proporcionados por reservas de glucógeno o triacilglicerol en el tejido o por sustratos transportados por la sangre, como glucosa y ácidos grasos. La glucólisis también produce ATP, pero no de manera tan eficiente como la fosforilación oxidativa. En algunos tipos de músculo esquelético, el glucógeno proporciona glucosa para la glucólisis que suministra energía para la contracción rápida, a corto plazo. Además, la energía la proporciona el fosfato de creatina (CP), que puede producir con rapidez una fuente de enlaces fosfato de alta energía para la resíntesis de ATP a partir de ADP. El ADP es producido por enzimas ATPasa que se ubican en la célula muscular. El CP no la usa de manera directa la ATPasa, sino que se utiliza para la regeneración rápida de ATP en el sitio de uso de este último. El ATP se emplea tanto para la contracción del músculo como para la relajación del mismo. La miosina ATPasa hidroliza ATP para proporcionar la energía para el deslizamiento del filamento de actina sobre el de miosina; el ATP también proporciona la energía para la eliminación de calcio desde el citosol por calcio ATPasas, de modo que pueda tener lugar la relajación. DIFERENCIAS DE LOS MÚSCULOS ESQUELÉTICO, CARDIACO Y LISO Los músculos esquelético y cardiaco tienen aspecto estriado debido a la disposición ordenada de las proteínas contráctiles actina y miosina. En el músculo liso, la contracción depende de dichas proteínas, pero no están dispuestas en un patrón tan organizado; por ende, no aparecen estriaciones. El músculo esquelético es el único tipo de músculo que es voluntario —el individuo decide en su mayor parte cuándo contraer el músculo esquelético, y el músculo tiene que ser activado por neuronas reguladas por el sistema nervioso central. El músculo cardiaco es involuntario; se contrae de manera espontánea. Los potenciales de acción son generados por células especializadas dentro del corazón mismo; así, es posible trasplantar el corazón de una persona a otra, y el corazón funciona de manera adecuada, incluso sin aferencias nerviosas en el corazón trasplantado del receptor. Sin embargo, la frecuencia de latidos del corazón, así como la fuerza de la contracción de las células cardiacas, pueden ser reguladas por el sistema nervioso autónomo (SNA; capítulo 19). El sistema nervioso simpático (SNS), componente del SNA, aumentará la frecuencia cardiaca, mientras que el sistema nervioso parasimpático (SNP) la disminuirá. El músculo liso también es involuntario. Tiene el potencial de contraerse a partir de muchos tipos de estímulos diferentes, pero no requiere aferencias nerviosas para que ocurra contracción. Incluso los cambios del potencial de membrana en reposo y el estiramiento del músculo pueden cambiar la fuerza de la contracción. El músculo liso, por lo general, no muestra contracciones seguidas por relajación completa como lo hacen los músculos esquelético y cardiaco, sino que más bien muestra aumento de la fuerza de contracción o disminución de la misma. Por ejemplo, si todo el músculo liso vascular que constituye los vasos sanguíneos de los órganos se relajaran por completo, el individuo entraría en choque; la presión arterial disminuiría hasta cifras peligrosamente bajas. Esto sucede en condiciones patológicas tan graves como lesión cerebral, que causa supresión de todo el control nervioso del músculo liso vascular y da lugar a choque neurogénico. La presión arterial no puede mantenerse si todo el músculo liso vascular en los vasos sanguíneos está relajado por completo. En el músculo liso, las gradaciones de la contracción están reguladas y afectadas por muchas influencias distintas, dependiendo de la ubicación del músculo liso y de la función del mismo. CALCIO Aunque se requiere calcio de manera uniforme para que el músculo se contraiga o para que aumente la fuerza de la contracción, las proteínas de unión a calcio en los tres tipos de músculo difieren, como lo hacen las fuentes de calcio. La troponina es la proteína de unión a calcio que inicia la contracción en los músculos esquelético y cardiaco. La calmodulina se une al calcio en el músculo liso e inicia aumentos de la fuerza de contracción. La actina y la miosina forman los puentes en los tres tipos de músculo. La fuente del calcio que inicia la contracción es diferente en los tres tipos de músculo. El calcio liberado a partir del SR por medio de receptores o canales de rianodina aumenta la concentración citosólica de calcio y la contracción empieza en el músculo esquelético (capítulo 9). En el músculo cardiaco, el calcio que se une a la troponina proviene tanto del SR como del espacio extracelular mediante canales de calcio sensibles a voltaje SL (receptores de dihidropiridina). Además, es el calcio que está entrando a la célula por medio de los canales de calcio el que activa la liberación de calcio desde el SR mediante los receptores o canales de rianodina (capítulo 10). En el músculo liso, el calcio puede entrar al citosol desde el líquido extracelular por medio de los canales de calcio sensibles a voltaje en el SL, y desde el SR mediante receptores activados por moléculas emisoras de señales desde vías receptoras de SL. El músculo liso también tiene otros receptores en el SL y el SR para movilización de calcio (capítulo 11). En los tres tipos de músculo, los aumentos del calcio citosólico inician los ciclos de fijaciones con puentes entre actina y miosina. PERIODO DE CONTRACCIÓN La evolución temporal de la contracción muscular es distinta en los músculos esquelético, cardiaco y liso. Las contracciones del músculo esquelético tardan varios milisegundos en ocurrir, las contracciones del músculo cardiaco tardan cientos de milisegundos, mientras que el músculo liso es mucho más lento, y pueden requerirse hasta minutos para que ocurran contracciones. Esta diferencia del tiempo de contracción se debe a la tasa de hidrólisis de ATP que ocurre en la cabeza de miosina. Las tasas rápidas de hidrólisis de ATP por la miosina ATPasa dan lugar a contracciones más rápidas, como en el músculo esquelético. Los músculos con tasas de hidrólisis de ATP más lentas muestran contracciones más lentas, como en el músculo liso. Las vías de emisión de señales que causan aumentos del calcio en el citosol pueden contribuir al retraso entre la señal y la contracción. CAMBIOS DE LA FUERZA DE CONTRACCIÓN La fuerza de contracción muscular puede alterarse en los tres tipos de músculo, pero por medios diferentes. La fosforilación de proteínas da lugar a contracciones más fuertes en los músculos tanto car- CAPÍTULO 8 Perspectiva general de la función muscular diaco (capítulo 10) como liso (capítulo 11), mientras que el músculo esquelético alcanza contracciones más fuertes al reclutar más células musculares o activar células musculares con una frecuencia más alta de activación nerviosa (capítulo 9). SIMILITUDES EN LOS MÚSCULOS ESQUELÉTICO, CARDIACO Y LISO Como se mencionó, la actina y miosina son las proteínas contráctiles involucradas en los ciclos de puentes en los tres tipos de músculo, aunque la distribución anatómica de la actina y la miosina es distinta en el músculo liso, en comparación con los músculos esquelético y cardiaco (no estriada en contraposición con estriada, respectivamente). La cabeza de miosina contiene un sitio de unión para actina. Este ciclo está bloqueado cuando la concentración de calcio es baja, pero está abierto cuando el calcio se une a la troponina (músculos esquelético y cardiaco) o calmodulina (músculo liso). Con la unión de actina y miosina, el sitio de ATPasa que se ubica en la cabeza de miosina puede liberar energía a partir del ATP para permitir el ciclo de los puentes, es decir, deslizamiento de actina a través de la miosina. Los tres tipos de músculo muestran la propiedad de incremento de la fuerza de contracción mediante aumento de la longitud del músculo antes de la contracción (en reposo); este fenómeno se llama relación longitud-tensión. Puesto que el músculo esquelético está fijo a huesos mediante tendones, las variaciones de la longitud de la célula en reposo son muy limitadas, y el músculo, por lo general, opera al máximo de la relación longitud-tensión. En el corazón, la célula muscular en reposo por lo general no está a la longitud óptima, de modo que hay reserva, es decir, pueden producirse contracciones más fuertes cuando la longitud en reposo es aumentada antes de la contracción. El músculo liso también muestra la relación longitud-tensión, pero otras influencias sobre el músculo pueden tener más importancia que los efectos de la longitud incrementada de la célula. Por ejemplo, en ciertos tipos de músculo liso vascular, cuando la célula es estirada, la célula responde con una magnitud aumentada de contracción. Este fenómeno se llama respuesta miogénica. En el tracto gastrointestinal, esto ocurre con la presencia de alimentos en el estómago y el intestino delgado. Los órganos huecos con funciones especializadas, como la vejiga urinaria y el útero pueden “estirarse”, pero no se estimula la contracción debido a otras influencias sobre la función del músculo. En el cuadro 8-1 se comparan los músculos esquelético, cardiaco y liso. En los tres capítulos posteriores se darán más detalles de los tipos de músculo individuales. RESUMEN DEL CAPÍTULO ■ ■ ■ Hay tres tipos de músculo en el organismo, que se clasifican por sus características morfológicas, función y mecanismos celulares de contracción —esquelético, cardiaco y liso. Todos los tipos de músculo requieren calcio para iniciar la contracción. El SL y el SR tienen funciones especializadas que aumentan el calcio citosólico para la contracción y eliminan calcio para la relajación. 81 CUADRO 8-1 Comparación de los músculos esquelético, cardiaco y liso. Esquelético Cardiaco Liso Aspecto Estriado Estriado No estriado Retículo sarcoplasmático Casi todo Menos Menos Voluntario Sí No No Proteína de unión a calcio Troponina Troponina Calmodulina Fuente de calcio SR SR Y SL SL y SR Inervación Neurona motora SNS; SNP SNS; SNP Duración de la contracción Milisegundos 100 ms 100 ms-minutos Fuerza de la contracción Reclutamiento Fosforilación; longitud-tensión Fosforilación; longitud-tensión Metabolismo Oxidativo, glucolítico Oxidativo Oxidativo Velocidad de reacción de la ATPasa Rápido Menos rápida Lenta SL, sarcolema; SNP, sistema nervioso parasimpático SNS, sistema nervioso simpático; SR, retículo sarcoplasmático. ■ ■ ■ ■ El músculo puede aumentar la fuerza de contracción al incrementar la longitud del músculo antes de la contracción —la relación longitud-tensión. La energía para la contracción se libera a partir del ATP por la miosina ATPasa. El músculo esquelético requiere aferencias nerviosas desde una neurona motora para iniciar la contracción (músculo voluntario). Los músculos cardiaco y liso pueden contraerse sin aferencias nerviosas, pero la fuerza de la contracción puede alterarse por aferencias provenientes del SNA —ramas simpática y parasimpática del SNA (músculo involuntario). PREGUNTAS DE ESTUDIO 1. ¿Cuál de las afirmaciones que siguen acerca del músculo es verdadera? A) la fuente de calcio para la contracción del músculo esquelético es sólo calcio que entra a la célula a través de los receptores de dihidropiridina B) la fuente de calcio para la contracción del músculo liso es sólo calcio que entra a la célula a través de los receptores de dihidropiridina C) la fuente de calcio para la contracción del músculo cardiaco es sólo calcio que entra a la célula a través de los receptores de dihidropiridina 82 SECCIÓN III Fisiología muscular D) la fuente de calcio para la contracción del músculo esquelético es sólo calcio que entra al citosol a través de los receptores de rianodina E) la fuente de calcio para la contracción del músculo cardiaco es sólo calcio que entra al citosol a través de los receptores de rianodina 2. ¿Cuál de las afirmaciones que siguen acerca del músculo es verdadera? A) los músculos tanto liso como cardiaco permanecen parcialmente contraídos en todo momento B) la contracción de los músculos tanto cardiaco como esquelético se inicia por la unión de calcio a la troponina C) los músculos tanto cardiaco como liso deben tener potenciales de acción para iniciar la contracción D) los músculos tanto cardiaco como liso inician la contracción por unión de calcio a troponina 3. ¿Cuál de las afirmaciones siguientes acerca del músculo es verdadera? A) el músculo esquelético puede aumentar la fuerza de la contracción al reclutar más unidades motoras B) el músculo cardiaco puede aumentar la fuerza de la contracción al reclutar más células musculares C) el músculo liso no puede cambiar la fuerza de la contracción D) el músculo cardiaco no puede cambiar la fuerza de la contracción 4. ¿Cuál de las afirmaciones que siguen acerca del músculo es verdadera? A) en los tres tipos de músculo (cardiaco, esquelético y liso) todas las células se contraen como una unidad B) los tres tipos de músculo están inervados por el sistema nervioso autónomo C) en los tres tipos de músculo, el calcio está involucrado en la contracción D) en los tres tipos de músculo, los antagonistas o bloqueadores de la dihidropiridina aumentan la fuerza de la contracción Estructura y función del músculo esquelético Kathleen H. McDonough C A P Í T U L O 9 O B J E T I V O S ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ Describir los procesos que tienen lugar en la unión neuromuscular. Explicar el acoplamiento excitación-contracción en el músculo esquelético. Describir la función de las proteínas que están involucradas en la contracción. Explicar qué sucede durante una contracción isométrica. Comprender qué ocurre durante una contracción isotónica. ¿De qué modo la carga afecta el acortamiento y la velocidad del acortamiento? Explicar cómo la fuerza de contracción de la fibra muscular puede aumentarse por suma y tétanos. Entender la relación longitud-tensión en el músculo esquelético. Explicar la unidad motora. Determinar cómo la fuerza de la contracción muscular total puede incrementarse mediante reclutamiento de unidades motoras. Comprender la relación fuerza-velocidad en el músculo esquelético; definir la base para la Vmáx. Describir los tres tipos distintos de fibras de músculo esquelético, y las bases para sus diferencias. Declarar cuándo se reclutan estas fibras. actina sobre el de miosina. La longitud de la banda A permanece igual (la miosina no se acorta), pero el tamaño de la banda I disminuye conforme se tira de la actina sobre la miosina. El deslizamiento de la actina sobre la miosina, con energía proporcionada por la miosina ATPasa que está ubicada en la cabeza de miosina, es la base molecular de la contracción del músculo esquelético (figura 9-2). El complejo se activa cuando la concentración de calcio en el citosol aumenta y se une al sitio de unión a calcio en la troponina. La troponina tiene tres componentes designados TnT, los cuales la fijan a la tropomiosina, TnI, que inhibe interacciones entre actina y miosina, y TnC, que se une al calcio. Cuando el calcio se une al TnC, hay un cambio conformacional en la posición de la troponina/tropomiosina, se elimina el obstáculo por TnI y tropomiosina, y permite que la actina y la cabeza de miosina interactúen, lo que hidroliza ATP para que suministre la energía para la contracción —deslizamiento de la actina sobre la miosina o paso por ciclos de puente—. Los puentes seguirán pasando por ciclos, es decir, las cabezas de miosina se unirán a sitios adyacentes en la actina y deslizarán la actina más sobre la ESTRUCTURA El músculo esquelético es distintivo debido a su estructura anatómica —estriaciones debidas al patrón regular de sarcómeros que están compuestos del posicionamiento ordenado de las proteínas actina y miosina—. En la figura 9-1 se muestran sarcómeros compuestos de alineaciones paralelas de filamentos gruesos (es decir, miosina) y filamentos delgados (esto es, actina, tropomiosina y troponina). La miosina constituye la banda A. La actina, junto con las otras dos proteínas, tropomiosina y troponina, constituye la banda I (porción del sarcómero donde la actina no se superpone con miosina). Parte del filamento de actina se superpone con el filamento de miosina, lo que permite que la interacción de estas dos proteínas inicie la contracción. El grado de superposición de filamentos gruesos y delgados es importante en la determinación de la cantidad de fuerza que el músculo esquelético y el músculo cardiaco, pueden generar. Las líneas Z representan los bordes del sarcómero y, durante el acortamiento, las líneas Z se acercan más una a otra conforme se tira del filamento de 83 84 SECCIÓN III Fisiología muscular Sarcómero a) Banda I Banda A Zona H b) Línea Z Línea Z Titina Filamento delgado Línea M Filamento grueso FIGURA 91 a) Corte aumentado de un sarcómero dentro de un músculo esquelético, que muestra el patrón de estriaciones debido a la orientación de los filamentos de actina y miosina. b) Dibujo de los componentes de los sarcómeros de una banda Z a otra, que muestra la proteína estructural titina, los filamentos gruesos (miosina) y los filamentos delgados (actina, tropomiosina y troponina). (Reproducida con autorización de Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology, 11th ed. McGraw-Hill, 2008.) Filamento grueso Puente Filamento delgado a) Sitios de unión a actina Sitios de unión a ATP Tropomiosina Troponina Cadenas ligeras Cadenas pesadas Actina Puente Miosina b) FIGURA 92 a) Dibujo del filamento grueso con las cabezas de miosina o puentes extendiéndose desde el filamento grueso. También se muestra la estructura torcida de los filamentos delgados. b) Aumento de la miosina y actina que muestra los tres componentes del filamento delgado —actina, tropomiosina y troponina— y las cadenas pesada y ligera de la miosina. Note los sitios de unión a actina y a ATP en la miosina. Los sitios de unión a actina son bloqueados por la tropomiosina cuando la concentración de calcio en el citosol es baja. Con la unión de calcio a troponina, la tropomiosina es alejada, y el sitio de unión para actina está disponible. La energía para el deslizamiento del filamento de actina a través del filamento de miosina es proporcionada por el ATP hidrolizado por la miosina ATPasa que se ubica en la cabeza de miosina. (Reproducida con autorización de Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology, 11th ed. McGraw-Hill, 2008.) CAPÍTULO 9 Estructura y función del músculo esquelético miosina, hasta que se termina la contracción por eliminación de calcio. El paso por ciclos de puentes da lugar a desarrollo de tensión, o acortamiento, o una combinación de ambos, dependiendo de la carga sobre el músculo. Si la carga es demasiado grande, habrá una contracción isométrica en la cual se desarrolla tensión del músculo, pero no acortamiento del mismo. Si la carga es menor, habrá una contracción isotónica en la cual el músculo se acorta después de que se desarrolla tensión (capítulo 10). Otras proteínas están involucradas en el mantenimiento de la estructura precisa de los sarcómeros. La titina, una proteína estructural grande en la célula de músculo esquelético, se extiende desde la línea Z hasta el centro del sarcómero, y estabiliza la estructura. Otro complejo proteínico grande consta de distrofina y varias glucoproteínas. Este complejo es esencial en la fijación del sarcómero, en particular, actina, al sarcolema (SL) y la matriz extracelular, de nuevo para mantener la estructura de los sarcómeros y la estabilidad de los mismos. El gen que codifica para el complejo de distrofina es grande y está sujeto a mutaciones que dan por resultado trastornos del músculo esquelético que se conocen como distrofia muscular. Un síntoma de esta enfermedad es la debilidad muscular progresiva debido a pérdida de la integridad estructural apropiada de las fibras musculares. La distrofia muscular de 1 2 + – Duchenne es un tipo de distrofia en la cual hay falta completa de la proteína distrofina, lo que da lugar a declinación rápida de la función del músculo esquelético, y muerte temprana. UNIÓN NEUROMUSCULAR Las células o fibras de músculo esquelético, por lo general, se extienden desde un tendón al otro tendón que fija el músculo a los huesos. El músculo esquelético se clasifica como músculo voluntario porque el sistema nervioso central ordena su contracción —el individuo puede controlar los músculos a voluntad—. De este modo, la inervación del músculo esquelético es esencial para la activación de la contracción. Cada fibra es activada por una neurona motora, mientras que una neurona motora puede inervar varias fibras musculares, lo cual forma una unidad motora. Cuando una unidad motora es activada, todas las fibras inervadas por esa neurona motora se contraerán. Las neuronas motoras de la médula espinal o del tallo encefálico, en respuesta a potenciales de acción que viajan por el axón hacia la célula de músculo esquelético, liberan el neurotransmisor acetilcolina (figura 9-3) en la unión neuromuscular. La cantidad de acetilcolina liberada es proporcional a la frecuencia de potenciales de acción. Potencial de acción de neurona motora Vesícula de acetilcolina El Ca2+ entra a canales sensibles a voltaje 8 Potencial de acción propagado en la membrana plasmática muscular Canales de Na+ sensibles a voltaje Liberación de acetilcolina 3 + – + – + + – + – – 9 85 Degradación de acetilcolina 4 + + – La unión a acetilcolina abre canales iónicos + + + + – – – – 5 Entrada de Na+ + + + – – – – – 7 Receptor de acetilcolina Acetilcolinesterasa Placa terminal motora + + 6 + + + – – Inicio del potencial de acción de la fibra muscular Corriente local entre la placa terminal despolarizada y la membrana plasmática muscular adyacente FIGURA 93 La unión neuromuscular es la parte especializada de la célula muscular —placa terminal motora— en la cual la neurona motora libera el neurotransmisor acetilcolina para activar la célula o fibra muscular. Los eventos en la unión se listan en orden cronológico. Note que cada fibra muscular recibe impulsos a partir de sólo una neurona motora, y que todas las fibras que reciben aferencias provenientes de esa neurona motora constituyen la unidad motora y se contraerán en sincronía. (Reproducida con autorización de Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology, 11th ed. McGraw-Hill, 2008.) 86 SECCIÓN III Fisiología muscular La acetilcolina se difunde a través de la hendidura sináptica y se une a un receptor colinérgico —el receptor nicotínico— sobre la membrana de la célula muscular (SL). La parte del SL del músculo que se asocia con la unión neuromuscular se llama placa terminal motora. Dentro de la hendidura está la enzima acetilcolinesterasa, que puede hidrolizar acetilcolina no unida y, así, limita la activación de los receptores nicotínicos de la membrana de la célula muscular. Este receptor es un canal que permite el flujo de sodio y potasio. El movimiento iónico que predomina es sodio que entra a la célula muscular, lo cual origina despolarización parcial de la membrana celular en la hendidura sináptica —un potencial de placa terminal (capítulo 7)—. Puesto que las neuronas motoras sólo causan despolarización de la membrana postsináptica, y no necesariamente un potencial de acción, el cambio es similar a un potencial postsináptico excitador (EPSP) que ocurre en neuronas. ACOPLAMIENTO EXCITACIÓN CONTRACCIÓN La despolarización es conducida al SL fuera de la unión neuromuscular, y si es fuerte inducirá un potencial de acción. El potencial de acción se transmite a lo largo del SL, hacia los túbulos T, que son invaginaciones del SL, las cuales permiten que la membrana celular entre en contacto estrecho con un sistema de membrana intracelular llamado retículo sarcoplasmático (SR, del inglés sarcoplasmic reticulum; figura 9-4). En el músculo esquelético, el túbulo T hace contacto con dos componentes (cisternas o sacos laterales) del SR que forman una tríada. El calcio se libera desde receptores de rianodina que se ubican en las cisternas cuando el túbulo T se despolariza durante un potencial de acción. Los receptores de dihidropiridina (DHP) también se conocen como canales de calcio, en el SL causan un cambio conformacional en los receptores de rianodina, lo que provoca que se abran y permite que el calcio se difunda desde el SR hacia el citosol. La contracción se inicia cuando la concentración de calcio en el citosol alcanza una cifra crítica y se une al TnC. En el músculo esquelético, todo el calcio que se usa para la contracción se libera desde el SR. Para que ocurra relajación, el calcio debe devolverse al SR. El SR consta de componentes longitudinales, así como del componente de cisterna (figura 9-4). La porción longitudinal del SR contiene la enzima calcio ATPasa, que se denomina SERCA. La SERCA tiene una Vmáx alto y, con la energía proveniente del ATP, bombea calcio contra su gradiente de concentración hacia el SR. Proteínas como la calsecuestrina dentro del SR se unen al calcio, y proporcionan una función de almacenamiento en el SR, pero también mantienen una concentración de calcio libre óptima, de modo que el gradiente de calcio para bombear calcio afuera del citosol y de regreso hacia el SR no es excesivo. Este proceso en el cual un potencial de acción lleva calcio aumentado, que da pie a la contracción, se llama acoplamiento excitación-contracción. Debido a la complejidad de la unión neuromuscular pueden ocurrir muchos estados morbosos cuando hay disfunción. Por ejemplo, los gases nerviosos inhiben la acetilcolinesterasa, lo que da por resultado activación continua de los receptores nicotínicos, y activación continua del músculo esquelético. Por último, las células ya no pueden generar potenciales de acción porque las células permanecen despolarizadas, y los canales de ion sodio que en circunstancias normales se abrirían e iniciarían la despolarización, se desactivan. Sarcolema Túbulo T Ca ATPasa Cisterna del retículo sarcoplasmático Receptor de rianodina, canal Receptor de dihidropiridina FIGURA 94 La conexión entre los túbulos T del sarcolema (SL) y las cisternas del retículo sarcoplasmático (SR) es el mecanismo para el acoplamiento del potencial de acción que viaja a lo largo del SL a la liberación de calcio desde el SR. El potencial de acción altera la conexión entre los canales de calcio (receptores de dihidropiridina) en el SL y los receptores de rianodina en el SR, lo que permite abertura de los canales de calcio del SR y liberación de calcio hacia el citosol. Esto va seguido de debilidad muscular y, puesto que el diafragma contiene músculo esquelético, la insuficiencia respiratoria lleva a la muerte. Enfermedades autoinmunitarias, como la miastenia grave, pueden dar lugar a producción de anticuerpos contra el receptor colinérgico nicotínico. La unión de anticuerpos a los receptores da por resultado emisión de señales o comunicación alterada entre la neurona motora y las fibras de músculo esquelético. Las contracciones quedan alteradas, y con el tiempo toda la estructura de la placa terminal motora se deteriora. La degeneración de la neurona motora, que se requiere para iniciar la contracción, da lugar a enfermedades como la esclerosis lateral amiotrófica (ALS o enfermedad de Lou Gehrig). La neurona motora disminuye de tamaño y se degenera, lo que lleva a desnervación de las células musculares y da por resultado alteración de la capacidad del músculo esquelético para contraerse; por último las células musculares se atrofian. Un síntoma temprano de la ALS es la debilidad muscular. FUNCIÓN TIPOS DE CONTRACCIONES La contracción puede ocurrir en dos modalidades: isométrica e isotónica, y combinaciones de ambas. Isométrico, como su nombre lo indica, se refiere a contracciones en las cuales la longitud (“métrico”) del músculo permanece igual (“iso”), pero la tensión o fuerza aumenta. En la figura 9-5 se muestra un esquema del aparato para medir la salida de contracciones isométricas. Una tira de músculo esquelético delgado se suspende entre un transductor de fuerza y una barra no movible. Puesto que el músculo está fijo en ambos extremos, cuando el músculo se estimula, el paso por ciclos de puentes sólo da lugar a desarrollo de tensión (dina/cm) o fuerza (dina). La longitud del músculo no cambia durante la contracción. CAPÍTULO 9 Estructura y función del músculo esquelético Contracciones isométricas 87 Transductor de fuerza Contracciones isotónicas Músculo 100 75 Estimulador Longitud 50 del músculo Estimulador 25 0 contracciones musculares isométricas. No se permite que el músculo se acorte. La tensión pasiva sobre el músculo como una función de la longitud del músculo en reposo se mide con un transductor de fuerza, y a continuación el músculo es estimulado para que se contraiga. Isotónico se refiere a contracciones en las cuales la tensión (tono) permanece igual, pero la longitud cambia. Sin embargo, antes del acortamiento el músculo debe aumentar la tensión o la fuerza para exceder la carga contra la cual se eleva o se contrae; así, la contracción consta de desarrollo de tensión seguida por acortamiento. En la figura 9-6 se muestra el aparato para medir contracciones isotónicas. El cambio de la longitud del músculo (acortamiento) puede medirse después de que el músculo se estimula. Hay dos cargas sobre el músculo: 1) la precarga que establece la longitud del músculo en reposo y 2) la poscarga que el músculo no detecta sino hasta después de que empieza la contracción. En el protocolo, la precarga se añade a la tira de músculo, se establece la longitud pasiva o en reposo, y después la barra horizontal se coloca bajo el músculo, de modo que la adición de más peso, la poscarga, no permite que el músculo se alargue más. Cuando el estimulador excita el músculo, se quita la barra y la contracción muscular hace que el músculo genere fuerza para igualar la poscarga y después acortamiento. En la figura 9-7 se muestra un modelo para la contracción del músculo esquelético. El músculo consta del elemento contráctil (CE; las proteínas contráctiles actina y miosina) y una serie de componentes elásticos. La carga puede considerarse el peso que el músculo debe levantar en una contracción isotónica. Note que antes del acortamiento, el tamaño del CE disminuye, es decir, los puentes están pasando por ciclos y tirando de la actina a través de la miosina, pero el músculo entero no se acorta —el paso por ciclos de puentes genera tensión (figura 9-7, B). Cuando la tensión o fuerza coincide con la carga (poscarga), el resto del paso por ciclos de puentes (contracción) da por resultado acortamiento del músculo (figura 9-7, C). Advierta que la poscarga determina cuánta tensión tendrá que generar el músculo antes del acortamiento. Una carga más pesada requerirá más desarrollo de tensión, y una carga más ligera requerirá menos desarrollo de tensión. Con una carga más pesada y más ten- contracciones isotónicas. La tensión pasiva es establecida por la precarga, y se mide la longitud del músculo. Se coloca una barra bajo el músculo de modo que cuando se añade la poscarga, el músculo no se alarga (no detecta la poscarga). En el momento de la estimulación, se quita la barra y el músculo desarrolla tensión para sólo igualar la poscarga. Durante el resto de la contracción, la tensión permanece constante y el músculo se acorta. Se miden la longitud del músculo y la tasa de acortamiento. Acortamiento FIGURA 95 Preparación de músculo aislado para estudiar FIGURA 96 Preparación de músculo aislado para estudiar B Tensión Transductor de fuerza Carga (precarga) Poscarga B A C CE CE C CE SE Carga SE SE L Tiempo Estimulación L L L = Carga FIGURA 97 Un modelo de músculo esquelético consta del elemento contráctil (CE) constituido por los filamentos grueso y delgado, y el componente elástico en serie (SE) que consiste de los componentes no contráctiles del músculo. La fase A es el músculo en reposo. Usando el modelo para representar una contracción isotónica, después de estimulación, el músculo desarrolla tensión (fase B) y estira el componente elástico en serie, es decir, se desarrolla tensión (para igualar la carga), pero el músculo entero no se acorta. Note que el elemento contráctil se acorta, es decir, los puentes pasan por ciclos y se tiran del filamento de actina sobre la miosina, pero el músculo entero no se está acortando. En el punto C, la tensión desarrollada por el elemento contráctil que estira el componente elástico en serie justo excede la poscarga, y durante el resto de la contracción el paso por ciclos de puentes en realidad acorta todo el músculo. La carga sobre el músculo determina qué tanta tensión tendrá que desarrollar el músculo para acortarse y levantar la carga. (Reproducida con autorización, de Sonnenblick EH: The Myocardial Cell: Structure, Function and Modification. Briller SA, Conn HL (editors). University of Pennsylvania Press, 1966.) 88 SECCIÓN III Fisiología muscular sión por desarrollar, el músculo mostrará menos acortamiento. Con una carga más ligera y menos tensión por desarrollar, el músculo presentará más acortamiento. Cuando la poscarga se grafica en el eje x, y la velocidad de acortamiento se grafica en el eje y, se demuestra una relación inversa —la curva de fuerza-velocidad (figura 9-8). En el lugar donde la curva se interseca con el eje x, no hay acortamiento (velocidad de acortamiento cero) —ésta es una contracción isométrica—, se desarrolla fuerza máxima. Si se disminuye la poscarga —círculo rojo— debe desarrollarse menos fuerza y ocurre algo de acortamiento y, por ende, la velocidad de acortamiento se puede representar. Si la poscarga se vuelve a disminuir, se desarrolla aún menos fuerza y ocurre aún más acortamiento, y la velocidad de acortamiento aumenta. En la intersección del eje y, hay la velocidad de acortamiento máxima —Vmáx—. Note la línea discontinua que conecta la curva al eje y —esto denota que la intersección es una extrapolación de la curva— por lo que es imposible estudiar una contracción en un músculo con carga cero; por ende, Vmáx es un estimado de la velocidad de acortamiento máxima. Otro hecho por notar es que el acortamiento y la velocidad de acortamiento cambian en la misma dirección —los aumentos del acortamiento ocurren con incrementos de la velocidad de acortamiento—. La relación fuerza-velocidad también se comen- Vmáx Velocidad tará en el capítulo 10 con referencia a la contracción del músculo cardiaco. REGULACIÓN DE LA CONTRACCIÓN, LONGITUDTENSIÓN El tipo de contracción, isométrica en contraposición con isotónica, está determinado por las condiciones de carga sobre el músculo. Si no se permite que el músculo se acorte, el desarrollo de tensión es el resultado total del paso por ciclos de puentes que da por resultado contracción isométrica. Por ejemplo, tirar de un objeto inmovible da lugar a una contracción isométrica —el músculo desarrolla tensión, pero no puede acortarse—. La cantidad de fuerza que se genera durante la contracción (tirón) la determina la proporción de calcio que se libera desde el SR. En circunstancias normales, la cantidad de calcio que se libera en respuesta a un potencial de acción es máxima en fibras de músculo esquelético. La longitud (precarga) del músculo antes de la contracción también afecta la fuerza de la contracción. La longitud de las fibras musculares antes de la contracción determina qué tanta superposición habrá entre la actina y la miosina y, así, cuántos puentes pueden formarse. Dado que la energía para la contracción la libera la actividad de la miosina ATPasa, alterar el número de puentes que interactúan altera la cantidad de miosina ATPasa que es activada y, así, la porción de ATP que será hidrolizado para proporcionar energía para la contracción y relajación. Esto tiene un efecto importante sobre la fuerza de la contracción. La longitud del músculo (precarga) afecta la tensión desarrollada, la tensión pasiva y la tensión total (figura 9-9). En el músculo esquelético, la tensión pasiva es baja hasta el punto Po en el cual empieza a aumentar de manera considerable. La tensión total aumenta en función de la longitud del músculo al igual que la tensión activa o desarrollada. La tensión activa es la tensión que se desarrolla durante la contracción por paso por ciclos de Po Total Fuerza o carga FIGURA 98 La curva de fuerza-velocidad es generada a partir del estudio de músculo aislado durante contracciones isotónicas. Para generar una forma típica, la precarga sobre el músculo se mantiene constante, es decir, la longitud en reposo es la misma para cada tensión (contracción) estudiada, pero la poscarga varía. En la intersección del eje x, la mayor poscarga, no hay acortamiento —esto representa una tensión (contracción) isométrica máxima (Po)—. A medida que la carga disminuye —el punto rojo—, debe desarrollarse menos tensión para igualar la poscarga y, por ende, puede ocurrir algo de acortamiento. Con más acortamiento hay mayor velocidad inicial de acortamiento que se grafica en el eje y. Con un decremento adicional de la poscarga hasta el punto rojo, hay aún menos tensión desarrollada, y puede ocurrir aún más acortamiento, de modo que hay una mayor velocidad de acortamiento. El punto verde representa una poscarga aún más ligera y, por ende, una velocidad de acortamiento aún mayor. La curva es extrapolada hasta la intersección del eje y que da la velocidad de acortamiento máxima (Vmáx). Este es un punto teórico porque el músculo no puede estudiarse en condiciones de carga cero. Activa o desarrollada Tensión, dinas/cm Pasiva Po Longitud, mm FIGURA 99 Relación entre la longitud del músculo (establecida por la precarga sobre el músculo aislado) y la tensión que puede medirse. La tensión activa o desarrollada es la diferencia entre la tensión total y la tensión pasiva. Es la tensión que el músculo produce durante la contracción. A la Po el músculo está en la longitud óptima para dar la mayor tensión —la tensión isométrica máxima. 89 CAPÍTULO 9 Estructura y función del músculo esquelético puentes y, por ende, es la diferencia entre la tensión total y la tensión pasiva. La tensión pasiva se debe a las propiedades estructurales del músculo esquelético. El músculo esquelético muestra la relación longitud-tensión, pero en el cuerpo, puesto que la mayor parte del músculo esquelético está fijo al hueso por tendones, la longitud óptima por lo general la establecen las características anatómicas. Contracción tetánica Suma A B C D E Tensión REGULACIÓN DE LA FUERZA DE CONTRACCIÓN EN EL MÚSCULO ESQUELÉTICO RECLUTAMIENTO, SUMA Y TÉTANOS La manera fisiológica de que el músculo esquelético intacto aumente la tensión es por medio de cambios del patrón de estimulación por las neuronas motoras. Reclutamiento espacial se refiere a números aumentados de neuronas motoras que se activan y, por ende, más unidades motoras que se contraen. Reclutamiento temporal se refiere a un número aumentado de potenciales de acción en una neurona motora, lo que afecta la contracción de las fibras musculares dentro de esa unidad motora. Dentro de un músculo, por lo general sólo un pequeño porcentaje de las células o fibras musculares se contraerá en cualquier momento, pero la contracción de cada fibra será máxima. Todas las fibras musculares inervadas por la misma neurona motora se contraerán al mismo tiempo. La fuerza de la contracción de todo el músculo aumenta si más neuronas motoras son activadas y, por ende, más fibras musculares son estimuladas para que se contraigan —reclutamiento espacial—. El orden del reclutamiento de unidades motoras se comentará más adelante con la presentación de los tipos de fibras musculares. El reclutamiento temporal se produce por aumento del número de potenciales de acción en la neurona motora. En la figura 9-10, la curva A muestra la contracción o tirón en respuesta a un estímulo. La activación más rápida (más potenciales de acción por segundo) libera de manera repetitiva acetilcolina para activar receptores nicotínicos, lo que da más potenciales de acción en la membrana muscular. Si dos estímulos están suficientemente separados (p. ej., retraso de 300 ms, como se muestra en B en la figura 9-10), ocurren dos contracciones idénticas separadas. Cuando los dos estímulos están separados alrededor de 40 a 50 ms, la contracción muscular parece ser un tirón, pero la fuerza de la contracción es mayor que la que generó un estímulo único (curva D, figura 9-10); esta respuesta se llama suma. El mecanismo para la suma es que la segunda contracción empieza antes del inicio de la relajación de la primera contracción. Por consiguiente, la contracción total es el desarrollo de tensión medible sin gasto de energía en superar el componente elástico en serie o la resistencia a la contracción por todos los “elementos no contráctiles” presentes en el músculo. Si los estímulos están separados más de 40 a 50 ms (curva C), la primera contracción empieza a desaparecer antes del inicio de la segunda contracción, lo que da un aspecto bifásico a las contracciones. El retraso óptimo entre dos estímulos puede variar en diferentes tipos de músculo esquelético, pero la capacidad para aumentar la fuerza por medio de suma está presente en todo el músculo esquelético. A medida que el retraso entre los estímulos se hace cada vez más pequeño, la contracción se hace cada vez más débil hasta que la contracción “sumada” será idéntica a la contracción iniciada por estímulo único —todas las fibras se Tirón Estímulo único Retraso de 300 ms Retraso de 120 ms Retraso de 40 a 50 ms 60 Retraso estíde mulos/s 1 ms FIGURA 910 Tensión muscular desarrollada durante contracciones efectuadas bajo diferentes patrones de estimulación. (A) Con un estímulo único, ocurre un tirón. (B) Con dos estímulos con 300 ms de separación, ocurren dos tirones idénticos. (C) Cuando el segundo estímulo ocurre antes de la relajación completa desde el primer tirón, la segunda contracción muestra más desarrollo de tensión. (D) Cuando los dos estímulos están separados alrededor de 40 a 50 ms, parece haber sólo una contracción, pero la tensión es 2 a 3 veces mayor que la que se produce con un estímulo. Cuando el músculo es estimulado con una sucesión rápida de estímulos (60 estímulos/s), ocurre tétanos —se desarrolla la mayor cantidad de tensión, y no hay relajación. Durante este patrón de estimulación, la liberación de calcio con cada estímulo sobrepasa los mecanismos de captación de calcio, de modo que el calcio citosólico permanece alto y no ocurre relajación. Dependiendo del tipo de músculo, la tensión permanecerá alta hasta que la estimulación termine, o hasta que se desarrolle fatiga y el músculo ya no pueda mantener la tensión. (E) Cuando los dos estímulos están separados alrededor de 1 ms, la contracción tiene aspecto idéntico a la contracción dada después de un estímulo —el músculo no puede responder al segundo estímulo porque está en el periodo refractario— la membrana de la célula muscular carece de capacidad de respuesta a un estímulo normal. habrán hecho refractarias al segundo estímulo— con respuesta sólo al primer estímulo. Esto por lo general ocurre con estímulos que están separados 1 a 2 ms (E). La tensión máxima que puede desarrollarse ocurre durante las contracciones tetánicas (figura 9-10). La base para este incremento de la tensión es que hay tantos potenciales de acción (p. ej., 60/s) que los mecanismos de liberación de calcio que ocurren con cada potencial de acción sobrepasan los mecanismos de captación de calcio; así, la concentración citosólica de calcio permanece alta de manera continua, y el músculo no se relaja entre estímulos. Los puentes siguen pasando por ciclos, sea hasta que la estimulación cesa y la concentración citosólica de calcio disminuye, o hasta que las células se fatigan. Tanto la suma como el tétanos (contracciones tetánicas) son ejemplos de reclutamiento temporal —las mismas fibras las estimulan, para que se contraigan, las mismas neuronas motoras—, pero la frecuencia de estimulación por la neurona altera la respuesta del músculo. En resumen, la fuerza de contracción de un músculo intacto 90 SECCIÓN III Fisiología muscular constituido por muchas unidades motoras diferentes puede aumentarse al: 1) incrementar el número de neuronas motoras activadas, lo que aumenta el número de unidades motoras que se contraen y 2) incrementar la frecuencia de potenciales de acción de la neurona motora, lo que provoca suma o tétanos de las fibras musculares en la unidad motora. TIPOS DE FIBRA Como se mencionó, la fuerza de la contracción del músculo esquelético se puede aumentar mediante reclutamiento espacial; este último ocurre cuando más neuronas motoras participan en una contracción, lo que “recluta” más unidades motoras, es decir, más fibras musculares para contraerse. Hay tres tipos básicos de fibras musculares en el músculo esquelético —tipo I, tipo IIa y tipo IIb (cuadro 9-1)—. Antes se les llamaban músculo rojo y blanco por el color imbuido por la presencia de mioglobina y muchas mitocondrias en el músculo rojo, y poca mioglobina en el músculo blanco. Puesto que el músculo rojo tiene muchas mitocondrias, cuenta con la capacidad, mediante producción de ATP por fosforilación oxidativa, para sostener contracciones durante periodos prolongados. Los tipos de fibra tienen diferentes diámetros al igual que las neuronas motoras que los inervan. El patrón de reclutamiento espacial está regido por el tamaño de las fibras musculares; las fibras de menor tamaño son reclutadas con mayor facilidad (reclutamiento más temprano), y las fibras de mayor tamaño son las últimas en ser reclutadas. Las fibras tipo I tienen el diámetro de menor tamaño y están inervadas por neuronas motoras que también son las de diámetro más pequeño; esto hace que tanto las neuronas como las fibras sean fáciles de activar. Estas fibras pequeñas que son reclutadas en etapas más tempranas tienen una capacidad oxidativa alta y pueden desempeñar trabajo durante periodos prolongados sin fatigarse. El flujo sanguíneo adecuado y la concentración alta de mitocondrias para metabolismo oxidativo permiten que estas fibras se contraigan durante horas; estas fibras se han llamado de tirón lento porque su actividad de miosina ATPasa es baja; también contienen mioglobina, una proteína que contiene hem, que se une al oxígeno y, por ende, puede servir como una reserva de oxígeno que puede usarse cuando la fosforilación oxidativa ocurre a tasas altas para apoyar tasas de contracción altas. En el pasado estas fibras tipo I se clasificaron como fibras “rojas” porque la concentración alta de mioglobina da color a las fibras musculares. Las fibras tipo II tienen una actividad de miosina ATPasa más alta y, por ende, una velocidad de contracción más rápida. Hay dos subtipos en este grupo —las fibras tipo IIa son fibras con un tirón rápido y capacidad tanto oxidativa como glucolítica, y las fibras tipo IIb son fibras con tirón rápido, pero que dependen, en su mayor parte, de la glucólisis para la producción de ATP—. Las fibras tipo IIb tienen concentraciones altas de las enzimas involucradas en la glucólisis; estas fibras tienen el diámetro más grande y son las últimas en ser reclutadas. Están inervadas por neuronas motoras de gran diámetro que requieren un mayor estímulo para generar un potencial de acción, lo que hace que sean las últimas en ser reclutadas. Tienen más probabilidades de fatigarse que los otros tipos de fibras debido a la dependencia de glucógeno como un sustrato para ATP para proporcionar la energía para la contracción. El aporte de glucógeno es CUADRO 9-1 Comparación de los tipos de fibra (célula) del músculo esquelético. Tipo I Tipo IIa Tipo IIb Metabolismo Oxidativo Oxidativo/glucolítico Glucolítico Tirón Lento Intermedio Rápido Mitocondrias Abundantes Intermedias Pocas Mioglobina Abundante Abundante Pocas Color Rojo Rojo Blanco Glucógeno Poco Intermedio Abundante Tasa de hidrólisis de ATP por la miosina ATPasa La más baja La más rápida La más rápida Rapidez de contracción La más lenta Intermedia La más rápida Flujo sanguíneo Grande Intermedio Bajo Fatiga No con facilidad Intermedia De inicio rápido Fuerza La menor Intermedia La mayor Tamaño de la neurona motora El más pequeño Intermedio El más grande Tamaño de la fibra El más pequeño Intermedio El más grande Reclutamiento Primero Segundo Al final Tensión total La menor Intermedia La más grande CAPÍTULO 9 Estructura y función del músculo esquelético limitado, y puesto que tienen un riego sanguíneo escaso, la glucosa puede no estar disponible con facilidad. Si la glucólisis da por resultado producción de ácido láctico porque no hay oxígeno disponible, las células se fatigarán en cuestión de minutos y disminuirán el desarrollo de tensión a pesar de activación repetida de neurona motora. Estas fibras antes se clasificaban como fibras “blancas” porque tenían menos mitocondrias, mioglobina y flujo sanguíneo —todo lo cual tiende a conferir el color rojo a las fibras tipo I—. Las fibras tipo IIa son de tamaño intermedio y, por ende, son reclutadas después de que las fibras lentas tipo I son activadas. Las fibras IIa pueden usar tanto glucólisis como fosforilación oxidativa para su aporte de energía y, por consiguiente, también muestran un tiempo intermedio para que ocurra fatiga. La actividad de ATPasa y, en consecuencia, la contracción, son rápidas como en las fibras tipo IIb, y el tiempo para que ocurra fatiga es intermedio. La capacidad de fosforilación oxidativa para proporcionar algo del ATP para la contracción prolonga el tiempo de contracción sostenida antes de que haya fatiga. En general, el diámetro de la fibra muscular es indicativo de la cantidad de actina y miosina en la fibra. Por tanto, las fibras de mayor diámetro tienen más actina y miosina, más puentes que pueden formar, y pueden desarrollar más tensión. Las fibras de menor tamaño desarrollan menos tensión debido a cantidades menores de actina y miosina pero, de nuevo, pueden contraerse durante un periodo prolongado debido a su abundante riego sanguíneo y capacidad oxidativa. Casi todos los músculos en el organismo están hechos de combinaciones de los tres tipos de fibras musculares, con un predominio de fibras rápidas o lentas. Los músculos involucrados en el mantenimiento de la postura deben tener capacidad duradera para contraerse y no fatigarse, de modo que éstos tienen una cantidad mayor de las fibras de tirón lento, tipo I. Por supuesto, en el mantenimiento de la postura, no todas las fibras musculares estarán contraídas en cualquier momento, sino que diferentes unidades motoras se encargarán de la contracción de manera cíclica. Los músculos que están involucrados en cambios rápidos, como los movimientos de los ojos, son predominantemente fibras tipo IIb —contracciones rápidas que no pueden sostenerse durante periodos prolongados—. Muchos músculos tienen cantidades intermedias de los distintos tipos de fibra; por ejemplo, las personas que realizan actividades prolongadas, como carreras de resistencia, tienen fibras oxidativas de tirón más lento en los músculos que se usan al correr. Los corredores que hacen sprint tienen más fibras glucolíticas, de tirón rápido, que son mejores para periodos de actividad breves y súbitos, pero no para actividad sostenida. examen físico, el médico nota que todas las variables medidas estuvieron dentro de límites normales, excepto el movimiento del ojo izquierdo. El movimiento lateral estuvo alterado, y ocurrió ptosis (caída del párpado) con los movimientos oculares rápidos y repetidos. El médico sospecha miastenia grave y solicita análisis para confirmar el diagnóstico. La miastenia grave es una enfermedad autoinmune en la cual el sistema inmunitario produce anticuerpos contra el receptor nicotínico. Al inicio, el defecto se manifiesta en unidades motoras pequeñas, sobre todo en los músculos oculares para el movimiento de los ojos. La activación de neuronas motoras rápidas para contracciones musculares rápidas para el movimiento de los ojos lleva a la liberación de menos acetilcolina (la producción es menor que la liberación). En individuos normales, hay receptores adecuados para compensar la cantidad disminuida de acetilcolina liberada. Con la miastenia grave, anticuerpos unidos a los receptores evitan la unión a acetilcolina, lo que lleva a contracciones musculares alteradas. El reposo puede reabastecer las reservas de acetilcolina. Los anticuerpos unidos al receptor nicotínico parecen desencadenar una respuesta inmunitaria y degeneración de la placa terminal motora muscular. Con más producción de anticuerpos, más unidades musculares quedan afectadas y a la postre pueden llevar a debilidad de músculos grandes, incluso función alterada de los músculos respiratorios. Los tratamientos para disminuir la producción de anticuerpos, a menudo exacerbados por la glándula timo, comprenden extirpación del timo y tratamiento con fármacos inmunosupresores, como corticosteroides. También se usan inhibidores de la colinesterasa porque inhiben la enzima que hidroliza la acetilcolina en la unión neuromuscular, lo que mantiene concentraciones más altas de acetilcolina y mayor estimulación de la placa terminal motora. RESUMEN DEL CAPÍTULO ■ ■ ■ CORRELACIÓN CLÍNICA Durante el mes anterior, una mujer de 45 años de edad nota que se ha sentido muy cansada después de trabajar; también percibe que el párpado izquierdo empieza a caer al final del día. En forma gradual advierte que el párpado empieza a caer incluso al final del día laboral, si ese día ha sido en particular estresante. Asimismo, experimenta fatiga cada vez más intensa, pero estos dos problemas desaparecen tras un sueño nocturno reparador. La mujer está preocupada y hace una cita con su médico. En el 91 ■ ■ ■ ■ ■ Las células de músculo esquelético también se llaman fibras. Las células de músculo esquelético tienen una sección especializada del SL que se conoce como la placa terminal motora, donde la neurona motora forma una sinapsis con el músculo. La acetilcolina es el neurotransmisor, y los receptores nicotínicos sobre el SL se unen a la acetilcolina y aumentan el flujo de entrada de sodio, lo que causa despolarización parcial y, por último, un potencial de acción en el SL adyacente. El potencial de acción viaja por las invaginaciones del SL (túbulos T) y, por medio de los receptores de dihidropiridina, hace que los receptores de rianodina se abran y liberen calcio. La troponina se une al calcio y empieza el proceso de contracción. La neurona motora puede inervar más de una fibra de músculo esquelético —la neurona motora y las fibras que inerva se llaman unidad motora—. Todas estas células musculares se contraen al mismo tiempo. La contracción puede ser isométrica o isotónica. En las contracciones isotónicas, la carga determina qué tanta tensión o fuerza debe desarrollar el músculo antes de que pueda ocurrir la fase de acortamiento de la contracción. El músculo esquelético puede aumentar la fuerza de la contracción al reclutar más unidades motoras (reclutamiento espacial). 92 ■ ■ ■ ■ SECCIÓN III Fisiología muscular Las unidades motoras que tienen las neuronas y fibras de menor diámetro son reclutadas con mayor facilidad (primero); son las fibras tipo I que son muy oxidativas y tienen una tasa baja de miosina ATPasa y, por ende, muestran contracción más lenta. Las fibras tipo II tienen una tasa alta de miosina ATPasa y, por consiguiente, contracción más rápida. Las fibras tipo IIa son tanto glucolíticas como oxidativas, y son reclutadas en segundo lugar. Las fibras tipo IIb son glucolíticas, y las últimas en ser reclutadas (las fibras y las neuronas tienen el mayor diámetro). Las fibras tipo II se fatigan con mayor rapidez que las fibras tipo I; las fibras tipo IIb se fatigan más rápido —en algunos minutos después de estimulación repetida. El músculo esquelético también puede aumentar la fuerza de la contracción con activación más rápida de la neurona motora —suma y tétanos. PREGUNTAS DE ESTUDIO 1. ¿Cuál de las afirmaciones que siguen acerca de la contracción del músculo esquelético es correcta? A) La liberación de acetilcolina en la unión neuromuscular inicia un potencial de acción en la placa terminal motora B) La acetilcolina se une a un receptor nicotínico en la membrana postsináptica C) La despolarización en el músculo esquelético se produce por flujo de entrada de calcio a través de canales de calcio sensibles a voltaje (receptores de dihidropiridina) D) La despolarización de la fibra muscular no es esencial para la contracción del músculo esquelético E) La norepinefrina que activa receptores adrenérgicos causa aumento de la fuerza de contracción 2. ¿Cuál de las afirmaciones que siguen acerca de la contracción muscular es verdadera para el músculo esquelético? A) Todas las células tienen potencial de marcapasos B) La fuerza de la contracción se correlaciona con el grado de fosforilación de las cadenas ligeras de miosina C) La fuerza de la contracción aumenta al reclutar más unidades motoras D) Todas las células musculares tienen capacidad oxidativa alta debido a la presencia abundante de mitocondrias y mioglobina 3. ¿Cuál de las afirmaciones que siguen acerca de la contracción muscular es verdadera para el músculo esquelético? A) En el cuerpo, la fuerza de la contracción es alterada fisiológicamente al cambiar la longitud de la célula en reposo desde 25 hasta 100% de la longitud máxima B) La fuerza de la contracción es alterada fisiológicamente por medio de alteración de la frecuencia de activación de neurona motora C) No puede ocurrir tétanos porque el potencial de acción muscular mantiene a la célula refractaria a estímulos que tienen menos de un segundo de separación D) La contracción muscular sólo consta de desarrollo de tensión Estructura y función del músculo cardiaco C A P Í T U L O 10 Kathleen H. McDonough O B J E T I V O S ■ ■ ■ ■ ■ ■ ■ Comprender el proceso de acoplamiento excitación-contracción en el músculo cardiaco, y cómo difiere del que se observa en el músculo esquelético. Definir los efectos de cambios de la longitud de la célula en reposo sobre el desarrollo de tensión muscular, es decir, la relación longitud-tensión. Describir la secuencia de eventos en las contracciones isotónicas —desarrollo de tensión y acortamiento. Explicar los efectos de la poscarga sobre las contracciones isotónicas en el músculo cardiaco. Determinar los efectos de cambios de la longitud de la célula en reposo sobre contracciones isotónicas a diferentes poscargas, esto es, explicar la curva de fuerza-velocidad. Describir los efectos de la contractilidad aumentada sobre la curva de fuerza-velocidad. Explicar los términos precarga, poscarga, contractilidad, fuerza y tensión. conducción presente en el corazón, y lo transmite a través de células especializadas para conducir potenciales de acción con rapidez. Las células de músculo cardiaco tienen uniones intercelulares comunicantes (conexiones comunicantes) a través de las cuales las células comunican información acerca del potencial de membrana —es decir, si una célula se despolariza, las células adyacentes también se despolarizarán debido a la comunicación a través de las uniones intercelulares comunicantes—. Así, todos los miocitos cardiacos en las aurículas se contraen juntos, y después todos los miocitos en el ventrículo hacen lo mismo (capítulo 23). Debido a esta contracción unificada de los ventrículos (o de las aurículas), se dice que el corazón es un sincitio funcional. Puesto que todas las células musculares ventriculares se contraen juntas, el tipo de reclutamiento espacial no existe en el corazón. El corazón depende de otros mecanismos para aumentar la fuerza de contracción. INTRODUCCIÓN El músculo cardiaco, al igual que el esquelético, es estriado debido a la estructura ordenada de los filamentos de actina y miosina y las proteínas accesorias que estabilizan el sarcómero. Al igual que el músculo esquelético tipo I, el músculo cardiaco parece ser de color rojo por el alto contenido de mitocondrias y mioglobina, y por su riego sanguíneo. El corazón usa grandes cantidades de ATP al latir 60 a 100 veces/ min (en condiciones en reposo normales) durante toda la vida del adulto normal, y la fosforilación oxidativa es la principal fuente de ese ATP, de ahí la concentración alta de mioglobina y el contenido mitocondrial grande. Hay estimados de que el fondo común de ATP miocárdico se recambia cada 10 s. El corazón es capaz de usar cualquier sustrato que se le proporcione en la sangre, y la captación depende de la concentración de esos sustratos, como glucosa, piruvato, lactato, ácidos grasos libres y cuerpos cetónicos. En circunstancias normales, la oxidación de ácidos grasos proporciona 60 a 90% del ATP usado por el corazón adulto. Al igual que en el músculo esquelético, el calcio es esencial para la contracción, y lo proporciona el acoplamiento excitación-contracción. Aunque el músculo cardiaco puede contraerse de manera espontánea debido a actividad de marcapaso en el nodo sinoauricular (SA), las células musculares individuales (miocitos) sólo se contraen cuando un potencial de acción lo inicia el sistema de ACOPLAMIENTO EXCITACIÓNCONTRACCIÓN Las células de músculo cardiaco se contraen cuando la concentración de calcio en las células aumenta desde alrededor de 10–7 M (0.1 μM) hasta 10–6 a 10–5 M (1 a 10 μM). La concentración de calcio presente en el citosol para iniciar la contracción tiene un profundo 93 94 SECCIÓN III Fisiología muscular efecto sobre la fuerza de la contracción (contractilidad). El acoplamiento excitación-contracción en el músculo cardiaco varía un poco del proceso que se observa en el músculo esquelético. Las características anatómicas de la interacción sarcolema (SL)-retículo sarcoplasmático (SR) son diferentes (se forman díadas en lugar de tríadas como en el músculo esquelético). Hay menos SR en el músculo cardiaco, de modo que el proceso de liberación de calcio depende de la entrada de calcio hacia la célula cardiaca a través de los canales sensibles a voltaje (receptores de dihidropiridina), los cuales se abren cuando el potencial de membrana alcanza aproximadamente –40 mV. Estos canales de calcio también se llaman “lentos” o canales de calcio tipo L porque se abren con más lentitud que los canales de sodio, y permanecen abiertos más tiempo, por lo general alrededor de 200 a 300 ms. Por ende, el potencial de acción en las células ventriculares cardiacas es mucho más prolongado que en el músculo esquelético, en el cual los canales de calcio en realidad no se abren (capítulo 9). La entrada de calcio a través de canales de calcio del SL es esencial para que ocurra contracción. La falta de calcio en el líquido extracelular evitaría que el corazón se contrajera. El proceso de acoplamiento excitación-contracción lo inician las células marcapaso en el nodo SA que generan, de manera espontánea, potenciales de acción (que se conocen como potenciales de acción lentos porque carecen de canales de sodio rápidos, y la despolarización se debe a la entrada de calcio a través de los canales de calcio lentos). Los potenciales de acción son transmitidos por las fibras de conducción auriculares atravesando las válvulas auriculoventriculares y, por último, hacia el sistema de conducción en los ventrículos. Todas las células de músculo ventricular se despolarizan al mismo tiempo debido al flujo de entrada rápido de sodio en favor de su gradiente electroquímico (concentración más alta de sodio fuera de la célula y potencial de membrana negativo en el interior del SL) a través de canales de sodio rápidos del SL. Cuando el potencial de membrana alcanza ~ –40 mV, los canales de calcio lentos se abren, lo que permite que el calcio se difunda en favor de su gradiente de concentración hacia el citosol. Parte de este calcio causa abertura de canales (receptores) de rianodina en el SR, y se difunde calcio hacia afuera del SR en favor de su gradiente de concentración. Parte del calcio que proviene del SL se une a la troponina, como lo hace todo el calcio liberado desde el SR. La unión de calcio a troponina da lugar a un tipo similar de interacción de actina y miosina y paso por ciclos de puentes, a los que ocurren en el músculo esquelético. La relajación sucede cuando la concentración de calcio en el citosol es disminuida por la calcio ATPasa en la parte longitudinal del SR que bombea calcio de regreso hacia el SR. También hay calsecuestrina en el músculo cardiaco, que sirve como un “sumidero” para calcio. Cuando la concentración de calcio se reduce, éste se difunde desde la troponina y las células se relajan. Otras dos proteínas están involucradas en la eliminación de calcio desde la célula cardiaca. Puesto que el calcio entra a la célula con cada potencial de acción, debe haber mecanismos para eliminar calcio, o el contenido de calcio en la célula aumentaría con cada latido cardiaco. El SL contiene una calcio ATPasa, la cual tiene afinidad alta por el calcio y que, por ende, puede bombear calcio hacia afuera de la célula, quizá, incluso durante la diástole. La otra proteína es el intercambiador de sodio-calcio, que opera con base en el gradiente de ion sodio. La concentración de ion sodio es mayor fuera de la célula que en la célula. Por medio del intercambiador, el ion sodio entra a la célula y se elimina ion calcio de la célula. Tres iones sodio entran por cada ion calcio que sale de la célula. La manipulación del gradiente de sodio puede tener efectos importantes sobre la extrusión de calcio desde la célula y, así, afectar la contracción. Dado que la concentración de calcio cambia durante cada potencial de acción, hay cierta evidencia de que los aumentos de la frecuencia cardiaca (un mayor número de potenciales de acción por minuto) pueden incrementar la disponibilidad de calcio para la contracción, lo que aumenta la cantidad de tensión que puede generarse. Este fenómeno se llama fenómeno de la escalera o treppe. Fisiológicamente, la frecuencia cardiaca es alterada por modulación de la tasa de activación del nodo SA por el sistema nervioso autónomo (SNA) y, como se verá más adelante, el componente del sistema nervioso simpático (SNS) del SNA aumenta no sólo la frecuencia cardiaca, sino también la contractilidad. En consecuencia, el papel fisiológico del fenómeno de la escalera es difícil de evaluar independiente de la modulación de la frecuencia cardiaca y la contractilidad del corazón por el SNS. Hay otras dos variaciones de la contracción que ocurren en el músculo cardiaco, que no suceden en el músculo esquelético. La fosforilación de proteínas contráctiles altera la fuerza de la contracción en el corazón. El corazón tiene mucha capacidad de respuesta al SNS —el componente de “lucha o huye” del SNA. Con la activación del SNS, receptores β-adrenérgicos en las células de músculo cardiaco son activados, y un esquema de emisión de señales intracelulares da por resultado la producción de cAMP y activación de proteína cinasa A. A continuación hay fosforilación de proteínas. Varias proteínas involucradas en la contracción son fosforiladas, y su actividad es alterada. Los canales de calcio del SL son fosforilados y permiten que entre más calcio a la célula, y la fuerza de la contracción es aumentada (mejora la contractilidad). Una proteína, conocida como fosfolambán, inhibe la calcio ATPasa del SR; cuando el fosfolambán es fosforilado, ejerce menos inhibición de la ATPasa, de modo que la captación de calcio aumenta. No parece ocurrir fosforilación de los canales de calcio en el músculo esquelético, en el cual la cantidad máxima de calcio se libera durante cada potencial de acción y, por consiguiente, no se puede aumentar. Recuérdese que el músculo esquelético tiene dos cisternas de SR en conjunto con el túbulo T, mientras que el músculo cardiaco sólo tiene una cisterna asociada con el túbulo T. El músculo esquelético no parece tener un fosfolambán funcional, de modo que la actividad de calcio ATPasa siempre opera a su capacidad máxima. El incremento de la cantidad de calcio que entra al citosol es un mecanismo importante para aumentar la fuerza de la contracción (contractilidad); la eliminación de calcio más rápido para relajación es un mecanismo importante cuando la frecuencia cardiaca se incrementa con estimulación por el SNS, y hay menos tiempo durante el ciclo de contracción-relajación. CONTRACCIÓN —LONGITUDTENSIÓN, CONTRACCIONES ISOMÉTRICAS La fuerza de la contracción en el músculo cardiaco puede alterarse con cambios de la longitud inicial o en reposo de las células musculares (precarga) similar al fenómeno en el músculo esquelético. El músculo cardiaco, a diferencia del músculo esquelético, puede tener cambios fisiológicos de la longitud de las células musculares. Por ejemplo, cuando el volumen en el ventrículo al final de la diástole (la Tensión, dinas/cm CAPÍTULO 10 Estructura y función del músculo cardiaco Activa o desarrollada Pasiva-músculo cardiaco Pasiva-músculo esquelético Po Longitud, mm FIGURA 101 La relación longitud-tensión en el músculo cardiaco es un poco diferente de la que se observa en el músculo esquelético —sobre todo debido a la presencia de tensión pasiva a longitudes más cortas. Esto se debe en parte a las diferencias anatómicas en la estructura del músculo esquelético (todas las fibras en paralelo) y el músculo cardiaco (las fibras existen en un patrón tipo tejido de canasta), así como a las propiedades de los componentes no contráctiles en el músculo esquelético en contraposición con el músculo cardiaco. Note que en el músculo esquelético las fibras, por lo general, operan en el punto azul —la longitud en reposo es óptima porque casi todo el músculo esquelético lo sostienen en su sitio los huesos, y la longitud en reposo no puede variar mucho—. En circunstancias normales el músculo cardiaco opera a longitud más baja (punto rojo) que la óptima y, por ende, tiene capacidad de reserva para aumentar el desarrollo de tensión, es decir, tener contracciones más fuertes, cuando se incrementa la longitud en reposo. En el corazón intacto, la longitud en reposo de las células cardiacas la establece el volumen en el ventrículo al final de la diástole (el estado relajado del músculo cardiaco). fase de relajación del ciclo cardiaco) es cambiado, la longitud de la célula muscular se cambia en la misma dirección. El volumen al final de la diástole ventricular aumentado da lugar a incremento de la longitud de la célula muscular ventricular antes del inicio de la contracción. En circunstancias normales el corazón opera a longitud celular o precarga más baja que la máxima (figura 10-1, círculo rojo), mientras que el músculo esquelético por lo general funciona a cargas máximas (círculo azul). Note que las propiedades de tensión pasivas del corazón difieren de las que se observan en el músculo esquelético. El músculo esquelético no aumenta la tensión pasiva sino hasta que la longitud de la célula muscular está cerca a la longitud que da la tensión activa máxima. El músculo cardiaco tiene tensión pasiva, incluso a longitudes de célula bajas; estas diferencias se deben a la disposición anatómica de las células musculares con los componentes no contráctiles en el músculo. El músculo esquelético es más distensible que el músculo cardiaco. En la figura 10-1, los efectos de aumentos de la precarga se muestran por medio de contracciones isométricas, es decir, se desarrolla más tensión por mayor longitud de la célula en reposo. El principio de la relación longitud-tensión, como en el músculo esquelético, es que el cambio de la longitud de la célula y el sarcómero altera el grado de superposición de los filamen- 95 tos de actina y miosina y, por ende, aumenta el potencial para que se formen puentes. Las modificaciones en la longitud en reposo de todo el músculo se asocian con cambios proporcionales en la longitud del sarcómero individual. El desarrollo de tensión máxima ocurre a longitudes del sarcómero de 2.2 a 2.3 μm. A longitudes del sarcómero más cortas, los filamentos delgados que se oponen pueden superponerse entre sí e interferir con la interacción con la miosina. A longitudes de sarcómero largas, la superposición puede ser insuficiente para la formación óptima de puentes. La mayor interacción de puentes lleva a una contracción más fuerte. Otros dos factores pueden contribuir al fenómeno de longitudtensión en el músculo cardiaco. El segundo mecanismo puede ser el resultado de un cambio (dependiente de la longitud) de la sensibilidad de los miofilamentos al calcio. Para una concentración de calcio citosólica similar, un músculo menos estirado desarrolla menos fuerza que una preparación de músculo cardiaco más estirado (más largo); este cambio de la sensibilidad al calcio ocurre sin retraso inmediatamente después de un cambio de longitud. La sensibilidad de las proteínas contráctiles, de manera específica troponina C, parece aumentar a mayores longitudes en reposo. Por último, hay cierta evidencia de que la cantidad de calcio liberada a partir del SR es mayor a longitudes en reposo más largas. Qué tanto estos dos factores contribuyen al mayor desarrollo de tensión está abierto a especulación, porque los estudios para demostrar estos dos efectos de la longitud sobre la dinámica del calcio por lo general se realizan en células u orgánulos aislados. En resumen, el corazón, por lo general opera a precargas más bajas que la máxima y, por ende, tiene reserva —el aumento de la longitud del músculo puede tener un profundo efecto sobre la fuerza de contracción que permite al corazón satisfacer las demandas de trabajo aumentado, como sucede durante el ejercicio. CONTRACCIÓN —FUERZA, VELOCIDAD, CONTRACCIONES ISOTÓNICAS Los efectos de la precarga alterada sobre la función del corazón también pueden observarse con contracciones isotónicas que representan una mejor coincidencia con las contracciones fisiológicas del corazón como una bomba. El ventrículo izquierdo debe desarrollar tensión (presión) para igualar la poscarga (presión aórtica) a fin de abrir la válvula aórtica y después permitir que la fase de acortamiento de la contracción bombee sangre (volumen sistólico) hacia la aorta. Recuerde que en la exposición sobre el músculo esquelético se señaló que hay una relación inversa entre la poscarga y la velocidad de acortamiento y, por ende, entre la poscarga y el acortamiento. La mayor poscarga da lugar a menos acortamiento. Al usar contracciones isotónicas, pueden analizarse los efectos de la precarga aumentada, es decir, más paso por ciclos de puentes, sobre la curva de fuerza-velocidad. Cuando el acortamiento y la velocidad de acortamiento se miden en función de la poscarga, las poscargas más altas dan por resultado menos acortamiento (figuras 9-8 y 10-2, curva negra). Si la precarga se aumenta desde la longitud 1 (L1) hasta la longitud 2 (L2), y se estudian las mismas contracciones sujetas a poscarga desde la precarga más alta, la velocidad de acortamiento (y el acortamiento) es mayor para cada poscarga. Si más puentes pueden interactuar, hay más actividad de miosina ATPasa y, por ende, más 96 SECCIÓN III Fisiología muscular energía que proviene de hidrólisis de ATP disponible para la contracción. Los puentes desarrollan la mayor tensión necesaria para igualar la poscarga mayor, y más energía está disponible para que ocurra más acortamiento y haya una mayor velocidad de acortamiento (curva azul, etiquetada L2). Note que la tensión isométrica máxima (intersección del eje x) está aumentada, pero la Vmáx no está incrementada, en comparación con la Vmáx en L1. La precarga sólo desvía la tensión máxima, no la velocidad de acortamiento máxima. AUMENTOS DE LA FUERZA DE CONTRACCIÓN EN EL MÚSCULO CARDIACO Como se mencionó, el músculo cardiaco puede aumentar la fuerza de la contracción cuando se incrementa la precarga según se demuestra por la relación longitud-tensión. Otra manera en que el músculo cardiaco puede incrementar la fuerza de la contracción es mediante aumentos del calcio citosólico, lo que da lugar a incremento de la contractilidad. La contractilidad aumenta la velocidad del paso por ciclos de puentes; por ende, incrementar la contractilidad puede alterar el acortamiento y la velocidad de acortamiento. Al comparar la curva de fuerza-velocidad durante la contractilidad aumentada (curva roja en la figura 10-2), es posible notar que la contractilidad creciente hace que toda la curva de fuerza-velocidad se desvíe hacia la derecha —tanto la tensión isométrica máxima (intersección en el eje x) como la Vmáx (intersección extrapolada en el eje y) aumentan—. Tal incremento de la contractilidad debe compararse con la curva que se le genera a la misma precarga (L1), la curva negra. Por lo general, los aumentos de la contractilidad dan por resultado contracciones más rápidas, de modo que los índices de celeridad de desarrollo de tensión o velocidad de acortamiento máxima (Vmáx) se usan para indicar incrementos de la contractilidad. La figura demuestra que hay dos maneras de aumentar la velocidad de acortamiento a la misma poscarga (los tres puntos en la figura); una es al incrementar la precarga (curva azul), y la otra es al aumentar la contractilidad (curva roja). Sin embargo, los mecanismos mediante los cuales las contracciones son más fuertes son diferentes. La superposición más óptima de los filamentos de actina y miosina modera el efecto de la precarga, mientras que más calcio citosólico para inducir el paso por ciclos de puentes más rápido media el efecto de la contractilidad. Los efectos de aumentar la contractilidad también pueden demostrarse al analizar la relación longitud-tensión. En la figura 10-3, la estimulación de nervios simpáticos que van al corazón da lugar a un cambio de la relación longitud-tensión hacia arriba y hacia la izquierda. Esto indica que para cualquier longitud en reposo dada del músculo cardiaco, la tensión que puede desarrollarse es mayor como resultado de estimulación del SNS. El mecanismo es el incremento de Vmáx ↑Contractilidad Velocidad L1 ↑Precarga o longitud -L2 L1 Po Fuerza o carga FIGURA 102 La curva de fuerza-velocidad en el músculo cardiaco puede alterarse por cambios de la longitud de la célula en reposo y por cambios de la contractilidad. L1 representa la longitud de la célula en reposo más corta, y L2 una mayor longitud de la célula en reposo. Al comparar el músculo a la misma poscarga (punto negro en contraposición con punto azul), el músculo puede acortarse más si la contracción empieza desde una precarga o longitud de la célula en reposo (L2) mayor. En ambas contracciones, la poscarga establece la tensión desarrollada. Note que la curva para precarga aumentada interseca el eje x más hacia la derecha —la mayor longitud en reposo permite que haya mayor tensión isométrica máxima (la relación longitud-tensión)—. Si se estudia el músculo en L1, y se administra un fármaco que aumente la contractilidad, toda la curva de fuerza-velocidad se desvía hacia arriba y hacia la derecha desde la curva negra hacia la curva roja —tanto la Po como la Vmáx están aumentadas—. Más calcio da por resultado contracciones más fuertes y una mayor velocidad de contracción, es decir, una mayor velocidad de paso por ciclos de puentes. Al comparar el punto negro con el punto rojo, cuando la contractilidad es mayor, el músculo puede desarrollar la misma tensión para igualar la carga, y hay más capacidad para acortarse y una mayor velocidad de acortamiento. Por ende, los cambios de la Vmáx indican cambios de la contractilidad. Los cambios de la Po pueden producirse por modificaciones de la precarga o por cambios de la contractilidad. CAPÍTULO 10 Estructura y función del músculo cardiaco Tensión desarrollada ↑SNS FIGURA 103 Efectos de cambios de la contractilidad sobre la relación longitud-tensión. Con la estimulación simpática (SNS) del músculo cardiaco, la contractilidad aumenta y la tensión desarrollada es mayor a cada longitud de la célula en reposo. calcio que se produce por la activación de receptores β-adrenérgicos con la producción consiguiente de AMP cíclico y activación de la fosforilación de los canales de calcio del SL por la proteína cinasa A. La importancia de estos dos mecanismos se hará muy obvia en las exposiciones sobre el sistema cardiovascular en la Sección V. RESUMEN DEL CAPÍTULO ■ ■ ■ ■ Los nervios simpáticos modulan la fuerza de la contracción (contractilidad) de las células cardiacas PREGUNTAS DE ESTUDIO Longitud ■ ■ 97 El acoplamiento excitación-contracción en el músculo cardiaco es similar al que se observa en el músculo esquelético, excepto porque debe entrar calcio a la célula del músculo cardiaco a través de los canales de calcio sensibles a voltaje para causar liberación de calcio desde los canales de rianodina del SR. En el músculo cardiaco, la fuerza de la contracción puede alterarse por cambios de la longitud de la célula en reposo (longitud-tensión) y por modificaciones de la contractilidad. Los índices de contractilidad, como la velocidad de acortamiento máxima (Vmáx), son aumentados por agonistas β-adrenérgicos. Puesto que el corazón tiene células con potencial de marcapaso, no se requiere inervación del corazón para que ocurra contracción. Las células ventriculares se contraen al mismo tiempo (un sincitio funcional) debido al sistema de conducción y a las uniones intercelulares comunicantes (conexiones comunicantes) entre las células cardiacas. Los nervios simpáticos y parasimpáticos modulan la frecuencia de latidos intrínseca. 1. ¿Cuál de las afirmaciones que siguen acerca de la contracción del músculo cardiaco es correcta? A) La liberación de acetilcolina en la unión neuromuscular inicia un potencial de acción en la membrana postsináptica B) La acetilcolina se une a un receptor nicotínico sobre la membrana postsináptica C) La despolarización de la fibra muscular no es esencial para la contracción del músculo cardiaco D) La activación de receptores adrenérgicos por norepinefrina causa incremento de la fuerza de contracción 2. ¿Cuál de las afirmaciones que siguen acerca de la contracción muscular es verdadera para el músculo cardiaco? A) Todas las células en el corazón se contraen a su propia frecuencia B) La fuerza de la contracción es independiente del grado de fosforilación de las proteínas celulares C) La fuerza de la contracción es aumentada al reclutar más unidades motoras D) Todas las células musculares tienen una capacidad oxidativa alta debido a la presencia abundante de mitocondrias y mioglobina 3. ¿Cuál de las afirmaciones que siguen acerca de la contracción muscular es verdadera para el músculo cardiaco? A) La fuerza de la contracción es alterada fisiológicamente al cambiar la longitud de la célula en reposo desde alrededor de 25 hasta 100% de la longitud máxima B) La fuerza de la contracción es alterada fisiológicamente al alterar la frecuencia de activación de la neurona motora C) El tétanos ocurre porque el potencial de acción muscular mantiene la célula refractaria a estímulos separados menos de un segundo D) La contracción muscular sólo consta de desarrollo de tensión 4. ¿Cuál de las afirmaciones que siguen acerca de la contracción muscular es verdadera para el músculo cardiaco? A) La Po (tensión isométrica máxima) es alterada tanto por la contractilidad como por la relación longitud-tensión B) En la curva de fuerza-velocidad, la Vmáx es alterada tanto por la contractilidad como por la relación longitud-tensión C) La poscarga determina cuántos puentes pueden interactuar durante una contracción D) La precarga determina el estado de fosforilación de la cadena ligera de miosina Estructura y función del músculo liso C A P Í T U L O 11 Kathleen H. McDonough O B J E T I V O S ■ ■ ■ ■ ■ Explicar el proceso de contracción en el músculo liso y compararlo con los de los músculos esquelético y cardiaco. Describir cómo el músculo liso puede activarse para inducir una contracción o para cambiar la fuerza de la misma. Comprender la relación entre el potencial de membrana del músculo liso vascular, los canales de calcio sensibles a voltaje, y la fuerza de contracción. Definir la diferencia entre músculo liso de múltiples unidades y unitario. Entender los términos que siguen y su función en el músculo liso: calmodulina, cadena ligera de miosina cinasa, y cadena ligera de miosina fosfatasa. INTRODUCCIÓN CONTRACCIÓN El músculo liso constituye las paredes de casi todos los órganos huecos del cuerpo, excepto el corazón. Como tal, la función y el control de la contracción del músculo liso variarán dependiendo del órgano en el cual se ubique y la función de ese órgano o sistema. Por ejemplo, el músculo liso en el tracto gastrointestinal se activará no sólo por estimulación mecánica por la presencia de alimentos en dicho tracto, sino también por sus aferencias nerviosas e influencias hormonales. El músculo liso en el útero mostrará respuesta de manera diferente durante el desarrollo de un embrión/feto que durante el ciclo menstrual normal. Las hormonas y las aferencias nerviosas, incluso, cambiarán las características morfológicas de músculo liso durante el embarazo, y harán que el útero trabaje como una unidad más que como células musculares independientes en el útero no gestante. La actividad de miosina ATPasa en el músculo liso tiene una tasa de hidrólisis de ATP mucho más lenta (10 a 100 veces más baja que la del músculo esquelético); por ende, las contracciones son mucho más lentas y a veces el modo de contracción da lugar a incrementos y decrementos de la fuerza de contracción más que a la relajación completa después de una contracción, como ocurre en los músculos esquelético y cardiaco. El proceso contráctil general es uniforme en todos los tipos de músculo liso. Un aumento del calcio en el citosol da por resultado la unión de calcio a una proteína de unión a calcio, la calmodulina (figura 11-1); este complejo se unirá a, y activará, la cadena ligera de miosina cinasa (MLCK) que, a su vez, fosforila la cadena ligera de miosina que se localiza en la cabeza de la miosina. En el músculo liso, la cadena ligera de miosina debe estar fosforilada para que la actina y miosina formen puentes e inicien el paso por ciclos de puentes o la contracción. La relajación o el desarrollo de tensión disminuido requiere desfosforilación de la cadena ligera de miosina por la cadena ligera de miosina fosfatasa. El equilibrio de la fosforilación y desfosforilación es importante para regular el desarrollo de tensión en el músculo liso porque la cinasa y la fosfatasa siempre están activas. El incremento del calcio citosólico inclina la balanza hacia más actividad de cinasa y, por ende, más desarrollo de tensión. La concentración más baja de calcio inclina la balanza hacia menos cinasa y, por consiguiente, más actividad de fosfatasa y menos desarrollo de tensión. Hay otros mecanismos para aumentar y disminuir la actividad de la cinasa y la fosfatasa. Por ejemplo, la fosforilación de la enzima MLCK reduce su actividad, lo que aminora la fosforilación de la 99 100 SECCIÓN III Fisiología muscular Célula de músculo liso Contracciones MLCK inactiva ↑ Ca2+ Citosólico Ca calmodulina MLCK activa Cadena ligera de miosina fosfatasa ATP Cadena ligera de miosina ~P Paso por ciclos de puentes Cadena ligera de miosina Relajación FIGURA 111 Esquema de los pasos en la contracción del músculo liso. Al igual que en otros tipos de músculo, el calcio inicia la contracción. El calcio se une a la calmodulina que activa la cadena ligera de miosina cinasa para que fosforile la cadena ligera de miosina. En el momento de la fosforilación, la miosina puede interactuar con la actina, lo que da por resultado paso por ciclos de puentes. Las fosfatasas pueden desfosforilar la cadena ligera de miosina, lo que lleva a relajación o menos desarrollo de tensión. El equilibrio de las actividades de cinasa y fosfatasa determina la magnitud del desarrollo de tensión en el músculo liso. La fosforilación tanto de la cinasa como de la fosfatasa lleva a decremento de su actividad —una da lugar a contracción más débil, y la otra, a contracción más fuerte. miosina y da por resultado más relajación. Esto ocurre cuando un receptor específico, el receptor β2-adrenérgico, en el sarcolema (SL) del músculo liso vascular y del músculo liso bronquial se activa, e incrementa la concentración intracelular de cAMP. La activación subsiguiente de la proteína cinasa A fosforila la MLCK y disminuye su actividad. El óxido nítrico causa una relajación similar del músculo liso, aunque la cinasa que fosforila la MLCK es la proteína cinasa G que es activada por el GMP cíclico. La regulación de la fosfatasa también es importante. Por ejemplo, la fosforilación de la cadena ligera de miosina fosfatasa disminuye su actividad, y da lugar a menos desfosforilación y, por tanto, más fosforilación de las cadenas ligeras de miosina y más contracción. La vía de la Ro cinasa lleva a fosforilación de la fosfatasa. Hay varios tipos de regulación de la MLC cinasa y fosfatasa que alteran las propiedades de contracción del músculo liso y son más específicos para la función particular de cada órgano y, por ende, se comentarán en las secciones específicas para cada órgano de este libro. ENERGÍA PARA LA CONTRACCIÓN Y RELAJACIÓN El ATP que se usa en la contracción y relajación del músculo liso se produce por medio de fosforilación oxidativa. Los sustratos, como la glucosa y los ácidos grasos, se proporcionan en la sangre, y el proceso oxidativo mitocondrial produce energía adecuada para las contracciones más lentas que ocurren en el músculo liso debido a la tasa más baja de enzima miosina ATPasa. Una adaptación interesante del músculo liso asegura que puedan ocurrir contracciones sostenidas a una utilización de ATP más baja que la predicha. El músculo liso puede mantener la tensión por medio de un fenómeno que se llama estado con pestillo. Se cree que esto es importante en los músculos esfínter donde el desarrollo de tensión debe ocurrir durante periodos prolongados que en teoría utilizarían grandes cantidades de ATP. El estado con pestillo parece ocurrir porque los puentes no se disocian con mucha rapidez pese al hecho de que la cadena ligera de miosina es desfosforilada; así, se minimiza el gasto de energía. Se desconoce el mecanismo exacto mediante el cual ocurre el estado con pestillo; sin embargo, la importancia fisiológica es notoria —mantenimiento de tensión con muy poco gasto de energía. VASCULAR EN CONTRAPOSICIÓN CON VISCERAL; MULTIUNITARIO EN CONTRAPOSICIÓN CON UNITARIO El músculo liso puede dividirse en visceral y vascular —el músculo visceral constituye las paredes de casi todos los órganos huecos, y el vascular las paredes de los vasos sanguíneos—. El músculo liso vascular, y hasta cierto grado el músculo liso visceral también pueden dividirse en dos tipos de células —multiunitario y unitarias (cuadro 11-1)—. Estos dos tipos de células musculares tienen caracterís- CUADRO 11-1 Comparación de los tipos de célula de músculo liso. Multiunitario Unitario Funcional Unidades individuales Sincitio Inervación Sí Poca Uniones intercelulares comunicantes Pocas Sí Respuesta al estiramiento Poca Sí Respuesta al SNS Sí Poca Control de la contracción Factores centrales o neurales Factores locales Ejemplos Músculo liso de las vías respiratorias Vasos sanguíneos de pequeño calibre CAPÍTULO 11 Estructura y función del músculo liso 101 Fibra nerviosa del sistema nervioso autónomo Varicosidad Hoja de células Mitocondria Vesículas sinápticas Varicosidades FIGURA 112 Patrón de inervación del músculo liso. Note que el nervio tiene múltiples ramas, y varicosidades en cada una de las ramas. El neurotransmisor se libera en las varicosidades y se difunde hacia el músculo liso. La unión al receptor apropiado da por resultado la modulación neural de la contracción del músculo liso. (Reproducida con autorización de Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology, 11th ed. McGraw-Hill, 2008.) ticas singulares que contribuyen a la variedad de funciones del músculo liso. El músculo liso multiunitario consta de células que actúan como unidades independientes —son inervadas y pueden responder a nervios de los sistemas nerviosos simpático y parasimpático—. Estos tipos de células tienen muy pocas uniones intercelulares comunicantes (conexiones comunicantes) y, por ende, la activación de una célula no lleva por fuerza a la activación de células en yuxtaposición a la célula activada. Otras células que reciben las mismas aferencias nerviosas mostrarán respuesta, pero sólo porque el nervio libera neurotransmisores a partir de varicosidades (figura 11-2) que sueltan al neurotransmisor cerca de la membrana de la célula muscular. Note que el mismo nervio liberará al neurotransmisor hacia muchas células. El neurotransmisor liberado se difunde hacia la membrana de la célula muscular y se une a receptores apropiados —no hay placa terminal motora especializada sobre la membrana de la célula muscular, sino sólo la presencia de receptores—. Los nervios tanto simpáticos como parasimpáticos pueden inervar el mismo músculo liso, y causar efectos opuestos sobre las células (véase más adelante). Por otro lado, el músculo unitario tiene muchas uniones intercelulares comunicantes (capítulo 3), de modo que la activación de una célula lleva con rapidez a la activación de células yuxtapuestas a esa célula. De este modo, las células se contraen como una “unidad”. Estas células, por lo general, tienen poca inervación y muestran una respuesta al estiramiento, o sea, las células aumentarán la tensión en respuesta al estiramiento, una propiedad que se comentará con mayor detalle en las secciones V y VIII. En el cuadro 11-1 se listan las propiedades del músculo liso multiunitario en contraposición con unitario. MÉTODOS DE ESTIMULACIÓN El músculo liso puede estimularse para que se contraiga o para que altere la fuerza de una contracción mediante estímulos diferentes —potenciales de acción, cambios del potencial de membrana que no alcanzan un potencial de acción, activación de receptores que inician una red de emisión de señales intracelular, activación de receptores que son canales iónicos, y el estiramiento, por sí mismo—. Los estímulos exactos que alteran la contracción del músculo liso pueden diferir en diversos órganos, e incluso en dos tipos diferentes de músculo —unitario y multiunitario—. En la figura 11-3 se muestran los efectos de la estimulación nerviosa simpática y parasimpática sobre el potencial de membrana del músculo liso visceral. La acetilcolina, el neurotransmisor del sistema nervioso parasimpático, por lo general hace que el potencial de membrana se torne menos negativo y que ocurran espigas (potenciales de acción) —lo cual genera más actividad contráctil—. La estimulación simpática, por lo general, da por resultado lo opuesto —potencial de membrana más negativo y actividad contráctil disminuida y relajación resultantes—. Advierta que el estiramiento también propicia más espigas de potencial de acción y, después, más contracción; el alimento en el intestino induce actividad contráctil aumentada del intestino. Otros estímulos llevan a la activación de músculo liso o la relajación del mismo. En la figura 11-4 se muestran una célula de músculo liso y algunos de los muchos mecanismos involucrados en el desencadenamiento de contracción y relajación. El calcio puede proporcionarse tanto por flujo de entrada de calcio desde el líquido extracelular a través de los canales de calcio sensibles a voltaje tipo L en el SL, y liberación de calcio desde el retículo sarcoplasmático (SR); sin embargo, el SR, que es menos abundante que en los músculos esquelético y cardiaco, puede liberar calcio por medio de activación de receptores de IP3. Los receptores de IP3 son canales similares a los receptores de rianodina en los otros dos tipos de músculo, y cuando el canal se abre, el calcio se difunde en favor de su gradiente de concentración hacia el citosol para iniciar contracción. El IP3 es un producto de la activación (mediada por receptor) de la fosfolipasa C (PLC) que hidroliza el fosfatidilinositol (PIP2). El diacilglicerol, el otro producto, activa la proteína cinasa C. Hay varios 102 SECCIÓN III Fisiología muscular Acetilcolina, estimulación parasimpática, frío, estiramiento mV 0 –50 Potencial de membrana Epinefrina, estimulación simpática FIGURA 113 Ejemplos de estímulos como el SNS y SNP sobre el potencial de membrana y la generación de potencial de acción del músculo liso. A potenciales de membrana más negativos, como con la estimulación con epinefrina o por el SNS, no hay potenciales de acción, y el músculo se hace más relajado. A potenciales de acción menos negativos, ocurren potenciales de acción y el músculo tiene más probabilidades de tener más tensión o tono. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) receptores diferentes sobre el músculo liso que están enlazados a la vía de PLC, lo que da por resultado contracciones más fuertes. Algunos de éstos incluyen receptores α-adrenérgicos que se unen a la norepinefrina, receptores muscarínicos que se unen a la acetilcolina, y receptores de endotelina específicos que se unen a la endotelina 1. El flujo de entrada de calcio a través de los canales de calcio sensibles a voltaje puede ser modulado por el potencial de membrana en reposo que es una función del movimiento de K+. La abertura de los canales de K (por calcio) o el cierre de los canales de K (por ATP) altera el potencial de membrana. La hiperpolarización de la célula (por abertura de los canales de K) da lugar a cierre de los canales de calcio sensibles a voltaje y relajación o dilatación si la célula es una célula de músculo liso vascular. La despolarización de la célula (que no es suficiente para generar un potencial de acción) causa abertura de los canales de calcio sensibles a voltaje, lo cual lleva a contracción (constricción de músculo liso si la célula es músculo liso vascular). Nótese también la presencia de canales operados por almacenamiento (SOC) en el SL. Estos canales permiten la entrada de calcio hacia el músculo liso para reabastecer las reservas de calcio del SR. No están claros los mecanismos que detectan concentración disminuida de calcio en el SR, y la comunicación entre el SR y el SL, pero parecen ser eficaces para mantener reservas suficientes de calcio en la célula de músculo liso; por último, las células de músculo liso tienen canales operados por receptor. Estos receptores son canales que permiten movimientos iónicos. Los canales purinérgicos representan este tipo de control —el ATP, que es una purina, abre este tipo de canal, lo que permite que entre calcio a la célula y promueve la contracción—. En el músculo liso vascular renal, la adenosina, que se considera un vasodilatador, se une al ROC, lo que permite la entrada de calcio y contracción. En la figura 11-4 no se muestra la modulación de la contracción del músculo liso que ocurre por productos provenientes de las otras células asociadas con el músculo liso; por ejemplo, las células endoteliales que revisten los vasos sanguíneos liberan varios factores que modulan el desarrollo de fuerza en el músculo liso vascular. Como se mencionó, la acetilcolina, que activa receptores muscarínicos en el Célula de músculo liso Receptor PLC PIP2 IP3 Receptor de IP3 SR Liberación de Ca2+ + ROC DAG PKC SOC Proteínas ~P Canal de calcio sensible a voltaje ATP (cierra el canal de K) Ca2+ (abre el canal de K) Canales FIGURA 114 Una célula de músculo liso con algunas de las muchas influencias sobre la contracción. Las contracciones pueden iniciarse por potenciales de acción, por receptores que se acoplan a la fosfolipasa C, y por alteraciones en el estado abierto de los canales de calcio sensibles a voltaje que son sensibles al potencial de membrana según está controlado por movimientos de potasio a través de la membrana. Los SOC son los canales operados por almacenamiento que se abren cuando las reservas de calcio en el SR son bajas. Los ROC son canales operados por receptor —que tiene capacidad de respuesta sobre todo a agentes como la adenosina y el ATP—. La liberación de calcio desde el SR, o la entrada de calcio a través de canales de calcio sensibles a voltaje, lleva a la unión a calmodulina y, por último, a contracción. CAPÍTULO 11 Estructura y función del músculo liso músculo liso vascular, puede causar contracción mediante un mecanismo de PLC. Sin embargo, la unión de acetilcolina a receptores muscarínicos sobre células endoteliales causa la producción de óxido nítrico que se difunde hacia la célula de músculo liso vascular, y activa la guanilato ciclasa para que produzca cGMP. El cGMP activa la proteína cinasa G que fosforila la MLCK y disminuye la contracción. Así, el sitio en el cual la acetilcolina se une al receptor (músculo liso en contraposición con célula endotelial) determina la respuesta. En el organismo, debido a las características anatómicas de las células endoteliales y las células de músculo liso vascular, y la presencia de acetilcolinesterasa, la acetilcolina proveniente de las varicosidades parasimpáticas predominantemente liberaría acetilcolina que se uniría a receptores muscarínicos sobre células endoteliales, lo que daría por resultado relajación o dilatación. Otras respuestas biológicas del músculo liso a la estimulación también son específicas para sitio. Por ejemplo, el músculo liso visceral en el tracto gastrointestinal se hace latente con la estimulación nerviosa simpática (figura 11-3), mientras que el músculo liso vascular en los vasos sanguíneos del tracto gastrointestinal desarrolla contracción más fuerte cuando es estimulado por nervios simpáticos. Esta respuesta específica para sitio se debe al tipo de receptores sobre las células —los receptores β-adrenérgicos causan relajación en respuesta a la estimulación simpática en el músculo liso visceral, mientras que los α-adrenérgicos provocan contracción más fuerte en respuesta a estimulación simpática en el músculo liso vascular. En resumen, el músculo liso es el músculo más diverso en el organismo. Sus funciones dependen, en mayor grado, del tejido en el cual se encuentra; por ende, se presentarán detalles más específicos acerca de la función del músculo liso en las Secciones V y VII a IX. RESUMEN DEL CAPÍTULO ■ ■ ■ ■ ■ ■ En el músculo liso, la proteína de unión a calcio es la calmodulina en lugar de la troponina, como en los músculos esquelético y cardiaco. El complejo de calcio-calmodulina activa la MLCK. La contracción depende de fosforilación de la cadena ligera de miosina por MLCK, lo que permite la unión de la miosina a la actina. La MLCK fosfatasa elimina el fosfato desde la cadena ligera de miosina, lo que da lugar a decremento de la fuerza de contracción. Muchos estímulos diferentes pueden inducir contracción o aumentar la fuerza de la contracción del músculo liso —canales de calcio sensibles a voltaje, canales de potasio sensibles a voltaje, canales operados por receptor, SOC, y acoplamiento farmacomecánico mediado por receptor. Incluso el estiramiento puede activar la contracción del músculo liso. La actividad de miosina ATPasa es más baja en el músculo liso; da por resultado las contracciones, o los cambios del desarrollo de tensión, más lentos. ■ ■ 103 Las células de músculo liso multiunitario están inervadas, tienen pocas uniones intercelulares comunicantes (conexiones comunicantes), y se contraen de manera individual. Las células de músculo liso unitarias pueden mostrar respuesta al estiramiento y tienen uniones intercelulares comunicantes que les permiten contraerse como una “unidad”. PREGUNTAS DE ESTUDIO 1. ¿Cuál de las afirmaciones que siguen acerca del músculo liso es verdadera? A) La fosforilación de cadenas ligeras de miosina se requiere para la contracción B) La inhibición de la cadena ligera de miosina cinasa aumenta la fuerza de la contracción C) La inhibición de la cadena ligera de miosina fosfatasa disminuye la contracción D) La estimulación de las células de músculo liso por óxido nítrico incrementará la contracción 2. ¿Cuál de las afirmaciones que siguen acerca de la contracción del músculo liso es correcta? A) La liberación de acetilcolina en la unión neuromuscular inicia un potencial de acción en la membrana postsináptica B) La acetilcolina se une a un receptor nicotínico en la membrana postsináptica C) La despolarización de la fibra muscular no es esencial para la contracción de músculo liso D) La activación de receptores adrenérgicos por norepinefrina siempre causa aumento de la fuerza de contracción en todas las células de músculo liso 3. ¿Cuál de las afirmaciones que siguen acerca de la contracción muscular es verdadera para el músculo liso? A) Todas las células tienen potencial de marcapasos B) La fuerza de la contracción se correlaciona con el grado de fosforilación de las cadenas ligeras de miosina C) La fuerza de la contracción es aumentada al reclutar más unidades motoras D) Todas las células musculares tienen una velocidad alta de miosina ATPasa y, por ende, una contracción rápida 4. ¿Cuál de las afirmaciones que siguen acerca de la contracción muscular es verdadera para el músculo liso? A) La fuerza de la contracción no puede alterarse de manera fisiológica al cambiar la longitud de la célula en reposo desde 25 hasta 100% de la longitud máxima B) La fuerza de contracción se altera de manera fisiológica al modificar la frecuencia de activación de la neurona motora C) La fuerza de la contracción puede cambiarse al modificar el equilibrio de las actividades de cadena ligera de miosina cinasa y fosfatasa D) La contracción muscular ocurre como contracciones seguidas por relajación completa de la célula SECCIÓN IV FISIOLOGÍA DEL SISTEMA NERVIOSO CENTRAL/NEURAL Introducción al sistema nervioso Susan M. Barman C A P Í T U L O 12 O B J E T I V O S ■ ■ ■ ■ ■ ■ ■ Nombrar los diversos tipos de glía y sus funciones. Nombrar las partes de una neurona y sus funciones. Describir el papel de la mielina en la conducción nerviosa. Listar los tipos de fibras nerviosas que se encuentran en el sistema nervioso de mamíferos. Describir la organización general de neuronas talámicas, corticales y de la formación reticular. Describir la función de las neurotrofinas. Comparar la regeneración nerviosa periférica y central. INTRODUCCIÓN El sistema nervioso puede dividirse en dos partes: el sistema nervioso central (SNC), que está compuesto del cerebro y la médula espinal, y el sistema nervioso periférico, que está compuesto de nervios que conectan el SNC a músculos, glándulas y órganos de los sentidos. Las neuronas son los bloques de construcción básicos del sistema nervioso. El cerebro del ser humano contiene alrededor de 1011 (100 mil millones) de neuronas. También contiene 10 a 50 veces este número de células gliales o glía. El SNC es un órgano complejo; se ha calculado que 40% de los genes del ser humano participa, al menos hasta cierto grado, en su formación. ELEMENTOS CELULARES EN EL SNC CÉLULAS GLIALES Glía es la palabra griega para “pegamento”; durante muchos años se creyó que la glía funcionaba sólo como tejido conjuntivo; sin embar- go, estas células ahora se reconocen por su función en la comunicación dentro del SNC, en asociación con las neuronas. A diferencia de las neuronas, las células gliales todavía pasan por división celular durante su etapa adulta, y su capacidad para proliferar es en particular notoria tras lesión cerebral. Hay dos tipos principales de glía: microglía y macroglía. La microglía son células recolectoras que semejan macrófagos tisulares y eliminan restos producidos por lesión, infección y enfermedad. La microglía surge a partir de macrófagos fuera del SNC, y no se relaciona desde los puntos de vista fisiológico y embrionario con otros tipos de células neurales. Hay tres tipos de macroglía: oligodendrocitos, células de Schwann y astrocitos (figura 12-1). Los oligodendrocitos y las células de Schwann están involucrados en la formación de mielina alrededor de axones en el SNC y el sistema nervioso periférico. Los astrocitos, que se encuentran en todo el cerebro, son de dos subtipos. Los astrocitos fibrosos, los cuales contienen muchos filamentos intermedios, se encuentran sobre todo en la sustancia blanca. Los astrocitos protoplasmáticos se localizan en la sustancia gris y tienen un cito- 105 106 SECCIÓN IV Fisiología del sistema nervioso central/neural A Oligodendrocito Oligodendrocito en la sustancia blanca B Célula de Schwann C Astrocito Oligodendrocitos perineurales Nodos de Ranvier Capilar Neurona Pie terminal Capas de mielina Axones Célula de Schwann Pie terminal Núcleo Lengua interna Astrocito fibroso Axón Neurona FIGURA 121 Principales tipos de células gliales en el sistema nervioso. A) Los oligodendrocitos son pequeños, con pocas prolongaciones. Los que están en la sustancia blanca proporcionan mielina, y los que están en la sustancia gris apoyan las neuronas. B) Las células de Schwann proporcionan mielina al sistema nervioso periférico. Cada célula forma un segmento de vaina de mielina de alrededor de 1 mm de largo; la vaina adopta su forma a medida que la lengua interna de la célula de Schwann gira varias veces alrededor del axón, y lo envuelve en capas concéntricas. Los intervalos entre segmentos de mielina son los nodos de Ranvier. C) Los astrocitos son las células gliales más comunes en el SNC, y se caracterizan por su forma estrellada. Hacen contacto tanto con capilares como con neuronas, y se cree que tienen una función nutritiva. También están involucrados en la formación de la barrera hematoencefálica. (Reproducida con autorización de Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000.) plasma granular. Ambos tipos de astrocitos envían prolongaciones hacia vasos sanguíneos, donde inducen a los capilares para que formen las uniones intercelulares herméticas (zonas de oclusión) que constituyen la barrera hematoencefálica. Esta última barrera evita la difusión de moléculas grandes o hidrofílicas (p. ej., proteínas) hacia el líquido cefalorraquídeo y el cerebro, mientras que permite la difusión de moléculas pequeñas. Los astrocitos también envían prolongaciones que envuelven sinapsis y la superficie de células nerviosas. Los astrocitos protoplasmáticos tienen un potencial de membrana que varía con la concentración externa de K+, pero no generan potenciales propagados. Ayudan a mantener la concentración apropiada de iones y neurotransmisores al captar K+ y los neurotransmisores glutamato y γ-aminobutirato (GABA). NEURONAS Las neuronas en el SNC de mamíferos tienen muchas formas y tamaños. Casi todas presentan las mismas partes de la neurona motora espinal típica (figura 12-2). El cuerpo celular (soma) contiene el núcleo, y es el centro metabólico de la neurona. Las dendritas se extienden hacia afuera desde el cuerpo celular, y tienen extensas arborizaciones. En particular en las cortezas cerebral y cerebelosa, las dendritas tienen pequeñas prolongaciones con abultamientos llamadas espinas dendríticas. Una neurona típica tiene un axón fibroso largo que se origina a partir de un área engrosada del cuerpo celular, el montículo del axón. La primera porción del axón se llama el segmento inicial. El axón se divide en terminales presinápticas, cada una de las cuales termina en varias protuberancias sinápticas que también se conocen como botones terminales o botones. Contienen gránulos o vesículas que almacenan los transmisores sinápticos secretados por los nervios. Con base en el número de prolongaciones que emanan del cuerpo celular, las neuronas pueden clasificarse como unipolares, bipolares y multipolares (figura 12-3). Los axones de muchas neuronas están mielinizados, es decir, adquieren una vaina de mielina, un complejo de proteína-lípido que está envuelto alrededor del axón (figura 12-2). En el sistema nervioso periférico, la mielina se forma cuando una célula de Schwann envuelve el axón con su membrana. Esto puede ocurrir hasta 100 veces, lo que da por resultado muchas capas de mielina alrededor de un axón (figura 12-1). La mielina, a continuación, se compacta cuando las porciones extracelulares de una proteína de membrana, llamada proteína cero (P0), se fijan a las porciones extracelulares de P0 en la membrana que muestra aposición. Diversas mutaciones en el gen que codifica para P0 causan neuropatías periféricas. La vaina de mielina envuelve el axón excepto en su terminación y en los nodos de Ranvier, constricciones de 1 μm periódicas que están separadas alrededor de 1 mm (figura 12-2). La función aislante de la mielina es crucial para la conducción saltatoria de potenciales de acción (capítulo 6). Algunas neuronas tienen axones no mielinizados, es decir, sólo están rodeados por células de Schwann sin la envoltura de la membrana de la célula de Schwann que produce mielina alrededor del axón. Dentro del SNC, las células que forman la mielina son oligodendrocitos (figura 12-1). A diferencia de la célula de Schwann, que forma la mielina en una neurona única, los oligodendrocitos emiten múltiples prolongaciones que forman mielina en muchos axones CAPÍTULO 12 Introducción al sistema nervioso 107 Cuerpo celular (soma) Segmento inicial del axón Nodo de Ranvier Célula de Schwann Montículo del axón Núcleo Botones terminales Dendritas FIGURA 122 Neurona motora con un axón mielinizado. Una neurona motora está compuesta de un cuerpo celular (soma) con un núcleo, varias prolongaciones llamadas dendritas, y un axón fibroso largo que se origina a partir del montículo del axón. La primera porción del axón se llama segmento inicial. Una vaina de mielina se forma a partir de células de Schwann, y rodea el axón, excepto en su terminación y en los nodos de Ranvier. Los botones terminales se ubican en las terminaciones terminales. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) vecinos. En la esclerosis múltiple (MS), una enfermedad autoinmunitaria discapacitante ocurre destrucción de mielina en placas en el SNC. La pérdida de la mielina se asocia con retraso de la conducción o bloqueo de la misma en los axones desmielinizados. SISTEMA NERVIOSO PERIFÉRICO El sistema nervioso periférico transmite información desde el SNC hacia los órganos efectores en todo el cuerpo. Contiene 12 pares de nervios craneales y 31 pares de nervios espinales. Los nervios craneales tienen funciones sensorial y motora bien definidas (cuadro 12-1). Muchas de estas funciones se describen de manera individual con mayor detalle en capítulos posteriores de esta sección. Los nervios espinales reciben su nombre con base en el nivel vertebral a partir del cual sale el nervio (cervicales, torácicos, lumbares, sacros y coccígeos). Estos nervios comprenden fibras motoras y sensitivas de músculos, la piel y glándulas en todo el organismo. TIPOS Y FUNCIÓN DE FIBRAS NERVIOSAS La velocidad de conducción axonal es la rapidez con la cual un potencial de acción viaja a lo largo del axón. En general, hay una relación directa entre el diámetro de una fibra nerviosa dada y su rapidez de conducción. Los neurólogos suelen usar pruebas de conducción nerviosa en el diagnóstico de algunas enfermedades. La velocidad de conducción axonal y otras características han llevado a la clasificación de fibras nerviosas (cuadro 12-2). Las fibras nerviosas de mamífero se dividen en tres grupos principales (A, B y C); el grupo A se subdivide en fibras α, β, γ y δ. En el cuadro 12-2 se listan los diversos tipos de fibras con sus diámetros, características eléctricas y funciones. Los axones grandes se relacionan con la sen- sación propioceptiva, la función motora somática, el tacto consciente y la presión, mientras que los axones de menor tamaño ayudan a las sensaciones de dolor y temperatura, y la función del sistema nervioso autónomo. Las fibras C de la raíz dorsal conducen algunos impulsos generados por receptores de tacto y otros receptores cutáneos, además de impulsos generados por receptores de dolor y de temperatura. También se usa un sistema numérico (Ia, Ib, II, III, IV) para clasificar fibras sensoriales. En el cuadro 12-3 se muestra una comparación del sistema con números y el sistema con letras. Además de variaciones de la rapidez de conducción y el diámetro de la fibra, las diversas clases de fibras en nervios periféricos difieren en su sensibilidad a la hipoxia y a anestésicos (cuadro 12-4). Este hecho tiene importancia clínica, así como fisiológica; por ejemplo, los anestésicos locales deprimen la transmisión en fibras del grupo C antes de que afecten fibras de tacto del grupo A. Por el contrario, la presión sobre un nervio puede causar pérdida de la conducción en fibras motoras, de tacto y de presión de diámetro grande, mientras que la sensación de dolor permanece relativamente intacta. Esto, a veces, se observa en individuos que duermen con los brazos bajo la cabeza durante periodos prolongados, lo que causa compresión de los nervios en los brazos. Debido a la asociación de sueño profundo con intoxicación alcohólica, el síndrome es más común los fines de semana, y ha adquirido el curioso nombre de “parálisis del sábado por la noche” o “del domingo por la mañana”. ORGANIZACIÓN DEL TÁLAMO, LA CORTEZA CEREBRAL Y LA FORMACIÓN RETICULAR El tálamo es un conjunto grande de grupos neuronales dentro del diencéfalo; participa en funciones sensoriales, motoras y límbicas que se describirán en capítulos posteriores de esta sección. Casi toda la información que llega a la corteza cerebral primero la procesa el tálamo, lo que lleva a que se le llame la “compuerta” hacia la corteza. 108 SECCIÓN IV Fisiología del sistema nervioso central/neural A Célula unipolar B Célula bipolar C Célula seudounipolar Dendritas Axón periférico que va hacia la piel y el músculo Dendrita Cuerpo celular Axón Cuerpo celular Prolongación bifurcada única Axón Axón central Cuerpo celular Terminales de axón Neurona de invertebrado Célula bipolar de la retina Célula ganglionar de la raíz dorsal D Tres tipos de células multipolares Dendritas Dendrita apical Cuerpo celular Cuerpo celular Dendrita basal Axón Dendritas Axón Neurona motora de la médula espinal Célula piramidal del hipocampo Cuerpo celular Axón Célula de Purkinje del cerebelo FIGURA 123 Diferentes tipos de neuronas en el sistema nervioso de mamíferos. A) Las neuronas unipolares tienen una prolongación, con diferentes segmentos que sirven como superficies receptivas y terminales liberadoras. B) Las neuronas bipolares tienen dos prolongaciones especializadas: una dendrita que lleva información a la célula, y un axón que transmite información desde la célula. C) Algunas neuronas sensoriales están en una subclase de células bipolares llamadas células seudounipolares. A medida que se desarrolla la célula, una prolongación única se divide en dos, ambas de las cuales funcionan como axones —uno que va hacia la piel o músculo, y otro que va hacia la médula espinal—. D) Las células multipolares tienen un axón y muchas dendritas. Los ejemplos comprenden las neuronas motoras, las células piramidales del hipocampo con dendritas en el vértice y la base, y células de Purkinje cerebelosas con un árbol dendrítico extenso en un plano único. (Adaptada de Ramón Y Cajal: Histology., 10th ed. Baltimore: Wood, 1933.) El tálamo se divide en núcleos que se proyectan de manera difusa hacia regiones amplias de la neocorteza, y núcleos que se proyectan hacia porciones específicas de la neocorteza y el sistema límbico. Los primeros son los núcleos de la línea media e intralaminar. Los segundos comprenden los núcleos de transmisión sensorial específicos, y los núcleos que se relacionan con mecanismos de control eferentes. Los núcleos de transmisión sensorial específicos comprenden los cuerpos geniculados medial y lateral, que transmiten impulsos auditivos y visuales a las cortezas auditiva y visual, y el lateral posterior ventral (VPL) y posteromedial ventral, que transmiten información somatosensorial a la circunvolución central posterior. Los núcleos ventral anterior y ventral lateral se relacionan con la función motora; reciben aferencias provenientes de los ganglios basales y el cerebelo, y se proyectan hacia la corteza motora. Los núcleos anteriores reciben aferencias desde los cuerpos mamilares y se proyectan hacia la corteza límbica (memoria y emoción). Casi todas las neuronas talámicas son excitadoras y liberan glutamato. Las neuronas del núcleo reticular talámico son inhibidoras y liberan GABA; modulan las respuestas de otras neuronas talámicas a aferencias que provienen de la corteza. La neocorteza está dispuesta en seis capas (figura 12-4). El tipo neuronal más común es la célula piramidal con un árbol dendrítico vertical extenso (figura 12-5) que puede extenderse a la superficie cortical. Sus cuerpos celulares pueden encontrarse en todas las capas corticales, excepto en la capa I. Los axones de estas células emiten colaterales recurrentes que giran de regreso, y sinapsis en las porcio- CAPÍTULO 12 Introducción al sistema nervioso 109 CUADRO 12-1 Funciones de los nervios craneales. Nervio craneal Tipo Función I. Olfatorio Sensorial Olfato II. Óptico Sensorial Visión III. Oculomotor Motor Movimientos oculares hacia arriba, hacia abajo y mediales; diámetro de la pupila; forma del cristalino IV. Troclear Motor Movimiento de los ojos hacia abajo y laterales V. Trigémino Motor Sensorial Masticación Propiocepción proveniente de la piel y de los músculos de la cara VI. Abducens Motor Movimientos oculares laterales VII. Facial Motor Sensorial Expresión facial; secreciones de las glándulas salivales Sensación de la piel del conducto auditivo externo; gusto desde los dos tercios anteriores de la lengua VIII. Vestibulococlear Sensorial Audición; sensación de movimiento IX. Glosofaríngeo Motor Sensorial Deglución; secreciones de las glándulas salivales parótidas Gusto desde el tercio posterior de la lengua; barorreceptor y quimiorreceptores X. Vago Motor Músculos esqueléticos de la laringe y la faringe; músculo liso y glándulas en la orofaringe, la laringe, el tórax y el abdomen Receptores en el tórax y el abdomen; gusto proveniente de la parte posterior de la lengua y la cavidad oral Sensorial XI. Accesorio Motor Músculos esqueléticos en el cuello XII. Hipogloso Motor Músculo esquelético de la lengua nes superficiales de los árboles dendríticos. Las fibras aferentes que provienen de los núcleos específicos del tálamo terminan sobre todo en la capa cortical IV, y las aferentes inespecíficas están distribuidas hacia las capas I a IV. Las neuronas piramidales son las únicas neuronas de proyección de la corteza, y son neuronas excitadoras que liberan glutamato. Los otros tipos de células corticales son neuronas de circuito local (interneuronas) que se clasifican con base en su forma, patrón de proyec- ción y neurotransmisor. Las interneuronas inhibidoras (células en cesta o células en candelabro) liberan GABA. Las células en cesta explican casi todas las sinapsis inhibidoras sobre los somas y las dendritas piramidales. Las células en candelabro son una fuente potente de inhibición de neuronas piramidales porque terminan sobre el segmento inicial del axón de la célula piramidal. Sus botones terminales forman hileras verticales cortas que semejan candeleros, lo que explica su nombre. Las células estrelladas espinosas, interneuronas CUADRO 12-2 Clasificación de fibras nerviosas de mamífero.a Tipo de fibra Función Diámetro de la fibra (μm) Velocidad de conducción (ms) Duración de la espiga (ms) Periodo refractario absoluto (ms) A α Propiocepción; motora somática 12–20 70–120 β Tacto, presión 5–12 30–70 γ Motora a husos musculares 3–6 15–30 δ Dolor, frío, tacto 2–5 12–30 Sistema nervioso autónomo preganglionar <3 Raíz dorsal Dolor, temperatura, algo de mecanorrecepción Simpática Simpática posganglionar B 0.4–0.5 0.4–1 3–15 1.2 1.2 0.4–1.2 0.5–2 2 2 0.3–1.3 0.7–2.3 2 2 C aLas fibras A y B son mielinizadas; las fibras C son no mielinizadas. 110 SECCIÓN IV Fisiología del sistema nervioso central/neural CUADRO 12-3 Clasificación numérica a veces usada para fibras sensoriales. Superficie de la piamadre Número Origen Tipo de fibra Ia Huso muscular, terminación anuloespiral Aα Ib Órgano tendinoso de Golgi Aα II Huso muscular, terminación en ramillete de flores; tacto, presión Aβ III Receptores de dolor y frío; algunos receptores de tacto Aδ IV Receptores de dolor, temperatura y otros Raíz dorsal C excitadoras que liberan glutamato, se sitúan en la capa IV, y son un receptor importante de información sensorial que surge del hipotálamo. Además de organizarse en capas, la corteza también está organizada en columnas. Las neuronas dentro de una columna tienen propiedades de respuesta similares, lo que sugiere que comprenden una red de procesamiento local (p. ej., columnas de orientación y de dominancia ocular en la corteza visual; capítulo 15). La formación reticular es una red neuronal compleja en la parte central del bulbo raquídeo y el mesencéfalo; está constituida de diversas agrupaciones neurales y fibras ascendentes y descendentes. Contiene los cuerpos celulares y las fibras de muchos de los sistemas serotonérgicos, noradrenérgicos, adrenérgicos y colinérgicos. Algunas de las fibras descendentes en ella inhiben la transmisión en vías sensoriales y motoras en la médula espinal; diversas áreas reticulares y las vías que vienen de ellas se relacionan con espasticidad (resistencia aumentada al estiramiento pasivo de un músculo) y ajuste de reflejos de estiramiento. El sistema activador reticular (RAS) es una vía polisináptica compleja que surge a partir de la formación reticular del tallo encefálico, con proyecciones hacia los núcleos intralaminar y reticular del tálamo que, a su vez, se proyectan de manera difusa e inespecífica a regiones amplias de la corteza (figura 12-6). Fibras colaterales se canalizan hacia ella no sólo desde los tractos sensoriales ascendentes largos, sino también desde los sistemas trigeminal, auditivo, visual y olfatorio. La complejidad de la red neural y el grado de convergencia en ella suprimen la especificidad de modalidad, y casi todas las neu- CUADRO 12-4 Susceptibilidad relativa de fibras nerviosas A, B y C de mamífero al bloqueo de la conducción producido por diversos agentes. Susceptibilidad a Más susceptible Intermedia Menos susceptible Hipoxia B A C Presión A B C Anestésicos locales C B A I Capa molecular II Capa de células granulares externa III Capa de células piramidales externa IV Capa de células granulares interna V Capa de células piramidales interna VI Capa multiforme Tinción de Nissl Tinción de Golgi Tinción de Weigert Sustancia blanca FIGURA 124 Estructura de la corteza cerebral. Las capas corticales se indican mediante los números. La tinción de Golgi muestra cuerpos y dendritas de célula neuronal, la tinción de Nissl muestra cuerpos celulares, y la tinción de la vaina de miosina, de Weigert, muestra fibras nerviosas mielinizadas. (Modificada con autorización de Ranson SW, Clark SL: The Anatomy of the Nervous System, 10th ed. Saunders, 1959.) ronas reticulares se activan con igual facilidad por estímulos sensoriales diferentes. Por ende, el sistema es inespecífico, mientras que las vías sensoriales clásicas son específicas por cuanto las fibras en ellas son activadas por sólo un tipo de estimulación sensorial. El RAS se relaciona con la conciencia y el sueño (capítulo 20). NEUROTROFINAS: APOYO TRÓFICO DE NEURONAS Las proteínas necesarias para la supervivencia de neuronas y el crecimiento de las mismas se llaman neurotrofinas. Muchas son producto de los músculos u otras estructuras que las neuronas inervan, pero otras las producen los astrocitos. Estas proteínas se unen a receptores en las terminaciones de una neurona. Son internalizadas y después transportadas por medio de transporte retrógrado hacia el cuerpo celular neuronal, donde fomentan la producción de proteínas que se asocian con el desarrollo, el crecimiento y la supervivencia neuronales. Otras neurotrofinas se producen en neuronas y se CAPÍTULO 12 Introducción al sistema nervioso B 111 mantenimiento de las mismas. Se encuentra en muchos tejidos diferentes. El NGF es captado por neuronas y transportado de manera retrógrada desde las terminaciones de las neuronas hacia su cuerpo celular. También está presente en el cerebro, y el crecimiento y el mantenimiento de neuronas colinérgicas en el prosencéfalo basal y el cuerpo estriado parecen depender de él. C A D DEGENERACIÓN Y REGENERACIÓN AXONALES B Axón FIGURA 125 Célula piramidal neocortical, que muestra la distribución de neuronas que terminan en ella. A) Aferentes inespecíficos desde la formación reticular y el tálamo; B) colaterales recurrentes de axones de células piramidales; C) fibras comisurales provenientes de sitios de imagen en espejo en el hemisferio contralateral; D) aferentes específicos que provienen de núcleos de transmisión sensorial talámicos. (Modificada con autorización de Chow KL, Leiman AL: The structural and functional organization of the neocortex. Neurosci Res Program Bull 1970;8(2):157-220.) transportan de una manera anterógrada a la terminación nerviosa, donde mantienen la integridad de la neurona postsináptica. La primera neurotrofina que se caracterizó fue el factor de crecimiento nervioso (NGF), una proteína que es necesaria para el crecimiento de neuronas simpáticas y algunas neuronas sensoriales, y el Cuando un nervio motor que va al músculo esquelético se corta y se permite que se degenere, el músculo se hará en extremo sensible a la acetilcolina, el transmisor liberado en la terminación nerviosa. Esta supersensibilidad (o hipersensibilidad) por desnervación de la estructura postsináptica al transmisor previamente secretado por las terminaciones del axón se debe, en gran parte, a la síntesis o activación de más receptores; esa y otras reacciones desencadenadas por daño de un axón se resumen en la figura 12-7. La degeneración ortógrada (degeneración walleriana) y la degeneración retrógrada del muñón del axón hasta el colateral más cercano (colateral de sostenimiento) ocurren por lo regular cuando se corta un nervio. Asimismo, sucede una serie de cambios en el cuerpo celular, incluso un decremento de la sustancia de Nissl (cromatolisis). El daño de nervio periférico con frecuencia es reversible. Si bien el axón se degenerará en posición distal al daño, los elementos conjuntivos del muñón distal a menudo sobreviven. Ocurre brote axonal desde el muñón proximal, y crece hacia la terminación nerviosa. Rama de axón (colateral de sostenimiento) Receptor Degeneración retrógrada Receptor hipersensible Sitio de lesión Corteza X Reacción retrógrada: cromatolisis Núcleos intralaminares del tálamo Brote Degeneración regenerativo ortógrada (walleriana) FIGURA 127 Resumen de cambios que ocurren en una neurona ascendente en el mesencéfalo del ser humano, sus proyecciones en los núcleos intralaminares del tálamo, y las eferencias desde los núcleos intralaminares hacia muchas partes de la corteza cerebral. La activación de estas áreas se muestra con estudios de PET cuando los sujetos cambian desde un estado despierto relajado hacia una tarea que demanda atención. (Reproducida con autorización de Barrett KE, y la estructura que inerva cuando su axón es aplastado o cortado en el punto marcado con la X. La hipersensibilidad de la estructura postsináptica al transmisor previamente secretado por el axón ocurre, en su mayor parte, por la síntesis o activación de más receptores. Hay degeneración tanto ortógrada (walleriana) desde el punto de daño hasta la terminal, y degeneración retrógrada del muñón del axón hasta el colateral más cercano (colateral de sostenimiento). También ocurren cambios en el cuerpo celular, entre ellos cromatolisis. El nervio empieza otra vez a crecer, con proyección de múltiples ramas pequeñas a lo largo de la trayectoria del axón previamente seguida (brote regenerativo). Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s McGraw-Hill Medical, 2009.) Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) Formación reticular del mesencéfalo FIGURA 126 Diagrama que muestra el sistema reticular 112 SECCIÓN IV Fisiología del sistema nervioso central/neural Esto depende de factores promotores del crecimiento secretados por células de Schwann que atraen axones hacia el muñón distal. Moléculas de adhesión de la superfamilia de la inmunoglobulina promueven el crecimiento del axón a lo largo de las membranas celulares y las matrices extracelulares. Moléculas inhibidoras en el perineurio, la vaina de tejido conjuntivo que cubre un fascículo nervioso, aseguran que los axones que se regeneran crezcan en una trayectoria correcta. Los muñones distales desnervados son capaces de aumentar la producción de neurotrofinas que promueven el crecimiento. Una vez que el axón regenerado alcanza su blanco, se forma una nueva conexión funcional. La regeneración permite que haya recuperación considerable, aunque incompleta; por ejemplo, el control motor fino puede quedar alterado de manera permanente porque algunas neuronas motoras son guiadas hacia una fibra muscular inapropiada. La recuperación de nervios periféricos luego de daño sobrepasa, con mucho, la de las vías del SNC. El muñón proximal de un axón dañado en el SNC formará brotes cortos, pero la recuperación del muñón distante es rara, y los axones dañados tienen pocas probabilidades de formar sinapsis nuevas. Esto se debe a que las neuronas del SNC carecen de las sustancias químicas promotoras del crecimiento necesarias para la regeneración. De hecho, la mielina del SNC es un potente inhibidor del crecimiento axonal. Asimismo, después de lesión del SNC, varios eventos —proliferación astrocítica, activación de microglía, formación de tejido cicatrizal, inflamación e invasión de células inmunes— proporcionan un ambiente inapropiado para la regeneración. Así, el tratamiento de lesiones del cerebro y de la médula espinal con frecuencia se enfoca en la rehabilitación, más que en revertir el daño nervioso. Investigadores tratan de identificar maneras de iniciar el crecimiento axonal y mantenerlo, dirigir los axones que se regeneran para que se reconecten con sus neuronas blanco, y reconstituir los circuitos neuronales originales. evocados visuales presenta una respuesta retrasada (conducción lentificada). Se emite el diagnóstico de esclerosis múltiple (MS), una enfermedad autoinmunitaria que afecta a más de tres millones de personas en todo el mundo, por lo general de 20 a 50 años de edad, y que afecta a mujeres alrededor de dos veces más que a varones. En la MS, anticuerpos y leucocitos del sistema inmunitario atacan la mielina, lo que causa inflamación y lesión de la vaina y, por último, de los nervios que rodea. La pérdida de la mielina lleva a escape de K+ a través de los canales sensibles a voltaje, hiperpolarización, y fracaso para conducir potenciales de acción. Los déficit pueden incluir debilidad muscular, fatiga, coordinación disminuida, lenguaje cercenado, visión borrosa o nublada, disfunción de la vejiga urinaria y alteraciones sensoriales. Los síntomas suelen exacerbarse por temperatura corporal o ambiente aumentada. Casi siempre, aparecen episodios transitorios de manera repentina, duran algunas semanas o meses, y después desaparecen poco a poco. Episodios subsiguientes pueden aparecer años más tarde, y finalmente no ocurre recuperación completa. Otros pacientes tienen una forma progresiva de la enfermedad en la cual no hay periodos de remisión. Los diagnósticos de MS, por lo general, se retrasan hasta que hay múltiples episodios con déficits separados en el tiempo y el espacio. Las pruebas de conducción nerviosa permiten detectar conducción lentificada en vías motoras y sensoriales. El análisis del líquido cefalorraquídeo permite detectar la presencia de bandas oligoclonales indicativas de una reacción inmunitaria anormal contra la mielina. La evaluación más definitiva es la MRI para detectar múltiples áreas del cerebro con tejido cicatrizal (esclerótico). Aunque no hay curación para la MS, algunos fármacos, como el β-interferón y los corticosteroides que suprimen la respuesta inmunitaria pueden reducir la gravedad de la enfermedad y lentificar la progresión de la misma. CORRELACIÓN CLÍNICA Una maestra de escuela de 27 años de edad se despierta una mañana con dolor intenso en el ojo izquierdo y visión borrosa (neuritis óptica). Está fuera del país (EUA) en unas vacaciones de verano, y decide esperar hasta que regrese a casa para acudir con su médico. En el transcurso de los dos días siguientes, el dolor y la pérdida visual empeoran. No obstante, para el momento en que regresa del viaje, 10 días más tarde, los síntomas se han abatido lo suficiente como para que ella decida que es innecesario acudir con su médico. Unos ocho meses más tarde presenta un inicio repentino de debilidad en la pierna derecha después de un día difícil en el aula; decide relajarse con un baño en tina, pero esto exacerba los síntomas. El problema progresa con rapidez hasta el grado en que no puede caminar. Tres días más tarde, es atendida por su médico, y le informa también el incidente que ocurrió mientras estuvo de vacaciones en verano. Se le solicita una resonancia magnética (MRI) del cerebro y una prueba de potenciales evocados visuales. Alrededor de una semana más tarde, la paciente nota mejoría importante, pero su médico le notifica que la MRI mostró múltiples lesiones en la sustancia blanca periventricular, y que la prueba de potenciales RESUMEN DEL CAPÍTULO ■ ■ ■ ■ ■ Las células de la glía son abundantes en el SNC. La microglía son células recolectoras. La macroglía comprende oligodendrocitos, células de Schwann y astrocitos. Los dos primeros están involucrados en la formación de mielina. Los astrocitos ayudan a mantener la concentración apropiada de iones y neurotransmisores en el SNC. Las neuronas están compuestas de un cuerpo celular (soma) que es el centro metabólico de la neurona, dendritas que se extienden hacia afuera desde el cuerpo celular y emiten extensas arborizaciones, y un axón fibroso largo que se origina desde un área un poco engrosada del cuerpo celular, el montículo del axón. Los axones de muchas neuronas adquieren una vaina de mielina, un complejo de proteína-lípido que está envuelto alrededor del axón. La mielina es un aislante eficaz, y la despolarización en axones mielinizados salta desde un nodo de Ranvier hacia el siguiente (conducción saltatoria). Las fibras nerviosas se dividen en diferentes categorías con base en su diámetro axonal, velocidad de conducción, y función. Los núcleos talámicos que se proyectan hacia regiones amplias de la neocorteza son los núcleos de la línea media e intralaminar, y los que se proyectan hacia áreas específicas comprenden los núcleos de transmisión sensorial específicos. CAPÍTULO 12 Introducción al sistema nervioso ■ ■ ■ La neocorteza está dispuesta en seis capas; el tipo neuronal más común es la célula piramidal cuyo cuerpo celular se sitúa en todas las capas, excepto en la capa I. Las neurotrofinas son producidas por astrocitos y trasladadas mediante transporte retrógrado hacia el cuerpo de la célula neuronal, donde fomentan la producción de proteínas asociadas con el desarrollo, el crecimiento y la supervivencia neuronales. Después de daño de un nervio periférico, las células de Schwann secretan un factor promotor del crecimiento que atrae el muñón proximal del axón hacia el muñón distal, lo que permite que haya regeneración. En el SNC, la regeneración neural es alterada por factores, como la proliferación astrocítica, formación de tejido cicatrizal, e inflamación. PREGUNTAS DE ESTUDIO 1. La distancia desde un electrodo estimulador hasta el electrodo de registro es de 4.5 cm. Cuando se estimula el axón, el periodo latente es de 1.5 ms. ¿Cuál es la velocidad de conducción del axón? A) 15 ms B) 30 ms C) 40 ms D) 67.5 ms E) Esto no puede determinarse a partir de la información proporcionada. 2. ¿Cuál de las respuestas que siguen tiene la velocidad de conducción más lenta? A) fibras Aα B) fibras Aβ C) fibras Aγ 113 D) fibras B E) fibras C 3. Un varón se queda dormido con un brazo bajo la cabeza. Cuando se despierta, este brazo está paralizado, pero siente hormigueos, y la sensación de dolor aún está intacta. La razón de la pérdida de la función motora sin pérdida de la sensación de dolor es que en los nervios que van a este brazo: A) las fibras A son más susceptibles a la hipoxia que las fibras B. B) las fibras A son más sensibles a la presión que las fibras C. C) las fibras C son más sensibles a la presión que las fibras A. D) el sueño afecta más los nervios motores que los nervios sensitivos. E) los nervios sensitivos están más cerca del hueso que los nervios motores y, por ende, son menos afectados por la presión. 4. El tálamo A) está organizado hacia seis capas. B) no transmite información auditiva y visual a la neocorteza. C) es un componente del sistema activador reticular. D) contiene neuronas que se proyectan de manera difusa en toda la neocorteza. E) es un componente del tallo encefálico. 5. ¿Cuál de las afirmaciones que siguen acerca de la neocorteza es falsa? A) está organizada en seis capas B) el tipo neuronal más común es la célula piramidal C) recibe aferencias directas desde el tálamo D) contiene un grupo de interneuronas inhibidoras llamadas células en cesto E) contiene un grupo de interneuronas excitadoras llamadas células en candelabro Sistemas sensoriales generales: tacto, dolor y temperatura C A P Í T U L O 13 Susan M. Barman O B J E T I V O S ■ ■ ■ ■ ■ Listar los sentidos comunes y sus receptores. Explicar los términos hiperalgesia y alodinia. Exponer la codificación sensorial. Comparar la vía que media las aferencias sensoriales provenientes de los sentidos del tacto, propioceptivo y vibratorio con la que media la información proveniente de receptores de dolor y de termorreceptores. Describir los mecanismos para modular la transmisión en las vías del dolor. INTRODUCCIÓN La información acerca de los ambientes interno y externo activa el sistema nervioso central (SNC) por medio de receptores sensoriales, los cuales son transductores que convierten diversas formas de energía en potenciales de acción en neuronas. Las características de algunos de estos receptores, y la manera en que generan impulsos en neuronas aferentes, se consideraron en el capítulo 5. Los receptores cutáneos para tacto y presión son mecanorreceptores. Los estímulos en potencia perjudiciales, como dolor, calor y frío extremos están mediados por nociceptores. Los quimiorreceptores son estimulados por un cambio de la composición química del ambiente en el cual están situados; éstos incluyen receptores para el gusto y el olfato, así como receptores viscerales, como los que son sensibles a cambios de la concentración plasmática de O2, pH y osmolalidad. Los fotorreceptores son aquellos que están en los bastones y conos en la retina, que muestran respuesta a la luz. Este capítulo se enfocará principalmente en receptores cutáneos y la transmisión en vías somatosensoriales que median el tacto y la propiocepción (vía de la columna dorsal-lemnisco medial), y dolor y temperatura (tracto espinotalámico). RECEPTORES SENSORIALES MECANORRECEPTORES CUTÁNEOS Los receptores sensoriales pueden ser terminaciones dendríticas especializadas de fibras nerviosas aferentes, y a menudo se relacio- nan con células no neurales que los rodean, lo que forma un órgano de sentido. El tacto y la presión son detectados mediante cuatro tipos de mecanorreceptores (figura 13-1). Los corpúsculos de Meissner son dendritas encapsuladas en tejido conjuntivo, y muestran respuesta a cambios de la textura y vibraciones lentas. Las células de Merkel son terminaciones dendríticas expandidas, y muestran respuesta a presión y tacto sostenidos. Los corpúsculos de Ruffini son terminaciones dendríticas agrandadas con cápsulas alargadas, y muestran respuesta a presión sostenida. Los corpúsculos de Pacini constan de terminaciones dendríticas no mielinizadas de una fibra nerviosa sensorial, encapsuladas por láminas concéntricas de tejido conjuntivo que dan al órgano el aspecto de una cebolla perla; estos receptores muestran respuesta a presión profunda y vibración rápida. NOCICEPTORES Y TERMORRECEPTORES Las sensaciones de dolor y temperatura surgen a partir de dendritas no mielinizadas de neuronas sensoriales ubicadas alrededor de folículos pilosos en toda la piel glabra y pilosa, así como en tejido profundo. Los impulsos provenientes de nociceptores (dolor) son transmitidos por medio de dos tipos de fibra. Un sistema comprende fibras Aδ con mielinización delgada que conducen a tasas de 12 a 30 ms. El otro consta de fibras C no mielinizadas que conducen a tasas bajas de 0.5 a 2 ms. Los termorreceptores también abarcan los dos tipos de fibra que siguen: los receptores de frío están en terminaciones dendríticas de fibras Aδ y fibras C, mientras que los receptores de calor están en fibras C. Los nociceptores mecánicos muestran respuesta a 115 116 SECCIÓN IV Fisiología del sistema nervioso central/neural A Modalidad Tacto Receptores Corpúsculo de Meissner Células de Merkel Corpúsculo de Pacini Terminaciones de Ruffini B Localización Campo receptivo C Intensidad y evolución temporal Serie de espigas neurales Estímulo FIGURA 131 Los sistemas sensoriales codifican para cuatro atributos elementales de estímulos: modalidad, ubicación (campo receptivo), intensidad y duración (cronología). A) La mano del humano tiene cuatro tipos de mecanorreceptores; su activación combinada produce la sensación de contacto con un objeto. La activación selectiva de células de Merkel y de terminaciones de Ruffini causa sensación de presión estable; la activación selectiva de corpúsculos de Meissner y de Pacini origina hormigueo y sensación vibratoria. B) La localización de un estímulo es codificada por la distribución espacial de la población de receptores activados. Un receptor sólo se activa cuando se toca la piel cerca de sus terminales sensoriales. Estos campos receptivos de mecanorreceptores (mostrados como áreas rojas en las yemas) difieren de tamaño en respuesta al tacto. Las células de Merkel y los corpúsculos de Meissner proporcionan la localización más precisa puesto que tienen los campos receptivos de menor tamaño y son más sensibles a presión aplicada mediante una sonda pequeña. C) La intensidad del estímulo es señalada por tasas de activación de receptores individuales; la duración del estímulo es señalada por la evolución temporal de la activación. Las series de espigas indican potenciales de acción desencadenados por presión ejercida con una sonda pequeña en el centro de cada campo receptivo. Los corpúsculos de Meissner y de Pacini se adecuan con rapidez; los otros se adaptan de modo lento. (Reproducida con autorización de Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000.) la presión fuerte. Los nociceptores térmicos son activados por temperatura de la piel de más de 45 °C o por frío intenso. Los nociceptores químicamente sensibles muestran respuesta a diversos agentes, como la bradicinina, histamina, acidez alta e irritantes ambientales. Los nociceptores polimodales muestran respuesta a combinaciones de estos estímulos. La International Association for the Study of Pain (IASP) define el dolor como “una experiencia sensorial y emocional desagradable relacionada con daño tisular real o potencial...” esto difiere de la nocicepción, definida por la IASP como la actividad inconsciente inducida por un estímulo perjudicial aplicado a receptores de los sentidos. El dolor suele clasificarse como fisiológico (o agudo) y patológico (o crónico), que incluye los dolores inflamatorio y neuropático. El dolor agudo típicamente tiene un inicio repentino y disminuye durante el proceso de curación. Puede considerarse “dolor bueno” porque sirve como un importante mecanismo protector. El reflejo de retirada es un ejemplo de esta función protectora del dolor (capí- tulo 14). El dolor crónico puede considerarse “dolor malo” porque persiste mucho tiempo después de la recuperación luego de una lesión, y a menudo es resistente a los analgésicos comunes, incluso antiinflamatorios no esteroideos (AINE) y opiáceos. Puede producirse por lesión nerviosa, incluso neuropatía diabética, daño nervioso inducido por toxina, e isquemia. El dolor con frecuencia se acompaña de hiperalgesia, una respuesta exagerada a un estímulo nocivo y alodinia, una sensación de dolor en respuesta a un estímulo inocuo. Un ejemplo de esto es la sensación dolorosa que produce un baño con agua caliente cuando la piel está dañada por quemadura solar. La hiperalgesia y la alodinia significan sensibilidad aumentada de fibras aferentes nociceptivas. En la figura 13-2 se muestra cómo sustancias químicas liberadas en el sitio de lesión pueden activar más nociceptores, lo que lleva a dolor inflamatorio. Las células lesionadas liberan sustancias químicas como K+ que despolariza terminales nerviosas, lo que hace que los nociceptores tengan mayor capacidad de respuesta. Las células lesionadas también liberan bradicinina y CAPÍTULO 13 Sistemas sensoriales generales: tacto, dolor y temperatura 117 Mastocitos CGRP Sustancia P Histamina Neurona de ganglio de la raíz dorsal Bradicinina Lesión 5-HT Prostaglandina K+ CGRP Sustancia P Vaso sanguíneo Médula espinal FIGURA 132 En respuesta a la lesión tisular, los mediadores químicos pueden sensibilizar nociceptores y activarlos. Estos factores contribuyen a la hiperalgesia y la alodinia. La lesión de tejido libera bradicinina y prostaglandinas que sensibilizan o activan nociceptores, que a su vez liberan sustancia P y péptido relacionado con el gen de la calcitonina (CGRP). La sustancia P actúa sobre los mastocitos (células cebadas) y causa desgranulación y liberación de histamina, que activa nociceptores. Causa extravasación de plasma y el CGRP dilata vasos sanguíneos; el edema resultante provoca liberación adicional de bradicinina. La serotonina (5-HT) es liberada a partir de plaquetas y activa nociceptores. (Reproducida con autorización de Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science. McGraw-Hill, 2000.) sustancia P, que pueden sensibilizar más terminales nociceptivas. La histamina es liberada a partir de células cebadas, la serotonina (5-HT) a partir de plaquetas, el péptido relacionado con el gen de la calcitonina (CGRP) a partir de terminales nerviosas, y las prostaglandinas a partir de membranas celulares; todos contribuyen al proceso inflamatorio y activan los nociceptores o los sensibilizan. Algunas sustancias liberadas actúan al liberar otra (p. ej., la bradicinina activa fibras tanto Aδ como C, e incrementa la síntesis de prostaglandinas y la liberación de las mismas). La prostaglandina E2 (un metabolito del ácido araquidónico, que se produce por la acción de la ciclooxigenasa) es liberada a partir de células dañadas, y produce hiperalgesia. Esta es la razón por la cual el ácido acetilsalicílico (aspirina) y otros AINE (inhibidores de la ciclooxigenasa) alivian el dolor. RECEPTORES SENSORIALES EN MÚSCULOS ESQUELÉTICOS Y ARTICULACIONES Los músculos esqueléticos contienen receptores llamados husos musculares y órganos tendinosos de Golgi que son importantes para la propiocepción. Desempeñan funciones importantes en el control motor, y se describen en el capítulo 14. Asimismo, los músculos contienen nociceptores que muestran respuesta a presión y a la liberación de metabolitos durante la isquemia. También las articulaciones de las extremidades contienen mecanorreceptores (corpúsculos de Pacini y de Ruffini) y nociceptores. CODIFICACIÓN SENSORIAL La conversión de un estímulo de receptor en una sensación reconocible se denomina codificación sensorial. Todos los sistemas senso- riales codifican para cuatro atributos elementales de un estímulo: modalidad, ubicación, intensidad y duración, los cuales se muestran para la modalidad de tacto en la figura 13-1. La modalidad es el tipo de energía transmitida por el estímulo. La forma particular de energía a la cual un receptor es más sensible se llama estímulo adecuado. La ubicación es el sitio del cuerpo o el espacio donde se originó el estímulo. Una unidad sensorial es un axón sensorial único y todas sus ramas periféricas; el campo receptivo de una unidad sensorial es la distribución espacial desde la cual un estímulo produce una respuesta en esa unidad (figura 13-1). Uno de los mecanismos más importantes que permiten la localización de un sitio de estímulo es la inhibición lateral. La actividad que surge a partir de neuronas sensoriales cuyos receptores se encuentran en el borde periférico del estímulo es inhibida en comparación con la de las neuronas sensoriales en el centro del estímulo. Así, la inhibición lateral eleva el contraste entre el centro y la periferia de un área estimulada, e incrementa la capacidad del cerebro para localizar una aferencia sensorial. La inhibición lateral subyace la evaluación neurológica llamada la prueba de discriminación de dos puntos, que se usa para probar la integridad del sistema de la columna dorsal (lemnisco medial), la vía central para el tacto y la propiocepción. En este procedimiento, las dos puntas en un par de calibradores son colocadas de manera simultánea sobre la piel, y se determina la distancia mínima entre las dos puntas del calibrador que puede percibirse como puntos de estimulación separados. Esto se denomina el umbral de discriminación de dos puntos, y es una medida de la agudeza táctil. Si la distancia es muy pequeña, cada punta del calibrador está tocando el campo receptivo de sólo una neurona sensorial. Si la distancia entre los puntos de estimulación es menor que este umbral, sólo puede sentirse un punto de estimulación. La magnitud de los umbrales de discriminación de dos puntos varía de un lugar a otro en el cuerpo, y es de menor tamaño donde los receptores de tacto son más abun- 118 SECCIÓN IV Fisiología del sistema nervioso central/neural dantes. Por ejemplo, los puntos de estímulo en la espalda deben estar separados al menos 65 mm antes de que puedan distinguirse como separados, mientras que en las yemas de los dedos, dos estímulos son reconocidos si están separados por apenas dos mm. La intensidad es señalada por la amplitud de la respuesta o frecuencia de generación de potencial de acción. Duración se refiere al tiempo desde el inicio hasta el final de una respuesta en el receptor. Si un estímulo de fuerza constante se aplica a un receptor, la frecuencia de los potenciales de acción en su nervio sensorial declina con el tiempo. Este fenómeno se conoce como adaptación o desensibilización. El grado al cual ocurre adaptación varía de un sentido a otro. Los receptores pueden clasificarse en receptores de adaptación rápida (fásicos) y de adaptación lenta (tónicos). Esto se muestra para diferentes tipos de receptores de tacto en la figura 13-1. A las columnas dorsales Mecanorreceptores Aβ Mecanorreceptores Nociceptores Aδ Receptores de frío I Nociceptores II Termorreceptores III Mecanorreceptores C IV V VI VII VÍAS SOMATOSENSORIALES ASTA DORSAL El asta dorsal en la médula espinal se divide con base en características histológicas en láminas I a VII; I es la más superficial y VII es la de mayor profundidad. La lámina II y parte de la III constituyen la sustancia gelatinosa, el área cerca de la parte superior de cada asta dorsal. Tres tipos de fibras aferentes primarias (con cuerpos celulares en los ganglios de la raíz dorsal) median la sensación cutánea: 1) fibras Aα y Aβ mielinizadas que transmiten impulsos generados por estímulos mecánicos; 2) fibras Aδ mielinizadas pequeñas que transmiten impulsos desde receptores de frío, nociceptores o mecanorreceptores, y 3) fibras C no mielinizadas que transmiten principalmente dolor y temperatura. La figura 13-3 muestra la distribución ordenada de estas fibras en diversas capas del asta dorsal. VÍA DE LA COLUMNA DORSAL En la figura 13-4 se ilustran las principales vías directas hacia la corteza cerebral para el tacto, el sentido vibratorio y la propiocepción (sentido de posición). Las fibras que median estas sensaciones ascienden de manera ipsilateral en las columnas dorsales al bulbo raquídeo, donde hacen sinapsis en los núcleos gracilis y cuneatus. Las neuronas de segundo orden provenientes de estos núcleos cruzan la línea media, ascienden en el lemnisco medial y finalizan en el núcleo lateral posterior ventral (VPL) contralateral y núcleos de transmisión sensorial específicos relacionados del tálamo; ese sistema ascendente se llama columna dorsal o sistema de lemnisco medial. Las fibras dentro de la vía de la columna dorsal están unidas en el tallo encefálico por fibras que median sensación provenientes de la cabeza. El tacto y la propiocepción se transmiten en su mayor parte por los núcleos sensorial principal y mesencefálico del nervio trigémino. Dentro de las columnas dorsales, las fibras que surgen a partir de niveles diferentes de la médula espinal están organizadas de manera somatotópica. Específicamente, las fibras de la parte sacra de la médula espinal tienen ubicación más medial, y las de la parte cervical de la médula espinal, más lateral. Esta disposición continúa en el bulbo raquídeo con representación de la parte baja del cuerpo (p. ej., el pie) en el núcleo gracilis, y representación de la parte alta del cuer- FIGURA 133 Representación esquemática de las terminaciones de los tres tipos de neuronas aferentes primarias en las diversas capas del asta dorsal de la médula espinal. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) po (p. ej., dedo de la mano) en el núcleo cuneatus. El lemnisco medial está organizado en dirección dorsal a ventral, lo que representa el cuello al pie. La organización somatotópica continúa por el tálamo y la corteza. Neuronas talámicas del VPL que transportan información sensorial se proyectan de una manera muy específica a las dos áreas sensoriales somáticas de la corteza (figura 13-5): el área sensorial somática I (SI) en la circunvolución central posterior y el área sensorial somática II (SII) en la pared de la cisura de Silvio. Además, SI se proyecta a SII. SI corresponde a las áreas 3, 2 y 1 de Brodmann. La disposición de proyecciones a SI es tal que las partes del cuerpo están representadas en orden a lo largo de la circunvolución central posterior; las piernas están en la parte superior y la cabeza en el pie de la circunvolución (figura 13-5). En la circunvolución central posterior no sólo existe localización detallada de las fibras que provienen de las diversas partes del cuerpo, sino que además el tamaño del área receptora cortical para impulsos que provienen de una zona particular del cuerpo es proporcional al uso de la parte. Los tamaños relativos de las áreas receptoras corticales se muestran de manera notoria en la figura 13-6, en la cual las proporciones del homúnculo están deformadas para que correspondan al tamaño de las áreas receptoras corticales para cada una. Nótese que las áreas corticales para la sensación proveniente del tronco y la espalda son pequeñas, mientras que las áreas muy grandes están relacionadas con impulsos que provienen de la mano y las partes de la boca relacionadas con el lenguaje. SII está ubicada en la pared superior de la cisura de Silvio, la cisura que separa el lóbulo temporal de los lóbulos frontal y parietal. La cabeza está representada en el extremo inferior de la circunvolución central posterior, y los pies en la parte inferior de la cisura de Silvio. La representación de las partes del cuerpo no es tan completa o detallada como lo es en la circunvolución central posterior. CAPÍTULO 13 Sistemas sensoriales generales: tacto, dolor y temperatura 119 Circunvolución poscentral Axones de neuronas de tercer orden Tálamo Corteza cerebral Tracto del lemnisco medial (axones de neuronas de segundo orden) Médula oblongada (bulbo raquídeo) Fascículo cuneatus (axones de neuronas sensoriales de primer orden) Tracto espinotalámico lateral (axones de neuronas de segundo orden) Receptor de estiramiento articular (propioceptor) Receptor de dolor Médula espinal Fascículo gracilis (axones de neuronas sensoriales de primer orden) Receptor de tacto a) Axones de neuronas de primer orden (que no forman parte del tracto espinotalámico) b) Receptor de temperatura FIGURA 134 Tractos ascendentes que conducen información sensorial de receptores periféricos a la corteza cerebral. a) Vía de la columna dorsal que media el tacto, el sentido vibratorio y la propiocepción. b) Tracto espinotalámico ventrolateral que media el dolor y la temperatura. (Reproducida con autorización de Fox SI: Human Physiology. McGraw-Hill, 2008.) Sl 6 8 9 4 3 1 2 Corteza parietal posterior Tronco Mano Sll Cara Lengua Auditiva al su Vi FIGURA 135 Áreas del cerebro relacionadas con la sensación somática, y algunas de las áreas receptoras corticales para otras modalidades sensoriales en el cerebro humano. Los números son los de las áreas corticales de Brodmann. El área auditiva primaria en realidad está situada en la cisura de Silvio en la parte superior de la circunvolución temporal superior, y por lo normal no es visible en una vista lateral de la corteza. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) TRACTO ESPINOTALÁMICO VENTROLATERAL Las fibras provenientes de nociceptores y termorreceptores hacen sinapsis en neuronas en el asta dorsal (figura 13-3). Los axones provenientes de estas neuronas cruzan la línea media y ascienden en el cuadrante ventrolateral de la médula espinal, donde forman el tracto espinotalámico lateral (figura 13-4). Las fibras dentro de este tracto hacen sinapsis con los núcleos VPL. Otras neuronas del asta dorsal que reciben aferencias nociceptivas hacen sinapsis en la formación reticular del tallo encefálico (vía espinorreticular) y después se proyectan hacia el núcleo centrolateral del tálamo. Estudios con tomografía por emisión de positrones (PET) e imágenes de resonancia magnética funcionales (fMRI) en humanos normales muestran que el dolor activa las áreas corticales SI, SII y la circunvolución cingulada en el lado opuesto del estímulo. Asimismo, la corteza mediofrontal y la corteza insular son activadas. Estas tecnologías fueron importantes para distinguir dos componentes de las vías de dolor. Desde los núcleos VPL en el tálamo, se proyectan 120 SECCIÓN IV Fisiología del sistema nervioso central/neural Brazo Codo razo Anteb eca Muñ o Man ue ñiq Me lar u An dio Me ice d r Ín lga Pu O Na jo r Ca iz ra Lab io su pe Labios Hombro Cabeza Cuello Tronco Cadera Piern a MORFINA Y ENCEFALINAS e Pi ie p del dos itales Gen De rior Labio inferior Dientes, encías y mandíbula Lengua Faringe Intraabdominal FIGURA 136 Homúnculo sensorial, dibujado sobre un corte coronal a través de la circunvolución poscentral. (Reproducida con autorización de Penfield W, Rasmussen G: The Cerebral Cortex of Man. Macmillan, 1950.) fibras a SI y SII. Esto se llama tracto neoespinotalámico, y se encarga de la conciencia inmediata de la sensación dolorosa, y la conciencia de la ubicación del estímulo nocivo. La vía que incluye sinapsis en la formación reticular del tallo encefálico y el núcleo talámico centrolateral se proyecta hacia el lóbulo frontal, el sistema límbico y la ínsula. Esto se conoce como el tracto paleoespinotalámico, y media la respuesta emocional al dolor. En el SNC, la sensación visceral viaja a lo largo de las mismas vías que la sensación somática en los tractos espinotalámicos y las radiaciones talámicas, y las áreas receptoras corticales para la sensación visceral están entremezcladas con las áreas receptoras somáticas. Esto quizá contribuye al fenómeno llamado dolor referido. La irritación de un órgano visceral produce dolor que se siente no en ese sitio, sino en una estructura somática. Se dice que ese dolor es referido a la estructura somática. Tal vez el ejemplo mejor conocido es el dolor cardiaco referido a la cara interna del brazo izquierdo. MODULACIÓN DE LA TRANSMISIÓN DEL DOLOR Muchas personas han aprendido a partir de la experiencia práctica que tocar o sacudir un área lesionada disminuye el dolor debido a la lesión. El alivio suele depender de inhibición de las vías del dolor en la compuerta del asta dorsal por estimulación de fibras aferentes de tacto-presión de diámetro grande. En la figura 13-3 se ilustra que las fibras colaterales provenientes de estas fibras aferentes mielinizadas hacen sinapsis en el asta dorsal. Dichas fibras colaterales pueden modificar las aferencias provenientes de terminales aferentes nociceptivas que también hacen sinapsis en el asta dorsal; esta última recibe el nombre de hipótesis del control de compuerta. La morfina es uno de los agentes analgésicos más eficaces. Los receptores que se unen a la morfina y las morfinas endógenas, los péptidos opioides, se encuentran en el mesencéfalo, el tallo encefálico y la médula espinal. Hay al menos tres sitios en los cuales los opioides pueden actuar para producir analgesia: en la periferia, en el sitio de una lesión; en el asta dorsal, donde fibras nociceptivas hacen sinapsis sobre células de ganglio de la raíz dorsal, y en sitios más rostrales en el tallo encefálico. En la figura 13-7 se muestran diversos modos de acción de opiáceos para disminuir la transmisión en vías de dolor. Los receptores opioides se encuentran situados sobre células de ganglio de la raíz dorsal y sobre fibras de nervios aferentes. En la periferia, la inflamación causa la producción de péptidos opioides por células inmunitarias, y éstos pueden actuar sobre los receptores en las fibras nerviosas aferentes para reducir el dolor que de otro modo se sentiría. Los receptores opioides en la región del asta dorsal quizá actúen de manera presináptica para disminuir la liberación de sustancia P. Las inyecciones de morfina en la sustancia gris periacueductal (PAG) del mesencéfalo alivian el dolor al activar vías descendentes que producen inhibición de la transmisión aferente primaria en el asta dorsal. Esta activación puede ocurrir por medio de proyecciones desde la PAG hacia el núcleo del rafe magno, y fibras serotoninérgicas descendentes provenientes de este núcleo median la inhibición. La acupuntura en un sitio distante del lugar de un dolor tal vez actúe al liberar endorfinas en el cerebro; la acupuntura en el sitio del dolor parece actuar principalmente de la misma manera que tocarse o sacudirse (mecanismo de control de compuerta). EXAMEN NEUROLÓGICO El componente sensorial de un examen neurológico incluye una evaluación de algunas modalidades sensoriales, entre ellas tacto, propiocepción, sentido vibratorio y dolor. La integridad de la vía del dolor se evalúa al estimular la piel con un alfiler y preguntar si percibe el estímulo como agudo. Para probar la propiocepción, un médico sostiene un dedo del paciente (el dedo gordo, un dedo de la mano o un dedo del pie) y, con el sujeto con los ojos cerrados, pregunta si está moviendo el dedo hacia arriba o hacia abajo. La sensibilidad vibratoria se prueba al aplicar un diapasón que está vibrando (128 Hz) sobre la piel en la yema de un dedo de la mano, en la punta del dedo gordo, o en prominencias óseas de los dedos del pie. La respuesta normal sería una sensación de “zumbido”. La sensación es más evidente sobre huesos. Un patrón de estímulos con presión rítmica es interpretado como vibración. Los impulsos de los cuales depende la sensación de vibración son transportados en las columnas dorsales. La degeneración de esta parte de la médula espinal ocurre en la diabetes mellitus mal controlada, la anemia perniciosa y las deficiencias de vitamina B12. El aumento del umbral para estímulos vibratorios es un síntoma temprano de esta degeneración. La sensación vibratoria y la propiocepción están estrechamente relacionadas; cuando una está disminuida, también lo está la otra. CAPÍTULO 13 Sistemas sensoriales generales: tacto, dolor y temperatura 121 A Norepinefrina Serotonina Nociceptor ENK Neurona de proyección B1 Aferencia sensorial B2 Aferencia sensorial + opiáceos/opioides Control Control Opiáceo Morfina Nociceptor Glutamato Neuropéptidos Glutamato Neuropéptidos Encefalina Ca2+ Morfina Encefalina Ca2+ Aferencia nula Aferencia nula + opiáceos Encefalina Aferencia sensorial Control Aferencia sensorial + opiáceos Control Encefalina Neurona de proyección FIGURA 137 Interneuronas de circuito local en el asta dorsal superficial de la médula espinal integran vías descendente y aferentes. A) Posibles interacciones de fibras aferentes nociceptivas, interneuronas y fibras descendentes en el asta dorsal. Las fibras nociceptivas terminan en neuronas de proyección espinotalámicas de segundo orden. Las interneuronas que contienen encefalina (ENK) ejercen acciones inhibidoras tanto presinápticas como postsinápticas. Las neuronas serotonérgicas y noradrenérgicas en el tallo encefálico activan interneuronas opioides y suprimen la actividad de neuronas de proyección espinotalámicas. B1) La activación de nociceptores libera glutamato y neuropéptidos desde terminales sensoriales, lo que despolariza neuronas de proyección y las activa. B2) Los opiáceos aminoran el flujo de entrada de Ca2+ lo que lleva a un decremento de la duración de potenciales de acción de nociceptor y una liberación disminuida de transmisor. Asimismo, hiperpolarizan la membrana de neuronas del asta dorsal al activar la conductancia de K+ y disminuir la amplitud del EPSP (capítulo 7) producido por estimulación de nociceptores. (Reproducida con autorización de Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000.) La estereognosia es la percepción de la forma y naturaleza de un objeto sin verlo. Los individuos sanos pueden identificar con facilidad objetos como llaves y monedas de diversas denominaciones. Esta capacidad depende de sensaciones de tacto y presión relativamente intactas, y está alterada cuando hay daño de las columnas dorsales. La incapacidad para identificar un objeto mediante tacto se llama agnosia táctil. La estereognosia alterada es un signo temprano de daño de la corteza cerebral, y a veces ocurre en ausencia de defec- to detectable alguno de la sensación de tacto y presión cuando hay una lesión en el lóbulo parietal en posición posterior a la circunvolución central posterior. También la estereognosia puede expresarse por el fracaso para identificar un objeto al verlo (agnosia visual), la incapacidad para identificar sonidos o palabras (agnosia auditiva) o color (agnosia de color), o la incapacidad para identificar la ubicación de una extremidad o la posición de la misma (agnosia de posición). 122 SECCIÓN IV Fisiología del sistema nervioso central/neural CORRELACIÓN CLÍNICA Una mujer de 55 años de edad, ejecutiva en una corporación grande, presentó una sensación de ardor en la palma de la mano derecha alrededor de seis meses antes. También notó hormigueo y entumecimiento en el pulgar y los dedos índice y medio derechos. Estos síntomas aparecieron después de pasar muchas y largas horas frente a la computadora preparando documentos del reporte anual para la corporación. Al inicio, los síntomas fueron más prominentes por la noche, e interrumpían el sueño de la paciente. El problema se ha intensificado a últimas fechas, y ahora tiene dolor en la muñeca derecha y dificultad para tomar objetos pequeños desde el escritorio. Visitó a su médico porque el trabajo en la computadora se había hecho cada vez más difícil. El médico llevó a cabo varias pruebas diagnósticas sencillas; cuando ejerció presión sobre el nervio mediano en la muñeca, la paciente experimentó una sensación parecida a descarga eléctrica (signo de Tinel). Cuando el médico le pidió que mantuviera los antebrazos hacia arriba al apuntar con los dedos hacia abajo y presionar los dorsos de las manos uno contra otro, en el transcurso de un minuto la mujer sintió hormigueo y entumecimiento creciente en los dedos de la mano (signo de Phalen). Las pruebas de conducción nerviosa indicaron conducción lentificada en el nervio mediano. Se diagnosticó síndrome del túnel carpiano, que se debe a compresión (quizá debido a inflamación) del nervio mediano que pasa por el túnel. Es más prevaleciente en mujeres que en varones, y se diagnostica por lo regular en individuos que usan las muñecas en actividades repetitivas (operadores de computadora, cajeros, músicos, pintores). Alrededor de 3% de las mujeres y 2% de los varones tienen probabilidades de que se les diagnostique este síndrome durante su vida. El nervio mediano proporciona información sensitiva proveniente del pulgar, y de los dedos índice y anular, y los nueve tendones que flexionan los dedos de las manos. El síndrome se caracteriza por dolor, parestesias y debilidad en la distribución del nervio mediano. El dolor en la muñeca o la mano, o el entumecimiento y hormigueo de los dedos de la mano (excepto el dedo meñique, que no está inervado por el nervio mediano) a menudo son los primeros síntomas. Los pacientes a veces informan debilidad en la mano, y una tendencia a dejar caer cosas. Los síntomas a menudo aparecen primero por la noche más que durante la actividad. El mejor tratamiento suele ser la colocación de una férula en la muñeca, AINE o corticosteroides. Si el dolor persiste, puede requerirse intervención quirúrgica. RESUMEN DEL CAPÍTULO ■ ■ Los receptores sensoriales por lo común se clasifican como mecanorreceptores, nociceptores, quimiorreceptores o fotorreceptores. El tacto y la presión son detectados por cuatro tipos de mecanorreceptores: corpúsculos de Meissner (que muestran respuesta a cambios de la textura y vibraciones lentas), las células de Merkel (que responden a presión y tacto sostenidos), los corpúsculos de ■ ■ ■ ■ ■ ■ Ruffini (que muestran respuesta a presión sostenida) y los corpúsculos de Pacini (que responden a presión profunda y vibraciones rápidas). Los nociceptores y termorreceptores son terminaciones nerviosas libres sobre fibras no mielinizadas o un poco mielinizadas en piel con pelo y glabra, y tejidos profundos. La hiperalgesia es una respuesta exagerada a un estímulo nocivo; la alodinia es una sensación de dolor en respuesta a un estímulo inocuo. La conversión de un estímulo de receptor en una sensación reconocible se denomina codificación sensorial. Todos los sistemas sensoriales codifican para cuatro atributos elementales de un estímulo: modalidad, ubicación, intensidad y duración. El tacto discriminativo, la propiocepción y las sensaciones vibratorias son transmitidos por medio de la vía de la columna dorsal (lemnisco medial) a SI. Las sensaciones de dolor y temperatura son mediadas por medio del tracto espinotalámico ventrolateral a SI. Las vías descendentes desde la PAG mesencefálica inhiben la transmisión en vías nociceptivas. Esta vía descendente incluye una sinapsis en el núcleo del rafe y la liberación de opiáceos endógenos. La morfina es un agente antinociceptivo eficaz que se une a receptores opiáceos endógenos en el mesencéfalo, el tallo encefálico y la médula espinal. PREGUNTAS DE ESTUDIO 1. Los corpúsculos de Pacini: A) son un tipo de termorreceptor. B) por lo general están inervados por fibras nerviosas Aδ. C) son receptores de tacto que se adaptan con rapidez. D) son receptores de tacto que se adaptan de manera lenta. E) son nociceptores. 2. La adaptación a un estímulo sensorial produce: A) una sensación disminuida cuando se suspenden otros tipos de estímulos sensoriales. B) una sensación más intensa cuando un estímulo dado se aplica de forma repetitiva. C) una sensación localizada a la mano cuando se estimulan los nervios del plexo braquial. D) una sensación disminuida cuando un estímulo dado se aplica repetidas veces con el tiempo. E) una tasa de activación disminuida en el nervio sensorial del receptor cuando se dirige la atención a otro tema. 3. ¿Los sistemas sensoriales codifican para cuál de los atributos que siguen de un estímulo? A) modalidad, ubicación, intensidad y duración B) umbral, campo receptivo, adaptación y discriminación C) tacto, gusto, audición y olfato D) umbral, lateralidad, sensación y duración E) sensibilización, discriminación, energía y proyección 4. Los termorreceptores: A) sólo son activados por frío o calor intenso. B) están ubicados sobre capas superficiales de la piel C) son un subtipo de nociceptor. D) están en terminaciones dendríticas de fibras Aδ y fibras C. E) todas las anteriores. CAPÍTULO 13 Sistemas sensoriales generales: tacto, dolor y temperatura 5. Una mujer de 50 años de edad es objeto de un examen neurológico que muestra pérdida de la sensibilidad al dolor y la temperatura, el sentido vibratorio y la propiocepción en ambas piernas. Estos síntomas podrían explicarse por: A) un tumor en la vía del lemnisco medial en la parte sacra de la médula espinal. B) una neuropatía periférica. C) un tumor grande en el asta dorsal sacra. D) un tumor grande que afecta la circunvolución paracentral posterior. E) un tumor grande en los núcleos talámicos ventrales posterolateral y posteromedial. 123 Reflejos espinales C A P Í T U L O 14 Susan M. Barman O B J E T I V O S ■ ■ ■ ■ ■ Definir los componentes de un arco reflejo. Describir los husos musculares y su función en el reflejo de estiramiento. Explicar las funciones de los órganos tendinosos de Golgi como parte de un sistema de retroacción que mantiene la fuerza muscular. Definir la inervación recíproca, el reflejo de estiramiento inverso, y el clono. Describir los efectos a corto y largo plazos de la lesión de la médula espinal sobre los reflejos espinales. INTRODUCCIÓN La unidad básica de la actividad refleja integrada es el arco reflejo. Este arco consta de un órgano de sentido, una neurona aferente, sinapsis dentro de una estación integradora central, una neurona eferente, y un órgano efector. Las neuronas aferentes entran al sistema nervioso central (SNC) por medio de las raíces dorsales espinales o los nervios craneales, y tienen su cuerpo celular en los ganglios de la raíz dorsal o en los ganglios homólogos para los nervios craneales. Las fibras eferentes salen del SNC por medio de las raíces ventrales espinales o nervios craneales motores correspondientes. La actividad en el arco reflejo empieza en un receptor sensorial con un potencial generador cuya magnitud es proporcional a la fuerza del estímulo (figura 14-1). Esto genera potenciales de acción de todo o nada en el nervio aferente; el número de potenciales de acción es proporcional al tamaño del potencial generador. En el SNC, las respuestas de nuevo son graduadas en términos de potenciales postsinápticos excitadores (EPSP) y potenciales postsinápticos inhibidores (IPSP) en las uniones sinápticas (capítulo 7). Se generan respuestas de todo o nada en el nervio eferente; cuando éstas llegan al órgano efector, de nuevo establecen una respuesta graduada. Cuando el efector es músculo liso, las respuestas se suman para producir potenciales de acción en el músculo liso, pero cuando el efector es músculo esquelético, la respuesta graduada es adecuada para producir potenciales de acción que desencadenan contracción muscular. La actividad dentro del arco reflejo es modificada por las múltiples aferencias que convergen en las neuronas eferentes o en cualquier estación sináptica dentro del asa refleja. El arco reflejo más simple es el que tiene una sinapsis única entre las neuronas aferentes y eferentes. Dichos arcos son monosinápticos, y los reflejos que ocurren en ellos se llaman reflejos monosinápticos. Los arcos reflejos en los cuales hay una o más interneuronas interpuestas entre las neuronas aferentes y eferentes se llaman reflejos polisinápticos. Puede haber desde dos, hasta cientos de sinapsis en un arco reflejo polisináptico. Como es evidente a partir de la descripción que se presenta más adelante, la actividad refleja se estereotipa y especifica en términos tanto del estímulo como de la respuesta; un estímulo particular desencadena una respuesta particular. El hecho de que las respuestas reflejas se estereotipen no excluye la posibilidad de que sean modificadas por la experiencia. Los reflejos son adaptables y pueden cambiarse para realizar tareas motoras y mantener el equilibrio. Las aferencias descendentes que provienen de regiones más altas del cerebro desempeñan un papel importante en la modulación de reflejos espinales y la adaptación de los mismos. REFLEJO MONOSINÁPTICO: EL REFLEJO DE ESTIRAMIENTO Cuando un músculo esquelético con inervación intacta es estirado, se contrae. Esta respuesta se llama reflejo de estiramiento. El estímulo que inicia el reflejo es estiramiento del músculo, y la respuesta es contracción del mismo músculo. El órgano de sentido (receptor) es una pequeña estructura que se parece a huso o fusiforme encapsulada que se llama el huso muscular, el cual se ubica dentro de la parte 125 126 SECCIÓN IV Fisiología del sistema nervioso central/neural Órgano de sentido Potencial generador Neurona aferente Sinapsis Potenciales de acción EPSP (e IPSP) Neurona eferente Potenciales de acción Unión neuromuscular Músculo Potenciales de placa terminal Potenciales de acción FIGURA 141 Arco reflejo. En el receptor y en el SNC ocurre una respuesta graduada, no propagada, proporcional a la magnitud del estímulo. La respuesta en la unión neuromuscular también es graduada, aunque en condiciones normales siempre es suficientemente grande como para producir una respuesta en el músculo esquelético. Por otro lado, en las porciones del arco especializadas para la transmisión (axones aferentes y eferentes, membrana muscular), las respuestas son potenciales de acción de todo o nada. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) carnosa del músculo. Los impulsos que se originan desde el huso se transmiten al SNC por medio de fibras sensoriales rápidas (grupo Ia) que pasan de manera directa a las neuronas motoras que inervan el mismo músculo. El reflejo de estiramiento es el reflejo monosináptico mejor conocido y estudiado, y se tipifica por el reflejo rotuliano. Un golpe en el tendón rotuliano desencadena el reflejo rotuliano, un reflejo de estiramiento del músculo cuádriceps femoral, porque el golpe sobre el tendón estira el músculo. El reflejo rotuliano es un ejemplo de un reflejo tendinoso profundo en un examen neurológico. La falta del reflejo rotuliano puede significar una anormalidad en cualquier sitio dentro del arco reflejo, como el huso muscular, las fibras nerviosas aferentes Ia, o las neuronas motoras del músculo cuádriceps. La causa más común es una neuropatía periférica que puede depender de diabetes mellitus, alcoholismo y toxinas, entre otras. Un reflejo hiperactivo puede significar una interrupción de vías corticoespinales inhibidoras y otras vías descendentes que influyen sobre el arco reflejo. ESTRUCTURA DE LOS HUSOS MUSCULARES En la figura 14-2A se ilustra la composición de un huso muscular y su inervación. Cada huso muscular tiene tres elementos esenciales: 1) un grupo de fibras musculares intrafusales especializadas, con extremos polares contráctiles y un centro no contráctil, 2) nervios aferentes mielinizados de diámetro grande (tipos Ia y II) que se originan en la porción central de las fibras intrafusales y 3) nervios eferentes mielinizados de diámetro pequeño que inervan las regiones contráctiles polares de las fibras intrafusales. Es importante entender la relación de estos elementos entre sí y con el músculo esquelético mismo para apreciar el papel de este órgano de sentido en la emisión de señales de cambios de la longitud del músculo en el cual se localiza. Los cambios de la longitud del músculo se asocian con modificaciones en el ángulo de la articulación; así, los husos musculares proporcionan información sobre la posición (esto es, propiocepción). Las fibras intrafusales se ubican en paralelo a las fibras extrafusales (las unidades contráctiles regulares del músculo) con los extremos de la cápsula del huso fijos a los tendones en ambos extremos del músculo. No contribuyen a la fuerza contráctil general del músculo, sino más bien desempeñan una función sensorial. Hay dos tipos de fibras intrafusales en husos musculares de mamífero. El primer tipo contiene muchos núcleos en un área central dilatada, y se llama fibra de bolsa nuclear (figura 14-2B). Hay dos subtipos de fibras de bolsa nuclear, dinámica y estática. Típicamente, hay 2 o 3 fibras de bolsa nuclear por cada huso. El segundo tipo de fibra intrafusal, la fibra de cadena nuclear, es más delgada y más corta, y carece de una bolsa definida. Cada huso tiene alrededor de cinco fibras de cadena nuclear. Hay dos clases de terminaciones sensoriales en cada huso, una terminación primaria (grupo Ia) única, y hasta ocho terminaciones secundarias (grupo II). La fibra aferente es envuelta por el centro de las fibras de bolsa nuclear dinámicas y estáticas, y las fibras de cadena nuclear. Las fibras sensoriales del grupo II se sitúan adyacentes a los centros de la bolsa nuclear estática y las fibras de la cadena nuclear; estas fibras no inervan las fibras de bolsa nuclear dinámicas. Las fibras aferentes Ia son muy sensibles a la velocidad del cambio de la longitud del músculo durante un estiramiento (respuesta dinámica); así, proporcionan información acerca de la rapidez de los movimientos, y permiten que se hagan movimientos correctivos rápidos. La actividad de estado estable (tónica) de fibras aferentes del grupo Ia y II proporciona información sobre la longitud de estado estable del músculo (respuesta estática). En el trazo superior en la figura 14-2C se muestran los componentes dinámico y estático de actividad en una fibra aferente durante estiramiento muscular. Note que descargan con mayor rapidez mientras al músculo lo estiran (área sombreada de los gráficos), y con menos rapidez durante estiramiento sostenido. Los husos tienen su propia inervación motora eferente que se llaman neuronas motoras γ. Son fibras de diámetro pequeño (3 a 6 μm), y constituyen alrededor de 30% de las fibras en las raíces ventrales. Hay dos tipos de neuronas motoras γ: dinámicas, que proporcionan las fibras de bolsa nuclear dinámica, y estáticas, que suministran las fibras de bolsa nuclear estática y las fibras de cadena nuclear. La activación de neuronas motoras γ dinámicas aumenta la CAPÍTULO 14 Reflejos espinales Fibras musculares intrafusales B Fibras intrafusales del huso muscular Fibra de bolsa nuclear dinámica Fibra de bolsa nuclear estática C Respuesta de una fibra sensorial Ia a activación selectiva de neuronas motoras 200 Imp/s A Huso muscular Respuesta dinámica Respuesta de estado estable Fibras de cadena nuclear Cápsula 127 0 Estiramiento solo 200 Axones eferentes 0 Estática Estimula fibra γ estática 200 Dinámica Terminaciones motoras γ Imp/s Axones aferentes Ia 0 Estiramiento Terminaciones sensoriales Imp/s II Estimula fibra γ dinámica 6 0 0.2 s FIGURA 142 Huso muscular de mamífero. A) Diagrama de los principales componentes del huso muscular de mamífero, incluso fibras musculares intrafusales, terminaciones de fibra sensorial aferente, y fibras motoras eferentes (neuronas motoras γ). B) Tres tipos de fibras musculares intrafusales: bolsa nuclear dinámica, bolsa nuclear estática y fibras de cadena nuclear. Una fibra aferente Ia única inerva los tres tipos de fibras para formar una terminación sensorial primaria. Una fibra sensorial del grupo II inerva fibras de cadena nuclear y de bolsa estática para formar una terminación sensorial secundaria. Las neuronas motoras γ dinámicas inervan fibras de bolsa dinámica; las neuronas motoras γ estáticas inervan combinaciones de fibras de cadena nuclear y de bolsa estática. C) Comparación del patrón de descarga de actividad aferente Ia durante estiramiento solo y durante estimulación de neuronas motoras γ estáticas o dinámicas. Sin estimulación de γ, las fibras aferentes Ia muestran una respuesta dinámica pequeña al estiramiento muscular, y un aumento modesto de la activación de estado estable. Cuando las neuronas motoras γ estáticas son activadas, la respuesta de estado estable aumenta y la respuesta dinámica disminuye. Cuando las neuronas motoras γ dinámicas son activadas, la respuesta dinámica es notoriamente aumentada, pero la respuesta de estado estable vuelve de manera gradual a su magnitud original. (Reproducida con autorización de Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000.) sensibilidad dinámica de las terminaciones aferentes del grupo Ia. La activación de las neuronas motoras γ estáticas incrementa el nivel de actividad tónica en las terminaciones aferentes de los grupos, tanto Ia como II, disminuye la sensibilidad dinámica de las fibras aferentes del grupo Ia, y puede evitar el silenciamiento de fibras aferentes Ia durante el estiramiento muscular (figura 14-2C). CONEXIONES CENTRALES DE FIBRAS AFERENTES Las fibras aferentes del grupo Ia terminan de manera directa sobre neuronas motoras α, e inervan las fibras extrafusales del mismo músculo (figura 14-3). El tiempo que transcurre entre la aplicación del estímulo y la respuesta se llama el tiempo de reacción. En seres humanos, el tiempo de reacción para un reflejo de estiramiento como el reflejo rotuliano es de 19 a 24 ms. Puesto que se conocen las velocidades de conducción de los tipos de fibras aferentes y eferentes, y puede medirse la distancia desde el músculo hasta la médula espinal, es posible calcular qué tanto del tiempo de reacción ocupó la conducción hacia la médula espinal y desde esta última. Cuando este valor se resta del tiempo de reacción, el restante, que se conoce como retraso central, es el tiempo que la actividad refleja tarda en cruzar la médula espinal. En seres humanos, el retraso central para el reflejo rotuliano es de 0.6 a 0.9 ms. Puesto que el retraso sináptico mínimo es de 0.5 ms, sólo una sinapsis podría haber sido cruzada. Los husos musculares también hacen conexiones que causan contracción muscular por medio de vías polisinápticas, y las fibras aferentes involucradas, quizá, son las de las terminaciones del grupo II secundarias. FUNCIÓN DE LOS HUSOS MUSCULARES Cuando el huso muscular es estirado, se deforman sus terminaciones sensoriales, y se generan potenciales de receptor; éstos, a su vez, establecen potenciales de acción en las fibras sensoriales a una frecuencia proporcional al grado de estiramiento. Dado que el huso está en paralelo con las fibras extrafusales, cuando el músculo es estirado de manera pasiva, los husos también son estirados, lo que se denomina “cargar el huso”, lo anterior inicia contracción refleja de las fibras extrafusales en el músculo esquelético. Por otro lado, las 128 SECCIÓN IV Fisiología del sistema nervioso central/neural Huso Raíz dorsal Tendón Fibra extrafusal Nervio sensorial Interneurona que libera mediador inhibidor Fibra Ib proveniente del órgano tendinoso de Golgi Fibra Ia proveniente del huso muscular Impulsos en el nervio sensorial Neurona motora Músculo en reposo Raíz ventral Placa terminal motora sobre fibra extrafusal Músculo estirado FIGURA 14-3 Diagrama que ilustra las vías de las cuales dependen el reflejo de estiramiento y el reflejo de estiramiento inverso. El estiramiento estimula el huso muscular, que activa fibras aferentes Ia que excitan la neurona motora. También estimula el órgano tendinoso de Golgi, el cual activa fibras aferentes Ib que excitan una interneurona que libera el mediador inhibidor glicina. Con el estiramiento fuerte, la hiperpolarización resultante de la neurona motora es tan grande que deja de descargar. (Reproducida con autorización Músculo contraído de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill, 2009.) fibras aferentes del huso muscular dejan de activarse de manera característica cuando se hace que el músculo se contraiga con estimulación eléctrica de las neuronas motoras α que van a las fibras extrafusales, porque el músculo se acorta mientras el huso está descargado (figura 14-4). Así, el huso y sus conexiones reflejas constituyen un dispositivo de retroacción que opera para mantener la longitud del músculo; si el músculo es estirado, la descarga del huso aumenta y se produce acortamiento reflejo, mientras que si el músculo es acortado sin un cambio de la descarga de neurona motora γ, la actividad aferente del huso disminuye, y el músculo se relaja. Cuando ocurre un reflejo de estiramiento, los músculos que antagonizan la acción del músculo involucrado (antagonistas) se relajan. Este fenómeno se debe a la inervación recíproca. Los impulsos en las fibras aferentes, que provienen de los husos musculares del músculo protagonista, causan inhibición postsináptica de las neuronas motoras α que van hacia los antagonistas. La vía que media este efecto es bisináptica. Una fibra colateral que se origina de cada fibra aferente Ia pasa en la médula espinal hacia una interneurona inhibidora, la cual hace sinapsis en una neurona motora que inerva los músculos antagonistas. EFECTOS DE LA DESCARGA DE NEURONA MOTORA γ La estimulación de neuronas motoras γ hace que los extremos contráctiles de las fibras intrafusales se acorten y, por ende, estira la por- Descarga eferente γ aumentada Descarga eferente γ aumentada —músculo estirado FIGURA 144 Efecto de diversas condiciones sobre la descarga del huso muscular. Cuando todo el músculo es estirado, el huso muscular también es estirado, y sus terminaciones sensoriales son activadas a una frecuencia proporcional al grado de estiramiento (“carga del huso”). Las fibras aferentes del huso dejan de activarse cuando el músculo se contrae (“descarga del huso”). La estimulación de neuronas motoras γ hace que las terminaciones contráctiles de las fibras intrafusales se acorten. Esto estira la región de la bolsa nuclear, lo que inicia impulsos en fibras sensoriales. Si todo el músculo es estirado durante la estimulación de las neuronas motoras γ, la tasa de descarga en fibras sensoriales aumenta más. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) CAPÍTULO 14 Reflejos espinales ción de la bolsa nuclear de los husos, lo que deforma las terminaciones e inicia impulsos en las fibras aferentes Ia (figura 14-4). Lo anterior puede llevar a la contracción refleja del músculo. Así, es posible hacer que el músculo esquelético se contraiga por medio de estimulación de las neuronas motoras α que inervan las fibras extrafusales o las neuronas motoras γ que inician contracción de manera indirecta por medio del reflejo de estiramiento. La actividad aumentada de neurona motora γ incrementa la sensibilidad del huso durante estiramiento. En respuesta a aferencias excitadoras descendentes que van hacia circuitos motores espinales, las neuronas motoras tanto α como γ son activadas. Debido a esta “coactivación α-γ”, las fibras intrafusales y extrafusales se acortan juntas, y puede ocurrir actividad aferente del huso durante todo el periodo de contracción muscular. De esta manera, el huso permanece capaz de responder al estiramiento y ajustar de manera refleja la descarga de neurona motora α. Las aferencias desde varias regiones del cerebro hacia las neuronas motoras γ influyen sobre la sensibilidad de los husos musculares. Así, el umbral de los reflejos de estiramiento en diversas partes del cuerpo se puede ajustar y cambiar para satisfacer las necesidades de control postural. La ansiedad causa un aumento de la descarga de neurona motora γ, lo cual puede explicar la aparición de reflejos tendinosos hiperactivos en pacientes ansiosos. Asimismo, el movimiento inesperado se asocia con una mayor descarga. 129 aferentes Ib terminan en la médula espinal en interneuronas inhibidoras que, a su vez, terminan de manera directa sobre las neuronas motoras α (figura 14-3). También hacen conexiones excitadoras con neuronas motoras α que proporcionan antagonistas al músculo. A diferencia de los husos, los órganos tendinosos de Golgi están en serie con las fibras musculares. Así, son estimulados tanto por estiramiento pasivo como por contracción activa del músculo. El umbral de los órganos tendinosos de Golgi es bajo. El grado de estimulación por estiramiento pasivo no es grande porque las fibras musculares más elásticas absorben gran parte del estiramiento, y esta es la razón por la cual se requiere un estiramiento fuerte para producir relajación. Empero, la descarga se produce, de manera regular, por contracción del músculo y, así, el órgano tendinoso de Golgi funciona como un transductor en un circuito de retroacción que regula la fuerza muscular de una manera análoga al circuito de retroacción del huso que regula la longitud del músculo. La importancia de las terminaciones primarias en los husos y los órganos tendinosos de Golgi en la regulación de la velocidad de la contracción muscular, la longitud del músculo y la fuerza muscular, se ilustra por el hecho de que el corte de los nervios aferentes que van a un brazo hace que la extremidad cuelgue laxamente en un estado semiparalizado. La organización del sistema se muestra en la figura 14-6. TONO MUSCULAR REFLEJO DE ESTIRAMIENTO INVERSO Hasta cierto grado, mientras más se estira un músculo, más fuerte es la contracción refleja. Sin embargo, cuando la tensión se hace grande, la contracción cesa de repente y el músculo se relaja. Esta relajación en respuesta a estiramiento fuerte se llama reflejo de estiramiento inverso. El receptor para el reflejo de estiramiento inverso es el órgano tendinoso de Golgi (figura 14-5); este órgano consta de un conjunto que se parece a una red de terminaciones nerviosas con protuberancias entre los fascículos de un tendón. Hay de 3 a 25 fibras musculares por cada órgano tendinoso. Las fibras sensoriales que provienen de los órganos tendinosos de Golgi forman el grupo Ib de fibras nerviosas mielinizadas, de conducción rápida. La estimulación de estas fibras aferentes Ib lleva a la producción de IPSP en las neuronas motoras α que inervan el músculo, desde las cuales surgen las fibras. Las hebras Fibra nerviosa La resistencia de un músculo al estiramiento a menudo se denomina su tono. Si se corta un nervio motor que inerva un músculo esquelético, el músculo ofrece muy poca resistencia, y se dice que está flácido. Un músculo hipertónico (espástico) es aquel en el cual la resistencia al estiramiento es alta por reflejos de estiramiento hiperactivos. En algún sitio entre los estados de flacidez y espasticidad está el área poco definida del tono normal. Los músculos, en general, son hipotónicos cuando la tasa de descarga de neurona motora γ es baja, e hipertónicos cuando es alta. Cuando los músculos son hipertónicos, se observa la secuencia de estiramiento moderado → contracción muscular, y estiramiento fuerte → relajación muscular. La flexión pasiva del codo, por ejemplo, encuentra resistencia inmediata como un resultado del reflejo de estiramiento en el músculo tríceps. El estiramiento adicional activa el reflejo de estiramiento inverso. La resistencia a la flexión de pronto se colapsa, y el brazo se flexiona. La flexión pasiva continua Fascículos tendinosos Fibras musculares Órgano de Golgi, que muestra ramificación de fibrillas nerviosas FIGURA 145 Órgano tendinoso de Golgi. (Reproducida con autorización de Gray H [editor]: Gray’s Anatomy of the Human Body, 29th ed. Lea & Febiger, 1973.) 130 SECCIÓN IV Fisiología del sistema nervioso central/neural Señal de control interneuronal Interneuronas Retroacción de fuerza Órganos tendinosos Alteraciones internas – Señal control α + α Señal eferente Músculo + Fuerzas externas Fuerza muscular + – Longitud del músculo Carga Longitud y velocidad Señal de control dinámica γ Señal de control estática γ Retroacción de longitud y velocidad Husos γd γs FIGURA 146 Diagrama de bloques del sistema de control motor periférico. Retroacción no neural desde el músculo (“longitud y velocidad”) que limita la longitud y la velocidad por medio de las propiedades mecánicas inherentes del músculo. γd, neuronas motoras γ dinámicas; γs, neuronas motoras γ estáticas. (Reproducida con autorización de Houk J: Medical Physiology, 13th ed. In: Mountcastle VB (editor). Mosby, 1974.) estira el músculo de nuevo, y la secuencia puede repetirse; esta secuencia de resistencia seguida por dar de sí cuando una extremidad es movida de manera pasiva se conoce como fenómeno de la navaja de muelle por su semejanza con el cierre de una navaja de bolsillo. El clono es la aparición de contracciones rítmicas, repetitivas, regulares, de un músculo sujeto a estiramiento repentino y sostenido. El clono sostenido con cinco o más contracciones se considera anormal. Durante un examen neurológico, el clono del tobillo puede iniciarse por dorsiflexión rápida, enérgica y sostenida del pie, y la respuesta es la flexión plantar rítmica en el tobillo. El clono también puede ocurrir después de alteración de las aferencias corticales descendentes hacia una interneurona inhibidora glicinérgica espinal que se conoce como la célula de Renshaw. Esta célula recibe aferencias excitadoras que provienen de neuronas motoras α por medio de colaterales de axón (y, a su vez, la célula de Renshaw inhibe la neurona motora). Además, fibras corticales que activan flexores del tobillo tienen contacto con células de Renshaw (así como con interneuronas inhibidoras activadas por fibras aferentes Ia) que inhiben los extensores del tobillo antagonistas. Este circuito evita la estimulación refleja de los extensores cuando los flexores son activos. Por ende, cuando hay daño de las fibras corticales descendentes (lesión de neurona motora superior), no hay inhibición de antagonistas. El resultado es contracción secuencial repetitiva de flexores y extensores del tobillo (clono). El clono puede observarse en pacientes con esclerosis lateral amiotrófica, apoplejía, esclerosis múltiple, daño de la médula espinal y encefalopatía hepática. REFLEJOS POLISINÁPTICOS: EL REFLEJO DE RETIRADA El reflejo de retirada es un reflejo polisináptico típico que ocurre en respuesta a una estimulación dolorosa de la piel o de los tejidos sub- cutáneos y músculo. La respuesta es contracción de músculo flexor e inhibición de músculos extensores, de modo que la parte del cuerpo estimulada es flexionada y retirada del estímulo. Cuando se aplica un estímulo fuerte a una extremidad, la respuesta no sólo incluye flexión y retirada de esa extremidad, sino también extensión de la extremidad opuesta; esta respuesta extensora cruzada forma parte del reflejo de retirada. Las respuestas flexoras pueden producirse por estimulación inocua de la piel o por estiramiento del músculo, pero las respuestas flexoras fuertes con retirada sólo son iniciadas por estímulos que son nocivos o al menos en potencia perjudiciales (estímulos nociceptivos). La flexión de la extremidad estimulada la aleja de la fuente de irritación, y la extensión de la otra extremidad apoya el cuerpo. Conforme aumenta la fuerza de un estímulo nocivo, el tiempo de reacción se acorta. La facilitación espacial y temporal ocurre en sinapsis en la vía polisináptica. Los estímulos más fuertes producen más potenciales de acción por segundo en las ramas activas, y hacen que más ramas se hagan activas; por ende, la suma de los EPSP a la magnitud de activación ocurre con mayor rapidez. Otra característica de la respuesta de retirada es el hecho de que la estimulación submáxima de cualesquiera de los nervios sensoriales de una extremidad nunca produce una contracción tan fuerte de los músculos flexores como la desencadenada por la estimulación eléctrica directa de los músculos mismos. Esto indica que los impulsos aferentes fraccionan el fondo común de neuronas motoras α, es decir, cada aferencia va a sólo parte del fondo común de neuronas motoras que inervan los flexores de esa extremidad particular. Por otro lado, si todas las aferencias sensoriales se separan y estimulan una después de la otra, la suma de la tensión que se desarrolla por estimulación de cada una es mayor que la que se produce por la estimulación directa del músculo o la estimulación de todas las aferencias a la vez. Esto indica que los diversos impulsos aferentes comparten algunas de las neuronas motoras, y que ocurre oclusión cuando todas las aferencias se estimulan a la vez. CAPÍTULO 14 Reflejos espinales INTEGRACIÓN ESPINAL Las neuronas motoras α espinales que inervan las fibras extrafusales en músculos esqueléticos son el lado eferente de muchos arcos reflejos. Todas las influencias neurales que afectan la contracción muscular se canalizan a través de ellos hacia los músculos, y, por ende, se llaman la vía común final. La superficie de la neurona motora α promedio y sus dendritas dan cabida a alrededor de 10 000 protuberancias sinápticas, lo que permite que haya muchas aferencias sinápticas. Al menos cinco aferencias van desde el mismo segmento espinal hasta una neurona motora espinal típica. Además de éstas, hay aferencias excitadoras e inhibidoras, por lo general transmitidas por medio de interneuronas, desde otros niveles de la médula espinal, y múltiples tractos descendentes largos desde el cerebro. Todas estas vías convergen en la vía común final y determinan la actividad de la misma. LESIÓN DE LA MÉDULA ESPINAL Un componente clave de los exámenes neurológicos comprende una evaluación de la integridad de los reflejos espinales. Las anormalidades de los reflejos con frecuencia orientan hacia la ubicación de una lesión de la médula espinal (SCI). Los defectos después de SCI varían, por supuesto, y dependen del nivel de la lesión y la gravedad de la misma. La transección de la médula espinal va seguida por un periodo de choque espinal durante el cual todas las respuestas reflejas espinales están muy deprimidas. Después, las respuestas reflejas regresan y se hacen hiperactivas. En seres humanos, el choque espinal, por lo general, dura un mínimo de dos semanas. El cese del bombardeo tónico de neuronas espinales por impulsos excitadores en vías descendentes, sin duda, desempeña una función en el choque espinal. La recuperación de la excitabilidad refleja que puede deberse al desarrollo de hipersensibilidad por desnervación a los mediadores liberados por las terminaciones excitadoras espinales restantes (capítulo 12). Otra posibilidad es el brote de colaterales desde neuronas existentes, con la formación de terminaciones excitadoras adicionales sobre interneuronas y neuronas motoras. La primera respuesta refleja en aparecer a medida que desaparece el choque espinal, suele ser una contracción leve de los flexores y aductores de la pierna en respuesta a un estímulo nocivo. En algunos pacientes, el reflejo rotuliano se recupera primero. Una vez que los reflejos espinales empiezan a reaparecer tras choque espinal, su umbral disminuye a un ritmo constante. mal a pinchazo y pellizco en las extremidades superiores y la pierna derecha, pero fue incapaz de detectar los estímulos nocivos que se le aplicaron en la pierna izquierda. También perdió la sensación de tacto y vibración en la pierna derecha, pero la sensación fue normal en la pierna izquierda y las extremidades superiores. Hubo poco movimiento espontáneo, si es que hubo alguno, en la pierna derecha, aunque todas las otras extremidades parecieron tener movimiento normal. Una resonancia magnética mostró que el lado derecho de la médula espinal estuvo gravemente dañado en el nivel torácico 10. Se estima que la incidencia anual en todo el mundo de SCI es de 10 a 83 por millón de habitantes. Las causas principales son accidentes vehiculares, violencia y lesiones deportivas. Alrededor de 52% de los casos de SCI da lugar a cuadriplejía, y alrededor de 42% lleva a paraplejía. La edad media de pacientes que sufren una SCI es de 33 años, y los varones superan en número a las mujeres por casi 4:1. La lesión de este paciente llevó a una hemisección de la médula espinal en el nivel torácico 10. Tal lesión causa un cuadro clínico característico que refleja daño de las vías sensorial ascendente (vía de la columna dorsal, tracto espinotalámico ventrolateral) y motora descendente (tracto corticoespinal), que se llama síndrome de BrownSéquard. La lesión del fascículo gracilis o del fascículo cuneatus lleva a pérdida ipsilateral del tacto discriminativo, la vibración y la propiocepción por debajo del nivel de la lesión. La pérdida del tracto espinotalámico lleva a pérdida de la sensación de dolor y temperatura contralateral, empezando uno o dos segmentos por abajo de la lesión. El daño del tracto corticoespinal produce debilidad y espasticidad en ciertos grupos musculares en el mismo lado del cuerpo. Aunque una hemisección espinal precisa es rara, el síndrome es común porque puede producirse por tumor de la médula espinal, traumatismo, enfermedad de disco degenerativa, e isquemia. RESUMEN DEL CAPÍTULO ■ ■ CORRELACIÓN CLÍNICA Un varón de 21 años de edad, estudiante de Medicina, fue herido con arma blanca en la espalda durante un intento de asalto. Un testigo llamó al número de emergencias y, cuando los paramédicos llegaron, el estudiante no podía mover la pierna derecha; fue trasladado con rapidez a la sala de urgencias del hospital local. Además del examen del sitio de lesión y del tratamiento del mismo, se le practicó un examen neurológico. El reflejo del huso muscular fue normal en ambos brazos y en la pierna izquierda, pero fue hiperactivo en la pierna derecha. Tuvo sensación nor- 131 ■ ■ Un arco reflejo consta de un órgano de sentido, una neurona aferente, una o más sinapsis dentro de una estación integradora central, una neurona eferente, y una respuesta efectora. Un huso muscular es un grupo de fibras musculares intrafusales especializadas, con extremos polares contráctiles y un centro no contráctil que se sitúa en paralelo a las fibras musculares extrafusales, y está inervado por fibras aferentes tipos Ia y II y neuronas motoras γ. El estiramiento del músculo activa el huso muscular para que se inicie contracción refleja de las fibras musculares extrafusales en el mismo músculo (reflejo de estiramiento). Un órgano tendinoso de Golgi es un conjunto parecido a una red de terminaciones nerviosas con protuberancias entre los fascículos de un tendón, que se ubica en serie con las fibras musculares extrafusales, e inervado por fibras aferentes tipo Ib. Son estimulados tanto por estiramiento pasivo como por contracción activa del músculo para relajar el músculo (reflejo de estiramiento inverso), y funcionan como un transductor para regular la fuerza muscular. Un colateral proveniente de una fibra aferente Ia se ramifica para terminar en una interneurona inhibidora que hace sinapsis sobre un músculo antagonista (inervación recíproca) para relajar ese músculo cuando el agonista se contrae. El clono es la aparición de 132 ■ SECCIÓN IV Fisiología del sistema nervioso central/neural contracciones rítmicas y regulares de un músculo sujeto a estiramiento repentino y sostenido. La transección de la médula espinal va seguida de un periodo de choque espinal durante el cual todos los reflejos están deprimidos. A esto le sigue un periodo de reflejos hiperactivos. PREGUNTAS DE ESTUDIO 1. El reflejo de estiramiento inverso: A) tiene un umbral más bajo que el del reflejo de estiramiento. B) es un reflejo monosináptico. C) es una relajación de un músculo en respuesta a un estiramiento fuerte del músculo. D) tiene al huso muscular como su receptor. E) requiere la descarga de neuronas centrales que liberan acetilcolina. 2. Cuando la descarga de neurona motora γ aumenta al mismo tiempo que la descarga de neurona motora α al músculo: A) tiene lugar inhibición expedita de la descarga en fibras aferentes Ia del huso. B) la contracción del músculo es prolongada. C) el músculo no se contraerá. D) el músculo no se relajará. E) el número de impulsos en fibras aferentes Ia del huso es mayor que cuando la descarga α sola está aumentada. 3. ¿Cuál de las que siguen no es una característica de un reflejo? A) modificación por impulsos que provienen de diversas partes del SNC B) puede comprender contracción simultánea de algunos músculos y relajación de otros C) está crónicamente suprimida después de transección de la médula espinal D) siempre comprende transmisión a través de al menos una sinapsis E) suele ocurrir sin percepción consciente 4. Los reflejos de retirada no: A) son iniciados por estímulos nociceptivos. B) son un ejemplo de un reflejo disináptico. C) son prolongados si el estímulo es fuerte. D) son un ejemplo de un reflejo flexor. E) se acompañan por la misma respuesta en ambos lados del cuerpo. Sentidos especiales I: visión C A P Í T U L O 15 Susan M. Barman O B J E T I V O S ■ ■ ■ ■ ■ ■ Describir las diversas partes del ojo y listar las funciones de cada una. Explicar cómo los rayos de luz en el ambiente son llevados a un foco en la retina, y el papel de la acomodación en este proceso. Definir los términos que siguen: hipermetropía (hiperopía), miopía, astigmatismo, presbiopía (presbicia) y estrabismo. Describir las respuestas eléctricas que producen los bastones y los conos, y explicar cómo se producen estas respuestas. Trazar las vías neurales que transmiten información visual desde los bastones y conos hacia la corteza visual. Nombrar los cuatro tipos de movimientos oculares, y la función de cada uno. INTRODUCCIÓN Los ojos son órganos complejos de los sentidos. Dentro de su cubierta protectora, cada ojo tiene una capa de receptores, un sistema de lentes que enfoca la luz en estos receptores, y un sistema de nervios que conduce los impulsos desde los receptores hasta el cerebro. La manera en que estos componentes operan para producir imágenes visuales conscientes es el tema de este capítulo. CARACTERÍSTICAS ANATÓMICAS DEL OJO Las principales estructuras del ojo se muestran en la figura 15-1. La capa protectora externa del globo ocular, la esclerótica, está modificada en posición anterior para formar la córnea transparente, a través de la cual los rayos de luz entran al ojo. Dentro de la esclerótica está la coroides, una capa que contiene muchos de los vasos sanguíneos que nutren las estructuras en el globo ocular. El revestimiento de los dos tercios posteriores de la coroides es la retina, el tejido neural que contiene las células receptoras. El cristalino es una estructura transparente sostenida en su sitio por el ligamento suspensorio del cristalino (zónula) circular que está fijo a la parte anterior engrosada de la coroides, el cuerpo ciliar. El cuerpo ciliar contiene fibras musculares circulares y longitudinales que se fijan cerca de la unión corneoescleral. Enfrente del cristalino está el iris pigmentado y opaco, la porción colorada del ojo, que contiene fibras musculares circulares que constriñen la pupila, y fibras radiales que la dilatan. Las variaciones del diámetro de la pupila pueden producir un cambio de hasta cinco veces la cantidad de luz que llega a la retina. El espacio entre el cristalino y la retina está lleno con un material gelatinoso transparente que se conoce como humor vítreo. El humor acuoso, un líquido transparente que nutre la córnea y el cristalino, se produce en el cuerpo ciliar mediante difusión y transporte activo desde el plasma. Fluye a través de la pupila y llena la cámara anterior del ojo. En circunstancias normales se reabsorbe a través del canal de Schlemm, un canal venoso en la unión entre el iris y la córnea (ángulo de la cámara anterior). La obstrucción de esta salida lleva a incremento de la presión intraocular. Una causa de presión aumentada es la permeabilidad disminuida a través de la red trabecular, el tejido que está alrededor de la base de la córnea, que drena el humor acuoso desde el ojo (glaucoma de ángulo abierto), y otro es el movimiento del iris hacia adelante, que oblitera el ángulo (glaucoma de ángulo cerrado). El glaucoma puede tratarse con bloqueadores β-adrenérgicos o inhibidores de la anhidrasa carbónica, ambos de los cuales disminuyen la producción de humor acuoso, o con agonistas colinérgicos, que aumentan el flujo de salida de dicho humor. 133 134 SECCIÓN IV Fisiología del sistema nervioso central/neural Músculo recto superior Esclerótica Conjuntiva Coroides Cuerpo ciliar Cámara posterior Cámara anterior Retina Fóvea central Cavidad anterior Córnea Arteria central Vena central Pupila Cristalino Iris Cámara posterior Nervio óptico Fibras zonulares del ligamento suspensorio Cámara vítrea (cavidad posterior) Músculo recto inferior FIGURA 151 Características anatómicas internas del ojo. (Reproducida con autorización de Fox SI: Human Physiology. McGraw-Hill, 2008.) El ojo está bien protegido contra lesión por las paredes óseas de la órbita. La córnea es humedecida y mantenida transparente por lágrimas que van desde la glándula lagrimal en la porción superior de cada órbita a través de la superficie del ojo para vaciarse por medio del conducto lagrimal hacia la nariz. El parpadeo ayuda a mantener húmeda la córnea. RETINA La retina se extiende en posición anterior casi hasta el cuerpo ciliar. Se organiza en 10 capas, y contiene bastones y conos, que son los receptores visuales, más cuatro tipos de neuronas: células bipolares, ganglionares, horizontales y amacrinas (figura 15-2). Los bastones y conos, que están cerca de la coroides, hacen sinapsis con células bipolares, y estas últimas hacen sinapsis con células ganglionares. Los axones de las células ganglionares convergen, y salen del ojo como el nervio óptico. Las células horizontales conectan las células receptoras a las otras células receptoras en la capa plexiforme externa. Las células amacrinas unen las células ganglionares una a otra en la capa plexiforme interna por medio de prolongaciones que tienen longitud y patrones variables. Uniones intercelulares comunicantes (conexiones comunicantes) también conectan las neuronas retinianas una con otra. La capa receptora de la retina descansa sobre el epitelio pigmentario cerca de la coroides, de modo que deben pasar rayos de luz a través de la célula ganglionar y de las capas de células bipolares para alcanzar los bastones y los conos. El epitelio pigmentario absorbe rayos de luz, lo que evita el reflejo de rayos de regreso a través de la retina; esa reflexión produciría imágenes visuales borrosas. El nervio óptico sale del ojo, y los vasos sanguíneos retinianos entran a él, en un punto que se encuentra a 3 mm en posición medial al polo posterior del globo, y un poco por arriba del mismo; esta región es visible a través del oftalmoscopio como el disco óptico. No hay receptores visuales sobre el disco y, en consecuencia, es un punto ciego. Cerca del polo posterior del ojo hay una mancha pigmentada amarillenta, la mancha amarilla (mácula lútea), la cual marca la localización de la fóvea central, una porción adelgazada, sin bastones, de la retina. En ella, los conos están densamente empacados, y cada uno hace sinapsis con una célula bipolar única que, a su vez, hace sinapsis sobre una célula ganglionar única, lo que proporciona una vía directa al cerebro. Hay muy pocas células suprayacentes y no hay vasos sanguíneos; así, la fóvea es el punto donde la agudeza visual es mayor. Cuando la atención es atraída hacia un objeto, o se fija en un objeto, los ojos se mueven de modo que los rayos de luz que llegan desde el objeto caigan en la fóvea. RECEPTORES VISUALES EN LA RETINA Los bastones se encargan de la visión en luz baja (visión nocturna), y sólo proporcionan visión en negro y blanco. La visión a color depende de los conos. Cada bastón y cono se divide en un segmento externo, un segmento interno, que incluye una región nuclear, y una zona sináptica (figura 15-3). Los segmentos externos son cilios modificados y están hechos de pilas regulares de sáculos o discos aplanados compuestos de membrana. Estos sáculos y discos contienen los compuestos fotosensitivos que reaccionan a la luz, e inician potenciales de acción en las vías visuales. Los segmentos internos son ricos en mitocondrias. Los bastones se llaman así por el aspecto delgado, parecido a un bastón, de sus segmentos externos. Los conos, por lo general, tienen segmentos internos gruesos y segmentos externos cónicos, aunque sus características morfológicas varían de un sitio a otro en la retina. En los conos, los sáculos se forman en los segmentos externos mediante pliegues hacia adentro de la membra- CAPÍTULO 15 Sentidos especiales I: visión 135 Epitelio pigmentario Bastón y cono Segmentos externos Segmentos internos Capa nuclear externa C C R C R R C R R Capa plexiforme externa H Capa nuclear interna MB FB FB RB MB RB A A Capa plexiforme interna Capa de células ganglionares MG DG MG DG Fibras del nervio óptico FIGURA 152 Componentes neurales de la porción extrafoveal de la retina. La dirección de la luz es desde la parte inferior hacia la parte superior de la figura. C, cono; R, bastón; MB, RB y FB, células bipolares enanas, bastón y planas; DG y MG, células ganglionares difusas y enanas; H, células horizontales; A, células amacrinas. (Modificada con autorización de Dowling JE, Boycott BB: Organization of the primate retina: electron microscopy. Proc R Soc Lond B 1966;166:80-111.) na celular, pero en los bastones los discos están separados de la membrana celular. En las porciones extrafoveales de la retina predominan los bastones (figura 15-4), y hay bastante convergencia. Las células bipolares planas (figura 15-2) hacen contacto sináptico con varios conos, y las células bipolares de bastón hacen contacto sináptico con varios bastones. Dado que en cada ojo de ser humano hay alrededor de seis millones de conos y 120 millones de bastones, pero en cada nervio óptico sólo hay 1.2 millones de fibras nerviosas, la convergencia general de receptores a través de células bipolares sobre células ganglionares es de alrededor de 105:1. Sin embargo, hay divergencia desde este punto en adelante. Hay dos veces más fibras en los tractos geniculocalcarinos que en los nervios ópticos, y en la corteza visual, el número de neuronas que se relaciona con la visión es 1000 veces el número de fibras en los nervios ópticos. EL MECANISMO DE FORMACIÓN DE IMAGEN Los ojos convierten energía en el espectro visible en potenciales de acción en el nervio óptico. Las imágenes de objetos en el ambiente se enfocan en la retina. Los rayos de luz que llegan a la retina generan potenciales en los bastones y conos. Los impulsos que se inician en la retina son conducidos hacia la corteza cerebral, donde producen la sensación de visión. 136 SECCIÓN IV Fisiología del sistema nervioso central/neural Bastón Cono Membrana plasmática 30 nm Discos Segmento externo Sacos Cuello ciliar Mitocondrias Segmento interno Núcleo Terminal sináptica FIGURA 153 Diagrama esquemático de un bastón y un cono. Cada bastón y cono se divide en un segmento externo, un segmento interno con una región nuclear, y una zona sináptica. Los sáculos y discos en el segmento externo contienen compuestos fotosensitivos que reaccionan a la luz para iniciar potenciales de acción en las vías visuales. (Reproducida con autorización de Lamb TD: Electrical responses of photoreceptors. In: Recent Advances in Physiology, No.10. Baker PF [editor]. Churchill Livingstone, 1984.) Los rayos de luz se desvían cuando pasan desde un medio de una densidad hacia un medio de una densidad diferente, excepto cuando llegan perpendiculares a la interfaz (figura 15-5). El desvío de rayos de luz se llama refracción, y es el mecanismo que permite enfocar una imagen exacta sobre la retina. Rayos de luz paralelos que llegan a un lente biconvexo son refractados hacia un punto detrás del lente. En el ojo, la luz en realidad se refracta en la superficie anterior de la córnea y en las superficies anterior y posterior del cristalino. El proceso de refracción puede representarse en un diagrama al dibujar los rayos de luz como si toda la refracción ocurriera en la superficie anterior de la córnea (figura 15-5). La imagen en la retina está invertida. Las conexiones de los receptores de la retina son tales que desde el nacimiento cualquier imagen invertida sobre la retina se ve en la posición correcta y se proyecta hacia el campo visual en el lado opuesto al área de la retina estimulada; esta percepción es innata y está presente en los lactantes. DEFECTOS COMUNES DEL MECANISMO DE FORMACIÓN DE IMAGEN En algunos individuos, el globo ocular es más corto que lo normal, y los rayos de luz paralelos son llevados a un foco que está detrás de la retina. Esta anormalidad se llama hipermetropía (hiperopía) (figura 15-6). La acomodación sostenida (enfoque debido a contracción del músculo ciliar), aun cuando se ven objetos distantes, puede compensar de manera parcial el defecto, pero el esfuerzo muscular prolongado es agotador y puede causar cefaleas y visión borrosa. El defecto se puede corregir con el uso de anteojos con lentes convexos, que ayudan al poder refractivo del ojo al acortar la distancia focal. En la miopía, el diámetro anteroposterior del globo ocular es demasiado largo (figura 15-6). La forma del ojo parece estar determinada en parte por la refracción presentada a él. En los adultos jóvenes, el trabajo extenso involucrado en actividades como estudiar, acelera la aparición de miopía. Este defecto puede corregirse con anteojos con lentes bicóncavos, que hacen que los rayos de luz paralelos diverjan un poco antes de llegar al ojo. El astigmatismo es un padecimiento común en el cual la curvatura de la córnea no es uniforme. Cuando la curvatura en un meridiano es diferente de la que hay en otros, los rayos de luz en ese meridiano son refractados hacia un foco distinto, de modo que parte de la imagen retiniana es borrosa. El astigmatismo, por lo general, Conos Bastones 1600 1200 800 Punto ciego Número de bastones o conos en un área de 0.0069 mm2 2000 400 0 100° 80° 60° 40° 20° 0° Retina nasal Fóvea Distancia desde la fóvea 20° 40° 60° Retina temporal 80° FIGURA 154 Densidad de bastones y conos a lo largo del meridiano horizontal a través de la retina del ser humano. Un gráfico de la agudeza relativa de la visión en las diversas partes del ojo adaptado a la luz sería paralelo a la curva de densidad de cono; un gráfico similar de la agudeza relativa del ojo adaptado a la oscuridad sería paralelo a la curva de densidad de bastón. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) CAPÍTULO 15 Sentidos especiales I: visión 137 a) Vidrio Aire Refracción Fuente de luz puntiforme Refracción nula Refracción b) b' a a' b FIGURA 155 Enfoque de fuentes de luz puntiformes. a) Cuando rayos de luz divergentes entran en un medio denso a un ángulo a su superficie convexa, la refracción los desvía hacia adentro. b) Refracción de luz por el sistema de lentes. En aras de la sencillez, la refracción sólo se muestra en la superficie corneal (sitio de mayor refracción) aunque también ocurre en el cristalino y en otros sitios. La luz que llega desde a (arriba) y b (abajo) es desviada en direcciones opuestas, lo que hace que b’ esté por arriba de a’ en la retina. (Reproducida con autorización de Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology, 11th ed. McGraw-Hill, 2008.) puede corregirse con lentes cilíndricos colocados de tal manera que igualan la refracción en todos los meridianos. El estrabismo es una alineación inadecuada de los ojos, con frecuencia se debe a problemas con los músculos oculares; es uno de los problemas oculares más comunes en niños; afecta a alrededor de 4% de los menores de seis años. Se caracteriza por giro hacia adentro, afuera, arriba o abajo, de uno o ambos ojos. El estrabismo también suele denominarse “bizquera”. Ocurre cuando las imágenes visuales no caen sobre puntos retinianos correspondientes. Cuando las imágenes visuales caen de manera crónica en puntos que no corresponden en las dos retinas en niños de corta edad, una es suprimida (escotoma de supresión). ACOMODACIÓN Cuando el músculo ciliar está relajado, rayos de luz paralelos que llegan al ojo ópticamente normal (emetrópico) son llevados a un foco en la retina. Mientras se mantenga esta relación, los rayos que provienen de objetos que están a menos de 6 m del observador son llevados a un foco por detrás de la retina y, en consecuencia, los objetos se ven borrosos. El problema de llevar rayos que divergen desde objetos cercanos hasta un foco en la retina puede resolverse al aumentar la curvatura del cristalino, un proceso que se llama acomodación. En reposo, el cristalino lo mantienen bajo tensión los ligamentos del cristalino, y se tira de él para que adopte una forma aplanada. El músculo ciliar se contrae cuando se dirige la vista a un objeto cercano; esto disminuye la distancia entre los bordes del cuerpo ciliar, y relaja los ligamentos del cristalino, de modo que el cristalino adopta una forma más convexa (figura 15-7). El grado al cual la curvatura del cristalino puede aumentarse es limitado, y los rayos de luz provenientes de un objeto muy cerca del individuo no pueden llevarse a un foco en la retina, incluso con el mayor de los esfuerzos. El punto más cercano al ojo en el cual un objeto puede llevarse hacia un foco claro por medio de acomodación se llama punto de visión cercana. Debido a la dureza creciente del cristalino, dicho punto retrocede en el transcurso de la vida, de manera lenta al principio y después con rapidez con la edad, desde 9 cm a los 10 años de edad hasta 83 cm a los 60 años de edad. Para el momento en que un individuo saludable llega a los 40 a 45 años de edad, la pérdida de la acomodación, por lo general, es suficiente para dificultar la lectura y el trabajo de cerca. Esta afección, que se conoce como presbiopía o presbicia, puede corregirse al usar anteojos con lentes convexos. 138 SECCIÓN IV Fisiología del sistema nervioso central/neural a) Vista normal (el objeto lejano está claro) Miopía (globo ocular demasiado largo) Miopía corregida FIGURA 157 Acomodación. Las líneas continuas representan la forma del cristalino, el iris y el cuerpo ciliar en reposo; las líneas discontinuas representan la forma durante la acomodación. Los músculos ciliares se contraen cuando la mirada se dirige a un objeto cercano, lo que disminuye la distancia entre los bordes del cuerpo ciliar y relaja los ligamentos del cristalino, y éste se hace más convexo. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) b) Vista normal (el objeto cercano está claro) Hipermetropía (globo ocular demasiado corto) co. La liberación del transmisor sináptico es constante en la oscuridad. Cuando llega luz al segmento externo, las reacciones que se inician cierran algunos de los canales de Na+, y el resultado es un potencial de receptor hiperpolarizante. La hiperpolarización reduce la liberación del transmisor sináptico, y esto genera una señal en las células bipolares que lleva a potenciales de acción en las células ganglionares. Los potenciales de acción son transmitidos al cerebro. Hipermetropía corregida Na+ FIGURA 156 Defectos comunes del sistema óptico del ojo. a) y b) en la hipermetropía (hiperopía) el globo ocular es demasiado corto y los rayos de luz llegan a un foco detrás de la retina. Un lente convexo corrige esto al sumarse al poder refractivo del cristalino. En la miopía el globo ocular es demasiado largo, y los rayos de luz se enfocan enfrente de la retina. La colocación de un lente bicóncavo enfrente del ojo hace que los rayos de luz diverjan un poco antes de llegar al ojo, de modo que son llevados a un foco sobre la retina. (Reproducida con autorización de K+ K+ Na+ Na+ Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology, 11th ed. McGraw-Hill, 2008.) EL MECANISMO FOTORRECEPTOR BASE IÓNICA DE LOS POTENCIALES DE FOTORRECEPTOR Los canales de Na+ en los segmentos externos de los bastones y conos están abiertos en la oscuridad, de modo que fluye corriente desde el segmento interno hacia el externo (figura 15-8). La corriente también fluye hacia la terminación sináptica del fotorreceptor. La Na+, K+-ATPasa en el segmento interno mantiene el equilibrio ióni- Oscuridad Luz FIGURA 158 Efecto de la luz sobre el flujo de corriente en receptores visuales. En la oscuridad, los canales de Na+ en el segmento externo los mantiene abiertos el cGMP. La luz lleva a incremento de la conversión de cGMP en 5’-GMP, y algunos de los canales se cierran; esto produce hiperpolarización de la terminal sináptica del fotorreceptor. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) CAPÍTULO 15 Sentidos especiales I: visión Luz incidente Cambio estructural en el retineno1 de fotopigmento Cambio conformacional de fotopigmento Membrana del segmento externo Rodopsina cGMP intracelular disminuido Cierre de canales de Na+ Hiperpolarización Liberación disminuida de transmisor sináptico Respuesta en células bipolares y otros elementos neurales FIGURA 159 Secuencia de eventos involucrados en la fototransducción en bastones y conos. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) COMPUESTOS FOTOSENSITIVOS Los compuestos fotosensitivos en los bastones y conos de los ojos están hechos de una proteína que se llama opsina y retinal, el aldehído de la vitamina A. El pigmento fotosensible en los bastones se llama rodopsina, uno de los muchos receptores acoplados a proteínas G; su opsina se conoce como escotopsina. La rodopsina tiene una sensibilidad máxima a la luz a una longitud de onda de 505 nm. En la figura 15-9 se resume la secuencia de eventos en fotorreceptores mediante los cuales la luz incidente produce una señal en la siguiente unidad neural sucesiva en la retina. La luz activa la rodopsina que a continuación activa la proteína G heterotrimérica asociada, la transducina (figura 15-10). La proteína G intercambia GDP por GTP, y la subunidad alfa se separa; esta subunidad permanece activa hasta que su actividad de GTPasa intrínseca hidroliza el GTP. La subunidad alfa activa la cGMP fosfodiesterasa, que convierte cGMP en 5′-GMP. En circunstancias normales, el cGMP actúa de manera directa sobre canales de Na+ para mantenerlos en la posición abierta, de modo que la declinación de la concentración citoplasmática de cGMP hace que algunos canales de Na+ se cierren; esto produce el potencial hiperpolarizante. Esa cascada de reacciones ocurre con mucha rapidez y amplifica la señal de luz. La amplificación ayuda a explicar la notoria sensibilidad de los fotorreceptores de bastones; estos receptores son capaces de producir una respuesta detectable a una cantidad tan pequeña como un fotón de luz. cGMP Transducina fosfodiesterasa Disco Activación de transducina Activación de fosfodiesterasa 139 GTP 5'-GMP cGMP Na+ Luz Canal sensible a cGMP Segmento externo del bastón ECF FIGURA 1510 Pasos iniciales en la fototransducción en bastones. La luz activa la rodopsina, que estimula la transducina para unirse a GTP. Esto activa la fosfodiesterasa, la cual cataliza la conversión de cGMP en 5’-GMP. El decremento resultante de la concentración citoplasmática de cGMP hace que los canales iónicos sensibles a cGMP se cierren. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) Los receptores de cono ayudan a la visión a color y muestran respuesta máxima a la luz a longitudes de onda de 440, 535 y 565 nm. La opsina de cono semeja a la rodopsina. La membrana celular de los conos está invaginada para formar los sáculos, pero los conos no tienen discos intracelulares separados como los que hay en los bastones. Los detalles de las respuestas de los conos a la luz son similares a los que se observan en bastones. PROCESAMIENTO DE LA INFORMACIÓN VISUAL EN LA RETINA Una característica de las células bipolares y ganglionares es que muestran mejor respuesta a estímulos pequeños, circulares y que, dentro de su campo receptivo, un anillo de luz alrededor del centro (iluminación de la periferia) inhibe la respuesta al punto central (figura 15-11). El centro puede ser excitador con un borde inhibitorio (una célula “con centro encendido”) o inhibitorio con periferia excitadora (una célula con “centro apagado”). La inhibición de la respuesta del centro por la periferia quizá se debe a retroacción inhibitoria desde un fotorreceptor a otro mediada por medio de células horizontales. La inhibición de la respuesta a la iluminación central por un aumento de la iluminación periférica es un ejemplo de inhibición lateral, en la cual la activación de una unidad neural particular se asocia con inhibición de la actividad de unidades cercanas. Es un fenómeno general en sistemas sensoriales, y ayuda a agudizar los bordes de un estímulo y mejorar la discriminación. VÍAS VISUALES Los axones de las células ganglionares retinianas pasan en el nervio óptico y el tracto óptico para terminar en el cuerpo geniculado lateral en el tálamo (figura 15-12). Las fibras de cada hemirretina 140 SECCIÓN IV Fisiología del sistema nervioso central/neural Campo de centro encendido Campo de centro apagado Luz Luz Iluminación central Iluminación periférica FIGURA 1511 Respuestas de células ganglionares retinianas a la luz en las porciones de sus campos receptivos indicados en blanco. A un lado de cada diagrama de campo receptivo hay un diagrama de la respuesta de la célula ganglionar, indicada por potenciales de acción registrados fuera de la célula. Note que en tres de las cuatro situaciones, hay descarga aumentada cuando se apaga la luz. (Adaptada con autorización de Kuffler SW: Discharge patterns and functional organizations of mammalian retina, J Neurophysiol. 1953;16(1):37-68.) Campo temporal Campo nasal IZQUIERDO DERECHO A Ojo izquierdo Ojo derecho B Célula ganglionar B A C Nervio óptico Quiasma óptico Región pretectal Tracto óptico C D Cuerpo geniculado lateral Tracto geniculocalcarino D Corteza occipital FIGURA 1512 Vías visuales. La transección de las vías en los sitios indicados por las letras causa los defectos de campo visual que se muestran en los diagramas a la derecha. Las fibras de la mitad nasal de cada retina se decusan en el quiasma óptico, de modo que las fibras en los tractos ópticos son las provenientes de la mitad temporal de una retina y la mitad nasal de la otra. Una lesión que interrumpe un nervio óptico causa ceguera en ese ojo (A). Una lesión en un tracto óptico causa ceguera en la mitad del campo visual (C) y se llama hemianopsia (media ceguera) homónima (mismo lado de ambos campos visuales). Las lesiones que afectan el quiasma óptico destruyen fibras provenientes de ambas hemirretinas nasales y producen una hemianopsia heterónima (lados opuestos de los campos visuales) (B). Las lesiones occipitales pueden respetar las fibras que provienen de la mácula (como en D) por la separación en el cerebro de estas fibras de las otras que ayudan a la visión (figura 15-13). (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) CAPÍTULO 15 Sentidos especiales I: visión 141 Cuadrante periférico superior de la retina Cuadrante superior de la mácula Nervios ópticos Quiasma óptico Cuadrante inferior de la mácula Cuadrante periférico inferior de la retina Núcleo geniculado lateral C Dorsal I Tractos ópticos FIGURA 513 Vista medial del hemisferio cerebral derecho del ser humano, que muestra la proyección de la retina sobre la corteza visual primaria (área de Brodmann 17; también conocida como V1) en la corteza occipital alrededor de la cisura calcarina. Las fibras geniculocalcarinas que se originan de la mitad medial del geniculado lateral terminan en el labio superior de la cisura calcarina, y las que provienen de la mitad lateral finalizan en el labio inferior. Las fibras provenientes del cuerpo geniculado lateral que transmiten visión macular se separan de las que transmiten visión periférica, y terminan en posición más posterior sobre los labios de la cisura calcarina. (Reproducida C I I C 5 Ventral 1 2 Vía magnocelular 3 6 4 Vía parvocelular con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) Corteza visual primaria (área 17) nasal se decusan en el quiasma óptico. En el cuerpo geniculado, las fibras que provienen de la mitad nasal de una retina, y la mitad temporal de la otra, hacen sinapsis sobre las células cuyos axones forman el tracto geniculocalcarino; este tracto pasa hacia el lóbulo occipital de la corteza cerebral. Los efectos de lesiones en estas vías sobre la función visual se comentan más adelante. El área receptora visual primaria (corteza visual primaria, área de Brodmann 17; también conocida como V1) está situada a los lados de la cisura calcarina (figura 15-13). La organización de la corteza visual primaria se comenta más adelante. Los axones de células ganglionares retinianas proyectan una representación espacial detallada de la retina en el cuerpo geniculado lateral. Cada cuerpo geniculado contiene seis capas bien definidas (figura 15-14). Las capas 3 a 6 tienen células parvocelulares pequeñas, y las capas 1 y 2 tienen células magnocelulares grandes. En cada lado, las capas 1, 4 y 6 reciben aferencias que provienen del ojo contralateral; las capas 2, 3 y 5 reciben aferencias que se derivan del ojo ipsilateral. En cada capa, hay una representación punto por punto precisa de la retina, y las seis capas están en registro, de modo que a lo largo de una línea perpendicular a las capas, los campos receptivos de las células en cada capa son casi idénticos. Sólo 10 a 20% de las aferencias hacia el núcleo geniculado lateral procede de la retina; aferencias importantes también se originan de la corteza visual y otras regiones del cerebro. La vía de retroacción desde la corteza visual está involucrada en el procesamiento visual que se relaciona con la percepción de orientación y movimiento. Hay dos clases de células ganglionares retinianas: células magno o M grandes, que se relacionan con el movimiento y la estereopsis, y células parvo o P pequeñas, que están relacionadas con el color, la textura y la forma. Las células ganglionares M y P se proyectan hacia las porciones magnocelular y parvocelular del geniculado lateral, respectivamente. Desde el núcleo geniculado lateral, una vía magnocelular y una vía parvocelular se proyectan hacia la corteza visual. La FIGURA 1514 Proyecciones de célula ganglionar desde la hemirretina derecha de cada ojo hacia el cuerpo geniculado lateral derecho y desde este núcleo hacia la corteza visual primaria derecha. Advierta las seis capas del cuerpo geniculado. Las células ganglionares P se proyectan hacia las capas 3 a 6, y las células ganglionares M, hacia las capas 1 y 2. Los ojos ipsilateral (I) y contralateral (C) se proyectan hacia capas alternas. No se muestran las células del área interlaminar, que se proyectan por medio de un componente separado de la vía P hacia grumos en la corteza visual. (Modificada con autorización de Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000.) vía magnocelular, desde las capas 1 y 2, lleva señales para la detección de movimiento, profundidad y titileo. La vía parvocelular, de las capas 3 a 6, lleva señales para la visión a color, textura, forma y detalle fino. Las células en la región interlaminar del núcleo geniculado lateral también reciben aferencias provenientes de células ganglionares P, quizá por medio de dendritas de células interlaminares que penetran en las capas parvocelulares. Se proyectan por medio de un componente separado de la vía P a los “grumos” en la corteza visual; son agrupaciones de células de alrededor de 0.2 mm de diámetro que, a diferencia de las células vecinas, contienen una concentración alta de la enzima mitocondrial citocromo oxidasa. CORTEZA VISUAL PRIMARIA El cuerpo geniculado lateral proyecta una representación punto por punto sobre la corteza visual primaria (figura 15-13). Al igual que el resto de la neocorteza, la corteza visual tiene seis capas. Los axones del núcleo geniculado lateral que forman la vía magnocelular terminan en la capa 4. Muchos de los axones que forman la vía parvocelu- 142 SECCIÓN IV Fisiología del sistema nervioso central/neural lar también terminan en la capa 4; empero, los axones que provienen de la región interlaminar terminan en las capas 2 y 3. Las capas 2 y 3 de la corteza contienen grumos; están dispuestos en un mosaico en la corteza visual y se relacionan con la visión a color. Con todo, la vía parvocelular también lleva datos oponentes a color a la parte profunda de la capa 4. Al igual que las células ganglionares, las neuronas del geniculado lateral y las neuronas en la capa 4 de la corteza visual muestran respuesta a estímulos en sus campos receptivos con centros encendidos y periferia inhibidora, o centros apagados y periferia excitadora. Una barra de luz que cubre el centro es un estímulo eficaz para ellas porque estimula todo el centro y relativamente poco de la periferia. Aun así, la barra no tiene orientación preferida y, como un estímulo, es igual de eficaz a cualquier ángulo. Las respuestas de las neuronas en otras capas de la corteza visual son a todas luces distintas. Las células simples muestran respuesta a barras de luz, líneas o bordes, pero sólo cuando tienen una orientación particular. Cuando una barra de luz es rota tan poco como 10° desde la orientación preferida, la tasa de activación de la célula simple por lo general disminuye, y si el estímulo se rota mucho más, la respuesta desaparece. Las células complejas, que semejan células simples por cuanto requieren una orientación preferida de un estímulo lineal, son menos dependientes de la localización de un estímulo en el campo visual que las células simples y que las células en la capa 4. Con frecuencia muestran respuesta de manera máxima cuando un estímulo lineal es movido en dirección lateral sin un cambio de su orientación. Pueden recibir aferencias provenientes de las células simples. La corteza visual está dispuesta en columnas verticales que se relacionan con la orientación (columnas de orientación). Cada una mide alrededor de 1 mm de diámetro. De cualquier modo, las preferencias de orientación de columnas vecinas difieren de una manera sistemática; al moverse de una columna a otra a través de la corteza, ocurren cambios secuenciales en la preferencia de orientación, de 5° a 10°. Así, parece probable que para cada campo receptivo de célula ganglionar en el campo visual, hay un conjunto de columnas en un área pequeña de la corteza visual que representan las posibles orientaciones preferidas a intervalos pequeños en los 360° completos. Las células simples y complejas se llaman detectores de característica porque muestran respuesta a ciertas características del estímulo y las analizan. Otra particularidad de la corteza visual es la presencia de columnas de dominancia ocular. Las células geniculadas y las células en la capa 4 reciben aferencias que provienen de sólo un ojo, y las células de la capa 4 alternan con células que reciben aferencias desde el otro ojo. Alrededor de la mitad de las células simples y complejas reciben una aferencia proveniente de ambos ojos. Las aferencias son idénticas o casi idénticas en términos de la porción del campo visual involucrado y la orientación preferida. No obstante, difieren en fuerza, de modo que entre las células a las cuales llega la aferencia desde el ojo ipsilateral o el contralateral, hay un espectro de células influidas en diferentes grados por ambos ojos. EFECTO DE LESIONES EN LAS VÍAS ÓPTICAS Las lesiones a lo largo de las vías ópticas desde los ojos hasta el cerebro pueden localizarse con un alto grado de exactitud por los efectos que producen en los campos visuales (figura 15-12). Las fibras que se producen de la mitad nasal de cada retina se decusan en el quiasma óptico, de modo que las fibras en los tractos ópticos son las de la mitad temporal de una retina y la mitad nasal de la otra. Puesto que cada tracto óptico origina la mitad del campo de visión, una lesión de un nervio óptico causa ceguera en ese ojo, pero una lesión en un tracto óptico provoca ceguera en la mitad del campo visual. Este defecto se clasifica como hemianopsia (media ceguera) homónima (mismo lado de ambos campos visuales). Las lesiones que afectan el quiasma óptico (p. ej., tumores hipofisarios) causan alteración de las fibras provenientes de ambas hemirretinas nasales, y producen una hemianopsia heterónima (lados opuestos de los campos visuales). Puesto que las fibras que provienen de las máculas se localizan en posición posterior en el quiasma óptico, aparecen escotomas hemianópsicos antes de que la visión en las dos hemirretinas se pierda por completo. Los defectos selectivos del campo visual se subclasifican como bitemporales, binasales y derecho o izquierdo. Las fibras del nervio óptico que se derivan de los cuadrantes superiores de la retina, los cuales ayudan a la visión en la mitad inferior del campo visual, terminan en la mitad medial del cuerpo geniculado lateral, mientras que las fibras provenientes de los cuadrantes inferiores de la retina finalizan en la mitad lateral. Las fibras geniculocalcarinas que se originan en la mitad medial del geniculado lateral terminan en el labio superior de la cisura calcarina, y las que provienen de la mitad lateral finalizan en el labio inferior. Las fibras provenientes del cuerpo geniculado lateral que ayudan a la visión macular se separan de las que ayudan a la visión periférica y terminan en posición más posterior en los labios de la cisura calcarina (figura 15-13). Debido a su disposición anatómica, las lesiones en el lóbulo occipital pueden producir defectos de campo visual cuadránticos separados (cuadrantes superior e inferior de cada medio campo visual). La preservación macular (esto es, pérdida de la visión periférica con visión macular intacta) también es común en presencia de lesiones occipitales (figura 15-12) porque la representación macular es separada de la de los campos periféricos y muy grande en comparación con la de estos últimos campos. Por ende, las lesiones occipitales deben extenderse distancias considerables para destruir la visión macular, así como la periférica. La destrucción bilateral de la corteza occipital en seres humanos causa ceguera subjetiva. VISIÓN A COLOR Los colores tienen tres atributos: tono, intensidad y saturación (grado de libertad de dilución con blanco). Para cualquier color hay un color complementario que, cuando se mezcla de manera apropiada con él, produce una sensación de blanco. El negro es la sensación producida por la ausencia de luz, pero quizá es una sensación positiva porque el ojo ciego no “ve negro”; más bien, “ve nada”. La sensación de blanco, cualquier color espectral e incluso el color extraespectral, púrpura, puede producirse al mezclar diversas proporciones de luz roja (longitud de onda de 723 a 647 nm), luz verde (575 a 492 nm), y luz azul (492 a 450 nm); por ende, el rojo, verde y azul se llaman colores primarios. Asimismo, el color que se percibe depende, en parte, del color de los otros objetos en el campo visual. De este modo, por ejemplo, un objeto rojo se observa como rojo si el campo está iluminado con luz verde o azul, pero como rosado claro o blanco si el campo está iluminado con luz roja. CAPÍTULO 15 Sentidos especiales I: visión El color está mediado por células ganglionares que restan o suman aferencias provenientes de un tipo de cono a las aferencias que provienen de otro tipo. El procesamiento en las células ganglionares y el núcleo geniculado lateral produce impulsos que pasan a lo largo de tres tipos de vías neurales, las cuales se proyectan a V1: una vía rojaverde que señala diferencias entre respuestas de cono L y M, una vía azul-amarilla que señala diferencias entre respuestas de cono S y la suma de respuestas de cono L y M, y una vía de luminancia que señala la suma de respuestas de conos L y M; estas vías se proyectan hacia los grumos y la porción profunda de la capa 4 de V1. Desde los grumos y la capa 4, la información sobre color se proyecta hacia V8. Sin embargo, se desconoce cómo V8 convierte las aferencias de color en la sensación de color. La ceguera al color es con frecuencia una afección hereditaria en la cual los individuos son incapaces de distinguir ciertos colores. El tipo más común es un déficit de visión de color rojo-verde, una afección ligada con el sexo por mecanismos genéticos que ocurren en alrededor de 8% de los varones y 0.4% de las mujeres. Los déficits de visión de color azul-amarillo son menos comunes, y no muestran selectividad de género. La ceguera al color, por lo general, se debe a una falta hereditaria de conos para colores específicos. También puede ocurrir en individuos con lesiones del área V8 de la corteza visual. REFLEJO LUMINOSO PUPILAR Cuando se dirige luz a un ojo, la pupila se constriñe (reflejo luminoso pupilar). Las fibras del nervio óptico que llevan los impulsos que inician estas respuestas pupilares salen de los nervios ópticos cerca de los cuerpos geniculados laterales. A cada lado, entran en el 143 mesencéfalo por medio del brazo del colículo superior, y terminan en el núcleo pretectal. Desde este núcleo, las neuronas de segundo orden se proyectan hacia el núcleo de Edinger-Westphal ipsilateral y contralateral. Las neuronas de tercer orden pasan desde este núcleo hacia el ganglio ciliar en el nervio oculomotor, y las neuronas de cuarto orden pasan desde este ganglio hacia el cuerpo ciliar. MOVIMIENTOS OCULARES El ojo es movido dentro de la órbita por seis músculos oculares (figura 15-15) inervados por los nervios oculomotor, troclear y abducens (craneales). Dado que los músculos oblicuos tiran en dirección medial, sus acciones varían con la posición del ojo. Cuando el ojo se gira en dirección nasal, el oblicuo inferior lo eleva, y el oblicuo superior lo deprime. Cuando se gira en dirección lateral, el recto superior lo eleva y el recto inferior lo deprime. Dado que gran parte del campo visual es binocular, se necesita un muy alto orden de coordinación de los movimientos de los dos ojos para que las imágenes visuales caigan en todo momento en puntos correspondientes en las dos retinas y para que se evite diplopía (visión doble). Hay cuatro tipos de movimientos oculares, cada uno controlado por un sistema neural diferente, pero que comparte la misma vía común final, las neuronas motoras que inervan los músculos oculares externos. Las sacadas, movimientos con sacudidas repentinos, ocurren conforme la mirada se desvía desde un objeto hacia otro. Llevan nuevos objetos de interés sobre la fóvea, y reducen la adaptación en la vía visual que ocurriría si la mirada se fijara en un objeto único durante periodos prolongados. Los movimientos de búsqueda sua- FIGURA 1515 Músculos extraoculares que se encargan de las seis posiciones cardinales de la mirada. El ojo es puesto en aducción por el recto medial y en abducción por el recto lateral. El ojo en aducción es elevado por el oblicuo inferior y deprimido por el oblicuo superior; el ojo en abducción es elevado por el recto superior y deprimido por el recto inferior. (Reproducida con autorización de Squire LR, et al. [editors]: Fundamental Neuroscience, 3rd ed. Academic Press, 2008.) 144 SECCIÓN IV Fisiología del sistema nervioso central/neural ves son movimientos de los ojos de seguimiento conforme siguen objetos que se mueven. Los movimientos vestibulares, ajustes que ocurren en respuesta a estímulos iniciados en los canales semicirculares, mantienen la fijación visual a medida que la cabeza se mueve. Los movimientos de convergencia llevan los ejes visuales uno hacia otro conforme se enfoca la atención en objetos cerca del observador. Queda de manifiesto la similitud con un sistema artificial de seguimiento sobre una plataforma inestable, como un barco: los movimientos sacádicos buscan blancos visuales, los movimientos de búsqueda los siguen a medida que se mueven, y los movimientos vestibulares estabilizan el dispositivo de rastreo conforme se mueve la plataforma en la cual el dispositivo está montado (esto es, la cabeza). Las sacadas son programadas en la corteza frontal y los colículos superiores, y siguen movimientos en el cerebelo. CORRELACIÓN CLÍNICA Un vendedor de 33 años de edad conducía su automóvil en un área rural y no se percató de la ciclista que iba en la acera a la derecha de su automóvil. Dio vuelta a la derecha en la esquina, y por accidente golpeó a la joven; por fortuna, la mujer no quedó lesionada, pero él recordó que algunos días antes mientras caminaba por una acera, no notó un perro que se le acercaba desde la izquierda. Ahora está consciente de que tiene visión periférica reducida. Hace una cita con un oftalmólogo para un examen de campo visual. Los resultados mostraron que tiene visión reducida en la mitad temporal del campo visual de ambos ojos. Cuando se le preguntó si había notado una incidencia aumentada de cefaleas recientemente, dijo que sí. El paciente había creído que las cefaleas se debían a estrés reciente en su empleo. Los resultados del examen de campo visual indicaron hemianopsia bitemporal (un tipo de hemianopsia heterónima); esto significa daño en el quiasma óptico, el cual lleva axones de células ganglionares de las mitades nasales de la retina, que transmiten información visual desde los campos visuales temporales. Una causa común de este defecto es un tumor de la hipófisis, como un adenoma hipofisario. La glándula hipófisis se sitúa en posición ventral al quiasma óptico. Cuando el tumor crece, ejerce presión sobre el quiasma óptico. A menudo ocurren cefaleas y lívido disminuida como resultado del tumor que típicamente es benigno. Una resonancia magnética de la hipófisis en este paciente reveló un tumor que ejercía presión sobre el quiasma óptico. El tratamiento médico hizo que el tumor disminuyera de tamaño, y los campos visuales se restituyeron a lo normal. (En el capítulo 62 se presentan más detalles acerca de tumores de la parte anterior de la hipófisis.) RESUMEN DEL CAPÍTULO ■ ■ Las principales partes del ojo son la esclerótica (cubierta protectora), la córnea (transferencia de rayos de luz), la coroides (nutrición), retina (células receptoras), cristalino e iris. La desviación de rayos de luz (refracción) permite enfocar una imagen exacta sobre la retina. La luz es refractada en la superficie ■ ■ ■ ■ ■ anterior de la córnea y en las superficies anterior y posterior del cristalino. Para llevar rayos que divergen desde objetos cercanos hacia un foco en la retina, se aumenta la curvatura del cristalino, un proceso que se llama acomodación. En la hipermetropía (hiperopía), el globo ocular es demasiado corto y los rayos de luz llegan a un foco detrás de la retina. En la miopía, el diámetro anteroposterior del globo ocular es muy largo. El astigmatismo es una afección común en la cual la curvatura de la córnea no es uniforme. La presbiopía es la pérdida de la acomodación para la visión cercana. El estrabismo es una alineación inadecuada de los ojos, por lo general, se debe a problemas con los músculos oculares. Los canales de Na+ en los segmentos externos de los bastones y conos están abiertos en la oscuridad, de modo que fluye corriente desde el segmento interno hacia el externo. Cuando la luz incide en el segmento exterior, algunos de los canales de Na+ se cierran, y las células son hiperpolarizadas. Las neuronas en la capa 4 de la corteza visual muestran respuesta a estímulos en sus campos receptivos con centros encendidos y periferia inhibidora o centros apagados y periferia excitadora. Las neuronas en otras capas se llaman células simples si muestran respuesta a barras de luz, líneas, o bordes, pero sólo cuando tienen una orientación particular. Las células complejas también requieren una orientación preferida de un estímulo lineal, pero dependen menos de la ubicación de un estímulo en el campo visual. La vía visual va de los bastones y conos a células bipolares, a las células ganglionares, y luego a través del tracto óptico al cuerpo geniculado lateral del tálamo, en el lóbulo occipital de la corteza cerebral. Las fibras provenientes de cada hemirretina nasal se decusan en el quiasma óptico; las fibras de la mitad nasal de una retina y la mitad temporal de la otra hacen sinapsis sobre las células cuyos axones forman el tracto geniculocalcarino. Las sacadas (movimientos con sacudidas súbitos) ocurren a medida que la mirada se desvía de un objeto a otro, y reducen la adaptación en la vía visual que ocurriría si se fijara la mirada en un objeto único durante periodos prolongados. Los movimientos de búsqueda suaves son de rastreo de los ojos conforme siguen objetos en movimiento. Los movimientos vestibulares ocurren en respuesta a estímulos en los canales semicirculares para mantener la fijación visual a medida que se mueve la cabeza. Los movimientos de convergencia llevan los ejes visuales uno hacia otro conforme la atención se enfoca en objetos cerca del observador. PREGUNTAS DE ESTUDIO 1. Un examen visual en un varón de 80 años de edad muestra que tiene capacidad reducida para ver objetos en los cuadrantes superior e inferior de los campos visuales izquierdos de ambos ojos, pero queda algo de visión en las regiones centrales del campo visual. El diagnóstico es: A) escotoma central. B) hemianopsia heterónima con preservación macular. C) lesión del quiasma óptico. D) hemianopsia homónima con preservación macular. E) retinopatía. 2. La acomodación visual comprende: A) tensión aumentada sobre los ligamentos del cristalino. B) un decremento de la curvatura del cristalino. C) relajación del músculo esfínter del iris. CAPÍTULO 15 Sentidos especiales I: visión D) contracción del músculo ciliar. E) presión intraocular aumentada. 3. La fóvea del ojo: A) tiene el umbral de luz más bajo. B) es la región de mayor agudeza visual. C) sólo contiene conos. D) sólo contiene bastones. E) está situada sobre la cabeza del nervio óptico. 4. ¿Cuál de las partes que siguen del ojo tiene la mayor concentración de bastones? A) cuerpo ciliar B) iris C) disco óptico D) fóvea E) región extrafoveal 5. La secuencia de eventos correcta involucrada en la fototransducción en bastones y conos en respuesta a la luz es: A) activación de la transducina, decremento de la liberación de glutamato, cambios estructurales de la rodopsina, cierre de canales de Na+, y disminución del cGMP intracelular. B) liberación disminuida de glutamato, activación de transducina, cierre de canales de Na+, disminución del cGMP intracelular, y cambios estructurales en la rodopsina. 145 C) cambios estructurales en la rodopsina, decremento del cGMP intracelular, liberación disminuida de glutamato, cierre de canales de Na+ y activación de transducina. D) cambios estructurales en la rodopsina, activación de la transducina, decremento del cGMP intracelular, cierre de los canales de Na+ y liberación disminuida de glutamato. E) activación de transducina, cambios estructurales en la rodopsina, cierre de los canales de Na+, decremento del cGMP intracelular, y liberación disminuida de glutamato. 6. La vitamina A es un precursor para la síntesis de: A) transducina. B) retinales. C) el pigmento del iris. D) escotopsina. E) humor acuoso. 7. ¿Cuál de los que siguen no está involucrado en la visión a color? A) activación de una vía que señala diferencias entre las respuestas de cono S y la suma de las respuestas de conos L y M B) capas geniculadas 3 a 6 C) vía P D) área V3A de la corteza visual E) área V8 de la corteza visual Sentidos especiales II: audición y equilibrio C A P Í T U L O 16 Susan M. Barman O B J E T I V O S ■ ■ ■ ■ ■ ■ ■ Describir los componentes de los oídos externo, medio e interno y sus funciones. Describir la manera en que los movimientos de moléculas en el aire se convierten en impulsos generados en células ciliadas en la cóclea. Trazar la trayectoria de impulsos auditivos en las vías neurales desde las células ciliadas cocleares hasta la corteza auditiva y comentar la función de esta última. Explicar cómo el tono y el volumen se codifican en las vías auditivas. Describir las diversas formas de sordera y las pruebas para su diagnóstico. Explicar cómo los receptores en los canales semicirculares detectan aceleración rotacional, y cómo los receptores en el sáculo y el utrículo detectan aceleración lineal. Listar las principales aferencias sensoriales que proporcionan la información que se sintetiza en el cerebro hacia el sentido de posición en el espacio. INTRODUCCIÓN Los receptores para la audición y el equilibrio se encuentran en el oído. El oído externo, el oído medio y la cóclea del oído interno se relacionan con la audición. Los canales semicirculares, el utrículo y el sáculo del oído interno se relacionan con el equilibrio. Los receptores en los canales semicirculares (células ciliadas) detectan aceleración rotacional, los receptores en el utrículo detectan aceleración lineal en la dirección horizontal, y los receptores en el sáculo detectan aceleración lineal en la dirección vertical. CARACTERÍSTICAS ANATÓMICAS DEL OÍDO OÍDOS EXTERNO Y MEDIO hacia la membrana timpánica (tímpano). El oído medio es una cavidad llena de aire en el hueso temporal que se abre por medio de la tuba auditiva (trompa de Eustaquio) hacia la nasofaringe, y a través de esta última hacia el exterior. La tuba, por lo general, está cerrada, pero durante la deglución, la masticación y los bostezos se abre, e iguala la presión de aire a ambos lados del tímpano. Los tres osículos o huesecillos del oído (martillo, yunque y estribo) están en el oído medio (figura 16-2). El manubrio (mango del martillo) está fijo a la parte posterior de la membrana timpánica. Su cabeza está fija a la pared del oído medio, y su prolongación corta está fija al yunque, que se articula con la cabeza del estribo. La base del estribo está fija mediante un ligamento anular a las paredes de la ventana oval. Dos músculos esqueléticos pequeños (tensor del tímpano y estapedio) se ubican en el oído medio. La contracción del primero tira del manubrio del martillo en dirección medial y disminuye las vibraciones de la membrana timpánica; la contracción del segundo tira de la base del estribo para separarla de la ventana oval. El oído externo canaliza ondas de sonido hacia el meato auditivo externo (figura 16-1). Las ondas de sonido pasan hacia adentro, 147 148 SECCIÓN IV Fisiología del sistema nervioso central/neural Oído externo Oído medio Oído interno Canales semicirculares Nervio facial Hélix Nervio vestibular Nervio coclear Pabellón de la oreja Cóclea Hueso temporal Ventana redonda Meato auditivo externo Cavidad timpánica Tuba auditiva Osículos del oído Lóbulo de la oreja Membrana timpánica FIGURA 161 Estructuras de las porciones externa, media e interna del oído humano. En aras de la claridad, la cóclea se giró un poco, y los músculos del oído medio se omitieron. (Reproducida con autorización de Fox SI: Human Physiology. McGraw-Hill, 2008.) Pirámide Hueso temporal Ático timpánico Músculo estapedio Tendón del músculo estapedio Osículos: Martillo Tendón del músculo tensor del tímpano Yunque Estribo Ventana oval Tendón del músculo estapedio Ventana redonda Pirámide Músculo tensor del tímpano Membrana timpánica Cavidad timpánica Tuba auditiva (trompa de Eustaquio) FIGURA 162 Vista medial del oído medio. Se indican las ubicaciones de los músculos auditivos fijos a los osículos del oído medio (Reproducida con autorización de Fox SI: Human Physiology. McGraw-Hill, 2008.) CAPÍTULO 16 Sentidos especiales II: audición y equilibrio OÍDO INTERNO Y CÓCLEA El oído interno (laberinto) consta de dos partes, una dentro de la otra. El laberinto óseo es una serie de canales en el hueso temporal. Dentro de estos canales, rodeado por un líquido (perilinfa), se localiza el laberinto membranoso (figura 16-3) que está lleno con un líquido rico en K+ (endolinfa). No hay comunicación entre los espacios llenos con endolinfa y los que están llenos con perilinfa. La porción coclear del laberinto es un tubo en espiral que, en seres humanos, tiene 35 mm de largo y realiza alrededor de 2.75 vueltas. La membrana basilar y la membrana de Reissner se dividen en tres cámaras o escalas (figura 16-4). La escala vestibular superior y la escala timpánica inferior contienen perilinfa y se comunican entre sí en el vértice de la cóclea por medio de una abertura pequeña (helicotrema). En la base de la cóclea, la escala vestibular termina en la ventana oval, que es cerrada por la base del estribo. La escala timpánica termina en la ventana redonda, un agujero en la pared medial del oído medio que es cerrado por la membrana timpánica secundaria flexible. La escala media es continua con el laberinto membranoso, y no se comunica con las otras dos escalas. 149 El órgano de Corti contiene los receptores auditivos (células ciliadas) cuyas prolongaciones perforan la lámina reticular que se apoya en las células pilares o bastones de Corti (figura 16-4). Las células ciliadas están dispuestas en cuatro hileras: tres hileras de células ciliadas externas en posición lateral al túnel formado por los bastones de Corti, y una hilera de células ciliadas internas en posición medial al túnel. La membrana tectorial cubre las hileras de células ciliadas; las puntas de los pelos de las células externas están embebidas en esta membrana. Los cuerpos celulares de las neuronas sensoriales se ubican en el ganglio espiral dentro del modiolo; ~95% de estas neuronas sensoriales inerva células ciliadas internas, ~5% inerva células ciliadas externas, y cada neurona sensorial inerva varias células ciliadas externas. En contraste, casi todas las fibras eferentes en el nervio auditivo terminan en las células ciliadas externas. Los axones de neuronas aferentes que inervan células ciliadas forman la división auditiva (coclear) del octavo par craneal. Los canales semicirculares están orientados en los tres planos. Dentro de los canales óseos, los canales membranosos están suspendidos en la perilinfa. Una estructura receptora (cresta ampollar) se sitúa en el extremo expandido (ampolla) de cada uno de los canales Cúpula Canal semicircular VIII Membrana tectorial Membrana basilar Membrana otolítica Cóclea Sáculo FIGURA 163 Esquema del oído interno humano que muestra el laberinto membranoso con agrandamientos de las estructuras en las cuales están embebidas las células ciliadas. El laberinto membranoso está suspendido en perilinfa y lleno con endolinfa rica en K+ que baña los receptores. Las células ciliadas (oscurecidas para resaltarlas) están en diferentes disposiciones características de los órganos receptores. Los canales semicirculares son sensibles a la aceleración angular que desvía la cúpula gelatinosa y las células ciliadas asociadas. En la cóclea, las células ciliadas hacen una espiral a lo largo de la membrana basilar en el órgano de Corti. Los sonidos transportados por el aire ponen en movimiento el tímpano, lo cual se transmite a la cóclea por los huesos del oído medio. Esto flexiona la membrana hacia arriba y hacia abajo. Las células ciliadas en el órgano de Corti son estimuladas por movimiento de corte. Los órganos otolíticos (sáculo y utrículo) son sensibles a la aceleración lineal en los planos vertical y horizontal. Las células ciliadas están fijas a la membrana otolítica. VIII, octavo par craneal, con divisiones auditiva y vestibular. (Adaptada con autorización de Hudspeth AJ. How the ear’s works work. Nature.1989;341:397. Copyright 1989 by Macmillan Magazines.) 150 SECCIÓN IV Fisiología del sistema nervioso central/neural Estría vascular Escala vestibular Membrana de Reissner Membrana tectorial Ligamento espiral Escala media Limbo Prominencia espiral Ganglio espiral Escala timpánica Ligamento espiral Modiolo Lámina espiral Órgano de Corti Células ciliadas externas Lámina reticular Membrana tectorial Célula ciliada interna IPC Membrana basilar CD Arco RECEPTORES AUDITIVOS: CÉLULAS CILIADAS Las células ciliadas en el órgano de Corti emiten señales auditivas, las células ciliadas en el utrículo producen señales de aceleración horizontal, las células ciliadas en el sáculo emiten señales de aceleración vertical, y un parche en cada uno de los tres canales semicirculares produce señales de aceleración rotacional. Estas células ciliadas tienen una estructura común (figura 16-5). Cada una está embebida en un epitelio constituido de células de sostén, con el extremo basal en contacto estrecho con neuronas aferentes. Hay 30 a 150 prolongaciones o cilios en forma de bastón que se proyectan desde el extremo apical. Excepto en la cóclea, uno de éstos, el cinocilio, es un cilio verdadero, pero no móvil, con nueve pares de microtúbulos alrededor de su circunferencia y un par central de microtúbulos. Es una de las prolongaciones de mayor tamaño, y tiene un extremo en maza. El cinocilio se pierde de las células ciliadas de la cóclea en adultos; sin embargo, las otras prolongaciones (estereocilios) se encuentran en todas las células ciliadas. Tienen centros compuestos de filamentos paralelos de actina que está cubierta con isoformas de miosina. Dentro de la agrupación de prolongaciones en cada célula hay una estructura ordenada. A lo largo de un eje hacia el cinocilio, la altura de los estereocilios aumenta poco a poco; a lo largo del eje perpendicular, todos los estereocilios tienen la misma altura. Túnel RESPUESTAS ELÉCTRICAS Fibras nerviosas Habénula perforada Célula pilar Membrana (bastón de Corti) basilar Lámina espiral FIGURA 164 Arriba: corte transversal de la cóclea, que muestra el órgano de Corti y las tres escalas de la cóclea. Abajo: estructura del órgano de Corti, como aparece en la vuelta basal de la cóclea. CD, células falángicas externas (células de Deiters) que apoyan las células ciliadas externas; IPC, célula falángica interna que apoya la célula ciliada interna. (Reproducida con autorización de Pickels JO: An Introduction to the Physiology of Hearing, 2nd ed. Academic Press, 1988.) membranosos. Cada cresta consta de células ciliadas y células de sostén (sustentaculares) rematadas por una partición gelatinosa (cúpula), que encierra la ampolla (figura 16-3). Las prolongaciones de las células ciliadas están embebidas en la cúpula, y la base de las células ciliadas tiene contacto con las fibras aferentes de la división vestibular del octavo par craneal. Dentro de cada laberinto membranoso hay un órgano otolítico (mácula). Otra mácula se localiza en la pared del sáculo en una posición semivertical. Las máculas contienen células de sostén y células ciliadas, rematadas por una membrana otolítica en la cual están embebidos cristales de carbonato de calcio, los otolitos (figura 16-3), que también se llaman otoconias o polvo auditivo. Las prolongaciones de las células ciliadas están embebidas en la membrana. Las fibras nerviosas que provienen de las células ciliadas se unen a las que provienen de las crestas en la división vestibular del octavo par craneal. El potencial de membrana en reposo de las células ciliadas es de alrededor de –60 mV. Cuando se empujan los estereocilios hacia el cinocilio, el potencial de membrana disminuye a alrededor de –50 mV. Las prolongaciones ciliares proporcionan un mecanismo para generar cambios del potencial de membrana proporcionales a la dirección y la distancia que el cilio se mueve. Cuando el fascículo de prolongaciones es empujado en la dirección opuesta, la célula es hiperpolarizada. El desplazamiento de las prolongaciones en una dirección perpendicular a este eje no proporciona cambio del potencial de membrana, y el desplazamiento de las prolongaciones en direcciones que son intermedias entre estas dos direcciones produce despolarización o hiperpolarización que es proporcional al grado al cual la dirección es hacia el cinocilio o en dirección contraria a este último. GÉNESIS DE POTENCIALES DE ACCIÓN EN FIBRAS NERVIOSAS AFERENTES Prolongaciones muy finas, que se conocen como uniones de punta (figura 16-6) atan la punta de cada estereocilio al lado de su vecino más alto, y en la unión hay canales catiónicos mecanosensitivos. Si se empujan los estereocilios más cortos hacia los más altos, el tiempo abierto de los canales aumenta, entran K+ y Ca2+ a través del canal y producen despolarización. A continuación, un motor molecular en el vecino más alto puede mover el canal hacia la base, lo que libera la tensión en la unión de punta. Esto hace que el canal se cierre y permite la restitución de un estado en reposo. La despolarización de células ciliadas hace que liberen un neurotransmisor que inicia la despolarización de neuronas aferentes vecinas. CAPÍTULO 16 Sentidos especiales II: audición y equilibrio 151 OM OM OL K S RC A SC E FIGURA 165 Estructura de una célula ciliada en el sáculo. Izquierda: las células ciliadas en el laberinto membranoso del oído tienen una estructura común, y cada una está dentro de un epitelio de células de sostén (SC) rematadas por una membrana otolítica (OM) embebida con los otolitos (OL). Prolongaciones en forma de bastón, o células ciliadas (RC) que se proyectan desde el extremo apical, están en contacto con fibras nerviosas aferentes (A) y eferentes (E). Excepto en la cóclea, una de éstas, el cinocilio (K) es un cilio verdadero, pero no móvil con nueve pares de microtúbulos alrededor de su circunferencia y un par central de microtúbulos. Las otras prolongaciones, los estereocilios (S) se encuentran en todas las células ciliadas; tienen centros de filamentos de actina cubiertos con isoformas de miosina. Hay una estructura ordenada dentro de la agrupación de prolongaciones en cada célula. A lo largo de un eje hacia el cinocilio, la altura de los estereocilios aumenta de manera progresiva; a lo largo del eje perpendicular, todos los estereocilios tienen la misma altura. (Reproducida con autorización de Hillman DE: Morphology of peripheral and central vestibular systems. In: Frog Neurobiology. Llinas R, Precht W (editors). Springer, 1976.) Derecha: fotomicrografía electrónica de barrido de prolongaciones sobre una célula ciliada en el sáculo. Se quitó la membrana otolítica. Las proyecciones pequeñas alrededor de la célula ciliada son microvellosidades sobre células de sostén. (Cortesía de A.J. Hudspeth.) Miosina Ca2+ K+ Unión de punta FIGURA 166 Representación esquemática de la función de las uniones de punta en las respuestas de células ciliadas. Cuando un estereocilio es empujado hacia un estereocilio más alto, la unión de punta es estirada y abre un canal iónico en su vecino más alto. A continuación un motor molecular mueve el canal en dirección descendente por el estereocilio más alto, de modo que se libera la tensión sobre la unión de punta. Cuando los pelos vuelven a su posición en reposo, el motor se mueve de regreso hacia arriba por el estereocilio. (Modificada con autorización de Kandel ER, Schwartz JH, Jessel TM [editors]: Principles of Neuroscience, 4th ed. McGraw-Hill, 2000.) 152 SECCIÓN IV Fisiología del sistema nervioso central/neural SL y SV Escala vestibular Na+ 150 K+ 5 Cl– 125 Na+ 1 K+ 150 – Cl 130 Órgano de Corti Na+ 150 K+ 3 Cl– 125 Escala timpánica FIGURA 167 Composición iónica (mmol/L) de la perilinfa en la escala vestibular, la endolinfa en la escala media y la perilinfa en la escala timpánica. SL, ligamento espiral; SV, estría vascular. La flecha discontinua indica la vía mediante la cual el K+ se recicla desde las células ciliadas hacia las células de sostén y al ligamento espiral, y después es secretado de regreso hacia la endolinfa por células en la estría vascular. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) El K+ que entra a las células ciliadas por medio de los canales catiónicos mecanosensitivos es reciclado (figura 16-7). Entra a las células de sostén y después pasa a otras células de sostén mediante uniones intercelulares herméticas (zonas de oclusión). En la cóclea, por último, llega a la estría vascular y es secretado de regreso hacia la endolinfa, lo cual completa el ciclo. Las prolongaciones de las células ciliadas se proyectan hacia la endolinfa y las bases están bañadas en perilinfa. La perilinfa está formada principalmente de plasma; la endolinfa está formada, en la escala media, por la estría vascular, y tiene una concentración alta de K+ y baja de Na+. Las células en la estría vascular tienen una concentración alta de Na+, K+-ATPasa. AUDICIÓN ONDAS DE SONIDO El sonido es la sensación que se produce cuando vibraciones de moléculas en el ambiente externo llegan a la membrana timpánica. El volumen de un sonido se correlaciona con la amplitud de una onda de sonido, y su tono, con su frecuencia (número de ondas por unidad de tiempo). La amplitud de una onda de sonido se expresa en una escala relativa, que se conoce como escala de decibel. La intensidad de un sonido en beles es el logaritmo de la proporción entre la intensidad de ese sonido y un sonido estándar. Un valor de 0 dB no significa la ausencia de sonido; más bien, es una magnitud de sonido cuya intensidad es igual a la de un estándar. El rango de 0 a 160 dB desde la presión umbral hasta una presión que es en potencia perjudicial para el órgano de Corti en realidad representa una variación de 107 veces la presión del sonido. Un rango de 120 a 160 dB (p. ej., armas de fuego, martillo neumático, avión a chorro al despegar) es doloroso, 90 a 100 dB (metro subterráneo, bombo de batería, motosierra, segadora de césped) es en extremo alto, 60 a 80 dB (reloj despertador, tráfico intenso, lavavajillas, conversación) es muy fuerte, 40 a 50 dB (p. ej., lluvia moderada, ruido ambiente normal) es moderado, y 30 dB (susurro, biblioteca) es leve. Las frecuencias de sonido audibles para el ser humano varían desde alrededor de 20 hasta 20000 ciclos por segundo (cps, Hz). El rango disminuye con la edad; en especial surge dificultad para detectar sonidos de frecuencia más alta. El umbral del oído humano varía con el tono del sonido; la mayor sensibilidad está en el rango de 1000 a 4000 Hz. El tono de la voz masculina y femenina promedio en conversación es de 120 y 250 Hz, respectivamente. El número de tonos que un individuo promedio puede distinguir es de alrededor de 2 000, pero los músicos entrenados pueden mejorar esta cifra. TRANSMISIÓN DE SONIDO El oído convierte ondas de sonido en el ambiente en potenciales de acción en los nervios auditivos. El tímpano y los osículos del oído transforman las ondas en movimientos de la base del estribo. Estos movimientos establecen ondas en el líquido del oído interno. La acción de las ondas sobre el órgano de Corti genera potenciales de acción en el nervio. La membrana timpánica se mueve hacia adentro y hacia afuera en respuesta a los cambios de presión producidos por ondas de sonido sobre su superficie externa. Así, la membrana funciona como un resonador que reproduce las vibraciones de la fuente de sonido. Deja de vibrar casi de inmediato cuando la onda de sonido cesa. Los movimientos de la membrana timpánica son impartidos al manubrio. El martillo se mece sobre un eje por medio de la unión de sus prolongaciones larga y corta, de modo que la prolongación corta transmite las vibraciones del manubrio hacia el yunque. El yunque se mueve de tal manera que las vibraciones se transmiten a la cabeza del estribo. Los movimientos de la cabeza del estribo hacen oscilar su base hacia adelante y hacia atrás como una puerta con bisagra en el borde posterior de la ventana oval. Los osículos del oído funcionan como un sistema de palancas que convierte las vibraciones resonantes de la membrana timpánica en movimientos del estribo contra la escala vestibular de la cóclea llena de perilinfa (figura 16-8). Este sistema aumenta la presión del sonido que llega a la ventana oval, porque la acción de palanca del martillo y el yunque multiplica 1.3 veces la fuerza, y el área de la membrana timpánica es mucho mayor que el área de la base del estribo. Cuando los músculos del oído medio (tensor del tímpano y estapedio) se contraen, el manubrio del martillo tira hacia adentro y la base del estribo empuja hacia afuera (figura 16-2), lo que disminuye la transmisión de sonido. Los sonidos fuertes inician el reflejo timpánico, que contrae los músculos del oído medio para evitar que ondas de sonido fuertes causen estimulación excesiva de los receptores auditivos. CONDUCCIÓN ÓSEA Y AÉREA La conducción osicular es la conducción normal de ondas de sonido al líquido en el oído medio a través de la membrana timpánica y CAPÍTULO 16 Sentidos especiales II: audición y equilibrio POTENCIALES DE ACCIÓN EN FIBRAS NERVIOSAS AUDITIVAS Martillo Yunque Pivote Estribo Ventana oval 153 Membrana de Reissner Órgano de Corti Ventana redonda Membrana basilar Tuba auditiva FIGURA 168 Representación esquemática de los osículos del oído y la manera en que su movimiento traduce movimientos de la membrana timpánica hacia una onda en el líquido del oído interno. La onda se disipa en la ventana redonda. Los movimientos de los osículos, el laberinto membranoso y la ventana redonda se indican con líneas discontinuas. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Las células ciliadas internas son las células sensoriales primarias que generan potenciales de acción en nervios auditivos, y son estimuladas por los movimientos de líquido antes mencionados. Las células ciliadas externas muestran respuesta al sonido, pero la despolarización las hace cortas, y la hiperpolarización las hace alargadas. Esto lo realizan en una parte muy flexible de la membrana basal, y esta acción aumenta la amplitud de los sonidos y la claridad de los mismos. La frecuencia de los potenciales de acción en las fibras del nervio auditivo es proporcional al volumen de los estímulos de sonido. El principal determinante del tono percibido cuando una onda de sonido llega al oído es el lugar del órgano de Corti, que se estimula al máximo. La onda viajera establecida por un tono produce depresión máxima de la membrana basilar y, en consecuencia, estimulación de receptor máxima, en un punto. La distancia entre este punto y el estribo guarda relación inversa con el tono del sonido; los tonos bajos producen estimulación máxima en el vértice de la cóclea, y los tonos altos causan estimulación máxima en la base. VÍA CENTRAL Medical, 2009.) los osículos del oído. Las ondas de sonido también inician vibraciones de la membrana timpánica secundaria que cierra la ventana redonda; este proceso, poco importante en la audición normal, se llama conducción aérea. La conducción ósea es la transmisión de vibraciones de los huesos del cráneo al líquido del oído interno. Esto desempeña un papel en la transmisión de ruidos en extremo fuertes. También ocurre conducción ósea considerable cuando se aplica de manera directa al cráneo un diapasón que vibra. ONDAS VIAJERAS Los movimientos de la base del estribo establecen una serie de ondas que viajan en la perilinfa de la escala vestibular. Las paredes óseas de la escala vestibular son rígidas, pero la membrana de Reissner es flexible. La membrana basilar no está bajo tensión, y también es deprimida con facilidad hacia la escala timpánica por los máximos de ondas en la escala vestibular. Los desplazamientos del líquido en la escala timpánica son disipados hacia el aire en la ventana redonda. El sonido deforma la membrana basilar, y el sitio en el cual esta deformación es máxima está determinado por la frecuencia de la onda de sonido. Las partes superiores de las células ciliadas en el órgano de Corti se mantienen rígidas por la lámina reticular, y las prolongaciones de las células ciliadas externas están embebidas en la membrana tectorial (figura 16-4). Cuando el estribo se mueve, ambas membranas lo hacen en la misma dirección, pero tienen bisagras en ejes diferentes, de modo que un movimiento de corte flexiona los cilios. Las prolongaciones de las células ciliadas internas no están fijas a la membrana tectorial, sino que las flexiona el líquido que se mueve entre la membrana y las células ciliadas subyacentes. Las fibras aferentes en la división auditiva del octavo par craneal terminan en los núcleos cocleares dorsal y ventral (figura 16-9). Desde ahí, impulsos auditivos pasan por diversas rutas a los colículos inferiores, los centros para los reflejos auditivos, y mediante el cuerpo geniculado medial en el tálamo a la corteza auditiva. Otros impulsos entran a la formación reticular. La información que proviene de ambos oídos converge en cada oliva superior, y más allá de esto, casi todas las neuronas muestran respuesta a aferencias provenientes de ambos lados. La corteza auditiva primaria es el área de Brodmann 41. Los tonos bajos se representan en posición anterolateral, y los tonos altos en posición posteromedial en la corteza auditiva. En la corteza auditiva primaria, casi todas las neuronas muestran respuesta a aferencias que se originan en ambos oídos, pero tiras de células son estimuladas por aferencias que proceden del oído contralateral, e inhibidas por aferencias que provienen del oído ipsilateral. Hay otras áreas receptoras auditivas, del mismo modo que hay varias áreas receptoras para la sensación cutánea. Las áreas de asociación auditivas adyacentes a las áreas receptoras auditivas primarias están difundidas. El fascículo olivo-coclear es un fascículo prominente de fibras eferentes en cada nervio auditivo, que surge a partir de complejos olivares superiores tanto ipsilateral como contralateral, y termina alrededor de las bases de las células ciliadas externas del órgano de Corti. LOCALIZACIÓN DEL SONIDO La determinación de la dirección desde la cual emana un sonido en el plano horizontal depende de detectar la diferencia de tiempo entre la llegada del estímulo en los dos oídos y el desfase consecuente de la fase de las ondas de sonido a ambos lados; también depende del hecho de que el sonido es más fuerte en el lado más cercano a la 154 SECCIÓN IV Fisiología del sistema nervioso central/neural fuente. La diferencia de tiempo detectable, que puede ser de apenas 20 μs, es el factor más importante a frecuencias por debajo de 3 000 Hz, y la diferencia de volumen es el factor más significativo a frecuencias por arriba de 3 000 Hz. Neuronas en la corteza auditiva que reciben aferencias que se originan en ambos oídos muestran respuesta máxima o mínima cuando el tiempo de llegada de un estímulo en un oído es retrasado por un periodo fijo respecto del tiempo de llegada al otro oído. Este periodo fijo varía de una neurona a otra. Los sonidos que vienen de manera directa de enfrente del individuo difieren en calidad de los que se derivan desde atrás porque cada pabellón auricular (la porción visible del oído externo) está girada un poco hacia adelante. Asimismo, los reflejos de ondas de sonido desde la superficie del pabellón auricular cambian conforme los sonidos se mueven hacia arriba o hacia abajo; el cambio en las ondas de sonido es el factor primario en la localización de sonidos en el plano vertical. Las lesiones de la corteza auditiva alteran la localización del sonido. par craneal o dentro de vías auditivas centrales. Puede alterar la capacidad para oír ciertos tonos, mientras que otros no quedan afectados. Los antibióticos aminoglucósidos, como la estreptomicina y la gentamicina, obstruyen los canales mecanosensitivos en los estereocilios de células ciliadas, y pueden hacer que las células se degeneren, lo que produce pérdida neurosensorial de la audición, y función vestibular anormal. El daño de las células ciliadas externas por exposición prolongada al ruido se asocia con pérdida de la audición. Otras causas son tumores del octavo par craneal, y el ángulo cerebelopontino, y daño vascular en el bulbo raquídeo. Las sorderas de conducción y neurosensorial pueden diferenciarse mediante pruebas simples con un diapasón. En el cuadro 16-1 se esbozan tres de estas pruebas, nombradas en honor de quienes las crearon. Las pruebas de Weber y de Schwabach demuestran el importante efecto enmascarador del ruido ambiental sobre el umbral auditivo. SISTEMA VESTIBULAR SORDERA La pérdida de la audición es el defecto sensorial más común en seres humanos. La presbiacusia, la pérdida auditiva gradual asociada con el envejecimiento, afecta a más de una tercera parte de las personas de más de 75 años de edad, y quizá se debe a pérdida acumulativa gradual de células ciliadas y neuronas. La pérdida de la audición casi siempre es un trastorno multifactorial causado por factores tanto genéticos como ambientales. Sordera de conducción se refiere a transmisión de sonido alterada en el oído externo o medio, y tiene repercusiones sobre todas las frecuencias de sonido. Las causas de sordera de conducción son presencia de tapones de cera (cerumen) o cuerpos extraños en los conductos auditivos externos, acumulación de líquido debida a otitis externa (inflamación del oído externo, u “oído del nadador”) u otitis media (inflamación del oído medio, perforación del tímpano, y otosclerosis, en la cual se resorbe hueso y queda reemplazado por hueso esclerótico que crece sobre la ventana oval. La sordera neurosensorial por lo general se debe a la pérdida de células ciliadas cocleares, pero también a problemas con el octavo El sistema vestibular se divide en el aparato vestibular y los núcleos vestibulares centrales. El aparato vestibular dentro del oído interno detecta el movimiento y la posición de la cabeza, y transduce esta información hacia una señal neural. Los núcleos vestibulares se relacionan con el mantenimiento de la posición de la cabeza en el espacio; los tractos que descienden desde estos núcleos median los ajustes de la cabeza sobre el cuello y de la cabeza sobre el cuerpo. Los ganglios vestibulares contienen los cuerpos celulares de las neuronas que inervan las crestas y las máculas. Cada nervio vestibular termina en el núcleo vestibular ipsilateral y en el lóbulo floculonodular del cerebelo (figura 16-9). Las fibras que provienen de los canales semicirculares terminan en las divisiones superior y medial del núcleo vestibular, y se proyectan sobre todo hacia núcleos que controlan el movimiento ocular. Las fibras que se originan en el utrículo y en el sáculo terminan en el núcleo de Deiters, que se proyecta hacia la médula espinal. Los núcleos vestibulares también se proyectan hacia el tálamo, y desde ahí hacia la corteza somatosensorial primaria. Las conexiones ascendentes hacia los núcleos de los nervios craneales se relacionan con los movimientos de los ojos. CUADRO 16-1 Pruebas comunes con un diapasón para distinguir entre sordera neurosensorial y de conducción. Weber Rinne Schwabach Método La base del diapasón vibratorio se coloca sobre el vértex del cráneo. La base del diapasón vibratorio se coloca sobre la apófisis mastoides hasta que el sujeto ya no lo escucha, y después se mantiene en el aire. Conducción ósea del paciente comparada con la de un sujeto sano. Normal Oye igual en ambos lados. Oye vibración en el aire después de que la conducción ósea termina. Sordera de conducción (un oído) Sonido más fuerte en el oído enfermo porque el efecto del ruido ambiental que produce enmascarado está ausente en el lado enfermo. Las vibraciones en el aire no se escuchan después de que la conducción ósea termina. Conducción ósea mejor que la normal (un defecto de conducción excluye ruido que produce enmascarado). Sordera neurosensorial (un oído) Sonido más fuerte en el oído normal. Vibración oída en el aire después de que la conducción ósea termina, en tanto la sordera nerviosa sea parcial. Conducción ósea peor que la normal. CAPÍTULO 16 Sentidos especiales II: audición y equilibrio 155 Hacia la corteza somatosensorial Hacia la corteza (circunvolución temporal superior) Tálamo Tálamo Cuerpo geniculado medial III IV Hacia el cerebelo Pineal Colículo inferior Formación reticular Fascículo longitudinal medial VI IV ventrículo Núcleos cocleares dorsal y ventral Ganglio vestibular Ganglio espiral Bulbo raquídeo Olivas superiores Núcleos vestibulares: superior, lateral (de Deiters), medial, espinal Tracto vestíbulo espinal lateral Tractos vestíbuloespinales anteriores Desde el utrículo, canales semicirculares Desde la cóclea AUDITIVO VESTIBULAR FIGURA 169 Diagrama simplificado de las vías auditiva (izquierda) y vestibular (derecha) principales, superpuestas en una vista dorsal del tallo encefálico. Se quitaron el cerebelo y la corteza cerebral. III, IV y VI son el tercer, cuarto y sexto nervios craneales. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) RESPUESTA A LA ACELERACIÓN ROTACIONAL La aceleración rotacional en el plano de un canal semicircular dado estimula su cresta. La endolinfa es desplazada en una dirección opuesta a la rotación, y el líquido empuja la cúpula, y la deforma. Esto flexiona las prolongaciones de las células ciliadas (figura 16-3). Cuando se alcanza una rapidez de rotación constante, el líquido gira al mismo ritmo que el cuerpo, y la cúpula oscila de regreso hacia la posición erecta. Cuando se suspende la rotación, la desaceleración produce desplazamiento de la endolinfa en la dirección de la rotación, y la cúpula se deforma en una dirección opuesta a la que hubo durante la aceleración. Vuelve a la posición media en 25 a 30 s. El movimiento de la cúpula en una dirección aumenta la actividad de nervios desde la cresta, y el movimiento en la dirección opuesta inhibe la actividad neural. La rotación causa estimulación máxima de los canales semicirculares en el plano de rotación. Puesto que los canales en un lado de la cabeza son una imagen en espejo de los que se encuentran en el otro lado, la endolinfa se desplaza hacia la ampolla en un lado y en dirección contraria a la misma en el otro lado. El patrón de estimulación que llega al cerebro varía con la dirección de la rotación y con el plano de la misma. El nistagmo es el movimiento a sacudidas, característico de los ojos, que se observa al principio de un periodo de rotación y al final. Es un reflejo que mantiene la fijación visual en puntos estacionarios mientras el cuerpo rota. Cuando empieza la rotación, los ojos se mueven con lentitud en una dirección opuesta a la dirección de rotación, y mantienen la fijación visual (reflejo vestíbulo-ocular). Al final de este movimiento, los ojos regresan con rapidez a un nuevo punto de fijación, y después se mueven lentamente en la otra dirección. El componente lento se inicia con impulsos que provienen de los laberintos vestibulares; el componente rápido lo desencadena un centro en el tallo encefálico. Por convención, la dirección del movimiento de los ojos en el nistagmo la identifica la dirección del componente rápido. Esta última es la misma que la de la rotación; el nistagmo posrotatorio, que se debe a desplazamiento de la cúpula, es en la dirección opuesta. El nistagmo puede observarse en reposo en pacientes con lesiones del tallo encefálico; puede usarse estimulación calórica para probar la función del laberinto vestibular. Los canales semicirculares se estimulan al instilar agua caliente (40 °C) o fría (30 °C) en el oído. La diferencia de temperatura establece corrientes de convección en la endolinfa, con movimiento consiguiente de la cúpula. En sujetos normales, el agua caliente causa nistagmo que va hacia el estímulo, y el agua fría induce nistagmo que va hacia el oído opuesto. 156 SECCIÓN IV Fisiología del sistema nervioso central/neural RESPUESTAS A LA ACELERACIÓN LINEAL Las máculas utricular y sacular muestran respuesta a la aceleración horizontal y vertical. Los otolitos son más densos que la endolinfa, y la aceleración en cualquier dirección hace que sean desplazados en la dirección opuesta, lo que deforma las prolongaciones de las células ciliadas y genera actividad neural. Las máculas también descargan tónicamente en ausencia de movimiento de la cabeza, debido a la influencia de la fuerza de gravedad sobre los otolitos. Los reflejos de enderezamiento laberínticos son una serie de respuestas integradas en núcleos del mesencéfalo a inclinación de la cabeza. La respuesta es una contracción compensatoria de los músculos del cuello para mantener el nivel de la cabeza. Estos reflejos estabilizan la cabeza y mantienen los ojos fijos en blancos visuales a pesar de movimientos del cuerpo. La percepción consciente de movimiento y el suministro de parte de la información necesaria para la orientación en el espacio quizá dependen de impulsos vestibulares que llegan a la corteza cerebral. El vértigo es la sensación de rotación en ausencia de rotación real, y es un síntoma prominente cuando un laberinto está inflamado. tado que dura varias horas. Los síntomas pueden ocurrir de manera repentina y recurrir a diario o muy rara vez. La pérdida de la audición al principio es transitoria, pero puede hacerse permanente. La fisiopatología tal vez involucre una reacción inmunitaria. Una respuesta inflamatoria puede aumentar el volumen de líquido dentro del laberinto membranoso, lo que hace que se rompa y permite que la endolinfa y la perilinfa se mezclen. No hay curación para la enfermedad de Ménière, pero los síntomas se pueden controlar al reducir la retención de líquido por medio de cambios de la dieta (dieta baja en sal o sin sal, consumo nulo de cafeína y de alcohol) o medicación. RESUMEN DEL CAPÍTULO ■ ■ ORIENTACIÓN ESPACIAL La orientación en el espacio depende de las aferencias que provienen de los receptores vestibulares, así como de indicios visuales, información que se origina en propioceptores en cápsulas articulares, y receptores de tacto y presión cutáneos. Estas cuatro aferencias se sintetizan en un ámbito cortical hacia un cuadro continuo de la orientación del individuo en el ambiente. ■ ■ ■ CORRELACIÓN CLÍNICA Una mujer de 26 años de edad acudió al consultorio de su médico de atención primaria debido a una serie reciente de episodios en los cuales sintió mareo intenso, tinnitus (zumbidos de oídos), náuseas y vómitos. Durante el último episodio, se percató de que apenas podía oír a su hija llamándola desde la habitación de junto. Cuando fue interrogada por el ayudante del médico, recordó que mientras estaba en la universidad había presentado episodios similares, aunque menos graves, de estos síntomas. En particular, experimentó tinnitus, vértigo y náuseas en varias ocasiones. Dado que los síntomas ocurrieron de manera esporádica y duraron sólo horas o un día, no buscó orientación médica. Los episodios más recientes la hicieron temer que le estuviera pasando algo grave. En una prueba de audición se detectó audición reducida en un oído. Con base en los síntomas, su médico sospechó enfermedad de Ménière. Más tarde la atendieron un otorrinolaringólogo y un neurólogo para excluir otras causas de los síntomas. La enfermedad de Ménière es una anormalidad del oído interno que causa vértigo o mareo intenso, tinnitus, pérdida fluctuante de la audición, y la sensación de presión o dolor en el oído afec- ■ ■ El oído externo canaliza ondas de sonido hacia el meato auditivo externo y la membrana timpánica. Desde ahí, las ondas de sonido pasan a través de tres osículos del oído (martillo, yunque y estribo) en el oído medio. El oído interno, o laberinto, contiene la cóclea y el órgano de Corti. Las células ciliadas en el órgano de Corti emiten señales de audición. Los estereocilios proporcionan un mecanismo para generar cambios del potencial de membrana proporcionales a la dirección y la distancia en que el pelo se mueve. El sonido es la sensación que se produce cuando vibraciones longitudinales de moléculas de aire llegan a la membrana timpánica. La actividad dentro de la vía auditiva pasa desde las fibras aferentes del octavo par craneal hacia los núcleos cocleares dorsal y ventral, los colículos inferiores, el cuerpo geniculado medial talámico, y después hacia la corteza auditiva. El volumen se correlaciona con la amplitud de una onda de sonido, y el tono con la frecuencia. La sordera de conducción se debe a la transmisión de sonido alterada en el oído medio, y repercute sobre todas las frecuencias de sonido. La sordera neurosensorial se debe a la pérdida de las células ciliadas cocleares o a daño del octavo par craneal o de las vías auditivas centrales. La aceleración rotacional estimula la cresta en el canal semicircular; ello desplaza la endolinfa en una dirección opuesta a la dirección de la rotación, lo cual deforma la cúpula y flexiona la célula ciliada. El utrículo responde a la aceleración horizontal, y el sáculo, a la aceleración vertical. La aceleración en cualquier dirección desplaza los otolitos, lo que deforma las prolongaciones de las células ciliadas y genera actividad neural. La orientación espacial depende de aferencias desde receptores vestibulares, indicios visuales, propioceptores en cápsulas articulares, y receptores de tacto y presión cutáneos. PREGUNTAS DE ESTUDIO 1. Un varón de 40 años de edad, empleado como trabajador de la construcción de caminos durante cerca de 20 años, acudió con su médico para reportar que tenía dificultad para oír durante conversaciones normales. Una prueba de Weber mostró que el sonido proveniente de un diapasón que vibraba se localizó en el oído derecho. Una prueba de Schwabach evidenció conducción ósea por debajo de lo normal. Una prueba de Rinne reveló que las conducciones tanto aérea como ósea fueron anormales, pero la CAPÍTULO 16 Sentidos especiales II: audición y equilibrio conducción de aire duró más tiempo que la conducción ósea. El diagnóstico fue: A) sordera sensorial en ambos oídos. B) sordera de conducción en el oído derecho. C) sordera sensorial en el oído derecho. D) sordera de conducción en el oído izquierdo. E) sordera neurosensorial en el oído izquierdo. 2. ¿Cuál sería el diagnóstico si un paciente tuviera los siguientes resultados de prueba? La prueba de Weber mostró que el sonido de un diapasón vibratorio fue más fuerte que lo normal; la prueba de Schwabach evidenció que la conducción ósea fue mejor que lo normal, y la prueba de Rinne reveló que la conducción aérea no duró más que la conducción ósea: A) sordera sensorial en ambos oídos. B) sordera de conducción en ambos oídos. C) audición normal. D) sordera tanto sensorial como de conducción. E) un posible tumor en el octavo par craneal. 3. El nistagmo posrotacional se origina por movimiento continuo de: A) el humor acuoso sobre el cuerpo ciliar en el ojo. B) el líquido cefalorraquídeo sobre los núcleos vestibulares. C) endolinfa en los canales semicirculares, con flexión consiguiente de la cúpula y estimulación de células ciliadas. D) endolinfa hacia el helicotrema. E) perilinfa sobre células ciliadas con prolongaciones embebidas en la membrana tectorial. 4. Algunas enfermedades dañan las células ciliadas en el oído. Cuando el daño de las células ciliadas externas es mayor que el daño de las células ciliadas internas: A) la percepción de la aceleración vertical está alterada. B) la concentración de K+ en la endolinfa está disminuida. 157 C) la concentración de K+ en la perilinfa está disminuida. D) hay pérdida intensa de la audición. E) las células ciliadas afectadas no se acortan cuando quedan expuestas a sonido. 5. ¿Cuáles de los que siguen están pareados de manera incorrecta? A) membrana timpánica:manubrio del martillo B) helicotrema:vértice de la cóclea C) base del estribo:ventana oval D) otolitos:canales semicirculares E) membrana basilar:órgano de Corti 6. La dirección del nistagmo es vertical cuando un sujeto es rotado: A) después de introducir agua caliente en un oído. B) con la cabeza inclinada hacia atrás. C) después de introducir agua fría en ambos oídos. D) con la cabeza inclinada hacia los lados. E) después de corte de un nervio vestibular. 7. En el utrículo, las uniones de punta en las células ciliadas están involucradas en: A) la formación de perilinfa. B) despolarización de la estría vascular. C) movimientos de la membrana basal. D) percepción de sonido. E) regulación de canales iónicos mecanosensitivos. Sentidos especiales III: olfato y gusto Susan M. Barman C A P Í T U L O 17 O B J E T I V O S ■ ■ ■ ■ ■ ■ Describir las características básicas del epitelio olfatorio y el bulbo olfatorio. Explicar la transducción de señal en receptores odorantes. Esbozar la vía mediante la cual los impulsos generados en el epitelio olfatorio llegan a la corteza olfatoria. Describir la ubicación de las papilas gustativas y su composición celular. Nombrar los cinco principales receptores gustativos y sus mecanismos de transducción de señal. Delinear las vías mediante las cuales los impulsos generados en receptores gustativos llegan a la corteza insular. INTRODUCCIÓN El olfato y el gusto se clasifican como sentidos viscerales por su asociación estrecha con la función gastrointestinal. Desde el punto de vista fisiológico, se relacionan entre sí; los sabores de diversos alimentos son, en gran parte, una combinación de su sabor y olor. Esto explica por qué la comida puede saber “diferente” si se tiene un resfriado que deprime el sentido del olfato. Los receptores tanto olfatorios como gustativos son quimiorreceptores que estimulan moléculas en solución que se localizan en el moco de la nariz, y en la saliva de la boca. OLFATO EPITELIO OLFATORIO Y BULBOS OLFATORIOS Una porción especializada de la mucosa nasal, el epitelio olfatorio pigmentado, amarillento (figura 17-1), contiene 10 a 20 millones de neuronas sensoriales olfatorias bipolares que se entremezclan con células de sostén (sustentaculares) parecidas a glía y células madre basales. El epitelio olfatorio es el lugar del cuerpo donde el sistema nervioso está más cerca del mundo externo. Cada neurona tiene una dendrita gruesa y corta que se proyecta hacia la cavidad nasal, donde termina en una protuberancia que contiene 10 a 20 cilios (figura 17-2). Los cilios son prolongaciones no mielinizadas que contienen receptores odorantes. Los axones de las neuronas sensoriales olfatorias pasan a través de la placa cribiforme del hueso etmoides y entran en los bulbos olfatorios (figura 17-1). Las células madre basales generan nuevas neuronas sensoriales olfatorias según se requiera para reemplazar las que quedan dañadas por exposición al ambiente. En los bulbos olfatorios, los axones de las neuronas sensoriales olfatorias (primer nervio craneal) entran en contacto con las dendritas primarias de las células mitrales y las células en penacho (figura 17-3) para formar unidades sinápticas separadas de manera anatómica, las cuales se denominan glomérulos olfatorios. Ambos tipos de neuronas envían axones hacia la corteza olfatoria. Los bulbos olfatorios también contienen células periglomerulares, que son neuronas inhibitorias, las cuales conectan un glomérulo a otro, y células granulares, que no tienen axones y hacen sinapsis recíprocas con las dendritas laterales de las células mitrales y en penacho. En estas sinapsis, la célula mitral o en penacho sale de la célula granular al liberar glutamato y la célula granular, a su vez, inhibe la célula mitral o en penacho al liberar ácido γ-aminobutírico (GABA). CORTEZA OLFATORIA El axón de las células mitrales y en penacho pasa en dirección posterior a través de la estría olfatoria lateral para terminar en dendritas apicales de células piramidales en cinco regiones de la corteza olfatoria: núcleo olfatorio anterior, tubérculo olfatorio, corteza piriforme, amígdala, y corteza entorrinal (figura 17-4). Desde estas 159 160 SECCIÓN IV Fisiología del sistema nervioso central/neural Bulbo olfatorio Bulbo olfatorio Placa cribiforme Epitelio olfatorio Neuronas sensoriales olfatorias FIGURA 171 Neuronas sensoriales olfatorias embebidas dentro del epitelio olfatorio en la fosa posterior dorsal de la cavidad nasal. Estas neuronas proyectan axones hacia el bulbo olfatorio del cerebro, una pequeña estructura ovoide que descansa sobre la placa cribiforme del hueso etmoides. (Reproducida con autorización de Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000.) regiones, la información viaja de manera directa a la corteza cerebral o por medio del tálamo a la corteza orbitofrontal. La discriminación consciente de olores depende de la vía hacia la corteza orbitofrontal. La activación orbitofrontal por lo general es mayor en el lado derecho que en el izquierdo; así, la representación cortical del olfato es asimétrica. La vía hacia la amígdala está involucrada con las respuestas emocionales a estímulos olfatorios, y la vía hacia la corteza entorrinal se relaciona con recuerdos olfatorios. Hacia la corteza olfatoria Hacia el bulbo olfatorio Células basales Axón Neurona sensorial olfatoria Dendrita Célula de sostén Moco Cilios FIGURA 172 Estructura del epitelio olfatorio. Hay tres tipos de células: neuronas sensoriales olfatorias, células de sostén y células madre basales en la base del epitelio. Cada neurona sensorial tiene una dendrita que se proyecta hacia la superficie epitelial. Muchos cilios sobresalen hacia la capa mucosa que reviste la luz nasal. Un axón único se proyecta desde cada neurona hacia el bulbo olfatorio. Los odorantes se unen a receptores odorantes específicos sobre los cilios, e inician una cascada de eventos que llevan a la generación de potenciales de acción en el axón sensorial. (Modificada con autorización de Kandel ER, Schwartz JH, Jessell FIGURA 173 Circuitos neurales básicos en el bulbo TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000.) and processing of odor molecular information. Science. 1999;286(5440):711–715.) olfatorio. Note que las células receptoras olfatorias con un tipo de receptor odorante se proyectan hacia un glomérulo olfatorio (OG), y células receptoras olfatorias con otro tipo de receptor se proyectan hacia un glomérulo olfatorio diferente. CP, placa cribiforme; PG, célula periglomerular; M, célula mitral; T, célula en penacho; Gr, célula granular. Flechas blancas, sinapsis excitatorias; flechas negras, sinapsis inhibitorias. (Adaptada con autorización de Mori K, Nagao H, Yoshihara Y. The olfactory bulb: Coding CAPÍTULO 17 Sentidos especiales III: olfato y gusto Célula mitral Bulbo olfatorio accesorio 161 Tracto olfatorio lateral Célula mitral Célula en penacho Bulbo olfatorio Órgano vomeronasal Tubérculo olfatorio Núcleo olfatorio anterior Corteza piriforme Amígdala Corteza entorrinal Epitelio olfatorio Hipotálamo Bulbo olfatorio contralateral Hipocampo Tálamo Corteza orbitofrontal Corteza frontal FIGURA 174 Diagrama de la vía olfatoria. La información la transmiten desde el bulbo olfatorio axones de neuronas de transmisión mitrales y en penacho en el tracto olfatorio lateral. Las células mitrales se proyectan hacia cinco regiones de la corteza olfatoria: núcleo olfatorio anterior, tubérculo olfatorio, corteza piriforme, y partes de la amígdala y de la corteza entorrinal. Las células en penacho se proyectan hacia el núcleo olfatorio anterior y el tubérculo olfatorio; las células mitrales en el bulbo olfatorio accesorio sólo se proyectan hacia la amígdala. La discriminación consciente del olor depende de la neocorteza (cortezas orbitofrontal y frontal). Los aspectos emotivos de la olfacción se derivan de proyecciones límbicas (amígdala e hipotálamo). En roedores y algunos mamíferos, un órgano vomeronasal bien desarrollado se relaciona con la percepción de olores que actúan como feromonas; sus receptores se proyectan hacia el bulbo olfatorio accesorio. (Reproducida con autorización de Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000.) DISCRIMINACIÓN OLFATORIA Y TRANSDUCCIÓN DE SEÑAL El epitelio olfatorio está cubierto por una capa delgada de moco que secretan las células de sostén y glándulas de Bowman, las cuales yacen por debajo del epitelio. El moco baña los receptores odorantes sobre los cilios y proporciona el ambiente molecular e iónico apropiado para la detección de olor. Las moléculas productoras de olor por lo general son pequeñas; contienen 3 a 20 átomos de carbono, y moléculas con el mismo número de átomos de carbono, pero distintas configuraciones estructurales tienen olores diferentes. La hidrosolubilidad y liposolubilidad altas son características de las sustancias que tienen olores fuertes. La anosmia (incapacidad para oler) y la hiposmia o hipestesia (sensibilidad olfatoria disminuida) pueden producirse por congestión nasal simple o por daño de los nervios olfatorios debido a fracturas de la placa cribiforme, neuroblastomas o meningiomas, o infecciones (como abscesos). El envejecimiento también se asocia con anormalidades de la sensación del olfato; más de 75% de los seres humanos de más de 80 años de edad tiene capacidad alterada para identificar olores. Los genes que codifican para alrededor de 1000 tipos diferentes de receptores odorantes constituyen la familia de genes de mayor tamaño hasta ahora descrita en mamíferos. Las secuencias de aminoácidos de receptores odorantes son diversas, pero todos los receptores odorantes están acoplados a proteínas G heterotriméricas. Cuando una molécula odorante se une a su receptor, las subunidades de proteína G (α, β, γ) se disocian (figura 17-5). La subunidad α activa la adenilato ciclasa para que catalice la producción de cAMP, la cual actúa como un segundo mensajero para abrir canales catiónicos, lo que causa una corriente de Ca2+ dirigida hacia adentro. Esto produce el potencial de receptor graduado, que a continuación lleva a un potencial de acción en el nervio olfatorio. Aunque hay millones de neuronas sensoriales olfatorias, cada una expresa sólo 1 de los 1 000 receptores odorantes diferentes. Cada neurona se proyecta hacia 1 o 2 glomérulos (figura 17-3). Esto proporciona un mapa bidimensional bien definido en el bulbo olfatorio que es singular para el odorante. Las células mitrales con sus glomérulos se proyectan hacia distintas partes de la corteza olfatoria. Los glomérulos olfatorios muestran inhibición lateral mediada por células periglomerulares y granulares. Esto agudiza señales olfatorias y las enfoca. Además, el potencial de campo extracelular en cada glomérulo oscila, y las células granulares pueden regular la frecuencia de la oscilación. Se desconoce la función exacta de la oscilación, pero quizá también ayude a enfocar las señales olfatorias que llegan a la corteza. GUSTO PAPILAS GUSTATIVAS El órgano del sentido especializado para el gusto (gustación) consta de alrededor de 10000 papilas gustativas. Hay cuatro tipos de células morfológicamente distintos dentro de cada papila gustativa: células basales, células oscuras, células claras y células intermedias 162 SECCIÓN IV Fisiología del sistema nervioso central/neural Odorante Adenilato ciclasa Receptor odorante Canal de Na+/Ca2+ Proteínas G a) Odorante Receptor odorante Adenilato ciclasa FIGURA 175 Transducción de señal en un receptor odorante. Los receptores olfatorios son receptores acoplados a proteína G que se disocian en el momento de la unión al odorante. La subunidad α de las proteínas G activa la adenilato ciclasa para que catalice la producción de cAMP. El cAMP actúa como un segundo mensajero para abrir canales catiónicos. La difusión hacia adentro de Na+ y Ca2+ produce despolarización. (Reproducida con Ca2+ Na+ Canal de Na+/Ca2+ cAMP ATP cAMP b) autorización de Fox SI: Human Physiology. McGraw-Hill, 2008.) (figura 17-6). Los tres últimos tipos de células se denominan células gustativas tipos I, II y III. Son las neuronas sensoriales que muestran respuesta a estímulos gustativos. Las terminaciones apicales de las células gustativas tienen microvellosidades, las cuales se proyectan hacia el poro gustativo, una pequeña abertura en la superficie dorsal de la lengua donde las células gustativas son expuestas al contenido oral. Cada papila gustativa está inervada por alrededor de 50 fibras nerviosas. Por el contrario, cada fibra nerviosa recibe aferencias que provienen de un promedio de cinco papilas gustativas. Las células basales surgen a partir de las células epiteliales que rodean la papila gustativa. Se diferencian hacia células gustativas nuevas, y las células antiguas son reemplazadas en un tiempo medio de alrededor de 10 días. Si se corta el nervio sensorial, las papilas gustativas que inerva se degeneran y, por último, desaparecen. Las papilas gustativas se ubican en la mucosa de la epiglotis, el paladar y la faringe, y en las paredes de las papilas de la lengua (figura 17-6). Las papilas fungiformes son estructuras redondeadas más numerosas cerca de la punta de la lengua; las papilas circunvaladas son estructuras prominentes dispuestas en V sobre el dorso de la lengua; las papilas foliadas están en el borde posterior de la lengua. Cada papila fungiforme tiene hasta cinco papilas gustativas, casi todas se localizan en la parte superior de la papila; cada papila valada y foliada contiene hasta 100 papilas gustativas, en su mayor parte se ubican a lo largo de los lados de las papilas. VÍAS GUSTATIVAS Las fibras nerviosas sensoriales que provienen de las papilas gustativas sobre los dos tercios anteriores de la lengua viajan en la rama cuerda del tímpano del nervio facial, y las que se generan en el tercio posterior de la lengua llegan al tallo encefálico por medio del nervio glosofaríngeo (figura 17-7). Las fibras que provienen de otras áreas que no son la lengua (p. ej., la faringe) llegan al tallo encefálico por medio del nervio vago. A cada lado, las fibras gustativas mielinizadas, pero de conducción lenta en estos tres nervios se unen en la porción gustativa del núcleo del tracto solitario (NTS) en el bulbo raquídeo (médula oblongada) (figura 17-7). Desde ahí, axones de neuronas de segundo orden ascienden en el lemnisco medial ipsilateral y pasan de manera directa hacia el núcleo ventral posteromedial del tálamo, desde el cual se proyectan fibras hacia la ínsula anterior y el opérculo frontal en la corteza cerebral ipsilateral. Esta región es rostral al área de la cara de la circunvolución central posterior, que puede ser el área que media la percepción consciente del gusto y de la discriminación del gusto. MODALIDADES, RECEPTORES Y TRANSDUCCIÓN GUSTATIVOS Se requiere un cambio de 30% de la concentración de la sustancia que se prueba antes de que se detecte una diferencia de intensidad. Una proteína que se une a moléculas productoras de gusto la produce la glándula de Ebner, la cual secreta moco hacia la hendidura alrededor de papilas valadas. La ageusia (falta del sentido del gusto) y la hipogeusia (sensibilidad disminuida del gusto) pueden originarse por daño del nervio lingual o glosofaríngeo. Trastornos neurológicos como el schwannoma vestibular, la parálisis de Bell, la disautonomía familiar, esclerosis múltiple y ciertas infecciones (p. ej., meningoencefalopatía ameboide primaria) también pueden causar problemas con la sensibilidad del gusto. La ageusia puede ser un efecto adverso de diversos fármacos o de deficiencia de vitamina B3 o de cinc. El envejecimiento y el abuso del consumo de tabaco contribuyen de igual manera a gusto disminuido. La disgeusia o CAPÍTULO 17 Sentidos especiales III: olfato y gusto Nervio cuerda del tímpano (VII) 163 Poro gustativo Saliva Nervio glosofaríngeo (IX) Célula epitelial Célula gustativa Circunvalada Célula basal Nervio aferente gustativo Glándula serosa A Hacia ganglio sensorial Foliada C Papila gustativa B Fungiforme FIGURA 176 Papilas gustativas ubicadas en las papilas (papillae) de la lengua del ser humano. A) Las papilas gustativas en los dos tercios anteriores de la lengua están inervadas por la rama cuerda del tímpano del nervio facial; las que están en el tercio posterior de la lengua están inervadas por la rama lingual del nervio glosofaríngeo. B) Los tres principales tipos de papilas (circunvaladas, foliadas y fungiformes) se sitúan en partes específicas de la lengua. C) Las papilas gustativas están compuestas de células madre basales y tres tipos de células gustativas (oscuras, claras e intermedias). Las células gustativas se extienden desde la base de la papila gustativa hasta el poro gustativo, donde microvellosidades tienen contacto con sustancias, que tienen sabor, disueltas en la saliva y el moco. (Modificada con autorización de Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000). parageusia (percepción desagradable del gusto) causa un sabor metálico, salado, repugnante o rancio. Los seres humanos tienen cinco sabores básicos establecidos: dulce, ácido, amargo, salado y umami. Todas las sustancias con sabor son detectadas a partir de todas las partes de la lengua y estructuras adyacentes. Los nervios aferentes que van al NTS contienen fibras que provienen de todos los tipos de receptores gustativos, sin alguna localización clara de tipos. Un quinto sentido del gusto, el umami, se ha añadido a los cuatro sabores clásicos; este sabor lo desencadena el glutamato, y en particular el glutamato monosódico (MSG), el cual se usa de manera muy extensa en la cocina asiática. El sabor es agradable y dulce, pero difiere del sabor dulce estándar. En la figura 17-8 se ilustra la transducción de señal en receptores gustativos. El sabor salado lo desencadena el NaCl. El gusto sensible a sal está mediado por un canal selectivo para Na+ que se conoce como ENaC, el canal de sodio epitelial sensible a amilorida. La entrada de Na+ hacia los receptores de sal despolariza la membrana, lo cual genera el potencial de receptor. En seres humanos, la sensibilidad del sabor salado a la amilorida es menos pronunciada que en algunas especies, lo que sugiere que hay mecanismos adicionales para activar receptores sensibles a sal. El sabor ácido lo originan protones (iones H+). Los ENaC permiten la entrada de protones y tal vez contribuyan a la sensación del sabor ácido. Los iones H+ también pueden unirse a un canal sensible a K+, y bloquearlo. El decremento de la permeabilidad al K+ puede despolarizar la membrana. Un canal catiónico sensible a nucleótidos cíclicos, activado por hiperpolarización, y otros mecanismos, puede contribuir a la transducción del sabor ácido. Las sustancias que saben dulce también actúan por medio de la proteína G gustducina. A la familia T1R3 de receptores acoplados a proteína G la expresan alrededor de 20% de las células gustativas, algunas de las cuales también expresan gustducina. Los azúcares saben dulce, pero también los compuestos como la sacarina, que tienen una estructura por completo diferente. Los azúcares naturales, como la sacarosa y los edulcorantes sintéticos, pueden actuar por medio de diferentes receptores sobre la gustducina. Al igual que los receptores con capacidad de respuesta a lo amargo, los receptores con capacidad de respuesta a lo dulce actúan por medio del metabolismo de nucleótidos cíclicos y fosfato de inositol. El sabor amargo lo producen diversos compuestos no relacionados. Muchos de éstos son venenos, y el sabor amargo sirve como un aviso para evitarlos. Algunos compuestos amargos se unen a canales selectivos para K+ y los bloquean. Muchos receptores enlazados a proteína G en el genoma del ser humano son receptores gustativos (familia T2R), y los estimulan sustancias amargas, como la estricnina. En algunos casos, estos receptores se acoplan a la proteína G heterotrimérica, gustducina. La gustducina disminuye el cAMP y aumenta la formación de fosfatos de inositol que podrían llevar a despolarización. Algunos compuestos amargos son permeables a la membrana y pueden no involucrar proteínas G; la quinina es un ejemplo. El sabor umami se debe a la activación de un receptor de glutamato metabotrópico truncado, mGluR4, en las papilas gustativas. No se ha establecido la manera en que la activación del receptor produce despolarización. El glutamato en los alimentos también puede activar receptores de glutamato ionotrópicos para despolarizar receptores de umami. 164 SECCIÓN IV Fisiología del sistema nervioso central/neural Corteza gustativa (ínsula anterior opérculo frontal) Núcleo ventral medial posterior del tálamo Ganglio geniculado Cuerda del tímpano N. VII Lengua Glosofaríngeo Núcleo del tracto solitario N. IX Ganglio petroso N. X Área gustativa Ganglio nodoso Faringe FIGURA 177 Diagrama de las vías gustativas. Las señales que provienen de las papilas gustativas viajan por medio de nervios diferentes hacia áreas gustativas del núcleo del tracto solitario que transmite la información hacia el tálamo; el tálamo se proyecta hacia la corteza gustativa. N. VII, N. IX y N. X son el séptimo, noveno y décimo pares craneales, respectivamente. (Modificada con autorización de Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000.) Salado α Agrio Receptor de dulce predicho γ Amargo Umami (L-glutamato) N X N N C N C ENaC, otros N C N C ENaC, HCN, otros C T1R3 (locus sac) C Familia T2R, otros C Sabor mGluR4 FIGURA 178 Transducción de señal en receptores gustativos. El sabor sensible a sal está mediado por un canal selectivo para Na+ (ENaC); el sabor ácido por iones H+ permeables a ENaC; el sabor umami por glutamato que actúa sobre un receptor metabotrópico, mGluR4; el sabor amargo está mediado por la familia T2R de receptores acoplados a proteína G; el sabor dulce quizá depende de la familia T1R3 de receptores acoplados a proteína G que se acoplan a la proteína G gustducina. (Adaptada con autorización de Lindemann B. Receptors and transduction in taste. Nature. 2001;413:219.) CAPÍTULO 17 Sentidos especiales III: olfato y gusto CORRELACIÓN CLÍNICA Un niño de 10 años de edad iba en el asiento delantero para el pasajero de un automóvil conducido por su padre. Se le cayó su reproductor de MP3, y soltó su cinturón de seguridad para recuperarlo. En ese momento, el automóvil fue golpeado por detrás por un motociclista que iba a exceso de velocidad. Se golpeó con fuerza la nariz contra el tablero de instrumentos. Lo llevaron a la sala de urgencias de un hospital cercano. Una radiografía mostró que se había fracturado el hueso etmoides, el cual separa la cavidad nasal del cerebro. Después de este accidente, perdió el sentido del olfato (anosmia), y su sentido del gusto también estuvo disminuido. El nervio olfatorio corre a través del hueso etmoides. El nervio puede quedar dañado cuando el hueso se fractura, lo que da lugar a la pérdida de la capacidad para oler. Por la estrecha relación entre el gusto y el olfato, la anosmia se asocia con reducción de la sensibilidad al gusto (hipogeusia). Las causas importantes de anosmia son infección de las vías respiratorias superiores, pólipos nasales, traumatismo encefálico, tumores del lóbulo frontal, toxinas, y uso prolongado de descongestionantes nasales. La anosmia, por lo general, es permanente cuando el nervio olfatorio u otros elementos neurales en la vía neural olfatoria quedan dañados. Además de ser incapaz de experimentar el placer de aromas agradables y toda la gama de sabores, los individuos con anosmia tienen riesgo porque son incapaces de detectar el olor de peligros como fugas de gas, incendios y alimento echado a perder. RESUMEN DEL CAPÍTULO ■ ■ ■ ■ ■ ■ ■ Las neuronas sensoriales olfatorias, las células de sostén (sustentaculares) y las células madre basales se encuentran en el epitelio olfatorio dentro de la porción superior de la cavidad nasal. Los cilios que se sitúan en la protuberancia dendrítica de la neurona sensorial olfatoria contienen receptores odorantes que se acoplan a proteínas G heterotriméricas. Los axones de neuronas sensoriales olfatorias hacen contacto con las dendritas de células mitrales y en penacho en los bulbos olfatorios, y forman glomérulos olfatorios. La información que proviene del bulbo olfatorio viaja por medio de la estría olfatoria lateral a la corteza olfatoria, incluso el núcleo olfatorio anterior, el tubérculo olfatorio, la corteza piriforme, amígdala y corteza entorrinal. Las papilas gustativas son los órganos de los sentidos especializados para el gusto, y están compuestas de células madre basales y células gustativas tipos I, II y III que quizá representen diversas etapas de diferenciación de células gustativas en desarrollo. Se ubican en la mucosa de la epiglotis, el paladar y la faringe, y en las paredes de papilas de la lengua. Hay receptores gustativos para sabores dulce, ácido, amargo, salado y umami. Los mecanismos de transducción de señal comprenden el paso por canales iónicos, unión a canales iónicos y bloqueo de los mismos, y sistemas de segundo mensajero. Los aferentes que provienen de papilas gustativas en la lengua viajan por medio del séptimo, noveno y décimo pares craneales 165 para hacer sinapsis en el NTS. Desde ahí, los axones ascienden por medio del lemnisco medial ipsilateral al núcleo ventral posteromedial del tálamo, y hacia la ínsula anterior y el opérculo frontal en la corteza cerebral ipsilateral. PREGUNTAS DE ESTUDIO 1. Los receptores odorantes se sitúan: A) en el bulbo olfatorio. B) sobre dendritas de células mitrales y en penacho. C) sobre neuronas que se proyectan de manera directa a la corteza olfatoria. D) sobre neuronas en el epitelio olfatorio que se proyectan hacia células mitrales y desde ahí de manera directa a la corteza olfatoria. E) sobre células sustentaculares que se proyectan al bulbo olfatorio. 2. Los receptores gustativos: A) para los sabores dulce, ácido, amargo, salado y umami están separados espacialmente sobre la superficie de la lengua. B) son sinónimos de las papilas gustativas. C) son un tipo de quimiorreceptor. D) están inervados por aferentes en los nervios facial, trigémino y glosofaríngeo. E) todas las anteriores son correctas. 3. ¿Cuál de los que siguen no aumenta la capacidad para discriminar muchos olores diferentes? A) muchos receptores distintos B) patrón de receptores olfatorios activados por un odorante dado C) proyección de diferentes axones de célula mitral a distintas partes del cerebro D) procesamiento neural en la amígdala E) inhibición lateral 4. ¿Cuáles de los que siguen están pareados de manera incorrecta? A) ENaC:agrio B) gustducina α:sabor amargo C) núcleo del tracto solitario:gusto D) papilas fungiformes:olor E) glándulas de Ebner:agudeza gustativa 5. ¿Cuál de las que siguen es verdadera acerca de la transmisión olfatoria? A) una neurona sensorial olfatoria expresa una amplia gama de receptores odorantes B) la inhibición lateral dentro de los glomérulos olfatorios reduce la capacidad para distinguir entre diferentes tipos de receptores odorantes C) la discriminación consciente de olores depende de la vía a la corteza orbitofrontal D) la olfacción se relaciona, de manera estrecha, con la gustación porque los receptores odorantes y gustativos usan las mismas vías centrales E) todas las anteriores son correctas 6. ¿Cuál de las que siguen es falsa acerca de la sensación gustativa? A) las fibras nerviosas sensoriales que provienen de las papilas gustativas en los dos tercios anteriores de la lengua viajan en la rama cuerda del tímpano del nervio facial B) las fibras nerviosas sensoriales que se originan en las papilas gustativas en el tercio posterior de la lengua viajan en la rama petrosa del nervio glosofaríngeo 166 SECCIÓN IV Fisiología del sistema nervioso central/neural C) la vía que proviene de las papilas gustativas en el lado izquierdo de la lengua es transmitida de manera ipsilateral a la corteza cerebral D) las células sustentaculares en las papilas gustativas sirven como células madre para permitir el crecimiento de nuevas papilas gustativas E) la vía que proviene de receptores gustativos incluye sinapsis en el núcleo del tracto solitario en el tallo encefálico y el núcleo ventral medial posterior en el tálamo Control de la postura y el movimiento C A P Í T U L O 18 Susan M. Barman O B J E T I V O S ■ ■ ■ ■ ■ ■ Describir cómo los movimientos hábiles se planean y se llevan a cabo. Identificar las partes del sistema nervioso central que regulan la postura. Definir la rigidez de descerebración y de descorticación. Explicar la función de los ganglios basales en el control del movimiento. Describir los síntomas de la enfermedad de Parkinson. Comentar las funciones del cerebelo y las anormalidades neurológicas que producen enfermedades de esta parte del encéfalo. INTRODUCCIÓN La actividad motora somática depende del patrón y de la tasa de descarga de las neuronas motoras espinales y neuronas homólogas en los núcleos motores de los pares craneales. Estas neuronas, la vía común final al músculo esquelético, reciben aferencias desde una gama de vías descendentes, otras neuronas espinales y aferentes periféricos. La integración de estas múltiples aferencias regula la postura del cuerpo y hace posible el movimiento coordinado. Las aferencias desencadenan actividad voluntaria, ajustan la postura del cuerpo para proporcionar un trasfondo estable para el movimiento y coordinan la acción de diversos músculos para hacer los movimientos suaves y precisos. El movimiento voluntario se planea en la corteza, los ganglios basales y en la parte lateral del cerebelo (figura 18-1). Los ganglios basales y el cerebelo canalizan información hacia las cortezas premotora y motora por medio del tálamo. La postura se ajusta de manera continua tanto antes del movimiento como durante el mismo por medio de vías del tallo encefálico descendentes y aferentes periféricos. El movimiento lo suavizan y coordinan las conexiones de porciones mediales e intermedias del cerebelo. Los ganglios basales y la parte lateral del cerebelo forman parte de un circuito de retroacción hacia las cortezas premotora y motora que se relaciona con la planeación del movimiento voluntario y la organización del mismo. CONTROL DE MÚSCULOS AXIALES Y DISTALES Dentro del tallo encefálico y la médula espinal las vías y neuronas que controlan los músculos esqueléticos del tronco y las porciones proximales de las extremidades se localizan en posición medial o ventral. Las vías y las neuronas que tienen que ver con el control de los músculos esqueléticos en las porciones distales de las extremidades se ubican lateralmente. Los músculos axiales tienen que ver con ajustes posturales y movimientos gruesos; los músculos distales de las extremidades median movimientos hábiles y finos. Por ejemplo, neuronas en la porción medial del asta ventral inervan músculos proximales de las extremidades, en particular los flexores, y neuronas del asta ventral lateral inervan músculos distales de las extremidades. De modo similar, el tracto corticoespinal ventral y las vías del tallo encefálico descendentes mediales (tractos rubroespinal, reticuloespinal, tectoespinal y vestibuloespinal) se relacionan con ajustes de músculos proximales y la postura, y los tractos corticoespinal lateral y rubroespinal con músculos distales de las extremidades y, en particular en el caso del tracto corticoespinal, con movimientos voluntarios hábiles. 167 168 SECCIÓN IV Fisiología del sistema nervioso central/neural Planea Ejecuta Ganglios basales Idea Áreas de asociación corticales Cortezas premotora y motora Cerebelo lateral Movimiento Cerebelo intermedio FIGURA 181 Control del movimiento voluntario. Las órdenes para el movimiento voluntario se originan en áreas de asociación corticales. La corteza, los ganglios basales y el cerebelo funcionan de manera cooperativa para planear los movimientos. El movimiento ejecutado por la corteza se transmite por medio de los tractos corticoespinales y los tractos corticobulbares hacia neuronas motoras. El cerebelo proporciona retroacción para ajustar el movimiento y suavizarlo. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) TRACTOS CORTICOESPINAL Y CORTICOBULBAR Los axones de las neuronas que provienen de la corteza motora, los cuales se proyectan hacia neuronas motoras espinales forman los tractos corticoespinales, un fascículo grande de alrededor de un millón de fibras. Alrededor de 80% de estas fibras cruza la línea media en las pirámides bulbares o del bulbo raquídeo para formar el tracto corticoespinal lateral (figura 18-2). El otro 20% forma el tracto corticoespinal ventral, que no cruza la línea media sino hasta que llega al nivel de la médula espinal en la cual termina. Las neuronas del tracto corticoespinal lateral hacen conexiones monosinápticas con neuronas motoras, en especial las que se relacionan con movimientos hábiles, y sobre interneuronas espinales. La trayectoria desde la corteza hacia la médula espinal pasa a través de la corona radiada hacia el extremo posterior de la cápsula interna. Dentro del mesencéfalo fibras del tracto corticoespinal atraviesan el pedúnculo cerebral y la protuberancia anular (puente de Varolio) basilar hasta que llegan a las pirámides bulbares en su camino a la médula espinal. El tracto corticobulbar está compuesto de las fibras que pasan desde la corteza motora hacia neuronas motoras en los núcleos trigeminal, facial e hipogloso. Las neuronas corticobulbares terminan, sea de manera directa sobre los núcleos de pares craneales o sobre sus interneuronas antecedentes dentro del tallo encefálico. Sus axones pasan por la rodilla de la cápsula interna, el pedúnculo cerebral (en posición medial a neuronas del tracto corticoespinal), y descienden con fibras del tracto corticoespinal en la protuberancia anular y el bulbo raquídeo. El sistema motor puede dividirse en neuronas motoras inferior y superior. Las neuronas motoras inferiores son neuronas motoras espinales y craneales que inervan de manera directa músculos esqueléticos. Las neuronas motoras superiores son aquellas en la corteza y el tallo encefálico que activan las neuronas motoras inferiores. Las respuestas fisiopatológicas al daño de neuronas motoras inferior y superior son muy distintivas. El daño de neuronas motoras inferiores se asocia con parálisis flácida, atrofia muscular, fasciculaciones (contracciones musculares visibles que aparecen como movimiento bajo la piel), hipotonía (tono muscular disminuido), e hiporreflexia o arreflexia. El daño de neuronas motoras superiores hace que los músculos se tornen débiles y flá- Circunvolución precentral (área 4, etcétera) Tracto corticoespinal Cápsula interna Decusación de las pirámides Pirámides Tracto corticoespinal ventral (20% de las fibras) Interneurona Tracto corticoespinal lateral (80% de las fibras) Célula del asta anterior Nervio espinal Músculo distal Músculo proximal FIGURA 182 Tractos corticoespinales. Este tracto se origina en la circunvolución precentral y pasa a través de la cápsula interna. Casi todas las fibras se decusan en las pirámides y descienden en la sustancia blanca lateral de la médula espinal para formar la división lateral del tracto que puede hacer conexiones monosinápticas con neuronas motoras espinales. La división ventral del tracto permanece sin cruzarse hasta que llega a la médula espinal, donde los axones terminan en interneuronas espinales antecedentes a neuronas motoras. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) do sd el pie Ma no ado y ce di r Ín ulga llo P ue ja C e C ar cul oo b a glo Car rp Pá CORTEZA MOTORA Y MOVIMIENTO VOLUNTARIO Labios Área motora Corteza motora Corteza sensorial complementaria somática primaria Corteza premotora Corteza parietal posterior 4 6 3,1,2 5 7 7 Corteza prefrontal FIGURA 183 Una vista de la corteza cerebral del ser humano, que muestra la corteza motora (área 4 de Brodmann) y otras áreas que se relacionan con el control del movimiento voluntario, junto con los números asignados a las regiones por Brodmann. (Reproducida con autorización de Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000.) IÓ IC AC MA ST SA díbu Len la gua ció n LIVA CI Ó N Man De glu Las neuronas de los tractos corticoespinal y corticobulbar tienen forma piramidal y se localizan en la capa V de la corteza cerebral (capítulo 12). En la figura 18-3 se muestran las principales regiones corticales involucradas en el control motor. Alrededor de 31% de las neuronas del tracto corticoespinal proviene de la corteza motora primaria (M1; área 4 de Brodmann) en la circunvolución precentral del lóbulo frontal, y se extiende hacia el surco central. La corteza premotora y la corteza motora complementaria (área 6 de Brodmann) explican 29% de las neuronas del tracto corticoespinal. El área premotora es anterior a la circunvolución precentral, en la superficie cortical lateral y medial, y el área motora complementaria está por encima del banco superior del surco cingulado sobre la cara medial del hemisferio. El otro 40% de las neuronas del tracto corticoespinal se origina en el lóbulo parietal (áreas 5, 7 de Brodmann) y el área somatosensorial primaria (áreas 3, 1, 2 de Brodmann) en la circunvolución poscentral. Las diversas partes del cuerpo están representadas en M1; los pies están en la parte superior de la circunvolución, y la cara en la parte inferior (figura 18-4). El área facial se representa de manera bilateral, pero el resto de la representación es unilateral; el área motora cortical controla la musculatura en el lado opuesto del cuerpo. El tamaño N VOCALIZACIÓ De 169 Me ñ An ique M ula ed r io Tronco Cadera lo illa il Rod Tob cidos, pero por último lleva a espasticidad, hipertonía (aumento de la resistencia al movimiento pasivo), reflejos de estiramiento hiperactivos, y reflejo extensor plantar anormal (signo de Babinski). El signo de Babinski es dorsiflexión del dedo gordo y separación y extensión de los otros dedos cuando se estimula la cara lateral de la planta del pie. En adultos, la respuesta normal a esta estimulación es flexión plantar en todos los dedos del pie. Es valioso en la localización de procesos morbosos, pero se desconoce su importancia fisiopatológica. Hombro Codo Muñeca CAPÍTULO 18 Control de la postura y el movimiento FIGURA 184 Homúnculo motor. La figura representa, en un corte coronal de la circunvolución precentral, la ubicación de la representación cortical de las diversas partes. El tamaño de las diversas partes es proporcional al área cortical dedicada a ellas. Compárese con la figura 13-6. (Reproducida con autorización de Penfield W, Rasmussen G: The Cerebral Cortex of Man. Macmillan, 1950.) de la representación cortical de cada parte del cuerpo es proporcional al número de neuronas corticoespinales que inervan la musculatura de esa región del cuerpo, y su papel en el movimiento voluntario fino. De este modo, las áreas involucradas en el lenguaje y en los movimientos de la mano son en especial grandes. Una organización somatotópica continúa a lo largo de las vías corticoespinal y corticobulbar. Las células en las áreas motoras corticales están dispuestas en columnas. Las neuronas en varias columnas corticales se proyectan hacia el mismo músculo; asimismo, las células en cada columna reciben aferencias sensoriales extensas que provienen del área periférica en la cual producen movimiento, y proporciona la base para el control del movimiento por retroacción. El área motora complementaria (figura 18-3), que se proyecta a M1, también contiene un mapa del cuerpo, pero es menos preciso que en M1. Está involucrada en la organización de secuencias motoras o la planeación de las mismas, mientras que M1 ejecuta los movimientos. Cuando los seres humanos cuentan para sí mismos sin hablar, la corteza motora está latente, pero cuando dicen los números en voz alta, el flujo sanguíneo aumenta en M1 y en el área motora complementaria. Así, tanto M1 como el área motora complementaria están involucrados en el movimiento voluntario cuando los movimientos que se realizan son complejos y comprenden planeación. La corteza premotora (figura 18-3), que también contiene un mapa somatotópico, recibe aferencias que provienen de regiones sensoriales de la corteza parietal y se proyectan a M1, la médula espinal y la formación reticular del tallo encefálico. Esta región se relaciona con el establecimiento de la postura al inicio de un movimiento planeado, y con la preparación del individuo para el movimiento. Está involucrada en su mayor parte en el control de los músculos proximales de las extremidades necesarios para orientar el cuerpo para el movimiento. 170 SECCIÓN IV Fisiología del sistema nervioso central/neural Además de proporcionar fibras que corren en los tractos corticoespinal y corticobulbar, el área sensorial somática y porciones del lóbulo parietal posterior se proyectan hacia el área premotora. Algunas de las neuronas en el área 5 (figura 18-3) se relacionan con la dirección de las manos hacia un objeto y la manipulación del mismo, mientras que algunas de las neuronas en el área 7 tienen que ver con la coordinación mano-ojo. VÍAS MEDIAL Y LATERAL DEL TALLO ENCEFÁLICO: POSTURA Y MOVIMIENTO VOLUNTARIO Como se mencionó, las neuronas motoras espinales están organizadas de tal manera que las que inervan los músculos más proximales se sitúan en posición más medial, y las que inervan los músculos más distales lo están en posición más lateral. Esta organización también se refleja en las vías descendentes del tallo encefálico (figura 18-5). Las vías mediales del tallo encefálico, que funcionan junto con el tracto corticoespinal ventral, son los tractos reticuloespinal, vestibuloespinal y tectoespinal de la protuberancia anular y el bulbo raquídeo. Estas vías descienden en las columnas ventrales ipsilaterales de A Vías del tallo encefálico mediales la médula espinal y terminan de manera predominante en interneuronas en la parte ventromedial del asta dorsal para controlar músculos axiales y proximales. Algunas neuronas de la vía medial hacen sinapsis de modo directo sobre neuronas motoras que controlan músculos axiales. Los tractos vestibuloespinales medial y lateral se describieron en el capítulo 16. El tracto medial se origina en los núcleos vestibulares medial e inferior, y se proyecta de manera bilateral hacia neuronas motoras espinales cervicales que controlan la musculatura del cuello. El tracto lateral se origina en los núcleos vestibulares laterales y se proyecta de manera ipsilateral hacia neuronas en todos los niveles espinales. Activa neuronas motoras para los músculos antigravedad (p. ej., extensores de la parte proximal de las extremidades) para controlar la postura y el equilibrio. Los tractos reticuloespinales de la protuberancia anular y el bulbo raquídeo se proyectan hacia todos los niveles espinales. Están involucrados en el mantenimiento de la postura y en la modulación del tono muscular, en especial por medio de una aferencia hacia neuronas motoras γ. Las neuronas reticuloespinales de la protuberancia anular son de manera particular excitatorias, y las neuronas reticuloespinales del bulbo raquídeo son sobre todo inhibitorias. El tracto tectoespinal se origina en el colículo superior del mesencéfalo. Se B Vías del tallo encefálico laterales Tectum Núcleo rojo (parte magnocelular) Formación reticular medial Tracto tectoespinal Núcleos vestibulares lateral y medial Tracto reticuloespinal Tractos vestibuloespinales Tracto rubroespinal FIGURA 185 Vías del tallo encefálico descendentes medial y lateral involucradas en el control motor. A) Las vías mediales (reticuloespinal, vestibuloespinal y tectoespinal) terminan en el área ventromedial de la sustancia gris espinal, y controlan los músculos axiales y proximales. B) La vía lateral (rubroespinal) termina en el área dorsolateral de la sustancia gris espinal y controla los músculos distales. (Reproducida con autorización de Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000.) CAPÍTULO 18 Control de la postura y el movimiento proyecta hacia la parte cervical de la médula espinal contralateral para controlar los movimientos de la cabeza y los ojos. El principal control de los músculos distales es desde el tracto corticoespinal lateral, pero neuronas dentro del núcleo rojo del mesencéfalo cruzan la línea media y se proyectan hacia interneuronas en la parte dorsal lateral del asta ventral espinal para también influir sobre neuronas motoras que controlan músculos distales de las extremidades. Este tracto rubroespinal excita neuronas motoras flexoras e inhibe neuronas motoras extensoras. DESCEREBRACIÓN Y DESCODIFICACIÓN Una transección completa del tallo encefálico entre los colículos superior e inferior permite que las vías del tallo encefálico funcionen de manera independiente de sus aferencias que provienen de estructuras cerebrales más altas. Esto se llama descerebración mediocolicular, y se diagrama en la figura 18-6 con la línea discontinua marcada con A. Esta lesión interrumpe todas las aferencias que provienen de la corteza y el núcleo rojo hacia músculos distales de las extremidades. Las vías reticuloespinales excitadora e inhibidora (sobre todo para músculos extensores posturales) permanecen intactas. La dominancia del impulso desde vías sensoriales ascendentes hacia la vía reticuloespinal excitatoria lleva a rigidez de descerebración, que se caracteriza por hiperactividad en músculos extensores en las cuatro extremidades. Esto semeja lo que surge después de hernia uncal, que se produce por una lesión supratentorial, como se observa en pacientes con tumores grandes o con una hemorragia en el hemisferio cerebral. Después de descerebración, el corte de raíces dorsales que van a una extremidad (línea discontinua marcada con B en la figura 18-6) elimina la hiperactividad de los músculos extensores, lo que sugiere que la rigidez de descerebración es espasticidad debida a facilitación del reflejo de estiramiento miotático. Las aferencias excitadoras desde la vía reticuloespinal activan neuronas motoras γ que activan de manera indirecta neuronas motoras α (por medio de la actividad aferente del huso; capítulo 14). Esto se llama el asa γ. La rigidez de descerebración también puede llevar a activación directa de neuronas motoras α. Si en un animal descerebrado se extirpa el lóbulo anterior del cerebelo (línea discontinua marcada con C en la figura 18-6), se exagera la hiperactividad de los músculos extensores (rigidez de descerebelación). Este corte elimina la inhibición cortical del núcleo fastigial cerebeloso, y de manera secundaria aumenta la excitación de núcleos vestibulares. Esta rigidez no se revierte al cortar las raíces dorsales. La eliminación de la corteza cerebral (descorticación; línea discontinua marcada con D en la figura 18-6) produce rigidez de descorticación, que se caracteriza por flexión de las extremidades superiores en el codo, e hiperactividad extensora de las extremidades inferiores. La flexión puede explicarse por excitación rubroespinal de músculos flexores en las extremidades superiores; la hiperextensión de las extremidades inferiores se debe a los mismos cambios que ocurren después de descerebración mediocolicular. La rigidez de descorticación se observa en el lado hemipléjico en seres humanos después de hemorragias o trombosis en la cápsula interna. Un 60% de las hemorragias intracerebrales ocurre en la cápsula interna, y 10%, cada una, en la corteza cerebral, la protuberancia anular, el tálamo y el cerebelo. 171 GANGLIOS BASALES Los ganglios basales están compuestos de núcleo caudado, putamen, globo pálido, y de manera funcional se relacionan núcleo subtalámico y sustancia negra (figura 18-7). El globo pálido se divide en segmentos externo e interno (GPe y GPi). La sustancia negra se divide en pars compacta y pars reticulada. El núcleo caudado y el putamen se llaman, en conjunto, cuerpo estriado; el putamen y el globo pálido forman el núcleo lenticular. Las principales aferencias hacia los ganglios basales terminan en el cuerpo estriado (figura 18-8). Incluyen la vía corticoestriada excitadora que proviene de M1 y la corteza premotora. También hay una proyección desde núcleos intralaminares del tálamo hacia el cuerpo estriado (vía talamoestriada). Las conexiones entre las partes de los ganglios basales comprenden una proyección nigroestriatal dopaminérgica desde la pars compacta de la sustancia negra hasta el cuerpo estriado, y una proyección GABAérgica correspondiente desde el cuerpo estriado a la pars reticulada de la sustancia negra. El cuerpo estriado se proyecta tanto al GPe como al GPi. El GPe se proyecta hacia el núcleo subtalámico, que a su vez se proyecta hacia el GPe y al GPi. La principal eferencia desde los ganglios basales es desde el GPi por medio del fascículo talámico hasta los núcleos lateral ventral, anterior ventral y centromediano del tálamo. Desde los núcleos talámicos, las fibras se proyectan hacia la corteza prefrontal y premotora. La sustancia negra también se proyecta hacia el tálamo. La principal característica de las conexiones de los ganglios basales es que la corteza cerebral se proyecta hacia el cuerpo estriado, el cuerpo estriado hacia el GPi, el GPi hacia el tálamo, y el tálamo de regreso a la corteza, lo que completa un asa. Las eferencias desde el GPi hacia el tálamo son inhibitorias, mientras que las aferencias desde el tálamo hacia la corteza cerebral son excitatorias. FUNCIÓN Los ganglios basales están involucrados en la planeación del movimiento voluntario y la programación del mismo (figura 18-1). Influyen sobre la corteza motora mediante el tálamo. Asimismo, el GPi se proyecta hacia núcleos del tallo encefálico y desde ahí hacia neuronas motoras en el tallo encefálico y la médula espinal. Tres vías bioquímicas en los ganglios basales en circunstancias normales operan de una manera equilibrada (figura 18-8): 1) el sistema dopaminérgico nigroestriatal, 2) el sistema colinérgico intraestriatal y 3) el sistema GABAérgico, que se proyecta desde el cuerpo estriado hacia el globo pálido y la sustancia negra. Cuando una o más de estas vías se hace disfuncional, ocurren anormalidades motoras características. Las enfermedades de los ganglios basales provocan dos tipos generales de trastornos: hipercinéticos e hipocinéticos. Las enfermedades hipercinéticas son aquellas en las cuales el movimiento es excesivo y anormal, incluso temblor, corea, atetosis y balismo. Las anormalidades hipocinéticas comprenden acinesia y bradicinesia. La corea se caracteriza por movimientos de “baile” rápidos e involuntarios. La atetosis por movimientos con contorsiones lentos y continuos. Los movimientos coreiformes y atetósicos se han comparado con el inicio de movimientos voluntarios que ocurren de una manera involuntaria y desorganizada. En el balismo ocurren movimientos sin sentido, intensos y violentos, involuntarios. La acinesia 172 SECCIÓN IV Fisiología del sistema nervioso central/neural Células de Purkinje Fibras corticorrubrales Corteza cerebral Fibras corticorreticulares Núcleos interpuesto y lateral Núcleo fastigial Fibras cerebelorrubrales Núcleo rojo s ale pin es ulo stib ve ras Fib Fibras rubroespinales Núcleos vestibulares Fibras corticoespinales Fibras reticuloespinales Fibras reticuloespinales RF excitador RF inhibidor del del puente bulbo raquídeo Raíz posterior y ganglio Fibras sensoriales ascendentes vía ALS Huso muscular Músculo extensor Neurona motora γ extensora Neurona motora α flexora Neurona motora α extensora Raíz anterior Lesión/respuesta motora Músculo flexor A = Rigidez extensora en todas las extremidades, rigidez/postura de descerebración A + B = Relajación de la rigidez extensora en la extremidad con raíz seccionada A + C = Aumento leve de la rigidez de descerebración en comparación con A A + C + B = Relajación nula de la rigidez de descerebración D = Flexión de las extremidades superiores, extensión de las extremidades inferiores, rigidez/postura de descorticación FIGURA 186 Circuito que representa lesiones producidas en animales experimentales para replicar déficits por descerebración y por descorticación observados en seres humanos. Las transecciones bilaterales se indican mediante las líneas discontinuas A-D. La descerebración está a nivel mediocolicular (A), la descorticación es rostral a las raíces dorsales, colículo superior, cortadas para una extremidad (B), y extirpación del lóbulo anterior del cerebelo (C). El objetivo fue identificar sustratos anatómicos, de los cuales depende la rigidez/postura de descerebración o de descorticación que se observa en seres humanos con lesiones que aíslan el prosencéfalo del tallo encefálico o separan el tallo encefálico rostral del caudal, y la médula espinal. (Reproducida con autorización de Haines DE [editor]: Fundamental Neuroscience for Basic and Clinical Applications, 3rd ed. Elsevier, 2006.) es la dificultad para iniciar el movimiento, y el movimiento espontáneo disminuido. La bradicinesia es la lentitud del movimiento. ENFERMEDAD DE PARKINSON La enfermedad de Parkinson tiene características tanto hipocinéticas como hipercinéticas. Fue la primera enfermedad que se identificó como consecuencia de una deficiencia de un neurotransmisor específico; se debe a la degeneración de neuronas dopaminérgicas en la pars compacta de la sustancia negra. Es una de las enfermedades neurodegenerativas más comunes; se estima que ocurre en 1 a 2% de los individuos de más de 65 años de edad. Los síntomas aparecen cuando 60 a 80% de las neuronas dopaminérgicas nigroestriatales se degeneran. CAPÍTULO 18 Control de la postura y el movimiento 173 Núcleo caudado Tálamo Tálamo Cápsula interna Ventrículo lateral Núcleo amigdaloide Núcleo caudado Putamen y globo pálido Putamen Vista lateral Núcleo caudado Globo pálido: Segmento externo Segmento interno Tálamo Cá ps ula Sustancia negra inte rna Globo pálido Amígdala Cola del núcleo caudado Putamen Núcleo subtalámico Corte frontal Corte horizontal FIGURA 187 Ganglios basales. Los ganglios basales están compuestos del núcleo caudado, el putamen y el globo pálido, y los que se relacionan de manera funcional con el núcleo subtalámico y sustancia negra. El corte frontal (coronal) muestra la ubicación de los ganglios basales en relación con estructuras circundantes. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) Las características hipocinéticas de la enfermedad de Parkinson son acinesia y bradicinesia, y las características hipercinéticas son rigidez en rueda dentada y temblor en reposo. La falta de actividad motora y la dificultad para el inicio de movimientos voluntarios son notorias. En la enfermedad de Parkinson faltan movimientos incons- Corteza cerebral Glu Globo pálido, ES GABA Cuerpo estriado (acetilcolina) GABA Glu GABA Glu Núcleo subtalámico Tallo encefálico GABA y PPN médula espinal GABA Globo pálido, IS SNPR GABA DA SNPC GABA Tálamo cientes normales, como oscilar los brazos al caminar, las expresiones faciales, y los movimientos nerviosos. La rigidez es diferente de la espasticidad porque la descarga de neurona motora aumenta hacia los músculos tanto agonista como antagonista. El movimiento pasivo de una extremidad encuentra una resistencia plástica, que se siente como si estuviera muerta, que se compara con flexionar un tubo de plomo y, por ende, se llama rigidez en tubo de plomo. A veces tiene lugar una serie de “detenciones” durante el movimiento pasivo (rigidez en rueda dentada), pero está ausente la pérdida repentina de la resistencia que se observa en una extremidad espástica. El temblor, que está presente en reposo y desaparece con la actividad, se debe a contracciones regulares, alternantes, de músculos antagonistas. Un tratamiento común para la enfermedad de Parkinson es la administración de l-DOPA (levodopa). A diferencia de la dopamina, este precursor de la dopamina cruza la barrera hematoencefálica y ayuda a reparar la deficiencia de dopamina. Sin embargo, la degeneración de estas neuronas continúa y en 5 a 7 años los efectos benéficos de la l-DOPA, por lo general, desaparecen. FIGURA 188 Diagrama de las principales conexiones de los ganglios basales. Las líneas continuas indican vías excitatorias, y las líneas discontinuas, vías inhibitorias. Los transmisores están indicados en las vías, cuando se conocen. Glu, glutamato; DA, dopamina. La acetilcolina es el transmisor producido por interneuronas en el cuerpo estriado. SNPR, pars reticulada de la sustancia negra; SNPC, pars compacta de la sustancia negra; ES, segmento externo; IS, segmento interno; PPN, núcleos pedunculopontinos. El núcleo subtalámico también se proyecta hacia la pars compacta de la sustancia negra; esta vía se ha omitido en aras de la claridad. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) CEREBELO El cerebelo está superpuesto sobre los principales sistemas sensoriales y motores en el tallo encefálico, y se conecta a este último por medio de los pedúnculos superior, medio e inferior. Desde un punto de vista funcional, el cerebelo se divide en tres partes (figura 18-9). El vestibulocerebelo tiene conexiones vestibulares y se conecta con el equilibrio y los movimientos de los ojos. El espinocerebelo recibe aferencias propioceptivas desde el cuerpo, así como una copia del “plan motor” que proviene de la corteza motora. Funciona para 174 SECCIÓN IV Fisiología del sistema nervioso central/neural Espinocerebelo Hacia sistemas descendentes mediales Hacia sistemas descendentes laterales lateral al vermis se proyecta hacia las regiones del tallo encefálico, las cuales se relacionan con el control de los músculos distales de las extremidades (vías laterales del tallo encefálico). Ejecución motora ORGANIZACIÓN CELULAR DEL CEREBELO Hacia las cortezas Planeación motora y motora premotora Hacia Equilibrio y núcleos vestibulares movimientos oculares Cerebrocerebelo Vestibulocerebelo FIGURA 189 Divisiones funcionales del cerebelo. (Modificada con autorización de Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000.) suavizar los movimientos activos y para coordinarlos. El cerebrocerebelo en el hemisferio lateral interactúa con la corteza motora en la planeación y la programación de movimientos. La porción medial del cerebelo se llama vermis y se proyecta al área del tallo encefálico que se relaciona con el control de los músculos axiales y proximales de las extremidades (vías del tallo encefálico medial). El área justo La corteza cerebelosa contiene cinco tipos de neuronas: células de Purkinje, granulosas, en cesto, estrelladas y de Golgi (figura 18-10). Las células de Purkinje figuran entre las neuronas de mayor tamaño en el SNC, con arborización dendrítica extensa. Son las únicas neuronas eferentes de la corteza cerebelosa, y sus acciones sinápticas son inhibitorias mediante la liberación de GABA. Las células granulosas inervan las células de Purkinje; sus axones se bifurcan y forman una T. Las ramas de la T son rectas y recorren largas distancias; así, se llaman fibras paralelas. Las fibras paralelas hacen contacto sináptico con las dendritas de muchas células de Purkinje, y liberan glutamato (un neurotransmisor excitador). Los otros tres tipos de neuronas en la corteza cerebelosa son interneuronas inhibitorias que liberan GABA. Las células en cesto reciben aferencias que provienen de las fibras paralelas, y cada una se proyecta hacia muchas células de Purkinje (figura 18-10). Sus axones forman un cesto alrededor del cuerpo celular y el montículo del axón de cada célula de Purkinje que inervan. Las células estrelladas son similares a las células en cesto, pero tienen ubicación más superficial. Las dendritas de las células de Golgi reciben aferencias provenientes de las fibras paralelas. Su cuerpo celular recibe aferencias 5 Célula estrellada Fibras paralelas: axones de células granulosas Axón 1 Célula de Purkinje Capa molecular 1 Capa de Purkinje Axón Capa granular 4 Célula granulosa 2 5 4 3 2 Célula de Golgi 3 Célula en cesto Axones FIGURA 1810 Ubicación y estructura de cinco tipos neuronales en la corteza cerebelosa. Los dibujos se basan en preparaciones teñidas con Golgi. Las células de Purkinje (1) tienen prolongaciones alineadas en un plano; sus axones son la única eferencia desde el cerebelo. Los axones de células granulosas (4) atraviesan y hacen conexiones con prolongaciones de células de Purkinje en la capa molecular. Las células de Golgi (2), en cesto (3), y estrelladas (5) tienen posición, forma, patrón de ramificación y conexiones sinápticas, características. (Reproducida con autorización de Kuffler SW, Nicholls JG, Martin AR: From Neuron to Brain, 2nd ed. Sinauer, 1984.) CAPÍTULO 18 Control de la postura y el movimiento ENFERMEDAD DEL CEREBELO Fibra paralela Corteza cerebelosa + + + + BC GC PC + GR – + – Fibra trepadora – Núcleos profundos NC + + + 175 Fibra musgosa Otras aferencias + FIGURA 1811 Diagrama de conexiones neurales en el cerebelo. Los signos de más (+) y de menos (–) indican si las terminaciones son excitatorias o inhibitorias. BC, célula en cesto; GC, célula de Golgi; GR, célula granulosa; NC, célula en núcleo profundo; PC, célula de Purkinje. Nótese que las PC y las BC son inhibitorias. Las conexiones de las células estrelladas, que no se muestran, son similares a las de las células en cesto, excepto porque terminan en su mayor parte en dendritas de célula de Purkinje. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) desde las fibras musgosas, y sus axones se proyectan hacia las dendritas de las células granulosas. Las aferencias primarias al cerebelo son las fibras musgosas y trepadoras, ambas son excitatorias (figura 18-11). Las fibras trepadoras transmiten aferencias propioceptivas que provienen de una fuente única, los núcleos olivares inferiores. Las fibras musgosas proporcionan aferencias propioceptivas, así como aferencias desde la corteza cerebral por medio de los núcleos de la protuberancia anular. Los circuitos fundamentales de la corteza cerebelosa son sencillos (figura 18-11). Las aferencias de fibras trepadoras ejercen un fuerte efecto excitatorio sobre una célula de Purkinje única, y las aferencias de fibra musgosas ejercen un efecto excitatorio sobre muchas células de Purkinje por medio de las células granulosas. A las células en cesto y a las células estrelladas también las excitan células granulosas por medio de las fibras paralelas, y sus eferencias inhiben la descarga de células de Purkinje (inhibición por anteroacción). A las células de Golgi las excitan los colaterales de fibras musgosas, colaterales de células de Purkinje, y fibras paralelas, e inhiben la transmisión desde fibras musgosas hacia células granulosas. Las eferencias de las células de Purkinje son inhibitorias para los núcleos cerebelosos profundos. Estos núcleos también reciben aferencias excitatorias por medio de colaterales que provienen de las fibras musgosas y trepadoras. Así, casi todos los circuitos cerebelosos parecen relacionarse sólo con la modulación o la cronología de las eferencias excitatorias de los núcleos cerebrales profundos del tallo encefálico y el tálamo. El daño del cerebelo lleva a varias anormalidades características, entre ellas hipotonía, ataxia y temblor de intención. Casi todas las anormalidades se manifiestan durante el movimiento. La ataxia notoria se caracteriza como descoordinación debida a errores del ritmo, el rango, la fuerza y la dirección del movimiento. La ataxia no sólo se manifiesta por la marcha “de embriaguez”, inestable, de base amplia de los pacientes, sino también por defectos de los movimientos hábiles que se involucran en la producción del lenguaje. El individuo hace pausas entre las palabras y sílabas, un fenómeno que se denomina lenguaje escandido. Los movimientos voluntarios también son anormales. Por ejemplo, si se intenta tocar un objeto con un dedo, el dedo pasa de largo a uno u otro lado. Esta dismetría inicia con prontitud una acción correctiva gruesa, pero la corrección pasa de largo al otro lado. En consecuencia, el dedo oscila hacia atrás y hacia adelante. Esta oscilación es el temblor de intención propio de la enfermedad cerebelosa. Otra característica de la enfermedad cerebelosa es la incapacidad para suspender el movimiento con prontitud. Por ejemplo, la flexión del antebrazo contra resistencia es contenida con rapidez cuando la fuerza de resistencia se suspende de manera repentina. El paciente con enfermedad cerebelosa no puede suspender el movimiento de la extremidad, y el antebrazo vuela hacia atrás en un arco amplio (fenómeno de rebote); esta es una de las razones por las cuales estos pacientes muestran disdiadococinesia, la incapacidad para efectuar movimientos opuestos que alternan con rapidez, como la pronación y supinación repetidas de las manos. Por último, los pacientes con enfermedad cerebelosa tienen dificultad para realizar acciones que comprenden el movimiento simultáneo en más de una articulación. Separan esos movimientos y los llevan a cabo en una articulación a la vez (descomposición del movimiento). Las anormalidades motoras que se asocian con daño cerebeloso varían, según la región afectada. Las principales disfunciones que se observan después de daño del vestíbulocerebelo son ataxia, desequilibrio y nistagmo. El daño de la vermis y del núcleo fastigial (parte del espinocerebelo) lleva a alteraciones del control de los músculos axiales y del tronco durante intentos de postura antigravedad, y lenguaje escandido. La degeneración de esta porción del cerebelo puede producirse por deficiencia de tiamina en individuos alcohólicos o desnutridos. Las principales disfunciones que se observan después de daño del cerebrocerebelo son retrasos del inicio de movimientos y descomposición del movimiento. CORRELACIÓN CLÍNICA Hace alrededor de dos años, a los 34 años de edad, un varón presentó debilidad progresiva en la pierna derecha y, por último, todo su lado derecho se debilitó. Fue incapaz de continuar su trabajo como electricista. Experimentó calambres en el músculo de la pantorrilla derecha y contracciones musculares espasmódicas en el brazo y la pierna. Un examen neurológico reveló atrofia muscular, fasciculaciones (contracciones musculares espasmódicas que aparecen como movimientos bajo la piel), e hipotonía de músculos en el brazo y la pierna. También tuvo hiporreflexia notoria. Las pruebas de función sensorial y cognitiva resultaron 176 SECCIÓN IV Fisiología del sistema nervioso central/neural ■ normales. Todos los signos fueron indicativos de una enfermedad de neurona motora inferior que afectó múltiples niveles de la médula espinal. Se le diagnosticó esclerosis lateral amiotrófica (ALS). En el transcurso del año siguiente, la enfermedad progresó hasta el grado en que tuvo dificultad para deglutir (disfagia), de modo que tuvo que alimentarse por medio de una sonda gástrica. Hace unos seis meses presentó dificultad para respirar y fue colocado en un ventilador. Hace una semana murió por neumonía. La ALS es una degeneración progresiva selectiva de neuronas motoras α. “Amiotrófica” significa “falta de nutrición del músculo” y describe la atrofia que presentan los músculos debido al desuso. “Esclerosis” se refiere a la dureza que se siente cuando un patólogo examina la médula espinal en la autopsia; la dureza se debe a la proliferación de astrocitos y formación de tejido cicatrizal en las columnas laterales de la médula espinal. Esta enfermedad mortal también se conoce como enfermedad de Lou Gehrig en reconocimiento de un famoso jugador de béisbol estadounidense que murió por ella. Se estima que la incidencia anual de ALS en todo el mundo es de 0.5 a 3 casos por 100 000 habitantes. Casi todos los casos son esporádicos, pero 5 a 10% es familiar. En 40% de los casos familiares hay una mutación en el gen que codifica para Cu/Zn superóxido dismutasa (SOD-1) en el cromosoma 21. El SOD es un recolector de radicales libres que reduce el estrés oxidativo. Un gen SOD-1 defectuoso permite que se acumulen radicales libres y maten neuronas. La enfermedad no tiene fronteras raciales, socioeconómicas ni étnicas. La esperanza de vida de los pacientes con ALS, por lo general, es de 3 a 5 años después del diagnóstico. La ALS se diagnostica con frecuencia a mediana edad, y afecta más a varones que a mujeres. No están claras las causas de la ALS, pero tal vez incluyan virus, neurotoxinas, metales pesados, defectos del DNA (en especial en la ALS familiar), anormalidades del sistema inmunitario, y anormalidades enzimáticas. No hay curación para la ALS; los tratamientos (p. ej., terapias física y ocupacional) se enfocan en aliviar los síntomas y mantener la calidad de vida. RESUMEN DEL CAPÍTULO ■ ■ ■ El tracto corticoespinal ventral y las vías del tallo encefálico descendentes mediales (tractos tectoespinal, reticuloespinal y vestibuloespinal) regulan los músculos proximales y la postura. Los tractos corticoespinal y rubroespinal laterales controlan los músculos distales de las extremidades y los movimientos voluntarios hábiles. La rigidez de descerebración lleva a hiperactividad en músculos extensores en las cuatro extremidades; en realidad es espasticidad debida a la facilitación del reflejo de estiramiento miotático. La postura de descorticación o rigidez de descorticación es flexión de las extremidades superiores en el codo, e hiperactividad extensora en las extremidades inferiores. Los ganglios basales incluyen el núcleo caudado, el putamen, el globo pálido, núcleo subtalámico y sustancia negra. Las conexiones entre las partes de los ganglios basales incluyen una proyección nigroestriatal dopaminérgica desde la sustancia negra hacia el cuerpo estriado, y una proyección GABAérgica desde el cuerpo estriado hacia la sustancia negra. ■ ■ La enfermedad de Parkinson se debe a degeneración de las neuronas dopaminérgicas nigroestriatales, y se caracteriza por acinesia, bradicinesia, rigidez en rueda dentada y temblor en reposo. La corteza cerebelosa contiene cinco tipos de neuronas: células de Purkinje, granulosas, en cesto, estrelladas y de Golgi. Las dos principales aferencias hacia la corteza cerebral son fibras trepadoras y fibras musgosas. Las células de Purkinje son la única eferencia desde la corteza cerebelosa y, por lo general, se proyectan hacia los núcleos profundos. El daño del cerebelo lleva a varias anormalidades características, entre ellas hipotonía, ataxia y temblor de intención. PREGUNTAS DE ESTUDIO 1. Una función primaria de los ganglios basales es: A) integración sensorial. B) memoria a corto plazo. C) planeación del movimiento voluntario. D) control neuroendocrino. E) sueño de ondas lentas. 2. El efecto terapéutico de la l-DOPA en pacientes con enfermedad de Parkinson por último desaparece porque: A) aparecen anticuerpos contra los receptores de dopamina. B) crecen vías inhibitorias hacia los ganglios basales desde el lóbulo frontal. C) las neuronas colinérgicas en el cuerpo estriado se degeneran. D) la acción normal del factor de crecimiento nervioso (NGF) es alterada. E) las neuronas dopaminérgicas en la sustancia negra se siguen degenerando. 3. La actividad neural aumentada antes de un movimiento voluntario hábil se observa primero en: A) neuronas motoras espinales. B) corteza motora precentral. C) mesencéfalo. D) cerebelo. E) áreas de asociación cortical. 4. Después de caer por un tramo de escaleras, se encuentra que una mujer joven tiene pérdida parcial del movimiento voluntario en el lado derecho del cuerpo, y pérdida de la sensación de dolor y temperatura en el lado izquierdo, por debajo de la región mediotorácica. Es probable que tenga una lesión con transección de: A) la mitad izquierda de la médula espinal en la región lumbar. B) la mitad izquierda de la médula espinal en la región torácica superior. C) vías sensoriales y motoras en el lado derecho de la protuberancia anular. D) la mitad derecha de la médula espinal en la región torácica superior. E) la mitad dorsal de la médula espinal en la región torácica superior. 5. ¿Cuáles de los que siguen no están pareados correctamente? A) vías del tallo encefálico mediales:control de músculos axiales y proximales B) vía del tallo encefálico lateral:tracto rubroespinal C) enfermedad de neurona motora superior:hipotonía D) enfermedad cerebelosa:temblor de intención E) rigidez de descerebración:hiperactividad de músculos extensores Sistema nervioso autónomo C A P Í T U L O 19 Susan M. Barman O B J E T I V O S ■ ■ ■ ■ ■ Describir la ubicación de los cuerpos celulares y las trayectorias axonales de neuronas simpáticas y parasimpáticas preganglionares. Determinar la ubicación y las trayectorias de neuronas simpáticas y parasimpáticas posganglionares. Nombrar los neurotransmisores que son liberados por neuronas preganglionares y posganglionares del sistema nervioso autónomo. Listar las principales funciones del sistema nervioso autónomo. Identificar algunas de las aferencias neurales hacia neuronas simpáticas y parasimpáticas. INTRODUCCIÓN El sistema nervioso autónomo (SNA) es uno de los sistemas de control de los cuales depende la homeostasis, y es la fuente de inervación de órganos que no son el músculo esquelético. Las terminales nerviosas están ubicadas en el músculo liso (p. ej., vasos sanguíneos, pared del intestino, vejiga urinaria), el músculo cardiaco y glándulas (p. ej., glándulas sudoríparas, glándulas salivales). Si bien la supervivencia es posible sin un SNA, la capacidad para adaptarse a factores ambientales que generan estrés y otros desafíos está gravemente comprometida en enfermedades que afectan este componente del sistema nervioso. El SNA tiene dos divisiones principales: sistemas nerviosos simpático y parasimpático, que clásicamente, es definido por las neuronas preganglionares y posganglionares dentro de las divisiones simpática y parasimpática. Una definición más moderna del SNA toma en cuenta las vías descendentes desde varias regiones del prosencéfalo y el tallo encefálico, así como vías aferentes viscerales que establecen la magnitud de actividad en nervios simpáticos y parasimpáticos. ORGANIZACIÓN ANATÓMICA DE LAS EFERENCIAS DEL SNA En la figura 19-1 se comparan algunas características fundamentales de la inervación del músculo esquelético y la inervación del músculo liso, el músculo cardiaco y las glándulas. Las neuronas motoras α sirven como la vía común final que enlaza el sistema nervioso central (SNC) a músculos esqueléticos (capítulo 14). De modo similar, las neuronas simpáticas y parasimpáticas sirven como la vía común final desde el SNC hacia blancos viscerales, sin embargo, la porción periférica del SNA está compuesta de dos neuronas: neuronas preganglionares y posganglionares. Los cuerpos celulares de las neuronas preganglionares están situados en la columna intermediolateral (IML) de la médula espinal, y en núcleos motores de algunos nervios craneales. En contraste con las neuronas motoras α de diámetro grande y conducción rápida, los axones preganglionares son fibras de diámetro pequeño y conducción relativamente lenta. Un axón preganglionar diverge hacia un promedio de alrededor de nueve neuronas posganglionares, lo que hace difusas las eferencias del SNA. Los axones de las neuronas posganglionares son en su mayor parte fibras C no mielinizadas, y terminan en los efectores viscerales. DIVISIÓN SIMPÁTICA En contraste con las neuronas motoras α que están ubicadas en todos los segmentos espinales, las neuronas preganglionares simpáticas están situadas en la IML de sólo el primer segmento torácico al tercer o cuarto segmentos lumbares. Esa es la razón por la cual el sistema nervioso simpático a veces se llama la división toracolumbar del SNA. Los axones de las neuronas preganglionares simpáticas 177 178 SECCIÓN IV Fisiología del sistema nervioso central/neural Sistema nervioso somático CNS Órgano efector ACh Sistema nervioso autónomo: división parasimpática CNS Órgano efector Ganglio ACh Sistema nervioso autónomo: división simpática CNS Órgano efector ACh Ganglio ACh NE (por medio del torrente sanguíneo) Médula suprarrenal Órgano efector Epi (también NE, DA, péptidos) FIGURA 191 Comparación de la organización periférica y los transmisores liberados por los sistemas nerviosos (SN) somatomotor y autónomo. ACh, acetilcolina; DA, dopamina; NE, norepinefrina; Epi, epinefrina. (Reproducida con autorización de Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology. McGraw-Hill, 2008.) abandonan la médula espinal al nivel al cual sus cuerpos celulares están ubicados, y salen a través de la raíz ventral junto con axones de neuronas motoras α y γ (figura 19-2), a continuación se separan de la raíz ventral mediante los ramos comunicantes blancos, y se proyectan hacia el ganglio paravertebral simpático adyacente, donde algunas de ellas terminan en el cuerpo celular de neuronas posganglionares. Los ganglios paravertebrales están situados adyacentes a cada segmento torácico y lumbar superior; además, hay algunos ganglios adyacentes a los segmentos espinales cervical y sacro. Los ganglios paravertebrales forman la cadena simpática (o el tronco simpático) a ambos lados. Los ganglios están conectados entre sí por medio de axones de neuronas preganglionares que viajan en dirección rostral o caudal para terminar en neuronas posganglionares situadas a cierta distancia. Esta disposición se muestra en las figuras 19-2 y 19-3. Algunas neuronas preganglionares pasan a través de la cadena simpática y terminan en neuronas posganglionares ubicadas en ganglios prevertebrales (o colaterales) cerca de las vísceras, incluso los ganglios celiaco, mesentérico superior y mesentérico inferior (figura 19-3). También hay neuronas preganglionares cuyos axones terminan directamente sobre un órgano efector, la médula de la glándula suprarrenal. Los axones de algunas neuronas posganglionares abandonan los ganglios de la cadena y vuelven a entrar en los nervios espinales por medio de los ramos comunicantes grises, y se distribuyen a efectores autónomos en las áreas inervadas por estos nervios espinales (figura 19-2). Estos nervios simpáticos posganglionares terminan sobre músculo liso de vasos sanguíneos y folículos pilosos, y en glándulas sudoríparas en las extremidades. Otras fibras posganglionares salen de los ganglios de la cadena, entran en la cavidad torácica y terminan en órganos viscerales. Las fibras posganglionares de los ganglios prevertebrales también terminan en blancos viscerales. DIVISIÓN PARASIMPÁTICA El sistema nervioso parasimpático a veces se llama la división craneosacra del SNA debido a la ubicación de sus neuronas preganglionares (figura 19-3). Los nervios parasimpáticos inervan las estructuras viscerales de la cabeza por medio de los nervios oculomotor, facial y glosofaríngeo, y las que están en el tórax y la parte alta del abdomen mediante los nervios vagos. Las eferencias sacras inervan las vísceras pélvicas por medio de ramas del segundo a cuarto nervios espinales sacros. Las fibras preganglionares parasimpáticas hacen sinapsis sobre células ganglionares agrupadas dentro de las paredes de órganos viscerales; así, estas fibras posganglionares parasimpáticas son muy cortas. TRANSMISIÓN QUÍMICA EN UNIONES DEL SISTEMA NERVIOSO AUTÓNOMO ACETILCOLINA Y NOREPINEFRINA Los principales agentes transmisores liberados por nervios del SNA son acetilcolina y norepinefrina (figura 19-1). Las neuronas que son colinérgicas (esto es, que liberan acetilcolina) incluyen: 1) todas las neuronas preganglionares, 2) todas las neuronas posganglionares parasimpáticas, 3) neuronas posganglionares simpáticas que inervan glándulas sudoríparas y 4) neuronas posganglionares simpáticas que terminan en vasos sanguíneos en algunos músculos esqueléticos y CAPÍTULO 19 Sistema nervioso autónomo Médula espinal Neurona preganglionar simpática Raíz dorsal Ganglio de la raíz dorsal Tractos espinotalámico y espinorreticular 179 Nervio espinal Raíz ventral Ramo comunicante gris Axón preganglionar Ramo comunicante blanco Neurona posganglionar simpática Ganglio prevertebral Ganglio paravertebral simpático Tronco simpático Hacia células blanco Desde receptores viscerales FIGURA 192 Proyección de fibras preganglionares y posganglionares simpáticas. El dibujo muestra la parte torácica de la médula espinal, ganglios paravertebrales y prevertebrales. Las neuronas preganglionares se muestran en color rojo, las neuronas posganglionares en azul oscuro, las vías sensoriales aferentes en azul, y las interneuronas en negro. (Reproducida con autorización de Boron WF, Boulpaep EL: Medical Physiology. Elsevier, 2005.) producen vasodilatación cuando son estimuladas (nervios vasodilatadores simpáticos). Las neuronas posganglionares simpáticas restantes son noradrenérgicas (esto es, liberan norepinefrina). La médula suprarrenal es en esencia un ganglio simpático en el cual las células posganglionares han perdido sus axones y secretan norepinefrina y epinefrina directamente hacia el torrente sanguíneo. La transmisión en ganglios del SNA está mediada principalmente por receptores colinérgicos nicotínicos N2 que son bloqueados por hexametonio, mientras que la transmisión en la unión neuromuscular es por medio de receptores colinérgicos nicotínicos N1, que son bloqueados por el D-tubocurare. La liberación de acetilcolina a partir de fibras posganglionares actúa sobre receptores muscarínicos, que son bloqueados por la atropina. La liberación de norepinefrina desde fibras posganglionares simpáticas actúa sobre adrenorreceptores α1, 𝛂2, 𝛃1 o 𝛃2, dependiendo del órgano blanco. En el cuadro 19-1 se muestran los tipos de receptores en diversas uniones dentro del SNA. Además de estos neurotransmisores “clásicos”, algunas fibras del SNA liberan neuropéptidos. Las vesículas granuladas pequeñas en neuronas noradrenérgicas posganglionares contienen ATP y norepinefrina, y las vesículas granuladas grandes contienen péptido Y. La estimulación de baja frecuencia puede promover la liberación de ATP, mientras que la estimulación de alta frecuencia puede causar liberación de neuropéptido Y. Las vísceras contienen receptores purinérgicos, y el ATP puede ser un mediador en el SNA junto con la norepinefrina. RESPUESTAS DE ÓRGANOS EFECTORES A IMPULSOS DE NERVIOS DEL SISTEMA NERVIOSO AUTÓNOMO En la figura 19-3 y el cuadro 19-1 se indican los efectos de la estimulación de las fibras nerviosas posganglionares noradrenérgicas y colinérgicas. La liberación de acetilcolina hacia músculo liso de algunos órganos lleva a la contracción (p. ej., las paredes del tracto gastrointestinal), y la liberación hacia otros órganos lleva a relajación (p. ej., esfínteres en el tracto gastrointestinal). Para algunos blancos inervados por el SNA, es posible cambiar desde contracción hacia relajación al cambiar desde activación del sistema nervioso parasimpático hacia activación del sistema nervioso simpático. Así sucede para los muchos órganos que reciben inervación doble con efectos antagonistas, entre ellos el tracto digestivo, las vías respiratorias y la 180 SECCIÓN IV Fisiología del sistema nervioso central/neural División simpática División parasimpática Dilata la pupila y eleva el párpado Cuerpo ciliar Estimula la salivación Modula el tono de vasos sanguíneos Ganglio ciliar Ganglio pterigopalatino CNIX Ganglio submandibular Relaja vías Causa respiratorias erección del pelo Cervical Estimula la secreción por glándulas Acelera sudoríparas los latidos del corazón Inhibe la digestión Torácica Secreta lágrimas CNVII Ganglio cervical superior Craneal CNIII Constriñe la pupila CNX Ganglio ótico Constriñe las vías respiratorias Desacelera los latidos del corazón Cervical Estimula la digestión Torácico Ganglio celiaco Vesícula biliar Estimula la producción de glucosa y su liberación Lumbar Craneal Estimula la vesícula biliar para que libere bilis Lumbar Ganglio aorticorrenal Estimula la secreción de epinefrina Ganglio mesentérico superior Sacra Cadena simpática Ganglio mesentérico inferior Ganglios prevertebrales Nervios esplácnicos pélvicos Dilata vasos sanguíneos en los intestinos y el recto Relaja la vejiga urinaria Sacro Contrae la vejiga urinaria Neuronas simpáticas Relaja el esfínter urinario Plexo pélvico Constriñe el esfínter urinario Estimula la eyaculación Estimula la contracción de músculo liso Preganglionares Estimula la erección del pene Posganglionares Varón Neuronas parasimpáticas Preganglionares Estimula la congestión y las secreciones Mujer Posganglionares FIGURA 193 Organización de los sistemas nerviosos simpático (izquierda) y parasimpático (derecha). Las neuronas simpáticas y parasimpáticas preganglionares se muestran en rojo y anaranjado, respectivamente; las neuronas simpáticas y parasimpáticas posganglionares en azul y verde, respectivamente. (Reproducida con autorización de Boron WF, Boulpaep EL: Medical Physiology. Elsevier, 2005.) 181 CAPÍTULO 19 Sistema nervioso autónomo CUADRO 19-1 Respuestas de algunos órganos efectores a la actividad de nervios del sistema nervioso autónomo. Sistema nervioso simpático Órganos efectores Sistema nervioso parasimpático Tipo de receptor Respuesta Músculo radial del iris —a α1 Contracción (midriasis) Músculo esfínter del iris Contracción (miosis) — Músculo ciliar Contracción para visión cercana — Ojos Corazón Nodo SA Disminuye la frecuencia cardiaca β1 Aumenta la frecuencia cardíaca Aurículas y ventrículos Disminuye la contractilidad β1, β2 Aumenta la contractilidad Nodo AV y fibras de Purkinje Disminuye la velocidad de conducción β1, β2 Aumenta la velocidad de conducción — α1, α2 Constricción β2 Dilatación Arteriolas Coronarias Piel — α1, α2 Constricción Músculo esquelético — α1 Constricción β2, Muscarínico Dilatación Vísceras abdominales — α1 Constricción Glándulas salivales Dilatación α1, α2 Constricción Renal — α1 Constricción — α1, α2 Constricción β2 Dilatación Venas sistémicas Pulmones Músculo bronquial Contracción β2 Relajación Glándulas bronquiales Estimulación α1 Inhibición β2 Estimulación Estómago Motilidad y tono Aumentos α1, α2, β2 Disminuye Esfínteres Relajación α1 Contracción Secreción Estimulación Desconocido Inhibición Motilidad y tono Aumentos α1, α2, β1, β2 Disminuye Esfínteres Relajación α1 Contracción (por lo general) Secreción Estimulación α2 Inhibición Vesícula biliar Contracción β2 Relajación Detrusr Contracción β2 Relajación Esfínter Relajación α1 Contracción Intestino Vejiga urinaria (Continúa) 182 SECCIÓN IV Fisiología del sistema nervioso central/neural CUADRO 19-1 Respuestas de algunos órganos efectores a la actividad de nervios del sistema nervioso autónomo. (Continuación) Sistema nervioso simpático Órganos efectores Sistema nervioso parasimpático Tipo de receptor Respuesta Útero Variable α1 Contracción (en gestación) β2 Relajación α1 Eyaculación α1 Contracción α1 Secreción leve, localizadab Muscarínico Secreción abundante, diluida, generalizada — α1, β2 Glucogenólisis Glándulas exocrinas Aumenta la secreción α Disminuye la secreción Glándulas endocrinas — α2 Inhibe la secreción Secreción profusa, acuosa α1 Secreción viscosa y espesa β Secreción de amilasa Órganos sexuales masculinos Erección Piel Músculos pilomotores — Glándulas sudoríparas Hígado Páncreas Glándulas salivales Glándulas lagrimales Secreción Tejido adiposo — a Un guión significa que estas células no están inervadas por esta división del SNA. b En las palmas y en algunos otros sitios (“sudoración adrenérgica”). — α2, β3 Lipólisis Modificado con autorización de Hardman JG, Limbird LE, Gilman AG [editors]: Goodman and Gilman’s The Pharmacological Basis of Therapeutics, 10th ed. McGraw-Hill, 2001. vejiga urinaria, el corazón es otro ejemplo de un órgano con control antagonista doble. La estimulación de nervios simpáticos aumenta la frecuencia cardiaca, y la de nervios parasimpáticos la disminuye. En otros casos, los efectos de la activación simpática y parasimpática son complementarios, un ejemplo es la inervación de las glándulas salivales, ya que, la activación parasimpática causa liberación de saliva acuosa, mientras que la activación simpática causa la producción de saliva espesa, viscosa. Las dos divisiones del SNA actúan de una manera sinérgica o cooperadora en el control de algunas funciones, un ejemplo es el control del diámetro de la pupila en el ojo, de manera que, las inervaciones tanto simpática como parasimpática son excitatorias, pero la primera contrae el músculo radial para dilatar la pupila (midriasis) y la segunda contrae el músculo esfínter para constreñirla (miosis). Otro ejemplo son las acciones sinérgicas de estos nervios sobre la función sexual, la activación de nervios parasimpáticos que inervan el pene aumenta el flujo de sangre y lleva a erección, mientras que la activación de nervios simpáticos que inervan los genitales masculinos causa eyaculación. También hay varios órganos inervados por sólo una división del SNA, como la médula suprarrenal, casi todos los vasos sanguíneos, los músculos pilomotores en la piel (folículos pilosos) y las glándulas sudoríparas, que están inervados exclusivamente por nervios simpáticos, por otro lado, el músculo lagrimal (la glándula lagrimal), el músculo ciliar (para acomodación para visión cercana), y la glándula salival sublingual están inervados exclusivamente por nervios parasimpáticos. El sistema nervioso parasimpático se relaciona en su mayor parte con los aspectos vegetativos de la vida cotidiana, y a veces se llama el sistema nervioso anabólico, por ejemplo, la acción parasimpática favorece la digestión y absorción de alimentos al incrementar la actividad de la musculatura intestinal, aumentar la secreción gástrica y relajar el esfínter pilórico. El sistema nervioso simpático puede preparar a un individuo para que afronte una emergencia, y puede llamarse el sistema nervioso catabólico. La actividad simpática dilata las pupilas (lo que permite que entre más luz a los ojos), acelera los latidos del corazón y aumenta la presión arterial (lo que proporciona mejor riego de los órganos vitales y los músculos), y constriñe los vasos sanguíneos en la piel (lo que limita el sangrado desde heridas), así como también lleva a incremento de las concentraciones plasmáticas de glucosa y ácidos grasos libres (lo que proporciona más energía). CAPÍTULO 19 Sistema nervioso autónomo CONTROL REFLEJO Y CENTRAL DE LA ACTIVIDAD DEL SISTEMA NERVIOSO AUTÓNOMO Como sucede para las neuronas motoras α, la actividad de los nervios del SNA depende de los reflejos (p. ej., reflejos barorreceptor y quimiorreceptor) y de aferencias excitatorias e inhibitorias descendentes que provienen de varias regiones del cerebro, por ejemplo, las neuronas simpáticas preganglionares en la IML reciben aferencias excitatorias desde la parte ventrolateral rostral del bulbo raquídeo y el núcleo paraventricular del hipotálamo, y aferencias inhibitorias desde neuronas del rafe del bulbo raquídeo. Además de estas vías directas hacia la IML, muchas regiones del cerebro emiten impulsos hacia estas vías para regular la actividad de nervios del SNA, éstas comprenden la parte ventrolateral caudal del bulbo raquídeo, el núcleo del tracto solitario, y el locus cerúleo, esto es análogo al control de la función somatomotora por áreas como los ganglios basales y el cerebelo. En los capítulos 29 y 30 se describen los mecanismos reflejos y centrales que regulan los nervios del SNA en su relación con el control del sistema cardiovascular en salud y enfermedad. El hipotálamo a menudo se considera una importante área central de control del SNA, de hecho, muchos de los mecanismos complejos de dicho sistema que mantienen la homeostasis son integrados en el hipotálamo, que también funciona con el sistema límbico como una unidad que regula la conducta emocional e instintiva, se interconecta con núcleos en el mesencéfalo, la protuberancia anular (puente de Varolio) y el bulbo raquídeo para regular la actividad del SNA. Las respuestas de este último sistema desencadenadas por activación del hipotálamo forman parte de fenómenos complejos como el consumo de alimentos, las emociones como la ira, y respuestas al estrés. CORRELACIÓN CLÍNICA Un profesor de biología de 67 años de edad, jubilado, súbitamente se desplomó en su hogar cuando se levantó luego de estar en una posición reclinada, su esposa llamó una ambulancia y reportó que el paciente estaba pálido pero que nunca perdió el conocimiento. Fue llevado a un hospital local, donde la presión arterial fue normal en una posición reclinada, pero disminuyó hasta cifras hipotensivas cuando se puso de pie. Cuando se le interrogó, reportó que empezó a experimentar disfunción eréctil unos seis meses atrás, también había notado dificultad para adaptarse a temperaturas ambientales alteradas, asimismo, había estado experimentando episodios de incontinencia urinaria y el médico detectó un grado leve de ataxia (marcha inestable) y temblor. Después de una serie de exámenes neurológicos, se le diagnosticó síndrome de Shy-Drager que es un subtipo de atrofia de múltiples sistemas (MSA) en la cual domina la insuficiencia del SNA. La MSA es un trastorno neurodegenerativo asociado con insuficiencia del SNA debida a pérdida de neuronas del SNA preganglionares en la médula espinal y el tallo encefálico. En ausencia de un SNA, es difícil regular la temperatura corporal, el equilibrio de líquidos y electrólitos, y la presión arterial, la 183 MSA también puede presentarse con déficit del cerebelo, los ganglios basales, el locus cerúleo, el núcleo olivar inferior y el tracto piramidal. Se define como un trastorno esporádico, progresivo, de inicio en la adultez, que se caracteriza por disfunción del SNA, parkinsonismo y ataxia cerebelosa en cualquier combinación. El sello patológico característico de la MSA son inclusiones citoplasmáticas y nucleares en oligodendrocitos y neuronas en áreas motoras y del SNA centrales, también hay disminución de marcadores monoaminérgicos, colinérgicos y peptidérgicos en varias regiones del cerebro y en el líquido cefalorraquídeo. La actividad simpática y la concentración plasmática de norepinefrina basales son normales en pacientes con MSA, pero no aumentan en respuesta a ponerse de pie u otros estímulos, lo cual lleva a hipotensión ortostática (presión arterial baja al ponerse de pie) grave, que puede llevar a mareo, oscurecimiento de la visión y desmayo debido a riego insuficiente del cerebro, además, la MSA se acompaña de disfunción parasimpática, incluso disfunción urinaria y sexual. La MSA se diagnostica más a menudo en individuos de 50 a 70 años de edad, y afecta más a varones que a mujeres, la disfunción eréctil suele ser el primer síntoma de la enfermedad, aunque también hay anormalidades del reflejo barorreceptor y de los mecanismos de control respiratorio. Si bien con frecuencia los primeros síntomas son anormalidades del SNA, 75% de los pacientes con MSA también experimenta alteraciones motoras. No hay cura para la MSA, el tratamiento se dirige a asegurar comodidad al paciente y mantener las funciones corporales durante tanto tiempo como sea posible. RESUMEN DEL CAPÍTULO ■ ■ ■ ■ ■ Las neuronas simpáticas preganglionares están situadas en la IML de la parte toracolumbar de la médula espinal, y se proyectan hacia neuronas posganglionares en los ganglios paravertebrales y prevertebrales o la médula suprarrenal. Las neuronas parasimpáticas preganglionares están situadas en núcleos motores de los pares craneales III, VII, IX y X y la IML sacra. Las terminales nerviosas de neuronas posganglionares están ubicadas en el músculo liso (p. ej., vasos sanguíneos, pared del intestino, vejiga urinaria), músculo cardiaco y glándulas (p. ej., glándulas sudoríparas, glándulas salivales). La acetilcolina es liberada en terminales nerviosas de neuronas preganglionares, neuronas parasimpáticas posganglionares, y algunas neuronas simpáticas posganglionares (glándulas sudoríparas, fibras vasodilatadoras simpáticas). Las neuronas posganglionares simpáticas restantes liberan norepinefrina. La actividad simpática puede preparar al individuo para afrontar una emergencia al acelerar los latidos del corazón, aumentar la presión arterial (riego de los órganos vitales), y constreñir los vasos sanguíneos de la piel (limita el sangrado a partir de heridas). La actividad parasimpática está relacionada con los aspectos vegetativos de la vida cotidiana, y favorece la digestión y absorción de alimentos al aumentar la actividad de la musculatura intestinal, incrementar la secreción gástrica, y relajar el esfínter pilórico. La actividad de los nervios del SNA depende tanto de reflejos (p. ej., reflejos barorreceptor y quimiorreceptor) como de 184 SECCIÓN IV Fisiología del sistema nervioso central/neural aferencias excitatorias e inhibitorias descendentes que provienen de varias regiones del cerebro, incluso el hipotálamo y el tallo encefálico. PREGUNTAS DE ESTUDIO 1. La administración de un antagonista de los receptores β-adrenérgicos no se esperaría que: A) disminuyera la velocidad de conducción en las fibras de Purkinje del corazón. B) disminuyera la frecuencia cardiaca. C) disminuyera la fuerza de la contracción cardiaca. D) relajara el músculo detrusor de la vejiga. E) contrajera músculos lisos bronquiales. 2. La actividad nerviosa simpática: A) es esencial para la supervivencia. B) causa contracción de algunos músculos lisos y relajación de otros. C) causa relajación del músculo radial del ojo para dilatar la pupila. D) relaja el músculo liso de la pared gastrointestinal y el esfínter gastrointestinal. E) todas las anteriores. 3. La actividad nerviosa parasimpática: A) relaja casi todo el músculo liso vascular. B) sólo afecta músculos lisos y glándulas. C) causa contracción del músculo radial del ojo para permitir la acomodación para visión cercana. D) contrae el músculo liso de la pared gastrointestinal y relaja el esfínter gastrointestinal. E) todas las anteriores. 4. ¿Cuál de los que siguen están correctamente pareados? A) nodo sinoauricular (SA):receptores colinérgicos nicotínicos B) ganglios del SNA:receptores colinérgicos muscarínicos C) músculo liso pilomotor:receptores β2-adrenérgicos D) vasculatura de algunos músculos esqueléticos: receptores colinérgicos muscarínicos E) glándulas sudoríparas:receptores α2-adrenérgicos Actividad eléctrica del cerebro, estados de sueño-vigilia y ritmos circadianos C A P Í T U L O 20 Susan M. Barman O B J E T I V O S ■ ■ ■ ■ ■ ■ Listar los principales usos clínicos del electroencefalograma. Describir los tipos primarios de ritmos del cerebro. Resumir las características conductuales y electrofisiológicas de cada una de las etapas del sueño sin movimiento rápido de los ojos y con movimiento rápido de los ojos. Describir el patrón de sueño nocturno normal en adultos, y las variaciones de este patrón desde el nacimiento hasta la vejez. Comentar el ritmo circadiano y el papel de los núcleos supraquiasmáticos en su regulación. Describir la regulación diurna de la síntesis de melatonina a partir de serotonina en la glándula pineal, y su secreción hacia el torrente sanguíneo. INTRODUCCIÓN El espectro de estados conductuales varía desde sueño profundo, sueño ligero, sueño con movimiento ocular rápido (REM), y los dos estados de vigilia: vigilia relajada y vigilia con atención concentrada. Patrones específicos de actividad eléctrica del cerebro se correlacionan con cada uno de estos estados, algunos procesos morbosos llevan a cambios en estos patrones de actividad. El despertar puede producirse por estimulación sensorial y por impulsos que ascienden en el centro reticular del mesencéfalo. Muchas de estas actividades son fluctuaciones rítmicas que tienen aproximadamente 24 h de duración (ritmo circadiano). ELECTROENCEFALOGRAMA EPILEPSIA El electroencefalograma (EEG) registrado a partir del cuero cabelludo es una medida de la suma de potenciales postsinápticos dendríticos en neuronas corticales subyacentes. El EEG es valioso en la localización de procesos patológicos en el cerebro. La epilepsia es un síndrome con múltiples causas y se caracteriza por cambios tanto conductuales como en el EEG. En algunas formas, ocurren patrones en el EEG característicos durante crisis convulsivas; entre los ataques a menudo es difícil demostrar anormalidades. Las crisis pueden dividirse en parciales (focales) y generalizadas. Las crisis parciales se originan en un pequeño grupo de neuronas y pueden producirse por lesión encefálica, infección cerebral, apoplejía, o tumor y; los síntomas dependen del foco convulsivo, y se subdividen en crisis parciales simples (sin pérdida del conocimiento) y complejas (con alteración del conocimiento). Un ejemplo de una crisis parcial son los movimientos con sacudidas localizados en una mano, que progresan hacia movimientos clónicos de todo el brazo. Las auras típicamente preceden al inicio de una crisis parcial, e incluyen sensaciones anormales, y el tiempo después de la crisis hasta que regresa la función neurológica normal se llama el periodo posictal. Las crisis generalizadas se asocian con actividad eléctrica diseminada y comprenden ambos hemisferios de manera simultánea y; se subdividen en categorías convulsiva y no convulsiva dependiendo de si ocurren movimientos tónicos o clónicos. Las crisis de ausencia (antes llamadas crisis de pequeño mal) son una de las formas de crisis generalizadas no convulsivas caracterizadas por pérdida momentánea del conocimiento. La crisis generalizada convulsiva más común es la convulsión tónico-clónica (antes llamada crisis de gran mal), que se asocia con inicio repentino de contracción de los músculos de las extremidades (fase tónica) que dura alrededor de 30 segundos, seguida por una 185 186 SECCIÓN IV Fisiología del sistema nervioso central/neural a) Ritmo alfa (relajado, con los ojos cerrados) b) Ritmo beta (alerta) ritmo beta (figura 20-1). Este fenómeno se llama bloqueo alfa (o despertamiento o respuesta alertadora) y puede ser producido por cualquier forma de estimulación sensorial o concentración mental (p. ej., resolución de problemas aritméticos). También se ha llamado desincronización, porque representa el rompimiento de la actividad neural altamente sincronizada, sin embargo, la actividad del EEG rápida que se observa en el estado de alerta también está sincronizada, pero a una tasa más alta, por ende, el término desincronización es desorientador. Las oscilaciones gamma de 30 a 80 Hz a menudo se observan cuando un individuo es despertado y enfoca la atención en algo, esto puede quedar reemplazado por actividad rápida y regular cuando el individuo inicia actividad motora en respuesta al estímulo. Tiempo FIGURA 201 Registros EEG que muestran los ritmos alfa y beta. a) y b) Cuando la atención se enfoca en algo, el ritmo alfa de 8 a 13 Hz es reemplazado por una actividad de bajo voltaje, de 13 a 30 Hz, irregular, el ritmo beta. (Reproducida con autorización de Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology, 11th ed. McGraw-Hill, 2008.) fase con sacudidas simétricas de las extremidades como resultado de contracción y relajación alternantes (fase clónica) que dura 1 a 2 min, hay actividad en el ECG rápida durante la fase tónica, en tanto en el momento de cada sacudida clónica ocurren ondas lentas, cada una precedida por una espiga y durante cierto tiempo después de un ataque hay ondas lentas. RITMOS CEREBRALES EN LA VIGILIA Y EL SUEÑO En seres humanos adultos que están despiertos pero en reposo con la mente divagando y los ojos cerrados, el componente más prominente del EEG es un patrón de ondas bastante regular a una frecuencia de 8 a 13 Hz y amplitud de 50 a 100 μV cuando se registra a partir del cuero cabelludo, este patrón es el ritmo alfa (figura 20-1), es más notorio en los lóbulos parietal y occipital, y se asocia con atención disminuida. Cuando la atención se enfoca en algo, el ritmo alfa queda reemplazado por una actividad de bajo voltaje, de 13 a 30 Hz, irregular; el Despierto Etapa del sueño 1 2 ETAPAS DEL SUEÑO Hay dos clases de sueño: sueño con REM y no-REM (NREM) o sueño de ondas lentas. El sueño REM se llama así debido a los movimientos oculares característicos que ocurren durante esta etapa del sueño. El NREM se divide en cuatro etapas (figura 20-2): una persona que queda dormida entra en sueño en etapa 1, y el EEG muestra un patrón de frecuencia mixta, de bajo voltaje, en esta etapa temprana de sueño de ondas lentas puede observarse un ritmo theta (4 a 7 Hz), durante todo el NREM, hay algo de actividad del músculo esquelético, pero no movimientos oculares. La etapa 2 se caracteriza por la aparición de husos de sueño (12 a 14 Hz) y complejos K bifásicos de alto voltaje ocasionales. En la etapa 3, un ritmo delta de alta amplitud (0.5 a 4 Hz) domina las ondas del ECG. La lentitud máxima se observa con ondas grandes en la etapa 4, de manera que, la característica del sueño profundo es un patrón de ondas lentas rítmicas, que indica sincronización notoria, y se denomina sueño de ondas lentas. Mientras que los ritmos theta y delta son normales durante el sueño, su aparición durante la vigilia es un signo de disfunción cerebral. Las ondas lentas de amplitud alta que se observan en el EEG durante el sueño quedan reemplazadas de manera periódica por actividad EEG de bajo voltaje, rápida, que semeja la que se observa en el estado despierto (figura 20-2), sin embargo, el sueño no se interrumpe; de hecho, en el umbral para el estado de alerta por estímulos sensoriales, está aumentado, y durante esta etapa del sueño 3 4 REM EOG EMG EEG 50 μV 1s FIGURA 202 Actividad EEG y muscular durante diversas etapas del ciclo de sueño-vigilia. El NREM tiene cuatro etapas. La etapa 1 se caracteriza por disminución leve del EEG. La etapa 2 tiene complejos K de amplitud alta y husos. Las etapas 3 y 4 son ondas delta lentas, de amplitud alta. El sueño de REM se caracteriza por movimientos de los ojos, pérdida del tono muscular, y un patrón de actividad de amplitud baja y frecuencia alta. La actividad de voltaje más alto en los trazos EOG durante las etapas 2 y 3 refleja actividad EEG de amplitud alta en las áreas prefrontales más que movimientos de los ojos. EOG, electrooculografía que registra los movimientos oculares; EMG, electromiografía que registra la actividad del músculo esquelético. (Reproducida con autorización de Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000.) 187 CAPÍTULO 20 Actividad eléctrica del cerebro, estados de sueño-vigilia y ritmos circadianos Etapas del sueño Despierto REM 1 2 3 4 Despierto TRASTORNOS DEL SUEÑO La narcolepsia es un trastorno neurológico crónico causado por la incapacidad del cerebro para regular normalmente los ciclos de sueño-vigilia. Hay una pérdida repentina del tono de los músculos voluntarios (cataplexia), una necesidad irresistible final de dormir durante las horas diurnas, y a veces episodios breves de parálisis total al principio del sueño o al final del mismo. La narcolepsia se caracteriza por un inicio repentino de sueño de REM, a diferencia del sueño normal que empieza con sueño, de ondas lentas (NREM). El cerebro de seres humanos con narcolepsia a menudo contiene menos neuronas productoras de hipocretina (orexina) en el hipotálamo, un ataque inmunitario contra estas neuronas puede llevar a su degeneración. La apnea obstructiva del sueño (OSA) es un trastorno común del sueño que comprende cese periódico del flujo de aire durante el sueño, y causa estructura anormal del sueño e hipersomnolencia diurna. La apnea del sueño central es un raro trastorno del sueño que 3 4 5 6 7 3 4 5 6 7 3 4 5 Horas de sueño 6 7 Adultos jóvenes 2 3 4 1 Etapas del sueño En una noche de sueño típica, un adulto joven primero entra en NREM, pasa por las etapas 1 y 2, y pasa 70 a 100 min en las etapas 3 y 4, a continuación el sueño se hace más ligero, y le sigue un periodo de REM. Este ciclo se repite a intervalos de alrededor de 90 min durante toda la noche (figura 20-3), y los ciclos son similares, aunque hay menos sueño en etapas 3 y 4 y más sueño durante el REM hacia la mañana. Ocurren cuatro a seis periodos de REM por cada noche. Si los seres humanos son despertados cada vez que muestran sueño de REM y después se les permite que duerman sin interrupción, muestran una cantidad mucho mayor de sueño de REM que lo normal durante algunas noches. Varios estudios implican que el sueño se necesita para mantener el equilibrio metabólico-calórico, el equilibrio térmico y la competencia inmunitaria. 2 REM 1 Despierto DISTRIBUCIÓN DE LAS ETAPAS DEL SUEÑO Niños 1 Etapas del sueño ocurren movimientos errantes y rápidos de los ojos, lo que explica su nombre. Otra característica del sueño REM es la aparición de potenciales fásicos grandes que se originan en las neuronas colinérgicas en la protuberancia anular (puente de Varolio) y pasan rápidamente hacia el cuerpo geniculado lateral y desde ahí hacia la corteza occipital. Se llaman espigas pontogeniculooccipitales (PGO). El tono de los músculos esqueléticos en el cuello está notoriamente reducido durante el sueño de REM. Los seres humanos despertados durante el sueño REM a menudo reportan que estaban soñando, no así los despertados desde sueño de ondas lentas, de este modo, el sueño de REM y los sueños están estrechamente relacionados. La tomografía por emisión de positrones (PET) de seres humanos en sueño de REM muestra actividad aumentada en el área pontina, la amígdala y la circunvolución cingulada anterior, pero actividad disminuida en las cortezas prefrontal y parietal. La actividad en áreas de asociación visual está aumentada, pero hay una disminución en la corteza visual primaria, esto es congruente con emoción aumentada y operación de un sistema neural cerrado separado de las áreas que relacionan la actividad cerebral con el mundo externo. 2 Ancianos REM 1 2 3 4 1 2 FIGURA 203 Ciclos de sueño normal a diversas edades. Las áreas de color más oscuro indican sueño de REM. (Reproducida con autorización de Kales AM, Kales JD. Sleep disorders. Recent findings in the diagnosis and treatment of disturbed sleep. N Engl J Med. 1974; 290:487.) ocurre cuando el cerebro temporalmente deja de enviar señales a las neuronas motoras frénicas que controlan el diafragma. El sonambulismo, la enuresis nocturna y los terrores nocturnos se denominan parasomnias, que son trastornos del sueño asociados con despertamiento desde NREM y REM. Los episodios de sonambulismo son más comunes en niños que en adultos, predominan en varones, y pueden durar varios minutos, los sonámbulos caminan con los ojos abiertos y evitan obstáculos, pero cuando se les despierta no pueden recordar los episodios. RITMOS CIRCADIANOS Y EL CICLO DEL SUEÑO-VIGILIA Casi todas las células vivas, si no es que todas, en vegetales y animales tienen fluctuaciones rítmicas de su función en un ciclo circadiano. En circunstancias normales, quedan sincronizadas con el ciclo de luz diurno-nocturno en el ambiente. Si no son sincronizadas, los ciclos se desfasan progresivamente del ciclo de luz-oscuridad porque duran más de 24 h o menos. El proceso de sincronización casi siempre depende de los núcleos supraquiasmáticos (SCN) que están 188 SECCIÓN IV Fisiología del sistema nervioso central/neural CH3O Melatonina H H H O C C N C CH3 H H N H Glándula pineal Día Inhibición Neuronas simpáticas Noche Estimulación Tracto retinohipotalámico Núcleo supraquiasmático (el “reloj biológico”) Ganglio cervical superior FIGURA 204 Secreción de melatonina. Las fibras retinohipotalámicas hacen sinapsis en los núcleos supraquiasmáticos (SCN), y hay conexiones desde los SCN hacia neuronas preganglionares simpáticas en la médula espinal que se proyectan hacia el ganglio cervical superior. Neuronas posganglionares se proyectan desde este ganglio hacia la glándula pineal que secreta melatonina. La actividad cíclica del SCN establece un ritmo circadiano para la liberación de melatonina. Este ritmo es sincronizado con ciclos de luz/oscuridad por neuronas en la retina. (Reproducida con autorización de Fox SI: Human Physiology. McGraw-Hill, 2008.) situados por arriba del quiasma óptico (figura 20-4). El SCN recibe información acerca del ciclo de luz-oscuridad mediante una vía neural especial, el tracto retinohipotalámico. Fibras eferentes que provienen del SCN inician señales neurales y humorales que sincronizan una amplia variedad de ritmos circadianos, incluso el ciclo del sueño-vigilia y la secreción de la hormona pineal melatonina. La exposición a luz brillante puede adelantar el ciclo del sueñovigilia, retrasarlo o tener efecto nulo sobre el mismo, dependiendo de la hora del día a la cual se experimenta. Durante las horas diurnas habituales carece de efecto, pero justo después de que cae la noche retrasa el inicio del periodo de sueño, y justo antes del amanecer acelera el inicio del siguiente periodo de sueño. Las inyecciones de melatonina tienen efectos similares. MECANISMOS NEUROQUÍMICOS QUE PROMUEVEN EL SUEÑO Y EL DESPERTAMIENTO Las transiciones entre sueño y vigilia manifiestan un ritmo circadiano que consta de un promedio de 8 h de sueño y 16 h de vigilia. Núcleos tanto en el tallo encefálico como en el hipotálamo son cru- ciales para las transiciones entre estos estados de conocimiento. El sistema activador reticular (RAS) del tallo encefálico está compuesto de varios grupos de neuronas que liberan norepinefrina, serotonina o acetilcolina. En el caso de las neuronas del prosencéfalo involucradas en el control de los ciclos de sueño-vigilia, las neuronas preópticas en el hipotálamo liberan GABA, y las neuronas hipotalámicas posteriores liberan histamina. Una teoría respecto a la base para las transiciones desde el sueño hacia la vigilia comprende actividad recíproca alternante de diferentes grupos de neuronas del RAS. En este modelo (figura 20-5), la vigilia y el sueño de REM se encuentran en extremos opuestos. Cuando la actividad de neuronas que contienen norepinefrina y que contienen serotonina (locus cerúleo y núcleos del rafe) es dominante, hay actividad reducida en neuronas que contienen acetilcolina en la formación reticular pontina. Este patrón de actividad contribuye a la aparición del estado de conciencia. El inverso de este patrón lleva al sueño de REM. Cuando hay un equilibrio más uniforme de la actividad de neuronas aminérgicas y colinérgicas, ocurre el NREM. Además, una liberación aumentada de GABA y reducida de histamina incrementa la probabilidad de NREM por medio de desactivación del tálamo y la corteza. La vigilia ocurre cuando la liberación de GABA es reducida y la liberación de histamina es aumentada. CAPÍTULO 20 Actividad eléctrica del cerebro, estados de sueño-vigilia y ritmos circadianos Núcleos del tallo encefálico que forman parte del sistema activador reticular Norepinefrina y serotonina Norepinefrina y serotonina Acetilcolina Acetilcolina Estado de alerta Sueño NREM Activación del tálamo y la corteza Activación del tálamo y la corteza Histamina Histamina GABA GABA Sueño REM 189 coordinar eventos con el ciclo de luz-oscuridad en el ambiente. La síntesis de melatonina y su secreción están aumentadas durante el periodo oscuro del día, y se mantienen a una cifra baja durante las horas diurnas, esta variación diurna de la secreción es desencadenada por norepinefrina secretada por los nervios simpáticos posganglionares que inervan la glándula pineal (figura 20-4). La norepinefrina actúa por medio de receptores beta-adrenérgicos para incrementar el cAMP intracelular, y el cAMP a su vez produce un notorio aumento de la actividad de N-acetiltransferasa. Esto da lugar a incremento de la síntesis de melatonina y la secreción de la misma. La melatonina circulante es metabolizada con rapidez en el hígado mediante 6-hidroxilación seguida por conjugación, y más de 90% de la melatonina que aparece en la orina está en la forma de conjugados 6-hidroxi, y de 6-sulfatoximelatonina. El mecanismo mediante el cual el cerebro metaboliza melatonina quizá comprende la división del núcleo indol. La descarga de los nervios simpáticos hacia la pineal es sincronizada con el ciclo de luz-oscuridad en el ambiente por medio de las fibras nerviosas retinohipotalámicas que van al SCN. Desde el hipotálamo, vías descendentes convergen en neuronas simpáticas preganglionares que a su vez inervan el ganglio cervical superior, el sitio de origen de las neuronas posganglionares que van hacia la glándula pineal. CORRELACIÓN CLÍNICA Hipotálamo con centros circadianos y homeostáticos FIGURA 205 Un modelo de cómo la actividad alternante de neuronas del tallo encefálico e hipotalámicas puede influir sobre los diferentes estados de conocimiento. (Reproducida con autorización de Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology, 11th ed. McGraw-Hill, 2008.) MELATONINA Y EL ESTADO DE SUEÑOVIGILIA La liberación de melatonina a partir de la ricamente vascularizada glándula pineal también desempeña un papel en los mecanismos del sueño (figura 20-4). La pineal surge del techo del tercer ventrículo en el diencéfalo, y está encapsulada por las meninges. El estroma pineal contiene glía y pinealocitos con características que sugieren que tienen una función secretora, al igual que otras glándulas endocrinas, tiene capilares fenestrados altamente permeables. En lactantes, la glándula pineal es grande y las células tienden a estar dispuestas en alvéolos, empieza a involucionar antes de la pubertad, y aparecen pequeñas concreciones de fosfato y carbonato de calcio (arena pineal) en el tejido. Dado que las concreciones son radiopacas, la glándula pineal a menudo es visible en radiografías del cráneo en adultos. El desplazamiento de una pineal calcificada desde su posición normal indica la presencia de una lesión ocupadora de espacio, como un tumor en el cerebro. La melatonina es sintetizada por pinealocitos y secretada hacia la sangre y el líquido cefalorraquídeo. El cambio diurno de la secreción de melatonina tal vez funcione como una señal cronológica para Un varón de 47 años de edad, científico investigador en un Instituto de biotecnología siempre ha sido querido por su actitud positiva y su alto nivel de energía. Recientemente, sus colegas han notado que se pone nervioso si las cosas no salen bien en el laboratorio, y parece estar cansado todo el tiempo, su esposa también ha estado preocupada respecto a que está más malhumorado que lo normal, lo acompaña al consultorio de un médico para su reconocimiento médico anual. Cuando el médico interroga al paciente respecto a su patrón de sueño, él insiste en que no ha habido cambio, pero su esposa menciona que se ha estado despertando repentinamente por la noche y haciendo ruidos muy fuertes, como respiración entrecortada. El paciente es remitido a una clínica del sueño, donde se le practica una polisomnografía (PSG) que registra la actividad del cerebro, los movimientos oculares, los movimientos del cuerpo, la respiración y la frecuencia cardiaca, así como la saturación de oxígeno en sangre en el transcurso de un ciclo de sueño. Los resultados de la PSG confirmaron la sospecha del médico, de que el científico tenía apnea obstructiva del sueño (OSA). La OSA es la causa más común de somnolencia diurna (hipersomnolencia) debida a sueño fragmentado por la noche, y afecta a alrededor de 24% de los varones y 9% de las mujeres de mediana edad en Estados Unidos. La respiración cesa durante más de 10 s durante episodios frecuentes de obstrucción de las vías respiratorias superiores (en especial la faringe) debido a reducción del tono muscular en dichas vías a pesar de actividad continua de los músculos de la inspiración. La apnea causa despertamientos breves para restablecer el tono de las vías respiratorias superiores. Los ronquidos son una queja común del paciente, y la OSA a menudo se asocia con obesidad. En reali- 190 SECCIÓN IV Fisiología del sistema nervioso central/neural dad no hay una disminución del tiempo de sueño total, pero los individuos con OSA experimentan un tiempo mucho mayor de NREM en etapa 1 (desde un promedio de 10% del sueño total hasta 30 a 50%) y una notoria reducción del sueño de ondas lentas (NREM en etapas 3 y 4). La fisiopatología de la OSA comprende tanto una reducción del tono neuromuscular al principio del sueño como un cambio del impulso respiratorio central. Un tratamiento común de la OSA es el uso de una mascarilla de presión positiva continua en las vías respiratorias (CPAP) durante el sueño a fin de prevenir oclusiones de las vías respiratorias. RESUMEN DEL CAPÍTULO ■ ■ ■ ■ ■ ■ ■ ■ El EEG tiene cierta utilidad para localizar procesos patológicos, y es útil para caracterizar diferentes tipos de epilepsia. Los principales ritmos en el EEG son oscilaciones alfa (8 a 13 Hz), beta (13 a 30 Hz), theta (4 a 7 Hz), delta (0.5 a 4 Hz) y gamma (30 a 80 Hz). Durante todo el NREM, hay algo de actividad del músculo esquelético, un ritmo theta puede observarse durante la etapa 1 del sueño. La etapa 2 se caracteriza por la aparición de husos de sueño y complejos K ocasionales. En la etapa 3, un ritmo delta es dominante. En la etapa 4 se observa disminución máxima con ondas lentas grandes. El sueño de REM se caracteriza por actividad EEG de bajo voltaje y alta frecuencia, y movimientos errantes y rápidos de los ojos. Un adulto joven típicamente pasa por las etapas 1 y 2, y pasa 70 a 100 min en las etapas 3 y 4. El sueño después se hace ligero, y le sigue un periodo de REM. Este ciclo se repite a intervalos de 90 min durante toda la noche. Las transiciones desde sueño hacia vigilia pueden comprender actividad recíproca alternante de diferentes grupos de neuronas del RAS. Cuando la actividad de neuronas que contienen norepinefrina y serotonina es dominante, la actividad en neuronas que contienen acetilcolina está reducida, lo que lleva a la aparición de vigilia. El inverso de este patrón lleva a sueño de REM. Asimismo, la vigilia ocurre cuando se reduce la liberación de GABA y aumenta la de histamina. La sincronización de procesos biológicos al ciclo de luz-oscuridad está regulada por el SCN. El cambio diurno de la secreción de melatonina a partir de serotonina en la glándula pineal tal vez funcione como una señal de cronología para coordinar eventos con el ciclo de luz-oscuridad, incluso el ciclo de sueño-vigilia. PREGUNTAS DE ESTUDIO 1. En un adulto sano, alerta, sentado con los ojos cerrados, el ritmo EEG dominante que se observa con electrodos sobre los lóbulos occipitales es: A) delta (0.5 a 4 Hz). B) theta (4 a 7 Hz). C) alfa (8 a 13 Hz). D) beta (13 a 30 Hz). E) actividad rápida, irregular, de bajo voltaje. 2. ¿Cuál de los patrones que siguen, de cambios de neurotransmisores o neuromoduladores centrales, se asocia con la transición desde NREM hacia vigilia? A) disminución de norepinefrina, aumento de serotonina, aumento de acetilcolina, disminución de histamina, y disminución de GABA B) disminución de norepinefrina, aumento de serotonina, aumento de acetilcolina, disminución de histamina, y aumento de GABA C) disminución de norepinefrina, disminución de serotonina, aumento de acetilcolina, aumento de histamina y aumento de GABA D) aumento de norepinefrina, aumento de serotonina, disminución de acetilcolina, aumento de histamina y disminución de GABA E) aumento de norepinefrina, disminución de serotonina, disminución de acetilcolina, aumento de histamina, y disminución de GABA 3. Un ritmo gamma (30 a 80 Hz): A) es característico de la actividad convulsiva. B) se observa en un individuo que está despierto pero no enfocado. C) puede ser un mecanismo para conjuntar información sensorial hacia una percepción y acción únicos. D) también se llama una respuesta alertadora. E) es característico de la apnea obstructiva del sueño. 4. La secreción de melatonina probablemente no sería aumentada por: A) estimulación de los ganglios cervicales superiores. B) una reducción de la luz solar. C) administración de norepinefrina por vía intravenosa lenta. D) estimulación del nervio óptico. E) aumento de la actividad de N-acetiltransferasa. 5. Las crisis de ausencia son una forma de: A) crisis generalizadas no convulsivas acompañadas de pérdida momentánea del conocimiento. B) crisis parciales complejas acompañadas de pérdida momentánea del conocimiento. C) crisis generalizadas no convulsivas sin pérdida del conocimiento. D) crisis parciales simples sin pérdida del conocimiento. E) crisis generalizadas convulsivas acompañadas de pérdida momentánea del conocimiento. 6. La narcolepsia es desencadenada por anormalidades en: A) los músculos esqueléticos. B) la médula oblongada (bulbo raquídeo). C) el hipotálamo. D) el bulbo olfatorio. E) la neocorteza. Aprendizaje, memoria, lenguaje y habla Susan M. Barman C A P Í T U L O 21 O B J E T I V O S ■ ■ ■ ■ ■ ■ Describir los diversos tipos de memoria a largo plazo. Definir plasticidad sináptica, potenciación a largo plazo, depresión a largo plazo, habituación y sensibilización, y sus papeles en el aprendizaje y la memoria. Listar las partes del cerebro involucradas en la memoria, y su papel en el procesamiento de memoria y el almacenamiento de la misma. Describir las anormalidades de la estructura y función del cerebro que se encuentran en la enfermedad de Alzheimer. Definir los términos hemisferio categórico y hemisferio representacional. Explicar las diferencias entre afasia fluente y no fluente. INTRODUCCIÓN La disponibilidad difundida de la tomografía por emisión de positrones (PET), imagen de resonancia magnética funcional (fMRI) y técnicas relacionadas ha desencadenado una revolución en el entendimiento de la función del cerebro. La PET a menudo se usa para medir el metabolismo de glucosa local, que es proporcional a la actividad neural, y la fMRI se usa para medir cantidades locales de sangre oxigenada. Estas técnicas hacen posible determinar la actividad en diversas partes del cerebro en sujetos sanos y en aquellos con diversas enfermedades. Se han usado para estudiar no sólo respuestas sencillas, sino también aspectos complejos del aprendizaje, la memoria y la percepción. En la figura 21-1 se muestra un ejemplo del uso de PET para estudiar las funciones de la corteza cerebral en el procesamiento de palabras. Diferentes porciones de la corteza son activadas cuando una persona está oyendo, viendo, hablando o generando palabras. declarativa se asocia con la conciencia (o al menos apreciación, y su retención depende del hipocampo y otras partes de los lóbulos temporales mediales del cerebro. La memoria implícita o no declarativa no comprende apreciación, y su retención por lo general no involucra procesamiento en el hipocampo. APRENDIZAJE Y MEMORIA Aprendizaje es la adquisición de información que hace posible alterar la conducta con base en la experiencia, y memoria es la retención de esa información y el almacenamiento de la misma. Las dos obviamente están estrechamente relacionadas, y deben considerarse juntas. Desde un punto de vista fisiológico, la memoria se divide en formas explícita e implícita (figura 21-2). La memoria explícita o FIGURA 211 Imágenes de áreas activas del cerebro en un varón (izquierda) y en una mujer (derecha) durante una tarea de lenguaje. Note que los varones usan sólo un lado del cerebro, mientras que las mujeres usan ambos lados del cerebro cuando se está procesando lenguaje. (Reproducida con autorización de Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology. McGraw-Hill, 2008.) 191 192 SECCIÓN IV Fisiología del sistema nervioso central/neural Dos formas de memoria a largo plazo Implícita (no declarativa) Explícita (declarativa) Hechos (semántica) Eventos (episódica) Imprimación Procedural (habilidades y hábitos) FIGURA 212 Formas de memoria a largo plazo. (Modificada con autorización de Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000.) Aprendizaje asociativo: condicionamiento clásico y operante Aprendizaje no asociativo: habituación y sensibilización Respuestas Musculatura emocionales esquelética Lóbulo temporal medial Hipocampo La memoria explícita se divide en memoria episódica para eventos y memoria semántica para hechos (p. ej., palabras, reglas y lenguaje). Los recuerdos explícitos inicialmente requeridos para actividades como andar en bicicleta pueden hacerse implícitos una vez que la tarea se aprende a fondo. La memoria implícita se subdivide en cuatro tipos. La memoria procedural incluye habilidades y hábitos, que, una vez adquiridos, se hacen inconscientes y automáticos. Imprimación es la facilitación del reconocimiento de palabras u objetos mediante exposición previa a ellos. Un ejemplo es el recuerdo mejorado de una palabra cuando se presentan las primeras letras de ella. En el aprendizaje no asociativo, se aprende acerca de un estímulo único. En el aprendizaje asociativo, se aprende acerca de la relación de un estímulo con otro. La memoria explícita y muchas formas de memoria implícita comprenden: 1) memoria a corto plazo, que dura segundos a horas, durante la cual el procesamiento en el hipocampo y en otros sitios lleva a cambios a largo plazo de la fuerza sináptica y 2) memoria a largo plazo, que almacena recuerdos durante años. Durante la memoria a corto plazo, los rastros de memoria están sujetos a alteración por traumatismos y diversos fármacos, mientras que los rastros de memoria a largo plazo son notoriamente resistentes a la alteración. La memoria de trabajo es una forma de memoria a corto plazo que mantiene disponible la información, por lo general durante periodos muy cortos, mientras que el individuo planea acción con base en ella. ELASTICIDAD SINÁPTICA Y APRENDIZAJE La clave para la memoria es la alteración de la fuerza de conexiones sinápticas seleccionadas. En todos los casos, salvo en los más simples, la alteración comprende activación de genes y síntesis de proteína, esto ocurre durante el cambio desde memoria de trabajo a corto plazo hacia memoria a largo plazo, si una intervención ocurre demasiado pronto después de una sesión de entrenamiento, la adquisición de memoria a largo plazo queda alterada. Esto es ejemplificado por la pérdida de memoria para los eventos que preceden inmediatamente a una conmoción cerebral o a terapia electroconvulsiva (amnesia retrógrada). Neocorteza Cuerpo estriado Amígdala Cerebelo Vías reflejas Los cambios de la función sináptica a corto y largo plazos pueden ocurrir como resultado del antecedente de descarga en una sinapsis, es decir, la conducción sináptica se puede fortalecer o debilitar con base en la experiencia; estos cambios, que pueden ser presinápticos o postsinápticos, despiertan gran interés porque representan formas de aprendizaje y memoria. Una forma de cambio plástico es la potenciación postetánica, la producción de potenciales postsinápticos aumentados en respuesta a estimulación. Este aumento dura hasta 60 segundos, y ocurre tras una breve serie de estímulos (tetanizante) en una neurona presináptica. La estimulación hace que se acumule Ca2+ en la neurona presináptica a un grado tal que los sitios de unión intracelulares que mantienen bajo el Ca2+ citoplasmático quedan abrumados. La habituación es una forma de aprendizaje simple en la cual un estímulo neuronal es repetido muchas veces. La primera vez que se aplica es nuevo y desencadena una reacción (la respuesta de “¿qué es?”); sin embargo, conforme se repite, desencadena una respuesta eléctrica cada vez menor, y finalmente, el sujeto queda habituado al estímulo y hace caso omiso del mismo, esto se asocia con liberación disminuida de neurotransmisor desde la terminal presináptica debido a Ca2+ disminuido. El decremento del Ca2+ intracelular se debe a una desactivación gradual de canales de Ca2+, que puede ser a corto plazo, o puede ser prolongado si la exposición al estímulo benigno se repite muchas veces. La habituación es un ejemplo clásico de aprendizaje no asociativo. Sensibilización es la aparición prolongada de respuestas postsinápticas aumentadas después de que un estímulo al cual un individuo ha quedado habituado es pareado una o varias veces con un estímulo nocivo, se debe a facilitación presináptica y puede ocurrir como una respuesta transitoria, y si se refuerza mediante aparejamientos adicionales del estímulo nocivo y el estímulo inicial, puede mostrar características de memoria a corto o largo plazos. La prolongación a corto plazo de la sensibilización se debe a un cambio (mediado por Ca2+) de la adenilil ciclasa que aumenta la producción de cAMP. La potenciación a largo plazo (LTP) comprende síntesis de proteína y crecimiento de las neuronas presináptica, postsináptica y sus conexiones. La LTP es un aumento persistente, que aparece con rapidez, de la respuesta de potencial postsináptica a la estimulación presináptica tras un periodo breve de estimulación rápidamente repetida de la neurona CAPÍTULO 21 Aprendizaje, memoria, lenguaje y habla presináptica, semeja la potenciación postetánica, pero es mucho más prolongada y puede durar días. A diferencia de la potenciación postetánica, es iniciada por un aumento del Ca2+ intracelular en la neurona postsináptica más que en la presináptica, ocurre en muchas partes del SNC, pero se ha estudiado con mayor detalle en el hipocampo, donde hay dos formas: LTP de fibra musgosa, que es presináptica e independiente de receptores N-metil-D-aspartato (NMDA), y LTP colateral de Schaffer, que es postsináptica y dependiente de receptores NMDA. La base hipotética para esta última forma se resume en la figura 21-3. La base de la LTP de fibra musgosa parece incluir cAMP e Ih, un canal catiónico activado por hiperpolarización. La depresión a largo plazo (LTD) se encuentra en todo el cerebro en las mismas fibras que la LTP. Se caracteriza por un decremento de la fuerza sináptica, que es producida por estimulación más lenta de neuronas presinápticas y se asocia con un aumento de menor magnitud del Ca2+ intracelular que el que ocurre en la LTP. 193 Corteza prefrontal Hipocampo Corteza parahipocampal FIGURA 214 Áreas relacionadas con la codificación de recuerdos explícitos. La corteza prefrontal y la corteza parahipocampal del cerebro son activas durante el codificado de recuerdos. (Adaptada con autorización de Rugg Russ MD. Memories are made of this. Science.1998;281(5380):1151-1152.) MEMORIA DE TRABAJO Mg2+ Ca2+ Na+ Glu PS NMDA AMPA P Ca2+ AMPA CaM CaM kII FIGURA 21-3 Producción de potenciación a largo plazo (LTP) en colaterales de Schaffer en el hipotálamo. El glutamato (Glu) liberado a partir de la neurona presináptica se une a receptores α-amino-3hidroxil-5-metil-4-isoxazol-propionato (AMPA) y N-metil-D-aspartato (NMDA) en la membrana de la neurona postsináptica. La despolarización desencadenada por activación de los receptores AMPA alivia el bloqueo de Mg2+ en el canal del receptor NMDA, y entra Ca2+ a la neurona con Na+. El incremento del Ca2+ citoplasmático activa la calmodulina (CaM), que a su vez activa la Ca2+/calmodulina cinasa II (CaM kII). La cinasa fosforila los receptores AMPA (P), lo que aumenta su conductancia, y mueve más receptores AMPA hacia la membrana de la célula sináptica desde sitios de almacenamiento citoplasmáticos. Además, una señal química (PS) puede pasar a la neurona presináptica, lo que produce un incremento a largo plazo de la liberación cuántica de glutamato. (Cortesía de R. Nicoll.) Las áreas de memoria de trabajo están conectadas al hipocampo y las porciones parahipocampales adyacentes de la corteza temporal medial (figura 21-4). La destrucción bilateral del hipocampo ventral, o enfermedad de Alzheimer (véase más adelante) y procesos morbosos similares que destruyen sus neuronas CA1, causan defectos notorios en la memoria a corto plazo. Los individuos con esa destrucción tienen memorias de trabajo y remota intactas, sus procesos de memoria implícita por lo general están intactos. Tienen rendimiento adecuado respecto a memoria consciente en tanto se concentran en lo que están haciendo, empero, si son distraídos durante incluso un periodo breve, se pierde toda la memoria de lo que estaban haciendo, y de lo que se proponían hacer. Así, son capaces de aprendizaje nuevo y retienen recuerdos previos a la lesión, antiguos, pero no pueden formar nuevos recuerdos a largo plazo. El hipocampo está estrechamente asociado con la corteza parahipocampal suprayacente en el lóbulo frontal medial (figura 21-4). Cuando los sujetos recuerdan palabras, la actividad en su lóbulo frontal izquierdo y su corteza parahipocampal izquierda aumenta, pero cuando recuerdan imágenes o escenas, la actividad tiene lugar en su lóbulo frontal derecho y en la corteza parahipocampal en ambos lados. Las conexiones en el hipocampo que van hacia el diencéfalo también están involucradas en la memoria. Algunas personas con daño cerebral relacionado con alcoholismo presentan deterioro de la memoria reciente, y la pérdida de memoria se correlaciona bien con la presencia de cambios patológicos en los cuerpos mamilares, que tienen conexiones eferentes extensas con el hipocampo por medio del fórnix. Los cuerpos mamilares se proyectan hacia el tálamo anterior por medio del tracto mamilotalámico. Desde el tálamo, las fibras relacionadas con la memoria se proyectan hacia la corteza prefrontal y desde ahí hacia el prosencéfalo basal, desde este último, una 194 SECCIÓN IV Fisiología del sistema nervioso central/neural proyección colinérgica difusa va a toda la neocorteza, la amígdala y el hipocampo desde el núcleo basal de Meynert. En la enfermedad de Alzheimer ocurre pérdida grave de estas fibras. La amígdala está estrechamente asociada con el hipocampo, y se relaciona con la codificación de recuerdos cargados emocionalmente, y con el recuerdo de los mismos. Durante la recuperación de recuerdos aterradores, los ritmos theta de la amígdala y el hipocampo se sincronizan. En sujetos sanos, eventos asociados con emociones fuertes son recordados mejor que los eventos sin una carga emocional, pero en pacientes con lesiones bilaterales de la amígdala, falta esta diferencia. MEMORIA A LARGO PLAZO Mientras que el proceso de codificación para la memoria explícita a corto plazo involucra el hipocampo, los recuerdos a largo plazo son almacenados en diversas partes de la neocorteza. Diversas partes de los recuerdos —visuales, olfatorias, auditivas, etc.— están ubicadas en las regiones corticales relacionadas con estas funciones, y los fragmentos se enlazan mediante cambios a largo plazo de la fuerza de transmisión en uniones sinápticas importantes de modo que todos los componentes son llevados a la conciencia cuando se rememora el recuerdo. Una vez que se han establecido recuerdos a largo plazo, se pueden recordar o tener acceso a ellos por medio de muchas asociaciones diferentes, por ejemplo, el recuerdo de una escena vivida puede ser evocado no sólo por una escena similar, sino también por un sonido u olor asociado con la escena, así, cada recuerdo almacenado debe tener múltiples rutas, y muchos recuerdos tienen un componente emocional. ENFERMEDAD DE ALZHEIMER Y DEMENCIA SENIL La enfermedad de Alzheimer es el trastorno neurodegenerativo relacionado más comúnmente con la edad. La declinación de la memoria inicialmente se manifiesta como una pérdida de la memoria episódica, que obstaculiza el recuerdo de eventos recientes, esta pérdida de la memoria a corto plazo va seguida por pérdida general de funciones cognitivas y otras funciones cerebrales, la necesidad de cuidado constante, y finalmente, la muerte. Los datos citopatológicos característicos de la enfermedad son marañas neurofibrilares intracelulares, compuestas en parte de formas hiperfosforiladas de la proteína tau que normalmente se une a microtúbulos, y placas seniles extracelulares, que tienen un centro de péptidos β-amiloides (Aβ) rodeados por fibras nerviosas alteradas y células gliales reactivas. En la figura 21-5 se compara una célula nerviosa normal con una que muestra anormalidades asociadas con enfermedad de Alzheimer. Los péptidos Aβ son productos de una proteína normal, la proteína precursora de amiloide (APP), una proteína transmembrana que se proyecta hacia el líquido extracelular (ECF) desde todas las células nerviosas, esta proteína es hidrolizada en tres sitios diferentes por las secretasas α, β y γ, respectivamente. Cuando la APP es hidrolizada por secretasa α, se producen productos peptídicos no tóxicos, con todo, cuando es hidrolizada por las secretasas β y γ, se producen polipéptidos con 40 a 42 aminoácidos; la longitud real varía debido a variación del sitio en el cual la secretasa γ, corta la cadena de proteína. Estos polipéptidos son tóxicos; el más tóxico es Aβσ1-42. Los polipéptidos forman agregados extracelulares, que pueden pegarse a receptores α-amino-3-hidroxilo-5-metil-4-isoxazol-propionato (AMPA) y canales de ion Ca2+, lo que aumenta el flujo de entrada de Ca2+, Terminales nerviosas A Normal B Enfermedad de Alzheimer Hebras de neuropilo Marañas neurofibrilares Aβ (fibrilar) Filamentos helicoidales pareados Orgánulos membranosos anormales Neuritas Placa senil FIGURA 215 Comparación de una neurona normal A) con una que muestra anormalidades asociadas con enfermedad de Alzheimer B). (Reproducida con autorización de Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000.) CAPÍTULO 21 Aprendizaje, memoria, lenguaje y habla 195 también inician una respuesta inflamatoria, con producción de marañas intracelulares, y las células dañadas finalmente mueren. Un dato interesante que puede tener implicaciones fisiológicas amplias es que las actividades mentales laboriosas frecuentes, como hacer crucigramas difíciles y jugar juegos de mesa, retrasan el inicio de la demencia cognitiva debida a enfermedad de Alzheimer y enfermedad vascular. Aún se desconoce la explicación para este fenómeno de “úselo o piérdalo”, pero ciertamente sugiere que el hipocampo y sus conexiones tienen plasticidad como otras partes del cerebro. En pacientes con esquizofrenia, un trastorno que se caracteriza por sentido de realidad deformado, los estudios de MRI muestran volúmenes reducidos de sustancia gris en el lado izquierdo del hipocampo anterior, la amígdala y la circunvolución parahipocampal, y en la circunvolución temporal posterosuperior. El grado de reducción en la circunvolución temporal superior izquierda se correlaciona con el grado de pensamiento trastornado en la enfermedad, y también hay anormalidades manifiestas de los sistemas dopaminérgicos y el flujo sanguíneo cerebral en esta enfermedad. LENGUAJE Y HABLA FISIOLOGÍA DEL LENGUAJE La memoria y el aprendizaje son funciones de partes grandes del cerebro, pero los centros que controlan algunas de las otras “funciones superiores del sistema nervioso”, en particular los mecanismos relacionados con el lenguaje, están más o menos localizados en la neocorteza. Las funciones de lenguaje del ser humano dependen más de un hemisferio cerebral que del otro, esto se llama el hemisferio dominante, y está relacionado con categorización y simbolización. El otro hemisferio no está menos desarrollado o es “no dominante”; en lugar de eso, está especializado en el área de relaciones espaciotemporales, es este hemisferio el que está relacionado, por ejemplo, con la identificación de objetos por su forma, y desempeña un papel primario en el reconocimiento de caras. Esto contribuye al concepto de especialización complementaria de los hemisferios, una para procesos secuenciales-analíticos (el hemisferio categórico) y una para relaciones visuoespaciales (el hemisferio representacional). El hemisferio categórico está relacionado con funciones de lenguaje, las lesiones en dicho hemisferio producen trastornos del lenguaje. En contraste, las lesiones en el hemisferio representacional llevan a astereognosia, la incapacidad para identificar objetos al palparlos. La especialización hemisférica se relaciona con el uso preferente de una de las manos. En 96% de los individuos diestros, que constituyen 91% de la población humana, el hemisferio izquierdo es el hemisferio dominante o categórico, y en el otro 4% el hemisferio derecho es dominante. En 70% de los zurdos, el hemisferio izquierdo también es el hemisferio categórico; en 15%, el hemisferio derecho es el hemisferio categórico, y en 15% no hay una lateralización clara. Las discapacidades de aprendizaje, como la dislexia (una capacidad alterada para aprender a leer) son 12 veces más comunes en los zurdos que en los diestros, posiblemente porque alguna anormalidad fundamental en el hemisferio izquierdo llevó a un cambio del uso preferente de una de las manos en etapas tempranas del desarrollo. Los talentos espaciales de los zurdos están bastante por arriba del promedio puesto que un número desproporcionadamente grande de artistas, músicos y matemáticos es zurdo. Algunas diferencias anatómicas entre los dos hemisferios tal vez se correlacionan con las diferencias funcionales. El plano temporal, un área de la circunvolución temporal superior que está involucrada en el procesamiento auditivo relacionado con el lenguaje, regularmente es de mayor tamaño en el lado izquierdo que en el derecho. Estudios de imágenes muestran que otras porciones de la superficie superior del lóbulo temporal izquierdo son de mayor tamaño en individuos diestros, el lóbulo frontal normalmente es más grueso que el izquierdo, y el lóbulo occipital izquierdo es más amplio y sobresale a través de la línea media. Las áreas del cerebro primarias relacionadas con el lenguaje están ubicadas a lo largo de la cisura de Silvio (surco cerebral lateral) del hemisferio categórico, y cerca de la misma. Una región en el extremo posterior de la circunvolución temporal superior llamada área de Wernicke (figura 21-6) se relaciona con la comprensión de información auditiva y visual. Se proyecta por medio del fascículo arqueado al área de Broca (área 44) del lóbulo frontal inmediatamente enfrente del extremo inferior de la corteza motora. El área de Broca procesa la información recibida desde el área de Wernicke hacia un patrón detallado y coordinado para vocalización, y después proyecta el patrón por medio de un área de articulación del lenguaje en la ínsula a la corteza motora, que inicia los movimientos apropiados de los labios, la lengua y la laringe para producir habla. En la figura 21-7 se muestra la secuencia probable de eventos que ocurre cuando un sujeto dice en voz alta el nombre de un objeto visual. La circunvolución angular detrás del área de Wernicke procesa información proveniente de palabras que son leídas, de tal manera que pueden convertirse en formas auditivas de las palabras en el área de Wernicke. En individuos que aprenden un segundo idioma durante la adultez, la fMRI revela que la porción del área de Broca relacionada con él está adyacente al área relacionada con el idioma materno, pero separada de la misma. No obstante, en niños que aprenden dos idiomas al principio de la vida, sólo un área única está involucrada con ambos, por supuesto, se sabe bien que los niños llegan a dominar un segundo idioma con mayor facilidad que los adultos. Fascículo arqueado Área de Broca Circunvolución angular Área de Wernicke FIGURA 216 Ubicación de algunas de las áreas en el hemisferio categórico que están relacionadas con funciones de lenguaje. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) 196 SECCIÓN IV Fisiología del sistema nervioso central/neural Izquierdo Derecho Área de Broca tida al área de Wernicke, el resultado es una enfermedad llamada afasia anómica. RECONOCIMIENTO DE ROSTROS Área facial de la corteza motora (área 4) 6 Área de Wernicke (área 22) Fascículo arqueado 5 Desde el núcleo geniculado lateral 4 Circunvolución angular (área 39) Áreas corticales visuales de orden superior (área 18) 1 3 2 Una parte importante de las aferencias visuales va al lóbulo temporal inferior, donde se almacenan representaciones de objetos, en particular caras. Las caras tienen importancia particular para distinguir entre amigos y enemigos, y el estado emocional de quienes son vistos; el almacenamiento y el reconocimiento de caras están más fuertemente representados en el lóbulo temporal inferior derecho en individuos diestros, y las lesiones en esta área causan prosopagnosia, la incapacidad para reconocer caras. Los pacientes que tienen esta anormalidad pueden reconocer formas y reproducirlas, pueden reconocer a las personas por su voz, y muchos de ellos muestran respuestas del SNA cuando ven caras familiares en contraposición con no familiares, empero, no pueden identificar las caras que ven. Corteza visual primaria (área 17) FIGURA 217 Trayectoria tomada por impulsos cuando un sujeto nombra un objeto visual proyectado en una sección horizontal del cerebro humano. (Reproducida con autorización de Barrett KE, Barman SM, Boitano S, Brooks H: Ganong’s Review of Medical Physiology, 23rd ed. McGraw-Hill Medical, 2009.) TRASTORNOS DEL LENGUAJE Las afasias son anormalidades de las funciones del lenguaje que no se deben a defectos de la visión o la audición, ni a parálisis motora, se originan por lesiones en el hemisferio categórico, y la causa más común es la embolia o trombosis de un vaso sanguíneo cerebral. Una clasificación conveniente las divide en afasias fluente, no fluente y anómica. En una forma de afasia fluente, la lesión está en el área de Wernike, el habla en sí es normal, y a veces el paciente habla excesivamente; sin embargo, lo que dice está lleno de jerga y neologismos que tienen poco sentido, el paciente tampoco logra comprender el significado de palabras habladas o escritas, de modo que otros aspectos del uso de lenguaje están alterados. Otra forma de afasia fluente es un padecimiento en el cual los pacientes pueden hablar relativamente bien y tienen buena comprensión auditiva, pero no pueden colocar partes de palabras juntas o evocar palabras, parece deberse a lesiones en la corteza auditiva (áreas 40, 41 y 42 de Brodmann) y alrededor de la misma. En la afasia no fluente, la lesión está en el área de Broca. El habla es lenta, y a los pacientes les resulta difícil encontrar palabras. Los pacientes con daño grave de esta área están limitados a dos o tres palabras con las cuales expresan toda la gama de significado y emoción, a veces las palabras retenidas son las que se estuvieron hablando en el momento de la lesión o de la enfermedad vascular aguda que causó la afasia. Cuando una lesión daña la circunvolución angular en el hemisferio categórico sin afectar el área de Wernicke o de Broca, no hay dificultad con el habla o con el entendimiento de información auditiva; en lugar de eso, hay problema para comprender lenguaje escrito o imágenes, porque la información visual no es procesada y transmi- CORRELACIÓN CLÍNICA Un niño de nueve años de edad empezó a tener crisis convulsivas epilépticas después de un accidente en bicicleta. Un electroencefalograma (EEG) mostró que las crisis se originaron en ambos lóbulos temporales, tuvo muchas crisis parciales y varias crisis tónico-clónicas hacia los 16 años de edad, la aparición frecuente de crisis convulsivas y las lagunas de conocimiento hicieron muy difícil que completara la enseñanza secundaria, a pesar de sus habilidades intelectuales, también fue incapaz de conservar un empleo como un obrero de montaje. A los 27 años de edad, se le practicó una intervención quirúrgica experimental en el cerebro para extirpar la amígdala, porciones grandes de la formación hipocampal, y porciones del área de asociación de la corteza temporal. Las crisis convulsivas se controlaron mejor después de la operación, y se redujeron a sólo alrededor de una crisis convulsiva importante al año, con todo, la operación llevó a déficits de memoria devastadores. Ha mantenido memoria a largo plazo para eventos que ocurrieron antes de la operación, pero sufre amnesia anterógrada, y su memoria a corto plazo está intacta, pero es incapaz de llevar eventos nuevos a la memoria a largo plazo. Tiene memoria procedural normal, y puede aprender nuevos acertijos y habilidades motoras, este es un paciente real en quien se practicó la intervención quirúrgica experimental en 1953. Este caso ha sido estudiado por muchos científicos, y ha llevado a un mayor entendimiento del enlace entre el lóbulo temporal y la memoria declarativa; su caso es el primero que llamó la atención respecto al papel crucial de los lóbulos temporales en la formación de recuerdos a largo plazo, y que implicó esta región en la conversión de recuerdos a corto plazo en recuerdos a largo plazo. Investigación posterior mostró que el hipocampo es la estructura primaria dentro del lóbulo temporal involucrada en esta conversión, dado que retuvo recuerdos de antes de la intervención quirúrgica, este caso también muestra que el hipocampo no está involucrado en el almacenamiento de memoria declarativa. Una fascinante grabación de audio por la National Public Radio del decenio de 1990-1999 CAPÍTULO 21 Aprendizaje, memoria, lenguaje y habla del paciente hablando a científicos fue publicada en 2007 y está disponible en el sitio http://www.npr.org/templates/story/story. php?storyId=7584970. RESUMEN DEL CAPÍTULO ■ ■ ■ ■ ■ ■ La memoria a largo plazo se divide en explícita (declarativa) e implícita (no declarativa). La explícita se subdivide en semántica y episódica. La implícita se subdivide en imprimación, procedural, aprendizaje asociativo y aprendizaje no asociativo. La plasticidad sináptica es la capacidad del tejido neural para cambiar según se refleja por LTP (una efectividad aumentada de la actividad sináptica) o LTD (una efectividad reducida de la actividad sináptica) después de uso continuo. Las estructuras hipocampales y otras estructuras del lóbulo temporal y la corteza de asociación están involucradas en la memoria declarativa. La enfermedad de Alzheimer se caracteriza por pérdida progresiva de la memoria a corto plazo, seguida por pérdida general de la función cognitiva. Los datos citopatológicos característicos de la enfermedad de Alzheimer son marañas neurofibrilares intracelulares y placas seniles extracelulares. Los hemisferios categórico y representacional son para procesos secuenciales-analíticos y relaciones visuoespaciales, respectivamente. Las lesiones en el hemisferio categórico producen trastornos del lenguaje, mientras que las lesiones en el hemisferio representacional producen astereognosia. Las afasias son anormalidades de las funciones del lenguaje, y se originan por lesiones en el hemisferio categórico. Se clasifican como fluente (área de Wernicke; corteza auditiva), no fluente (área de Broca), y anómica (circunvolución angular) con base en la ubicación de las lesiones cerebrales. PREGUNTAS DE ESTUDIO 1. El hemisferio representacional: A) es el hemisferio cerebral derecho en la mayoría de los individuos diestros. B) es el hemisferio cerebral izquierdo en la mayoría de los individuos zurdos. C) incluye parte del cerebro relacionada con funciones del lenguaje. D) es el sitio de lesiones en la mayoría de los pacientes con afasia. E) es morfológicamente idéntico al hemisferio no representacional opuesto. 197 2. Los efectos de la pérdida bilateral de la función hipocampal son: A) desaparición de recuerdos remotos. B) pérdida de la memoria de trabajo. C) pérdida de la capacidad para codificar eventos del pasado reciente en la memoria a largo plazo. D) pérdida de la capacidad para recordar caras y formas, pero no de la capacidad para recordar palabras impresas o habladas. E) producción de respuestas emocionales inapropiadas cuando se recuerdan fenómenos del pasado reciente. 3. ¿Cuáles de las que siguen están pareadas de manera incorrecta? A) lesión del lóbulo parietal del hemisferio representacional:inatención y abandono unilateral B) pérdida de neuronas colinérgicas en el núcleo basal de Meynert y áreas relacionadas del prosencéfalo:pérdida de la memoria reciente C) lesiones de los cuerpos mamilares: pérdida de la memoria reciente D) lesión de la circunvolución angular en el hemisferio categórico:afasia no fluente E) lesión del área de Broca en el hemisferio categórico:habla lenta 4. El hemisferio representacional es mejor que el hemisferio categórico en: A) funciones de lenguaje. B) reconocimiento de objetos por su forma. C) entendimiento de palabras impresas. D) entendimiento de palabras habladas. E) cálculos matemáticos. 5. Una lesión del área de Wernicke (el extremo posterior de la circunvolución temporal superior) en el hemisferio categórico hace que los pacientes: A) pierdan la memoria a corto plazo. B) hablen con una voz lenta y con pausas. C) experimenten déjà vu. D) hablen rápidamente pero con poco sentido. E) pierdan la capacidad para reconocer caras. 6. ¿Cuál de los que siguen más probablemente no está involucrado en la producción de LTP? A) cAMP B) Ca2+ C) receptores NMDA D) hiperpolarización de membrana E) despolarización de membrana SECCIÓN V FISIOLOGÍA CARDIOVASCULAR Perspectiva general del sistema cardiovascular Lois Jane Heller y David E. Mohrman O B J E T I V O S ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ Definir el papel del sistema cardiovascular en la homeostasis general del organismo. Identificar los principales compartimientos de líquido corporal y decir el volumen aproximado de cada uno. Hacer un diagrama de las vías de flujo de sangre entre el corazón y otros órganos corporales importantes. Declarar la relación entre el flujo sanguíneo, la presión arterial y la resistencia vascular. Predecir los cambios relativos de flujo a través de un tubo causados por cambios de la longitud del tubo, el radio del tubo, la viscosidad del líquido y la diferencia de presión. Identificar las cámaras y válvulas del corazón, y describir la vía de flujo a través del corazón. Definir el gasto cardiaco. Describir la vía de la propagación del potencial de acción en el corazón. Listar cinco factores esenciales para la acción de bombeo ventricular apropiada. Enunciar la relación entre el llenado ventricular y el gasto cardiaco (ley de Starling del corazón) y describir su importancia en el control del gasto cardiaco. Identificar la distribución de nervios simpáticos y parasimpáticos en el corazón, y listar los efectos básicos de estos nervios sobre la función cardiaca. Listar los principales tipos de vasos en un lecho vascular y describir las diferencias morfológicas entre ellos. Describir las características anatómicas y funciones básicas de los diferentes tipos de vasos. Identificar los principales mecanismos en el control vascular y la distribución del flujo sanguíneo. Describir la composición básica del líquido y las porciones celulares de la sangre, y listar los eventos asociados con la coagulación de la sangre. 199 C A P Í T U L O 22 200 SECCIÓN V Fisiología cardiovascular FUNCIÓN HOMEOSTÁTICA DEL SISTEMA CARDIOVASCULAR Tres componentes de los fluidos acuosos, conocidos en conjunto como el agua corporal total, explican alrededor de 60% del peso corporal, esta agua está distribuida entre los compartimientos intracelular, intersticial y plasma (figura 22-1); note que alrededor de dos terceras partes del agua corporal está contenida dentro de células y se comunica con el líquido intersticial a través de las membranas plasmáticas de células. Del líquido que está afuera de las células (es decir, el líquido extracelular), sólo una pequeña cantidad, el volumen plasmático, circula dentro del sistema cardiovascular. La sangre está compuesta de plasma y aproximadamente un volumen igual de elementos formes (principalmente eritrocitos). El líquido plasmático circulante se comunica con el líquido intersticial a través de las paredes de vasos capilares de pequeño calibre dentro de los órganos. El líquido intersticial es el ambiente inmediato de células individuales, las cuales deben extraer sus nutrientes del líquido intersticial y liberan sus productos hacia este último, sin embargo, el líquido intersticial no puede considerarse un reservorio grande para nutrientes o un sumidero grande para productos metabólicos porque su volumen es de menos de la mitad que el de las células a las que sirve. En consecuencia, el bienestar de células individuales depende mucho de los mecanismos homeostáticos que regulan la composición del líquido intersticial, esta tarea se logra mediante exposición continua del líquido intersticial a líquido plasmático circulante “fresco”. Conforme la sangre pasa a través de capilares, se intercambian solutos entre el plasma y el líquido intersticial mediante el proceso de difusión. El resultado neto de la difusión transcapilar siempre es que el líquido intersticial tiende a adoptar la composición de la sangre que llega, por ejemplo, si la concentración de ion potasio en el intersticio de un músculo esquelético particular fuera más alta que la que hay en el plasma que está entrando al músculo, el potasio se difundiría hacia la sangre conforme pasara por los capilares del músculo, mas dado que esto elimina potasio del líquido intersticial, la concentración de ion potasio intersticial disminuiría, y dejaría de disminuir cuando ya no ocurriera movimiento neto de potasio hacia capilares, es decir, cuando la concentración intersticial alcanzara la del plasma que está llegando. Tres condiciones son esenciales para que este mecanismo circulatorio controle con eficacia la composición del líquido intersticial: 1) debe haber control adecuado del flujo sanguíneo a través de los capilares tisulares, 2) la composición química de la sangre que está llegando (o arterial) debe controlarse para que sea la que es óptima en el líquido intersticial y 3) las distancias de difusión deben ser PULMONES HEMICARDIO DERECHO HEMICARDIO IZQUIERDO ÓRGANOS CORPORALES CAPILARES Células Compartimiento de plasma circulante ≈3 L Compartimiento intersticial (ambiente interno) ≈12 L Compartimiento intracelular ≈30 L FIGURA 221 Principales compartimientos de líquido corporal, con volúmenes promedio indicados para un ser humano de 70 kg. El agua corporal total es de alrededor de 60% del peso corporal. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) CAPÍTULO 22 Perspectiva general del sistema cardiovascular 100% 201 Pulmones Bomba cardiaca derecha Bomba cardiaca izquierda 100% 100% Músculo cardiaco Cerebro Músculo esquelético Hueso 3% 14% 15% 5% VENAS ARTERIAS Sistema gastrointestinal, bazo Hígado Riñón Piel Otro 21% 6% 22% 6% 8% FIGURA 222 Circuitos cardiovasculares que indican la distribución porcentual del gasto cardiaco hacia diversos sistemas en un individuo en reposo. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) cortas. En la figura 22-1 se muestra cómo el sistema de transporte cardiovascular opera para lograr estas tareas. Como se comentó, las sustancias son transportadas entre las células en el plasma en vasos capilares dentro de órganos mediante el proceso de difusión. Este transporte ocurre en distancias en extremo pequeñas porque casi ninguna célula en el organismo está ubicada a más de alrededor de 10 μm de un capilar, en esas distancias microscópicas, la difusión es un proceso muy rápido; empero, es un mecanismo muy inadecuado mediante el cual mover sustancias desde los capilares de un órgano, como los pulmones, hacia los capilares de otro órgano que puede estar a 1 m o más de distancia. Por consiguiente, las sustancias son transportadas entre órganos mediante el proceso de convección, por medio del cual las sustancias se mueven junto con el flujo sanguíneo simplemente porque están disueltas o por lo demás contenidas dentro de la sangre. Las distancias relativas involucradas en el transporte cardiovascular no se ilustran bien en la figura 22-1; si la figura se dibujara a escala, de modo que 2.5 cm (1 pulgada) representara la distancia desde los capilares hasta las células dentro de un músculo de la pantorrilla, los capilares en los pulmones tendrían que estar ubicados ¡a alrededor de 2.4 kilómetros (1.5 millas) de distancia! En la figura 22-2 se ilustra la disposición funcional general del sistema cardiovascular, puesto que en esta figura se expresa un punto de vista funcional más que anatómico, el corazón aparece en tres lugares: como la bomba cardiaca derecha, como la bomba cardiaca izquierda, y como el tejido de músculo cardiaco. Es una práctica común considerar al sistema cardiovascular como: 1) la circulación pulmonar, compuesta de la bomba cardiaca derecha y los pulmones y 2) la circulación sistémica, en la cual la bomba cardiaca izquierda proporciona sangre a los órganos sistémicos (todas las estructuras excepto la porción de intercambio de gases de los pulmones. Las circulaciones pulmonar y sistémica están dispuestas en serie, es decir, una después de la otra, en consecuencia, los hemicardios derecho e izquierdo deben bombear, cada uno, el mismo volumen de sangre cada minuto, esta cantidad se llama el gasto cardiaco; de manera que un gasto cardiaco de 5 a 6 l/min es normal para un individuo en reposo. Los órganos sistémicos en general están dispuestos en paralelo (esto es, lado a lado) dentro del sistema cardiovascular (figura 22-2), esta disposición paralela tiene dos consecuencias importantes; en primer lugar, casi todos los órganos sistémicos reciben sangre de composición idéntica —aquella que acaba de salir de los pulmones y se conoce como sangre arterial—; en segundo lugar, el flujo a través de cualesquiera de los órganos sistémicos puede controlarse de manera independiente del flujo a través de los otros órganos, así, por ejemplo, la respuesta cardiovascular al ejercicio de todo el cuerpo 202 SECCIÓN V Fisiología cardiovascular puede comprender aumento del flujo sanguíneo a través de algunos órganos, pero flujo sanguíneo disminuido o sin cambios a través de otros. Muchos de los órganos en el cuerpo ayudan a desempeñar la tarea de reacondicionar de manera continua la sangre circulante en el sistema cardiovascular, los órganos que se comunican con el ambiente externo, como los pulmones, desempeñan papeles clave, como es evidente en la disposición que se muestra en la figura 22-2, la sangre que ha pasado a través de un órgano sistémico regresa al hemicardio derecho y es bombeada a través de los pulmones, donde se intercambian oxígeno y dióxido de carbono, así, la composición de gases de la sangre siempre se reacondiciona inmediatamente después de salir de un órgano sistémico. Al igual que los pulmones, muchos de los órganos sistémicos también sirven para reacondicionar la composición de la sangre, aunque los circuitos de flujo impiden que lo hagan cada vez que la sangre completa un circuito, por ejemplo, los riñones ajustan la composición de electrólitos de la sangre que está pasando por ellos. Dado que la sangre acondicionada por los riñones, se mezcla libremente con toda la sangre circulante, y puesto que los electrólitos y el agua pasan libremente a través de casi todas las paredes capilares, los riñones controlan el equilibrio de electrólitos de todo el ambiente interno; para lograr esto, es necesario que una unidad dada de sangre pase a menudo a través de los riñones, de hecho, los riñones (en condiciones de reposo) normalmente reciben alrededor de una quinta parte del gasto cardiaco. Esto excede mucho la cantidad de flujo que se necesita para satisfacer las necesidades de nutrientes del tejido renal, esta situación es común a órganos que tienen una función de acondicionamiento de la sangre. Los órganos que acondicionan la sangre también pueden soportar, al menos temporalmente, reducciones graves del flujo sanguíneo, por ejemplo, la piel puede tolerar fácilmente una reducción del flujo sanguíneo cuando es necesario para conservar el calor corporal (capítulo 70). Casi todos los órganos abdominales grandes también caen dentro de esta categoría, la razón es simplemente que debido a sus funciones de acondicionamiento de la sangre, su flujo sanguíneo normal es mucho mayor que el que se requiere para mantener sus necesidades metabólicas basales. El cerebro, el músculo cardiaco y los músculos esqueléticos tipifican órganos en los cuales la sangre fluye principalmente para satisfacer las necesidades metabólicas del tejido. No reacondicionan la sangre para el beneficio de cualquier otro órgano. El flujo hacia el cerebro y el músculo cardiaco normalmente sólo es un poco mayor que el que se requiere para su metabolismo, y no toleran bien las interrupciones del flujo sanguíneo; puede ocurrir pérdida del conocimiento en el transcurso de algunos segundos después de suspensión del flujo cerebral, y puede sobrevenir daño cerebral permanente en apenas 4 min sin flujo. De modo similar, el músculo cardiaco (miocardio) normalmente consume aproximadamente 75% del oxígeno que se le aporta, y la capacidad de bombeo del corazón empieza a deteriorarse en cuestión de latidos de una interrupción del flujo coronario. La tarea de proporcionar flujo sanguíneo adecuado al cerebro y el músculo cardiaco recibe una prioridad alta en la operación general del sistema cardiovascular (véase más adelante). LA FÍSICA BÁSICA DEL FLUJO SANGUÍNEO Como se esbozó, la tarea de mantener la homeostasis intersticial requiere que una cantidad suficiente de sangre fluya de manera continua a través de los millones de capilares en el organismo, en un individuo en reposo, esto asciende a un gasto cardiaco de alrededor de 5 l/min (aproximadamente 302.8 L [80 galones]/hora). Durante la vida cotidiana de las personas, los índices metabólicos y, por ende, los requerimientos de flujo sanguíneo en diferentes órganos y regiones en todo el cuerpo cambian de un momento a otro. Así, el sistema cardiovascular debe ajustar de manera continua tanto la magnitud del gasto cardiaco como la manera en que este último es distribuido hacia diferentes partes del cuerpo. Una clave de las más importantes para comprender cómo opera el sistema cardiovascular es por medio de entendimiento de la relación entre los factores físicos que determinan la tasa de flujo de líquido a través de un tubo. El tubo que se muestra en la figura 22-3 podría representar un segmento de cualquier vaso sanguíneo en el cuerpo, tiene una cierta longitud (L) y un cierto radio interno (r) a través del cual fluye sangre. El líquido fluye a través del tubo sólo cuando las presiones en el líquido en los extremos de entrada y de salida (Pi y Po) son desiguales, es decir, cuando hay una diferencia de presión (ΔP) entre los extremos, ya que, proporcionan la fuerza impulsora para el flujo, y puesto que aparece fricción entre el líquido en movimiento y las paredes estacionarias de un tubo, los vasos tienden a ofrecer resistencia al movimiento de líquido a través de ellos. Esta resistencia vascular es una medida de la dificultad para hacer fluir líquido a través del tubo, es decir, qué tanta diferencia de presión se requiere para causar un cierto flujo. La relación importante entre flujo, dife- Longitud (L) Radio (r ) Pi Po · Flujo (Q ) Presión a la entrada ΔP = Pi < Po FIGURA 223 Factores que influyen sobre el flujo de líquido a través de un tubo. Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) Presión a la salida (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular CAPÍTULO 22 Perspectiva general del sistema cardiovascular rencia de presión y resistencia se describe mediante la ecuación de flujo básica como sigue: Diferencia de presión Flujo = _________________ Resistencia (1) ΔP Q· = ___ R · donde Q es el flujo (volumen/tiempo), ΔP es la diferencia de presión (mmHg), y R la resistencia al flujo (mmHg × tiempo/volumen). La ecuación de flujo básica puede aplicarse no sólo a un tubo único, sino también a redes complejas de tubos, por ejemplo, al lecho vascular de un órgano o toda la circulación sistémica; de manera que el flujo sanguíneo a través del cerebro está determinado por la diferencia de presión entre las arterias y las venas cerebrales, dividida entre la resistencia general al flujo a través de los vasos en el lecho vascular cerebral. A partir de la ecuación de flujo básica debe ser evidente que sólo hay dos maneras en las cuales puede cambiarse el flujo de sangre a través de cualquier órgano: 1) al cambiar la diferencia de presión a través de su lecho vascular o 2) al cambiar su resistencia vascular. Más a menudo, es el cambio de la resistencia vascular de un órgano lo que hace que el flujo a través del órgano cambie. La resistencia al flujo a través de un tubo cilíndrico depende de varios factores, entre ellos el radio del tubo y su longitud, y la viscosidad del líquido que está fluyendo por él. Estos factores influyen sobre la resistencia al flujo como sigue: 8 Lη πr R = ____ 4 (2) donde r es el radio interno del tubo, L es la longitud del tubo, y η es la viscosidad del líquido. Note que la resistencia es inversamente proporcional al radio interno del tubo a la cuarta potencia. Así, incluso cambios pequeños del radio interno de un tubo tienen una influencia muy grande sobre su resistencia al flujo, por ejemplo, disminuir a la mitad, el radio interno de un tubo aumentará 16 veces su resistencia al flujo. Las ecuaciones (1) y (2) pueden combinarse hacia una expresión conocida como la ecuación de Poiseuille (ecuación [3]), que incluye todos los términos que influyen sobre el flujo a través de un vaso cilíndrico: · π r4 Q = ΔP ____ 8 Lη (3) De nuevo, advierta que sólo ocurre flujo cuando hay una diferencia de presión, entonces, no sorprende que la presión arterial sea una variable cardiovascular de importancia extrema y regulada con sumo cuidado, también nótese de nuevo que, para cualquier diferencia de presión dada, el radio del tubo tiene una influencia muy grande sobre el flujo a través de un tubo. Por consiguiente, es lógico que los flujos de sangre por órganos estén regulados principalmente mediante cambios del radio de vasos sanguíneos dentro de órganos. Mientras que la longitud del vaso y la viscosidad de la sangre son factores que influyen sobre la resistencia vascular, no son variables que se puedan manipular fácilmente para el propósito de control del flujo sanguíneo momento a momento. En lo que se refiere al sistema cardiovascular general (figuras 22-1 y 22-2), es posible concluir que la sangre fluye a través de los vasos dentro de un órgano sólo porque hay una diferencia de presión entre la sangre en las arterias que riegan el órgano y las venas que lo drenan. El trabajo primario de la bomba cardiaca es mantener la presión 203 dentro de las arterias más alta que la que hay dentro de las venas. Normalmente, la presión promedio en arterias sistémicas es de alrededor de 100 mmHg, y en venas sistémicas se aproxima a 0 mmHg. Por tanto, puesto que la diferencia de presión (ΔP) es idéntica a través de todos los órganos sistémicos, el gasto cardiaco está distribuido entre los diversos órganos sistémicos únicamente con base en sus resistencias individuales al flujo, dado que la sangre fluye a lo largo de la vía de menos resistencia, los órganos con resistencia relativamente baja reciben flujo relativamente alto. CORAZÓN ACCIÓN DE BOMBEO El corazón yace en el centro de la cavidad torácica suspendido por sus fijaciones a los grandes vasos dentro de un saco fibroso delgado llamado el pericardio. Una pequeña cantidad de líquido en el saco lubrica la superficie del corazón y permite que se mueva libremente durante la contracción y la relajación. El flujo sanguíneo a través de todos los órganos es pasivo y sólo ocurre porque la acción de bombeo del corazón mantiene más alta la presión arterial que la presión venosa. La bomba cardiaca derecha proporciona la energía necesaria para mover sangre a través de los vasos pulmonares, y la bomba cardiaca izquierda proporciona la energía para mover sangre a través de los órganos sistémicos. La cantidad de sangre bombeada por minuto desde cada ventrículo (el gasto cardiaco, CO) depende del volumen de sangre eyectada por cada latido (el volumen sistólico, SV) y el número de latidos cardiacos por minuto (la frecuencia cardiaca, HR) como sigue: CO[volumen/min] = SV[volumen/latido] × HR[latidos/min] (4) A partir de la relación anterior debe ser evidente que todas las influencias sobre el gasto cardiaco deben actuar al cambiar la frecuencia cardiaca o el volumen sistólico. Estas influencias se describirán con detalle en capítulos subsiguientes. En la figura 22-4 se indica la vía de flujo sanguíneo a través de las cámaras del corazón. La sangre venosa regresa desde los órganos sistémicos hacia la aurícula derecha por medio de las venas cavas superior e inferior, pasa a través de la válvula tricúspide hacia el ventrículo derecho, y desde ahí es bombeada a través de la válvula pulmonar hacia la circulación pulmonar por medio de las arterias pulmonares. La sangre venosa pulmonar oxigenada fluye en las venas pulmonares hacia la aurícula izquierda, y pasa a través de la válvula mitral hacia el ventrículo izquierdo, desde ahí es bombeada a través de la válvula aórtica hacia la aorta para ser distribuida hacia los órganos sistémicos. Aunque las características de anatomía macroscópica de la bomba cardiaca derecha difieren un poco de las de la bomba cardiaca izquierda, los principios de bombeo son idénticos, cada bomba consta de un ventrículo, que es una cámara cerrada rodeada por una pared muscular (figura 22-5). Las válvulas están diseñadas estructuralmente para permitir el flujo en sólo una dirección, y se abren y cierran de manera pasiva en respuesta a la dirección de las diferencias de presión a través de ellas. La acción de bombeo ventricular ocurre porque el volumen de la cámara intraventricular cambia cíclicamente por contracción y relajación rítmicas y sincronizadas 204 SECCIÓN V Fisiología cardiovascular Vena cava superior hace que la presión dentro de la cámara aumente; tan pronto como la presión ventricular excede la presión en la arteria pulmonar (bomba derecha) o en la aorta (bomba izquierda), se fuerza la sangre hacia afuera de la cámara a través de la válvula de salida (pulmonar o aórtica) (figura 22-5). Esta fase del ciclo cardiaco durante la cual las células de músculo ventricular se están contrayendo se llama sístole. Dado que la presión es más alta en el ventrículo que en la aurícula durante la sístole, la válvula de entrada o auriculoventricular (AV) (tricúspide o mitral) se cierra, cuando las células de músculo ventricular se relajan, la presión en el ventrículo disminuye por debajo de la que hay en la aurícula, la válvula AV se abre, y el ventrículo se vuelve a llenar con sangre (figura 22-5, derecha), esta porción del ciclo cardiaco se llama diástole. La válvula de salida se cierra durante la diástole porque la presión arterial es mayor que la presión intraventricular. Después del periodo de llenado diastólico, se inicia la fase sistólica de un nuevo ciclo cardiaco. Aorta Arteria pulmonar Aurícula derecha Venas pulmonares Válvula pulmonar Aurícula izquierda Válvula aórtica Válvula mitral Válvula tricúspide Ventrículo izquierdo Vena cava inferior EXCITACIÓN La acción de bombeo eficiente del corazón requiere una coordinación precisa de la contracción de millones de células de músculo cardiaco individuales y la contracción de cada célula es desencadenada, cuando un impulso excitatorio eléctrico (potencial de acción) avanza sobre su membrana. La coordinación apropiada de la actividad contráctil de las células de músculo cardiaco individuales se logra principalmente por medio de la conducción de potenciales de acción desde una célula hacia la siguiente por medio de uniones intercelulares comunicantes (conexiones comunicantes) que conectan todas las células del corazón en un sincitio funcional (es decir, que actúan como una unidad sincrónica), además, las células musculares en ciertas áreas del corazón están adaptadas de manera especí- Ventrículo derecho FIGURA 224 Trayectoria del flujo de sangre a través del corazón. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) de las células de músculo cardiaco individuales que yacen en una orientación circunferencial dentro de la pared ventricular. Cuando las células del músculo ventricular se están contrayendo, generan una tensión circunferencial en las paredes ventriculares que SÍSTOLE VENTRICULAR Aurícula DIÁSTOLE VENTRICULAR Válvula de salida Válvula de entrada Pared ventricular FIGURA 225 Acción de bombeo ventricular. Books/McGraw-Hill, 2006.) Cámara intraventricular (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical CAPÍTULO 22 Perspectiva general del sistema cardiovascular 205 4. Las contracciones musculares deben ser enérgicas (no insuficientes). 5. Los ventrículos deben llenarse de manera adecuada durante la diástole. Nodo sinoauricular Nodo auriculoventricular Músculo auricular Cartílago Haz de His Rama izquierda del fascículo Rama derecha del fascículo Músculo ventricular FIGURA 226 Sistema de conducción eléctrica del corazón. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th En los capítulos subsiguientes, se estudiará con detalle cómo los requerimientos antes mencionados se satisfacen en el corazón normal. CONTROL DEL CORAZÓN Y DEL GASTO CARDIACO Llenado diastólico A medida que el llenado cardiaco aumenta durante la diástole, el volumen eyectado durante la sístole también se incrementa, como consecuencia (figura 22-7), con otros factores iguales, el volumen sistólico aumenta conforme se incrementa el volumen cardiaco al final de la diástole. Este fenómeno (ley de Starling del corazón) es una propiedad intrínseca del músculo cardiaco, y es uno de los reguladores primarios del gasto cardiaco. Los mecanismos que originan este fenómeno dependen en su mayor parte de la relación longitudtensión de las células de músculo cardiaco, y se describirán con detalle en capítulos subsiguientes. Influencias nerviosas del sistema nervioso autónomo fica para controlar la frecuencia de la excitación cardiaca, la vía de conducción, y la tasa de la propagación de impulso a través de diversas regiones del corazón. En la figura 22-6 se muestran los principales componentes de este sistema de excitación y conducción especializado. Incluyen el nodo sinoauricular (nodo SA), el nodo auriculoventricular (AV), el haz de His, y sus ramas derecha e izquierda constituidas por células especializadas llamadas fibras de Purkinje. El nodo SA contiene células especializadas que en circunstancias normales funcionan como el marcapaso del corazón e inician el potencial de acción que es conducido a través del corazón. El nodo AV contiene células de conducción lenta que normalmente funcionan para crear un pequeño retraso entre la contracción auricular y la contracción ventricular. Las fibras de Purkinje están especializadas para la conducción rápida, y aseguran que todas las células ventriculares se contraigan casi en el mismo instante. Si bien el corazón puede latir de manera inherente por sí solo, la función cardiaca puede estar influida profundamente por aferencias nerviosas que provienen de las divisiones tanto simpática como parasimpática del sistema nervioso autónomo (SNA), estas aferencias permiten modificar el bombeo del corazón según sea apropiado para satisfacer las necesidades homeostáticas cambiantes del organismo. Todas las porciones del corazón están ricamente inervadas por fibras simpáticas adrenérgicas. Cuando están activos, estos nervios simpáticos liberan norepinefrina sobre células cardiacas, la cual inte- REQUERIMIENTOS PARA LA OPERACIÓN EFICAZ Para que haya acción de bombeo ventricular efectiva y eficiente, el corazón debe estar funcionando de manera apropiada en cinco aspectos básicos: 1. Las contracciones de células de músculo cardiaco individuales deben ocurrir a intervalos regulares y estar sincronizadas (no ser arrítmicas). 2. Las válvulas deben abrirse por completo (no presentar estenosis). 3. Las válvulas no deben tener escape (no deben ser insuficientes ni presentar regurgitación). Volumen sistólico ed. New York: Lange Medical Books/McGraw-Hill, 2006.) Volumen ventricular al final de la diástole FIGURA 227 Ley de Starling del corazón. (Reproducida con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) 206 SECCIÓN V Fisiología cardiovascular ractúa con receptores β1-adrenérgicos sobre células de músculo cardiaco para aumentar la frecuencia cardiaca, la velocidad de conducción del potencial de acción, la fuerza de contracción y las frecuencias de contracción y relajación, es decir que, en general, la activación simpática actúa para aumentar el bombeo cardiaco. Fibras nerviosas parasimpáticas colinérgicas viajan al corazón por medio del nervio vago e inervan el nodo SA, el nodo AV y el músculo auricular. Cuando están activos, estos nervios parasimpáticos liberan acetilcolina sobre células de músculo cardiaco, la cual interactúa con receptores muscarínicos sobre células de músculo cardiaco para disminuir la frecuencia cardiaca (nodo SA) y la velocidad de conducción del potencial de acción (nodo AV). Los nervios parasimpáticos también pueden actuar para reducir la fuerza de la contracción de las células musculares auriculares (no ventriculares), de manera que, en general, la activación parasimpática actúa para disminuir el bombeo cardiaco. Por lo general un incremento de la actividad nerviosa parasimpática se acompaña de una reducción de la actividad nerviosa simpática, y viceversa. VASCULATURA La sangre que es eyectada hacia la aorta por el hemicardio izquierdo pasa de manera consecutiva a través de muchos tipos diferentes de vasos antes de que regrese al hemicardio derecho. Las principales ARTERIAS ARTERIOLAS clasificaciones de vasos son arterias, arteriolas, capilares, vénulas y venas (figura 22-8). Estos segmentos vasculares consecutivos se distinguen uno de otro por diferencias en dimensiones físicas, características morfológicas y función. Algo que los vasos sanguíneos tienen en común es que están revestidos con una capa continua de células endoteliales, de hecho, esto es cierto para todo el sistema circulatorio, incluso las cámaras cardiacas y aun las hojuelas valvulares. En la figura 22-8 se muestran algunas características físicas representativas para cada uno de los principales tipos de vasos, no obstante, es necesario percatarse de que el lecho vascular es un continuo, y que la transición desde un tipo de segmento vascular hacia otro no es súbita. El área de corte transversal total a través del cual fluye sangre a cualquier nivel particular en el sistema vascular es igual a la suma de las áreas de corte transversal de todos los vasos individuales dispuestos en paralelo a ese nivel. El número y los valores de área de corte transversal totales presentados en la figura 22-8 son estimados para toda la circulación sistémica. Las arterias son vasos de pared gruesa que contienen, además de algo de músculo liso, un componente grande de fibras de elastina y colágeno. Es principalmente debido a las fibras de elastina, que pueden estirarse hasta dos veces su longitud sin carga, las arterias se pueden expandir para aceptar y almacenar temporalmente algo de la sangre eyectada por el corazón durante la sístole y, después, mediante retroceso pasivo, proporcionar esta sangre a los órganos torrente CAPILARES VÉNULAS VENAS Válvulas unidireccionales Aorta Venas cava Diámetro interno 2.5 cm 0.4 cm 30 μm 5 μm 70 μm 0.5 cm 3 cm Grosor de la pared 2 mm 1 mm 20 μm 1 μm 7 μm 0.5 mm 1.5 mm Número 1 160 5 × 107 1010 108 200 2 400 cm2 4500 cm2 4000 cm2 40 cm2 18 cm2 Área de corte 4.5 cm2 20 cm2 transversal total FIGURA 228 Características estructurales del sistema vascular periférico. Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular CAPÍTULO 22 Perspectiva general del sistema cardiovascular abajo durante la diástole. La aorta es la arteria más grande y tiene un diámetro interno (luminal) de alrededor de 25 mm. El diámetro arterial disminuye con cada ramificación consecutiva, y las arterias de menor calibre tienen diámetro de aproximadamente 0.1 mm. El patrón de ramificación arterial consecutivo causa un aumento exponencial del número de arterias, así, mientras el calibre de vasos individuales se hace progresivamente menor, el área de corte transversal disponible para el flujo sanguíneo dentro del sistema arterial aumenta a varias veces la que hay en la aorta. Las arterias a menudo se denominan vasos conducto porque tienen resistencia relativamente baja y no cambiante al flujo. Las arteriolas son de menor calibre y están estructuradas de modo diferente que las arterias; en proporción al tamaño de la luz, las arteriolas tienen paredes mucho más gruesas, con más músculo liso y menos material elástico que las arterias. Dado que las arteriolas son tan musculares, su diámetro puede cambiar de manera activa para regular el flujo sanguíneo a través de órganos periféricos, y a pesar de su tamaño diminuto, las arteriolas son tan numerosas que en paralelo su área de corte transversal colectiva es de tamaño mucho mayor que el que hay a cualquier nivel en las arterias. Las arteriolas a menudo se denominan vasos de resistencia debido a su resistencia alta y cambiable, que regula el flujo sanguíneo periférico a través de órganos individuales. Los capilares son los vasos de menor calibre en la vasculatura, de hecho, los eritrocitos con diámetro de alrededor de 7 μm deben deformarse para pasar a través de ellos. La pared capilar consta de una capa única de células endoteliales, que separan la sangre del líquido intersticial por sólo alrededor de 1 μm. Los capilares carecen de músculo liso y, así, de la capacidad para cambiar su diámetro de manera activa. Son tan numerosos que el área de corte transversal colectiva total de todos los capilares en los órganos sistémicos es más de 1000 veces la que hay en la raíz de la aorta. Dado que los capilares tienen alrededor de 0.5 mm de longitud, puede calcularse el área de superficie total disponible para intercambio de material entre la sangre y el líquido intersticial, por arriba de los 100 m2 y, por obvias razones, los capilares se consideran los vasos de intercambio del sistema cardiovascular. Además de la difusión transcapilar de solutos que ocurre a través de las paredes de estos vasos, a veces puede haber movimientos netos de líquido (volumen) hacia los capilares, o hacia afuera de los capilares, o en ambas direcciones, por ejemplo, la tumefacción de tejido (edema) es un resultado de movimiento neto de líquido desde el plasma hacia el espacio intersticial. Después de abandonar los capilares, la sangre es recolectada en vénulas y venas y devuelta al corazón. Los vasos venosos tienen paredes muy delgadas en proporción a su diámetro, sus paredes contienen músculo liso, y los diámetros de vasos venosos pueden cambiar de manera activa. Debido a sus paredes delgadas, los vasos venosos son bastante distensibles, por consiguiente, su diámetro cambia de manera pasiva en respuesta a cambios pequeños de la presión de distensión transmural (esto es, la diferencia entre las presiones interna y externa a través de la pared del vaso). Los vasos venosos, en especial los de mayor diámetro, también tienen válvulas unidireccionales que evitan el flujo en reversa, estas válvulas tienen especial importancia en la operación del sistema cardiovascular al permanecer de pie y durante el ejercicio (véase más adelante). Resulta ser que las vénulas y venas periféricas normalmente contienen más de 50% del volumen sanguíneo total, en consecuencia, comúnmente se consideran vasos de capacitancia. Lo que es más importante, los cambios del volumen 207 venoso influyen mucho sobre el llenado cardiaco y, por ende, sobre el bombeo del corazón, así, las venas periféricas en realidad desempeñan un papel de importancia extrema en el control del gasto cardiaco. CONTROL DE VASOS SANGUÍNEOS El flujo sanguíneo a través de lechos vasculares individuales está profundamente influido por cambios de la actividad de los nervios simpáticos que inervan arteriolas, estos nervios liberan norepinefrina a partir de sus terminaciones, que interactúa con receptores α-adrenérgicos sobre las células de músculo liso para causar contracción y, así, constricción arteriolar. La reducción del diámetro arteriolar aumenta la resistencia vascular y disminuye el flujo sanguíneo, estas fibras neurales proporcionan el medio más importante de control reflejo de la resistencia vascular y el flujo sanguíneo en órganos. El músculo liso arteriolar también tiene gran capacidad de respuesta a cambios en las condiciones químicas locales dentro de un órgano que acompañan a cambios del índice metabólico del órgano. Por razones que se comentarán más adelante, el índice metabólico tisular aumentado lleva a dilatación arteriolar y flujo sanguíneo tisular aumentado. Las vénulas y venas también están ricamente inervadas por nervios simpáticos, y se constriñen cuando estos nervios son activados, el mecanismo es el mismo que el que está involucrado con las arteriolas. Así, la actividad nerviosa simpática aumentada se acompaña de volumen venoso disminuido, la importancia de este fenómeno es que la constricción venosa tiende a aumentar el llenado cardiaco y, por ende, el gasto cardiaco mediante la ley de Starling del corazón. No hay control neural o metabólico local importante de los vasos arteriales o capilares. SANGRE La sangre es un líquido complejo que sirve como el medio para transportar sustancias entre los tejidos del organismo, y desempeña también muchas otras funciones. En circunstancias normales, alrededor de 40% del volumen de sangre entera es ocupado por células sanguíneas que están suspendidas en el plasma, que explica el resto del volumen. La fracción del volumen sanguíneo ocupada por células es un parámetro importante en clínica llamado el hematocrito: Hematocrito = volumen celular volumen sanguíneo total (5) CÉLULAS SANGUÍNEAS La sangre contiene tres tipos generales de “elementos formes”: eritrocitos, leucocitos y plaquetas (cuadro 22-1). Todos se forman en la médula ósea a partir de una célula madre común. Los eritrocitos son con mucho los más abundantes, están especializados para transportar oxígeno desde los pulmones hacia otros tejidos al unir oxígeno a la hemoglobina, una proteína hem que contiene hierro concentrado dentro de los eritrocitos. Debido a la presencia de hemoglobina, la sangre puede transportar 50 a 60 veces la cantidad de oxígeno que el 208 SECCIÓN V Fisiología cardiovascular CUADRO 22-1 Valores normales de eritrocitos, leucocitos y plaquetas en la sangre de un ser humano adulto.a Eritrocitos 4.0 a 5.5 millones/μl de sangre Plaquetas 130 000 a 400 000/μl de sangre Leucocitos 4000 a 10 000/μl de sangre Tipo de leucocito Por ciento de leucocitos totales Función primaria Neutrófilos 50 a 70 Fagocitosis Eosinófilos 1a4 Reacciones de hipersensibilidad alérgicas Basófilos 0 a 0.75 Reacción de hipersensibilidad alérgica Monocitos 2a8 Fagocitosis y producción de anticuerpos Linfocitos 15 a 40 Producción de anticuerpos e inmunidad mediada por células Granulocitos polimorfonucleares a Los rangos de referencia normales varían un poco con la edad, el género y la raza. También pueden variar de un laboratorio a otro. El tema se hace más confuso porque se usan diversas mediciones de unidad para reportar los valores en la sangre, de modo que es necesario ser cauto al interpretar datos. plasma solo podría transportar. Además, la capacidad amortiguadora de ion hidrógeno de la hemoglobina tiene importancia vital para la capacidad de la sangre para transportar dióxido de carbono. Una fracción pequeña pero importante de las células en la sangre son leucocitos, que están involucrados en procesos inmunitarios y tienen papeles específicos (cuadro 22-1). Las plaquetas son fragmentos de célula pequeños que son importantes en el proceso de coagulación de la sangre. PLASMA El plasma es el componente líquido de la sangre y, como se indica en el cuadro 22-2, es una solución compleja de electrólitos y proteínas. El suero es el líquido que se obtiene a partir de una muestra de sangre después de que se ha permitido que se coagule. Para todos los propósitos prácticos, la composición del suero es muy similar a la del plasma, excepto porque no contiene ninguna de las proteínas de la coagulación. Los electrólitos inorgánicos (iones como sodio, potasio, cloruro y bicarbonato) son los solutos más concentrados en el plasma, de éstos, el sodio y el cloruro son con mucho los más abundantes y, por ende, son la principal causa de la osmolaridad normal del plasma de alrededor de 300 mOsm/l. En una primera aproximación, el plasma es una solución 150 mM de cloruro de sodio, esa solución se llama solución salina isotónica, y tiene muchos usos clínicos como un líquido que es compatible con las células. El plasma por lo normal contiene muchas proteínas diferentes, y casi todas las proteínas plasmáticas pueden clasificarse como albúminas, globulinas o fibrinógeno con base en diferentes características físicas y químicas que se usan para separarlas. Se han identificado más de 100 proteínas plasmáticas distintas, y cada una probablemente desempeña alguna función específica. Muchas proteínas plasmáticas están involucradas en la coagulación de la sangre o en reacciones inmunitarias/de defensa, y muchas otras son proteínas transportadoras importantes para diversas sustancias, entre ellas ácidos grasos, hierro, cobre y ciertas hormonas. Las proteínas no cruzan fácilmente las paredes capilares y, en general, sus concentraciones plasmáticas son mucho mayores que sus concentraciones en el líquido intersticial, como se comentará, las proteínas plasmáticas desempeñan un papel osmótico importante en el movimiento de líquido transcapilar y, así, la distribución del volumen extracelular entre los compartimientos de plasma e intersticial. La albúmina desempeña un papel de especial importancia a este respecto, simplemente porque es con mucho la más abundante de las proteínas plasmáticas. El plasma también sirve como el vehículo para transportar nutrientes y productos de desecho, así, una muestra de plasma contiene muchas moléculas orgánicas pequeñas, como glucosa, aminoácidos, urea, creatinina y ácido úrico, cuyos valores medidos son útiles en el diagnóstico clínico. HEMOSTASIA Dondequiera que ocurra daño de un vaso sanguíneo, se inician diversos procesos dirigidos a prevenir la salida de sangre del espacio vascular o suspenderla. Los tres procesos primarios se resumen en los apuntes que siguen: 1. Agregación plaquetaria y formación de tapón: ocurren como resultado de los siguientes pasos: A) lesión de vaso con daño endotelial y exposición de colágeno; B) adherencia de plaquetas al colágeno (mediada por el factor de Von Willebrand); C) cambio de la forma de las plaquetas (desde discos hacia esferas espinosas); D) desgranulación plaquetaria con liberación de los siguientes: i) difosfato de adenosina, que hace que la agregación plaquetaria “tapone” la solución de continuidad, ii) tromboxano, que causa vasoconstricción, adhesión y agregación plaquetarias. CAPÍTULO 22 Perspectiva general del sistema cardiovascular 209 CUADRO 22-2 Constituyentes normales del plasma de ser humano adulto. Clase Cationes Constituyente Sodio 3.5 a 5.0 mEq/l (Ca2+) 4.3 a 5.2 mEq/l Magnesio Hierro (Mg2+) 1.2 a 1.8 mEq/l 60 a 160 μg/dl (Fe3+) Hidrógeno Aniones 136 a 145 mEq/l (K+) Potasio Calcio Rango de concentración normal (Na+) Cloruro (H+) 35 a 45 nmol/l (pH 7.35 a 7.45) (Cl–) 98 a 106 mEq/l –) Bicarbonato (HCO3 23 a 28 mEq/l Lactato 0.67 a 1.8 mEq/l 2– Proteínas Fosfato (HPO4 en su mayor parte) 3.0 a 4.5 mg/l Total (7% del peso del plasma) 6 a 8 g/dl Albúmina 3.4 a 5.0 g/dl Globulinas 2.2 a 4.0 g/dl Fibrinógeno 0.3 g/dl (Nota: La aspirina y otros inhibidores de la ciclooxigenasa son anticoagulantes porque evitan la formación de tromboxano.) 2. Vasoconstricción local: mediada en su mayor parte por tromboxano, pero también puede ser inducida por liberación local de otras señales químicas que constriñen vasos locales y reducen el flujo sanguíneo. 3. Coagulación de la sangre: la formación de un gel sólido constituido de la proteína, fibrina, plaquetas, y células sanguíneas atrapadas. El paso crucial en la coagulación de la sangre es la formación de trombina a partir de protrombina, que a continuación cataliza la conversión de fibrinógeno en fibrina. El coágulo final es estabilizado mediante enlaces covalentes entre cadenas de fibrina catalizados por el factor XIIIa (cuya formación es catalizada por trombina). La cascada de reacciones que ocurre desde la lesión de un vaso hasta la formación de trombina es como sigue: 1. lesión de vaso o daño de tejido con exposición de la sangre a células subendoteliales que liberan tromboplastina (“factor tisular”). 2. la proteína plasmática factor VII se une al factor tisular, que la convierte en una forma activada, el factor VIIa. 3. el factor VIIa cataliza la conversión de los factores tanto IX como X en formas activadas, IXa y Xa, respectivamente. 4. IXa también ayuda a convertir el factor X en Xa (factor Stuart). 5. Xa convierte la protrombina en trombina. 6. La trombina: A) activa las plaquetas (las hace pegajosas, induce desgranulación, promueve la fijación de diversos factores que participan en la coagulación); B) convierte el fibrinógeno en fibrina; C) recluta la “vía intrínseca”, que amplifica la formación adicional de factor Xa y facilita la conversión de protrombina en trombina al promover las reacciones que siguen: i) conversión de factor XI en su forma activada, XIa, que a continuación convierte el factor IX en IXa, que después se fija a plaquetas activadas y convierte el factor X en Xa, ii) conversión del factor VIII (que falta en personas con hemofilia) en su forma activada, VIIIa, que se fija a plaquetas activadas y acelera la conversión de factor X en Xa, iii) conversión del factor V en su forma activada, Va, que se fija a plaquetas activadas y acelera la conversión de protrombina en trombina. Varios agentes usados en clínica como anticoagulantes interfieren con diversos pasos en este proceso de coagulación. El dicumarol y la warfarina bloquean la actividad de la vitamina K, que es necesaria para la síntesis de muchos de los factores de la coagulación por el hígado. La heparina activa una proteína plasmática llamada antitrombina III que, a su vez, desactiva la trombina y varios de los otros factores de la coagulación. Dado que el calcio es un importante cofactor de la coagulación, los quelantes del calcio, como EDTA, oxalato y citrato se usan para evitar que la sangre almacenada se coagule. También se dispone de diversos agentes trombolíticos modelados con base en el activador del plasminógeno tisular (tPA) endógeno, que promueven la disolución del coágulo de fibrina después de que se forma. Estos agentes promueven la formación de plasmina a partir de plasminógeno que ataca enzimáticamente el coágulo, y lo convierte en péptidos solubles. 210 SECCIÓN V Fisiología cardiovascular ■ CORRELACIÓN CLÍNICA Un banquero de inversiones de 45 años de edad se desmayó en el centro comercial, mas estuvo consciente para el momento en que los paramédicos llegaron algunos minutos más tarde, pero fue llevado a la sala de urgencias para evaluación. Reportó que a últimas fechas había estado muy ocupado y un poco estresado, pero que por lo demás estaba sano. Tenía aspecto pálido y sintió un poco de náuseas y mareo justo antes del episodio, pero no recordó haber caído ni cualquier otra cosa sino hasta que el paramédico estaba inclinado sobre él en el suelo. Los datos físicos incluyeron: peso = 90 kg, altura = 1.77 m (5’10”), presión arterial = 130/85 mmHg, frecuencia cardiaca = 85 latidos/min, ruidos cardiacos y respiratorios normales, reflejos normales y cognición normal. Se obtuvo un electrocardiograma y se encontró que estaba normal. Se obtuvieron muestras de sangre para medir marcadores de infarto de miocardio e insuficiencia cardiaca y se encontró también que estaban normales. Fue admitido al hospital y se vigiló el ECG durante toda la noche, sin datos importantes. Los episodios de síncope (desmayo) inexplicables pueden ser el resultado de un problema nervioso (crisis convulsivas) o un decremento importante del flujo sanguíneo hacia el cerebro. Esto último puede originarse por fracaso del corazón para mantener suficiente gasto cardiaco (debido a una arritmia, insuficiencia valvular, o infarto que afecta una cantidad importante de la masa miocárdica), o una falla dentro de la vasculatura sistémica sea para mantener suficiente actividad contráctil (debido a supresión de las influencias nerviosas simpáticas, como en el síncope vasovagal) o para proporcionar suficiente riego sanguíneo arterial al cerebro (debido a oclusiones graves, a menudo transitorias, de la arteria carótida, que desencadenan un ataque isquémico transitorio [TIA]). Se efectuaron pruebas diagnósticas para evaluar estas posibilidades. Se colocó en el paciente un monitor Holter que registró su ECG durante un periodo de 24 h, y se encontró que fue normal. Siguiendo el proceso de eliminación, se emitió el diagnóstico de síncope vasovagal que por lo general se debe a un incremento de las eferencias vagales (parasimpáticas) hacia el corazón (capítulos 23 y 29). En la mayor parte de los casos de síncope vasovagal no se requiere tratamiento. RESUMEN DEL CAPÍTULO ■ ■ ■ ■ La función primaria del sistema cardiovascular es mantener la homeostasis del líquido intersticial. La ley física que rige la operación cardiovascular es que el flujo a través de cualquier segmento es igual a la diferencia de presión a través de ese segmento dividida por su resistencia al flujo, es decir, Q. = ΔP/R. El corazón bombea sangre mediante llenado con sangre y eyección de esta última, rítmicos, desde las cámaras ventriculares que tienen válvulas de entrada y salida unidireccionales pasivas. Los cambios de la frecuencia cardiaca y del volumen sistólico (y, por ende, el gasto cardiaco) pueden acompañarse de alteraciones del llenado ventricular y de la actividad nerviosa del sistema nervioso autónomo hacia el corazón. ■ ■ ■ El flujo de sangre a través de órganos individuales está regulado por cambios en el diámetro de sus arteriolas. Los cambios del diámetro arteriolar pueden lograrse mediante alteraciones de la actividad nerviosa simpática y por variaciones en las condiciones locales. La sangre es una suspensión compleja de eritrocitos, leucocitos y plaquetas en plasma que es ideal para transportar gases, sales, nutrientes y moléculas de desecho por todo el sistema. La hemostasia comprende agregación plaquetaria, vasoconstricción local y coagulación de la sangre. PREGUNTAS DE ESTUDIO 1. Usted necesita determinar la dosis correcta de un fármaco IV que sólo se distribuye dentro del espacio extracelular. ¿Cuál de los valores que siguen sería el estimado más cercano del volumen del líquido extracelular de un varón adulto joven y sano que pesa 100 kg (220 libras)? A) 3 L B) 5 L C) 8 L D) 10 L E) 20 L 2. Usted ha transfundido 1 L de sangre a su paciente deshidratado; en cualquier instante, ¿en cuál segmento del lecho vascular se encontrará casi toda esta sangre? A) arterias B) arteriolas C) capilares D) venas E) aurícula derecha 3. ¿Cuál de los que siguen producirá el mayor incremento del flujo sanguíneo a través de músculos que están haciendo ejercicio? A) disminución a la mitad de la longitud de los capilares B) disminución a la mitad de la viscosidad de la sangre C) duplicación del radio de las vénulas D) duplicación de la diferencia de presión arterial a través del lecho E) duplicación del radio de las arteriolas 4. Un individuo ha tenido “gripe” durante tres días, con vómitos y diarrea graves sin aumentar su ingestión de líquido. ¿Cómo es probable que esto influya sobre su hematocrito? A) efecto nulo B) más alto que lo normal C) más bajo que lo normal D) es imposible de predecir E) lo elimina 5. Calcule el gasto cardiaco a partir de los datos que siguen: presión en la arteria pulmonar = 20 mmHg; presión en la aurícula izquierda = 5 mmHg; resistencia vascular pulmonar = 3 mmHg por l/min. A) 3 l/min. B) 5 l/min. C) 4 l/min. D) 15 l/min. E) 60 mmHg. Células de músculo cardiaco C A P Í T U L O 23 Lois Jane Heller y David E. Mohrman O B J E T I V O S ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ Describir las características de los potenciales de reposo cardiacos y potenciales de acción cardiacos de respuesta “rápida” y “lenta”. Identificar los periodos refractarios del ciclo eléctrico de la célula cardiaca. Definir el potencial umbral y describir la interacción entre el estado de los canales iónicos y el potencial de membrana durante la fase de despolarización del potencial de acción. Comprender el potencial marcapaso y describir la base para la actividad eléctrica rítmica de las células cardiacas. Listar las fases del ciclo eléctrico de la célula cardiaca y enunciar las alteraciones de la permeabilidad de membrana de las cuales depende cada fase. Describir las uniones intercelulares comunicantes (conexiones comunicantes) y su papel en la excitación cardiaca. Explicar la vía normal de la conducción del potencial de acción a través del corazón. Indicar la cronología de la excitación eléctrica de diversas áreas del corazón, e identificar las formas características del potencial de acción y las velocidades de conducción en cada una de las partes principales del sistema de conducción. Enunciar la relación entre eventos eléctricos de excitación cardiaca y las ondas P, QRS y T, el intervalo PR y el segmento ST del electrocardiograma. Señalar cómo los potenciales diastólicos de células marcapaso pueden ser alterados para producir cambios en la frecuencia cardiaca. Describir cómo los nervios simpáticos y parasimpáticos cardiacos alteran la frecuencia cardiaca y la conducción de potenciales de acción cardiacos. Definir los términos cronotrópico y dromotrópico. Determinar el proceso de excitación-contracción en el músculo cardiaco, y describirlo. Comprender las contracciones isométrica, isotónica y con poscarga del músculo cardiaco. Identificar la influencia de la precarga alterada sobre las capacidades productoras de tensión, y de acortamiento, del músculo cardiaco. Describir la influencia de la poscarga alterada sobre las capacidades de acortamiento del músculo cardiaco. Definir los términos contractilidad y estado inotrópico, y describir la influencia de la contractilidad alterada sobre las capacidades productoras de tensión, y de acortamiento del músculo cardiaco. Comprender el efecto de la actividad nerviosa simpática alterada sobre el estado inotrópico cardiaco. Señalar las relaciones entre el volumen ventricular y la longitud del músculo ventricular, y entre la presión intraventricular y la tensión muscular; explicar la ley de Laplace. 211 212 SECCIÓN V Fisiología cardiovascular CARACTERÍSTICAS DE LAS CÉLULAS DE MÚSCULO CARDIACO La coordinación de la actividad de las células de músculo (miocitos) cardiaco depende de un estímulo eléctrico que es iniciado regularmente a una frecuencia apropiada, y conducido de manera fiable por todo el corazón. La acción de bombeo mecánica depende de una contracción robusta de las células musculares que da lugar a ciclos repetitivos de desarrollo de tensión, acortamiento y relajación; además, debe haber disponibles mecanismos para ajustar las características de excitación y contracción para satisfacer las demandas cambiantes sobre el sistema circulatorio. PROPIEDADES ELÉCTRICAS La contracción de los miocitos cardiacos es desencadenada por potenciales de acción que ocurren en sobre la membrana celular. Los potenciales de acción cardiacos difieren mucho de los del músculo esquelético en tres aspectos importantes que promueven la excitación rítmica sincrónica del corazón: 1) pueden ser autogenerados; 2) pueden ser conducidos directamente de célula a célula, y 3) tienen una duración larga, lo que impide la fusión contracciones musculares de individuales. Para entender estas propiedades eléctricas especiales del músculo cardiaco, y cómo la función cardiaca depende de ellas, deben revisarse con sumo cuidado las propiedades eléctricas básicas de las membranas celulares excitables descritas en los capítulos 3 y 6. RESUMEN DE POTENCIALES DE MEMBRANA DE CÉLULAS CARDIACAS En reposo, las membranas de las células cardiacas son más permeables al potasio que a cualquier otro ion, dado que la concentración de potasio dentro de las células es significativamente más alta que la del líquido intersticial (150 mM en contraposición con 4 mM, respectivamente), la difusión de potasio hacia afuera a favor de su gradiente de concentración es equilibrada por la generación de un potencial de membrana (esto es, el potencial de equilibrio de potasio). Los gradientes tanto eléctrico como de concentración favorecen la entrada de Na+ y Ca2+ a las células en reposo; sin embargo, la permeabilidad muy baja de la membrana en reposo al Na+ y Ca2+, en combinación con una bomba de sodio que requiere energía y que extruye Na+ desde la célula, evitan que el Na+ y el Ca2+ se acumulen gradualmente dentro de la célula en reposo. El fuerte gradiente de sodio promueve la eliminación de Ca2+ desde el citoplasma por medio del intercambiador de sodio/calcio. Los potenciales de acción de células de diferentes regiones del corazón no son idénticos, sino que tienen características variables que dependen de diferencias de los patrones de cambios en sus permeabilidades iónicas. Algunas células cardiacas tienen la capacidad para actuar como marcapaso y para iniciar de manera espontánea potenciales de acción, no así las células de músculo cardiaco ordinarias (excepto en condiciones poco comunes). En la figura 23-1 se muestran las características eléctricas de membrana básicas de una célula de músculo cardiaco ordinaria y de una célula tipo marcapaso cardiaco. Los potenciales de acción provenientes de estos tipos de células se denominan potenciales de acción de “respuesta rápida” y de “respuesta lenta”, respectivamente. Como se muestra en el panel A de esta figura, los potenciales de acción de respuesta rápida se caracterizan por una rápida despolarización (fase 0) con un exceso sustancial (voltaje interno positivo), una rápida reversión de potencias en exceso (fase 1), una meseta larga (fase 2) y una repolarización (fase 3) a un estado alto (es decir, de alto potencial negativo) de membrana en reposo (fase 4). En comparación, los potenciales de acción de respuesta lenta se caracterizan por una fase de despolarización inicial más lenta (fase 0), una amplitud menor de sobreimpulso, una fase de meseta más corta y menos estable (fase 2), y una repolarización (fase 3) hacia un potencial de “reposo” inestable, lentamente despolarizante (fase 4) (figura 23-1B). El potencial de reposo inestable que se observa en células marcapaso con potenciales de acción de respuesta lenta se denomina de manera variada la despolarización fase 4, despolarización diastólica, o potencial marcapaso. Esas células por lo general se encuentran en los nodos sinoauricular (SA) y auriculoventricular (AV). Como se indica en la figura 23-1A, las células están en un estado refractario absoluto durante casi todo el potencial de acción (es decir, no pueden ser estimuladas para que activen otro potencial de acción). Cerca del final del potencial de acción, la membrana es relativamente refractaria, y sólo puede volver a ser excitada por un estímulo de magnitud mayor que lo normal, justo después del potencial de acción, la membrana es transitoriamente hiperexcitable, y se dice que está en un periodo “vulnerable” o “supranormal”. Durante potenciales de acción lentos ocurren alteraciones similares de la excitabilidad de membrana, pero en la actualidad no están bien caracterizadas. Recuerde que el potencial de membrana de cualquier célula en cualquier instante dado, depende de la permeabilidad relativa de la membrana celular a iones específicos al igual que en todas las células excitables, los potenciales de acción de células cardiacas son el resultado de cambios transitorios de la permeabilidad iónica de la membrana celular que son desencadenados por una despolarización inicial. Los paneles C y D de la figura 23-1 indican los cambios de las permeabilidades de membrana a K+, Na+ y Ca2+ que producen las diversas fases de los potenciales de acción de respuesta rápida y lenta. Note que durante la fase de reposo las membranas de ambos tipos de células están dominadas por una permeabilidad más alta al K+ que al Na+ o al Ca2+, por ende, los potenciales de membrana están más cerca al potencial de equilibrio de potasio durante este periodo que al potencial de equilibrio de cualquier otro ion. En las células tipo marcapaso, se cree que al menos tres mecanismos contribuyen a la despolarización lenta de la membrana que se observa durante el intervalo diastólico: 1) hay un decremento lento progresivo de la permeabilidad de la membrana al K+; 2) hay un aumento lento de la permeabilidad al Na+ y 3) hay un incremento leve de la permeabilidad de la membrana a iones calcio. Cuando el potencial de membrana se despolariza hasta el potencial umbral en cualquier tipo de célula, se desencadenan alteraciones rápidas e importantes de la permeabilidad de la membrana a iones específicos y, una vez iniciados, estos cambios de permeabilidad no se pueden suspender, y proceden hasta que se completan. La fase de aumento rápido característico del potencial de acción de respuesta rápida es resultado de un incremento repentino de la permeabilidad al Na+. Este periodo de permeabilidad muy alta al CAPÍTULO 23 Células de músculo cardiaco B Potenciales de acción de respuesta rápida Potenciales de acción de respuesta lenta Fase 2 Fase 0 0 –50 3 Fase Potencial transmembrana (mV) A 213 e Fas 4 Fase 4 –100 Periodo refractario absoluto Periodo refractario relativo Periodo supranormal Permeabilidad de membrana relativa C D 10.0 Na+ K Ca2+ Ca2+ + K + 1.0 Na+ 0.1 0 0.15 Tiempo (s) 0.30 0 0.15 Tiempo (s) 0.30 FIGURA 231 Evolución temporal de los cambios de potencial de membrana y permeabilidad a iones que ocurren durante potenciales de acción de “respuesta rápida” (A y C) y de “respuesta lenta” (B y D). (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) sodio (fase 0) es breve (figura 23-1C) y va seguido por un incremento breve de la permeabilidad al potasio (que no se muestra en la figura 23-1C). Esta corriente breve explica la repolarización muy temprana que sigue de inmediato a la fase de aumento inicial del potencial de acción (fase 1). El desarrollo y el mantenimiento de un estado de meseta despolarizado (fase 2) depende de: 1) una reducción sostenida de la permeabilidad al K+; 2) un aumento lento y sostenido de la permeabilidad de membrana a Ca2+ y 3) acción electrogénica de un intercambiador de Na+/Ca2+ en el cual, tres iones Na+ entran a la célula en intercambio por un Ca2+ único, que sale de la célula. En células con el potencial de acción de respuesta lenta, la corriente de entrada rápida inicial es pequeña (o incluso no existe), por consiguiente, la fase de aumento lento de estos potenciales de acción es principalmente el resultado de un movimiento hacia adentro de iones Ca2+. En ambos tipos de células, la membrana es repolarizada (fase 3) a su potencial de reposo original a medida que la permeabilidad al K+ aumenta y las permeabilidades al Ca2+ y Na+ vuelven a sus valores en reposo bajos. Recuérdese que los cambios de permeabilidad suavemente graduados que producen potenciales de acción son el resultado neto de alteraciones en cada uno de los muchos canales iónicos individuales dentro de la membrana plasmática de una célula única (capítulo 6). En el cuadro 23-1 se resumen algunas de las principales corrientes y tipos de canal (sensibles tanto a voltaje como a ligando) involucrados en la actividad eléctrica de las células cardiacas. Algunos de los canales sensibles a voltaje muestran respuesta a un cambio del potencial de membrana sostenido, de inicio repentino, con sólo un periodo breve de activación, empero, los cambios en el potencial de membrana de inicio más lento pero de la misma magnitud pueden no activar estos canales en absoluto, para explicar esa conducta, se postula que estos canales tienen dos “compuertas” que operan de manera independiente —una compuerta de activación y una compuerta de inactivación— ambas deben estar abiertas para que el canal como un todo se abra. Estas dos compuertas muestran respuesta a cambios del potencial de membrana, pero lo hacen con sensibilidades a voltaje y evoluciones temporales diferentes. Con una despolarización repentina hasta el umbral, las compuertas de activación de canal de sodio se abren y dentro de algunos milisegundos sus compuertas de inactivación se cierran. En células marcapaso con despolarización diastólica lenta, las compuertas de inactivación de los canales de sodio se cierran antes de que las compuertas de activación tengan una oportunidad de abrirse, cuando se alcanza el umbral, sólo el canal de calcio está disponible para abrirse, lo que explica la fase de aumento lento inicial de los potenciales de acción en células marcapaso. El cierre de la compuerta de inactivación de canal de calcio se retrasa más de 100 ms hasta cerca del final de la 214 SECCIÓN V Fisiología cardiovascular CUADRO 23-1 Características de canales iónicos cardiacos importantes en orden de su participación en un potencial de acción. Corriente Canal Mecanismo de compuerta iK1 Canal Kir+ (rectificador interno) Voltaje Mantiene la permeabilidad al K+ alta durante la fase 4. Su descomposición contribuye a la despolarización diastólica. Su supresión durante las fases 0 a 2 contribuye a la meseta. iNa Canal de Na+ (rápido) Voltaje Explica la fase 0 del potencial de acción. La desactivación puede contribuir a la fase 1 del potencial de acción. Ito Canal de K+ (hacia afuera transitorio) Voltaje Contribuye a la fase 1 del potencial de acción. iCa Canal de Ca2+ (canales hacia adentro lentos, L) Ambos Contribuye a la fase 2 del potencial de acción. La desactivación puede contribuir a la fase 3 del potencial de acción. Es aumentado por estimulación simpática y agentes β-adrenérgicos. iK Canal de K+ (rectificador tardío) Voltaje Causa la fase 3 del potencial de acción. Es aumentado por Ca2+ intracelular aumentado. iKATP Canal de K+ (sensible a ATP) Ligando Incrementa la permeabilidad al K+ cuando el [ATP] es bajo. iKACh Canal de K+ (activado por acetilcolina) Ligando Los efectos de la estimulación vagal dependen de él. Disminuye la despolarización diastólica (y la frecuencia cardiaca). Hiperpolariza el potencial de membrana en reposo. Acorta la fase 2 del potencial de acción. (“curioso”, “funny”) Na+ (corriente marcapaso) Ambos Es activado por hiperpolarización y contribuye a la despolarización diastólica. Es aumentado por la estimulación simpática y agentes β-adrenérgicos. Es suprimido por estimulación vagal. Papel funcional fase de meseta. Las compuertas de inactivación en los canales de sodio y de calcio permanecen cerradas hasta que la membrana se repolariza. Esto explica el periodo refractario prolongado de células de músculo cardiaco. Múltiples factores influyen sobre la operación de canales de K+, algunos de los cuales se resumen en el cuadro 23-1, por ejemplo, la concentración intracelular alta de Ca2+ contribuye a la activación de algunos canales de K+ durante la repolarización. Aunque las células en ciertas áreas del corazón típicamente tienen potenciales de acción de tipo rápido, y las células en otras áreas normalmente tienen potenciales de acción de tipo lento, es importante reconocer que todas las células cardiacas son en potencia capaces de tener uno u otro tipo de potencial de acción dependiendo de sus potenciales de membrana en reposo y de la rapidez con la cual se despolarizan hasta el potencial umbral. La despolarización rápida hasta el potencial umbral generalmente es un evento forzado en una célula por la aparición de un potencial de acción en una célula adyacente. La despolarización lenta hasta el umbral ocurre cuando una célula pierde de manera espontánea y gradual por sí mismas su polarización de reposo, lo que normalmente sólo sucede en el nodo SA. Una despolarización moderada crónica de la membrana en reposo (causada, p. ej., por concentración extracelular moderadamente alta de K+) puede desactivar los canales de sodio rápidos (esto es, evitar que se abran) sin inactivar los canales de calcio lentos. En estas condiciones, los potenciales de acción de todas las células cardiacas serán del tipo lento. Con todo, las despolarizaciones sostenidas grandes pueden inactivar los canales tanto rápido como lento y, así, hacer a las células de músculo cardiaco por completo inexcitables. CONDUCCIÓN DE POTENCIALES DE ACCIÓN CARDIACOS Los potenciales de acción son conducidos sobre la superficie de células individuales porque la despolarización activa, en cualquier área de la membrana produce corrientes locales en los líquidos intracelular y extracelular que despolarizan de manera pasiva áreas de la membrana inmediatamente adyacentes hasta su umbral de voltaje para despolarización activa. En el corazón, las células de músculo cardiaco están conectadas una a otra en sus porciones terminales por estructuras llamadas discos intercalados. Estos discos contienen: 1) fijaciones mecánicas firmes entre membranas celulares adyacentes por medio de proteínas llamadas adherinas en estructuras denominadas desmosomas y 2) conexiones eléctricas de resistencia baja entre células adyacentes a través de canales formados por una proteína llamada conexina en estructuras denominadas uniones intercelulares comunicantes (conexiones comunicantes). En la figura 23-2 se muestra de manera esquemática cómo estas uniones intercelulares comunicantes permiten la propagación del potencial de acción de una célula a otra. Las células B, C y D se muestran en la fase de reposo con cargas más negativas en el interior que en el exterior. La célula A se muestra en la fase de meseta en un potencial de acción, y tiene más cargas positivas adentro que afuera. Debido a las uniones intercelulares comunicantes, la atracción electrostática puede causar un flujo de corriente (movimiento iónico) local entre la membrana despolarizada de una célula A activa y la membrana polarizada de la célula B en CAPÍTULO 23 Células de músculo cardiaco – + + + + + – – – – – –+ ++ Célula B en reposo – – – – – + + + – – – – – – + + + + + + – – – – – – – – + + + + + + + Célula A con potencial de acción + + + + + + + – – – – – – – + + + + – – + + – – – – + – + + – – + – 215 + + + – – – Célula C en reposo – – – – – + + + + + + + + + + + + – – – – – – – – Célula D – –+ + en reposo – – – – – – + + + – – – – + + + + – – + + + + – – Unión intercelular comunicante (nexo) FIGURA 232 Corrientes locales y conducción célula a célula de potenciales de acción de células de músculo cardiaco. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) reposo, como lo indican las flechas en la figura. Este movimiento iónico despolariza la membrana de la célula B. Una vez que las corrientes locales provenientes de la célula A activa despolarizan la membrana de la célula B cerca de la unión intercelular comunicante hasta la magnitud umbral, se desencadenará un potencial de acción en ese sitio, y será conducido sobre la célula B. Dado que la célula B se ramifica (una característica morfológica común de las fibras de músculo cardiaco), su potencial de acción evocará potenciales de acción sobre células C y D. Este proceso se continúa en todo el miocardio. De este modo, un potencial de acción iniciado en cualquier sitio en el miocardio será conducido de célula a célula por todo el miocardio. La rapidez con la cual un potencial de acción se propaga a través de una región de tejido cardiaco se llama velocidad de conducción, que varía considerablemente en distintas áreas en el corazón, y está determinada por tres variables: 1. La velocidad de conducción depende de manera directa del diámetro de la fibra muscular involucrada. Así, la conducción por células de diámetro pequeño en el nodo AV es significativamente más lenta que la conducción sobre células de diámetro grande en el sistema de Purkinje ventricular. 2. La velocidad de conducción también depende de manera directa de la intensidad de las corrientes despolarizantes locales, que a su vez está determinada de manera directa por la tasa de aumento del potencial de acción. La despolarización rápida favorece la conducción rápida. 3. La velocidad de conducción depende de las propiedades de capacidad o resistencia o ambas, de las membranas celulares, las uniones intercelulares comunicantes, y el citoplasma. Las características eléctricas de las uniones intercelulares comunicantes pueden estar influidas por condiciones externas que promueven la fosforilación/desfosforilación de las proteínas conexina. En la figura 23-3 se muestran detalles de las consecuencias generales del sistema de conducción cardiaco. Como se mencionó, las adaptaciones eléctricas específicas de diversas células en el corazón se reflejan en la forma característica de sus potenciales de acción que –100 Músculo auricular mV Nodo SA A B Músculo auricular C Nodo AV D Fibra de Purkinje E Músculo ventricular Músculo ventricular F G R Onda P Onda T ECG Q S Intervalo PR Segmento ST Intervalo QT 1.0 s FIGURA 233 Actividad eléctrica del corazón: registros de voltaje de célula única (trazos A a G) y electrocardiograma de la derivación II. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) 216 SECCIÓN V Fisiología cardiovascular se muestran en la mitad derecha de la figura 23-3. Note que los potenciales de acción que se muestran en la figura 23-3 han sido posicionados para indicar el momento en el cual el impulso eléctrico que se origina en el nodo SA llega a otras áreas del corazón. Las células del nodo SA actúan como el marcapaso normal del corazón, y determinan la frecuencia cardiaca, esto se debe a que la despolarización diastólica espontánea de la membrana en reposo es más rápida en las células del nodo SA, y alcanzan su potencial umbral antes que las células en otros lugares del corazón. El potencial de acción iniciado por una célula del nodo SA primero se disemina de manera progresiva a través de la pared auricular. En la figura 23-3 se muestran potenciales de acción de células en dos regiones diferentes de las aurículas: una cerca del nodo SA y una a mayor distancia de dicho nodo, ambas células tienen potenciales de acción de forma similar, pero su desplazamiento temporal refleja el hecho de que se requiere cierto tiempo para que el impulso se disemine sobre las aurículas. La conducción del potencial de acción se disminuye mucho conforme pasa por el nodo AV, esto se debe al tamaño pequeño de las células del nodo AV y la tasa lenta de aumento de sus potenciales de acción. Puesto que el nodo AV retrasa la transferencia de la excitación cardiaca desde las aurículas hacia los ventrículos, la contracción auricular puede contribuir al llenado ventricular justo antes de que los ventrículos se contraigan, advierta también que las células del nodo AV tienen una despolarización espontánea más rápida durante el periodo en reposo que otras células del corazón, excepto las del nodo SA. El nodo AV a veces se denomina un marcapaso latente, y en muchas situaciones patológicas controla el ritmo cardiaco (más que el nodo SA). Debido a los potenciales de acción que aumentan de manera rápida y otros factores, como células de diámetro grande la conducción eléctrica es en extremo rápida en las fibras de Purkinje, esto permite que el sistema de Purkinje transfiera casi al unísono el impulso cardiaco hacia células en muchas áreas del ventrículo. En la figura 23-3 se muestran potenciales de acción provenientes de células musculares en dos áreas del ventrículo. Debido a la velocidad de conducción alta en el tejido ventricular, sólo hay una discrepancia pequeña en su tiempo de inicio. Note que las células ventriculares que son las últimas en despolarizarse tienen potenciales de acción de duración más breve y, así, son las primeras en repolarizarse. La importancia fisiológica de esta conducta inesperada no está clara, pero influye sobre los electrocardiogramas que se comentarán en el capítulo 25. ELECTROCARDIOGRAMAS Los campos de potencial eléctrico causados por la actividad eléctrica del corazón se extienden a través del líquido extracelular del cuerpo, y pueden medirse con electrodos colocados sobre la superficie del cuerpo. La electrocardiografía proporciona un registro de cómo la diferencia de voltaje entre dos puntos sobre la superficie del cuerpo cambia con el tiempo como resultado de los eventos eléctricos del ciclo cardiaco. En cualquier instante del ciclo cardiaco, el electrocardiograma indica el campo eléctrico neto que es la suma de muchos campos eléctricos débiles que están siendo producidos por los cambios de voltaje que están ocurriendo en células cardiacas individuales. Cuando un gran número de células se están despolarizando o repolarizando de manera simultánea, se observan voltajes grandes en el electrocardiograma, puesto que el impulso eléctrico se disemi- na a través del tejido cardiaco en una vía constante, el patrón temporal de cambio de voltaje registrado entre dos puntos de la superficie del cuerpo también es constante y se repetirá por sí mismo con cada ciclo cardiaco. El trazo inferior de la figura 23-3 representa un registro típico de los cambios de voltaje normalmente medidos entre el brazo derecho y la pierna izquierda conforme el corazón pasa por dos ciclos de excitación eléctrica; este registro se llama un electrocardiograma de la derivación II. Las principales características de un electrocardiograma son la onda P, el complejo QRS y la onda T. La onda P corresponde a la despolarización auricular, el complejo QRS a la despolarización ventricular, y la onda T a la repolarización ventricular. CONTROL DE LA FRECUENCIA CARDIACA Las contracciones rítmicas normales del corazón ocurren debido a la actividad de marcapaso eléctrico espontánea (automaticidad) de las células en el nodo SA. El intervalo entre latidos cardiacos (y, así, la frecuencia cardiaca) está determinado por el tiempo que requieren las membranas de estas células marcapaso en despolarizarse de manera espontánea hasta la cifra umbral. Las células del nodo SA se activan a una frecuencia espontánea o intrínseca (≈ 100 latidos/min) en ausencia de cualquier influencia externa. Las dos influencias externas más importantes sobre la automaticidad de las células del nodo SA provienen del sistema nervioso autónomo (SNA) (capítulo 19). Las fibras provenientes de las divisiones tanto simpática como parasimpática del SNA terminan sobre células en el nodo SA, y estas fibras pueden modificar la frecuencia cardiaca intrínseca. La activación de los nervios simpáticos cardiacos (que aumentan el tono simpático del corazón) incrementa la frecuencia cardiaca. El aumento del tono parasimpático del corazón disminuye la frecuencia cardiaca. Los nervios parasimpáticos y simpáticos influyen sobre la frecuencia cardiaca al alterar el curso de la despolarización espontánea del potencial de reposo en células marcapaso SA (figura 23-4). Las fibras parasimpáticas cardiacas, que viajan al corazón por los nervios vagos, liberan la sustancia transmisora acetilcolina sobre células del nodo SA. La acetilcolina aumenta la permeabilidad de la membrana en reposo al K+, y disminuye la permeabilidad diastólica al Na+. El proceso de emisión de señales involucra interacción de la acetilcolina con receptores muscarínicos en la membrana de células del nodo SA, que a su vez están enlazados con proteínas G inhibitorias, Gi. La activación de Gi tiene dos efectos: 1) un incremento de la permeabilidad al K+ que se produce por una abertura aumentada de los canales KAch y 2) una supresión de la adenilato ciclasa que lleva a un decremento de la concentración intracelular de monofosfato de adenosina cíclico (cAMP) que reduce la corriente marcapaso que va hacia adentro transportada por Na+ (if ). Estos cambios de permeabilidad tienen dos efectos sobre el potencial de reposo de células marcapaso cardiacas: 1) causan una hiperpolarización inicial del potencial de membrana en reposo al llevarlo más cerca al potencial de equilibrio de K+ y 2) hacen más lenta la frecuencia de despolarización espontánea de la membrana en reposo (figura 23-4). Estos dos efectos aumentan el tiempo entre latidos al prolongar el tiempo que se requiere para que la membrana en reposo se despolarice hasta la cifra umbral. Puesto que normalmente hay cierta actividad tónica CAPÍTULO 23 Células de músculo cardiaco 217 –50 Tono pa rasimpá tico –30 PROPIEDADES MECÁNICAS Intrínseco –10 Tono simpático Potencial de membrana (mV) nodo AV y pueden influir sobre el tiempo entre las ondas P y R (el intervalo PR). –70 Potencial umbral –90 Tiempo FIGURA 234 Efecto de los tonos simpático y parasimpático sobre el potencial marcapaso. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) continua de nervios parasimpáticos cardiacos, la frecuencia cardiaca en reposo normal es de aproximadamente 70 latidos/min. Los nervios simpáticos liberan la sustancia transmisora norepinefrina sobre células cardiacas. Además de otros efectos que se comentan más adelante, la norepinefrina aumenta las corrientes hacia adentro llevadas por el Na+ (if ) y por el Ca2+ durante el intervalo diastólico. El proceso de emisión de señales involucra interacción de norepinefrina con receptores β1-adrenérgicos en la membrana de células del nodo SA, que a su vez están enlazados con proteínas G estimulantes Gs. La activación de Gs aumenta la actividad de la adenilato ciclasa, lo que lleva a un incremento del cAMP intracelular que aumenta la probabilidad de estado abierto del canal de corriente de Na+ marcapaso (if ). Estos cambios aumentan la frecuencia cardiaca al incrementar la tasa de despolarización diastólica (figura 23-4). Además de los nervios simpáticos y parasimpáticos, hay muchos factores (por lo general menos importantes) que pueden alterar la frecuencia cardiaca, éstos incluyen varios iones y varias hormonas circulantes, así como influencias físicas, como la temperatura y la distensión de la pared auricular. Todos actúan al alterar el tiempo requerido para que la membrana en reposo se despolarice hasta el potencial umbral, por ejemplo, una concentración anormalmente alta de Ca2+ en el líquido extracelular tiende a disminuir la frecuencia cardiaca al cambiar el potencial umbral. Se dice que los factores que aumentan la frecuencia cardiaca tienen un efecto cronotrópico positivo. Los que disminuyen la frecuencia cardiaca tienen un efecto cronotrópico negativo. Un aumento de la actividad simpática también incrementa la velocidad de conducción del potencial de acción (tiene un efecto dromotrópico positivo), mientras que un aumento de la actividad parasimpática disminuye la velocidad de conducción (tiene un efecto dromotrópico negativo). Estos efectos son más notables en el La contracción de la célula de músculo cardiaco es iniciada por el potencial de acción que actúa sobre organelos intracelulares para evocar generación de tensión, o acortamiento, o ambos, de la célula. Se recomienda al lector que revise meticulosamente los materiales que se presentan en los capítulos 9 y 10 sobre detalles celulares específicos que describen la contracción de los músculos esquelético y cardiaco. ACOPLAMIENTO EXCITACIÓN CONTRACCIÓN El evento importante en el acoplamiento excitación-contracción en el músculo cardiaco es un aumento notorio de la concentración de Ca2+ libre intracelular desde menos de 0.1 μM hasta una cifra tan alta como 100 μM. Cuando la onda de despolarización pasa sobre la membrana de la célula muscular, se libera Ca2+ desde el retículo sarcoplasmático (SR) hacia el líquido intracelular. El desencadenante específico es un pequeño incremento localizado, de la concentración de calcio que desencadena una liberación masiva de calcio desde el SR. Aunque la cantidad de Ca2+ que entra a la célula durante un potencial de acción único es bastante pequeña en comparación con la que se libera desde el SR, no sólo es esencial para desencadenar la liberación de calcio desde el SR, sino que también lo es para mantener concentración adecuada de Ca2+ en las reservas intracelulares a largo plazo. El proceso contráctil iniciado por el incremento de la concentración intracelular de calcio se describió en los capítulos 9 y 10, recuerde que el acoplamiento excitación-contracción en el músculo cardiaco difiere del que se observa en el músculo esquelético por cuanto puede ser modulado; un desencadenante de potencial de acción único en el músculo cardiaco puede producir diferentes intensidades de interacción actina-miosina (contracción) que puede resultar de un disparo del potencial de acción única en el músculo cardiaco. El mecanismo para esto depende en gran parte de variaciones de la cantidad de Ca2+ que llega a los miofilamentos y, por ende, el número de puentes activados durante la contracción. Esta capacidad del músculo cardiaco para variar su fuerza contráctil; es decir, cambiar su contractilidad, tiene importancia extrema para la función cardiaca (véase más adelante en este capítulo). RELAJACIÓN Los procesos que participan en la reducción del Ca2+ intracelular que termina la contracción comprenden: 1) captación activa de ~80% del calcio de regreso hacia el SR por la acción de bombas Ca2+ATPasa, 2) extrusión activa de ~5% del calcio desde la célula por medio de bombas Ca2+-ATPasa sarcolémicas y 3) intercambio pasivo de ~15% del calcio con sodio extracelular por medio del intercambiador de Na+-Ca2+ ubicado en el sarcolema. El intercambiador de Na+-Ca2+ es activado por el gradiente de sodio a través del sarcolema, que a su vez es mantenido por la Na+/K+-ATPasa. Este inter- 218 SECCIÓN V Fisiología cardiovascular cambiador es electrogénico por cuanto tres iones Na+ entran a la célula en intercambio por cada ion Ca2+ que sale. Este movimiento hacia adentro neto de carga positiva tal vez contribuya al mantenimiento de la fase de meseta del potencial de acción. El glucósido cardiaco digital, disminuye la bomba de Na+/K+ y, así, reduce el gradiente de sodio, lo que da por resultado un incremento del Ca2+ intracelular que queda secuestrado en el SR. Este mecanismo contribuye de manera importante al efecto positivo de los glucósidos cardiacos sobre la fuerza contráctil del corazón insuficiente. La duración de la contracción de la célula de músculo cardiaco es aproximadamente la misma que la de su potencial de acción, por consiguiente, el periodo refractario eléctrico de una célula de músculo cardiaco no termina sino hasta que se completa la respuesta mecánica. La relajación mecánica acompaña la repolarización eléctrica, como consecuencia, las células de músculo cardiaco no pueden ser activadas con suficiente rapidez para causar un estado fusionado (tetánico) de contracción prolongada, esto es afortunado porque la contracción y relajación intermitentes son esenciales para la acción de bombeo del corazón. La interacción de puentes que ocurre después de que un músculo es activado para que se contraiga imparte al músculo el potencial para desarrollar fuerza, o acortarse, o ambas (capítulos 9 y 10). El hecho de si hace una, la otra o alguna combinación de ambas, depende principalmente de lo que es permitido que suceda por las restricciones externas impuestas sobre el músculo durante la contracción. Las células musculares en la pared ventricular operan bajo diferentes restricciones durante distintas fases de cada ciclo cardiaco, y pasan por contracciones tanto isométrica como isotónica. CONTRACCIONES ISOMÉTRICAS: RELACIONES DE LONGITUDTENSIÓN Recuerde que la fuerza contráctil isométrica máxima del músculo cardiaco está fuertemente influida por la longitud inicial del músculo (figura 23-5). Sarcómero 2 1 MECÁNICA DE LA CÉLULA DE MÚSCULO CARDIACO 3 4 5 En reposo En contracción En reposo En reposo Tensión muscular En contracción 6 4 2 Tensión activa 5 5 1 6 En contracción 1 3 3 Tensión en reposo Estímulo Tiempo Tensión muscular 6 Tensión isométrica máxima 4 5 Tensión activa 2 Tensión en reposo 3 1 Lmáx Longitud del músculo FIGURA 235 Contracciones isométricas y el efecto de la longitud del músculo sobre la tensión en reposo y el desarrollo de tensión activa. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) CAPÍTULO 23 Células de músculo cardiaco El panel superior muestra el arreglo experimental para medir la fuerza muscular en reposo y durante contracción, a tres longitudes diferentes. El panel de en medio muestra registros de tiempo de tensiones musculares registradas a cada una de las tres longitudes en respuesta a un estímulo externo, y el panel inferior muestra un gráfico de los resultados de tensión en reposo y máxima, graficados contra la longitud del músculo. La influencia dependiente de la longitud sobre la tensión en reposo del músculo cardiaco es representada por la curva inferior en el gráfico de la figura 23-5. Cuando un músculo es estimulado para que se contraiga mientras su longitud se mantiene constante (es decir, contracción isométrica) desarrolla tensión activa. La tensión total ejercida por un músculo durante la contracción es la suma de las tensiones activa y en reposo, y es representada por la curva superior en la figura 23-5. Se muestra que el desarrollo de tensión activa es máximo a alguna longitud intermedia denominada Lmáx. En circunstancias normales, el músculo cardiaco opera a longitudes bastante por debajo de la Lmáx, de modo que aumentar la longitud muscular incrementa la tensión desarrollada durante una contracción isométrica. Los mecanismos involucrados en la relación entre la longitud del músculo cardiaco y la tensión desarrollada se comentan en el capítulo 10. El punto importante es que la dependencia de desarrollo de tensión activa y acortamiento sobre la longitud del músculo es una propiedad fundamental del músculo cardiaco que tiene efectos en extremo poderosos sobre la función del corazón. Contracción isotónica CONTRACCIONES ISOTÓNICA Y CON POSCARGA Durante una contracción isotónica (“carga fija”), el músculo se acorta contra una carga constante (figura 23-6). Cuando se suspende un peso de 1 g desde un músculo en reposo, dará lugar a alguna longitud del músculo en reposo específica, determinada por la curva de longitud-tensión en reposo del músculo. Si el músculo fuera a contraerse de manera isométrica a esta longitud, sería capaz de generar una cierta cantidad de tensión, por ejemplo, 4.5 g según lo indica la línea discontinua en el gráfico de la figura 23-6, sin embargo, no se generará una tensión contráctil de 4.5 g cuando se levanta un peso de 1 g. Cuando un músculo tiene potencial contráctil en exceso de la tensión que está desarrollando en realidad, se acorta, así, en una contracción isotónica, la longitud del músculo disminuye a tensión constante, como lo ilustra la flecha horizontal desde el punto 1 hacia el punto 3 en la figura 23-6. No obstante, a medida que el músculo se acorta, su potencial contráctil disminuye de manera inherente, como lo indica la pendiente hacia abajo de la curva de tensión isométrica máxima en la figura 23-6. Existe alguna longitud corta a la cual el músculo es capaz de generar sólo 1 g de tensión, y cuando se alcanza esta longitud, el acortamiento debe cesar. De este modo, la curva en el diagrama de longitud-tensión del músculo cardiaco que indica cuánta tensión isométrica puede desarrollar un Contracción con poscarga Isométrica Isotónica 3 2 1 5 1g 1 1g 1g 4 2g de poscarga 1g En reposo 1g 1g 3g de carga total 2g 2g 2g En contracción En reposo 5 Tensión muscular (g) 219 En contracción Tensión isométrica máxima 4 5 3 4 2 1 Tensión en reposo 3 2 1 Longitud del músculo FIGURA 236 Relación de las contracciones isotónica y con poscarga con el diagrama de longitud-tensión del músculo cardiaco (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) SECCIÓN V Fisiología cardiovascular CONTRACTILIDAD DEL MÚSCULO CARDIACO Varios factores además de la longitud inicial del músculo pueden afectar el potencial generador de tensión del músculo cardiaco. Se dice que cualquier intervención que aumente la tensión isométrica máxima que un músculo puede desarrollar a una longitud fija, aumenta la contractilidad del músculo cardiaco. Se dice que ese agente tiene un efecto inotrópico positivo sobre el corazón. El regulador fisiológico más importante de la contractilidad del músculo cardiaco es la norepinefrina. Cuando nervios simpáticos liberan norepinefrina sobre células de músculo cardiaco, no sólo tiene el efecto cronotrópico sobre la frecuencia cardiaca ya comentada, sino también un efecto inotrópico positivo pronunciado que hace que las células del músculo cardiaco se contraigan con mayor rapidez y energía. El efecto positivo de la norepinefrina sobre el potencial generador de tensión isométrica se ilustra en la figura 23-7A. En presencia de norepinefrina, el músculo cardiaco desarrollará, a cada longitud, más tensión isométrica, lo que aumenta la curva de tensión isométrica máxima en el gráfico de longitud-tensión del músculo cardiaco. La norepinefrina aumenta la contractilidad del músculo cardiaco porque mejora la fuerza de la contracción muscular aun cuando la A Contracción isométrica n Co 5 NE Tensión isométrica máxima 3 NE 4 Si n Tensión muscular (g) 2 1 o Con si n NE Tensión en reposo Longitud del músculo Contracción con poscarga B 5 4 3 n Co S músculo a diversas longitudes también establece el límite sobre qué tan lejos puede proceder el acortamiento muscular con diferentes cargas. La figura 23-6 también muestra una contracción isotónica con poscarga, en la cual la carga sobre el músculo en reposo (la precarga) y la carga sobre el músculo durante la contracción (la carga total) son diferentes. En el ejemplo de la figura 23-6, la precarga es igual a 1 g, y puesto que durante la contracción se agrega un peso de 2 g adicional (la poscarga), la carga total es igual a 3 g. Puesto que la precarga determina la longitud del músculo en reposo, las dos contracciones isotónicas mostradas en la figura 23-6 empiezan desde la misma longitud, empero, debido al arreglo de carga diferente, el músculo con poscarga debe aumentar su tensión total a 3 g antes de que pueda acortarse. Esta tensión inicial se desarrollará de manera isométrica, y puede representarse como en dirección del punto 1 al punto 4 en el diagrama de longitud-tensión, una vez que el músculo genera suficiente tensión para igualar la carga total, su salida de tensión es fijada en 3 g, y ahora se acortará de manera isotónica porque su potencial contráctil aún excede su salida de tensión. Este acortamiento isotónico es representado como un movimiento horizontal en el diagrama de longitud-tensión a lo largo de la línea desde el punto 4 hacia el punto 5. Como en cualquier contracción isotónica, el acortamiento debe cesar cuando el potencial de producción de tensión del músculo es disminuido lo suficiente por el cambio de longitud para que sea igual a la carga sobre el músculo. Note que el músculo con poscarga se acorta menos que el músculo sin poscarga aun cuando ambos músculos empezaron a contraerse a la misma longitud inicial, los aumentos de la poscarga disminuirán más el acortamiento del músculo. Los factores que afectan la magnitud del acortamiento del músculo cardiaco durante una contracción con poscarga despiertan interés especial porque el volumen sistólico está determinado por qué tanto se acorta el músculo cardiaco en estas condiciones. Tensión muscular (g) 220 Tensión isométrica máxima NE in NE Más acortamiento con NE 2 1 o Con sin Tensión en reposo NE Longitud del músculo FIGURA 237 Efecto de la norepinefrina (NE) sobre el generador de tensión isométrica (A) y con poscarga (B) contracciones de músculo cardiaco. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) longitud es constante. Los cambios de la contractilidad y la longitud inicial pueden ocurrir de manera simultánea pero, por definición, un cambio de la contractilidad debe comprender un cambio desde una curva de longitud-tensión isométrica máxima hacia otra. En la figura 23-7B se muestra cómo la elevación de la curva de longitud-tensión isométrica máxima con norepinefrina incrementa la cantidad de acortamiento en contracciones de músculo cardiaco con poscarga. Con precarga y carga total constantes, ocurre más acortamiento en presencia de norepinefrina que en su ausencia, esto se debe a que cuando la contractilidad es aumentada, el potencial generador de tensión es igual a la carga total, a una longitud más corta del músculo. Advierta que la norepinefrina carece de efecto sobre la relación longitud-tensión en reposo del músculo cardiaco, es decir, causa acortamiento aumentado al cambiar la longitud final del músculo, pero no la inicial, asociada con contracciones con poscarga. El mecanismo celular del efecto de la norepinefrina sobre la contractilidad está mediado por su interacción con un receptor β1-adrenérgico. La vía de emisión de señales involucra una activación de la proteína Gs-cAMP-proteína cinasa A, que a continuación fosforila el canal de Ca2+, lo que incrementa la corriente de calcio hacia adentro durante la meseta del potencial de acción. Este aumento del flujo de entrada de calcio no sólo contribuye a la magnitud del CAPÍTULO 23 Células de músculo cardiaco incremento del Ca2+ intracelular para un latido dado, sino que también carga las reservas de calcio internas, lo que permite que se libere más durante despolarizaciones subsiguientes; este incremento del Ca2+ libre durante la activación permite que se formen más puentes, aumenta la velocidad de recambio de puentes, y permite que se desarrolle más tensión a un ritmo más rápido. Dado que la norepinefrina también causa fosforilación de la proteína reguladora, fosfolambán, sobre la bomba Ca2+-ATPasa del retículo sarcoplasmático, la tasa de reatrapamiento de calcio hacia el SR es mejorada, y la tasa de relajación también es aumentada. Esto se llama un efecto lusitrópico positivo. Además de reatrapamiento más rápido de calcio por el SR, también hay un decremento (inducido por norepinefrina) de la duración del potencial de acción, este efecto se logra mediante una alteración del canal de potasio, que ocurre en respuesta al [Ca2+] intracelular elevado que aumenta la permeabilidad al potasio, termina la fase de meseta del potencial de acción, y contribuye a la relajación temprana. (Ese acortamiento del intervalo sistólico es útil en presencia de frecuencias cardiacas altas que por lo demás podrían comprometer de manera importante el tiempo de llenado diastólico.) Se ha mostrado que la actividad parasimpática aumentada tiene un efecto inotrópico negativo pequeño sobre el corazón. En las aurículas, donde este efecto es más pronunciado, se cree que el efecto inotrópico negativo se debe a un decremento de la duración del potencial de acción, y de la cantidad de Ca2+ que entra a la célula durante el potencial de acción. Los cambios de la frecuencia cardiaca también influyen sobre la contractilidad cardiaca. Recuérdese que una pequeña cantidad del Ca2+ extracelular entra a la célula durante la fase de meseta de cada potencial de acción. A medida que aumenta la frecuencia cardiaca, entra más Ca2+ a las células por cada minuto, hay una acumulación de Ca2+ intracelular, y se libera una mayor cantidad de Ca2+ hacia el sarcoplasma con cada potencial de acción. De este modo, un incremento repentino de la frecuencia de latidos va seguido por un aumento progresivo de la fuerza contráctil a una meseta más alta (la relación de fuerza-frecuencia). RELACIÓN DE LA MECÁNICA DE LAS CÉLULAS DEL MÚSCULO CARDIACO CON LA FUNCIÓN VENTRICULAR Ciertos factores geométricos dictan cómo las relaciones de longitudtensión de las fibras de músculo cardiaco en la pared ventricular determinan las relaciones de volumen y presión de la cámara ventricular. Las relaciones reales son complejas porque la forma del ventrículo es compleja y a menudo se modela como un cilindro o una esfera, aunque su forma real yace en algún sitio entre ambos. Puesto que las células de músculo cardiaco están orientadas de manera circunferencial en la pared ventricular, uno u otro modelo puede usarse para ilustrar tres puntos funcionales importantes: 1. Un aumento del volumen ventricular causa un incremento de la circunferencia ventricular y, por ende, un aumento de la longitud de las células de músculo cardiaco individuales; así, la magnitud de llenado diastólico del ventrículo determina la “precarga”. 221 2. A cualquier volumen ventricular dado, un incremento de la tensión de células de músculo cardiaco individuales en la pared causa un aumento de la presión intraventricular. 3. Conforme el volumen ventricular disminuye (esto es, a medida que se reduce el radio ventricular), las células musculares en las paredes ventriculares requieren una menor fuerza total (colectiva) para producir cualquier presión intraventricular dada. El último punto es un reflejo de la ley de Laplace que establece la relación física que debe haber entre la tensión total de pared y la presión interna en cualquier vaso hueco con paredes contenedoras circulares. Independientemente de si el ventrículo se contempla como un cilindro hueco o una esfera hueca, la ley de Laplace dice que la tensión de pared total (T) depende tanto de la presión intraventricular (P) como de su radio interno (r) como sigue: T = P × r. Una implicación de la ley de Laplace es que las células musculares en la pared ventricular tienen un trabajo un poco más fácil de producir presión interna al final de la sístole (cuando el radio es pequeño) que al principio de la sístole (cuando el radio es grande). Lo que es más importante, la ley de Laplace tiene gran importancia clínica en algunas situaciones patológicas. CORRELACIÓN CLÍNICA Un anciano es llevado a la sala de urgencias por su hija, ella reporta que el paciente a últimas fechas se quejó de debilidad intensa, fatiga, algo de mareo, y que parece estar un poco desorientado, este estado apareció sólo unos días antes, y no parece estar mejorando ni empeorando; por lo demás el paciente ha estado sano y gusta del baile tradicional. El paciente está alerta y tiene capacidad de respuesta, pero está muy débil y pálido, la presión arterial en el momento de la admisión es de 100/60 mmHg, y la frecuencia cardiaca es de 41 latidos/min. Un electrocardiograma obtenido en el momento de la admisión verifica la bradicardia (frecuencia cardiaca lenta) con una frecuencia ventricular de 41 latidos/min y una frecuencia auricular de 95 latidos/min. No hay signos en el ECG de isquemia cardiaca ni de infarto de miocardio, pero la aparición de ondas P es muy rápida y no se correlaciona con aparición lenta de ondas QRS. Este paciente tiene un bloqueo cardiaco de tercer grado (total) del nodo AV en el cual los potenciales de acción que se originan en el nodo SA no se conducen a través del nodo AV hacia los ventrículos. Los ventrículos están siendo conducidos a un ritmo de “escape” establecido por un marcapaso ubicado por debajo del nodo AV que se activa a una tasa, que es significativamente más lenta que la de las células ganglionares SA marcapaso. Las aurículas están latiendo mucho más rápido debido a activación simpática (desencadenada por la presión arterial baja), pero las señales no son conducidas a través del bloqueo del nodo AV. Los síntomas de debilidad, fatiga, mareo y desorientación son un resultado de la presión arterial baja que, a su vez, depende de la frecuencia ventricular baja y gasto cardiaco reducido. Las causas son toxicidades farmacológicas (p. ej., bloqueadores β-adrenérgicos, bloqueadores de los canales de calcio), alteraciones metabólicas (p. ej., hiperpotasemia), infartos de pared anterior de miocardio (esto es, isquemia sep- 222 SECCIÓN V Fisiología cardiovascular tal), y cardiomiopatía (p. ej., por una infección viral). Se efectuarán pruebas diagnósticas para determinar cuál podría ser la causa de su afección. El tratamiento de este paciente comprenderá implantación de un marcapaso cardiaco; éste es una pila implantada en el tejido subcutáneo que envía estímulos repetitivos a través de electrodos colocados tanto en la aurícula como en el ventrículo derechos a través de la vena cava. Este marcapaso de cámara doble, sincronizará la frecuencia de latidos auriculares y ventriculares a una frecuencia y ritmo más normales, y quizá restituirá el estado circulatorio y los niveles de actividad del paciente de regreso a lo normal. RESUMEN DEL CAPÍTULO ■ ■ ■ ■ ■ ■ ■ ■ Los potenciales de membrana de miocitos cardiacos son un resultado de la permeabilidad relativa de la membrana a diversos iones y sus diferencias de concentración a través de la membrana. Los potenciales de acción de miocitos cardiacos son un resultado de cambios de la permeabilidad de membrana a diversos iones. Los potenciales de acción de miocitos cardiacos tienen fases de meseta largas que generan periodos refractarios largos e impiden contracciones sumadas o tetánicas. Los potenciales de acción son generados de manera espontánea por células marcapaso en el nodo SA, y son conducidos de una célula a otra por medio de uniones intercelulares comunicantes (conexiones comunicantes) en todo el corazón. La tasa de despolarización diastólica espontánea de las células del nodo SA (y por tanto el ritmo cardiaco) está modulada por el sistema nervioso autónomo. La excitación del miocito cardiaco inicia la contracción al aumentar la concentración de calcio citosólico que activa el aparato contráctil. La respuesta mecánica del miocito depende de la precarga (determinada por la longitud en reposo inicial), la poscarga (determinada por la tensión que debe desarrollarse), y la contractilidad (el grado de activación del aparato contráctil dependiente de la cantidad de calcio liberada en el momento de la activación). Las relaciones longitud-tensión del miocito cardiaco se correlacionan con cambios del volumen y la presión en el ventrículo intacto. PREGUNTAS DE ESTUDIO 1. ¿Un fármaco que promueve la activación temprana del canal de K+ “rectificador tardío” (Ik) en el músculo cardiaco hará cuál de los siguientes? A) el potencial de reposo estará aumentado (hiperpolarizado) B) la duración del potencial de acción estará disminuida C) la amplitud máxima del potencial de acción estará disminuida D) la velocidad de conducción del potencial de acción estará aumentada E) el periodo refractario absoluto estará prolongado 2. La velocidad de conducción del potencial de acción en el tejido de músculo cardiaco está influida por todos los siguientes, excepto: A) el diámetro de la célula. B) el potencial de membrana en reposo. C) la concentración extracelular de potasio. D) la tasa de aumento (fase 0) del potencial de acción. E) la duración de la fase de meseta (fase 2) del potencial de acción. 3. La ruta primaria de eliminación del [Ca2+] del sarcoplasma durante la relajación de una célula de músculo cardiaco es mediante: A) transporte activo hacia afuera de la célula. B) intercambio pasivo con sodio extracelular. C) transporte activo hacia el retículo sarcoplasmático. D) atrapamiento de calcio por troponina en los miofilamentos. E) movimiento pasivo hacia afuera de la célula por medio de canales de calcio tipo L. 4. ¿Una estrategia terapéutica para mejorar la cantidad de acortamiento activo de músculo cardiaco podría incluir cuál de las que siguen? A) disminuir la precarga B) disminuir la poscarga C) disminuir la contractilidad D) administrar un agente cronotrópico negativo E) administrar un agente inotrópico negativo 5. Su paciente accidentalmente ha recibido una inyección en bolo grande de cloruro de potasio y muere, ¿por qué? A) el músculo cardiaco es despolarizado y el corazón se ha detenido en diástole B) el músculo cardiaco es despolarizado y el corazón se ha detenido en sístole C) el músculo cardiaco es hiperpolarizado y el corazón se ha detenido en diástole D) la conducción del potencial de acción está acelerada y el corazón está fibrilando E) las uniones intercelulares comunicantes entre las células de músculo cardiaco están alteradas La bomba cardiaca C A P Í T U L O 24 Lois Jane Heller y David E. Mohrman O B J E T I V O S ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ Correlacionar los eventos electrocardiográficos con los mecánicos durante el ciclo cardiaco. Listar las principales fases del ciclo cardiaco delineadas por la abertura y el cierre valvulares. Describir los cambios de presión y de volumen en aurículas, ventrículos y aorta durante cada fase del ciclo cardiaco. Definir y enunciar los valores normales para 1) los volúmenes ventricular al final de la diástole, al final de la sístole, sistólico, las presiones diastólica y sistólica máxima, y 2) las presiones diastólica aórtica, sistólica y del pulso. Mencionar similitudes y diferencias entre eventos mecánicos en las bombas cardiacas izquierda y derecha. Describir el origen de los ruidos cardiacos. Hacer un diagrama de la relación entre la presión y el volumen ventriculares izquierdos durante el ciclo cardiaco. Definir gasto e índice cardiacos. Mencionar la relación entre gasto cardiaco, frecuencia cardiaca y volumen sistólico. Identificar los principales determinantes del volumen sistólico. Enunciar la ley de Starling del corazón. Predecir el efecto de la precarga ventricular alterada sobre el volumen sistólico y la relación presión-volumen ventricular. Predecir el efecto de la poscarga ventricular alterada sobre el volumen sistólico y la relación presión-volumen ventricular. Predecir el efecto de la contractilidad (estado inotrópico) ventricular alterada sobre el volumen sistólico y la relación de presión-volumen ventricular. Dibujar una familia de curvas de función cardiaca describiendo la relación entre la presión de llenado y el gasto cardiaco en diversas magnitudes de tono simpático. Resumir el efecto de la estimulación neural simpática sobre la función cardiaca. Listar los determinantes del consumo miocárdico de oxígeno. 223 224 SECCIÓN V Fisiología cardiovascular En este capítulo se describen: 1) las características mecánicas básicas de la bomba cardiaca, 2) los factores que influyen sobre el gasto cardiaco, o lo regulan, o ambos y 3) las fuentes de energía y los costos de energía requeridos para la actividad miocárdica. CICLO CARDIACO BOMBA IZQUIERDA La función mecánica del corazón puede describirse por los cambios de presión, volumen y flujo que ocurren dentro de él durante un ciclo cardiaco; el cual se define como una secuencia completa de contracción y relajación. Los eventos mecánicos normales de un ciclo de la bomba cardiaca izquierda se correlacionan en la figura 24-1. Esta importante figura resume mucha información y debe estudiarse de forma cuidadosa. DIÁSTOLE VENTRICULAR La fase diastólica del ciclo cardiaco empieza con la abertura de las válvulas auriculoventriculares (AV). (A menos que se indique lo contrario, sístole y diástole denotan las fases de operación ventricular.) La válvula mitral se abre cuando la presión ventricular izquierda disminuye por debajo de la presión auricular izquierda y empieza el periodo de llenado del ventrículo (figura 24-1). La sangre que se había acumulado previamente en la aurícula detrás de la válvula mitral cerrada se vacía con rapidez hacia el ventrículo, y esto causa un decremento inicial de la presión auricular. Más tarde, las presiones en ambas cámaras aumentan lentamente juntas conforme la aurícula y el ventrículo se siguen llenando de manera pasiva con sangre que regresa al corazón a través de las venas. La contracción auricular inicia cerca del final de la diástole ventricular por la despolarización de las células musculares auriculares, que causa la onda P del electrocardiograma. A medida que las células A B Fase del ciclo cardiaco C D E R T P Derivación II del electrocardiograma Auricular Contracción muscular Ventricular Incisura Presión sistólica Presión aórtica 120 80 Presión del pulso Presión diastólica Presión (mmHg) Presión en el ventrículo izquierdo 40 Presión en la aurícula izquierda 0 Cerrada Válvula aórtica Válvula mitral S3 Ruidos cardiacos 120 Abierta S4 Volumen al final de la diástole Volumen del ventrículo izquierdo (ml)) Cerrada Abierta Cerrada S1 S2 Volumen sistólico Volumen al final de la sístole 60 FIGURA 241 Ciclo cardiaco —hemicardio izquierdo. Fases del ciclo cardiaco: A) diástole; B) sístole que se divide en tres periodos; C) contracción isovolumétrica; D) eyección y E) relajación isovolumétrica. (Modificada con autorización de Mohrman DE, Flujo de salida Flujo aórtico 0 0 0.2 0.4 Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) Tiempo (s) Flujo de entrada 0.6 0.8 Abierta S3 CAPÍTULO 24 La bomba cardiaca musculares auriculares desarrollan tensión y se acortan, la presión auricular se eleva, y una cantidad adicional de sangre es forzada hacia el ventrículo. A frecuencias cardiacas normales, la contracción auricular no es esencial para el llenado ventricular adecuado. Esto es evidente en la figura 24-1, que muestra que el ventrículo casi ha alcanzado su máximo (volumen al final de la diástole) antes de que empiece la contracción auricular. La contracción auricular desempeña una función cada vez más importante en el llenado ventricular conforme la frecuencia cardiaca aumenta porque el intervalo entre latidos para el llenado pasivo se hace progresivamente más corto. Note que durante toda la diástole, las presiones auricular y ventricular son casi iguales; esto es debido a que una válvula mitral abierta normal ofrece muy poca resistencia al flujo y, así, sólo se necesita una diferencia de presión auricular-ventricular muy pequeña para producir llenado ventricular. SÍSTOLE VENTRICULAR La sístole ventricular empieza cuando el potencial de acción pasa por el nodo AV y se propaga el músculo ventricular, lo que da lugar al complejo QRS del electrocardiograma. La contracción de las células musculares ventriculares hace que la presión intraventricular aumente por arriba de la que hay en la aurícula, lo que produce cierre repentino de la válvula AV. La presión en el ventrículo izquierdo se sigue incrementando mucho a medida que la contracción ventricular se intensifica. Cuando la presión en el ventrículo izquierdo excede la que hay en la aorta, la válvula aórtica se abre. El periodo entre el cierre de la válvula mitral y la abertura de la válvula aórtica se denomina la fase de contracción isovolumétrica porque, durante este intervalo, el ventrículo es una cámara cerrada con un volumen fijo. La eyección ventricular empieza con la abertura de la válvula aórtica. Al principio de la eyección entra sangre a la aorta con rapidez y hace que la presión aumente en ese sitio. La presión se eleva de manera simultánea tanto en el ventrículo como en la aorta conforme las células musculares ventriculares se siguen contrayendo al principio de la sístole; este intervalo a menudo se llama el periodo de eyección rápida. Las presiones en el ventrículo izquierdo y la aorta finalmente alcanzan un máximo llamado presión sistólica máxima. En este punto, la fuerza de la contracción del músculo ventricular empieza a bajar. El acortamiento del músculo y la eyección continúan, pero a un ritmo reducido. La presión aórtica empieza a disminuir porque la sangre está saliendo de la aorta y de las arterias de gran calibre con una rapidez mayor que la de la entrada de sangre desde el ventrículo izquierdo. Durante toda la eyección, hay diferencias de presión muy pequeñas entre el ventrículo izquierdo y la aorta porque el orificio de la válvula aórtica es tan grande que ofrece muy poca resistencia al flujo. Al final, la fuerza de la contracción ventricular se reduce hasta el grado en que la presión intraventricular disminuye por debajo de la presión aórtica. Esto causa cierre repentino de la válvula aórtica. Aparece un descenso, llamado la incisura o muesca dicrótica, en el trazo de presión aórtica porque un volumen pequeño de sangre aórtica debe fluir en dirección retrógrada para llenar las hojuelas de la válvula aórtica a medida que se cierran. Después del cierre de la válvula aórtica, la presión intraventricular disminuye con rapidez conforme el músculo ventricular se relaja. Durante un intervalo breve, llamado la fase de relajación isovolumétrica, la válvula 225 mitral también se cierra; por último, la presión intraventricular disminuye por debajo de la presión auricular, la válvula AV se abre, y empieza un nuevo ciclo cardiaco. Note que la presión auricular crece de manera progresiva durante la sístole ventricular porque la sangre sigue regresando al corazón y llena la aurícula. La presión auricular aumentada al final de la sístole promueve llenado ventricular rápido una vez que la válvula AV se abre para empezar el siguiente ciclo cardiaco. El ventrículo ha alcanzado su mínimo (volumen al final de la sístole) en el momento del cierre de la válvula aórtica. La cantidad de sangre eyectada desde el ventrículo durante un latido único, el volumen sistólico, es igual al volumen ventricular al final de la diástole menos el volumen ventricular al final de la sístole. Durante la fase temprana más rápida de eyección sistólica, la aorta se distiende porque el hemicardio izquierdo está introduciendo más sangre en ella que la que está saliendo hacia los órganos sistémicos; esa distensión es causada por la presión creciente dentro de la aorta. Durante la fase de debilitación más tardía de la eyección cardiaca, sucede lo contrario. El resultado general es que la presión aórtica alcanza un valor máximo (presión sistólica) cerca de la mitad de la sístole ventricular. Durante la diástole, la presión arterial es mantenida por el retroceso elástico de las paredes de la aorta y otras arterias de gran calibre. Sin embargo, la presión aórtica disminuye de manera gradual durante la diástole a medida que la aorta lleva sangre a los lechos vasculares sistémicos. La presión aórtica más baja, que se alcanza al final de la diástole, se llama presión diastólica. La diferencia entre la presión diastólica y la sistólica máxima en la aorta se llama presión de pulso arterial. Los valores típicos para las presiones sistólica y diastólica en la aorta son de 120 y 80 mmHg, respectivamente, con una presión de pulso de 40 mmHg. A una frecuencia cardiaca en reposo normal de alrededor de 70 latidos/min, el corazón pasa alrededor de dos terceras partes del ciclo cardiaco en diástole, y una tercera parte en sístole. Cuando aumenta la frecuencia cardiaca, los intervalos tanto diastólico como sistólico se hacen más breves. Las duraciones de potenciales de acción se acortan, y la velocidad de conducción se incrementa. Las tasas de contracción y relajación también se elevan. Este acortamiento del intervalo sistólico tiende a disminuir los efectos adversos potenciales de incrementos de la frecuencia cardiaca sobre el tiempo de llenado diastólico. BOMBA DERECHA Dado que en todo el corazón sólo hay un sistema de excitación eléctrica, eventos mecánicos similares ocurren casi de manera simultánea tanto en el hemicardio izquierdo como en el derecho. Ambos ventrículos tienen periodos sistólico y diastólico sincrónicos, y las válvulas del hemicardio derecho, por lo normal se abren y se cierran casi al unísono. Puesto que los dos lados del corazón están dispuestos en serie en la circulación, deben bombear la misma cantidad de sangre y, por consiguiente, deben tener el mismo gasto cardiaco. La principal diferencia entre las bombas derecha e izquierda ocurre en la magnitud de la presión sistólica máxima. Las presiones desarrolladas por el hemicardio derecho (figura 24-2) son de forma considerable más bajas que las que hay en el hemicardio izquierdo (figura 24-1). Esto se debe a que los vasos pulmonares ofrecen resistencia considerablemente menor al flujo sanguíneo que la ofrecida 226 SECCIÓN V Fisiología cardiovascular Derivación II del electrocardiograma Presión en la arteria pulmonar Presión (mmHg) 25 20 15 10 5 Presión en la aurícula derecha a c v Presión en el ventrículo derecho 0 0.0 0.2 0.4 0.6 Tiempo (s) 0.8 FIGURA 242 Ciclo cardiaco —hemicardio derecho. 1.0 (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) de manera colectiva por los órganos sistémicos. Por ende, se requiere menos presión arterial para impulsar el gasto cardiaco a través de los pulmones que a través de los órganos sistémicos. Las presiones sistólica y diastólica en la arteria pulmonar típicas son de 25 y 8 mmHg, respectivamente. Las pulsaciones de presión que ocurren en la aurícula derecha son transmitidas de modo retrógrado hacia las venas de gran calibre cerca del corazón. Estas pulsaciones, que se muestran en el trazo de presión auricular de la figura 24-2, pueden visualizarse en el cuello sobre las venas yugulares en un individuo en decúbito. Se denominan en conjunto el pulso venoso yugular, y pueden proporcionar información útil en clínica acerca del corazón. La contracción auricular produce el primer máximo de presión, llamado onda a. La onda c, que le sigue poco después, coincide con el inicio de la sístole ventricular, y es provocada por un abultamiento inicial de la válvula tricúspide hacia la aurícula derecha. La presión en la aurícula derecha disminuye después de la onda c debido a relajación auricular y un desplazamiento hacia abajo de la válvula tricúspide durante el vaciamiento ventricular. A continuación, la presión en la aurícula derecha empieza a aumentar hacia un tercer máximo, la onda v, conforme las venas centrales y la aurícula derecha se llenan detrás de una válvula tricúspide cerrada con sangre que regresa al corazón desde los órganos periféricos. Con la abertura de la válvula tricúspide en el momento de la conclusión de la sístole ventricular, la presión en la aurícula derecha vuelve a disminuir a medida que la sangre pasa hacia el ventrículo derecho relajado. Poco después, la presión en la aurícula derecha empieza a aumentar una vez más hacia la siguiente onda a conforme la sangre que está regresando llena las venas centrales, la aurícula derecha y el ventrículo derecho durante la diástole. RUIDOS CARDIACOS En la figura 24-1 se incluye un registro de los ruidos del corazón, que ocurren en el ciclo cardiaco. Estos ruidos por lo normal se escuchan mediante auscultación con un estetoscopio colocado sobre el tórax. El primer ruido cardiaco, S1, ocurre al principio de la sístole debido al cierre repentino de las válvulas AV, que produce vibraciones de las estructuras cardiacas y la sangre en las cámaras ventriculares. S1 puede auscultarse con mayor claridad al colocar el estetoscopio sobre el vértice del corazón. Note que este ruido ocurre de inmediato después del complejo QRS del electrocardiograma. El segundo ruido cardiaco, S2, surge por el cierre de las válvulas aórtica y pulmonar al principio del periodo de relajación isovolumétrica; este ruido se ausculta cerca del momento de la onda T en el electrocardiograma. La válvula pulmonar por lo general se cierra un poco después que la válvula aórtica. Dado que esta discrepancia es elevada durante la fase inspiratoria del ciclo respiratorio, la inspiración causa lo que se denomina desdoblamiento fisiológico del segundo ruido cardiaco. La discrepancia del cierre de la válvula durante la inspiración puede variar de 30 a 60 ms. Uno de los factores que lleva a eyección prolongada del ventrículo derecho durante la inspiración es que la presión intratorácica disminuida que acompaña a la inspiración aumenta de manera transitoria el retorno venoso y el llenado diastólico del hemicardio derecho. Por razones que se detallarán más adelante en este capítulo, este volumen de llenado extra será eyectado, pero se requiere un poco de tiempo extra. En circunstancias normales el tercer y cuarto ruidos cardiacos (figura 24-1), no están presentes; sin embargo, cuando lo están, junto con S1 y S2, producen lo que se llama ritmos de galope (que semejan el sonido de un caballo galopando). Cuando está presente, el tercer ruido cardiaco ocurre poco después de S2 durante el periodo de llenado ventricular pasivo rápido y, en combinación con los ruidos cardiacos S1 y S2, produce lo que se llama el ritmo de galope ventricular. Aunque S3 a veces puede detectarse en niños normales, se ausculta más comúnmente en pacientes con insuficiencia ventricular izquierda. El cuarto ruido cardiaco, que en ocasiones se ausculta poco antes de S1, se relaciona con la contracción auricular y llenado activo rápido del ventrículo. Así, la combinación de ruidos S1, S2 y S4 produce lo que se llama ritmo de galope auricular. La presencia de S4 a menudo indica rigidez diastólica ventricular aumentada, que puede ocurrir con varios estados morbosos cardiacos. RELACIONES DE PRESIÓNVOLUMEN Y LONGITUDTENSIÓN DEL CICLO CARDIACO La presión y el volumen intraventriculares están íntimamente enlazados a la tensión y longitud de las células de músculo cardiaco en la pared ventricular. En la figura 24-3A y B. se observa la correspondencia entre un asa de presión-volumen ventricular y un asa de longitud-tensión del músculo cardiaco durante un ciclo cardiaco. Está claro que la conducta de longitud-tensión del músculo cardiaco es la base subyacente para la función ventricular. Cada fase importante del ciclo cardiaco ventricular tiene una fase correspondiente de cambio de longitud y tensión del músculo cardiaco. Por ejemplo, durante el llenado ventricular diastólico, el incremento progresivo de la presión ventricular distiende el músculo cardiaco en reposo a longitudes mayores a lo largo de su curva de longitud-tensión en reposo, y causa un aumento correspondiente de la tensión del músculo. La presión ventricular al final de la diástole se denomina precarga ventricular porque establece el volumen ventricular al final de la diástole y, por ende, la longitud en reposo de las fibras del músculo cardiaco al final de la diástole. CAPÍTULO 24 La bomba cardiaca 227 A Presión intraventricular (mmHg) 120 Eyección Alcanza el volumen al final de la sístole La válvula aórtica se abre 80 Sístole Relajación isovolumétrica Contracción isovolumétrica Se abre la válvula mitral Llenado diastólico Alcanza el volumen al final de la diástole 130 Volumen sistólico Volumen intraventricular (ml) 60 B Tensión del músculo Acortamiento Alcanza la longitud al final de la sístole Activa Desarrollo de tensión isométrica Relajación isométrica Distensión pasiva Alcanza la longitud al final de la diástole FIGURA 243 Ciclo de presión-volumen ventricular A), y ciclo de longitud-tensión correspondiente del músculo cardiaco B). (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Longitud del músculo Al inicio de la sístole, las células de músculo ventricular desarrollan tensión de manera isométrica (durante la fase de contracción isovolumétrica) y la presión intraventricular se incrementa en consecuencia. Después de que la presión intraventricular aumenta lo suficiente como para abrir la válvula de salida, la eyección ventricular empieza como una consecuencia del acortamiento del músculo ventricular. La presión arterial sistémica a menudo se denomina poscarga ventricular porque determina la tensión que las fibras de músculo cardiaco deben desarrollar antes de que puedan acortarse. Cabe hacer notar que otros factores que influyen sobre la tensión de pared real requerida para eyectar sangre desde el ventrículo (como el volumen al final de la diástole, la velocidad de contracción, la viscosidad de la sangre) contribuyen a la poscarga ventricular, pero los autores optan por hacer caso omiso de ellos en este punto. Durante la eyección cardiaca, el músculo cardiaco está generando de forma simultánea tensión y acortamiento activos (esto es, una contracción isotónica con poscarga). El volumen sistólico está determinado por qué tanto las células de músculo ventricular son capaces de acortarse durante la contracción. Esto, como se comentó, depende de la relación longitud-tensión de las células de músculo cardiaco y la carga contra la cual se están acortando. Una vez que el acortamiento cesa y la válvula de salida se cierra, las células de músculo cardiaco se relajan de manera isométrica. La tensión de pared ven- Medical Books/McGraw-Hill, 2006.) tricular y la presión intraventricular disminuyen al unísono durante la relajación isovolumétrica. DETERMINANTES DEL GASTO CARDIACO El gasto cardiaco (litros de sangre bombeados por cada uno de los ventrículos por minuto) es una variable cardiovascular de importancia extrema que es ajustada continuamente de modo que el sistema cardiovascular opere para satisfacer las necesidades circulatorias del cuerpo momento a momento. Al pasar desde reposo hacia ejercicio extenuante, por ejemplo, el gasto cardiaco de una persona promedio aumentará desde casi 5.5 hasta quizá 15 l/min. El gasto cardiaco extra proporciona a los músculos esqueléticos que están haciendo ejercicio el aporte nutricional adicional necesario para sostener un índice metabólico aumentado. Para entender la respuesta del sistema cardiovascular no sólo al ejercicio, sino también a todas las otras demandas fisiológicas o patológicas impuestas sobre él, es necesario entender qué determina el gasto cardiaco y, por ende, qué lo controla. El gasto cardiaco es el producto de la frecuencia cardiaca y el volumen sistólico (CO = HR × SV) (capítulo 22). Por consiguiente, todos SECCIÓN V Fisiología cardiovascular los cambios del gasto cardiaco deben ser producidos por cambios de la frecuencia cardiaca, o del volumen sistólico o de ambos. Los factores que influyen sobre la frecuencia cardiaca lo hacen al alterar las características de despolarización diastólica de las células marcapasos (véase capítulo 23; figura 23-4). Recuérdese que las variaciones de la actividad de los nervios simpáticos y parasimpáticos que van a las células del nodo sinoauricular (SA) constituyen los reguladores más importantes de la frecuencia cardiaca. Los aumentos de la actividad simpática incrementan la frecuencia cardiaca, mientras que los de la actividad parasimpática la disminuyen. Estas aferencias neurales tienen efectos inmediatos (en cuestión de un latido) y, por ende, pueden causar ajustes muy rápidos del gasto cardiaco. A Tensión del músculo (g) 228 5 Tensión isométrica máxima 4 3 Más acortamiento 2 Tensión en reposo 1 Precarga de mayor magnitud Longitud del músculo INFLUENCIAS SOBRE EL VOLUMEN SISTÓLICO EFECTO DE CAMBIOS DE LA PRECARGA VENTRICULAR: LEY DE STARLING DEL CORAZÓN El volumen de sangre que el corazón eyecta con cada latido puede variar de modo significativo. Uno de los factores de mayor importancia de los cuales dependen estas variaciones del volumen sistólico es la magnitud del llenado cardiaco durante la diástole. Este concepto fue introducido en el capítulo 22 (véase figura 22-7), y se conoce como la ley de Starling del corazón. Para revisar (y para volver a recalcar su importancia), esta ley establece que, ante igualdad de otros factores, el volumen sistólico aumenta a medida que lo hace el llenado cardiaco. Como se muestra a continuación, este fenómeno se basa en las propiedades mecánicas intrínsecas del músculo miocárdico. En la figura 24-4A se ilustra cómo el aumento de la precarga del músculo incrementará la magnitud de acortamiento durante una contracción subsiguiente con una carga total fija. Recuérdese a partir de la naturaleza de la relación longitud-tensión en reposo, que la precarga aumentada necesariamente se acompaña de incremento de la longitud inicial de la fibra muscular. Cuando un músculo empieza desde una longitud mayor, tiene más distancia para acortarse antes de que llegue a la longitud a la cual su capacidad de generación de tensión ya no es mayor que la carga que está sobre éste (véase capítulo 23). Las células de músculo cardiaco muestran la misma conducta cuando están operando en la pared ventricular. Los incrementos de la precarga ventricular elevan casi por igual tanto el volumen al final de la diástole como el volumen sistólico (figura 24-4B). La relación precisa entre precarga cardiaca (presión de llenado cardiaco) y el volumen al final de la diástole tiene consecuencias fisiológicas y clínicas de importancia especial. Aunque la relación es un poco curvilínea, en especial a presiones de llenado muy altas, es casi lineal en el rango de operación normal del corazón. La pendiente baja de esta relación indica la increíble distensibilidad del ventrículo normal durante la diástole. (Por ejemplo, ¡un cambio de la presión de llenado de sólo 1 mmHg por lo normal cambiará el volumen al final de la diástole alrededor de 25 ml!) Una forma de insuficiencia cardiaca se llama insuficiencia diastólica, y se caracteriza por un ventrículo anormalmente rígido y una Presión del LV (mmHg) B 120 80 Más volumen sistólico 40 Precarga ventricular de mayor magnitud 120 60 Volumen del LV (ml) FIGURA 244 Efecto de cambios de la precarga sobre el acortamiento del músculo cardiaco durante contracciones con poscarga (A) y sobre el volumen sistólico ventricular (B). (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) alteración de la relación entre la presión de llenado cardiaco y el volumen al final de la diástole. En esta situación, el llenado diastólico es limitado, el volumen sistólico está reducido, el gasto cardiaco es insuficiente, y la función del sistema cardiovascular está comprometida. Debe observarse en la figura 24-4A que el incremento de la precarga aumenta la longitud inicial del músculo sin cambiar de manera importante la longitud final a la cual el músculo se acorta contra una carga total constante. Así, el aumento de la presión de llenado ventricular eleva el volumen sistólico principalmente al disminuir el volumen al final de la diástole. Esto no se acompaña de una alteración importante del volumen al final de la sístole (figura 24-4B). EFECTOS DE CAMBIOS DE LA POSCARGA VENTRICULAR En la figura 24-5A se muestra cómo la poscarga aumentada, a precarga constante, tiene un efecto negativo sobre el acortamiento del músculo cardiaco porque el músculo no puede acortarse más allá de la longitud a la cual su potencial generador de tensión isométrica máximo es igual a la carga total sobre él. Así, el acortamiento debe cesar a una longitud de músculo mayor cuando la poscarga es incrementada. CAPÍTULO 24 La bomba cardiaca abajo y está aplanada, de modo que el efecto de la poscarga sobre el volumen al final de la sístole está muy exagerado. Tensión muscular (g) A Tensión isométrica máxima 5 4 Menos acortamiento 3 2 1 Tensión en reposo Carga total de mayor magnitud Longitud del músculo B 120 Presión del LV (mmHg) 229 80 Volumen sistólico más bajo 40 Poscarga ventricular de mayor magnitud EFECTO DE CAMBIOS DE LA CONTRACTILIDAD DEL MÚSCULO CARDIACO Recuérdese que la activación del sistema nervioso simpático da lugar a liberación de norepinefrina a partir de nervios simpáticos cardiacos, lo cual aumenta la contractilidad de las células de músculo cardiaco individuales. Esto da lugar a un cambio hacia arriba de la curva de longitud-tensión isométrica máxima. Ese cambio dará por resultado un incremento del acortamiento de un músculo que se está contrayendo con precarga y carga total constantes (figura 24-6A). De este modo, la norepinefrina liberada por estimulación de nervios simpáticos incrementará el volumen sistólico ventricular al disminuir el volumen al final de la sístole sin influir de manera directa sobre el volumen al final de la diástole (figura 24-6B). Además de estos cambios de la magnitud del acortamiento de miocitos, un incremento de la contractilidad también causará un aumento de las tasas de desarrollo de tensión, y de acortamiento, de miocitos. Esto dará lugar a un alza de la tasa de desarrollo de presión isovolumétrica (dP/dt) y la tasa de eyección durante la sístole. 60 120 Volumen del LV (ml) Contracción con poscarga A de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) En circunstancias normales, la poscarga ventricular media es casi constante, porque los mecanismos de control cardiovascular descritos en el capítulo 29 mantienen la presión arterial media dentro de límites estrechos. En muchas situaciones patológicas, como hipertensión (presión arterial alta) y obstrucción de la válvula aórtica, la poscarga ventricular anormalmente alta ejerce una influencia adversa sobre la función ventricular. Cuando ocurre esto, el volumen sistólico puede estar disminuido como se muestra por los cambios en el asa de presión-volumen indicados mediante la línea discontinua en la figura 24-5B. En estas condiciones, nótese que el volumen sistólico está disminuido porque el volumen al final de la sístole está aumentado. La relación entre presión al final de la sístole y volumen al final de la sístole obtenida a una precarga constante, pero poscargas diferentes se indica mediante la línea discontinua en la figura 24-5B. En un corazón que funciona de forma normal, el efecto de cambios de la poscarga sobre el volumen al final de la sístole (y, por ende, el volumen sistólico) es bastante pequeño (alrededor de 0.5 ml/mmHg). Sin embargo, en la insuficiencia cardiaca sistólica, la línea de presión-volumen al final de la sístole está desviada hacia in Co NE Más acortamiento Tensión en reposo E 3 NE N 4 n S 5 Tensión isométrica máxima 2 n Co 1 in os Longitud del músculo B Presión del LV (mmHg) acortamiento del músculo cardiaco durante contracciones con poscarga A) y sobre el volumen sistólico ventricular B). La línea discontinua muestra cómo un incremento de la poscarga disminuye el volumen sistólico. La línea punteada muestra la relación entre la presión al final de la sístole y el volumen al final de la sístole obtenida a una precarga constante, pero a poscargas diferentes. (Modificada con autorización Tensión del músculo (g) FIGURA 245 Efectos de cambios de la poscarga sobre el Co nN E Sin NE 120 80 Volumen sistólico aumentado 40 60 120 Volumen del LV (ml) FIGURA 246 Efecto de la norepinefrina (NE) sobre las contracciones con poscarga sobre el músculo cardiaco (A) y sobre el volumen sistólico ventricular (B). (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGrawHill, 2006.) 230 SECCIÓN V Fisiología cardiovascular – MAGNITUD DE ACTIVIDAD DE NERVIO PARASIMPÁTICO CARDIACO Cronotrópica FRECUENCIA negativa CARDIACA + Cronotrópica positiva MAGNITUD DE ACTIVIDAD DE NERVIO SIMPÁTICO CARDIACO + GASTO CARDIACO + Contractilidad (inotrópica positiva) PRESIÓN ARTERIAL + FIGURA 247 Influencias sobre el gasto cardiaco. (Modificada con autorización de Mohrman DE, Poscarga – PRESIÓN DE LLENADO Precarga Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange En la figura 24-7 se resumen las principales influencias sobre el gasto cardiaco. La frecuencia cardiaca es controlada por influencias cronotrópicas sobre la actividad eléctrica espontánea de células del nodo SA. Los nervios parasimpáticos cardiacos tienen un efecto cronotrópico negativo, y los nervios simpáticos tienen un efecto cronotrópico positivo sobre el nodo SA. El volumen sistólico es controlado por influencias sobre el rendimiento contráctil del músculo cardiaco ventricular —en particular su grado de acortamiento en la situación con poscarga—. Las tres influencias distintas sobre el volumen sistólico son contractilidad, precarga y poscarga. La actividad aumentada de nervios simpáticos cardiacos tiende a incrementar el volumen sistólico al elevar la contractilidad del músculo cardiaco (un efecto inotrópico positivo). La presión arterial aumentada tiende a disminuir el volumen sistólico al incrementar la poscarga sobre las fibras de músculo cardiaco. La presión de llenado ventricular aumentada incrementa el volumen al final de la diástole, lo cual tiende a aumentar el volumen sistólico por medio de la ley de Starling del corazón. En este punto tiene importancia reconocer que tanto la frecuencia cardiaca como el volumen sistólico están sujetos a más de una influencia. Así, el hecho de que la contractilidad aumentada tiende a incrementar el volumen sistólico no debe considerarse que signifique que en el sistema cardiovascular intacto, el volumen sistólico siempre es alto cuando la contractilidad es alta. Después de pérdida de sangre causada por hemorragia, por ejemplo, el volumen sistólico puede ser bajo a pesar de una magnitud alta de actividad de nervios simpáticos y contractilidad aumentada. Puesto que la presión arterial es normal o baja después de hemorragia, el volumen sistólico bajo relacionado con pérdida grave de sangre debe ser (y lo es) el resultado de presión de llenado cardiaco baja. CURVAS DE FUNCIÓN CARDIACA Una manera muy útil de resumir las influencias sobre la función cardiaca y las interacciones entre ellas es mediante curvas de función cardiaca (figura 24-8). La presión de llenado cardiaco (“precarga cardiaca”) es graficada como la variable independiente en el eje horizontal en esta figura, y el gasto cardiaco como la variable dependiente en el eje vertical. Cada curva en esta figura muestra el efecto de cambios de la precarga cardiaca sobre el gasto cardiaco a una magnitud constante de actividad de nervios simpáticos cardiacos. Diferentes curvas se usan a diferentes niveles de actividad de nervios Magnitudes de actividad de nervios simpáticos cardiacos Muy aumentada 10.0 Gasto cardiaco (l/min) RESUMEN DE DETERMINANTES DEL GASTO CARDIACO + (ley de Starling) Medical Books/McGraw-Hill, 2006.) La insuficiencia sistólica se caracteriza por capacidad gravemente deprimida de las células de músculo cardiaco para producir tensión y acortarse. En estas circunstancias, la contracción sistólica es limitada, el volumen sistólico está reducido, el gasto cardiaco es insuficiente, y la función del sistema cardiovascular está comprometida. VOLUMEN SISTÓLICO Moderadamente aumentada 8.0 Normal C B 6.0 Disminuida A 4.0 2.0 0 2.0 4.0 6.0 Presión de llenado cardiaco (mmHg) FIGURA 248 Influencia de nervios simpáticos cardiacos sobre las curvas de función cardiaca. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) CAPÍTULO 24 La bomba cardiaca simpáticos cardiacos. De este modo, en la figura 24-8 se muestra cómo la presión de llenado cardiaco y la magnitud de actividad de nervios simpáticos cardiacos interactúan para determinar el gasto cardiaco. Cuando la presión de llenado cardiaco es de 2 mmHg y la actividad de los nervios simpáticos cardiacos es normal, el corazón operará en el punto A y tendrá un gasto cardiaco de 5 l/min. Cada curva única en la figura 24-8 muestra cómo el gasto cardiaco sería modificado por cambios de la presión de llenado cardiaco si la actividad de nervios simpáticos cardiacos se mantuviera a una magnitud fija. Por ejemplo, si la actividad de nervios simpáticos cardiacos permaneciera normal, aumentar la presión de llenado cardiaco de 2 a 4 mmHg haría que el corazón cambie su operación desde el punto A hacia el punto B en el diagrama de función cardiaca. En este caso, el gasto cardiaco aumentaría desde 5 hasta 7 l/min sólo como resultado de la presión de llenado aumentada (ley de Starling) por otro lado, si la presión de llenado cardiaco se fijara en 2 mmHg mientras la actividad de los nervios simpáticos cardiacos estuviera moderadamente aumentada desde lo normal, el corazón cambiaría desde operar en el punto A hacia operar en el punto C. De nuevo, el gasto cardiaco aumentaría desde 5 hasta 7 l/min. Sin embargo, en este caso el gasto cardiaco no se incrementa por medio del mecanismo dependiente de longitud porque la presión de llenado cardiaco no cambió. El gasto cardiaco aumenta a presión de llenado constante con un incremento de la actividad simpática cardiaca por dos razones. En primer lugar, el aumento en la actividad de nervios simpáticos cardiacos incrementa la frecuencia cardiaca. En segundo lugar, pero igual de importante, el aumento en la actividad de nervios simpáticos incrementa el volumen sistólico al elevar la contractilidad cardiaca. De este modo, los gráficos de función cardiaca consolidan el conocimiento de muchos mecanismos de control cardiaco, y son más útiles para describir cómo el corazón interactúa con otros elementos en el sistema cardiovascular. Además, estos gráficos vuelven a hacer hincapié en el importante punto de que un cambio de la presión de llenado cardiaca, sólo tendrá un efecto muy potente sobre el gasto cardiaco a cualquier magnitud de actividad simpática. RESUMEN DE INFLUENCIAS NEURALES SOBRE LA FUNCIÓN CARDIACA Los efectos del sistema nervioso simpático sobre las propiedades eléctricas y mecánicas del músculo cardiaco y, así, sobre la capacidad de bombeo cardiaca, son iniciados por la interacción de la norepinefrina con receptores β1-adrenérgicos en las células de músculo cardiaco. Esto da lugar a una cascada de eventos que comprenden la activación de Gs de la adenilato ciclasa, formación de cAMP y activación de la proteína cinasa A con fosforilación subsiguiente de muchas moléculas que desempeñan funciones reguladoras en procesos intracelulares. Estos eventos celulares se combinan para evocar las mejorías siguientes de las capacidades de bombeo del corazón: 1. un aumento de la frecuencia cardiaca (efecto cronotrópico positivo) por activación de la corriente de sodio hacia adentro if en células del nodo SA; 2. un decremento de la duración del potencial de acción cardiaco por activación temprana de la corriente iK tardía en miocitos 231 cardiacos, lo que minimiza el efecto perjudicial de frecuencias cardiacas altas sobre el tiempo de llenado diastólico; 3. Un incremento de la tasa de conducción del potencial de acción, en particular evidente en el nodo AV (efecto dromotrópico positivo), al alterar la conductividad de uniones intercelulares comunicantes (conexiones comunicantes); 4. un aumento de la contractilidad cardiaca (efecto inotrópico positivo) al activar la corriente iCa2+ e incrementar la liberación de Ca2+ desde el retículo sarcoplasmático, lo que aumenta la capacidad contráctil del músculo cardiaco a cualquier precarga dada. 5. Un aumento de la tasa de relajación cardiaca (efecto lusitrópico positivo) al incrementar la captación de Ca2+ por el retículo sarcoplasmático, lo que también ayuda a minimizar el efecto perjudicial de las frecuencias cardiacas altas sobre el tiempo de llenado diastólico. Casi todas las influencias de las catecolaminas sobre el corazón son un resultado de incrementos de la actividad neural simpática. Aunque las catecolaminas circulantes de origen suprarrenal en potencia pueden evocar efectos similares, sus concentraciones por lo normal son tan bajas que sus contribuciones son insignificantes. Fármacos específicos llamados bloqueadores (antagonistas) de receptores β1-adrenérgicos pueden bloquear todos los efectos de las catecolaminas que provienen de cualquier fuente sobre el músculo cardiaco. Estos fármacos suelen ser útiles en el tratamiento de enfermedad de arteria coronaria para contrarrestar demandas metabólicas aumentadas impuestas sobre el corazón por la actividad de nervios simpáticos. Los incrementos de la actividad simpática también pueden tener influencias indirectas sobre la función cardiaca, que son una consecuencia de alteraciones en el tono arteriolar y venoso inducidas por actividad simpática (esto es, alteraciones de la poscarga y la precarga, respectivamente (véanse los capítulos subsiguientes). DETERMINANTES DEL CONSUMO MIOCÁRDICO DE OXÍGENO En muchas situaciones patológicas, como aterosclerosis coronaria grave, los requerimientos de oxígeno del tejido miocárdico pueden exceder la capacidad del flujo sanguíneo coronario para suministrar oxígeno al músculo cardiaco. Esta desproporción puede dar lugar a dolor o molestias retrosternales intensos llamados angina de pecho. Tiene importancia entender qué factores determinan los costos de energía y, por ende, el consumo miocárdico de oxígeno, porque la reducción de la demanda de oxígeno suele tener beneficio clínico importante para el enfermo. Dado que el corazón obtiene su energía casi por completo a partir del metabolismo aeróbico, el consumo miocárdico de oxígeno guarda relación directa con el uso miocárdico de ATP. El metabolismo basal del tejido cardiaco (la energía consumida en procesos celulares que no son contracción, como bombeo de iones dependiente de energía) por lo normal explica alrededor de 25% del uso miocárdico de ATP y, en consecuencia, del consumo miocárdico de oxígeno en un individuo en reposo. Los procesos relacionados con la contracción muscular explican casi 75% del uso miocárdico de energía. Principalmente, esto refleja división de ATP vinculada con paso de 232 SECCIÓN V Fisiología cardiovascular puentes por ciclos durante Las fases de contracción isovolumétrica y eyección del ciclo cardiaco. También se usa algo de ATP para secuestro de Ca2+ al final de cada contracción. La energía que se gasta durante la fase de contracción isovolumétrica del ciclo cardiaco explica la porción más grande (~50%) del consumo miocárdico total de oxígeno pese al hecho de que el corazón no hace trabajo externo durante este periodo. La energía necesaria para contracción isovolumétrica depende principalmente de la presión intraventricular que debe desarrollarse durante este tiempo, es decir, de la poscarga cardiaca. La poscarga cardiaca, entonces, es un determinante importante del consumo miocárdico de oxígeno. Las reducciones de la poscarga cardiaca pueden producir reducciones importantes en clínica de los requerimientos miocárdicos de energía y, por ende, del consumo miocárdico de oxígeno. La utilización de energía durante la contracción isovolumétrica en realidad tiene relación más directa con el desarrollo de tensión de pared isométrica que con el desarrollo de presión intraventricular. Recuérdese que la tensión de pared se relaciona con la presión intraventricular y con el radio ventricular por medio de la ley de Laplace (T = P × r). En consecuencia, las reducciones de la precarga cardiaca (esto es, volumen al final de la diástole, radio) también tenderán a reducir la energía requerida para la contracción isovolumétrica. Es durante la fase de eyección del ciclo cardiaco que el corazón en realidad desempeña trabajo externo, y la energía que éste gasta durante la eyección depende de qué tanto trabajo externo está haciendo. En un sistema de fluido, el trabajo (fuerza × distancia) es igual a la presión (fuerza/distancia2) × volumen (distancia3). El trabajo físico externo efectuado por el ventrículo izquierdo en un latido, es decir, el trabajo sistólico, es igual al área encerrada por el asa de presión-volumen del ventrículo izquierdo (figura 24-3A). El trabajo sistólico es aumentado sea por un incremento del volumen sistólico (trabajo de “volumen” aumentado) o por un aumento de la poscarga (trabajo de “presión” aumentado). En términos de utilización de ATP y consumo de oxígeno, los incrementos del trabajo de presión del corazón son más costosos que los aumentos del trabajo de volumen. Así, las reducciones de la poscarga son en especial útiles en la reducción de los requerimientos miocárdicos de oxígeno para hacer trabajo externo. Los cambios de la contractilidad miocárdica pueden tener importantes consecuencias sobre el requerimiento de oxígeno para el metabolismo basal, la generación de tensión de pared isovolúmica, y el trabajo externo. Las células de músculo cardiaco usan más energía en el desarrollo rápido de una tensión dada y acortamiento por una cantidad dada, que en hacer lo mismo más lentamente. Con la contractilidad aumentada, también se gasta más energía en el transporte activo de Ca2+. El resultado neto de estas influencias a menudo se denomina efecto de “gasto de energía” de la contractilidad aumentada. La frecuencia cardiaca es uno de los determinantes de mayor importancia del consumo miocárdico de oxígeno porque los costos de energía por minuto deben ser iguales al costo de energía por cada latido, multiplicado por el número de latidos por minuto. En general, se ha encontrado que es más eficiente (esto es, se requiere menos oxígeno) lograr un gasto cardiaco dado con frecuencia cardiaca baja y volumen sistólico alto que con frecuencia cardiaca alta y volumen sistólico bajo. Esto de nuevo parece relacionarse con el costo de energía relativamente alto de la fase de desarrollo de presión isovolumétrica del ciclo cardiaco. Mientras menos presión (tensión de pared) se desarrolla, y mientras menos a menudo ocurre desarrollo de presión, menos energía se usa. CORRELACIÓN CLÍNICA Una mujer de 40 años de edad acude a la sala de urgencias debido al inicio repentino de debilidad y mareo alrededor de una hora antes. Además, reporta una sensación de aleteo en el tórax y la garganta. El examen revela frecuencia cardiaca rápida (165 latidos/min; taquicardia) y regular, y la presión arterial es baja, de 80/60 mmHg. El ECG muestra taquicardia relacionada con un complejo QRS estrecho. Después de intentos por restituir las frecuencias normales al dar masaje en el cuello en la ubicación de los senos carotídeos, y al colocar una compresa fría sobre su cara, se le administra una inyección de adenosina por vía intravenosa que convirtió la frecuencia cardiaca a 80 latidos/minuto, y la presión arterial a 130/85 mmHg. La molestia primaria de debilidad y mareo sugiere una disminución del flujo sanguíneo sistémico, incluso de la circulación cerebral, causado por la presión arterial baja. Esto podría depender de un decremento sea del gasto cardiaco o de la resistencia periférica total. En esta paciente, la frecuencia cardiaca muy rápida acortó gravemente la duración de la diástole y, como resultado, el llenado cardiaco estuvo significativamente alterado. Debido a la ley de Starling del corazón que relaciona el volumen al final de la diástole con el volumen sistólico, el decremento del llenado da lugar a una disminución del volumen sistólico. A pesar de la frecuencia cardiaca rápida, el volumen sistólico bajo produce un decremento importante del gasto cardiaco. La estrategia terapéutica comprende intentos por disminuir la frecuencia cardiaca. El ECG sugiere una taquicardia supraventricular con un foco ectópico auricular que se activa con rapidez o un circuito del nodo AV de reentrada que excita rápidamente el sistema de conducción ventricular normal (véanse más detalles en el capítulo 25.) Las maneras mecánicas de interrumpir estos procesos comprenden estrategias para aumentar la activación vagal hacia el tejido auricular al 1) dar masaje en el cuello en la región del seno carotídeo (masaje carotídeo), estirar los barorreceptores arteriales y hacer maniobras para que los centros cardiovasculares del bulbo raquídeo crean que la presión arterial está alta (capítulo 29) o 2) instituir el reflejo de buceo con una compresa fría en la cara, en el cual se induce bradicardia por activación de fibras aferentes del nervio trigémino al cerebro (capítulo 71). La estrategia farmacológica primaria empleada en la sala de urgencias comprendió inyección de un bolo de adenosina por vía intravenosa. Esto puede tener una influencia transitoria sobre la conducción supraventricular dentro del corazón, y a menudo puede interrumpir la vía de reentrada anormal. La adenosina fue eficaz en esta paciente. Si no lo hubiera sido, fármacos como los bloqueadores de receptores β-adrenérgicos o los bloqueadores de los canales de calcio pueden ser eficaces. Se remitió a la paciente con un cardiólogo quien realizará pruebas extensas de su función cardiaca para tratar de identificar la fuente anatómica del ritmo cardiaco anormal. CAPÍTULO 24 La bomba cardiaca RESUMEN DEL CAPÍTULO ■ ■ ■ ■ ■ ■ ■ ■ El bombeo de sangre eficaz por el corazón requiere llenado de las cámaras, excitación y contracción de las células de músculo cardiaco, generación de presión dentro de las cámaras, abertura y cierre de las válvulas cardiacas, y movimiento unidireccional de sangre a través de las cámaras hacia la aorta o la arteria pulmonar, coordinados. Excepto por las presiones de eyección más bajas, los eventos del lado derecho del corazón son idénticos a los del lado izquierdo. Los ruidos cardiacos relacionados con movimientos valvulares y detectados en la auscultación pueden usarse para identificar los inicios de las fases diastólica y sistólica del ciclo cardiaco. Los eventos de un ciclo cardiaco ventricular único pueden desplegarse como registros de cambios eléctricos, mecánicos, de presión, de ruido o de flujo contra el tiempo, o como un registro del volumen contra la presión. El gasto cardiaco se define como la cantidad de sangre bombeada por uno u otro de los ventrículos por minuto, y está determinado por el producto de la frecuencia cardiaca y el volumen sistólico. El volumen sistólico puede ser alterado por cambios de la precarga (llenado) ventricular, la poscarga ventricular (presión arterial), o la contractilidad del músculo cardiaco, o todos o una combinación de los anteriores. Una curva de función cardiaca describe la relación entre llenado ventricular y gasto cardiaco, y puede ser cambiada hacia arriba (izquierda) o hacia abajo (derecha) por cambios de la actividad simpática al corazón, o por cambios de la contractilidad del músculo cardiaco. La energía para la contracción del músculo cardiaco se deriva principalmente de vías metabólicas aeróbicas de modo que el trabajo cardiaco está estrechamente relacionado con el consumo miocárdico de oxígeno. PREGUNTAS DE ESTUDIO 1. Usted está auscultando los ruidos cardiacos de su paciente, y ha identificado los intervalos sistólico y diastólico. Cuatro de los estados que se listan a continuación existen durante la misma fase del ciclo cardiaco, y uno no. ¿Cuál es el que no? A) la válvula mitral está abierta B) está ocurriendo el segmento ST del ECG C) la onda “v” del pulso venoso yugular acaba de ocurrir 233 D) el volumen ventricular está aumentando con rapidez E) la presión aórtica está disminuyendo 2. Su paciente tiene arteriopatía coronaria con oclusión importante de la arteria coronaria descendente anterior izquierda. Aunque todos los que siguen aumentarán el trabajo cardiaco, ¿Cuál de los que siguen más probablemente dará lugar a un ataque de angina de pecho? A) recuencias cardiacas altas B) presiones arteriales altas C) volumen al final de la diástole aumentado D) presión arterial baja E) contractilidad cardiaca aumentada 3. Se está administrando un fármaco que bloquea los receptores β1-adrenérgicos cardiacos. ¿Cuáles serán las consecuencias directas de este fármaco sobre el corazón? A) la frecuencia cardiaca aumentará B) el intervalo PR del ECG se acortará C) las demandas metabólicas se reducirán D) la tasa de flujo coronario aumentará E) la contractilidad cardiaca aumentará 4. Se coloca una cánula en la arteria pulmonar (PA) de un individuo sano normal, y se reporta que la presión es de 25/8 mmHg. ¿Cuál de las que siguen se puede suponer a partir de estos valores? A) la presión sistólica del RV es de 25 mmHg; la presión diastólica de la PA es de 8 mmHg B) la presión sistólica del RV es de 25 mmHg; la presión en la aurícula izquierda es de 8 mmHg C) la presión sistólica en el RV debe ser de mucho más de 25 mmHg D) la presión diastólica en el RV debe ser de mucho más de 8 mmHg E) la presión capilar pulmonar es de 8 mmHg 5. Un individuo tiene una disfunción del sistema nervioso autónomo en la cual la actividad nerviosa parasimpática cardiaca aumenta de manera repentina, mientras que la actividad simpática cardiaca disminuye. ¿Cuál de las situaciones que siguen es más probable que ocurra? A) el volumen del LV al final de la diástole aumentará B) el gasto cardiaco aumentará C) la frecuencia cardiaca aumentará D) el volumen del LV al final de la sístole disminuirá E) el flujo sanguíneo coronario aumentará Evaluaciones de la función cardiaca C A P Í T U L O 25 Lois Jane Heller y David E. Mohrman O B J E T I V O S ■ ■ ■ ■ ■ ■ ■ ■ ■ Declarar la relación entre eventos eléctricos de excitación cardiaca y las ondas P, QRS y T, los intervalos PR y QT, y el segmento ST del electrocardiograma. Enunciar las convenciones electrocardiográficas básicas de Einthoven y, dados los datos, determinar el eje eléctrico medio del corazón. Describir el electrocardiograma de 12 derivaciones estándar. Detectar arritmias cardiacas comunes a partir del electrocardiograma, identificar sus bases fisiológicas y describir sus consecuencias fisiológicas. Calcular el gasto cardiaco usando el principio de Fick. Reconocer imágenes ecocardiográficas obtenidas durante el ciclo cardiaco. Definir la fracción de eyección e identificar los métodos de visualización que se usan para determinarla. Definir la relación presión-volumen al final de la sístole. Listar anormalidades valvulares estenóticas y regurgitantes para el hemicardio izquierdo, y describir sus consecuencias en términos de presiones intracardiaca y arterial, patrones de flujo, y ruidos cardiacos que las acompañan. TÉCNICAS PARA EVALUAR LA FUNCIÓN CARDIACA Se dispone de diversos métodos para evaluar la función cardiaca. Algunos de éstos son no invasivos (p. ej., electrocardiografía para evaluar las características eléctricas, auscultación del tórax para evaluar la función de válvulas, y ecocardiografía para visualizar la acción de bombeo mecánica), y otros requieren diversos tipos de instrumentación invasiva. En este capítulo se proporcionarán una perspectiva general breve de algunos de estos instrumentos clínicos de uso común, y una introducción a algunas de las anormalidades funcionales cardiacas comunes. MEDICIÓN DE LA EXCITACIÓN CARDIACA —EL ELECTROCARDIOGRAMA El electrocardiograma es un recurso clínico poderoso que se usa para evaluar la frecuencia de los latidos del corazón, el ritmo y las características de conducción del tejido cardiaco. El electrocardiograma es el resultado de corrientes propagadas a través del líquido extracelular, que son generadas por la diseminación de la onda de excitación por todo el corazón (véase capítulo 23). Electrodos que se colocan sobre la superficie del cuerpo registran las diferencias de potencial pequeñas entre diversos sitios de registro, que varían durante la evolución temporal del ciclo cardiaco. 235 236 SECCIÓN V Fisiología cardiovascular R +1 mV Voltaje +0.5 T P 0 Q –0.5 S Segmento Segmento Intervalo ST PR QRS Intervalo PR Intervalo QT Tiempo FIGURA 251 Electrocardiograma típico. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) En la figura 25-1 se muestra un registro electrocardiográfico típico de voltajes registrados en la superficie del cuerpo entre la pierna izquierda y el brazo derecho (llamado “derivación II”). Las principales características del electrocardiograma son las ondas P, QRS y T que se producen por la despolarización auricular, despolarización ventricular y repolarización ventricular, respectivamente. El periodo que transcurre desde el inicio de la onda P hasta el inicio del complejo QRS se llama intervalo PR e indica el tiempo que se requiere para que un potencial de acción se disemine por las aurículas y el nodo auriculoventricular (AV). Durante la última porción del intervalo PR (segmento PR), no se detectan voltajes sobre la superficie corporal. Esto se debe a que las células musculares auriculares están despolarizadas (en la fase de meseta de sus potenciales de acción), las células ventriculares aún están en reposo, y el campo eléctrico establecido por el potencial de acción que progresa a través del nodo AV pequeño no es suficientemente intenso como para que se detecte. La duración del intervalo PR normal varía de 120 a 200 ms. Poco después de que el impulso cardiaco sale del nodo AV y pasa hacia el sistema de Purkinje de conducción rápida, todas las células musculares ventriculares se despolarizan con rapidez y causan el complejo QRS. La onda R es el evento de mayor tamaño en el electrocardiograma porque las células musculares ventriculares son muy numerosas, y porque se despolarizan casi al unísono. El complejo QRS normal dura entre 60 y 100 ms. (La repolarización de células auriculares también está ocurriendo en el mismo periodo durante el cual la despolarización ventricular genera el complejo QRS en el electrocardiograma [véase figura 23-3]. La repolarización auricular no es evidente en el electrocardiograma porque es un evento poco sincronizado en una masa relativamente pequeña de tejido cardiaco, y está por completo ensombrecido por los eventos eléctricos importantes que ocurren en los ventrículos en este momento.) El complejo QRS va seguido por el segmento ST. En circunstancias normales, no se miden potenciales eléctricos sobre la superficie del cuerpo durante el segmento ST porque no están ocurriendo cambios rápidos en el potencial de membrana en cualquiera de las células del corazón; las células auriculares ya han vuelto a la fase de reposo, mientras que las células musculares ventriculares están en la fase de meseta de sus potenciales de acción. (Sin embargo, la lesión miocárdica o el flujo sanguíneo inadecuado puede producir elevaciones o depresiones en el segmento ST.) Cuando las células ventriculares empiezan a repolarizarse, aparece de nuevo un voltaje sobre la superficie del cuerpo, y se mide como la onda T del electrocardiograma. La onda T es más amplia y no tan grande como la onda R porque la repolarización ventricular es menos sincrónica que la despolarización. Cuando concluye la onda T, todas las células del corazón se encuentran en el estado de reposo. El intervalo QT aproxima a grandes rasgos la duración de la despolarización de miocitos ventriculares y, así el periodo de sístole ventricular. A una frecuencia cardiaca normal de 60 latidos/min, el intervalo QT por lo normal es de menos de 380 ms. No se mide potencial en la superficie corporal sino hasta que el nodo sinoauricular (SA) genera el siguiente impulso. La operación del sistema de conducción especializado es un factor primario en la determinación del patrón electrocardiográfico normal; por ejemplo, el tiempo de transmisión del nodo AV determina el intervalo PR. Asimismo, la eficacia del sistema de Purkinje ventricular en la sincronización de la despolarización ventricular se refleja en la magnitud grande y la duración breve del complejo QRS. Recuerde que casi todas las células de músculo cardiaco son inherentemente capaces de tener ritmicidad, y que todas las células cardiacas están interconectadas eléctricamente por medio de uniones intercelulares comunicantes (conexiones comunicantes). Así, un ritmo cardiaco funcional puede ocurrir, y a menudo ocurre, sin la participación de parte del sistema de conducción especializado o de todo ese sistema. Empero, esa situación es anormal, y la existencia de vías de conducción anormales produce un electrocardiograma anormal. CONVENCIONES ELECTROCARDIOGRÁFICAS BÁSICAS El registro de electrocardiogramas es un procedimiento diagnóstico sistemático estandarizado por la aplicación universal de ciertas convenciones. Las convenciones para registrar electrocardiogramas y analizarlos a partir de las tres derivaciones bipolares estándar de las extremidades, se describen de forma breve aquí, y se resumen en la figura 25-2. Los electrodos de registro se colocan en ambos brazos y en la pierna izquierda —por lo general en las muñecas y el tobillo—. Los brazos y las piernas actúan como extensiones conductivas desde el cuerpo, y las mediciones de voltaje en realidad están entre puntos que forman un triángulo equilátero sobre el tórax. Esto se llama triángulo de Einthoven, en honor del fisiólogo holandés que lo ideó a finales del siglo XIX. Cualquier trazo electrocardiográfico único es un registro de la diferencia de voltaje medida entre cualesquiera de los dos vértices del triángulo de Einthoven. Un ejemplo del electrocardiograma de la derivación II medido entre el brazo derecho y la pierna izquierda ya se ha mostrado en la figura 25-2. De modo similar, los electrocardiogramas de la derivación I y de la derivación III representan mediciones de voltaje obtenidas a lo largo de los otros dos lados del triángulo de Einthoven (figura 25-2). Los símbolos + y – en la figura 25-2 indican convenciones de polaridad que se han adoptado de manera universal. Por ejemplo, una deflexión hacia arriba en el electrocardiograma de la derivación II (como ocurre por CAPÍTULO 25 Evaluaciones de la función cardiaca II + er i +D i ón ac r iv Selector y amplificador de derivación va ció n e –D Registrador de gráfico Brazo izquierdo – Derivación I + III – Brazo derecho 237 Pierna izquierda Escala de voltaje 10 mm hacia arriba = +1 mV Velocidad del papel FIGURA 252 Convenciones electrocardiográficas 25 mm = 1 s de Einthoven. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) partir de dos derivaciones electrocardiográficas. Para entender estos análisis electrocardiográficos más complejos, es necesario examinar la manera en que los voltajes aparecen sobre la superficie del cuerpo como resultado de la actividad eléctrica cardiaca. lo normal durante las ondas P, R y T) indica que hay una polaridad eléctrica en ese instante entre los electrodos de la pierna izquierda y del hombro derecho; el electrodo de la pierna izquierda es positivo. Por el contrario, una deflexión descendente en un registro de la derivación II indica que existe una polaridad eléctrica entre este par de electrodos, pero esta vez el electrodo en la pierna izquierda está registrando cargas negativas en lugar de positivas. Se han establecido convenciones de polaridad similares para los registros de la derivación I y de la derivación III, y se indican por los símbolos + y − en la figura 25-2. Además, el equipo de registro electrocardiográfico por lo general está estandarizado de modo que una deflexión de 1 cm en el eje vertical siempre representa una diferencia de potencial de 1 mV, y que un segmento de 25 mm del trazo horizontal de cualquier registro electrocardiográfico representa 1 s. Casi todos los registros electrocardiográficos contienen señales de calibración, de modo que pueden detectarse con facilidad frecuencias y amplitudes de onda anormales. Muchas anormalidades eléctricas cardiacas pueden detectarse en registros provenientes de una derivación electrocardiográfica única (véase la sección que sigue). Con todo, cierta información útil en clínica sólo se puede recabar al combinar la información obtenida a Células despolarizadas Nodo SA - + - + - - - + + +++ Aurículas RA – – DIPOLOS CARDIACOS Y REGISTROS ELECTROCARDIOGRÁFICOS En la figura 25-3 se ilustra la conceptualización de Einthoven de cómo la actividad eléctrica cardiaca causa diferencias de potencial sobre la superficie del cuerpo. En este ejemplo, el corazón es mostrado en un instante en la fase de despolarización auricular. El impulso cardiaco, después de haber surgido en el nodo SA, se está diseminando como un frente de onda de despolarización por todo el tejido auricular. En cada punto a lo largo de este frente de onda de actividad eléctrica, existe una separación de carga pequeña en el líquido extracelular entre membranas polarizadas (exterior positivo) y membranas despolarizadas (exterior negativo). Así, el frente de onda puede considerarse una serie de dipolos eléctricos individuales (regiones de separación de carga). Cada dipolo individual está orien- + I LA – Frente de onda de actividad eléctrica Dipolo neto III II Ventrículos + + LL FIGURA 253 Dipolo cardiaco neto durante despolarización auricular y sus componentes en las derivaciones de las extremidades. RA, aurícula derecha; LA, aurícula izquierda; LL, pierna izquierda. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) 238 SECCIÓN V Fisiología cardiovascular tado en la dirección del movimiento de frente de onda local. La flecha negra grande en la figura 25-3 representa el dipolo neto creado por las contribuciones sumadas de todos los dipolos individuales distribuidos a lo largo del frente de onda de despolarización auricular. El líquido extracelular salado actúa como un excelente conductor, y permite que estos dipolos netos instantáneos generados sobre la superficie del músculo cardiaco se registren por medio de electrodos en la superficie del cuerpo. El dipolo neto que se presenta en cualquier instante está orientado (esto es, apunta) hacia la dirección general del movimiento de frente de onda en ese instante. La magnitud o fuerza del dipolo (representada aquí mediante la longitud de la flecha) está determinada por: 1) la extensión del frente de onda (esto es, cuántas células se están despolarizando de manera simultánea en el instante en cuestión) y 2) la constancia de la orientación entre dipolos individuales en puntos diferentes en el frente de onda (dipolos con la misma orientación se refuerzan entre sí; los dipolos con orientación opuesta se cancelan uno a otro). El dipolo neto en el ejemplo de la figura 25-3 hace que la porción inferior izquierda del cuerpo en general sea positiva respecto a la porción superior derecha. Este dipolo particular hará que existan voltajes positivos en las tres derivaciones de las extremidades del electrocardiograma. Esto puede deducirse a partir del triángulo de Einthoven al observar que el dipolo neto tiene algún componente que orienta en la dirección positiva de las derivaciones I, II y III (figura 25-3, mitad derecha). El componente que un dipolo cardiaco tiene en una derivación de electrocardiograma dada se encuentra al trazar líneas perpendiculares desde el lado apropiado del triángulo de Einthoven hacia la punta y la cola del dipolo (figura 25-3). (Puede ser útil pensar en el componente de cada derivación como la “sombra” emitida por el dipolo sobre esa derivación como resultado de un “sol” localizado bastante más allá del ángulo del triángulo de Einthoven que es opuesto a la derivación.) Note que el dipolo en este ejemplo es más paralelo a la derivación II y, por ende, tiene un componente grande en la dirección de dicha derivación. Así, creará un voltaje de mayor magnitud en la derivación II que en las derivaciones I o III. Este dipolo tiene un componente más bien pequeño en la derivación III porque está orientado casi perpendicular a dicha derivación. La configuración de la derivación de extremidad puede considerarse una manera de ver la actividad eléctrica del corazón desde tres perspectivas (o ejes) diferentes. El vector que representa la fuerza y reorientación del dipolo instantáneo del corazón es el objeto bajo observación, y su aspecto depende de la posición desde la cual se ve. El voltaje instantáneo medido en el eje de la derivación I, por ejemplo, indica cómo el dipolo que está siendo generado por la actividad eléctrica del corazón en ese instante aparece cuando se ve directamente desde arriba. Un dipolo cardiaco que está orientado de manera horizontal parece grande en la derivación I, mientras que un dipolo cardiaco orientado de forma vertical, independientemente de qué tan grande sea, no produce voltaje en la derivación I. Así, es necesario tener vistas desde dos direcciones para establecer la magnitud del dipolo del corazón y la orientación del mismo. Un dipolo orientado verticalmente sería invisible en la derivación I, pero sería evidente si se ve desde la perspectiva de la derivación II o III. Es importante reconocer que el ejemplo de la figura 25-3 sólo se refiere a un instante durante la despolarización auricular. La magnitud y orientación del dipolo cardiaco neto cambian continuamente en el transcurso de la despolarización auricular. La naturaleza de estos cambios determinará la forma de la onda T en cada una de las derivaciones del electrocardiograma. La onda P termina cuando la onda de despolarización llega al borde no muscular entre las aurículas y los ventrículos, y el número de dipolos individuales se hace muy pequeño. En este momento, el impulso cardiaco aún se está transmitiendo de manera lenta hacia los ventrículos a través del nodo AV. Aun así, la actividad eléctrica en el nodo AV involucra tan pocas células que no genera dipolo cardiaco neto detectable. Así, no se miden voltajes en la superficie del cuerpo durante un breve periodo después de la onda P. Un dipolo cardiaco neto sólo reaparece cuando la despolarización completa su paso por el nodo AV, entra al sistema de Purkinje, y empieza su paso rápido sobre las células musculares ventriculares. Dado que las fibras de Purkinje terminan en el tabique intraventricular y en las capas endomiocárdicas en el vértice de los ventrículos, la despolarización ventricular ocurre primero en estas áreas y después procede hacia afuera y hacia arriba a través del miocardio ventricular. DESPOLARIZACIÓN VENTRICULAR Y EL COMPLEJO QRS El complejo QRS del electrocardiograma depende de los cambios rápidos y grandes de la magnitud y la dirección de los dipolos cardiacos netos que existen durante la despolarización ventricular. El proceso normal se ilustra en la figura 25-4. La despolarización ventricular inicial por lo general ocurre en el lado izquierdo del tabique intraventricular según se muestra con un diagrama en el panel superior de la figura. El análisis del dipolo cardiaco formado por esta despolarización ventricular inicial con la ayuda del triángulo de Einthoven muestra que este dipolo tiene un componente negativo en la derivación I, un componente negativo pequeño en la derivación II y un componente positivo en la derivación III. El panel superior derecho muestra las deflexiones reales en cada una de las derivaciones electrocardiográficas de las extremidades que serán producidas por este dipolo. Note que es posible que un dipolo cardiaco dado produzca deflexiones opuestas en diferentes derivaciones. Por ejemplo, en la figura 25-4, las ondas Q aparecen en las derivaciones I y II, pero no en la derivación III. La segunda hilera de paneles en la figura 25-4 muestra los ventrículos durante el instante en la despolarización ventricular en que el número de dipolos individuales es mayor, o su orientación es más similar o ambos. Esta fase genera el dipolo cardiaco neto grande, del cual depende la onda R del electrocardiograma. En la figura 25-4, este dipolo cardiaco neto es casi paralelo a la derivación II. Como se indica, ese dipolo produce ondas R positivas grandes en las tres derivaciones de las extremidades. La tercera hilera de diagramas en la figura 25-4 muestra la situación cerca del final de la diseminación de la despolarización a través de los ventrículos e indica cómo el dipolo cardiaco neto pequeño presente en este momento produce la onda S. Nótese que la onda S no aparece de forma necesaria en todas las derivaciones del electrocardiograma (como en la derivación I de este ejemplo). La hilera inferior de diagramas en la figura 25-4 muestra que durante el segmento ST, todas las células musculares ventriculares están en un estado despolarizado. No hay ondas de actividad eléctrica moviéndose por el tejido del corazón. En consecuencia, no existe dipolo neto en este momento, y no presentan diferencias de voltaje CAPÍTULO 25 Evaluaciones de la función cardiaca – + I 239 I – – II III II Onda Q – III + + + I – – II Onda R II III + – I + III + I – I – II Onda S II III + + III I – + I – – II Segmento ST II III FIGURA 254 Despolarización ventricular y la generación del complejo QRS. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular + + III Physiology, 6th ed. New York: Lange Medical Books/ McGraw-Hill, 2006.) entre puntos sobre la superficie del cuerpo. Todos los trazos electrocardiográficos serán planos al nivel isoeléctrico (voltaje cero). EJE ELÉCTRICO MEDIO Y DESVIACIONES DEL EJE REPOLARIZACIÓN VENTRICULAR Y LA ONDA T La orientación del dipolo cardiaco durante la fase más intensa de despolarización ventricular (esto es, en el instante en que la onda R alcanza su máximo) se llama eje eléctrico medio del corazón. Se usa en clínica como un indicador de si la despolarización ventricular está procediendo en vías normales. El eje eléctrico medio se reporta en grados de acuerdo con la convención indicada en la figura 25-5. Nótese que la dirección hacia abajo es designada como más 90° en este sistema de coordenadas polares lo que describe la orientación del eje en el plano frontal (un plano vertical que divide el cuerpo en secciones anterior y posterior). Como se indicó, un eje eléctrico medio que yace en cualquier lugar en el cuadrante inferior izquierdo del paciente es considerado normal (entre 0° y 90°). Existe una desviación del eje hacia la izquierda cuando el eje eléctrico medio cae en el cuadrante superior izquierdo del paciente, y puede indicar cualesquiera de los varios estados, como un desplazamiento físico del corazón hacia la izquierda, hipertrofia del ventrículo izquierdo, o pérdida de la actividad eléctrica en parte del ventrículo derecho (p. ej., después de un infarto). Una desviación del eje hacia la derecha existe cuando el eje eléctrico medio cae en el cuadrante inferior derecho del paciente, y puede indicar, entre varios estados, un desplazamiento físico del corazón hacia la derecha, hipertrofia ven- La onda T por lo normal es positiva en la derivación II al igual que la onda R (figura 25-1). Esto indica que el dipolo cardiaco neto generado durante la repolarización ventricular está orientado en la misma dirección general que la que se presenta durante la despolarización ventricular. Esto tal vez sea un poco sorprendente. De cualquier modo, recuérdese que las últimas células ventriculares en despolarizarse son las primeras en repolarizarse (véase figura 24-3). Las razones de esto no se entienden bien, pero el resultado es que el frente de onda de actividad eléctrica durante la repolarización ventricular tiende a volver a trazar, en dirección inversa, la trayectoria seguida durante la despolarización ventricular. Por ende, el dipolo que se forma durante la repolarización tiene la misma polaridad que el que se forma durante la despolarización. Esta vía de propagación del frente de onda revertida durante la repolarización ventricular da lugar a una onda T positiva registrada, por ejemplo, en la derivación II. La onda T es más amplia y de menor tamaño que la onda R porque la repolarización de las células musculares ventriculares no está tan bien sincronizada que su despolarización. 240 SECCIÓN V Fisiología cardiovascular RA LA <90 grados 180 0 grados Desviación del eje hacia la derecha > +90 grados grados Desviación del eje hacia la izquierda < 0 grados Rango de cero a +90 grados normal +90 grados LL FIGURA 255 Eje eléctrico medio y desviaciones del eje. RA, aurícula derecha; LA, aurícula izquierda; LL, pierna izquierda. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) tricular derecha o pérdida de la actividad eléctrica en parte del ventrículo izquierdo. El eje eléctrico medio del corazón puede determinarse a partir del electrocardiograma. El proceso comprende determinar cuál orientación de dipolo neta única producirá las amplitudes de la onda R registradas en cualesquiera de las dos derivaciones. Por ejemplo, si las ondas R en las derivaciones II y III son positivas (hacia arriba) y de igual magnitud, el eje eléctrico medio debe ser de +90°. Como debe ser obvio, en este caso, la amplitud de la onda R en la derivación I será de cero. De manera alternativa, es posible explorar los registros electrocardiográficos para el trazo de derivación con las ondas R de mayor tamaño y después deducir que el eje eléctrico medio debe ser casi paralelo a esa derivación. En la figura 25-4, por ejemplo, la onda R más grande ocurre en la derivación II; la cual tiene una orientación de +60°, que está muy cerca del eje eléctrico medio real en este ejemplo. EL ELECTROCARDIOGRAMA DE 12 DERIVACIONES ESTÁNDAR El electrocardiograma clínico estándar involucra mediciones de voltaje registradas a partir de 12 derivaciones diferentes. Tres de éstas son las derivaciones bipolares de las extremidades I, II y III, que ya se comentaron. Las otras nueve son derivaciones unipolares. Tres de estas derivaciones se generan al usar los electrodos de las extremidades, dos de los electrodos son conectados eléctricamente para formar un electrodo indiferente, mientras que en el tercer electrodo de extremidad se hace el polo positivo del par. Los registros realizados a partir de estos electrodos se llaman derivaciones unipolares aumentadas de las extremidades. El registro de voltaje obtenido entre el electrodo en el brazo derecho y el electrodo indiferente se llama un electrocardiograma de la derivación aVR. De modo similar, la derivación aVL se registra a partir del electrodo en el brazo izquierdo, y la derivación aVF se registra a partir del electrodo en la pierna izquierda. Las derivaciones estándar de las extremidades (I, II y III) y las unipolares aumentadas de las extremidades (aVR, aVL y aVF) registran la actividad eléctrica del corazón como aparece desde seis “perspectivas” diferentes. Los ejes para las derivaciones I, II y III son los de los lados del triángulo de Einthoven, mientras que aquéllos para aVR, aVL y aVF son especificados por líneas trazadas desde el centro del triángulo de Einthoven hacia cada uno de los vértices (figura 25-6A). Estas seis derivaciones de las extremidades pueden considerarse el sistema de referencia hexaxial para observar los vectores cardiacos en el plano frontal (figura 25-6B). Las otras seis derivaciones del electrocardiograma de 12 derivaciones estándar también son derivaciones unipolares que “analizan” las proyecciones del vector eléctrico en el plano transverso (un plano horizontal que divide el cuerpo en segmentos superior e inferior). Estos potenciales se obtienen al colocar un electrodo adicional (explorador) en seis posiciones especificadas en la pared del tórax (figura 25-6C). El electrodo indiferente en este caso se forma al conectar eléctricamente los electrodos de las extremidades. Estas derivaciones se identifican como derivaciones precordiales o torácicas, y se designan V1 a V6. Como se muestra en la figura 25-6, la onda de excitación ventricular se aleja desde V1, lo que da por resultado una deflexión descendente. La onda de excitación ventricular se dirige hacia V6, lo que da lugar a una deflexión ascendente. En resumen, el electrocardiograma es un poderoso recurso para evaluar las características de excitación cardiaca. No obstante, debe reconocerse que el ECG no proporciona evidencia directa de la eficacia de bombeo mecánico. Por ejemplo, una válvula cardiaca con escape por lo general no tendrá consecuencias electrocardiográficas directas, pero puede influir de manera adversa sobre la capacidad de bombeo del corazón. EXCITACIÓN Y RITMICIDAD CARDIACAS ANORMALES El material que se presenta aquí es una introducción a las anormalidades más comunes de la frecuencia y el ritmo cardiacos, con hincapié en las consecuencias fisiológicas primarias de estas situaciones anormales. Muchos problemas de excitación cardiacos pueden diagnosticarse a partir de la información en una derivación única de un electrocardiograma. El trazo electrocardiográfico de la derivación II en la parte superior de la figura 25-7 es identificado como un ritmo sinusal normal con base en las características que siguen: 1) la frecuencia de complejos QRS es ~1/s, lo que indica una frecuencia normal de 60 latidos/min; 2) la forma del complejo QRS es normal para la derivación II y su duración es de menos de 120 ms, lo que indica despolarización rápida de los ventrículos mediante vías de conducción normales; 3) cada complejo QRS va precedido por una onda P de configuración apropiada, lo que indica el origen de la excitación en el nodo SA; 4) el intervalo PR es de menos de 200 ms, lo que señala retraso apropiado de la conducción de la propagación del impulso a través del nodo AV; 5) el intervalo QT es de menos de la mitad del intervalo R a R, lo que indica repolarización ventricular normal, y 6) no hay ondas P extra, lo que muestra que no hay bloqueo de la conducción del nodo AV. Los trazos electrocardiográficos subsiguientes en las figuras 25-7 y 25-9 representan irregularidades que se encuentran por lo común en la práctica clínica. El examen de cada uno de estos trazos con las características anteriores en mente ayudará en el diagnóstico diferencial. Las consecuencias fisiológicas de la excitación y conducción anormales en el corazón dependen de si la anormalidad eléctrica evoca CAPÍTULO 25 Evaluaciones de la función cardiaca A B RA – – I + aV aV L + R + LA – – – – aVF II aVR – 241 C – – + aVL + – + I III – – + + + LL + III + aVF V1 + II V6 V2 V3 V4 V5 D FIGURA 256 El electrocardiograma de 12 derivaciones estándar. A y B) Derivaciones en el plano frontal. C) Posiciones de electrodos para las derivaciones precordiales en el plano transverso. D) Un ECG de 12 derivaciones. La línea inferior es una tira de ritmo obtenida a partir de la derivación V1. RA, aurícula derecha; LA, aurícula izquierda; LL, pierna izquierda. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006. La figura 25-6 D es cortesía del Dr. David Gutterman. una taquicardia, que limitará el tiempo para el llenado cardiaco entre los latidos; evoca una bradicardia, que es inadecuada para apoyar gasto cardiaco suficiente, o disminuye la coordinación de la contracción de los miocitos, lo que reducirá el volumen sistólico (SV). ANORMALIDADES SUPRAVENTRICULARES Los trazos 2 a 6 por debajo del trazo normal en la figura 25-7 representan arritmias supraventriculares típicas (esto es, que se originan en las aurículas o en el nodo AV). La taquicardia supraventricular (que se observa en el trazo 2 de la figura 25-7, y a veces se llama taquicardia auricular paroxística) ocurre cuando las aurículas son excitadas anormalmente e impulsan los ventrículos a una frecuencia muy rápida. Estos paroxismos pueden empezar de manera repentina, duran entre algunos minutos y horas, y después, de manera igual de repentina, desaparecen y la frecuencia cardiaca se revierte a lo normal. Los complejos QRS parecen normales (aunque frecuentes) en la taquicardia auricular paroxística simple porque las vías de conducción ventriculares operan normalmente. Las ondas P y T pueden estar superpuestas debido a la frecuencia cardiaca alta. Los brotes de estas arritmias pueden acompañarse de presión arterial baja y mareo porque la frecuencia cardiaca en extremo alta no permite que haya suficiente tiempo diastólico para el llenado ventricular. Existen dos mecanismos que pueden explicar la taquicardia supraventricular. En primer lugar, una región auricular, por lo general fuera del nodo SA, puede hacerse irritable (quizá debido a la interrupción local del flujo sanguíneo) y empezar a activar con rapidez para asumir la función de marcapasos para todo el corazón. Esa región marcapasos anormal se llama foco ectópico. De manera alternativa, la conducción auricular puede quedar alterada de modo que una onda de excitación única no desaparece, sino que viaja de manera continua alrededor de algún asa de conducción auricular anormal. En este caso, la actividad continua en el asa de conducción puede impulsar las aurículas y el nodo AV a una frecuencia muy alta. Este proceso autosostenido se llama fenómeno de reentrada, y se representa en un diagrama en la figura 25-8. Esta situación puede aparecer como resultado de repolarización anormal y periodos refractarios alterados en áreas locales del miocardio. El aleteo auricular es una forma especial de taquicardia de origen auricular en la cual una vía de reentrada grande impulsa las aurículas a frecuencias muy rápidas (250 a 300 latidos/min) y los periodos refractarios normales del tejido del nodo AV son abrumados. Así, la frecuencia 242 SECCIÓN V Fisiología cardiovascular 1. Ritmo sinusal normal 2. Taquicardia supraventricular 3. Bloqueo de primer grado 2:1 4:1 4. Bloqueo de segundo grado 5. Bloqueo de tercer grado 6. Fibrilación auricular 1 mV FIGURA 257 Arritmias 1s supraventriculares. (Reproducida con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) ventricular a menudo es alguna proporción fija de la frecuencia auricular (2:1, 4:1), con frecuencias a menudo de 150 a 220 latidos/min. Los bloqueos de la conducción ocurren en el nodo AV y por lo general representan conducción alterada a través del tejido. En el bloqueo cardiaco de primer grado (trazo 3 de la figura 25-7), la única anormalidad eléctrica es conducción excepcionalmente lenta a través del nodo AV. Este estado se detecta por un intervalo PR anormalmente largo (> 0.2 s). Por lo demás, el electrocardiograma puede resultar normal. A frecuencias cardiacas normales, los efectos fisiológicos de un bloqueo de primer grado por lo regular son intrascendentes. Sin embargo, el peligro es que la conducción lenta puede deteriorarse hacia una interrupción real de la conducción. Se dice que hay un bloqueo cardiaco de segundo grado (trazo 4 de la figura 25-7) cuando algunos de los impulsos auriculares, pero no todos, son transmitidos a través del nodo AV hacia el ventrículo. Los impulsos son bloqueados en el nodo AV si las células de la región aún están en un periodo refractario desde una excitación previa. La situación se agrava por frecuencias auriculares altas y conducción más baja que lo normal a través de la región del nodo AV. En el bloqueo de segundo grado, algunas ondas P, pero no todas, se acompañan de complejos QRS y ondas T correspondientes. La frecuencia auricular a menudo es más rápida que la ventricular por una cierta proporción (p. ej., 2:1, 3:1, 4:1). Este estado puede no representar un problema clínico grave en tanto la frecuencia ventricular sea suficiente para satisfacer las necesidades de bombeo. En el bloqueo cardiaco de tercer grado (trazo 5 de la figura 25-7) no se transmiten impulsos a través del nodo AV. En este evento, algún área en los ventrículos —a menudo en el fascículo común o ramas del fascículo cerca de la salida del nodo AV— asume la función de marcapasos para el tejido ventricular. Las frecuencias auricular y ventricular son por completo independientes, y las ondas P y los complejos QRS están por completo disociados en el electrocardiograma. La frecuencia ventricular quizá es más lenta que lo normal (bradicardia), y a veces es suficientemente lenta como para alterar el gasto cardiaco. La fibrilación auricular (trazo 6 de la figura 25-7) se caracteriza por una pérdida completa de la sincronía por lo normal cercana de las fases de excitación y de reposo entre las células auriculares. Células en áreas diferentes de las aurículas se despolarizan, repolarizan y son excitadas de nuevo al azar. En consecuencia, no aparecen ondas P en el electrocardiograma, aunque puede haber ondas pequeñas rápidas e irregulares manifiestas durante toda la diástole. La frecuencia ventricular a menudo es muy irregular en la fibrilación auricular porque los impulsos entran al nodo AV desde las aurículas en momentos impredecibles. La fibrilación es un proceso que se sostiene por sí solo. No se entienden bien los mecanismos que están por detrás de ella, pero se cree que los impulsos progresan repetidas veces alrededor de vías de conducción irregulares (a veces llamadas vías circulares, lo que implica un fenómeno de reentrada como se describió antes, y como se muestra en la figura 25-8). Empero, dado que la contracción auricular por lo general desempeña una función insignificante en el llenado ventricular, la mayoría de los pacientes puede tolerar bien la fibrilación auricular en tanto la frecuencia ventricular sea suficiente para mantener el gasto cardiaco. El peligro real con la fibrilación auricular yace en la tendencia de la sangre a formar coágulos en las aurículas en ausencia de la contracción auricular vigorosa y coordinada normal. Estos coágulos suelen fragmentarse, salir del corazón y alojarse en arterias de pequeño calibre en toda la circulación sistémica o pulmonar. Estos émbolos pueden tener efectos devastadores sobre la función de órganos cruciales. En consecuencia, se usa terapia anticoagulante como profilaxis para pacientes con fibrilación auricular. Vía normal Vía de reentrada FIGURA 258 Vías de excitación cardiaca normal y de reentrada (circular). (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) CAPÍTULO 25 Evaluaciones de la función cardiaca 243 1. Ritmo sinusal normal 2. Bloqueo de rama del fascículo 3. Contracción ventricular prematura 4. Taquicardia ventricular 5. Síndrome de QT largo con torsades de pointes FIGURA 259 Arritmias ventriculares. (Modificada con autorización de Mohrman DE, Heller LJ: 6. Fibrilación ventricular 1 mV 1s ANORMALIDADES VENTRICULARES Los trazos 2 a 6 por debajo del trazo normal en la figura 25-9 muestran anormalidades eléctricas ventriculares típicas. Los bloqueos de conducción llamados bloqueos o hemibloqueos de rama del fascículo (trazo 2 de la figura 25-9) pueden ocurrir en una u otra de las ramas del sistema de Purkinje del tabique intraventricular a menudo como resultado de un infarto del miocardio. La despolarización ventricular es menos sincrónica que lo normal en la mitad del corazón con el sistema de Purkinje no funcional. Esto da por resultado una ampliación del complejo QRS (>0.12 s) porque se requiere más tiempo para que se complete la despolarización ventricular célula-célula del lado bloqueado. Los efectos fisiológicos directos de los bloqueos de rama del fascículo por lo general carecen de importancia. Las contracciones ventriculares prematuras (PVC; trazo 3 de la figura 25-9) se originan por potenciales de acción iniciados por un foco ectópico en el ventrículo, y propagados desde éste. Como resultado, el ventrículo se despolariza y se contrae antes de lo que normalmente lo haría. Una PVC a menudo va seguida por un latido omitido (lo que se llama una pausa compensadora) porque las células ventriculares aún son refractarias cuando surge el siguiente impulso normal desde el nodo SA. El patrón de despolarización ventricular muy anormal de una PVC produce las deflexiones de gran amplitud y de larga duración en el electrocardiograma. Las formas de los registros electrocardiográficos de estos latidos extra son muy variables, y dependen del sitio ectópico de su origen, y de las vías de despolarización involucradas. El volumen de sangre eyectado por el latido prematuro en sí es más pequeño que lo normal (si es que se eyecta un volumen), mientras que el volumen sistólico del latido que sigue a la pausa compensadora es mayor que el normal. Esto se debe en parte a las diferencias de los tiempos de llenado, y en parte a un fenómeno inherente del músculo cardiaco llamado potenciación posextrasistólica. Dado que las PVC ocurren en ocasiones en la mayoría de los individuos y, aunque a veces son alarmantes para quien las está experimentando, no son peligrosas. Con todo, la aparición frecuente de PVC, puede ser una señal de posible daño miocárdico o problemas de perfusión, y puede llevar a taquicardia ventricular o incluso fibrilación ventricular (véase más adelante). Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) La taquicardia ventricular (trazo 4 de la figura 25-9) ocurre cuando los ventrículos son impulsados a frecuencias altas, por lo general por impulsos que se originan a partir de un foco ectópico ventricular. La taquicardia ventricular es una afección muy grave. No sólo hay limitación del tiempo de llenado diastólico por la frecuencia rápida, sino que las vías de excitación anormales también hacen que la contracción ventricular sea menos sincrónica y, por ende, menos eficaz que lo normal. Además, la taquicardia ventricular a menudo precede a la fibrilación ventricular. Los intervalos QT prolongados (lado izquierdo del trazo 5 en la figura 25-9) son un resultado de repolarización tardía de miocitos ventriculares, lo cual puede deberse a abertura inapropiada de canales de sodio o cierre prolongado de canales de potasio durante la fase de meseta del potencial de acción. Aunque el intervalo QT normal varía con la frecuencia cardiaca, por lo normal es de menos de 40% de la duración del ciclo cardiaco (excepto a frecuencias cardiacas muy altas). El síndrome de QT largo se identifica cuando el intervalo QT es de más de 50% de la duración del ciclo. Puede ser de origen genético (mutaciones que influyen sobre diversos canales iónicos involucrados en la excitabilidad cardiaca), puede ser adquirido por diversas alteraciones de electrólitos (concentraciones sanguíneas bajas de Ca2+, Mg2+ o K+), o puede ser inducido por varios fármacos (entre ellos algunos antiarrítmicos). La prolongación del periodo refractario del miocito, que acompaña al síndrome de QT largo, extiende el periodo vulnerable durante el cual estímulos extra pueden desencadenar taquicardia o fibrilación. Los pacientes con síndrome de QT largo están predispuestos a un tipo en particular peligroso de taquicardia ventricular llamada torsades de pointes (“torsión de puntos”, como se muestra en el lado derecho del trazo 5 en la figura 25-9). Esto difiere de la taquicardia ventricular ordinaria por cuanto los complejos eléctricos ventriculares varían de amplitud de manera cíclica alrededor de la basal, y pueden deteriorarse con rapidez hacia fibrilación ventricular. En la fibrilación ventricular (trazo 6 de la figura 25-9), diversas áreas del ventrículo son excitadas y se contraen de manera asincrónica. Los mecanismos son similares a los que se observan en la fibrilación auricular. El ventrículo es en especial susceptible a fibrilación siempre que ocurre una excitación prematura al final de la onda T de la excitación previa, es decir, cuando casi todas las células ventriculares 244 SECCIÓN V Fisiología cardiovascular Presión del LV (mmHg) A Contractilidad aumentada Contractilidad normal 120 Poscarga aumentada 80 120 60 Volumen del LV (ml) Presión del LV (mmHg) 80 Co ntr a no ctilid rm al ad Individuo normal 120 Ẋtc Q̇ = ________ [X] _ [X] Precarga aumentada 40 B algunas suposiciones importantes basadas en técnicas no invasivas. En uno de los métodos más exactos para medir el gasto cardiaco mediante métodos invasivos se emplea el principio de Fick, que se comenta con mayor detalle en el capítulo 26. En resumen, éste establece que la cantidad de una sustancia consumida por los tejidos, Xtc, es igual a lo que ingresa menos lo que egresa (que es la diferencia de concentración arteriovenosa en la sustancia ([X]a – [X]v) tantas veces como la tasa de flujo sanguíneo, Q. ). Esta relación puede despejarse de manera algebraica para resolver el flujo sanguíneo, como sigue: Paciente con insuficiencia cardiaca d da tili da c i a ntr inu Co ism d Sin tratamiento Tratado con reductor de la poscarga 40 60 120 180 Volumen del LV (ml) FIGURA 2510 Relaciones de presión-volumen al final de la sístole del ventrículo izquierdo. A) El efecto de contractilidad aumentada desplaza la línea hacia arriba y hacia la izquierda. B) El efecto de la insuficiencia cardiaca sistólica desplaza la línea hacia abajo y hacia la derecha. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) se encuentran en el periodo “hiperexcitable” o “vulnerable” de su ciclo eléctrico. Además, dado que algunas células están repolarizadas y algunas aún son refractarias, pueden desencadenarse con facilidad vías circulares en este momento. Dado que en presencia de fibrilación ventricular no hay acción de bombeo, la situación es mortal a menos que se corrija con rapidez por medio de conversión cardiaca (por lo común llamada desfibrilación o cardioversión eléctrica externa). Durante este procedimiento, la aplicación artificial de corrientes grandes a todo el corazón (por medio de electrodos en forma de paleta aplicados a través del tórax) puede ser eficaz para despolarizar todas las células del corazón simultáneamente y, así, permitir que se restablezca una vía de excitación normal. Es necesario administrar reanimación cardiopulmonar (CPR) hasta que se logra una desfibrilación. MEDICIÓN DEL GASTO CARDIACO Principio de Fick: la medición del gasto cardiaco no es una tarea simple, y por lo general comprende alguna maniobra invasiva o a (1) v Un método común para determinar el gasto cardiaco es usar el principio de Fick para calcular el flujo colectivo a través de los órganos sistémicos a partir de: 1) la tasa de consumo de oxígeno de todo el cuerpo (X. tc); 2) el contenido de oxígeno de la sangre arterial ([X]a) y 3) la concentración de oxígeno en sangre venosa mixta ([X]v). De los valores que se requieren para este cálculo, el contenido de oxígeno de la sangre venosa mixta es el más difícil de obtener. En general, la muestra para medición de oxígeno en sangre venosa debe obtenerse a partir de un catéter venoso colocado en la arteria pulmonar a fin de asegurarse de que es una muestra mezclada de sangre venosa que proviene de todos los órganos sistémicos. El cálculo del gasto cardiaco a partir del principio de Fick se observa mejor mediante un ejemplo. Supóngase que un paciente está consumiendo 250 ml de O2/min cuando la sangre arterial sistémica contiene 200 ml de O2/L y la sangre del ventrículo derecho contiene 150 ml de O2/L. Esto significa que, en promedio, cada litro de sangre pierde 50 ml de O2 conforme pasa por los órganos sistémicos. Para que se consuman 250 ml de O2 por minuto, deben pasar 5 L de sangre por la circulación sistémica cada minuto: Q. = 250 [ml de O2/min] 200 – 150 [ml de O2/L de sangre] = 5 [L de sangre/min] (2) Aunque el uso del principio de Fick como se describió proporciona el estándar para la determinación del gasto cardiaco, existen varias otras técnicas que proporcionan buenos estimados del gasto cardiaco. Las técnicas de dilución de indicador comprenden la inyección de una cantidad conocida de indicador (colorante o un bolo térmico) hacia la sangre que está entrando al hemicardio derecho, y se colocan detectores apropiados para registrar de manera continua la concentración de indicador en la sangre conforme sale del hemicardio derecho. La dilución del indicador es proporcional al gasto cardiaco. Otras técnicas para obtener imágenes del corazón (ecocardiografía, angiografía ventricular, ventriculografía con radionúclidos) pueden usarse para estimar el volumen sistólico, el gasto cardiaco y otros índices de la función ventricular, como se describe a continuación. El índice cardiaco es el gasto cardiaco corregido para el tamaño del individuo. Por ejemplo, el gasto cardiaco de una mujer de 50 kg será significativamente más bajo que el de un varón de 90 kg. Aun así, se ha encontrado que el gasto cardiaco se correlaciona mejor con el área de superficie corporal que con el peso corporal. Por ende, es común expresar el gasto cardiaco por metro cuadrado de área de superficie. En condiciones en reposo, el índice cardiaco por lo normal es de alrededor de 3 (l/min)/m2. CAPÍTULO 25 Evaluaciones de la función cardiaca ESTIMACIONES DE LA CONTRACTILIDAD CARDIACA A menudo es importante evaluar la función cardiaca de un individuo sin usar procedimientos invasivos mayores. Los avances en algunas técnicas han hecho posible obtener imágenes bidimensionales y tridimensionales del corazón durante todo el ciclo cardiaco. El análisis visual o auxiliado por computadora de esas imágenes proporciona información útil en la evaluación clínica de la función cardiaca. La ecocardiografía es la más usada de las diversas técnicas de obtención de imágenes disponibles en la actualidad. Esta técnica no invasiva se basa en el hecho de que las ondas de sonido se reflejan de regreso hacia la fuente cuando encuentran cambios repentinos de la densidad del medio a través del cual viajan. Un transductor, colocado en sitios específicos en el tórax, genera impulsos de ondas ultrasónicas y detecta ondas reflejadas desde las interfases de tejido cardiaco. Mientras más tiempo transcurre entre la transmisión de la onda y la llegada del reflejo, a mayor profundidad se encuentra la estructura en el tórax. Esa información puede reconstruirse de diversas maneras mediante computadora para producir una imagen continua del corazón y sus cámaras durante todo el ciclo cardiaco. La ecocardiografía es en especial idónea para detectar operación anormal de válvulas cardiacas o de la función contráctil en porciones de las paredes del corazón. También puede proporcionar estimados de los volúmenes de las cámaras cardiacas en diferentes momentos del ciclo cardiaco, que se usan de diversos modos para evaluar la función del corazón. La fracción de eyección (EF) es una medición clínica en extremo útil que puede calcularse a partir de un ecocardiograma. Se define como la proporción entre volumen sistólico (SV) y el volumen al final de la diástole (EDV): SV EF = ____ EDV (3) A partir de las imágenes pueden hacerse estimados de los volúmenes al final de la diástole y de la sístole, y calcular el SV. La EF por lo común se expresa como un porcentaje, y en circunstancias normales varía desde 55 hasta 80% (media 67%) en condiciones de reposo. Las EF de menos de 55% indican contractilidad miocárdica deprimida. La relación presión-volumen al final de la sístole es otra técnica útil en clínica para evaluar la contractilidad cardiaca. El volumen al final de la sístole para un ciclo cardiaco dado se estima mediante una de las técnicas de obtención de imágenes antes descritas, mientras que la presión al final de la sístole para ese ciclo cardiaco puede obtenerse a partir de la presión arterial registrada en el punto del cierre de la válvula aórtica (la incisura). Pueden obtenerse valores para varios ciclos cardiacos durante administración de un vasoconstrictor por vía intravenosa lenta (que aumenta la poscarga), y graficar los datos como en la figura 25-10 en el contexto de asas de presiónvolumen ventricular generales. Como se muestra, los aumentos de la contractilidad miocárdica se relacionan con rotación a la izquierda de la relación presión-volumen al final de la sístole. Este método para evaluar la función cardiaca tiene particular importancia porque proporciona un estimado de la contractilidad que es independiente del EDV (precarga). (Recuérdese que los incrementos de la precarga causan aumentos del SV sin cambiar el volumen al final de la sístole 245 [véase figura 24-4]. Así, sólo las alteraciones de la contractilidad causarán cambios de la relación presión-volumen al final de la sístole.) Las líneas de presión-volumen al final de la sístole tanto “normales” como “con contractilidad aumentada” casi se proyectan al origen a presión cero, volumen cero (figura 25-10A). Así, es posible obtener un estimado clínico razonable de la pendiente de la relación presiónvolumen al final de la sístole (léase “contractilidad miocárdica” a partir de una medición única de la presión y el volumen al final de la sístole. Esto evita la necesidad de efectuar múltiples pruebas caras con administraciones de vasodilatador o vasoconstrictor por vía intravenosa lenta. Un decremento de la contractilidad (como puede originarse por enfermedad cardiaca) se relaciona con un cambio hacia abajo de la relación presión-volumen al final de la sístole, y se conoce como insuficiencia cardiaca sistólica. En esta situación, los aumentos del impulso simpático tienen influencia limitada sobre el gasto cardiaco. Parte del proceso compensador comprende un aumento importante de la retención de líquido en el cuerpo que da lugar a un incremento del volumen sanguíneo circulante y del EDV ventricular (capítulo 29). Un asa de presión-volumen del ventrículo izquierdo que describe los eventos de un ciclo cardiaco de un corazón insuficiente (figura 25-10B) está muy desplazada a la derecha de la de un corazón normal. El paciente no tratado descrito en esta figura está en serios problemas con SV y EF reducidos, y presión de llenado alta con posible congestión vascular pulmonar. Además, la pendiente de la línea que describe la relación presión-volumen al final de la sístole muestra desplazamiento hacia abajo y está menos empinada, lo que indica la contractilidad reducida del músculo cardiaco. No obstante, debido a esta relación más plana, las reducciones pequeñas de la poscarga cardiaca (esto es, presión arterial) producirán incrementos considerables de la EF y el SV que ayudarán de manera significativa a este paciente. FUNCIÓN ANORMAL DE VÁLVULAS CARDIACAS La acción de bombeo del corazón está alterada cuando las válvulas no funcionan de manera apropiada. Varias técnicas, que varían desde la auscultación simple (escuchar los ruidos cardiacos) hasta ecocardiografía o cateterismo cardiaco, se emplean para obtener información acerca de la naturaleza de estas funciones inadecuadas de válvulas y la magnitud de las mismas. A menudo, los defectos de válvula cardiaca se acompañan de ruidos cardiacos anormales llamados soplos. Estos ruidos se originan por gradientes de presión anormales y patrones de flujo sanguíneo turbulentos que se presentan durante el ciclo cardiaco. En general, cuando una válvula no se abre por completo (o sea, muestra estenosis), la cámara torrente arriba de la válvula tiene que desarrollar más presión durante su fase sistólica para obtener un flujo dado a través de la válvula. Este aumento del trabajo de “presión” inducirá hipertrofia de las células de músculo cardiaco y engrosamiento de las paredes de esa cámara. (Esto es análogo a los músculos esqueléticos hipertrofiados del levantador de pesas que hace trabajo isométrico o de tensión alta.) Cuando una válvula no se cierra por completo (es decir, muestra insuficiencia, regurgitación o incompetencia), el flujo sanguíneo regurgitante representa un volumen adicional que debe eyectarse para obtener suficiente flujo anterógrado 246 SECCIÓN V Fisiología cardiovascular hacia afuera del ventrículo y hacia los tejidos. Este aumento del trabajo de “volumen” a menudo lleva a dilatación de la cámara, pero no a un incremento del grosor de la pared. (Esto es análogo a los músculos esqueléticos no hipertrofiados, pero con buen tono del corredor de larga distancia que hace trabajo isotónico o de acortamiento.) Una segunda generalidad acerca de las anormalidades valvulares es que cuando existe un aumento de la presión auricular como resultado de estenosis o regurgitación de válvula AV, esto dará por resultado presiones más altas en los lechos capilares torrente arriba. Si las presiones hidrostáticas capilares están aumentadas, surgirá edema de tejidos con consecuencias negativas considerables sobre la función de esos órganos torrente arriba. En la figura 25-11 se presenta una breve perspectiva general de cuatro de los defectos valvulares comunes que influyen sobre la fun- ción del ventrículo izquierdo. El lector debe notar que pueden ocurrir anormalidades estenóticas y regurgitantes similares en las válvulas del ventrículo derecho, con consecuencias similares sobre la función de dicho ventrículo. ESTENOSIS AÓRTICA En la figura 25-11A se muestran algunas características de la estenosis aórtica. En circunstancias normales, la válvula aórtica se abre de modo amplio y ofrece una vía de resistencia muy baja a través de la cual la sangre abandona el ventrículo izquierdo. Si esta abertura queda estrechada (estenótica), la resistencia al flujo a través de la válvula aumenta. Puede requerirse una diferencia de presión importante A B 150 Presión aórtica 100 Presión en el ventrículo izquierdo Presión en la aurícula izquierda 50 0 ECG Fonocardiograma C D Presión aórtica 100 Presión en el ventrículo izquierdo Presión en la aurícula izquierda 50 0 ECG Fonocardiograma FIGURA 2511 Anormalidades valvulares comunes. A) Estenosis aórtica. B) Estenosis mitral. C) Regurgitación (insuficiencia) aórtica. D) Insuficiencia mitral. (Modificada con autorización de Mohrman DE, Heller LJ: Cardiovascular Physiology, 6th ed. New York: Lange Medical Books/McGraw-Hill, 2006.) CAPÍTULO 25 Evaluaciones de la función cardiaca entre el ventrículo izquierdo y la aorta para eyectar sangre a través de una válvula aórtica estenótica. Las presiones intraventriculares pueden aumentar hasta cifras muy altas durante la sístole mientras la presión aórtica se incrementa de forma más lenta que lo normal hasta un valor sistólico que es subnormal (figura 25-11A). La presión del pulso por lo general es baja en la estenosis aórtica. El desarrollo de presión intraventricular alta es un estímulo fuerte para hipertrofia de células de músculo cardiaco, y la estenosis aórtica siempre se acompaña de un incremento de la masa muscular del ventrículo izquierdo. Esto tiende a producir una desviación del eje eléctrico hacia la izquierda. (El eje eléctrico medio caerá en el cuadrante superior derecho de la figura 25-5.) La sangre que se está eyectando a través del orificio estrechado suele alcanzar velocidades muy altas, y puede ocurrir flujo turbulento conforme la sangre entra a la aorta. Este flujo turbulento anormal puede auscultarse como un soplo sistólico (o de eyección) con un estetoscopio colocado de manera apropiada. La consecuencia fisiológica primaria de la estenosis aórtica es una poscarga ventricular alta que se origina por restricción del tracto de salida. Esto impone una carga de trabajo de presión aumentada sobre el ventrículo izquierdo. ESTENOSIS MITRAL En la figura 25-11B se presentan algunas características de la estenosis mitral. Una diferencia de presión de más de algunos milímetros de mercurio a través de la válvula mitral durante la diástole es claramente anormal e indica que esta válvula es estenótica. La resistencia alta exige una diferencia de presión aumentada para lograr flujo normal a través de la válvula (Q. = ΔP/R). En consecuencia, en la estenosis mitral hay incremento de la presión en la aurícula izquierda (figura 25-11B). La carga de trabajo alta de la aurícula izquierda puede inducir hipertrofia del músculo de dicha aurícula. La presión elevada en la aurícula izquierda se refleja de regreso hacia el lecho pulmonar y, si es suficientemente alta, causa edema pulmonar y congestión vascular pulmonar. A menudo puede auscultarse un soplo diastólico relacionado con flujo turbulento a través de la válvula mitral estenótica. Las consecuencias fisiológicas primarias de la estenosis mitral son aumentos de la presión de la aurícula izquierda y de la presión capilar pulmonar. Esto último puede causar interferencia con el intercambio de gases normal en los pulmones, lo que lleva a disnea (falta de aliento). INSUFICIENCIA AÓRTICA En la figura 25-11C se muestran las características típicas de la regurgitación (insuficiencia, incompetencia) aórtica. Cuando las hojuelas