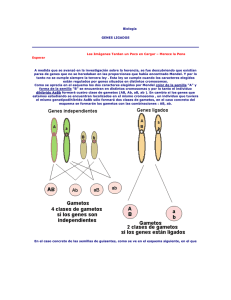
Exposición Formación de Gametos para el cuadro de Punnet - Encontrar todas las combinaciones posibles y no trabajar de más. Cuando un organismo hace gametos, cada gameto recibe solo una copia del gen, que es seleccionada al azar. Esto se conoce como la ley de la segregación. Frecuencia de Recombinación Ligamiento Genético Cuando los genes están en cromosomas separados o muy lejos en el mismo cromosoma se segregan independientemente. Es decir, cuando los genes van hacia los gametos, el alelo que se recibe para un gen no afecta al alelo que se recibe para el otro. En un organismo heterocigoto doble (AaBb), esto resulta en l a formación de los 4 tipos posibles de gametos con igual frecuencia, 25%. Los genes en cromosomas separados se segregan independientemente debido a la orientación aleatoria de los pares de cromosomas homólogos durante la meiosis. Los cromosomas homólogos son cromosomas apareados que portan los mismos genes, pero pueden tener alelos diferentes de esos genes. Un miembro de cada par homólogo viene de la madre de un organismo, el otro viene de su padre. Cuando estamos siguiendo dos genes, esto resulta en cuatro tipos de gametos que se producen con la misma frecuencia. Cuando los genes están en el mismo cromosoma pero muy separados, se segregan independientemente debido al entrecruzamiento (recombinación homóloga). Proceso en el cual los cromosomas homólogos intercambian fragmentos equivalentes de forma aleatoria. El entrecruzamiento puede poner combinar nuevos alelos en el mismo cromosoma, lo que causa que vayan hacia el mismo gameto. Cuando los genes están muy separados, el entrecruzamiento sucede con la frecuencia suficiente para que todos los tipos de gametos se produzcan con 25% de frecuencia. Cuando los genes están muy juntos en el mismo cromosoma, aún sucede el entrecruzamiento, pero el resultado (en términos de tipos de gametos producidos) es diferente. En lugar de segregarse independientemente, los genes tienden a "mantenerse juntos" durante la meiosis. Es decir, los alelos de los genes que ya están juntos en un cromosoma tenderán a transmitirse como una unidad a los gametos. En este caso, los genes están ligados. Los tipos comunes de gametos contienen configuraciones parentales de alelos, es decir, los que ya estaban juntos en el cromosoma en el organismo antes de la meiosis (o sea, en el cromosoma que obtuvo de sus padres). Los tipos raros de gametos contienen configuraciones recombinantes de alelos: los que solo pueden formarse si ocurre recombinación (entrecruzamiento) entre los genes. ¿Por qué los tipos de gametos recombinantes son raros? La razón básica es que los entrecruzamientos entre dos genes que están muy juntos no son muy comunes. Los entrecruzamientos durante la meiosis ocurren en posiciones más o menos aleatorias a lo largo del cromosoma, entonces la frecuencia de entrecruzamientos entre dos genes depende de la distancia entre ellos. Gracias a esta relación, podemos usar la frecuencia de sucesos de recombinación entre dos genes (es decir, su grado de ligamiento genético) para estimar su distancia relativa en el cromosoma. Dos genes muy juntos tendrán muy pocos sucesos de recombinación y estarán fuertemente ligados, mientras que dos genes que están ligeramente más separados tendrán más sucesos de recombinación y estarán ligados con menor frecuencia. Frecuencia Recombinante Supongamos que estamos interesados en ver si dos genes en la mosca de la fruta (Drosophila) están ligados entre ellos y, si es así, qué tan fuertemente ligados están. En nuestro ejemplo, los genes son: - El gen púrpura, con un alelo dominante pr+ que especifica ojos rojos normales y un alelo recesivo pr que especifica ojos púrpura. - El gen vestigial con un alelo dominante vg+ que especifica alas largas normales y un alelo recesivo vg que especifica alas cortas "vestigiales". Empezar cruzando dos moscas homocigotas como se muestra a continuación: Este cruzamiento nos da exactamente lo que necesitamos para observar la recombinación: una mosca que es heterocigota para los genes púrpura y vestigial, en la cual sabemos claramente qué alelos están juntos en un solo cromosoma. Para hacerlo, podemos cruzar una mosca heterocigota doble con un sujeto de prueba, una mosca que es homocigota recesiva para todos los genes de interés (en este caso, los alelos pr y vg). El propósito de usar un sujeto de prueba es asegurar que los alelos proporcionados por un padre no probador determinen completamente el fenotipo, o apariencia, de la descendencia. Cuando cruzamos nuestra mosca de interés con una de prueba, podemos "leer" directamente el genotipo de cada gameto de la apariencia física de la descendencia. Una mosca hembra heterocigota doble produce cuatro tipos diferentes de óvulos, cada uno de los cuales se combina con un espermatozoide de la mosca de prueba masculina. En este cruzamiento se producen cuatro clases fenotípicas (basadas en la apariencia) diferentes de descendientes, y cada una corresponde a un gameto particular de la madre: Las cuatro clases de descendientes no se producen en cantidades iguales, lo que nos dice que los genes púrpura y vestigial están ligados. Como esperamos para los genes ligados, las configuraciones de cromosomas parentales están sobrerepresentadas en la descendencia, mientras que las configuraciones de cromosomas recombinantes están subrepresentadas. Para medir el ligamiento cuantitativamente, podemos calcular la frecuencia de recombinación (FR) entre los genes púrpura y vestigial: Podemos identificar estas moscas como clases recombinantes por dos razones: una, sabemos por la serie de cruzamientos que realizamos que deben haber heredado un cromosoma de su madre que haya experimentado un suceso de recombinación; y dos, son las clases subrepresentadas (relativo a las clases sobrerepresentadas parentales). La frecuencia de recombinación de esos genes es de 10.7%. Una forma en que las frecuencias de recombinación se han usado históricamente es para construir mapas de ligamiento, mapas cromosómicos basados en las frecuencias de recombinación. De hecho, el estudio del ligamiento ayudó a los primeros genetistas a establecer que los cromosomas eran realmente lineales y que cada gen tenía su lugar específico en un cromosoma. La frecuencia de recombinación no es una medida directa de qué tan separados físicamente están los genes en los cromosomas. Sin embargo, proporciona una estimación o aproximación de la distancia física. Así, podemos decir que un par de genes con una frecuencia de recombinación más grande están probablemente más separados, mientras que un par con una frecuencia de recombinación menor probablemente están más juntos. Es importante notar que la frecuencia de recombinación es como máximo 50% (que corresponde a los genes no ligados o que se segregan independientemente). Es decir, 50% es la frecuencia de recombinación más grande que mediremos directamente entre los genes. También se puede comparar las frecuencias de recombinación para averiguar el orden de los genes en un cromosoma. Por ejemplo, supongamos que tenemos tres genes A, B y C, y queremos conocer su orden en el cromosoma (¿ABC? ¿ACB? ¿CAB?). Si observamos las frecuencias de recombinación entres los tres pares posibles de genes (AC, AB, BC), podemos saber qué genes se encuentran más separados y qué otro gen se encuentra en medio.