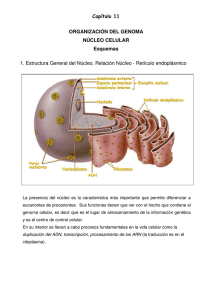
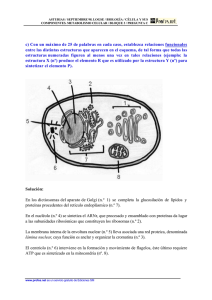
Núcleo La envoltura nuclear y tráfico entre el núcleo y el citoplasma Las membranas nucleares actúan como una barrera selectiva que impide el libre paso de las moléculas entre el interior nuclear y el citoplasma, manteniéndolos como dos compartimentos metabólicamente independientes. (a) (b) (a) Micrografía electrónica de un núcleo de célula HeLa en interfase. La heterocromatina es evidente alrededor de la superficie interna de la envoltura nuclear. Pueden verse dos nucléolos prominentes y algunos grumos de cromatina diseminados en el nucleoplasma. (b) Esquema que muestra algunos de los principales componentes del núcleo. (A: TOMADA DE WERNER W. FRANKE, INT. REV. CYTOL. (SUPL) 4:130, 1974.) Los únicos canales en la envuelta nuclear están representados por los complejos de poro nucleares, que permiten un intercambio controlado de moléculas entre el núcleo y el citoplasma. El tráfico selectivo de proteínas y ARN a través de los complejos de poro nucleares no sólo mantiene la composición interna del núcleo sino que tiene un papel clave en la regulación de la expresión génica. La envuelta nuclear posee una estructura compleja, constituida por membranas nucleares, la lámina nuclear en su cara interna y por los complejos de poro nucleares. La membrana nuclear externa se continúa con la membrana del retículo endoplásmico, por lo que hay una comunicación directa entre el espacio intermembrana y el lumen del retículo endoplásmico. La membrana nuclear interna tiene proteínas únicas que son específicas para el núcleo, como aquellas que unen la matriz nuclear de láminas La envoltura nuclear. (a) Esquema que muestra la membrana doble, el complejo del poro nuclear, la lámina nuclear y la continuidad de la membrana externa con el retículo endoplásmico rugoso. Ambas membranas de la envoltura nuclear contienen su propio complemento distintivo de proteínas. (b) Micrografía electrónica de un corte a través de una porción de la envoltura nuclear de una (a) célula de raíz de cebolla. Nótense la doble membrana (NM) con un espacio interpuesto, el complejo del poro nuclear (NPC) y la heterocromatina relacionada (HC) que no se extiende en la región de los poros nucleares. (B: TOMADA DE WERNER W. FRANKE ET AL., J. CELL BIOL. 91:47S, 1981; MEDIANTE AUTORIZACIÓN DE (b) DERECHOS RESERVADOS DE LA ROCKEFELLER UNIVERSITY PRESS.) 10-50 nm La lámina nuclear es el soporte mecánico de la envoltura nuclear. (a) FIGURA 12-3 La lámina nuclear. Núcleo de una célula humana cultivada que se tiñó con anticuerpos fluorescentes para revelar la lámina nuclear (rojo), que se encuentra en la superficie interna de la envoltura nuclear. La matriz nuclear está teñida de verde. las membranas interna y externa se unen en los complejos de poro nuclear, siendo los únicos canales que permiten el paso de pequeñas moléculas polares y de macromoléculas a través de la envuelta nuclear Subyacente a la membrana nuclear interna se localiza la lámina nuclear, una red fibrosa que proporciona soporte estructural al núcleo Enfermedades de la lámina nuclear MUTACIONES EN LA LÁMINA A ESTÁN ASOCIADOAS AL SÍNDROME DE LA PROGENIE DE HUTCHINSON – GILFORD (HGPS) QUE SE CARACTERIZA POR EL ENVEJECIMINETO PREMATURO Y LA MUERTE DURANTE LA ADOLESCENCIA A CAUSA DE UN ATAQUE CARDIACO O APOPLEJIA (c) Estas micrografías muestran el núcleo dentro de un fibroblasto que se cultivó a partir de un paciente con HGPS (hilera inferior) o un sujeto sano (hilera superior). Las células se tiñeron para mostrar la proteína lámina A (columna izquierda), para DNA (columna media) o se muestran en estado vivo con el microscopio óptico con contraste de fase (columna derecha). El núcleo celular del paciente con HGPS es anormal por la presencia de una proteína lámina A trunca en la lámina nuclear. (A: TOMADA DE H. MA, A.J. SIEGEL Y R. BEREZNEY; J. CELL BIOL. 146:535, 1999; AUTORIZACIÓN DE DERECHOS DE THE ROCKEFELLER UNIVERSITY PRESS. B: REIMPRESA CON AUTORIZACIÓN DE U. AEBI, J. COHN, L. BUHLE Y L. GERACE, NATURE 323:561, 1986, © COPYRIGHT 12986, MACMILLAN MAGAZINES LIMITED. C: TOMADA DE ANNA MATTOUT ET AL., POR CORTESÍA DE ROBERT D. GOLDMAN, CURR. OPIN. CELL BIOL. 18:338, 2006.) La lámina nuclear está compuesta de proteínas fibrosas de 60 a 80 kilodaltons (kd) denominadas lamininas junto a algunas proteínas asociadas Las lamininas son un tipo de proteínas de los filamentos intermedios; las otras clases se encuentran en el citoesqueleto Las células de mamíferos poseen tres genes de laminina, denominados A, B y C, que codifican al menos siete proteínas diferentes. Al igual que las otras proteínas de los filamentos intermedios, las láminas se ensamblan entre ellas para formar filamentos Las láminas interaccionan con proteínas de la membrana nuclear interna, como la emerina y el receptor de laminina B, mediando su unión a la envuelta nuclear y localizando y organizándolas en el interior nuclear La lámina nuclear también se une a la cromatina a través de las histonas H2A y H2B además de a otras proteínas cromatínicas. (a) Micrografía electrónica de un extremo del citoplasma nuclear de un oocito de rana tomado minutos después de la inyección con partículas de oro cubiertas con una proteína que en condiciones normales se encuentra en el núcleo. Se observa que estas partículas pasan a través del centro del poro nuclear (flechas) en su ruta desde el citoplasma hacia el núcleo. (b) En una magnificación mucho más definida puede verse que las partículas de oro se organizan en un arreglo lineal dentro de cada poro. A TRAVÉS DEL PORO NUCLEAR PASAN Proteínas al núcleo RNA mensajeros y t-RNA y las subunidades del ribosoma del núcleo al citosol. Sn RNA que van en ambas direcciones (a) (b) Complejo del poro nuclear Los complejos del poro nuclear son los únicos canales a través de los cuales pueden viajar pequeñas moléculas polares, iones y macromoléculas (proteínas y ARN) entre el núcleo y el citoplasma El complejo del poro nuclear es una estructura muy grande con un diámetro de aproximadamente 120 nm y un peso molecular estimado de aproximadamente 125 millones de daltons —unas 30 veces el tamaño de un ribosoma Mediante el control del tráfico de moléculas entre el núcleo y el citoplasma, el complejo del poro nuclear tiene un papel fundamental en la fisiología de todas las células eucariotas. 1,- Las moléculas de ARN que son sintetizadas en el núcleo deben ser exportadas de manera eficiente al citoplasma, donde intervienen en la síntesis de proteínas. 2,- las proteínas necesarias para las funciones nucleares (p. ej., factores de transcripción) deben entrar en el núcleo procedentes de los lugares de síntesis en el citoplasma. Muchas proteínas sufren un trasiego continuo entre el núcleo y el citoplasma. Las moléculas pueden pasar a través del complejo del poro nuclear mediante uno de dos mecanismos diferentes Las moléculas pequeñas y algunas proteínas con un peso molecular inferior a 20-40 kDa pasan libremente a través de la envuelta nuclear en ambas direcciones: del citoplasma al núcleo o del núcleo al citoplasma indistintamente. Figura 9.6 Tráfico molecular a través de los complejos del poro nuclear. Las moléculas pequeñas son capaces de atravesar rápidamente los canales abiertos del complejo del poro nuclear por difusión pasiva. Por el contrario, las macromoléculas (proteínas y ARN) son transportadas por un mecanismo selectivo dependientes de energía. Estas moléculas difunden de manera pasiva a través de los canales acuosos abiertos, los cuales tienen un diámetro estimado de aproximadamente 9 nm, en el interior del complejo del poro nuclear La mayoría de las proteínas y ARN, sin embargo, no son capaces de pasar por estos canales abiertos. Estas macromoléculas atraviesan el poro central de unos 10-40 nm del complejo del poro nuclear mediante un proceso activo, en el que las proteínas y los ARN adecuados son reconocidos y transportados selectivamente en una dirección específica (del núcleo al citoplasma o del citoplasma al núcleo). Señales de localización nuclear Estas proteínas son las responsables de todas las características de la estructura y de la función del genoma; incluyen las histonas, las ADN polimerasas, las ARN polimerasas, factores de transcripción, factores de splicing y muchas otras. Estas proteínas se etiquetan para ser destinadas al núcleo con secuencias de aminoácidos específicas, denominadas señales de localización nuclear, que son reconocidas por los receptores de transporte nuclear y dirigen la translocación de proteínas a través del complejo del poro nuclear. Importinas Los receptores de transporte nuclear conocidos como importinas (debido a su actividad transportadora de proteínas al núcleo) reconocen las señales de localización nuclear. El movimiento de macromoléculas a través de poro nuclear se controla por una proteína denominada Ran. Ran representa una de las diversas clases de proteínas de bajo peso molecular cuya conformación y actividad están reguladas por la unión y la hidrólisis de GTP El importe de proteínas a través del complejo del poro nuclear comienza cuando una importina específica se une a la señal de localización nuclear de una proteína transportadora en el citoplasma Algunas proteínas permanecen en el interior del núcleo una vez transportadas desde el citoplasma, pero muchas otras viajan continuamente entre el núcleo y el citoplasma. Algunas de estas proteínas actúan como transportadores (carriers) de otras moléculas, como los ARN; otras coordinan las funciones nucleares y citoplasmáticas (p. ej., regulando la actividad de los factores de transcripción). Las proteínas se etiquetan para ser exportadas del núcleo mediante una secuencia de aminoácidos específica, llamada señal de exportación nuclear. Al igual que las señales de localización nuclear las señales de exportación nuclear son reconocidas por receptores en el interior del núcleo exportinas, que dirigen el transporte de las proteínas a través del complejo del poro nuclear al citoplasma. Al igual que las importinas, muchas exportinas pertenecen a una familia de receptores de transporte nuclear denominados carioferinas Regulación del transporte de proteínas al núcleo Los factores de transcripción sólo son funcionales cuando están presentes en el núcleo, por lo que la regulación de su transporte al núcleo es otra forma de control de la expresión génica. En uno de los mecanismos de regulación, los factores de transcripción u otras proteínas) se asocian con proteínas citoplasmáticas que enmascaran las señales de localización nuclear; puesto que ya no se reconocen estas señales, las proteínas permanecen en el citoplasma Transporte de ARN Mientras que muchas proteínas son transportadas selectivamente desde el citoplasma al núcleo, la mayoría de los ARN son exportados desde el núcleo al citoplasma. Puesto que las proteínas se sintetizan en el citoplasma, la salida de los ARNm, ARNr, ARNt y microARN (ARNmi) es un proceso fundamental en la expresión génica en las células eucariotas. Las importinas del tipo carioferina y las exportinas transportan la mayoría de ARNt, ARNr, ARNmi y los ARN pequeños nucleares en un modo dependiente de Ran/GTP. Los ARNm son exportados por un complejo de dos proteínas (el «exportador del ARNm. Este transporte de ARNm parece ser independiente de Ran Organización interna del núcleo En las células animales una matriz laxa de lamininas nucleares se extiende desde la lámina nuclear hacia el interior del núcleo. Estas lamininas sirven como puntos de unión para la cromatina y organizan otras proteínas en cuerpos nucleares. La cromatina en el interior nuclear se organiza en grandes lazos de ADN, y regiones específicas de estos lazos se encuentran unidas a la matriz de laminina mediante proteínas de unión a laminina presentes en la cromatina Nucléolo y procesamiento del ARNr La subestructura que más destaca en el núcleo es el nucléolo que es el sitio donde tiene lugar la transcripción y el procesamiento del ARNr, y el ensamblaje de los ribosomas. Las células necesitan una gran cantidad de ribosomas para satisfacer la necesidad de síntesis de proteínas. Por ejemplo, las células de mamífero en continuo crecimiento contienen entre 5 y 10 millones de ribosomas, que deben sintetizarse cada vez que la célula se divide. El nucléolo es una fábrica de producción de ribosomas, diseñada para cubrir las necesidades de producción a gran escala de los ARNr y de ensamblaje de las subunidades ribosómicas. Evidencias recientes sugieren que los nucléolos también poseen un papel más general en la modificación del ARN y que varios tipos de ARN entran y salen del nucléolo en estadios específicos de su procesamiento Ensamblaje de ribosomas