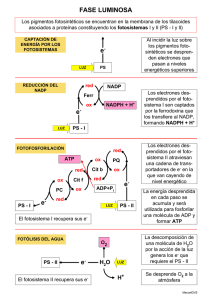
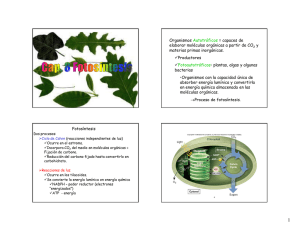
Integrantes: - Yahaira Clavijo - Sebastián Chile - David Chávez - Bryan Changuan Objetivos Aprender las etapas del proceso bioquímico mas importante de la atmósfera, responsable de la transformación de la energía luminosa en energía química, necesaria y utilizada por los seres vivos. Conocer el mecanismo bioquímico por el cual las plantas verdes, algas y algunas bacterias son capaces de transformar el dióxido de carbono y agua en carbohidratos y oxigeno. Estudiar las reacciones de la fase luminosa de la fotosíntesis Antecedentes Fosforilación oxidativa Las mitocondrias Cuatro complejos generan la mayor NADH-Q parte de ATP oxidoreductasa, mediante la succinato-Q acción combinada reductasa, Qde las reacciones citocromo c del ciclo del ácido oxidorreductasa, cítrico y la y citocromo c fosforilación oxidasa. oxidativa. Cuatro complejos Aproximadamente NADH-Q 30 moléculas de oxidoreductasa, ATP se generan succinato-Q cuando una reductasa, Qmolécula de citocromo c glucosa se oxida oxidorreductasa, por completo a y citocromo c CO2 y H2O oxidasa. Fotosíntesis Introducción • 60% es realizada por algas y bacterias La radiación electromagnét ica → energía química Autótrofos • Recuperar parte de esta energía de la glucosa sintetizada a través de la vía glucolítica y el metabolismo aeróbico Reacciones luminosas • Poder reductor y ATP Reacciones • Ciclo de oscuras Clavin o RIL La fotosíntesis transforma la energía lumínica en energía química Fotosíntesis Respiración celular Generar e− de alta energía Generar una fuerza protónmotriz Transformar esta fuerza en ATP Se requieren e− de alta energía Formar NADPH Generar ATP ¿Cómo podemos generar electrones de alta energía sin utilizar combustible químico? Impulsar los e− desde un estado de baja energía a uno alto Se pueden capturar los electrones excitados Producir el poder reductor Cadena de transporte de e− y una fuerza protón-motriz Síntesis de ATP La luz es absorbida y la energía se utiliza para conducir electrones del agua para generar NADPH y para conducir protones a través de una membrana. Estos protones regresan a través de la ATP sintasa para producir ATP Fotosíntesis en plantas Fotosistema I Forma NADPH Deficiente e− Se genera un gradiente de protones transmembrana Fotosistema II Oxida el agua Síntesis de ATP Reemplaza los e− perdidos Cloroplastos Estroma Tilacoides Grana Lamelas del estroma La fotosíntesis tiene lugar en los cloroplastos • Transcurren las reacciones fotosintéticas • Da lugar a las reacciones oxidoreducción generando la fuerza protónmotriz • Agrupación de tilacoides • Regiones de la membrana tilacoidal, conecta los diferentes grana Los procesos iniciales de la fotosíntesis tiene lugar en las membranas tilacoidales Proteínas captadoras de luz, centros de reacción, cadena de transporte de electrones, ATP sintasa 40% galactolipidos, 4% sulfolipidos, 10% fosfolipidos Impermeable a moléculas e iones Estroma: enzimas que usan el NADPH y ATP para transformar CO2 en azúcar Las células de las hojas contienen entre 1 y 100 cloroplastos Los cloroplastos surgieron de un proceso endosimbiótico Contiene su propio ADN No son autónomos Procesos de endosimbiosis algas rojas y pardas provienen de otro suceso más Plantas superiores y algas verdes Ancestro de cianobacterias Genoma cloroplasto < cianobacteria Características comunes Relación dependiente Anabaena La absorción de la luz por la clorofila induce la transferencia de electrones Clorofila a Hidrofóbico y esterificado Tetrapirrol sustituido Un fitol(V) Un anillo pirrol reducido (R) un anillo de 5Cfusionad o al anillo(A) Son Polienos Absorben energía solar más rápidamente Coeficiente máximo de extinción molar: 𝜀 = 105 𝑀−1 𝑐𝑚−1 ¿Qué sucede cuando una molécula de pigmento tal como la clorofila absorbe la luz? La energía de la luz excita a un e− desde su nivel basal a un nivel energético superior Centro de reacción Maximizar la separación y dificultar el retorno Retorna a su estado basal Puede pasar desde la molécula inicial al aceptor de e− Puede producir otros compuestos Separación de forma inducida Un par especial de clorofilas inicia la separación de cargas Centro de reacción fotosintético bacteriano (Rhodopseudomonas viridis) Subunidad C •Un tipo de citocromo c con cuatro hemos tipo c •Localización: cara externa membrana celular Subunidad L Subunidad M •Constituyen el núcleo estructural •Se les asocia: 4 moléculas Bacterioclorofila b (BChl-b) 2 moléculas Bacteriofeofitina b (BPh) 2 quinonas (Q_A y Q_B) Subunidad H •Localización: citoplasma de la membrana Un par especial de clorofilas inicia la separación de cargas Bacterioclorofilas Son fotoreceptores similares a las clorofilas Presenta en su centro dos protones en ves de un ion magnesio El par de moléculas de BChl-b se denomina par especial debido a su papel fundamental en la fotosíntesis. Un par especial de clorofilas inicia la separación de cargas Absorción por el dímero L-M de dos moléculas de BChl-b • El par especial presenta un máximo de absorción a 960nm Transferencia de un electrón • Desde P960 hacia BPh Aparición de cargas • Carga positiva en P960 y negativa en BPh QA transfiere rápidamente el electrón lejos de BPh • Antes de que el electrón vuelva a P960 para neutralizar las cargas Un par especial de clorofilas inicia la separación de cargas Para evitar la recombinación de cargas estas se transfieren de BPh a QA y de P960 a uno de los hemos de la subunidad citocromo. Captura de dos protones del citoplasma •Contribuye a la formación del gradiente de protones a través de la membrana celular La reducción completa de QB desde Q a QH2 •Tiene lugar tras la absorción de un segundo fotón y la transferencia de un segundo electrón desde un par especial QA pasa el electrón a QB El flujo cíclico de electrones reduce el citocromo de centro de reacción QH2 entra en la reserva Q QH2 se reoxida a Q por el complejo bc1 Complejo bc1 transfiere electrones hasta Cit C2 Estos electrones fluyen hacia la subunidad citocromo El gradiente de protones generado en el curso de este ciclo promueve la generación de ATP mediante la acción de la ATP sintasa EN LA FOTOSINTESIS PRODUCTORA DE OXÍGENO, DOS FOTOSISTEMAS GENERAN UN GRADIENTE DE PROTONES Y NADPH La fotosíntesis depende de la actuación conjunta de dos complejos fotosensibles PS I par especial P700, utiliza electrones para crear NADPH PS II par especial P680, se genera O2 y electrones, procedentes de dos moléculas de agua El citocromo bf genera un gradiente de protones El fotosistema II transfiere electrones del agua a la plastoquinona y genera un gradiente de protones Oxidada (Q), Reducida (𝑄𝐻2 ) 2𝑄 + 2𝐻2 𝑂 𝐿𝑢𝑧 𝑂2 + 2𝑄𝐻2 Fotosíntesis II. Plastoquinona pasa de una forma oxidada a reducida (plastoquinol) Parecido al centro de reacción bacteriano Subunidades adicionales se unen a más de 30 moléculas de clorofila El fotosistema II transfiere electrones del agua a la plastoquinona y genera un gradiente de protones Flujo electrónico análogo al que ocurre en el sistema bacteriano Plastoquinona unida al centro QA Plastoquinona desplazable unida al centro QB P680 es un oxidante muy fuerte que extra electrones de las moléculas de agua unidas al centro de manganeso El fotosistema II transfiere electrones del agua a la plastoquinona y genera un gradiente de protones El centro de manganeso, en su estado reducido, oxida dos moléculas de agua para generar una molécula de oxígeno. Los cuatro electrones extraídos del agua se utiliza para reducir dos moléculas de Q hasta QH2 El fotosistema II transfiere electrones del agua a la plastoquinona y genera un gradiente de protones Sentido de gradientes de protones Estroma. Dos protones capturados por cada molécula de plastoquinona Lumen. Cuatro protones generados en la oxidación del agua Se genera un gradiente de protones Más protones en el lumen que en el estroma El citocromo bf conecta el fotosistema II al fotosistema I ē fluyen del F.S.I al F.S.II Complejo citocromo bf. Cataliza transferencia de ē del QH2 a la PC • El plastoquinol (QH2) se oxida a plastoquinona (Q), un ē a la vez. • ē →Fe-S→ plastocianina (Pc) a su forma reducida. • Citocromo bf reduce una segunda molécula Q→QH2 , tomando 2 protones de un lado de la membrana para reoxidar el QH2 y liberar los protones al otro lado . • Los protones se liberan en el lumen del tilcoide y se toman del estroma (↑∆p). Fotosistema I utiliza la energía de la luz para generar ferredoxina reducida, un potente reductor Fotosistema I, cataliza etapa final de las reacciones de la fase luminosa ē se transfiere a la ferredoxina→ ē NADP+ Par de moléculas de clorofila a P700(+) inicia separación de carga fotoinduciada. Se neutraliza mediante la transferencia de ē La ferredoxina-NADP+ reductasa convierte el NADP+ en NADPH Ferredoxina 2 moléculas de ferredoxina Porta 1 ē disponible NADPH Reductor con 2 ē disponibles Usado en procesos biosintéticos Grupo prostético Estroma de la membrana (∆p) La cooperación entre fotosistema I y fotosistema II genera un flujo de electrones desde el H2O al NADP+ La síntesis de ATP es impulsada por un gradiente de protones a través de la membrana tilacoide • Jagendorf demostró que los cloroplastos sintetizan ATP en la obscuridad, →∆ pH en la membrana tilacoide. • fuerza protón-motriz → formación de ATP. • Se liberan protones en el tilacoide del lumen o se captan del estroma (∆p) . • Membrana tilacoide es impermeable a los protones. Pero es permeable a Cl- y Mg2+ (neutralidad eléctrica). La ATP sintasa de los cloroplastos se parece mucho a la mitocondrial y a la procariótica Fuerza protón-motriz → ATP Complejo CF1-CF0 CF0 conduce protones a través de la membrana tilacoidal CF1 cataliza la formación de ATP Los protones fluyen del lumen a través del enzima hacia el estroma o matriz donde se sintetiza ATP. El flujo cíclico de electrones a través del fotosistema I da lugar a la producción de ATP en vez de NADPH Relación elevada NADPH – NADP+ Existe otra fuente de ē para generar NADPH NADP+ no acepta ē de la ferredoxina reducida ē (P700) siguen una vía alterna que no termina en NADPH FOSFORILACIÓN CÍCLICA LA ABSORCIÓN DE 8 FOTONES PRODUCE 1 MOLÉCULA DE O2, 2 NADPH Y 3 ATP Absorción 4 fotones 1 molécula O2 4 protones (lumen tilacoide) Mediante ciclo Q del complejo citocromo bf Oxidan 2 plastoquinol Liberan 8 protones (lumen) Absorción 4 fotones + LA ABSORCIÓN DE 8 FOTONES PRODUCE 1 MOLÉCULA DE O2, 2 NADPH Y 3 ATP ê de 4 moléculas plastocianina 4 moléculas ferredoxina 2 moléculas NADPH + + 2𝐻2 𝑂 + 2𝑁𝐴𝐷𝑃+ + 10𝐻𝑒𝑠𝑡𝑟𝑜𝑚𝑎 → 𝑂2 + 2𝑁𝐴𝐷𝑃𝐻 + 12𝐻𝑙𝑢𝑚𝑒𝑛 12 protones liberados en el lumen fluyen a través de la ATP sintasa 2𝑁𝐴𝐷𝑃+ + 3𝐴𝐷𝑃−3 + 3𝑃𝑖 −2 + 𝐻 → 𝑂2 + 2𝑁𝐴𝐷𝑃𝐻 + 3𝐴𝑇𝑃−4 + 𝐻2 𝑂 LA ABSORCIÓN DE 8 FOTONES PRODUCE 1 MOLÉCULA DE O2, 2 NADPH Y 3 ATP 8 fotones 3 moléculas ATP Fotofosforilación cíclica, forma productiva de generar ATP Absorción 4 fotones 8 protones (lumen) 2 moléculas ATP LOS PIGMENTOS AUXILIARES CANALIZAN LA ENERGÍA HACIA LOS CENTROS DE REACCIÓN Primera Segunda - Moléculas clorofila a, longitudes de onda específicas. - 450nm y 650nm - Fotones absorbidos, pasan al cloroplasto sin ser absorbidos. LA TRASNFERENCIA DE ENERGÍA POR RESONANCIA PERMITE QUE ESTA SE DESPLACE DEL LUGAR INICIAL DE ABSORCIÓN AL CENTRO DE REACCIÓN ¿Cómo se conduce la energía desde un pigmento auxiliar al centro de reacción? Transferencia de energía por resonancia Transferencia de energía por resonancia LOS COMPLEJOS CAPTADORES DE LUZ CONTIENEN CLOROFILAS Y CAROTENOIDES Clorofila b y carotenoides Clorofila b Carotenoides • Captación de la luz • Conducción de la energía hacia el centro de reacción • Diferencia de la clorofila a, grupo formilo en lugar de metilo • 450nm y 500nm • 400nm y 500nm • Función protectora COMPONENTES DE LA FOTOSÍNTESIS ESTAN MUY ORGANIZADOS Membrana tilacoidal Superpuestas (apiladas) Fotosistema II No superpuestas (sin apilar) Directamente con el estroma del cloroplasto Fotosistema I y ATP sintasa ¿Cuál es el significado funcional de esta diferenciación lateral del sistema membranoso de los tilacoides? MUCHOS HERBICIDAS INHIBEN LAS REACCIONES DE LA FASE LUMINOSA DE LA FOTOSÍNTESIS Interfieren con funciones del fotosistema II o I Fotosistema II: bloquean flujo de electrones Fotosistema I: desvían electrones de su destino final Inhibidores fotosistema II derivados de la urea (diurón) y triazina (atrazina) Inhibidor fotosistema I: paraquat LA CAPACIDAD DE TRANSFROMAR LA ENERGÍA LUMINOSA EN ENERGÍA QUÍMICA ES MUY ANTIGUA Moléculas con azufre reducido H2S, dadores de electrones 𝑪𝑶𝟐 + 𝟐𝑯𝟐 𝑺 → 𝑪𝑯𝟐 𝑶 + 𝟐𝑺 + 𝑯𝟐 𝑶 Fotosíntesis evolucionó en el dominio Bacteria Ubiquinona-citocromo c oxidorreductasa y citocromo bf, presentes en cadena de transporte de e respiratoria y fotosintética CONCLUSIONES Los electrones generan el poder reductor, una fuerza protón-motriz lo que conduce a la síntesis de ATP La fase luminosa se divide en dos fotosistemas muy importantes ya que si uno de estos se para o cambia cambiaría por completo la fase luminosa de la fotosíntesis Las observaciones del sistema de las bacterias fotosintetizadoras serán aplicables a los sistemas de las plantas El fotosistema II incrementa la absorción de energía y su transferencia al centro de reacción La cooperación entre fotosistema I y fotosistema II genera un flujo de electrones desde el H2O al NADP+ Los productos de las reacciones de la fase luminosa se disponen o se encuentran en el lugar correcto para que posteriormente ocurran las reacciones de la fase oscura Los pigmentos auxiliares canalizan la energía hacia los centros de reacción mediante resonancia y aumentan la eficacia de la captación de luz Los fotosistemas presentan características estructurales comunes que sugieren un origen evolutivo común, por lo tanto el aparato fotosintético evoluciono a partir de un sistema ancestral de traducción de energía ARTÍCULO CIENTÍFICO REVISTA AÑO FACTOR DE IMPACTO INSTITUCIÓN AUTOR ARTÍCULO CIENTÍFICO • International Journal of Clinical and Experimental Medicine • 15 Agosto 2015 • 9, 1277 • Escuela China de Medicina, Universidad Beijing de Medicina • Zhi-Rong Sun, Nan-Nan Zhu, Li-Li Cheng TEMA Comparación de la fotosíntesis y los parámetros fluorescentes entre Dendrobium officinale y Dendrobium loddigesii. OBJETIVO GENERAL - Investigar y comparar la fotosíntesis y parámetros fluorescentes entre Dendrobium officinale y Dendrobium loddigesii, para proporcionar información útil para el cultivo artificial de estos cultivos. OBJETIVO ESPECÍFICO - Analizar la fotosíntesis y las características de la clorofila de fluorescencia para evaluar la capacidad fotosintética de la planta y la eficiencia. - Investigar la relación entre las características de crecimiento y desarrollo y las características fotosintéticas, junto con los estudios anteriores. - El contenido de clorofila es importante para adaptarse al medio ambiente y utilizar los recursos. - Los contenidos globales de clorofila a y la clorofila b fueron: D. loddigesii > D. officinale RESULTADOS RESULTADOS Tasa fotosintética y la frecuencia respiratoria - Tasa de fotosíntesis aparente ( APR) fue el principal factor para reflejar la capacidad fotosintética . - APR general de D. loddigesii > Dendrobium officinale - Tasa de fotosíntesis neta ( NPR) denotan la capacidad de acumulación de materia orgánica en planta. - NPR de D. loddigesii >D. officinale - La frecuencia respiratoria de D. loddigesii < D. officinale La actividad química del fotosistema ↑ en el consumo de oxígeno en PSI (8-10 meses) en comparación con la de 6 meses en D. loddigesii Rolfe. D. officinale Kimura ↑ el oxígeno consume en PSI. el consumo de oxígeno en PSI en el D. officinale > D. loddigesii Rolfe (8-10 meses), la transmisión de electrones elevada en el D. officinale Kimura. La liberación de oxígeno en PSII de D. officinale Kimura > D. loddigesii Rolfe en cada mes. Conclusiones - En este estudio, no se observaron diferencias en los parámetros de fotosíntesis y fluorescencia de la plántula de D. loddigesii y D. officinale. - Este estudio podría proporcionar información útil para el cultivo y el uso de estos cultivares en la medicina tradicional china.